
INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
1
1. INTRODUÇÃO E OBJECTIVOS
A enzima β-galactosidase (EC.3.2.1.23), comummente conhecida como
lactase, é a enzima que catalisa a hidrólise dos resíduos terminais não redutores
dos resíduos β-D-galactose em β-D-galactosideos. Convencionalmente, a sua
aplicação principal é a hidrólise da lactose de leite e derivados, particularmente
soro de leite. Recentemente, têm sido extensivamente estudadas β-
galactosidases com actividade transglicolítica para a produção de alimentos
funcionais (1,2,3).
São muitos os organismos que sintetizam naturalmente a β-
galactosidase, incluído microorganismos, plantas e células animais.
Normalmente, são mais utilizadas em indústria as enzimas extraídas de
Aspergillus spp e de Kluyveromyces spp. uma vez que são obtidas a partir de
culturas destes microrganismos com produtividade e rendimento aceitáveis,
tendo a vantagem de que os produtos são “geralmente reconhecidos como
seguros” (“GRAS”) para consumo humano, o que é crítico para aplicações
relacionadas com alimentação (4).
Uma das principais razões que tem levado ao estudo intensivo desta
enzima e das suas diversas aplicações é a possibilidade da obtenção de
alimentos sem lactose e que, por isso, possam ser consumidos por pessoas
intolerantes a este dissacarídeo, sem prejuízo do restante aporte nutricional
benéfico fornecido por tais alimentos quando inseridos numa alimentação
equilibrada.
1.1. Lactose
A lactose (figura 1), juntamente com a maltose e a sacarose, é um dos
dissacarídeos fisiologicamente mais importantes. Os dissacarídeos são açúcares
formados por dois resíduos de monossacarídeos unidos por uma ligação

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
2
glicosídica. Os seus nomes químicos baseiam-se nos seus monossacarídeos
constituintes.
A principal fonte de lactose é o leite, numa percentagem que varia entre 4,2
e os 4,9 deste açúcar (5) (tabela 1).
Figura 1. Molécula de lactose
Tabela 1. Percentagem de lactose em leite e derivados (5)
Alimento Quantidade de lactose (g/100 g
de alimento) Leite de vaca esterilizado gordo 4,5
Leite de vaca esterilizado meio-gordo 4,7
Leite de vaca esterilizado magro 4,7
Leite de vaca pasteurizado gordo 4,7
Leite de vaca UHT gordo 4,7
Leite de vaca UHT meio-gordo 4,9
Leite de vaca UHT magro 4,9
Leite de cabra cru 4,6
Leite de ovelha cru 4,2
Leite humano 7,5
Nata maturada pasteurizada com 30% de gordura 2,7
Queijo flamengo com 30% ou 45% de gordura 0,2
Requeijão com 13% de proteína 5,1
Queijo fresco 1,5
Iogurte natural sólido meio-gordo 5,0
Iogurte açucarado batido meio-gordo 6,4
Iogurte líquido meio-gordo 3,9
A solubilidade e o poder edulcorante da lactose são baixos quando
comparado com outros açúcares, incluindo a glucose, galactose, frutose e
sacarose (6). Este açúcar higroscópico tem uma forte tendência para absorver
sabores e odores e tem um efeito laxativo quando consumido em grandes
quantidades. Contudo, a hidrólise da lactose diminuí os problemas de
precipitação e aumenta o seu poder edulcorante (4).

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
3
A hidrólise da lactose em glucose e galactose pela β-galactosidase é um
processo importante na indústria alimentar devido ao seu potencial efeito
benéfico na assimilação de produtos que contêm lactose por pessoas com
dificuldade ou intolerância (7,8). Tem também aplicações tecnológicas e
industriais, tais como a elaboração de leite com sabor doce, de boa
aceitabilidade, e que pode ser consumido por indivíduos intolerantes à lactose,
formação de galactooligossacáridos através de reacções de transglicosilação
durante a hidrólise da lactose que devido ao seu efeito pré-biótico favorecem o
crescimento de bactérias benéficas da microflora intestinal (1,9); melhoramento
nas características tecnológicas e sensoriais de alimentos que contêm lactose
hidrolisada proveniente de leite ou soro de leite, tais como aumento da
solubilidade (evitando a cristalização da lactose e o aspecto arenoso em gelados
e produtos condensados ou em pó) e aumento do poder edulcorante com
menor valor calórico dos produtos. Muitos produtos com baixo teor em lactose
(como por exemplo: leite sem lactose ou com percentagem reduzida de lactose,
gelados elaborados com leite sem lactose, queijos curados) têm aplicações na
alimentação humana e alimentação animal (10).
Para além do leite, a principal fonte de lactose é o soro de queijo, que é o
principal subproduto da indústria queijeira. Cerca de 9 litros de soro de queijo
são gerados durante a produção de 1 Kg de queijo (4). Estima-se que todos os
anos 145 x 106 toneladas de soro de queijo e 6 x 106 toneladas de lactose sejam
produzidos mundialmente. Infelizmente, nos últimos cinquenta anos, metade
da produção anual não foi transformada em subprodutos mas deitados
directamente em habitats aquosos. Esta situação cria enormes problemas
ambientais e necessita de novas estratégias para a recuperação de subprodutos
(11). A carga orgânica do soro de queijo é elevada principalmente devido ao seu
conteúdo em lactose, que, juntamente com as elevadas quantidade de soro de
queijo que são geradas, faz com que este subproduto se torne um problema
ambiental e são requeridas soluções para a sua revalorização. Deste modo, a

