
Experimental & Applied Acarology, 16 (1992) 331-344 Elsevier Science Publishers B.V., Amsterdam
ACARI 650
331
Cuticular hydrocarbons from Varroa jacobsoni
J.L. Nation, M.T. Sanford and K. Milne Department of Entomology & Nematology, University of Florida, Gainesville, FL, USA
ABSTRACT
Nation, J.L., Sanford, M.T. and Milne, K., 1992. Cuticular hydrocarbons from Varroajacobsoni. Exp. AppL Acarol., 16: 331-344.
The cuticular hydrocarbons from Varroa mites collected in Florida and Italy and their honeybee hosts were characterized by gas chromatography (GC) and by coupled GC-mass spectrometry (MS). A Varroa mite has a hydrocarbon pattern that is qualitatively identical to that of its honeybee host. Mites and all stages of honeybees share a characteristic pattern of prominant alkane components in- cluding heneicosane, tricosane, pentacosane, heptacosane, nonacosane and hentriacontane. Adult honey bees and mites from adult bees also have as major components the alkenes 8-nC31:1, 10-nC31:1 and 10-nC33:1, while pupae and mites from pupae have only traces of these alkenes. Coupled GC-MS analysis after addition of dimethyl disulfide (DMDS) was used to determine the location of the dou- ble bond in the three prominent alkenes above. The question as to whether the mites simpiy acquire the hydrocarbons from their hosts or possibly synthesize them was not resolved. No characteristic differences between the hydrocarbons of mites from Italy and from Florida were found. We speculate that the hydrocarbons help the mites avoid desiccation and may help integrate the mites into honey- bee colony life.
INTRODUCTION
The rapid introduction of the honeybee mite, Varroa jacobsoni Oudemans, to commercial honeybee, Apis mellifera L. colonies over most of Asia, Europe and North America, and concern about its economic impact upon apiculture, have led to numerous studies of its biology and control. The mites are rela- tively large, typically measuring 1.5-2.0 mm wide by 1.0-1.8 mm long (Den- mark et al., 1991 ). They suck body fluids from adults, larvae and pupae with stylet-like mouthparts. Fuchs (1990) found that drone brood cells contained about 8.3 times as many mites as worker brood cells. Le Conte et al. (1989)
Correspondence to: J.L. Nation, Dept. of Entomol. & Nematol., Bldg. 970, Hull Rd., Univ. of Florida, Gainesville, FL 32611-0620, USA. Florida Agricultural Experiment Station Journal Series No. R-02473.
0168-8162/93/$06.00 �9 1993 Elsevier Science Publishers B.V. All rights reserved.

332 J.L. NATION ET AL.
found that mites were attracted to extracts of honeybee larvae, and that the extracts contained methyl and ethyl esters of commonly occurring fatty acids. In a laboratory olfactometer the mites were attracted to several esters, but principally methyl palmitate. The exact role these esters may play, however, in honeybee and mite ecology are uncertain.
It is not known whether all the Varroa mites on honey bees constitute a single species. Griffiths et al. (1983 ) suggested that there may be more than a single species involved. Delfinado-Baker and Aggarwal ( 1987 ) believe that V. jacobsoni and V. underwoodi co-exist in Apis cerana Fabr. colonies in Ne- pal. Grobov et al. (1980) described morphological variations in mites taken from A. mellifera colonies in the USSR, Japan and West Germany.
The hydrocarbons on the cuticle of some insects are species specific and have proven useful in taxonomic and systematic studies. Carlson and Service (1980) and Carlson and Walsh (1981) were able to differentiate species complexes of mosquitoes and West African black flies, respectively, by gas chromatographic analyses of cuticle hydrocarbons. Carlson and Yocom (1986) presented evidence for species specificity in some tephritid fruit flies. Lockey ( 1981, 1982) found species specificity in the hydrocarbons from bee- tles in the family Tenebrionidae. Carlson (1988a) and Castner and Nation (1984, 1986) have similarly found species-specific hydrocarbons in cock- roaches and mole crickets, respectively. Blomquist et al. (1976) showed spe- cies specificity in gryllid crickets. Carlson and Bolten (1984), Lavine et al. ( 1988 ) and Brand et al. ( 1991 ) proposed methods for the identification of Africanized and European honeybees based upon hydrocarbon analyses. Ex- cept for our preliminary report (Nation et al., 1991 ), we have not found data on hydrocarbon analyses from mites, possibly because mites are often ex- tremely small, making chemical analyses more difficult.
We present here the results of gas chromatographic analyses of the hydro- carbons from the honeybee mite, V. jacobsoni, collected from several sites in Italy and from Florida, and show comparisons with the hydrocarbons ex- tracted from their hosts, A. mellifera.
MATERIALS AND METHODS
Mites and bees Mites were collected from colonies at the University of Florida, Gaines-
ville, FL. Mites from pupae were collected by opening individual brood cells and removing the pupae. Pupae selected were in the purple-eye stage of de- velopment. Bees and mites that definitely could be associated with each other were coded so that the GC analysis of a mite and its host could be correlated. Usually only one mite per pupa was found. Mites from adult bees were col- lected by shaking a frame of adult worker bees into a Kraft paper bag, freezing them, and then dumping them into a pan in the laboratory. Individual bees

CUTICULAR HYDROCARBONS FROM VARROA 333
were observed under a 10 • dissecting scope for the presence of mites. Some mites were found on the surface of the paper bag and on the pan, and could not be associated with any particular bee, but were nevertheless analyzed. Each mite was placed in a glass vessel made from the tip of a Pasteur pipet that was sealed in a flame, and cut to about 3 cm length. The mite was pushed with a needle to the sealed end, 10/~1 of pentane (HPLC grade) was added and the vessel was placed in a vial with a screw cap and stored at - 30~ until analysis. If necessary at the time of analysis, an additional 10 /d of pentane was added to cover the mite. Individual bees were kept frozen in scintillation vials, and about 2 ml solvent added at least 5 min prior to GC analysis. Sol- vent extracts from bees were passed through 0.5 g silicic acid (BioSil HA, minus 325 mesh; BioRad Corp.) in a Pasteur pipet to remove oxygenated compounds (Carlson and Bolten, 1984; Castner and Nation, 1986; Smith, 1990a). Hydrocarbons were eluted with the void volume of the column and an additional 1 ml of solvent was used to wash the column. Extracts of mites were not passed through the silicic acid column because of the extremely small volume available from each mite ( 10 #1 or less).
