FACULDADE DE ENGENHARIA QUÍMICA DE LORENA DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
DETECÇÃO DE SIDERÓFOROS EM FUNGOS DECOMPOSITORES DE MADEIRA
Dissertação de mestrado apresentada como parte das exigências para a obtenção do título de Mestre em Biotecnologia Industrial
Banca examinadora:
Dr8 Adriane Maria Ferreira Milagres Dr8 Maricilda Palandi Mello Dr. André Luis Ferraz
Estudante: Tranferido da Biblioteca do DEBIQ para a Bilblioteca
Universitária em Junbo/2004 Proc. oº 202/04
Diovana Aparecida dos Santos Napoleão
Lorena-SP-Brasil 1999

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
DETECÇÃO DE SIDERÓFOROS EM FUNGOS DECOMPOSITORES DE MADEIRA
Este exemplar corresponde a versão final da dissertação de mestrado aprovada pela banca examinadora
M ria Ferreira Milagres Orientadora - Presi ente da Banca Examinadora
LORENA - SP - BRASIL 1999

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
DETECÇÃO DE SIDERÓFOROS EM FUNGOS DECOMPOSITORES DE MADEIRA
Este exemplar corresponde a versão final da dissertação de mestrado aprovada pela banca examinadora
e ria Ferreira Milagres Orientadora - Presidente da Banca Examinadora
Lorena-SP-Brasil 1999

Ao Agnaldo pelo carinho e incentivo em todos os momentos da minha vida.

ii
AGRADECIMENTOS
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP)
pelo apoio financeiro, possibilitando desta forma a realização deste trabalho.
À Prof Drª Maricilda Palandi Mello pela participação e sugestões na
avaliação deste trabalho.
À Prof Drª Adriane Maria Ferreira Milagres pela orientação e críticas
deste trabalho.
À Drª Angela Machuca pelas sugestões imprescindíveis no
desenvolvimento deste trabalho.
À Prof Drª Heizir de Castro Ferreira pelas críticas e sugestões que
objetivaram meu enriquecimento profissional e pessoal. Sou grata pelo carinho e
'·" amizade nos ensinamentos para o meu aprimoramento.
- --,..,
Ao Prof. Dr. André Luis Ferraz pela participação e valiosas discussões
que permitiram o desenvolvimento deste trabalho.
Ao amigo Djalma pelo auxílio e solicitude em todos os momentos da
realização deste trabalho.
À amiga Carmen Lúcia pelo convivío e auxílio prestado durante o
desenvolvimento deste trabalho.
À todos os integrantes do Departamento de Biotecnologia que
colaboraram com a realização deste trabalho.

iii
CONTEÚDO Página
1 - INTRODUÇÃ0 01
2 -REVISÃO BIBLIOGRÁFICA 04
2.1 - DEGRADAÇÃO BIOLÓGICA DA MADEIRA. 04
2.1.1 - FUNGOS DE DECOMPOSIÇÃO BRANDA (soft-rot) 06
2.1.2- FUNGOS DE DECOMPOSIÇÃO PARDA (brown-rot) 06
2.1.3- FUNGOS DE DECOMPOSIÇÃO BRANCA (white-rot) 07
2.2-SISTEMAS NÃO-ENZIMÁTICOS ENVOLVIDOS NA BIODEGRADAÇÃO DA
MADEIRA 08
2.3- SIDERÓFOROS 11
2.3. 1 - CONDIÇÕES DE CULTIVO PARA OBTENÇÃO DE SIDERÔFOROS 12
2.3.2- TIPOS ESTRUTURAIS DE SIDERÔFOROS 13
2.4 - MÉTODOS PARA DETECÇÃO DE SIDERÓFOROS 19
2.4. 1 - ENSAIO COM O REAGENTE CAS 19
2.4.2-ENSAIO DE CSÁKY 20
2.4.3- ENSAIO DE ARNOW 21
2.4.4- MÉTODO DE LOWRY 21
2.4.5- MÉTODO ELETROFORÉTIC0 22
3 - OBJÉTIVOS 23
4- MATERIAL E MÉTODOS 24
4.1 - MICRORGANISMOS UTILIZADOS 24

iv
4.2 - MEIO DE MANUTENÇÃO DOS FUNGOS 24
4.3 - DETECÇÃO DE SIDERÓFOROS EM MEIO SÓLID0 : 25
4.3.1 - MEDIDA DE CRESCIMENTO FÚNGICO E DA VELOCIDADE DA
REAÇÃO CASEM MEIO SÓLID0 27
4.4 - PRODUÇÃO DE SIDERÓFOROS EM MEIO LÍQUID0 28
4.5 - MÉTODOS QUÍMICOS 28
4. 5. 1 - MÉTODO CAS 28
4.5.2- MÉTODO DE ARNOW 29
4.5.3- MÉTODO DE CSÁKY 30
4.5.4- MÉTODO DE LOWRY MODIFICADO PARA DETECÇÃO DE ÁCIDOS
HIDROXÂMICOS 31
4.6 - TESTE DE ATIVIDADE OXIDATIVA. 31
4.7 - DETERMINAÇÃO DE PROTEÍNA. 32
4.8 - SEPARAÇÃO DOS SOBRENADANTES FÚNGICOS DE ACORDO COM A
MASSA MOLAR 32
4.9 - CROMATOGRAFIA EM SEPHADEX G-25 35
4.10 - CROMATOGRAFIA EM SEPHADEX G-10 35
i;.- 4.11 - DETERMINAÇÃO DE AÇÚCARES REDUTORES 36
4.12 - ESTUDO DOS INTERFERENTES DOS ENSAIOS DE DETERMINAÇÃO DE
SIDERÓFOROS 36
5 - RESULTADOS E DISCUSSÃ0 37
5.1 - DETECÇÃO DE SIDERÓFOROS EM MEIO SÓLID0 37
5.2 - PRODUÇÃO DE SIDERÓFOROS EM MEIO LÍQUID0 42

