
1
Profesor Patrocinante Dr. Jaime Figueroa V. Instituto de Bioquímica y Microbiología Facultad de Ciencias
BÚSQUEDA Y ANÁLISIS DE GENES IMPLICADOS EN
RESISTENCIA A ANTIBIÓTICOS EN CEPAS NACIONALES DE Piscirickettsia salmonis
Tesis de Grado presentada como
parte de los requisitos para optar al
Título Profesional de Bioquímico
CARLOS JAVIER CARTES HENRÍQUEZ
VALDIVIA – CHILE
2013

2

3
AGRADECIMIENTOS
En primer lugar agradecer al Dr. Jaime Figueroa, por permitirme realizar esta tesis en su
laboratorio y aceptarme desde mis inicios en el laboratorio haciendo taller, además por
tener siempre buena disposición y paciencia para enseñarme y claro por toda su simpatía y
buenas historias.
Tambien agradecer a todo el equipo y amigos del Laboratorio de Biología Molecular de
Peces, Denise, pancha, nano, Adolflow, Willy, lolenza, mati, conchita, pablo, mary,
marchota, y Felipe, por su ayuda siempre que la necesité, su buena onda diaria, bullying y
multiples tallas que hicieron que la estadía en el laboratorio fuera muy agradable, divertida
y que no sea solo algo rutinario, además de los muy buenos carretes. Tambien a los que ya
no forman parte del lab, pero que si me ayudaron en mi formación cuando recién llegué a
aprender como Clara, alexei, tami y barbara.
Finalmente agradecer a mi familia, mis padres y hermana que siempre me apoyaron en mis
decisiones y en los momentos difíciles, además de mis amigos.
Este trabajo de tesis fue financiado con el patrocinio de los proyectos FONDECYT
1130069, FONDAP 11150027.

i
Indice general
1. Resumen ......................................................................................................................... 1
Abstract ............................................................................................................................... 2
2. Introducción ................................................................................................................... 3
2.1 Piscirickettsia salmonis ........................................................................................... 3
2.1.1 Generalidades ......................................................................................................... 3
2.1.2 Patología y signos clínicos de la enfermedad......................................................... 6
2.1.3 Mecanismos de transmisión ................................................................................... 8
2.1.4 Diagnóstico y tratamiento ...................................................................................... 8
2.2 Antimicrobianos .......................................................................................................... 10
2.2.1 Generalidades ....................................................................................................... 10
2.2.2 Florfenicol ............................................................................................................ 14
2.2.3 Oxitetraciclina ...................................................................................................... 15
2.3 Resistencia a antibióticos ............................................................................................ 16
2.3.1 Generalidades ....................................................................................................... 16
2.3.2 Resistencia a Oxitetraciclina ................................................................................ 18
2.3.3 Resistencia a Florfenicol ...................................................................................... 20
2.4 Antibiogramas ............................................................................................................. 22
2.5 Hipótesis y objetivos ................................................................................................... 24

ii
2.5.1 Hipótesis ............................................................................................................... 24
2.5.2 Objetivos .............................................................................................................. 24
3. Materiales y métodos ...................................................................................................... 25
3.1 Materiales .................................................................................................................... 25
3.1.1 Reactivos .............................................................................................................. 25
3.1.2 Soluciones ............................................................................................................ 26
3.1.3 Kit de reactivos..................................................................................................... 27
3.1.4 Material biológico ................................................................................................ 27
3.1.5 Equipos ................................................................................................................. 27
3.1.6 Software y herramientas on-line .......................................................................... 30
3.2 Metodología ................................................................................................................ 31
3.2.1 Identificación in-silico de genes de resistencia a antibióticos en las distintas cepas
de P. salmonis ............................................................................................................... 31
3.2.2 Medio de cultivo celular ....................................................................................... 32
3.2.3 Infección de cultivo celular con P. salmonis........................................................ 32
3.2.4 Cultivo líquido libre de células ............................................................................ 33
3.2.5 Cuantificación de P. salmonis .............................................................................. 33
3.2.6 Identificación P. salmonis .................................................................................... 34
3.2.7 Cultivo E. coli ATCC 25922 ................................................................................ 35
3.2.8 Diseño y estandarización de la CIM .................................................................... 35

iii
3.2.9 Cinéticas de expresión .......................................................................................... 41
3.2.10 Extracción de DNA genómico ........................................................................... 43
3.2.11 Extracción de RNA total de P. salmonis ............................................................ 43
3.2.12 Sintesis de cDNA ............................................................................................... 44
3.2.13 Análisis de la variación de expresión de los genes de P. salmonis mediante PCR
en tiempo real (qPCR) ................................................................................................... 45
4. Resultados .................................................................................................................... 49
4.1. Análisis bioinformático de genomas de P. salmonis ................................................. 49
4.1.1. Análisis comparativo in-silico entre genomas disponibles de cepas nacionales de
P. salmonis y la cepa tipo LF-89 con respecto a genes de resistencia a antibióticos .... 49
4.1.2. Comparación de secuencia de genes encontrados entre las cepas ...................... 55
4.1.3 Homología de secuencias analizadas con secuencias de otras especies bacterianas
....................................................................................................................................... 79
4.1.4 Caracterización del entorno génico de genes asociados a resistencia a antibióticos
en cepas LF-89, IBM-009, IBM-034 ............................................................................ 81
4.2 Determinación de las CIM de Oxitetraciclina y Florfenicol para cepas de P. salmonis
LF-89, IBM-009 e IBM-034 ............................................................................................. 84
4.2.1 Identificación de P. salmonis ............................................................................... 84
4.2.2 Determinación de la CIM para Oxitetraciclina en E. coli ATCC 25922 ............. 86
4.2.3 Determinación de la CIM para Oxitetraciclina y Florfenicol en cepa tipo LF-89,
IBM-034 e IBM-009 de P. salmonis ............................................................................. 86

iv
4.2.4 Determinación de la CIM para Florfenicol en E. coli ATCC 25922 ................... 88
4.2.5 Determinación de la CIM para Florfenicol en cepa tipo LF-89, IBM-034 e IBM-
009 de P. salmonis ........................................................................................................ 88
4.2.5 Efectos del Metanol y Etanol en las determinaciones de CIM ............................ 92
4.3.1 Estandarización de partidores diseñados para PCR en tiempo real ..................... 95
4.3.2 Evaluación de la cinética de expresión para genes de resistencia a antibióticos . 99
5. Discusión .................................................................................................................... 112
5.1 Identificación y análisis de genes posiblemente involucrados en resistencia a
Oxitetraciclina y Florfenicol ........................................................................................... 112
5.2 Determinación de sensibilidad y resistencia a Oxitetraciclina y Florfenicol en cepas
de P. salmonis ................................................................................................................. 119
5.3 Cinéticas de expresión .............................................................................................. 123
6. Referencias:................................................................................................................ 129

v
Indice de Figuras
Figura 1. Cladograma de la relación filogenética entre P. salmonis y microorganismos de
las divisiones alfa y gama proteobacterias.. ........................................................................... 5
Figura 2. Distribución de antimicrobianos utilizados en fase de agua de mar según
principio activo, periodo Enero-Septiembre de 2011 ........................................................... 13
Figura 3. Secuencia nucleotídica codificante para Tet E de P. salmonis. .......................... 56
Figura 4. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
putativa Tet E de P. salmonis.. ............................................................................................. 58
Figura 5. Secuencia nucleotídica codificante para Tet C de P. salmonis. ........................... 60
Figura 6. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
putativa Tet C de P. salmonis.. ............................................................................................. 62
Figura 7. Secuencia nucleotídica codificante para Bomba transportadora de tetraciclina de
P. salmonis. .......................................................................................................................... 64
Figura 8. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
putativa para Bomba transportadora de Tetraciclina de P. salmonis. .................................. 66

vi
Figura 9. Secuencia nucleotídica codificante para Bomba transportadora del tipo Bcr A
(Bcr/Cfla) de P. salmonis. .................................................................................................... 68
Figura 10. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
Bomba transportadora del tipo Bcr A (Bcr/Cfla) putativa de P. salmonis. .......................... 70
Figura 11. Secuencia nucleotídica codificante para Bomba transportadora del tipo Bcr 2
(Bcr/Cfla) de P. salmonis. .................................................................................................... 72
Figura 12. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
Bomba transportadora del tipo Bcr 2 (Bcr/Cfla) putativa de P. salmonis. ........................... 74
Figura 13. Secuencia nucleotídica codificante para Transportador de
Clorfanfenicol/Florfenicol de P. salmonis. .......................................................................... 76
Figura 14. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
transportadora de Cloranfenicol/Florfenicol putativa de P. salmonis.. ................................ 78
Figura 15. Analisis del entorno génico para los genes codificante para Bcr/Cfla y Tet C en
P. salmonis ........................................................................................................................... 83
Figura 16. Identificación de P. salmonis. ............................................................................ 85

vii
Figura 17. CIM de Oxitetraciclina para E. coli cepa ATCC 25922 y P. salmonis cepa LF-
89, IBM-034 e IBM-009.. ..................................................................................................... 87
Figura 18. CIM de Florfenicol para E. coli cepa ATCC 25922 y P. salmonis cepa LF-89,
IBM-034 e IBM-009. ............................................................................................................ 89
Figura 19. Efecto del Metanol y Etanol en el crecimiento de P. salmonis.......................... 93
Figura 20. Estandarización de partidores para genes de resistencia a Oxitetraciclina.. ...... 96
Figura 21. Estandarización de partidores para genes de resistencia a Florfenicol. ............. 98
Figura 22. Gráfico de cinética de expresión de Tet E. ...................................................... 101
Figura 23. Gráfico de cinética de expresión de Tet C. ...................................................... 103
Figura 24. Gráfico de cinética de expresión de bomba transportadora de Tetraciclina. ... 105
Figura 25. Gráfico de cinética de expresión de transportador Bcr/Cfla. ........................... 107
Figura 26. Gráfico de cinética de expresión de transportador del tipo Bcr2. ................... 109
Figura 27. Gráfico de cinética de expresión de transportador de Cloranfenicol/Florfenicol.
............................................................................................................................................ 111

viii
Indice de tablas
Tabla I. Volumen de medio, bacteria y antibióticos utilizados para preparar las distintas
concentraciones montadas en la técnica. .............................................................................. 37
Tabla II. Distribución de las concentraciones de cada antibiótico a analizar en la placa
ELISA de 96 pocillos. .......................................................................................................... 38
Tabla III.Concentración de antibiótico y tiempo de tratamiento en cepas de P. salmonis
LF-89, IBM-034 e IBM-009. ................................................................................................ 42
Tabla IV. Secuencia de partidores utilizados para RT-qPCR y su producto esperado. ...... 46
Tabla V.Genes putativos de resistencia a Oxitetraciclina encontrados en 9 cepas nacionales
de P. salmonis. ...................................................................................................................... 50
Tabla VI.Genes putativos de resistencia a Florfenicol encontrados en 9 cepas nacionales de
P. salmonis. .......................................................................................................................... 52
Tabla VII. Genes putativos de resistencia múltiple a drogas encontrados en 9 cepas
nacionales de P. salmonis. .................................................................................................... 54
Tabla VIII. Análisis in-silico de identidad aminoacídica de las proteínas putativas para
resistencia a Oxitetraciclina y Florfenicol con otras especies bacterianas. .......................... 80

ix
Tabla IX. Resumen de los resultados de CIM para Oxitetraciclina y Florfenicol en las
cepas estudiadas de P. salmonis ........................................................................................... 91

x
Lista de abreviaturas
ATCC: Colección Americana de Tipos de Cultivo
Buffer SB: Buffer Sodio-Borato
CFU: Colony-forming unit
CIM: Concentración Inhibitoria Mínima
DEPC: Dietilpirocarbonato
LB: Lysogeny Broth
Medio MH: Medio Mueller-Hinton
OD: Optical Density
Pb: Pares de base
PBS: Tampón fosfato salino
P. salmonis: Pisciricketssia salmonis
SBF: Suero Bovino Fetal
SHK-1: Salmon Head Kidney-1
SRS: Sindrome Rickettsial Salmonideo

1
1. Resumen
Piscirickettsia salmonis es el patógeno causante de la septicemia rickettsial salmonidea
(SRS) o piscirickettsiosis. Esta infección sistémica afecta principalmente a cultivos
salmonideos en el sur de Chile, llevando a grandes pérdidas económicas en la acuicultura
chilena.
Actualmente en la industria, para tratar P. salmonis se utilizan principalmente los
antibióticos Oxitetraciclina y Florfenicol, por lo que es de fundamental importancia el
conocimiento acerca del grado de resistencia o sensibilidad que cada uno de los subtipos de
P. salmonis presentan hacia estos, así como qué genes podrían estar involucrados en
conferirla.
El objetivo del presente estudio fue identificar genes que podrían estar involucrados en
conferir resistencia a estos antibióticos en distintas cepas de P. salmonis aisladas en Chile,
además de realizar estudios de sensibilidad antimicrobiana hacia estos y un posterior
análisis de expresión para 6 de los genes encontrados, realizando cinéticas frente a distintos
tratamientos con Oxitetraciclina y Florfenicol, las cuales se analizaron mediante RT-qPCR.
Los resultados experimentales permitieron la identificación in-silico de la secuencia
codificante de diversos genes posiblemente involucrados en conferir resistencia para los
antibióticos en estudio, además se pudieron observar diferencias en el genoma de las cepas.
El estudio de sensibilidad antimicrobiana permitió determinar que no todas las cepas
estudiadas presentan el mismo grado de resistencia para estos antibióticos, observándose
diferencias claras entre ellas. Finalmente el análisis de la expresión de estos genes frente a
tratamientos con los antibióticos en estudio permitió observar un aumento en la expresión
de estos en las primeras horas de desafío, lo que permite concluir que estarían actuando en
respuesta a los antibióticos, aumentando su expresión al ser sometidos a estos.

2
Abstract
Piscirickettsia salmonis is the pathogen that causes the Salmonid Rickettsial Septicemia
(SRS) or Pirickettsiosis. This systemic infection mainly affects salmonid farming in the
south of Chile, leading to important economic losses in the Chilean aquaculture.
Currently in the industry, Oxytetracycline and Florfenicol are primarily used to treat P.
salmonis, so it is critical knowledge about the degree of resistance or susceptibility that the
diferent strains of this bacteria has to these antimicrobials and also the genes that could be
involve in conferring this resistance.
The aim of this study was to identified genes that could be involve in conferring resistance
to these antibiotics in diferent P. salmonis strains isolated in Chile, also make tests about
the susceptibility to Oxytetracycline and Florfenicol and then analyze gene expression of
six of the genes founded, making kinetics against different treatments of this antibiotics,
wich were analyzed by RT-qPCR.
The experimental results allowed to indetify in-silico the coding sequence of genes that
could be involve in conferring resistance to the antibiotics in study and also genomics
differences could be seen among the strains. The susceptibility test to antibiotics allowed
determining that the strains in study doesn’t have the same resistance to these drugs and
have clear differences between them. Finally the analysis of gene expression showed that
these genes are up regulated during the first hours post treatment with antibiotics, leading to
the conclusion that these genes are acting in response to antibiotics, by an up regulation in
response to these.

3
2. Introducción
2.1 Piscirickettsia salmonis
2.1.1 Generalidades
Piscirickettsia salmonis gen. nov., sp. nov., es una bacteria patógena, la cual fue
descrita por Bravo y Campos en el año 1989, información que fue complementada por
Fryer et al. en 1992, ellos se refirieron a esta bacteria como un patógeno que era capaz de
infectar a Salmón Coho (Oncorhynchus kisutch), en las costas del sur de Chile y
denominaron a la enfermedad Pisciricketsiosis (Fryer et al.,1992), una enfermedad hoy
endémica que causa grandes pérdidas en la producción de salmones. En Chile se estiman
pérdidas en la acuicultura cercanas a los $100 millones anuales atribuidas a SRS (McCarthy
et al., 2008). Si bien la enfermedad fue reportada por primera vez en salmón Coho, su
infectividad ha sido demostrada en otras especies de salmonídeos cultivadas, como salmón
del Atlántico (Salmo salar), salmón Chinook (Oncorhynchus tshawytscha) y trucha
Arcoíris (Oncorhynchus mykiss) desde el sur de Chile al hemisferio norte (Rojas et al.,
2009).
El patógeno P. salmonis se ha ubicado en la clase de las Gamaproteobacterias (Fryer
et al., 2002) en el orden Thiotrichales, Familia Piscirickettsiacaea, Género Piscirickettsia
según la secuenciación del gen 16S rRNA (Fryer et al., 1992), secuenciación que permitió
un gran avance en dilucidar la taxonomía de esta bacteria. Dentro del género de las
Gamaproteobacterias se encuentran también los géneros Francisella, Coxiella y Legionella,

4
por lo que son estos géneros los más relacionados filogenéticamente con P. salmonis, como
se observa en la figura 1.
En un comienzo P. salmonis fue descrita como un patógeno intracelular obligado,
aunque actualmente se describe como intracelular facultativo, debido a la capacidad de
crecer en medios artificiales libres de células. Es una bacteria aerobia, Gram negativa,
pleomórfica aunque por lo general presenta morfología cocoide o en anillos, es inmóvil, no
encapsulada y presenta un diámetro aproximado entre 0.2 y 1.5µm (Bravo & Campos,
1989; Rojas et al., 2007). Al ser examinada por microscopia electrónica exhibe una típica
estructura protoplasmática de un procarionte y la pared celular de una bacteria Gram
negativa (Cvitanich et al., 1991). La bacteria se suele replicar por fisión binaria, unida a la
membrana de vacuolas citoplasmáticas en células hospederas de peces infectados o en
líneas celulares de peces, induciendo su efecto citopático característico (Mauel y Miller,
2002; Fryer y Hedrick, 2003). Descripciones realizadas por microscopia electrónica de
transmisión indican que este microorganismo posee dos membranas en su superficie, una
externa ondulada y otra interna cercana, esta descripción es característica de
microorganismos Rickettsiales, por lo que en un comienzo se solía clasificar junto a estos
organismos (Fryer et al., 1990). El DNA sería de tipo fibrilar y se localizaría en la región
central del microorganismo, según análisis por microscopia electrónica (Fryer et al., 1990).
In vitro se ha visto que su temperatura óptima para replicación es de 15-18°C, retardándose
sobre 20°C o bajo 10°C y no observándose por sobre los 25°C (Fryer et al., 1990).
Este patógeno que infecta principalmente a salmónidos entre otras especies
acuáticas presenta una amplia distribución, observándose en salmónidos del noreste y
noroeste del océano pacifico, además del noroeste del océano atlántico.

5
Figura 1. Cladograma de la relación filogenética entre P. salmonis y microorganismos
de las divisiones alfa y gama proteobacterias. Se utilizaron las secuencias del gen 16S
rRNA disponibles en la base de datos NCBI y se utilizó el algoritmo UPGMA (Adolfo Isla,
Laboratorio de Biología Molecular de Peces, Instituto de Bioquímica y Microbiología,
UACh).

6
2.1.2 Patología y signos clínicos de la enfermedad
La Septicemia Rickettsial Salmonidea (SRS) o pisciricketsiosis causada por
P. salmonis es una infección sistémica, afectando principalmente los cultivos de
salmonideos en el sur de Chile (Rojas et al., 2008), dentro de los signos patológicos
presentes en los salmónidos infectados se puede observar una coloración corporal más
oscura, además de inapetencia, nado letárgico y se suele observar que nadan cerca de las
superficies (Branson & Diaz-Munoz 1991; Cvitanich et al., 1991; Brocklebank et al., 1992;
Olsen et al.,1997; Evelyn et al., 1998).
También se pueden llegar a ver pequeñas manchas blancas que pueden progresar y
llevar a ulceras. Quizás el signo externo más observado durante la infección son branquias
pálidas resultantes de una anemia significativa, debido a que también se observa un
descenso del hematocrito (Branson & Diaz-Munoz 1991; Cvitanich et al., 1991). Otras
lesiones en la piel que se han descrito son erosiones y extensas áreas descamadas (Larenas
et al., 1995). Como características internas de la infección se ve un riñón hinchado y
descolorido, además de un bazo alargado. Hemorragias viscerales, estomacales, de vejiga y
musculares también pueden llegar a ser observadas (Schäfer et al., 1990; Cvitanich et al.,
1991).También se pueden observar hígados pálidos en los peces más afectados y nódulos
circulares color crema de 5-6 mm de diámetro (Branson & Diaz-Munoz 1991; Cvitanich et
al., 1991; Olsen et al., 1997).
Dentro de las características microscópicas de la patogenia se puede apreciar
necrosis en los hepatocitos del hígado y de las células hematopoyéticas en el parénquima
del riñón y bazo, observable en la fase más aguda de la enfermedad. La lámina propia del
intestino también es otro sitio común de necrosis, hemorragias e inflamación crónica. La

7
necrosis es seguida de una inflamación granulomatosa que puede ser observada en el
intersticio del riñón y bazo. Esta pérdida de células hematopoyéticas lleva a la anemia
característica de la piscirickettsiosis. También se puede evidenciar necrosis vascular y
perivascular en el hígado. Entre las áreas de necrosis, los macrófagos se pueden encontrar
albergando agregados intracelulares de P. salmonis. Esto puede ir acompañado de
endocarditis, peritonitis, pancreatitis y branquiatitis con cambios vasculares similares a los
observados en hígado y órganos hematopoyéticos. En algunos casos también se ha visto
involucrado el ovario, en salmones Coho infectados (Cvitanich et al., 1991). Recientemente
se ha podido purificar P. salmonis desde cerebro de salmón Coho infectado en Chile
(Skarmeta et al., 2000). Se vio que el comportamiento inusual en el nado de peces
infectados puede ser producto de infecciones de la P. salmonis en cerebro, de cepas
altamente virulentas.
La infección inicial posiblemente comienza cuando las barreras físicas de la piel y/o
las branquias son traspasadas por la bacteria (Almendras et al., 1997; Smith et al., 1999),
una vez que la bacteria se va replicando comienza el descoloramiento de las áreas
infectadas que posteriormente progresan a ulceras. Tras la dispersión de la bacteria a través
de las branquias se inician las infecciones locales que llevan a que se expandan vía
sanguínea a numerosos órganos (Almendras et al., 1997).
Se ha comprobado la supervivencia y multiplicación de P. salmonis en macrófagos
y monocitos, sin un daño evidente a las células hospederas. La capacidad de la bacteria para
sobrevivir y multiplicarse dentro de los fagocitos puede ser un mecanismo para ser
inasequible para el sistema inmune innato del salmónido y así evadir la primera línea de
defensa contra patógenos. De esta manera la supervivencia y proliferación de la bacteria
dentro de los macrófagos y monocitos puede contribuir al establecimiento de la

8
piscirickettsiosis. Una combinación del crecimiento intracelular de la bacteria y la
regulación de la muerte de la célula huésped, evitando así la destrucción del patógeno,
puede considerarse la base de la patogénesis de la P. salmonis (McCarthy et al., 2008).
2.1.3 Mecanismos de transmisión
El principal tipo de transmisión del patógeno P. salmonis en una infección natural
sería la transmisión horizontal (por contacto directo o indirecto entre peces) y no necesita
de un vector. Se han desarrollado trabajos en lo que se postula la transmisión vertical
(Larenas et al., 2003) mediante inoculación de P. salmonis en machos y hembras de trucha
arcoíris, sin embargo esta demostración solo se ha hecho in vitro, siendo además
observaciones poco concluyentes y no se ha observado en condiciones naturales.
El reservorio para la bacteria es desconocido, pero podría incluir una o más especies de
peces u otro animal acuático (Almendras et al., 1997). Mediante algunos reportes se ha
indicado que el principal agente de transmisión de esta bacteria serían tanto las orinas como
las heces de peces infectados, otros autores han señalado que este patógeno ingresaría al
nuevo hospedero a través de las branquias y/o heridas en la piel (Smith et al., 1999). Se ha
descrito también que P. salmonis podría utilizar isópodos hematófagos externos como
vectores de transmisión natural (Toranzo et al., 2005).
2.1.4 Diagnóstico y tratamiento
En lo que refiere al diagnóstico para la SRS, las alteraciones anátomo-patológicas
tanto macroscópicas como microscópicas previamente descritas no son suficientes para
obtener un diagnóstico definitivo de la enfermedad. Un diagnóstico presuntivo puede

