
Cyanopeptoline und Scytocyclamide: Zyklische
Peptide aus Scytonema hofmanni PCC 7110
Struktur und biologische Aktivität
Inaugural-Dissertation zur Erlangung der Doktorwürde
der Fakultät für Biologie der Albert-Ludwigs-Universität
Freiburg im Breisgau
vorgelegt von Apotheker Jan Christoph Grewe
aus Landau i. d. Pfalz
2005

Dekan: Prof. Dr. G. Fuchs
Leiter der Arbeit: Prof. Dr. J. Weckesser
Referent: Prof. Dr. J. Weckesser
Koreferent: PD Dr. A. Liszkay
Tag der Verkündung des Prüfungsergebnisses: 22.12.2005

I
Inhaltsverzeichnis Inhaltsverzeichnis........................................................................................................................I Chemikalien und Materialien ................................................................................................... IV Geräte .......................................................................................................................................VI Abkürzungen ...........................................................................................................................VII Zusammenfassung.................................................................................................................... IX 1. Einleitung ............................................................................................................................... 1
1.1. Cyanobakterien und sekundäre Stoffwechselprodukte ................................................... 1 1.2. Cyanobakterien................................................................................................................ 1 1.3. Symbiose ......................................................................................................................... 4 1.4. Sekundärmetabolismus.................................................................................................... 5 1.5. Ökologie der Sekundärstoffwechselprodukte ................................................................. 5 1.6 Biosynthese der Sekundärstoffwechselprodukte.............................................................. 6 1.7. Biologisch aktive Substanzen aus Cyanobakterien......................................................... 7
1.7.1 Cyanobakterielle Toxine ........................................................................................... 7 1.7.1.1. Hepatotoxine ..................................................................................................... 8 1.7.1.2. Neurotoxine....................................................................................................... 9 1.7.1.3. Dermatotoxine und hautreizende Toxine ........................................................ 10
1.7.2.Cyanobakterien als Quelle neuer Arzneistoffe........................................................ 11 1.7.2.1. Antimikrobielle Substanzen ............................................................................ 11 1.7.2.2. Zytostatika....................................................................................................... 13 1.7.2.3. Enzyminhibitoren ............................................................................................ 14 1.7.2.4. Weitere Substanzklassen ................................................................................. 16
2. Material und Methoden ........................................................................................................ 22 2.1. Kultivierung von Scytonema hofmanni PCC 7110....................................................... 22
2.1.1. Anzucht und Kulturmedium................................................................................... 22 2.1.2. Ernte der Kulturen und Lagerung........................................................................... 23
2.2. Peptid-Isolierung aus Scytonema hofmanni PCC 7110................................................. 23 2.2.1. Extraktion ............................................................................................................... 23 2.2.2. Festphasenchromatographie ................................................................................... 23
2.3. Hochleistungsflüssigchromatographie .......................................................................... 24 2.3.1. Analytische Umkehrphasenchromatographie ........................................................ 24 2.3.2. RP-HPLC im semipräparativen und präparativen Maßstab................................... 25
2.4. Identifizierung der Peptide und Reinheit ...................................................................... 27 2.4.1. Co-Chromatographie (Co-Elution) ........................................................................ 27
2.5. Strukturaufklärung der isolierten Substanzen ............................................................... 28 2.5.1. Ultraviolett-Spektroskopie ..................................................................................... 28 2.5.2. Infrarotspektroskopie ............................................................................................. 28 2.5.3. Massenspektroskopie ............................................................................................. 29 2.5.4. Elektrospray-Ionisations-Massenspektrometrie ..................................................... 29 2.5.5. Magnetische Kernresonanzspektroskopie .............................................................. 30 2.5.6. Gaschromatographie-Massenspektrometrie-Messungen ....................................... 30 2.5.7. Konfigurationbestimmung der Aminosäuren......................................................... 30
2.6. Enzymhemmtests .......................................................................................................... 31 2.6.1. Trypsin ................................................................................................................... 31 2.6.2. α–Chymotrypsin..................................................................................................... 31 2.6.3. Plasmin ................................................................................................................... 32 2.6.4. Papain ..................................................................................................................... 32 2.6.5. Elastase Typ IV ...................................................................................................... 32

II
2.6.6. Thrombin................................................................................................................ 32 2.6.7. Glutathion-S-Transferase ....................................................................................... 33 2.6.8. Carboxypeptidase A ............................................................................................... 33 2.6.9. Leucin-Aminopeptidase ......................................................................................... 33 2.6.10. Thermolysin ......................................................................................................... 33 2.6.11. Proteinphosphatase PP1A .................................................................................... 34
2.7. Testung auf antimikrobielle Aktivität ........................................................................... 34 2.8. Hämolysetest ................................................................................................................. 35 2.9. Toxizitätstest mit Kleinkrebsen (Crustaceen) ............................................................... 35 2.10. Zytotoxizitätsversuche ................................................................................................ 36
2.10.1. Zelllinien .............................................................................................................. 36 2.10.2. Trypanblaufärbung ............................................................................................... 36
2.11. Versuche zur Membranschädigung............................................................................. 37 2.11.1. Testung auf porenbildende Aktivität.................................................................... 37 2.11.2. 86Rubidium+-Efflux .............................................................................................. 37
2.12. Immunhistochemische Untersuchungen ..................................................................... 38 2.13. Testung auf Apoptose.................................................................................................. 38
2.13.1. Fluoreszenz-aktivierte Zell-Sortierung (FACS-Analysen) .................................. 38 2.13.1.1. Apoptose-Färbung mit Annexin V-FITC und Propidiumiodid..................... 39 2.13.1.2. Mitochondriales Membranpotential .............................................................. 39 2.13.1.3. Detektion reaktiver Sauerstoffspezies ........................................................... 39
3. Ergebnisse ............................................................................................................................ 40 3.1. Charakterisierung der Cyanopeptoline und Scytocyclamide ........................................ 40
3.1.1. Extraktion und Isolierung....................................................................................... 40 3.1.2. Strukturaufklärung von Cyanopeptolin 1060A ...................................................... 43
3.1.2.1. Ultraviolett-Spektroskopie .............................................................................. 43 3.1.2.2. Infrarotspektroskopie ...................................................................................... 43 3.1.2.3. Massenspektrometrie....................................................................................... 44 3.1.2.4. Magnetische Kernresonanzspektroskopie ....................................................... 45
3.1.3. Strukturaufklärung von Cyanopeptolin 1067A ...................................................... 49 3.1.3.1. Ultraviolett-Spektroskopie .............................................................................. 49 3.1.3.2. Infrarotspektroskopie ...................................................................................... 49 3.1.3.3. Massenspektrometrie....................................................................................... 50 3.1.3.4. Magnetische Kernresonanzspektroskopie ....................................................... 51
3.1.4. Strukturaufklärung von Cyanopeptolin 946A ........................................................ 53 3.1.4.1. Ultraviolett-Spektroskopie .............................................................................. 53 3.1.4.2. Massenspektrometrie....................................................................................... 53 3.1.4.3. Magnetische Kernresonanzspektroskopie ....................................................... 55
3.1.5. Strukturaufklärung von Scytocyclamid A.............................................................. 57 3.1.5.1. Ultraviolett-Spektroskopie .............................................................................. 57 3.1.5.2. Infrarotspektroskopie ...................................................................................... 58 3.1.5.3. Massenspektrometrie Teil I ............................................................................. 59 3.1.5.4. Magnetische Kernresonanzspektroskopie ....................................................... 59 3.1.5.5. Massenspektrometrie Teil II............................................................................ 62 3.1.5.6. Bestimmung der Aminosäurekonfiguration .................................................... 64
3.1.6. Strukturaufklärung von Scytocyclamid B .............................................................. 65 3.1.6.1. Ultraviolett-Spektroskopie .............................................................................. 65 3.1.6.2. Infrarotspektroskopie ...................................................................................... 66 3.1.6.3. Massenspektrometrie....................................................................................... 67 3.1.6.4. Magnetische Kernresonanzspektroskopie ....................................................... 69
3.1.7. Struktur von Scytocyclamid C ............................................................................... 71

III
3.1.7.1. Infrarotspektroskopie ...................................................................................... 71 3.1.7.2. Massenspektrometrie....................................................................................... 72 3.1.7.3. NMR-Analytik zu Scytocyclamid C ............................................................... 73
3.2. Biologische Aktivität..................................................................................................... 75 3.2.1. Enzyminhibitions-Assays....................................................................................... 75 3.2.2. Antimikrobielle Untersuchungen ........................................................................... 77 3.2.3. Toxizitätsversuche mit Kleinkrebsen ..................................................................... 77 3.2.4. Zelltoxizitäts-Assays .............................................................................................. 78 3.2.5. Porenbildende Aktivität.......................................................................................... 80 3.2.6. Membranschädigung .............................................................................................. 81 3.2.7. Hämolysetest .......................................................................................................... 81 3.2.8. Apoptose-Assays.................................................................................................... 82 3.2.9. Immunhistochemische Untersuchungen ................................................................ 84
4. Diskussion ............................................................................................................................ 86 4.1. Isolierung und Strukturaufklärung ................................................................................ 86 4.2. Biologische Aktivität..................................................................................................... 92
4.2.1. Enzymhemmwirkung ............................................................................................. 92 4.2.2. Testung auf antimikrobielle Aktivität .................................................................... 93 4.2.3. Toxizitäts-Assays ................................................................................................... 94
4.2.3.1. Toxizitätstest mit Primärkonsumenten............................................................ 94 4.2.3.2. Testung auf Zytotoxizität ................................................................................ 94
4.3. Ökologie cyanobakterieller Sekundärstoffwechselprodukte......................................... 98 4.4. Therapeutische Relevanz peptidischer Sekundärmetabolite ....................................... 100 4.5. Ausblick ...................................................................................................................... 103
Literatur.................................................................................................................................. 105 Danksagung............................................................................................................................ 119

IV
Chemikalien und Materialien Acetonitril Carl Roth GmbH & Co KG, Karlsruhe Alexa 488 Anti Maus / Kaninchen IgG Molecular Probes, Heidelberg Alexa 594-Phalloidin Molecular Probes, Heidelberg Ammoniumeisen-(III)-citrat Merck KGaA, Darmstadt Ammoniumhydrogencarbonat Carl Roth GmbH & Co KG, Karlsruhe Anti-alpha-Tubulin III Antikörper Sigma-Aldrich, Taufkirchen APS Biorad München BAPNA Sigma-Aldrich GmbH (Fluka), Taufkirchen Borsäure Merck KGaA, Darmstadt BSA, Fraktion V, Lyophilisat Paesel und Lorei, Hanau BST Medium dehydrated Difco Laboratories GmbH, Heidelberg Trypticase Peptone Becton Dickinson AG, Basel C18-Kartuschen (1000 mg) Macherey-Nagel GmbH & Co KG, Düren Calciumchlorid • 2H2O Merck KGaA, Darmstadt Carboxypeptidase A Merck KGaA, Darmstadt Carboxypeptidase A Inhibitor (potato tuber) Sigma-Aldrich GmbH, Taufkirchen CCCP Sigma-Aldrich GmbH, Taufkirchen Chromabond C18 Material Macherey-Nagel GmbH & Co KG, Düren Chromozym PL Sigma-Aldrich GmbH, Taufkirchen Chymotrypsin Sigma-Aldrich GmbH, Taufkirchen Citronensäure Merck KGaA, Darmstadt Cobaltnitrat • 6 H2O Merck KGaA, Darmstadt Columbia Blood Agar Base Difco Laboratories GmbH, Heidelberg CSB-Puffer Pharmakologie-Freiburg Cystein Sigma-Aldrich GmbH, Taufkirchen Dikalium-hydrogenphosphat • 3H2O Merck KGaA, Darmstadt Dinatrium - EDTA Sigma-Aldrich GmbH, Taufkirchen Dinatriumhydrogenphosphat x 2 H2O p.a. Merck KGaA, Darmstadt DMSO Sigma-Aldrich GmbH (Fluka), Taufkirchen DTT Fluka, Taufkirchen EGTA Sigma-Aldrich GmbH, Taufkirchen Elastase Sigma-Aldrich GmbH, Taufkirchen EMEM Pharmakologie-Freiburg Essigsäure Carl Roth GmbH & Co KG, Karlsruhe Essigsäure p.a. Merck KGaA, Darmstadt Ethanol Carl Roth GmbH & Co KG, Karlsruhe FCS Biochrom AG, Berlin DiOC6(3) Molecular Probes, Heidelberg Fluoresceinisothiocyanat-Annexin V BD Pharmingen, Heidelberg Glycerin Carl Roth GmbH & Co KG, Karlsruhe Glycin Carl Roth GmbH & Co KG, Karlsruhe H-Bouillon Merlin 3-M-HCl-Gas in MeOH Supelco, Bellefonte, PA,USA Hippuryl-L-phenylalanin Sigma-Aldrich GmbH, Taufkirchen Hydroethidin Molecular Probes, Heidelberg Kaliumchlorid p.a. Merck KGaA, Darmstadt Kaliumdihydrogenphosphat p.a. Merck KGaA, Darmstadt Kupfersulfat • 5 H2O Merck KGaA, Darmstadt D-FDAA Novabiochem, Läufelfingen, Schweiz L-FDAA Novabiochem, Läufelfingen, Schweiz Leucinaminopeptidase Sigma-Aldrich GmbH, Taufkirchen l-Leucin-p-Nitroanilid Sigma-Aldrich GmbH, Taufkirchen Magnesiumsulfat • 7 H2O Merck KGaA, Darmstadt Manganchlorid • 4 H2O Sigma-Aldrich GmbH, Taufkirchen McCoys-Medium Sigma-Aldrich GmbH, Taufkirchen Methanol Carl Roth GmbH & Co KG, Karlsruhe MH-Bouillon MacFarland Natriumacetat Sigma-Aldrich GmbH, Taufkirchen Na-Benzoyl-DL-arginin-4-nitroanilid Sigma-Aldrich GmbH, Taufkirchen

V
Natriumcarbonat Merck KGaA, Darmstadt Natriumchlorid Merck KGaA, Darmstadt Natriumdihydrogenphosphat x 2 H2O reinst Merck KGaA, Darmstadt Natriumhydrogencarbonat p.a. Merck KGaA, Darmstadt Natriummolybdat • 2 H2O Merck KGaA, Darmstadt Natriumnitrat Merck KGaA, Darmstadt Natriumphosphat Sigma-Aldrich GmbH, Taufkirchen Natriumsulfat Sigma-Aldrich GmbH, Taufkirchen p-Nitrophenolphosphat Sigma-Aldrich GmbH, Taufkirchen N-Succinyl-Ala-Ala-Ala-p-nitroanilid Sigma-Aldrich GmbH, Taufkirchen N-(3-[2-Furyl]acryloyl-Gly-Leu-amid Sigma-Aldrich GmbH, Taufkirchen N-Succinyl-Ala-Ala-Pro-Phe-p-nitroanilid Sigma-Aldrich GmbH, Taufkirchen Papain Sigma-Aldrich GmbH, Taufkirchen Paraformaldehyd Sigma-Aldrich GmbH, Taufkirchen PEG (MW 35.000) Sigma-Aldrich GmbH, Taufkirchen PIPES Sigma-Aldrich GmbH, Taufkirchen Plasmin Sigma-Aldrich GmbH, Taufkirchen Propidiumiodid BD Pharmingen, Heidelberg Proteinphosphatase PP1A Calbiochem, Bad-Soden QMA Kartuschen (1000 mg) Waters GmbH, Eschborn SA Kartuschen (500 mg) Macherey-Nagel GmbH & Co, KG Düren Salzsäure Carl Roth GmbH & Co KG, Karlsruhe Sephadex LH-20 Material Amersham Biosciences Europe GmbH Freiburg TFA Sigma-Aldrich GmbH (Fluka), Taufkirchen TFAA Fluka, Schweiz Thermolysin Sigma-Aldrich GmbH, Taufkirchen Thrombin Sigma-Aldrich GmbH, Taufkirchen Tosyl-Gly-Pro-Lys-p-nitroanilid Sigma-Aldrich GmbH, Taufkirchen Tris Sigma-Aldrich GmbH, Taufkirchen Tris-HCl Sigma-Aldrich GmbH, Taufkirchen Trypanblau Sigma-Aldrich GmbH (Fluka), Taufkirchen Trypsin Sigma-Aldrich GmbH, Taufkirchen Valinomycin Sigma-Aldrich GmbH, Taufkirchen Zinksulfat • 7 H2O Merck KGaA, Darmstadt

VI
Geräte Autoklav Webeco, Bad Schwartau Autosampler Vials Waters GmbH, Eschborn Chirasil-Val-Säule (Permabond-Chirasil-Val) Macherey-Nagel GmbH & Co. KG Düren CC 250/4 Nucleodur C18-5 Gravity (HPLC-S) Macherey-Nagel GmbH & Co. KG Düren SP 250/4 Nucleosil 100-5 C18 HPLC Säule Macherey-Nagel GmbH & Co. KG Düren SP 250/10 Nucleosil 100-7 C18 HPLC Säule Macherey-Nagel GmbH & Co. KG Düren SP 250/21 Nucleosil 100-7 C18 HPLC Säule Macherey-Nagel GmbH & Co. KG Düren FACS FACS-Calibur Becton/Dickinson Corp., Heidelberg Feinwaage R200D Sartorius AG, Göttingen Gefrierschränke Liebherr-Hausgeräte GmbH, Ochsenhausen HPLC 600 Controller Waters GmbH, Eschborn HPLC 717 Autosampler Waters GmbH, Eschborn HPLC 996 UV Detektor Waters GmbH, Eschborn HPLC Fraction Collector Waters GmbH, Eschborn IR-Spektrometer Spectrum One FT-IR-spectr. Perkin-Elmer, Boston, USA Lyophilisator lyovac 2 Leybold Heraeus GmbH, Hanau Magnetrührer Ikamag Ret IKA, Staufen Massenspektrometer (Maldi Voyager DE-PRO time of flight) PerSeptive BioSystems, Framingham, MS
Massenspektrometer (“LCQ ion trapter”) Finnigan Corp., San Jose Massenspektrometer (“LTQ-FT-ICR) Thermo Electron Corporation, Bremen Membranfilter 0,45 µm (Chromafil PET) Macherey-Nagel GmbH & Co. KG Düren Mikrotiterplatten Falcon, Becton Dickinson, 3072 Mikrotiterplatten (UV) Corning Incorporated 3635 costar, Corning, USA MR 5000 Elisa Reader Dynatech, Chantilly, Virginia NMR Spektrometer (Avance DMX500) Bruker AG, Rheinstetten NMR Spektrometer (Avance DRX500) Bruker AG, Rheinstetten NMR Spektrometer (Avance DRX600) Bruker AG, Rheinstetten Papierfilter 595 Schleicher & Schuell, Dassel PH-Meter 761 Calimatic Knick, Elektron. Messgeräte GmbH & Co Berlin Planktonnetz Hydro-Bios, Kiel-Holtenau Quarzküvetten (UV) Hellma, Müllheim Saugflasche Schott Glas, Duran®, Mainz Säulenofen Millipore GmbH, Eschborn Schütteltisch SWIP Edmund Bühler, Tübingen Szintillationszähler LKB Rackbeta 1209 LKB, Gräfelfing Speed Vac Heto-Holten GmbH, Wettenberg Steilbrustflaschen Schott Glas, Duran®, Mainz Sorvall RC-5B Zentrifuge DuPont Instruments, Bad Homburg Thamnotoxkit F MicroBiotests Inc., Nazareth, Belgien Thermomixer 5437 Eppendorf AG, Hamburg UV Spektrometer Varian Cary 100 Bio Cary, Cary, USA Vakuumfiltrationskammer Macherey-Nagel GmbH & Co. KG, Düren Vakuumpumpe Vaccubrand GmbH & Co KG, Wertheim Zentrifuge 5417 R Eppendorf AG, Hamburg

VII
Abkürzungen ACE Angiotensin converting enzyme Adda 3-amino-9-methoxy-10-phenyl-2,6,8 trimethyl deca,4,6 dienoic acid AFA Aphanizomenon flos aquae Ahp 3-Amino-6-hydroxy-2-oxo-piperidin Aoc Aminooctanoic acid ATCC American type culture collection BAPNA Benzoyl-L-arg-p-nitroanilid BST-Agar Bovine Somatotropin CCCP carbonyl-cyanide m-chlorophenylhydrazone COSY Correlated spectroscopy CSB Cytoskeleton stabilizing buffer Da Dalton Dhb Didehydrobutyrin DDT DiThiothreitol DiOC6(3) 3,3'-Dihexyloxacarbocyanine iodide DMSO Dimethylsulfoxid DNA Desoxyribonucleinsäure dubl Dublett EAWAG Eidgenössische Anstalt für Wasserversorgung EC Enzyme class ED50 Halbmaximale effektive Dosis EDTA Ethylendiamintetraessigsäure ELISA Enzyme linked immunosorbent assay EMEM Eagle’s minimum essential medium ESI Electrospray et al. Et alii FACS Fluorescence activated cell sorter FCS Fötales Kälber Serum FITC Fluoreszeinthiocyanat FSC Forward scatter FT Fourrier Transform FT-ICR Fourrier Transform- Ionenzyklotron gp Glykoprotein ha Hektar HAc Essigsäure H-Bouillon Hinton-Bouillon HE Hydroethidin Hiv Hydroxyisovaleriansäure HMBC Heternuclear Multiple-Bond Correlation Hmv Hydroxymethylvaleriansäure HPLC High performance liquid chromatography HSQC Heteronuclear single quantum coherence spectroscopy HyPro Hydroxyprolin IC50 Inhibition concentration; malbmaximale Hemmwirkung IgG Immunglobulin G IR Infrarot IUPAC International Union of Pure and Applied Chemistry LC-MS Liquid chromatography; Flüssigchromatographie LD50 Letale Dosis; Dosis die zum Tod von 50%der Vergleichspopulation führt LPS Lipopolysaccharid LTQ-FT Linear Trap Q – Fourrier Transform MALDI-TOF-MS Matrix assisted laser deionisation ionisation Time of flight Me Methyl- MeOH Methanol MH-Bouillon Müller-Hinton-Bouillon MHZ Megaherthz
VIII
MIC Minimal inhibierende Konzentration mmu Milli mass units NMTyr N-Methyl-Tyrosin MS Massenspektrometrie mult Multiplett NMR Nuclear magnetic resonance NOE Nuclear overhauser effect NRPS Nichtribosomale Polypeptid Synthetase OD Optische Dichte OMTyr O-Methyl-Tyrosin PBS Phosphate buffered saline (Phosphatpuffer) PCC Pasteur culture collection PEG Polyethylenglykol PI Propidiumiodid PIPES Piperazin-N,N´-bis[2-ethansulfonsäure] ppm Parts per million PSD Post source decay RNA Ribonucleinsäure ROESY Rotating Frame Overhauser Effect Spectroscopy RP-HPLC Reversed phase-HPLC; Umkehrphasen-HPLC sing Singulett SSC Sideward scatter T Tesla TFA Trifuoressigsäure TFAA Trifluoressigsäureanhydrid TISTR Thailand Institute of Scientific and Technological Research TOCSY Total correlated spectroscopy trip Triplett Tris Tris(hydroxymethyl)-aminomethan U/min Umdrehungen / Minute UV Ultraviolett v/v Volumenanteil (volume per volume)
Aminosäure Dreibuchstaben-Code Einbuchstaben-Code Alanin Ala A Arginin Arg R Asparagin Asn N Asparaginsäure Asp D Cystein Cys C Glutamin Gln Q Glutaminsäure Glu E Glycin Gly G Histidin His H Isoleucin Ile I Leucin Leu L Lysin Lys K Methionin Met M Phenylalanin Phe F Prolin Pro P Serin Ser S Threonin Thr T Tryptophan Trp W Tyrosin Tyr Y Valin Val V

IX
Zusammenfassung
Die vorliegende Arbeit beschreibt die Isolierung und Charakterisierung von Cyanopeptolinen
und Scytocyclamiden aus Zellen des Cyanobakteriums Scytonema hofmanni PCC 7110. Es
wurden drei Peptide vom Cyanopeptolin-Typ isoliert und mit den Scytocyclamiden A bis C
eine, für Scytonema hofmanni PCC 7110 bisher nicht beschriebene Peptidklasse identifiziert.
Die neu isolierten Cyanopeptoline wurden als Derivate mit Molekülmodifikationen zu den
von Matern et al. (2001 und 2003) beschriebenen Scyptolinen A und B, sowie Hofmannolin
identifiziert. Cyanopeptolin 1067A unterscheidet sich von Hofmannolin durch den Austausch
von Tyrosin gegen die basische Aminosäure Arginin auf N-terminaler Seite der Ahp-Gruppe
[2-Hydroxyisovaleryl-Glu-(Thr-Arg-Ahp-OMTyr-NMTyr-Val-O-)]. Mit Cyanopeptolin
1060A wurde eine weitere verwandte Struktur zu Hofmannolin aufgeklärt: anstatt 2-Hydroxy-
3-methylvaleriansäure findet man 2-Hydroxyisovaleriansäure als Acylrest am N-Terminus der
Seitenkette [2-Hydroxyisovaleryl-Glu-(Thr-Tyr-Ahp-OMTyr-NMTyr-Val-O-)]. Bei Cyano-
peptolin 946A [Butyryl-Ala-Thr-(Thr-Leu-Ahp-Thr-MTyr-Val-O-)] wurde das Fehlen der
Chlorierung am N-methylierten Tyrosin im Vergleich zu Scyptolin A belegt.
Scytocyclamid A gleicht zu großen Teilen Laxaphycin A, einem Peptid, welches erstmals bei
Anabaena laxa beschrieben wurde. Es ist ein zyklisches Undecapeptid [(βAoc-Gln-E-Dhb-
Hyp-Hse-Phe-Leu-Ile-Ile-Leu-Gly)]. Im Vergleich zu Laxaphycin A ist Homoserin gegen
Glutamin ausgetauscht. Zwei weitere Peptide aus dieser Substanzklasse wurden strukturell
aufgeklärt: Scytocyclamid B weist eine zu Laxaphycin D identische Masse und
Aminosäuresequenz auf. Scytocyclamid C unterscheidet sich von Scytocyclamid B durch das
Fehlen einer Hydroxylgruppe am Leucin. Beide sind zyklische Dodecapeptide: [(βAoc-Val-
Hle-Ala-Hle-Gln-NM-Ile-Has-Thr-Pro-Leu-Thr)] für Scytocyclamid B, bzw. [(βAoc-Val-Hle-
Ala-Leu-Gln-NM-Ile-Has-Thr-Pro-Leu-Thr)] für Scytocyclamid C. Die Strukturen der
bearbeiteten Peptide wurden durch Massenspektometrie, IR- und NMR-Spektroskopie gelöst.
Aufgrund der geringen Ausbeute musste bei den Cyanopeptolinen auf eine Konfigurations-
analyse verzichtet werden. Analog zu allen anderen beschriebenen Cyanopeptolinen liegt
jedoch vermutlich jeweils die L-Konformation der Aminosäuren vor. Bei Scytocyclamid A
wurde die Konfiguration durch GC-MS-Versuche an chiralen Phasen soweit möglich
bestimmt.
Die Cyanopeptoline sowie die Scytocyclamide wurden in verschiedenen Testsystemen auf
ihre antimikrobielle, enzyminhibierende, zytotoxische und toxische Wirkung überprüft. Eine

X
antimikrobielle Wirksamkeit wurde für keine der Substanzen festgestellt. Bei den
Enzymhemmtests hatte Cyanopeptolin 946A eine vergleichbare Wirkung auf Elastase (IC50
von 0,45 µg/ml entsprechend 0,47 µmol) wie Scyptolin A (0,16 µmol), bzw. Scyptolin B
(0,14 µmol). Bei allen anderen getesteten Enzymen wurde keine Wirkung beobachtet.
Cyanopeptolin 1060A konnte kein Zielenzym zugeordnet werden. Die Strukturvariante des
Cyanopeptolins 1067A mit der basischen Aminosäure Arginin neben der Ahp-Gruppe hatte
deutliche Wirkung gegenüber Trypsin (IC50 von 0,064 µg/ml entsprechend 60 nmol). Neben
Trypsin wurde auch Plasmin gehemmt (IC50 von 1,0 µg/ml entsprechend 0,94 µmol).
Für die Klasse der Scytocyclamide wurden toxische Effekte auf Primärkonsumenten belegt:
Im 24 h Toxizitätsassay hatten die genannten Substanzen bereits nach 2 h letale Wirkungen
bei Thamnocephalus platyurus (LD50 für Scytocyclamid B von 4 µmol und für
Scytocyclamid A von 12 µmol).
Bei den Tests zu einer möglichen Wirkung der isolierten Peptide auf humane Zelllinien waren
die Cyanopeptoline unwirksam, während Scytocyclamide eine ausgeprägte Zellzytotoxizität
aufwiesen. Bei den Zellzytotoxizitätstests wurden alle getesteten Zelllinien bei einer
Konzentration von 10 µg/ml (entsprechend ca. 8 µmol) gleichermaßen beeinflusst und die
Zellen abgetötet. Die Wirkung wurde neben Aufnahmen mit dem Phasenkontrastmikroskop
durch ein Differential-Interferenz-Kontrastmikroskop dokumentiert.
Bei den Ansätzen zur Frage nach der Ursache der Zytotoxizität wurden verschiedene
apoptotische Prozesse überprüft. Eine für Apoptose charakteristische Caspase-Aktivität oder
der Verlust des Mitochondrien-Membranpotentials wurden allerdings nicht bestätigt. Es gab
jedoch andere Hinweise auf mögliche Apoptose-Mechanismen, wie die Translozierung von
Phosphatidylserin von der Membraninnenseite auf die Membranaußenseite, welche mittels
Durchflusszytometrie verfolgt wurde. Eine mögliche Induktion der Apoptose könnte über
reaktive Sauerstoffspezies erfolgen, welche durch die Oxidierung von Hydroethidin zu
Ethidium nachgewiesen wurden.
Membranschädigung durch Porenbildung in künstlichen Membranen mit Diphytanoyl-
phosphatidylcholin als Membranbestandteil durch die isolierten Peptide wurde nicht
beobachtet. Bei cholesterolhaltigen Membranen wurde jedoch erhöhte Leitfähigkeit
gemessen. Die vermutete Membranschädigung wurde durch 86Rubidium+-Efflux-Experimente
an HT-29-Zellen belegt. Als weiterer zellschädigender Effekt wurde der Abbau von
α-Tubulin durch Scytocyclamide mittels Immunfluoreszenzfärbung nachgewiesen.

Einleitung 1
1. Einleitung
1.1. Cyanobakterien und sekundäre Stoffwechselprodukte
Naturprodukte sind eine wertvolle Quelle für biologisch aktive Verbindungen trotz der
Fortschritte in der synthetischen Chemie. Heute ermöglichen DNA-Sequenzdatenbanken neue
Zielstrukturen zu identifizieren. Nachdem man sich bei der Suche nach neuen Wirkstoffen
oder Leitstrukturen für neue Medikamente zunächst auf terrestrische Pflanzen und Tiere
beschränkte, wird mittlerweile das Gebiet der Mikroorganismen intensiv untersucht.
Meist sind es die Sekundärstoffwechselprodukte, die von besonderem Interesse sind. Sie sind
zwar für Wachstum und Entwicklung entbehrlich, wurden aber im Zuge der Evolution unter
dem Selektionsdruck der Umwelt optimiert und stellen ein großes Reservoir von
„Wirkstoffen“ dar. Auch die forschenden Pharmakonzerne bedienen sich dieses schier
unerschöpflichen Molekülbaukastens. Dabei gibt die traditionelle Medizin einzelner Völker
oft wichtige Hinweise bei der Suche nach potenten Verbindungen (Brunner, 2000). Hinweise
auf die Verwendung von Cyanobakterien finden sich beispielsweise aus der Zeit um 1500 v.
Chr. als Nostoc-Arten zur Behandlung von Gicht, Fisteln und verschiedenen Formen des
Krebs eingesetzt wurden (Burja et al., 2001). Neben der AFA-Alge (Aphanizomenon flos
aquae, kurz AFA) werden Nostoc sp. und Spirulina sp. als Nahrungsergänzungsmittel und
teilweise seit mehr als 1.000 Jahren als protein- und vitaminreiche Nahrungsquelle (Kay,
1991) eingesetzt. Seit mehreren Jahren werden cyanobakterielle Extrakte in die Screening-
Programme aufgenommen. Die stoffliche Vielfalt steht im Einklang mit der beobachteten
breiten Bioaktivität. Auf Grund der pharmakologischen Eigenschaften könnten solche
Substanzen, wie oben erwähnt, als Leitstrukturen zur Entwicklung neuer Pharmaka
Bedeutung erlangen (Radau, 2000).
1.2. Cyanobakterien
Cyanobakterien, früher auch Blaualgen oder Cyanophyceaen genannt, sind prokaryontische,
photoautotrophe Organismen, die keinen membranumhüllten Zellkern oder andere
membrangebundene Zellorganellen wie Chloroplasten oder Mitochondrien besitzen. Sie
besitzen Chlorophyll a, daneben zusätzlich Phycobiline, die im gelben und grünen
Spektralbereich (Phycocyanin, bzw. Phycoerythrin) Licht absorbieren. Phycobiline findet man

Einleitung 2
nur bei Cyanobakterien und Rotalgen. Sie sind es auch, welche die charakteristische Farbe
ausmachen.
Die Zugehörigkeit zu den Bakterien wurde unter anderem durch elektronenmikroskopische
Untersuchungen und 16s rRNA Analysen begründet. Cyanobakterien zählen zu den gram-
negativen Bakterien.
Die Einteilung der Cyanobakterien in Familien erfolgt derzeit noch nach morphologischen
Kriterien und nach der Vermehrungsweise (Anagnostidis et al., 1990; Baker et al., 1999). So
können Cyanobakterien einzellig, in Zellgruppen (Kolonien) oder als Zellfäden (Filamente)
auftreten. Sie vermehren sich durch Zwei- oder Mehrfachteilung oder durch wiederholte
Abgliederung endständiger Zellen. Es gibt Veröffentlichungen, welche eine Eingliederung der
Cyanobakterien unter dem „Bacteriological Code“ vornehmen. Bisher sind gerade einmal 13
cyanobakterielle Gattungen im „International Journal of Systematic and Evolutionary
Microbiology (IJSEM)/International Journal of Systematic Bacteriology (IJSB)“
vorgeschlagen worden; von diesen wurden aber nur fünf unter gültigen Gesichtspunkten
veröffentlicht (Oren, 2004). Beide Nomenklatursysteme werden im Augenblick parallel
angewandt, als Konsequenz daraus besitzen viele Cyanobakterienarten mehrere Synonyme
(Skulberg, 1993). In absehbarer Zeit ist eine neue Einteilung unter Berücksichtigung
genetischer Daten zu erwarten.
Zur Zeit werden 5 Sektionen unterschieden, wobei die Sektionen I (Chroococcales) und
II (Pleurocapsales) einzellige Formen umfassen, die sich auf Grund ihrer Zellteilung
unterscheiden, während die Sektionen III (Oscillatoriales), IV (Nostocales) und V
(Stigonematales) filamentöse Arten umfassen, welche sich durch Besitz von Heterocysten und
die Art der Differenzierung unterscheiden.
Fossile Funde (versteinerte Stromatoliten) belegen, dass die Vorläufer der heutigen
Cyanobakterien bereits vor 2,5 bis 3,5 Milliarden Jahren die Erde besiedelten und damit zu
den ältesten Organismen gehören, die zu einer sauerstoffbildenden Photosynthese fähig sind.
Stromatoliten sind calciumhaltige Strukturen, welche während starkem Wachstum von
Cyanobakterien gebildet werden können. Prominestestes Beispiel sind die Stromatoliten in
Shark Bay, Westaustralien.
Man nimmt heute an, dass Cyanobakterien im Proterozoikum eine der dominierenden
Lebensformen waren. Auch gilt es als gesichert, dass Cyanobakterien einen entscheidenden
Einfluss auf die Sauerstoffanreicherung der damals noch anaeroben Atmosphäre hatten.
Es gibt Evidenzen, die nahe legen, dass symbiontisch lebende Cyanobakterien sich evolutiv
zu Chloroplasten entwickelt haben (Stackebrandt, 1985): die innere Membran der

Einleitung 3
Chloroplasten enthält Transportsysteme und Enzyme, die denen von Cyanobakterien ähneln
(Douglas et al., 2001). Zum anderen ist die DNA der Chloroplasten ringförmig angeordnet
und nicht mit Histonen assoziiert. Die Organellen enthalten alle für die Transkription und
Translation benötigten Komponenten. Molekulargenetische Merkmale stützen diese
Endosymbiontentheorie (Martin et al., 1997; Kutschera und Niklas, 2005).
Neben der Fähigkeit zur oxygenen Photosynthese, besitzen manche Arten auch die
Fähigkeit Luftstickstoff zu Ammoniak zu reduzieren. Die Stickstofffixierung findet unter
Sauerstoffausschluss in spezialisierten Zellen, den Heterocysten, statt, welche eine verdickte
Zellwand besitzen und der Nitrogenase Schutz vor Sauerstoff bieten. Die Fähigkeit
Luftstickstoff zu fixieren, macht man sich heute beim Reisanbau zu nutze, indem man
Cyanobakterien als Düngemittel dem Boden zusetzt. Cyanobakterien sind auf der ganzen
Welt verbreitet und kommen terrestrisch, planktisch, benthisch, limnisch und marin vor.
Manche Arten sind in der Lage bestimmte Nischen zu ihrem Vorteil auszunutzen. So können
manche Arten zum Beispiel ihren Auftrieb im Wasser mittels Gasvakuolen steuern. Durch
diese Eigenschaften können sie sich an Licht- und Nahrungsverhältnisse optimal anpassen.
Unter bestimmten äußeren Bedingungen kann es zu Massenvermehrung von Cyanobakterien
kommen. Diese können dann als „Wasserblüten" an der Oberfläche von Gewässern
beobachtet werden. Wasserblüten wurden vor allem bei den Gattungen Aphanizomenon,
Microcystis, Anabaena, Nodularia, Nostoc, Lyngbya und Oscillatoria beschrieben.
Massenvermehrung wird insbesondere in den heißen Sommermonaten in eutrophen
Gewässern mit niedrigem Sauerstoffgehalt beobachtet. Insgesamt zeigen sich die
verschiedenen Gattungen als sehr anpassungs- und widerstandsfähige Organismen, welche
vorübergehende Austrocknung, Temperaturschwankungen, Sauerstoffmangel, oder
wechselndes Nährstoffangebot tolerieren können. Als photolithoautotrophe Bakterien sind
Cyanobakterien in der Lage bestimmte ökologische Nischen als Lebensräume zu nutzen,
unter anderem auch unter extremen Umweltbedingungen, wie in heißen Quellen, in der Wüste
und in arktischen Seen (Fay, 1983). Ein neuer Ansatzpunkt für ein Nischendasein wurde bei
dem Cyanobakterium Acaryochloris marina gefunden, welches anstatt Chlorophyll a und
Phycobilliproteine Chlorophyll d zur Photosynthese nutzt. Somit reichen geringere
Lichtintensitäten zur Energiegewinnung als bei Konkurrenten, welche Chlorophyll a nutzen,
aus (Kühl et al., 2005).

Einleitung 4
1.3. Symbiose
Einige Cyanobakterien leben in Symbiose mit verschiedenen Eukaryoten, dazu zählen
verschiedene Protisten, Pilze, Algen und Pflanzen (Sergeeva et al., 2002). Die Interaktionen
zwischen Wirt und Symbiont sind spezifisch. Ein Vorteil für den Wirt scheint in der Fähigkeit
der Cyanobakterien zu liegen, Stickstoff und Kohlendioxid fixieren zu können. Als
Gegenleistung bietet der Wirt eine Wachstumsumgebung, die die Cyanobakterien vor u. a. zu
hoher Lichtintensität oder vor Austrocknung schützt (Adams, 2000). Wie bereits erwähnt,
macht man sich aus agrarökonomischer Sicht die Fähigkeit der Stickstofffixierung von
Cyanobakterien in der Kultivierung von Reispflanzen zu Nutze. In diesem Zusammenhang
wurde auch eine spezifische Symbiose zwischen Azolla-Anabaena beschrieben, in welcher
der Stickstoffertrag von normalerweise 20-30 kg N ha-1 auf 600 kg N ha-1 erhöht war
(Vaishampayan et al., 2001). Azolla ist in den Reisfeldern Ostasiens verbreitet, wo ein
beachtlicher Teil des von ihm gebundenen Stickstoffs den Reispflanzen zugute kommt. Auch
marine Schwämme werden als Symbiosepartner beschrieben, wie etwa Dysidea herbacea mit
Oscillatoria spongeliae (Ridley et al., 2005). Die Geosiphon-Symbiose ist die einzige
bekannte Pilz-Cyanobakterien-Endosymbiose (Schüssler, 2001). Eine weitere Endosymbiose
findet man zwischen Nostoc sp. und dem Mammutblatt (Gunnera) (Meeks, 1998). In den
meisten Fällen jedoch sind es Symbiosen, bei denen die Bakterien extrazellulär vorliegen.
Flechten sind die am meisten verbreiteten Lebensgemeinschaften. Von den 15.000-20.000
Flechtenarten haben etwa 8 bis 15 % ein Cyanobakterium als Phycobiont, meistens Nostoc,
aber auch Calothrix, Scytonema und Fischerella (Adams, 2000).
Die Herkunft mancher Sekundärstoffwechselprodukte wird inzwischen kontrovers diskutiert.
Es konnte in einigen Fällen nachgewiesen werden, dass die ursprünglich einem Eukaryoten
zugeordnete Substanz auf ein in Symbiose lebendes Cyanobakterium zurückgeht. Die aus
dem im Indischen und Pazifischen Ozean heimischen Seehasen Dolabella auricularia
gewonnenen Dolastatine scheinen nicht von der marinen Schnecke produziert zu werden. Sie
gehen wahrscheinlich auf die mit der Nahrung eingenommenen Cyanobakterien zurück (Piel,
2004). Weiterhin ist oft ungeklärt, ob diese Metabolite nur als Produkt einer Symbiose
gebildet werden (Sings und Rinehart, 1996; Weckesser et al., 1996). Patellamide wurden
ursprünglich aus marinen Tunikaten Lissoclinum patella isoliert. Jetzt konnte schlüssig
nachgewiesen werden, dass die Biosynthese von Patellamide A und B über einen „microcin-
like pathway“ in Prochloron didemni, dem cyanobakteriellen Symbiont von Lissoclinum
patella erfolgt (Schmidt et al., 2005).

Einleitung 5
1.4. Sekundärmetabolismus
Unter Sekundärstoffwechselprodukten versteht man niedermolekulare Stoffwechselprodukte,
die für Wachstum und Vermehrung nicht essentiell sind. Man geht davon aus, dass sie
trotzdem einen Nutzen für den produzierenden Organismus bringen. Erst mit der Erforschung
der natürlichen Lebensbedingungen im Bereich des Mikrohabitats wird die Funktion der
Sekundärmetabolite erkennbar. Jedoch müssen die vielfach beschriebenen Bioaktivitäten
nicht mit der ökologischen Funktion übereinstimmen. Die Bildung ist von mehreren Faktoren
abhängig, wie Wachstumsbedingungen, Wachstumsphase, Induktion oder Repression der
Bildung. Verfügt ein Organismus über die notwendigen Gene zur Bildung der
Sekundärstoffwechselprodukte, so findet man häufig mehrere verschiedene, aber
strukturverwandte Produkte in unterschiedlichen Ausbeuten. Unter den
Sekundärstoffwechselprodukten finden sich eine Vielzahl von Stoffklassen, unter ihnen
Alkaloide, Polyketide oder nichtribosomal gebildete Peptide, manche davon sind toxisch. In
den folgenden Abschnitten wird eine Auswahl an Sekundärstoffwechselprodukten gegeben
ohne dabei Anspruch auf Vollständigkeit zu erheben.
1.5. Ökologie der Sekundärstoffwechselprodukte
Gesicherte Vorstellungen über die Funktion der Sekundärstoffwechselprodukte gibt es nur
wenige. Unter Einbeziehung der Lebensbedingungen in den Mikrohabitaten werden nur
manche Funktionen erklärbar. Viele publizierte Thesen beruhen daher auf Spekulationen.
Eine gängige These besagt, dass die Metabolite gebildet werden, um einen Selektionsvorteil
zu genießen und die ökologische Nische zu verteidigen. So wurde vor einiger Zeit eine
Serinprotease aus Anabaena beschrieben, welche unter Stickstoffmangel induziert wird (Yoon
und Golden, 1998). Diese Beobachtung steht im Einklang mit der Bildung von
Sekundärstoffwechselprodukten unter bestimmten Kulturbedingungen, bzw. nur in
bestimmten Entwicklungsphasen der Organismen oder Kulturen (Ruhephasen, oder
Stresssituationen). Vielfach sind exogene oder endogene Effektoren für die Existenz in
natürlichen Ökosystemen verantwortlich (Fritsche, 2002). Scytonemin, eine Substanz aus der
Ummantelung von Scytonema, wurde eine Lichtschutzfunktion zugeordnet. Als UV-Licht
absorbierende Substanz ermöglicht sie dem Cyanobakterium auch an Standorten hoher
Lichtintensität zu überleben, indem sie energiereiches Licht absorbieren und in unschädliche
Wärme umwandeln (Proteau et al., 1993).

Einleitung 6
Bei toxinbildenden Algen nimmt man an, dass das Toxin zur Abwehr von
Primärkonsumenten der Cyanobakterien produziert wird (Carmichael, 1992). Bei einem
Vergleich von käuflichen Proteasen (Isolate aus Rindern) mit Isolaten von Proteasen aus dem
Darm von Daphnien wurde belegt, dass die Proteaseinhibition in der natürlichen Umgebung
dazu führen kann, dass Cyanobakterien nicht von Fraßfeinden verdaut werden können
(Agrawal et al., 2005). Hierzu gehören die Ansätze, die eine Hemmung zellregulierender
Enzyme wie Proteinphosphatasen und endogenen Proteasen postulieren. Eine weitere
wirkungsvolle Abwehr von Fraßfeinden scheint die Produktion von ungenießbaren
Metaboliten zu sein, was dazu führt, dass die Fraßfeinde die Cyanobakterien wieder
„ausspucken“ (Nagle und Paul, 1999). Andere Autoren vermuten, dass die Substanzen als
Chelatoren potentiell toxischer Metalle fungieren (Humble et al., 1997).
1.6 Biosynthese der Sekundärstoffwechselprodukte
Aufgrund der Zusammensetzung der Peptid-Sekundärmetabolite lässt sich vermuten, dass sie
nichtribosomal synthetisiert werden. An Stelle von DNA und RNA als Informationsträger tritt
ein Protein-Multienzymkomplex. Dieser Biosynthesetyp kommt nur bei Bakterien und Pilzen
vor und ist für die Bildung der meisten peptidischen Sekundärmetabolite verantwortlich. Die
Synthese erfolgt nicht-ribosomal nach dem Thiotemplate-Mechanismus. Als Besonderheiten
findet man unter den Nichtribosomale-Peptid-Synthetase-Produkten Cyclopeptide,
D-Aminosäuren, nicht-proteinogene Aminosäuren, N-Methylierung, Einbau von
Hydroxysäuren (Depsipeptide), Einbau von Polyketideinheiten und
Aminosäuremodifikationen (z. B. oxidativ). Weitere Syntheseapparate wurden an Bacillus
untersucht. Dort sind sie für die Synthese der Peptidantibiotika Gramicidin S, Bacitracin und
Tyrocidin verantwortlich (Kleinkauf und von Döhren, 1996). Gene der Nichtribosomalen-
Peptid-Syntethase finden sich in den meisten cyanobakteriellen Gattungen (Christiansen,
2001). Durch Kristallstrukturen von NRPS-Domänen bekommt man einen immer genaueren
Einblick in die Biosynthese. Diese Kenntnisse sollen dazu genutzt werden in einem solchen
Biokatalysator einzelne Moleküle gezielt auszutauschen, um auf diese Weise
Peptidantibiotika mit neuen Eigenschaften zu gewinnen (Schwarzer et al., 2003; Dürfahrt und
Marahiel, 2005). Neben den Peptidsynthetasen sind auch Polyketidsynthasen charakterisiert
worden, sie sind für die Biosynthese von Polyketiden und Alkaloiden verantwortlich
(Dittmann et al., 2001). Bei Cyanobakterien ist die Biosynthese der toxischen Microcystin-
Peptide mit am genauesten untersucht (Tillett et al., 2000).

Einleitung 7
1.7. Biologisch aktive Substanzen aus Cyanobakterien
Auf der Suche nach neuen Quellen für bioaktive Substanzen werden immer wieder
Cyanobakterien als Untersuchungsgegenstand gewählt. Systematisches Screening nach
Bioaktivität zeigte, dass diese Mikroorganismen eine reiche Quelle an bisher unbekannten
bioaktiven Substanzen bietet (Patterson et al., 1991, 1993 und 1994; Honkanen et al., 1995;
Falch et al., 1995; Jaki et al., 1999). So offenbaren Cyanobakterien ein großes Potential für
die pharmazeutische Forschung, um eingangs erwähnte Leitsubstanzen mit ungewöhnlichen
oder unbekannten Strukturmerkmalen zu finden.
In den folgenden Abschnitten werden die Sekundärstoffwechselprodukte anhand ihrer
biologischen Aktivität unterschieden. Die Einteilung erfolgt nach den Wirkungseigenschaften
in pharmakologischen Klassen. Im Allgemeinen sind es Peptide, obwohl auch Ketide,
Makrolide oder Alkaloide genannt werden. Die hier behandelten Stoffwechselprodukte
spiegeln die große strukturelle Vielfalt an Substanzen aus dem Sekundärstoffwechsel wider.
Angesichts mehrerer hundert Substanzen, die bis heute strukturell aufgeklärt und
veröffentlicht wurden, erhebt die Auflistung jedoch keinen Anspruch auf Vollständigkeit.
1.7.1 Cyanobakterielle Toxine
Cyanotoxine umfassen ein breites Spektrum an algiziden, antimykotischen, antibakteriellen
aber auch tier- und humantoxischen Eigenschaften. Einige Beispiele sind im Folgenden
aufgelistet.
Zytotoxine und Biotoxine
Zytotoxine besitzen oftmals algizide, antimykotische, oder antibakterielle Eigenschaften, oder
besitzen Aktivität auf Zelllinien. Ihre Erforschung liegt in der Suche nach neuen
pharmazeutisch oder landwirtschaftlich nutzbaren Substanzen begründet. Einen Überblick zu
cyanobakteriellen Zytotoxinen gibt der Abschnitt „Cyanobakterien als Quelle neuer
Arzneistoffe“ (1.7.2.). Aus Cyanobakterien erhaltene Biotoxine mit hepatotoxischen,
neurotoxischen, dermatotoxischen und reizauslösenden Eigenschaften sind bekannt (Bartram
und Chorus, 1999).