V
5.3 - CARACTERIZAÇÃO DOS CALDOS DE CULTURAS DE UM FUNGO DE
DECOMPOSIÇÃO PARDA E UM DE DECOMPOSIÇÃO BRANCA. 49
5.3.1- DETECÇÃO PELO MÉTODO CAS 49
5.3.2 - DETECÇÃO DE SIDERÔFOROS POR MÉTODO BASEADOS NA
ESTRUTURA DOS COMPOSTOS 53
5.4 - OBTENÇÃO E ANÁLISE DE UMA FRAÇÃO DE BAIXA MASSA MOLAR DOS
EXTRATOS DE P. COCOS E T. VERSICOLOR PARA DETECÇÃO DE
SIDERÓFOROS 54
5.5 - CROMATOGRAFIA EM SEPHADEX G-25 58
5.6 - CROMATOGRAFIA EM SEPHADEX G-10 60
6 - CONCLUSÕES 64
7 - REVISÃO BIBLIOGRÁFICA 66
ANEXO

vi
LISTAS DE TABELAS Página
Tabela 1 - Crescimento de fungos e detecção de sideróforos em placas preparadas
meio a meio ~··········································································39
Tabela 2 - Reação CAS e atividade oxidativa de fungos em meio líquido 47
••
Tabela 3- Determinação de sideróforos nos extratos de P. cocos 54.
.. -· 1"'-
Tabela 4 - Determinaçào de sideróforos nos extratos de T versicolor , : 56

vii
LISTA DE FIGURAS Página
••
Figura 1: Estrutura da parede da célula vegetal mostrando as diferentes camadas
(esquerda) e a distribuição dos diferentes componentes dentro da parede (direita).
ML: lamela média; P: parede primária; S1 e Si parede secundária; T: parede terciária;
W: lúmen da célula de madeira. 05
Figura 2: Distribuição das enzimas lignocelulolíticas e dos agentes de baixa massa
molar durante a degradação da matriz lignocelulósica da madeira por fungo de
decomposição branca. Os agentes de baixa massa molar, oxalato, radical catiônico do
álcool veratrílico (AV• +) e outra espécies radicalares, são secretados pela hifa fúngica,
localizada no lúmen da célula, e difundem através da parede celular (EVANS et ai.,
1994) 10
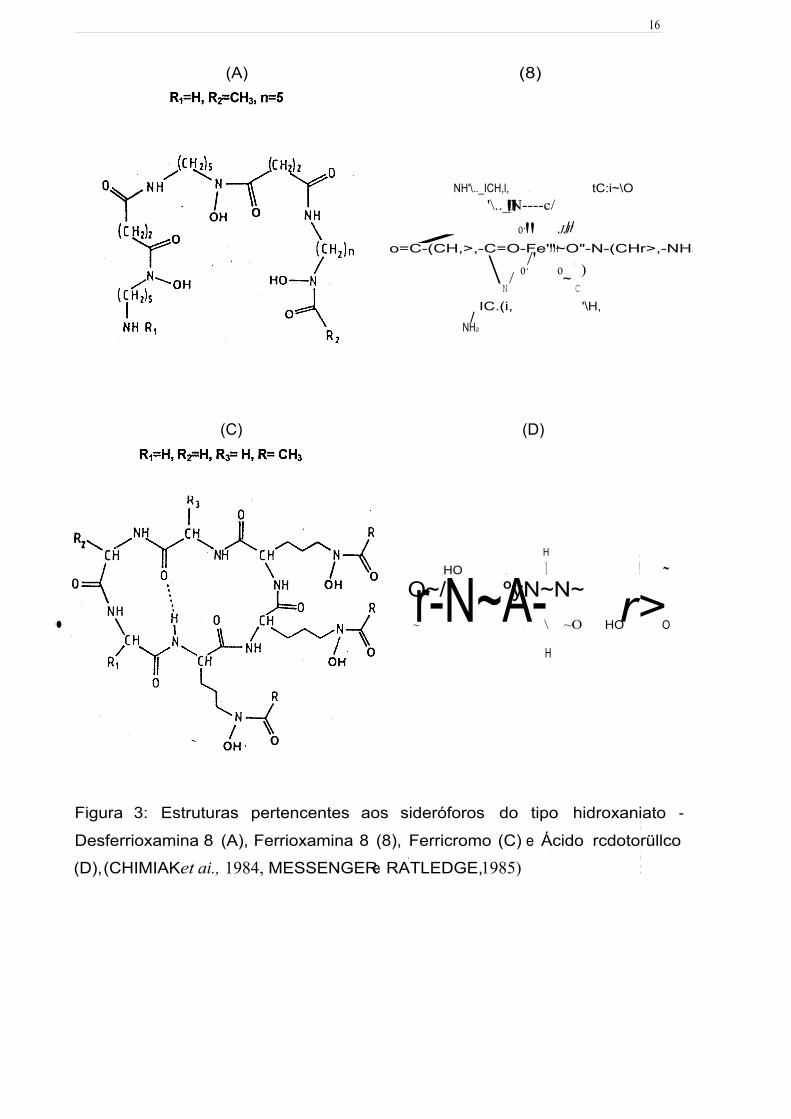
Figura 3: Estruturas pertencentes aos sideróforos do tipo hidroxamato -
Desferrioxamina B (A), Ferrioxamina B (B), Ferricromo (C) e Ácido rodotorúlico (D),
(CHIMIAKeta/., 1984, MESSENGER e RATLEDGE, 1985) 16
Figura 4: Estruturas pertencentes aos sideróforos do tipo catecolato .- Enterobactina (A),
Enteroquelina (B), Agrobactina A (C), Vibriobactina (D) (CHIMIAK et ai., 1984,
MESSENGER e RATLEDGE, 1985) 17
Figura 5: Estruturas pertencentes aos sideróforos do tipo misto, Pseudobactina (A),
Pioverdina (B), (CHIMIAKeta/., 1984) 18

viii
Figura 6: Etapas de preparo das placas pelo método de cobertura (A) e pelo método
meio a meio (8) : 25
Figura 7: Meio sólido contendo ágar-CAS/ ágar-ME inoculado com : A- P. pini,
8- P. chrysosporium, C~ T. versicolor, D- M. tremellosus, E- T. aurantiacus,
F- L. sulfureus 40
Figura 8: Cinética da reação CAS produzida por extratos extracelulares de
P. chrysosporium, P. medula-panis e P. coccineus 44
Figura 9: Cinética da reação CAS produzidos por extratos celulares de
P. cocos, L. su/fureus e T. aurantiacus 45
Figura 10: Cinética da reação CAS (-+-) e do pH (-•-) dos sobrenadantes de cultivo
do P. cocos , 50
Figura 11: Cinética da reação CAS (-+-) e do pH (-•-) dos sobrenadantes de cultivo
do T. versicolor 52
Figura 12: Cromatografia em Sephadex G-25 das frações do caldo extracelular do P.
cocos, caldo concentrado por liofilização (A), fração de baixa massa molar (PM < 5
KDa) (8). A coluna foi eluída com água deionizada, com um fluxo de 0,8 ml/min e
frações de 1,5 ml foram coletadas. A coluna foi calibrada com blue dextran PM =
~000 KDa (V0) = 1.5 ml, cobalamina PM = 1355,4 g/mol (V1) = 24 ml e riboflavina
PM = 376,37 g/mol (V2) = 39 ml. 59

