
UNIVERSIDADE ESTADUAL DE CAMPINAS
FACULDADE DE ODONTOLOGIA DE PIRACICABA
CARLOS LANE FOGAÇA
AVALIAÇÃO DO EFEITO DO DOADOR DE NITROXILA, SAL
ANGELIS, NA NOCICEPÇÃO INDUZIDA PELA FORMALINA
NA ARTICULAÇÃO TEMPOROMANDIBULAR DE RATOS.
PIRACICABA
2017

CARLOS LANE FOGAÇA
AVALIAÇÃO DO EFEITO DO DOADOR DE NITROXILA, SAL
ANGELIS, NA NOCICEPÇÃO INDUZIDA PELA FORMALINA
NA ARTICULAÇÃO TEMPOROMANDIBULAR DE RATOS.
Tese apresentada à Faculdade de
Odontologia de Piracicaba da
Universidade Estadual de Campinas
como parte dos requisitos exigidos para
a obtenção do título de Doutor em
Odontologia, na área de Fisiologia Oral.
Orientadora: Profa. Dra. Juliana Trindade Clemente Napimoga
ESTE EXEMPLAR CORRESPONDE À
VERSÃO FINAL DA TESE DEFENDIDA
PELO ALUNO CARLOS LANE FOGAÇA
E ORIENTADO PELA PROFª. DRª.
JULIANA TRINDADE CLEMENTE
NAPIMOGA.
PIRACICABA
2017

Agência(s) de fomento e nº(s) de processo(s): Não se aplica.
Ficha catalográfica Universidade Estadual de Campinas
Biblioteca da Faculdade de Odontologia de Piracicaba Marilene Girello - CRB 8/6159
Fogaça, Carlos Lane, 1955 -
F687a Fog Avaliação do efeito do doador de nitroxila, Sal Angelis, na nocicepção induzida pela formalina na articulação temporomandibular de ratos / Carlos Lane Fogaça. – Piracicaba, SP: [s.n.], 2017. Fog
Orientador: Juliana Trindade Clemente Napimoga. Fog Tese (doutorado) – Universidade Estadual de Campinas, Faculdade de Odontologia de Piracicaba. Fog
1. Articulação temporomandibular. 2. Dor. I. Clemente-Napimoga, Juliana Trindade, 1978-. II. Universidade Estadual de Campinas. Faculdade de Odontologia de Piracicaba. III. Título.
Informações para Biblioteca Digital
Título em outro idioma: Evaluation of nitroxila donor effect, Angelis Salt, on the
formalin induced nociception in temporomandibular joint of rats
Palavras-chave em inglês:
Temporomandibular joint
Pain
Área de concentração: Fisiologia Oral
Titulação: Doutor em Odontologia
Banca examinadora:
Juliana Trindade Clemente Napimoga [Orientador]
Giovana Tófoli Moniz
Marcelo Sperandio
Cínthia Pereira Machado Tabchoury
Sidney Figueroba Raimundo
Data de defesa: 20-07-2017
Programa de Pós-Graduação: Odontologia


DEDICATÓRIA Dedico esse trabalho,
A Deus,
Por estar sempre presente me abençoando e iluminando meus caminhos, para
alcançar grandes vitórias.
Aos meus amados filhos,
Lucas, Filipe e Jessica
Pela adorável companhia e amor.
Aos meus pais,
Clementina e Herculano (In memoriam)
Pelo amor e apoio em todos os momentos da minha vida. Vocês sempre serão de
importância eterna para mim.
Aos meus queridos irmãos,
Cinara (In memoriam) e Diógenes
Pela maior cumplicidade, incentivo e, sempre torcendo por mim.

AGRADECIMENTO ESPECIAL A minha orientadora,
Profa. Dra. Juliana Trindade Clemente Napimoga
Agradeço imensamente pela oportunidade de ter possibilitado a realização
de um grande sonho, e pela orientação soberba e suprema, ter me orientado com
tamanha competência, sabedoria e confiança compreendendo minhas dificuldades e
estando sempre disposta a contribuir... ETERNO OBRIGADO!!
Aos meus colegas de doutorado,
Monaliza Lamana, Cristina Gomes de Macedo e Henrique Ballassini Abdalla
pelos ensinamentos laboratoriais e experimentos com animais, e por toda ajuda
oferecida, estando presentes em todos os momentos, muito obrigado! Desejo muito
sucesso a vocês!
A Maricele Deola, pelo carinho, companheirismo, incentivo e amizade.

AGRADECIMENTOS
À Universidade Estadual de Campinas (UNICAMP), em nome do seu
magnífico Reitor José Tadeu Jorge, e à Faculdade de Odontologia de Piracicaba
(FOP), em nome de seu Diretor Guilherme Elias Pessanha Henriques, pelo
privilégio em ser aluno desta instituição e por oferecer uma excelente qualidade de
ensino e infraestrutura para a realização desse trabalho.
À Universidade Comunitária Região de Chapecó – SC, em nome do seu
magnífico Reitor Prof. Dr. Cláudio Alcides Jacoski, e em nome do seu Diretor do
Centro de Ciências da Saúde Prof. Dr. Altamir Trevisan Dutra e ao Coordenador de
Curso Odontologia Prof. Dr. Diogo Alexander Oliveira, pelo privilégio em ser
comtemplado com o convênio DINTER entre a UNOCHAPECÓ e a FOP-UNICAMP
para a realização deste doutorado.
A Srª. Eliete Riguetto, secretária do Departamento de Ciências
Fisiológicas da FOP-UNICAMP e Srª Maria Eliza dos Santos, secretária do
Programa de Pós-Graduação em Odontologia, obrigada pela atenção e carinho.
Ao Carlos Alberto Feliciano, pela amizade e ajuda prestada em todos os
momentos.
Aos meus grandes amigos e colegas de consultório Andreia Soletti e
Giovani Soletti, pela amizade e por toda ajuda oferecida, estando presentes em
todos os momentos, sejam aqueles bons ou ruins, muito obrigado!
Aos professores do Departamento de Ciências Fisiológicas da
Faculdade de Odontologia de Piracicaba, pela oportunidade da realização deste
sonho e pelo exemplo de dedicação ao ensino e a pesquisa, e em especial à Profª.
Cínthia Pereira Machado Tabchoury, muito obrigada pelo constante apoio e
motivação oferecidos para a conclusão deste trabalho.
Enfim, agradeço imensamente a todos que direta ou indiretamente tiveram
grande importância para a conclusão de mais esta etapa. Obrigado pelas palavras de

apoio e incentivo, a contribuição de cada um de vocês foi de essencial importância.
Desejo sucesso e felicidade a cada pessoa que hoje faz parte da minha vida e que,
de uma forma ou outra, são peças fundamentais para o sucesso e continuação do
meu trajeto.

EPÍGRAFE
“Por maior que seja, a dificuldade, jamais desanime. O nosso pior momento na vida
é sempre o momento de melhorar”.
Chico Xavier

RESUMO
Este estudo avaliou o efeito periférico do doador de nitroxila, Sal Angelis (SA),
na dor induzida pela formalina na articulação temporomandibular de ratos
(ATM) e o papel dos receptores opioides mu (µ), delta (δ) e capa (κ) neste efeito.
Para realização deste estudo foram utilizados ratos machos Wistar (± 250 g,
n=4/grupo, perfazendo um total de 11 grupos). A injeção intra-articular de SA (1.5,
15, 50 ou 150 µg/ATM) reduziu significativamente o comportamento nociceptivo
induzido pela formalina 1,5% na ATM dos animais (p<0,05: ANOVA, teste de
Tukey). O pré-tratamento dos animais com o antagonista não seletivo dos
receptores opioides (naloxona, 10 µg/ATM); ou o antagonista de receptores μ-
opioide (CTOP, 120 µg/ATM); ou antagonista do receptor δ-opioide (ICI174,864;
30 µg/TMJ); ou antagonista do receptor κ-opioide (Nor-BNI, 200 µg/ATM)
reduziram significativamente o efeito antinociceptivo induzido pelo SA na ATM
(p<0,05: ANOVA, teste de Tukey), no entanto, este efeito é independente da
ativação da via intracelular óxido nítrico/ guanosina monofosfofato cíclico
(NO/GMPc). O tratamento da ATM com o SA foi capaz de diminuir
significativamente a migração de células inflamatórias e liberação da citocina
pró-inflamatória TNF-α induzida pela formalina (p<0,05: ANOVA, teste Tukey),
apesar de não ter alterado a expressão da molécula de adesão ICAM-1 e do fator
de decaimento de aceleração CD55 (p>0,05: ANOVA, teste de Tukey). Os
resultados sugerem que o SA quando administrado diretamente na ATM de
ratos apresenta um potente efeito analgésico e anti-inflamatório mediado pelos
receptores opioides, no entanto, independente da ativação da via intra-celular
ON/GMPc.
Palavras-chave: sal angelis, dor, articulação temporomandibular

ABSTRACT
The aim of this investigation was evaluate the peripheral antinociceptive effect of
the nitroxyl donor, Angeli’s Salt (AS) in the temporomandibular joint (TMJ) of rats
and the participation of opioid receptors mµ (µ), delta (δ) and kappa (κ) in this
effect. For that, Male Wistar rats were used (± 250g, n=4/group, in a total of 11
groups). Intra-TMJ injection of AS (1.5, 15, 50 or 150µg/TMJ) significantly reduced
nociceptive behavioral responses induced by formalin (p<0.05: ANOVA, Tukey’s
test). Pretreatment with a non-selective antagonist of opioid receptors (naloxone,
10µg/TMJ); or antagonist of µ-opioid receptor (CTOP, 120µg/TMJ); or antagonist
of δ-opioid receptor (ICI174,864; 30 µg/TMJ); or antagonist of κ-opioid receptor
(Nor-BNI, 200µg/TMJ) significantly reduced the peripheral antinociceptive effect
induced by intra-TMJ injection of AS (p<0.05: ANOVA, Tukey’s test). On the
other hand, this effect is independent of the activation of the intra-cellular nitric
oxide/cyclic guanosine monophosphate (NO/cGMP) pathway. The intra-TMJ
injection of AS (50µg/TMJ) significantly reduced leukocyte migration and the
release of pro-inflammatory cytokine TNF-α (p<0.05: ANOVA, Tukey`s test), but
not modulated the expression of intercellular adhesion molecule 1 (ICAM-1) and
decay-accelerating factor CD55 (p>0.05). The results suggest that AS induces a
potential antinociceptive and anti-inflammatory effect mediated by opioid
receptors, but independent of the activation of intracellular NO/cGMP pathway.
Keywords: angelis salt, pain, temporomandibular joint.