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
4
bioconversão da lactose do soro de queijo por β-galactosidase desempenha um
papel fundamental por ampliar as aplicações do soro de queijo reduzindo mais
de 75% a poluição e gerando produtos de interesse para alimentação animal,
nutrição humana e confeitaria, uso farmacêutico e agrícola (4,11).
1.2 Intolerância à lactose
A intolerância à lactose é mais comum a um hidrato de carbono e pode
afectar pessoas de todos os grupos etários, apesar de ocorrer com mais
frequência em idades avançadas. A intolerância à lactose é causada pela
deficiência em lactase (β-galactosidase), a enzima que digere o açúcar do leite, a
lactose (12).
No organismo humano, a β-galactosidase está localizada nas
microvilosidades do intestino delgado (13). Devido aos baixos níveis da enzima
no intestino, grande parte da população demostra intolerância à lactose e, em
consequência, tem dificuldade em digerir leite e lacticínios (14).
A lactose é um dissacarídeo formado por glucose e galactose que se
encontra em grande abundância no leite. Nos seres humanos, a intolerância à
lactose ou a má-absorção de lactose é um problema comum. Está estimado que
mais de 70% da população adulta no mundo tenha problemas a digerir a
lactose, resultado da reduzida ou falta de actividade da β-galactosidase no
intestino delgado (13). Geralmente, a actividade da enzima lactase diminui
exponencialmente ao longo da vida (15), atingindo cerca de 10% do valor à
nascença. Mesmo em adultos que retêm níveis elevados de lactase (75% - 85%
dos caucasianos adultos) a quantidade de lactase é cerca de metade quando
comparada com as outras sacaridases tais como a sucrase, a α-dextrinase ou a
glucoamilase (12).
A diminuição ou falta de lactase é, geralmente, denominada por
hipolactasia. Há três tipos de hipolactásia: hipolactásia primária, hipolactásia