ix
Figura 13: Cromatografia em Sephadex G-25 das frações· do caldo extracelular do
T. versicolor, caldo concentrado por liofilização (A), fração de baixa massa molar (PM
< 5 KDa) (8). A coluna foi eluída com água deionizada, com um fluxo de 0,8 mL/min
e frações de 1,5 ml foram coletadas. A coluna foi calibrada com blue dextran
PM = 2000 KDa (Vo) = 15 ml, cobalamina PM = 1355,4 g/mol (V1) = 24 ml e
riboflavina PM = 376,37 g/mol (V2) = 39 ml. 61
Figura 14: Cromatografia em Sephadex G-1 O das frações do caldo do P. cocos de
baixa massa molar (PM < 5 kDa). A coluna foi calibrada com cobalamina
PM = 1355,4 g/mol (V0 = V1) = 24 ml e riboflavina PM = 376,37 g/mol (V2) = 39
mL ; 63
Figura 15: Cromatografia em Sephadex G-1 O das frações do caldo do T. versicolor
de baixa massa molar (PM < 5 kDa). A coluna foi calibrada com cobalamina
PM = 1355,4 g/mol (Vo = V1) = 24 ml e riboflavina PM = 376,37 g/mol (V2) = 39
ml , 63

RESUMO
Detecção de sideróforos em fungos decompositores de madeira. Diovana A S. Napoleão. Dissertação de Mestrado. Programa de Pós-Graduação em
. Biotecnologia Industrial, Departamento de Biotecnologia, Faculdade de Engenharia Química de Lorena, Orientador: Drª Adriane Maria Ferreira Milagres (Departamento de Biotecnologia, FAENQUIL, CP 116, 12600-000, Lorena, SP, Brasil). Banca Examinadora: Drª Maricilda Palandi Mello e Dr. André Luís Ferraz. Abril de 1 999
•
Neste trabalho foi estudada a produção de sideróforos, em meio sólido e líquido, utilizando 2 cepas de decomposição parda (P. cocos e L. sulfureus}, 6 cepas de decomposição branca (P. coccineus, P. medula-panis, P. pini, M. tremellosus, P. chrysosporium, T. versicolor) e um ascomiceto, T. aurantíacus. Diversas metodologias foram empregadas na detecção desses compostos. complexantes de ferro. O cultivo em meio sólido foi realizado em placas contendo meio extrato de malte suplementado com o reagente CAS. Entretanto, alguns fungos cresceram pouco e outros não cresceram. Esse fato foi associado à presença do detergente HDTMA, altamente tóxico, utilizado na preparação deste meio. A fim de evitar a inibição do crescimento dos fungos no meio CAS, uma adaptação desse método foi proposta utilizando placas contendo metade ágar-extrato de malte e metade de ágar-CAS, sendo as cepas fúngicas inoculadas na metade contendo ágar-extrato de malte. Quando os fungos foram inoculados nesse meio de detecção de sideróforos, foi possível observar um bom crescimento de todas as cepas e também a mudança de cor do ágar-CAS, indicando a produção de sideróforos. Em meio líquido, P. cocos, L. sulfureus e T. aurantíacus produziram as maiores reações CAS. M. tremellosus e r. versicolor não reagiram ao CAS e os demais fungos produziram uma reação CAS intermediária, porém, não alteraram a cor da reação. Essa diferença entre as reações CAS observadas em meio sólido e líquido pode ser atribuída a diferentes fatores associados à concentração de sideróforos .. A partir desses resultados foram escolhidas 2 cepas fúngicas, uma pertencente à classe de decomposição parda (P. cocos) e a outra de decomposição branca ( T. versicolor) para obtenção de de uma fração de baixa massa molar dos extratos fúngicos e detecção de sideróforos. Nas culturas de P. cocos foi detectada a maior reação CAS e observou-se que o pH do meio reduziu de 5,5 para 2,8. A redução do pH foi atribuída à presença do ácido oxálico, o qual também reage com 'o CAS e intensifica a reação. T. versicolor não reagiu com o CAS, porém foi detectada a presença de compostos de baixa massa molar ( < 5 kDa) com atividade oxidativa em seus extratos extracelulares. A determinação e caracterização de compostos de baixa masa molar foi realizada através dos métodos CAS, Arnow, Csáky e· o método adaptado de Lowry. O extrato de P. cocos ( < 5 kDa) reagiu a todos os métodos de detecção de sideróforos sugerindo a presença de sideróforos dos tipos catecolato e hidroxamato. O ultrafiltrado de T. versicolor reagiu aos métodos de Csáky e Lowry, sugerindo uma estrutura de um composto do tipo hidroxamato.

ABSTRACT
The production of siderophore in solid and liquid media was studied, using two brown-rot strains (P. cocos and L. sulfureus), 6 white-rot strains (P. coccineus, P. medula-panis, P. pini, M. tremellosus, P. chrysosporium, T. versicolor) and one ascomycete T. auarantiacus. Several methodologies were employed to detect these iron chelant compounds. The cultivation in solid medium was performed in plates, with malt extract agar medium supplemented with CAS reagent. However, most fungi grew poorly or did not grow. This fact was associated to the presence of the detergente HDTMA, used in preparation of the CAS medium. To solve this problem, a modification of the CAS-agar plate assay was made by incorporating the CAS blue dye in a medium with no contact with the fungi tested. Half of each plate used in our experiments was filled with malte extract agar and the other half with CAS-blue-agar. When the fungi were inoculated in this medium it was possible to see a good growth of all strains and the calor change of agar-CAS, wich indicates production of siderophores. ln liquid medium, P. cocos, L. sulfureus and T. aúrantiacus produced the most intense CAS reaction. M. tremellosus and T. versicolor did not react to CAS, and the other fungi produced an intermediate CAS reaction, but did not alter the reaction colour. This difference between the CAS reactions observed in solid and liquid media can be attributed to different factors associated with the concentration of siderophores. Based on these results a choice of two fungai strains has been made, one belonging to the brown-rot class (P. cocos) and the other to the white-rot class (T. versicolor), in arder to obtain a low molecular weight fraction from fungai extracts and siderophore detection. P. cocos produced the highest CAS reaction simultaneously reducing the. pH of the malt extract medium from 5,5 to 2,8. The pH decrease was attributed to the presence of oxálic acid, wich could have reacted to CAS and intensified the reaction. T. versicolor did not react to CAS; however, the presence of low molecular weight compounds ( < 5 kDa) with oxidizing activity was detected in its extracellular extracts. The determination and characterization of low molecular weight compounds was performed employing CAS method, the methods of Arnow and Csáky and the adapted method of Lowry. P. cocos extract (< 5 kDa) reacted to all the methods for siderophore detection suggesting the presence of two types of siderophore: catechol and hidroxamate. T. versicolor ultrafiltrate reacted to Csáky's and Lowry's methods suggesting the structure of a compound of the hidroxamate type.

