Naturwissenschaften 81, 42-43 (1994) © Springer-Verlag 1994
Bees Perceive Illusionary Distance Information from Rotating Spirals W. H. Kirchner and J. Lengler
Theodor-Boveri-Institut ffir Biowissenschaften der Universitfit, Lehrstuhl fiir Verhaltensphysiologie und Soziobiologie, D-97074 Wfirzburg
Spatial vision, reconstruction of the three-dimensional world from the two- dimensional images on the retinae and estimation of distances of objects in the visual field, is in vertebrates mainly mediated by stereopsis, distance- dependent convergence of the eyes, and accommodation of the lenses. In insects, there a r e r a r e cases, in which stereopsis is used as it was shown in the praying mantis [8]. Generally, however, the spa- tial resolution of insect vision limits the use of stereopsis to distances not longer than one or two lengths of the insect's body [2]. Insects also do not have mov- able eyes and cannot adjust the focal length of their eyes to object distances. Thus, they have to rely on mechanisms different to those used by humans and other vertebrates to estimate distances. Exner [3] and Horridge [4] postulated that motion parallax should help moving insects to extract distance information from retinal images. Recently, more and more experimental evidence has been found to support this idea. In particular, is has been shown that freely flying honeybees estimate visual distances by using the speed of lateral image motion [6, 10]. Bees can learn to distinguish otherwise similar flower-like objects by the height above a background [7, 9]. It was therefore argued that the parallax between object and background is used to estimate distances, when bees fly at small distances over objects [7, 9]. How- ever, it is also possible that the apparent expansion and contraction of objects in the visual field during approach and departure are used to estimate distances. We therefore conducted an experiment to determine whether bees use looming as a cue to depth. In our experimental approach we used rotating spirals, which remain constant in size, but show either centrifugal or centripetal movement of contrast boundaries. This experimental layout also permitted us to distinguish between two alternative mechanisms of
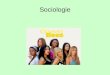
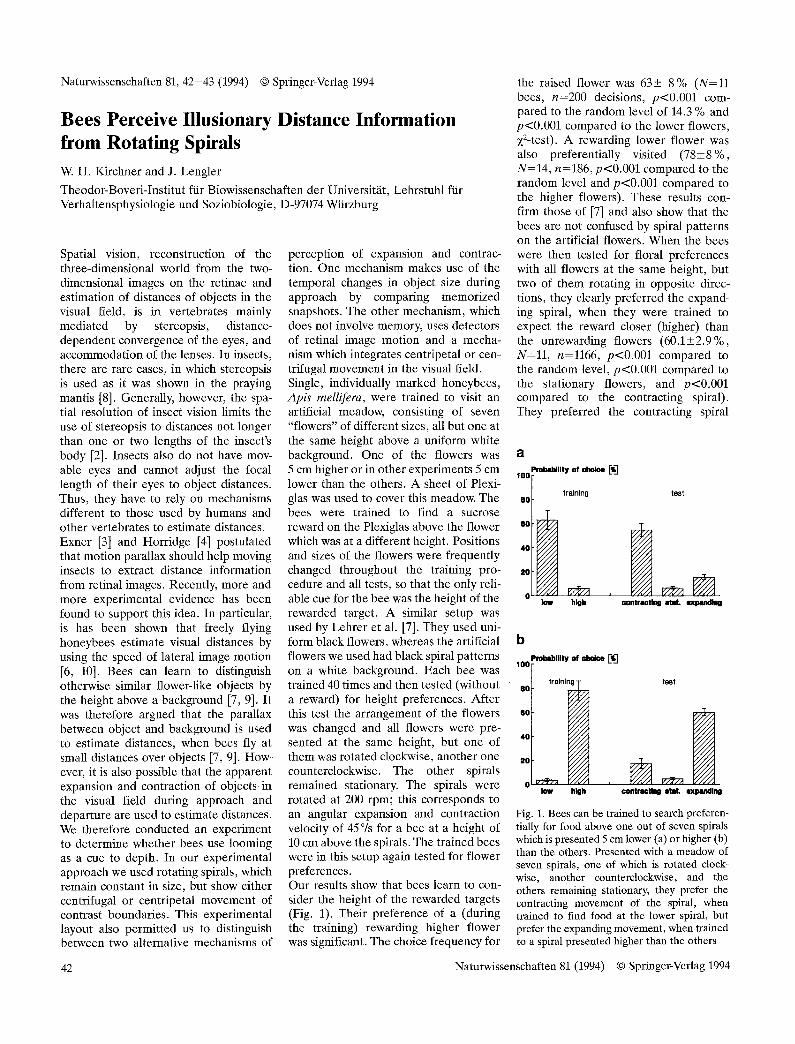
perception of expansion and contrac- tion. One mechanism makes use of the temporal changes in object size during approach by comparing memorized snapshots. The other mechanism, which does not involve memory, uses detectors of retinal image motion and a mecha- nism which integrates centripetal or cen- trifugal movement in the visual field. Single, individually marked honeybees, Apis mellifera, were trained to visit an artificial meadow, consisting of seven "flowers" of different sizes, all but one at the same height above a uniform white background. One of the flowers was 5 cm higher or in other experiments 5 cm lower than the others. A sheet of Plexi- glas was used to cover this meadow. The bees were trained to find a sucrose reward on the Plexiglas above the flower which was at a different height. Positions and sizes of the flowers were frequently changed throughout the training pro- cedure and all tests, so that the only reli- able cue for the bee was the height of the rewarded target. A similar setup was used by Lehrer et al. [7]. They used uni- form black flowers, whereas the artificial flowers we used had black spiral patterns on a white background. Each bee was trained 40 times and then tested (without a reward) for height preferences. After this test the arrangement of the flowers was changed and all flowers were pre- sented at the same height, but one of them was rotated clockwise, another one counterclockwise. The other spirals remained stationary. The spirals were rotated at 200 rpm; this corresponds to an angular expansion and contraction velocity of 45% for a bee at a height of 10 cm above the spirals. The trained bees were in this setup again tested for flower preferences. Our results show that bees learn to con- sider the height of the rewarded targets (Fig. 1). Their preference of a (during the training) rewarding higher flower was significant. The choice frequency for