9
realizarse en cortes histológicos o impresiones de tejido mediante la visualización del
patógeno dentro de macrófagos o hepatocitos, un diagnóstico confirmatorio se obtiene
aislando la bacteria en cultivos celulares (San Martin et al., 2010).
Para un diagnóstico confirmatorio se obtiene P. salmonis desde cultivos celulares, aunque
actualmente también se puede aislar esta bacteria en medios enriquecidos, sin la necesidad
de usar cultivos celulares. Otras pruebas utilizadas son detección mediante frotis o
impresiones de tejidos teñidos con tinción de Gram, Giemsa, Naranja de acridina o azul de
metileno. Si bien estas técnicas histoquímicas son ampliamente usadas no son específicas
(Almendras et al. 1997). Existen métodos más específicos para la detección de P. salmonis
como la inmunofluorescencia indirecta (IFAT) o la inmunohistoquímica (IHC), las cuales
son más específicas ya que se basan en el uso de anticuerpos. Los tejidos utilizados para un
diagnóstico adecuado son riñón, hígado y sangre, recolectados en infecciones clínicas o
subclínicas. Sin embargo otra técnica ampliamente utilizada, rápida, especifica y altamente
sensible es el PCR, la que permite una detección de P. salmonis a través de su DNA
genómico. Además esta técnica permite un análisis sobre las características taxonómicas y
ecológicas, poder determinar hospederos naturales, distribución geográfica entre otras
(Mauel et al., 1996).
En lo que se refiere al tratamiento, actualmente en el mercado nacional existen vacunas
autorizadas para el control de esta bacteria. En casos activos de esta enfermedad la
administración de antibióticos sigue siendo la principal vía de control para esta patología
(San Martín et al., 2010). Dentro de los tratamientos existentes en el registro de
medicamentos veterinarios se encuentran los antimicrobianos Oxitetraciclina, Florfenicol,
Ácido Oxolínico y Flumequina. También existen otros antibióticos que sin embargo no

10
están autorizados y no son usados actualmente en la industria, como por ejemplo
Rifampicina, Doxiciclina, o Minociclina.
2.2 Antimicrobianos
2.2.1 Generalidades
La principal herramienta usada actualmente en Chile para combatir infecciones
bacterianas y parasitarias en la salmonicultura son los antibióticos y los antiparasitarios. En
este ámbito estas infecciones son consideradas como una importante causa de pérdidas en
la productividad, debido a morbilidad y mortalidad de peces, reducción en los niveles de
producción, alteraciones reproductivas, altos costos en el control de estas, entre otras (San
Martín et al., 2010). Es necesario tener un conocimiento acerca de los antibióticos usados y
así también conocer el status de resistencia actual en la población bacteriana, debido a que
un buen uso de los antimicrobianos garantiza además de una eficacia clínica, una
disminución o incluso evitar la resistencia que los patógenos pueden adquirir ante estos,
además de disminuir su impacto ambiental y permitiendo así la entrega de un producto
inocuo al consumidor. Por lo descrito anteriormente es que establecer un uso prudente y
correcto de los antibióticos es fundamental, por lo que para la elección adecuada de este se
deben considerar los factores propios de la bacteria, del fármaco y del huésped. Es decir se
debe tener en consideración la etiología de la enfermedad, la sensibilidad que las bacterias
presenten a los antimicrobianos, la farmacocinética del antibiótico, el impacto ambiental
que produzca, etc.
Al tratarse de una bacteria intracelular, el tratamiento contra P. salmonis no es simple, por
lo que el mejor tratamiento sigue siendo la prevención y tomar medidas generales para el

11
control de las infecciones. Tanto para la prevención como para el tratamiento se han
utilizado diversas vacunas y antimicrobianos, desde los 90’s se han hecho ensayos in vitro,
donde se ha podido observar sensibilidad a Estreptomicina, Gentamicina, Tetraciclina,
Cloranfenicol, Eritromicina, Oxitetraciclina, Claritromicina, Sarafloxacina y resistencia a
Penicilina, Penicilina G y Espectinomicina (Fryer et al., 1990; Cvitanich et al., 1991;
Almendras et al., 1997), sin embargo se ha visto a través del tiempo que esta sensibilidad a
los antibióticos nombrados disminuye, probablemente debido a resistencia adquirida por la
bacteria (Cassigoli et al., 1994; Fryer et al., 2003). Actualmente los antimicrobianos más
utilizados son Ácido Oxolínico, Flumequina, Oxitetraciclina y Florfenicol los cuales son
administrados oralmente a los peces, sin embargo en los últimos años, en la industria
agropecuaria se ha visto una disminución en el uso de los dos primeros y un aumento en el
uso de Oxitetraciclina y Florfenicol.
Dentro de otras medidas preventivas para evitar un brote de Piscirickettsiosis también se
encuentran practicas simples como eliminar peces ya muertos o enfermos, evitar el estrés
excesivo en los peces en cultivo, disminuir la biomasa, entre otras (Almendras et al., 1997).
Según el informe sanitario del servicio nacional de pesca y acuicultura entregado el 2012
(Sernapesca), Piscirickettsiosis es la causa infecciosa mayormente asociada a las
mortalidades de las principales especies salmonideas producidas en el sur de Chile, además
presenta el mayor porcentaje de diagnósticos de laboratorio en centros marinos. Frente a
esta situación en diciembre de 2012 se estableció el programa específico de vigilancia y
control de Piscirickettsiosis, que está orientado a disminuir el impacto de la enfermedad,
mediante su detección temprana y aplicando medidas de control oportunas y graduales
(Sernapesca 2012).

12
Según datos entregados por Sernapesca en el año 2011, para el tratamiento de SRS se
destina un 79% del total de antimicrobianos usados en fase de agua de mar, siendo de estos
un 50% Florfenicol y un 47% Oxitetraciclina, porcentajes que se pueden observar en la
Figura 2 (Sernapesca 2011), es decir son los antimicrobianos más usados en Chile para esta
combatir esta enfermedad y por esa razón serán estudiados en esta tesis.

13
Figura 2. Distribución de antimicrobianos utilizados en fase de agua de mar según
principio activo, periodo Enero-Septiembre de 2011 (Informe sanitario de acuicultura,
SERNAPESCA 2011).

14
2.2.2 Florfenicol
Florfenicol es un antimicrobiano sintético perteneciente a la familia de los fenicoles,
es un análogo fluorado del Cloranfenicol, derivado de Tianfenicol. Es un antimicrobiano de
amplio espectro, similar al de Cloranfenicol y con un espectro mayor al de Tianfenicol de
acuerdo a pruebas in vitro (Wang et al., 2000). Es altamente lipofílico y su mecanismo de
acción consiste en inhibir la síntesis proteica de la bacteria, mediante inhibición de la
peptidil transferasa por unión reversible a la subunidad ribosomal 50 S, tratándose de un
antimicrobiano bacteriostático (Cannon et al., 1990). Es un compuesto neutro y liposoluble,
característica que le permite atravesar con facilidad las barreras celulares y por ende
alcanzar concentraciones intracelulares efectivas y difundir por todo el organismo con
rapidez, ya que se absorbe rápidamente. Debido a que presenta un espectro bacteriano hacia
las Gram negativas (además de Gram positivas) y a su capacidad de penetrar a nivel
intracelular es usado en infecciones causadas por P. salmonis. En cuanto a la vía de
administración de este antimicrobiano a los salmones, es preferentemente oral, aunque
también se puede administrar por vía parenteral.
Debido a que presenta un espectro más amplio y a que Cloranfenicol está limitado por su
toxicidad, Florfenicol podría ser un buen agente microbiano para reemplazar a
Cloranfenicol. Por otra parte se ha visto que cepas bacterianas que son resistentes a
Cloranfenicol son susceptibles a Florfenicol (Neu et al., 1980).

15
2.2.3 Oxitetraciclina
Oxitetraciclina o terramicina como se le llama comercialmente, es un
antimicrobiano perteneciente a la familia de las tetraciclinas, este antimicrobiano es de
amplio espectro exhibiendo actividad contra un amplio rango de bacterias Gram +/-. Es
bacterioestático y moderadamente lipofílico, pero lo suficiente para poder atravesar con
facilidad las barreras celulares y distribuirse por todo el organismo. Su mecanismo de
acción consiste en inhibir, en forma reversible, la síntesis de proteínas impidiendo la unión
del aminoacil-tRNA al sitio A del ribosoma bacteriano, uniéndose directamente a la
proteína S7 de la subunidad 30 S (Goldman et al., 1983). Para el caso de las bacterias Gram
negativas como P. salmonis, atraviesan la membrana externa a través de las porinas OmpF
y OmpC mediante difusión pasiva, asociado a un catión positivo (probablemente magnesio)
formando un complejo, este complejo se acumula en el periplasma donde se disocia,
permitiendo que la molécula no cargada de Oxitetraciclina difunda por la bicapa lipídica de
la membrana interna (Chopra et al., 2001; Michalova et al., 2004). Este antibiótico se aisló
en 1948 a partir de S. rimosus y según el orden de descubrimiento de las Tetraciclinas, las
Oxitetraciclinas se clasifican en el grupo de la primera generación de Tetraciclinas (Jara et
al., 2007).
Sus propiedades antimicrobianas favorables, además de no ocasionar efectos adversos han
llevado a que sean ampliamente usadas en terapias de infecciones tanto en humanos como
otros animales (Chopra et al., 2001). La vía de administración de este antimicrobiano en
salmones es preferentemente oral, pero también se pueden administrar por vía parenteral.
Se ha observado que la presencia de cationes divalentes como Fe+2
, Ca+2
o Mg+2
en los

16
alimentos disminuye la absorción de este fármaco, debido a que estos iones tienen un efecto
quelante que alteran la biodisponibilidad (San Martín et al., 2010).
2.3 Resistencia a antibióticos
2.3.1 Generalidades
La resistencia es una respuesta genético-evolutiva de las poblaciones bacterianas
expuestas a un estrés ambiental severo y continuo, como lo son las aplicaciones frecuentes
y/o continuas de antimicrobianos (San Martín et al., 2010). La evolución de la resistencia a
antibióticos encuentra su sustrato biológico en la variación genética producida por el
material genético bacteriano a través de fenómenos de mutación, intercambio de material
genético entre bacterias y la subsecuente selección de estas variantes por la presencia de
antibióticos en su entorno (Levin et al., 2000; Courvalin et al., 2001; Bushman et al., 2002;
Summers, 2002). Existen diversos mecanismos genéticos que permiten esta evolución,
estos incluyen los mecanismos de transformación, transducción, conjugación (intercambio
genético entre bacterias) además de transposición e integración (intercambio genético en la
misma bacteria) (Courvalin et al., 2001; Bushman et al., 2002), es decir este intercambio
genético puede ocurrir tanto entre bacterias patógenas como no patógenas, ya estén
estrechamente relacionadas o de diferentes orígenes evolutivos y/o fuentes ecológicas.
La rápida expresión de la variación genética en bacterias se ve favorecida por su rápida
multiplicación que produce un alto número de generaciones en un corto tiempo y también
por la haploidia de su material genético (Bushman et al., 2002), además el hecho de la
aparición de la resistencia a antibióticos, su selección y su diseminación en el ambiente

17
también es producto de el alto número de poblaciones bacterianas presentes en el ambiente
y la habilidad que tienen estas para poder ser transportadas a distintos nichos ecológicos a
través de diversos tipos de vectores.
Estos cambios evolutivos en las bacterias por lo general no son susceptibles de
modificación, por tanto la prevención de que se produzcan estas resistencias debería
centrarse en modificar las presiones selectivas de este fenómeno, teniendo como ejemplo
principal el del uso indiscriminado de antibióticos en las diversas actividades humanas
(Levy, 2001, Swartz, 2002).
Por esta razón es necesario supervisar el uso correcto de antibióticos tanto en medicina
veterinaria como humana, es decir mantener un uso responsable de estos y así evitar
detonar este proceso evolutivo que en consecuencia lleva a la resistencia bacteriana, y de
esta manera evitar una serie de problemas biológicos, sanitarios y económicos.
Dentro de estos problemas se encuentran el aumento tanto de la mortalidad como de la
morbilidad de las enfermedades producidas por estas bacterias resistentes, lo que conlleva a
un aumento importante de los costos de su tratamiento y su prevención (Cabello et al.,
2003). En el caso de la acuicultura, el costo económico de la resistencia a antibióticos va
más allá de tan solo la medicina y la salud humana, ya que bacterias patógenas para peces
que presenten resistencia a diversos tratamientos llevan a grandes pérdidas económicas en
la acuicultura misma.
Que actualmente el problema de la resistencia a antibióticos en Chile sea un problema serio
de salud pública, con proyecciones sanitarias y económicas muy importantes ha sido
resultado del uso indiscriminado de antibióticos en la medicina humana, veterinaria y en la
agricultura (Cabello et al., 2003). Se ha dado cuenta de la presencia de bacterias

18
potencialmente patógenas con resistencia múltiple a antibióticos en el entorno acuático que
rodea varios recintos de la acuicultura salmonera (Miranda et al., 2002).
En Chile hay un importante uso de antibióticos en la industria acuícola, sin embargo hay un
muy bajo número de publicaciones científicas respecto de los patógenos frecuentemente
aislados y de las susceptibilidades que estos tienen a los diversos antibióticos (Miranda et
al., 2001). Esta ausencia de información sugiere que el uso de antibióticos en la acuicultura
nacional no está basado en la regla dorada de la terapia antibiótica, la cual especifica que el
uso de un antibiótico debe estar sujeto a un conocimiento de la especie bacteriana y de su
susceptibilidad hacia ese compuesto (Cabello et al., 2003). Como manera de poder adoptar
de forma oportuna y eficaz estrategias efectivas de control hacia la resistencia a
antibióticos es importante determinar de manera prematura cambios en los patrones de
sensibilidad bacteriana a los antimicrobianos usados en su tratamiento, de manera de poder
determinar cómo está afectando actualmente este problema a la salmonicultura en Chile.
2.3.2 Resistencia a Oxitetraciclina
Existen mecanismos genéticos y bioquímicos que explican la resistencia a
Oxitetraciclina en bacterias. En 1980 Méndez y su equipo examinaron por primera vez
determinantes genéticos de resistencia a tetraciclinas desde plásmidos en miembros de
Enterobacteriaceae y Pseudomonadaceae (Méndez et al., 1980).
Dos genes se consideran de la misma clase y tienen la misma designación si tienen una
identidad en su secuencia aminoacídica mayor a 80% y son considerados distintos genes sin
tienen menos de un 79% de identidad en su secuencia aminoacídica (Levy et al., 1999).

19
Estos genes que confieren resistencia a Tetraciclinas y Oxitetraciclinas se denominan genes
tet y otr, no existe diferencia inherente entre un gen de resistencia a tetraciclina y uno de
resistencia a Oxitetraciclina. La mayoría de estos genes codifica para bombas de flujo de
flujo de salida (efflux proteins), pero también existen los que codifican para genes de
protección ribosomal o protección enzimática (Ridenhour et al., 1996; Roberts et al., 1996).
En cuanto a las proteínas de flujo de salida, son las proteínas tet mejor estudiadas y
pertenecen a la superfamilia MFS (Major Facilitator Superfamily) (Paulsen et al., 1996).
Estos genes codifican para proteínas transmembrana de alrededor de 46 kDa que exportan
las moléculas de Tetraciclina y/o Oxitetraciclina desde la célula reduciendo de esta manera
la concentración intracelular de la droga y protegiendo así los ribosomas. Se encuentran
presentes en Gram - y Gram +. Se han dividido en seis grupos de acuerdo a su identidad
aminoacídica.
Presentan entre 12 y 14 alfa hélices transmemabrana y presentan similitudes con otras
proteínas de flujo de salida involucradas en resistencia múltiple a drogas, resistencia a
cloranfenicol y quinolonas, entre otras (Chopra et al., 1991; Levy et al., 1992).
También existen genes que codifican para otro tipo de resistencia a tetraciclinas,
denominadas proteínas de protección ribosomal, las cuales son proteínas citoplasmáticas
que protegen a los ribosomas de la acción de estos antimicrobianos. Estas proteínas
presentan homología a los factores de elongación EF-Tu y EF-G (Sánchez-Pescador et al.,
1988; Taylor et al., 1996) siendo mayor esta homología en el extremo amino terminal que
contiene el dominio de unión a GTP. Estas proteínas se unen al ribosoma lo que causa una
alteración en la conformación de este que previene a la molécula de tetraciclina de unirse al
ribosoma, sin alterar o detener la síntesis de proteínas, luego se disocia para que los factores
de elongación puedan unirse ya que poseen sitios de unión sobrelapados en el ribosoma

20
(Chopra et al., 2001). Confieren un mayor espectro de resistencia al que se ve en bacterias
que presentan proteínas de flujo de salida. También se han visto genes (aunque muy pocos)
que codifican para proteínas de inactivación enzimática de Tetraciclinas. Esta es una
proteína citoplasmática de 44 kDa que modifica químicamente la molécula de tetraciclina
en presencia de oxígeno y NADPH (Speer et al., 1991).
2.3.3 Resistencia a Florfenicol
A través de los años las bacterias han desarrollado mecanismos para evitar los
efectos inhibitorios de Florfenicol y Cloranfenicol. El primer y mayor tipo de resistencia
que se ha visto principalmente para cloranfenicol es la de inactivación enzimática por
acetilación de la molécula del antimicrobiano, mediante distintos tipos de cloranfenicol
acetiltransferasas (CATs) (Murray et al., 1997). Sin embargo se ha visto que las CATs solo
presentan actividad acetiltransferasa hacia Cloranfenicol, Tianfenicol y Azidamfenicol,
pero no hacia Florfenicol, esto debido a que la modificación se produce en el C-3 de la
molécula, sitio aceptor que esta estructuralmente modificado en Florfenicol con respecto a
Cloranfenicol. Trabajos han demostrado que bacterias que poseen resistencia a
Cloranfenicol solo basado en CATs son susceptibles a Florfenicol (Cannon et al., 1990).
También existen otros tipos de resistencia, como lo son las bombas exportadoras de
Cloranfenicol y Florfenicol, se ha visto que esta exportación puede ser por transportadores
específicos o bien por transportadores múltiples de drogas. Los transportadores específicos
por lo general aportan un mayor nivel de resistencia que los de múltiples de drogas, se
conocen alrededor de 8 grupos de transportadores específicos denominados de E-1 hasta E-
8, en los cuales los grupos E-3 y E-4 se ha visto que aportan resistencia a Florfenicol y

21
Cloranfenicol a la vez. Se han descrito en la literatura genes pertenecientes al grupo E-3
entre los que destacan pp-flo, cmlA-like, floSt, flo y floR. Otro tipo de resistencia a
Cloranfenicol y Florfenicol está dada mediante inactivación por fosfotransferasas,
mutaciones puntuales en el sitio objetivo (target) y disminución de permeabilidad de la
membrana externa (Shaw, 1983; Murray et al., 1997). Muchos de los genes que codifican
para alguno de estos tipos de resistencia se encuentran en elementos móviles en el genoma,
como plásmidos, transposones o cassettes y por lo general se encuentran acompañados de
otros genes de resistencia a algún otro antimicrobiano o a metales pesados (Schwarz et al.,
2004).
Además de estos dos tipos de resistencia específicos para Oxitetraciclina y Florfenicol
descritos en los puntos 1.3.2 y 1.3.3 respectivamente, existen bombas exportadoras de
resistencia múltiple a drogas, estas proteínas confieren una resistencia simultánea a distintas
drogas, lo que se denomina fenotipo MDR (Multi Drugs Resistance). Estas bombas
transportadoras actúan sinérgicamente con otros mecanismos de resistencia para proveer un
elevado nivel de resistencia en la bacteria.
Tanto las bombas transportadoras específicas para un tipo de drogas o bien transportadoras
múltiples de drogas, son proteínas de membrana y han sido caracterizadas en cinco
familias, estas son: superfamilia ABC (ATP-Binding Cassette), superfamilia MFS (Major
Facilitator Superfamily), familia MATE (toxic compound extension), familia SMR (Small
Multidrug Resistance) y superfamilia RND (Resistance-Nodulation-Division) (Li et al.,
2009).

22
2.4 Antibiogramas
Por las razones que se señalaron previamente una selección apropiada de los
antibióticos a utilizar contra determinado microorganismo es de fundamental importancia,
dentro de esta elección un rol sumamente importante juegan las pruebas de sensibilidad a
antibióticos. El objetivo de estos estudios de sensibilidad es evaluar de manera in vitro la
respuesta de un microorganismo determinado a uno o varios antibióticos. Para evaluar este
parámetro existen diversos métodos, siendo los principales el método de kirby-bauer o
prueba de difusión en agar y las pruebas de dilución en placas o pruebas de concentraciones
inhibitorias mínimas. La concentración inhibitoria mínima (MIC) se refiere a la
concentración mínima de antibiótico in vitro necesario para evitar el crecimiento de la
bacteria, de esta manera se seleccionan aquellos antibióticos a los que la bacteria demuestra
ser sensible.
El método de Kirby-Bauer consiste en crecer las bacterias en medios agares adecuados que
permitan su crecimiento, en el cual se colocan discos de antibióticos con distintas
concentraciones. Tras horas adecuadas de incubación, dependiendo de la sensibilidad de la
bacteria hacia el antimicrobiano presente en el disco se verá la formación de halos
dependiendo si el crecimiento de la bacteria es inhibido o no. Este método entrega datos
cualitativos acerca de la sensibilidad de la bacteria hacia ese antimicrobiano, lo que se
clasifica en sensible, intermedio y resistente, tras la medición del halo en mm. (Bauer &
Kirby et al., 1996).
El método de diluciones en placas consiste en determinar las concentraciones inhibitorias
mínimas o MIC (Minimun Inhibitory Concentration), es decir la concentración más baja a
la cual un agente microbiano evita el crecimiento visible de microorganismos (Miller,

23
Walker et al, 2005). El método de dilución en microplacas fue el utilizado en esta tesis y
presenta ventajas por sobre la prueba de difusión en gel de agar, en las que destacan que
además de entregar datos cualitativos, entrega datos cuantitativos. Otra ventaja que presenta
es que permite el trabajo con bacterias que crecen más lento o son de naturaleza más
fastidiosa (Miller, Walker et al., 2005), y es la prueba de elección frente a las características
de P. salmonis (Fryer et al., 2003). Otra ventaja es que presenta mejor poder predictivo de
la eficacia in vivo que las pruebas de difusión en gel de agar (Salomon et al., 1998).

24
2.5 Hipótesis y objetivos
2.5.1 Hipótesis
Las diferencias genómicas tanto de presencia, como de expresión de distintos genes de
resistencia a antibióticos en las cepas nacionales de P. salmonis, determinan la sensibilidad
que estas cepas presenten a diferentes antibióticos, y este factor está eventualmente
relacionado con su patogenicidad.
2.5.2 Objetivos
Objetivo General:
Buscar genes determinantes en la resistencia a antibióticos en los genomas disponibles de
cepas nacionales de P. salmonis.
Objetivos Específicos:
1- Analizar comparativamente in silico los genomas disponibles de las cepas
nacionales con la cepa tipo LF-89 respecto a genes de resistencia a antibióticos.
2- Caracterizar los genes asociados a resistencia a antibióticos encontrados en el
objetivo 1 de 3 cepas de P. salmonis.
3- Establecer un sistema de prueba de sensibilidad/resistencia a antibióticos de las 3
cepas seleccionadas mediante curvas de crecimiento/inhibición en cultivo líquido.
4- Cuantificar y comparar patrones de expresión de genes de resistencia a antibióticos
de las 3 cepas de P. salmonis mediante RT-qPCR.