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
5
secundária e alactásia congénita. A hipolactásia primária é a mais comum e
corresponde ao decréscimo dos níveis da enzima verificado ao longo da vida em
certas populações e indivíduos. Segue-se a hipolactásia secundária ou adquirida
que pode ter origem numa doença gastrointestinal que danificou as
microvilosidades ou que pode surgir como consequência de uma infecção do
intestino delgado, afecções inflamatórias, HIV ou malnutrição sendo que nas
crianças é geralmente secundária a infecções virais ou bacterianas. A alactásia
congénita é o tipo mais raro pois corresponde a ausência completa da produção
de lactose (12,13).
Em todos os casos, a lactose não é hidrolisada em galactose e glucose no
intestino delgado, passando para o cólon onde as bactérias da microflora do
cólon fermentam a lactose originando ácidos gordos de cadeia curta e gases,
dióxido de carbono e hidrogénio, causando sintomas como dor abdominal,
náuseas, diarreia e flatulência. A extensão destes sintomas é variável de
indivíduo para indivíduo. O consumo de pequenas quantidades poderá ser
tolerado por alguns indivíduos pois deverá ter poucas consequências uma vez
que os ácidos gordos de cadeia curta são prontamente absorvidos e os gases
podem ser absorvidos ou expelidos. Grandes quantidades, normalmente
superiores a 12 gramas, consumidas num único alimento (quantidade
tipicamente encontrada em 240 ml de leite) poderão resultar numa maior
entrada de substrato no cólon do que aquela que consegue ser eliminada por
um processo normal.
A actividade da lactase pode também ser retardada ou diminuída após
nutrição parentérica prolongada. A má digestão da lactose com todos estes
sintomas também pode ocorrer em adultos com o Síndrome do Cólon Irritável
ou em crianças com dores abdominais recorrentes.
A deficiência em lactase é geralmente diagnosticada com base em:
1. Historial dos sintomas gastrointestinais que ocorrem após ingestão
de leite;
INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
6
2. Teste para níveis anormais de hidrogénio no hálito;
3. Teste de anormal tolerância à lactose – o teste de tolerância à
lactose foi originalmente baseado numa dose oral de lactose de 50g.
Recentemente, doses menores do que 50g de lactose foram usadas de modo a
aproximar o real consumo de lactose em lacticínios (12,16).
Os sintomas da intolerância à lactose são aliviados pela redução do
consumo de alimentos que contenham lactose. Uma dieta completamente
isenta de lactose poderá não ser necessária nas pessoas com deficiência em
lactase uma vez que o grau de intolerância à lactose é muito variável de
individuo para individuo e a maioria dos pacientes não requer uma total evicção
da lactose da sua dieta. Assim sendo, os produtos lácteos não deverão ser
totalmente eliminados da alimentação uma vez que estes fornecem nutrientes
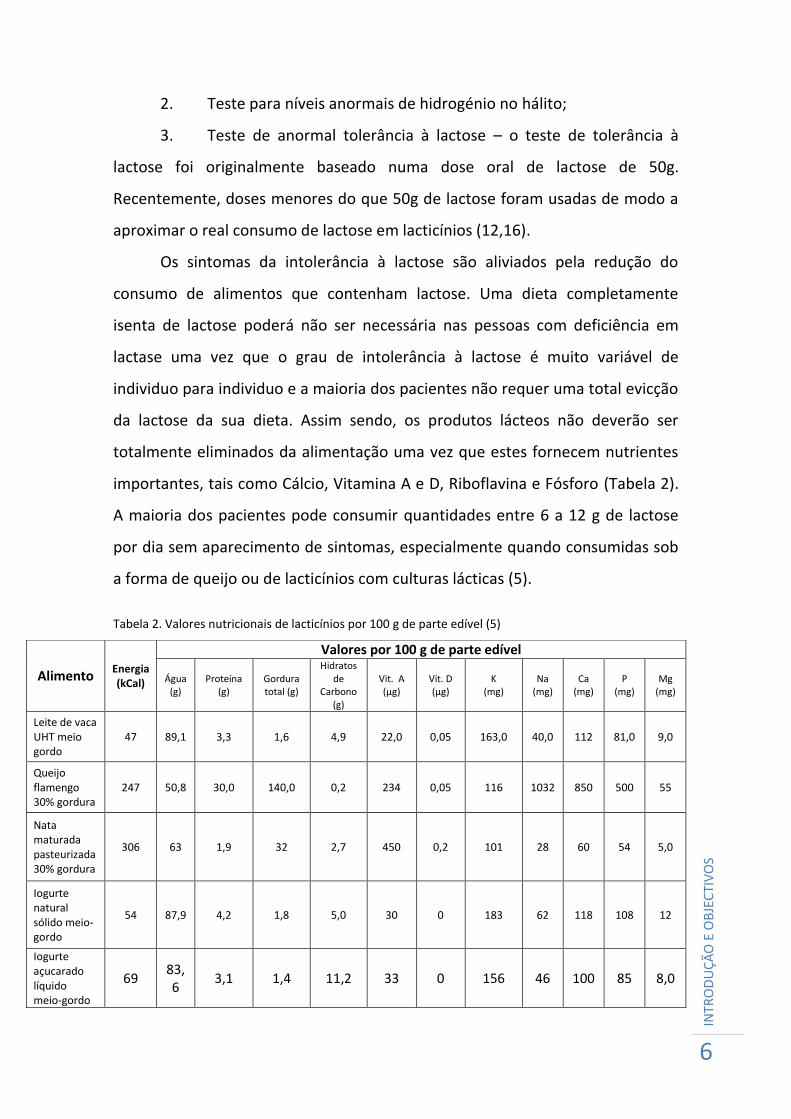
importantes, tais como Cálcio, Vitamina A e D, Riboflavina e Fósforo (Tabela 2).
A maioria dos pacientes pode consumir quantidades entre 6 a 12 g de lactose
por dia sem aparecimento de sintomas, especialmente quando consumidas sob
a forma de queijo ou de lacticínios com culturas lácticas (5).
Tabela 2. Valores nutricionais de lacticínios por 100 g de parte edível (5)
Alimento Energia (kCal)
Valores por 100 g de parte edível
Água (g)
Proteína (g)
Gordura total (g)
Hidratos de
Carbono (g)
Vit. A (µg)
Vit. D (µg)
K (mg)
Na (mg)
Ca (mg)
P (mg)
Mg (mg)
Leite de vaca UHT meio gordo
47 89,1 3,3 1,6 4,9 22,0 0,05 163,0 40,0 112 81,0 9,0
Queijo flamengo 30% gordura
247 50,8 30,0 140,0 0,2 234 0,05 116 1032 850 500 55
Nata maturada pasteurizada 30% gordura
306 63 1,9 32 2,7 450 0,2 101 28 60 54 5,0
Iogurte natural sólido meio-gordo
54 87,9 4,2 1,8 5,0 30 0 183 62 118 108 12
Iogurte açucarado líquido meio-gordo
69 83,6
3,1 1,4 11,2 33 0 156 46 100 85 8,0