1 - INTRODUÇÃO
Nos últimos anos têm-se procurado esclarecer os mecanismos pelos
quais os fungos penetram e degradam os componentes da parede celular da madeira.
Numerosas enzimas oxidativas envolvidas nesse processo foram isoladas e
caracterizadas, porém, ainda permanece pouco conhecido o seu modo de atuação que
~m combinação a outros metabólitos processam a degradação da complexa matriz
lignocelulósica (JELLISON et ai., 1991, GOODELL et ai., 1997).
'--1 EVANS et ai. (1991 ), utilizando técnicas de imunocitoquímica e
microscopia eletrônica mostraram a localização das enzimas lignocelulolíticas
(celulases, lignina peroxidase, manganês peroxidase, lacase) em madeira degradada
por fungos de decomposição branca. Os autores postularam a hipótese de que
nenhuma destas enzimas iniciariam o processo de degradação da madeira, uma vez
que estas não foram encontradas na parede celular nos estágios iniciais da
decomposição. Uma das razões para isso é o limitado acesso das enzimas aos
respectivos substratos devido aos seus tamanhos que as impedem de penetrarem na
parede celular intacta. Assim, parece pouco provável que haja difusão das enzimas se
a parede não estiver parcialmente modificada (HIGHLEY, 1980, KOENIGS, 1974).
O processo de degradação de um substrato lignocelulósico como a
madeira ocorre pela ação combinada de uma variedade de sistemas enzimáticos e
alguns agentes não enzimáticos de baixa massa molar, recentemente descritos na
literatura, produzidos extracelularmente por fungos. Através destes sistemas, os fungos
degradariam as macromoléculas a componentes menores e solúveis, os quais
poderiam ser incorporados no metabolismo do microrganismo (ERIKSSON et ai.,
1990).

2
A degradação da lignina presente na madeira é um processo oxidativo,
onde participam diversas enzimas oxidativas dos tipos lacases e peroxidases com
geração de radicais livres. Dentre os fungos que participam na decomposição da
madeira os basidiomicetos são os principais responsáveis pela degradação da parede
celular (FEKETE et ai., 1989).
O ferro está presente como componente da estrutura das enzimas
portadoras do grupo heme, particularmente das peroxidases envolvidas na
decomposição branca das madeiras. Por outro lado, organismos classificados como de
decomposição parda necessitam do elemento Fe para degradação da celulose, que
• ocorre através da catálise do Sistema Fenton (Fe2+IH202). O ferro e outros metais, tal
como o manganês, são elementos essenciais para o metabolismo e crescimento dos
fungos. O ferro existe na natureza no estado de oxidação Fe(III), de forma insolúvel o
qual não é prontamente assimilável pelos microrganismos. Para solubilizar e sequestrar
o ferro e outros metais de transição, muitos microrganismos utilizam um sistema próprio
e eficiente de aquisição de metais. Compostos de baixa massa molar (500-1000 Da)
produzidos por uma variedade de microrganismos,. conhecidos como sideróforos, são
descritos na literatura como um dos sistemas mais bem compreendidos para quelar e
solubilizar o ferro.
Os sideróforos podem ser quimicamente classificados em 2 categorias
principais, hidroxamatos e catecolatos, A detecção de sideróforos em meios de cultura
pode ser realizada através de diferentes procedimentos, os quais incluem métodos
inespecíficos como o método CAS (SCHWYN e NEILANDS, 1987) e a reação com
cloreto de ferro (HOLZBERG e ARTIS, 1983). Métodos químicos específicos como o
teste de Csáky (CSÁKY, 1948) e o ensaio colorimétrico com o perclorato férrico
(ATKIN, 1968) são usados para detecção de estruturas de tipo hidroxamato, e o teste
de Arnow (ARNOW, 1937) para detecção de sideróforos do tipo catecolato.
Alguns estudos relacionam a produção desses complexantes com o
processo de degradação da madeira ou de outros substratos lignocelulósicos (DANIEL,
1994; JELLISON et ai., 1990; GOODELL et ai., 1995; GOODELL et ai., 1997).

Os sideróforos possuem capacidade de catalisar reações de óxido-redução,
dependendo do pH, devido à presença de metal na molécula (HIDER, 1984). Assim,
através da formação de complexos com Mn ou Fe poderiam participar de reações de • despolimerização da matriz lignocelulósica, permitindo o aumento do tamanho dos
poros.
Por analogia com outros sistemas, é provável que os fungos de
decomposição branca e parda produzam sideróforos em ambientes pobres em ferro,
porém, ainda é necessário verificar se há diferenças na estrutura ou no modo de ação
• entre os sideróforos produzidos por essas categorias de fungos. Foi dentro deste
contexto que os objetivos do presente trabalho centraram-se em detectar sideróforos na
fração de baixa massa molar ( < 5 KDa) de fungos de decomposição branca e parda,
visando sua caracterização química.
.. -· ~

2 - REVISÃO BIBLIOGRÁFICA
2.1 - Degradação biológica da madeira
A degradação biológica da madeira por ação de fungos envolve enzimas
específicas que são excretadas das células fúngicas e atacam os componentes da
parede celular das fibras da madeira.
Os mecanismos de colonização, invasão do lúmen das células, secreção
de enzimas que degradam a lignina e os polissacarídeos da madeira, são conhecidos,
em parte, devido a estudos por microscopia eletrônica (DANIEL et ai., 1989, BARRASA
et ai., 1992, DANIEL, 1994, SREBOTNIK e MESSNER, 1994). A degradação fúngica
da parede da célula vegetal, geralmente extende-se desde o lúmen da célula,
atravessando as diferentes camadas da parede até alcançar a lamela média. Um
esquema representativo da parede da célula vegetal é ilustrado na Figura 1.
Diversas classes ou espécies de fungos provocam diferentes tipos de
degradação na madeira. Tal fato faz com que sejam distinguidas três categorias de
fungos: Fungos de decomposição branda (soft-rot), decomposição parda (brown-rot) e
decomposição branca (white-rot).