25
3. Materiales y métodos
3.1 Materiales
3.1.1 Reactivos
Merck: Aceite de inmersión, Ácido clorhídrico, Albumina de suero bovino, Caldo Mueller-
Hinton, Cloroformo, Etanol, EDTA, Glicina, GoTaq Flexi, GoTaq qPCR Master, Inhibidor
de RNAsa, Isopropanol, M-MLV Transcriptasa reversa, Hidroxido de Sodio, Cloruro de
Potasio.
Winkler: Albumina de suero bovino, Cloruro de sodio, Fosfato de Potasio monobásico,
Sacarosa.
Promega: Random Primer, GoTaq Flexi, M-MLV Reverse Transcriptase, Nucleótidos
(dNTPs), Inhibidor RNasa, RQ1 RNAse-Free DNAse, GoTaq qPCR Master Mix.
UsBio: Tris base, L-cisteina.
Becton Dickinson: Tinción de Gram, Bacto Agar, Bacto Triptona.
Fermentas: Marcador de peso molecular para DNA.
Ambion: Agua DEPC, Kit RiboPure-Bacteria, DNase I (RNase Free), Trizol.

26
Thermo Scientific: Medio Leibovitz L-15
Invitrogen: Syber-Safe, Tripsina 0,5%
Arquimed: Etanol
Sigma: Glucosa, Inhibidor de proteasas, Oxytetracycline Hydrochloride, Florfenicol.
Promega: Kit wizard.
Lonza: Agarosa.
3.1.2 Soluciones
Buffer SB 25X: 10 mM NaOH, ajustado a pH 8 con ácido bórico.
Caldo Mueller-Hinton: Mueller-Hinton 21g/L autoclavado.
Medio suplementado: Medio L-15 suplementado con 10% de suero bovino fetal.
Medio Caso suplementado: Medio Caso suplementado con L-Cisteina, suero bovino fetal y
cloruro de fierro.
Medio LB-Agar: medio LB, Bacto-agar 15 g/L. Autoclavado

27
PBS: NaCl 136.89 mM, KCl 2.68 mM, Na2HPO4 10.14 mM, KH2PO4 1,76 mM, pH 7.4.
3.1.3 Kit de reactivos
Extraccion genómico wizard, Promega (USA).
GoTaq® qPCR Master Mix, Promega (USA).
Genomic DNA purification Kit, Promega (USA).
LightCycler® FastStart DNA Marter SYBR Green I, Roche (USA).
3.1.4 Material biológico
Microorganismo patógeno intracelular facultativo Pisciricketssia salmonis cepa LF-89
(ATCC VB 1361), IBM-034 e IBM-009.
Cepa tipo E. coli American Type Culture Collection (ATCC) N° 25922.
Línea Celular derivada de leucocitos de riñón anterior de Salmo salar, SHK-1.
3.1.5 Equipos
Agitador Orbital: Ika big-squid, Ikamäleon.
Autoclave: Orthman.

28
Balanza: Satorius TE4101.
Baño Termo-regulado: N-Biotec.
Cámara de electroforesis: Biorad Mini-Protean III.
Cámara de electroforesis horizontal: Labnet Enduro.
Cámara de Flujo Laminar: Nuaire NU425-400E.
Cámara de Transferencia: Labnet.
Centrifuga: Sigma 2-16 multigene.
Centrifuga termorregulada: Sigma 2-16pk.
Centrifuga, Boeco C-28A. 35
Centrifuga Hitachi Himac CR22GII, High Speed Refrigerated
28
Centrifuga Hitachi Himac CP20WX, Preparative Ultracentrifuge
Espectrofotómetro: Thermo Scientific Evulution 60
Espectrofotómetro de placas de ELISA: BioTek Synergy 2
Fuente de poder: Biorad Power PacTM Universal Power Supply
Fuente de poder: Enduro E0303
Homogenizador de Vidrio Potter-Elvehjem 5mL Weathon Science
Incubador: Zhicheng ZSD 1270

29
Incubador-Shaker: Zhicheng ZHWY-200B
Microcentrifuga: Sigma 1-14
Microondas: Somela Faney WT1700
Micropipetas: Axygen, Labnet.
Microscopio: LW-Scientific I4 Series
Microscopio: Olympus BKX41.
Nanoview: General Electric.
pH metro: InoLab.
Propipeta Fastpette v2: Labnet.
Shaker: Zhcheng, ZHWY-200B
Termoblock: Labnet
Termociclador: Eppendorf Mastercycler personal.
Termociclador en gradiente: Multigene, Labnet.
Termociclador qPCR: Stratagene MX 3000P y MX 3005.
Termociclador: Eppendorf Mastercycler personal.
Transiluminador UV: Syngene, INGENIUS.
Vortex: Brarnstead International.

30
3.1.6 Software y herramientas on-line
Libre Office 3.5
Microsoft Office Excel 2010
SigmaPlot 11.0
MxPro
PerlPrimer
ARDB (Antibiotic Resistance Genes Database)
GenDB 2.2
MAUVE
http://www.ncbi.nlm.nih.gov/ National Center for
Biotechnology Information (NCBI)

31
3.2 Metodología
3.2.1 Identificación in-silico de genes de resistencia a antibióticos en las distintas cepas
de P. salmonis
Se utilizó la herramienta bioinformática GenDB para realizar un análisis in-silico en los
genomas de las cepas de P. salmonis LF-89, IBM-009 e IBM-034 con el fin de rastrear
genes putativos de resistencia a antibióticos. Este análisis fue complementado mediante la
comparación de las secuencias nucleotídicas y aminoacídicas de los posibles genes
encontrados con genes de resistencia a antibióticos registrados en la base de datos ARBD y
en NCBI. Para confirmar la presencia de los genes encontrados en estas cepas se realizaron
alineamientos de estos genes utilizando un sitio web exclusivo para realizar BLAST entre
cepas de P. salmonis.
Tras obtener las secuencias codificantes de estos genes se estudiaron los dominios
conservados de cada una utilizando la herramienta bioinformática on-line SMART, junto
con la aplicación Conserved Domains and Protein Clasification (NCBI).
El porcentaje de identidad de estas secuencias con secuencias de otras especies bacterianas
fue analizada utilizando la información presente en la base de datos de NCBI, utilizando la
herramienta BLAST, donde se determinaron las secuencias con mayor identidad
aminoacídica, query coverage y un e value menor.
Para rastrear el entorno génico de cada uno de los genes evaluados se utilizó la herramienta
bioinformática GenDB 2.2 analizando estos genes para cada cepa en estudio.

32
3.2.2 Medio de cultivo celular
El cultivo de la línea celular SHK-1 fue realizado en botellas de cultivo de 25 y 150 cm2, se
utilizó medio de cultivo Leibovitz´s L-15 suplementado con SBF 10%. Las células fueron
mantenidas en estufa incubadora a una temperatura entre 15-17ºC.
Las células fueron subcultivadas cada 15 días y sembrando en las botellas de cultivo células
con 70%-80% de confluencia. Al alcanzar este porcentaje de confluencia las células se
encuentran listas para ser infectadas con P. salmonis.
3.2.3 Infección de cultivo celular con P. salmonis
Se despertaron las cepas congeladas a -80°C de P. salmonis utilizando la línea celular
SHK-1 en medio de cultivo Leibovitz´s L-15 suplementado con SBF 10%. Para esto se
descongelaron alícuotas de P. salmonis (cepa LF-89, IBM-034 o IBM-009), una vez
descongeladas rápidamente se centrifugaron a 5.000 xg por 7 minutos y posteriormente se
eliminó el sobrenadante. Luego se procedió a lavar las células, resuspendiéndolas
cuidadosamente con 500µL de medio L-15 10% SBF y tras esto centrifugándolas a 4.000
xg por 10 minutos. Se eliminó el sobrenadante y nuevamente se procedió a resuspender con
500µL de medio L-15 10% SBF y con esta suspensión bacteriana se infectó una botella de
25 que contenían la línea células SHK-1. Se realizó la misma metodología para cada
una de las tres cepas utilizadas en esta tesis. La botella fue incubada a 20°C por 5 días
aprox. dependiendo de la cepa bacteriana y su rapidez de infección.

33
3.2.4 Cultivo líquido libre de células
El cultivo de P. Salmonis cepa LF-89, IBM-034 e IBM-009 en medio liquido libre de
celulas, fue realizado en medio CASO suplementado. Para esto tras aproximadamente 5
días de infección celular, al observar evidente efecto citopático, las bacterias se liberaron al
medio, se tomaron alícuotas de 150 µL y 200 µL del medio y fueron agregadas a 8 µL de
cultivo líquido CASO suplementado en tubos Falcon de 50mL. Cada tubo Falcon con
medio líquido con bacterias fue almacenado en un incubador shaker a 18°C protegido de la
luz a 150rpm, durante aproximadamente 4-5 días dependiendo de la cepa, hasta que estas
alcanzaran un OD600 de 0,6-0,8. Este medio fue preparado de acuerdo a la descripción
realizada por Vera et al 2012.
3.2.5 Cuantificación de P. salmonis
Las bacterias crecidas en medio líquido libre de células fueron cuantificadas por
turbidimetría, para esto se tomó una alícuota de 1µL de cada tubo y se midió su OD a
600nm. Como blanco se utilizó medio CASO suplementado. Para el cálculo del número de
bacterias se utilizó la fórmula de McFarldand (McFarland 1907)
OD 600nm 0,132 1.5x108 CFU/mL
OD 600nm muestra X CFU/mL

34
3.2.6 Identificación P. salmonis
3.2.6.1 Tinción de Gram
Se tomó 1µL de bacteria crecida en medio líquido y se centrifugo a 4.000 xg por 5 minutos,
se eliminó el sobrenadante y el precipitado de bacterias se resuspendió con PBS 1X, para
realizar la tinción de Gram, tomando una alícuota de 10-20µL y se depositó en un porta
objetos. Una vez fijado, el frotis fue cubierto con tinción primaria (cristal violeta) por 2
minutos, luego se removió con agua destilada para posteriormente agregar Lugol el que se
mantuvo sobre la muestra por 1 minuto. Se retiró lavando con agua destilada y se agregó el
agente decolorante (alcohol) por 15 segundos y finalmente se agregó la Safranina y se
realizó la tinción por 30 segundos. Se lavó con agua destilada y se dejó secar para
finalmente examinar el frotis bajo un lente de inmersión con aceite (100X) corroborando la
morfología bacteriana y si son Gram positivos o Gram negativos en el caso de P. salmonis
se tratan de cocos Gram negativos.
3.2.6.2 PCR confirmatorio
Se tomó una alícuota un 1µL de la cepa correspondiente, a la cual se le realizó extracción
de DNA genómico según el kit Wizard de Promega y luego se verificó mediante PCR
confirmatorio según Mauel (Mauel et al., 1996). Los productos de este PCR anidado se
fragmentaron en una corrida electroforética en un gel de agarosa al 1% y fueron
visualizados con luz UV para corroborar que eran del tamaño deseado (500pb).

35
3.2.7 Cultivo E. coli ATCC 25922
La bacteria fue crecida en una placa de agar LB. Se pasó una colonia a 5mL de medio
Mueller-Hinton y se dejó crecer a 37°C por 24 horas previas a su uso. Se evaluó mediante
Gram que se tratara de bacilos Gram negativos.
3.2.8 Diseño y estandarización de la CIM
3.2.8.1 Preparación de antibióticos
Los antibióticos utilizados en esta tesis fueron Oxitetraciclina y Flofenicol, se prepararon
soluciones stock de 8mg/mL para ambos antibióticos, para posteriormente realizar las
distintas diluciones necesarias para el experimento. Se pesó la cantidad correspondiente de
Oxitetraciclina y se disolvió en metanol absoluto. Para Florfenicol se hizo lo mismo pero se
disolvió en etanol 95%. Una vez preparados ambos antibióticos se almacenaron a -20°C
para evitar su deterioro siguiendo las condiciones de almacenamiento descritas por Sigma.
Se mantuvieron fuera de la luz y su antigüedad no debe superar las dos semanas.
3.2.8.2 Montaje de la técnica en E. coli ATCC 25922
La cepa fue crecida en medio Mueller-Hinton hasta alcanzar una absorbancia de 0,8 a
600nm, posteriormente se realizaron microdiluciones de esta cepa basándose en el método
de diluciones en microplacas para determinar la CIM en bacterias acuáticas (Miller &
Walker, 2005).

36
De la solución stock de cada antibiótico (8 mg/mL) se procedió a preparar las distintas
soluciones patrones de antibióticos (80 µg/mL y 0,8 µg/mL), estas soluciones fueron
utilizadas para preparar la concentración necesaria de antibiótico en cada uno de los
pocillos, tal como se especifica en la tabla I.

37
Tabla I. Volumen de medio, bacteria y antibióticos utilizados para preparar las distintas
concentraciones montadas en la técnica.
Concentración final de AB
(µg/ml) Medio MH (µl) Antibiótico (µl) Bacteria (µl) Stock de AB usado
(µg/ml)
Control s/ AB (+) 180 0 20 -
Control s/bacteria (-) 200 0 0 -
0,02 175 5 20 0,8
0,04 170 10 20 0,8
0,08 160 20 20 0,8
0,125 155 25 20 80
0,25 167,5 12,5 20 80
0,5 155 25 20 80
1 130 50 20 80
2 170 10 20 80
4 160 20 20 80
8 140 40 20 80

38
Para este experimento se utilizaron placas ELISA de 96 pocillos estériles con tapa, se
evaluaron concentraciones desde 0,02 µg/mL hasta 8 µg/mL, además de un control positivo
sin antibiótico y un control negativo que solo contiene medio. El diseño de este
experimento se detalla en la tabla II. Los cultivos fueron crecidos a 37°C por 19 horas tal
cómo se especifica en la publicación de Miller. Posteriormente se leyeron las absorbancias
a 600nm en el lector de microplacas utilizando el software asociado al equipo para analizar
los datos los que posteriormente fueron graficados utilizando el software SigmaPlot 11.0.

39
Tabla II. Distribución de las concentraciones de cada antibiótico a analizar en la placa
ELISA de 96 pocillos.
1 2 3 4 5 6 7 8 9 10 11 12
Oxitetraciclina
A 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
B 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
C 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
D 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
Florfenicol
E 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
F 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
G 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +
H 8 4 2 1 0,5 0,25 0,125 0,08 0,04 0,02 Ctrl - Ctrl +

40
3.2.8.3 Montaje de la técnica en P. salmonis
Tras ser infectadas las células de la línea celular SHK-1 como se describe en el punto 3.2.3,
se tomaron alícuotas de esta y se pasaron a medio líquido como se describe en el punto
3.2.4, estas se incubaron a 18°C hasta que alcanzaron absorbancias de 0,8 a 600nm. Estas
muestras fueron identificadas mediante Gram y PCR confirmatorio para ver que se trataba
de P. salmonis. Posteriormente las muestras se diluyeron con medio CASO hasta alcanzar
absorbancias de 0,08 y se procedió a realizar el experimento al igual que en el montaje de la
técnica para E. coli. Microdiluciones en placas ELISA de 96 pocillos estériles con tapa,
preparadas de la misma manera en cómo se describe en la Tabla 1, con la diferencia que se
utilizó medio CASO suplementado. La distribución en la placa es la misma que se muestra
en la Tabla 2, las cuales fueron incubadas a 18°C por 5 días, para su posterior lectura en el
lector de microplacas y su análisis correspondiente mediante SigmaPlot 11.0.
3.2.8.4 Análisis del efecto de los solventes metanol y etanol 95% en los resultados de la
CIM
El efecto que puedan generan los solventes de los antibióticos, metanol y etanol 95% en el
crecimiento de la bacteria también fue evaluado, se utilizó el mismo montaje descrito en las
tablas 1 y 2, con la diferencia fundamental que en lugar del antibiótico se utilizó agua
destilada estéril.

41
3.2.9 Cinéticas de expresión
Para evaluar los cambios en la expresión de determinados genes que sufren las distintas
cepas de P. salmonis en el tiempo tras ser tratadas con distintas concentraciones de
antibióticos se diseñó experimentalmente un ensayo de cinética en el tiempo desafiando a
las bacterias a dos concentraciones distintas tanto de Oxitetraciclina como de Florfenicol.
Se crecieron las bacterias en medio CASO suplementado a 18°C hasta que alcanzaran una
absorbancia de 0,6-0,8 a 600nm, una vez alcanzada esta fase exponencial se trataron con
los antibióticos correspondientes tal como se describe en diseño experimental en la tabla
III. En cada punto se colectaron alícuotas de 1µL para realizar una posterior extracción de
RNA y síntesis de cDNA con el fin de evaluar finalmente los cambios de expresión en el
tiempo frente a distintas concentraciones de antibiótico mediante RT-qPCR.
Estas cinéticas fueron realizadas por separado para las cepas LF-89, IBM-034 e IBM-009.

42
Tabla III.Concentración de antibiótico y tiempo de tratamiento en cepas de P. salmonis
LF-89, IBM-034 e IBM-009.
Antibiótico Concentración
(µg/mL) Tiempo
- - 0h
Oxitetraciclina 0,02 2h
Oxitetraciclina 0,02 4h
Oxitetraciclina 0,02 6h
Oxitetraciclina 0,08 2h
Oxitetraciclina 0,08 4h
Oxitetraciclina 0,08 6h
- - 0h
Florfenicol 0,02 2h
Florfenicol 0,02 4h
Florfenicol 0,02 6h
Florfenicol 0,08 2h
Florfenicol 0,08 4h
Florfenicol 0,08 6h

43
3.2.10 Extracción de DNA genómico
Se realizó extracción de DNA genómico desde P. salmonis cepa LF-89, IBM-034 e IBM-
009 crecidas en medio CASO suplementado, cuando se encontraban en un OD 0,6
utilizando el Kit Wizard de Promega, de acuerdo a las indicaciones del fabricante.
3.2.11 Extracción de RNA total de P. salmonis
Transcurrido los tiempos de las cinéticas (Tabla III), se realizó la extracción de RNA total
para cada punto de la cinética de las cepas de P. salmonis evaluadas crecidas en medio
Caso suplementado, en un OD600 de 0,6-0.8. Se colecto 1mL de este medio que se
centrifugó a 14.000 xg por 3 minutos. Posteriormente se realizó extracción de RNA total
desde el sedimento celular, utilizando el Kit de Trizol. El sedimento celular fue
resuspendido en 1mL de Trizol, luego se agregaron 0,2mL de cloroformo mezclando
suavemente e incubando por 3 minutos a temperatura ambiente. Se centrifugó a 12.000 xg
por 15 minutos a 4°C, transfiriendo la fase acuosa a un nuevo tubo y precipitando con
0,6mL de isopropanol (+ glicógeno), se incubó por 5 minutos a temperatura ambiente y se
centrifugó a máxima velocidad por 10 minutos a 4°C. El pellet fue lavado con etanol 70% y
posteriormente se centrifugó a 7.500 xg por 5 minutos a 4°C (paso que se repitió 2 veces) y
finalmente el sedimento se dejó secar por 15 minutos a temperatura ambiente para luego
resuspenderlo con agua DEPC.
El RNA fue cuantificado por espectrofotometria a 260nm utilizando el equipo NanoVue
(General Electric).
Luego de realizar la extracción de RNA total, se procedió al enriquecimiento de la muestra,
eliminando el DNA genómico presente en la muestra, para ello se utilizó la DNAsa

44
(Ambion) realizando tratamientos de una hora, de acuerdo a las indicaciones del fabricante.
Para confirmar la eliminación del gDNA se realizó una reacción de PCR, amplificando el
gen GROE de P. salmonis, realizando una reacción previa al paso de tratamiento con
DNAsa y una posterior a este, de manera que se espera que para el producto amplificado
para el templado post tratamiento con DNAsa no aparezca, a diferencia del producto del
templado previo al tratamiento, demostrando así que en la muestra solo hay RNA y no
gDNA.
3.2.12 Sintesis de cDNA
La síntesis de la primera hebra de cDNA, fue realizada por transcripción reversa (RT-PCR),
utilizando M-MLV Reverse Transcriptase (Promega) y Random Primer (Promega) de
acuerdo a las indicaciones del fabricante. Se hicieron en un volumen de 20µL, agregando
1µL de mix de dNTPs (10mM), 2µL de random primers (500µg/mL), 1µg de RNA total y
agua libre de nucleasas hasta un volumen final de 14,5µL. La mezcla se dejó incubar por 10
minutos a 60°C y luego se enfrió directamente en hielo previniendo así la formación de
estructuras secundarias. Luego se procedió a añadir 1µL de transcriptasa reversa, 4µL de
buffer de transcriptasa reversa 5X y 0,5µL de inhibidor de RNAsas. La mezcla se incubó
por 60 minutos a 37°C seguido por 10 minutos a 70°C. Finalmente Los cDNA fueron
almacenados a -20°C.