LISTA DE ABREVIATURAS E SIGLAS
5-HT - Serotonina
AINEs - Anti-inflamatórias não-estereoidais
AKT - Proteína quinase B
AMP - Adenosina Monofosfato cíclico
ATM - Articulação Temporomandibular
ATP - Adenosina trifosfato
DTM - Disfunção Temporomandibular
GMPc - Guanosina monofosfato cíclico
HNO - Doadores de nitroxila
IL-1 - Interleucina 1
IL-6 - Interleucina 6
IL-12 - Interleucina 12
IL-17 - Interleucina 17
K+
ATP - Canais de potássio sensíveis ao ATP
ON - Óxido Nítrico
PI3Kγ - Fosfatidilinositol 3-quinase isoforma gama
PKA - Proteína quinase A
PKC - Proteína quinase C
PKG - Proteinoquinase dependente do GMPc
SNC - Sistema Nervoso Central
TNF-α - Fator de necrose tumoral alpha
SA - Sal de Angelis

SUMÁRIO
1. INTRODUÇÃO
14
2. REVISÃO DA LITERATURA
16
3. PROPOSIÇÃO
21
4. MATERIAL E MÉTODOS
22
5. RESULTADOS
28
6. DISCUSSÃO
35
7. CONCLUSÃO
40
REFERÊNCIAS
41
ANEXO 1- Certificado do Comitê de Ética no Uso de Animais 48

14
1. INTRODUÇÃO
A dor crônica é uma das principais razões para a busca de tratamentos
de desordens temporomandibulares (DTM) responsável por causar incapacidades
físicas e psicológicas (Su Lobbezoo et al., 2017; Prasad et al., 2016). A etiologia
das DTMs é multifatorial envolvendo fatores biomecânicos, neuromusculares,
biopsicológicos e neurobiológicos (Prasad et al., 2016). Os fatores neurobiológicos
das DTMs ainda não estão bem esclarecidos, especialmente os mecanismos
envolvidos na transição da dor aguda para o estado crônico após injúrias de
tecidos/ nervos periféricos orofaciais. O controle da dor decorrente de DTMs ainda
é um grande desafio para clínicos e pacientes (Cairns et al., 2010).
A dor é um dos sinais clássicos de processo inflamatório
apresentando como denominador comum à sensibilização dos receptores
nociceptivos aferentes primários. Decorrente de estímulos inflamatórios ou lesões
teciduais, a liberação de citocinas e quimiocinas pró e anti-inflamatórias
desencadeiam a liberação de prostanóides e aminas simpatomiméticas que por
sua vez atuam diretamente nos nociceptores causando a hipernocicepção, a
redução do limiar de excitabilidade devido à modulação de canais de sódio
voltagem-dependentes (Khasar et al. 1999, Verri et al, 2006, Quinteiro et al., 2012;
Quinteiro et al., 2014). Com o melhor entendimento nos mecanismos envolvidos
nas condições dolorosas, diversas terapias antiálgicas têm sido propostas.
O óxido nítrico (ON) é um sinalizador intracelular envolvido na
transmissão nociceptiva, que tem despertado bastante interesse no meio
científico. O ON é um gás altamente difusível que contribui para uma variedade
de funções biológicas tais como respostas imunológicas (Langrehr et al., 1993)
e sinalizações neurais (Brenman, 1997) através da via intracelular L-
Arginina/ON/GMPc/K+ATP
(Ferreira et al., 1991; Cunha et al., 2010). Na dor, o ON
tem apresentado um duplo efeito, ou seja, pode induzir ou reduzir a dor
dependendo da sua concentração e tecido onde é liberado (Cury et al., 2011).
Sobre o efeito antinociceptivo do ON, tem sido demonstrado que no tecido
periférico doadores ou substâncias que aumentam a concentração neuronal
de ON resultam no bloqueio da hiperalgesia inflamatória (Cury et al., 2011).
O nitroxil (HNO), produzido a partir do ON reduzido em um elétron,
recebeu, até recentemente, menor atenção do que as espécies oxidadas, no

15
entanto, estudos recentes revelaram aspectos importantes da química do HNO,
que lhe conferem importância biológica. A simplicidade estrutural dessa espécie
triatômica contrasta com a complexidade de algumas de suas características e
propriedades químicas fundamentais (Fukuto et al., 2005).
Para o estudo dos efeitos do HNO são usados compostos doadores
desta espécie. Entre estes, o que está melhor caracterizado é o trioxodinitrato
de sódio (Na2N2O3) também chamado de Sal de Angelis (SA). Este, além de ser
usado em terapias anti-alcoólicas, também tem propriedades vasodilatadoras.
Similarmente ao ON, doadores de nitroxila (HNO) também ativam a guanilil ciclase
que atua como um receptor intracelular para NO (Fukuto et al.,2005).
Em particular, em um estudo recente, o doador de nitroxila
denominado Sal Angelis, inibiu a hiperalgesia inflamatória induzida na pata de
ratos por diferentes agentes inflamatórios, através da inibição da produção e
liberação de citocinas pró-inflamatórias e ativação periférica da via intracelular
GMPc/PKG/K+ATP (Zaperlon et al., 2013).
O tratamento das condições dolorosas ainda é considerado um desafio
para clínicos, pacientes e pesquisadores. Sendo assim, considerando que ativação
periférica da via intracelular L-Arginina/ON/GMPc está envolvida no efeito
antinociceptivo de diferentes drogas na ATM (Clemente et al., 2004; Clemente-
Napimoga et al., 2009; Pena-dos-Santos et al., 2009), este estudo apresenta como
proposta investigar o efeito do doador de nitroxila Sal Angelis na hiperalgesia
inflamatória induzida pela formalina na ATM de ratos assim como os mecanismos
envolvidos neste processo.

16
2. REVISÃO DA LITERATURA
2.1. As condições dolorosas decorrentes da Articulação Temporomandibular:
A Articulação Temporomandibular (ATM) é uma articulação dupla
bilateral, que se movimenta interdependente. É através dela que a mandíbula, único
osso móvel do crânio, liga-se à base craniana. Constitui a parte terminal do osso da
mandíbula e está intimamente relacionada com o crânio através do osso temporal
(Douglas, 1998). Sendo assim, pode-se considerar que a ATM é uma das
articulações mais utilizadas no corpo humano, anatomicamente complexa, que
possibilita a mandíbula executar variados movimentos durante a mastigação,
deglutição e fala (Amantéa et al., 2004; Nunes e Maciel, 2005; Ingawalé e Goswami,
2009).
A ATM, assim como outras articulações sinoviais do corpo humano, a
ATM pode ser afetada por doenças inflamatórias, traumáticas, infecciosas,
congênitas, e neoplásicas. As alterações que acometem a ATM e musculatura
adjacente são denominadas de Disfunções Temporomandibulares (DTMs). As DTMs
podem ser definidas como um conjunto de desordens musculoesqueléticas e
neuromusculares que afetam a ATM e tecidos circundantes, caracterizadas por
limitação dos movimentos articulares, dor e ruído articular (Leeuw et al, 2013, Graff-
Radford e Bassiur, 2014). Em particular, as condições dolorosas decorrentes da
ATM resultam de processos inflamatórios (Kopp, 2001), que progridem para quadros
de dor inflamatória persistentes resultando em dor orofacial crônica. Por este motivo,
as condições dolorosas decorrentes da ATM são consideradas denominador comum
para o desenvolvimento de dor orofacial crônica (Cairns, 2010; Leeuw, 2010; Sessle,
2011).
A dor é um dos sinais clássicos do processo inflamatório, sendo resultado
da liberação de várias citocinas pró-inflamatórias, em particular do fator de necrose
tumoral alfa (TNF-α) e das interleucinas IL-1, IL-6, IL- 12, e IL-17, que contribuem
para uma remodelação articular assim como uma degradação da cartilagem (Vernal
et al., 2008). A liberação destas citocinas facilita a liberação de prostanóides e
aminas simpatomiméticas que promovem a sensibilização de nociceptores
periféricos desta região (Raja et al., 1988; Alstergrem e Kopp, 2000; Nordahl et al.,
2000; Kopp, 2001; Oliveira et al., 2005) e de neurônios nociceptivos centrais do
complexo sensório-nuclear trigeminal do tronco encefálico (Iwata et al, 1999; Sessle,

17
2000; Dubner e Ren, 2004).
O sistema de receptores nociceptivos nos tecidos faciais é representado
por um arranjo plexiforme e de terminações livres de fibras nervosas amielínicas.
Essas fibras, chamadas terminações nervosas livres, estão distribuídas pela pele,
tecido subcutâneo, fáscia, tecido adiposo, camada adventícia dos vasos sanguíneos,
periósteo, músculos, cápsula articular e ligamentos da ATM. Em circunstâncias
normais, este sistema está inativo, mas não totalmente. Muitas dessas terminações
nervosas livres atuam como nociceptores, respondendo a estímulos nocivos (Sessle,
1995).
Quando há exposição aos fluidos dos tecidos circunvizinhos ou
substâncias irritantes como ácido lático, bradicinina, histamina, íons potássio e
outras substâncias liberadas de tecidos inflamados e metabolicamente anormais; as
terminações nervosas livres tem sua atividade aferente acentuada quando são
excitadas por aplicação de forças mecânicas, térmicas, elétricas ou químicas (Milan,
1999). Quando esses nociceptores são excitados, enviam informações ao Sistema
Nervoso Central (SNC) para que identifique a intensidade, qualidade, localização e
duração desse estímulo (Sessle, 1995).
Frente a uma lesão, citocinas pro- e anti-inflamatórias e quimiocinas são
liberadas no local da lesão tanto por células residentes (como macrófagos, por
exemplo), quanto pelas células do próprio infiltrado inflamatório, que irão
desencadear a ativação de mediadores inflamatórios responsáveis pela dor
inflamatória. Nos tecidos que envolvem a ATM, tem sido demonstrado o importante
papel da cascata de hiperalgesia inflamatória, descrita por Verri e colaboradores em
2006, desencadeada pelas citocinas pró-inflamatórias TNF-alpha (TNF-α) e IL-1 beta
(IL-1β) (Rodrigues et al., 2006; Pena-dos-Santos et al., 2009; Quinteiro et al., 2012;
Silva Quinteiro et al., 2014). Uma vez liberadas, as citocinas pró-inflamatórias TNF-α
e IL-1β são responsáveis pela liberação de citocinas (IL-6, IL-12 e IL-18) e
quimiocinas (CINC-1 e KC), as quais resultam na liberação de prostanóides (em
particular a prostaglandina E2) e aminas simpatomiméticas. Os prostanóides e
aminas simpatomiméticas agem diretamente nos nociceptores, preferencialmente
das fibras C aferentes primárias. Uma vez ativados, desencadeiam a ativação das
vias de segundos mensageiros, como a proteína quinase A e proteína kinase C
(Quinteiro et al, 2012), resultando na fosforilação de canais de sódio voltagem
dependente, responsáveis pelo aumento da excitabilidade das membranas