Einleitung 8
1.7.1.1. Hepatotoxine
Die cyanobakteriellen Peptide vom Microcystin-Typ stellen die am besten untersuchte
Gruppe an cyanobakterielen Sekundärmetaboliten dar. Das Interesse an der Charakterisierung,
Toxizität und den zu Grunde liegenden Mechanismen liegt an ihrer potentiellen Gefahr für
Tier und Mensch sowie an dem Umstand, dass sie von Cyanobakterienarten wie Microcystis
aeruginosa gebildet werden, die besonders häufig vorkommen (Bartram und Chorus, 1999
1999). Neben Microcystis-Arten wurden Microcystine auch in Anabaena-, Nostoc- und
Planktothrix-Arten. identifiziert. Microcystine sind Hepatotoxine. Mit LD50-Werten zwischen
50 und 250 µg/kg Körpergewicht (i.p. Maustest) ist die Toxizität deutlich höher als bei
Cyanid (LD50 1 mg/kg). Microcystine werden über den Gallensäure-Transporter in die
Leberzellen eingeschleust. Die toxische Wirkung wird durch irreversible Hemmung der
Proteinphosphatasen 1 und 2A hervorgerufen. Dadurch kommt es zu einem Ungleichgewicht
zwischen Phosphatasen und Kinasen und einer Hyperphosphorelierung von Proteinen im
Zytosol, bzw. des Zytoskeletons. Durch die übermäßige Proteinphosphorelierung kommt es
zu einem Zusammenbruch des Zytoskeletts und zum Ausfall der Leberfunktionen.
Wahrscheinlich wirken Microcystine und Nodularine nicht nur primär auf
Proteinphosphatasen, sondern haben noch andere toxische Effekte (Börner, 2001). Dieser
Verdacht scheint sich nach neueren Veröffentlichungen zu bestätigen. Im Zusammenhang mit
Vergiftungserscheinungen durch Hepatotoxine wurde die Möglichkeit der Apoptoseinduktion
untersucht. Hierbei wurde eine Zellschädigung über verschiedene Wege der
Apoptoseeinleitung gezeigt (Chen et al., 2005).
Untersuchungen an Tieren und Zellkulturen zeigen, dass Microcystine in subletalen Dosen als
Tumorpromotoren wirksam sein können (Nishiwaki-Matsushima et al., 1992). In diesem
Zusammenhang steht auch die Beobachtung, dass in China in Gebieten mit einem zwar
niedrigen, aber stetigen Gehalt an Microcystinen im Trinkwasser eine erhöhte Rate am
Auftreten von Leberkrebs beobachtet wird (Kuiper-Goodman et al., 1999).
Bisher wurden mehr als 60 Strukturvarianten der Microcystine beschrieben. Alle
Microcystine besitzen die ungewöhnliche Aminosäure Adda (3-Amino-9-methoxy-2,6,8-
trimethyl-10-phenyldeka-4,6-diensäure), sowie D-iso-Glutamat, N-Methyldehydroalanin,
D-Alanin, und D-erythro-β-methyl-iso-Aspartat. Die ungewöhnliche Aminosäure Adda
bewirkt ein charakteristisches UV-Spektrum mit einem Absorptionsmaximum bei 238 nm.

Einleitung 9
Nodularine werden von Nodularia spumigena gebildet. Sie zeigen eine den Microcystinen
vergleichbare Bioaktivität und werden ebenfalls zu den Hepatotoxinen gezählt. Im Gegensatz
zu Microcystinen bestehen Nodularine nur aus fünf Aminosäuren. Wie Microcystine enthalten
sie ebenfalls Adda, D-MeAsp und D-Glu, zusätzlich noch L-Arg und N-Methyldehydro-α-
aminobutyrat.
Ein weiteres Lebertoxin, Cylindrospermopsin, wurde aus Cylindrospermopsis raciborskii,
Umezakia natans und Aphanizomenon ovalisporum isoliert. Cylindrospermopsin ist ein
Alkaloid mit einer zyklischen Guanidingruppe und ruft Schädigungen an Leber, Niere,
Thymus und Herz hervor (Moore et al., 1993.
1.7.1.2. Neurotoxine
Cyanobakterien sind in der Lage Alkaloide als Sekundärstoffwechselprodukte zu bilden.
Unter den von ihnen gebildeten Alkaloiden finden sich Neurotoxine. Im Gegensatz zu den
Hepatotoxinen, werden die Neurotoxine von den Zellen in die Umgebung abgegeben
(Exotoxine). Das erste charakterisierte Cyanobakterientoxin, Anatoxin-A, wird von Anabaena
gebildet. Anatoxin-A zeigt agonistische Wirkung an cholinergen Rezeptoren der Postsynapse
(Carmichael et al., 1979). Des Weiteren ist die Affinität zum Rezeptor im Vergleich zu
Acetylcholin bis zu 8-fach stärker ausgeprägt. Anatoxin-A löst eine Dauerpolarisation aus und
führt zu massiven Muskelkrämpfen. Acetylcholinesterase ist nicht in der Lage das Toxin
abzubauen. Dies führt dazu, dass sich unmittelbar Vergiftungserscheinungen zeigen, die zum
Erstickungstod binnen weniger Minuten führen können. Aus diesem Grund wird Anatoxin-A
NH
O
NHN
ONH
O
OMe
CO2H
NH
O
ONH
O
NH
ONH
NH2
NHCO2HMicrocystin LR
NH
N NH NH NH
O3SOOH
O
O
H
H H
Cylindrospermopsin

Einleitung 10
häufig auch als „very fast death factor“ (VFDF) bezeichnet (Draisci et al., 2001). Die LD50-
Werte für das Toxin liegen zwischen 200 und 250µg/kg Körpergewicht.
Die Tetrahydropurinalkaloide Saxitoxin und Neosaxitoxin (frühere Bezeichnung:
Aphantoxine) sind die bislang als giftigste bekannten Cyanobakterientoxine. Sie wirken über
eine Unterbrechung der Erregungsleitung indem sie Natriumionenkänale blockieren. Der Tod
tritt durch Lähmung der Atemmuskulatur und Ersticken ein. Die letale Dosis liegt bei 10µg/kg
Körpergewicht. Erstmals wurden diese Aphantoxine aus Aphanizomenon flos-aquae isoliert
(Mahmood & Carmichael 1986). Saxitoxine sind die Ursache für oft tödlich verlaufendes
„paralytic shellfish poisoning“. Saxitoxine werden in Muscheln stark angereichert und deren
Verzehr kann dann unbehandelt tödlich verlaufen. Vermehrt wird dieses Phänomen während
oder nach Auftreten massiver Wasserblüten („red tides“) beobachtet (Ueno und Nagata,
1994).
1.7.1.3. Dermatotoxine und hautreizende Toxine
Ein dem Teleocidin A-1 aus Streptomyces Arten identisches Molekül wurde aus Lyngbya
majuscula isoliert (Cardellina et al., 1979). Lyngbyatoxin A gehört zu den Indolalkaloiden.
Durch Aktivierung der Proteinkinase C bewirkt es eine verstärkte Phosphorelierung von
Serin- und Threoninresten eukaryotischer Proteine (Osborne et al., 2001). In dem vermehrten
Auftreten von Kontaktdermatiden beim Baden in Gewässern um Hawaii und zeitgleich
auftretenden Wasserblüten wird ein Zusammenhang mit Lyngbyatoxin gesehen. Die
Kontaktdermatitis wird auch als „swimmers itch“ bezeichnet (Mundt und Teuscher, 1988).
Lipopolysaccharide sind integrale Membranbestandteile gram-negativer Bakterien,
einschließlich Cyanobakterien. Hinsichtlich der Wirkung cyanobakterieller
Lipopolysaccharide auf Menschen und andere Organismen wurden kaum Untersuchungen
durchgeführt (Duy et al., 2000). Cyanobakterielle Endotoxine scheinen eine geringere
NHN
NH
NH
ONH2
O
NH OHOH
NH
HONH
Anatoxin-A Saxitoxin

Einleitung 11
Toxizität aufzuweisen als LPS von Salmonella (Bell und Codd, 1996). Hautreizungen werden
unter anderem auch auf Lipopolysaccharide zurückgeführt (Börner, 2001).
1.7.2.Cyanobakterien als Quelle neuer Arzneistoffe
Bei der Suche nach möglichst bisher unbekannten Substanzen bedient man sich auch der
Cyanobakterien. Unter den charakterisierten Substanzklassen aus Cyanobakterien finden sich
die unterschiedlichsten Bioaktivitäten. In Kenntnis der Wirkungsweise und der Struktur-
Wirkungsbeziehung der natürlichen Substanzen versucht man Strukturanaloga mit stärkeren
und spezifischen Aktivitäten zu entwickeln (Burja et al., 2001).
1.7.2.1. Antimikrobielle Substanzen
Einige cyanobakterielle zyklische Peptide weisen antimikrobielle Aktivität auf. Es gibt zum
Beispiel zahlreiche Veröffentlichungen zur Produktion von antifungalen Inhaltsstoffen aus
Cyanobakterien und Algen. Meistens wird jedoch nach möglichem Einsatz als Arzneimittel
gesucht. Untersuchungen zur Wirksamkeit gegen pathogene Pilze wurden nur wenige
durchgeführt (Nagle, 2002). Bereits Anfang der 90iger Jahre wurde cyanobakterielle Peptide
mit antifungalen Eigenschaften beschrieben. Unter ihnen die Laxaphycine, welche aus dem
Extrakt von Anabaena laxa isoliert wurden (Frankmölle et al., 1992). Laxaphycine sind
zyklische Peptide aus 10, bzw. 12 Aminosäuren. Interessanterweise wurde die antifungale
Wirkung nur bei Kombination beider Laxaphycin-Typen beobachtet. Die Einzelsubstanzen
wiesen keine Aktivität auf. Vergleichbare Ergebnisse wurden für die strukturell verwandten
Lobocyclamide aus Lyngbya confervoides (MacMillan et al., 2002) und Hormothamnine aus
Hormothamnion enteromorphoides (Gerwick et al., 1989) beschrieben. Hormothamnin A
besitzt eine geringe antimikrobielle Wirkung gegen Bacillus subtilis und Pseudomonas
aeruginosa. Es unterscheidet sich strukturell von Laxaphycin A durch die Geometrie der
Doppelbindung der Dehydrobutyryl-Einheit.

Einleitung 12
Neben den Laxaphycinen wurden eine Reihe weiterer zyklischer Peptide mit antifungalen
Eigenschaften beschrieben, wie Scytonemin A aus Scytonema sp. (Helms et al., 1988),
Schizotrin A aus Schizotrix sp. (Pergament et al., 1994), Majusculamid C aus Lyngbya
majuscula (Carter et al., 1984) sowie später aus einem Schwamm der Gattung Pilocaulis
(Williams et al., 1993). Die von Williams beschriebene Struktur lässt eine symbiotische
Ursache vermuten. Majusculamid C wurde zur Bekämpfung von fungalen
Pflanzenpathogenen, wie resistente Arten von Phytophthora infestans und Plasmopora
viticola, patentiert (Moore & Mynderse 1982, US Patent).
Weitere antifungale Wirkstoffe wurden aus Cyanobakterien beschrieben, unter ihnen
Lyngbyabellin B aus Lyngbya majuscula. Es hat antifungale Wirkung gegen Candida
albicans (Milligan et al., 2000). Ebenso wurden zwei neue zyklische Peptide aus Tolypothrix
byssoidea (EAWAG 195), Tolybyssidin A und B, mit moderater antifungaler Wirkung gegen
Candida albicans isoliert (Jaki et al., 2001).
NH
ONH
NH
NH
OH
O
OO
O
NOH
O
NH
ONH
ONH
ONH O
NHNH
O
OH
N
O
NHOH O
NH NH
O
O
NH
OH
O NH
O
NHOH
NH ONH2
O
O
N
O
NH
OO
NH
OH
O
NH2
OH
O
NH
Laxaphycin A Laxaphycin D
Majusculamid C
NO
N
NH
O
O
NH
ONH
ON
ONH
O
O O
O
NH
O
OMe

Einleitung 13
Nicht nur Peptide haben antibiotische Eigenschaften: Fischerellin A aus Fischerella musciola,
hat wachstumsinhibierende Eigenschaften gegenüber anderen Cyanobakterien (MIC 14 nM
gegen Synechococcus PCC 6911, Hagmann et al., 1996). Die Wirkung beruht auf der
Hemmung des Photosystems II. Daneben besitzt Fischerellin A antifungale und herbizide
Aktivität.
1.7.2.2. Zytostatika
Zu den zytostatischen Substanzen gehören die Cryptophycine. Sie wurden erstmals aus
Nostoc sp. ATCC 53787 isoliert (Hirsch et al., 1990, US Patent). Bei den Cryptophycinen
wurde eine potente in vitro Aktivität (ng/ml bis pg/ml) gegen Tumorzellen festgestellt.
Cryptophycine greifen in die Tubulinpolymerisation ein. Sie zählen zu den zyklischen
Depsipeptiden. Ihre Sequenz setzt sich aus jeweils einer δ- und α-Hydroxysäure, sowie einer
α- und β-Aminosäure zusammen. Neben der zytostatischen wurde antifungale Wirkung
beschrieben. Ein aus Nostoc sp. GSV 224 isoliertes Cryptophycin-24 hat identische Struktur
zu Arenastatin A aus dem Schwamm Dysidea arenaria (Kobayashi et al., 1994).
Es gibt viele Sekundärmetabolite, welche auf zytotoxische Eigenschaften untersucht wurden.
In der Hoffnung neue Leitsubstanzen für antitumorale, respektive antivirale Wirkstoffe zu
finden, wurde Mitte der 80iger Jahre wurde ein breit angelegtes Screening auf Zytotoxizität
vom „National Cancer Institute“ durchgeführt (Boyd, 1985). Durch die vielfachen
Veröffentlichungen gehören Zytotoxizitätsassays oft zu den Standarduntersuchungen bei der
Beschreibung einer Bioaktivität. Bei einem kürzlich durchgeführten biologischen Screening
von terrestrischen und Süßwasser-Cyanobakterien auf antimikrobielle Aktivität, „Brine
Shrimp Lethality“ und Zytotoxizität wurden von 22 Proben lipophile und hydrophile Extrakte
hergestellt und untersucht. Zytotoxizität gegen KB-Zellen (human nasopharyngal carcinoma)
R R´ X
Cryptophycin-1 Me H Cl
Cryptophycin-2 Me H H
Cryptophycin-24 H H H
Cryptophycin-52 Me Me Cl
N
N
O
O HFischerellin A
O
NH
ONH
O
OOO
R´R
X
OMe
O

Einleitung 14
wurde für 38,6 % der Extrakte belegt (Mian et al., 2003). Eine weitere Substanz, der viel
Aufmerksamkeit gewidmet wurde, war Dolastatin aus dem Seehasen Dolabella auricularia
(Kap. 1.3.). Unter den Dolastatinen finden sich Strukturen vom Cyanopeptolin-Typ, sowie
weitere zyklische Depsipeptide. Zurzeit befinden sich Dolastatin 10 und Derivate in
verschiedenen Phasen klinischer Studien (de Jonge et al., 2005), unter anderem zum Einsatz
gegen metastasierenden Prostatakrebs (Vaishampayan et al., 2000).
Obyanamid aus dem marinen Cyanobakterium Lyngbya confervoides ist ein zyklisches
Depsipeptid, das auf KB und LoVo (human colorectal adenocarcinoma) Zellen zytotoxisch
wirkt (Williams et al., 2002).
Hapalosin, ebenfalls ein zyklisches Depsipeptid, hat eine MDR-Umkehr („multi drug
resistance“) mit einer Effektivität, welche die von Verapamil übersteigt. Aus diesem Grund
wurden Molekülmodifikationen vorgenommen, um die Effektivität in der Tumortherapie zu
untersuchen (Hermann et al., 2001).
Zytotoxische Effekte wurden auch für die Laxaphycine A und B auf CCRF-CEM (human
lymphoblastic cells) (Gbankoto et al., 2005) und für Wewakpeptine, einem zyklischen
Depsipeptid aus Lyngbya semiplena (Han et al., 2005) beschrieben.
1.7.2.3. Enzyminhibitoren
Unter den biologisch aktiven Substanzen befinden sich viele mit enzyminhibitorischer
Wirkung. Ein erstes Beispiel der später als Cyanopeptoline klassifierten Depsipeptide
(Weckesser et al., 1996) war das erwähnte Dolastatin 13 (Pettit, 1997). Auf Grund
struktureller Ähnlichkeit zu cyanobakteriellen Depsipeptiden geht man heute davon aus, dass
Dolastatin auf cyanobakteriellen Ursprung zurückzuführen ist (Rinehart et al.,1990). Den
vielfältigen Cyanopeptolinen gemeinsam ist ein 19-gliedriger Ring aus 6 Bausteinen. Der
Ring schließt konservierte Aminosäuren ein, sowie Variationsmöglichkeiten an den anderen
Positionen. Des Weiteren ergibt sich die Strukturvielfalt aus Modifikationen in der
Seitenkette. Als charakteristisches Merkmal enthalten die Cyanopeptoline einen Sechserring
im Ringgerüst, ein Piperidinderivat: Die Ahp-Gruppe (3-Amino-6-hydroxy-2-oxo-piperidin)
kann als zyklisches Glutaminsäurealdehyd interpretiert werden. Neben dieser ungewöhnlichen
Gruppe ist der Ringschluss durch eine Esterbindung charakteristisch. Bis auf eine Ausnahme
wird der Ringschluss über die Aminosäure Threonin geknüpft. Bei den Nostopeptinen A und
B befindet sich an dieser Stelle ein Hydroxymethylprolin (Okino et al., 1997). Die
Aminosäuren, die durch Lactonisierung den Ringschluss vermitteln, sind relativ konserviert.
Bisher wurden nur die Aminosäuren Valin oder Leucin an dieser Stelle gefunden (R1). An

Einleitung 15
Position R2 des Ringsystems findet man nur N-methylierte aromatische Aminosäuren. An
Position R4 dominieren basische Aminosäuren mit großer Varianz. Gleiches gilt für die
Position upstream der Ahp-Gruppe. Die für die Ringstruktur verantwortlichen Aminosäuren
liegen alle in der L-Konformation vor. Die Peptidbindungen upstream der Ahp-Gruppe (R2
und R3) sind cis-, alle anderen trans-konfiguriert. Weitere konservierte Strukturmerkmale
sind die Verknüpfung der Seitenkette über die am Ringschluss beteiligte Aminosäure
Threonin in Form einer Acylierung. Der N-Terminus ist durch eine Säure maskiert. Es finden
sich viele strukturelle Unterschiede durch die Varianz der Säure.
In absehbarer Zeit ist eine Novellierung der Nomenklatur zyklischer Peptide zu erwarten
(Moss, 2005). Bis jetzt sind die Vorschläge aber noch nicht durch die IUPAC veröffentlicht
worden, weshalb in dieser Arbeit die Bezeichnung analog zu vorangegangenen Arbeiten
erfolgt.
Bis heute wurden mehr als 70 Varianten der Cyanopeptoline veröffentlicht. Cyanopeptoline
haben je nach strukturellen Eigenschaften Enzymhemmung gegen verschiedene Proteasen:
unter ihnen Elastase, Trypsin, Thrombin, Chymotrypsin, Papain und Plasmin. Es konnte
jedoch nicht immer eine Bioaktivität im Sinne einer Enzymhemmung beschrieben werden.
Neben der Hemmung von Enzymen gibt es Hinweise auf toxische Effekte auf
Primärkonsumenten, wie Daphnien (Jakobi et al., 1995 und Agrawal et al., 2005).
Eine weitere Gruppe der Cyanopeptoline bilden die Microviridine, welche erstmals aus
Microcystis viridis isoliert wurden (Ishitsuka et al., 1990). Beispiele dieser Substanzklasse
wurden als Hemmstoffe der Tyrosinase, andere als Hemmstoffe der Elastase, bzw. des
Chymotrypsins beschrieben (Murakami et al., 1997a). Im Gegensatz zu den Cyanopeptolinen,
welche ebenfalls Serin-Proteaseinhibitoren sein können, sind es Tetradecapeptide mit
N
O
NHO
O
O
NH
O
NH
ONH
N
OO
NH O
OH
OH
OHO
R4
Ahp
R 3
X - L-Thr R 2
O R 1
Schematischer Aufbau eines Cyanopeptolins Cyanopeptolin 963A

Einleitung 16
trizyklischer Struktur. Microviridin J wirkt als Proteaseinhibitor im Häutungszyklus von
Daphnia sp. mit letaler Folge (Rohrlack et al., 2004).
Anabaenopeptin A wurde erstmalig als Nebenkomponente bei Anabaena flos-aquae NRC
525-17 gefunden (Harada et al., 1995). Dieses Peptid, bestehend aus zyklischen Strukturen,
hat als Besonderheit einen Ringschluss, welcher durch die ϖ-Aminogruppe eines Lysins und
der Carboxygruppe eines Phenylalanins vermittelt wird. Charakteristisch ist die Verknüpfung
der Seitenkette mit dem Ringgerüst über eine Ureido-Funktion.
Neben den zyklischen Peptiden aus Cyanobakterien gibt es unter den Proteasehemmstoffen
auch lineare Strukturen, wie in der Gruppe der Aeruginosine. Sie wurden erstmals aus
Microcystis aeruginosa isoliert (Murakami et al., 1994). Aeruginosine weisen ein breites
Wirkungsspektrum gegen unterschiedliche Serinproteasen auf. Besonders ausgeprägt ist die
Inhibition von Thrombin durch Aeruginosin 102-A mit einem IC50-Wert von 0,04 µg/ml
(Matsuda et al., 1996). Charakteristisch ist der Azabizyklus, welcher auf ein Octahydroindol
zurückgeht. Chemisch zählen die Aeruginosine zu den natürlichen Guanidinderivaten.
1.7.2.4. Weitere Substanzklassen
Das Interesse an Bluthochdruck- und Stärkungsmitteln für das Herz ist verständlicherweise
groß. Terrestrische Anabaena sp. produzieren ein chloridhaltiges zyklisches Dekapeptid,
Anabaenopeptin F
NH
NHO
N O
NH
OO
NH
O NH
NH
OO OH
NH NH2
NH
OH
Aeruginosin 98-B
N
ONH
ONH
O
OH
OH
NH NH
NH2
HO3SO
H
H

Einleitung 17
Puwainaphycin C, welches inotrope Wirkung, ohne chronotrope Begleiteigenschaften zu
zeigen, aufweist. Die ED50 von Puwainaphycin liegt bei 0,2 ppm. Die Strukturanaloga
Puwainaphycin A, B, D und E, welche als Nebenprodukte isoliert wurden, besitzen
verringerte Aktivität (Moore et al., 1989; Gregson et al., 1992).
Scytonemin A (Struktur siehe Kap. 1.7.2.4.), ein Hauptmetabolit aus Scytonema sp. (strain U-
3-3) hat Calcium-antagonistische Eigenschaften. Microginine, lineare Peptide aus Microcystis
aeruginos wurden als Inhibitoren von ACE („angiotensin-converting-enzyme“) mit IC50-
Werten von 7µg/ml beschrieben (Okino et al., 1993).
Zwei weitere lineare Lipopeptide wurden aus Lyngbya majuscula isoliert. Die Microcoline A
und B haben starke immunsuppressive Aktivität und sind stärker wirksam in der
Unterdrückung der Lymphozyten-Proliferation als Cyclosporin A (Koehn et al., 1992; Zhang
et al., 1997).
Scytonema hofmanni PCC 7110
Die Gattung Syctonema gehört in die Ordnung der Nostocales und in die Familie der
Scytonemataceae (Komarek und Anagnostidis, 1989). Die Trichome von Scytonema sind
isopolar und mit einer Hülle umschlossen. Des Weiteren zeichnet sie sich durch das
Vorhandensein von falschen Verzweigungen aus. Scytonema besitzt Heterocysten und kann
Luftstickstoff fixieren.
Abb. 1: a: schematische Darstellung einer falschen Verzweigung b: Trichom von Scytonema: falsche Verweigung und Heterozyste (verdickte, bräunliche Zelle).
Microcolin A
NN
N
O
O
NH
OAc
N
O
O OO
OH
a) b)

Einleitung 18
Scytonema Arten kommen vor allem terrestrisch vor. Sie formen raue, filzige Matten auf
feuchten Felsen, Holz und Boden (Bold und Wynne, 1985). Man findet Besiedelung auch im
Periphyton von Seen (Hawes et al., 1994) oder entlang der Meeresküste (unter anderem in
den Everglades bei Florida).
Aus Scytonema hofmanni PCC 7110 wurden bisher drei peptidische Substanzen
charakterisiert (Matern et al., 2001 und 2003). Es handelt sich um zyklische Peptide vom
Cyanopeptolin-Typ. Die Scyptoline (Scyptolin A und B) besitzen ein identisches Ringgerüst
und eine Variation in der Seitenkette. Bei Hofmannolin sind sowohl Ringgerüst, als auch die
Seitenkette modifiziert.
Für eine weitere Substanz wurde ein Strukturvorschlag anhand der Fragmentspektren der
Massenbestimmung gegeben. Weiterführende Versuche zur Strukturaufklärung und
Bestimmung der Bioaktivität von P12,8 wurden von Matern (2002) nicht unternommen.
Strukturvorschlag P12,8
N
O
NHO
O
O
NH
O
NH
NH
OH
N
OO
NH O
OHOH
OH
O
OH O
NH
NH2
NH
N
O
NHO
O
O
NH
O
NH
ONH
OH
N
OO
NH O
OH
Cl
OH
OR
NH
O
N
O
NH
OH
O
O
O
NH
ONH
OH O
OOH
NH
OH
N
OO
NH O
OH
O
ONH
O
R
Scyptolin A und B (+R) Hofmannolin

Einleitung 19
Weder den Scyptolinen A und B, noch Hofmannolin konnte in verschiedenen Testsystemen
antimikrobielle Wirksamkeit attestiert werden. Bei der Prüfung auf enzyminhibitorische
Wirkung auf Proteasen wurde für die Scyptoline eine signifikante Hemmung der Serin-
Protease Elastase (IC50 von 0,16 µg/ml) beschrieben. In geringem Umfang (IC50 > 500 µg/ml)
wurde auch eine Beeinflussung von Trypsin und der Cystein-Protease Papain beobachtet,
wohingegen keine Wirkung auf die Serin-Proteasen Thrombin und Plasmin erhalten wurde.
Die enzyminhibitorische Wirkung auf Elastase wurde durch kristallographische
Untersuchungen auf molekularer Ebene beschrieben (Matern et al., 2003).
Neben den durch Matern et al. (2001 und 2003) beschriebenen Cyanopeptolinen gibt es nur
wenige Veröffentlichungen, welche die Gattung Scytonema als Untersuchungsgegenstand
wählten: Neben den Cyanopeptolinen wurden aus Scytonema sp. antibiotisch wirksame
Substanzen beschrieben: Scytonemin A aus Scytonema sp. (strain U-3-3) hat nur schwache
Aktivität gegen ein breites Spektrum an Bakterien und Pilzen. Daneben besitzt es stark
calciumantagonistische Eigenschaften und nur schwache Zytotoxizität auf CCRF-CEM-
Zellen (Helms et al., 1988).
Eine weitere antibiotisch wirksame Substanz aus Scytonema sp. TISTR 8208 ist von
Chetsumon et al. (1995) beschrieben worden: ein zyklisches Peptid mit einem breiten
Wirkspektrum gegen sowohl gram-positive und gram-negative Keime, ebenso gegen
pathogene Hefen und Dermatophyten. Eine Struktur wurde nicht veröffentlicht, aber anhand
erster NMR-Daten wurde Ähnlichkeit zu Scytonemin A vermutet.
Scytonemin A
NOO
NHOH
NHO
ONH
OH
O
NH
O
OH
O
OHO
NH
NH
O
O
NH
OH
ONH
O
N
N
O
OH
OH
O NH

Einleitung 20
Als weiterer bioaktiver Stoff wurde Cyanobacterin aus Scytonema hofmanni isoliert (Gleason
et al., 1991). Es findet Verwendung als Herbizid, bzw. Algizid. Aus Scytonema varium wurde
ein potentes anti-HIV-Protein isoliert. Scytovirin bindet spezifisch an das hoch glycosylierte
virale Hüllprotein gp120, gp160 und gp41 des HI-Virus, jedoch nicht an den zellularen CD4
Rezeptor oder andere getestete Proteine (Bokesch et al., 2003). Makrolide, wie Scytophycine
und Tolytoxine, isoliert aus Scytonema sp., haben zytotoxische und antimykotische
Eigenschaften (Ishibashi et al., 1986 und Carmeli et al., 1990).
O
O
OH O
O
OMe
OMe
OH O OMe
OMeN H
O
Scytophycin A

Einleitung 21
Zielsetzung
Ziel der vorgelegten Arbeit war es, axenisch kultivierte Zellen des Cyanobakterienstamms
Scytonema hofmanni PCC 7110 hinsichtlich der Expression an Sekundärstoffwechsel-
produkten näher zu charakterisieren. Neben den aus diesem Stamm bereits bekannten
Peptiden Scyptolin A und B sowie Hofmannolin beobachtet man in HPLC-
Chromatogrammen bisher nicht identifizierte Signale, deren Strukturen aufgeklärt werden
sollten. Auf Grund ähnlicher Retentionszeiten und UV-Spektren wurden zum einen
Cyanopeptolin-Strukturen unter den Signalen vermutet, zum anderen schloss die Analyse
Peaks ein, deren Retentionzeiten und UV-Spektren dies nicht zwingend vermuten ließen. Für
die Strukturaufklärung waren die Peptide in präparativer Menge zu isolieren und mit den
derzeit verfügbaren chemischen Methoden zu charakterisieren.
Es sollte die bereits für Cyanopeptoline beschriebene inhibitorische Wirkung auf Serin- und
Cysteinproteasen geprüft werden. Auch sollten zuvor nicht geprüfte Enzymhemmtests
eingesetzt werden, darunter Metalloproteasen. Der Fokus sollte auch mögliche antimikrobielle
Aktivitäten einschließen. Des Weiteren sollten die neu isolierten Stoffe, wie auch die
Scyptoline und Hofmannolin auf Zytotoxizität geprüft werden. Bei positivem Befund sollten
weiterführende, auf den Wirkungsmechanismus zielende Versuche, wie z. B. Testung auf
Apoptose, Porenbildung, Membranschädigung etc. durchgeführt werden. Mit der Testung auf
Toxizität gegenüber Primärkonsumenten aus der Klasse der Crustaceen (z. B.
Thamnocephalus platyurus und Daphnia magna) sollte in der vorgelegten Arbeit auch ein
Beitrag zur möglichen Bedeutung der cyanobakteriellen Peptide geleistet werden, da sich die
Befunde mehren, dass diese Sekundärmetabolite eine ökologische Funktion haben.

Material und Methoden 22
2. Material und Methoden
2.1. Kultivierung von Scytonema hofmanni PCC 7110
2.1.1. Anzucht und Kulturmedium Zur Kultivierung von Stamm- und Batchkulturen von Scytonema hofmanni PCC 7110 wurde
BG 11-Medium (Tabelle 1) verwendet. Ausgehend von den Stammlösungen wurden
Konzentrate für jeweils 5 l BG11-Medium hergestellt und portionsweise bei -20 °C
eingefroren.
Tabelle 1: Zusammensetzung des BG11-Mediums (Rippka et al., 1979) und der Stammlösung Komponenten Konz. (g/l) Konz. (mM) Stammlösung (g/l) NaNO3 1,5 17,7 150 K2HPO4 • 3H2O 0,04 0,18 40 MgSO4 • 7 H2O 0,075 0,3 75 CaCl2 • 2H2O 0,036 0,25 36 Citronensäure 0,006 0,03 6 Ammoniumeisen-(III)-citrat 0,006 0,03 6 Na2-EDTA 0,001 0,0027 1 Na2CO3 0,02 0,38 20 Spurenelement-Lösung 1 ml - - Tabelle 2: Spurenelemente-Stammlösung Komponenten Komz. (g/l) H3BO3 2,860 MnCl2 • 4 H2O 1,810 ZnSO4 • 7 H2O 0,222 Na2MoO4 • 2 H2O 0,390 CuSO4 • 5 H2O 0,079 Co(NO3)2 • 6 H2O 0,0494 Zum Herstellen der Medien der Batch-Kulturen wurden die Aliquote aufgetaut und mit Aqua
dest. ad 7 l aufgefüllt und auf pH 7,8 eingestellt. Anschließend wurden die Medien bei 121 °C
3 h autoklaviert. Nach Abkühlen wurden die Medien unter sterilen Bedingungen mit den
Vorkulturen angeimpft. Die Anzucht erfolgte photoautotroph als Flüssigkultur bei 26 °C unter
24 h Beleuchtung mit Weißlicht (1.000 Lux). Um ausreichende Belüftung zu gewährleisten,
wurde das Kulturmedium mit Druckluft (ca. 0,5 bar) begast. Zusätzlich wurden die Kulturen
mittels Magnetrührer (Ikamag Ret, Staufen) durchmischt. Bei den Vorkulturen wurde die
Durchmischung durch Verwendung eines Schütteltischs (Swip, Edmund Bühler) mit
100 Upm gewährleistet, während auf eine zusätzliche Belüftung verzichtet wurde. Die
Kulturen wurden unter den genannten Bedingungen für 8-10 Wochen im Brutraum kultiviert.

Material und Methoden 23
2.1.2. Ernte der Kulturen und Lagerung Dank der fädigen Morphologie von Scytonema hofmanni PCC 7110 war es möglich die
Biomasse nach 8-10 wöchigem Wachstum über ein Planktonnetz mit der Maschengröße
55 µm zu ernten. Die feuchte Biomasse wurde bei -20 °C tiefgefroren und lyophilisiert
(Lyovac 2, Leybold-Heraeus, Hanau). Die getrockneten Zellen wurden bei -20 °C bis zur
weiteren Bearbeitung gelagert.
2.2. Peptid-Isolierung aus Scytonema hofmanni PCC 7110
2.2.1. Extraktion Das Extraktionsschema wurde anfangs unverändert zu Matern et al. (2001) angewandt. Da
jedoch Peptide mit unterschiedlichen Substanzeigenschaften isoliert werden sollten, wurde
das Extraktionsschema abgewandelt. Nach Vorversuchen wurden einige Parameter geändert,
so dass sich folgender Extraktionsweg herauskristallisierte:
Lyophilisiertes Zellmaterial (ca. 6-10 g) wurde in einem Mörser fein pulverisiert und in einen
Messbecher überführt. Die Zellen wurden 15 min mit ca.150 ml 50 % (v/v) Methanol, gefolgt
von weiteren zwei Extraktionschritten mit jeweils 100 ml 50 % (v/v) Methanol, extrahiert.
Nach jedem Extraktionsschritt wurde bei 9.000 U/min 20 min zentrifugiert (Sorvall RC-5B,
Rotor: GSA; DuPont Instruments, Bad Homburg). Die Überstände wurden dekantiert,
vereinigt und über eine Porzellanfritte mit doppeltem Papierfilter (Schleicher & Schuell, 595
Rundfilter, Durchmesser 110 mm) filtriert. Das Filtrat wurde nochmals bei 15.000 U/min 20
min (Rotor: SS34) zentrifugiert. Der Extrakt wurde bis zur weiteren Verarbeitung bei 4 °C
gelagert.
2.2.2. Festphasenchromatographie Zur Vorbereitung der C18-Chromatographie wurde 20 g pulverförmiges C18-Material in
200 ml 90 % (v/v)Methanol suspendiert und ca. 3 h zum Quellen stehen gelassen. Das
gequollene Säulenmaterial wurde in eine Glassäule (Sigma, Taufkirchen; Länge 25 cm,
Durchmesser 2,8 cm) gefüllt und mit einem Säulenvolumen 90 % (v/v) Methanol gewaschen
und mit zwei Säulenvolumina 10 % (v/v) Methanol equilibriert. Der Rohextrakt wurde auf
10 % bis 20 % (v/v) Methanol eingestellt und auf die Säule aufgetragen. Zusätzlich wurde
Vakuum (Vaccubrand GmbH & Co KG, Wertheim) an die Säule angelegt, um einen

Material und Methoden 24
gleichmäßigen Durchfluss zu gewährleisten. Der Durchbruch wurde mittels HPLC-Analytik
auf etwaige Peptide getestet. Bei Abwesenheit von Peptiden konnte der Durchbruch
verworfen werden. Durch drei Elutionsschritte wurden die an das Säulenmaterial gebundenen
Stoffe eluiert. Dazu wurden 30 %, 60 % und 90 % (v/v) Methanol verwendet, die Eluate
wurden jeweils separat gesammelt. Um die Qualität der Extraktion, wie auch der Elution
beurteilen zu können, wurden Proben genommen und mittels HPLC-Analytik untersucht.
Proben mit einem Methanolgehalt größer 50 % wurden mit Aqua dest. 1:2 verdünnt. Alle
Proben wurden mit einem Membranfilter (Chromafil PET, Durchmesser 0,45 µm, Macherey
Nagel, Düren) filtriert und in HPLC-Vials überführt.
Abb. 2: Extraktionsschema zur Isolierung von Peptiden aus Scytonema hofmanni PCC 7110
2.3. Hochleistungsflüssigchromatographie
2.3.1. Analytische Umkehrphasenchromatographie Die verschiedenen Eluate, der Durchbruch sowie bereits isolierte Peptide wurden mittels RP-
HPLC analysiert und auf Reinheit geprüft. Dazu wurden die isolierten, getrockneten Peptide
wieder in wenig 50 % (v/v) Methanol aufgenommen, die Proben der Extraktion wurden bei
einem Methanolgehalt > 50 % mit Aqua dest. 1:2 verdünnt. Alle Proben wurden mit einem
Membranfilter (Chromafil PET, Durchmesser 0,45 µm, Macherey Nagel) filtriert und
LyophilisiertesZellmaterial
MethanolischerRohextrakt
Umkehrphasen -Chromatographie
30% MeOH 50% MeOH 75% MeOH 90% MeOH
HPLC - Analytik
50% Methanol
Elution mit
Wasch-Lösung: keine Peptide
Hofmannolin, Cyanopeptolin 1042A
Scyptolin A, B, Cyanopeptoline 946A, 1067A
Scytocyclamide A, B, C
Analytisch / Semipräparativ / Präparativ
Material und Methoden 25
zusätzlich bei 14.000 U/min 5 min zentrifugiert (Eppendorf Centrifuge 5417R, Hamburg). Für
einen analytischen HPLC-Lauf wurden 50 µl Probe mit Hilfe eines Autosamplers (Waters
717, Eschborn) auf eine RP-HPLC-Säule (SP 250/4 Nucleosil 100-5 C18, bzw. CC250/4
Nucleodur C18 Gravity, 5 µm, Macherey-Nagel, Düren) aufgetragen. Die gesamte aus
Einzelmodulen aufgebaute HPLC-Anlage musste vor Beginn der Analysen konditioniert und
programmiert werden.
Zur Konditionierung der Säule, wie auch für die Analysen wurden Gradienten nach Lawton et
al. (1994) benutzt (Tabelle 3). Es wurde jeweils eine Methodenprogrammierung für die
Konditionierung (Lawton Ende MS) und eine für die Analyse (Lawton Anfang MS) benutzt.
Das Laufmittel bestand aus den Komponenten Aqua bidest. (A), Acetonitril (B) und
Acetonitril mit 0,5 % Trifluoressigsäure (C). Um eventuelle Analysenrückstände aus der
Säule zu spülen, wurden die Säulen in wechselnden zeitlichen Abständen zusätzlich mit
100 % Methanol gespült.
Tabelle 3: Gradientenzusammensetzung für HPLC-Läufe nach Lawton et al. (1994) Lawton Anfang MS (Method Set)
Zeit (min) Flussrate (ml / min) A % B % C % curve
0 1 70 20 10 0
10 1 65 25 10 6
40 1 30 60 10 6
42 1 0 90 10 6
Lawton Ende MS (Method Set)
Zeit (min) Flussrate (ml / min) A % B % C % curve
0 1 0 90 10 0
1 1 70 20 10 6
9 1 70 20 10 6
2.3.2. RP-HPLC im semipräparativen und präparativen Maßstab Um die gewünschten Peptide in veritablen Mengen isolieren zu können, musste ein scale-up
auf größere Injektionskapazitäten vorgenommen werden. Die peptidhaltigen Fraktionen der
C18-Eluate wurden dazu zunächst im Rotationsverdampfer (Rotavapor, Büchi, Flawil)
eingeengt und nochmals bei 15.000 U/min 10 min zentrifugiert (Sorvall RC-5B, Rotor: GSA,
DuPont Instruments, Bad Homburg). Vor der Auftrennung wurden die Proben zusätzlich
durch einen Membranfilter (Chromafil PET, Durchmesser 0.45 µm, Macherey-Nagel, Düren)

Material und Methoden 26
filtriert. Für die verschiedenen Trennläufe wurden die Proben mit Hilfe eines Autosamplers
(Waters 717, Eschborn) auf eine RP-HPLC-Säule (SP 250/10 Nucleosil 100-7 C18, bzw. SP
250/21 Nucleosil 100-7 C18, Macherey-Nagel, Düren) aufgetragen. Je nach Säulenart wurde
die Flussrate ausgewählt. Bei semipräparativer Auftrennung (SP 250/10-Säule) betrug sie
5 ml/min und bei präparativen Bedingungen 9 ml/min. Je nach Trennleistung der Säulen
musste die Fließmittelzusammensetzung variiert werden. Des Weiteren waren zusätzliche
Reinjektionen notwendig sofern nach Vereinigung der jeweils gleichen Peptide keine
Reinsubstanzen erhalten wurden, oder aber wenn zuerst eine Vortrennung vorgenommen
wurde. Als Fließmittel wurden Aqua bidest., Acetonitril, sowie Acetonitril mit 0,5 % (v/v)
TFA in unterschiedlichen Mengenverhältnissen eingesetzt. Die Menge an Acetonitril mit
0,5 % (v/v) TFA war als einzige konstant und betrug zu jeder Zeit 10 % (v/v) an der
Fließmittelzusammensetzung. Dies gewährleistete, dass der Gehalt an TFA konstant bei
0,05 % (v/v) lag. Die HPLC-Läufe wurden sowohl unter isokratischen Bedingungen als auch
unter einem Gradienten durchgeführt. Um die optimalen Trennbedingungen zu finden,
wurden die peptidhaltigen Fraktionen zuerst einem Gradientenlauf unterzogen, dessen
einziger Unterschied zur analytischen Auftrennung in der Programmierung der Flussrate sich
widerspiegelte. Anhand der erhaltenen Chromatogramme wurden die neuen
Trennbedingungen festgelegt. Entweder bediente man sich eines Gradienten oder man wählte
eine isokratische Fließmittelzusammensetzung analog der Retentionszeit weniger 5-7 % des
organischen Anteils des Fließmittels. Bei unzureichender Reinheit war bei Reinjektion
meistens eine isokratische Fließmittelzusammensetzung gewählt worden, die je nach
Trennanforderung jedes Mal neu bestimmt werden musste.
Die einzelnen Peptidfraktionen wurden im „peak mode“ mit einem Fraktionensammler
(Waters Fraction Collector, Eschborn) gesammelt. Beispielhaft werden die angewendeten
Gradienten zur Fließmittelbestimmung, sowie die zur Vortrennung genutzten
Programmierungen angegeben (Tabelle 4).
Tabelle 4: Gradient für semipräparative Läufe Lawton Anfang 5ml MS (Method Set)
Zeit (min) Flussrate (ml / min) A % B % C % curve
0 5 70 20 10 0
10 5 65 25 10 6
40 5 30 60 10 6
42 5 0 90 10 6

Material und Methoden 27
Diese Fraktion wurde jedoch nicht ausschließlich über eine Gradienten-Trennung
aufgearbeitet. Vielmehr wurden auch isokratische Fließmittelzusammensetzungen eingesetzt
(A: 60 %, B: 30 %, C: 10 % oder aber A: 50 %, B: 40 %, C: 10 %). Es gab keine allgemein
verlässliche Programmierung.
Tabelle 5: Gradient für C18 Eluat mit 75 % (v/v) Methanol C18E75_Gradient MS (Method Set)
Zeit (min) Flussrate (ml / min) A % B % C % curve
0 5 60 30 10
5 5 70 20 10 11
20 5 30 60 10 6
Tabelle 6: Gradient für C18 Eluat mit 90 % (v/v) Methanol C18E90_Gradient MS (Method Set)
Zeit (min) Flussrate (ml / min) A % B % C % curve
0 5 60 30 10 0
10 5 40 50 10 6
15 5 20 70 10 6
25 5 20 70 10 6
2.4. Identifizierung der Peptide und Reinheit Die einzelnen Peptide wurden anhand der Retentionszeiten im Chromatogramm zugeordnet.
Bei Verschiebung der Retentionszeiten, z. B. durch nachlassende Trennbodenzahl der Säule,
wurde versucht die Zuordnung anhand Peakhöhe, Peakform und anhand der aufgenommen
UV-Spektren vorzunehmen (Maxima, Minima und Aussehen). War eine genaue Zuordnung
nicht möglich, so wurde entweder eine Massenbestimmung der gesammelten Fraktion
veranlasst, oder man verglich die gesammelte Fraktion mit bereits eindeutig identifizierten
Peptiden.
2.4.1. Co-Chromatographie (Co-Elution) Um Reinsubstanzen mit bereits isolierten Substanzen ähnlicher Retentionszeit vergleichen zu
können, bedient man sich der Co-Chromatographie (Co-Elution). Hierzu werden zunächst in
direkt aufeinander folgenden getrennten Läufen die Retentionszeiten von Probe und Referenz
verglichen. Stimmen die Retentionszeiten im Rahmen der üblichen Verschiebung überein,

Material und Methoden 28
erstellt man eine Mischung aus Probe und Referenz, wobei beachtet werden muss, dass beide
Lösungen im selben Konzentrationsbereich liegen. Andernfalls kann eine Überlagerung der
Signale stattfinden und eventuelle Unterschiede in den Retentionszeiten überdeckt werden.
Die Auswertung der so co-eluierten Proben erfolgt einmal anhand der Peaksymmetrie und
zum anderen mittels UV-Spektren-Vergleich von Peakschultern und Peakmaximum. Wenn
der Peak des Gemischs symmetrisch ist und nur eine deutliche Spitze aufweist und wenn auch
die UV-Spektren sich decken, so liegen bei den beiden untersuchten Proben die gleichen
Verbindungen vor.
2.5. Strukturaufklärung der isolierten Substanzen
2.5.1. Ultraviolett-Spektroskopie UV-Spektren der gereinigten Substanzen wurden spektrometrisch (Varian Cary 100 Bio UV-
visible Spectrophotometer, Cary, USA) im Wellenlängenbereich von 195 bis 350 nm
aufgenommen. Zur Messung wurden Quarzküvetten (Hellma, Müllheim) mit der Schichtdicke
von 1 cm verwendet. Die Substanzen wurden in Methanol p.a. gelöst und auf eine
Konzentration von 1 mg/ml eingestellt. Als Referenz wurde Methanol p.a. verwendet.
2.5.2. Infrarotspektroskopie Molekülschwingungen und -rotationen werden durch Absorption von Strahlung im infraroten
Bereich des elektromagnetischen Spektrums angeregt. Es gibt zwei Möglichkeiten, solche
Molekülschwingungen oder -rotationen zu messen:
- direkt als Absorption im Infrarot-Spektrum oder
- indirekt als Streustrahlung im Raman-Spektrum
Im Institut für Pharmazeutische Wissenschaften (Universität Freiburg) wurden Messungen an
einem Fourier-Transform-IR-Spektrometer (Spectrum One FTIR-Spectrometer, Perkin-Elmer,
Boston) direkte Absorptionsmessungen vorgenommen. Die einzelnen Peptide wurden in
lyophilisierter Form auf einen Diamantkristall aufgetragen und im Transmissionsmodus
vermessen. Die Spektren wurden über einen Wellenlängenbereich von 4.000 bis 650 cm-1
aufgezeichnet.

Material und Methoden 29
2.5.3. Massenspektroskopie Die MS-Analysen wurden zum einen von Dr. M. Erhard (AnagnosTec GmbH, Luckenwalde),
zum anderen in der Arbeitsgruppe von Prof. W. Haehnel (Universität Freiburg, Institut für
Biologie II) zusammen mit Frau B. Knapp sowie zusammen mit Frau Dr. J. Blom an der
Limnologischen Station der Universität Zürich durchgeführt. Hieraus ergaben sich
verschiedene Durchführungsprotokolle.
Im Folgenden wird die Vorgehensweise bei der Analyse mittels Matrix unterstützter Laser
Desorptions/Ionisations-Flugzeit-Massenspektroskopie (MALDI-TOF-MS) beschrieben.
Für MALDI-TOF-MS wurden die Peptide in 10 µl Aqua dest./Ethanol/Acetonitril (1/1/1)
gelöst. Alle Proben wurden in einer gesättigten α-Cyano-4-hydroxyzimtsäure-Matrix (gelöst
in 50 % Acetonitril mit 0,03 % TFA(v/v)) analysiert. Je 1 µl Matrix wurde mit 1µl Probe
gemischt und direkt auf die Platte aufgetragen. Das Molekulargewicht der Peptide wurde mit
einem „Maldi Voyager Elite time-of-flight Massenspektrometer“ (PerSeptive BioSystems,
Farmingham, MS) ermittelt. Das Licht des N-Lasers hatte eine Wellenlänge von 337 nm, die
Beschleunigungsspannung betrug 20 kV. Die Messungen wurden im „delayed extraction
mode“ durchgeführt und erlaubten so die Bestimmung der monoisotopischen Massen. Ein
Filter für Massen < 400 verbesserte die Messungen, indem die stärksten Matrix-Ionen
ausgeblendet wurden. Die Spektren wurden in der Betriebsart für Erfassung und Beugung
positiver Ionen aufgezeichnet („positive ion detection mode“ und „reflector mode“). Für die
„post-source-decay“ (PSD)-Messungen wurde die Reflektorspannung schrittweise
herabgesetzt. Es wurden zwölf spektrale Segmente gemessen und danach addiert (Erhard et
al., 1999).
2.5.4. Elektrospray-Ionisations-Massenspektrometrie Alle Versuche mit Elektrospray-Ionisations-Massenspektrometrie wurden an einem Thermo
Finnigan LTQ-FT-Massenspektrometer (Thermo Electron Corporation, Bremen) mit ESI-
Quelle durchgeführt. Die Ionisierung erfolgte über Elektrospray, die Detektion über eine
lineare Ionenfalle kombiniert mit einem Fourier Transform Ion Cyclotron Resonanz
Massenspektrometer (FTICR-MS; supraleitender Magnet mit 7 Tesla). Die gereinigten
Peptide wurden in 70 % (v/v) Methanol mit 0,1 % Essigsäure gelöst (0,01 mg/ml). Die
Flussrate wurde auf 5 µl/min eingestellt.