45
3.2.13 Análisis de la variación de expresión de los genes de P. salmonis mediante PCR
en tiempo real (qPCR)
3.2.13.1 Diseño de partidores para qPCR
Tras obtener las secuencias para los genes codificantes para Tet E, Tet C, Bomba
transportadora de tetraciclinas, Bcr A, Bcr2 y Transportador de Flofenicol mediante las
herramientas bioinformáticas descritas para P. salmonis, se procedio a diseñar partidores
heterólogos para qPCR para estos genes mencionados, utilizando el software PerlPrimer.
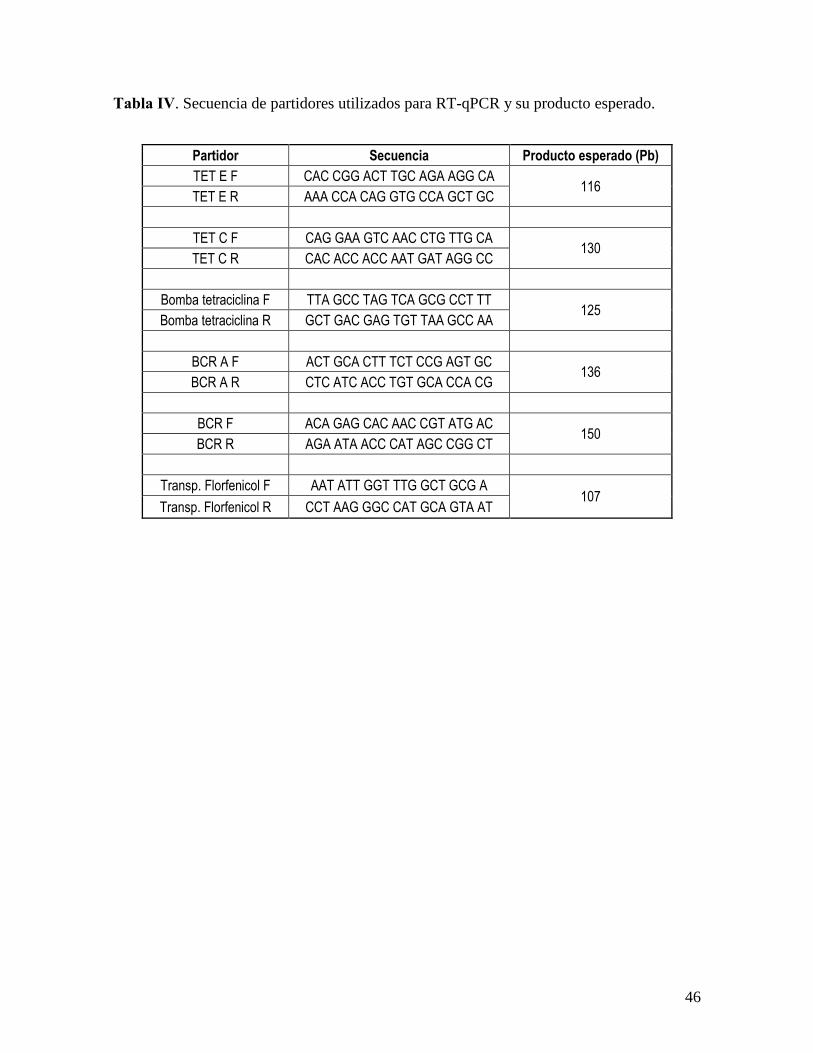
Los partidores se describen a continuación en la Tabla IV.
Una vez diseñados los partidores se procedio a evaluar las diferencias de expresión de estos
genes en las tres cepas evaluadas de P. salmonis.
46
Tabla IV. Secuencia de partidores utilizados para RT-qPCR y su producto esperado.
Partidor Secuencia Producto esperado (Pb)
TET E F CAC CGG ACT TGC AGA AGG CA 116
TET E R AAA CCA CAG GTG CCA GCT GC
TET C F CAG GAA GTC AAC CTG TTG CA 130
TET C R CAC ACC ACC AAT GAT AGG CC
Bomba tetraciclina F TTA GCC TAG TCA GCG CCT TT 125
Bomba tetraciclina R GCT GAC GAG TGT TAA GCC AA
BCR A F ACT GCA CTT TCT CCG AGT GC 136
BCR A R CTC ATC ACC TGT GCA CCA CG
BCR F ACA GAG CAC AAC CGT ATG AC 150
BCR R AGA ATA ACC CAT AGC CGG CT
Transp. Florfenicol F AAT ATT GGT TTG GCT GCG A 107
Transp. Florfenicol R CCT AAG GGC CAT GCA GTA AT

47
3.2.13.2 Condiciones Experimentales
Una vez crecidas las bacterias hasta su fase exponencial (OD=0,6) en medio CASO
suplementdo se evaluó la variación de la expresión de los seis genes descritos. La
extracción de RNA total se realizó en cada punto de la cinética descrita en el punto 3.2.9
(Tabla 3) es decir a 2, 4 y 6 horas tratando las bacterias con cuatro condiciones distintas:
Oxitetraciclina (0,2 ug/mL y 0,8 ug/mL) y Florfenicol (0,2 ul/mL y 0,8 ug/mL), siendo el
tiempo 0h además utilizado como control para la expresión de estos genes en estado basal
sin presencia de antibiotico en el medio. Una vez extraído el RNA (como se describe en el
punto 3.2.11) se procedió a sintetisar el cDNA como se describe en el punto 3.2.12.
3.1.13.3 Ensayo qPCR
La evaluación de la expresión de los genes previamente identificados en los genomas de
las cepas de P. salmonis fue realizada utilizando como templado el cDNA sintetizado para
cada punto de la cinética previamente descrita. La reacción se realizó utilizando el kit de
tiempo real GoTaq qPCR Master Mix (Promega) de acuerdo a las indicaciones del
fabricante. La reaccion de qPCR fue realizada en el termociclador de tiempo real Stratagene
MX3000P y MX3005. El equipo se programó con los siguientes ciclos: denaturación inicial
a 95ºC por 10 min, 45 ciclos de 94ºC por 20 s, 60ºC por 15s y 72ºC por 20 s. Además se
agregó en cada corrida un paso para elaborar la curva de disociación, con 95ºC por 20s,
63ºC por 10s y 95ºC por 1s. La fluorescencia fue medida al final de cada uno de los 45
ciclos y durante la última rampla de aumento de temperatura para la elaboración de la curva
de disociación. Como gen normalizador se utilizaron el gen RpoD. Cada cuantificación se

48
realizó por triplicado. La línea base para el cálculo del ciclo umbral (ct) de cada muestra
fue calculada por el programa MxPRO (Stratagene) utilizando un algoritmo LMS (Least-
Mean-Square algorithm) y el mismo programa entregó los datos de ct. La cuantificación
relativa se hizo según la fórmula del "ΔΔct" (Pfaffl, 2001) y se graficaron los datos y se les
realizó la prueba estadística T de Student, usando el programa Sigmaplot 11.0,
seleccionando los p<0.01 como altamente significativo (**) y p<0.05 como significativo
(*).

49
4. Resultados
4.1. Análisis bioinformático de genomas de P. salmonis
4.1.1. Análisis comparativo in-silico entre genomas disponibles de cepas nacionales de
P. salmonis y la cepa tipo LF-89 con respecto a genes de resistencia a antibióticos
Recientemente se secuenciaron genomas de 9 cepas de Pisciricketssia salmonis aisladas en
el sur de Chile. Utilizando el software GenDB se realizó un análisis in-silico de estos 9
genomas con el fin de identificar posibles genes involucrados en la resistencia a los
antibióticos Oxitetraciclina y Florfenicol. Mediante este análisis se identificaron tres genes
putativos de resistencia a Oxitetraciclina. Dos de ellos presentes en todas las cepas, y un
tercer gen que se vio ausente tanto en la cepa tipo LF-89 (IBM-028) como en las cepas
pertenecientes a los genogrupos I y II (Tabla V). Para poder identificar estos genes
putativos y darle una nomenclatura, el software realizó un alineamiento por BLASTN de la
secuencia nucleotidica de estos tres genes hacia la base de datos NCBI dándoles una
notación, se indicó una nomenclatura putativa hacia los genes condificantes para Tet E, Tet
C y un gen que posiblemente codifica para una bomba transportadora de Tetraciclina y
Oxitetraciclina previamente descritos para otras especies.

50
Tabla V. Genes putativos de resistencia a Oxitetraciclina encontrados en 9 cepas
nacionales de P. salmonis. Se muestran los genes putativos posiblemente involucrados en
conferir resistencia a Oxitetraciclilana encontrados mediante el software GenDB 2.2 en las
9 cepas de aislados nacionales de P. salmonis. Presencia (+) o ausencia (-). Genogrupos
obtenidos del paper en preparación de Figueroa et al 2013.
Cepa Genogrupo tet E tet C Bomba tetraciclina
IBM-028 II + - +
IBM-004 I + - +
IBM-009 VIII + + +
IBM-012 II + + +
IBM-014 VI + + +
IBM-023 II + + +
IBM-029 ? + + +
IBM-030 II + + +
IBM-034 I + - +

51
Mediante este mismo análisis in silico se identificaron cinco genes putativos que
posiblemente estén implicados en la resistencia a Florfenicol. Estos cinco genes se
encontraron presentes en los 9 genomas (Tabla VI).
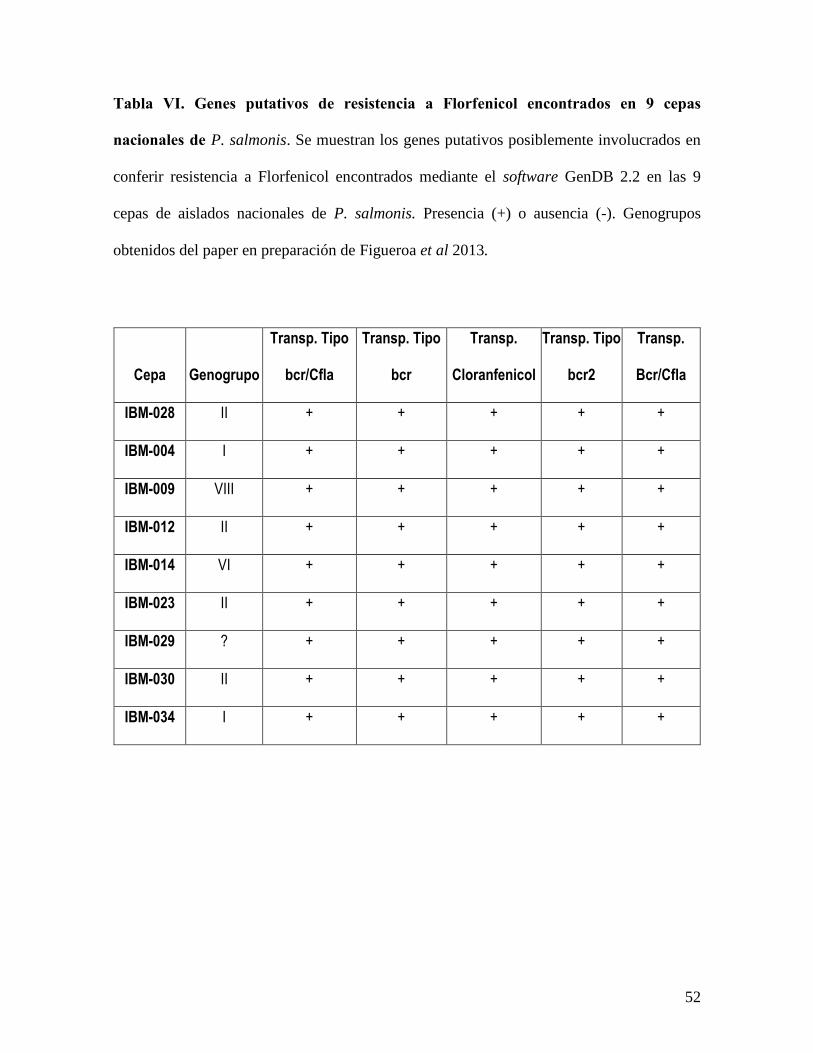
52
Tabla VI. Genes putativos de resistencia a Florfenicol encontrados en 9 cepas
nacionales de P. salmonis. Se muestran los genes putativos posiblemente involucrados en
conferir resistencia a Florfenicol encontrados mediante el software GenDB 2.2 en las 9
cepas de aislados nacionales de P. salmonis. Presencia (+) o ausencia (-). Genogrupos
obtenidos del paper en preparación de Figueroa et al 2013.
Cepa Genogrupo
Transp. Tipo
bcr/Cfla
Transp. Tipo
bcr
Transp.
Cloranfenicol
Transp. Tipo
bcr2
Transp.
Bcr/Cfla
IBM-028 II + + + + +
IBM-004 I + + + + +
IBM-009 VIII + + + + +
IBM-012 II + + + + +
IBM-014 VI + + + + +
IBM-023 II + + + + +
IBM-029 ? + + + + +
IBM-030 II + + + + +
IBM-034 I + + + + +

53
Se realizó el mismo procedimiento anteriormente utilizado, para poder identificar y darle
una nomenclatura a estos genes. Finalmente se encontraron cinco posibles genes que
podrían estar involucrados en conferir resistencia múltiple a drogas (Tabla VII), estos genes
se encuentran presentes en las 9 cepas, con la excepción de un putativo transportador de la
familia MFS el que se encuentra ausente en los genomas tanto de la cepa tipo LF-89 (IBM-
028) como en las cepas pertenecientes a los genogrupos I y II, además un putativo
transportador mdtD que solo se encuentra presente en estas tres cepas. Los genes
encontrados mediante este análisis in silico se resumen en la tabla VII

54
Tabla VII. Genes putativos de resistencia múltiple a drogas encontrados en 9 cepas
nacionales de P. salmonis. Se muestran los genes putativos posiblemente involucrados en
conferir resistencia múltiple a drogas encontrados mediante el software GenDB 2.2 en las 9
cepas de aislados nacionales de P. salmonis. Presencia (+) o ausencia (-). Genogrupos
obtenidos del paper en preparación de Figueroa et al 2013.
Cepa Genogrupo Transp. MFS Transp. MFS Transp. DHA2
Transp.
Drug/met.
Transp.
mdtD
IBM-028 II + - + + +
IBM-004 I + - + + +
IBM-009 VIII + + + + -
IBM-012 II + + + + -
IBM-014 VI + + + + -
IBM-023 II + + + + -
IBM-029 ? + + + + -
IBM-030 II + + + + -
IBM-034 I + - + + +

55
4.1.2. Comparación de secuencia de genes encontrados entre las cepas
De acuerdo a los genes putativos encontrados en cada cepa, se seleccionaron tres genes que
posiblemente otorguen resistencia a Oxitetraciclina y tres genes que posiblemente otorguen
resistencia a Florfenicol. Los genes seleccionados fueron los codificantes para Tet E, Tet C,
Bomba transportadora de tetraciclina, Transportador Bcr/CflA, Bcr 2 y transportador de
cloranfenicol. Utilizando el software GenDB 2.2 se realizó la identificación in-silico de las
secuencias codificantes de estos genes.
4.1.2.1. Análisis in-silico de la secuencia codificante para Tet E
La secuencia codificante del gen tet e obtenida con la herramienta GenDB 2.2 se indica en
la figura 3. La secuencia fue alineada utilizando el software BioEdit.

56
Tet E
1 ttg acc caa gca aag caa caa ctc atc gcc att tta tta atc acc 45
46 tta ctt ggc ttc atc ggt ttg gca att cct tat ccg att ctt gct 90
91 ccg atg ctg ctg aaa agt cag gct ttc tcc tta cat acg cat gcc 135
136 acg cgt atg ctt tgg tac ggt gca cta ctt gcc gcc tac cct atc 180
181 ggt cag ttt ttg ggc tct gct att tta ggt gat ctt tca gat cat 225
226 tgg ggg cga cgt aaa att tta att ctt tct tta cta cta aca acc 270
271 atc ggt tat gca ata agt gca att gcc gtt tat caa cat tat ctt 315
316 ggg ctt ttt ctt ttt gct cga tta ttc acc ggc ctt gca gaa ggc 360
361 aat att gcc att tgt cgt gct gcg gca gca gac ctt agc cca aag 405
406 att agt aaa tac acc agt ttt ggt tta atc aat gca gct ggc acc 450
451 tgt ggt ttt atc atc ggc ccc att ctc gct ggt gtt ttt tca gat 495
496 aaa acg att tac cta ggt ttt agc tat gct aca cca ttt ttc att 540
541 gcc tta tta ctc agc tta ttc acg tta atc gcc act tat ttt ttc 585
586 ttt act gaa acc ttt aat cca atc aaa aat aaa gct cat caa gca 630
631 ttt tct tta ttt agt tta ctg att act gcc atg aat aat atc att 675
676 ttc act gta cgg ctt cag cat att aaa tgg cta cta ttc agt gct 720
721 cta cta ctt atg ttc agt gta gat tta acc tat caa ttc gca ccg 765
766 ctg ttt ctt aca gcc cat tgg caa atg acc agt ctt gcg att gct 810
811 aaa aca tta ttt gta ctg tgg gtc agt att att att ggc gaa gtt 855
856 tgg tta gta cca cta ctt gct aaa cgc ttc aat aaa tat aaa ctt 900
901 att ttc aat agc acc att gct tta tca ata ttt ctt atc cta atg 945
946 atc aca act act tat tta gca cta tta tat gca gcc tta ttt ttg 990
991 ctt ggt ata ttc att gct tta tca acc aca ctt gca acc att ctt 1035
1036 att tcc gat gcc gtt aac cct cat att caa ggt cgt att tta ggg 1080
1081 gca ttt agc tca ttg cgt gtc ctt ggt gat gca atc ggt gca att 1125
1126 act ggt gat tta tgt gcc att ttt tat cca gca gcg cca tta tgg 1170
1171 cta gcg gta agt atg ctt att att tca tta ata tta tta ctt gtg 1215
1216 act aaa aat aaa cgt aaa cta taa 1239
Figura 3. Secuencia nucleotídica codificante para Tet E de P. salmonis. Se muestra la
secuencia nucleotídica del gen que codifica para Tet E. Alineada mediante el software
BioEdit.

57
Posteriormente la secuencia aminoacídica fue analizada utilizando la herramienta on-line
SMART indicando dos dominios conservados, correspondientes al dominio MFS_1 (Major
facilitator superfamily domain) además de un dominio transmembrana, los cuales se
indican en la figura 4. Además mediante la aplicación Conserved Domains and Protein
Clasification (NCBI) se pudieron observar ambos dominios, además de dominios putativos
correspondientes a poros translocadores de sustrato y cierto dominio característico en
proteínas transportadoras de drogas.

58
Figura 4. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
putativa Tet E de P. salmonis. A) Secuencia aminoacídica de la proteína Tet E, en colores
se destacan los dominios reconocidos con el análisis SMART. B) Esquema SMART de la
proteína Tet E, los colores corresponden a los dominios identificados e indicados en la
secuencia aminoacídica. Verde: Dominio MFS 1 (Major facilitator superfamily domain).
Azul: Dominio trasmembrana.

59
4.1.2.2 Análisis in-silico de la secuencia codificante para Tet C
La secuencia codificante del gen tet c obtenida con la herramienta GenDB 2.2 se indica en
la figura 5. La secuencia nucleotídica fue alineada utilizando el software BioEdit.

60
Tet C
1 atg tta aat aaa aat tca ttg ttt tct atg ctg cca tta ttt ctc 45
46 gtt ttg ttt att gat ggt atg ggc ttg gga cta tta ttc cct att 90
91 tta aac ggt gtg att gtt gat ccg aca tca gac ttt tta ttc cat 135
136 acc gta tcg aca ggg acg aga gaa atc att tat ggt ctt att gtc 180
181 ggt atc ttt atg atc tgc tgg ttt ttt ggt gcg gca att cta ggt 225
226 gat tta tct gat acc att ggt cgt aag aaa gca ctt atg att tgt 270
271 tta att gga tca ttt ata ggc tat tta ttg tca gcg att tca att 315
316 atg atg cat agc ttg act ttg ctt att tta ggt cga gtg atc gct 360
361 ggt ttt act gca gga agt caa cct gtt gca caa gcg gct att gtt 405
406 gat gtg agc tct gaa gag cac aaa aca cga aat att ggc ttt att 450
451 tta tta tct gtt tca ttg gga ttt gtg tgt ggg cct atc att ggt 495
496 ggt gtg cta tcg gat agt caa atc tat agt aat ttt aat tat gat 540
541 act cca cta tat ttt gct tct atc atc tca tta att aat gca att 585
586 ttg ctg ggc gtt ctt ttt aaa gaa act ttt tat aaa aca gaa aaa 630
631 gtt aaa ata aaa tta cat cat gcc ata aat ata ttt att tca gct 675
676 ttt ggt cac aag aaa att tgt aag cta gcg att gtc atg ctg gtg 720
721 atg att ttc ggc tgg agt aat tat ttt tct ttt att tct gtt tat 765
766 atg aca aaa aat tat cac ttt aat acc ttt gaa aac tcg atg ttc 810
811 tta gcg gtt atg ggt gtg ggc ttt agc atc ggt tgt ggc tat gtc 855
856 gtt gag tgg tgt gct aag cgc tat tta ctg agt cat acg gtg cgt 900
901 att atg ttg ctg ctc act ggt att tca gta ttg ctg aca ttg ctc 945
946 att cat aat gtc tat acg tgt tat tta ttt gct cta tta atc ggt 990
991 tta acg tta gcg gtg gct tat tcg agt att att gcg ctt ttt tca 1035
1036 aat caa gtg gga gca gat gag caa ggt tgg gtg atg ggt gtc acc 1080
1081 ggc tct att atg gca tta tgc ttt ggt tta aca tct ttt ggc aca 1125
1126 ggg gtg att gca agt ttt gct ggt gct agt att cca att gtg ttt 1170
1171 gcg gtg att ggc tta tta gca agt ggg ggg ctg ttg tat tta ttt 1215
1216 cgc ttt gaa gaa act aaa taa 1236
Figura 5. Secuencia nucleotídica codificante para Tet C de P. salmonis. Se muestra la
secuencia nucleotídica del gen alineada mediante el software BioEdit.

61
Posteriormente la secuencia aminoacídica fue analizada utilizando la herramienta on-line
SMART indicando dos dominios conservados, correspondientes al dominio MFS_1 (Major
facilitator superfamily domain) además de un dominio transmembrana, los cuales se
indican en la figura 6. Además mediante la aplicación Conserved Domains and Protein
Clasification (NCBI) se pudieron observar ambos dominios además, de dominios putativos
correspondientes a poros translocadores de sustrato.

62
Figura 6. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
putativa Tet C de P. salmonis. A) Secuencia aminoacídica de la proteína Tet C, en colores
se destacan los dominios reconocidos con el análisis SMART. B) Esquema SMART de la
proteína Tet C, los colores corresponden a los dominios identificados e indicados en la
secuencia aminoacídica. Verde: Dominio MFS 1 (Major facilitator superfamily domain).
Azul: Dominio trasmembrana.

63
4.1.2.3 Análisis in-silico para secuencia codificante para bomba transportadora de
tetraciclina
La secuencia codificante del gen obtenida con la herramienta GenDB 2.2 se indica en la
figura 7.

64
Bomba transportadora de tetraciclina
1 atg ctt ata aaa tcc ata gaa cac ttt gtt att aaa gac agt tta 45
46 cta aaa tta ttc act tta tat tct gtt att ttt cta ggc tct ctt 90
91 ggc tat ttt atc gtg atg ccg gtg tat gca gag cta atc ctt agc 135
136 cct ata caa ggc tta ctg cct gag cac gtt gat ata aca acc cgg 180
181 cgt att ata ttt agc cta gtc agc gcc ttt gcc ccc cta tta act 225
226 ctt tgt att gct ccg ttt att ggc cat tta tct gac caa ctt ggc 270
271 cgc aaa aaa gta ctc ttg ttt tgt ctt ggc tta aca ctc gtc agc 315
316 ttt cta ctc cct att acc gcc att gcc atc agc agc atg act ctc 360
361 ttg ttt att agc aat gct atc aat agc att gca tct gcc agt caa 405
406 ccc atc gcc cag gct gct tta acg gat atc agc aca ggg cgc atg 450
451 aaa gct tac tta atg gga cta tcc gcc tca agc atg aca gca gca 495
496 atg tca ttt ggt ccg gca ctc ggc agc tat tta atc aag gtc agc 540
541 ttt gat gct aca ttt tgg tgt agc gca ctt ttc aca gta att aat 585
586 att att ttg gtc tat tta act ttt tca cca gaa aaa aaa cga ata 630
631 aaa gct gca aaa aca cct agc ttg tta tct tca ttc aag gct tac 675
676 tct ccc ctc gca gtg acc atg ctt ata ctg gtc ttt tta tgc caa 720
721 ctg gcg tgg agc agc tac tat caa aac att aac tac aca gtc atc 765
766 agt atc tgg cac cta cca atc agc tct aac ctc tac caa caa cta 810
811 atg ctg atc att ggc tta agt atg att tta tct cta ctt att ctg 855
856 tac cca gct att att cga att atc acg ata ata caa tgc ttt aag 900
901 ctt tgc ctt atc att gca act tta tcg atg cta gta ata agc cta 945
946 acg cat cag atc tgg ctc tat aaa ata att ctt att cct tta tct 990
991 agc agc att gcc atg act tac cct tgc tat atg tct att tta tcg 1035
1036 aat gct cta act aag cgc cat caa ggc tca ggc att gca ctg gct 1080
1081 gga ggg ctt atg ggc atc agc tgg aca ctc agt ggc tct ttc agt 1125
1126 ggg ttg atc gcc aac atc aac ctt cac tta gcc aac tgg cta gca 1170
1171 gtc att atc cta ata ttg gca ctt ttt tta cta ttt atc aca atc 1215
1216 aaa aaa acg gtc caa aaa agc aaa tgt ctt tac att taa 1254
Figura 7. Secuencia nucleotídica codificante para Bomba transportadora de
tetraciclina de P. salmonis. Se muestra la secuencia nucleotídica del gen, alineada
mediante la mediante el software BioEdit.

65
La secuencia nucleotídica fue alineada mediante el software BioEdit. Posteriormente la
secuencia aminoacídica fue analizada utilizando la herramienta on-line SMART indicando
dos dominios conservados, correspondientes al dominio MFS_1 (Major facilitator
superfamily domain) además de un dominio transmembrana, los cuales se indican en la
figura 8. Además mediante la aplicación Conserved Domains and Protein Clasification
(NCBI) se pudieron observar ambos dominios además de dominios putativos
correspondientes a poros translocadores de sustrato y cierto dominio característico en
proteínas transportadoras de drogas.

66
Figura 8. Análisis in-silico de los dominios de la secuencia aminoacídica de la proteína
putativa para Bomba transportadora de Tetraciclina de P. salmonis. A) Secuencia
aminoacídica de la proteína, en colores se destacan los dominios reconocidos con el análisis
SMART. B) Esquema SMART de la proteína, los colores corresponden a los dominios
identificados e indicados en la secuencia aminoacídica. Verde: Dominio MFS 1 (Major
facilitator superfamily domain). Azul: Dominio trasmembrana.