18
neuronais (Verri el al., 2006).
É importante evidenciar o fato de que os tecidos adjacentes a ATM são
mais sensíveis à liberação das aminas simpatomiméticas e PGE2 quando
comparado a tecidos cutâneos (Rodrigues et al., 2006). O componente simpático
das condições dolorosas inflamatórias explica a menor efetividade dos anti-
inflamatórios não esteroidais nestas condições. Considerando que a ATM recebe
uma rica inervação simpática (Widenfalk and Wiberg, 1990; Yoshino et al., 1998;
Kido et al., 2001) e os nociceptores localizados nas fibras C nociceptivas primárias
são mais sensíveis a liberação das aminas simpáticas (Rodrigues et al., 2006), em
acréscimo as particularidades anatômicas da ATM, é compreensível a taxa de
insucessos na terapêutica das condições dolorosas decorrentes da ATM.
2.2. Óxido Nítrico e o Sal Angelis:
O óxido nítrico (ON) é um gás difusível com a capacidade de difundir-se
livremente através das membranas celulares e, ao contrário de outros
neurotransmissores clássicos, o ON não pode ser regulado pelo processo de
armazenamento, liberação e reabsorção. Em vez disso, a biossíntese de ON é
controlada dinamicamente com o intuito de fornecer os níveis adequados de acordo
com a demanda tecidual (Yun et al., 1997; Kumar et al., 2017 ).
O óxido nítrico está envolvido em diferentes processos fisiológicos do
corpo através da modulação de diferentes reações bioquímicas como
neurotransmissão, plasticidade sináptica, regulação da expressão gênica, percepção
da dor e depressão (Kumar et al., 2017).
A síntese de ON utiliza como percussor a L-arginina, um aminoácido que
é convertido em outro aminoácido, a L-citrulina, sendo essa reação catalisada por
enzimas chamadas de ON sintases [ONS] (Zaperlon et al., 2013). Muitas pesquisas
vêm sendo desenvolvidas na intenção de desvendar e/ou estabelecer o mecanismo
de ação do óxido nítrico e suas principais respostas. Sabe-se que o ON é capaz de
ativar a enzima guanilil ciclase solúvel (GC), possivelmente pela reação com um
átomo de ferro em uma estrutura heme, que então catalisa a conversão do trifosfato
de guanosina (GTP) em monofosfato de guanosina cíclico (GMPc) (Staurengo-
Ferrari et al., 2014).
Considerando a ação do ON no sistema nervoso, muitas linhas de
pesquisa tem demonstrado um efeito “dual” do óxido nítrico no sistema nociceptivo,

19
podendo tanto induzir como reduzir a dor dependendo do sítio de ação e da
quantidade disponibilizada nos tecidos (Souza et al., 2001; Cunha et al., 2010;
Staurengo-Ferrari et al., 2014). Além disso, uma série de compostos analgésicos são
dependentes do ON para promover seu efeito antinociceptivo, tais como os opioides,
alguns anti-inflamatórios não-esteroides, dipirona e diclofenaco. (Staurengo-Ferrari
et al., 2014).
Na ATM, tem sido demonstrado que a administração local de diferentes
drogas anti-inflamatórias, tais como o agonista do receptor κ-opioide U50,488
(Clemente et al., 2004; Clemente-Napimoga et al., 2009) e a 15-deoxy-12,14-
prostaglandin J2 (15d-PGJ2) (Pena-dos-Santos et al., 2009; Macedo et al., 2016)
resultam em uma resposta analgésica através da ativação da via intracelular
ON/GMPc.
Até recentemente, os efeitos biológicos do ON vinham sendo atribuídos
diretamente à molécula de ON exclusivamente. No entanto, ON pode ser tanto
oxidado (ON+) como reduzido (ON-), sendo a espécie um-elétron reduzida do ON
denominada de nitroxil (Irvine et al., 2008).
O nitroxil (HNO) é a forma reduzida em um elétron e próton do ON, e
recentemente tem se destacado pelas suas distintas ações farmacológicas e
vantagens terapêuticas. Evidências tem demonstrado que perifericamente os
doadores de HNO bloqueiam diretamente a hiperalgesia inflamatória (Cury et al.,
2011) e modulam a neurotransmissão da dor (Cunha et al., 2010), conferindo assim
ao HNO importante função biológica.
O sal de Angelis (SA), ou trioxodinitrato de sódio (Na2N2O3) é o doador
sintético mais comumente utilizado no estudo biológico do HNO (Paolocci et al.,
2007). Tem sido demonstrado que o SA apresenta efeito antinociceptivo em modelos
de hiperalgesia mecânica induzida por uma variedade de estímulos como
carragenina, LPS, citocinas e PGE2. Este efeito é resultado da inibição da produção
de citocinas pró-inflamatórias induzida por carragenina, provocando uma
antinocicepção periférica (Staurengo-Ferrari et al., 2014). Neste sentido, o efeito
antinociceptivo de SA em modelos comportamentais de hiperalgesia mecânica,
térmica e neuropatia é dependente da ativação da via antinociceptiva do
GMPc/PKG/K+ATP (Zaperlon et al., 2013; Staurengo-Ferrari et al., 2014; Longhi-
Balbinot et al., 2016).
Apesar de altos níveis de ON ser detectado no fluido sinovial de pacientes

20
com condições dolorosas na ATM (Anbar and Gratt 1998; Suenaga et al. 2001; Wahl
et al. 2003), o papel do ON na dor inflamatória da ATM não está claro. Considerando
que a ativação periférica da via intracelular ON/GMPc está envolvida no efeito
antinociceptivo de diferentes drogas na ATM (Clemente et al., 2004; Clemente-
Napimoga et al., 2009; Pena-dos-Santos et al., 2009), este estudo apresenta como
proposta investigar o efeito do doador de nitroxila Sal Angelis na hipernocicepção
inflamatória induzida pela formalina na ATM de ratos assim como os mecanismos
envolvidos neste processo.

21
3. PROPOSIÇÃO
Este estudo apresenta como proposta avaliar o efeito do doador de nitroxila Sal
Angelis na hipernocicepção inflamatória induzida pela formalina na ATM de ratos
assim como os mecanismos envolvidos neste processo, através:
da avaliação do efeito do Sal Angelis no comportamento nociceptivos induzido
pela formalina na ATM de ratos;
da avaliação do efeito do Sal Angelis na migração de células inflamatórias e
extravasamento plasmáticos induzidos pela formalina na ATM de ratos;
da avaliação do efeito do Sal Angelis na liberação das citocinas pro-
inflamatórias (TNF-α e IL-1β), liberação do neuropeptídeo Substância P, da
expressão da molécula de adesão (ICAM-1) e do fator de aceleração de
decaimento (CD55);
da avaliação da participação dos receptores opioides (mu, delta e capa) e da
ativação da via intracelular GMPc/PKG/K+ATP no efeito antinociceptivo
induzido pelo Sal Angelis na ATM de ratos.

22
4. MATERIAL E MÉTODOS
4.1. Animais
Para a realização deste trabalho foram utilizados ratos machos Wistar (±
250g, n=4 animais por grupo, total de 11 grupos) provenientes do CEMIB (Centro
Multidisciplinar para Investigação Biológica na Área da Ciência em Animais de
Laboratório) e mantidos no Biotério da FOP – UNICAMP, em gaiolas plásticas (5 por
gaiola) contendo maravalha, em ambiente com controle de luminosidade (ciclos
claro/escuro de 12h) com alimentação e água, ad libitum. Todos os procedimentos
experimentais foram aprovados pelo Comitê de Ética em Experimentação Animal da
Universidade Estadual de Campinas (CEUA #3101-1) e estão de acordo com as
diretrizes determinadas pelo Conselho Nacional de Controle de Experimentação
Animal (CONCEA) e pela Associação Internacional para Estudo da Dor (IASP), em
animais conscientes (Zimmermann, 1983). Para minimizar o estresse durante as
sessões experimentais, os animais foram previamente manipulados pelo
pesquisador por um período de sete dias. Para retirada de tecidos os animais foram
eutanasiados por aprofundamento de anestesia (Isoflurano 5%).
4.2. Drogas
Trioxodinitrato de sódio – Sal Angelis (SA) sintetizado e utilizado como
descrito anteriormente (Smith e Hein, 1960). Naloxona, inibidor não seletivo de
receptores opioides; nor-binaltorphiminie (Nor-BNI), antagonista do receptor κ-
opioide; N,N-diallyl-Tyr-Aib-Aib-Phe-Leu (ICI174,864), antagonista do receptor δ-
opioide; D-Phe-Cys-Tyr-D-Trp-Orn-Thr-Pen-Thr-NH2 (CTOP), antagonista do
receptor μ-opioide; ODQ, inibidor da guanilil sintase; KT5823, inibidor da PKG;
glibenclamida, bloqueador de canal de K+ATP e uma solução aquosa de 37% de
formaldeído foram obtidos da Sigma-Aldrich (St. Louis, MO, USA). Todas as drogas
foram diluídas em solução de cloreto de sódio a 0,9%. A solução de formalina foi
preparada a partir da solução aquosa de 37% de formaldeído e diluída em NaCl
0,9% até uma concentração final de 1,5% (Roveroni et al., 2001).
4.3. Injeção intra-articular na ATM
Os animais foram anestesiados por 30 segundos através da inalação de