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
7
Os indivíduos que possuem intolerância à lactose deverão ser informados
de que isto não significa que sejam alérgicos ao leite ou lacticínios, uma vez que
este tipo de alergia está, geralmente, relacionado com a parte proteica destes
produtos.
Dado isto, há um mercado considerável para leite e derivados sem
lactose, que podem ser obtidos por hidrólise enzimática usando β-galactosidase
(4).
1.3. β-galactosidase
A enzima β-galactosidase é a enzima que hidrolisa a lactose nos seus
monómeros, glucose e galactose (Figura 2), sendo por esse motivo, também
denominada por lactase.
Figura 2. Hidrólise da lactose em glucose e galactose
A enzima β-galactosidase (Figura 3) pode ser obtida a partir de uma
grande variedade de fontes, tais como microrganismos, plantas e animais. No
entanto, de acordo com a sua origem, as suas propriedades diferem,
essencialmente, no que respeitante às condições óptimas de aplicação da
enzima, sobretudo na gama de pH (17). Enzimas de plantas e animais têm pouco
valor comercial mas várias fontes microbiológicas de β-galactosidase são de
grande interesse tecnológico. Os microrganismos oferecem várias vantagens
relativamente às outras fontes tais como fácil manuseamento, maior taxa de
multiplicação e maior produção. Como resultado do interesse comercial na β-

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
8
galactosidase, um grande número de microrganismos têm sido testados como
fontes potenciais desta enzima (14,18).
Figura 3. β-galactosidase (http://www.sprintechnologies.com/sprin/index.jsp?idPagina=44)
1.3.1. Fontes microbianas da enzima
1.3.1.1. Enzimas bacterianas
A enzima β-galactosidase pode ser produzida por um vasto número de
bactérias, mas as bactérias Streptococcus thermophilus e Bacillus
stearothermophilus são consideradas as fontes primordiais bacterianas desta
enzima (14).
Apesar da enzima β-galactosidase proveniente de Escherichia coli ser a
mais extensivamente estudada, o seu uso industrial é dificultado pelo facto de a
sua aplicação não ser considerada segura em alimentos, ou seja, de este
microrganismo não possuir estatuto “GRAS”. No entanto, a β-galactosidase
proveniente desta bactéria está comercialmente disponível para fins analíticos
(4).

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
9
As bactérias ácido-lácticas (incluindo o grupo do lactococci,
streptococci e lactobacilli) e as bifidobactérias (tabela 3), são organismos
reconhecidos como seguros, ou seja, que possuem estatuto “GRAS” e que foram
identificados como boas fontes de β-galactosidase, especialmente para
aplicação em alimentos funcionais (4,19,20).
1.3.1.2. Enzimas de fungos
A gama de pH óptimo das enzimas fúngicas é de 2,5 a 5,4, o que faz com
que estas enzimas sejam apropriadas para o processamento de soro de leite
ácido. A temperatura óptima para estas enzimas é alta e podem ser tipicamente
usadas a temperaturas superiores a 50ºC.
A principal fonte fúngica desta enzima é Aspergillus niger (14) (tabela 3).
A purificação de β-galactosidase de diferentes origens fúngicas tem sido
realizada através de variadas técnicas de purificação, tais como peneiramento
molecular, permuta iónica e técnicas cromatográficas.
1.3.1.3. Enzimas de leveduras
As leveduras têm sido consideradas fontes importantes de β-
galactosidase do ponto de vista industrial.
Com um pH óptimo próximo da neutralidade, as enzimas provenientes de
leveduras são bastante apropriadas para hidrólise de lactose no leite, e são
vastamente aceites como seguras para uso em alimentos (21). Tem sido
realizada muita investigação relativamente à produção de β-galactosidase de
diferentes estirpes de leveduras devido ao seu potencial (22).
As principais leveduras utilizadas para a produção de β-galactosidase são
Kluyveromyces lactis, Kluyveromyces marxianus e Kluyveromyces fragilis (14)
(Tabela 3).

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
10
Tabela 3. Fontes microbianas de β-galactosidase (14)
Fonte Microorganismo
Bactérias
Alicyclobacillus acidocaldarius subsp. Rittmannii
Arthrobacter sp.
Bacillus acidocaldarius, B. circulans, B. coagulans, B.subtilis, B. megaterum, B. stearothermophilus
Bacteriodes polypragmatus
Bifidobacterium bifidum, B. infantis
Clostridium acetobutylicum, C. thermosulfurogens
Corynebacterium murisepticum
Escherichia coli
Klebsiella pneumoniae
Lactobacillus acidophilus, L. bulgaricus,L. helviticus, L.kefiranofaciens, L. lactis, L. sporogenes, L. themophilus, L.delbrueckii
Leuconostoc citrovorum
Pediococcus acidilacti, P. pento
Propioionibacterium shermanii
Pseudomonas fluorescens
Pseudoalteromonas haloplanktis
Streptococcus cremoris, S. lactis, S. thermophius
Sulfolobus solfatarius
Thermoanaerobacter sp.
Thermus rubus, T. aquaticus
Trichoderma reesei
Vibrio cholera
Xanthomonas campestris
Fungos
Alternaria alternate, A. palmi
Aspergillus foelidis, A. fonsecaeus, A. fonsecaeus, A. Carbonarius, A. Oryzae
Auerobasidium pullulans
Curvularia inaequalis
Fusarium monilliforme, F. oxysporum
Mucor meihei, M. pusillus
Neurospora crassa
Penicillum canescens, P. chrysogenum, P. expansum
Saccharopolyspora rectivergula
Scopulariapsis sp
Streptomyces violaceus
Leveduras
Bullera singularis
Candida pseudotropicalis
Saccharomyces anamensis, S. lactis, S. fragilis
Kluyveromyces bulgaricus, K. fragilis, K. lactis, K. marxianus
Tradicionalmente, as β-galactosidase mais utilizadas na indústria são as
provenientes de Aspergillus spp e de Kluyveromyces spp porque podem ser
obtidas prontamente e com elevado rendimento de culturas destes