67
4.1.2.4 Análisis in-silico para secuencia codificante para bomba del tipo Bcr A
(Bcr/Clfa)
La secuencia codificante del gen obtenida con la herramienta GenDB 2.2 se indican en la
figura 9. La secuencia nucleotídica fue alineada mediante el software BioEdit.

68
Bcr/Cfla
1 atg gtt gca gcc aat gcg att gtt cgc gat atc tat gaa ggt gag 45
46 cgt ggt gca cag gtg atg agc tat att tca agt gcg gtg gct tta 90
91 gcc ccg gtt ttt gca cct ttt att ggt agt tat tta tcc tgg cag 135
136 atg acc ttt tgg gtg ctg gct ggc tgt ggt gtc tta ttg tgc ttg 180
181 caa atg ttg ctg ggt ttg caa gag act tgt gtt agg cgt cag gtg 225
226 aac ttg ctt aac ttt aaa agc tgt tta gct gtg tat cgt cag tta 270
271 att ttt cat cga caa ttt ttt ctc cat agt ata att agt cta tgc 315
316 agc ttt agt att tta ttt tta ttt tta ttt ttt tct ttt tcg cct 360
361 tat tta ttt att gat cag tta ggc tta gct aag acc gag ttt ggt 405
406 tat tgg ttt ggt tta aat gcc ctg gtg ttg atg gtt atg agc ttt 450
451 att ggt ggc ttt ggt gtg aaa tat gtc agc att cgc ttt tgg atg 495
496 ggt tta gga gtg ttg tgt atg tta gtt ggc ggt ttt acg atg atg 540
541 att tgg cta tat tgc agc acg agt tta agt ttg acc gcg att atg 585
586 ctg cca atg ttt atg att agt gca ggc atc gcc tgg acg gta tct 630
631 cct gct gtt gta ggt gca atg atg cct ttt ggt cat att tct ggt 675
676 tct gcc gcg gct ctt tta ggc tgt att cgc ttt gca ggc tct gcg 720
721 ctg ttg gtc acc ttg atg atg act tta ggg ccc acc agt gct gtg 765
766 tat ttt gct agt tgt atc tgc agc ttg gct ttg att gca ttg ttt 810
811 ggt tgt att ttc att att aaa agc agg cag act gtt gta tat tga 855
Figura 9. Secuencia nucleotídica codificante para Bomba transportadora del tipo Bcr
A (Bcr/Cfla) de P. salmonis. Se muestra la secuencia nucleotídica del gen, alineada
mediante la mediante el software BioEdit.

69
Posteriormente la secuencia aminoacídica fue analizada utilizando la herramienta on-line
SMART indicando dos dominios conservados, correspondientes al dominio MFS_1 (Major
facilitator superfamily domain) además de un dominio transmembrana, los cuales se
indican en la figura 10. Además mediante la aplicación Conserved Domains and Protein
Clasification (NCBI) se pudieron observar ambos dominios además de dominios putativos
correspondientes a dominios caracteristicos de proteínas transportadoras de drogas y un
dominio de la sub familia Bcr/CflA (transportadora de drogas).
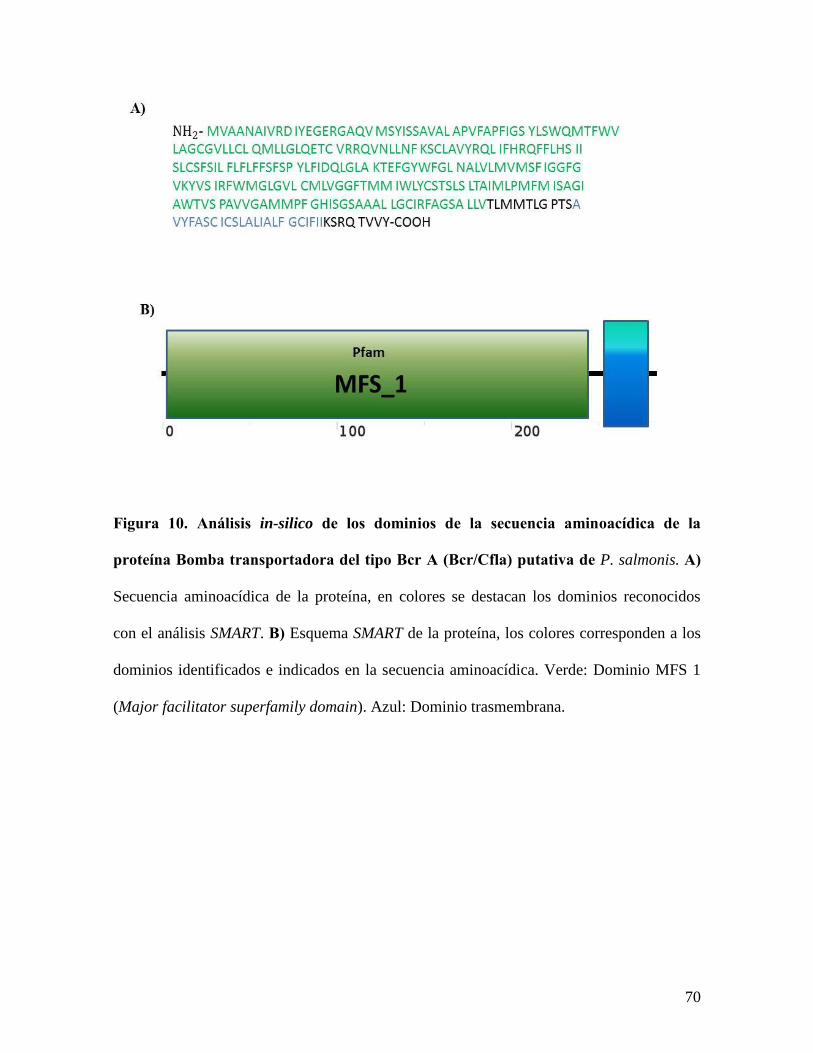
70
Figura 10. Análisis in-silico de los dominios de la secuencia aminoacídica de la
proteína Bomba transportadora del tipo Bcr A (Bcr/Cfla) putativa de P. salmonis. A)
Secuencia aminoacídica de la proteína, en colores se destacan los dominios reconocidos
con el análisis SMART. B) Esquema SMART de la proteína, los colores corresponden a los
dominios identificados e indicados en la secuencia aminoacídica. Verde: Dominio MFS 1
(Major facilitator superfamily domain). Azul: Dominio trasmembrana.

71
4.1.2.5 Análisis in-silico para secuencia codificante para bomba del tipo Bcr 2
(Bcr/Cfla)
La secuencia codificante del gen obtenida con la herramienta GenDB 2.2 se indica en la
figura 11. La secuencia nucleotídica fue alineada mediante el software BioEdit.

72
Bcr 2
1 atg act gtg acc aca cca tgc aac cga gca aaa ctc aca ctc ttt 45
46 gtc tca gtc att gcc gtt atc gcc gca gca ata gaa acc gac tta 90
91 ttt att cca agc tta ccg gcg atg caa cac tat tat ttg gca tct 135
136 tcc aac caa att cag ctc ctg atc agc atg aat ttt tta ggg ctt 180
181 tgc atc tca agc cta ttt tat ggt cca ctc gca gat agc tat ggc 225
226 cgc aag cct atc tta ctg acc ggc ctt gtt atc ttt gtt ttt agc 270
271 gct gtt ggc tgt gta ttt tca ctt tca tta agc aat att atc ttc 315
316 tgg cgt ttt att gaa ggc atc ggc agc ggt gcc tgc ttt gtc gtc 360
361 cct gga gca gtt atc ttt gac ttg tat agt aaa gaa gca gct gca 405
406 cgt att tta ggc acc ctc aac agc ata act aca att gct atg gca 450
451 gcc tca cct tta gct ggt ggt ttt att cac act caa ttt ggc tgg 495
496 cgt gct aat ttt tta ttt tta gct ttt atc gcc ata ctc gcc ttg 540
541 ctt ctt gct att ttt ctc att aaa gag tcg ctc cct aaa aaa aac 585
586 aga act ccc ctg cac atc aaa tcg att acg cat ggc tat tat cga 630
631 ctg ata acc cat act gaa tca ctt agc tac tta ctg att atc tgc 675
676 tgt gta ttt ggc gct tac atg gtc tat att tcg gga ctg tct tta 720
721 att ttt gtc aat cat ttg cac att tca gag tca gcc ttc cct tac 765
766 tat caa ggt gcg gtc tta ctt gct ttt gct gca atc agc atg gca 810
811 tgt gga cgt atc att cat ttc tta ggt atg aaa aag gcc tta cat 855
856 tac agc act att acc atg gct att ggc gct att tta tta ttg ttc 900
901 acc ggg tta ttt ttc gct aac tca gcc ttg ctc atc acc att acc 945
946 atg tcg att ttt gcc gca ggt gtc gct tta atg cta acc att atc 990
991 ttt ggt aac tat atg gaa gtg att cca gaa att aaa ggt atc gcc 1035
1036 tcc gcc tta tgt aat gcc cta cgc ctc tgt gct gca gca tta ttc 1080
1081 gtt ggt ctt gcc ggt ttg ctc ttt acc gga acc att acg cct att 1125
1126 gcc att ttt atc ttt act ctg gca atc atc agc aca gta gtt tta 1170
1171 gtc tgg ctg caa aga aaa aaa tta cac caa caa ccc aaa cac cca 1215
1216 tct cac tct cat caa agt agt cat tca acc acc tga 1251
Figura 11. Secuencia nucleotídica codificante para Bomba transportadora del tipo Bcr
2 (Bcr/Cfla) de P. salmonis. Se muestra la secuencia nucleotídica del gen, alineada
mediante la mediante la herramienta bioinformática ExPasy y el software BioEdit.

73
Posteriormente la secuencia aminoacídica fue analizada utilizando la herramienta on-line
SMART un dominio conservado el que corresponde al dominio MFS_1 (Major facilitator
superfamily domain), se indica en la figura 12. Además mediante la aplicación Conserved
Domains and Protein Clasification (NCBI) se pudo observar este dominio, además de
dominios putativos correspondientes a poros translocadores de sustrato, un dominio
putativo específico de proteínas de resistencia a drogas y un dominio putativo de la
subfamilia de transportadores Bcr/Cfla.

74
Figura 12. Análisis in-silico de los dominios de la secuencia aminoacídica de la
proteína Bomba transportadora del tipo Bcr 2 (Bcr/Cfla) putativa de P. salmonis. A)
Secuencia aminoacídica de la proteína, en colores se destacan los dominios reconocidos
con el análisis SMART. B) Esquema SMART de la proteína, los colores corresponden a los
dominios identificados e indicados en la secuencia aminoacídica. Verde: Dominio MFS 1
(Major facilitator superfamily domain).

75
4.1.2.6 Análisis in-silico para secuencia codificante para bomba transportadora de
cloranfenicol/florfenicol
La secuencia codificante del gen obtenida con la herramienta GenDB 2.2 se indica en la
figura 13. La secuencia nucleotídica fue alineada mediante el software BioEdit.

76
Transportador cloranfenicol/florfenicol
1 atg ttc ttt att ata atc ggc ctt ata gtt tct gga gtg tac ttt 45
46 aat aaa ata ata ata ttg aga tta ctt atg aat cga ctt ttt att 90
91 gcg ctt ttt gtg aat ctt gtt att cct ttg ggg gga atg agt aca 135
136 gac att tat tta cca tca ctg cct gcg acg gca gac tat ttt ggc 180
181 tca tca gat aca ctg gta caa att acc gtc acg ctg ttc act tta 225
226 ggg tta gga gtg ggt cag ata gtc gct ggg ccg atc tca gat gct 270
271 tta ggc cgt aaa aat ccc ttt tta ttt ggg atg tta tta cag ttg 315
316 acc tct gtt ttg ttg atc tta aat acc agc tca ttg aca aca ttg 360
361 att gtt gta cgg ctt ttt caa ggt ttt ggc gct gcg ttt atg atg 405
406 gtg cct gtt cgc gct atg tta agc gat gtg ttt gat ggt gca gcg 450
451 tta aaa aaa caa tat acg tat att acc gca tct ttt gct ctt ggg 495
496 ccc atc gtt gcc cct ttt att ggt ggc tat ttg cag cat tat att 540
541 ggc tgg caa gct aac ttt tac ttt gtt ctt gtt tat ttg gct ttt 585
586 ttg aca aca gtt gct tta gtc gtt tta aaa gaa acc agt acg act 630
631 aaa aaa gca ttt tcg ctt gtt cat gtg agt tca agt tat atc acc 675
676 gtt tta aaa aat aaa tta ttt tta acg gga act att tta att agt 720
721 ttt ttt ctt ggg tat gtc gct ata ttt aat gtt gtc ggt ccg ttt 765
766 gtt att cag gat ata ttg gag cgg tcg gct att ttt tat ggt tat 810
811 att gct tta tta atg ggt ttt gct tgg ttt tct ggt aat atg acc 855
856 agt cgg tta ttt ttc aaa ttt tct cga cag tct aaa tca aat aca 900
901 gcg ctt tta tgt atg ctg att gct gct gtt gct atg ttt agt att 945
946 agt ttt agt act atc agt tta ctc aag ttt att gcg cct ata ttt 990
991 att atg att ttt tgt ggg gga ttt tta ttt cct att tat gtt ggg 1035
1036 gaa tgt tta tct tta ttt aaa gaa caa gcg gca tca gcc aat ggt 1080
1081 cta tta ttt tcc tcg att tgg att gtg ttt agt tta ttt tca ttt 1125
1126 att gct gct ttt ttg aaa gtt tat tct ctg ctg ccg att gct agt 1170
1171 tgt tat tta att ttg acc ttg atg att tgc tgt tgg ttt ttg ttc 1215
1216 gtt gca aga aag cat tga 1233
Figura 13. Secuencia nucleotídica codificante para Transportador de
Clorfanfenicol/Florfenicol de P. salmonis. Se muestra la secuencia nucleotídica del gen,
alineada mediante la mediante el software BioEdit.

77
Posteriormente la secuencia aminoacídica fue analizada utilizando la herramienta on-line
SMART indicando dos dominios conservados, correspondientes al dominio MFS_1 (Major
facilitator superfamily domain) y dos dominios transmembrana los que se indican en la
figura 14. Además mediante la aplicación Conserved Domains and Protein Clasification
(NCBI) se pudieron observar ambos dominios además de dominios putativos
correspondientes a poros translocadores de sustrato, un dominio putativo específico de
proteínas de resistencia a drogas y un dominio putativo de la subfamilia de transportadores
Bcr/Cfla.

78
Figura 14. Análisis in-silico de los dominios de la secuencia aminoacídica de la
proteína transportadora de Cloranfenicol/Florfenicol putativa de P. salmonis. A)
Secuencia aminoacídica de la proteína, en colores se destacan los dominios reconocidos
con el análisis SMART. B) Esquema SMART de la proteína, los colores corresponden a los
dominios identificados e indicados en la secuencia aminoacídica. Verde: Dominio MFS 1
(Major facilitator superfamily domain). Azul: Dominio trasmembrana.

79
4.1.3 Homología de secuencias analizadas con secuencias de otras especies bacterianas
Los porcentajes de identidad de las secuencias aminoacídicas obtenidas como se explicó
previamente, fueron comparadas con la información disponible en la base de datos de
NCBI mediante la herramienta bioinformática BLAST, esto se realizó con el fin de
encontrar las secuencias con mayor homología aminoacídica de otras especies bacterianas
presentes en esta base de datos y determinar de esta manera si estas proteínas se encuentran
conservadas en otra especie o si se ven altas homologías en bacterias filogenéticamente
relacionadas. Los datos obteniedos con los mayores porcentajes de homología son
indicados en tabla VIII.

80
Tabla VIII. Análisis in-silico de identidad aminoacídica de las proteínas putativas
para resistencia a Oxitetraciclina y Florfenicol con otras especies bacterianas. Se
muestran los mayores porcentajes de homología con respecto a secuencias de NCBI.
Prot. Putativa BLASTP Microorganismo
Id. Sec. Aac. Query Coverage
Tet E Drug. Transp. Legionella drancourtii 36% 91%
Tet. Efflux transporter Algoriphagus sp. 33% 92%
Drug. Transp. MFS Legionella Longbeachae 35% 99%
Tet C TET(E) E. coli 50% 95%
Drug. Transp. Coxiella Burnetti 40% 100%
Drug. Transp. Coxiella Burnetti 40% 100%
B. Tetraciclina TET(E) E. coli 67% 58%
Transp. MFS
Photobacterium damselae 36% 96%
TET(E) Fibrisoma limi 25% 88%
Bcr/Cfla florR E. coli 30% 87%
Clor. resist. Protein Vibrio anguillarum 33% 87%
Transp. sflR Legionella pneumophila 31% 87%
Exp. Flor. Vibrio nigripulchritudo 31% 90%
Bcr2/Cfla Flor effux pump Legionella pneumophila 31% 87%
Flor effux pump Legionella longbeachae 30% 87%
Transp. Multidrogas Aeromonas veronii 35% 90%
Transp. Drogas Bcr Ferrimonas balearica 37% 90%
Transp. Clr/Flor Transp. Multidrogas Coxiella burnetti 46% 90%
Transp. Drogas Bcr Coxiella burnetti 39% 88%
FlorR E. coli 25% 84%

81
4.1.4 Caracterización del entorno génico de genes asociados a resistencia a antibióticos
en cepas LF-89, IBM-009, IBM-034
Ya encontrados los genes putativos para resistencia a los antimicrobianos Oxitetraciclina y
Florfenicol presentes en los genomas analizados de P. salmonis, se procedió a realizar una
caracterización del entorno génico en el que se encuentran cada uno de estos genes, con el
fin de determinar si hay genes determinantes que permitan dilucidar el funcionamiento de
estos o bien si están contenidos junto a otros genes de resistencia o virulencia. Este análisis
se realizó utilizando el software GenDB 2.2.
Analizando el entorno génico del gen putativo tet e, presente en todas las cepas estudiadas,
se observó la presencia de dos transportadores MFS (Major Facilitator Family) ubicados
muy cerca de este en el genoma, estos son genes que podrían estar involucrados en
resistencia a drogas. Además según análisis bioinformaticos a islas de patogenicidad
presentes en P. salmonis (Lagos, 2013) se observó que este gen esta presente en la “Isla de
patogenicidad 2 IBM-034”, la cual se encuentra presente en todas las cepas analizadas de P.
salmonis.
En el entorno génico del gen putativo tet c ausente en la cepa LF-89 e IBM-034, se observó
la presencia de una transposasa cercana a este gen, además de ciertos genes pertenecientes
al sistema de secreción Dot/Icm.
Tras analizar el entorno génico del gen putativo codificante para una bomba transportadora
de Tetraciclina, el cual se encuentra presente en todas las cepas, se observó la presencia de
un gen codificante para una proteína putativa transportadora de la familia MFS, ademas de
un gen codificante para un transportador ABC y una ligasa A.

82
Se observó que ubicados en el genoma muy cerca del gen codificante para el transportador
Bcr A (Bcr/Clfa) se encontraban transportadores MFS y otro transportador putativo de
drogas, ademas de una transposasa y otro tipos de transportadores.
Se observó que ubicados en el genoma muy cerca del gen codificante para el transportador
Bcr A (Bcr/Clfa) se encontraban transportadores MFS y otro transportador putativo de
drogas, ademas de una transposasa y otro tipos de transportadores.
En el entorno génico del gen Bcr2 (Bcr/Cfla) se observaron integrasas y otras bombas
transportadoras, no para anitbióticos, pero si de metales pesados.
Finalmente en el entorno génico del gen codificante para el Transportador de Florfenicol, se
observó otro gen codificante para una proteína de resistencia multiple a drogas ubicadas
muy cerca de este gen (ychE), y una transposasa.
Mediante el software MAUVE (Multiple Genome Alignment) se analizaron los los entornos
génicos de estos genes, para todos los genes se pudo observar que en las tres cepas se
encontraba el mismo bloque de homología (el gen se encuentra en el mismo entorno
génico), esto no se observó sin embargo para el gen condificante para Tet C, donde solo se
observó en la cepa IBM-009 ratificando lo observado previamente. En la figura 15 se
esquematizan los dos casos obtenidos, en el cual se puede observar el mismo bloque de
homología en las tres cepas y en el segundo caso solo el gen en una cepa y sin bloque de
homología.

83
Figura 15. Analisis del entorno génico para los genes codificante para Bcr/Cfla y Tet
C en P. salmonis A) Bcr/Cfla: Se observa el mismo bloque de homología (Naranjo) para
las tres cepas estudiadas. B) Tet C: Se observa que Tet C se encuentra solo en IBM-009 y
que no hay bloque de homología cercano a este gen con las otras cepas. Anlizado mediante
software MAUVE.

84
4.2 Determinación de las CIM de Oxitetraciclina y Florfenicol para cepas de P.
salmonis LF-89, IBM-009 e IBM-034
4.2.1 Identificación de P. salmonis
Previo a trabajar con la bacteria, se realizó una confirmación de la identidad de E. coli y de
P. salmonis. Para la identificación de E. coli se realizó una tinción por Gram, donde el
resultado indicó que se trataba de bacilos gram negativos. Para la identificación de P.
salmonis se realizó una tinción de Gram, donde el resultado indicó que se trataba de cocos
gram negativos, morfología característica de la bacteria. Ademas para confirmar la
identidad de P. salmonis, se realizaron extracciones de DNA genómico para posteriormente
realizar el PCR confirmatorio para P. salmonis tal como se especifica en el punto 3.2.6.2 de
materiales y métodos. Los resultados obtenidos para las tinciones de Gram y para el PCR
confirmatorio se muestran en la figura 16 (A y B).
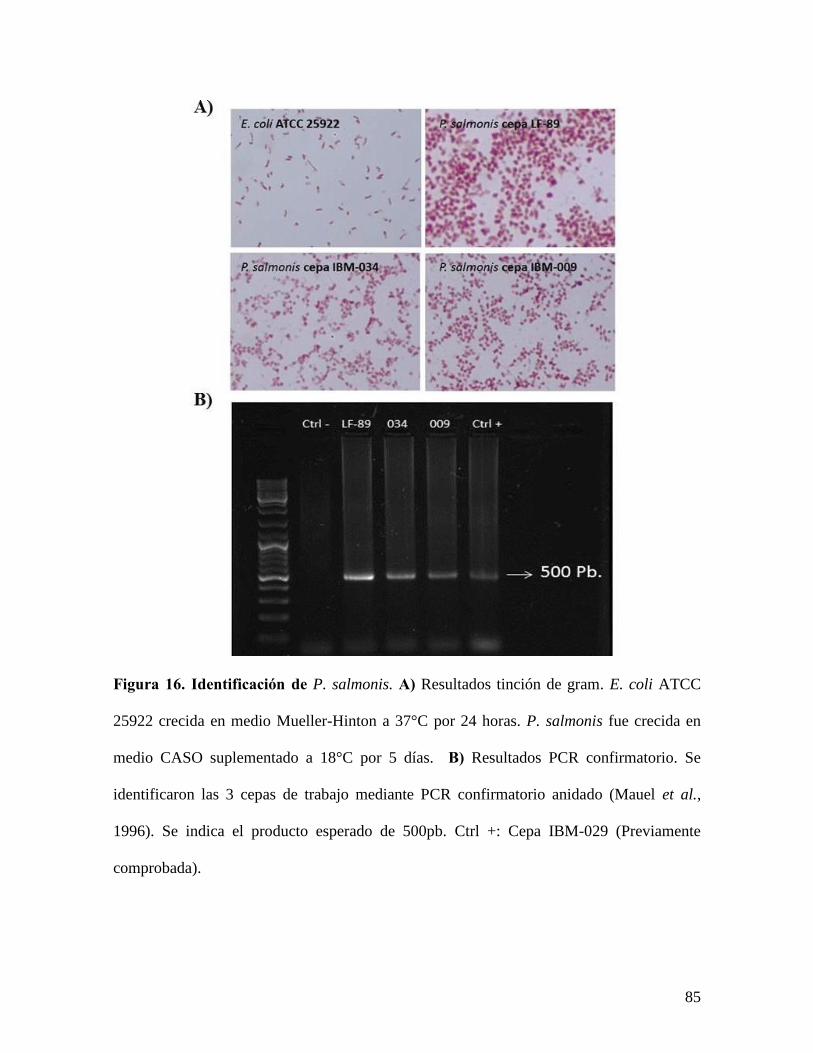
85
Figura 16. Identificación de P. salmonis. A) Resultados tinción de gram. E. coli ATCC
25922 crecida en medio Mueller-Hinton a 37°C por 24 horas. P. salmonis fue crecida en
medio CASO suplementado a 18°C por 5 días. B) Resultados PCR confirmatorio. Se
identificaron las 3 cepas de trabajo mediante PCR confirmatorio anidado (Mauel et al.,
1996). Se indica el producto esperado de 500pb. Ctrl +: Cepa IBM-029 (Previamente
comprobada).