23
isoflurano (3 %) (Figura 1). Em seguida uma agulha calibre 30G, acoplada e uma
seringa Hamilton (50 μl) por meio de uma cânula de polietileno P20 foi introduzida na
ATM. O ponto de referência para a injeção intra-articular foi à borda do arco
zigomático na região póstero-lateral do côndilo. Os animais retornaram a consciência
aproximadamente 30 segundos após o término da inalação do anestésico (Roveroni
et al., 2001; Clemente et al., 2004).
Figura 1. Anestesia inalatória durante injeção intra-articular na ATM
4.4. Teste comportamental
Imediatamente após a injeção intra-articular, o animal já consciente, foi
recolocado na câmara de observação e as respostas comportamentais
caracterizadas pelo ato de coçar a região injetada com a pata dianteira ou traseira e
pelo ato de levantar reflexamente a cabeça foram quantificadas durante 45 min.,
divididos em 9 blocos de 5 min. O tempo em segundos que o animal permaneceu
coçando a região orofacial foi quantificado através da utilização de um cronômetro, e
o número de vezes que o animal levantou reflexamente a cabeça foi quantificado por
um contador de células (Roveroni et al., 2001; Clemente et al., 2004). Considerando
que o ato de levantar reflexamente a cabeça segue um padrão uniforme de 1 s de
duração, a intensidade da resposta nociceptiva foi quantificada somando-se esse
comportamento ao ato de coçar a região injetada como previamente padronizado
(Roveroni et al., 2001; Clemente et al., 2004). Imediatamente após as análises
comportamentais, os animais foram eutanasiados por aprofundamento de anestesia
(Isoflurano 5%) seguido de deslocamento cervical e amostras do tecido periarticular

24
e lavado intra-articular foram removidas para análises posteriores. Os tecidos
periarticulares foram removidos por uma dissecção dos músculos temporal e
masseter profundo com atenção aos pontos anatômicos (arco zigomático e
timpânica) até a exposição do processo condilar. As amostras representam todos os
tecidos que cercam o processo condilar: músculos mastigatórios (temporal posterior,
masseter, pterigoide externo) cartilagem articular, fibrocartilagem do disco e
ligamentos laterais. O tamanho da amostra padrão é de 1 x 1 x 0, 5 cm.
4.5. Delineamento experimental
Para o desenvolvimento deste trabalho foram realizados os seguintes
grupos experimentais:
4.5.1. Para avaliar o efeito do Sal Angelis na hipernocicepção inflamatório induzida
pela formalina na ATM de ratos:
GRUPO 1: Injeção intra-articular de salina (45 µl/ATM)
GRUPO 2: Injeção intra-articular de formalina 1,5% (15 µl/ATM) + salina (30
µl/ATM).
GRUPO 3 a 6: Pré-tratamento com uma injeção intra-articular de Sal Angelis (SA)
(1.5, 15, 50 ou 150 µg/15 µl/ATM) (Zarpelon et al., 2013) 15 minutos antes da injeção
intra-articular de Formalina 1,5% (15 µl/ATM) + salina (30 µl/ATM).
GRUPO 7: Pré-tratamento com uma injeção intra-articular de SA (150 ou 50 µg/15

25
µl/ATM) na ATM contralateral 15 minutos antes da injeção intra-articular de Formalina
1,5% (15 µl/ATM) + salina (30 µl/ATM).
4.5.2. Para avaliar a participação de receptores opioides endógenos e ativação da via
GMPc/ PKG/K+ATP no efeito antinociceptivo do Sal Angelis:
GRUPO 8: Pré-tratamento com uma injeção intra-articular do antagonista de
receptores opioides não seletivo Naloxona (10 µg/15 µl/ATM) (Pena-dos-Santos et
al., 2009) + SA (50 µg/15 µl/ATM) 15 minutos antes da injeção intra-articular de
Formalina 1,5% (15 µl/ATM).
GRUPO 9 a 11: Pré-tratamento com uma injeção intra-articular do antagonista do
receptor μ-opioide (CTOP 120 µg/15 µl/ATM); ou do antagonista do receptor κ-
opioide (Nor-BNI 200 µg/15 µl/ATM); ou do antagonista do receptor δ-opioide (ICI
174,864 30 µg/15 µl/ATM) (Pena-dos-Santos et al., 2009) + SA (50 µg/15 µl/ATM) 15
minutos antes da injeção intra-articular de Formalina 1,5% (15 µl/ATM).
GRUPO 12 a 14: Pré-tratamento com uma injeção intra-articular do inibidor da
guanilil sintase (ODQ 8 µg/15 µl/ATM); ou do inibidor da PKG (KT5823 1,5 µg/15
µl/ATM); ou do bloqueador de canal de K+ATP (glibenclamida 160 µg/15 µl/ATM)
(Zarpelon et al., 2013) + SA (50 µg/15 µl/ATM) 15 minutos antes da injeção intra-
articular de Formalina
1,5% (15 µl/ATM).

26
4.6. Quantificação da migração leucocitária
Para a avaliação da migração de neutrófilos, os animais foram mortos e
um lavado intra-articular (2x) com 5 µl de PBS contendo EDTA (1 mM) foi realizado.
O lavado (10 µl) foi diluído em 90 µl de PBS/EDTA e a partir deste, realizadas a
contagem total dos leucócitos. Para a contagem total de leucócitos, alíquotas de 10
µl do lavado articular foram diluídas em líquido de Turk (20 µl), na proporção de 1:2,
sendo a contagem total dos leucócitos realizada em câmara de Neubauer, com o
auxílio de microscópio óptico (aumento de 100x) e contador manual e expressa
como número de células x 104/cavidade articular (Quinteiro et al., 2014).
4.7. Extração de proteínas do tecido periarticular
Amostras de tecido periarticular foram homogeneizadas em 500 µl de
buffer apropriado contendo inibidores de protease (Ripa Lysis Buffer, Santa Cruz,
Biotechnology, Dallas, Texas, USA) seguida de centrifugação por 10 min/10,000
rpm/4 °C. A quantidade total de proteínas extraídas foi medida usando o kit BCA
(Thermo Scientific, Rockford, IL, USA). Os sobrenadantes foram armazenados a -
20°C e utilizados posteriormente para as análises através de ELISA e Western Blot.
4.8. Western Blot para avaliação da expressão da molécula de adesão celular
(ICAM-1) e do fator de aceleração de decaimento (CD55).
As proteínas do sobrenadante foram separadas por eletroforese em gel
de poliacrilamida SDS-PAGE 10% e transferidas para membranas de nitrocelulose.
As membranas foram incubadas "overnight" à 4°C com tampão de bloqueio [PBS 5%
(p/v) de leite desnatado e 0,1% Tween 20]. As membranas foram lavadas três vezes
com PBS 0,1% Tween 20. Em seguida foram incubadas em solução de PBS
contendo 5% de leite desnatado e 0,1% Tween 20 contendo anticorpo anti-ICAM,

27
anti-CD55 ou anti-α-tubulina (Santa Cruz) e GAPDH (Genetex). Após a lavagem, as
membranas foram incubadas com anticorpo secundário conjugado HRP e
novamente lavadas. As membranas foram então reveladas com kit de
quimioluminescência (ECL, Amershan Pharmacia Biotech, Little Chalfont, U.K.)
como descrito no manual de instruções. O controle negativo foi obtido pela omissão
do anticorpo primário.
4.9. Avaliação da liberação das citocinas pró-inflamatorias TNF-alpha e IL-
através ensaio imunoenzimático (ELISA).
As análises utilizando o método imunoenzimático foram realizadas de
acordo com as instruções do fabricante para cada kit. Foram utilizados os kits para
TNF-α (R&D Systems, USA), IL-1β (R&D System, USA) e Substância P (Fênix
Pharmaceuticals, USA).
4.10. Análise estatística
Os dados foram avaliados utilizando-se análise de variância a um critério
de avaliação (One-Way ANOVA). As comparações múltiplas foram feitas pelo teste
de Tukey. Para todos os testes o nível de significância foi estabelecido em 5%. O
programa GraphPad Prism 6.0 foi utilizado para a realização dos cálculos e
confecção dos gráficos.

28
5. RESULTADOS
5.1. Efeito periférico antinociceptivo do doador de nitroxila Sal Angelis (SA) na
ATM de ratos.
Os resultados demonstraram que as quatro doses do SA (1,5; 15; 50 ou
150 μg/ATM) reduziram significativamente (p<0,05: ANOVA, teste de Tukey) a
resposta nociceptiva induzida pela injeção intra-articular de formalina na ATM de
ratos (Figura 2). A aplicação intra-articular do SA (150 μg/ATM) na ATM
contralateral (ct) diminuiu significativamente a resposta comportamental induzida
pela formalina na ATM ipsilateral (p<0,05: ANOVA, teste de Tukey), demonstrando
um efeito sistêmico. A aplicação intra-articular do SA (50 μg/ATM) na ATM
contralateral não apresentou efeito na nocicepção induzida pela formalina na ATM
ipsilateral (p>0,05), demonstrando o efeito periférico desta dose, sendo assim, a
dose de 50 μg / ATM do SA foi selecionada para os experimentos posteriores
(Figura 2)
Figura 2. Avaliação do efeito antinociceptivo do SA na região da ATM.
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
+
**
**
*
##
Re
sp
os
ta n
oc
ice
pti
va
(s
)
1 .5 1 5 5 0 1 5 0 1 5 0 c t 5 0 c t
S A ( g /A T M )
1 ,5 % F o rm a lin a
N a C l
0 ,9 %
O símbolo (+) indica diferença estatística quando comparado ao grupo que recebeu
NaCl 0,9% (p<0,05: ANOVA, teste de Tukey) . O símbolo (*) indica reposta nociceptiva
significamente menor quando comparado a do grupo formalina 1,5%. O símbolo (#)
indica resposta nociceptiva significamente menor que a dos grupos de SA (1,5 e 15
μg/ATM). Os dados estão expressos como média ± DP. Fonte: Autoria própria