Mites from Italy were collected by beekeepers who had been instructed by one of us (MTS) to collect mites with clean forceps and to put each mite into a separate vial with a teflon-lined screw cap, which we had provided. Only mites were then mailed to Gainesville, FL. Mites from Brazil and Satsuma, FL, were collected by apiculturists under similar instructions. Mites were re- moved from shipment vials and placed into glass vessels prepared from Pas- teur pipets as described above and frozen until analysis.
Gas chromatography Samples and standards were analyzed on a 25-m• fused silica
capillary column with bonded polydimethylsiloxane coating (Alltech RSL- 150) in a Shimadzu G14-A gas chromatograph with capillary injector port and flame ionization detector (FID). Samples were injected in the splitless mode, with a purge flow after 30 s to remove solvent and sample components that had not entered the column. Initially, mites were analyzed with a tem- perature program suggested by Dr. David Carlson (pers. commun.) , which would allow detection of small as well as large hydrocarbons, as follows: The injector port temperature and the detector temperature were constant at 270 ~ and 320~ respectively. Helium was used as the carrier gas with a linear flow rate of 25 cm/min . Samples were injected in the splitless mode. The initial temperature of the column was 60 ~ and following injection the temperature was programmed to 240 ~ at 30 ~ C/min , and then programmed to 320 ~ at 10~ and held for a total run time of 45 min.
Small hydrocarbons (less than 20 carbons) were not detected, and a second temperature program was used to facilitate calculation of Kovats Index val- ues (Kovats, 1965 ) for sample components. Injector port and detector tem-

334 J,L. NATION ET AL.