86
4.2.2 Determinación de la CIM para Oxitetraciclina en E. coli ATCC 25922
Con el fin de estandarizar esta técnica y poder hacerla reproducible a otras cepas acuáticas
se realizó la prueba de sensibilidad a antibióticos por micro diluciones en placa para la cepa
E. coli ATCC 25922 desafiandola tanto a Oxitetraciclina. Tras crecer la cepa en medio
líquido se confirmo la identidad de la bacteria. Confirmada la identidad de la bacteria estas
fueron cultivadas en placas de ELISA de 96 pocillos con cantidades de medio, bacteria y
antibiótico como se describió en la tabla I y en el orden como se describió en la tabla II en
materiales y métodos. Estas placas fueron incubadas a 35°c por 19 horas aprox. Y
posteriormente se leyó su absorbancia a 600nm en el lector de placas. Se realizaron 8
replicas de muestra por placa, esto fue realizado por triplicado. El rango de CIM obtenido
fue de 1-4µg/mL, concordando con los rangos aceptados en la literatura.
4.2.3 Determinación de la CIM para Oxitetraciclina y Florfenicol en cepa tipo LF-89,
IBM-034 e IBM-009 de P. salmonis
Una vez estandarizada la técnica con E. coli ATCC 25922, se procedió a determinar la CIM
en la cepa tipo de P. salmonis LF-89, en la cepa IBM-034 y en la cepa IBM-009 frente a
Oxitetraciclina. Se creció la cepa en medio líquido y posteriormente se confirmó la
identidad de la bacteria. La técnica se realizó como se describe en la tabla I y en el orden
que se presenta en la tabla II. Las placas se incubaron por 5 días a 18°C para luego leer
absorbancias a 600nm en el lector de placas. Se realizaron 8 replicas de muestra por placa,
esto fue realizado por triplicado. Los resultados frente a Oxitetraciclina obtenidos para la
estandarización de la técnica con E. coli ATCC 25922 y para las cepas de P. salmonis LF-
89, IBM-034 e IBM-009 se grafican a continuación en la figura 17.

87
Figura 17. CIM de Oxitetraciclina para E. coli cepa ATCC 25922 y P. salmonis cepa
LF-89, IBM-034 e IBM-009. E. coli ATCC 25922 fue cultivada en medio líquido Mueller-
Hinton en placas de ELISA de 96 pocillos con distintas concentraciones de antibiótico,
incubadas durante 19 horas. Se leyó su absorbancia a 600nm obteniéndose un rango de
CIM de 1-4µg/mL. Las cepas de P. salmonis fueron cultivadas en medio líquido CASO
suplementado, en placas de ELISA de 96 pocillos con distintas concentraciones de
antibiótico, incubadas durante 5 días. Se leyó su absorbancia a 600nm. Las flechas azul y
roja en el grafico de LF-89 muestran la CIM para IBM-034 e IBM-009 respectivamente.

88
4.2.4 Determinación de la CIM para Florfenicol en E. coli ATCC 25922
Para poder estandarizar esta técnica, en este caso para Florfenicol y así hacerla
reproducible a otras cepas acuáticas se realizó la prueba de sensibilidad a antibióticos por
micro diluciones en placa para la cepa E. coli ATCC 25922 desafiandola esta vez
Florfenicol. La cepa se creció en medio líquido y se confirmo su identidad. Posteriormente
estas fueron cultivadas en placas de ELISA de 96 pocillos con cantidades de medio,
bacteria y antibiótico como se indica en en materiales y métodos tabla I y en el orden como
se describió en la tabla II. Las placas fueron incubadas a 35°c por 19 horas aprox. Y
posteriormente se leyó su absorbancia a 600nm en el lector de placas. Se realizaron 8
replicas de muestra por placa, esto fue realizado por triplicado. El rango de CIM obtenido
fue de 2-8µg/mL, concordando con los rangos aceptados en la literatura.
4.2.5 Determinación de la CIM para Florfenicol en cepa tipo LF-89, IBM-034 e IBM-
009 de P. salmonis
Una vez estandarizada la técnica con E. coli ATCC 25922, se determinó la CIM en la cepa
tipo de P. salmonis LF-89, en la cepa IBM-034 y en la cepa IBM-009 frente a Florfenicol.
Se creció la cepa en medio líquido y posteriormente se confirmó la identidad de la bacteria.
La técnica se llevo a cabo como se muestra en la tabla I, manteniendo el orden descrito en
la tabla II. Las placas se incubaron por 5 días a 18°C para luego leer absorbancias a 600nm
en el lector de placas. Se realizaron 8 replicas de muestra por placa, esto fue realizado por
triplicado.Los resultados frente a Florfenicol obtenidos para la estandarización de la técnica
con E. coli ATCC 25922 y para las cepas de P. salmonis LF-89, IBM-034 e IBM-009 se
grafican a continuación en la figura 18.

89
Figura 18. CIM de Florfenicol para E. coli cepa ATCC 25922 y P. salmonis cepa LF-
89, IBM-034 e IBM-009. E. coli ATCC 25922 se cultivó en medio líquido Mueller-Hinton
en placas de ELISA de 96 pocillos con distintas concentraciones de Florfenicol, incubadas
durante 19 horas. Se leyó su absorbancia a 600nm obteniéndose un rango de CIM de 2-
8µg/mL. Las cepas de P. salmonis fueron cultivadas en medio líquido CASO
suplementado, en placas de ELISA de 96 pocillos con distintas concentraciones de
Florfenicol, incubadas durante 5 días. Se leyó su absorbancia a 600nm. Las flechas azul y
roja en el grafico de LF-89 muestran la CIM para IBM-034 e IBM-009 respectivamente.

90
Los resultados obtenidos para los rangos de concentraciones inhibitorias mínimas en las
cepas LF-89, IBM-034 e IBM-009 de P. salmonis para los antibióticos estudiados
Oxitetraciclina y Florfenicol determinados mediante el método de micro diluciones en
placas se resumen en la tabla IX.

91
Tabla IX. Resumen de los resultados de CIM para Oxitetraciclina y Florfenicol en las
cepas estudiadas de P. salmonis. Se muestran los resultados obtenidos de CIM
previamente graficados para Oxitetraciclina y Florfenicol en las tres cepas analisadas.
Cepa Rango CIM (µg/mL) Antibiótico
LF-89 0,08-0,25 Oxitetraciclina
IBM-034 0,08-0,25 Oxitetraciclina
IBM-009 0,5-2 Oxitetraciclina
LF-89 0,08-0,25 Florfenicol
IBM-034 0,08-0,25 Florfenicol
IBM-009 2-8 Florfenicol

92
4.2.5 Efectos del Metanol y Etanol en las determinaciones de CIM
Con el fin de descartar posibles efectos que tengan los solventes utilizados para disolver los
antibióticos se evaluó el efecto que estos alcoholes en distintas concentraciones pudieran
tener en el crecimiento de P. salmonis. El montaje del método fue el mismo utilizado para
determinar las CIM anteriormente, con la diferencia fundamental de que el antibiótico fue
reemplazado por el solvente y con adición de agua destilada para llegar al volumen final de
las concentraciones desarolladas.
Una vez crecida la bacteria y confirmada su identidad, las placas se incubaron en las
mismas condiciones utilizadas para las CIM anteriores, es decir por 5 días a 18°c y
posteriormente leyendo absorbancia a 600nm.
Estos efectos se evaluaron tanto para metanol (solvente de Oxitetraciclina) y etanol
(solvente de Florfenicol) y para cada una de las cepas analizadas, es decir LF-89, IBM-034,
IBM-009. Los resultados se grafican a continuación en la figura 19.

93
Figura 19. Efecto del Metanol y Etanol en el crecimiento de P. salmonis. Las bacterias
fueron cultivadas en medio líquido CASO suplementado, en placas de ELISA de 96
pocillos con distintas concentraciones evaluando el efecto del solvente, incubadas durante
5 días a 18°C. Se leyó su absorbancia a 600nm.

94
4.3 Cinéticas de expresión
Con el fin de evaluar el cambio de expresión de los genes previamente caracterizados en las
3 cepas de P. salmonis estudiadas se realizaron cinéticas para evaluar su expresión a 3
tiempos (2 horas, 4 horas y 6 horas). La cinética se realizó tratando los cultivos de P.
salmonis con distintas concentraciones de antibióticos (Oxitetraciclina y Florfenicol) tal
como se describe en el diseño experimental en el punto 3.2.9. De esta manera se pudo
evaluar el cambio en la expresión de los distintos genes posiblemente involucrados en
resistencia a antibióticos en el tiempo, con respecto a su expresión basal (bacteria sin
tratamiento), con el fin de realizar una comparación entre cepas, ver tiempo de cambios de
expresión y que tanto varian tras los tratamientos a 2 concentraciones distintas.
Las distintas cinéticas se realizaron evaluando cada tiempo con su respectivo tratamiento
con n=3, transcurrido el tiempo correspondiente se recolectaron las bacterias en estudio, se
les extrajo el RNA, se cuantifico y se verificó que los RNAs estén libres de gDNA y
posteriormente se sintetizó cDNA. El cDNA sintetizado se utilizó para evaluar la expresión
de cada gen en estudio mediante PCR en tiempo real. Para esto fue necesario diseñar
partidores para cada gen estudiado los que se describen en la tabla IV de materiales y
métodos. Cada set de partidores fue estandarizado y evaluado previamente por PCR en
tiempo real para confirmar que solo se obtuviera un producto para cada uno y la ausencia
de dimerización de los partidores.

95
4.3.1 Estandarización de partidores diseñados para PCR en tiempo real
Previo a realizar las mediciones de expresión génica por PCR en tiempo real se evaluó a
una temperatura de 60°C cada uno de los set de partidores diseñados y se observó que todos
formaban producto único y ausencia de dimerización de partidores. En la figura 20 (A,B y
C) se muestra el gráfico de las curvas de disociación del producto de RT-qPCR para cada
set de partidores utilizado para los genes de resistencia a Oxitetraciclina, en donde la
ordenada corresponde a la derivada de la fluorescencia emitida por el producto y la abcisa a
la temperatura en grados celsuis.

96
Figura 20. Estandarización de partidores para genes de resistencia a Oxitetraciclina.
Curva de disociación de los productos de RT-qPCR realizada por el equipo Stratagene
MxPro 3005 mediante el software MxPro, luego finalizada la amplificación. Realizado en
triplicado. A) Tet E, B) Tet C, C) Bomba transportadora de Tetraciclina.

97
Estandarizados los partidores para los genes de resistencia a Oxitetraciclina, se procedió a
evaluar los genes utilizados para evaluar la resistencia a Florfenicol.
En la figura 21 (A,B y C) se muestra el gráfico de las curvas de disociación del producto de
RT-qPCR para cada set de partidores utilizado para los genes de resistencia a Florfenicol,
en donde la ordenada corresponde a la derivada de la fluorescencia emitida por el producto
y la abcisa a la temperatura en grados celsuis.

98
Figura 21. Estandarización de partidores para genes de resistencia a Florfenicol.
Curva de disociación de los productos de RT-qPCR realizada por el equipo Stratagene
MxPro 3005 mediante el software MxPro, luego finalizada la amplificación. Realizado en
triplicado. A) Bcr/Cfla B) Bcr2 (Bcr/Cfla) C) Transportador de Cloranfenicol/Florfenicol.

99
4.3.2 Evaluación de la cinética de expresión para genes de resistencia a antibióticos
Se realizó la cuantificación de la expresión relativa para los genes putativos codificantes
para Tet E, Tet C, Bomba de tetraciclina, Bcr/Cfla, Bcr2 y transportador de
Cloranfenicol/Florfenicol. La cuantificación se realizó utilizando la formula descrita en
materiales y métodos. Obtenido el resultado de la formula se calculó la media para la
expresión relativa de los controles (bacteria sin tratamiento) de las cinéticas que se
utilizaron como expresión basal para cada gen estudiado, la cual en el gráfico corresponde a
1 de razón de expresión, en el eje de las ordenadas. De este modo se pudo calcular la
relación entre la media de expresión relativa de cada tiempo con la media calculada para su
control correspondiente, lo que permite observar si la expresión del gen aumenta o
disminuye con respecto al control. Como gen normalizador se utilizó el gen constitutivo de
P. salmonis RpoD.
4.3.2.1 Evaluación de la expresión de Tet E
LF-89: Tras un tratamiento con 0,02µg/ml de Oxitetraciclina se observó un aumento
significativo a las 4h post tratamiento cercano a las 35 veces con respecto al control, a las
6h post tratamiento se observo una disminución en la expresión al nivel del control. Tras un
tratamiendo con 0,08µg/ml de Oxitetraciclina se vió un aumento de expresión a las 2h con
respecto al control cercano a las 35 veces el cual se mantuvo alto hasta las 6h post
tratamiento.

100
IBM-034: Tras un tratamiento con 0,02 µg/ml de Oxitetraciclina no se observó un aumento
en la expresión con respecto al control, el cual si se observó tras un tratamiento con
0,08µg/ml de Oxitetraciclina donde aumento en 5 veces a las 2h.
IBM-009: Tras un tratamiento con 0,02µg/ml de Oxitetraciclina se observó un aumento de
expresión de 4 veces con respecto al control a las 4h, disminuyendo a las 6h. No se vieron
diferencias tras tratamiento con 0,08µg/ml.
El gráfico de la cinética de expresión de Tet E para las 3 cepas en estudio se puede observar
en la figura 22.

101
Figura 22. Gráfico de cinética de expresión de Tet E. Se evaluó mediante RT-qPCR la
expresión relativa de Tet E en la cepa tipo LF-89, IBM-034 e IBM-009 tras tratamientos
con Oxitetraciclina con respecto al control. Como gen normalizador se utilizó el gen RpoD.
Se utilizó un n=3, las barras indican el error estándar. Se considera p<0.01 como altamente
significativo (**) y p<0.05 como significativo (*) según la prueba estadística T de Student.

102
4.3.2.2 Evaluación de la cinética de expresión para Tet C
LF-89: No se observó expresión de Tet C en esta cepa (no ct)
IBM-034: No se observó expresión de Tet C en esta cepa (no ct)
IBM-009: Tras un tratamiento con 0,02µg/ml de Oxitetraciclina se observó un aumento de
expresión del doble con respecto al control, el cual se observó a las 2h, disminuyendo en el
tiempo. Tras un tratamiento con 0,08µg/ml de Oxitetraciclina se observó un aumento de
expresión del doble con respecto al control, el cual disminuyo en el tiempo (observándose
un descenso mas lento que en el tratamiento con 0,02µg/ml).
El gráfico de la cinética de expresión de Tet C para las cepas en estudio se puede observar
en la figura 23.

103
Figura 23. Gráfico de cinética de expresión de Tet C. Se evaluó mediante RT-qPCR la
expresión relativa de Tet C en la cepa tipo LF-89, IBM-034 e IBM-009 tras tratamientos
con Oxitetraciclina con respecto al control. Como gen normalizador se utilizó el gen RpoD.
Se utilizó un n=3, las barras indican el error estándar. Se considera p<0.01 como altamente
significativo (**) y p<0.05 como significativo (*) según la prueba estadística T de Student.

104
4.3.2.3 Evaluación de la cinética de expresión para bomba transportadora de
Tetraciclina
LF-89: Tras un tratamiento con 0,02µg/ml de Oxitetraciclina se observó un aumento de
expresión cercano a las 170 veces tras 4h de tratamiento, observándose una disminución al
nivel del control a las 6h. Tras un tratamiento con 0,08µg/ml de Oxitetraciclina se observó
un aumento cercano a las 50 veces con respecto al control pero más rápido, viéndose el
cambio a las 2h, con una posterior disminución en el tiempo.
IBM-034: Tras un tratamiento con 0,02µg/ml de Oxitetraciclina se observó un aumento
significativo a las 4h post tratamiento, de 60 veces con respecto al control, disminuyendo a
las 6h pero aun manteniéndose sobre el control. Tras un tratamiento con 0,08µg/ml se vió
un aumento de expresión cercano a las 300 veces con respecto al control a las 2h,
disminuyendo a las 4h a una expresión cercana a las 50 veces por sobre el control
manteniéndose a las 6h.
IBM-009: Tras un tratamiento con 0,02µg/ml se observó un aumento de expresión de 5
veces a las 4h, volviendo a su nivel basal a las 6h. No se observaron cambios significativos
tras un tratamiento con 0,08µg/ml.
El gráfico de la cinética de expresión para bomba transportadora de Tetraciclina para las
cepas en estudio se puede observar en la figura 24.
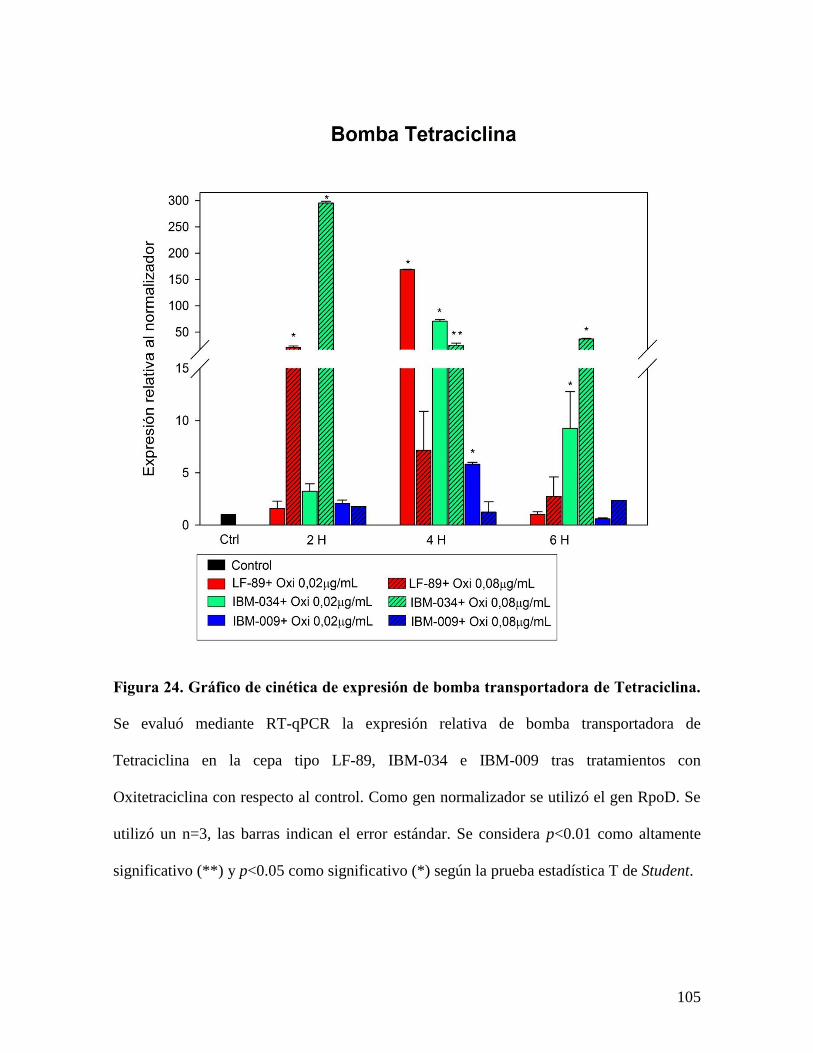
105
Figura 24. Gráfico de cinética de expresión de bomba transportadora de Tetraciclina.
Se evaluó mediante RT-qPCR la expresión relativa de bomba transportadora de
Tetraciclina en la cepa tipo LF-89, IBM-034 e IBM-009 tras tratamientos con
Oxitetraciclina con respecto al control. Como gen normalizador se utilizó el gen RpoD. Se
utilizó un n=3, las barras indican el error estándar. Se considera p<0.01 como altamente
significativo (**) y p<0.05 como significativo (*) según la prueba estadística T de Student.

106
4.3.2.4 Evaluación de la cinética de expresión para Bcr/Cfla
LF-89: Tras un tratamiento con 0,02µg/mL de Florfenicol se vió un aumento en la
expresión de una vez con respecto a la expresión basal, que se mantuvo por las primeras 4h
volviendo a su estado basal a las 6h. Tras un tratamiento con Florfenicol con 0,08µg/mL de
Florfenicol se vió un aumento significativo con respecto a la expresión basal de 8 veces a
las 2h post tratamiento el cual fue descendiendo en el tiempo llegando casi a niveles basales
a las 6h post tratamiento.
IBM-034: Tras un tratamiento con 0,02µg/mL de Florfenicol no se vieron grandes aumento
con respecto a la expresión basal hasta las 4h post tratamiento donde la expresión aumentó
1 vez por sobre el estado basal y a las 6h llegando a 2 veces por sobre la expresión basal.
Tras un tratamiento con 0,08µg/mL se vió un rápido aumento en la expresión de este gen
llegando a las 7 veces por sobre el estado basal a las 2h post tratamiento, el cual fue
disminuyendo en el tiempo.
IBM-009: Tras un tratamiento con 0,02µg/mL de Florfenicol no se vieron aumentos con
respecto a al expresión basal, los que si se observaron tras tratar con 0,08µg/mL de
Florfenicol donde se vió un aumento cercano a las 11 veces por sobre el estado basal a las
2h de tratamiento, descendiendo a estados basales ya a las 4h.
El gráfico de la cinética de expresión para el transportador Bcr/Cfla para las cepas en
estudio se puede observar en la figura 25.

107
Figura 25. Gráfico de cinética de expresión de transportador Bcr/Cfla. Se evaluó
mediante RT-qPCR la expresión relativa del transportador Bcr/Cfla en la cepa tipo LF-89,
IBM-034 e IBM-009 tras tratamientos con Florfenicol con respecto al control. Como gen
normalizador se utilizó el gen RpoD. Se utilizó un n=3, las barras indican el error estándar.
Se considera p<0.01 como altamente significativo (**) y p<0.05 como significativo (*)
según la prueba estadística T de Student.

108
4.3.2.5 Evaluación de la cinética de expresión para Transportador del tipo Bcr2
LF-89: No se observó expresión en esta cepa (no ct).
IBM-034: Tras un tratamiento con 0,02µg/mL de Florfenicol se observó un aumento de 1
vez con respecto a la expresión basal a las 4h post tratamiento. Tras un tratamiento con
0,08µg/mL se vió un aumento de 2 veces con respecto a la expresión basal a las 2h,
llegando hasta 4 veces tras 6h post tratamiento.
IBM-009: Tras un tratamiento con 0,02µg/mL prácticamente no se observó un aumento con
respecto a la expresión basal. Tras un tratamiento con 0,08µg/mL se observó un aumento
de 1 vez por sobre la expresión basal a las 2h, disminuyendo en el tiempo.
El gráfico de la cinética de expresión para el transportador del tipo Bcr2 para las cepas en
estudio se puede observar en la figura 26.