29
5.2. Avaliação da participação de receptores opioides no efeito antinociceptivo
do SA.
O pré-tratamento com a injeção intra-articular do antagonista não seletivo
dos receptores opioides (Naloxona, 10 μg/ATM), reduziu significativamente o efeito
do SA (50 µg/ATM) na nocicepção induzida pela formalina 1,5% (Figura 3). Este
resultado sugere a participação dos receptores opioides no efeito antinociceptivo do
SA, sendo assim, para avaliar qual receptor opioide estaria envolvido neste efeito os
animais foram tratados com o antagonista do receptor δ-opioide ICI 174,864 (30
µg/ATM), ou antagonista do receptor μ-opioide CTOP (120 µg/ATM), ou do
antagonista do receptor κ-opioide Nor-BNI (200 µg/ATM). Os resultados
demonstraram que os antagonistas dos receptores δ-, κ- e μ-opioide reduziram o
efeito antinociceptivo induzido pelo SA (50 µg/ATM) (Figura 3).
Figura 3. Avaliação da participação dos receptores opioides no efeito antinociceptivo
do SA na ATM de ratos.
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
Co
mp
ort
am
en
to n
oc
ice
pti
vo
(s
)
S A 5 0 g /T M J
F o rm a lin a 1 .5 %
N LXv e íc u lo IC I C T O P N o r-B N I
+
*
* *
*
*
#
# #
#
O símbolo (+) indica diferença estatística significativa maior quando comparado ao grupo
NaCl 0,9% (p<0.05: ANOVA, teste de Tukey). O símbolo (*) indica resposta
significativamente menor quando comparado a do grupo formalina 1,5%. O símbolo (#)
indica diferença estatística significativamente maior quando comparado ao grupo SA (50
μg/ATM). Os dados são expressos como média ± DP. Fonte: Autoria própria

30
5.3. Avaliação da participação da via GMPc/PKG/K+ATP no efeito antinociceptivo
do SA.
A coadministração do inibidor do GMPc ODQ (8 µg/15 µl), ou o inibidor da
proteína quinase G (PKG) KT5823 (1,5 µg/ATM), ou o inibidor do canal de potássio
sensível ao ATP (K+ATP) Glibenclamida (160 µg/ATM) com o SA, não modificou o
efeito antinociceptivo induzido pelo SA na resposta nociceptiva induzida pela injeção
intra-articular de Formalina 1,5% (Figura 4).
Figura 4. Avaliação da participação da via GMPc/ PKG/K+ATP no efeito antinociceptivo
do SA na ATM de ratos.
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
3 0 0
Re
sp
os
ta n
oc
ice
pti
va
(s
)
S A 5 0 g /A T M
1 ,5 % F o rm a lin a
O D Q K T 5 8 2 3 G lib
** *
*
O símbolo (*) indica resposta significativamente menor quando comparado a do grupo
formalina 1,5%. (p<0.05: ANOVA, teste de Tukey). Os dados são expressos como média ±
DP.

31
5.4. Avaliação do efeito do SA na migração leucocitária induzida pela formalina
na ATM de ratos.
O pré-tratamento com uma injeção intra-articular com o SA (1,5, 15, 50 e
150 µg/ATM) reduziu significativamente a migração de leucócitos induzida pela
injeção intra-articular de formalina 1,5% na ATM de ratos (Figura 5).
Figura 5. Avaliação do efeito do SA na migração leucocitária induzida pela formalina
na ATM de ratos.
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
Le
uc
óc
ito
s x
10
4c
el/
ca
vid
ad
e
1 ,5 1 5 5 0 1 5 0
S A ( g /A T M )
1 ,5 % F o rm a lin a
N a C l 0 ,9 %
+
*
*
* *# #
#
O símbolo (+) indica diferença estatística quando comparado ao grupo que recebeu NaCl
0,9% (p<0,05: ANOVA, Tukey test). O símbolo (*) indica migração leucocitária
significativamente menor quando comparado a do grupo formalina 1,5% (p<0,05: ANOVA,
Tukey test). O símbolo (#) indica migração leucocitária significativamente menor que a do
grupo SA 1,5 µg/ATM. Os dados são expressos como média ± DP. Fonte: autoria própria.

32
5.5. Efeito do SA na expressão da molécula de adesão ICAM-1 e do fator de
aceleração de decaimento CD 55.
O efeito do SA na migração leucocitária induzida pela injeção intra-
articular de formalina na ATM, não está associado a uma alteração da expressão da
ICAM-1 ou CD55 (Figura 6A e B).
Figura 6. Efeito do SA na expressão da molécula de adesão ICAM-1 e do fator de
aceleração de decaimento CD 55.
0 .0
0 .5
1 .0
1 .5
2 .0
Ex
pre
ss
ão
de
IC
AM
(D
O)
S A 5 0 g /A T M
1 ,5 % F o rm a lin a
N LX
1 0 g /A T M
A
Ex
pre
ss
ão
CD
55
(D
O)
0 . 0
0 .5
1 .0
1 .5
S A 5 0 g /A T M )
1 .5 % F o rm a lin a
N LX
1 0 g /A T M
B
A análise por Western Blot demonstrou que o tratamento com uma injeção intra-articular de
SA (50 μg/ATM) não alterou o nível proteico das moléculas ICAM-1 e CD55 (p>0,05:
ANOVA, Tukey test). Fonte: autoria própria.

33
5.6. Efeito do SA na liberação de TNF-α e IL-1β induzido pela formalina na ATM
de ratos.
Os resultados demonstraram que a injeção intra-articular de SA (50
µg/ATM) reduziu significativamente a liberação da citocina pró-inflamatórias TNF-α
(p<0,05: ANOVA, teste de Tukey), mas não de IL-1β (p>0,05), induzido pela
formalina 1,5% na ATM (Fig. 7A e B).
Figura 7. Efeito do SA na liberação de TNF-α e IL-1β induzido pela formalina na ATM.
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
N a C l 0 ,9 %
+
**
TN
F-
(p
g/m
l)
S A (5 0 g /A T M )
1 ,5 % F o rm a lin a
N LX
1 0 g /A T M
A
0
2 5 0
5 0 0
7 5 0
1 0 0 0
1 2 5 0
1 5 0 0
IL-1
(
pg
/mg
)
* *
*
N a C l 0 ,9 %
S A (5 0 g /A T M )
1 ,5 % F o rm a lin a
N LX
1 0 g /A T M
(A) O símbolo (+) indica liberação de TNF-α significativamente maior quando comparada a
do grupo NaCl 0,9% (p>0,05; ANOVA, Tukey test). O símbolo (*) indica liberação de TNF-α
significativamente menor quando comparado ao grupo formalina 1,5%. (B) O símbolo (*)
indica liberação de IL-1β significativamente maior quando comparada a do grupo NaCl
0,9% (p>0,05; ANOVA, Tukey test). Os dados são expressos como média ± DP. Fonte:
autoria própria.
34
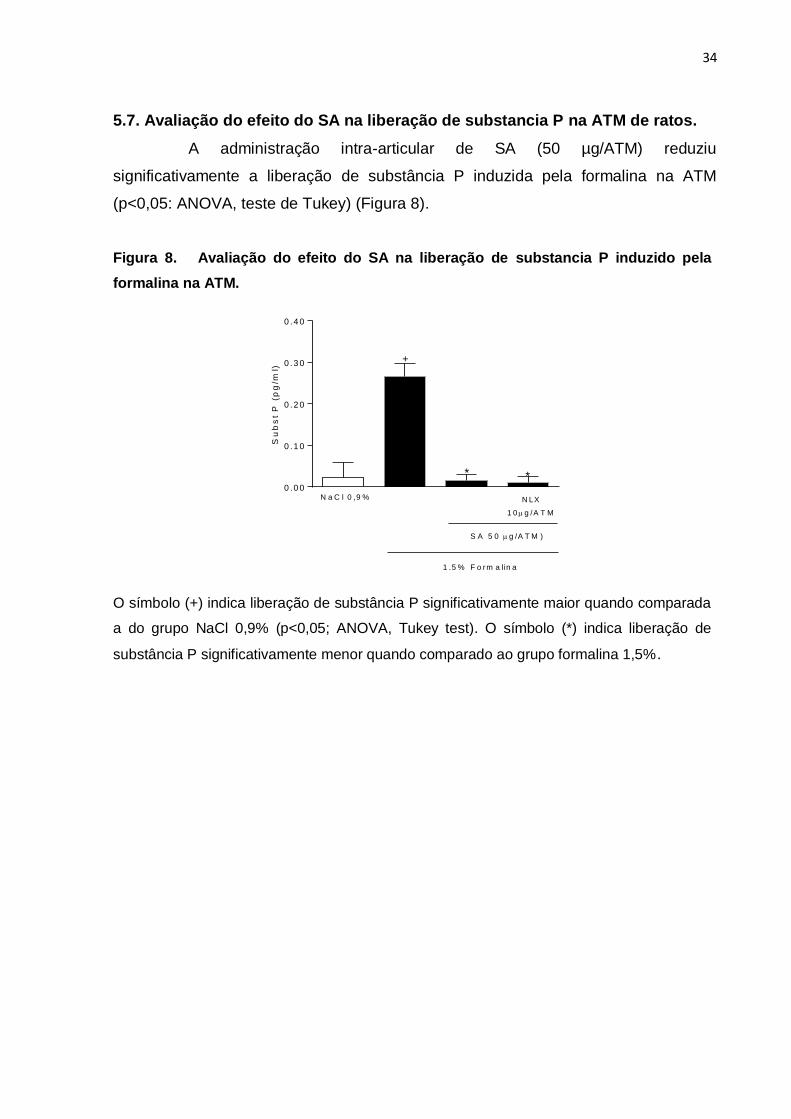
5.7. Avaliação do efeito do SA na liberação de substancia P na ATM de ratos.
A administração intra-articular de SA (50 µg/ATM) reduziu
significativamente a liberação de substância P induzida pela formalina na ATM
(p<0,05: ANOVA, teste de Tukey) (Figura 8).
Figura 8. Avaliação do efeito do SA na liberação de substancia P induzido pela
formalina na ATM. S
ub
st
P (
pg
/ml)
0 . 0 0
0 .1 0
0 .2 0
0 .3 0
0 .4 0
N a C l 0 ,9 %
S A 5 0 g /A T M )
1 .5 % F o rm a lin a
N LX
1 0 g /A T M
* *
+
O símbolo (+) indica liberação de substância P significativamente maior quando comparada
a do grupo NaCl 0,9% (p<0,05; ANOVA, Tukey test). O símbolo (*) indica liberação de
substância P significativamente menor quando comparado ao grupo formalina 1,5%.