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
11
microrganismos. Adicionalmente, produtos obtidos destes microrganismos são
“GRAS” para consumo humano o que é crítico na aplicação à produção de
alimentos.
No Kluyveromyces spp, a β-galactosidase é intracelular. Devido à sua
natureza intracelular, a enzima necessita de ser extraída da célula da levedura
pela disrupção da célula ou pela permeabilização da célula através de
substâncias químicas e/ou tratamentos mecânicos. São fontes fúngicas desta
enzima Kluveromyces lactis e Kluveromyces marximus (4). A lactose é primeiro
transportada para o interior da célula através de uma permease e depois
hidrolisada intracelularmente a glucose e galactose que seguirão a via glicolítica.
A enzima desta levedura tem um pH óptimo perto da neutralidade (6.0 –
7.0) e, como tal, tem uma ampla gama de aplicações, particularmente na
hidrólise de soro de queijo, que como já foi referido anteriormente, é o principal
subproduto da indústria queijeira.
A Lactozym 2600L, β-galactosidase utilizada neste estudo, é uma enzima
livre obtida a partir de Kluyveromyces lactis, disponibilizada comercialmente
pela Sigma-Aldrich®. Esta preparação enzimática encontra-se no estado liquido.
Será também utilizada a imibond galactosidase SPRIN®, uma preparação
de β-galactosidase proveniente de Kluyveromyces lactis, disponibilizada
comercialmente pela SPRIN® technologies, já imobilizada covalentemente numa
resina Amino Acrílica e que possuí um tamanho de partícula entre os 200-
500μm.
Kluyveromyces Lactis é um ascomiceto que pertence aos
endoascomycetales tendo sido anteriormente mal classificado como
Saccharomyces lactis e Kluyveromyces fragilis var. lactis. Tem sido objecto de
intensivo estudo taxonómico sendo que, hoje em dia, Kluyveromyces lactis é
considerado uma espécie única (23).
A sua propriedade de crescer na lactose como única fonte de carbono
distingue o K. lactis da maioria das outras leveduras e pode ser usada para

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
12
seleccionar isolados a partir de produtos da indústria de lacticínios. Por isso o
seu nome de levedura do leite ou láctea. No entanto, este microorganismo
pode usar uma variada gama de substratos para a respiração.
Possui um metabolismo respiratório fermentativo e a maioria das
estirpes pode crescer em fontes de carbono fermentáveis quando a via
respiratória não está acessível. No entanto, K. lactis é, aparentemente, um
aeróbio estrito, que não é capaz de crescer sob condições estritamente
anaeróbias.
A utilização de lactose é uma das principais características do K. lactis e
tem sido, por isso, bastante estudada. O metabolismo da lactose é induzido e
co-regulado com o metabolismo da galactose por um activador transcripcional,
Lac9p ou KlGal4p. Dois genes LAC4 e LAC12, que codificam a β-galactosidase e a
lactose premiase, respectivamente, são específicos para o metabolismo da
lactose (23).
A β-galactosidase de K. lactis (codificada pelo gene LAC4) tem um grande
interesse em biotecnologia na indústria alimentar e na reciclagem de soro de
queijo. A vantagem de K. lactis relativamente a A. Niger (microrganismo
também bastante utilizado como fonte desta enzima) é que a levedura K. lactis
produz maiores quantidades de unidades enzimáticas. Contudo, devido à sua
natureza intracelular, a produção industrial e a aplicação da β-galactosidase de
K.lactis são limitadas pelos custos associados ao processo extracção. Para
contornar este facto, duas estratégias têm sido adoptadas para a secreção de β-
galactosidase de K. lactis: usando leveduras de estirpes com capacidade de lise
espontânea ou através da utilização de sequências sinal de secreção heteróloga
para a produção extracelular. Nos dois casos, plasmídeos episomais são usados
para a clonagem e expressão de β-galactosidase de K. lactis recombinante (4).