peratures were the same as above, but the column (same as above) was ini- tially at 200~ and the temperature was programmed to rise at 4~ to 320~ and to hold for the duration of the run. The injection was splitless. The linear flow rate of helium carrier gas through the column was 25 cm/s. Data collection occurred with a Spectra Physics 4100 Integrator or with the Spectra Physics WINner Integrator. Representative samples from mites and bees were co-chromatographed with a standard series of n-alkanes (Alltech Associates) for comparison of retention times. Mass spectral analysis of rep- resentative samples and of derivatives of major alkenes were performed on a Perkin Elmer 8420 GC interfaced with a Finnigan Ion Trap Detector (ITD, Model 6210), with INCOS data collection software and a 80286 computer. Spectra were compared with the NBS mass spectral library of about 44 000 spectra. Dimethyl-disulfide addition was used to determine the location of double bonds in key alkenes in extracts from honey bees and Varroa mites (Dunkelblum et al., 1985; Carlson et al., 1989).
Stat&tical analys& The means from the ratio n-alkane/monomethyl branched alkane were
analyzed by one-way analysis of variance (ANOVA) and the means were sep- arated by calculating Fisher's least significant difference.
RESULTS
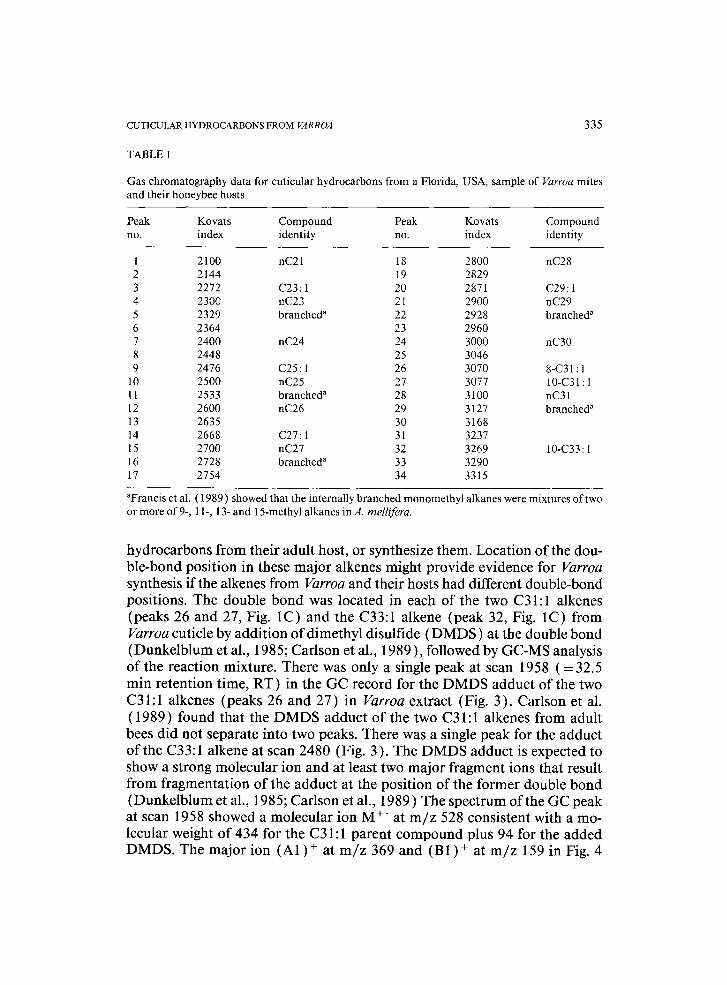
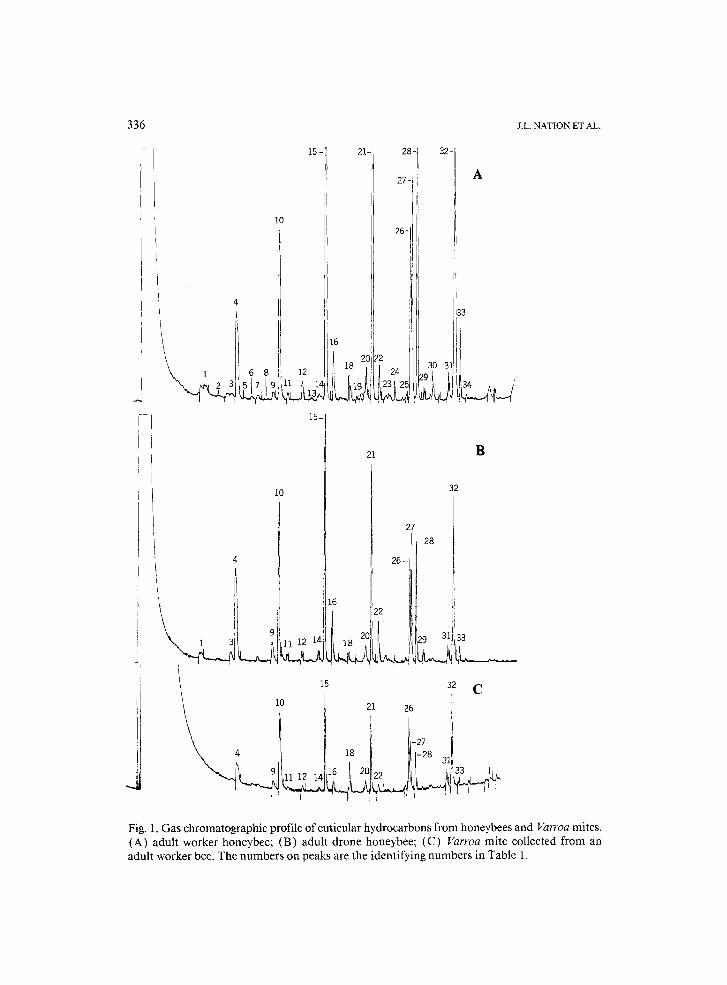
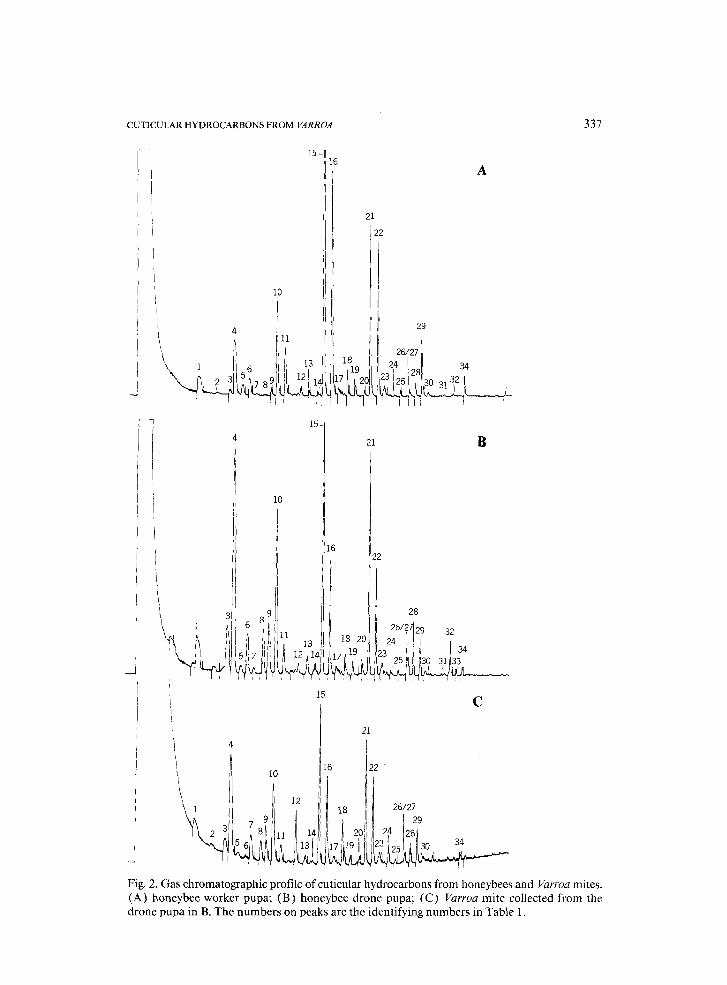
It was possible to analyze the hydrocarbons from single Varroa mites. Fig- ures 1 and 2 show representative gas chromatographic (GC) records from mites and bees, and Table 1 correlates peak numbers from the GC records with the calculated Kovats Index (KI) and compound identity when known.
The major hydrocarbons on the surface of Varroa mites taken from adult worker bees were n-alkanes with 23, 25, 27, 29 and 31 carbons, two unsatu- rated C31:1 hydrocarbons (peaks 26, 27 in Fig. 1 C) and an unsaturated C33:1 compound (peak 32 in Fig. 1C), and these were the same hydrocarbons on the cuticles of adult worker bees (Table 1 and Figs. 1A, B and 2A, B). Our results agreed with reports on bees from a number of workers (McDaniel et al., 1984; Francis et al., 1985, 1989; Carlson, 1988b; Smith, 1990b, 1991 ).