Material und Methoden 30
2.5.5. Magnetische Kernresonanzspektroskopie Alle NMR-Messungen wurden von L. Oberer (Novartis Pharma AG, Basel) durchgeführt. Die
Spektren wurden auf einem Bruker Avance (Bruker, Rheinstetten) DMX 500, bzw. DRX 500
Spektrometer bei 300 K, 500 MHZ und 11.746T sowie auf einem Bruker Avance DRX 600
bei 295 K, 600 MHZ und 14.095T aufgezeichnet, die mit selektiven TXI-Probenköpfen für 1D-13C Spektren und dreifach inversen Probenköpfen für die 1D-1H und 2D Spektren
ausgestattet sind. Die Peptide wurden in DMSO-d6 gelöst (< 2 mg/0,5ml), 1H und 13C
Resonanzverschiebungen wurden gegen DMSO-d6 (2,49 ppm bzw. 39,9 ppm) bestimmt. Es
wurden folgende NMR Experimente durchgeführt: 1D-1H, 1D-13C, 1H-1H-COSY, 1H-1H-
ROESY, 1H-1H-TOCSY, 1H-13C-COSY (HSQC).
2.5.6. Gaschromatographie-Massenspektrometrie-Messungen Zur Bestimmung der Konfiguration der Aminosäuren in Scytocyclamid A wurden 100 µg des
gereinigten Peptids und äquivalente Mengen an Aminosäurestandards mit 100 µl 3 M HCl-
Gas in Methanol (Supelco; Bellefonte, PA, USA) bei 110 °C über 24 h verestert. Die
Reaktionsgemische wurden getrocknet und mit Trifluoressigsäureanhydrid (TFAA; Fluka,
Schweiz) acyliert bei 80 °C über 1 h. Die gaschromatographischen Analysen wurden mit einer
Chirasil-Val-Säule (Permabond®-L-Chirasil-Val; 25m x 0,25mm; Macherey-Nagel, Düren)
durchgeführt. Die Trennung erfolgte 2 min bei 80 °C, gefolgt von einem
Temperaturgradienten von 80 °C auf 180 °C bei 8 °C min-1, sowie 10 min bei 180 °C. Die
Zuordnung der Enantiomeren erfolgte über Vergleich der Retentionzeiten mit denen der
Aminosäurestandards.
2.5.7. Konfigurationbestimmung der Aminosäuren Die Bestimmung erfolgte nach Marfey (1984). Zur Analyse der Aminosäuren wurden 100 µg
Peptid zunächst mit 6 N HCl bei 110 °C innerhalb 24 h hydrolysiert. Das resultierende
Aminosäurengemisch aus 100 µg Scytocyclamid A wurde mit 50 µl einer Lösung (10 mg /
ml) von 1-Fluor-2,4-Dinitrophenyl-5-L-Alaninamid (L-FDAA, Novabiochem, Läufelfingen,
Schweiz) in Aceton versetzt. Nach Zugabe von 100 µl 1M NaHCO3 wurde die Lösung 3 min
bei 80 °C erhitzt. Nach Abkühlen auf Raumtemperatur wurden 120 µl einer 1 M HCl und 300
µl 50 % (v/v) Acetonitril zugegeben. Die so derivatisierten Proben wurden auf einer C-18
Supelco-Sil ODS Umkehrphasensäule (4,6 x 250 mm; Supelco; Bellefonte, PA, USA)

Material und Methoden 31
untersucht. Ein weiterer Versuch wurde mit D-FDAA (D-FDAA, Novabiochem,
Läufelfingen, Schweiz), wobei die Probenaufbereitung und Versuchsdurchführung
beibehalten wurde.
2.6. Enzymhemmtests Die isolierten Peptide wurden in 50 % (v/v) MeOH gelöst (1 µg/µl) und für ein erstes
Aktivitäts-Screening in einer 1:100 Verdünnung eingesetzt. Bei einer nennenswerten
Hemmung wurden die Verdünnungsstufen angepasst, um eine IC50-Bestimmung
vorzunehmen. In diesen Fällen wurden anstatt Einzelbestimmungen Mehrfachbestimmungen
(dreifach, beziehungsweise vierfach Bestimmungen) durchgeführt. Die einzelnen
Verdünnungsstufen wurden mit den jeweiligen Enzymen inkubiert, mit Substrat versetzt, die
Änderung der Absorption wurde mit einem ELISA Reader (MR 5000 Elisa Reader, Dynatech,
Chantilly, Virginia) verfolgt. Alle Testansätze wurden im Mikrotiterplatten-Maßstab
durchgeführt. Des Weiteren wurden in den Testserien Positivkontrollen mit 50 % (v/v) MeOH
an Stelle der Peptidlösung und Blindproben ohne Enzym mitgeführt. Zur Überprüfung der
Funktionalität der entsprechenden Tests wurden zu Beginn der Untersuchungen käufliche
Inhibitoren der Enzyme eingesetzt.
2.6.1. Trypsin Trypsin (EC 3.4.21.4, Sigma, Taufkirchen) wurde in 1 mM HCl mit 20 mM CaCl2 (pH 3,1,
0,1 mg/ml) gelöst. 10 µl dieser Lösung wurden mit 10 µl der jeweiligen Peptidverdünnung
und 130 µl Puffer (0,1 M Tris-HCl, pH 8,2, mit 23 mM CaCl2) 10 min bei 25 °C vorinkubiert.
Danach wurden 150 µl Substrat (BAPNA, 0,3 mg/ml in Wasser) zugesetzt. Nach weiteren
3 min Inkubation wurde die Absorption bei 405 nm gemessen (Elfant et al., 1985).
2.6.2. α–Chymotrypsin α–Chymotrypsin (EC 3.4.21.1-2, Sigma, Taufkirchen) wurde in 100 mM Tris-HCl mit 10 mM
CaCl2 (pH 7,8, 0,2 mg/ml) gelöst. 50 µl dieser Lösung wurden mit 10 µl der jeweiligen
Peptidverdünnung und 190 µl Puffer (0,1 M Tris-HCl, pH 7,8, mit 10 mM CaCl2) 10 min bei
25 °C inkubiert. Danach wurden 50 µl Substratlösung zugegeben. Nach weiteren 10 min
Inkubation wurde die Absorption bei 405 nm gemessen (Delmar et al., 1979).

Material und Methoden 32
2.6.3. Plasmin Der Test wurde in modifizierter Form nach Lottenberg et al. (1981) durchgeführt. Plasmin
(EC 3.4.21.7, Sigma, Taufkirchen) wurde in einem Tris-Puffer (0.05 M Tris-HCl, pH 8.9,
0,5 mg/ml) gelöst. 10 µl dieser Lösung wurden mit 10 µl der jeweiligen Peptidverdünnung
und 170 µl Puffer 10 min bei 25 °C inkubiert. Danach wurde 10 µl Substrat (N-p-Tosyl-Gly-
Pro-Lys-p-nitroanilid, 1,1 mg/ml in Wasser) zugesetzt. Nach weiteren 3 min Inkubation
wurde die Absorption bei 405 nm gemessen.
2.6.4. Papain Papain (EC 3.4.22.2, Sigma, Taufkirchen) wurde in einem Natriumphosphat-Puffer (0,1 M
Natriumphosphat, pH 6,5, mit 1 mM EDTA, 100 µl/ml) aufgenommen. 10 µl dieser Lösung
wurden mit 10 µl der jeweiligen Peptidverdünnung und 170 µl Puffer, der zusätzlich 2 mM
Cystein enthielt, 10 min bei 40 °C vorinkubiert. Danach wurde 10 µl Substrat zugesetzt
(BAPNA, 4,2 mg/ml in DMSO). Nach weiteren 3 min Inkubation wurde die Absorption bei
405 nm gemessen (Barrett, 1981).
2.6.5. Elastase Typ IV Elastase (EC 3.4.21.36, Sigma, Taufkirchen) wurde in Tris-Puffer (0,2 M Tris-HCl, pH 8,0,
0,075 mg/ml) gelöst. 10 µl dieser Lösung wurden mit 10 µl der jeweiligen Peptidverdünnung
und 150 µl Puffer bei 30 °C 20 min inkubiert. Danach wurden 30 µl Substrat (N-succinyl-
Ala-Ala-Ala-p-nitroanilid, 18 mg/20 ml Puffer) zugesetzt. Nach weiteren 3 min Inkubation
wurde die Absorption bei 405 nm gemessen (Bieth et al., 1974).
2.6.6. Thrombin Der Test wurde in modifizierter Form nach Lottenberg et al. (1981) durchgeführt. Thrombin
(EC 3.4.21.5, Sigma, Taufkirchen) wurde in einem Tris-Puffer (0,2 M Tris-HCl, pH 8,0,
0,5 mg/ml) gelöst. 10 µl dieser Lösung wurden mit 10 µl der jeweiligen Peptidverdünnung
und 170 µl Puffer bei 25 °C 10 min inkubiert. Danach wurden 10 µl Substrat (N-p-Tosyl-Gly-
Pro-Lys-p-nitroanilid, 0,5 mg/ml in Wasser) zugesetzt. Nach weiteren 3 min Inkubation
wurde die Absorption bei 405 nm gemessen.

Material und Methoden 33
2.6.7. Glutathion-S-Transferase Der Test wurde in modifizierter Form nach Habig et al. (1974) durchgeführt. Glutathion-S-
Transferase (EC 2.5.1.18, Sigma, Taufkirchen) wurde in einem Kaliumphosphat-Puffer
(0,1 M Kaliumphosphat, pH 6,5, mit 1 mM EDTA, 1 unit/ml) gelöst. Zu 260 µl
Kaliumphosphat-Puffer (0,1 M Kaliumphosphat, pH 6,5, mit 1 mM EDTA) wurden 10 µl
einer Glutathion-Lösung (75 mM der reduzierten Form in Puffer), sowie 10 µl Substrat
(1-Chlor-2,4-Dinitrobenzen, 30 mM in 95 % (v/v) Ethanol) gegeben und bei 25 °C inkubiert
bis die Absorption bei 340 nm konstant war. Danach wurden 10 µl der Enzymlösung
zugegeben und die Absorptionsänderung über 5 Minuten verfolgt.
2.6.8. Carboxypeptidase A Carboxypeptidase A (EC 3.4.17.1, Sigma, Taufkirchen) wurde in eiskalter NaCl-Lösung (1
M, 0,5 mg/ml) gelöst. 50 µl dieser Lösung wurden mit 10 µl der jeweiligen Peptidverdünnung
und 140 µl Tris-Puffer (0,025 M Tris-HCl, pH 7,5, mit 0,5 M NaCl) versetzt und 30 min bei
Raumtemperatur inkubiert. Danach wurden 150 µl Substrat (Hippuryl-L-phenylalanin,
0,001 M in Tris-HCl mit NaCl) zugegeben und die Absorption bei 254 nm gemessen
(Bergmeyer et al., 1974).
2.6.9. Leucin-Aminopeptidase Leucin-Aminopeptidase (EC 1.1.1.42, Sigma, Taufkirchen) wurde in kaltem Aqua dest.
(2,4 µl/ml). 10 µl dieser Lösung wurden mit 10 µl der jeweiligen Peptidverdünnung und
270 µl Natriumphosphat-Puffer (0,05 M Natriumphosphat, pH 7,2, 7,2 mg/ml) 15 min bei
37 °C inkubiert. Danach wurden 10 µl Substrat (L-Leucin-p-Nitroanilid, 0,024 M in Methanol
100 %) zugegeben. Nach weiteren 30 min Inkubation bei 37 °C wurde die Absorption bei
405 nm gemessen (Pfleiderer et al., 1970).
2.6.10. Thermolysin Der Test wurde in modifizierter Form nach Holmquist et al. (1974) durchgeführt.
Thermolysin (EC 3.4.24.27, Sigma, Taufkirchen) wurde in Acetat-Puffer (Natriumacetat
10 mM, Calciumacetat 5 mM, 0,5 mg/ml) gelöst. 60 µl dieser Lösung wurden mit 10 µl der
jeweiligen Peptidverdünnung und 140 µl Tris-Puffer (Tris-HCl 0,05 M, mit CaCl2 0,01 M,
mit NaCl 0,1 M, pH 7,5, 0,5 mg/ml) 30 min bei Raumtemperatur inkubiert. Danach wurden

Material und Methoden 34
90 µl Substrat (N-(3-[2-Furyl]acryloyl)-Gly-Leu-amid in Tris-Puffer (Tris-HCl 0,05 M, mit
CaCl2 0,01 M, mit NaCl 0,1 M, pH 7,5, 0,5 mg/ml) zugegeben. Nach weiteren 3 min
Inkubation wurde die Absorption bei 405 nm gemessen.
2.6.11. Proteinphosphatase PP1A Der Test wurde in modifizierter Form nach An und Carmichael (1994) durchgeführt. Es
wurde im Enzympuffer gelöste Protein-Phosphatase (EC 3.1.3.16, Calbiochem, Bad Soden)
verwendet, welche vor Gebrauch mit Verdünnungspuffer (Tris-HCl 50 mM, 1 mg/ml BSA,
MgCl2 20 mM, MnCl2 1 mM, DTT 2 mM, pH 7,4) auf 0,1 u/10 µl eingestellt wurde. Es
wurden 10 µl der Phosphatase-Verdünnung mit 10 µl der jeweiligen Peptidverdünnung
10 min bei Raumtemperatur inkubiert. Danach wurden 180 µl des Reaktionspuffers (Tris-HCl
50 mM, MgCl2 20 mM, MnCl2 1 mM, DDT 1 mM, 0,5 mg/ml BSA, p-Nitrophenolphosphat
(Tris-Salz) pH 8,1)zugegeben. Nach weiteren 20 min wird Inkubation wurde die Absorption
bei 405 nm gemessen.
2.7. Testung auf antimikrobielle Aktivität Als Methode wurde Mikrodilution nach Chmel et al. (1980) gewählt. Hierzu wurden zunächst
Stammlösungen der Peptide (128 µg/ml) sowie eines Peptidgemischs (je 42,7 µg/ml)
hergestellt: die Peptide wurden in DMSO aufgenommen und bis zur gewünschten
Konzentration mit Mueller-Hinton-Bouillon verdünnt. Vor Beginn der Versuche wurde die
Sterilität der Peptide ausgetestet, indem die Peptidstammlösungen auf BST-Agar-Platten
aufgetragen und bei 36 °C für 48 h inkubiert wurden. Es wurde kein Wachstum beobachtet.
Die Stammlösung des Peptidgemischs wurde aufgrund einer Keimbelastung mit einem
Sterilfilter (0,2 µm, Schleicher & Schuell FP 030/3, Dassel) filtriert.
Aus den Keimsuspensionen in MH-Bouillon Mac Farland (Ausnahme: Streptococcus
pyogenes Gr. A in H-Bouillon FA Merlin) mit 0.5 = 1 x 108 Keime/ml wurde ein Inokulum
mit 5 x 105 Keime/ml hergestellt (1:200 Verdünnung).
Von den jeweiligen Stammlösungen (128 µg/ml) wurden 1:2 Verdünnungen bis zur
Endkonzentrationen von 0.125 µg/ml hergestellt. In alle Vertiefungen der 96er Deep-Well-
Platten wurden jeweils 50 µl MH-Bouillon vorgelegt, anschließend wurden 50 µl der
betreffenden Verdünnungen der Peptide zupipettiert. Die einzelnen Wells wurden mit 10 µl
der 1:200 Keimverdünnung beimpft, so dass eine weitere Keimverdünnung stattfand und die

Material und Methoden 35
Endkonzentration bei annähernd 5 x 104 Keime / ml lag. Die Platten wurden über Nacht im
Brutschrank bei 36 °C inkubiert und am nächsten Tag ausgewertet.
Tabelle 7: Medium für Sterilitätsassay: BST-Agar Komponenten Menge BST Medium dehydrated 36,8 g Aqua dest. ad 1000 ml pH 7,4 Tabelle 8: Medium für Mikrodilutionsassays: Mueller-Hinton-Bouillon Komponenten Menge ( g / l ) Rinder Infus 2,0 Stärke 1,5 Pepton aus Casein 17,5 pH 7,4 ± 0,2 Herstellung:
21 g der Bouillon-Mischung wurden in einem Liter destillierten Wasser suspendiert, gut
gemischt und unter häufigem Rühren/Schütteln erhitzt, bis die Komponenten vollständig
gelöst waren. Das Medium wurde bei 118-121 °C 15 min sterilisiert.
2.8. Hämolysetest Von den isolierten Peptiden wurde jeweils eine gesättigte Stammlösung in Ethanol hergestellt.
Die Stammlösungen wurden mit PBS-Puffer auf eine Ethanolkonzentration von 10 % (v/v)
verdünnt. Mögliche Auswirkungen wurden am Lichtmikroskop verfolgt. Hierzu wurden zu
einer Erythrozyten-Lösung (vom Schaf) jeweils 20 µl Peptid-Lösung zupipettiert. Zusätzlich
wurde eine Blutagarplatte mit jeweils 10 µl Peptidlösung beimpft und im Brutschrank 48 h
inkubiert.
2.9. Toxizitätstest mit Kleinkrebsen (Crustaceen) Hierzu wurde die Mortalität der Kleinkrebse nach 24 h Inkubation mit den jeweiligen
Peptiden untersucht (Centeno et al., 1995). Die Testbedingungen orientierten sich am
käuflichen Thamnotoxkit FTM (MicroBioTests Inc., Nazareth, Belgien) und dem
mitgelieferten Standardprotokoll. Es kamen die Cyanopeptoline Hofmannolin, Scyptolin A

Material und Methoden 36
und B, sowie Scytocyclamide A und B zum Einsatz. Die Versuche wurden in 24er
Zellkulturplatten durchgeführt, wobei folgende Peptidkonzentrationen eingesetzt wurden:
100, 50, 25, 10, 1 und 0,1 µM bei einem Volumen von jeweils 500 µl. In jedes Well sollten
wenn möglich mindestens 10 Crustaceen gegeben werden. Als Kontrollen wurden Wasser
und 1 % (v/v) MeOH mitgeführt. Nach 24 h Inkubation bei 20 °C im Dunkeln wurde die
Todesrate pro Well bestimmt.
Vergleichend wurden die Scytocyclamide A und B auf Ihre Wirkung auf Daphnia magna
getestet. Es wurden keine vollständigen Versuchsreihen durchgeführt, die Versuche sollten
lediglich einem Screening dienen. Es wurden jeweils zweimal die Konzentrationen 100, 50
und 10 µM auf eine Daphnie getestet.
2.10. Zytotoxizitätsversuche
2.10.1. Zelllinien Insgesamt wurden 6 Peptide (Scyptolin A / B, Hofmannolin sowie die Scytocyclamide A bis
C) aus Scytonema hofmanni PCC 7110 auf Wirksamkeit gegenüber verschiedenen Zelllinien
getestet. Für die Versuche wurden gebrauchsfertige Zelllinien zur Verfügung gestellt:
CaCo 2 Zellen aus humanen Colorectalem Adenokarzinom
HeLa humane Epithelzellen eines Zervixkarzinoms
Swiss 3T3 Zellen aus embryonalen Mäusefibroblasten
Ebl embryonale Rinderlungenzellen
RBL basophile Rattenleukämiezellen
LoVo Zellen aus humanen Colorectalem Adenokarzinom
2.10.2. Trypanblaufärbung Um die Lebensfähigkeit von Zellen zu prüfen, wird heute auf den einfach und schnell
anzuwendenden Routinetest der Trypanblaufärbung zurückgegriffen. Trypanblau ist ein
saurer Farbstoff, dessen Anion an Zellproteine bindet. Trypanblau dringt durch defekte
Zellmembranen toter Zellen in das Cytosol und färbt diese Zellen tiefblau. Lebende Zellen
erscheinen unter dem Mikroskop leuchtend hell. Jedoch ist bei zu langer Einwirkzeit des
Trypanblau ist ein Anstieg von toten Zellen zu beobachten, da dieser Farbstoff zytotoxisch
wirkt. Die Zellzahl abgetöteter Zellen wurde mit einem Hämozytometer nach Neubauer

Material und Methoden 37
bestimmt (Schichttiefe: 0,1 mm). Vor dem Zählen wurden die Zellen vom Flaschenboden
gelöst, abzentrifugiert und nochmals mit PBS-Puffer gewaschen. Die Zellen wurden in
frisches Medium aufgenommen und resuspendiert. Anschließend wurden 20 µl
Zellsuspension mit 20 µl Trypanblau-Lösung gut vermischt und auf die Zählkammer
aufgebracht.
2.11. Versuche zur Membranschädigung
2.11.1. Testung auf porenbildende Aktivität
„Lipid-bilayer“-Studien sind eine Möglichkeit, um durch Toxine verursachte Porenbildung in
einem in vitro-System zu untersuchen (Benz et al., 1978): Hierbei verwendet man einen mit
wässriger Lösung gefüllten Behälter, der durch eine Scheidewand in zwei Bereiche unterteilt
ist. In der Scheidewand befindet sich ein Loch mit einem Durchmesser von etwa einem
Millimeter. Über diesem Loch kann man nun eine Membran aus Lecithin entstehen lassen. Sie
ist etwa 5 nm dick und damit ungefähr genau so dick wie eine natürliche Membran und
besteht ausschließlich aus Diphytanoyl-Phosphatidylcholin und n-Dekan.
Abb. 3: Messapparatur zur Bestimmung der Membranleitfähigkeit; Versuchsbedingungen: 1 M KCl, pH 6, Membranen aus Diphytanoyl-Phosphatidylcholin/n-Dekan
2.11.2. 86Rubidium+-Efflux Scytocyclamid A und B wurden in DSMO gelöst (10 µg/µl). Zu einer Zellsuspension aus HT-
29-Zellen wurde 1 µCi 86Rb+/ml zugegeben (Barth et al., 2001). Die Zellen wurden in
24-Well-Platten mit 2 x 105/well ausgesät. Das Volumen des Inokulums betrug 500 µl. Nach

Material und Methoden 38
4 Tagen Wachstum wurden die Zellen zweimal mit PBS-Puffer gewaschen. Anschließend
wurden die Zellen in 500 µl Medium ohne FCS (McCoys-Medium, Sigma, Taufkirchen)
aufgenommen und mit 1,76 µl Peptid 15 min bei Raumtemperatur inkubiert. Aliquote der
Versuchsansätze wurden entnommen und der Rubidiumefflux durch Szintillationszählung
(LKB, Rackbeta 1209, Gräfelfing) bestimmt. Es wurden jeweils 3-fach Bestimmungen
durchgeführt. DMSO (1,76 µl) wurde als Kontrolle mitgeführt.
2.12. Immunhistochemische Untersuchungen HeLa-Zellen wurden in einem Zytoskelett stabilisierenden Puffer (CSB), welcher 80 mM
PIPES, 5 mM EGTA, 1 mM MgCl2, und 4 % (v/v) PEG (MW 35.000) enthielt mit 4 % (v/v)
Paraformaldehyd 12 min fixiert (Schaefer et al., 2002). Nach Waschen mit CSB-Puffer
wurden die Zellen 2 min mit 0,1 % (v/v) Triton X-100 in CSB permeabilisiert. „normal goat
serum“ wurde zugegeben um unspezifische Bindungen zu verhindern. Zur α-Tubulin III
Färbung wurden die Zellen mit monoklonalem Mäuse anti-α-Tubulin III Antikörper (1:4.000;
Sigma, Taufkirchen) inkubiert. Der resultierende Immunkomplex wurde mit Alexa 488
gekoppeltem F(ab´)2 Fragment eines anti-Mäuse, bzw. anti-Kaninchen IgG (Molecular
Probes, Heidelberg) visualisiert. F-actin-Färbung wurde mit Alexa 594 gekoppeltem
Phalloidin (Molecular Probes, Heidelberg) inkubiert und mit PBS-Puffer gewaschen.
2.13. Testung auf Apoptose
2.13.1. Fluoreszenz-aktivierte Zell-Sortierung (FACS-Analysen) Die FACS-Analysen wurden mit dem Becton /Dickinson FACS-Calibur (Becton / Dickinson
Corp., Heidelberg). Die verschiedenen Fluoreszenzkanäle wurden auf Vorwärtsstreulicht
(„forewardscatter“ FSC), Seitwärtsstreulicht („sidewardscatter“ SSC), Fluorescein-
Isothiocyanat (FITC), Hydroethidin (2-HE, Molecular Probes), Dihexyloxacarbocyanin-Iodid
(DiOC6(3) (Molecular Probes) und auf Propidiumiodid (PI) eingestellt. Die
Messwellenlängen wurden je nach Untersuchungsziel auf die passenden Wellenlängen
eingestellt. Die Fluoreszenzintensität war hierbei Maß für die Absorption bei der
Maximalwellenlänge des betreffenden Markers. Die Daten wurden in Form von
Häufigkeitshistogrammen akquiriert und mit CellQuest® (Becton / Dickinson Corp.,
Heidelberg) ausgewertet.

Material und Methoden 39
2.13.1.1. Apoptose-Färbung mit Annexin V-FITC und Propidiumiodid Die Apoptosebestimmung erfolgte mit Hilfe von FITC-markiertem Annexin V und
Propidiumiodid gemäß Protokoll (BD Pharmingen, Heidelberg). Die mit Scytocyclamid A
(10 µmol) für 5-6 h inkubierten Zellen (MEF-Zelllinie: embryonale Mäusefibroblasten)
wurden 1-2 Mal mit PBS-Puffer gewaschen und in 100 µl Annexin Bindungspuffer
aufgenommen. Dieser Puffer enthielt Ca2+-Ionen. Es folgte die Zugaben von 5 µl Annexin V
und 2 µl Propidiumiodid. Der Versuchsansatz wurde bei Raumtemperatur im Dunkeln 15 min
inkubiert. Anschließend wurden weitere 100 µl Annexinbindungspuffer zugegeben und
innerhalb von 1 h die Messungen durchgeführt.
2.13.1.2. Mitochondriales Membranpotential Die Messung des Membranpotentials wurde über die DiOC6(3)-Färbung am
Durchflusszytometer verfolgt. Die verwendeten Zellen (250.000) wurden abzentrifugiert, der
Überstand aus Medium (EMEM, unter Zusatz von 5 % (v/v) fötalem Kälberserum) wurde
verworfen. Anschließend resuspendierte man die Zellen in 200 µl Medium, welches 2 µM
Hydroethidin enthielt. Es wurde bei 37 °C, 5 % CO2 unter Lichtausschluß 15 min inkubiert
gefolgt von der Zugabe von 20 nM DiOC6(3) und nochmaliger Inkubation bei 37 °C, 5 %
CO2 unter Lichtausschluß für weitere 15 min. Eine Positivkontrolle wurde parallel dazu
bearbeitet: 250.000 Zellen wurden nach Zentrifugation in Medium, welches 200 µM CCCP
enthielt, resuspendiert und 1 h bei 37 °C inkubiert. Der Ansatz wurde abzentrifugiert, der
Überstand verworfen und die Zellen in 100 µl Medium, welches 40 nM DiOC6(3) enthielt
resuspendiert. Es folgte Inkubation bei 37 °C für 15 min. Es wurde auf 1 ml Gesamtvolumen
mit PBS-Puffer aufgefüllt und die Proben mit Durchflusszytometrie (Becton / Dickinson
Corp., Heidelberg) gemessen.
2.13.1.3. Detektion reaktiver Sauerstoffspezies Die Versuchsdurchführung ist bei „Mitochondrien Membranpotential“ des Methodenteils
(siehe 2.16.1.2.) angegeben. Es wurde lediglich auf einen anderen, für Ethidium geeigneten
Fluoreszenzlichtkanal umgestellt.

Ergebnisse 40
3. Ergebnisse In der vorliegenden Arbeit wurde das Spektrum an Peptid-Sekundärmetaboliten des
Cyanobakterienstamms Scytonema hofmanni PCC 7110 analytisch-chemisch sowie
hinsichtlich biologischer Wirksamkeit näher untersucht. Durch Matern (2002) waren bereits 3
Cyanopeptoline charakterisiert worden. Für ein weiteres Peptid wurde anhand von
Fragmentdaten aus der Massenspektrometrie eine Struktur vom Cyanopeptolin-Typ postuliert,
ohne dass diese durch weiterführende Analysen bestätigt werden konnte.
3.1. Charakterisierung der Cyanopeptoline und Scytocyclamide Das HPLC-Chromatogramm des Zell-Rohextraktes (Abb. 4) bestätigte zunächst die von
Matern et al. (2002) identifizierten Peptide. Neben diesen Signalen wurden jedoch weitere
bisher nicht näher untersuchte Signale erhalten. Die Signale mit Retentionszeiten > 25 min
hatten untypische UV-Spektren im Vergleich zu den bisher charakterisierten Peptiden aus
Scytonema hofmanni. Ausgehend von einem methanolischen Rohextrakt wurden in der
vorgelegten Arbeit die einzelnen Peptide über mehrere Schritte der Festphasen-
chromatographie isoliert. Die Strukturen der einzelnen Verbindungen wurden über UV- und
IR-Messungen, Massenspektrometrie und durch NMR-Analytik näher charakterisiert. Wie im
weiteren Verlauf der Arbeit belegt wird, erwiesen sich die zusätzlich identifizierten
Cyanopeptoline als potente Hemmstoffe neuer Zielenzyme aus der Klasse der Proteasen. Mit
den bisher unbekannten Scytocyclamiden wurden Peptide mit zytotoxischen Eigenschaften
identifiziert.
3.1.1. Extraktion und Isolierung Die Zellen aus Scytonema hofmanni PCC 7110 wurden mit 50 % (v/v) Methanol extrahiert,
und der so gewonnene Rohextrakt wurde durch RP-HPLC untersucht. Anhand der UV-
Absorptionsspektren einzelner Signale wurden von Matern (2002) nicht identifizierte bisher
nicht untersuchte Peptide vermutet (Abb. 4 und 5). Neben den für die Cyanopeptoline
bekannten UV-Spektren wurden bei den Signalen mit einer Retentionszeit > 25 min für
Cyanopeptoline untypische Spektren detektiert. Diese Peptide wurden in die Untersuchungen
mit einbezogen (Abb.5).
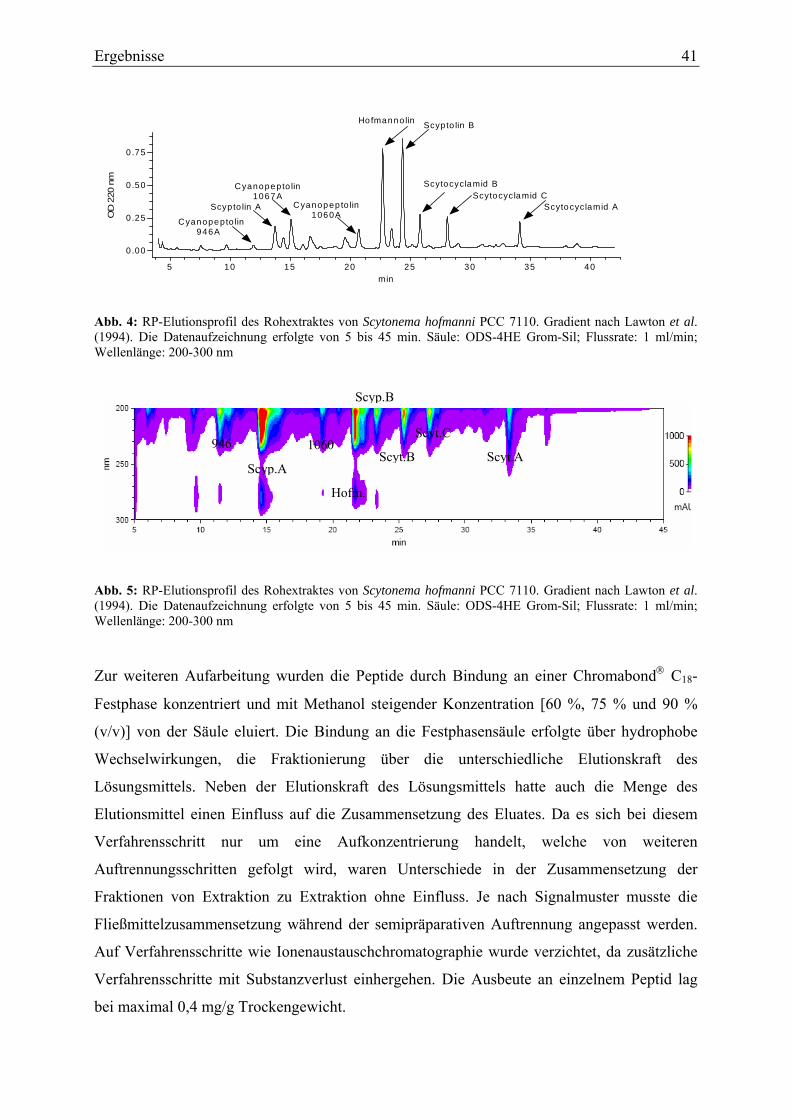
Ergebnisse 41
Abb. 4: RP-Elutionsprofil des Rohextraktes von Scytonema hofmanni PCC 7110. Gradient nach Lawton et al. (1994). Die Datenaufzeichnung erfolgte von 5 bis 45 min. Säule: ODS-4HE Grom-Sil; Flussrate: 1 ml/min; Wellenlänge: 200-300 nm
Abb. 5: RP-Elutionsprofil des Rohextraktes von Scytonema hofmanni PCC 7110. Gradient nach Lawton et al. (1994). Die Datenaufzeichnung erfolgte von 5 bis 45 min. Säule: ODS-4HE Grom-Sil; Flussrate: 1 ml/min; Wellenlänge: 200-300 nm Zur weiteren Aufarbeitung wurden die Peptide durch Bindung an einer Chromabond® C18-
Festphase konzentriert und mit Methanol steigender Konzentration [60 %, 75 % und 90 %
(v/v)] von der Säule eluiert. Die Bindung an die Festphasensäule erfolgte über hydrophobe
Wechselwirkungen, die Fraktionierung über die unterschiedliche Elutionskraft des
Lösungsmittels. Neben der Elutionskraft des Lösungsmittels hatte auch die Menge des
Elutionsmittel einen Einfluss auf die Zusammensetzung des Eluates. Da es sich bei diesem
Verfahrensschritt nur um eine Aufkonzentrierung handelt, welche von weiteren
Auftrennungsschritten gefolgt wird, waren Unterschiede in der Zusammensetzung der
Fraktionen von Extraktion zu Extraktion ohne Einfluss. Je nach Signalmuster musste die
Fließmittelzusammensetzung während der semipräparativen Auftrennung angepasst werden.
Auf Verfahrensschritte wie Ionenaustauschchromatographie wurde verzichtet, da zusätzliche
Verfahrensschritte mit Substanzverlust einhergehen. Die Ausbeute an einzelnem Peptid lag
bei maximal 0,4 mg/g Trockengewicht.
Scyt.C
Scyt.B Scyt.A
Scyp.B
Hofm.
1060
Scyp.A
946
5 10 15 20 25 30 35 40
0.00
0.25
0.50
0.75
Hofmanno lin Scypto lin B
Scypto lin A
Scytocyclamid BScytocyclamid C
Scytocyclamid A
Cyanopepto lin946A
Cyanopepto lin1067A
Cyanopep to lin1060A
min
OD
220
nm
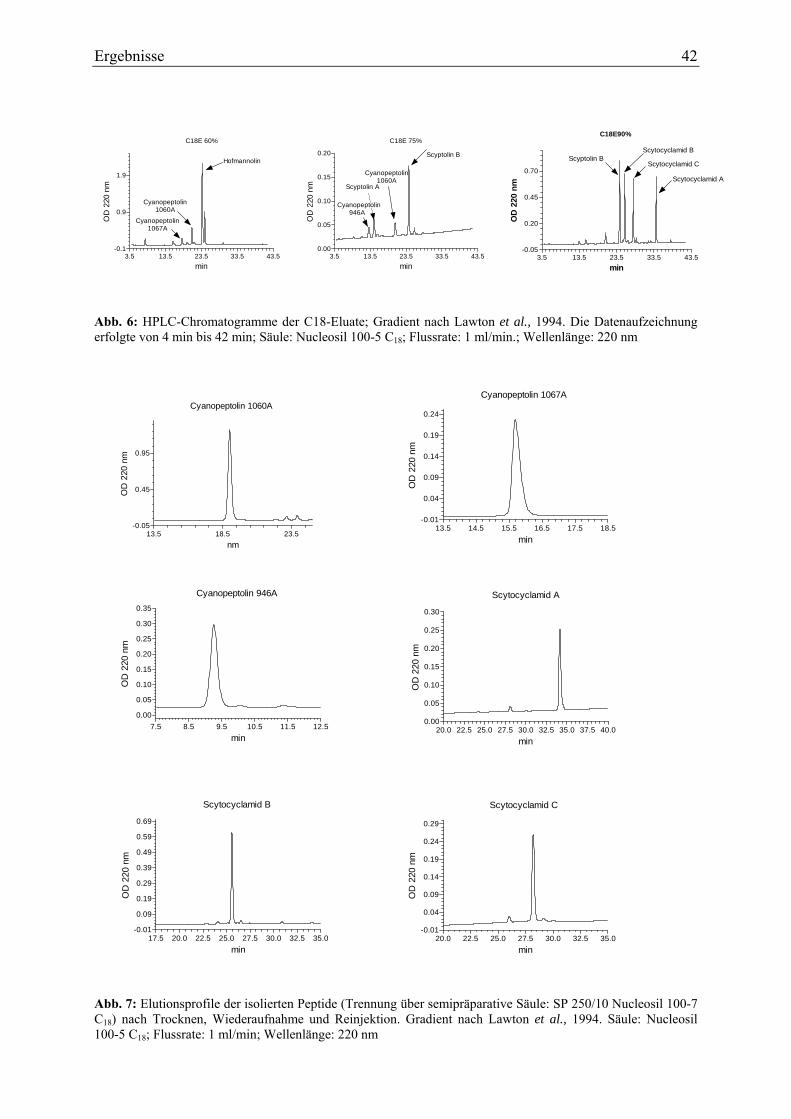
Ergebnisse 42
Abb. 6: HPLC-Chromatogramme der C18-Eluate; Gradient nach Lawton et al., 1994. Die Datenaufzeichnung erfolgte von 4 min bis 42 min; Säule: Nucleosil 100-5 C18; Flussrate: 1 ml/min.; Wellenlänge: 220 nm
Abb. 7: Elutionsprofile der isolierten Peptide (Trennung über semipräparative Säule: SP 250/10 Nucleosil 100-7 C18) nach Trocknen, Wiederaufnahme und Reinjektion. Gradient nach Lawton et al., 1994. Säule: Nucleosil 100-5 C18; Flussrate: 1 ml/min; Wellenlänge: 220 nm
C18E90%
3.5 13.5 23.5 33.5 43.5-0.05
0.20
0.45
0.70
Scytocyclamid B
Scytocyclamid C
Scytocyclamid A
Scyptolin B
min
OD
220
nm
C18E 75%
3.5 13.5 23.5 33.5 43.50.00
0.05
0.10
0.15
0.20 Scyptolin B
Cyanopeptolin1060A
Cyanopeptolin946A
Scyptolin A
min
OD
220
nm
C18E 60%
3.5 13.5 23.5 33.5 43.5-0.1
0.9
1.9
Hofmannolin
Cyanopeptolin1060A
Cyanopeptolin1067A
min
OD
220
nm
Cyanopeptolin 1060A
13.5 18.5 23.5-0.05
0.45
0.95
nm
OD
220
nm
Cyanopeptolin 1067A
13.5 14.5 15.5 16.5 17.5 18.5-0.01
0.04
0.09
0.14
0.19
0.24
min
OD
220
nm
Scytocyclamid B
17.5 20.0 22.5 25.0 27.5 30.0 32.5 35.0-0.01
0.09
0.19
0.29
0.39
0.49
0.59
0.69
min
OD
220
nm
Scytocyclamid A
20.0 22.5 25.0 27.5 30.0 32.5 35.0 37.5 40.00.00
0.05
0.10
0.15
0.20
0.25
0.30
min
OD
220
nm
Cyanopeptolin 946A
7.5 8.5 9.5 10.5 11.5 12.50.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
min
OD
220
nm
Scytocyclamid C
20.0 22.5 25.0 27.5 30.0 32.5 35.0-0.01
0.04
0.09
0.14
0.19
0.24
0.29
min
OD
220
nm
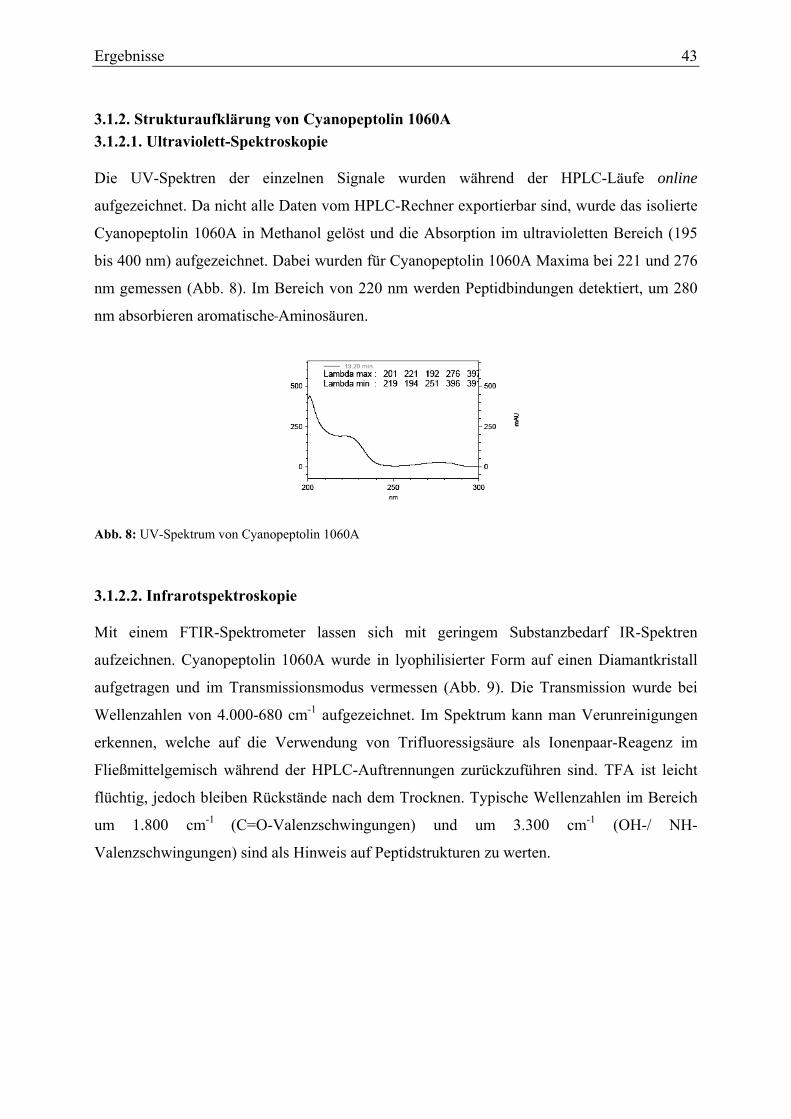
Ergebnisse 43
3.1.2. Strukturaufklärung von Cyanopeptolin 1060A 3.1.2.1. Ultraviolett-Spektroskopie Die UV-Spektren der einzelnen Signale wurden während der HPLC-Läufe online
aufgezeichnet. Da nicht alle Daten vom HPLC-Rechner exportierbar sind, wurde das isolierte
Cyanopeptolin 1060A in Methanol gelöst und die Absorption im ultravioletten Bereich (195
bis 400 nm) aufgezeichnet. Dabei wurden für Cyanopeptolin 1060A Maxima bei 221 und 276
nm gemessen (Abb. 8). Im Bereich von 220 nm werden Peptidbindungen detektiert, um 280
nm absorbieren aromatische Aminosäuren.
Abb. 8: UV-Spektrum von Cyanopeptolin 1060A
3.1.2.2. Infrarotspektroskopie Mit einem FTIR-Spektrometer lassen sich mit geringem Substanzbedarf IR-Spektren
aufzeichnen. Cyanopeptolin 1060A wurde in lyophilisierter Form auf einen Diamantkristall
aufgetragen und im Transmissionsmodus vermessen (Abb. 9). Die Transmission wurde bei
Wellenzahlen von 4.000-680 cm-1 aufgezeichnet. Im Spektrum kann man Verunreinigungen
erkennen, welche auf die Verwendung von Trifluoressigsäure als Ionenpaar-Reagenz im
Fließmittelgemisch während der HPLC-Auftrennungen zurückzuführen sind. TFA ist leicht
flüchtig, jedoch bleiben Rückstände nach dem Trocknen. Typische Wellenzahlen im Bereich
um 1.800 cm-1 (C=O-Valenzschwingungen) und um 3.300 cm-1 (OH-/ NH-
Valenzschwingungen) sind als Hinweis auf Peptidstrukturen zu werten.
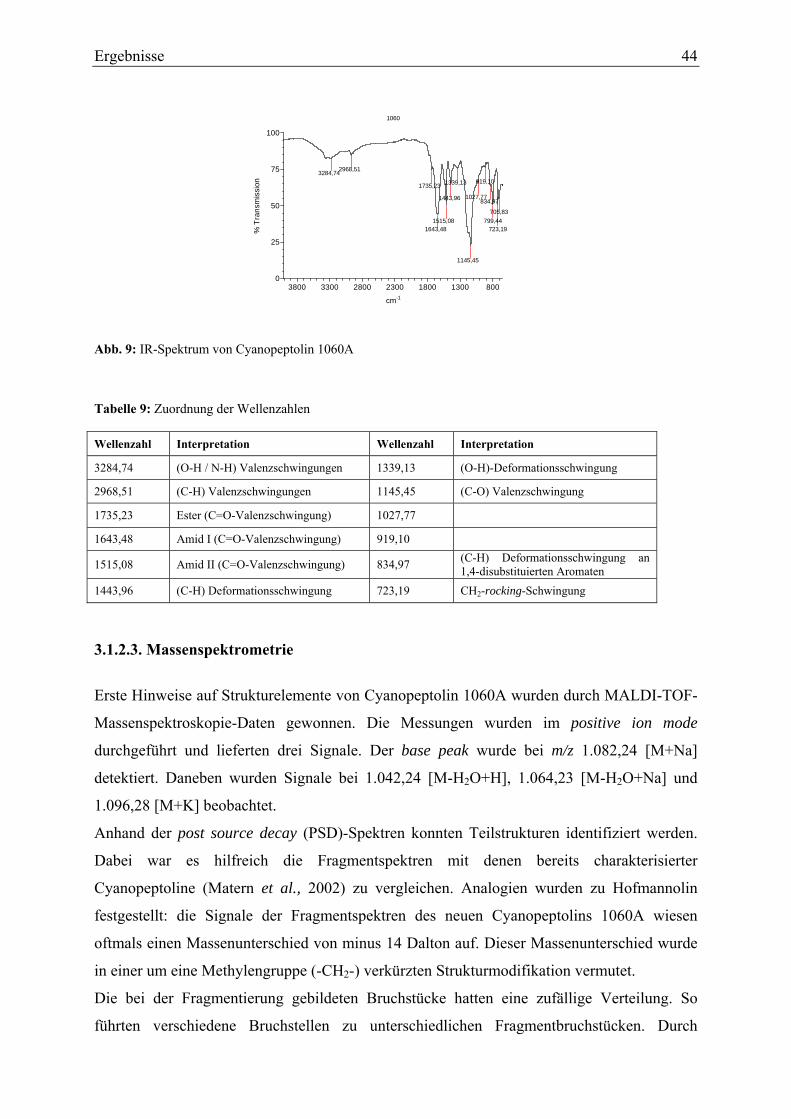
Ergebnisse 44
Abb. 9: IR-Spektrum von Cyanopeptolin 1060A
Tabelle 9: Zuordnung der Wellenzahlen Wellenzahl Interpretation Wellenzahl Interpretation
3284,74 (O-H / N-H) Valenzschwingungen 1339,13 (O-H)-Deformationsschwingung
2968,51 (C-H) Valenzschwingungen 1145,45 (C-O) Valenzschwingung
1735,23 Ester (C=O-Valenzschwingung) 1027,77
1643,48 Amid I (C=O-Valenzschwingung) 919,10
1515,08 Amid II (C=O-Valenzschwingung) 834,97 (C-H) Deformationsschwingung an 1,4-disubstituierten Aromaten
1443,96 (C-H) Deformationsschwingung 723,19 CH2-rocking-Schwingung
3.1.2.3. Massenspektrometrie
Erste Hinweise auf Strukturelemente von Cyanopeptolin 1060A wurden durch MALDI-TOF-
Massenspektroskopie-Daten gewonnen. Die Messungen wurden im positive ion mode
durchgeführt und lieferten drei Signale. Der base peak wurde bei m/z 1.082,24 [M+Na]
detektiert. Daneben wurden Signale bei 1.042,24 [M-H2O+H], 1.064,23 [M-H2O+Na] und
1.096,28 [M+K] beobachtet.
Anhand der post source decay (PSD)-Spektren konnten Teilstrukturen identifiziert werden.
Dabei war es hilfreich die Fragmentspektren mit denen bereits charakterisierter
Cyanopeptoline (Matern et al., 2002) zu vergleichen. Analogien wurden zu Hofmannolin
festgestellt: die Signale der Fragmentspektren des neuen Cyanopeptolins 1060A wiesen
oftmals einen Massenunterschied von minus 14 Dalton auf. Dieser Massenunterschied wurde
in einer um eine Methylengruppe (-CH2-) verkürzten Strukturmodifikation vermutet.
Die bei der Fragmentierung gebildeten Bruchstücke hatten eine zufällige Verteilung. So
führten verschiedene Bruchstellen zu unterschiedlichen Fragmentbruchstücken. Durch
1060
8001300180023002800330038000
25
50
75
100
3284,742968,51
705,83
1735,23
1515,081643,48
1443,96
1339,13
1145,45
919,10
834,97
799,44
1027,77
723,19
cm-1
% T
rans
mis
sion

Ergebnisse 45
Überschneidungen der Bruchstücke kann man Rückschlüsse auf die vollständige Sequenz
bekommen. Durch die Vergleiche der Fragmentspektren und die Zuordnungen der
Fragmentmassen zu bestimmten Bruchstücken konnte man ableiten, dass Cyanopeptolin
1060A das gleiche Ringgerüst aufweist wie Hofmannolin und dass die Molekülmodifikation
in der Seitenkette lokalisiert ist (siehe hierzu Abb. 11). Die Fragmentzuordnungen sind in
Tabelle 10 angegeben.
Tabelle 10: MALDI-TOF PSD-Fragmentzuordnung von Cyanopeptolin 1060A Sequenz Ion m/z Tyr Ahp OMTyr NMTyr Val Thr Glu Hiv M-H2O+H 1042 Tyr Ahp OMTyr NMTyr Val Thr Glu Hiv M-CO- H2O+H 1015 Tyr Ahp OMTyr NMTyr Thr Glu Hiv M-H2O+H 944 Tyr Ahp OMTyr Val Thr Glu Hiv M- H2O+H 865 Tyr Ahp OMTyr Val Thr Glu Hiv M-CO- H2O+H 837 Tyr Ahp OMTyr NMTyr Val Thr M-H2O+H 814 Tyr NMTyr Val Thr Glu Hiv M+H 770 NMTyr Val Thr Glu Hiv M+H 607 Ahp OMTyr NMTyr Val M+H 567 Ahp OMTyr NMTyr Val M- H2O+H 549 Tyr Thr Glu Hiv M- H2O+H 476 Ahp OMTyr NMTyr M-H2O+H 450 Tyr Ahp OMTyr M-H2O+H 436 Ahp OMTyr M-H2O+H 273 Ahp OMTyr M-CO- H2O+H 245 Val Thr M+H 200 NMTyr Immoniumion 150 Tyr Immoniumion 136 Tyr Seitenkette 107 Glu Seitenkette 102 Val Immoniumion 72 Es wurde eine Summenformel von C53H69N7O16 bestimmt. Die Struktur konnte jedoch anhand
der Massenzahlen nicht gelöst werden. Die Strukturmodifikation der Seitenkette musste daher
durch H-NMR-Messungen bestimmt werden.
3.1.2.4. Magnetische Kernresonanzspektroskopie
Aus den 1H-NMR-Spektren von Cyanopeptolin 1060A ließen sich 48 unterschiedliche
chemische Verschiebungen bestimmen, welchen 69 Protonen zugeordnet werden mussten.
Von den 69 Protonen waren 68 auf den Bereich von 0,5 ppm bis 9,5 ppm verteilt. Neben
diesen Resonanzsignalen war ein weiteres breites Signal mit hoher Linienbreite bei 12,1 ppm
detektiert worden. Linienverbreiterungen sind oftmals durch intermolekulare
Austauscheffekte zu erklären. Die chemische Verschiebung zu diesem Signal lässt vermuten,
dass es sich mit hoher Wahrscheinlichkeit um ein Proton einer Hydroxylgruppe handelt.
Durch weitere Messungen, wie TOCSY- und ROESY-Korrelationsmessungen, sowie durch