5
umu,;iio DA
FIBRA ' rlBRILAS DE CELULOSE
'Ml\!Rll I IGNIIIA.HH11Cfl lll OSF
flFMICfllllOSF
Figura 1: Estrutura da parede da célula vegetal mostrando as diferentes camadas
(esquerda) e a distribuição dos diferentes componentes dentro da parede
(direita).
ML: lamela média; P: parede primária; S1 e S2: parede secundária; T: parede
terciária; W: lúmen da célula de madeira

(í
2.1.1 - Fungos de decomposição branda (soft-rot)
Encontra-se neste grupo espécies de fungos pertencentes às classes
Ascomycotina e Deuteromycotina, e o termo utilizado para classificá-los deve-se ao fato
deles produzirem um amolecimento nas camadas superficiais da parede da célula •
vegetal, formando cavidades típicas na camada S2 (KIRK e FARREL, 1987,
BLANCHETTE, 1991 ).
O termo decomposição branda foi primeiramente usado por SAVORY,
(1954) para caracterizar a aparência tenra da superfície da madeira degradada. Os
fungos de decomposição branda degradam, preferencialmente, os polissacarídeos da ••
madeira, porém, podem provocar algumas pequenas transformações na lignina. A
presença destes fungos é mais comum em madeiras duras do que em moles,
provavelmente pelas diferenças estruturais entre elas (ERIKSSON, 1981, FENGEL e
WEGENER, 1984 ).
2.1.2 - Fungos de decomposição parda (brown-rot)
Os fungos deste grupo pertencem à classe Basidiomycotina e degradam
principalmente os polissacarídeos da parede celular com limitada degradação e/ou
transformação da lignina (ERIKSSON et ai., 1990). Este grupo de fungos é considerado
colonizador da madeira, durante os estágios iniciais da decomposição.
Há evidências de que há produção por parte destes fungos de pequenos
agentes capazes de difundir pelas camadas S3 e S2, onde são observadas inicialmente
as mudanças morfológicas. Entre tais agentes não-enzimáticos destacam-se os
compostos de baixa massa molar, sideróforos, (JELLISON et ai., 1991 ), ácido oxálico

------ ------·-··- ·-----------····---· ·-·--- ... -- --··· 7
·--- --·-----------------·----------~
(SHIMADA et et., 1994), glicopeptídeos (TANAKA et ai., 1993, HIRANO et ai., 1995) e
radicais hidroxila (OH•) (WOOD, 1994). Os radicais hidroxila (OH•), provavelmente
produzidos .através da reação de Fenton, seriam os responsáveis pelas modificações
observadas na lignina. Devido a seu enorme poder oxidativo, os radicais OH• são
capazes de despolimerizar celulose e atacar unidades fenólicas e não-fenólicas de
lignina (BACKA et ai., 1992, WOOD, 1994).
2.1.3 - Fungos de decomposição branca (white-rot)
Os fungos causadores da decomposição branca degradam
extensivamente todos os componentes da madeira, inclusive lignina (ANDER e
ERIKSSON, 1977). Trata-se de um grupo heterogêneo de organismos pertencentes à
classe Basidiomycotina e de algumas espécies da classe Ascomycotina, que
apresentam em comum a capacidade de produzir enzimas que oxidam compostos
fenólicos, incluindo muitos relacionados com a liqnina (ERIKSSON e KIRK, 1985).
Dentro deste grupo diferenciam-se os fungos que degradam
. polissacarídeos e lignina com igual eficiência, daqueles que degradam
preferencialmente lignina (BLANCHETTE, 1991, BLANCHETTE et a/., 1992).
O fungo mais extensivamente estudado como representante deste grupo
é o basidiomiceto Phanerochaete chrysosporium, produtor de altos níveis de enzimas
ligninolíticas e eficiente na degradação de lignina em madeira.

8
2.2 - Sistemas não-enzimáticos envolvidos na biodegradação da madeira
Os estudos de microscopia eletrônica revelaram que o processo de
decomposição da parede celular pode ocorrer sem o contato direto da hifa com a
célula, sugerindo a participação de sistemas enzimáticos e/ou não-enzimáticos
extracelulares, embora o mecanismo através do qual isto ocorre não seja bem
compreendido. O transporte das enzimas lignocelulolíticas até o substrato (parede
celular) pode estar relacionado com a camada da glucana que rodeia a hifa fúngica e
que cria um microambiente favorável às enzimas e à ocorrência de diferentes reações
(EVANS et ai., 1991, RUEL et ai., 1994). Atualmente existe muita controvérsia se,
• devido ao seu tamanho, as enzimas lignocelulolíticas conseguem penetrar na parede
da célula vegetal para agirem diretamente sobre os diferentes componentes ou se a
decomposição é iniciada por agentes não-enzimáticos de baixa massa molar que
podem difundir facilmente através da parede (EVANS et ai., 1991, 1994, JOSELEAU et
ai., 1994).
Por outro lado, ainda não foi possível relacionar a perda dos
componentes da madeira, durante o ataque fúngico, com a bateria de enzimas
produzidas por um determinado fungo no decorrer do processo, sugerindo que outros
sistemas não enzimáticos devam estar envolvidos na degradação (DANIEL et ai., 1989;
EVANS et ai., 1991, 1994; JOSELEAU et ai., 1994; HAMMEL et ai., 1995). A estrutura
altamente organizada da matriz lignocelulósica na parede da célula vegetal, constitui
uma barreira impermeável a diversos microrganismos e seus sistemas enzimáticos,
enquanto se mostra facilmente permeável à água e a outros pequenos agentes, de
baixa massa molar (COWLING, 1975). Com base nestas informações, um crescente
interesse voltou-se para a teoria dos mediadores de baixa massa molar, os quais
poderiam penetrar nos poros da parede celular e iniciar a degradação da lignina ou
celulose longe da hifa fúngica (HARVEY et ai., 1986). Posteriormente, após os
componentes lignocelulósicos da parede ficarem mais expostos, o processo de
degradação seria continuado pela ação enzimática de celulases, hemicelulases e
fenoloxidases.

9
EVANS et ai., (1991, 1994), utilizaram técnicas de imunocitoquímica para
mostrar por microscopia eletrônica a localização das enzimas que degradam os
componentes da madeira durante os diferentes estágios do processo. Os resultados do
estudo mostraram que a lignina peroxidase (LiP) e a lacase não estabelecem contato
com a parede da célula a menos que o· processo de decomposição esteja relativamente
avançado. Da mesma forma, endoglucanases e celobiohidrolases somente se ligam às
fíbrilas de celulose se suas extremidades estiverem expostas.
Com base nesses resultados os autores postularam a hipótese de que
nenhuma das enzimas celulases, lacases ou LiP, iniciariam oprocesso de degradação
oo madeira, uma vez que estas não foram localizadas na parede celular nos estágios
iniciais de decomposição. Os iniciadores do processo provavelmente são moléculas
pequenas que podem difundir desde a hifa e penetrar através dos poros da matriz
lignocelulósica. Assim, é proposto que os radicais livres de H202 iniciariam a quebra da
celulose, e o radical catiônico do álcool veratrílico seria o responsável pelo início da
degradação'da lignina (Figura 2).