The most characteristic difference between mites taken from pupae and mites taken from adult bees was in the quantity of C31:1 and C33:1 alkenes. Varroa taken from the purple-eye stage of drone pupae showed only traces of these alkenes (peaks 26, 27 and 32, Fig. 2C), while those taken from adult bees had a large quantity relative to other hydrocarbons. Significantly, only traces of these unsaturated hydrocarbons were found in cuticular extracts of both worker and drone pupae (Fig. 2A, B), on which immature Varroa de- velop, although we found no Varroa mites on worker pupae in this study. When Varroa mites leave the brood cell, they either acquire these unsaturated
CUTICULAR HYDROCARBONS FROM VARROA
TABLE 1
Gas chromatography data for cuticular hydrocarbons from a Florida, and their honeybee hosts
335
USA, sample of Varroa mites
Peak Kovats Compound Peak Kovats Compound no. index identity no. index identity
1 2100 nC21 18 2800 nC28 2 2144 19 2829 3 2272 C23:1 20 2871 C29:1 4 2300 nC23 21 2900 nC29 5 2329 branched a 22 2928 branched" 6 2364 23 2960 7 2400 nC24 24 3000 nC30 8 2448 25 3046 9 2476 C25:1 26 3070 8-C31:1
10 2500 nC25 27 3077 10-C31:1 11 2533 branched a 28 3100 nC31 12 2600 nC26 29 3127 branched a 13 2635 30 3168 14 2668 C27:1 31 3237 15 2700 nC27 32 3269 10-C33:1 16 2728 branched" 33 3290 17 2754 34 3315
"Francis et al. ( 1989 ) showed that the internally branched monomethyl alkanes were mixtures of two or more of 9-, 11-, 13- and 15-methyl alkanes in A. mellifera.
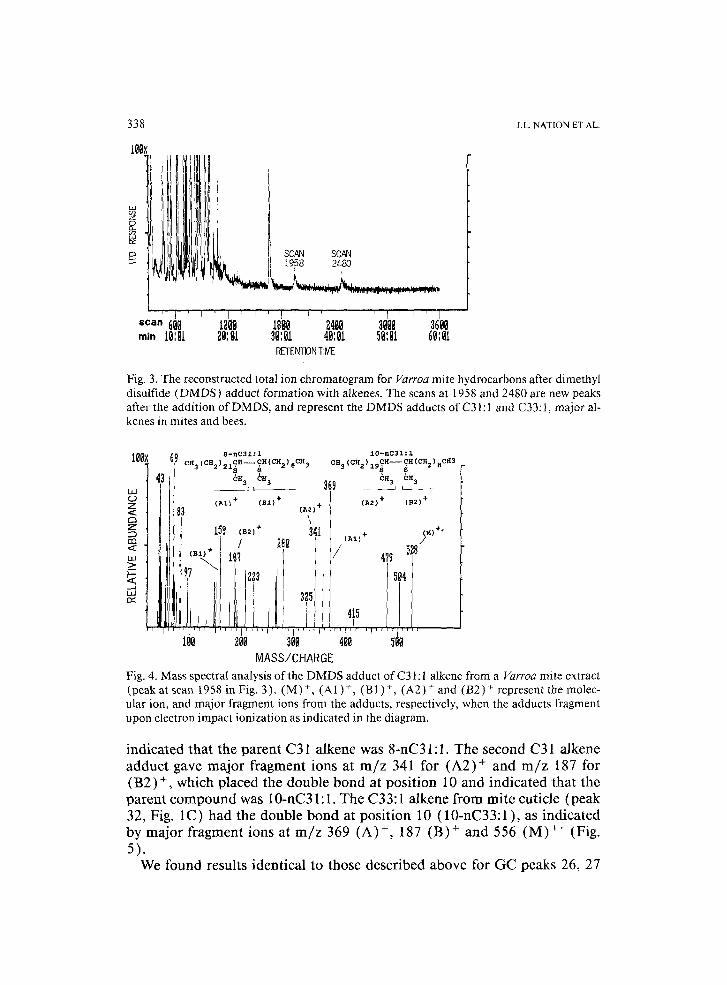
hydrocarbons from their adult host, or synthesize them. Location of the dou- ble-bond position in these major alkenes might provide evidence for Varroa synthesis if the alkenes from Varroa and their hosts had different double-bond positions. The double bond was located in each of the two C31:1 alkenes (peaks 26 and 27, Fig. 1C) and the C33:1 alkene (peak 32, Fig. 1C) from Varroa cuticle by addition of dimethyl disulfide (DMDS) at the double bond (Dunkelblum et al., 1985; Carlson et al., 1989 ), followed by GC-MS analysis of the reaction mixture. There was only a single peak at scan 1958 ( = 32.5 min retention time, RT) in the GC record for the DMDS adduct of the two C31:1 alkenes (peaks 26 and 27) in Varroa extract (Fig. 3). Carlson et al. (1989) found that the DMDS adduct of the two C31:1 alkenes from adult bees did not separate into two peaks. There was a single peak for the adduct of the C33:1 alkene at scan 2480 (Fig. 3). The DMDS adduct is expected to show a strong molecular ion and at least two major fragment ions that result from fragmentation of the adduct at the position of the former double bond (Dunkelblum et al., 1985; Carlson et al., 1989) The spectrum of the GC peak at scan 1958 showed a molecular ion M § at m / z 528 consistent with a mo- lecular weight of 434 for the C31:1 parent compound plus 94 for the added DMDS. The major ion (A1) § at m / z 369 and (BI) + at m / z 159 in Fig. 4
336
1
I
J.L. NATION ET AL.
15
10
20i
15-
21
10
2~ 32-
27" A
26-
11
i33
!4 ,293~ 3 ' 1 1 3 4
27 26_i 128
B
32
Ii I[li I 22 'IL ? 1121~ 18 20 9 3~ 33
15, s2 C 10 21 26
i i
Fig. 1. Gas chromatographic profile of cuticular hydrocarbons from honeybees and Varroa mites. (A) adult worker honeybee; (B) adult drone honeybee; (C) Varroa mite collected from an adult worker bee. The numbers on peaks are the identifying numbers in Table 1.