Ergebnisse 46
HSQC-Aufnahmen konnte dieses Signal der Säurefunktion von Glutaminsäure zugeordnet
werden (siehe hierzu Abb. 11).
In Analogie zu dem von Matern et al. (2003) gefundenen Hofmannolin konnten 7 Methyl-
und 12 Methingruppen bestimmt werden. Wie auch die Daten aus der Massenspektroskopie
nahe legten, konnten nur 7 anstatt der 8 in Hofmannolin gefundenen Methylengruppen
zugeordnet werden. Daneben wurden 12 aromatische Protonen detektiert. Die Auswertung der
HSQC-Korrelationen ergab, dass 59 Protonen an Kohlenstoff gebunden sind. Weitere
10 Protonen müssen in diesem Fall an andere Atome gebunden sein. Jeweils 2 Protonen
konnten phenolischen und olefinischen Hydroxylgruppen zugeordnet werden. Die
verbleibenden 5 Protonen konnten an Stickstoffatomen der Amidbindungen (Dubletts
zwischen 7,0 und 8,6 ppm) gefunden werden.
Es wurden 5 Signale ausgewertet, welche keine Aufspaltung aufwiesen. Durch Integration
konnten 2 dieser 5 Signale Methylgruppen zugeordnet werden, welche nicht in Nachbarschaft
zu anderen Kohlenstoffatomen stehen (2,75 und 3,72 ppm). Die anderen 3 Signale gehören zu
2 phenolischen OH-Gruppen (9,07 und und 9,37 ppm), sowie zur Säurefunktion der
Glutaminsäure (12,13 ppm) (siehe hierzu Abb. 11).
Neben den Singuletts der aromatischen Methoxygruppe und der N-Methylgruppe zeigen die
anderen Methylgruppen Duplizität durch die Nachbarschaft eines weiteren Protons. Alle 7
Methylengruppen sind an Kohlenstoff gebunden, wie aus den 1H bzw. 13C Verschiebungen
hervorgeht (Tabelle 11). Anhand der HSQC-Korelationen der Methingruppen lassen sich drei
an Sauerstoff und sieben an Stickstoff gebundene Methingruppen identifizieren.
Bei den aromatischen Aminosäuren mit insgesamt 12 Protonen ergeben sich durch para-
Substitution für 4 Protonen einer Aminosäure jeweils zwei chemisch äquivalente
Protonenpaare. Es handelt sich um ein AA`BB`-Spinsystem. Die Signale werden jeweils
durch die Nachbarschaft eines weiteren Protons in Dubletts aufgespaltet.
Die Unterschiede zu Hofmannolin zeigen sich in der Seitenkette. Durch die um eine
Methylengruppe verkürzte Seitenkette bekommt man keine Aufspaltung zu einem Triplett bei
0,93 ppm. In Einklang dazu stehen fünf Korrelationen aus dem TOCSY-Spektrum, welche
durch die Protonen tragende Gruppen verursacht werden (siehe hierzu Abb. 11).
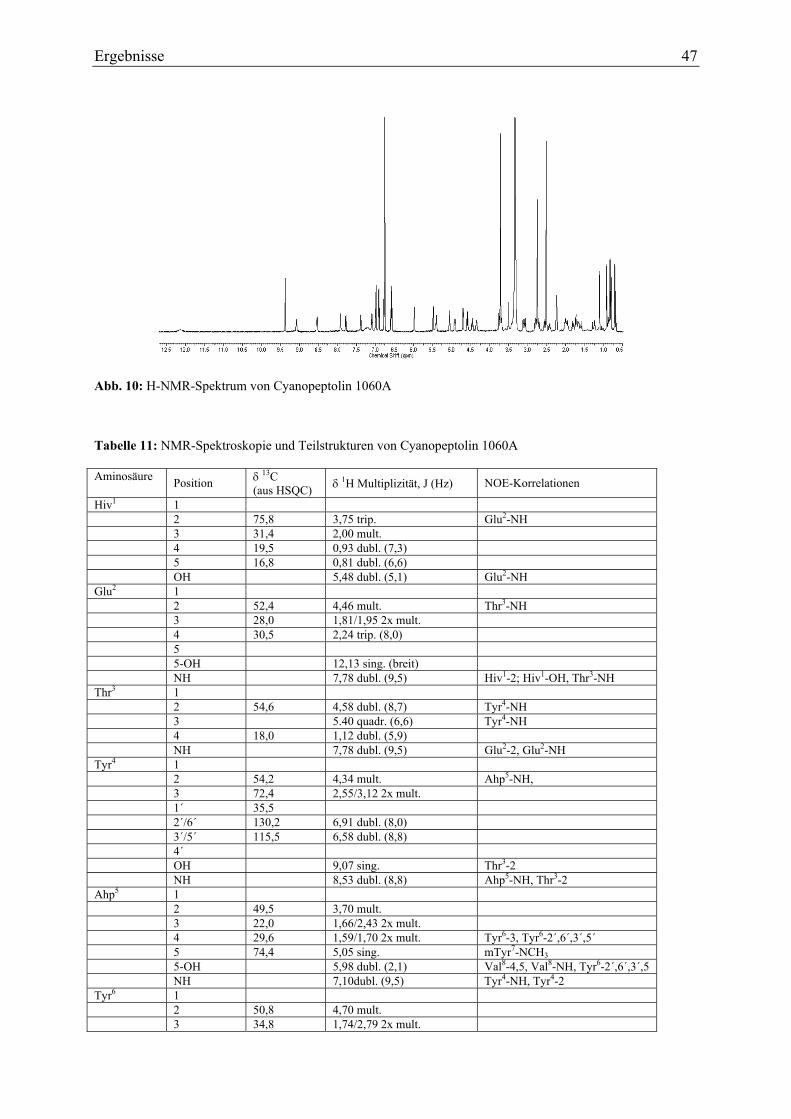
Ergebnisse 47
Abb. 10: H-NMR-Spektrum von Cyanopeptolin 1060A
Tabelle 11: NMR-Spektroskopie und Teilstrukturen von Cyanopeptolin 1060A Aminosäure Position δ 13C
(aus HSQC) δ 1H Multiplizität, J (Hz) NOE-Korrelationen
Hiv1 1 2 75,8 3,75 trip. Glu2-NH 3 31,4 2,00 mult. 4 19,5 0,93 dubl. (7,3) 5 16,8 0,81 dubl. (6,6) OH 5,48 dubl. (5,1) Glu2-NH Glu2 1 2 52,4 4,46 mult. Thr3-NH 3 28,0 1,81/1,95 2x mult. 4 30,5 2,24 trip. (8,0) 5 5-OH 12,13 sing. (breit) NH 7,78 dubl. (9,5) Hiv1-2; Hiv1-OH, Thr3-NH Thr3 1 2 54,6 4,58 dubl. (8,7) Tyr4-NH 3 5.40 quadr. (6,6) Tyr4-NH 4 18,0 1,12 dubl. (5,9) NH 7,78 dubl. (9,5) Glu2-2, Glu2-NH Tyr4 1 2 54,2 4,34 mult. Ahp5-NH, 3 72,4 2,55/3,12 2x mult. 1´ 35,5 2´/6´ 130,2 6,91 dubl. (8,0) 3´/5´ 115,5 6,58 dubl. (8,8) 4´ OH 9,07 sing. Thr3-2 NH 8,53 dubl. (8,8) Ahp5-NH, Thr3-2 Ahp5 1 2 49,5 3,70 mult. 3 22,0 1,66/2,43 2x mult. 4 29,6 1,59/1,70 2x mult. Tyr6-3, Tyr6-2´,6´,3´,5´ 5 74,4 5,05 sing. mTyr7-NCH3 5-OH 5,98 dubl. (2,1) Val8-4,5, Val8-NH, Tyr6-2´,6´,3´,5 NH 7,10dubl. (9,5) Tyr4-NH, Tyr4-2 Tyr6 1 2 50,8 4,70 mult. 3 34,8 1,74/2,79 2x mult.

Ergebnisse 48
1´ 2´/6´ 130,8 6,76 dubl. (8,0) Ahp5-5 3´/5´ 113,8 6,76 dubl. (8,0) Ahp5-5 4´ OCH3 55,4 3,72 sing. Ahp5-NH mTyr7 1 2 61,0 4,91 mult. 3 33,4 3,07 2x mult. Ahp5-4 2´/6´ 130,8 6,99 dubl. (8,8) Ahp5-4 3´/5´ 115,8 6,78 dubl. (8,0) 4´ OH 9,37 sing. NCH3 30,5 2,75 sing. Val8-4,5, Val8-NH Val8 1 2 56,4 4,69 mult. 3 31,4 2,02 mult. 4 19,5 0,84 dubl. (6,6)) mTyr7-NCH3 5 17,6 0,71 dubl. (6,6) mTyr7-NCH3, Ahp5-OH NH 7,39 dubl. (9,6) mTyr7-NCH3, Ahp5-OH Korrelationen innerhalb des Spinsystems einer Aminosäure werden nicht aufgeführt.
Aus den NMR-Daten geht hervor, dass Cyanopeptolin 1060A das gleiche Ringgerüst wie
Hofmannolin besitzt. Insoweit wurde der Strukturvorschlag aus der Massenspektrometrie
bestätigt. Des Weiteren wurde die Strukturmodifikation in der Seitenkette aufgeklärt.
Während in Hofmannolin der N-Terminus der Seitenkette durch 2-Hydroxy-3-
methylisovaleriansäure acyliert ist, betseht der Acylrest im neu isolierten Cyanopeptolin aus
2-Hydroxy-isovaleriansäure. Die NMR-Ergebnisse stimmen mit der postulierten
Summenformel C53H69N7O16 und der errechneten Massen von 1.060 Da, bzw. der Masse des
base peak von m/z 1.082 ([M+Na]) vollständig überein, so dass die Struktur von
Cyanopeptolin 1060A bestätigt wurde.
Abb. 11: Struktur von Cyanopeptolin 1060A aus Scytonema hofmanni, Hiv: 2-Hydroxy-isovaleriansäure, MTyr: N-Methyl-Tyrosin, OMTyr: O-Methyl-Tyrosin, Ahp: 3-Amino-6-Hydroxy-2-Oxo-Piperidin
N
O
NH
OH
O
O
O
NH
O
NH
OH O
OOH
NH
OH
N
OO
NH O
OH
O
Tyr4
Ahp5
OMTyr6
NMTyr7Val8
Glu2
Hiv1 Thr3
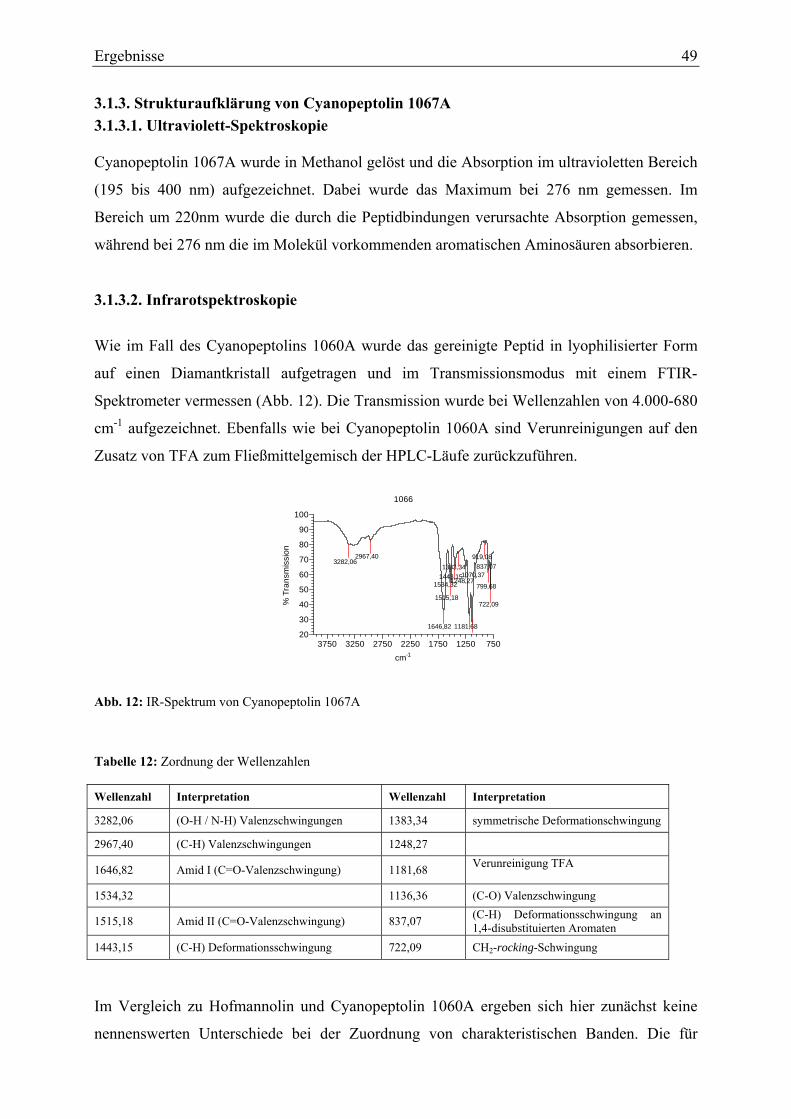
Ergebnisse 49
3.1.3. Strukturaufklärung von Cyanopeptolin 1067A 3.1.3.1. Ultraviolett-Spektroskopie Cyanopeptolin 1067A wurde in Methanol gelöst und die Absorption im ultravioletten Bereich
(195 bis 400 nm) aufgezeichnet. Dabei wurde das Maximum bei 276 nm gemessen. Im
Bereich um 220nm wurde die durch die Peptidbindungen verursachte Absorption gemessen,
während bei 276 nm die im Molekül vorkommenden aromatischen Aminosäuren absorbieren.
3.1.3.2. Infrarotspektroskopie
Wie im Fall des Cyanopeptolins 1060A wurde das gereinigte Peptid in lyophilisierter Form
auf einen Diamantkristall aufgetragen und im Transmissionsmodus mit einem FTIR-
Spektrometer vermessen (Abb. 12). Die Transmission wurde bei Wellenzahlen von 4.000-680
cm-1 aufgezeichnet. Ebenfalls wie bei Cyanopeptolin 1060A sind Verunreinigungen auf den
Zusatz von TFA zum Fließmittelgemisch der HPLC-Läufe zurückzuführen.
Abb. 12: IR-Spektrum von Cyanopeptolin 1067A
Tabelle 12: Zordnung der Wellenzahlen Wellenzahl Interpretation Wellenzahl Interpretation
3282,06 (O-H / N-H) Valenzschwingungen 1383,34 symmetrische Deformationschwingung
2967,40 (C-H) Valenzschwingungen 1248,27
1646,82 Amid I (C=O-Valenzschwingung) 1181,68 Verunreinigung TFA
1534,32 1136,36 (C-O) Valenzschwingung
1515,18 Amid II (C=O-Valenzschwingung) 837,07 (C-H) Deformationsschwingung an 1,4-disubstituierten Aromaten
1443,15 (C-H) Deformationsschwingung 722,09 CH2-rocking-Schwingung
Im Vergleich zu Hofmannolin und Cyanopeptolin 1060A ergeben sich hier zunächst keine
nennenswerten Unterschiede bei der Zuordnung von charakteristischen Banden. Die für
1066
75012501750225027503250375020
30
40
50
60
70
80
90
100
3282,062967,40
1646,82
1534,32
1515,18
1443,151383,34
1248,27
1181,68
1070,37
722,09
cm-1
% T
rans
mis
sion
799,68
837,07919,08
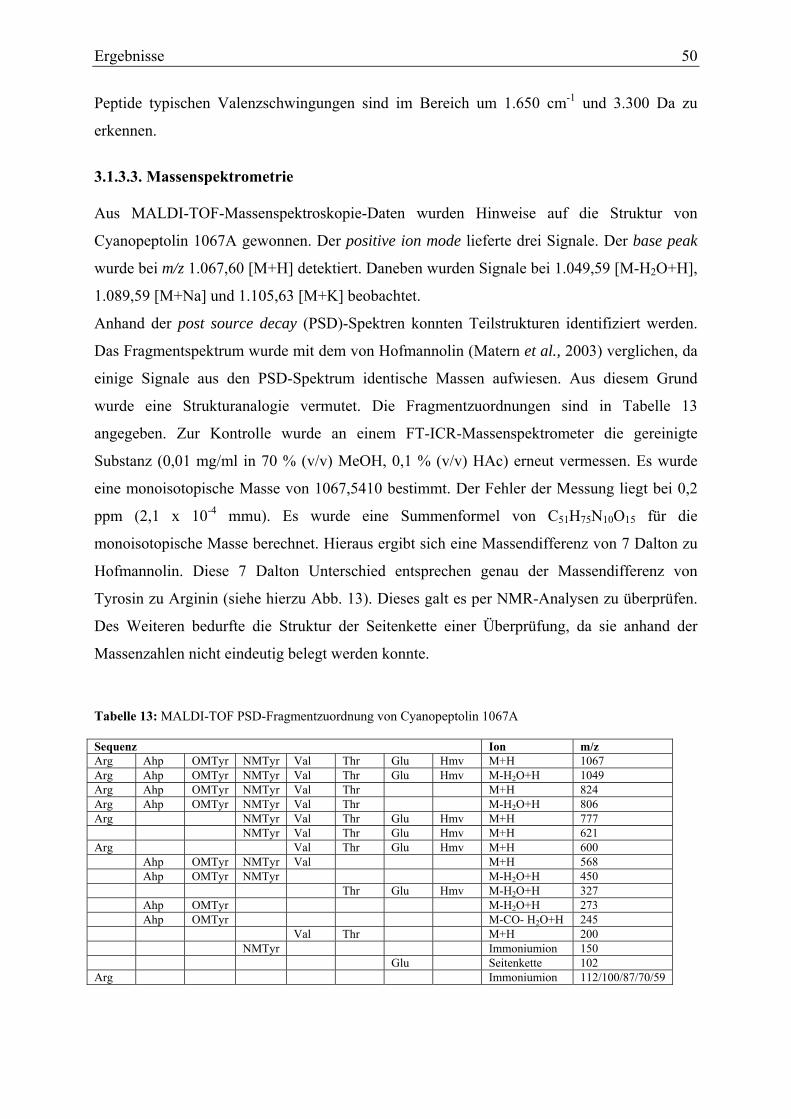
Ergebnisse 50
Peptide typischen Valenzschwingungen sind im Bereich um 1.650 cm-1 und 3.300 Da zu
erkennen.
3.1.3.3. Massenspektrometrie Aus MALDI-TOF-Massenspektroskopie-Daten wurden Hinweise auf die Struktur von
Cyanopeptolin 1067A gewonnen. Der positive ion mode lieferte drei Signale. Der base peak
wurde bei m/z 1.067,60 [M+H] detektiert. Daneben wurden Signale bei 1.049,59 [M-H2O+H],
1.089,59 [M+Na] und 1.105,63 [M+K] beobachtet.
Anhand der post source decay (PSD)-Spektren konnten Teilstrukturen identifiziert werden.
Das Fragmentspektrum wurde mit dem von Hofmannolin (Matern et al., 2003) verglichen, da
einige Signale aus den PSD-Spektrum identische Massen aufwiesen. Aus diesem Grund
wurde eine Strukturanalogie vermutet. Die Fragmentzuordnungen sind in Tabelle 13
angegeben. Zur Kontrolle wurde an einem FT-ICR-Massenspektrometer die gereinigte
Substanz (0,01 mg/ml in 70 % (v/v) MeOH, 0,1 % (v/v) HAc) erneut vermessen. Es wurde
eine monoisotopische Masse von 1067,5410 bestimmt. Der Fehler der Messung liegt bei 0,2
ppm (2,1 x 10-4 mmu). Es wurde eine Summenformel von C51H75N10O15 für die
monoisotopische Masse berechnet. Hieraus ergibt sich eine Massendifferenz von 7 Dalton zu
Hofmannolin. Diese 7 Dalton Unterschied entsprechen genau der Massendifferenz von
Tyrosin zu Arginin (siehe hierzu Abb. 13). Dieses galt es per NMR-Analysen zu überprüfen.
Des Weiteren bedurfte die Struktur der Seitenkette einer Überprüfung, da sie anhand der
Massenzahlen nicht eindeutig belegt werden konnte.
Tabelle 13: MALDI-TOF PSD-Fragmentzuordnung von Cyanopeptolin 1067A Sequenz Ion m/z Arg Ahp OMTyr NMTyr Val Thr Glu Hmv M+H 1067 Arg Ahp OMTyr NMTyr Val Thr Glu Hmv M-H2O+H 1049 Arg Ahp OMTyr NMTyr Val Thr M+H 824 Arg Ahp OMTyr NMTyr Val Thr M-H2O+H 806 Arg NMTyr Val Thr Glu Hmv M+H 777 NMTyr Val Thr Glu Hmv M+H 621 Arg Val Thr Glu Hmv M+H 600 Ahp OMTyr NMTyr Val M+H 568 Ahp OMTyr NMTyr M-H2O+H 450 Thr Glu Hmv M-H2O+H 327 Ahp OMTyr M-H2O+H 273 Ahp OMTyr M-CO- H2O+H 245 Val Thr M+H 200 NMTyr Immoniumion 150 Glu Seitenkette 102 Arg Immoniumion 112/100/87/70/59

Ergebnisse 51
3.1.3.4. Magnetische Kernresonanzspektroskopie
Die NMR-Messungen von Cyanopeptolin 1067A dokumentieren den Austausch einer
Aminosäure (siehe hierzu Abb.13). Es ergaben sich wenige Änderungen in den NMR-
Spektren im Vergleich zu Hofmannolin. Im Bereich von 6,5 bis 7,5 ppm fehlen die Signale,
welche bei Hofmannolin von Tyrosin hervorgerufen werden (Tabelle 14). Die Signale, welche
vom Peptidrückgrad gebildet werden unterscheiden sich kaum, da sich hier keine Änderungen
durch den Wechsel der Aminosäure bemerkbar machen. Man erhält allerdings eine
zusätzliche Signalgruppe, welche durch die sekundäre Amingruppe in der Seitenkette des
Arginins verursacht wird (7,73 ppm). Zusätzliche Signale erhält man durch zwei weitere
Methylengruppen (Arg4-C4/5) in Arginin (1,43 und 3,06ppm). Die NOE-Korellationen sind
mit denen von Tyrosin vergleichbar, Unterschiede ergeben sich nur innerhalb des
Spinsystems von Arginin. Die zur Seitenkette zugehörigen Signale von 2-Hydroxy-3-
methylvaleriansäure sind mit denen aus Hofmannolin kongruent (HSQC-Korrelationen, wie
H-NMR-Daten). Damit stimmen die Ergebnisse aus der NMR-Analytik mit dem
Strukturvorschlag aus der Massenspektrometrie überein, so dass die vorgeschlagene Struktur
(Abb. 13) von Cyanopeptolin 1067 A bestätigt wurde.
Abb.13: Struktur von Cyanopeptolin 1067A aus Scytonema hofmanni, Hmv: 2-Hydroxy-3-methylvaleriansäure, MTyr: N-Methyl-Tyrosin, OMTyr: O-Methyl-Tyrosin, Ahp: 3-Amino-6-Hydroxy-2-Oxo-Piperidin
N
O
NHO
O
O
NH
O
NH
OH O
OOH
NH
OH
N
OO
NH O
OH
ONH
NH
NH2
Thr3
Glu2
Hmv1
Val8
NMTyr7
Ahp5Arg4
OMTyr6
Ergebnisse 52
Abb. 14: H-NMR-Spektrum von Cyanopeptolin 1067A:
Tabelle 14: NMR-Spektroskopie und Teilstrukturen von Cyanopeptolin 1067A Aminosäure Position δ 13C
(aus HSQC) δ 1H Multiplizität, J (Hz) NOE-Korrelationen
Hmv1 1 2 75,4 3,80 dubl. (4,4) Glu2-NH 3 38,7 1,72 mult. Glu2-NH 4 23,9 1,38 mult., 1,12 mult. Glu2-NH 5 12,0 0,80 trip. (8,0) 6 15,9 0,88 Glu2-NH 2-OH 5,58 breit Glu2 1 2 52,1 4,55 mult. Thr3-NH, 3 28,6 1,98/1,88 2x mult. 4 30,4 2,26 trip. (8,0) 5 5-OH 2-NH 174,0 7,91 dubl. (8,7) Hmv1-2/3/4/6, Thr3-NH Thr3 1 2 55,3 4,61 dubl. (9,5) 3 72,4 5,45 quadr. (6,5) Arg4-NH, Ahp5-NH 4 18,3 1,17 dubl. (6,6) 2-NH 172,7 8,12 dubl. (8,8) Glu2-2, Glu2-NH Arg4 1 2 52,1 4,22 mult. Ahp5-NH 3 27,8 1,91 mult., 1,43 mult. 4 25,5 1,43 mult. Thr3-3 5 40,7 3,06 6(NH) 7,73 trip. (5,2) 7 NH, NH2 2-NH 169,5 8,57 dubl. (8,8) Thr3-3; Ahp5-NH Ahp5 1 2 49,1 3,71 mult. 3 22,0 2,41/1,62 2x mult. 4 29,7 1,72/1,60 2x mult. 5 74,3 5,06 sing. Tyr6-3, Tyr6-2´/6´ 5-OH 6,03 sing. 2-NH 170,3 7,16 dubl. (9,6) Arg4-2, Thr3-3, Arg4-NH Tyr6 1 2 51,0 4,70 mult. mTyr7-2, mTyr7-NCH3 3 34,8 2,79/1,75 2x mult. Ahp5-5
Ergebnisse 53
1´ 2´/6´ 131,0 6,74 sing. Ahp5-5 3´/5´ 113,8 6,74 sing. 4´ OCH3 55,3 3,70 sing. mTyr7 1 2 61,3 4,91 dubl. (11,7) Tyr6-2, Val8-NH 3 33,4 3,10/2,71 2x mult. 2´/6´ 131,0 6,99 dubl. (8,0) 3´/5´ 115,5 6,79 dubl. (8,1) 4´ 4´-OH 9,45 sing. NCH3 30,8 2,75 sing. Val8-NH, Tyr6-2, Val8-4 Val8 1 2 56,3 4,73 mult. 3 31,2 2,07 mult. 4 19,7 0,86 dubl. (7,3) mTyr7-NCH3 5 17,8 0,73 dubl. (6,6) 2-NH 169,5 7,45 dubl. (9,5) mTyr7-NCH3, mTyr7-2 Korrelationen innerhalb des Spinsystems einer Aminosäure werden nicht aufgeführt.
3.1.4. Strukturaufklärung von Cyanopeptolin 946A 3.1.4.1. Ultraviolett-Spektroskopie Mit Cyanopeptolin 946A wurde ein weiteres bisher unbekanntes Cyanopeptolin aus
Scytonema hofmanni PCC 7110 isoliert. Während der HPLC-Auftrennung zeigte
Cyanopeptolin 946A das für Cyanopeptoline typische UV-Absorptionspektrum. Die Maxima
(202 nm sowie 276nm) wurden aus den Datensätzen der HPLC-Auftrennung übernommen.
Absorption bei 220 nm weist auf Peptidbindung, 276 nm lässt aromatische Aminosäuren
vermuten.
3.1.4.2. Massenspektrometrie Erste Hinweise auf die Struktur von Cyanopeptolin 946A lieferten MALDI-TOF-
Massenspektroskopie-Daten. Hierzu wurde die Probe wiederum im positive ion mode
analysiert. Es wurden drei Signale detektiert. Der base peak wurde bei m/z 969,40 [M+Na]
detektiert. Daneben wurden Signale bei 951,39 [M-H2O+Na] und 929,39 [M-H2O+H]
beobachtet.
Anhand der post source decay (PSD)-Spektren konnten Teilstrukturen identifiziert werden.
Das Fragmentspektrum wurde mit denen der Scyptoline (Matern et al., 2001) verglichen, da
einige Signale aus den PSD-Spektrum identische Massen aufwiesen. Aus diesem Grund
wurde eine Strukturanalogie vermutet. Die Fragmentzuordnungen sind in Tabelle 15
angegeben. Zur Kontrolle wurde an einem FT-ICR-Massenspektrometer die gereinigte
Substanz (0,01 mg/ml in 70 % (v/v) MeOH, 0,1 % (v/v) HAc) erneut vermessen. Es wurde
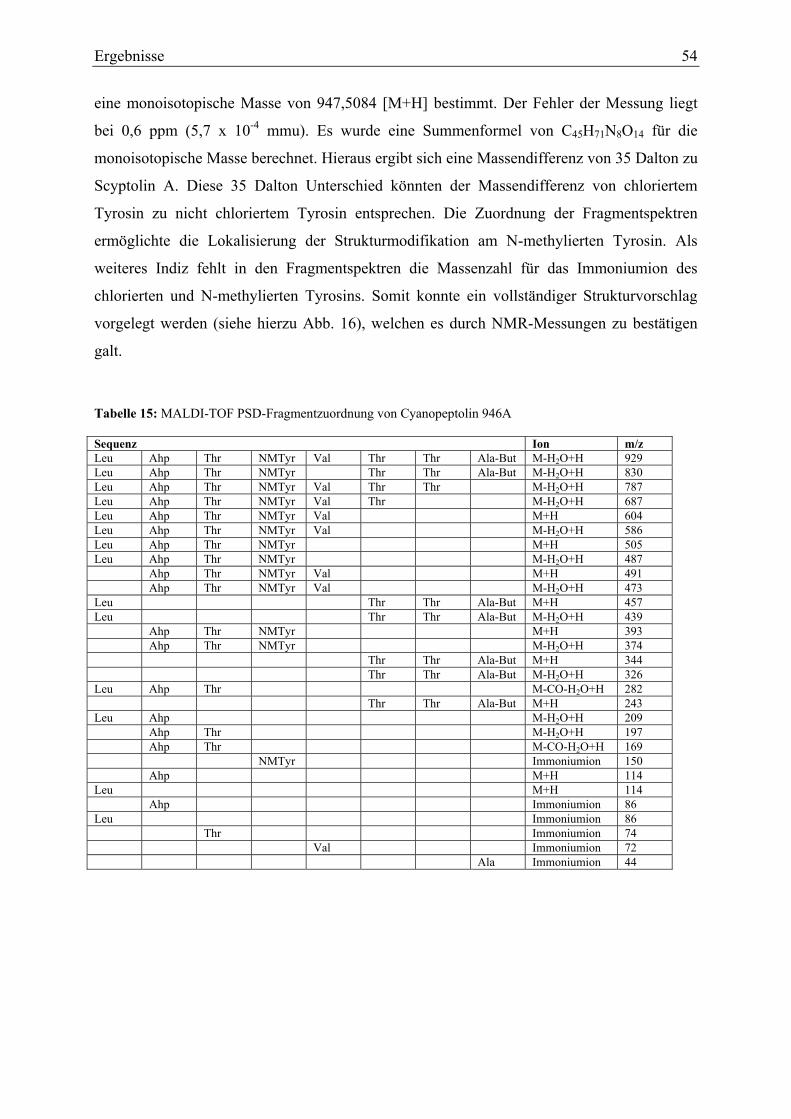
Ergebnisse 54
eine monoisotopische Masse von 947,5084 [M+H] bestimmt. Der Fehler der Messung liegt
bei 0,6 ppm (5,7 x 10-4 mmu). Es wurde eine Summenformel von C45H71N8O14 für die
monoisotopische Masse berechnet. Hieraus ergibt sich eine Massendifferenz von 35 Dalton zu
Scyptolin A. Diese 35 Dalton Unterschied könnten der Massendifferenz von chloriertem
Tyrosin zu nicht chloriertem Tyrosin entsprechen. Die Zuordnung der Fragmentspektren
ermöglichte die Lokalisierung der Strukturmodifikation am N-methylierten Tyrosin. Als
weiteres Indiz fehlt in den Fragmentspektren die Massenzahl für das Immoniumion des
chlorierten und N-methylierten Tyrosins. Somit konnte ein vollständiger Strukturvorschlag
vorgelegt werden (siehe hierzu Abb. 16), welchen es durch NMR-Messungen zu bestätigen
galt.
Tabelle 15: MALDI-TOF PSD-Fragmentzuordnung von Cyanopeptolin 946A Sequenz Ion m/z Leu Ahp Thr NMTyr Val Thr Thr Ala-But M-H2O+H 929 Leu Ahp Thr NMTyr Thr Thr Ala-But M-H2O+H 830 Leu Ahp Thr NMTyr Val Thr Thr M-H2O+H 787 Leu Ahp Thr NMTyr Val Thr M-H2O+H 687 Leu Ahp Thr NMTyr Val M+H 604 Leu Ahp Thr NMTyr Val M-H2O+H 586 Leu Ahp Thr NMTyr M+H 505 Leu Ahp Thr NMTyr M-H2O+H 487 Ahp Thr NMTyr Val M+H 491 Ahp Thr NMTyr Val M-H2O+H 473 Leu Thr Thr Ala-But M+H 457 Leu Thr Thr Ala-But M-H2O+H 439 Ahp Thr NMTyr M+H 393 Ahp Thr NMTyr M-H2O+H 374 Thr Thr Ala-But M+H 344 Thr Thr Ala-But M-H2O+H 326 Leu Ahp Thr M-CO-H2O+H 282 Thr Thr Ala-But M+H 243 Leu Ahp M-H2O+H 209 Ahp Thr M-H2O+H 197 Ahp Thr M-CO-H2O+H 169 NMTyr Immoniumion 150 Ahp M+H 114 Leu M+H 114 Ahp Immoniumion 86 Leu Immoniumion 86 Thr Immoniumion 74 Val Immoniumion 72 Ala Immoniumion 44

Ergebnisse 55
3.1.4.3. Magnetische Kernresonanzspektroskopie Die Integration des 1H-NMR Spektrums von Cyanopeptolin 946A lässt 70 Protonen
vermuten, welche in einem Bereich bis 11 ppm Resonanzsignale verursachten. Zusammen mit
den Daten aus HSQC-Messungen ließen sich 34 verschiedene C-H-Korrelationen bestehend
aus 10 Methyl-, 6-Methylen- und 18 Methingruppen unterscheiden. Neben diesen an
Kohlenstoffatome gebundenen Protonen gibt es weitere 10 Protonen, welche Resonanzsignale
verursachen. Eine Zuordnung zu 6 NH-Gruppen, einer phenolischen und drei alkoholischen
Funktionen konnte getroffen werden. Bei den Methylgruppen konnte man eine Unterteilung
anhand der Nachbarstrukturen vornehmen. Dabei handelt es sich um eine primäre
(Aufspaltung des Signals zum Triplett), acht sekundäre (Dubletts) und eine N-Methyl-Gruppe
(Singulett). Die chemischen Verschiebungen im 1H- und HSQC-Spektrum belegen, dass alle
sechs Methylengruppen an Kohlenstoff gebunden vorliegen. Bezüglich der Methingruppen
ließ sich die Aussage treffen, dass vier an Sauerstoff und acht an Stickstoff gebunden sind.
Die restlichen Methingruppen verteilen sich auf vier aromatische und zwei aliphatische
Gruppen. Bei der aromatischen Aminosäure ergeben sich durch para-Substitution für die 4
Protonen jeweils zwei chemisch äquivalente Protonenpaare. Es handelt sich um ein AA`BB`-
Spinsystem. Die Signale werden durch die Nachbarschaft eines weiteren Protons in Dubletts
aufgespaltet. Aus den Spinsystemen der TOCSY-Experimente und den 13C-Korrelationen in
den HSQC-Spektren konnten N-acyliertes Alanin, Leucin, Valin und Threonin Reste neben
O-acyliertem Threonin, N-alkyliertem Threonin und N-alkyliertem Tyrosin bestimmt werden
(siehe hierzu Abb. 16).
In Übereinstimmung mit Hauptstrukturmerkmalen für Cyanopeptoline lag das Ahp-
Ringsystem vor. Die NOE-Korrelationen dieser Teilstruktur entsprachen der Konfiguration
früherer Publikationen (Weckesser et al., 1996). Weitere Übereinstimmung zu bekannten
Cyanopeptolinen konnte durch α-α-NOE-Korrelationen bei der cis-Konfiguration der
Amidbindung zwischen Thr7 und N-Methyl-Tyrosin nachgewiesen werden, während die
restlichen Amidbindungen trans-konfiguriert vorliegen. HMBC-Messungen belegten die
Esterbrückenbildung von Val9 zu Thr4 entsprechend beobachtet von Matern et al. (2001 und
2003). Die Seitenkette mit der Butyryl-Gruppe ergab ein Spinsystem entsprechend einer
Propyleinheit. Die aus den NMR-Messungen gewonnenen Strukturdaten stimmen mit dem
aus den MALDI-TOF-Ergebnissen gewonnenen Strukturvorschlag vollständig überein. Somit
ergeben sich für Cyanopeptolin 946A ein Molekulargewicht von 946 Da und eine
Summenformel von C45H70N8O14.
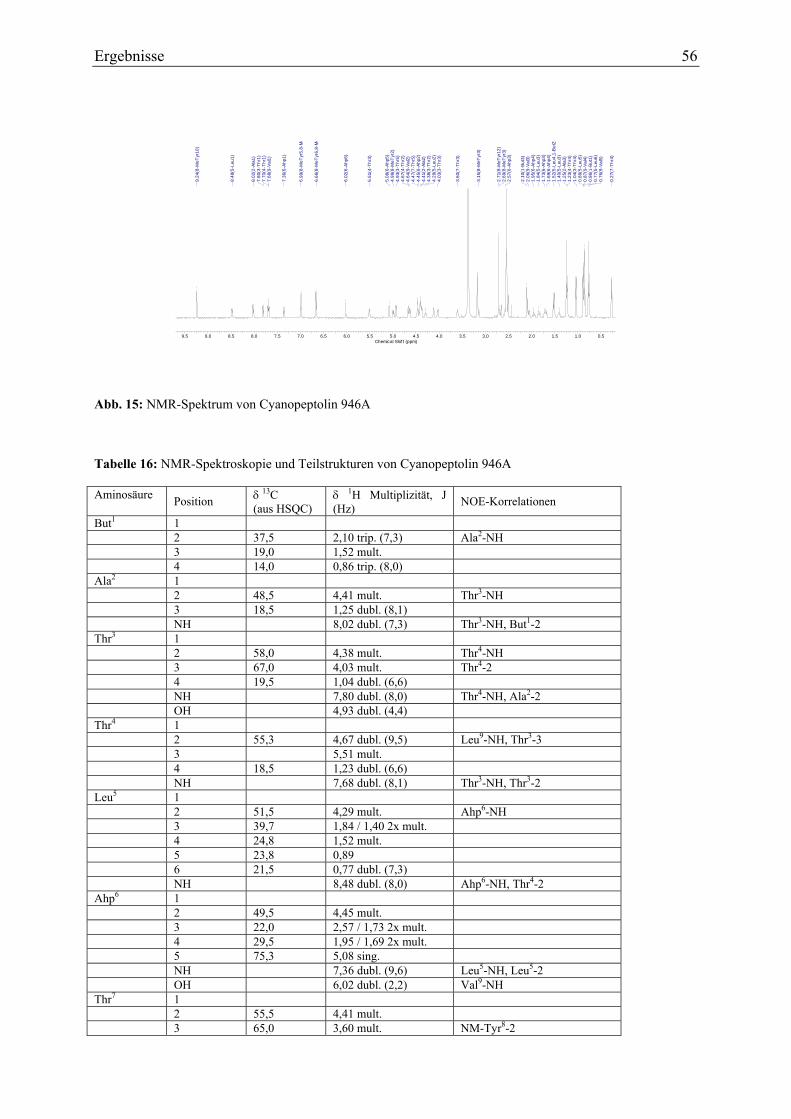
Ergebnisse 56
Abb. 15: NMR-Spektrum von Cyanopeptolin 946A
Tabelle 16: NMR-Spektroskopie und Teilstrukturen von Cyanopeptolin 946A Aminosäure Position δ 13C
(aus HSQC) δ 1H Multiplizität, J (Hz) NOE-Korrelationen
But1 1 2 37,5 2,10 trip. (7,3) Ala2-NH 3 19,0 1,52 mult. 4 14,0 0,86 trip. (8,0) Ala2 1 2 48,5 4,41 mult. Thr3-NH 3 18,5 1,25 dubl. (8,1) NH 8,02 dubl. (7,3) Thr3-NH, But1-2 Thr3 1 2 58,0 4,38 mult. Thr4-NH 3 67,0 4,03 mult. Thr4-2 4 19,5 1,04 dubl. (6,6) NH 7,80 dubl. (8,0) Thr4-NH, Ala2-2 OH 4,93 dubl. (4,4) Thr4 1 2 55,3 4,67 dubl. (9,5) Leu9-NH, Thr3-3 3 5,51 mult. 4 18,5 1,23 dubl. (6,6) NH 7,68 dubl. (8,1) Thr3-NH, Thr3-2 Leu5 1 2 51,5 4,29 mult. Ahp6-NH 3 39,7 1,84 / 1,40 2x mult. 4 24,8 1,52 mult. 5 23,8 0,89 6 21,5 0,77 dubl. (7,3) NH 8,48 dubl. (8,0) Ahp6-NH, Thr4-2 Ahp6 1 2 49,5 4,45 mult. 3 22,0 2,57 / 1,73 2x mult. 4 29,5 1,95 / 1,69 2x mult. 5 75,3 5,08 sing. NH 7,36 dubl. (9,6) Leu5-NH, Leu5-2 OH 6,02 dubl. (2,2) Val9-NH Thr7 1 2 55,5 4,41 mult. 3 65,0 3,60 mult. NM-Tyr8-2
9.5 9.0 8.5 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5Chemical Shif t (ppm)
9.24
(8-M
eTyr
10)
8.48
(5-L
eu1)
8.02
(2-A
la1)
7.80
(3-T
hr1)
7.70
(4-T
hr1)
7.68
(9-V
al1)
7.36
(6-A
hp1)
6.99
(8-M
eTyr
5,8-
Me
6.66
(8-M
eTyr
6,8-
Me
6.02
(6-A
hp6)
5.51
(4-T
hr3)
5.08
(6-A
hp5)
4.99
(8-M
eTyr
2)4.
93(3
-Thr
5)4.
67(4
-Thr
2)4.
64(9
-Val
2)4.
47(7
-Thr
5)4.
45(6
-Ahp
2)4.
41(2
-Ala
2)4.
38(3
-Thr
2)4.
29(5
-Leu
2)4.
03(3
-Thr
3)
3.60
(7-T
hr3)
3.15
(8-M
eTyr
3)
2.71
(8-M
eTyr
12)
2.69
(8-M
eTyr
3)2.
57(6
-Ahp
3)
2.10
(1-B
ut3)
2.06
(9-V
al3)
1.95
(6-A
hp4)
1.84
(5-L
eu3)
1.73
(6-A
hp3)
1.69
(6-A
hp4)
1.52
(5-L
eu4,
1-B
ut2)
1.40
(5-L
eu3)
1.25
(2-A
la3)
1.23
(4-T
hr4)
1.04
(3-T
hr4)
0.89
(5-L
eu5)
0.87
(9-V
al4)
0.86
(1-B
ut1)
0.77
(5-L
eu6)
0.76
(9-V
al5)
0.27
(7-T
hr4)

Ergebnisse 57
4 19,8 0,27 dubl. (5,9) OH 4,47 dubl. (4,4) NM-Tyr8 1 2 61,0 4,99 mult. Val9-NH Thr7-2 3 33,5 3,15 / 2,69 2x mult. 2´/6´ 132,0 6,99 dubl. (8,8) 3´/5´ 116,0 6,66 dubl. (8,1) 4´-OH 9,24 sing. NCH3 30,3 2,71 sing. Val9-NH Val9 1 2 56,5 4,64 mult. 3 31,0 2,06 mult. 4 19,8 0,87 mult. 5 18,0 0,76 dubl. (7,4) NH 7,68 dubl. (8,0) Ahp6-OH, NM-Tyr8-2 / -NCH3 Korrelationen innerhalb des Spinsystems einer Aminosäure werden nicht aufgeführt.
Abb. 16: Strukturvorschlag zu Cyanopeptolin 946A aus Scytonema hofmanni, But: Butyryl, NMTyr: N-Methyl-Tyrosin, Ahp: 3-Amino-6-Hydroxy-2-Oxo-Piperidin
Cyanopeptolin 946A hat die typischen Merkmale eines Cyanopeptolins, wie Ahp-Gruppe,
Lactonblidung über Threonin, 19-gliedrige Ringstruktur. Die Struktur unterscheidet sich von
der von Scyptolin A (Matern et al., 2001) durch Fehlen der Chlorierung an N-Methyltyrosin.
3.1.5. Strukturaufklärung von Scytocyclamid A 3.1.5.1. Ultraviolett-Spektroskopie Während der HPLC-Auftrennung wurde für Scytocyclamide A ein für cyanobakterielle
Peptide aus Scytonema hofmanni untypisches UV-Spektrum gemessen. Es waren keine
Maxima in dem für Peptide typischen Spektralbereich von 200 bis 300 nm sichtbar. Im
Bereich um 220 nm verursachen die Peptidbindungen im Molekül Absorption, im Bereich um
280 nm typischerweise aromatische Strukturelemente. Scytocyclamid A wies eine von 200 bis
250 nm sinkende Absorption auf. Bei 220 nm war kein Maximum messbar, jedoch wurde
N
O
NHO
O
O
NH
O
NH
ONH
OH
N
OO
NH O
OHOH
OH
NH
OBut1 Ala2 Thr3
Leu5 Ahp6
Thr7
Val9
NMTyr8
Thr4
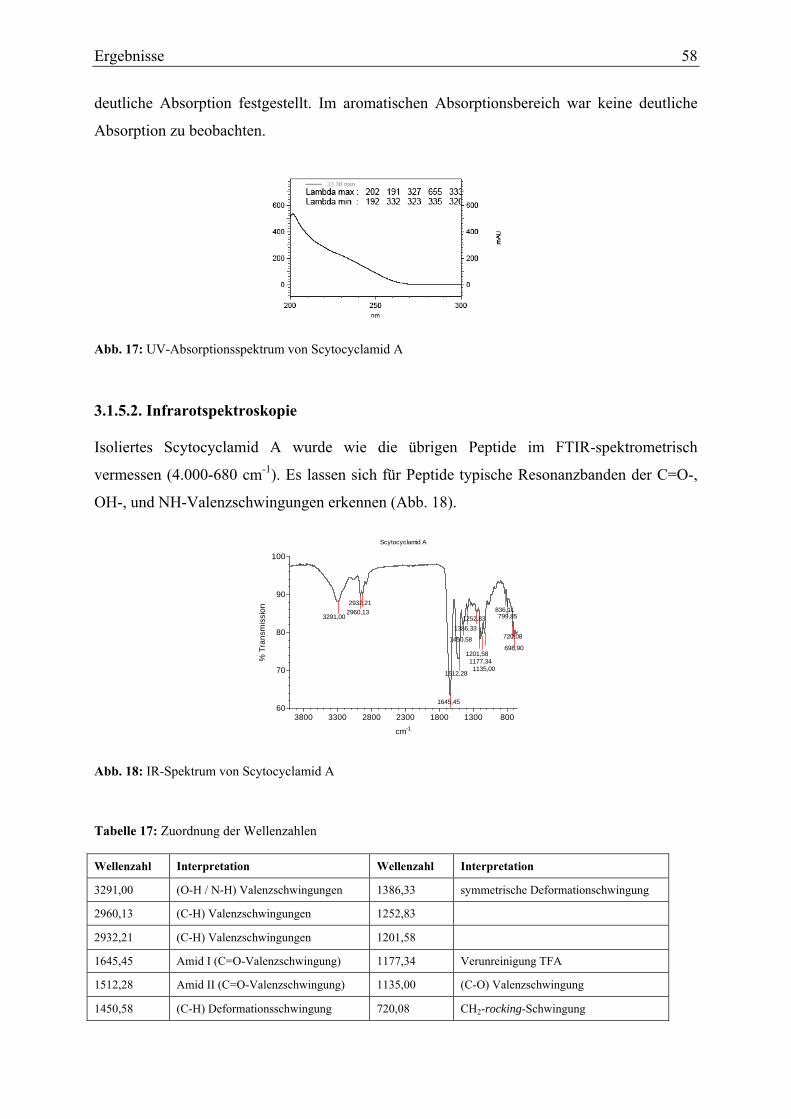
Ergebnisse 58
deutliche Absorption festgestellt. Im aromatischen Absorptionsbereich war keine deutliche
Absorption zu beobachten.
Abb. 17: UV-Absorptionsspektrum von Scytocyclamid A
3.1.5.2. Infrarotspektroskopie Isoliertes Scytocyclamid A wurde wie die übrigen Peptide im FTIR-spektrometrisch
vermessen (4.000-680 cm-1). Es lassen sich für Peptide typische Resonanzbanden der C=O-,
OH-, und NH-Valenzschwingungen erkennen (Abb. 18).
Abb. 18: IR-Spektrum von Scytocyclamid A Tabelle 17: Zuordnung der Wellenzahlen Wellenzahl Interpretation Wellenzahl Interpretation
3291,00 (O-H / N-H) Valenzschwingungen 1386,33 symmetrische Deformationschwingung
2960,13 (C-H) Valenzschwingungen 1252,83
2932,21 (C-H) Valenzschwingungen 1201,58
1645,45 Amid I (C=O-Valenzschwingung) 1177,34 Verunreinigung TFA
1512,28 Amid II (C=O-Valenzschwingung) 1135,00 (C-O) Valenzschwingung
1450,58 (C-H) Deformationsschwingung 720,08 CH2-rocking-Schwingung
Scytocyclamid A
80013001800230028003300380060
70
80
90
100
3291,002960,132932,21
1645,45
1512,28
1450,58
1386,331252,83
1201,581177,341135,00
836,11799,85
720,08
698,90
cm-1
% T
rans
mis
sion