10
.i Ugniilue, I Cduwes • icosidne, ·-----~~~~~------·------ ~--~~~~------~ ..
AV·+
O:ubto
Figura 2: Distribuição das enzimas lignocelulolíticas e dos agentes de baixa
massa molar durante a degradação da matriz lignocelulósica da madeira por
fungo de decomposição branca. Os agentes de baixa massa molar, oxalato,
radical catiônico do. álcool veratrílico (AV• +) e outra espécies radicalares, são
secretados pela hifa fúngica, localizada no lúmen da célula, e difundem através
da parede celular (EVANS et ai., 1994)

11
. Todos esses antecedentes, e muitos outros existentes na literatura;
reforçam a hipótese de que a degradação da matriz lignocelulósica da madeira não é
resultado somente da atividade enzimática, principalmente nos estágios iniciais, mas
também o resultado da ação de diversos agentes não-enzimáticos, de baixa massa
molar, A degradação não enzimática é provavelmente de maior significância para os
fungos de decomposição parda, uma vez que estes provocam uma despolimerização
muito rápida e eficiente da matriz lignocelulósica, que em um estágio inicial atinge a
camada S2 da parede, degradando-a extensamente, enquanto a camada S3, adjacente
à hifa fúngica, permanece intacta. No decorrer do processo, os poros da matriz vão
aumentando lentamente de tamanho. Porém, mesmo assim o tamanho destes, tanto
em madeira intacta quanto degradada, não supera os 3,8 nm de diâmetro, o qual
permite a entrada de moléculas com uma massa molar menor que 6.000 Da
• (FLUORNOYeta/., 1991, SREBOTNIKe MESSNER, 1991).
Entre os agentes de baixa massa molar até agora identificados e
relacionados à degradação dos componentes da parede celular na madeira, por fungos
de decomposição branca e parda, estão: o oxalato, álcool veratrílico, manganês,
sideróforos e os glicopeptídeos.
2.3 - Sideróforos
Sideróforos são compostos de baixa massa molar, ligantes específicos de
íon ferríco (Fe3+), sintetizados geralmente por bactérias anaeróbias facultativas,
aeróbias e por fungos (CHIMIAK et et., 1984).
O íon ferroso (Fe2+), é um elemento essencial no crescimento da maioria
dos microrganismos, e existe na natureza predominantemente como íon ferríco Fe3\
. insolúvel e não disponível para a sua assimilação por parte dos microrganismos
secretam compostos de baixa massa molar (300 - 1000 Da), sob a forma de
sideróforos.

12
Para so,l~biliza~ e r~tirar o íon ferrí~o (Fe3+) e ou~ros meta_;s de transição, algu~s fungo~.
e bactenas sintetizam e produzidos como ligantes livres (forma desfem) sendo (
complexado com o ferro presente no meio, resultando na formação do complex:J
férrico. O complexo é então utilizado para transportar o ferro para o interior da célula. . .
Os sideróforos apresentam uma forte afinidade pelo Fe3+ e a constante de
estabilidade dos complexos férricos compreendem uma faixa de 1025 - 1 O~
(MESSENGER e RATLEDGE, 1985).
Grande parte dos estudos com sideróforos está relacionada aos
mecanismos de captação de Fe(III), de transporte até o interior da célula microbiana, e
de regulação molecular da produção, principalmente em bactérias e fungos patógenos
de plantas e animais, onde estes sideróforos representam o agente de patogenicidade
ou fator de virulência (PAYNE, 1'988, MARTÍNEZ et ai., 1990, WOOLDRIDGE e
WILLIANS, 1993).
·• 2.3.1 - Condições de cultivo para obtenção de sideróforos
Os fenômenos metabólicos e bioquímicos que determinam o início da
excreção de sideróforos pelos microrganismos ocorrem apenas sob condições
especiais. A condição essencial para a produção dos sideróforos por microrganismos é
um ambiente pobre em ferro. Muitos microrganismos somente excretam os
complexantes quando a concentração de ferro no ambiente é menor que 1,0 µM
(BAGG e NEILANDS, 1987). Segundo NEILANDS, (1984) uma concentração- de ferro
acima de 1 O µM no meio de cultivo produz um bom crescimento microbiano, porém, o
rendimento de sideróforos é baixo.
A fonte de carbono também influencia na produção de sideróforos, sendo
que os melhores substratos indutores da produção de sideróforos são aqueles
metabolizados unicamente por vias aeróbias, como por exemplo o succinato e o citrato.
A degradação desses compostos requer a presença de enzimas que contém ferro em

13
sua estrutura e como consequência verifica-se uma redução mais rápida do ferro
intracelular (NEILANDS, 1984). Essas informações coincidem com os resultados
descritos por SHAH et .ai., (1983) onde a produção de sideróforos por Azospíríllum
lípoferum foi favorecida quando o ácido málico, succínio ou glucônico foram usados
como fontes de carbono, sob uma alta aeração.
TSCHIERSKE et ai., (1996), otimizaram a produção de sideróforos pelo
Cunnínghamella elegans DSM 1908 e utilizaram como fonte de carbono sacarose e sais minerais variados. O rendimento de rizoferrina no meio de fermentação foi superior
a 4 g/L. A suplementação do meio com diferentes precursores levaram à formação de 9 •
tipos de rizoferrinas diferentes no meio de fermentação.
2.3.2 - Tipos Estruturais de Sideróforos
Estruturalmente os sideróforos, podem ser classificados em dois grandes
grupos: os do tipo catecolato e os hidroxamatos.
Os sideróforos pertencentes à classe dos hidroxamatos são derivados da
ornitina, é os grupos acil encontrados nestas estruturas são derivados de vários ácidos,
entre eles, os mais comuns são o ácido acético e o anidromevalônico (HELM et ai.,
1994) (Figura 3).
Os hidroxamatos em sua maioria apresentam em sua estrutura 3 grupos
de hidroxamatos secundários: R-N{O)-C(=O)-R, onde R representa um derivado de
aminoácido ou a própria estrutura do aminoácido. Cada grupo de hidroxamato contém
dois átomos de oxigênio formando um ligante bidentado com o ferro. Todavia, cada
sideróforo deste tipo forma um complexo hexadentado octaedro com o Fe3+
(MESSENGER e RATLEDGE, 1985).
Os sideróforos pertencentes à classe dos hidroxamatos quando
complexados com Fe3+ absorvem fortemente entre 425 e 500 nm, porém, essa faixa de
comprimento de onda pode ser variável em função das diferentes estruturas dos
/