CUTICULAR HYDROCARBONS FROM VARROA 337
10
4
15-
,11
�9 ~ 15-
10
I6
16
A
21 22
26/27 1 2324 28 34
21 B
15
22
28 26/27~29 32
24 ' 34 23 5
C
21
I /
10 ~ 122
Fig. 2. Gas chromatographic profile of cuticular hydrocarbons from honeybees and Varroa mites. (A) honeybee worker pupa; (B) honeybee drone pupa; (C) Varroa mite collected from the drone pupa in B. The numbers on peaks are the identifying numbers in Table 1.
338 J.L. N A T I O N ET AL.
Ill
' l I scan ~l~
,•.)t SCAN SCAN
' I I 1 ' I '1 I ' I I' ' " i - " - - - ~200 18~0 24~B 30~ 3600
RETENTION TIME
Fig. 3. The reconstructed total ion chromatogram for Varroa mite hydrocarbons after dimethyl disulfide (DMDS) adduct formation with alkenes. The scans at 1958 and 2480 are new peaks after the addition of DMDS, and represent the DMDS adducts of C31:l and C33:1, major al- kenes in mites and bees.
100x 69
klA C3 Z < ~3 Z
a3 <
>__
8-nC31~1 10-nc31:1 CR (CH) CH~CH(CH-)6CH 3 CH3(CH2) CH--CH(CH )_CH3
~ ~3 369 ~H3 cx3
I (~I}+ (B~)+ (A2) + I (A2)+ (B2)+ ' \
. 34I [ 5 9 (e21 + ( (~, +"
t t 28~ I I ~'"+ /
115
t~O 200 30~ 488 500 MASS/CHARGE
Fig. 4. Mass spectral analysis of the DMDS adduct of C31:1 alkene from a Varroa mite extract (peak at scan 1958 in Fig, 3). (M) +, (A1)+, (BI)+, (A2) + and (B2) + represent the molec- ular ion, and major fragment ions from the adducts, respectively, when the adducts fragment upon electron impact ionization as indicated in the diagram.
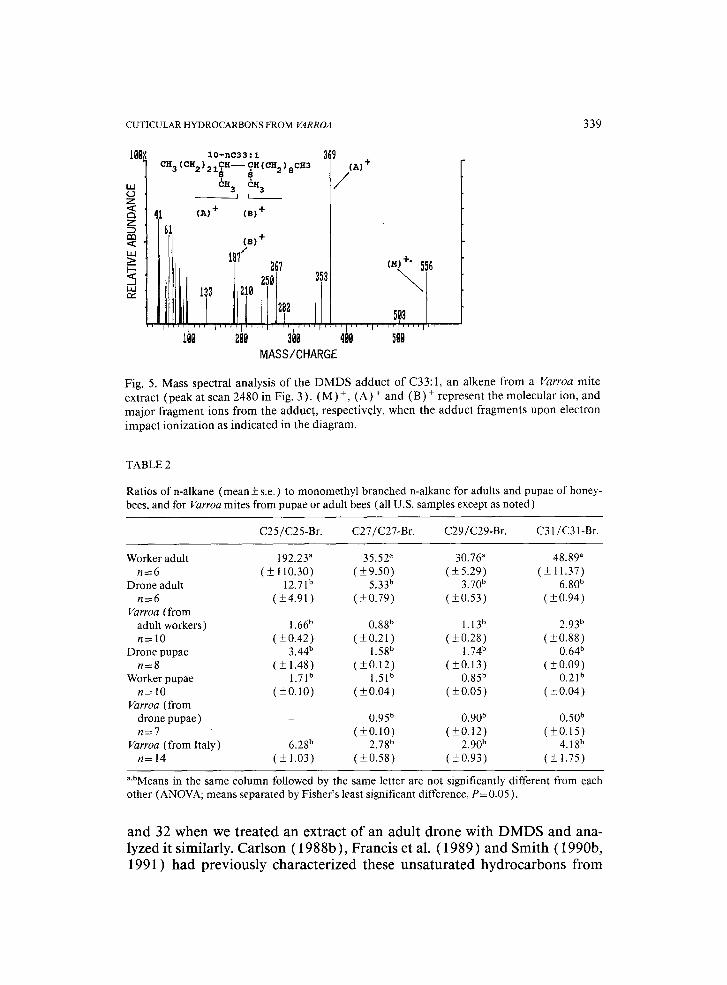
indicated that the parent C31 alkene was 8-nC31:1. The second C31 alkene adduct gave major fragment ions at m / z 341 for (A2) + and m / z 187 for (B2) +, which placed the double bond at position 10 and indicated that the parent compound was 10-nC31:1. The C33:1 alkene from mite cuticle (peak 32, Fig. 1C) had the double bond at position 10 (10-nC33:1 ), as indicated by major fragment ions at m / z 369 (A) +, 187 (B) + and 556 (M) + (Fig. 5).
We found results identical to those described above for GC peaks 26, 27
C U T I C U L A R H Y D R O C A R B O N S F R O M VARROA 339
100:
Z < C; 7
< uA >_
..,A
l O - n c 3 3 : 1 CH~ (CH.)..CH-- CH(CH )
2 z Z.L~ , 8 cH3
6n 3 cn 3 J t _ _
4 1 (~)+ (m +
61
1 8 7 / 267
13 20 250 3~
? i, l, i . . . . j . . . . ~ . . . . I . . . . i . . . . I " ' I '
1BO 200 30B MASS/CHARGE
369 (AI
/
(M) +" 556
' 1 . . . . I . . . . I . . . . J ' '
400 500
Fig. 5. Mass spectral analysis of the DMDS adduct of C33:1, an alkene from a Varroa mite extract (peak at scan 2480 in Fig. 3). (M)+, (A) + and (B) + represent the molecular ion, and major fragment ions from the adduct, respectively, when the adduct fragments upon electron impact ionization as indicated in the diagram.