Ergebnisse 59
3.1.5.3. Massenspektrometrie Teil I Scytocyclamid A wurde mittels MALDI-TOF-Massenspektroskopie analysiert. Im positive
ion mode wurden drei Signale detektiert. Der base peak wurde bei m/z 1.245,83 [M+Na]
detektiert. Daneben wurden Signale bei 1.223,83 [M+H] und 1.261,81 [M+K] beobachtet.
Anhand der post source decay (PSD)-Spektren wurden Strukturhinweise gesucht. So wie das
unbekannte UV-Spektrum zeigten auch die PSD-Spektren keine Ähnlichkeit zu bereits
isolierten Peptiden aus Scytonema hofmanni. Die Fragmentspektren zeigten jedoch für
Aminosäuren typische Massenzahlen, so dass man von einer Peptidstruktur ausgehen konnte
(siehe hierzu Abb. 24). Ausgehend vom Molekulargewicht von 1.223 Da für das
monoisotopische Signal [M+H] wurde eine Struktur mit 11 bis 12 Aminosäuren erwartet. Aus
dem PSD-Spektrum wurden Leucin, bzw. Isoleucin, Phenylalanin und Glycin als Bausteine
über die Fragmentmasse ihrer Immoniumionen abgeleitet. Weitere Fragmente konnten zuerst
nicht zugeordnet werden. Es wurden NMR-Untersuchungen vorgenommen um weitere
Strukturelemente zu identifizieren.
3.1.5.4. Magnetische Kernresonanzspektroskopie Umfangreiche NMR-Analysen (1H-NMR-, COSY-, ROESY-, HSQC-Messungen) in
DMSO-d6 ergaben eine Summenformel von C61H98N12O14 und ließen eine Zusammensetzung
aus 11 bis 12 Aminosäuren vermuten. Es wurden die folgenden Aminosäuren bestimmt: 2
Leucine, 2 Isoleucine, Homoserin, Glutamin, Phenylalanin, Glycin, 4-Hydroxyprolin,
Dehydrobutyrin sowie 3-Aminooctansäure. Die Sequenz der Aminosäuren wurde durch NOE-
Korrelationen aus dem ROESY-Spektrum bestimmt. Dabei wurden Sequenz-NOE-Effekte
zwischen den α-Protonen einer Aminosäure-Einheit und dem NH-Proton der nachfolgenden
Aminosäure beobachtet: zwischen 2-H von Homoserin und NH von Phenylalanin, 2-H von
Isoleucin8 und NH von Isoleucin9, von 2-H von Isoleucin9 und NH von Leucin10, zwischen
2-H von Leucin10 und NH von Gly11 sowie von 2-H von Gly11 und NH von Aoc1. Die
letztgenannte Korrelation spiegelt den Ringschluss wieder. Des Weiteren konnten NOE-
Korrelationen zwischen NH-Protonen benachbarter Aminosäuren beobachtet werden: Aoc1 zu
Gly11und Phe6 zu Leu7. Dehydrobutyrin besitzt kein α-Proton, jedoch konnte eine Kopplung
der Protonen der Methylgruppe zum α-Proton des Hydroxyprolins zugeordnet werden. Für
den Dehydrobutyryl-Rest kann aus starken NOEs zwischen dem NH und dem 3-H der Dhb-
Einheit die E-Konfiguration abgeleitet werden. Anhand der Auswertungen wurde die Struktur
cyclo-(βAoc-Gln-E-Dhb-Hyp-Hse-Phe-Leu-Ile-Ile-Leu-Gly) vorgeschlagen (siehe hierzu
Abb. 24).
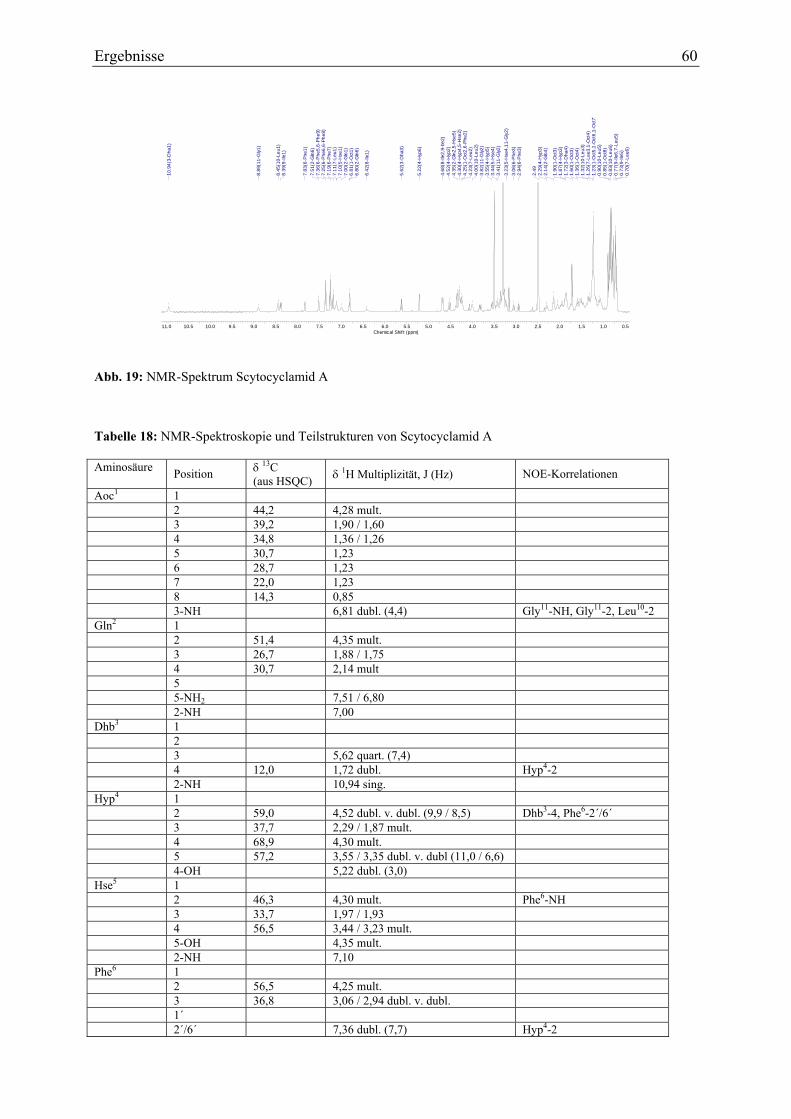
Ergebnisse 60
Abb. 19: NMR-Spektrum Scytocyclamid A
Tabelle 18: NMR-Spektroskopie und Teilstrukturen von Scytocyclamid A Aminosäure Position δ 13C
(aus HSQC) δ 1H Multiplizität, J (Hz) NOE-Korrelationen
Aoc1 1 2 44,2 4,28 mult. 3 39,2 1,90 / 1,60 4 34,8 1,36 / 1,26 5 30,7 1,23 6 28,7 1,23 7 22,0 1,23 8 14,3 0,85 3-NH 6,81 dubl. (4,4) Gly11-NH, Gly11-2, Leu10-2 Gln2 1 2 51,4 4,35 mult. 3 26,7 1,88 / 1,75 4 30,7 2,14 mult 5 5-NH2 7,51 / 6,80 2-NH 7,00 Dhb3 1 2 3 5,62 quart. (7,4) 4 12,0 1,72 dubl. Hyp4-2 2-NH 10,94 sing. Hyp4 1 2 59,0 4,52 dubl. v. dubl. (9,9 / 8,5) Dhb3-4, Phe6-2´/6´ 3 37,7 2,29 / 1,87 mult. 4 68,9 4,30 mult. 5 57,2 3,55 / 3,35 dubl. v. dubl (11,0 / 6,6) 4-OH 5,22 dubl. (3,0) Hse5 1 2 46,3 4,30 mult. Phe6-NH 3 33,7 1,97 / 1,93 4 56,5 3,44 / 3,23 mult. 5-OH 4,35 mult. 2-NH 7,10 Phe6 1 2 56,5 4,25 mult. 3 36,8 3,06 / 2,94 dubl. v. dubl. 1´ 2´/6´ 7,36 dubl. (7,7) Hyp4-2
11.0 10.5 10.0 9.5 9.0 8.5 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5Chemical Shif t (ppm)
10.9
4(3-
Dha
1)
8.89
(11-
Gly1
)
8.45
(10-
Leu1
)8.
39(9
-Ile1
)
7.83
(6-P
he1)
7.51
(2-G
ln6)
7.36
(6-P
he5,
6-Ph
e9)
7.25
(6-P
he6,
6-Ph
e8)
7.19
(6-P
he7)
7.11
(7-L
eu1)
7.10
(5-H
se1)
7.00
(2-G
ln1)
6.81
(1-O
ct1)
6.80
(2-G
ln6)
6.42
(8-Il
e1)
5.62
(3-D
ha3)
5.22
(4-H
yp6)
4.68
(8-Il
e2,9
-Ile2
)4.
52(4
-Hyp
2)4.
35(2
-Gln
2,5-
Hse
5)4.
30(4
-Hyp
4,5-
Hse
2)4.
25(1
-Oct
2,6-
Phe2
)4.
23(7
-Leu
2)4.
00(1
0-Le
u2)
3.82
(11-
Gly2
)3.
55(4
-Hyp
5)3.
44(5
-Hse
4)3.
41(1
1-G
ly2)
3.23
(5-H
se4,
11-G
ly2)
3.06
(6-P
he3)
2.94
(6-P
he3)
2.49
2.29
(4-H
yp3)
2.14
(2-G
ln4)
1.90
(1-O
ct3)
1.87
(4-H
yp3)
1.72
(3-D
ha4)
1.60
(1-O
ct3)
1.36
(1-O
ct4)
1.32
(10-
Leu3
)1.
26(7
-Leu
3,1-
Oct
4)1.
23(1
-Oct
5,1-
Oct
6,1-
Oct
7)0.
90(1
0-Le
u5)
0.85
(1-O
ct8)
0.83
(10-
Leu6
)0.
77(9
-Ile6
,7-L
eu5)
0.73
(9-Il
e5)
0.70
(7-L
eu6)
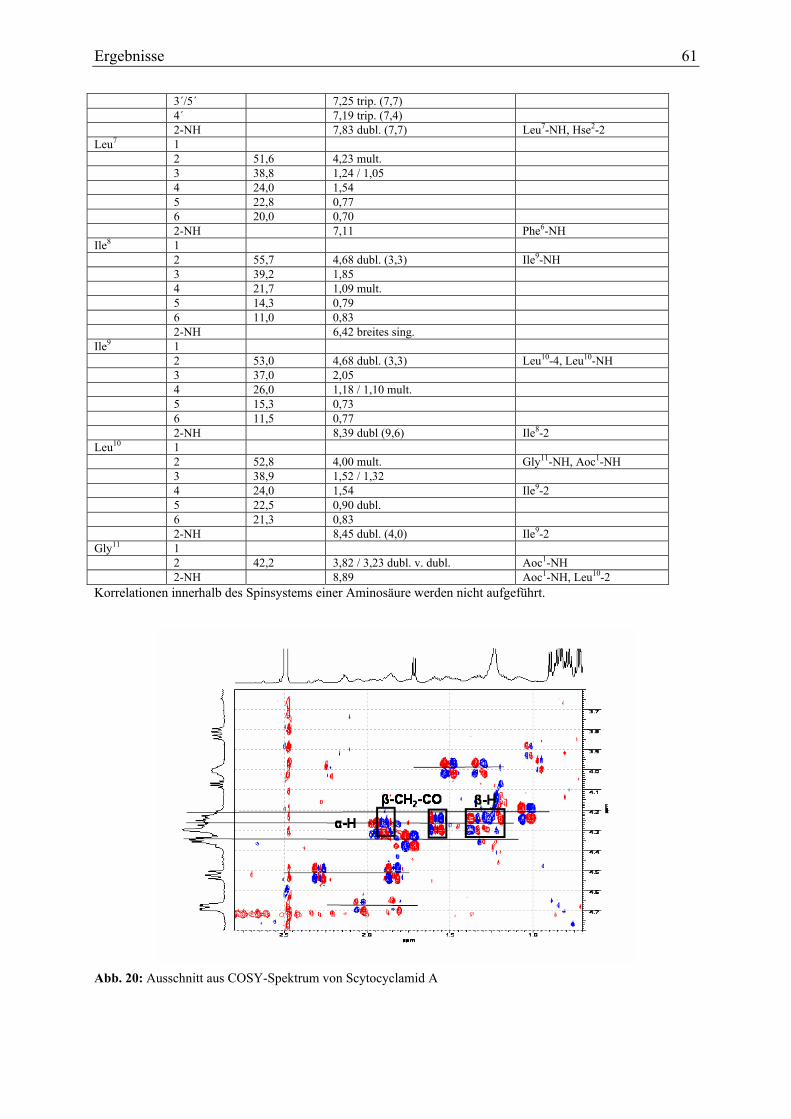
Ergebnisse 61
3´/5´ 7,25 trip. (7,7) 4´ 7,19 trip. (7,4) 2-NH 7,83 dubl. (7,7) Leu7-NH, Hse2-2 Leu7 1 2 51,6 4,23 mult. 3 38,8 1,24 / 1,05 4 24,0 1,54 5 22,8 0,77 6 20,0 0,70 2-NH 7,11 Phe6-NH Ile8 1 2 55,7 4,68 dubl. (3,3) Ile9-NH 3 39,2 1,85 4 21,7 1,09 mult. 5 14,3 0,79 6 11,0 0,83 2-NH 6,42 breites sing. Ile9 1 2 53,0 4,68 dubl. (3,3) Leu10-4, Leu10-NH 3 37,0 2,05 4 26,0 1,18 / 1,10 mult. 5 15,3 0,73 6 11,5 0,77 2-NH 8,39 dubl (9,6) Ile8-2 Leu10 1 2 52,8 4,00 mult. Gly11-NH, Aoc1-NH 3 38,9 1,52 / 1,32 4 24,0 1,54 Ile9-2 5 22,5 0,90 dubl. 6 21,3 0,83 2-NH 8,45 dubl. (4,0) Ile9-2 Gly11 1 2 42,2 3,82 / 3,23 dubl. v. dubl. Aoc1-NH 2-NH 8,89 Aoc1-NH, Leu10-2 Korrelationen innerhalb des Spinsystems einer Aminosäure werden nicht aufgeführt.
Abb. 20: Ausschnitt aus COSY-Spektrum von Scytocyclamid A

Ergebnisse 62
Abb. 21: Ausschnitt aus TOCSY-Spektrum von Scytocyclamid A. Aus den gezeigten Ausschnitten der COSY- und TOCSY-Spektren ließ sich die β-Stellung der Aminogruppe der Amino-Octansäure bestimmen.
Konfiguration von Hydroxyprolin in Scytocyclamid A
Starke NOE-Effekte werden zwischen 2-H und 3-H und zwischen 3-H`und 4-H beobachtet.
Anhand dieser NOE-Effekte lässt sich die Aussage treffen, dass das Hydroxyprolin-alpha
Proton auf die gleiche Seite schaut, wie das gamma-OH. Hieraus ergibt sich das trans-
Diastereomere, trans-4-Hydroxyprolin in der (2S 4R)-Konfiguration (Abb. 22).
Abb. 22: NOE-Korrelationen am Hydroxyprolin (s=strong, m=medium, w=weak)
3.1.5.5. Massenspektrometrie Teil II Anhand der Struktur aus den NMR-Daten wurde die vorgeschlagene Sequenz durch
Zuordnungen der Fragmentmassen aus der MALDI-TOF-Analyse von Scytocyclamid A
bestätigt. Aus den Zuordnungen der Massenzahlen in Tabelle 18 lassen sich unter anderem
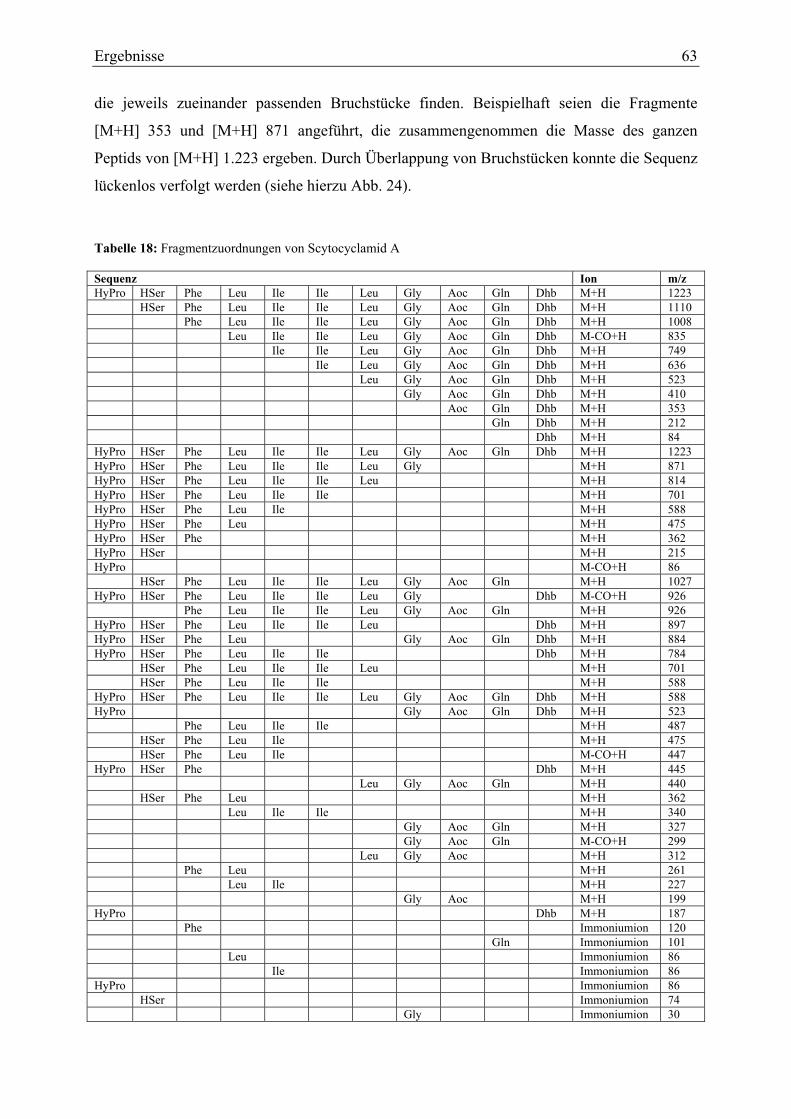
Ergebnisse 63
die jeweils zueinander passenden Bruchstücke finden. Beispielhaft seien die Fragmente
[M+H] 353 und [M+H] 871 angeführt, die zusammengenommen die Masse des ganzen
Peptids von [M+H] 1.223 ergeben. Durch Überlappung von Bruchstücken konnte die Sequenz
lückenlos verfolgt werden (siehe hierzu Abb. 24).
Tabelle 18: Fragmentzuordnungen von Scytocyclamid A Sequenz Ion m/z HyPro HSer Phe Leu Ile Ile Leu Gly Aoc Gln Dhb M+H 1223 HSer Phe Leu Ile Ile Leu Gly Aoc Gln Dhb M+H 1110 Phe Leu Ile Ile Leu Gly Aoc Gln Dhb M+H 1008 Leu Ile Ile Leu Gly Aoc Gln Dhb M-CO+H 835 Ile Ile Leu Gly Aoc Gln Dhb M+H 749 Ile Leu Gly Aoc Gln Dhb M+H 636 Leu Gly Aoc Gln Dhb M+H 523 Gly Aoc Gln Dhb M+H 410 Aoc Gln Dhb M+H 353 Gln Dhb M+H 212 Dhb M+H 84 HyPro HSer Phe Leu Ile Ile Leu Gly Aoc Gln Dhb M+H 1223 HyPro HSer Phe Leu Ile Ile Leu Gly M+H 871 HyPro HSer Phe Leu Ile Ile Leu M+H 814 HyPro HSer Phe Leu Ile Ile M+H 701 HyPro HSer Phe Leu Ile M+H 588 HyPro HSer Phe Leu M+H 475 HyPro HSer Phe M+H 362 HyPro HSer M+H 215 HyPro M-CO+H 86 HSer Phe Leu Ile Ile Leu Gly Aoc Gln M+H 1027 HyPro HSer Phe Leu Ile Ile Leu Gly Dhb M-CO+H 926 Phe Leu Ile Ile Leu Gly Aoc Gln M+H 926 HyPro HSer Phe Leu Ile Ile Leu Dhb M+H 897 HyPro HSer Phe Leu Gly Aoc Gln Dhb M+H 884 HyPro HSer Phe Leu Ile Ile Dhb M+H 784 HSer Phe Leu Ile Ile Leu M+H 701 HSer Phe Leu Ile Ile M+H 588 HyPro HSer Phe Leu Ile Ile Leu Gly Aoc Gln Dhb M+H 588 HyPro Gly Aoc Gln Dhb M+H 523 Phe Leu Ile Ile M+H 487 HSer Phe Leu Ile M+H 475 HSer Phe Leu Ile M-CO+H 447 HyPro HSer Phe Dhb M+H 445 Leu Gly Aoc Gln M+H 440 HSer Phe Leu M+H 362 Leu Ile Ile M+H 340 Gly Aoc Gln M+H 327 Gly Aoc Gln M-CO+H 299 Leu Gly Aoc M+H 312 Phe Leu M+H 261 Leu Ile M+H 227 Gly Aoc M+H 199 HyPro Dhb M+H 187 Phe Immoniumion 120 Gln Immoniumion 101 Leu Immoniumion 86 Ile Immoniumion 86 HyPro Immoniumion 86 HSer Immoniumion 74 Gly Immoniumion 30
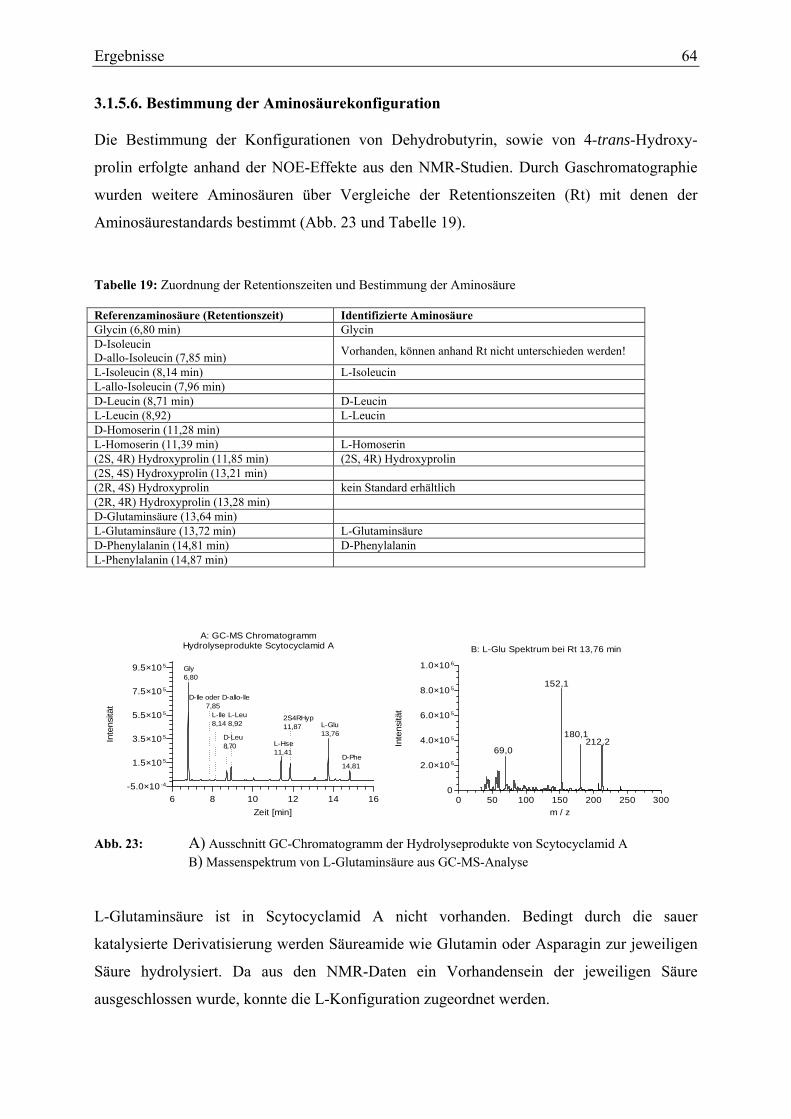
Ergebnisse 64
3.1.5.6. Bestimmung der Aminosäurekonfiguration Die Bestimmung der Konfigurationen von Dehydrobutyrin, sowie von 4-trans-Hydroxy-
prolin erfolgte anhand der NOE-Effekte aus den NMR-Studien. Durch Gaschromatographie
wurden weitere Aminosäuren über Vergleiche der Retentionszeiten (Rt) mit denen der
Aminosäurestandards bestimmt (Abb. 23 und Tabelle 19).
Tabelle 19: Zuordnung der Retentionszeiten und Bestimmung der Aminosäure Referenzaminosäure (Retentionszeit) Identifizierte Aminosäure Glycin (6,80 min) Glycin D-Isoleucin D-allo-Isoleucin (7,85 min) Vorhanden, können anhand Rt nicht unterschieden werden!
L-Isoleucin (8,14 min) L-Isoleucin L-allo-Isoleucin (7,96 min) D-Leucin (8,71 min) D-Leucin L-Leucin (8,92) L-Leucin D-Homoserin (11,28 min) L-Homoserin (11,39 min) L-Homoserin (2S, 4R) Hydroxyprolin (11,85 min) (2S, 4R) Hydroxyprolin (2S, 4S) Hydroxyprolin (13,21 min) (2R, 4S) Hydroxyprolin kein Standard erhältlich (2R, 4R) Hydroxyprolin (13,28 min) D-Glutaminsäure (13,64 min) L-Glutaminsäure (13,72 min) L-Glutaminsäure D-Phenylalanin (14,81 min) D-Phenylalanin L-Phenylalanin (14,87 min)
Abb. 23: A) Ausschnitt GC-Chromatogramm der Hydrolyseprodukte von Scytocyclamid A B) Massenspektrum von L-Glutaminsäure aus GC-MS-Analyse L-Glutaminsäure ist in Scytocyclamid A nicht vorhanden. Bedingt durch die sauer
katalysierte Derivatisierung werden Säureamide wie Glutamin oder Asparagin zur jeweiligen
Säure hydrolysiert. Da aus den NMR-Daten ein Vorhandensein der jeweiligen Säure
ausgeschlossen wurde, konnte die L-Konfiguration zugeordnet werden.
A: GC-MS ChromatogrammHydrolyseprodukte Scytocyclamid A
6 8 10 12 14 16-5.0×10 -4
1.5×10 5
3.5×10 5
5.5×10 5
7.5×10 5
9.5×10 5 Gly6,80
D-Ile oder D-allo-Ile7,85
L-Ile8,14
D-Leu8,70
L-Leu8,92
L-Hse11,41
2S4RHyp11,87 L-Glu
13,76
D-Phe14,81
Zeit [min]
Inte
nsitä
t
B: L-Glu Spektrum bei Rt 13,76 min
0 50 100 150 200 250 3000
2.0×10 5
4.0×10 5
6.0×10 5
8.0×10 5
1.0×10 6
69,0
152,1
180,1212,2
m / z
Inte
nsitä
t

Ergebnisse 65
D-Isoleucin und D-allo-Isoleucin wiesen die gleiche Retentionszeit auf, weshalb eine
Zuordnung nicht getroffen wurde. Um dieses Problem zu lösen, wurde versucht die
Konfiguration dieser beiden Aminosäuren über eine Aminosäurebestimmung nach Marfey zu
lösen (Marfey et al., 1984). Die Proben wurden dazu nach Derivatisierung mittels HPLC-
Messungen untersucht. Dies führte nicht zum Erfolg, so dass eine Zuordnung nicht getroffen
werden konnte. Aminooctansäure konnte mangels Referenzsubstanz nicht aufgeklärt werden.
Ebenso wenig war ein Standard für (2R, 4R) Hydroxyprolin zu bekommen, jedoch konnte die
Konfiguration über die NOE-Daten bestimmt werden (Abb. 22). Die restlichen Aminosäuren
wurden jeweils passenden Referenzsubstanzen zugeschrieben. Die genaue Position der
Diastereomeren Aminosäuren D- und L-Leucin, sowie der Isoleucine im Ringgerüst konnte
nur anhand der Vergleiche mit den Strukturanaloga aus Hormothamnion sp. und Lyngbya sp.
postuliert werden.
Abb. 24: Struktur von Scytocyclamid A
3.1.6. Strukturaufklärung von Scytocyclamid B 3.1.6.1. Ultraviolett-Spektroskopie Während der HPLC-Auftrennung zeigte Scytocyclamide B, ebenso wie Scytocyclamid A, ein
für die bisher aus Scytonema hofmanni isolierten Peptide untypisches UV-Spektrum. Im
Vergleich zu Scytocyclamid A nahm die Absorption im Bereich zwischen 200 und 250 nm
stärker ab und die Absorption hatte einen kurvenartigen Verlauf, während die Abnahme der
NH
ONH
NH
NH
O
OO
O
NOH
O
O
NH2
NH
ONH
ONH
ONH O
NHNH
O
OH
Aoc1
Gln2
Dhb3
Hyp4
Hse5Phe6
Leu7
Ile8
Ile9
Leu10 Gly11

Ergebnisse 66
Absorption bei Scytocyclamid A in diesem Bereich eher linear abnahm. Eine für aromatische
Aminosäuren typische Absorption um 280 nm wurde nicht beobachtet.
Abb. 25: UV-Spektrum von Scytocyclamid B.
3.1.6.2. Infrarotspektroskopie Nähere Informationen über Funktionalitäten von Scytocyclamid B wurden durch
Infrarotspektroskopie (4.000-680 cm-1) aufgezeichnet. Rückstände von Trifluoressigsäure
konnten zugeordnet werden. Aus den Zuordnungen der Wellenzahlen (Tabelle 20) ließ sich
auf das Vorhandensein von Amidbindungen schließen. Weitere charakteristische
Absorptionsbanden im hohen Wellenzahlbereich werden durch OH- und NH-
Valenzschwingungen verursacht und sind zusammen mit den C=O-Valenzschwingungen
typisch für Naturstoffe mit Peptid-Charakter.
Abb. 26: IR-Spektrum von Scytocyclamid B
Tabelle 20: Zuordnung der Wellenzahlen Wellenzahl Interpretation Wellenzahl Interpretation
3299,50 (O-H / N-H) Valenzschwingungen 1385,52 symmetrische Deformationschwingung
2966,70 (C-H) Valenzschwingungen 1293,73
Scytocyclamid B
50010001500200025003000350020
30
40
50
60
70
80
90
100
3299,502966,702936,322877,89
1780,39
1634,47
1522,42
1451,11
1385,52
1293,73
1202,06
1154,45
1059,42
838,57
699,56
721,61799,97
cm-1
% T
rans
mis
sion

Ergebnisse 67
2936,32 (C-H) Valenzschwingungen 1202,06
2877,89 1154,45 (C-O) Valenzschwingung
1780,39 1059,42
1634,47 Amid I (C=O-Valenzschwingung) 838,57
1522,42 Amid II (C=O-Valenzschwingung) 721,51 CH2-rocking-Schwingung
1451,11 (C-H) Deformationsschwingung 699,56
3.1.6.3. Massenspektrometrie Weitere Hinweise auf die Struktur von Scytocyclamid B wurden per MALDI-TOF-Analysen
gewonnen. Hierzu wurde die Probe im positive ion mode analysiert. Es wurden drei Signale
detektiert. Der base peak wurde bei m/z 1.367,79 [M+H] detektiert. Daneben wurde ein
Signal bei 1.389,84 [M+Na] beobachtet. Bei einer weiteren unabhängigen Proben-
Aufbereitung wurde zusätzlich die Masse von 1.405,70 [M+K] detektiert.
Ausgehend von einem Molekulargewicht von 1.367 Da für das monoisotopische Signal
[M+H] wurde eine Struktur mit 12 oder 13 Aminosäuren erwartet. Zur Bestätigung des
Peptidcharakters wurden post source decay (PSD)-Spektren aufgenommen. Die PSD-
Spektren hatten keine Ähnlichkeit zu bereits isolierten Peptiden aus Scytonema hofmanni.
Jedoch wurden Fragmentmassen mit für Aminosäuren typischen Massenzahlen detektiert, so
dass man von einer Peptidstruktur ausgehen konnte. Aus dem PSD-Spektrum konnten Leucin,
bzw. Isoleucin, ein methyliertes Leucin, bzw. methyliertes Isoleucin, Alanin, Valin, Threonin
und Prolin als Bausteine über die Fragmentmasse ihrer Immoniumionen abgeleitet werden.
Weitere Fragmente konntenzuerst nicht zugeordnet werden. Da zu dem
Untersuchungszeitpunkt die Struktur von Scytocyclamid A bereits bekannt war, wurde nach
Strukturanalogien gesucht. In die Suche wurden ebenfalls die verwandten Laxaphycine,
Hormothamnine und Lobocyclamide einbezogen. Bei den Laxaphycinen wurde eine Struktur
mit dem Nominalgewicht von 1.366 Da beschrieben (Frankmölle et al., 1992). Deshalb lag
das Augenmerk auf Sequenzvergleichen bei denen Peptidfragmente analog zu Laxaphycin D
konstruiert und mit den Fragmentmassen aus den PSD-Spektren verglichen wurden. Hieraus
ergaben sich zahlreiche Hinweise auf eine analoge Struktur. Die so getroffenen
Fragmentzuordnungen sind in Tabelle 21 angegeben. Somit konnte ein Strukturvorschlag für
die NMR-Analysen vorgelegt werden (Abb. 27). Zur Bestätigung der Strukturanalogie
wurden umfangreiche NMR-Analysen vorgenommen.
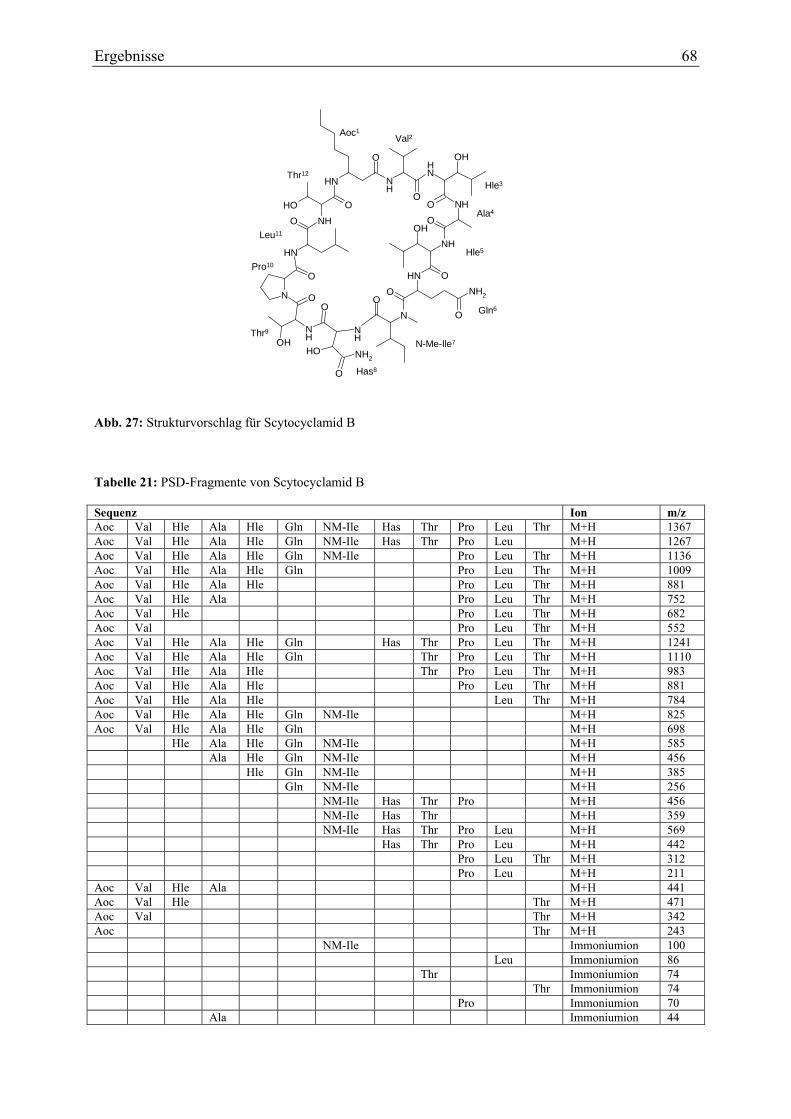
Ergebnisse 68
Abb. 27: Strukturvorschlag für Scytocyclamid B
Tabelle 21: PSD-Fragmente von Scytocyclamid B Sequenz Ion m/z Aoc Val Hle Ala Hle Gln NM-Ile Has Thr Pro Leu Thr M+H 1367 Aoc Val Hle Ala Hle Gln NM-Ile Has Thr Pro Leu M+H 1267 Aoc Val Hle Ala Hle Gln NM-Ile Pro Leu Thr M+H 1136 Aoc Val Hle Ala Hle Gln Pro Leu Thr M+H 1009 Aoc Val Hle Ala Hle Pro Leu Thr M+H 881 Aoc Val Hle Ala Pro Leu Thr M+H 752 Aoc Val Hle Pro Leu Thr M+H 682 Aoc Val Pro Leu Thr M+H 552 Aoc Val Hle Ala Hle Gln Has Thr Pro Leu Thr M+H 1241 Aoc Val Hle Ala Hle Gln Thr Pro Leu Thr M+H 1110 Aoc Val Hle Ala Hle Thr Pro Leu Thr M+H 983 Aoc Val Hle Ala Hle Pro Leu Thr M+H 881 Aoc Val Hle Ala Hle Leu Thr M+H 784 Aoc Val Hle Ala Hle Gln NM-Ile M+H 825 Aoc Val Hle Ala Hle Gln M+H 698 Hle Ala Hle Gln NM-Ile M+H 585 Ala Hle Gln NM-Ile M+H 456 Hle Gln NM-Ile M+H 385 Gln NM-Ile M+H 256 NM-Ile Has Thr Pro M+H 456 NM-Ile Has Thr M+H 359 NM-Ile Has Thr Pro Leu M+H 569 Has Thr Pro Leu M+H 442 Pro Leu Thr M+H 312 Pro Leu M+H 211 Aoc Val Hle Ala M+H 441 Aoc Val Hle Thr M+H 471 Aoc Val Thr M+H 342 Aoc Thr M+H 243 NM-Ile Immoniumion 100 Leu Immoniumion 86 Thr Immoniumion 74 Thr Immoniumion 74 Pro Immoniumion 70 Ala Immoniumion 44
N
O
NHOH O
NH NH
O
O
NH
OH
O NH
O
NHOH
NH ONH2
O
O
N
O
NH
OO
NH
OH
O
NH2
OH
O
NH
Aoc1Val2
Hle3
Ala4
Hle5
Gln6
N-Me-Ile7
Has8
Thr9
Pro10
Leu11
Thr12

Ergebnisse 69
3.1.6.4. Magnetische Kernresonanzspektroskopie Durch 1H-NMR-Analysen, sowie zweidimensionale 1H-NMR-Spektroskopie wurde der
Strukturvorschlag auf seine Richtigkeit überprüft. Erschwerend kam hinzu, dass
Scytocyclamid B als Konformerengemisch vorlag, was zu Duplizität einzelner Signale führte,
so dass nicht für alle chemischen Verschiebungen Zuordnungen zu einzelnen
Strukturelementen gemacht werden konnten (Tabelle 22). Über H, H-COSY werden skalar
gekoppelte Protonenpaare detektiert. Kopplungen über mehr als 3 Bindungen fallen schwach
aus, so dass ausschließlich vicinal und geminal gekoppelte Protonen erfasst wurden. Alle
miteinander koppelnden Protonen bilden ein Spinsystem. Die Identifizierung einzelner
Aminosäuren erfolgte über die Zuordnung ihrer charakteristischen Spinsysteme. Die Sequenz
der Aminosäuren wurde über H, H-ROESY-Experimente verifiziert. Hierbei wurden
Sequenz-NOE-Korrelationen vom Typ α-N, i , i+1 ausgewertet. Informationen über die
chemische Umgebung des Kohlenstoffatoms lieferten H, C-HSQC-Messungen, welche
direkte C-H-Korrelationen erfassen. Die chemische Verschiebung der Kohlenstoffatome
wurde aus den HSQC-Messungen bestimmt.
Die in der Massenspektrometrie nicht zu unterscheidenden Aminsäuren Leucin und Isoleucin,
sowie deren Strukturmodifikationen wurden durch die Aufspaltungsmuster der einzelnen
Signale, sowie zugehörigen Kopplungskonstanten bestimmt. Hydroxyasparagin und Glutamin
konnte durch Zuordnung von Signalen der Amidgruppen in den Seitenketten eindeutig von
ihren jeweiligen Säuren unterschieden werden. Ein Vergleich mit den publizierten Daten zu
Laxaphycin D (Frankmölle et al., 1992) zeigt keine Unregelmäßigkeiten oder Abweichungen.
Alle gewonnen Daten stützen den Strukturvorschlag aus der Massenspektrometrie, so dass der
Strukturvorschlag bestätigt werden kann (siehe hierzu Abb. 27).
Abb. 27: NMR-Spektrum Scytocyclamid B
8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0 0.5Chemical Shif t (ppm)
51.6622.9910.8613.302.412.947.222.6316.249.001.021.655.029.051.00
0.99
(12-
Thr4
)1.
04(9
-Thr
4)
1.31
(4-A
la3)
2.32
(1-O
ct3)
2.40
(1-O
ct3)
2.96
(7-M
e-Ile
1)
3.47
(5-H
le3,
3-H
le3)
3.65
(10-
Pro5
)3.
67(1
0-Pr
o5)
3.90
(9-T
hr3)
3.98
(12-
Thr3
)4.
08(2
-Val
2)4.
11(1
-Oct
2,12
-Thr
2)4.
20(4
-Ala
2)4.
30(5
-Hle
2)4.
34(8
-Has
3,10
-Pro
2,11
-Leu
2,3-
H4.
47(9
-Thr
2)4.
61(8
-Has
2,6-
Gln
2)4.
73(7
-Me-
Ile2)
4.82
(12-
Thr5
)4.
97(3
-Hle
7)4.
98(9
-Thr
5)5.
10(5
-Hle
7)
5.82
(8-H
as6)
6.88
(6-G
ln6)
7.24
(6-G
ln6)
7.30
(8-H
as5)
7.32
(9-T
hr1)
7.59
(1-O
ct1)
7.70
(8-H
as1,
5-H
le1)
7.77
(6-G
ln1)
7.80
(12-
Thr1
)7.
87(1
1-Le
u1)
7.88
(4-A
la1)
7.99
(3-H
le1)
8.21
(2-V
al1)
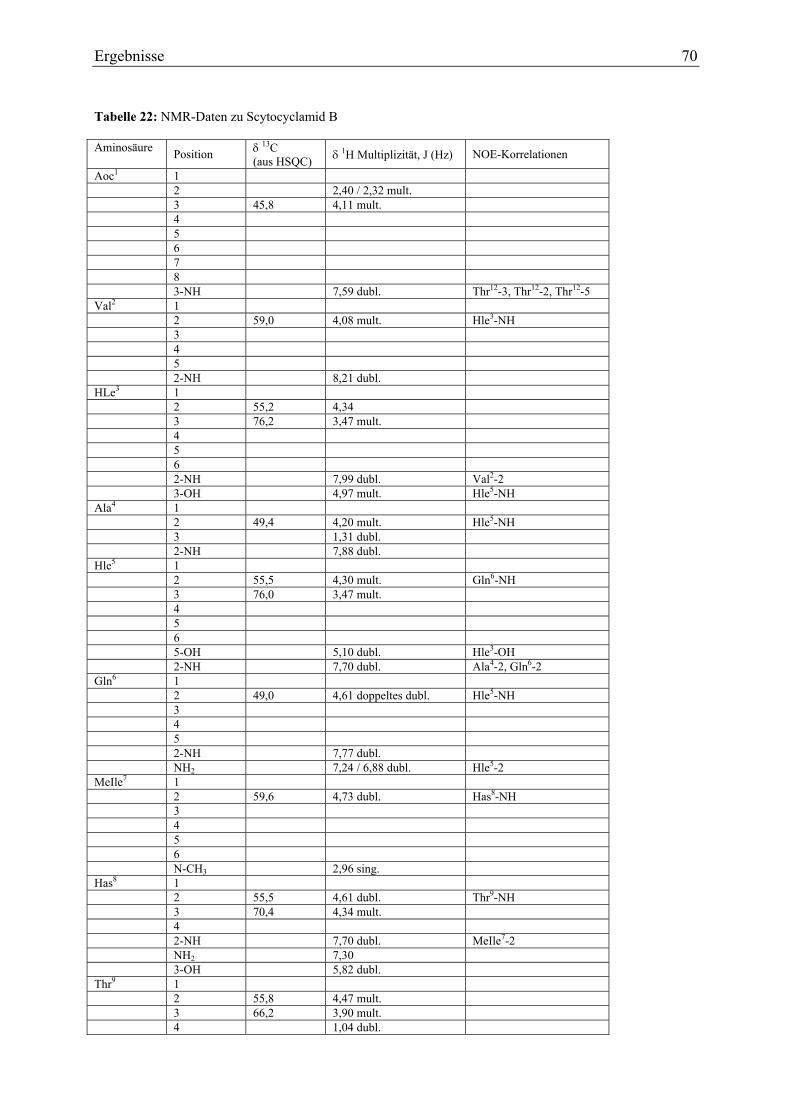
Ergebnisse 70
Tabelle 22: NMR-Daten zu Scytocyclamid B Aminosäure Position δ 13C
(aus HSQC) δ 1H Multiplizität, J (Hz) NOE-Korrelationen
Aoc1 1 2 2,40 / 2,32 mult. 3 45,8 4,11 mult. 4 5 6 7 8 3-NH 7,59 dubl. Thr12-3, Thr12-2, Thr12-5 Val2 1 2 59,0 4,08 mult. Hle3-NH 3 4 5 2-NH 8,21 dubl. HLe3 1 2 55,2 4,34 3 76,2 3,47 mult. 4 5 6 2-NH 7,99 dubl. Val2-2 3-OH 4,97 mult. Hle5-NH Ala4 1 2 49,4 4,20 mult. Hle5-NH 3 1,31 dubl. 2-NH 7,88 dubl. Hle5 1 2 55,5 4,30 mult. Gln6-NH 3 76,0 3,47 mult. 4 5 6 5-OH 5,10 dubl. Hle3-OH 2-NH 7,70 dubl. Ala4-2, Gln6-2 Gln6 1 2 49,0 4,61 doppeltes dubl. Hle5-NH 3 4 5 2-NH 7,77 dubl. NH2 7,24 / 6,88 dubl. Hle5-2 MeIle7 1 2 59,6 4,73 dubl. Has8-NH 3 4 5 6 N-CH3 2,96 sing. Has8 1 2 55,5 4,61 dubl. Thr9-NH 3 70,4 4,34 mult. 4 2-NH 7,70 dubl. MeIle7-2 NH2 7,30 3-OH 5,82 dubl. Thr9 1 2 55,8 4,47 mult. 3 66,2 3,90 mult. 4 1,04 dubl.