109
Figura 26. Gráfico de cinética de expresión de transportador del tipo Bcr2. Se evaluó
mediante RT-qPCR la expresión relativa del transportador del tipo Bcr2 en la cepa tipo LF-
89, IBM-034 e IBM-009 tras tratamientos con Florfenicol con respecto al control. Como
gen normalizador se utilizó el gen RpoD. Se utilizó un n=3, las barras indican el error
estándar. Se considera p<0.01 como altamente significativo (**) y p<0.05 como
significativo (*) según la prueba estadística T de Student.

110
4.3.2.6 Evaluación de la cinética de expresión para Transportador de
Cloranfenicol/Florfenicol
LF-89: No se observó expresión (no ct)
IBM-034: No se observó expresión (no ct)
IBM-009: Tras un tratamiento con 0,02µg/mL de Florfenicol se observó un aumento
significativo de 6 veces con respecto a su expresión basal a las 2h pos tratamiento, la que se
mantuvo alta a las 6h post tratamiento. Tras un tratamiento con 0,08µg/mL de Florfenicol
se observó un aumento de 5 veces a las 4h post tratamiento, el cual descendió en el tiempo.
El gráfico de la cinética de expresión para el Transportador de Cloranfenicol/Florfenicol
para las cepas en estudio se puede observar en la figura 27.

111
Figura 27. Gráfico de cinética de expresión de transportador de
Cloranfenicol/Florfenicol. Se evaluó mediante RT-qPCR la expresión relativa del
transportador de Cloranfenicol/Florfenicol en la cepa tipo LF-89, IBM-034 e IBM-009 tras
tratamientos con Florfenicol con respecto al control. Como gen normalizador se utilizó el
gen RpoD. Se utilizó un n=3, las barras indican el error estándar. Se considera p<0.01 como
altamente significativo (**) y p<0.05 como significativo (*) según la prueba estadística T
de Student.

112
5. Discusión
5.1 Identificación y análisis de genes posiblemente involucrados en resistencia a
Oxitetraciclina y Florfenicol
5.1.1 Tet E
Mediante los análisis in-silico de las cepas de P. salmonis realizados mediante el software
GenDB 2.2, se pudo identificar un gen codificante para una putativa proteína de resistencia
a Tetraciclinas y Oxitetraciclinas de 412 aá, según la anotación entregada por el software, la
cual se realiza por homolgía de su secuencia, se indicó que podría tratarse de Tet E, una
proteína transmembrana perteneciente a la familia de las Tet, que se encarga de conferir
resistencia a distintos tipos de Tetraciclinas (Chopra et al., 2001), exportando estas
moléculas hacia fuera de la bacteria evitando de esta manera que alcancen su concentración
letal. Este gen se encontró presente en todas las cepas analizadas de P. salmonis.
Mediante análisis de sus dominios utilizando la herramienta bioninformatica SMART se
pudo determinar dos dominios conservados, uno correspondiente a dominios
transmembrana, que corrobora el hecho de que se trate de una proteína transmembrana,
como es el caso de las bombas transportadoras de Tetraciclinas. Además se pudo
determinar un dominio conservado de la familia MFS (Major Facilitator Superfamily),
familia a la cual pertenecen las proteínas Tet (Paulsen et al., 1996), lo que corrobora los
indicios de que podría tratarse de una proteína de membrana que confiera resistencia a estas
moléculas antibacterianas.
Mediante el análisis de su secuencia aminoacídica realizado por BLAST hacia la base de
datos de NCBI se pudo observar que no existe una proteína que comparta una identidad

113
muy alta con esta, encontradose mayores identidades cercanas al 36%. Estas identidades
fueron las mas altas encontradas en esta base de datos, las que en su mayoría correspondían
a proteínas de resistencia a drogas de la familia MFS, en distintos tipos de Legionella, una
especie bacteriana cercana filogenéticamente a P. salmonis, también se pudo observar un
similar grado de identidad con una proteína transportadora para moléculas de Tetraciclinas.
El hecho de que las identidadas encontradas no superen el 40% permite descartar la
posibilidad asignarle como notación el nombre de una proteína o un gen ya existente en
otra especie bacteriana, dos genes se consideran de la misma clase y tienen la misma
designación si tienen una identidad en su secuencia aminoacídica mayor a 80% y son
considerados distintos genes si tienen menos de un 79% de identidad en su secuencia
aminoacídica (Levy et al., 1999), sin embargo que no contenga un alto porcentaje de
identidad aminoacídica también se debe a que presente aminoácidos equivalentes y no
necesariamente el mismo, sin afectar asi su naturaleza y función, como describe Mindlin et
al 2006.
Otra característica importante a destacar con respecto a este gen fue el hecho de que se
encuentra presente en la “Isla de patogenicidad 2 IBM-034”, según el trabajo realizado por
Lagos el 2013, la cual se encuentra presente en todas las cepas analizadas de P. salmonis, lo
que entrega indicios de como la bacteria pudo haber adquirido este gen de resistencia.
5.1.2 Tet C
Los análisis in-silico realizados por el software GenDB 2.2 hacia las cepas de P. salmonis
identificaron un gen codificante para una putativa de resistencia a Tetraciclina y
Oxitetracilina de 411 aá, dándole como notación el nombre putativo de Tet C. Al igual que
la proteína putativa Tet E (5.1.3) se trata de una proteína transmembrana involucrada en

114
conferir resistencia a distintos tipos de Tetraciclinas (Chopra et al., 2001), se descartó el
hecho de que se trate del mismo gen que el descrito en el punto 5.1.3 realizando un blast
entre ellas, otorgando una indentidad aminoacídica de 31%. Este gen no se vio presente en
todas las cepas de P. salmonis, observándose su ausencia en la cepa tipo LF-89
(perteneciente al genogrupo II, Figueroa et al., 2013), IBM-004 e IBM-034 (ambas
pertenecientes al genogrupo I, Figueroa et al., 2013).
Mediante la predicción de los dominios conservados para esta secuencia utilizando la
herramienta SMART se logró identificar un dominio conservado de proteínas
transmembrana, reafirmando asi que se trata de una proteína de este tipo. Además se
indetificó un dominio conservado de la familia MFS, familia a la cual pertenecen las
proteínas Tet, tal como lo describió Paulsen et al el año 2006, además de otras proteínas
que confieren resistencia a antibióticos.
El análisis de la secuencia aminoacídica de esta proteína mediante BLAST hacia la base de
datos del NCBI indicó mayores identidades aminoacídicas entre 40-50% con una proteína
de la familia Tet, además de otras proteínas transportadores de drogas presentes en
Coxiella, otra especie filogenéticamente cercana a P. salmonis. Sin embargo estos
porcentajes de identidad tampoco permiten afirmar que se trate de la misma proteína.
No se observó que este gen este presente en una isla de patogencidad, sin embargo en su
entorno génico se observó la presencia de transposasas, que podrían dar indicio de que el
gen podría formar parte de un transposon. El entorno génico de este gen en la cepa IBM-
009 no se observó en ninguna de las otras dos cepas analizadas, siendo el único de los
genes en estudio que no presenta un bloque de homología en estas cepas (datos analizados
mediante el software MAUVE).

115
5.1.3 Bomba transportadora de Tetraciclina
En el análisis in-silico mediante el software GenDB 2.2 se identificó un gen putativo
codificante para un transportador de tetraciclinas de 417 aá, asignándole esta notación,
indicándose que se podría tratar de otra proteína encargada en conferir resistencia a
Oxitetraciclina. Este gen se vio presente en todas las cepas analizadas de P. salmonis.
Realizando un análisis de sus dominios conservados utilizando la herramienta SMART se
pudo observar un dominio conservado de transmembrana reafirmando que se trata de una
proteína transmembrana, además se observó un dominio conservado característico de la
familia MFS, familia a la que pertenecen proteínas de transmembrana involucradas en
resistencia a Tetraciclinas y otras drogas, al igual que lo visto en los dos genes previamente
analizados en los puntos 5.1.1 y 5.1.2.
El análisis de la secuencia aminoacídica mediante BLAST hacia la base de datos de NCBI
indico mayores identidades aminoacídicas hacia proteínas de resistencia a Oxitetraciclina,
uno cercano al 60% y otros en el rango del 30% de identidad. Sin embargo con estos
porcentajes de identidad no se puede asegurar que se trate de la misma proteína.
No se observó que este gen forme parte de una isla de patogenicidad, sin embargo se
observaron otras bombas putativas para resistencia ubicadas cerca de este gen. Se pudo
observar que el entorno génico, comprendido por un bloque de homología génica analizado
mediante el software MAUVE es el mismo en las tres cepas analizadas.
Estos resultados indican que los tres genes de resistencia específica hacia oxitetraciclinas
encontrados en los genomas de estas cepas corresponden a bombas transportadoras de
oxitetraciclinas, algo que se ha podido ver en trabajo previos donde se ha descrito que en
bacterias gram negativas el mecanismo predominante de resistencia a tetraciclinas son las
bombas de flujo de drogas (Poole 2005). Además este tipo de resistencia a tetraciclinas por

116
bombas transportadoras también ha sido observada en otros patógenos de peces como
Photobacterium, Vibrio, Pseudomonas, Alteromonas, Citrobacter y Salmonella spp, visto
en trabajos previos por Furushita et al 2003.
5.1.4 Transportador del tipo Bcr/Cfla
En el análisis realizado mediante el software bioinformático GenDB 2.2 se identificó un
gen putativo codificante para una proteína de 417 aá transportadora de la familia Bcr/Cfla,
las que corresponden a proteínas transmembranas (12 segmentos transmembrana) las cuales
confieren resistencia a diversos antibióticos, entre ellos moléculas como Cloranfenicol y
Florfenicol. Este gen se vio presente para todas las cepas de P. salmonis. Según el análisis
de dominios conservados realizado mediante la herramienta SMART se observo un dominio
conservado MFS característico de esta superfamilia, a la cual corresponde la familia de
proteínas transportadoras de drogas Bcr/Cfla (Poole 2005). Además se pudo observar un
dominio conservado de transmembrana, reafirmando que se trata de una proteína de
membrana. Los análisis de la secuencia aminoacídica por BLAST hacia la base de datos de
NCBI no se determinaron proteínas con un alto porcentaje de identidad hacia esta proteína,
los mayores valores de identidad rondeaban los 30%, los que correspondieron a proteínas
de resistencia a Florfenicol, como transportadores del tipo FlorR. Al no observarse
porcentajes altos de identidad aminoacídica no se puede determinar si se trata
efectivamente de esas proteínas. Sin embargo se ha observado que proteínas
transportadoras de Florfenicol como CmlA, presentan el dominio conservado de la familia
MFS y dominios de la subfamilia Bcr/Cfla (NCBI).
En el entorno génico de este gen se observaron otros genes codificantes a proteínas de la
familia MFS y otros transportadores de drogas, indicando que estos se ubican muy cercanos

117
en el genoma, lo que se correlaciona con lo descrito en el trabajo de Poole en 2005, donde
se explica que los exportadores específicos para cloranfenicol y florfenicol de la familia
MFS por lo general son codificados en elementos móviles junto a otro genes de resistencia,
sin embargo no se observó que formara parte de una isla de patogenicidad. Este bloque de
homología en el que se encontraron estos genes esta presente en las tres cepas estudiadas,
manteniéndose conservado entre estas. Se han descrito clusters que contienen diversos
genes de resistencia y que por lo general están acompañados de elementos de movilidad
genética (como transposones o integrones) de esta manera se mueven en bloques entre
moléculas de DNA (Li et al., 1999), lo que puede explicar el hecho de que se encuentren
mas de un gen de resistencia en el mismo entorno génico.
5.1.5 Transportador del tipo Bcr2 (Bcr/Cfla)
Mediante el análisis bioinformatico realizado a través de GenDB 2.2, se identificó un gen
putativo codificante para una proteína de 416 aá, el programa le asignó como notación una
proteína transportadora del tipo Bcr2 (Bcr/Clfa), subfamilia que ha sido descrita como
proteínas transmembrana de resistencia a antibióticos, pertenecientes a la familia MFS. Este
gen se pudo ver presente en el genoma de todas las cepas analizadas de P. salmonis.
El análisis a sus dominios conservados mediante la herramienta bioinformática SMART
indicó un dominio característico de la familia MFS, corroborando en parte la notación
asignada por GenDB 2.2, los análisis de la secuencia aminoacídica realizados por BLAST a
la base de datos de NCBI indicaron mayores porcentajes de identidad entre 35-37% con
proteínas transportadoras de drogas de la familia MFS, tal como lo indicó el análisis por
GenDB 2.2, además también se pudieron observar identidades cercanas al 30% con
proteínas transportadoras de Florfenicol en especies de Legionella, especie

118
filogenéticamente cercana a P. salmonis, sin embargo las identidades observadas no son lo
suficientemente altas para decir que se trata de la misma proteína y no se puede clasificar
como una de ellas. El entorno génico para este gen fue el mismo en las 3 cepas analizadas,
mediante el software MAUVE se pudo observa un bloque de homología que contiene los
mismos genes, entre los que se pudieron observar integrasas y otras bombas transportadoras
de metales pesados, esto se correlaciona con la publicación previamente realizada por
Baker-Austin et al., 2006 donde se muestra que la asociación de la resistencia a antibióticos
y resistencia a metales pesados es muy frecuente en un mismo organismo, y que estos genes
suelen encontrarse en el mismo entorno génico móvil (plásmido, transposon, integron), lo
que puede explicar además la presencia de integrasas.
5.1.6 Transportador de Cloranfenicol/Florfenicol
Los análisis bioinformaticos mediante GenDB 2.2 identificaron un gen que codifica para
una proteína putativa transmembrana de 410 aá que actúa como transportador de
Cloranfenicol y Florfenicol. De todas las cepas analizadas, todas mostraron la presencia de
este gen.
Según un análisis a la secuencia codificante mediante la herramienta bioinformática
SMART se pudieron identificar dominios conservados en esta proteína los que
correspondieron a un dominio característico de la familia MFS, familia de la cual forman
parte numerosas proteínas de resistencia a drogas. Además se pudieron observar dos
dominios transmembrana, corroborando que se trata de una proteína de membrana, tal
como lo predicho por GenDB 2.2.
Según lo observado por los análisis por BLAST a la base de datos de NCBI se identificaron
mayores porcentajes de identidad aminoacídica con una proteína de resistencia a drogas

119
presente en Coxiella burnetti (46%) otra especie cercana filogenéticamente a P. salmonis.
Además se pudieron observar porcentajes de identidad menores con algunas proteínas de
resistencia a Florfenicol. Si bien estos porcentajes indican homología con estas proteínas,
no se puede determinar que correspondan a la misma, aunque comparte similitudes y
motivos de consensos con proteínas de este tipo. Según información presente en NCBI se
ha visto que proteínas transportadoras de Florfenicol como CmlA, presentan el dominio
conservado de la familia MFS y dominios de la subfamilia Bcr/Cfla.
El análisis a su entorno génico realizado mediante el software MAUVE indicó un bloque de
homología para las tres cepas analizadas, en la que se encuentra este gen. Dentro de este
bloque de homología conservado tambien se pudo observar otro gen putativo de resistencia
multiple a drogas, ademas de transposasas, por lo que una probable razón de la adquisición
de este gen es que se encuentre dentro de un transposon.
5.2 Determinación de sensibilidad y resistencia a Oxitetraciclina y Florfenicol en cepas
de P. salmonis
Importantes variaciones en los patrones de la resistencia in-vitro a antibióticos se ha visto
en aislados de distintas especies de salmones, zonas geográficas y ambientes acuáticos han
sido observadas en los últimos años (Mora 2010) mostrando un potencial riesgo de el
generamiento de resistencia antimicrobiana. Oxitetraciclina y Florfenicol son los dos
antibióticos mas utilizados para combatir P. salmonis en la industria salmonera en nuestro
país (Sernapesca 2011). En esta tesis se logró desarrollar un método para determinar el
nivel de resistencia que distintas cepas de P. salmonis presentan ante estos dos antibióticos,
basándose en un método de diluciones seriadas de ambos antibióticos en microplacas,

120
cultivando la bacteria en medio liquído (medio CASO suplementado). Mediante este
método se logró obtener datos confiables sobres las CIM (Concentración inhibitoria
mínima) para las distintas cepas estudiadas, desafiándolas a Oxitetraciclina y Florfenicol.
Las cepas utilizadas en esta tesis no habían sido estudiadas previamente y se compararon
con los resultados obtenidos para la cepa tipo LF-89.
Previo a este estudio para poder determinar la CIM en P. salmonis se debio estandarizar la
metodología, para esto se utilizó la cepa E. coli ATCC 25922, la cual se utiliza como cepa
control presentando rangos establecidos para estos antibióticos descritos en la literatura
(Miller et al., 2005). Esto sirve como control de calidad de la técnica, los rangos obtenidos
en esta tesis concuerdan con los rangos aceptados en la literatura, por lo que la técnica esta
bien estandarizada y se pudo proceder a trabajar con las cepas en estudio.
5.2.1 CIM para Oxitetracilina en P. salmonis
Se logró determinar la CIM de Oxitetraciclina para las cepas IBM-034 e IBM-009 de P.
salmonis realizando una comparación frente a la cepa tipo LF-89.
Los datos obtenidos permitieron determinar que la cepa IBM-034 (genogrupo I) presenta la
misma resistencia a Oxitetraciclina que la cepa de referencia LF-89 (genogrupo II) (0,08-
0,25µg/m). La cepa IBM-009 (genogrupo VIII) mostró ser más resistente a este antibiótico
(0,5-2µg/mL), debiéndose utilizar una mayor concentración de Oxitetraciclina para detener
su crecimiento. Estos resultados tienen concordancia con lo visto previamente mediante los
análisis in-silico a estas cepas, donde se pudo observar que las cepas pertenecientes al
genogrupo I (como IBM-034) y la cepa de referencia LF-89 presentaban un menor número
de genes putativos para resistencia a antibióticos, además ambas cepas mostraron ser muy

121
similares entre si, a diferencia de la cepa IBM-009 que mostró tener mayor cantidad de
genes de resistencia a antibióticos.
5.2.2 CIM para Florfenicol en P. salmonis
Se logró determinar la CIM de Florfenicol para las cepas IBM-034 e IBM-009 de P.
salmonis realizando una comparación frente a la cepa tipo LF-89.
Los rangos de CIM obtenidos para Florfenicol indicaron lo siguiente:
Los datos obtenidos para las CIM de Florfenicol indicaron que la cepa IBM-034
(genogrupo I) presenta la misma resistencia a Florfenicol que la cepa de referencia LF-89
(genogrupo II) (0,08-0,25µg/mL), además se determinó que la cepa IBM-009 presenta una
resistencia considerablemente mayor a este antibiótico que las dos descritas previamente
(2-8µg/mL), lo mismo que se observó para Oxitetraciclina. Esto nuevamente es
concordante con los datos observados in-silico donde se vio que esta cepa presenta mayor
número de genes de resistencia a antibióticos que la cepa tipo y las del genogrupo I. Esto
puede explicar en parte que la cepa IBM-009 presente mayor resistencia ante ambos
antibióticos estudiados.
Comparando ambos antibióticos en la cepa IBM-009 se pudo observar que esta cepa
presenta una mayor resistencia a Florfenicol que a Oxitetraciclina, estos datos concuerdan
con los obtenidos por Yañez et al 2013 donde también se pudo observar que las cepas
estudiadas eran mas susceptibles a Oxitetraciclina. Cabe destacar que esta diferencia no se
observó para la cepa de referencia LF-89 donde se determinó el mismo rango de resistencia
para ambos antibióticos, valores que son concordantes con los determinados en esta tesis
para LF-89.

122
Existen otros estudios previos realizados por Smith et al., en 1996 para determinar la CIM
de Oxitetraciclina, Cloranfenicol, Eritromicina entre otros antibióticos en 4 cepas de P.
salmonis, sin embargo ese estudio fue realizado creciendo las bacterias en células CHSE-
214 (aún no se desarollaba un cultivo líquido para crecer la bacteria). Por otra parte estos
valores se pueden comparar con especies bacterianas filogenéticamente relacionadas, como
lo visto por Bohle et al., 2009, donde se determinaron las CIM para Francisella
philomiragiase, obteniendo una CIM para oxitetraciclina de 0,5µg/mL y de 1µg/mL para
florfenicol, valores que se acercan a los obtenidos en esta tesis para las cepas de P.
salmonis. Además datos obtenidos por Soto et al., 2010 indicaron que para Francisella
asiatica otro patógeno de peces, indicaron que su CIM para florfenicol era de 2µg/mL, otro
valor dentro del rango de los obtenidos en esta tesis.
Como proyecciones a estos experimentos se deberían realizar las mismas pruebas para
determinar CIM pero creciendo estas cepas bacterianas intracelularmente, con el fin de
poder comparar como varía el grado de resistencia de estas bacterias creciendo en un medio
líquido libre de células y de manera intracelular (debido a que se trata de una bacteria
intracelular facultativa). Esta comparación permitiría obtener información de como abordar
este patógeno una vez que ya ha infectado al pez.
5.2.3 Efectos del Metanol y Etanol sobre las determinaciones de CIM
Para determinar si los solventes de los antibióticos utilizados influían o no en la
determinación de las CIM, se estudió el efecto tanto de metanol como de etanol en el
crecimiento de la bacteria, como se describe en los resultados 4.2.5. En ningún caso se
observó que el crecimiento de las distintas cepas de P. salmonis se viera afectado por estos
solventes, indicando así que la ausencia de crecimiento de P. salmonis en las pruebas

123
anteriores es netamente efecto del antibiótico al que la bacteria es expuesta y los solventes
de estos antibióticos no tienen interferencia en esto.
5.3 Cinéticas de expresión
Encontrados ciertos genes que podrían estar involucrados en conferir resistencia a
antibióticos en las cepas de P. salmonis, se seleccionaron 6 de estos para realizar una
investigación de como varía su expresión en respuesta a los tratamientos de la bacteria con
estos antibióticos, es decir se evaluó de que manera la bacteria responde del punto de vista
de la expresión génica al verse sometida a la presencia de estos antimicrobianos
(concentraciones de antibióticos a las cuales la bacteria aun es resistente).
Estudios previos hacia genes que codifican para proteínas transmembrana de resistencia a
Oxitetraciclina indican que la expresión de estos genes se ve estimulada en presencia de
moléculas de Tetraciclinas y/o Oxitetraciclinas. Esto se debe a que existen genes
represores que codifican para proteínas que se unen al promotor del gen de resistencia. En
presencia del antibiótico estas proteínas sufren un cambio conformacional al unirse las
moléculas de antibiótico, de este manera la proteína represora se disocia del promotor y se
produce un aumento en la expresión del gen de resistencia en respuesta a la presencia de las
moléculas del antibiótico (Berens et al., 2003).
Los datos obtenidos en este estudio mostraron que bajo tratamientos con Oxitetraciclinas
los tres genes putativos de resistencia para esta molécula antimicrobiana (Tet E, Tet C y
bomba transportadora de Tetraciclina/Oxitetraciclina) sufrieron un aumento en su expresión
con respecto a la expresión en estado basal (bacteria creciendo en medio liquído sin
estimulos con antimicrobianos).