TABLE 2
Ratios of n-alkane (mean_+ s.e.) to monomethyl branched n-alkane for adults and pupae of honey- bees, and for Varroa mites from pupae or adult bees (all U.S. samples except as noted)
C25/C25-Br. C27/C27-Br. C29/C29-Br. C31/C31-Br.
Worker adult 192.23" 35.52" 30.76 a 48.89 a n = 6 (_+ 110.30) (_+9.50) (_+5.29) (_+ 11.37)
Drone adult 12.7 lb 5.33 b 3.70 b 6.80 b n = 6 ( _+ 4.91 ) (_+0.79) (_+0.53) (_+0.94)
Varroa (from adult workers) 1.66 b 0.88 b 1.13 b 2.93 b
n = 10 (_+0.42) ( _+ 0.21 ) ( _+ 0.28) ( _+ 0.88) Drone pupae 3.44 b 1.58 b 1.74 b 0.64 b
n=8 (_+1.48) (_+0.12) (_+0.13) (_+0.09) Worker pupae 1.71 b 1.51 b 0.85 b 0.21 b
n= lO (_+0.10) (_+0.04) (_+0.05) (_+0.04) Varroa (from
drone pupae) - 0 . 9 5 b 0 . 9 0 b 0 . 5 0 8
n = 7 (_+0.10) (_+0.12) (_+0.15) Varroa (from Italy) 6.28 b 2.78 b 2.90 b 4.18 b
n = 1 4 (_+1.03) (_+0.58) (_+0.93) (_+1.75)
a,bMeans in the same column followed by the same letter are not significantly different from each other (ANOVA; means separated by Fisher's least significant difference, P= 0.05 ).
a n d 32 w h e n w e t r e a t e d a n e x t r a c t o f a n a d u l t d r o n e w i t h D M D S a n d a n a -
l y z e d i t s i m i l a r l y . C a r l s o n ( 1 9 8 8 b ) , F r a n c i s e t a l . ( 1 9 8 9 ) a n d S m i t h ( 1 9 9 0 b ,
1991 ) h a d p r e v i o u s l y c h a r a c t e r i z e d t h e s e u n s a t u r a t e d h y d r o c a r b o n s f r o m

340 J.L. NATION ET AL.
adult honeybee cuticles as 8-nC31:1, 10-nC31:l and 10-nC33:1. Thus, our data indicate that the double bonds are located at the same places in these three alkenes from mites and honeybees.
Peaks 11, 16, 22 and 29 in Fig. 1A and 1B represent internally branched monomethyl alkanes. Francis et al. (1989) showed that these internally branched monomethyl alkanes were mixtures of two or more of the 9-, 11-, 13- and 15-methyl alkanes in A. mellifera. Mites from Florida and from Italy showed no statistically significant differences in the ratio of n-alkane/mono- methyl branched alkanes, nor were they significantly different from worker and drone pupae, or adult drones (Table 2). The ratios were significantly different, however, between adult workers and the pupal stages of either workers or drones, adult drones, or mites. Adults had much less of the methyl branched alkanes relative to n-alkane homologues than pupae (Figs. 1 and 2, and Table 2 ). Our results agreed with previously published data showing that the ratios were characteristically different in adult workers and drones (Carl- son, 1988b; Francis et al., 1989; Carlson et al., 1991 ).
DISCUSSION
Studies on the cuticular lipids of insects are now quite extensive as indi- cated by a recent review (Lockey, 1988 ). The cuticular "wax" on the surface of insects consists of a wide variety of lipid types, including, but not limited to, n-alkanes, n-alkenes, branched-chain alkanes, alcohols, fatty acids, sterols, ethers, esters, and sometimes acylglycerols. The cuticular lipids of insects serve a great variety of functions, including semiochemical communication (How- ard and Blomquist, 1982; Bartelt and Jackson, 1984; Peschke, 1987), possi- ble protective mimicry (Howard et al., 1980; Vander Meer and Wojcik, 1982; Vander Meer et al., 1989), and water proofing (Jackson and Baker, 1970; Nelson, 1978; Lockey, 1988 ). In particular, the hydrocarbon components on the cuticle of honeybees have received extensive treatment (Carlson and Bol- ten, 1984; McDaniel et al., 1984; Carlson, 1988b; Francis et al., 1989; Smith, 1990b, 1991 ).
We found no evidence that the cuticular hydrocarbons of mites collected from Italy were different from those of mites collected in Florida. Our data show that the hydrocarbons found on the surface of Varroa mites from Flor- ida are qualitatively the same as those of the pupal hosts on which they de- velop. There are differences, however, between mites and adult workers in proportions of some of the hydrocarbons, especially in the amount of mono- methyl branched n-alkanes. Even though we found adult mites on worker adults, they never had the very large ratios for n-alkane/branched alkane that were characteristic of adult worker bees. This difference between mites and worker adults suggests that mites may have their own characteristic hydrocar- bon pattern and, if true, could indicate mite biosynthesis of hydrocarbons.

CUTICULAR HYDROCARBONS FROM VARROA 341
We have not studied the origin of Varroa hydrocarbons. The mites may acquire them by feeding upon the pupae on which they develop, by physical contact with bees or the comb wax (Blomquist et al., 1980), or possibly by synthesis. If Varroa mites acquire the hydrocarbons from their hosts rather than synthesizing them, then mites that have transferred to adult workers for some time should begin to show the very large ratios in the proportion of internally branched hydrocarbons relative to the n-alkanes of C25, C27, C29 and C31 characteristic of adult workers. A test of whether they synthesize the hydrocarbons or acquire them from their host may be possible by confining a number of mites on a population of worker adults that contain no drones for a known period of time, with interval sampling to monitor changes in the straight-chain/methyl branched ratio for C25, C27, C29 and C31 hydrocarbons.
If mites can be reared on a synthetic or artificial food, then a direct test of their ability to synthesize hydrocarbons should be possible by using radio- labeled lipid precursors. Possibly even the presence of hydrocarbons on their cuticles when reared artificially would be evidence of synthesis. Regardless of how the mites acquire their hydrocarbons, it seems probable that these lipids offer protection from desiccation. One of the functions of hydrocarbons on the cuticle of insects is to provide protection from excessive water loss.