Ergebnisse 71
2-NH 7,32 dubl. Has8-2 3-OH 4,98 mult. Pro10 1 2 59,5 4,34 mult. 3 4 5 47,0 3,67 / 3,65 dubl. Leu11 1 2 51,0 4,34 mult. Thr12-NH 3 4 5 6 2-NH 7,87 mult. Thr12 1 2 57,8 4,11 mult. Aoc1-NH 3 66,2 3,98 mult. Aoc1-NH 4 0,99 2-NH 7,80 dubl. Leu11-2 3-OH 4,82 dubl. Aoc1-NH Korrelationen innerhalb des Spinsystems einer Aminosäure werden nicht aufgeführt.
3.1.7. Struktur von Scytocyclamid C Erste Untersuchungen zu Scytocyclamid C ergaben große Ähnlichkeit zu Scytocyclamid B.
Dies galt für das Verhalten während der chromatographischen Aufreinigung. Beispielsweise
eluierten Scytocyclamid B und C bei ähnlichen Retentionszeiten (26 min / 28 min). Die UV-
Spektren waren identisch (ohne Abb.).
3.1.7.1. Infrarotspektroskopie Die Infrarotspektroskopie-Aufnahmen wurden unter den bereits beschriebenen Bedingungen
durchgeführt. Wie aus dem Spektrum ersichtlich ergaben sich zu Scytocyclamid B keine
nennenswerten Unterschiede, weshalb auf eine Zuordnung der Wellenzahlen an dieser Stelle
verzichtet wird.
Abb.28: Infrarotspektrum von Scytocyclamid C
Scytocyclamid C
65011501650215026503150365075
85
95
3284,962962,29
1641,86
1520,70
1450,72
1201,501137,36
838,23800,45
721,08
cm-1
% T
rans
mis
sion
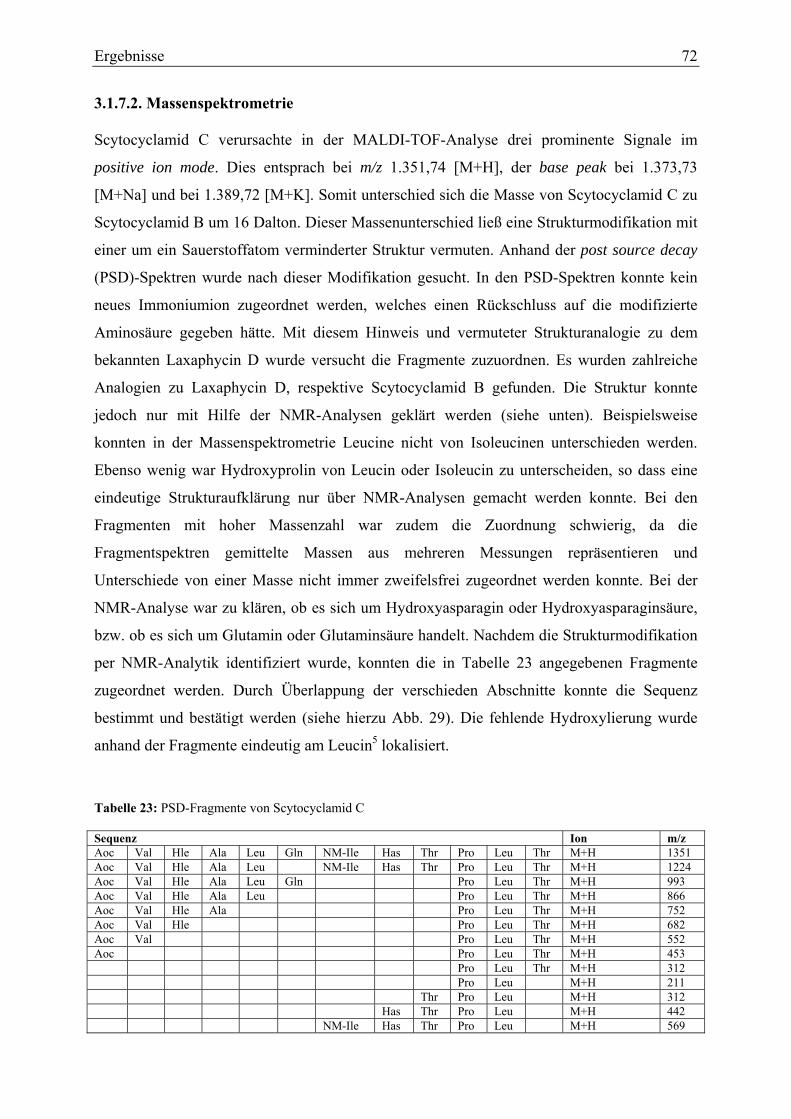
Ergebnisse 72
3.1.7.2. Massenspektrometrie Scytocyclamid C verursachte in der MALDI-TOF-Analyse drei prominente Signale im
positive ion mode. Dies entsprach bei m/z 1.351,74 [M+H], der base peak bei 1.373,73
[M+Na] und bei 1.389,72 [M+K]. Somit unterschied sich die Masse von Scytocyclamid C zu
Scytocyclamid B um 16 Dalton. Dieser Massenunterschied ließ eine Strukturmodifikation mit
einer um ein Sauerstoffatom verminderter Struktur vermuten. Anhand der post source decay
(PSD)-Spektren wurde nach dieser Modifikation gesucht. In den PSD-Spektren konnte kein
neues Immoniumion zugeordnet werden, welches einen Rückschluss auf die modifizierte
Aminosäure gegeben hätte. Mit diesem Hinweis und vermuteter Strukturanalogie zu dem
bekannten Laxaphycin D wurde versucht die Fragmente zuzuordnen. Es wurden zahlreiche
Analogien zu Laxaphycin D, respektive Scytocyclamid B gefunden. Die Struktur konnte
jedoch nur mit Hilfe der NMR-Analysen geklärt werden (siehe unten). Beispielsweise
konnten in der Massenspektrometrie Leucine nicht von Isoleucinen unterschieden werden.
Ebenso wenig war Hydroxyprolin von Leucin oder Isoleucin zu unterscheiden, so dass eine
eindeutige Strukturaufklärung nur über NMR-Analysen gemacht werden konnte. Bei den
Fragmenten mit hoher Massenzahl war zudem die Zuordnung schwierig, da die
Fragmentspektren gemittelte Massen aus mehreren Messungen repräsentieren und
Unterschiede von einer Masse nicht immer zweifelsfrei zugeordnet werden konnte. Bei der
NMR-Analyse war zu klären, ob es sich um Hydroxyasparagin oder Hydroxyasparaginsäure,
bzw. ob es sich um Glutamin oder Glutaminsäure handelt. Nachdem die Strukturmodifikation
per NMR-Analytik identifiziert wurde, konnten die in Tabelle 23 angegebenen Fragmente
zugeordnet werden. Durch Überlappung der verschieden Abschnitte konnte die Sequenz
bestimmt und bestätigt werden (siehe hierzu Abb. 29). Die fehlende Hydroxylierung wurde
anhand der Fragmente eindeutig am Leucin5 lokalisiert.
Tabelle 23: PSD-Fragmente von Scytocyclamid C Sequenz Ion m/z Aoc Val Hle Ala Leu Gln NM-Ile Has Thr Pro Leu Thr M+H 1351 Aoc Val Hle Ala Leu NM-Ile Has Thr Pro Leu Thr M+H 1224 Aoc Val Hle Ala Leu Gln Pro Leu Thr M+H 993 Aoc Val Hle Ala Leu Pro Leu Thr M+H 866 Aoc Val Hle Ala Pro Leu Thr M+H 752 Aoc Val Hle Pro Leu Thr M+H 682 Aoc Val Pro Leu Thr M+H 552 Aoc Pro Leu Thr M+H 453 Pro Leu Thr M+H 312 Pro Leu M+H 211 Thr Pro Leu M+H 312 Has Thr Pro Leu M+H 442 NM-Ile Has Thr Pro Leu M+H 569

Ergebnisse 73
NM-Ile Has Thr Pro M+H 456 NM-Ile Has Thr M+H 359 Gln NM-Ile M+H 256 Leu Gln NM-Ile M+H 369 Ala Leu Gln NM-Ile M+H 440 Hle Ala Leu Gln NM-Ile M+H 569 Hle Ala Leu Gln M+H 442 Hle Ala Leu Gln NM-Ile Has Thr M+H 800 Aoc Val Hle Ala Leu Gln NM-Ile Has Thr M+H 1040 Aoc Val Hle Ala Leu Gln NM-Ile Thr M+H 910 Aoc Val Hle Ala Leu Gln Thr M+H 783 Aoc Val Hle Ala Leu Gln M+H 682 Aoc Val Thr M+H 342 Aoc Thr M+H 243 NM-Ile Immoniumion 100 Leu Immoniumion 86 Leu Immoniumion 86 Thr Immoniumion 74 Thr Immoniumion 74 Val Immoniumion 72 Pro Immoniumion 70 Ala Immoniumion 44 3.1.7.3. NMR-Analytik zu Scytocyclamid C Das 1H-NMR-Spektrum zeigte die vermutete Analogie zu Scytocyclamid B (Abb. 29). Die
Zuordnung der chemischen Verschiebungen zu bestimmten Strukturelementen erfolgte nach
Auswertung der Korrelationsmessungen (TOCSY-, ROESY-, COSY-, HSQC-Messungen).
Die genauen Zuordnungen, sowie die Unterschiede zu Scytocyclamid B sind Tabelle 24 zu
entnehmen. Ebenso wie Scytocyclamid B lag Scytocyclamid C als Konformerengemisch vor,
so dass nicht alle Signale eindeutig zugeordnet werden konnten. Das Vorhandensein von
Hydroxyasparaginsäure und Glutaminsäure konnte durch Zuordnung von Signalen „freier“
Amidprotonen ausgeschlossen werden. Leucine, Isoleucine oder deren Analoga und
Hydroxyprolin konnten anhand ihrer typischen Spinmuster unterschieden, bzw.
ausgeschlossen werden.
Abb. 28: NMR-Spektrum Scytocyclamid C
8.5 8.0 7.5 7.0 6.5 6.0 5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5 1.0Chemical Shif t (ppm)
0.99
(12-
Thr4
)1.
05(9
-Thr
4)
1.25
(4-A
la3)
2.31
(1-O
ct3)
2.42
(1-O
ct3)
2.94
(7-M
e-Ile
1)
3.50
(3-H
le3)
3.67
(10-
Pro5
)3.
93(9
-Thr
3)3.
98(1
2-Th
r3)
4.02
(2-V
al2)
4.09
(1-O
ct2)
4.11
(12-
Thr2
)4.
17(5
-Leu
2)4.
18(4
-Ala
2)4.
32(1
1-Le
u2)
4.34
(3-H
le2)
4.35
(8-H
as3,
10-P
4.48
(9-T
hr2)
4.59
(6-G
ln2)
4.63
(8-H
as2)
4.70
(7-M
e-Ile
2)
6.82
(6-G
ln6)
7.25
(6-G
ln6)
7.33
(8-H
as5)
7.35
(9-T
hr1)
7.60
(8-H
as1)
7.65
(1-O
ct1)
7.72
(12-
Thr1
)7.
81(5
-Leu
1)7.
84(4
-Ala
1)7.
91(6
-Gln
1)7.
94(1
1-Le
u1)
8.02
(3-H
le1)
8.21
(2-V
al1)
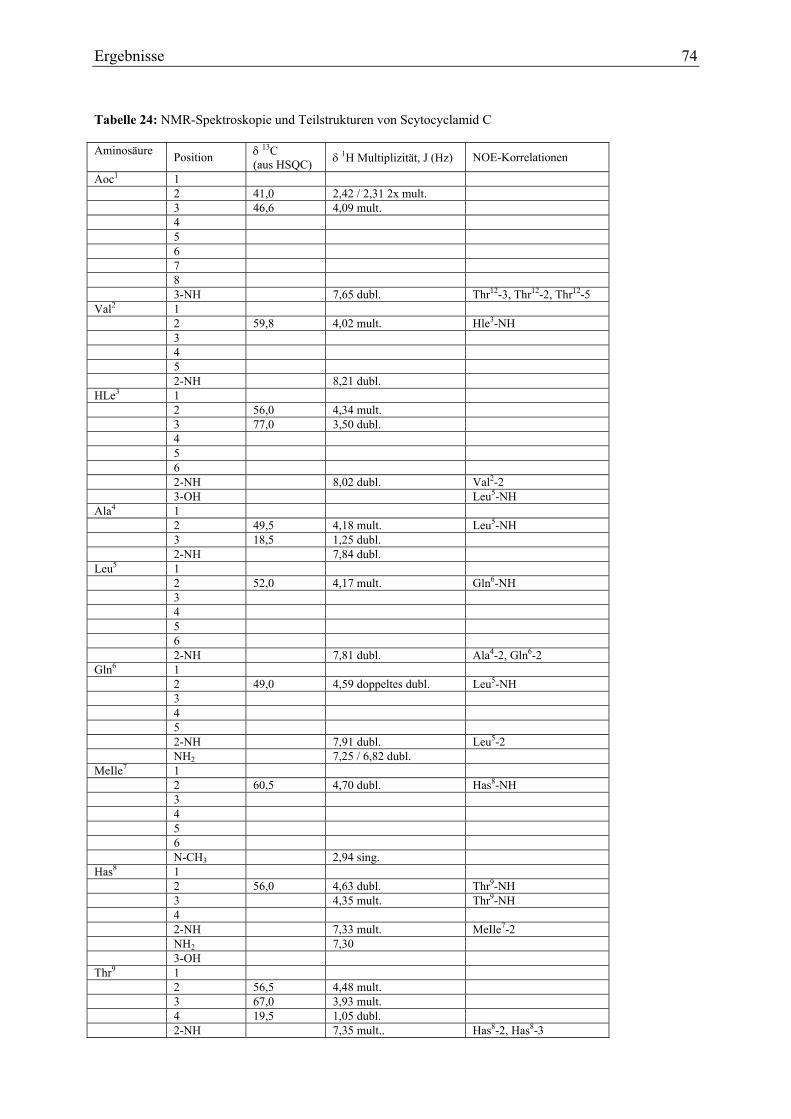
Ergebnisse 74
Tabelle 24: NMR-Spektroskopie und Teilstrukturen von Scytocyclamid C Aminosäure Position δ 13C
(aus HSQC) δ 1H Multiplizität, J (Hz) NOE-Korrelationen
Aoc1 1 2 41,0 2,42 / 2,31 2x mult. 3 46,6 4,09 mult. 4 5 6 7 8 3-NH 7,65 dubl. Thr12-3, Thr12-2, Thr12-5 Val2 1 2 59,8 4,02 mult. Hle3-NH 3 4 5 2-NH 8,21 dubl. HLe3 1 2 56,0 4,34 mult. 3 77,0 3,50 dubl. 4 5 6 2-NH 8,02 dubl. Val2-2 3-OH Leu5-NH Ala4 1 2 49,5 4,18 mult. Leu5-NH 3 18,5 1,25 dubl. 2-NH 7,84 dubl. Leu5 1 2 52,0 4,17 mult. Gln6-NH 3 4 5 6 2-NH 7,81 dubl. Ala4-2, Gln6-2 Gln6 1 2 49,0 4,59 doppeltes dubl. Leu5-NH 3 4 5 2-NH 7,91 dubl. Leu5-2 NH2 7,25 / 6,82 dubl. MeIle7 1 2 60,5 4,70 dubl. Has8-NH 3 4 5 6 N-CH3 2,94 sing. Has8 1 2 56,0 4,63 dubl. Thr9-NH 3 4,35 mult. Thr9-NH 4 2-NH 7,33 mult. MeIle7-2 NH2 7,30 3-OH Thr9 1 2 56,5 4,48 mult. 3 67,0 3,93 mult. 4 19,5 1,05 dubl. 2-NH 7,35 mult.. Has8-2, Has8-3

Ergebnisse 75
3-OH Pro10 1 2 4,35 mult. 3 4 5 48,0 3,67 mult.. Leu11 1 2 52,0 4,32 dubl. Thr12-NH 3 4 5 6 2-NH 7,94 dubl. Thr12 1 2 58,5 4,11 mult. Aoc1-NH 3 67,0 4,01 mult. Aoc1-NH 4 20,2 0,99 dubl. 2-NH 7,72 dubl. Leu11-2 3-OH Aoc1-NH Korrelationen innerhalb des Spinsystems einer Aminosäure werden nicht aufgeführt.
Aus den so gewonnen Daten ergibt sich folgende Struktur für Scytocyclamid C:
Abb. 29: Struktur von Scytocyclamid C
3.2. Biologische Aktivität
3.2.1. Enzyminhibitions-Assays Für einige Peptide aus dem Sekundärstoffwechselprodukte von Cyanobakterien ist potente
spezifische Hemmung proteolytischer Enzyme bekannt. Um das Wirkungsspektrum der aus
Scytonema hofmanni PCC 7110 isolierten Peptide auf Proteasen zu dokumentieren, wurde die
Hemmung der einzelnen isolierten Peptide auf verschiedene Serinproteasen (Trypsin,
N
O
NHOH O
NH NH
O
O
NH
OH
O NH
O
NH
NH ONH2
O
O
N
O
NH
OO
NH
OH
O
NH2
OH
O
NH
Aoc1Val2
Hle3
Ala4
Gln6
N-Me-Ile7
Has8
Thr9
Pro10
Leu11
Thr12
Leu5
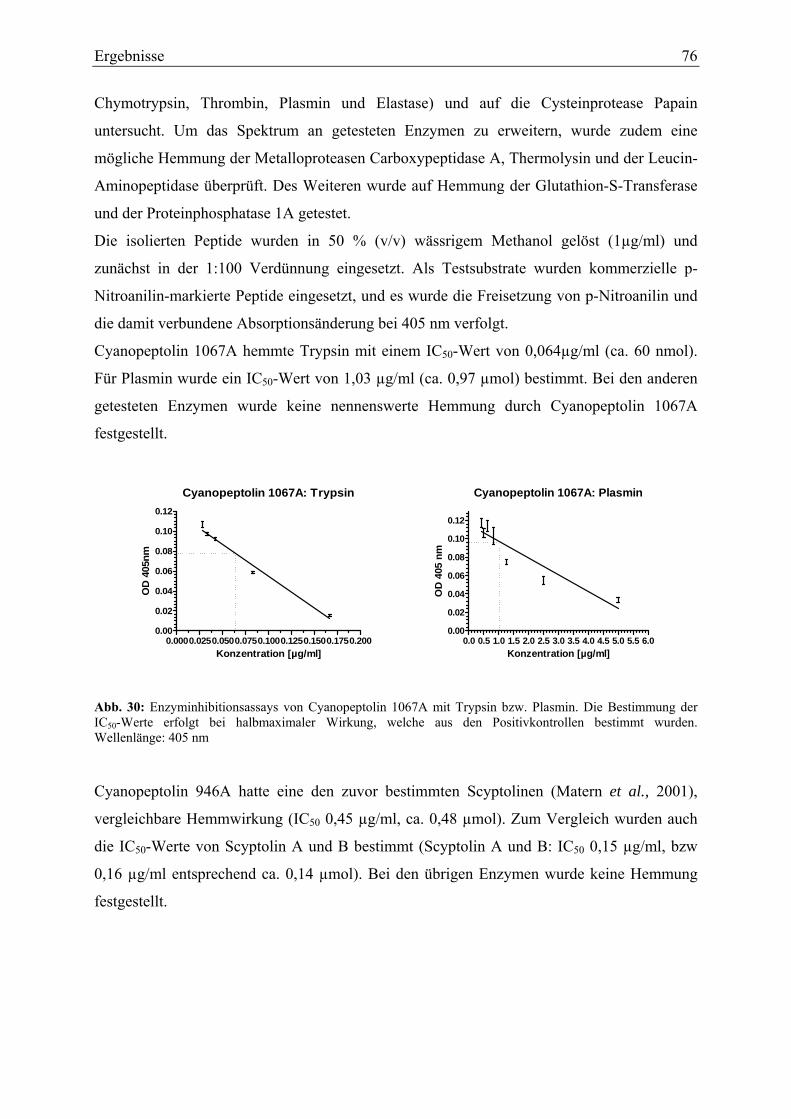
Ergebnisse 76
Chymotrypsin, Thrombin, Plasmin und Elastase) und auf die Cysteinprotease Papain
untersucht. Um das Spektrum an getesteten Enzymen zu erweitern, wurde zudem eine
mögliche Hemmung der Metalloproteasen Carboxypeptidase A, Thermolysin und der Leucin-
Aminopeptidase überprüft. Des Weiteren wurde auf Hemmung der Glutathion-S-Transferase
und der Proteinphosphatase 1A getestet.
Die isolierten Peptide wurden in 50 % (v/v) wässrigem Methanol gelöst (1µg/ml) und
zunächst in der 1:100 Verdünnung eingesetzt. Als Testsubstrate wurden kommerzielle p-
Nitroanilin-markierte Peptide eingesetzt, und es wurde die Freisetzung von p-Nitroanilin und
die damit verbundene Absorptionsänderung bei 405 nm verfolgt.
Cyanopeptolin 1067A hemmte Trypsin mit einem IC50-Wert von 0,064µg/ml (ca. 60 nmol).
Für Plasmin wurde ein IC50-Wert von 1,03 µg/ml (ca. 0,97 µmol) bestimmt. Bei den anderen
getesteten Enzymen wurde keine nennenswerte Hemmung durch Cyanopeptolin 1067A
festgestellt.
Abb. 30: Enzyminhibitionsassays von Cyanopeptolin 1067A mit Trypsin bzw. Plasmin. Die Bestimmung der IC50-Werte erfolgt bei halbmaximaler Wirkung, welche aus den Positivkontrollen bestimmt wurden. Wellenlänge: 405 nm Cyanopeptolin 946A hatte eine den zuvor bestimmten Scyptolinen (Matern et al., 2001),
vergleichbare Hemmwirkung (IC50 0,45 µg/ml, ca. 0,48 µmol). Zum Vergleich wurden auch
die IC50-Werte von Scyptolin A und B bestimmt (Scyptolin A und B: IC50 0,15 µg/ml, bzw
0,16 µg/ml entsprechend ca. 0,14 µmol). Bei den übrigen Enzymen wurde keine Hemmung
festgestellt.
Cyanopeptolin 1067A: Trypsin
0.0000.0250.0500.0750.1000.1250.1500.1750.2000.00
0.02
0.04
0.06
0.08
0.10
0.12
Konzentration [µg/ml]
OD
405
nm
Cyanopeptolin 1067A: Plasmin
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.00.00
0.02
0.04
0.06
0.08
0.10
0.12
Konzentration [µg/ml]
OD
405
nm
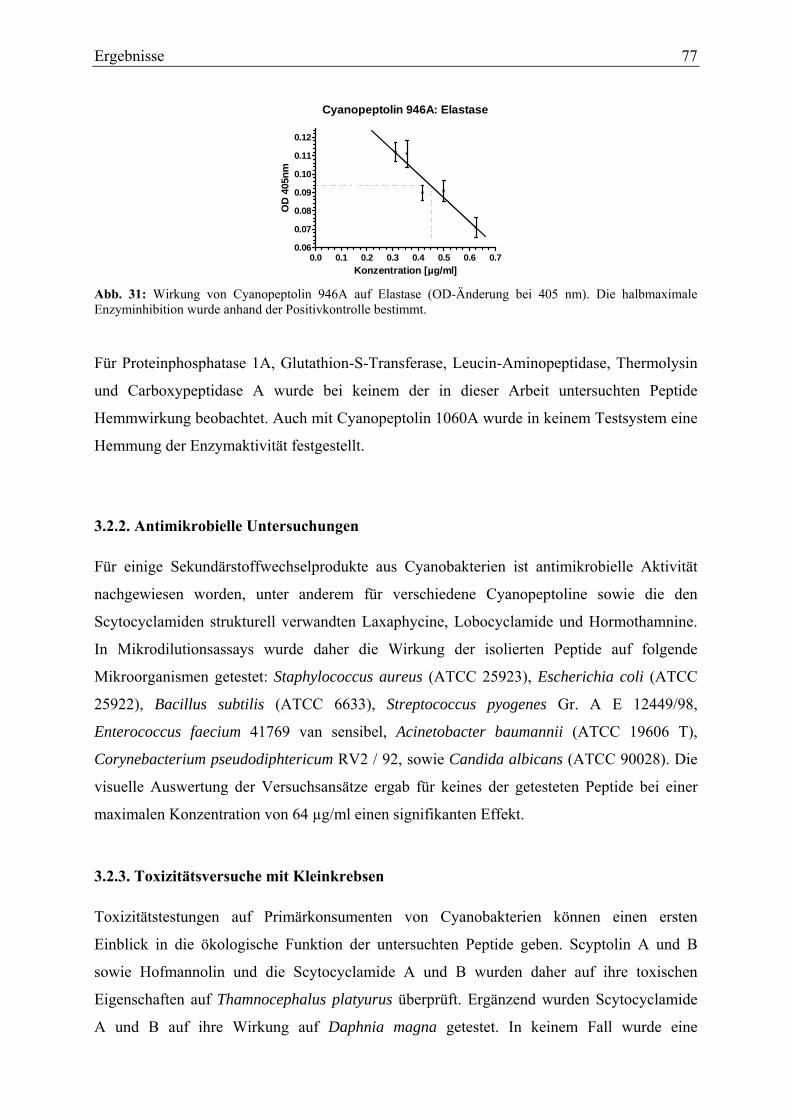
Ergebnisse 77
Abb. 31: Wirkung von Cyanopeptolin 946A auf Elastase (OD-Änderung bei 405 nm). Die halbmaximale Enzyminhibition wurde anhand der Positivkontrolle bestimmt. Für Proteinphosphatase 1A, Glutathion-S-Transferase, Leucin-Aminopeptidase, Thermolysin
und Carboxypeptidase A wurde bei keinem der in dieser Arbeit untersuchten Peptide
Hemmwirkung beobachtet. Auch mit Cyanopeptolin 1060A wurde in keinem Testsystem eine
Hemmung der Enzymaktivität festgestellt.
3.2.2. Antimikrobielle Untersuchungen Für einige Sekundärstoffwechselprodukte aus Cyanobakterien ist antimikrobielle Aktivität
nachgewiesen worden, unter anderem für verschiedene Cyanopeptoline sowie die den
Scytocyclamiden strukturell verwandten Laxaphycine, Lobocyclamide und Hormothamnine.
In Mikrodilutionsassays wurde daher die Wirkung der isolierten Peptide auf folgende
Mikroorganismen getestet: Staphylococcus aureus (ATCC 25923), Escherichia coli (ATCC
25922), Bacillus subtilis (ATCC 6633), Streptococcus pyogenes Gr. A E 12449/98,
Enterococcus faecium 41769 van sensibel, Acinetobacter baumannii (ATCC 19606 T),
Corynebacterium pseudodiphtericum RV2 / 92, sowie Candida albicans (ATCC 90028). Die
visuelle Auswertung der Versuchsansätze ergab für keines der getesteten Peptide bei einer
maximalen Konzentration von 64 µg/ml einen signifikanten Effekt.
3.2.3. Toxizitätsversuche mit Kleinkrebsen Toxizitätstestungen auf Primärkonsumenten von Cyanobakterien können einen ersten
Einblick in die ökologische Funktion der untersuchten Peptide geben. Scyptolin A und B
sowie Hofmannolin und die Scytocyclamide A und B wurden daher auf ihre toxischen
Eigenschaften auf Thamnocephalus platyurus überprüft. Ergänzend wurden Scytocyclamide
A und B auf ihre Wirkung auf Daphnia magna getestet. In keinem Fall wurde eine
Cyanopeptolin 946A: Elastase
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.70.06
0.07
0.08
0.09
0.10
0.11
0.12
Konzentration [µg/ml]O
D 4
05nm
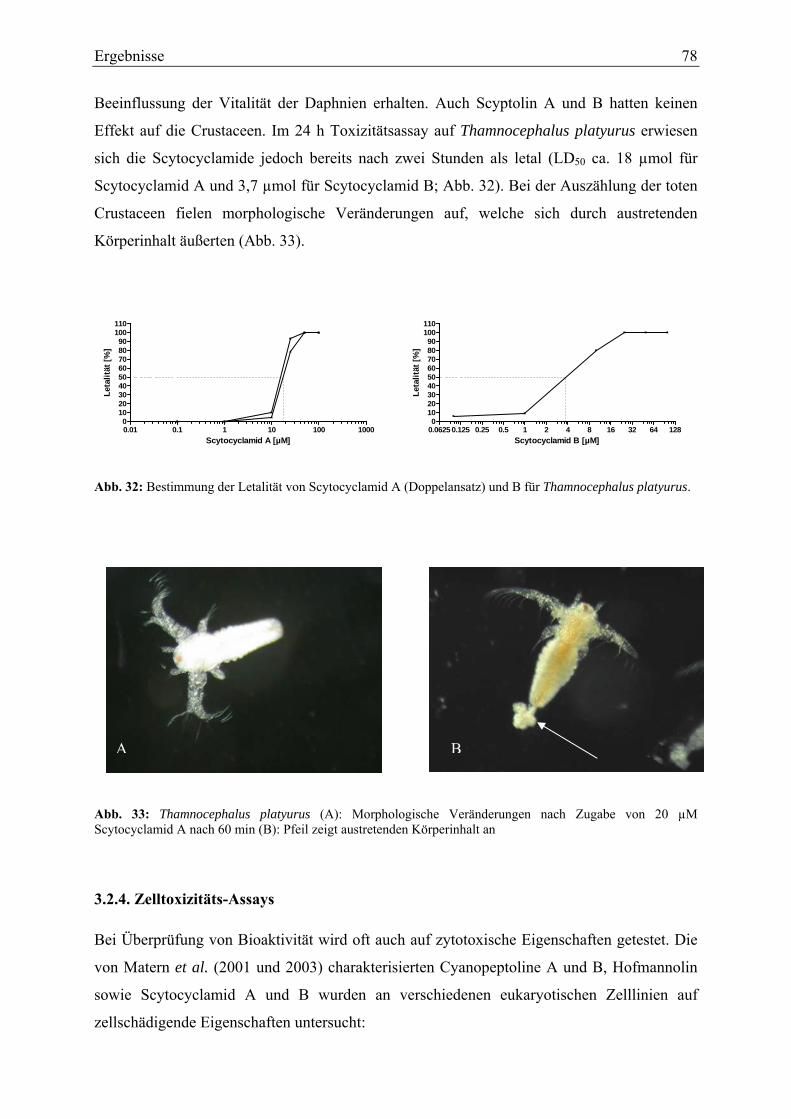
Ergebnisse 78
Beeinflussung der Vitalität der Daphnien erhalten. Auch Scyptolin A und B hatten keinen
Effekt auf die Crustaceen. Im 24 h Toxizitätsassay auf Thamnocephalus platyurus erwiesen
sich die Scytocyclamide jedoch bereits nach zwei Stunden als letal (LD50 ca. 18 µmol für
Scytocyclamid A und 3,7 µmol für Scytocyclamid B; Abb. 32). Bei der Auszählung der toten
Crustaceen fielen morphologische Veränderungen auf, welche sich durch austretenden
Körperinhalt äußerten (Abb. 33).
Abb. 32: Bestimmung der Letalität von Scytocyclamid A (Doppelansatz) und B für Thamnocephalus platyurus. Abb. 33: Thamnocephalus platyurus (A): Morphologische Veränderungen nach Zugabe von 20 µM Scytocyclamid A nach 60 min (B): Pfeil zeigt austretenden Körperinhalt an
3.2.4. Zelltoxizitäts-Assays Bei Überprüfung von Bioaktivität wird oft auch auf zytotoxische Eigenschaften getestet. Die
von Matern et al. (2001 und 2003) charakterisierten Cyanopeptoline A und B, Hofmannolin
sowie Scytocyclamid A und B wurden an verschiedenen eukaryotischen Zelllinien auf
zellschädigende Eigenschaften untersucht:
0.0625 0.125 0.25 0.5 1 2 4 8 16 32 64 1280
102030405060708090
100110
Scytocyclamid B [µM]
Leta
lität
[%]
0.01 0.1 1 10 100 10000
102030405060708090
100110
Scytocyclamid A [µM]
Leta
lität
[%]
A B

Ergebnisse 79
CaCo 2 adhärente epitheliale Zellen aus humanen colorectalem Adenokarzinom HeLa humane Epithelzellen eines Zervixkarzinoms Swiss 3T3 adhärente Zellen aus embryonalen Mäusefibroblasten Ebl embryonale Rinderlungenzellen Rbl basophile Rattenleukämiezellen LoVo Zellen aus humanen colorectalem Adenokarzinom 293 adhärente Fibroblasten aus humaner embryonaler Niere
Die Auswertung erfolgte visuell nach 30 min bzw. 2 h (anfangs nach 24 h) Inkubation mit
dem Lichtmikroskop. Auf eine Begutachtung nach 24 h wurde verzichtet, da alle zu
beobachtenden Veränderungen direkt nach Applikation der Peptide zu sehen waren. Die
Cyanopeptoline hatten keinen Einfluss auf die Viabilität der Zellen. Dagegen hatten
Scytocyclamide starke zellschädigende Eigenschaften: Bei visueller Beobachtung unter dem
Mikroskop ließen sich die Bildung von Vesikeln („blebbing“) und Veränderungen an der
Zellstruktur verfolgen. Zellschädigung wurde bis zu einer Konzentration von 10 µg/ml
beobachtet. In den 1:10 Verdünnungen waren keine eindeutigen Veränderungen zu erkennen.
Abb. 34: Hela-Zellen im Phasenkontrastmikroskop (Phase 2). Die Aufnahmen erfolgten direkt nach Applikation (10 µg/ml) der jeweiligen Scytocyclamide A, B und C (entsprechend dem Buchstabenkürzel) sowie Zusatz von 10 µl einer 0,5 %igen (v/v) Trypanblau-Lösung. Kontrolle: Ansatz mit 1 % (v/v) DMS.
Kontrolle A
B C
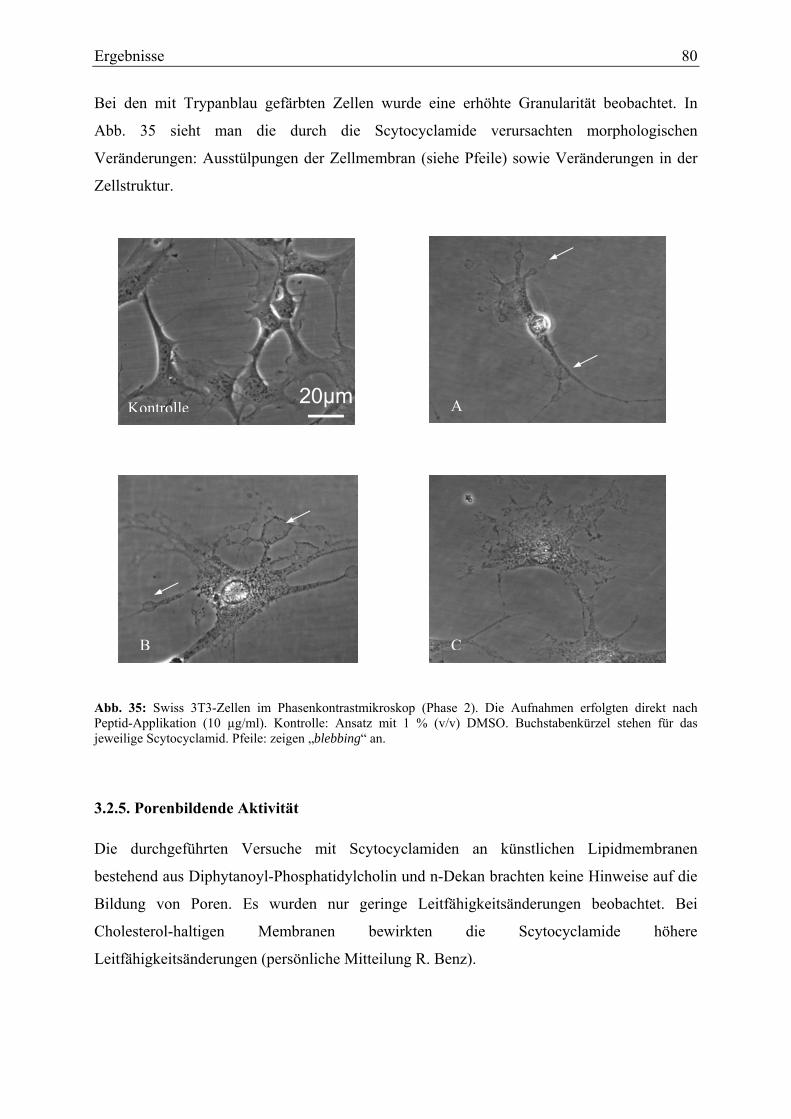
Ergebnisse 80
Bei den mit Trypanblau gefärbten Zellen wurde eine erhöhte Granularität beobachtet. In
Abb. 35 sieht man die durch die Scytocyclamide verursachten morphologischen
Veränderungen: Ausstülpungen der Zellmembran (siehe Pfeile) sowie Veränderungen in der
Zellstruktur.
Abb. 35: Swiss 3T3-Zellen im Phasenkontrastmikroskop (Phase 2). Die Aufnahmen erfolgten direkt nach Peptid-Applikation (10 µg/ml). Kontrolle: Ansatz mit 1 % (v/v) DMSO. Buchstabenkürzel stehen für das jeweilige Scytocyclamid. Pfeile: zeigen „blebbing“ an.
3.2.5. Porenbildende Aktivität Die durchgeführten Versuche mit Scytocyclamiden an künstlichen Lipidmembranen
bestehend aus Diphytanoyl-Phosphatidylcholin und n-Dekan brachten keine Hinweise auf die
Bildung von Poren. Es wurden nur geringe Leitfähigkeitsänderungen beobachtet. Bei
Cholesterol-haltigen Membranen bewirkten die Scytocyclamide höhere
Leitfähigkeitsänderungen (persönliche Mitteilung R. Benz).
A
Kontrolle
B
D
A
C
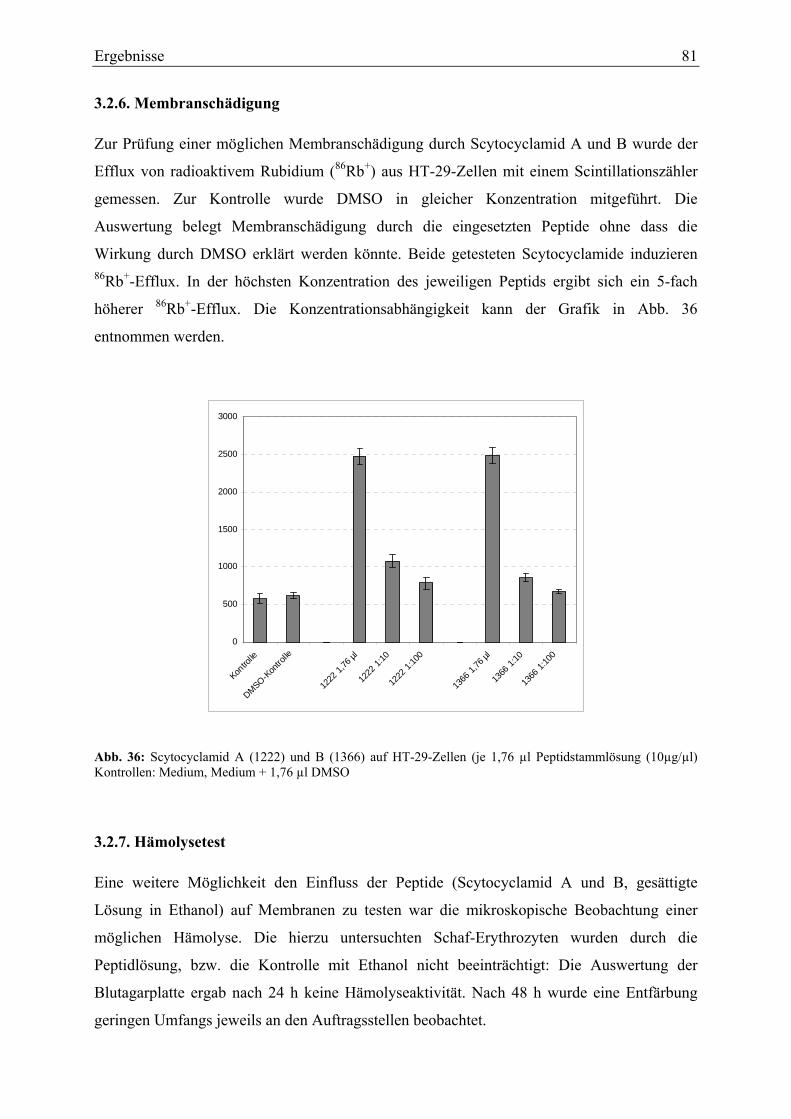
Ergebnisse 81
3.2.6. Membranschädigung Zur Prüfung einer möglichen Membranschädigung durch Scytocyclamid A und B wurde der
Efflux von radioaktivem Rubidium (86Rb+) aus HT-29-Zellen mit einem Scintillationszähler
gemessen. Zur Kontrolle wurde DMSO in gleicher Konzentration mitgeführt. Die
Auswertung belegt Membranschädigung durch die eingesetzten Peptide ohne dass die
Wirkung durch DMSO erklärt werden könnte. Beide getesteten Scytocyclamide induzieren 86Rb+-Efflux. In der höchsten Konzentration des jeweiligen Peptids ergibt sich ein 5-fach
höherer 86Rb+-Efflux. Die Konzentrationsabhängigkeit kann der Grafik in Abb. 36
entnommen werden.
Abb. 36: Scytocyclamid A (1222) und B (1366) auf HT-29-Zellen (je 1,76 µl Peptidstammlösung (10µg/µl) Kontrollen: Medium, Medium + 1,76 µl DMSO
3.2.7. Hämolysetest Eine weitere Möglichkeit den Einfluss der Peptide (Scytocyclamid A und B, gesättigte
Lösung in Ethanol) auf Membranen zu testen war die mikroskopische Beobachtung einer
möglichen Hämolyse. Die hierzu untersuchten Schaf-Erythrozyten wurden durch die
Peptidlösung, bzw. die Kontrolle mit Ethanol nicht beeinträchtigt: Die Auswertung der
Blutagarplatte ergab nach 24 h keine Hämolyseaktivität. Nach 48 h wurde eine Entfärbung
geringen Umfangs jeweils an den Auftragsstellen beobachtet.
0
500
1000
1500
2000
2500
3000
Kontro
lle
DMSO-Kon
trolle
1222
1,76
µl
1222
1:10
1222
1:10
0
1366
1,76
µl
1366
1:10
1366
1:10
0

Ergebnisse 82
3.2.8. Apoptose-Assays Bei den in dieser Arbeit durchgeführten Versuchen zur Apoptose wurde Scytocyclamid A an
embryonalen Mäusefibroblasten-Zellen (MEF-SV40) getestet. Im Phasenkontrastmikroskop
ließ sich die Bildung von Vesikeln (blebbing) beobachten, was ein erster Hinweis auf
apoptotische Vorgänge sein könnte. Blebbing ist ein charakteristisches Kennzeichen für
apoptotische Prozesse. Alle Versuche wurden durch Durchflusszytometrie analysiert. Bei
dieser Methode werden Zellen einzeln angesaugt und durch einen Laserstrahl geschossen.
Während der Detektion können gleichzeitig verschiedene Parameter überprüft werden:
Vorwärts- und Seitwärtsstreulicht, sowie die verschiedenen Fluoreszenzqualitäten. Auf Kanal
1 wurde Vorwärtsstreulicht, auf Kanal 2 Seitwärtsstreulicht gelegt. Diese beiden
Grundparameter ermöglichten eine Vorauswahl an lebenden, nicht geschädigten Zellen, da
sich im Untergang befindliche Zellen üblicherweise einen hohen Gehalt an Granula im
Vergleich zur ansonsten homogenen Zellpopulation aufweisen. Der höhere Gehalt an Granula
bewirkt eine geringere Durchlässigkeit für Lichtquanten und die Zellen zeigen praktisch keine
Seitwärtslichtstreuung. Durch Setzen eines Tores („Gates“) wurden die Zellen ausgewählt,
deren Eigenschaften in Bezug auf diese Parameter nicht lebendige Zellen ausschließt. Der
erste Datenplot jeder Versuchsreihe zeigt die Ausgangssituation der betreffenden
Zellpopulation ohne Fluoreszenzmarkierungen (Abb. 37). Beim direkten Vergleich der beiden
Ausgangspopulationen ergab sich eine sichtbare Veränderung der „Zellwolke“ nach Zugabe
von Scytocyclamid A. Dies machte sich in einer Intensitätsabnahme der
Vorwärtslichtstreuung bemerkbar, wohingegen eine Zunahme der Lichtstreuung im
Seitwärtsscatter beobachtet wurde.
Ein sehr früh nachzuweisendes Apoptose-Merkmal ist der Verlust der asymmetrischen
Verteilung von Phosphatidylserin in der Plasmamembran der Zelle. Annexin V ist ein
Phospholipid-bindendes Protein, das in Anwesenheit von Calcium große Affinität zu dem
Phospholipid Phosphatidylserin besitzt. Dieses Phospholipid transloziert in einem frühen
Stadium der Apoptose von der Innen- zur Außenseite der Zellmembran (Vermes, 2000). FITC
markiertes Annexin V bindet Ca2+-abhängig Phosphatidylserin und diese Bindung kann durch
FACS-Analyse quantifiziert werden. Dabei werden die Zellen mit Propidiumiodid (PI)
gegengefärbt. In Kombination mit Propidiumiodid, einem fluoreszierenden Lebendfarbstoff,
der DNA solcher Zellen färbt, die die Membranintegrität verloren haben, kann man lebende
(AV-/PI-), früh apoptotische (AV+/PI-) und spät apoptotische / tote Zellen (AV+/PI+)
voneinander unterscheiden. Im Versuchsansatz befindet sich bei den Kontrollzellen
Phospharidylserin fast ausschließlich in der dem Cytoplasma zugewandten

Ergebnisse 83
Phospholipiddoppelschicht. Beim Vergleich der Kontrollzellpopulation und den mit
Scytocyclamid A behandelten Zellen (2x 105 Zellen, 10 µM, 5-6 h Inkubation) im Annexin V-
Versuchsansatz wurde eine zusätzliche „Zellwolke“ beobachtet, welche durch
Propidiumiodid-positive/Annexin V positive Zellen verursacht wurde. Hier lagen
spätapoptotische oder nekrotische Zellen vor. Die Zunahme im rechten unteren Quadranten
war auf Zellen im frühen apoptotischen Stadium zurückzuführen, welche Propidiumiodid
negativ (= vital) sind.
Bei induzierter Apoptose kann als mögliches weiteres Charakteristikum eine Reduktion des
Mitochondrien Membranpotentials beobachtet werden. Mitochondrien mit normalen
Membranpotential lagern bedingt durch die Ladungsdifferenz verstärkt den Farbstoff 3,3´-
Dihexyloxacarbocyanin-Iodid (DiOC6(3) in ihrer Membran ein. Sind die Mitochondrien einer
Zelle depolarisiert, so bleibt die Akkumulation des Fluoreszenzfarbstoffes DiOC6(3) in den
Mitochondrien aus. Die dann gemessene Fluoreszenz ist deutlich niedriger als bei Zellen mit
nicht depolarisierten Zellen. Die Fluoreszenz sowohl der Vergleichspopulation als auch der
mit Scytocyclamid A behandelten Zellen wiesen unveränderte Fluoreszenz auf. Eine mögliche
Apoptoseinduktion verläuft ohne Verlust des Mitochondrien Membranpotentials
Ein weiterer Mechanismus über den zytotoxische Substanzen Apoptose induzieren können,
verläuft über die Bildung von reaktiven Sauerstoffspezies (ROS). Dies wurde mit 2-
Hydroethidin, einem typischen Marker für die Generierung von ROS untersucht. Durch
reaktive Sauerstoffspezies gebildetes Ethidium interkaliert in die zelluläre DNA und zeigt
Fluoreszenz. Diese Auswirkung lässt sich im linken oberen Quadranten des DiOC6(3) / 2-HE
Versuches verfolgen (Abb. 37) Die Fluoreszenzmarkierung führte zu keiner Fluoreszenz bei
den Kontrollzellen, während die mit Scytocyclamid A behandelte Fraktion eine deutliche
Fluoreszenzzunahme in Form einer zusätzlichen „Zellwolke“ im linken oberen Quadranten
zeigte, was auf die Aktivität reaktiver Sauerstoffspezies zurückzuführen war (Abb. 37).
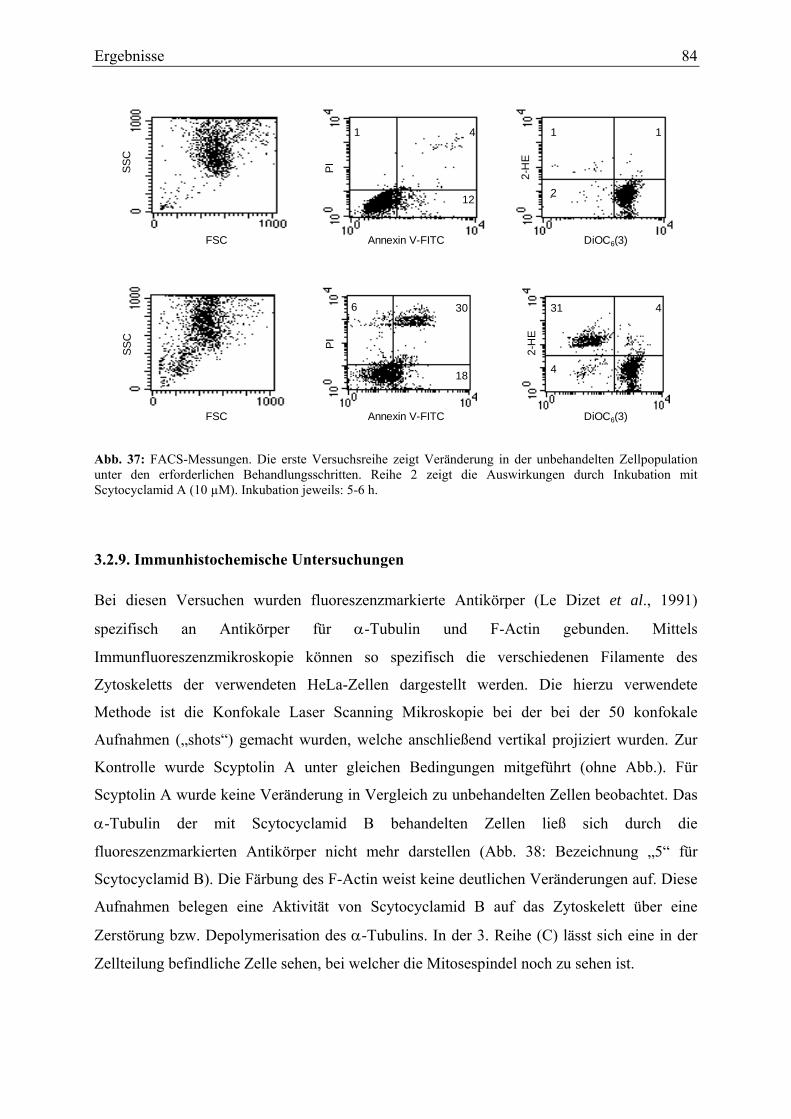
Ergebnisse 84
Abb. 37: FACS-Messungen. Die erste Versuchsreihe zeigt Veränderung in der unbehandelten Zellpopulation unter den erforderlichen Behandlungsschritten. Reihe 2 zeigt die Auswirkungen durch Inkubation mit Scytocyclamid A (10 µM). Inkubation jeweils: 5-6 h.
3.2.9. Immunhistochemische Untersuchungen Bei diesen Versuchen wurden fluoreszenzmarkierte Antikörper (Le Dizet et al., 1991)
spezifisch an Antikörper für α-Tubulin und F-Actin gebunden. Mittels
Immunfluoreszenzmikroskopie können so spezifisch die verschiedenen Filamente des
Zytoskeletts der verwendeten HeLa-Zellen dargestellt werden. Die hierzu verwendete
Methode ist die Konfokale Laser Scanning Mikroskopie bei der bei der 50 konfokale
Aufnahmen („shots“) gemacht wurden, welche anschließend vertikal projiziert wurden. Zur
Kontrolle wurde Scyptolin A unter gleichen Bedingungen mitgeführt (ohne Abb.). Für
Scyptolin A wurde keine Veränderung in Vergleich zu unbehandelten Zellen beobachtet. Das
α-Tubulin der mit Scytocyclamid B behandelten Zellen ließ sich durch die
fluoreszenzmarkierten Antikörper nicht mehr darstellen (Abb. 38: Bezeichnung „5“ für
Scytocyclamid B). Die Färbung des F-Actin weist keine deutlichen Veränderungen auf. Diese
Aufnahmen belegen eine Aktivität von Scytocyclamid B auf das Zytoskelett über eine
Zerstörung bzw. Depolymerisation des α-Tubulins. In der 3. Reihe (C) lässt sich eine in der
Zellteilung befindliche Zelle sehen, bei welcher die Mitosespindel noch zu sehen ist.
SSC
SSC
FSC
FSC Annexin V-FITC
Annexin V-FITC
PIPI
DiOC6(3)
DiOC6(3)
2-H
E2-
HE
1 11
12
4
2
31 4
418
306
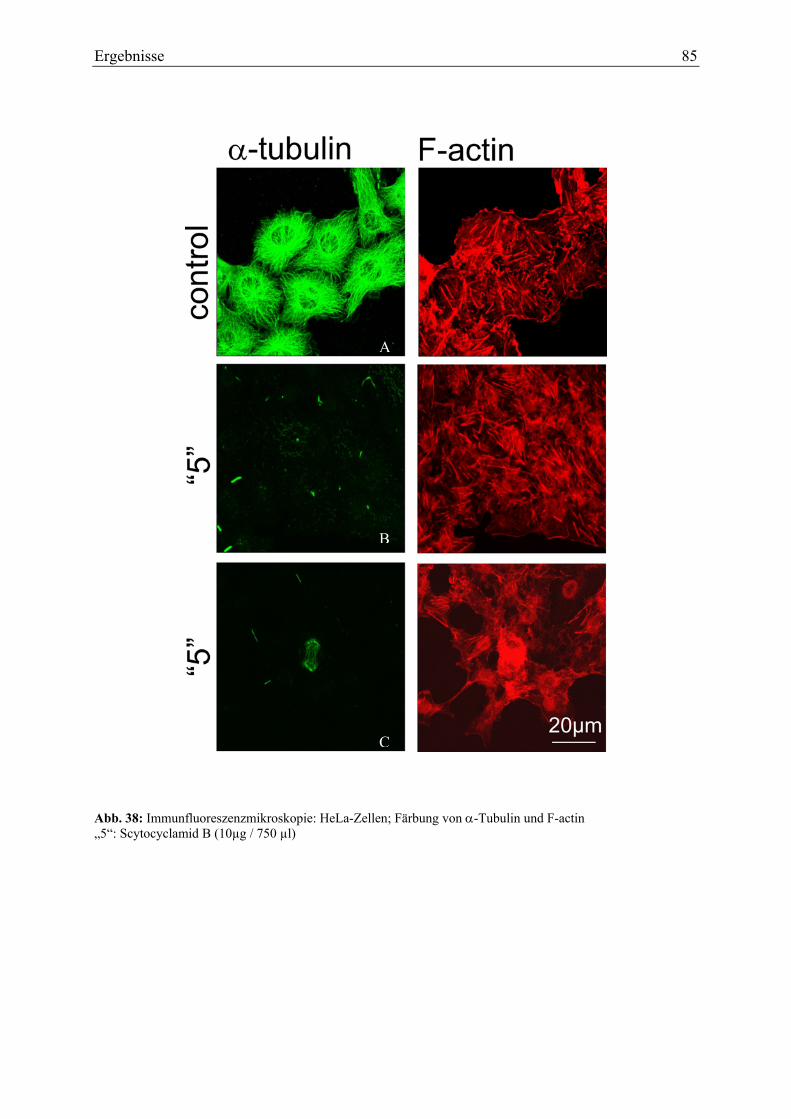
Ergebnisse 85
Abb. 38: Immunfluoreszenzmikroskopie: HeLa-Zellen; Färbung von α-Tubulin und F-actin „5“: Scytocyclamid B (10µg / 750 µl)
A
B
C