14
compostos e das condições de pH e temperatura em que o ensaio é realizado. Os
trihidroxamatos produzem uma cor alaranjada em pH 2,0 - 3,0, contudo, em pH inferior,
a cor é modificada gradualmente, tornando-se ligeiramente púrpura, e posteriormente
incolor. Os monohidroxamatos-Fe, tais como ácido acetohidroxâmico ou
benzohidroxâmico, apesentam a cor púrpura a pH 5,0 (absorção máxima em
500-520 nm) e a pH neutro estes ácidos hidroxâmicos são avermelhados (absorção
máxima em 420-450 nm) (CHIMIAK et ai., 1984).
Os sideróforos pertencentes ao grupo dos catecolatos possuem em sua
estrutura dois átomos de oxigênio para quelar o ferro, formando um complexo
octaédrico, como no caso dos sideróforos pertencentes ao grupo dos hidroxamatos.
Esses compostos exibem uma fluorescência no ultravioleta semelhante às
características observadas do ácido 2,3 dihidroxibenzóico e absorvem nos
comprimentos de onda de 320, 250 e 210 nm (MESSENGER e RATLEDGE, 1985)
(Figura 4).
Os catecolatos férricos apresentam-se com uma coloração vermelha-
púrpura a pH neutro, porém quando o ferro complexado é dissociado a solução
apresenta-se incolor (PAYNE, 1994).
Os sideróforos do tipo catecolato, são detectados principalmente em
bactérias (HIDER, 1984). LEONG e NEILANDS, (1982), realizaram um estudo em
bactérias gram-negativas patógenas provenientes de plantas e demonstraram que a
maior parte das cepas da A. tumefacciens e da A. radiobacter investigadas produziram
sideróforos do tipo catecolato.
Por outro lado, JELLISON et ai., 1990 isolaram um composto com
características de catecol a partir da cultura líquida do fungo de decomposição parda,
Gloephillum trabeum. Um sideróforos do tipo catecol foi também detectado na cultura
do fungo Ganoderma applanatum, através do método de Arnow, o qual é específico
para catecóis (PARRA et ai., 1998).
Os sideróforos podem ainda apresentar ambas estruturas de hidroxamato
e· catecolato e, ainda, há alguns tipos de estruturas detectadas e isoladas que não
correspondem a nenhuma destas classes (Figura 5). Os compostos do tipo misto são
constituídos por ácidos amino ou imino carboxílicos (CHIMIAK et ai., 1_984).

15
Os complexos de Fe3+ destes tipos de compostos não apresentam
transferência de carga dos hidroxamatos e catecolatos férricos, consequentemente,
estes compostos apresentam-se incolores. Além disto, estes compostos exibem uma
baixa absorção no ultravioleta, tornando-se mais difícil a sua quantificação .
••
16
(A)
(C)
·•
(8)
/
NH'\.._ICH,l, . tC:i~\O '\.._N----c/ " . // o· JJ' " ,,, o=C-(CH,>,-C=O-Fe'"~O"-N-(CHr>,-NH.
\ /' . o· o ) / ~ . N C IC.(i, '\H,
/ NH2
(D)
. H ,
:HO . 1 ! ~ O~/ ºyN~N~ r-N~A- r> ~ . \ ~O HO O
H
Figura 3: Estruturas pertencentes aos sideróforos do tipo hidroxaniato - 1 1
Desferrioxamina 8 (A), Ferrioxamina 8 (8), Ferricromo (C) e Ácido rcdotorüllco • 1
• . 1
(D), (CHIMIAK et ai., 1984, MESSENGER e RATLEDGE, 1985) :

17
(A) (8)
H ---ci--- -:?º _.....--tH, C..........._ O N O
1 c=o :o HO
(C) X=OH
(D)
X ~ •
ó OH Q_.._ >-l_ He . j o NH T - OH
~~NH O
' i 1
Figura 4: Estruturas pertencentes aos sideróforos do tipo catecolato -
Enterobactina (A), Enteroquelina (8), Agrobactina A (C) e Vibriobactina (D) 1
(CHIMIAK et ai., 1984, MESSENGER e RATLEDGE, 1985)

18
(A)
HO~o Ltla O OH D-allo-Thr
NH / NH "-L ·Ala
o
(8)
o
NhN:XXX· .::;,- 1 OH HO o ~ "~~
. HN··@··N OH ~
~O O NHH
NH ~ . 1 / HO....,___ .>, L·Lys
D·i5er )yN H L-TC
L·Arg L·Th(
""' . D- Ser-NH o
Figura 5: Estruturas pertencentes aos sideróforos do tipo misto, Pseudobactina
(A), Pioverdina (8), (CHIMIAK et ai., 1984)

19
2.4 - Métodos para detecção de sideróforos
2.4.1 - Ensaio com o reagente CAS
•• Uma técnica rápida e simples para a detecção de sideróforos em cultivo
de microrganismo é o ensaio com o reagente CAS (Cromo-Azurol-S).
O reagente CAS é utilizado para detecção de sideróforo através de uma
reação independente da estrutura do composto. O reagente CAS não mede a
concentração total de sideróforos numa cultura, mas a quantidade de sideróforos não
complexados presentes em um determinado momento. A condição para uma reação
positiva com o reagente CAS é o sideróforo não estar complexado, caso contrário, o
sideróforo não tem capacidade para retirar o ferro do CAS. O princípio do método pode
ser explicado pela reação:
·---~=-~~~~ .. ~.·~.ru~.;;;;;;;;iiiiiiiii;;;;;;;;;;;;;;;;;;;~;;;.;;;;;;;;;;;.;;;;~ ... ;;..;;;:;;;iiiiiiiiiiiiiiiiiiiiiiiiõiiiiiiiiiiiiiiiii,;ijjl
Corante - Fe3-a. + Ouelante'" ~ · Fe .. Quelante + Corante
(Azul) (Laranja)
Quando um quelante forte (por exemplo, sideróforo) é adicionado ao
corante complexado com ferro, o quelante capta o ferro do corante, formando um novo
complexo. Consequentemente, há liberação do corante livre de ferro, sendo essa
reação acompanhada pela mudança de cor. O reagente CAS é o corante
normalmente utilizado para esse tipo de reação, por apresentar uma extrema

20
sensibilidade. Para uma completa solubilização do CAS, é recomendado o seu uso
juntamente com um outro composto, como por exemplo o detergente HDTMA (brometo
de hexa-deciltrimetilamônio ). Em solução, essa reação pode ser por medida
espectrofotométrica a 630 nm (SCH)/VYN e NEILANDS, 1987).
2.4.2 - Ensaio de Csáky
O ensaio de Csáky é um método utilizado para detectar agentes
complexantes de Fe(III), pertencentes à classe dos hidroxamatos. Os compostos desta
classe apresentam em suas estruturas ligações de hidroxilamina, que são detectadas
através .do método de Csáky (CHIMIAK et ai., 1984).
O princípio deste método, baseia-se na oxidação da hidroxilamina a
nitrito, em meio contendo ácido acético e uma solução de iodo. Estima-se a formação
de nitrito através da adição do ácido sulfanílico e a.-naftilamina, reação esta que pode
ser acompanhada pela formação de um complexo colorido (CSÁKY, 1948).
Este método possibilita estimar a presença de hidroxilamina no meio
reacional, porém, não de outros compostos nitrogenados oxidados, pois a presença
destes compostos degradariam a estrutura da hidroxilamina, parcial ou totalmente,
dependendo da concentração. A adição do ácido sulfanílico ao meio impede que parte
da hidroxilamina comece a ser degradada durante a reação.
A vantagem deste ensaio está relacionada com sua sensibilidade para
detectar sideróforos do tipo hidroxamato (PAYNE, 1994).