124
El gen codificante para Tet E mostró un aumento significativo tras las 4h post tratamiento
de 0,02µg/mL de Oxitetraciclina en la cepa LF-89, en IBM-034 no se observó un aumento
en la expresión a esta concentración. En la cepa IBM-009 se observó un aumento, aunque
menor, con respecto al estado basal también tras las 4h post tratamiento. En las dos cepas
que se vieron aumentos se observó un descenso de la expresión tras 6h post tratamiento. Al
tratar la cepa IBM-034 con una mayor concentración de antibiótico (0,08µg/mL) si se
observó un aumento de expresión tras 2h post tratamiento.
Al analizar la cinética de expresión para Tet C, se pudo reafirmar lo visto mediante los
análisis in-silico, el que indicó que este gen no se encuentra en la cepa LF-89 y tampoco en
IMB-034, al no verse expresión mediante los análisis por qPCR, es decir el análisis por
qPCR permite afirmar que no es un problema de la secuenciación del genoma de la bacteria
que puede presentar la ausencia de un gen ya que no esta 100% completa, si no que
efectivamente el gen se encuentra ausente en esas cepas. Para el caso de la cepa IBM-009
donde si se había visto presencia del gen, se observó un aumento al doble en la expresión
tras 2h post tratamiento, este aumento fue disminuyendo en el tiempo. El aumento en la
expresión fue el mismo para las dos concentraciones de antibiótico evaluadas, sin embargo
al tratar con una concentración mayor se pudo observar que el descenso posterior de la
expresión fue más lento.
El análisis a la cinética de expresión para la bomba transportadora de Tetraciclinas mostró
los niveles de aumento de expresión más altos de estos 3 genes. Tanto para LF-89 como
para IBM-034 se vio un aumento significativo de expresión a las 4h post tratamiento, se
observó un descenso de esta expresión tras 6h post tratamiento. Al tratarse con una
concentración mayor de antibiótico (0,08µg/mL) se pudo ver que el aumento en la
expresión fue mas rápido, viéndose el mayor nivel de expresión a las 2h post tratamiento,

125
indicando que la bacteria responde mas rápido al verse sometida a una mayor cantidad de
antibiótico. En la cepa IBM-009 también se pudo observar un aumento de expresión con
respecto al estado basal tras las 4h post tratamiento, sin embargo fue un aumento
considerablemente menor en comparación a las otras dos cepas analisadas. Esto se puede
deber a que la cepa IBM-009 presente un estado basal de expresión para este gen mayor a
las dos primeras cepas, de manera que el aumento de expresión con respecto a su estado
basal no varía mucho, ya que según los análisis in-vitro de resistencia mostró ser una cepa
mas resistente, es decir mas adaptada que las otras dos.
Mediante estos análisis se pudo ver que en la mayoría de los casos la respuesta hacia el
desafío con Oxitetraciclina en cuanto a expresión génica era rápida, siempre en el rango de
las 2-4 horas, observándose ya un descenso de expresión tras las 6 horas post tratamiento,
probablemente debido a que la bacteria en este tiempo ya fue capaz de expulsar las
moléculas de antibiótico fuera de la célula. Estos resultados pueden correlacionarse con los
vistos por Berg et al., en 1988, donde se demostró que una sobre expresión de proteínas
integrales de membrana de resistencia a estos antimicrobianos era letal para E. coli,
probablemente debido a un colapso en el potencial de membrana (Eckert et al., 1989), por
lo que la expresión de estos genes debe ser estrictamente controlada en ausencia del
antibiótico.
Los datos obtenidos en la cinética bajo tratamientos con Florfenicol mostraron que los tres
genes putativos de resistencia hacia este antibiótico (transportador Bcr/Cfla, transportador
Bcr2/Cfla y transportador de Cloranfenicol/Florfenicol) aumentaron su expresión tras estos
tratamientos con respecto a su expresión basal. Estudios previos hacia proteínas
transportadoras de Cloranfenicol y Florfenicol como CmlA1, mostraron que la expresión de

126
estas podía ser inducida bajo concentraciones subinhibitorias de Cloranfenicol (George et
al., 2002).
El gen codificante para el transportador Bcr/Cfla se vio aumentado en las cepas LF-89 e
IBM-034 pero no en niveles significativos tras tratamientos con Florfenicol a 0,02µg/mL,
para la cepa IBM-009 no se vieron aumentos a esta concentración. Por el contrario tras
tratatamientos a una concetracion de 0,08µg/mL se observaron aumentos de la expresión de
este gen tras las 2h post tratamiento en las tres cepas estudiadas, expresión que fue
descendiendo hasta llegar a su nivel basal a las 6 horas post tratamiento, lo mismo que se
pudo observar previamente para los genes de resistencia a Oxitetraciclina. Esto indica que
para estimular la expresión de este gen y observar cambios significativos se necesitaron
concentraciones de 0,08µg/mL de Florfenicol.
El análisis para la cinética de expresión para el gen codificante al transportador Bcr2/Cfla
indicó que no hubo grandes aumentos de expresión tras tratamiento con Florfenicol. En la
cepa IBM-034 se observó un aumento de una vez por sobre la expresión basal a las 4h. Al
tratar con una concentración mayor de Florfenicol se observó el mismo aumento pero a las
2h post tratamiento. En la cepa IBM-009 se observaron los mismos resultados. Sin embargo
en la cepa LF-89 no se observó expresión de este gen, a diferencia de lo que se había visto
por análisis bioinformaticos que indicaban que esta cepa si presentaba el gen. Una
posibilidad de esta falta de expresión puede deberse a que este gen se encuentre como un
pseudogen en esta cepa, los pseudogenes son comúnmente producidos por pequeños
cambios en el marco de lectura producidos por pequeñas deleciones, inserciones, etc. (Lerat
et al 2005). Comparando dos secuencias homologas de un mismo gen presente en genomas
cercanamente relacionados como es el caso de estas cepas se pueden detectar pequeños
cambios en la secuencia que podrían dar pista si se trata efectivamente o no de un

127
pseudogen, comparando ambas secuencias se vio una identidad de un 96%, indicando que
hay pequeñas diferencias en sus secuencias, posiblemente estos cambios podrían estar en su
secuencia reguladora, sin embargo un mayor número de análisis debería ser realizado para
ver si efectivamente es este el caso o no. Otro motivo puede deberse a que este gen de la
cepa LF-89 presente mutaciones en su promotor, esto no pudo ser evaluado debido a que
mediante el análisis por GenDB 2.2 se analiza la secuencia codificante y no el promotor,
por lo que mayor número de análisis deberían ser realizados para este caso. Una tercera
opción podría ser probar otro set de partidores para este gen y ver si se ve o no expresión.
Finalmente para el gen que codificante para el transportador de Cloranfenicol/Florfenicol,
mediante las cinéticas de expresión se observó que en la cepa IBM-009 hubo un aumento
de expresión tras 2h de tratamiento con Florfenicol. Para la cepa LF-89 e IBM-034 no se
vio expresión para este gen. Al igual que para el gen codificante para la proteína Bcr2/Cfla
esto puede deberse a las opciones descritas para el caso anterior, estos genes mostraron
una identidad del 99%.
Cabe destacar que estos experimentos tanto de análisis in-silico, como de expresión génica,
son solo un comienzo para el análisis de resistencia a nivel genómico, como proyecciones a
esta investigación se deberían realizar experimentos que demuestren funcionalidad de las
proteínas para las que estos genes codifican y de esa manera ver si efectivamente son
determinantes en la resistencia.

128
Basandose en los resultados obtenidos en esta tesis se puede concluir que:
- Se pudo estandarizar un método efectivo de micro diluciones en placas para determinar la
concentración inhibitoria minima de P. salmonis tanto para Oxitetracicina como
Florfenicol, pudiendo crecer la bacteria en un medio líquido libre de células.
- La resistencia a los antibióticos Oxitetraciclina y Florfenicol varía entre las cepas de P.
salmonis estudiadas, siendo menos resistente la cepa tipo LF-89. Además esta variación de
resistencia parece estar íntimamente relacionada con el número de genes de resistencia a
antibióticos encontrados en sus genomas y a la expresión de estos.
- Los genes putativos para conferir resistencia a antibióticos, encontrados mediante análisis
in-silico en estas cepas si parecen tener relación directa con la resistencia a estos
antimicrobianos, lo que se vio tanto por análisis de homología aminoacídica de sus
secuencias como por análisis in-vitro.
- Oxitetraciclina y Florfenicol inducen efectivamente un aumento considerable en la
expresión de estos genes con respecto a su expresión basal. Sin embargo, se deberían
realizar análisis futuros a estos genes para ver si esta resistencia conferida es específica para
estos o bien son genes que actúan en conferir resistencia a un número mayor de
antibióticos.
- Se logró comparar genéticamente y fenotípicamente aislados nacionales de P. salmonis,
en su grado de resitencia, lo que es importante debido a que en el país no existen
demasiadas investigaciones para caracterizar completa y epidemiológicamente a las
bacterias resistentes y a los genes responsables de esta resistencia.

129
6. Referencias
Almendras F.E., Fuentealba I.C., Jones S.R.M., Markham F. & Spangler E. (1997)
Experimental infection and horizontal transmission of Piscirickettsia salmonis in
freshwater-raised Atlantic salmon, Salmo salar L. Journal of Fish Diseases 20, 409–418.
Baker-Austin C., Wright M., Stepanauskas R. & Mcarthur J.V. (2006) Co-selection of
antibiotic and metal resistance. TRENDS in Microbiology 14, 176-182.
Bauer, A., Kirby, W., Sherrisj, C. (1996). Antibiotic susceptibility testing by standardized
single disc method. American Journal Clinical Pathology. 45 : 493 – 496.
Bohle, H., Tapia, E., Martinez, A., Rozas, M., Figueroa, A., Bustos, P. (2009). Francisella
philomiragia, bacteria asociada con altas mortalidades en salmones del atlántico (salmo
salar) cultivados en balsas-jaulas en el lago Llanquihue. Arch Med Vet. 41: 237 – 244.
Branson E.J. & Diaz-Munoz D. (1991) Description of a new disease condition occurring in
farmed coho salmon, Oncorhynchus kisutch (Walbaum), in South America. Journal of Fish
Diseases 14, 147–156.
Bravo S. & Campos M. (1989) Coho salmon syndrome in Chile. Fish Health Section
Newsletter, American Fisheries Society 17, 3.

130
Brocklebank J.R., Speare D.J., Armstrong R.D. & Evelyn T. (1992) Septicemia suspected
to be caused by a rickettsia-like agent in farmed Atlantic salmon. Canadian Veterinary
Journal 33, 407–408.
Bushman, F. (2002). Lateral DNA Transfer, Mechanisms and Consequences. Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, New York.
Cabello, F. (2003). Antibióticos y acuicultura un análisis de sus potenciales impactos para
el medio ambiente, la salud humana y animal en Chile. APP.
17: 1 – 14.
Cabello, F. (2004). Antibióticos y acuicultura en Chile: consecuencias para la salud humana
y animal. Rev. Méd. Chile. 132: 1001 – 1006.
Cannon M., Hartford S., Davies J. (1990). A comparative study on the inhibitory actions of
chloramphenicol, thiamphenicol and some fluorinated derivatives. Antimicrob.
Chemotherpy, 26: 307-317.
Chopra I. and Roberts M. (2001). Tetracycline Antibiotics: Mode of Action, Applications,
Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiology and
molecular biology reviews. 65: 232–260.
Courvalin, P. and Trieu-Cuot, P. (2001). Minimizing potential resistance: The molecular
view. Clin. Infect. Dis.33(Suppl. 3): S138-S146.

131
Cvitanich J.D., Garate N. O. & Smith C.E. (1991) The isolation of a rickettsia-like
organism causing disease and mortality in Chilean salmonids and its confirmation by
Koch’s postulate. Journal of Fish Diseases 14, 121–145.
Evelyn T.P.T., Kent M.L., Poppe T.T. & Bustos P. (1998) Salmonid rickettsial septicemia.
In: Diseases of Seawater Netpen- Reared Salmonid Fishes (ed. by M.L. Kent & T.T.
Poppe), pp. 31–33. Dept. Fisheries and Oceans, Nanaimo, B.C.
Figueroa J., Rojas C., Ojeda N., Haussmann D., Jara C., Isla A., Torres E., Enriquez R.,
Yañez A., and Romero A.
(Paper en preparación) Genetic diversity among Piscirickettsia
salmonis isolates: classification through genomic approximation.
Fluit AC, Visser MR, Schmitz FJ. Molecular detection of antimicrobial resistance. Clin
Microbiol Rev 2001;14:836–71.
Furushita M., Shiba T., Maeda T. (2003) Similarity of tetracycline resistance genes isolated
from fish farm bacteria to those from clinical isolates. Appl Environ Microbiol. 69: 5336–
42.
Fryer J.L., Lannan C.N., Garcés L.H., Larenas J.J. & Smith P.A. (1990) Isolation of a
rickettsiales-like organism from disease coho salmon Oncorhynchus kisutch in Chile. Fish
Pathology 25, 107–114.

132
Fryer J.L., Lannan C.N., Giovannoni S.J., Wood N.D. (1992) Piscirickettsia salmonis gen.,
sp. nov., the causative agent of a epizootic disease in the Salmonid Fishes. Int. J. Syst.
Bacteriol. 42: 120-126.
Fryer J.L. (2002) Clarification of the systematics of Piscirickettsia salmonis. Fish Health
Section Newsletter, American Fisheries Society 30, 23.
Fryer J.L., Hedrick R P (2003). Review Piscirickettsia salmonis: a Gram-negative
intracellular bacterial pathogen of fish. J. Fish Dis. 26: 251-262.
George A., Hall R. (2002). Efflux of chloramphenicol by the CmlA1 protein. FEMS
Microbiology Letters 209:209-213.
Goldman RA, Hasan T, Hall CC, Strycharz WA, Cooperman BS. (1983).
Photoincorporation of tetracycline into Escherichia coli ribosomes. Identification of the
major proteins photolabeled by native tetracycline and tetracycline photoproducts and
implications for the inhibitory action of tetracycline on protein synthesis. Biochemistry.
22(2):359–368.
Jara M. A. (2007). Tetracyclines: A model of antimicrobial resistance. Avances en Ciencias
Veterinarias. 22: 49-55.
Larenas J., Hidalgo, L., Garcés, H., Fryer, J.L., Smith, P. (1995). Piscirickettsiosis:
Lesiones en salmón del Atlántico (Salmo salar) infectados naturalmente con
Piscirickettsia salmonis. Av. Cienc. Vet. 10: 53 - 58.

133
Larenas J.J., Bartholomew J., Troncoso O., Fernández S., Ledezma H., Sandoval N., Vera
P., Contreras J., Smith P. (2003). Experimental vertical transmission of Piscirickettsia
salmonis and in vitro study of attachment and mode of entrance into the fish ovum. Dis.
Aquat. Org. 56: 25-30.
Lerat E., Ochman H. (2005). Recognizing the pseudogenes in bacterial genomes. Nucleic
Acids Research. 33: 3125-3132.
Levin, B.R., and Bergstrom, C.T. (2000). Bacteria are different: Observations,
interpretations, speculations, and opinions about the mechanisms of adaptive evolution in
prokaryotes. Proc. Natl. Acad. Sci. USA 97: 6981-6985.
Levy, S. B., L. M. McMurry, T. M. Barbosa, V. Burdett, P. Courvalin, W. Hillen, M. C.
Roberts, J. I. Rood, and D. E. Taylor. (1999). Nomenclature for new tetracycline resistance
determinants. Antimicrob. Agents Chemother. 43: 1523–1524.
Levy, S.B. (2001). Antibiotic resistance: Consequences of inaction. Clin. Infect. Dis. 33
(Suppl.3):S124-S129.
Li X-Z, Nikaido H. (2009). Efflux-Mediated Drug Resistance in Bacteria: an Update.
Drugs. 69(12): 1555–1623

134
Mauel M.J., Giovannoni S.J., Fryer J.L. (1996). Development of polymerase chain reaction
assays for detection, identification and differentiation of Piscirickettsia salmonis. Dis Aquat
Org., 26: 189-195.
Mauel MJ., Miller D.L (2002). Piscirickettsiosis and piscirickettsiosis-like infections in
fish: a review. Vet. Microbiol. 87: 279-289.
McFarland. J., Amer. J. (1907). Sodium boric acid: a Tris-free, cooler conductive medium
for DNA electrophoresis. Med. Assoc. 14,1176.
Mendez, B., C. Tachibana and S. B. Levy. (1980). Heterogeneity of tetracycline resistance
determinants. Plasmid 3: 99–108.
Michalova, E., Novotna, P., Schlegelova, J. (2004). Tetracyclines in veterinary medicine
and bacterial resistance to them. Vet. Med. – Czech 49, (3): 79-100.
Miller, R. A., Walker, R. D., Carson, J., Coles, M., Coyne, R., Dalsgaard, I., Gieseker, C.,
Hsu, H. M., Mathers, J.J., Papapetropoulou, M., Petty, B., Teitzel, C., Reimschuessel, R.
(2005). Standardization of a broth microdilution susceptibility testing method to determine
minimum inhibitory concentrations of aquatic bacteria. Dis. Aquat. Org. 64: 211 – 222.
Mindlin S. Z., Petrova M. A., Bass I. A. & Gorlenko M. Zh. (2006). Origin, Evolution, and
Migration of Drug Resistance Genes. Russian Journal of Genetics. 42: 1257–1271.

135
Miranda, C. D., Judge, K., Crest, A. and Roberts, M. C. (2001). Distribution of tetracycline
resistance genes in bacteria from Chilean salmon farms. ICAAC Abstract.
Miranda, C.D. and Zemelman, R. (2002) Antimicrobial multiresistance in bacteria isolated
from freshwater Chilean salmon farms. Sci. Total Environ. 293: 207-218.
Mora J. (2010) Antibiogram analyses strains of Piscirickettsia salmonis isolated from
salmonid and trout, reared in the region of Los Ríos, Los lagos and Aysén during 2007-
2008. Bachelor Veterinary Science Thesis. Universidad de Concepción, Facultad de
Ciencias Veterinarias, Chile.
Murray, I.A. and Shaw, W.V. (1997). O-acetyltransferases for chloramphenicol and other
natural products. Antimicrob. Agents Chemother. 41: 1–6.
Neu, K. P. and Fu, H. C. (1980). In vitro activity of chloram- phenicol and thiamphenicol
analogs. Antimicrob. Agents Chemother. 18: 311–316.
Olsen A.B., Melby H.P., Speilberg L., Evensen O. & Hastein T.(1997) Piscirickettsia
salmonis infection in Atlantic salmon Salmo salar in Norway – epidemiological,
pathological and microbiological findings. Diseases of Aquatic Organisms 31, 35–48.
Paulsen, I. T., M. H. Brown, and R. A. Skurray. (1996). Proton-dependent multidrug efflux
systems. Microbiol. Rev. 60: 575–608.

136
Poole K. (2005). Efflux-mediated antimicrobial resistance. Journal of Antimicrobial
Chemotherapy. 56, 20–51
Ridenhour, M. B., H. M. Fletcher, J. E. Mortensen, and L. Daneo-Moore. (1996). A novel
tetracycline-resistant determinant, tet(U), is encoded on the plasmid pKQ10 in
Enterococcus faecium. Plasmid 35: 71–80.
Roberts, M. C. (1996). Tetracycline resistant determinants: mechanisms of action,
regulation of expression, genetic mobility and distribution. FEMS Microbiol. Rev. 19: 1–
24.
Rojas MV, Olivares J, Del Río R, Marshall SH. 2007. Characterization of a novel and
genetically different small infective form of Piscirickettsia salmonis. Microb Pathog
44:370–378.
Rojas V., Olivares P., del Río R. Marshall S. (2008) Characterization of a novel and
genetically different small infective variant of Piscirickettsia salmonis. Microb.
Pathogenesis. 44: 370-378.
Rojas V., Galanti N., Bols N., Marshall S. (2009) Productive infection of Piscirickettsia
salmonis in macrophages and monocyte-like cells from rainbow trout, a possible survival
strategy. J. Cell. Biochem. 108: 631-637.

137
Salomon, S. A., Watts JL, Aarestrup, F. M . (1998). Minimum inhibitory concentrations for
selected antimicrobial agents organisms isolated from the mammary glands of dairy heifers
in New Zealand and Denmark. Journal Dairy Science. 81: 570 - 578.
San Martín B., Tadaishi Y., Gallardo A. (2010). Manual de buenas prácticas en el uso de
antibióticos y antiparasitarios en la salmonicultura chilena. Servicio Nacional de Pesca.
Ministerio de Economía, Fomento y Turismo de Chile.
Sanchez-Pescador, R., J. T. Brown, M. Roberts, M. S. Urdea. (1988). Homology of the
TetM with translational elongation factors: implications for potential modes of tetM
conferred tetracycline resistance. Nucleic. Acids Res. 16: 1218.
Schäfer J.W., Alvarado V., Enriquez R. & Monra´s M. (1990) The ‘‘coho salmon syndrome
(CSS): a new disease in Chilean salmon, reared in sea water. Bulletin of the European
Association of Fish Pathologists 10, 130.
SERNAPEASCA. 2011. Informe sanitario de la acuicultura, período enero-septiembre
2011. Gobierno de Chile. (Servicio Nacional de Pesca, www.sernapesca.cl)
SERNAPESCA. 2012. Informe sanitario de salmonicultura en centros marinos año 2012.
Gobierno de Chile. (Servicio Nacional de Pesca, www.sernapesca.cl)
Shaw, W.V. (1983). Chloramphenicol acetyltransferase, enzymology and molecular
biology. Crit. Rev. Biochem. 14: 1–46.

138
Sheridan, R. P., and I. Chopra. (1991). Origin of tetracycline efflux proteins: conclusions
from nucleotide sequence analysis. Mol. Microbiol. 5: 895–900.
Shu-Peng HO., Tain-Yao HSU., Ming-Hui CHEN and Way-Shyan WANG. (2000).
Antibacterial Effect of Chloramphenicol, Thiamphenicol and Florfenicol against Aquatic
Animal Bacteria. J. Vet. Med. Sci. 62(5): 479–485.
Skarmeta A.M., Henriquez V., Zahr M., Orrego C. & Marshall S.H. (2000) Isolation of a
virulent Piscirickettsia salmonis from the brain of naturally infected coho salmon. Bulletin
of the European Association of Fish Pathologists 20, 261–263.
Smith P.A., Pizarro P., Ojeda P., Contreras J., Oyanedel S. & Larenas J. (1999) Routes of
entry of Piscirickettsia salmonis in rainbow trout Oncorhynchus mykiss. Diseases of
Aquatic Organisms 37, 165–172.
Soto E., Endris R. G. & Hawke J. P. (2010). In Vitro and In Vivo Efficacy of Florfenicol for
Treatment of Francisella asiatica Infection in Tilapia. Antimicrob Agents Chemother. 54:
4664–4670.
Speer, B. S., L. Bedzyk, and A. A. Salyers. (1991). Evidence that a novel tetracycline
resistance gene found on two Bacteroides transposons encodes an NADP-requiring
oxidoreductase. J. Bacteriol. 173: 176–183

139
Summers, A.O. (2002). Generally overlooked fundamentals of bacterial genetics and
ecology. Clin. Infect. Dis. 34 (Suppl. 3):S85-S92.
Swartz, M.N. (2002) Human diseases caused by foodborne pathogens of animal origin.
Clin. Infect. Dis. 34 (Suppl. 3):S111-S122.
Taylor, D. E., and A. Chau. (1996). Tetracycline resistance mediated by
ribosomal protection. Antimicrob. Agents Chemother. 40: 1–5.
Toranzo A., Magariños B., Romalde J. (2005). A review of the main bacterial fish diseases
in mariculture systems. Aquaculture. 246: 37-61.
Úna M. McCarthy, James E. Bron, Linton Brown, Fazel Pourahmad, Ian R. Bricknell, Kim
D. Thompson, Alexandra Adams, Anthony E. Ellis. (2008) Survival and replication of
Piscirickettsia salmonis in rainbow trout head kidney macrophages. Fish & Shellfish
Immunology 25, 477–484.
Vera T., Isla A., Cuevas A., Figueroa J., (2012). Un nuevo medio de cultivo para el
patógeno Piscirickettsia salmonis. Arch. Med. Vet. In prenss.

140
Yáñez, A. J., Valenzuela, K., Matzner, C., Olavarría, V., Figueroa, J., Avendaño-Herrera,
R. and Carcamo, J. G. (2013), Broth microdilution protocol for minimum inhibitory
concentration (MIC) determinations of the intracellular salmonid pathogen Piscirickettsia
salmonis to florfenicol and oxytetracycline. Journal of Fish Diseases.
doi: 10.1111/jfd.12144
Recommended

