the raised flower was 63+ 8% (N=11 bees, n=200 decisions, p<0.001 com- pared to the random level of 14.3 % and p<0.001 compared to the lower flowers, X2-test). A rewarding lower flower was also preferentially visited (78+8 %, N=14, n=186, p<0.001 compared to the random level and p<0.001 compared to the higher flowers). These results con- firm those of [7] and also show that the bees are not confused by spiral patterns on the artificial flowers. When the bees were then tested for floral preferences with all flowers at the same height, but two of them rotating in opposite direc- tions, they clearly preferred the expand- ing spiral, when they were trained to expect the reward closer (higher) than the unrewarding flowers (60.1+2.9%, N=11, n=1166, p<0.001 compared to the random level, p<0.001 compared to the stationary flowers, and p<0.001 compared to the contracting spiral). They preferred the contracting spiral
a
of choke i-qL-[ Probability 100
801 training
4O
2O
o r/Y~'A Imv high
test
oontrsotlflg stat. empandlng
b looPrObablllty of choice rir,~
low high
test
contrsotlrdl star. =n(pandlng
Fig. 1. Bees can be trained to search preferen- tially for food above one out of seven spirals which is presented 5 cm lower (a) or higher (b) than the others. Presented with a meadow of seven spirals, one of which is rotated clock- wise, another counterclockwise, and the others remaining stationary, they prefer the contracting movement of the spiral, when trained to find food at the lower spiral, but prefer the expanding movement, when trained to a spiral presented higher than the others
42 Naturwissenschaften 81 (1994) © Springer-Verlag 1994

after they had been rewarded on a flower further away (lower) than the unreward- ing flowers (54.8+5.5 %, N=14, n=865, p<0.001 compared to the random level, p<0.001 compared to the stationary flowers, and p<0.001 compared to the expanding spiral). The frequency of choices of the spiral rotating in the opposite direction was not statistically different from the random level (which is 14.3% with seven flowers), but signifi- cantly higher than the frequency of choices of the stationary flowers (p<0.005). These results show that bees consider expansion and contraction of the images of objects on their retinae during approach and departure and use them as a measure of distance. Since the size of the spirals remained constant during the tests, these results also show that the bees did not solve the problem by temporal comparisons of snapshots of objects, but by a mechanism which uses the move- ment of contrast boundaries to detect expansion and contraction. This suggests that the output of movement detectors may not only be integrated to detect homogeneous movement as it has been shown for the optomotor responses of flies, but that there may also be distinct units integrating centripetal and centrifu- gal components in the visual flow field as it has been suggested for the case of the landing reaction of flies [1]. An effect similar to the illusionary distance percep- tion from rotating spirals reported in the present study has been found in the sting- less bee species Trigona angustula [5], in which guard bees hovering in front of the