Specific mechanisms have been identified that provide some resistance to Varroa for various species and races of honeybees. Our work may be particu- larly relevant to that ofPeng et al. ( 1987 ), who showed that grooming behav- iour ofA. cerana worker bees caused them to remove mites from each other's bodies and from introduced brood ofA. mellifera workers. Given the cuticu- lar hydrocarbon differences among Africanized and European A. mellifera andA. cerana (Francis et al., 1985; Carlson, 1988b), it is possible that mites can be recognized based upon hydrocarbon patterns. Analyses of the cuticular hydrocarbons from A. cerana and of mites from A. cerana may prove interesting.
Vander Meer and Wojcik (1982) and Vander Meer et al. (1989) described the relationship of a commensal beetle and of an hymenopteran parasitoid, respectively, that live in fire ant colonies, despite the fact that the ants are notably fierce defenders of their nests and predators of other insects. Though there is no direct experimental evidence to explain the mechanism (s) of col- ony integration, both the beetle and the hymenopteran showed the same hy- drocarbon pattern and/or volatiles pattern of the ants.
Varroa hydrocarbon patterns are qualitatively identical to specific life- stages of European honeybees. The bees may have difficulty detecting the mites as foreign invaders because of the hydrocarbon pattern presented by the mites. The question of whether the mites simply acquire their host hydrocarbons or have their own biosynthetic capability takes on more significance.

342 J.L. NATION ET AL.
ACKNOWLEDGEMENTS
We thank NSF for Instrument Award # 8806823 and the Institute of Food and Agricultural Sciences (IFAS), University of Florida, for funds to pur- chase the GC-MS system. We are grateful to Italian beekeepers and apicultur- ists for collecting samples for us in Italy. We thank Jane Medley for assistance in preparing some of the figures, and Dr. David Carlson and Dr. Robert Van- der Meer, USDA/ARS, Gainesville, FL and Dr. Harvey Cromroy, Univ. of Florida, Gainesville, FL for helpful comments and criticism on early drafts.
REFERENCES
Bartelt, R.J. and Jackson, L.L., 1984. Hydrocarbon component of the Drosophila virilis (Dip- tera: Drosophilidae ) aggregation pheromone: (Z)- 10-heneicosene. Ann. Entomol. Soc. Am., 77:364-371.
Blomquist, G.J., Blailock, T.T. Scheetz, R.W. and Jackson, L.L., 1976. Cuticular lipids of in- sects - VII. Cuticular hydrocarbons of the crickets Acheta domesticus, Gryllus pen nsylvanicus and Nemobiusfasciatus. Comp. Biochem. Physiol., 54B: 381-386.
Blomquist, G.J., Chu, A.J. and Remaley, S., 1980. Biosynthesis of wax in the honeybee, Apis mellifera L. Insect Biochem., 10: 313-321.
Brand, H.M., Puleo, S.L. and Brand, E.E., 1991. Identification of the Africanized bee in South America by the composition of its wax. Bee Sci., 1:106-111.
Carlson, D.A., 1988a. Hydrocarbons for identification and phenetic comparisons: cockroaches, honey bees and tsetse flies. Fla. Entomol., 71: 333-345.
Carlson, D.A., 1988b. Africanized and European honey-bee drones and comb waxes: analysis of hydrocarbon components for identification. In: G.R. Needham, R.F. Page Jr., M. Delfi- nado-Baker and C.E. Bowman (Editors), Africanised Honey Bees and Bee Mites. Ellis Hor- wood Ltd./John Wiley, New York, NY, pp. 264-274.
Carlson, D.A. and Bolten, A.B., 1984. Identification of Africanized and European honey bees, using extracted hydrocarbons. Bull. Entomol. Soc. Am., 30: 32-35.
Carlson, D.A. and Service, M.W., 1980. Identification of mosquitoes of Anopheles gambiae species complex A and B by analysis of cuticular components. Science, 207:1089-1091.
Carlson, D.A. and Walsh, J.F., 1981. Identification of two West African.black flies (Diptera: Simuliidae) of the Simulium damnosum species complex by analysis of cuticular paraffins. Acta Tropica, 38: 235-239.
Carlson, D.A. and Yocom, S.R., 1986. Cuticular hydrocarbons from six species oftephritid fruit flies. Arch. Insect Biochem. Physiol., 3: 397-412.
Carlson, D.A., Roan, C-S., Yost, R.A. and Hector, J., 1989. Dimethyl disulfide derivatives of long chain alkenes, alkadienes, and alkatrienes for gas chromatography/mass spectrometry. Anal. Chem., 61 1564-1571.
Carlson, D.A., Roubik, D.W. and Milstrey, K., 1991. Distinctive hydrocarbons among giant honey bees, the Apis dorsata group (Hymenoptera: Apidae). Apidologie, 22:169-181.
Castner, J.L. and Nation, J.L., 1984. Cuticular lipids for species recognition of mole crickets (Orthoptera: Gryllotalpidae): I. Scapteriscus didactylus, Scapteriscus imitatus and Scapter- iscus vicinus. Fla. Entomol., 67:155-160.
Castner, J.L. and Nation, J.L., 1986. Cuticular lipids for species recognition of mole crickets (Orthoptera: Gryllotalpidae): II. Scapteriscus abbreviatus, S. acletus, S. vicinus, S. sp., and Neocurtilla hexadactyla. Arch. Insect Biochem. Physiol., 3: 127-134.

CUTICULAR HYDROCARBONS FROM VARROA 343
Delfinado-Baker, M. and Aggarwal, K., 1987. A new Varroa (Acari: Varroidae) from the nest ofApis cerana (Apidae). Int. J. Acarol., 13: 233-237.