Diskussion 86
4. Diskussion Die vorgelegte Arbeit sollte zur Kenntnis von Cyanobakterien als Quelle unbekannter
Sekundärstoffwechselprodukte mit biologischer Aktivität beitragen. Untersucht wurden
Peptidmetabolite aus axenisch kultivierten Scytonema hofmanni PCC 7110 Zellen. Die heute
verfügbaren und in der Arbeit angewendeten chemischen Methoden haben die
Strukturaufklärung von Molekülen selbst in Mikrogramm-Mengen erlaubt. Dadurch war es
möglich, auch zuvor unbekannte Peptide, die teils in nur niedriger zellulärer Konzentration
vorlagen und von Matern (2002) noch nicht identifiziert wurden, aufzuklären. Diese
einleitenden Worte sind hier vorangestellt, da die zelluläre Konzentration der im Folgenden
beschriebenen Peptide 0,04 % bezogen auf das Trockengewicht nicht überstieg. Die
Besprechung der Isolierung der großenteils neu detektierten und identifizierten
Cyanopeptoline und der Scytocyclamide erfolgt im nächsten Abschnitt gemeinsam, die
Strukturaufklärung nach Substanzklassen getrennt.
4.1. Isolierung und Strukturaufklärung Ein im Arbeitskreis etabliertes Reinigungsprotokoll zu Peptiden aus Scytonema hofmanni
(Matern et al., 2001) sieht die Herstellung eines Rohextraktes aus lyophilisierten Zellen mit
70 % (v/v) Methanol vor. Da jedoch die Extraktionsmethode jeweils für die zu isolierenden
Peptide individuell anzupassen war, wurden die verschiedenen Verfahrensschritte überprüft.
Vorversuche dienten dazu die optimale Methanolkonzentration für die Extraktion zu
ermitteln. Die Methanolkonzentration konnte von 70 % auf 50 % (v/v) ohne merkbaren
Verlust an dem jeweiligen Peptid reduziert werden. Andere Extraktionsmittel wie 5 % (v/v)
wässrige Essigsäure oder Acetonitril brachten keine Vorteile. Nach Herstellung des
Rohextraktes aus lyophylisiertem Zellmaterial sieht das Reinigungsprotokoll ein
Größenausschlussverfahren über Gelpermeationschromatographie an Sephadex® LH 20 vor.
Zur Peptidisolierung (Abb. 2) wurde jedoch immer der gesamte Durchbruch verwendet, so
dass unter den gewählten Bedingungen die Funktion des Größenausschluss nicht
gewährleistet war. LH-20-Material ist über C3-Gruppen modifiziert. Über diese
Strukturmodifikation könnte eine Trennung über schwach lipophile Stoffeigenschaften
vorgenommen werden. Die LH-20-Eluate zeigten nach HPLC-Analyse weder Peptide in den
Chromatogrammen noch ergaben sich Hinweise auf verbesserte Stofftrennung des LH-20-
Durchbruchs im Vergleich zum methanolischen Rohextrakt. Wie aus dem in dieser Arbeit

Diskussion 87
verwendeten Reinigungsprotokoll zu entnehmen ist, wurde daher auf diesen Verfahrensschritt
verzichtet. Der nächste Reinigungsschritt sieht eine Bindung der im Rohextrakt befindlichen
Peptide an eine Festphase vor. Über amphiphile Wechselwirkungen werden die Peptide
vollständig an Chromabond® C18-Material gebunden, so dass der Durchbruch keine Peptide
im Chromatogramm zeigt. Auf weitere Verfahrensschritte der
Ionenaustauschchromatographie (Matern et al., 2001 und 2003) wurde daher ebenfalls
verzichtet, da sie die Ausbeute an neu zu isolierenden Peptide nicht erhöhten. Bei den
Versuchen hierzu wurde unter anderem festgestellt, dass die Bindung der Peptide an die
Festphasenmaterialien nicht ausschließlich auf ionische Wechselwirkungen zurückzuführen
ist, sondern lipophile Stoffeigenschaften einschließt. Zur Kontrolle der Anwendung der
Ionenaustauschchromatographie wäre eine zusätzliche Elution mit Methanol nach erfolgter
Elution mit dem salzhaltigen Elutionsmittel sinnvoll.
Die in der Arbeit charakterisierten Cyanopeptoline und Scytocyclamide wurden direkt aus den
C18-Eluaten per RP-HPLC-Auftrennungen isoliert. Da sich teilweise die Retentionszeiten der
Cyanopeptoline von den Scytocyclamiden während der HPLC-Auftrennung deutlich
unterschieden, wurde eine Vortrennung während der Festphasenchromatopgraphie
vorgenommen. Über die Elution mit steigender Methanolkonzentration wurden Fraktionen
mit unterschiedlichem Peptidmuster erzeugt. Die isokratische Fließmittelzusammensetzung
wurde für optimale Trennergebnisse im präparativen Maßstab jeweils angepasst. Gering
wechselnde Peptidzusammensetzungen der einzelnen Fraktionen von Extraktion zu
Extraktion ließen den Schluss zu, dass neben der gesteigerten Elutionskraft durch den
erhöhten Anteil an organischem Lösungsmittel auch die Menge an Elutionsmittel Einfluss auf
die Peptidzusammensetzung der Eluate hat. Dies konnte jedoch problemlos über die
Programmierung der Fließmittelzusammensetzung während der präparativen Aufbereitung
ausgeglichen werden.
Die Struktur der isolierten Peptide wurden schließlich durch Methoden der UV-/IR-
Spektroskopie, MALDI-TOF-/FTICR-Massenspektrometrie sowie zweidimensionale 1H und 13C-NMR-Messungen einschließlich Korrelationsmessungen aufgeklärt.
Scytocyclamide: Hinweise auf die Sequenzbausteine der isolierten Scytocyclamide (Abb. 24,
27 und 29) wurden über die Fragmentmassen der Immoniumionen aus den PSD-Spektren der
MALDI-TOF-Analysen oder aus charakteristischen Spinsystemen in 2D-NMR-
Korrelationsmessungen bekommen (beispielhaft gezeigt in Abb. 20 und 21). Teilweise wurde

Diskussion 88
aus den NMR-Daten direkt die Konfiguration einzelner Bausteine abgeleitet, so im Fall der E-
Konfiguration des Dehydrobutyrylrestes in Scytocyclamid A. Auch wurde die trans-
Konfiguration des 4-Hydroxyprolins anhand der NOE-Korrelationen bestimmt. Zur weiteren
Konfigurationsbestimmung wurde Scytocyclamid A einer Hydrolyse unterworfen, um die
entstehenden Produkte nach Derivatisierung an chiralen Phasen per Gaschromatographie zu
identifizieren. Hierzu wurden käufliche Aminosäuren als Referenzsubstanzen verwendet.
Aminooctansäure konnte mangels Referenz nicht näher bestimmt werden. Als problematisch
erwies sich, dass manche Aminosäuren enantiomer (Leucin), bzw. als Diastereomere
(Isoleucin) vorkamen. Die Struktur von Scytocyclamid A beinhaltet 2 Leucine, wie auch 2
Isoleucine. Die gaschromatographischen Untersuchungen belegten das Vorhandensein von je
einem L- und D-Leucin sowie von L-Isoleucin und einem D-Isoleucin, bzw. einem D-allo-
Isoleucin. Anhand der Retentionszeiten konnten D-Isoleucin und D-allo-Isoleucin nicht
unterschieden werden. Weiterhin war es unmöglich die genaue Lokalisierung der betreffenden
Aminosäuren vorzunehmen. Hierzu hätte man Partialhydrolysen durchführen können, mit
Bestimmung und Aufreinigung der Bruchstücke. Bedingt durch den geringen Gehalt an
Scytocyclamiden (ca. 0,04 % der Zelltrockenmasse) wurde hierauf verzichtet.
Anfang der 90iger Jahre war Laxaphycin A, eine Substanz mit großer Strukturähnlichkeit
beschrieben worden (Frankmölle, et al., 1992). Scytocyclamid A unterscheidet sich
Laxaphycin A (Kap. 1.7.2.1.) durch den Produzenten sowie in der Aminosäuresequenz, bei
der Glutamin anstatt Homoserin an Position2 eingebaut ist. Ein zu Laxaphycin
strukturähnliches Peptid, Hormothamnin, wird von einem weiteren Cyanobakterium
(Hormothamnium enteromorphoides) produziert. Der Unterschied zu Laxaphycin A besteht in
der Z-Konfiguration des Dehydrobutyrins (Gerwick et al., 1989). Ein weiteres
strukturverwandtes Peptid ist Lobocyclamid aus Lyngbya converfoides (MacMillan et al.,
2002). Lobocyclamide besitzen an Position2 ein Serin, an Position6 Tyrosin anstatt
Phenylalanin. Wenn man die Konfiguration der Strukturverwandten Laxaphycin A,
Hormothamnin A und Lobocyclamid A vergleicht, scheinen die zuordnungsrelevanten
Aminosäuren eine konservierte Konfiguration aufzuweisen. Deshalb lässt sich die folgende
Struktur Scytocyclamid A postulieren:

Diskussion 89
Abb. 39: Struktur von Scytocyclamid A (postulierte Stereochemie an der Stereozentren der Aminosäuren in Position1,7,8,9,10) Konfigurationsanalysen zu Scytocyclamid B und C konnten nicht durchgeführt werden, da
ihre Strukturen erst kürzlich gelöst wurden. Mit den Scytocyclamiden A bis C konnten für
Scytonema hofmanni PCC 7110 bis dato unbekannte Peptide charakterisiert werden. Die
Namensgebung der zuvor genannten und zu den Scytocyclamiden strukturverwandten Peptide
orientierte sich jeweils am produzierenden Organismus. Die Bezeichnungen Scytophycin
(Patterson et al., 1994) und Scytonemin (Helms et al., 1988) waren bereits für andere
cyanobakterielle Inhaltsstoffe vergeben, weshalb hier Scytocyclamid gewählt wurde.
Scytocyclamid B besitzt eine identische Aminosäuresequenz zu Laxaphycin D (Frankmölle,
et al., 1992). Die Stereochemie von Scytocyclamid B wurde im Rahmen dieser Arbeit nicht
bestimmt. Jedoch lässt sich durch die von Scytocyclamid A zu Laxaphycin A bestehende
Strukturverwandschaft auch bei Scytocyclamid B eine zu Laxaphycin D identische
Stereochemie erwarten.
Bei den Scytocyclamiden sind untypische Aminosäuren, wie β-Aminooctansäure, Homoserin
sowie D-Aminosäuren am Aufbau beteiligt. Dies ist als Hinweis auf nichtribosomale
Biosynthese zu sehen. Neben den genannten untypischen Aminosäuren besitzt Scytocyclamid
A Dehydrobutyrin als Baustein. Dehydrobutyrin wurde auch bei den zuvor genannten
strukturverwandten Peptiden sowie in Microcystinen gefunden (Beattie et al., 1998). Des
Weiteren kennt man Dehydro-Aminosäuren von Lantibiotika. Lantibiotika wurden nach der
untypischen Aminosäure Lanthionin benannt. Diese Aminosäure treten in der Natur
normalerweise nicht auf (Sahl et al., 1995). Die chemische Reaktion, welche für die
NH
ONH
NH
NH
O
OO
O
NOH
O
O
NH2
NH
ONH
ONH
ONH O
NHNH
O
OH
Aoc1
Gln2
Dhb3
Hyp4
Hse5Phe6
Leu7
allo-Ile8
Ile9
Leu10 Gly11

Diskussion 90
chemische Synthese dieser Aminosäuren gebraucht wird wurde erstmals durch Ingram (1970)
postuliert. Dehydratisierung von Threonin führt zu Dehydrobutyrin. Es wäre denkbar, dass in
Scytocyclamid A Dehydrobutyrin ebenfalls aus Threonin gebildet wird. Anhand der
Massendaten, welche direkt aus dem Rohextrakt generiert wurden ergibt sich kein Hinweis
auf Threonin in der Aminosäuresequenz.
Cyanopeptoline: Das Molekulargewicht der Cyanopeptoline wurde über MALDI-TOF-MS
bzw. durch ein Verfahren mit gepulster Laserionisierung bestimmt, welches eigens für die
Detektion cyanobakterieller Peptide entwickelt worden war (Erhard et al., 1998). Im letzteren
Verfahren werden Mutterionen im Flug weiter fragmentiert, und man erhält post source decay
(PSD)-Spektren aus wenigen, teilweise überlappenden Fragmentionen. Ein Vergleich der
Fragmentionen der neu isolierten Peptide mit Fragmentmassen aus einer Fragmentdatenbank,
welche Angaben zu Molekulargewichten und charakteristischen PSD-Fragmenten für mehr
als 2.000 cyanobakterielle Peptide enthält brachte keine Strukturhinweise für die neu zu
charakterisierenden Peptide (Erhard, 1999). Strukturhinweise auf die neu identifizierten
Cyanopeptoline wurden hauptsächlich über Vergleiche ihrer PSD-Spektren mit denen von
Matern et al. (2002) bereits charakterisierter Cyanopeptoline gewonnen. So konnten damals
anhand der Massenunterschiede verschiedene Modifikationen vorgeschlagen werden, welche
über umfangreiche NMR-Analysen bestätigt bzw. gelöst wurden. Da in der jetzt vorgelegten
Arbeit die Ausbeute an den einzelnen Peptid bezogen auf das Trockengewicht der Zellen mit
max. 0,04 % sehr gering war, musste auf eine Konfigurationsanalyse verzichtet werden.
Bisher wurden für die Aminosäuren der Cyanopeptoline jedoch nur L-Konfigurationen
bestimmt, so dass für die neu charakterisierten Cyanopeptoline diese Konfiguration ebenfalls
postuliert wird. Diese Annahme wird durch eine von Matern et al. (2003) durchgeführte
Konfigurationsanalyse gestützt, bei welcher für Scyptolin A und B ausschließlich L-
konfigurierte Aminosäuren gefunden wurden. Die isolierten Cyanopeptoline weisen die
Charakteristika auf, welche sie in die Klasse der Cyanopeptoline subsumiert (Jakobi et al.,
1995): es sind zyklische Depsipeptide, bei denen der Ringschluß über eine Esterbindung zum
Lacton erfolgt. Die Ahp-Gruppe ist jeweils vorhanden.
Die in der Massenspektrometrie auftretenden Signale, welche [M-H2O+H]+ zugeordnet
wurden, gehen vermutlich auf Dehydratisierung unter Ausbildung einer Doppelbindung
zurück (Forchert, 1999). Dieser Effekt trat bei ESI-MS noch stärker in Erscheinung, so dass
hier von einer Fragmentierung in der Ionenquelle auszugehen ist. Ein Nachweis der

Diskussion 91
Wasserabspaltung konnte durch NMR-Analytik nicht gezeigt werden, so dass von einer
Quellenfragmentierung auszugehen ist.
Cyanopeptolin 946A wurde durch Einbau von nicht-chloriertem Tyrosin im Vergleich zu
Scyptolin A modifiziert. Veränderungen der Bioaktivität gegenüber den Scyptolinen wurden
nicht gefunden. Daher stellt sich die Frage, weshalb diese Strukturmodifikation gebildet wird.
Eine einfache Erklärung wäre, dass bei der Biosynthese im Multienzymkomplex der
Unterschied zwischen chlorierter und nicht-chlorierter Aminosäure nicht erkannt wird und der
geringe Gehalt an dieser Strukturmodifikation durch die „Fehlerrate“ zu erklären ist. Denkbar
wäre, dass die Halogenase, welche für die Chlorierung zuständig ist, ein gewisses
Fehlerpotential aufweist. Molekulargenetische Untersuchungen könnten hier Aufschluss
geben.
In der Arbeit von Matern et al. (2002) wird Scyptolin A als mögliches Artefakt von Scyptolin
B diskutiert. Würde Scyptolin A aus Scyptolin B durch Esterhydrolyse während der
Aufreinigung entstehen, so müsste eine Strukturvariation von Scyptolin B entstehen, welche
am Tyrosin nicht chloriert ist. Dieses Artefakt konnte mit LC-MS-Analysen, sowie durch
MALDI-TOF-Analysen nicht nachgewiesen werden. Auch die über C18-Festphasen
konzentrierten Eluate zeigen diese Masse nicht, so dass man zu dem Schluss kommen kann,
dass Scyptolin A nicht aus Scyptolin B entstanden ist. Jedoch wird durch diese Feststellung
eine Fehlsteuerung während der Biosynthese, welche die Produktion von Cyanopeptolin 946A
erklärt, unwahrscheinlich, da Scyptolin B in größerer Menge als Scyptolin A synthetisiert
wird. Somit müsste auch ein Artefakt mit der Masse von 1.070 Da zu finden sein, welches
Scyptolin B ohne Chlorierung entspricht. Insofern bleibt die Bedeutung der Synthese von
Strukturvarianten weiterhin spekulativ. Die Untersuchung der Biosynthese sollte in
absehbarer Zeit die Antwort geben, da molekulargenetische Untersuchungen zur Biosynthese
nichtribosomal gebildeter Sekundärstoffwechselprodukte im Fokus vieler Arbeitsgruppen
liegen (Chang et al., 2002, Hofmann et al., 2003, Grünewald et al., 2005). Neuere genetische
Studien bestätigen, dass Depsipeptide in Cyanobakterien über nichtribosomale Peptid-
Synthetasen gebildet werden. Peptid-Synthetasen wurden für Anabaenopeptilide (Rouhiainen
et al., 2000), Nostocyclopeptide (Becker et al., 2004) sowie Nostopeptolid A (Hofmann et al.,
2003) charakterisiert. Die modulare Struktur der Peptid-Synthetasen erleichtert den Austausch
von Aminosäurebausteinen innerhalb der Peptide und die Bildung einer großen Zahl an
Strukturvarianten mit unterschiedlichen Zielspezifitäten und Wirkungen (Mootz et al., 2000).
Cyanopeptolin 1060A hat bis auf eine Methylengruppe in der Seitenkette eine zu
Hofmannolin (Matern et al., 2001) identische Struktur. Eine Konfigurationsanalyse wurde

Diskussion 92
nicht durchgeführt, jedoch wird durch die Strukturanalogie von einer identischen
Konfiguration ausgegangen. Strukturmodifikationen in der Seitenkette sind typische
Erscheinungen bei Cyanopeptolinen. Besonders häufig ist der N-Terminus durch eine Säure
acyliert. So ist beispielsweise der N-Terminus von Cyanopeptolin 954 ist durch Essigsäure
acyliert (von Elert et al., 2005). Bei Tasipeptinen findet man Butyrylreste am N-Terminus
(Williams et al., 2003).
Cyanopeptolin 1067A besitzt strukturelle Ähnlichkeit zu Hofmannolin (Matern et al., 2001).
In Position4 findet sich jedoch bei Cyanopeptolin 1066 mit Arginin eine basische Aminosäure.
Cyanopeptoline mit Arginin zwischen der Ahp-Gruppe und Threonin sind häufig beobachtete
Strukturvarinaten: Cyanopeptolin A (Martin et al., 1993), Cyanopeptolin S und SS (Jakobi et
al., 1995 und 1996), Micropeptin 90 (Ishida et al., 1995), Micropeptin EI992 (Ploutno et al.,
2002), Oscillapeptin J (Blom et al., 2003) seien beispielhaft angeführt. Mit der Bestätigung
der Struktur von Cyanopeptolin 1067A durch NMR-Analysen konnte ein von Matern et al.
(2002) nur anhand der MS-Daten und Datenbankvergleich getroffener Strukturvorschlag: 3-
(4-Hydroxyphenyl)-lactyl-Glu-(Thr-Arg-Ahp-Leu/Ile-NMTyr-Leu/Ile-O-) widerlegt werden.
Für die Seitenkette wurde Glutaminsäure und als N-Acylrest para-Hydroxyphenylmilchsäure
mit dem Hinweis auf notwendige Überprüfung postuliert. Da die Struktur von Hofmannolin
bekannt war, konnte die Zuordnung der Fragmentmassen durch Vergleich mit den beiden
Spektren erfolgen. Viele der von Matern et al. zugeordneten Fragmentmassen wiesen
Übereinstimmung mit den gemessenen Massenzahlen auf, trotzdem waren der Acylrest der
Seitenkette sowie die Leucine, bzw. Isoleucine nicht in der Sequenz von Cyanopeptolin
1067A enthalten. Bei den Fragmentzuordnungen fehlte unter anderem das für den Ring ohne
Seitenkette charakteristische Fragment, welches normalerweise in den Fragmentspektren von
Cyanopeptolinen deutlich zu erkennen ist (siehe PSD-Fragmentzuordnungen Tabelle 10, 13
und 15)
4.2. Biologische Aktivität
4.2.1. Enzymhemmwirkung
Von den in dieser Arbeit charakterisierten Peptiden haben die Scytocyclamide A bis C keine
Enzymhemmung in den gewählten Aktivitätstests. Bei den Cyanopeptolinen ergeben sich
vergleichbare Aktivitäten zu den von Matern et al. (2001 und 2003) untersuchten Scyptolinen
A und B sowie Hofmannolin. So hemmt Cyanopeptolin 946A Elastase spezifisch und potent

Diskussion 93
(IC50 0,48 µM). Die Werte für die Scyptoline A und B liegen vergleichbar bei 0,14 µM
(Matern et al., 2001), was sich schon durch die strukturelle Ähnlichkeit vermuten ließ.
Anhand der von Matern et al. (2003) publizierten kristallographischen Daten lässt sich kein
Hinweis auf eine Beteiligung der Chloridfunktion an der Bindung an die Inhibitor-
Bindungsstellen im aktiven Zentrum des Enzyms feststellen. Mit den Erfahrungen aus der
Gesamtheit der für diese Arbeit durchgeführten Enzymhemmtests kann man die relativ
geringen Unterschiede (ca. dreifach) im Wirkungsgrad auf die unterschiedlichen
Enzymchargen und damit verbundenen Aktivitätsunterschiede zurückführen. Außerdem sind
Enzymhemmtests relativ empfindlich für Störungen.
Für Cyanopeptolin 1060A konnte trotz des erweiterten Spektrums an getesteten Enzymen
keine Enzymhemmwirkung nachgewiesen werden. Cyanopeptolin 1060A unterscheidet sich
von Hofmannolin nur durch eine um eine Methylengruppe verkürzte Seitenkette. Diese dürfte
auf eine mögliche Enzymhemmung keinen Einfluss haben. Hofmannolin ist mit Scyptolin B
das mengenmäßig am stärksten exprimierte Peptid in Scytonema hofmanni PCC 7110.
Postuliert man wie von Elert et al. (2005), dass das primäre Ziel der von Cyanobakterien
produzierten Proteasehemmstoffe die Verdauungsenzyme der Primärkonsumenten ist, dann
stellt sich die Frage, ob nicht alle Strukturverwandten der Cyanopeptoline Enzymhemmstoffe
sein müssten.
Für Cyanoeptolin 1067A konnte indes eine potente Hemmung von Trypsin (IC50 60 nM)
nachgewiesen werden. Plasmin wurde ebenfalls gehemmt (IC50-Wert von 0,97 µM). Da sich
die Strukturen von Cyanopeptolin 1067A und Hofmannolin nur durch den Austausch der
Aminosäure an Position4 unterscheiden, kann die unterschiedliche Wirkung auf die basische
Aminosäure zurückgeführt werden. Dieses Ergebnis steht in Einklang mit den bisher
publizierten Daten zu Cyanopeptolinen, welche ebenfalls an Position4 eine der basischen
Aminosäuren Arginin oder Lysin aufweisen (z. B. von Elert et al., 2005, Czarnecki et al.,
2005) und Hemmstoffe von Trypsin sind.
4.2.2. Testung auf antimikrobielle Aktivität
Von einigen Inhaltsstoffen aus Cyanobakterien ist auch antimikrobielle Aktivität bekannt
(Ostenvik et al., 1998). Inhaltsstoffe aus Scytonema sp. TISTR 8208 wurden als stark
wirksame antimikrobielle Substanzen mit Wirkung auf gram-positive wie gram-negative
Bakterien und pathogene Hefen beschrieben (Chetsumon et al., 1993). Zugehörige Strukturen
wurden jedoch nicht veröffentlicht. Auch wurden Laxaphycine mit antifungaler Wirkung

Diskussion 94
beschrieben, wobei Laxaphycin A nur in Kombination mit Laxaphycin B eine Wirkung
zugeschrieben werden konnte (Frankmölle et al., 1992). Dies waren Anhaltspunkte auch bei
den neu charakterisierten Scytocyclamiden eine Aktivität erwarten zu lassen. Jedoch wurde
hier bis zu einer maximalen Konzentration von 64 µg/ml keine Wirkung beobachtet.
4.2.3. Toxizitäts-Assays 4.2.3.1. Toxizitätstest mit Primärkonsumenten
Um eine mögliche ökologische Funktion der untersuchten Peptide zu überprüfen, wurden die
isolierten Peptide in ihrer toxischen Wirkung auf Primärkonsumenten getestet. In der Literatur
finden sich Hinweise, dass Cyanopeptoline toxische Wirkungen bei Primärkonsumenten
verursachen, so beispielsweise durch Oscillapeptin J, einem Cyanopeptolin, welches Arginin
als Aminosäurebaustein am N-Terminus der Ahp-Gruppe besitzt (Blom et al., 2003). Die hier
ebenfalls getesteten Scyptoline A und B sowie Hofmannolin hatten jedoch keine toxischen
Effekte bei Thamnocephalus platyurus. Da Cyanopeptolin 1067A zum Untersuchungszeit-
punkt noch nicht identifiziert war, blieb eine Antwort hier offen.
Die Scytocyclamide A und B, erwiesen sich gegenüber Thamnocephalus platyurus als hoch
toxisch, so wurde bereits nach 2 h im 24 h Toxizitätstest die letale Wirkung durch diese
Peptide beobachtet. Dieses Ergebnis steht im Einklang mit dem beschriebenen Fraßschutz bei
Laxaphycinen. Hormothamnion enteromorphoides scheint nicht als Nahrungsquelle für
Herbivore zu dienen, weshalb Fraßschutz verleihende Inhaltsstoffe vermutet wurden
(Pennings et al., 1997). In der untersuchten Spezies wurde Laxaphycin A als
Hauptkomponente bestimmt, jedoch kein Hormothamnin A. Für Laxaphycin A und auch für
eine Mischung der isolierten Peptide wurde Fraßschutz nachgewiesen. Bei dem hier
durchgeführtem Toxizitätstest wird der isolierte Wirkstoff zum Medium gegeben, so dass die
Crustaceen zwangsweise in Kontakt mit dem Wirkstoff kommen, während sie in der Natur
den Verzehr der cyanobakteriellen Zellen vermeiden können. Eine Wirkung der
Scytocyclamide A und B auf Daphnia magna wurde in dieser Arbeit zwar in Vorversuchen
beobachtet, die jedoch der Reproduzierung bedürfen.
4.2.3.2. Testung auf Zytotoxizität
Bereits Ende der 80iger, Anfang der 90iger Jahre wurden cyanobakterielle Inhaltsstoffe als
potentielle zytotoxische Substanzen beschrieben. Carmeli et al. (1990) beschreibt Tolytoxine
und Scytophycine mit zytotoxischen Eigenschaften. Diese Substanzen wurden aus 3

Diskussion 95
verschiedenen Scytonema-Spezies isoliert. Zur gleichen Zeit wurde ein breit angelegtes
Screening-Programm initiiert, welches Laborstämme von Cyanobakterien als Quelle neuer
antineoplastischer Substanzen als Untersuchungsgegenstand hatte (Patterson et al., 1991). Im
Verlauf dieser Untersuchungen wurden die Familien der Scytonemataceae und
Stigonemataceae als überaus produktive Erzeuger neuer zytotoxischer Substanzen
identifiziert. Die Testung der Cyanopeptoline (Scyptoline A und B sowie Hofmannolin) ergab
keine zytotoxische Wirkung gegenüber den getesteten Zelllinien. Meist wurde Zytotoxizität
nur als Routinetest durchgeführt, so dass in vielen Veröffentlichungen keine Hinweise auf
einen möglichen Wirkungsmechanismus gegeben werden. Untersuchungen zur Zytotoxizität
der Laxaphycine A und B ohne nähere Hinweise auf den Wirkungsmechanismus wurden
kürzlich durch Gbankoto et al. (2005) beschrieben.
Die in der vorgelegten Arbeit isolierten Scytocyclamide A bis C erwiesen sich in einer
Konzentration von ca. 10 µM als toxisch, wobei die Wirkung direkt nach Applikation
beobachtet wurde. Die visuelle Auswertung deutete auch auf morphologische Veränderungen
hin, bei denen blebbing beobachtet wurde (Abb. 35). Blebbing tritt bei apoptotischen
Vorgängen auf, weshalb unter anderem auch auf Apoptose getestet wurde. Die zelltoxische
Wirkung wurde an mehreren Zelllinien (Kap. 3.2.4.) belegt. Die Wirkung der Scytocyclamide
A bis C weist insofern keine Spezifität innerhalb der eingesetzten Zelllinien auf. Der erste
Eindruck ließ Zelllyse und eventuell eine Zerstörung der Zellmembran vermuten. Da die
Peptide in DMSO als Lösungsmittel gelöst waren (1µg/µl), wurde jeweils eine DMSO
Kontrolle mitgeführt (die Verdünnung des eingangs unverdünnten DMSO erfolgte durch die
Zugabe in die Kavität der Ansätze). Weitere Faktoren wie Einflüsse des Fließmittels aus den
HPLC-Läufen wurden ausgeschlossen: zum einen unterscheiden sich die Peptide in den
Retentionszeiten um bis zu 5 min, so dass Wechselwirkungen zwischen den Peptiden
ausgeschlossen werden konnten, zum anderen haben alle 3 Scytocyclamide zytotoxische
Wirkung. Cyanopeptoline, welche aus der gleichen Extraktion unter gleichen HPLC-
Bedingungen isoliert wurden, waren unwirksam. Zusätzlich wurden 2 Fließmittelfraktionen
mit unterschiedlichen Retentionszeiten zu den Scytocyclamiden gesammelt und unter
identischen Bedingungen aufbereitet und auf Wirkung getestet. Alle Kontrollen belegen, dass
die Scytocyclamide für die zytotoxische Wirkung verantwortlich sind. Da die Scytocyclamide
relativ hydrophob sind (sie eluieren erst bei hohem Anteil organischen Lösungsmittel von der
Säule), aber auch funktionelle Gruppen mit hydrophilem Charakter besitzen, wurde überlegt
inwieweit eine Detergenswirkung Ursache für die Zellschädigung sein kann. Um sich einen
Eindruck einer Detergenswirkung auf die getesteten Zellen zu machen, wurden Lösungen

Diskussion 96
verschiedener Detergentien (Tween 1 % und 0,02 %, Triton X 1 % und 0,04 %, Thesit 1 %
und 0,02 %, SDS 1 % und 0,4 %, Zwittergent 1 % und 0,2 %, Chaps 1 % und 0,5 % jeweils
Volumenprozent in H2O) auf ihre Wirkung auf HeLa-Zellen getestet. Die von den
Detergentien bewirkten Veränderungen der Zellmorphologie ließen im Vergleich mit der
Wirkung der Scytocyclamide A bis C keinen Rückschluss auf einen Zelltod durch
Detergenswirkung durch die Scytocyclamide zu. Eine weitere Überlegung war, ob eventuell
Porenbildung für die Wirkung in Frage kommt. Die Literatur beschreibt Membrankanäle
bildende Peptide mit Molekulargewichten zwischen 1.000 und 1.500 Da. Beispiele hierfür
sind das zyklische Gramicidin S (Gause et al., 1944 und Kiricsi et al., 2002) sowie
Valinomycin (z. B. Dolowy, 2001). In einem Vorversuch wurde deshalb mit Valinomycin (10
µM) ein Ionophor als Vergleichssubstanz bei den Zytotoxizitätsassays mitgeführt. Nach
Applikation des Valinomycins konnte keine direkte morphologische Veränderung an den
Zellen beobachtet werden, so dass auch hier keine zu den Scytocyclamiden A bis C
vergleichbare Wirkung vermutet wurde. Um einer möglichen Porenbildung als Mechanismus
für den durch Scytocyclamide A bis C ausgelösten Zelltod nachzugehen, wurden „lipid-
bilayer“ Ansätze durchgeführt. Eine geringfügig erhöhte Membranleitfähigkeit lieferte keinen
Hinweis auf Porenbildung bei Verwendung von Diphytanoyl-Phosphatidylcholin als
Membrankomponente.
Die strukturverwandten Laxaphycine besitzen neben der Zytotoxizität auf KB-Zellen
(Hautkrebs) und LoVo-Zellen (colorektales Adenokarzinom) (Frankmölle et al., 1992)
antifungale Wirkung u.a. gegen Aspergillus oryzae, Candida albicans, wobei Laxaphycin A
nur in Kombination mit Laxaphycin B wirkt (Bonnard et al., 1997). Die Scytocyclamide A
bis C haben dagegen keine antibakterielle und auch keine antifungale Wirkung (Kap. 3.2.2.).
Da Laxaphycin A als Reinsubstanz unwirksam gegen Pilzpathogene ist und nur in
Kombination mit Laxaphycin B wirkt, könnte das in Einklang mit den Ergebnissen zu
Scytocyclamiden stehen, welche als Reinsubstanzen und im Gemisch unwirksam sind und
bisher kein Synergiepartner gefunden wurde.
Die Scytocyclamide A bis C besitzen keine Wirkung gegenüber den getesteten
Mikroorganismen. Es wäre jedoch möglich, dass sie einen spezifischen Partner in der
Membran brauchen, um ihre Wirkung zu entfalten. Die antifungale Wirkung der Laxaphycine
könnte sich durch Ergosterol in der Zellmembran erklären lassen, Bakterien fehlt jedoch
Cholesterol in der Membran. Daher wurden ergänzend Versuche an „lipid bilayern“ mit
cholesterolhaltigen künstlichen Membranen durchgeführt (Kap. 3.2.5.), wobei für die
Scytocyclamide A bis C erhöhte Membranleitfähigkeit festgestellt wurde (persönliche

Diskussion 97
Mitteilung R. Benz, Universität Würzburg). Ob dies als Hinweis auf den
Wirkungsmechanismus zu werten ist, muss dennoch vorerst offen bleiben. Hierzu müssten
weitere valide Untersuchungen durchgeführt werden. Dies war im Rahmen dieser Arbeit nicht
mehr möglich. Um die vermutete Membranschädigung zu überprüfen, wurden Versuche zum 86Rb+-Efflux aus HT-29-Tumorzellen durchgeführt. Das Ergebnis belegte eindeutig erhöhte
Effluxraten, welche auf die Peptidwirkung zurückzuführen waren und Membranschädigung
belegen. Eine mitgeführte DMSO-Kontrolle schließt eine Beteiligung des Lösungsvermittlers
an der Membranschädigung aus.
Weiterführende Versuche wurden mit Scytocyclamid B unternommen, um die
morphologischen Veränderungen zu erklären. Mit Fluoreszenzfärbung wurde α-Tubulin und
F-actin spezifisch angefärbt (Abb. 38). Durch Phalloidin-fluoreszenzmarkierte Antikörper
(Alexa Fluor® 594 phalloidin+anti-Ratte oder anti-Maus IgG) wurde F-Aktin gefärbt. Es
ließen sich keine Veränderungen durch Scytocyclamid B am F-Aktin Zytoskelett beobachten.
Scytocyclamid B behandelte Zellen lassen keine α-Tubulinfärbung (Alexa Fluor® 488
F(ab')2+anti-α-tubulin III Antikörper) mehr erkennen. Dies legt den Schluss nahe, dass der
Wirkungsmechanismus der Zellschädigung zusätzlich über Tubulinabbau erfolgen könnte. In
der Literatur sind wenige cyanobakterielle Sekundärmetabolite beschrieben, welche auf
Tubulin wirken. So wurde von Patterson et al. (1993) das aus Scytonema sp. isolierte
Tolytoxin mit zytostatischer und antifungaler Wirkung beschrieben (siehe auch Kap. 1.7.2.4.):
Tolytoxin gehört zur Klasse der Makrolide. Die zytostatische Wirkung beruht auf der
Zerstörung der Mikrofilamente, während kein Effekt auf Mikrotubuli oder die
Intermediärfilamente beschrieben wurde. Aus Nostoc sp. wurde Cryptophycin isoliert (Kap.
1.7.2.4.). Cryptophycin wirkt als Inhibitor der Mitose und die Zellteilung wird unterbrochen.
Des Weiteren wird der Abbau von Mikrotubuli beschrieben (Chen et al., 1998). Dieses
Phänomen würde den hier beobachteten Effekten entsprechen, jedoch lassen die strukturellen
Unterschiede von Scytocyclamiden und Cryptophycinen keine Verallgemeinerung im Sinne
eines vergleichbaren Mechanismus zu. Das aus Lyngbya majuscula isolierte Curacin A besitzt
ebenfalls toxische Wirkungen auf Krebszellen, indem es in die Mitose eingreift. Curacin A
bindet dabei an die Colchicin-Bindungsstelle der Mikrotubuli (Verdier-Pinard et al., 1998)
Für Microcystine wird eine Zerstörung der Intermediärfilamente im Zusammenhang mit der
hepatotoxischen Wirkung gesehen (Ding et al., 2001). Bei der genannten Untersuchung
wurde ebenfalls die Bildung reaktiver Sauerstoffspezies durch die Gabe von Microcystin LR
beobachtet, gefolgt von der Alterung der Zytoskelettstrukturen. Chen et al. (1998) vermutet,
dass reaktive Sauerstoffspezies eine elementare Rolle in der durch Microcystin LR

Diskussion 98
induzierten Zerstörung des Zytoskeletts spielen. Versuche zur Apoptose (Kap. 3.2.8.) belegten
auch für Scytocyclamide die Bildung reaktiver Sauerstoffspezies. Eine direkte Verbindung
kann jedoch auch hier nicht hergestellt werden, da Microcystine weitere Effekte verursachen,
wie z. B. Hyperphosphorelierung nach Hemmung der Proteinphosphatase 1A.
Scytocyclamide hatten keine Hemmwirkung auf die Phosphatase (Kap. 3.2.1.). Weitere auf
den Wirkungsmechanismus zielende Versuche waren die Untersuchung apoptotischer
Charakteristika. Die beobachteten durch „blebbing“ verursachten „apoptotic bodies“ (Abb.
35) sind typisch für Apoptose. In der Literatur der letzten Jahre finden sich vermehrt
Untersuchungen apoptotischer Vorgänge, welche durch cyanobakterielle Inhaltsstoffe
induziert werden (Herfindal et al., 2005; Ding et al., 2003; Botha et al., 2004). Die hierzu in
der vorgelegten Arbeit durchgeführten Versuche (Kap. 3.2.8.) belegen, dass Scytocyclamid A
keine Caspase-Aktivität und kein Verlust des Mitochondrienmembranpotentials induziert.
Somit können diese Signaltransduktionswege zur Apoptoseeinleitung ausgeschlossen werden
(Kap. 3.2.8.). Die Daten aus den Versuchen zu reaktiven Sauerstoffspezies (Abb. 37) belegen,
dass reaktive Sauerstoffspezies in einen Apoptoseprozess involviert sein können.
Weiterführende Versuche sollten zur Bestätigung durchgeführt werden. So sollte die Bildung
reaktiver Sauerstofferstoffspezies durch die Zugabe von Antioxidantien wie N-Acetylcystein
zu unterbinden sein (Gottlieb et al., 2000). Membranschädigung und Apoptose schließen sich
nicht gegenseitig aus. Es wäre auch denkbar, dass mehrere Prozesse unabhängig voneinander
verlaufen. Apoptose wurde nach 5-6 h Inkubation gemessen, während die morphologischen
Veränderungen direkt nach Applikation beobachtet wurden. Dies würde für unabhängige
Prozesse sprechen. Bei den Filmaufnahmen zur Wirkung der Scytocyclamide A und B auf
HeLa und Ebl-Zellen mit einem Differential-Phasenkontrast-Mikroskop, bzw. mit einem
Phasenkontrastmikroskop konnte eine zellschädigende Wirkung in Bruchteilen von Sekunden,
bzw. in wenigen Sekunden verfolgt werden (siehe DVD im Anhang). Diese Beobachtungen
stehen wiederum in Einklang mit den Ergebnissen von der Fluoreszenzfärbung.
Weiterführende Versuche zur Klärung des Wirkungsmechanismus (ß-Tubulinfärbung, weitere
Immunohistochemische-Untersuchungen, Dosis-Wirkungsstudien, Involvierung von Ca2+-
Kanälen) könnten die bereits durchgeführten und diskutierten Experimente ergänzen.
4.3. Ökologie cyanobakterieller Sekundärstoffwechselprodukte Wie anfangs erwähnt, besiedeln Cyanobakterien viele verschiedene, teilweise extreme
Lebensräume. Die große Vielfalt an den von ihnen bewohnten ökologischen Nischen ist ein

Diskussion 99
möglicher Hinweis auf einen hohen Grad biologischer Adaptation, welcher es den
Cyanobakterien ermöglicht sich erfolgreich in der Natur zu behaupten. Mit ein Grund dafür
könnte ihre Fähigkeit sein, ein breites Spektrum an defensiven Metaboliten zu produzieren.
Diese Sekundärstoffwechselprodukte zeigen ein breites Spektrum biologischer Aktivitäten,
vor allem wachstumshemmende oder entwicklungshemmende Eigenschaften (Burja et al.,
2002). Unter den Sekundärstoffwechselprodukten aus Cyanobakterien finden sich z. B.
verschiedene Toxine (s. Kap. 1.7.1.). Sie gehören zu den meist untersuchten Stoffen, da von
ihnen potentielle gesundheitliche Gefahr ausgeht. Über die Funktion cyanobakterieller Toxine
gibt es, wie auch bei vielen anderen bioaktiven Sekundärmetaboliten von Bakterien, Algen
oder Pilzen, eher Spekulationen als gesicherte Erkenntnisse. Unter anderem wurde über die
Funktion von Peptiden als Speicherstoffe spekuliert, was jedoch vermutlich ausgeschlossen
werden kann, da auch unter Mangelbedingungen kein nennenswerter Abbau der Peptide
innerhalb der Zelle beobachtet wird (Chorus et al., 1999). Daneben wurde
Lichtschutzfunktion diskutiert, jedoch ist dies auch eher unwahrscheinlich, da es ebenso
Spezies gibt, welche keine Toxinbildung haben. In den meisten Fällen ist nicht einmal
bekannt, wie die Biosynthese bzw. die Akkumulation der Verbindungen reguliert wird, oder
ob es induzierende Faktoren gibt, welche dazu führen, dass manche Spezies toxinbildend ist,
während verwandte Spezies dieses Merkmal nicht aufweisen. Eine nahe liegende Bedeutung
für die Bildung von Toxinen könnte in der Abwehr von Feinden liegen (Vining, 1990). Dieser
Kontext könnte erklären, dass „schutzlose“ marine Invertebraten, welche sich kaum gegen
Feinde wehren oder vor ihnen fliehen können bioaktive Substanzen bilden, während das für
bewegliche oder gepanzerte Meeresbewohner nicht gilt (Pawlik, 1993). Die Abwehr von
Fraßfeinden kann sich in Form einer „chemischen Abwehr“ äußern. Die direkte Bekämpfung
von Nahrungskonkurrenten wäre über die Absonderung von Toxinen möglich, doch trotz
intensiver Forschung gibt es bisher keinen Beleg, dass die Toxine aus der Zelle exportiert
werden. Bisher gelten cyanobakterielle Toxine als Endotoxine (Watanabe et al., 1996).
Jüngste Daten von Sequenzanalysen des Gens mcyH lassen einen ABC-Transporter für dieses
Protein vermuten, welcher Verwandschaft zu Proteinen aufweist, die bei anderen Bakterien
für den Export von Peptiden zuständig sind (Boerner, 2001). Der Export konnte jedoch noch
nicht nachgewiesen werden. Das im Medium nachweisbare Toxin könnte ebenso durch
Zelllyse freigesetzt worden sein. Schwieriger als bei den toxischen Sekundärmetaboliten fällt
es eine Funktion für die nicht-toxischen Peptide zu postulieren. Viele der nicht-toxischen
Peptide sind Hemmstoffe von Proteasen (Fastner et al., 2001). Protease-Inhibition kann
weitreichende Folgen für den Fraßfeind haben. Bei Insekten konnte beispielsweise ein

Diskussion 100
Aminosäuredefizit nach Aufnahme eines Proteasehemmstoffes beobachtet werden (Jongsma
und Bolter, 1997). Cyanopeptolin SS wirkt auf Primärkonsumenten von Cyanobakterien, wie
Daphnien toxisch (Jakobi et al., 1996). Die in dieser Arbeit getesteten Cyanopeptoline aus
Scytonema hofmanni PCC 7110 hatten keinen Effekt auf Daphnien. Jedoch verursachen die
isolierten Peptide ausgeprägte Hemmung von Serin-Proteasen. Dies könnte durchaus auch
Effekte bei den Primärkonsumenten verursachen wie sie von Agrawal et al. (2005)
beschrieben wurden: Ein Zellextrakt von Microcystis sp. PCC 7806 verursachte Hemmung
der Verdauungsenzyme aus dem Darm von Daphnien. Ein Beitrag zu einer möglichen
ökologischen Funktion der Peptide aus Scytonema hofmanni PCC 7110 wurde in dieser Arbeit
durch die Toxizitätstests von Cyanopeptolinen und Scytocyclamiden geleistet. So wurde eine
toxische Wirkung der Scytocyclamide auf Thamnocephalus platyurus, einem Fraßfeind in
Süßwasser, festgestellt. Daphnien wurden durch die isolierten Scytocyclamide A bzw. B nicht
geschädigt. Für Oscillapeptin J wurde ebenfalls eine toxische Wirkung auf Thamnocephalus
beschrieben (Blom et al., 2003). Oscillapeptin J besitzt die Charakteristika eines
Cyanopeptolins, und in der Aminosäuresequenz ist Arginin mit dem N-Terminus der Ahp-
Gruppe verknüpft, ein Strukturmerkmal was auch in dem neu charakterisierten Cyanopeptolin
1067A zu finden ist. Eine Hemmung ganz anderer Art wurde kürzlich von Becher et al.
(2005) beschrieben: das einen Biofilm bildende Cyanobakterium Fischerella sp. (ATCC
43239) produziert Sekundärstoffwechselprodukte, welche insektizid auf benthnische Insekten
Larven wirkt.
Die besprochene strukturelle Vielfalt der Metabolite aus Cyanobakterien, sowie ihre
verschiedenen Lebensräume bieten somit viel Platz für weitere Spekulationen über die
Funktion dieser Stoffwechselprodukte. Die gefundenen Bioaktivitäten sind aber auch immer
vom benutzten Aktivitätstest abhängig, so dass die eigentliche Funktion nicht unbedingt mit
den beschriebenen Bioaktivitäten übereinstimmen muss. Die in dieser Arbeit angewandten
Aktivitätstests geben einen kleinen Eindruck möglicher Bioaktivität wider. Bei den
beschriebenen Bioaktivitäten muss beachtet werden, dass die in vitro Versuche nur geringe
Rückschlüsse auf die Funktion oder Wirkung unter natürlichen Bedingungen zu lassen.
4.4. Therapeutische Relevanz peptidischer Sekundärmetabolite Wie in der Einleitung angedeutet haben besonders Peptide und Polyketide aus Bakterien und
Pilzen ein breites Wirkungsspektrum, das von antibiotischen über zytostatische bis zu
immunsuppressiven Eigenschaften reicht. Charakteristisch sind zyklische Strukturen, der

Diskussion 101
Einbau nicht-proteinogener Aminosäuren und Fettsäuren sowie postsynthetische
Modifikationen (Sieber und Marahiel, 2003). Die in der vorgelegten Arbeit isolierten
Scytocyclamide aus Scytonema hofmanni PCC 7110 besitzen Ähnlichkeit zu Laxaphycinen
aus Anabaena laxa (Frankmölle et al., 1992), jedoch wurden im Gegensatz zu diesen weder
bei den Scytocyclamiden noch beim Peptidgemisch der 3 Scytocyclamide antibiotische
Wirkung festgestellt. Aus Scytonema sp. TISTR 8208 wurde von einer ca. 1.400 Da großen
Verbindung berichtet, welche antibiotische Wirksamkeit vergleichbar mit Tetracyclin gegen
gram-positive Bakterien, pathogene Hefen und filamentöse Pilze besitzt (Chetsumon et al.,
1998). Es gibt weitere cyanobakterielle Peptide, welche eine „multidrug resistance-reversing
activity“ besitzen. Beispiele hierfür sind die Cryptophycine aus Nostoc Arten oder das
zyklische Hexapeptid Dendroamid A aus Stigonema dendroideum (Smith und Zhang, 1996;
Ogino et al., 1996).
Neben antibiotischer Wirksamkeit wird vermehrt nach zytostatischen Eigenschaften gesucht.
Einige der aus Cyanobakterien isolierten Substanzen oder Derivate haben Eingang in
klinische Prüfphasen erlangt. Darunter auch Dolastatin-10, welches sich in Phase II der
klinischen Prüfung befindet, ebenso die synthetischen Analoga Cemadotin und ILX-651,
(Haefner, 2003). Dolastatine wirken antimitotisch und greifen in die Tubulinpolymerisation
ein. Einen anderen bisher einzigartigen Wirkungsmechanismus hat Kahalalide F, ein
zyklisches Depsipeptid. Kahalalide F gehört zu einer Familie neuartiger
dehydroaminobuttersäure-haltiger Peptide und verändert die Funktion der lysosomalen
Zellmembran (Jimeno et al. 2004) und führt dadurch zum Zelltod. Im Gegensatz zu anderen
Zytostatika wird hier die Erbinformation der Tumorzellen oder deren Proteinsynthese nicht
beeinflusst. Das Peptid wurde aus der vor Hawaii lebenden Meeresschnecke Elysia rufescens
gewonnen, jedoch wird die Herkunft der Substanz einer Grünalge (Bryopsis sp.)
zugeschrieben (Hamann et al., 1993) Für viele aus marinen Invertebraten gewonnenen
Substanzen wird auch eine mikrobielle Herkunft vermutet, wie das auch für die Dolastatine
gilt (Haefner, 2003). Die zytotoxischen Laxaphycine und die in der vorliegenden Arbeit neu
charakterisierten Scytocyclamide weisen ebenfalls Dehydroaminobuttersäure als Baustein auf,
so dass diese Peptide Leitstrukturcharakter besitzen könnten. Ein aus Lyngbya majuscula
isoliertes zyklisches Depsipeptid Apratoxin A besitzt starke in vitro Zytotoxizität auf humane
Tumorzelllinien im Bereich von 0,36 bis 0,52 nM, dagegen nur geringe in vivo Aktivität
gegen Darmkrebs und Brustkrebs (Luesch et al., 2001). Tubulin ist ein Angriffspunkt für
Zytostatika. So werden Colchicin aus der Herbstzeitlosen (Colchicum autumnale) und
Paclitaxel aus Taxus brevifolia (bzw. partialsynthetisch aus Baccatin III aus Taxus baccata) in

Diskussion 102
der Pharmakotherapie eingesetzt. Neue Leitsubstanzen sind von großem Interesse, da die
bekannten antimicrotubulären Substanzen gute Substrate für Effluxpumpen sind, welche den
Wirkstoff wieder aus der Zelle transportieren und somit die Wirksamkeit verringern und zur
Resistenzentwicklung beitragen (Endicott et al., 1989). Für die ebenfalls in dieser Arbeit
beschriebenen Cyanopeptoline konnte keine zytotoxische Aktivität beschrieben werden. In
der Literatur gibt es jedoch Hinweise, dass Cyanopeptoline zytotoxisch sein können.
Beispielhaft seien die aus Symploca sp. isolierten Tasipeptine genannt, welche ihre Wirkung
bis zu einem IC50-Wert von 0,82 µM entfalten (Williams et al., 2003). Cyanopeptoline sind
häufig Inhibitoren von Proteasen. Die neu charakterisierte Strukturvariante in Cyanopeptolin
1067A besitzt mit einem IC50-Wert von 60 nM starke Hemmwirkung auf Trypsin. Des
Weiteren wurden durch die in Scytonema hofmanni PCC 7110 gefundenen Cyanopeptoline
1067A, bzw. 946A Plasmin und Elastase gehemmt (Abb. 30 und 31). Proteasen spielen eine
Schlüsselrolle bei pathologischen Prozessen. Fehlregulation von Proteasen spielt eine Rolle
bei z. B. rheumatoider Arthritis, kardiovaskulären Erkrankungen, bakteriellen oder viralen
Infektionen, Krebs oder der Alzheimer Krankheit. In allen Organen unseres Körpers besitzen
Proteasen essentielle Funktionen (Lendeckel und Hooper, 2004 und 2005). Die
kristallographischen Studien mit cyanobakteriellen Peptiden und ihren Zielenzymen können
dazu beitragen die Struktur-Wirkungsbeziehungen zu erkennen und ggf. Leitsubstanzen für
potentielle Pharmaka zu entwickeln oder zu modifizieren. Bei den Cyanopeptolinen wurde
Ko-Kristallisation von A90720A aus Microchaete lokatensis mit Trypsin und bei Scytonema
hofmanni von Scyptolin A mit Elastase erreicht (Lee et al.,1994; Matern et al., 2003), bei
Microcystinen LR mit der Proteinphosphatase 1 (Goldberg et al., 1995), bei Aeruginosin 98B
mit Trypsin (Sandler et al., 1998). Anhand der gewonnen Informationen versucht man
synthetische Inhibitoren der Proteasen zu entwickeln (Walker et al., 2001). Es bleibt
anzumerken, dass Peptid-Substrate als Arzneistoffe für die Hemmung von Proteasen nur
bedingt geeignet sind, da sie eine gute orale Bioverfügbarkeit ausschließen. Bei einer
parenteralen Applikationsform muss eine geeignete Formulierung gefunden werden, welche
ausreichende Bioverfügbarkeit gewährleistet. Eine mögliche Anwendung als Arzneistoff setzt
voraus, dass keine hämolytische Aktivität vom Wirkstoff ausgeht. Der in dieser Arbeit
durchgeführte Versuch zur Hämolyse (Kap. 3.2.7.) kann diese nicht gänzlich ausschließen;
Hämolyse hätte jedoch bei mikroskopischer Beobachtung anhand des Aufplatzens der
Erythrozyten und dem Austreten des Blutfarbstoffs beobachtet werden müssen.