nest entrance respond to expanding and contracting patterns by moving away from and towards the entrance. Srinivasan and coworkers [7, 9], who demonstrated in a similar experiment for the first time that bees can discriminate between objects at different distances, concluded by exclusion that bees can estimate an object's distance by using its apparent motion across the retina when the background is featureless. Our results show that there is another cue, which bees can use (and might have used in the experiments described in [7] and [9]): retinal expansion and contraction of objects on approach and departure. It was shown [9] that a textured back- ground improves the distance discrim- ination performance of the bees signifi- cantly This indicates that retinal expan- sion and contraction cannot be the only cues bees use in the height dis- crimination paradigm, but that retinal object motion relative to the background is also being used. Since behavioral experiments using a different training paradigm [6, 10] unequivocally showed that freely flying bees make use of the angular velocity of retinal image motion to estimate visual distances, we have to conclude that three different computa- tional mechanisms are used by the bees to estimate depth, all of which, however, make use of visual flow-field informa- tion. Millions of years of evolution of the visual system of flying insects seem to have led, even in a single, particular spe- cies, to a number of algorithms making use of the output of the insects' move- ment detectors. This should not be too
surprising, since any new additional integration mechanism of the output of visual motion detectors requires no more than some additional synapses and a small set of specialized integrating neu- rons.
The experimental data presented here are from an undergraduate thesis of J. L., supervised by W. H. K. The authors thank C. Dreller, R. Gadagkar, M. V. Srinivasan, M. Wicklein, and J. Zeil for critically reading the manuscript.
Received October 5, 1993
1. Borst, A., Bahde, S.: Naturwissen- schaften 75, 265 (1988)
2. Burkhardt, D., Motte, I., de la: Ento- tool. Gener. 12, 221 (1987)
3. Exner, S.: Die Physiologic der facet- tierten Augen von Krebsen und Insek- ten. Leipzig 1891
4. Horridge, G. A.: Proc. R. Soc. Lond. B 229, 13 (1986)
5. Kelber, A., Zeil, J.: J. Comp. Physiol. A 167, 569 (1990)
6. Kirchner, W. H., Srinivasan, M. V.: Naturwissenschaften 76, 281 (1989)
7. Lehrer, M., Srinivasan, M. V., Zhang, S. W., Horridge, G. A.: Nature 332, 356 (1988)
8. Rossel, S.: ibid. 302, 821 (1983) 9. Srinivasan, M. V., Lehrer, M., Zhang,
S. W., Horridge, G. A.: J. Comp. Physiol. A. 165, 605 (1989)
10. Srinivasan, M. V., Lehrer, M., Kirch- her, W. H., Zhang, S. W.: Vis. Neurosci. 6, 519 (1991)
I I @LIBESPRECHUNGEN H a n d b u c h der Naturfarbstoffe . Vorkom- men - Verwendung - Nachweis. Von H. Schweppe. Landsberg: Ecomed 1992. 800 S., zahlr. Abb., DM 298,-. Die Nutzung von Naturfarbstoffen durch die Menschen war vor der Entwicklung der synthetischen Farbmittel fiber Jahr- tausende hinweg von groBer Bedeutung. Heute interessieren die Naturfarbstoffe aus kultur- und chemiehistorischer Sicht, bei der Analyse von gefiirbten histori-
schen Objekten wie Textilien oder Buch- malereien sowie entsprechenden arch/io- logischen Funden, bei der Restaurierung und Konservierung solcher Objekte, bei Verwendungen in kunsthandwerklichen Bereichen und sonstigen Anwendungen wie der Einf~rbung von Lebensmitteln und Kosmetika. Nicht zuletzt interes- siert sich auch allgemein die Naturstoff- chemie ftir die f~irbenden Stoffe aus der Pflanzen- und Tierwelt. Alle diese
Aspekte der Naturfarbstoffe werden im vorliegenden Handbuch yon H. Schweppe, einem Experten auf diesem Gebiet, umfassend behandelt; bisher hat es keine so /ibergreifende und aktuelle Darstellung dieses Gesamtgebietes gege- ben. Nach einer Einfiihrung befagt sich der Abschnitt II ausftihrlich und sehr infor- mativ mit der Geschichte der Verwen- dung yon F/irbepflanzen, Farbstoffinsek-
Naturwissenschaften 81 (1994) © Springer-Verlag 1994 43
Recommended