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
13
1.3.2. Aplicação Industrial
Apesar da enzima β-galactosidase ter inúmeras aplicações na indústria
alimentar e dos lacticínios, a moderada estabilidade da enzima livre é uma das
limitações que impede a implementação geral de biocatalisadores à escala
industrial. Para a produção de leite sem lactose, por exemplo, a enzima β-
galactosidase pode ser adicionada directamente ao leite inteiro, mas após o
processo estar finalizado, a enzima pode ser desactivada pelo tratamento por
calor. Uma vez que a enzima livre não pode ser reutilizada, este procedimento
não é economicamente viável (14).
Apesar de ainda haver quem utilize a enzima livre para hidrolisar a
lactose, o alto custo da enzima e a sua moderada estabilidade fazem com a
imobilização da β-galactosidase seja uma área de grande interesse devido aos
seus potenciais benefícios. A utilização de técnicas de imobilização é de
importância significativa do ponto de vista económico porque permite a
reutilização da enzima, o seu uso em operações contínuas, aumenta a
estabilidade enzimática e facilita a separação da enzima do substrato no final do
processo (4, 24). No entanto, a utilização de enzimas imobilizadas apresenta
ainda algumas limitações, incluindo a baixa taxa de recuperação da actividade
enzimática, a perda gradual da enzima durante o processo reaccional e uma
elevada resistência na transferência de massa entre algumas enzimas
imobilizadas e os substratos (4).
1.4. Métodos de Imobilização Enzimática
Vários tipos de suportes e técnicas têm sido utilizados e investigados para
a imobilização desta enzima mas apenas alguns foram ampliados e menos ainda
foram aplicados a nível industrial ou semi-industrial (14). Aparentemente a

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
14
adequação do suporte varia de enzima para enzima e consoante o objectivo do
seu uso sendo necessário optimizar para cada sistema (25).
A imobilização de células ou enzimas é definida como “a confinação física
de enzimas ou células ou a sua localização numa região ou espaço definido com
a retenção da sua actividade catalítica, que podem ser utilizadas repetida e
continuamente” (26).
O processo de imobilização deverá ser suave o suficiente de modo a que
a enzima não se desnature durante o processo de imobilização (27). A enzima
imobilizada é mais facilmente manipulada e facilita a separação do produto,
minimizando ou eliminando a contaminação de proteína no produto (28).
A tecnologia de imobilização de β-galactosidase é um processo efectivo
para a hidrólise bem-sucedida da lactose e ultrapassa os problemas associados
aos custos da enzima solúvel (21,25). Todavia, existem problemas associados à
enzima imobilizada tais como a contaminação microbiológica e a aderência de
proteínas. Contudo, para processos a longo termo usando enzimas imobilizadas,
é indispensável proceder a lavagens e pasteurizações periódicas (4).
Em estudos anteriores (29, 30, 31) a enzima foi imobilizada de diversos
modos, tais como:
1. Adsorção física;
2. Encapsulação;
3. Ligação covalente.
1.4.1. Adsorção física
A adsorção física (Figura 4) é considerada o método mais simples de
imobilização no qual uma enzima é imobilizada num transportador insolúvel em
água e o biocatalisador é mantido na superfície do transportador por forças
físicas (força de Van der Waals). Frequentemente, estão presentes forças

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
15
adicionais na interacção entre o transportador e o biocatalizador principalmente
interacções hidrofóbicas, pontes de hidrogénio e ligações heteropolares
(iónicas). Este método tem a vantagem de ser de simples realização e de ter
pouca influência na conformação do biocatalizador. No entanto, as
desvantagens desta técnica são a relativa fragilidade nas forças de ligação
adsortivas. Diferentes materiais de suporte inorgânicos (alumínio, sílica, vidro
poroso, terra diatomácea, argila, bentonite) e orgânicos (celulose, amido,
carvão activado e resinas iónicas como a amberlite, Sephadex, Dowex) podem
ser usados para adsorção enzimática. A adsorção da enzima também pode ser
estabilizada por tratamento com gluteraldeído (14,26).
1.4.2. A Encapsulação
A encapsulação é o método que visa colocar a enzima num invólucro de
pequenas dimensões (Figura 4). A encapsulação em membrana e em matriz
(incluindo microencapsulação) são os principais métodos de encapsulação. A
principal vantagem da técnica de encapsulação é a simplicidade pela qual as
partículas esféricas são obtidas pela extrusão de uma suspensão de polímero-
célula num meio contendo iões de carga positiva ou através de polimerização
térmica. Para além disso, as esferas formadas, particularmente as esferas de
alginato, são transparente e, normalmente, mecanicamente estáveis. A principal
limitação desta técnica para a imobilização de enzimas é a possível lenta perda
de enzima durante o uso continuado. Contudo, melhoramentos poderão ser
realizados através da utilização de procedimentos de ligação apropriados. As
matrizes usadas são normalmente feitas de polímeros tais como o alginato de
cálcio, agar, k-carregenina, poliacrilamida, quitosano e colagénio. No entanto,
algumas matrizes sólidas como o carvão activado e a terra de diatomácea,
também podem ser usadas para imobilização. As membranas normalmente