35
6. DISCUSSÃO
O Óxido Nítrico (ON), um gás difusível natural, é produzido a partir da
conversão de L-arginina para L-citrulina através de isoformas de ON sintetases,
estando envolvido na regulação de várias funções fisiopatológicas (Cunha et al.,
2010; Paolocci et al., 2007). O nitroxil (HNO) é a forma reduzida em um elétron e
protonada do ON, recentemente vem se destacando como um novo congênere
redox, com distintas ações farmacológicas e vantagens terapêuticas. Evidências tem
demonstrado que perifericamente os doadores de HNO bloqueiam diretamente a
hiperalgesia inflamatória (Cury et al., 2011) e modulam a neurotransmissão da dor
(Cunha et al., 2010), conferindo assim ao HNO importante função biológica.
O Sal de Angeli (SA), ou trioxodinitrato de sódio (Na2N2O3) é o doador
sintético mais comumente utilizado no estudo biológico do HNO (Paolocci et al.,
2007). Tem sido demonstrado que o SA apresenta efeito antinociceptivo em modelos
de hiperalgesia mecânica induzida por uma variedade de estímulos como
carragenina, LPS, citocinas e PGE2 (Zarpelon et al., 2013). Corroborando com estes
estudos, os resultados deste trabalho demonstraram que o pré-tratamento com SA
foi capaz de reverter a hiperalgesia inflamatória induzida pela formalina na
articulação temporomandibular de ratos.
Em um estudo prévio, foi demonstrado que o SA inibiu a hiperalgesia
inflamatória induzida pela carragenina na pata de ratos através da ativação da via
intra-celular GMPc/PKG/K+ATP de forma independente da ativação de receptores
opioides periféricos (Zaperlon et al., 2013). No entanto, nossos resultados
demonstraram que o efeito antinociceptivo induzido pelo SA na ATM de ratos, pelo
menos em parte, é dependente de receptores opioides endógenos, subtipos capa,
delta e mμ; localizados na fibras C-nociceptivas aferentes primárias.
Diferenças entre tecidos subcutâneos e profundos, como os que
envolvem a ATM já vem sendo descrito pela literatura. Inicialmente, a inervação
predominante dos dois tecidos apresentam diferentes subconjuntos de fibras
neuronais primárias. Considerando que os tecidos profundos apresentam maior
excitabilidade de fibras nervosas centrais quando comparados aos tecidos
subcutâneos, pode-se concluir que distúrbios sensoriais podem resultar em mais
condições dolorosas em tecidos profundos quando comparados a tecidos
superficiais (Imbe et al., 2001). Sendo assim, é válido hipotetizar que os sistemas de

36
modulação de dor, como por exemplo, o sistema opioide, pode ser mais efetivo nos
tecidos profundos como os tecidos adjacentes da ATM.
O sistema opioide tem sido descrito como mediador do efeito
antinociceptivo de diferentes drogas quando administradas diretamente na ATM
(Clemente et al., 2004; Pena-dos-Santos et al., 2009; Araújo et al., 2017; Coura et
al., 2017). Uma vez ativados, os receptores opioides localizados nas fibras C-
nociceptivas primárias resultam na ativação da via intra-celular ON/GMPc/
PKG/K+ATP resultando em hiperpolarização destas células e consequentemente
analgesia (Clemente et al., 2009; Pena-dos-Santos et al., 2009). Sendo assim, seria
possível sugerir que o efeito antinociceptivo induzido pelo SA na ATM seria mediado
por ativação de receptores opioides (conforme demonstrado por este estudo) e da
via intra-celular GMPc/ PKG/K+ATP, como previamente descrito (Zarpelon et al.,
2013).
Os resultados do nosso estudo demonstraram que a administração de
inibidores seletivos para GMPc, PKG e canais K+ATP, não foram capazes de alterar a
resposta analgésica do SA (Figura 3), ou seja, o efeito antinociceptivo do SA na ATM
é dependente da ativação de receptores opioides mas não da ativação da via intra-
celular GMPc/ PKG/K+ATP. Frente a estes resultados pode-se sugerir que o SA possa
estar atuando diretamente em células residentes, como macrófagos, por exemplo,
liberando peptídeos opioides endógenos, e estes por sua vez, ativando os
receptores opioides periféricos das fibras C-nociceptivas primárias (Macedo et al.,
2016). Recentemente foi demonstrado que o SA é capaz de promover alteração
fenotípica em macrófagos, do tipo M1 (classicamente ativados), com características
pró-inflamatórias, para M2 (alternativamente ativados), de caráter resolutivo do
processo inflamatório através da liberação de peptídeos opioides endógenos
(Andrews et al., 2016).
A ativação de receptores opioides por peptídeos opioides endógenos está
associada com a ativação de outras vias intracelulares. Sabe-se que a ativação dos
receptores kappa-opóides (KOR) induz uma ativação de canais de potássio
simultaneamente com uma ação inibitória dos canais de cálcio, de forma a
hiperpolarizar a membrana neural (Bruchas et al., 2010). Além dessa ação direta
sobre os canais iônicos, a ativação dos receptores KOR leva a uma inativação da
síntese de AMPcíclico, inibindo a ativação da cascata de sinalização neuronal, como
por exemplo, ativação das proteinoquinases PKA e PKC, que são vinculadas a

37
processos pró-nociceptivos (Minneman & Iverson 1976; Bruchas et al., 2010).
O processo inflamatório na articulação temporomandibular é resultado da
liberação de citocinas pró-inflamatórias (Kopp, 2001), as quais contribuem para a
remodelação da articulação e degradação da cartilagem (Vernal et al., 2008). No
endotélio, o TNF-α e outras citocinas podem induzir uma regulação aumentada de
moléculas de adesão em leucócitos e células endoteliais, levando ao aumento da
migração do lúmen vascular para o tecido conjuntivo, potencializando a quimiotaxia
inflamatória (Moen et al., 2005).
Dentro desse contexto foi levantada a hipótese de que o efeito
antinociceptivo do SA poderia estar associado a um efeito anti-inflamatório.
Inicialmente os resultados demonstram que o SA reduziu significativamente a
migração de leucócitos induzida pela injeção intra-articular de formalina (Figura 4).
Sabe-se que a migração de leucócitos pode ser essencial para o aumento da
permeabilidade de parede do vaso, formação de exsudato e indução de dor induzida
por certos estímulos inflamatórios (Cunha et al., 2008). A adesão e transmigração do
leucócito dos vasos sanguíneos para dentro dos tecidos são reguladas pela ligação
de moléculas de adesão complementar encontradas na superfície do endotélio e
mediadores químicos.
A ICAM é uma molécula de adesão intercelular continuamente presente
em baixas concentrações nas membranas dos leucócitos e células endoteliais,
podendo ser induzida por interleucina-1 (IL-1) e fator de necrose tumoral (TNF), e é
expressa pelo endotélio vascular, macrófagos e linfócitos. Quando ativados, os
leucócitos se ligam às células endoteliais através de ICAM-1 e, em seguida migram
para os tecidos. (Zarbock et al., 2011). Considerando que o mecanismo do processo
inflamatório envolve adesão de células inflamatórias e sua transmigração mediada
pelos leucócitos do tipo β2 integrinas, os quais se interagem com imunoglobulinas
como a ICAM, levantamos a hipótese de que o efeito anti-inflamatório SA se deve,
pelo menos em parte, pela diminuição da expressão da ICAM. No entanto, nossos
resultados demonstraram que o SA não interfere na expressão da ICAM durante o
processo inflamatório induzido pela formalina na ATM de ratos (Figura 5A).
Apesar da ICAM-1 estar presente na maioria das células endoteliais e
estar diretamente envolvida na adesão e transmigração de células durante o
processo inflamatório, outros mediadores como o Fator de Aceleração de
Decaimento (também conhecido como CD55) também interferem neste processo. Ao

38
contrário da ICAM-1, o CD55 é uma molécula anti-adesiva que promove a
depuração da ligação dos leucócitos com as células endoteliais. Tem sido
demonstrado que existe uma interação entre o CD55 e a ICAM-1 (Shafren et al.,
2000), e esta interação limita a ligação de leucócitos com a ICAM-1. Sendo assim,
foi avaliado se a redução da migração de células inflamatórias pelo SA poderia estar
associada ao aumento da expressão da molécula CD55. Contrariando a hipótese, os
nossos resultados demonstraram que não há diferença na expressão do CD 55 entre
os grupos testados (Figura 5B).
Os trabalhos de Cunha et al. (1992) e Zarpelon et al. (2013) mostraram
que a inibição da migração de células polimorfonucleares leva a inibição da
sequência de liberação de mediadores da hipernocicepção inflamatória iniciados
pelo TNF-α, responsáveis.pela liberação de IL-1β e quimiocinas (Zarpelon et al,
2013).
Tem sido demonstrado que após a indução de um processo inflamatório,
o TNF- α é a primeira citocina a ser liberada, de modo que é considerada a citocina-
chave do processo inflamatório (Cunha et al., 1992). Nossos resultados
demonstraram que pré- tratamento com SA reduziu significativamente a liberação de
TNF- α, mas por outro lado não foi capaz de diminuir a liberação de IL-1β induzida
pela injeção intra-articular de formalina 1,5%.
O processo inflamatório envolve uma série de mediadores químicos
liberados no local da lesão que tem por finalidade sensibilizar os nociceptores.
Alguns dos principais mediadores inflamatórios que compõem a “sopa inflamatória”
são peptídeos (bradicinina), lipídios (prostaglandinas), neurotransmissores
(serotonina [5-HT] e ATP) e neurotrofinas (NGF). Estes mediadores são capazes de
sensibilizar/excitar os nociceptores aferentes primários, tendo como principal função
a transmissão aferente para o corno dorsal da medula espinhal. Além disso, essa
sensibilização permite através de uma função eferente dos nociceptores
denominada transporte antidrômico, iniciar o processo de inflamação neurogênica.
Neste processo, dois neurotransmissores têm grande destaque, Substância P (SP) e
do Peptídeo relacionado ao gene da calcitonina (CGRP), pois desempenham
funções cruciais para a manutenção do processo inflamatório como aumentar o fluxo
sanguíneo local e a permeabilidade vascular, promovendo a ativação de células
leucocitárias (mastócitos e neutrófilos), que contribuem para a manutenção deste
processo inflamatório (Snijdelaar et al., 2000; Julius D, Basbaum AI. 2001). Neste