Denmark, H.A., Cromroy, H.L. and Cutts, L., 1991. Varroa mite, Varroajacobsoni Oudemans (Acari: Varroidae). Entomol. Circ. No. 347, Fla. Dept. Agric. & Consumer Serv., Div. of Plant Industry, Gainesville, FL, 4 pp.
Dunkelblum, E., Tan, S.H. and Silk, P.J., 1985. Double-bond location in monounsaturated fatty acids by dimethyl disulfide derivatization and mass spectrometry: Application to analysis of fatty acids in pheromone glands of four Lepidoptera. J. Chem. Ecol., 11: 265-277.
Francis, B.R., Blanton, W.E. and Nunamaker, R.A., 1985. Extractable surface hydrocarbons of workers and drones of the genus Apis. J. Apic. Res., 24:13-26.
Francis, B.R., Blanton, W.E., Littlefield, J.L. and Nunamaker, R.A., 1989. Hydrocarbons of the cuticle and hemolymph of the adult honey bee (Hymenoptera: Apidae). Ann. Entomol. Soc. Am., 82: 486-494.
Fuchs, S., 1990. Preference for drone brood cells by Varroa jacobsoni Oud in colonies of Apis mellifera carnica. Apidologie, 21: 193-199.
Griffiths, D.A., Gray, J. and Pegazzano, F., 1983. Varroa - The acarologists' view. In: R. Cav- alloro (Editor), Varroajacobsoni Oud. affecting Honey Bees: Present Status and Needs. Proc. Meet. Ec Experts' Group, Wageningen. A.A. Balkema, Rotterdam, pp. 79-83.
Grobov, O.F., Poulenetz, N.M. and Sofronova, S.L., 1980. La variabilit6 g6ographique des di- mensions du scutum dorsal des femelles de Varroa jacobsoni Oudemans. In: Le XXVIIe Congr6s Int. Apic., Athene, 1979. Apimondia, Bucharest, pp. 381-382.
Howard, R.W. and Blomquist, G.J., 1982. Chemical ecology and bio-chemistry of insect hydro- carbons. Ann. Rev. Entomol., 27: 149-172.
Howard, R.W., McDaniel, C.A. and Blomquist, G.J., 1980. Chemical mimicry as an integrating mechanism: cuticular hydrocarbons of a termitophile and its host. Science, 210:431-433.
Jackson, L.T. and Baker, G.L., 1970. Cuticular lipids of insects. Lipids, 5: 239-246. Kovats, E., 1965. Gas chromatographic characterization of organic substances in the retention
index system. Adv. Chromatogr., 1: 229-247. Lavine, B.K., Carlson, D.A., Henry, D. and Jurs, P.C., 1988. Taxonomy based on chemical
constitution: differentiation of Africanized honey-bees from European honey-bees. J. Che- mometrics, 2: 29-37.
Le Conte, Y., Arnold, G., Trouiller, J., Masson, C., Chappe, B. and Ourisson, G., 1989. Attrac- tion of the parasitic mite Varroa to the drone larvae of honey bees by simple aliphatic esters. Science, 245: 638-639.
Lockey, K.H., 1981. Cuticular hydrocarbons of adult Cylindrinotus laevioctostriatus (Goetze) and Phylan gibbus (Fabricius) (Coleoptera: Tenebrionidae). Insect Biochem., 11:549-561.
Lockey, K.H., 1982. Hydrocarbons of adult Onymacris plana (Peringuey) and Onymacris ru- gatipennis (Haag) (Coleoptera: Tenebrionidae). Insect Biochem., 12: 69-81.
Lockey, K.H., 1988. Lipids of the insect cuticle: origin, composition and function. Comp. Biochem. Physiol., 89B: 595-645.
McDaniel, C.A., Howard, R.W., Blomquist, G.J. and Collins, A.M., 1984. Hydrocarbons of the cuticle, sting apparatus and sting shaft of Apis mellifera L. Identification and preliminary evaluation as chemotaxonomic characters. Sociobiology, 8:287-298.
Nation, J.L., Sanford, M.T. and Milne, K., 1991. Comparison of cuticular hydrocarbons from Varroa mites and honey bees. Am. Bee J., 131: 778-779.
Nelson, D.R., 1978. Long-chain methyl-branched hydrocarbons: Occurrence, biosynthesis, and function. Adv. Insect Physiol., 13: 1-33.
Peng, Y.S., Fang, Y., Xu, S., Ge, L. and Nasr, M.E., 1987. Response of foster Asian honey bee (Apis cerana) colonies to the brood of European honey bees (Apis mellifera) infested with parasitic mite, Varroajacobsoni. J. Invert. Pathol., 49: 259-264.

344 J.L. NATION ET AL.
Peschke, K., 1987. Cuticular hydrocarbons regulate mate recognition, male aggression, and fe- male choice of the rove beetle, Aleochara curtula. J. Chem. Ecol., 13:1993-2008.
Smith, R-K., 1990a. Principles of chemical analysis of bee hydrocarbons. Bee Sci., 1:15-17. Smith, R-K., 1990b. Chemotaxonomy of honey bees (Apis mellifera L. ). Part 1: European and
African workers. Bee Sci., 1: 23-32. Smith, R-K., 1991. Chemotaxonomy of honey bees (Apis mellifera L.). Part 2: Africanized
workers. Bee Sci., 1: 82-94. Vander Meer, R.K and Wojcik, D.P., 1982. Chemical mimicry in the myrmecophilous beetle
Myrmecaphodius excavaticollis. Science, 18: 806-808. Vander Meer, R.K., Jouvenaz, D.P. and Wojcik, D.P., 1989. Chemical mimicry in a parasitoid
(Hymenoptera: Eucharitidae) of fire ants (Hymenoptera: Formicidae). J. Chem. Ecol., 15: 2247-2261.
Recommended

