Diskussion 103
4.5. Ausblick Die Ergebnisse der vorgelegten Arbeit lassen Spielraum für vielfältige Folgeuntersuchungen
und Ergänzungen. So war es im Rahmen dieser Arbeit nicht mehr möglich weitere
Untersuchungen zur Konfigurationsbestimmung durchzuführen. Dabei wäre vor allem das
zuletzt aufgeklärte Scytocyclamide B von besonderem Interesse, da hier eine identische
Aminosäuresequenz zu Laxaphycin D gefunden wurde. Auch wären molekulargenetische
Untersuchungen von Interesse, welche eine mögliche Verwandschaft von Scytonema
hofmanni zu Anabaena laxa, welche die Laxaphycine bilden, belegen könnten. Dies gilt
insbesondere für die NRPS-Gene.
Für die Laxaphycine wurde eine antifungale Wirkung beschrieben (Frankmölle et al., 1992),
welche bei den hier untersuchten Scytocyclamiden nicht belegt werden konnte. Jedoch ist zu
bedenken, dass diese Wirkung nur in Form eines Synergie-Effektes zu Stande kam und
Laxaphycin A als Reinsubstanz nicht antifungal wirksam ist. Es stellt sich nun die Frage, ob
auch Scytonema Peptide produziert, die antifungale Wirkung induzieren können.
In Bezug auf einen möglichen Mechanismus zur Zytotoxizität wurden viele Ansatzpunkte
durch die in dieser Arbeit durchgeführten Versuche gefunden, welche weiterer
Untersuchungen bedürfen und ein breites Feld für Folgeversuche bieten: Es wurden
Fluoreszenzfärbungen mit α-Tubulin und F-actin durchgeführt, so dass ergänzend geprüft
werden müsste, ob ß-Tubulin ebenfalls abgebaut wird. Des Weiteren bietet sich hier die
Möglichkeit Dosis-Wirkungs-Beziehungen näher zu untersuchen. Fluoreszenzfärbungen den
Zellkern betreffend wurden ebenfalls nicht vorgenommen. In Zusammenhang mit der
Tubulindepolymerisation ist auch ein möglicher Zusammenhang mit der Membranschädigung
zu sehen, da hohe intrazelluläre Ca2+-Konzentrationen zu Tubulinabbau führen können
(O´Brien et al. 1997). Auch wäre eine Beteiligung von Ca2+-Kanälen denkbar, welche in
diesem Zusammenhang untersucht werden müssten. Ebenso müssten die zur Apoptose
durchgeführten Versuche ergänzt werden, um eine Zelltoxizität über einen
Apoptosemechanismus zu bestätigen oder zu widerlegen. So müsste man beispielsweise die
Wirkung reaktiver Sauerstoffspezies durch die Zugabe von Antioxidantien oder
Radikalfängern mildern können.
Die Hemmung von Trypsin mit Cyanopeptolin 1067A im nmol-Bereich würde eine
Untersuchung der Hemmwirkung auf molekularer Ebene ermöglichen, so dass hier
Kristallisationsversuche angestrebt werden könnten. Dies würde einen Vergleich zu der
Wirkung von Scyptolin A auf die Serin-Protease Elastase zulassen (Matern et al., 2003).

Diskussion 104
Eine Fortführung der Toxizitätsversuche auf Thamnocephalus platyurus mit Cyanopeptolin
1067A wäre ebenfalls interessant, da die aus Scytonema hofmanni isolierten Cyanopeptolin
(Scyptolin A und B sowie Hofmannolin) keine Wirkung zeigten (Kap. 3.2.3.), Cyanopeptolin
1067A jedoch strukturelle Ähnlichkeit zu dem wirksamen Oscillapeptin J besitzt (Blom et al.,
2003). Auch könnte eine ökologische Funktion der Peptide durch Versuche zur Entwicklung
von Fischen und Amphibien (Oberemm et al., 1999) untersucht werden.

Literatur 105
Literatur Adams DG. (2000) Symbiotic interactions. In: The Ecology of Cyanobacteria. Whitton BA. Potts M. (eds.) Kluwer Academic Publishers, Netherlands : 523-61 Agrawal MK. Zitt A. Bagchi D. Weckesser J. Bagchi SN. von Elert E. (2005) Characterization of proteases in guts of Daphnia magna and their inhibition by Microcystis aeruginosa PCC 7806. Environmental Toxicology. 20(3): 314-22 An J. Carmichael WW. (1994) Use of a colorimetric protein phosphatase inhibition assay and enzyme linked immunosorbent assay for the study of microcystins and nodularins. Toxicon. 32(12): 1495-507 Anagnostidis K. Komarek J. (1990) Modern Approach to the Classification System of Cyanophytes 5. Stigonematales. Algological Studies. 86: 1-74 Baker PD Fabbro DL. (1999) A guide to the identification of common Blue-Green algae (Cyanoprokaryotes) in Australian freshwaters. CRCFE and MDBC Identification Guide No.25 (Edited by J. Sonnerman and J. H. Hawking. Cooperative Research Centre for Freshwater Ecology: Thurgoona, NSW) Barrett AJ. (1981) Cystatin, the egg white inhibitor of cysteine proteinases. Methods in. Enzymology 80: 771-7 Barth H. Pfeifer G. Hofmann F. Maier E. Benz R. Aktories K. (2001) Low pH-induced formation of ion channels by clostridium difficile toxin B in target cells. Journal of Biological Chemistry. 276(14): 10670-6 Bartram J. Chorus I. (1999) Toxic Cyanobacteria in water: A guide to their public health consequences, monitoring and management, E & FN Spon, London & New York Beattie KA. Kaya K. Sano t. Codd GA. (1998) Three dehydrobutyrine-containing microcystins from Nostoc. Phytochemistry (Oxford). 47(7): 1289-1292 Becher PG. Juettner F. (2005) Insecticidal compounds of the biofilm-forming cyanobacterium Fischerella sp. (ATCC 43239). Environmental Toxicology. 20(3): 363-72 Becker JE. Moore RE. Moore BS. (2004) Cloning, sequencing, and biochemical characterization of the nostocyclopeptide biosynthetic gene cluster: molecular basis for imine macrocyclization. Gene. 325: 35-42 Bell SG. Codd GA. (1996) Detection, analysis and risk assessment of cyanobacterial toxins, p. 109-122. In: Hester RE. Harrison RM. (eds.), Agricultural chemicals and the environment. Issues in Environmental Science and Technology no. 5. Royal Society of Chemistry, Cambridge, United Kingdom Benz R. Janko K. Boos W. Lauger P. (1978) Formation of large, ion-permeable membrane channels by the matrix protein (porin) of Escherichia coli. Biochimica et Biophysica Acta. 511(3): 305-19

Literatur 106
Bergmeyer HU. Gawehn K. Grassl M. (1974) In: Methods of Enzymatic Analysis (Bergmeyer, HU., ed) Volume I, 2nd ed., 436-437, Academic Press, Inc. New York, NY Bieth J. Frechin JC. (1974) Elastase inhibitors as impurities in commercial preparations of Kunitz soybean trypsin inhibitor. Biochimica et Biophysica Acta. 364(1): 97-102 Blom JF. Bister B. Bischoff D. Nicholson G. Jung G. Suessmuth RD. Juettner F. (2003) Oscillapeptin J, a new grazer toxin of the freshwater cyanobacterium Planktothrix rubescens. Journal of Natural Products. 66(3): 431-4 Boerner T. (2001) Neue bioaktive Verbindungen: die Toxine der Cyanobakterien. Biologie in unserer Zeit 31(2): 108-15 Bokesch HR. O'Keefe BR. McKee TC. Pannell LK. Patterson GM. Gardella RS. Sowder RC 2nd. Turpin J. Watson K. Buckheit RW Jr. Boyd MR. (2003) A potent novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Biochemistry. 42(9): 2578-84 Bold HC. Wynne MJ. (1985) Introduction to the Algae. Prentice-Hall, Inc., Englewood Cliffs, New Jersey Bonnard, I.; Rolland, M.; Francisco, C.; Banaigs, B. (1997) Total structure and biological properties of laxaphycins A and B, cyclic lipopeptides from the marine cyanobacterium Lyngbya majuscula. Letters in Peptide Science. 4(4-5-6): 289-292 Botha N. Gehringer MM. Downing TG. van de Venter M. Shephard EG. (2004) The role of microcystin-LR in the induction of apoptosis and oxidative stress in CaCo2 cells. Toxicon. 43(1): 85-92 Boyd JD. (1985) DCT board approves new cancer program, natural product concepts. Cancer Letters (11): 4-5 Brunner U. (2000) Ethnobotanik und Naturstoffscreening. Pharmazeutische Zeitung (37) Burja AM. Abou-Mansour E. Banaigs B. Payri C. Burgess JG. Wright PC. (2002) Culture of the marine cyanobacterium, Lyngbya majuscula (Oscillatoriaceae), for bioprocess intensified production of cyclic and linear lipopeptides. Journal of Microbiological Methods. 48(2-3): 207-19 Burja AM. Banaigs B. Abou-Mansour E. Burgess JG. Wright PC. (2001) Marine cyanobacteria: A prolific source of natural products. Tetrahedron. 57(46): 9347-77 Cardellina JH Marner FJ. Moore RE. (1979) Seaweed dermatitis: structure of lyngbyatoxin A. Science. 204(4389): 193-5 Carmeli S. Moore RE. Patterson GM. (1990) Tolytoxin and new scytophycins from three species of Scytonema. Journal of Natural Products. 53(6): 1533-42 Carmichael WW. Biggs DF. Peterson MA. (1979) Pharmacology of anatoxin-a, produced by the freshwater cyanophyte Anabaena flos-aquae NRC-44-1. Toxicon. 17(3): 229-36

Literatur 107
Carmichael WW. (1992) Cyanobacteria secondary metabolites-the cyanotoxins. Journal of Applied Bacteriology. 72(6): 445-59 Carter OC. Moore RE. Mynderse J. Niemczura WP. Todd JS. (1984) Structure of majusculamide C, a cyclic depsipeptide from Lyngbya majuscula. Journal of Organic Chemistry (49): 236-41. Centeno MD. (1995) Cyst-based toxicity tests: 9. The potential of Thamnocephalus platyurus as test species in comparison with Streptocephalus proboscideus (Crustacea, Branchiopoda, Anostraca). Environmental Toxicology and Water Quality 10(4): 275-82 Chang Z. Flatt P. Gerwick WH. Nguyen VA. Willis CL. Sherman DH. (2002) The barbamide biosynthetic gene cluster: a novel marine cyanobacterial system of mixed polyketide synthase (PKS)-non-ribosomal peptide synthetase (NRPS) origin involving an unusual trichloroleucyl starter unit. Gene. 296(1-2): 235-47 Chen BD. Nakeff A. Valeriote F. (1998) Cellular uptake of a novel cytotoxic agent, cryptophycin-52, by human THP-1 leukemia cells and H-125 lung tumor cells. International Journal of Cancer. 77(6): 869-73 Chen T. Wang Q. Cui J. Yang W. Shi Q. Hua Z. Ji J. Shen P. (2005) Induction of apoptosis in mouse liver by microcystin-LR: a combined transcriptomic, proteomic, and simulation strategy. Molecular & Cellular Proteomics. 4(7): 958-74 Chetsumon A. Fujieda K. Hirata K. Yagi K. Miura Y. (1993) Optimization of antibiotic production by the cyanobacterium Scytonema sp. TISTR 8208 immobilized on polyurethane foam. Journal of Applied Phycology. 5(6): 615-22 Chetsumon A. Maeda I. Umeda F. Yagi K. Miura Y. Mizoguchi T. (1994) Antibiotic production by the immobilized cyanobacterium, Scytonema sp. TISTR 8208, in a seaweed-type photobioreactor. Journal of Applied Phycology. 6(5-6): 539-43 Chetsumon A. Maeda I. Umeda F. Yagi K. Miura Y. Mizoguchi T. (1995) Continuous antibiotic production by an immobilized cyanobacterium in a seaweed-type bioreactor. Journal of Applied Phycology. 7(2): 135-9 Chetsumon A. Umeda F. Maeda I. Yagi K. Mizoguchi T. Miura Y. (1998) Broad spectrum and mode of action of an antibiotic produced by Scytonema sp. TISTR 8208 in a seaweed-type bioreactor. Applied Biochemistry & Biotechnology. 70-72: 249-56 Chmel H. Louria DB. (1980) Antifungal antibiotics: mechanism of action, resistance, susceptibility testing, and assays of activity in biological fluids. In: Antibiotics in Laboratory Medicine, Lorian V. (ed), Baltimore, Williams & Wilkins, 73-113 Chorus I. Bartram J. (1999) Toxic Cyanobacteria in water: A guide to their public health consequences, monitoring and management, E & FN Spon, London & New York Christiansen G. Dittmann E. Via Ordorika L. Rippka R. Herdman M. Boerner T. (2001) Nonribosomal peptide synthetase genes occur in most cyanobacterial genera as evidenced by their distribution in axenic strains of the PCC. Archives of Microbiology. 176(6):452-8

Literatur 108
Czarnecki O. HenningM. Lippert I. Welker M. (2005) Identification of peptide metabolites of Microcystis (Cyanobacteria) that inhibit trypsin-like activity in planktonic herbivorous Daphnia (Cladocera). Environmental Microbiology (0)0 Chen BD. Nakeff A. Valeriote F. (1998) Cellular uptake of a novel cytotoxic agent, cryptophycin-52, by human THP-1 leukemia cells and H-125 lung tumor cells. International Journal of Cancer. 77(6): 869-73 de Jonge MJA.van der Gaast A. Planting AST. van Doorn L. Lems A. Boot I. Wanders J. Satomi M. Verweij J. (2005) Phase I and Pharmacokinetic Study of the Dolastatin 10 Analogue TZT-1027, Given on Days 1 and 8 of a 3-Week Cycle in Patients with Advanced Solid Tumors. Clinical Cancer Research (11): 3806-13 DelMar EG, Largman C, Brodrick JW, Geokas MC. (1979) A sensitive new substrate for chymotrypsin. Analytical Biochemistry 99(2): 316-20. Demain AL.(2000) Microbial natural products: A past with a future. In: Biodiversity: New leads for the pharmaceutical and agrochemical industries. (Wrigley SK. Hayes MA. Thomas R. Chrystal EJT. Nicholson N. eds.) Royal Society of Chemistry (257): 3-16 Ding WX. Shen HM. Ong CN. (2001) Critical role of reactive oxygen species formation in microcystin-induced cytoskeleton disruption in primary cultured hepatocytes. Journal of Toxicology & Environmental Health Part A. 64(6): 507-19 Ding WX. Nam Ong C. (2003) Role of oxidative stress and mitochondrial changes in cyanobacteria-induced apoptosis and hepatotoxicity. FEMS Microbiology Letters. 220(1): 1-7 Dittmann E. Neilan BA. Boerner T. (2001) Molecular biology of peptide and polyketide biosynthesis in cyanobacteria. Applied Microbiology & Biotechnology. 57(4): 467-73 Dolowy, K. (2001) Environmental toxins and ion channels Cellular & Molecular Biology Letters. 6(2A). 343-347 Douglas S. Zauner S. Fraunholz M. Beaton M. Penny S. Deng LT. Wu X. Reith M. Cavalier-Smith T. Maier UG. (2001) The highly reduced genome of an enslaved algal nucleus. Nature. 410(6832): 1091-6 Draisci R. Ferretti E. Palleschi L. Marchiafava C. (2001) Identification of anatoxins in blue-green algae food supplements using liquid chromatography-tandem mass spectrometry. Food Additives & Contaminants. 18(6): 525-31 Duerfahrt T. Marahiel MA. (2005) Functional and structural basis for targeted modification of non-ribosomal peptide synthetases. Ernst Schering Research Foundation Workshop. (51): 79-106 Duerfahrt T. Marahiel MA. (2005) Peptidantibiotika vom molekularen Fließband. Nachrichten aus der Chemie 53: 507-13 Duy TN. Lam PK. Shaw GR. Connell DW. (2000) Toxicology and risk assessment of freshwater cyanobacterial (blue-green algal) toxins in water. Reviews of Environmental Contamination & Toxicology. 163: 113-85

Literatur 109
Endicott JA. Ling V. (1989) The biochemistry of P-glycoprotein-mediated multidrug resistance. [Review] [192 refs] Annual Review of Biochemistry. 58: 137-71 Elfant M. Bryant L. Starcher B. (1985) Isolation and characterization of a proteinase inhibitor from marama beans. Proceedings of the Society for Experimental Biology & Medicine. 180(2): 329-33 Erhard M. von Doehren H. Jungblut PR. (1999) Rapid identification of the new anabaenopeptin G from Planktothrix agardhii HUB 011 using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Communications in Mass Spectrometry. 13(5): 337-43 Erhard M. (1999) Matrix assisted laser desorption / ionisation time of flight Massenspektrometrie von Cyanobakterien und Untersuchungen zur Biosynthese und Funktion der Microcystine. Dissertation, Technische Universität Berlin Falch BS. Koenig GM. Wright AD. Sticher O. Angerhofer CK. Pezzuto JM. Bachmann H. (1995) Biological activities of cyanobacteria: evaluation of extracts and pure compounds. Planta Medica. 61(4): 321-8 Fastner J. Erhard M. von Doehren H. (2001) Determination of oligopeptide diversity within a natural population of Microcystis spp. (cyanobacteria) by typing single colonies by matrix-assisted laser desorption ionization-time of flight mass spectrometry. Applied & Environmental Microbiology. 67(11): 5069-76 Fay P. (1983) The blue-greens. In: The Institute of Biology's Studies in Biology 160, E. Arnold, London Forchert A. (1999) Isolierung und Charakterisierung neuer Peptidstrukturen aus Cyanobakterien der Gattung Planktothrix und Microcystis sp. Dissertation, Technische Universität Berlin Frankmoelle WP. Larsen LK. Caplan FR. Patterson GM. Knubel G. Levine IA. Moore RE. (1992) Antifungal cyclic peptides from the terrestrial blue-green alga Anabaena laxa. I. Isolation and biological properties. Journal of Antibiotics. 45(9): 1451-7 Frankmoelle WP. Knubel G. Moore RE. Patterson GM. (1992) Antifungal cyclic peptides from the terrestrial blue-green alga Anabaena laxa. II. Structures of laxaphycins A, B, D and E. Journal of Antibiotics. 45(9): 1458-66 Fritsche W. Laplace F. (2002) Mikrobiologie. Spektrum Akademischer Verlag, Heidelberg Gause GG. Brazhnikova MG. (1944) Gramicidin S and its use in the treatment of infected wounds. Nature 154: 703 Gbankoto A. Vigo J. Dramane K. Banaigs B. Aina E. Salmon JM. (2005) Cytotoxic effect of Laxaphycins A and B on human lymphoblastic cells (CCRF-CEM) using digitised videomicrofluorometry. In Vivo. 19(3): 577-82

Literatur 110
Gerwick WH. Mrozek C. Moghaddam MF. Agarwal SK. (1989) Novel cytotoxic peptides from the tropical marine cyanobacterium Hormothamnion enteromorphoides. 1. Discovery, isolation and initial chemical and biological characterization of the hormothamnins from wild and cultured material. Experientia. 45(2): 115-21 Gleason FK. (1991) Structure and activity of an herbicidal natural product isolated from the cyanobacterium Scytonema hofmanni In: Bioactive compounds from marine organisms: with emphasis on the Indian Ocean (Thomson MF. Sarojini R. Nagabhushanam R. eds.) Indo-United States Symposium, Goa, India XVII+410P. A. A. Balkema: Rotterdam, Netherlands. Illus Maps. 1991. 37-42 Goldberg J. Huang HB. Kwon YG. Greengard P. Nairn AC. Kuriyan J. (1995) Three-dimensional structure of the catalytic subunit of protein serine/threonine phosphatase-1. Nature. 376(6543): 745-53 Gottlieb E. Vander Heiden MG. Thompson CB. (2000) Bcl-xL prevents the initial decrease in mitochondrial membrane potential and subsequent reactive oxygen species production during tumor necrosis factor alpha-induced apoptosis. Molecular & Cellular Biology. 20(15): 5680-5689 Gregson JM. Chen JL. Patterson GML. Moore RE. (1992) Structures of puwainaphycins A-E. Tetrahedron. 48(18): 3727-3734 Gruenewald J. Kopp F. Mahlert C. Linne U. Sieber SA. Marahiel MA. (2005) Fluorescence resonance energy transfer as a probe of peptide cyclization catalyzed by nonribosomal thioesterase domains. Chemistry & Biology. 12(8): 873-81. Habig WB. Pabst MJ. Jakoby WB. (1974) Glutathione-S-transferases: The first enzymatic step in mercapturic acid formation. Journal of Biological Chemistry (249): 71309 Haefner B. (2003) Drugs from the deep: marine natural products as drug candidates. Drug Discovery Today. 8(12): 536-44 Hagmann L. Jüttner F. (1996): Fischerellin A, a novel photosystem-II-inhibiting allelochemical of the cyanobacterium Fischerella muscicola with antifungal and herbicidal activity. Tetrahedron Letter. (37): 6539-42 Hamann, Mark T; Scheuer, Paul J. (1993) Kahalalide F: A bioactive depsipeptide from the sacoglossan mollusk Elysia rufescens and the green alga Bryopsis sp. Journal of the American Chemical Society. 115(13). 5825-5826. Han B. Goeger D. Maier CS. Gerwick WH. (2005) The wewakpeptins, cyclic depsipeptides from a Papua New Guinea collection of the marine cyanobacterium Lyngbya semiplena. Journal of Organic Chemistry. 70(8): 3133-9 Harada KI. Fujii K. Shimada T. Suzuki M. Sano H. Adachi K. Carmichael WW. (1995) Two Cyclic Peptides, Anabaenopeptins, A Third Group of Bioactive Compounds from the Cyanobacterium Anabaena flos-aquae NRC 525-17. Tetrahedron Letters. 36(9): 1511-14 Hawes I. Smith R. (1994) Seasonal dynamics of epilithic periphyton in oligotrophic Lake Taupo, New Zealand. New Zealand Journal of Marine & Freshwater Research. 28(1): 1-12

Literatur 111
Helms GL. Moore RE. Niemczura WP. Patterson GML. (1988) Scytonemin A, a novel calcium antagonist from a blue-green alga. Journal of Organic Chemistry. 53: 1298-1307 Herfindal L. Oftedal L. Selheim F. Wahlsten M. Sivonen K. Doskeland SO. (2005) A high proportion of Baltic Sea benthic cyanobacterial isolates contain apoptogens able to induce rapid death of isolated rat hepatocytes. Toxicon. 46(3): 252-60 Hermann C. Giammasi C. Geyer A. Maier ME. (2001) Syntheses of hapalosin analogs by solid-phase assembly of acyclic precursors. Tetrahedron. 57(43): 8999-9010 Hirsch, CF. Liesch JM. Salvatore MJ. Schwartz RE. Sesin DF. (1990) Antifungal fermentation product and method. U.S. patent 4,946,835. Hoffmann D. Hevel JM. Moore RE. Moore BS. (2003) Sequence analysis and biochemical characterization of the nostopeptolide A biosynthetic gene cluster from Nostoc sp. GSV224. Gene. 311: 171-80 Holmquist B. Bunning P. Riordan JF. (1979) A continuous spectrophotometric assay for angiotensin converting enzyme. Analytical Biochemistry. 95(2):540-8 Honkanen RE. Caplan FR. Baker KK. Baldwin CL. Bobzin SC Bolis CM. Cabrera GM. Johnson LA. Jung JH. Larsen LK. Levine IA Moore RE. Nelson CS. Patterson GML Tschappat KD. Tuang GD. Boynton AL. Arment AR. An J. Carmichael WW. Rodland KD Magun BE. Lewin RA. (1995) Protein phosphatase activity in extracts of cultured blue-green algae (Cyanophyta). Journal of Phycology. 31(3): 478-86 Hooper NM. Lendeckel U. (Eds.) Series:Proteases in Biology and Disease. Vol. 1-4 (2004, 2005) Springer Verlag. Humble AV. Gadd GM. Codd GA. (1997) Binding of copper and zinc to three cyanobacterial microcystins quantified by differential pulse polarography Water Research. 31(7): 1679-86 Ingram L. (1970) A ribosomal mechanism for synthesis of peptides related to nisin. Biochimica et Biophysica Acta. 224(1):263-5 Ishibashi M. Moore RE. Patterson GML. Xu CF. Clardy J. (1986) Scytophycins, cytotoxic and antimycotic agents from the Cyanophyte Scytonema-Pseudohofmanni. Journal of Organic Chemistry 51: 5300–06 Ishida K. Murakami M. Matsuda H. Yamaguchi K.(1995) Micropeptin 90, a plasmin and trypsin inhibitor from the blue-green alga Microcystis aeruginosa (NIES-90). Tetrahedron Letters. 36(20): 3535-8 Ishitsuka M. Kusumi T. Kakisawa H. Kaya K. Watanabe MM. (1990) Microviridin a novel tricyclic depsipeptide from the toxic cyanobacterium Microcystis viridis. Journal of the American Chemical Society. 112(22): 8180-8182 Jaki B. Orjala J. Burgi HR. Sticher O. (1999) Biological screening of cyanobacteria for antimicrobial and molluscicidal activity, brine shrimp lethality, and cytotoxicity Pharmaceutical Biology. 37(2): 138-43

Literatur 112
Jaki B. Zerbe O. Heilmann J. Sticher O. (2001) Two novel cyclic peptides with antifungal activity from the cyanobacterium Tolypothrix byssoidea (EAWAG 195). Journal of Natural Products. 64(2): 154-8 Jakobi C. Oberer L. Quiquerez C. Koenig WA. Weckesser J. (1995) Cyanopeptolin S, a sulfate-containing depsipeptide from a water bloom of Microcystis sp. FEMS Microbiology Letters. 129(2-3): 129-33 Jakobi C. Rinehart KL. Neuber R. Mez K. Weckesser J. (1996) Cyanopeptolin SS, a disulphated depsipeptide from a water bloom: Structural elucidation and biological activities. Phycologia. 35(6 SUPPL.): 111-116 Jimeno J. Faircloth G. Fernández Sousa-Faro JM. Scheuer P. Rinehart K. (2004) New Marine Derived Anticancer Therapeutics - A Journey from the Sea to Clinical Trials. Marine Drugs 1, 14-29 Jongsma MA. Bolter C. (1997) The adaptation of insects to plant protease inhibitors. Journal of Insect Physiology. 43(10): 885-895 Kay RA. (1991) Microalgae as food and supplement. Critical Reviews in Food Science & Nutrition. 30(6): 555-73 Kiricsi M. Prenner EJ. Jelokhani-Niaraki M. Lewis RN. Hodges RS. McElhaney RN. (2002) The effects of ring-size analogs of the antimicrobial peptide gramicidin S on phospholipid bilayer model membranes and on the growth of Acholeplasma laidlawii B. European Journal of Biochemistry. 269(23): 5911-20 Kleinkauf H, Von Doehren H. (1996) A nonribosomal system of peptide biosynthesis. European Journal of Biochemistry 236(2): 335-51 Kobayashi M. Kurosu M. Ohyabu N. Wang W. Fujii S. Kitagawa I. (1994) The absolute stereostructure of arenastatin A, a potent cytotoxic depsipeptide from the Okinawan marine sponge Dysidea arenaria. Chemical & Pharmaceutical Bulletin (Tokyo) 42(10): 2196-98 Koehn FE. Longley RE. Reed JK. (1992) Microcolins A and B, new immunosuppressive peptides from the blue-green alga Lyngbya majuscula. Journal of Natural Products. 55(5): 613-9 Komarek J. Anagnostidis K. (1989) Modern Approach to the Classification System of Cyanophytes 4. Nostocales. Algological Studies. 82(3): 247-345 Kuehl M. Chen M. Ralph PJ. Schreiber U. Larkum AW. (2005) Ecology: a niche for cyanobacteria containing chlorophyll d. Nature. 433(7028): 820 Kuiper-Goodman T. Falconer I. Fitzgerald J. (1999) In: Chorus I. Bartram J. (1999) Toxic Cyanobacteria in water: A guide to their public health consequences, monitoring and management, Chapter 4: human health aspects. E & FN Spon, London & New York Kutschera U. Niklas KJ. (2005) Endosymbiosis, cell evolution, and speciation. Theory in Biosciences, 124(1): 1-24

Literatur 113
Lawton LA. Edwards C. Codd GA. (1994) Extraction and high-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst. 119(7): 1525-30 Le Dizet M. Piperno G. (1991) Detecion of acetylated alpha-tubulin by specific antibodies. Methods in Enzymology (196): 264-74 Lee AY. Smitka TA. Bonjouklian R. Clardy J. (1994) Atomic structure of the trypsin-A90720A complex: a unified approach to structure and function. Chemistry & Biology. 1(2): 113-7 Lendeckel U. Hooper NM. (Series Editors) Series:Proteases in Biology and Disease. Vol. 1-4 (2004, 2005) Springer Verlag. Lottenberg R. Christensen U. Jackson CM. Coleman PL. (1981) Assay of coagulation proteases using peptide chromogenic and fluorogenic substrates. Methods in Enzymology. 80: 341-61 Luesch H. Yoshida WY. Moore RE. Paul VJ. Corbett TH. (2001) Total structure determination of apratoxin A, a potent novel cytotoxin from the marine cyanobacterium Lyngbya majuscula. Journal of the American Chemical Society. 123(23): 5418-23 MacMillan JB. Ernst-Russell MA. de Ropp JS. Molinski TF. (2002) Lobocyclamides A-C, lipopeptides from a cryptic cyanobacterial mat containing Lyngbya confervoides. Journal of Organic Chemistry. 67(23): 8210-5 MacMillan JB. Molinski TF. (2002) Lobocyclamide B from Lyngbya confervoides. Configuration and asymmetric synthesis of beta-hydroxy-alpha-amino acids by (-)-sparteine-mediated aldol addition. Organic Letters. 4(11): 1883-6 Mahmood NA. Carmichael WW. (1986) Paralytic shellfish poisons produced by the freshwater cyanobacterium Aphanizomenon flos-aquae NH-5. Toxicon. 24(2): 175-86 Marfey P. (1984) Determination of D-amino acids using a bifunctional reagent 1,5-difluoro-2,4-dinitrobenzene. Carlsberg Research Communications (49): 591-96 Martin C. Oberer L. Ino T. Koenig WA. Busch M. Weckesser J. (1993) Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystis sp. PCC 7806. Journal of Antibiotics. 46(10): 1550-6 Martin W. Stoebe B. Goremykin V. Hapsmann S. Hasegawa M. Kowallik KV. (1997) Gene transfer to the nucleus and the evolution of chloroplasts. Nature. 393(6681): 162-5 Matern U. Oberer L. Falchetto RA. Erhard M. Koenig WA. Herdman M. Weckesser J. (2001) Scyptolin A and B, cyclic depsipeptides from axenic cultures of Scytonema hofmanni PCC 7110. Phytochemistry (Oxford). 58(7): 1087-95 Matern U. (2002) Zyklische Peptide aus Scytonema hofmanni bzw. Nostoc punctiforme: Struktur und biologische Aktivität. Dissertation, Albert-Ludwigs-Universität Freiburg

Literatur 114
Matern U. Oberer L. Erhard M. Herdman M. Weckesser J. (2003) Hofmannolin, a cyanopeptolin from Scytonema hofmanni PCC 7110. Phytochemistry. 64(6): 1061-7 Matsuda H. Okino T. Murakami M. Yamaguchi K. (1996) Aeruginosins 102-A and B, new thrombin inhibitors from the cyanobacterium Microcystis viridis (NIES-102). Tetrahedron. 52(46): 14501-14506 Meeks JC. (1998) Symbiosis between nitrogen-fixing cyanobacteria and plants Bioscience. 48(4): 266-76 Mian P. Heilmann J. Burgi HR. Sticher O. (2003) Biological screening of terrestrial and freshwater cyanobacteria for antimicrobial activity, brine shrimp lethality, and cytotoxicity. Pharmaceutical Biology. 41(4): 243-7 Milligan KE. Marquez BL. Williamson RT. Gerwick WH. (2000) Lyngbyabellin B, a toxic and antifungal secondary metabolite from the marine cyanobacterium Lyngbya majuscula. Journal of Natural Products. 63(10): 1440-3 Moore RE. Bornemann V. Niemczura WP. Gregson GM. Chen JL. Norton TR. Patterson GML. Helms GL.(1989) Puwainaphycin C a Cardioactive Cyclic Peptide from the Blue-Green Alga Anabaena BQ-16-1 Use of Two-Dimensional Carbon-13-Carbon-13 and Carbon-13-Nitrogen-15-Correlation Spectroscopy in Sequencing the Amino Acid Units. Journal of the American Chemical Society. 111(16): 6128-6132 Moore RE. Mynderse JS. (1982) Majusculamide C United States Patent 4342751 Moore RE. Ohtani I. Moore BS. de Koning CB. Yoshida WY. Runnegar MT. Carmichael WW. (1993) Cyanobacterial Toxins. Gazzetta Chimica Italiana (123): 329-36 Mootz HD. Schwarzer D. Marahiel MA. (2000) Construction of hybrid peptide synthetases by module and domain fusions. Proceedings of the National Academy of Sciences of the United States of America. 97(11): 5848-53 Moss GP (2005) Nomenclature of Cyclic Peptides. Chemistry international-The News Magazine of the international Union of Pure and Applied Chemistry (IUPAC). 27(2) Mullaney PF. Van Dilla MA. Coulter JR. Dean PN. (1969) Cell sizing: a light scattering photometer for rapid volume determination. Review of Scientific Instruments. 40(8): 1029-32 Mundt S. Teuscher E. (1988) Blue algae as a source of pharmacologically active compounds. Pharmazie. 43(12): 809-15 Murakami M. Okita Y. Matsuda H. Okino T. Yamaguchi K. (1994) Aeruginosin 298-A, a thrombin and trypsin inhibitor from the blue-green alga Microcystis aeruginosa (NIES-298) Tetrahedron Letters. 35(19): 3129-3132 Murakami M. Sun Q. Ishida K. Matsuda H. Okino T. Yamaguchi K. (1997) Microviridins, elastase inhibitors from the cyanobacterium Nostoc minutum (NIES-26). Phytochemistry (Oxford). 45(6): 1197-1202

Literatur 115
Nagle DG. Paul VJ. (1999) Production of secondary metabolites by filamentous tropical marine cyanobacteria: Ecological functions of the compounds. Journal of Phycology. 35(6 Suppl.): 1412-21 Nagle, DG.; Wedge, DE.; (2002); Antifungal properties of cyanobacteria and algae: Ecological and agricultural implications. In: Chemical ecology of plants: Allelopathy in aquatic and terrestrial ecosystems. Inderjit, M., Azim U. (Editors): 7-32. Nishiwaki-Matsushima R. Nishiwaki S. Ohta T. Yoshizawa S. Suganuma M. Harada K. Watanabe MF. Fujiki H. (1991) Structure-function relationships of microcystins, liver tumor promoters, in interaction with protein phosphatase. Japanese Journal of Cancer Research. 82(9): 993-6 Oberemm A. Becker J. Codd GA. Steinberg C. (1999) Effects of cyanobacterial toxins and aqueous crude extracts of cyanobacteria on the development of fish and amphibians Environmental Toxicology. 14(1): 77-88 O'Brien ET. Salmon ED. Erickson HP. (1997) How calcium causes microtubule depolymerization. Cell Motility & the Cytoskeleton. 36(2): 125-35 Ogino J. Moore RE. Patterson GM. Smith CD. (1996) Dendroamides, new cyclic hexapeptides from a blue-green alga. Multidrug-resistance reversing activity of dendroamide A. Journal of Natural Products. 59(6): 581-6 Okino T. Matsuda H. Murakami M. Yamaguchi K. (1993) Microginin, an angiotensin-converting enzyme inhibitor from the blue-green alga Microcystis aeruginosa. Tetrahedron Letters. 34(3): 501-4 Okino T. Sun Q. Matsuda H. Murakami M. Yamaguchi K. (1997) Nostopeptins A and B, elastase inhibitors from the cyanobacterium Nostoc minutum. Journal of Natural Products 60: 158-61 Oren A. (2004) A proposal for further integration of the cyanobacteria under the Bacteriological Code. International Journal of Systematic & Evolutionary Microbiology. 54(Pt 5): 1895-902 Osborne NJ. Webb PM. Shaw GR. (2001) The toxins of Lyngbya majuscula and their human and ecological health effects. Environment International. 27(5): 381-92 Ostensvik O. Skulberg OM. Underdal B. Hormazabal V. (1998) Antibacterial properties of extracts from selected planktonic freshwater cyanobacteria-a comparative study of bacterial bioassays. Journal of Applied Microbiology. 84(6): 1117-24 Patterson GML. Balwin CL. Bolis CM. Caplan FR. Karuso H. Larsen LK. Levine IA. Moore RE. Nelson CS. (1991) Antineoplastic activity of cultures Blue-Green Algae (Cyanophyta). Journal of Phycology. 27(4): 530-6. Patterson GML. Baker KK. Baldwin CL. Bolis CM. Caplan FR. Larsen LK. Levine IA. Moore RE. Nelson CS. Al E. (1993) Antiviral activity of cultured blue-green algae (Cyanophyta). Journal of Phycology. 29(1): 125-30

Literatur 116
Patterson GM. Smith CD. Kimura LH. Britton BA. Carmeli S. (1993) Action of tolytoxin on cell morphology, cytoskeletal organization, and actin polymerization. Cell Motility & the Cytoskeleton. 24(1): 39-48 Patterson GML. Larsen LK. Moore RE. (1994) Bioactive natural products from blue-green algae. Journal of Applied Phycology. 6(2): 151-7 Patterson GM. Bolis CM. (1994) Scytophycin production by axenic cultures of the cyanobacterium Scytonema ocellatum. Natural Toxins. 2(5): 280-5 Pawlik JR. (1993) Marine Invertebrate Chemical Defenses. Chemical Reviews, 93: 1911-22 Pennings, SC.; Pablo, SR.; Paul, VJ. (1997) Chemical defenses of the tropical, benthic marine cyanobacterium Hormothamnion enteromorphoides: Diverse consumers and synergisms Limnology & Oceanography. 42(5): 911-17 Pergament I. Carmeli S. (1994) Schizotrin A; a novel antimicrobial cyclic peptide from a cyanobacterium. Tetrahedron Letters. 35(45): 8473-76 Pettit GR. (1997) The dolastatins. Fortschritte der Chemie Organischer Naturstoffe. 70: 1-79 Pfleiderer G. (1970) Methods in Enzymology, XIX, 514-21 Piel J. (2004) Metabolites from symbiotic bacteria. Natural Product Reports. 21(4): 519-38 Ploutno A. Shoshan M. Carmeli S. (2002) Three novel protease inhibitors from a natural bloom of the cyanobacterium Microcystis aeruginosa. Journal of Natural Products. 65(7): 973-8 Proteau PJ. Gerwick WH. Garcia-Pichel F. Castenholz R. (1993) The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Experientia. 49(9):825-9 Radau G. (2000) Serine proteases inhibiting cyanopeptides. Pharmazie. 55(8):555-60 Ridley CP. Bergquist PR. Harper MK. Faulkner DJ. Hooper JN. Haygood MG. (2005) Speciation and biosynthetic variation in four dictyoceratid sponges and their cyanobacterial symbiont, Oscillatoria spongeliae. Chemistry & Biology. 12(3): 397-406 Rinehart KL. Holt TG. Fregeau NL. Keifer PA. Wilson GR. Perun TJ Jr. Sakai R. Thompson AG. Stroh JG. Shield LS. (1990) Bioactive compounds from aquatic and terrestrial sources. Journal of Natural Products. 53(4): 771-92 Rippka R. Deruelles J. Waterbury JB. Herdman M. Stanier RY. (1979). Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Journal of General Microbiology 111: 1-61 Rohrlack T. Christoffersen K. Kaebernick M. Neilan BA. (2004) Cyanobacterial protease inhibitor microviridin J causes a lethal molting disruption in Daphnia pulicaria. Applied & Environmental Microbiology. 70(8): 5047-50

Literatur 117
Rouhiainen L. Paulin L. Suomalainen S. Hyytiainen H. Buikema W. Haselkorn R. Sivonen K. (2000) Genes encoding synthetases of cyclic depsipeptides, anabaenopeptilides, in Anabaena strain 90. Molecular Microbiology. 37(1): 156-67 Sahl HG. Jack RW. Bierbaum G. (1995) Biosynthesis and biological activities of lantibiotics with unique post-translational modifications. European Journal of Biochemistry. 230(3): 827-53 Sandler B., Murakami M., Clardy, J. (1998) Atomic Structure of the Trypsin-Aeruginosin 98-B Complex. J. Am. Chem. Soc., 120 (3): 595-96 Schaefer AW. Kabir N. Forscher P. (2002) Filopodia and actin arcs guide the assembly and transport of two populations of microtubules with unique dynamic parameters in neuronal growth cones. Journal of Cell Biology. 158(1): 139-52 Schmidt EW. Nelson JT. Rasko DA. Sudek S. Eisen JA. Haygood MG. Ravel J. (2005) Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proceedings of the National Academy of Sciences of the United States of America. 102(20): 7315-20 Schuessler A. Kluge M. (2001) Geosiphon pyriforme, an endocytosymbiosis between fungus and cyanobacteria, and its meaning as a model system for arbuscular mycorrhizal research. In: Hock B. The Mycota: A comprehensive treatise on fungi as experimental systems for basic and applied research. Fungal associations. 151-161 Sergeeva E. Liaimer A. Bergman B. (2002) Evidence for production of the phytohormone indole-3-acetic acid by cyanobacteria. Planta. 215(2): 229-38 Sieber SA. Marahiel MA. (2003) Learning from nature's drug factories: nonribosomal synthesis of macrocyclic peptides. Journal of Bacteriology. 185(24): 7036-43 Sings HL. Rinehart KL. (1996) Compounds produced from potential tunicate-blue-green algal symbiosis: A review. Journal of Industrial Microbiology & Biotechnology. 17(5-6): 385-96 Skulberg, O.M., W.W. Carmichael, G.A. Codd and R. Skulberg (1993). Taxonomy of toxic cyanophyceae (cyanobacteria). In: Algal Toxins in Seafood and Drinking Water. I.R. Falconer, Edt. Academic Press, London. 145-164 Smith CD. Zhang X. (1996) Mechanism of action cryptophycin. Interaction with the Vinca alkaloid domain of tubulin. Journal of Biological Chemistry. 271(11): 6192-8 Stackebrandt E. (1985) Phylogeny and phylogenetic classification of prokaryotes. In: Schleifer KH. Stackebrandt E. (eds.) The evolution of prokaryotes. Academic press Inc. London 309-34 Tillett D. Dittmann E. Erhard M. von Doehren H. Boerner T. Neilan BA. (2000) Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: an integrated peptide-polyketide synthetase system. Chemistry & Biology. 7(10): 753-64 Ueno Y. Nagata S. (1994) Natural poisons: (1) Algae toxins. Journal of Toxicological Sciences. 19(3): App75-84

Literatur 118
Vaishampayan A. Sinha RP. Haeder DP. Dey T. Gupta AK. Bhan U. Rao AL. (2001) Cyanobacterial biofertilizers in rice agriculture. Botanical Review. 67(4): 453-516 Vaishampayan U. Glode M. Du W. Kraft A. Hudes G. Wright J. Hussain M. (2000) Phase II study of dolastatin-10 in patients with hormone-refractory metastatic prostate adenocarcinoma. Clinical Cancer Research. 6(11): 4205-8 Verdier-Pinard P. Lai JY. Yoo HD. Yu J. Marquez B. Nagle DG. Nambu M. White JD. Falck JR. Gerwick WH. Day BW. Hamel E. (1998) Structure-activity analysis of the interaction of curacin A, the potent colchicine site antimitotic agent, with tubulin and effects of analogs on the growth of MCF-7 breast cancer cells. Molecular Pharmacology. 53(1): 62-76 von Elert E. Oberer L. Merkel P. Huhn T. Blom JF (2005) Cyanopeptolin 954, a Chlorine-Containing Chymotrypsin Inhibitor of Microcystis aeruginosa NIVA Cya 43. Journal of Natural Products 68(9): 1324-27 Vining LC. Functions of secondary metabolites. (1990) Annual Review of Microbiology. 44: 395-427 Walker B. Lynas JF. Strategies for the inhibition of serine proteases. (2001) Cellular & Molecular Life Sciences. 58(4): 596-624 Watanabe M. Hirada K. Carmichael WW. Fujiki H. (Eds). (1996). Toxic Microcystis. Boca Raton, FL: CRC Press. Weckesser J. Martin C. Jakobi C. (1996) Cyanopeptolins, depsipeptides from cyanobacteria Systematic & Applied Microbiology. 19(2): 133-138 Williams DE. Burgoyne DL. Rettig SJ. Andersen RJ. Fathi-Afshar ZR. Allen TM. (1993) The isolation of majusculamide C from the sponge Ptilocaulis trachys collected in Enewetak and determination of the absolute configuration of the 2-methyl-3-aminopentanoic acid residue. Journal of Natural Products 56(4): 545-51 Williams PG. Yoshida WY. Moore RE. Paul VJ. (2002) Isolation and structure determination of obyanamide, a novel cytotoxic cyclic depsipeptide from the marine cyanobacterium Lyngbya confervoides. Journal of Natural Products. 65(1): 29-31 Williams PG. Yoshida WY. Moore RE. Paul VJ. (2003) Tasipeptins A and B: new cytotoxic depsipeptides from the marine cyanobacterium Symploca sp. Journal of Natural Products. 66(5): 620-4 Yoon HS. Golden JW. (1998) Heterocyst pattern formation controlled by a diffusible peptide. Science. 282(5390): 935-8 Zhang LH. Longley RE. Koehn FE. (1997) Antiproliferative and immunosuppressive properties of microcolin A, a marine-derived lipopeptide. Life Sciences. 60(10):751-62

Danksagung 119
Danksagung Die vorliegende Arbeit entstand am Institut für Biologie II in der Abteilung für Mikrobiologie, der Albert-Ludwigs-Universität Freiburg in der Arbeitsgruppe von Prof. Dr. J. Weckesser. Herrn Prof. Dr. J. Weckesser danke ich für die Überlassung des überaus interessanten Themas, die wissenschaftliche Betreuung dieser Arbeit und seine Unterstützung. Allen früheren und jetzigen Mitarbeitern im Labor „Weckesser“ danke ich für die Hilfsbereitschaft, Diskussionsbereitschaft…: Ute, Ximena, Manuel, Andreas und Karin. Ein spezieller Dank geht an Frau Silvia Radziwill für ihre stetige Hilfsbereitschaft und Unterstützung. Herrn Prof. Dr. G. Fuchs danke ich für die Aufnahme in das Graduiertenkolleg „Biochemie der Enzyme“, sowie für finanzielle Unterstützung und seine stetige Hilfsbereitschaft. Großer Dank gilt sämtlichen Kooperationspartnern, die zum Gelingen dieser Arbeit beigetragen haben: Lukas Oberer, Novartis Pharma AG, Basel: NMR-Analytik und Diksussionbereitschaft Dr. M. Erhard, AnagnosTec GmbH, Luckenwalde: MALDI-MS-Analysen Drs. G. Modjesch, F. Volk, P. Bissel, Pharmazeutisches Institut, Universität Freiburg: FT-IR-Messungen Prof. Dr. W. Haehnel, Dr. F. Drepper, Frau B. Knapp: Institut für Biologie II, Biochemie der Pflanzen, Universität Freiburg: MS-Workshop und LTQ-FTICR-MS-Messungen Prof. Jüttner, Frau Dr. J. Blom, Limnologische Station, Universität Zürich: LC-MS-, GC-MS-Messungen, Toxizitätsassays; Prof. Dr. M. Simon, Dr. J. Pardo, Max-Planck-Institut Freiburg: Apoptoseuntersuchungen Dr. Pelz, Frau A. Wittmer: Institut für medizinische Mikrobiologie und Hygiene, Universität Freiburg: Antimikrobielle Untersuchungen, Hämolyse Prof. Dr. R. Benz, Frau E. Maier: Biozentrum, Universität Würzburg: Untersuchungen an künstlichen Membranen Prof. Dr. K. Aktories, Frau Dr. G. Schmidt, Dr. J. Leemhuis, Dr. T. Giesemann, Institut für Pharmakologie, Universität Freiburg: Zelltoxizität, 86Rb+-Efflux, Immunhistochemische Untersuchungen, Mikroskopische Aufnahmen Hochschuldidaktikzentrum der Universitäten des Landes Baden-Württemberg: Fortbildungen Graduiertenkolleg „Biochemie der Enzyme“: Stipendium, Reisemittel, Fortbildung Allen nicht namentlich genannten Personen, die zum Gelingen dieser Arbeit beigetragen haben, danke ich für die entgegengebrachte Unterstützung (Werkstatt, Bibliothek, Labor „Fuchs“, Graduiertenkolleg,…)

Danksagung 120
Abschließend nochmals hervorheben und meinen besonderen Dank aussprechen möchte ich Prof. Dr. J. Weckesser und Prof. Dr. G. Fuchs, Lukas Oberer, Frau Dr. J. Blom, Frau Dr. G. Schmidt, Prof. Dr. M. Simon, Dr. J. Pardo, Dr. J. Leemhuis, Frau S. Radziwill und M. Kraft, die mich bestmöglichst unterstützt haben. Besonders danken möchte ich meinen Eltern, die mich immer unterstützt und meine Ausbildung gefördert haben sowie allen Freunden…und natürlich Tina für ihre Unterstützung.

Recommended

