39
contexto, nosso estudo demonstrou que o tratamento com SA foi capaz de inibir a
liberação de substância P no tecido periarticular da região temporomandibular de
ratos.
Frente a estes dados é possível sugerir que o efeito antinociceptivo do SA
na ATM está associado a ativação de células residentes, como macrófagos
residentes, resultando em dois efeitos: (1) aumentando a liberação de peptídeos
opioides endógenos, os quais teriam uma ação direta nos receptores opiodes
localizados nas fibras C-nociceptivas, hiperpolarizando estas células e reduzindo
uma ação antidrômica do neurônio, demonstrado através da redução de liberação de
SP, resultando em uma menor sinalização para a quimiotaxia inflamatória. (2)
diminuindo a liberação de TNF-α, o qual apresenta papel chave na sinalização da
migração leucocitária durante o processo inflamatório, além de reduzir vias de
segundos mensageiros relacionados com a hiperalgesia inflamatória (Verri et al.,
2006).
Este estudo demonstrou que a administração direta na ATM do doador de
nitroxila, Sal Angelis, foi capaz de induzir uma diminuição na hiperalgesia
inflamatória induzida pelo agente inflamatório formalina. Este efeito foi mediado pela
ativação de receptores opioides localizados nas fibras C-nociceptivas, diminuição da
migração de células inflamatórias e diminuição da liberação da citocina pró-
inflamatória TNF-α. Assim sendo o uso do Sal Angelis, surge como uma promessa
terapêutica como um potente efeito analgésico e anti-inflamatório mediado pelos
receptores opioides, mas independente da ativação da via intra-celular ON/GMPc.

40
7. CONCLUSÃO
Este estudo demonstrou que a administração direta na ATM do doador de
nitroxila, Sal Angelis, foi capaz de induzir uma diminuição na hiperalgesia
inflamatória e quimiotaxia inflamatória induzida pelo agente inflamatório formalina.
Este efeito foi mediado pela ativação de receptores opioides localizados nas fibras
C-nociceptivas, no entanto, independente da ativação da via intra-celular ON/GMPc.

41
8. REFERÊNCIAS*
Alstergrem P, Koop S. Prostaglandin E2 in temporomandibular joint synovial fluid and
its relation to pain and inflammatory disorders. J Oral Maxillofac Surg. 2000;
58(2)180-6.
Amantéa DV, Novaes AP, Campolongo GD, de Barros TP. A importância da
avaliação postural no paciente com disfunção da articulação temporomandibular.
Acta Ortop Bras. 2004; 12:155-9.
Anbar, M. and Gratt, B.M., The possible role of nitric oxide in the physiopathology of
pain associated with temporomandibular joint disorders, J OralMaxillofac Surg 1998;
56: 872-82.
Andrews KL, Lumsden NG, Farry J, Jefferis A-M, Kemp-Harper BK, Chin-Dusting
JPF. Nitroxyl: a vasodilator of human vessels that is not susceptible to tolerance. Clin
Sci. 2015; 129 (2):179–86.
Araújo IW, Chaves HV, Pachêco JM, Val DR, Vieira LV, Santos R, Freitas RS,
Rivanor RL, Monteiro VS, Clemente-Napimoga JT, Bezerra MM, Benevides NM.
Role of central opioid on the antinociceptive effect of sulfated polysaccharide from
the red seaweed Solieria filiformis in induced temporomandibular joint pain. Int
Immunopharmacol. 2017; 17(44):160-167.
Bouloux GF. Temporomandibular joint pain and synovial fluid analysis: a review of
the literature J Oral Maxillofac Surg. 2009; 67(11):2497-504.
Brenman JE. Synaptic signaling by nitric oxide, Curr. Opin. Neurobiol 1997; 7:374-
378.
Bruchas MR, Macey TA, Lowe JD, Chavkin C. Kappa opioid receptor activation of
p38 MAPK is GRK3- and arrestin-dependent in neurons and astrocytes. J Biol Chem.
2006; 281(26):18081-9.
Cairns, B.E. Pathophysiology of TMD pain--basic mechanisms and their implications
for pharmacotherapy.J. Oral Rehabil. 2010; 37:391-410.
* De acordo com as normas da UNICAMP/FOP, baseadas na padronização do International Committee of Medical Journal Editors - Vancouver Group. Abreviatura dos periódicos em conformidade com o PubMed.

42
Clemente, J.T., Parada, C.A., Veiga, M.C., Gear, R.W., Tambeli, C.H. Sexual
dimorphism in the antinociception mediated by kappa opioid receptors in the rat
temporomandibular joint. Neurosci. Lett. 2004; 372:250–255.
Clemente-Napimoga, J.T., Pellegrini-da-Silva, A., Ferreira, V.H., Napimoga, M.H.,
Parada, C.A., Tambeli, C.H. Gonadal hormones decrease temporomandibular joint
kappa mediated antinociception through a down- regulation in the expression of
kappa opioid receptors in the trigeminal ganglia. Eur. J. Pharmacol. 2009; 617:41-47.
Coura CO, Chaves HV, do Val DR, Vieira LV, Silveira FD, Dos Santos Lopes FM,
Gomes FI, Frota AF, Souza RB, Clemente-Napimoga JT, Bezerra MM, Benevides
NM. Int Mechanisms involved in antinociception induced by a polysulfated fraction
from seaweed Gracilaria cornea in the temporomandibular joint of rats. J Biol
Macromol. 2017; 97:76-84.
Cunha FQ, Poole S, Lorenzetti BB, Ferreira SH. The pivotal role of tumour necrosis
factor alpha in the development of inflammatory hyperalgesia. Br. J. Pharmacol.
1992; 107:660–664.
Cunha TM, Roman-Campos D, Lotufo CM, Duarte HL, Souza GR, Verri WA Jr,
Funez MI, Dias QM, Schivo IR, Domingues AC, Sachs D, Chiavegatto S, Teixeira
MM, Hothersall JS, Cruz JS, Cunha FQ, Ferreira SH. Morphine peripheral analgesia
depends on activation of the PI3Kgamma/AKT/nNOS/NO/KATP signaling pathway.
Proc. Natl. Acad. Sci. U. S. A. 2010; 107:4442-4447.
Cunha TM, Verri WA Jr, Schivo IR, Napimoga MH, Parada CA, Poole S, Teixeira
MM, Ferreira SH, Cunha FQ. Crucial role of neutrophils in the development of
mechanical inflammatory hypernociception.J Leukoc Biol. 2008; 83:824-832.
Cury Y, Picolo G, Gutierrez VP, Ferreira SH. Pain and analgesia: the dual effect of
nitric oxide in the nociceptive system. Nitric Oxide 2011; 25:243-257.
Douglas, CR. Fisiologia das Posturas Mandibulares. In: Patofisiologia oral: fisiologia
normal e patológica aplicada e odontologia e fonoaudiologia. v.1. São Paulo:
Pancast. 1998; p. 227-36.
Dubner R, Ren K. Brainstem mechanisms of persistent pain following injury. J Orofac
Pain. 2004; 18(4):299-305.
Ferreira SH, Duarte ID, Lorenzetti BB. The molecular mechanism of action of

43
peripheral morphine analgesia: stimulation of the cGMP system via nitric oxide
release. Eur. J. Pharmacol. 1991; 201:121-122.
Fukuto JM, Bartberger MD, Dutton AS, Paolocci N, Wink DA, Houk KN. The
physiological chemistry and biological activity of nitroxyl (HNO): the neglected,
misunderstood, and enigmatic nitrogen oxide. Chem Res Toxicol 2005; 18(5):790-
801.
Graff-Radford,S.B., Bassiur, J.P. Temporomandibular disorders and headaches.
Neurol. Clin. 2014; 32:525-537.
Imbe H, Iwata K, Zhou QQ, Zou S, Dubner R, Ren K. Orofacial deep and cutaneous
tissue inflammation and trigeminal neuronal activation. Implications for persistent
temporomandibular pain. Cells Tissues Organs. 2001; 169:238–247.
Ingawalé S, Goswami T. Temporomandibular joint: disorders, treatments, and
biomechanics. Ann Biomed Eng. 2009; 37(5):976-96.
Irvine JC, Ritchie RH, Favaloro JL, Andrews KL, Widdop RE, KempHarper BK.
Nitroxyl (HNO): the Cinderella of the nitric oxide story. Trends Pharmacol Sci. 2008;
29:601–608.
Iwata K, Tashiro A, Tsubol Y, Imai T., Sumino R., Morimoto,T., et al. Medullary dorsal
horn neuronal activity in rats with persistent temporomandibular joint and perioral
inflammation. J. Neurophysiol. 1999; 82: 1244-53.
Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001;
413:203–210.
Khasar SG, McCarter G, Levine JD. Epinephrine produces a adrenergic receptor-
mediated mechanical hyperalgesia and in vitro sensitization of rat nociceptors. J
Neurophysiol 1999; 81:1104–1112.
Kido, M.A.; Zhang, J.Q.; Muroya, H.; Yamaza, T.; Terada, Y. and Tanaka, T.:
Topography and distribution of sympathetic nerve fibers in the rat temporomandibular
joint: immunocytochemistry and ultrastructure. Anat Embryol (Berl). 2001; 203
(5):357-66.
Kopp S. Neuroendocrine, immune, and local responses related to
temporomandibular disorders. J Orofac Pain. 2001; 15:9–28.