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
16
utilizadas para encapsulação de enzimas são nylon, celulose, polisulfone e
poliacrilamida. (14,26)
Este método é o utilizado neste estudo.
1.4.3. Ligação Covalente
No método de ligação covalente há uma retenção da enzima numa
superfície de suporte através da formação de uma ligação covalente. As enzimas
ligam-se a um suporte através de certos grupos funcionais tais como o grupo
amina, carboxilo, hidroxilo e sulfidrilo. Estes grupos funcionais não deverão
estar no centro activo da enzima. É frequente que se recomende que a
imobilização seja realizada na presença de substrato ou de um inibidor
competitivo para que se proteja o sítio activo num suporte. Os grupos
funcionais no material de suporte são normalmente activados através de um
reagente químico tal como brometo cianogenico, carbodimida e gluteraldeído.
Cascas de ovo partidas em pequenos pedaços podem ser bons transportadores
para imobilização de β-galactosidase devido ao seu baixo custo, boa resistência
mecânica e resistência a ataques microbianos (14,26) (Figura 4).
Figura 4. Esquema dos vários métodos de imobilização. (26)

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
17
1.5.Cinética Enzimática
A cinética enzimática é o estudo da velocidade de uma reacção química
que ocorre na presença de uma enzima. Permite compreender melhor os
pormenores do mecanismo catalítico das enzimas, o papel das enzimas no
metabolismo, o controlo da actividade e os mecanismos de inibição.
O estudo cinético de uma enzima visa primariamente caracterizar, ou
seja descrever, a actividade dessa enzima. In vitro estuda-se a actividade da
enzima procurando saber que tipo de reacções pode catalisar, com que
substratos pode interagir e como se modifica essa actividade (qualitativa e/ou
quantitativamente) quando se fazem variar as condições experimentais. O valor
de pH, a temperatura, o tempo de incubação, as concentrações dos substratos,
de cofactores ou de outras substâncias (inibidores ou activadores) são exemplos
de condições experimentais que podem ser modificadas com o objectivo de
observar a variação da actividade da enzima (32).
Para que uma reacção enzimática se dê, a enzima (E) e o substrato (S)
têm de se encontrar e formar o complexo enzima-substrato (ES). Este equilíbrio
é expresso sob a forma:
E+S ES
A enzima pode então transformar o substrato num produto (P). Existe de
forma transitória um complexo EP, havendo então dissociação deste complexo
em enzima livre (E) e produto (P). Este mecanismo pode escrever-se na forma
simplificada:
E+S [ES] E+P

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
18
A reacção de formação de ES é normalmente mais rápida que a reacção
de dissociação de EP, ou seja, a libertação do produto da reacção é
normalmente um passo limitante da reacção.
O estudo de uma reacção enzimática in vitro não reflecte a verdadeira
situação das enzimas em células, mas possibilita a simplificação da
determinação de parâmetros cinéticos. Dentro de uma célula, o substrato pode
estar confinado a um dado compartimento e não sofrer facilmente difusão: por
exemplo, pode ser o produto de uma reacção anterior numa via metabólica,
pelo que passa directamente de uma enzima para a outra.
Nos ensaios in vitro é utilizada uma concentração de substrato maior que
a concentração de enzima de modo a possibilitar a monitorização da variação da
velocidade de reacção. Como S está a ser convertido em P, S decresce com o
tempo, à medida que o produto se acumula em solução. Como a concentração
de substrato influencia a formação do complexo ES (e portanto da formação de
produto), vai influenciar também a velocidade da reacção (32,33).
Nestas condições, a velocidade inicial medida (V0) depende apenas da
concentração de enzima. Um gráfico representativo de V0 em função da
concentração de substrato apresenta uma forma característica de semi-
hipérbole: a baixas concentrações de substrato, a variação de V0 com a
concentração de substrato é linear; à medida que se usam contracções de
substrato mais elevadas, V0 sofre uma variação cada vez menor, tendendo para
um valor limite. Este valor é denominado velocidade máxima (Vmáx).
Os primeiros estudos da cinética enzimática com base científica foram
feitos por Leonor Michaelis e Maud Menten, em 1913, que propuseram pela
primeira vez um mecanismo para as reacções catalisadas enzimaticamente. As
suas concepções foram baseadas nos factos observáveis quando um substrato
sofre uma transformação sob a acção catalítica de uma enzima:

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
19
• a velocidade aumenta hiperbolicamente com a concentração do
substrato, sendo a reacção de ordem zero quando a concentração do substrato
é elevada e de 1ª ordem quando a concentração é baixa.
• a velocidade aumenta linearmente com a concentração de enzima,
para qualquer valor constante da concentração de substrato.
A semi-hipérbole correspondente ao comportamento da velocidade de
uma enzima em função da concentração do substrato pode ser descrita
matematicamente pela equação de Michaelis-Menten:
V0=Vmax [S]/(KM+[S])
em que [S], V0 e Vmax são as grandezas anteriormente definidas e KM a
constante de Michaelis. Esta equação pode ser deduzida de forma intuitiva
considerando a forma como se desenrola a catálise enzimática (33)
É assumida, nesta dedução, a existência de um estado estacionário, em
que a concentração do complexo enzima-substrato permanece constante ao
longo do tempo.
A constante Km é expressa em unidades de concentração e tem um
significado tal que quando a concentração do substrato iguala o seu valor a
velocidade da reacção é igual a metade de Vmax (i.e., [S]½ = KM => V = ½Vmax).
A teoria do estado estacionário, introduzida por G. E. Briggs e J. B. S.
Haldane em 1925, considera que a velocidade a que o complexo ES se forma é
aproximadamente igual à velocidade da sua dissociação. Existe um curto
período de tempo, normalmente na ordem dos microssegundos, em que há
uma acumulação de ES – estado pré-estacionário. O estudo da cinética
enzimática envolvendo a teoria do estado estacionário é referida como cinética
do estado estacionário (33).
Também é assumido que a concentração de produto é insignificante no
início da reacção, logo, não é dada importância à extensão da reacção inversa

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
20
no equilíbrio entre substrato e produto, S P, considerando-se apenas a reacção
directa na dedução da equação. Assim, a reacção é descrita por:
E+S [ES] E+P
Como já referido, a formação do complexo limita a velocidade da
reacção, medida como V0:
V0 = k2[ES]
Se a concentração do complexo enzima-substrato fosse facilmente
mensurável, o valor de k2 seria simples de determinar directamente. No
entanto, esse não é o caso, e o valor de concentração do complexo enzima-
substrato tem de ser deduzido. Para tal, considere-se que a concentração do
total de enzima (Et) presente na reacção é o somatório da concentração de
enzima em complexo com o substrato e da concentração de enzima "livre" (E)
(não ligada a substrato):
[E]t = [E]l + [ES]
Embora o gráfico obtido directamente da equação de Michaelis-Menten
seja de interpretação relativamente simples, existem tratamentos matemáticos
que simplificam a representação gráfica da equação e permitem a obtenção
rápida de parâmetros cinéticos.
O tratamento mais conhecido é o de Lineweaver-Burk. A equação de
Michaelis-Menten pode ser transformada numa equação da recta, do tipo
y=ax+b:

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
21
Este tipo de tratamento matemático permite uma determinação mais
precisa de Vmax, um parâmetro que só se obtém por aproximação num gráfico
de Michaelis-Menten.
1.5.1. Inibição Enzimática
Quando a presença de uma substância na solução provoca a diminuição
da velocidade da reacção essa substância designa-se por Inibidor e a sua acção
pode ser reversível ou irreversível. A inibição reversível é caracterizada por um
equilíbrio estabelecido muito rapidamente entre enzima e inibidor. Os principais
tipos de inibição reversível são:
• a inibição competitiva, ocorre quando o inibidor compete com o
normal substrato para se ligar ao sítio activo catalítico.
• a inibição não-competitiva, ocorre quando o inibidor se liga à
enzima ou ao complexo enzima-substrato, não no sítio activo do enzima mas
num outro sítio não cataliticamente activo. Ao ligar-se forma complexos
inactivos, alterando o valor da constante catalítica.
• a inibição incompetitiva (caso muito particular de inibição mista),
ocorre quando, ), o inibidor não se combina com o enzima livre nem interfere
com a ligação entre enzima e substrato mas combina-se com o complexo
enzima-substrato para formar um complexo enzima-substrato-inibidor inactivo.
Estes tipos distintos de inibição reversível podem distinguir-se
experimentalmente pelos efeitos do inibidor na cinética da reacção e que se
podem analisar recorrendo à equação de Michaelis (32).

INTR
OD
UÇ
ÃO
E O
BJE
CTI
VO
S
22
1.6. Objectivos
O objectivo principal deste trabalho é estudar a enzima β-galactosidase,
proveniente de Kluyveromices lactis, na sua forma livre e imobilizada, com a
finalidade de produção de derivados de lactose de modo a melhorar as
características e as qualidades de leites, com aplicação na indústria.
Os objectivos específicos incluem:
1. Imobilização da β-galactosidase em alginato de cálcio.
2. Avaliação da actividade e estabilidade operacional da enzima em
diferentes condições operacionais, com avaliação dos parâmetros
cinéticos (e.g. temperatura, pH, concentração da enzima e do
substrato).
3. Comparação do desempenho enzimático nas diferentes formas (livre e
imobilizada) através da utilização de um substrato padrão.
4. Optimização dos parâmetros operacionais para aplicação em leite.
Recommended

