44
Kumar S, Tansley-Hancock O, Sedley W, Winston JS, Callaghan MF, Allen M, Cope
TE, Gander PE, Bamiou DE, Griffiths TD, The Brain Basis for Misophonia. Curr Biol.
2017;27(4):527-533.
Langrehr JM, Hoffman RA, Lancaster JR Jr, Simmons RL. Nitric oxide, a new
endogenous immunomodulator, Transplantation. 1993; 55:1205-1212.
Leeuw R, Eisenlohr-Moul T, Bertrand P. The association of smoking status with sleep
disturbance, psychological functioning, and pain severity in patients with
temporomandibular disorders. J Orofac Pain. 2013; 27(1):32-41.
Longhi-Balbinot DT, Rossaneis AC, Pinho-Ribeiro FA, Bertozzi MM, Cunha FQ,
Alves-Filho JC, Cunha TM, Peron JP, Miranda KM, Casagrande R, Verri WA Jr.The
nitroxyl donor, Angeli's salt, reduces chronic constriction injury-induced neuropathic
pain. Chem Biol Interact. 2016; 256:1-8.
Macedo CG, Napimoga MH, Rocha-Neto LM, Abdalla HB, Clemente-Napimoga JT.
The role of endogenous opioid peptides in the antinociceptive effect of 15-deoxy
(Δ12,14)-prostaglandin J2 in the temporomandibular joint. Prostaglandins Leukot.
Essent. Fatty Acids. 2 016; 110:27- 34.
Mackay CR. Moving targets: cell migration inhibitors as new anti-inflammatory
therapies. Nat Immunol. 2008; 9:988–998.
McDougall JJ. Peripheral analgesia: Hitting pain where it hurts. Biochimica et
Biophysica Acta. 2011; 1812:459–467.
Millan MJ. The induction of pain: an integrative review. Prog Neurobiol. Oxford. 1999;
57(1):1-164.
Minneman KP, Iversen IL. Enkephalin and opiate narcotics increase cyclic GMP
accumulation in slices of rat neostriatum. Nature. 1976; 262:313–314.
Moen A, Sol J, Sampimon O. Indication of transmission of BVDV in the absence of
persistently infected (PI) animals. Prev Vet Med. 2005; 72(1-2):93-98
Nordahl, S, Alstergren, P, Kopp, S. Tumor necrosis factor-alpha in synovial fluid and
plasma from patients with chronic connective tissue disease and its relation to
temporomanibular joint pain. J Oral Maxillo fac Surg. 2000; 58(5):525-30.
Nunes Junior PC, Maciel RL. Propriedades anatômicas e funcionais da ATM com

45
aplicabilidade no tratamento fisioterapêutico. Fisioter Bras. 2005; 6(5):381-387.
Oliveira MC, Parada CA, Veiga MC, Rodrigues LR, Barros SP, Tambeli CH.
Evidence for the involvement of endogenous ATP and P2X receptors in TMJ pain.
Eur J Pain. 2005; 9:87–93.
Ozan F, Polat S, Kara I, Küçük D, Polat HB. Prevalence study of signs and
symptoms of temporomandibular disorders in a Turkish population. J Contemp Dent
Pract. 2007 May1; 8(4):35-42.
Paolocci N, Jackson MI, Lopez BE, Miranda K, Tocchetti CG, Wink DA, Fukuto JM.
The pharmacology of nitroxyl (HNO) and its therapeutic potential: not just the Janus
face of NO. Pharmacology & therapeutics. 2007; 113(2):442-458.
Pena-Dos-Santos DR, Severino FP, Pereira SA. Activation of peripheral κ/δ opioid
receptors mediates 15-deoxy- ∆12, 14-prostaglandin J2 induced antinociception in rat
temporomandibular joint. Neuroscience. 2009; 163:1211-9.
Prasad S R, Kumar N R, Shruthi H R, Kalavathi S D. Temporomandibular pain. J
Oral Maxillofac Pathol 2016; 20:272-5
Quinteiro MS, Henrique Napimoga M, Gomes Macedo C, Furtado Freitas F, Balassini
Abdalla H, Bonfante R, Trindade Clemente-Napimoga J. 15-deoxy-Δ12,14-
prostaglandin J2 reduces albumin-induced arthritis in temporomandibular joint of rats.
Eur J Pharmacol. 2014 Oct 5; 740:58-65. doi: 10.1016/j.ejphar.2014.07.002. Epub
2014 Jul 10.
Quinteiro MS, Napimoga MH, Mesquita KP, Clemente-Napimoga JT. The indirect
antinociceptive mechanism of 15d-PGJ2 on rheumatoid arthritis-induced TMJ
inflammatory pain in rats. Eur J Pain. 2012; 16(8):1106-15.
Raja SN, Meyer RA, Campbell JN. Peripheral mechanisms of somatic pain.
Anesthesiology. 1988; 68 (4): 571-90.
Rodrigues LL, Oliveira MC, Pelegrini-da-Silva A, de Arruda Veiga MC, Parada CA,
Tambeli CH. Peripheral sympathetic component of the temporomandibular joint
inflammatory pain in rats. J. Pain. 2006; 7:929–936.
Roveroni RC, Parada CA, Cecilia M, Veiga FA, Tambeli CH. Development of a
behavioral model of TMJ pain in rats: The TMJ formalin test. Pain. 2001; 94:185–191.

46
Sessle BJ. Acute and chronic craniofacial pain: brainstem mechanisms of nociceptive
transmission and neuroplasticity, and their clinical correlates. Crit Rev Oral Biol Med.
2000; 11:57–91.
Sessle BJ. Brainstem mechanisms underlying craniofacial pain and its modulation.
Adv Pain Res Ther 1995; 22:413–421.
Sessle, BJ. Peripheral and central mechanisms of orofacial inflammatory pain. Int.
Rev. Neurobiol. 2011; 97:179-206.
Shafren, D. R., D. J. Dorahy, R. F. Thorne, R. D. Barry. J. Gen. Virol. 2000; 8:889–
894.
Silveira AM, Feltrin PP, Zanetti RV, Mautoni MC. Prevalence of patients harboring
temporomandibular disorders in an otorhinolaryngology department. Braz J
Otorhinolaryngol. 2007; 73(4):528-32.
Snijdelaar DG, Dirksen R, Slappendel R, Crul BJ. Substance P. European Journal of
Pain. 2000; 4:121-135.
Sousa AM, Prado WA. The dual effect of a nitric oxide donor in nociception. Brain
Res. 2001; 897:9-19.
Staurengo-Ferrari L, Zarpelon AC, Longhi-Balbinot DT, Marchesi M, Cunha TM,
Alves-Filho JC et al. Nitroxyl inhibits overt pain-like behavior in mice: Role of
cGMP/PKG/ATP-sensitive potassium channel signaling pathway. Pharmacological
Reports. 2014; 66(4):691-698.
Su, N., Lobbezoo, F., van Wijk, A., van der Heijden, G. J. M. G. and Visscher, C. M.,
Associations of pain intensity and pain-related disability with psychological and socio-
demographic factors in patients with temporomandibular disorders: a cross-sectional
study at a specialised dental clinic. J Oral Rehabil 2017, 44: 187–196.
Suenaga, S., Abeyama, K., Hamasaki, A., Mimura, T. and Noikura, T.,
Temporomandibular disorders: relationship between joint pain and effusion and nitric
oxide concentration in the joint fluid, Dentomaxillofac Radio 2001; 30 214-8.
Vernal R, Vela´squez E, Gamonal J, Garcia-Sanz JA, Silva A, Sanz M. Expression of
proinflammatory cytokines in osteoarthritis of the temporomandibular joint. Arch Oral
Biol. 2008; 53:910–915.

47
Verri WA Jr, Cunha TM, Parada CA, Poole S, Cunha FQ, Ferreira SH.
Hypernociceptive role of cytokines and chemokines: targets for analgesic
drugdevelopment? Pharmacol Ther. 2006; 112(1):116-38.
Verri WA Jr, Cunha TM, Parada CA, Poole S, Cunha FQ, Ferreira SH..
Hypernociceptive role of cytokines and chemokines: Targets for analgesic drug
development? Pharmacology & Therapeutics. 2006; 112:116–138.
Wahl SM, McCartney-Francis N, Chan J, Dionne R, Ta L, Orenstein JM. Nitric oxide
in experimental joint inflammation. Benefit or detriment? Cells Tissues Organs. 2003;
174:26-33.
Widenfalk B, Wiberg M: Origin of sympathetic and sensory innervation of the
temporo-mandibular joint. A retrograde axonal tracing study in the rat. Neurosci Lett.
1990; 109 (1-2):30-5.
Yoshino K; Kawagishi S, Amano N. Morphological characteristics of primary sensory
and post-synaptic sympathetic neurones supplying the temporomandibular joint in the
cat. Arch Oral Biol. 1998; 43 (9):679-86.
Yun CW, et al. G-protein coupled receptor from yeast Saccharomyces cerevisiae.
Biochem Biophys Res Commun. 1997; 240(2):287-92.
Zarbock A, Ley K, McEver RP, Hidalgo A. Leukocyte ligands for endothelial
selectins: specialized glycoconjugates that mediate rolling and signaling under flow.
Blood. 2011; 118:6743–6751.
Zarpelon AC, Souza GR, Cunha TM, Shivo IRS, Marchesi M, Casagrande R. The
nitroxyl donor, Angeli’s salt inhibits inflammatory hyperalgesia in rats.
Neuropharmacolog. 2013; 71:1-9.
Zimmermann, M. Ethical guidelines for investigations of experimental pain in
conscious animals. Pain. 1983; 16:109-110.

48
ANEXO 1 – Certificado do Comitê de Ética no Uso de Animais
Recommended

![UNIVERSIDADE DE SÃO PAULO · UNIVERSIDADE DE SÃO PAULO FACULDADE DE CIÊNCIAS FARMACÊUTICAS DE RIBEIRÃO PRETO Efeito vasodilatador do doador de óxido nítrico [Ru(terpy)(bdq)NO]+3](https://img.pdfslide.tips/doc/110x75/5e4f79d3586fe6262a5b3335/universidade-de-sfo-paulo-universidade-de-sfo-paulo-faculdade-de-cincias-farmacuticas.jpg)