21
2.4.3 - Ensaio de Arnow
O ensaio de Arnow é utilizado para detectar sideróforos do tipo catecolato
presentes em sobrenadantes de culturas líquidas.
Este método determina quantitativamente a presença do composto 3,4
·• dihidroxi-fenilalanina nas estruturas de sideróforos pertencentes à classe dos
catecolatos. As estruturas que contenham somente um grupo OH fenólico reagem
fracamente, ou por muitas vezes não apresentam reação ao método de Arnow. Porém,
compostos que apresentam em suas estruturas dois ou três grupos fenólicos reagem
fortemente, resultando na formação de um complexo colorido estável no mínimo 1 hora
(ARNOW, 1937).
A vantagem deste método é a sua especificidade na detecção de
sideróforos do tipo salicilato e dihidroxibenzoato pertencentes · a classe dos
catecolatos.
2.4.4 - Método de Lowry
Os complexantes de ferro que contém estruturas do tipo hidroxamato
pode ser analisados pelo método de Lowry, através de reações de oxidação e redução.
O princípio deste método baseia-se na interação do ácido hidroxâmico
com o reagente de Folin, resultando na formação de um complexo azul, sob condições
alcalinas. Porém, alguns compostos de natureza protéica podem interferir na execução
desta metodologia. Estes interferentes podem ser completamente removidos através de
uma solução saturada de sulfato de amônia e uma posterior extração com alcool
benzílíco. O procedimento executado nesta etapa apresenta uma recuperação dos
compostos complexantes de ferro de aproximadamente 95 % (SUBRAMANIAN et ai.,
1965).

22
O método apresenta algumas vantagens, tais como a simplicidade da
técnica e a sensibilidade na análise dos resultados obtidos.
2.4.5 - Método Eletroforético
A eletroforese em papel baseia-se na separação de partículas carregadas
• quando sob a ação de um campo elétrico. É uma técnica utilizada para a separação de
misturas de compostos biológicos, sendo muito eficiente na separação de misturas de
aminoácidos e peptídeos, mas também é utilizada para a separação de sideróforos.
O uso do papel como suporte para realização da eletroforese é o mais
simples e usado na separação de uma vasta gama de produtos carregados, muito
embora n~o se obtenha em alguns casos uma boa separação dos compostos, com a
utilização de altas voltagens. A amostra é aplicada no papel cortado no tamanho
conveniente, que é colocado na cuba eletroforética e previamente saturado com a
solução de corrida, e após o tempo estipulado, o papel é retirado e seco.
Os sideróforos do tipo catecolato poderão ser detectados na fita de papel
por fluorescência à luz ultra-violeta e os do tipo hidroxamato poderão ser detectados
após pulverização do papel com uma solução de cloreto de ferro (FEKETE et ai., 1983).

23
3 - OBJETIVOS
O objetivo principal deste trabalho foi detectar a produção de sideróforos
em culturas de fungos degradadores de madeira e verificar se os síderóforos
produzidos por fungos de decomposição branca e parda são químicamente
semelhantes e se apresentam capacidade oxídatíva.
Durante a realização deste trabalho foram cumpridos os seguintes
objetivos específicos:
=> Seleção de fungos produtores de sideróforos em meio sólido utilizando o reagente
CAS,
=> Detecção da produção de sideróforos e de atividade oxidativa pelos diferentes
fungos em culturas líquidas,
=> Seleção de um fungo de decomposição branca· e um de decomposição parda e
caracterização dos respectivos extratos,
=> Caracterização dos extratos de um fungo de decomposição branca e de um fungo
de decomposição parda.

24
4- MATERIAL E MÉTODOS
• 4.1 - Microrganismos utilizados
Foram utilizados espécies de fungos da classe Basidiomycotina. Duas
espécies da classe de decomposição parda ("Brown-rot"): Poria cocos (ATCC 62778) e
Leetipoiús sulfureus (ATCC 52600), e seis da classe de decomposição branca ("White-
rot"): Pycnoporus coccíneus (ATCC 32258), Poria medula-panis (ATCC 42463),
Phellinus pini (ATCC 12240), Merulius tremellosus (ATCC 48745), e as cepas do
Phanerochaete chrysosporium, Trametes versicolor pertencentes à coleção de cultura
do DEBIQ. . -. ,..
Uma espécie de fungo pertencente à classe Ascomycotina, Thermoascus
aurantiacus ( cepa brasileira), também foi utilizada neste estudo.
4.2 - Meio de manutenção dos fungos
Os fungos foram mantidos e repicados semanalmente em tubos de
ensaio contendo ágar-extrato de malte, 2%. A incubação foi realizada a 28ºC, com
exceção do T aurantíacus que foi incubado a 45ºC, durante 8 dias, ou até o máximo
crescimento micelial.

25
4.3 - Detecção de sideróforos em 'meio sólido
Para a detecção dos sideróforos em meio sólido foram utilizadas duas
modificações da metodologia de SCHWYN e NEILANDS (1987), visando o melhor
• crescimento de todas as cepas fúngicas. Um esquema do preparo dos meios
modificados é mostrado na Figura 6. .
1 2 6i?
3
A
B
º · wasw
2 E§
3
Figura 6: Etapas de preparo das placas pelo método de cobertura (A) e pelo método meio a meio (8)

26
Etapas de preparo das placas pelo método de cobertura (A)
Derramou-se o conteúdo dos tubos contendo 10 ml de meio ágar-CAS
(azul) nas placas de Petri, quando o meio de cultura solidificou nas placas foi
derramado 15 ml do meio ágar-ME 2% (amarelo) sobre o meio ágar-GAS. Em seguida
os fungos foram inoculados no centro das placas.
Etapas de preparo das placas pelo método meio a meio (8)
Derramou-se o conteúdo dos tubos contendo 20 ml do meio ágar-ME 2%
nas placas de Petri, porém, quando o meio de cultura solidificou cortou-se metade do
meio ágar-ME 2% contido na placa e adicionou-se 10 ml do meio ágar-CAS na metade
da placa vazia. Os fungos foram inoculados nas placas meio a meio na metade que
continha o meio ágar-ME 2%.
Para o preparo do meio ágar-CAS, foram dissolvidos: 60,50 mg do
reagen~e cromo azurol S (CAS) em 50 ml de água desionizada e misturados com 10
ml de uma solução de Fe(III) (1,0 ml FeC'3.6 H20 em HCI 10 mM). Sob constante
agitação, este meio foi lentamente adicionado à solução de brometo hexadecil trimeti!
amônia (HDTMA) (72,9 mg/ 40 mLde água deionizada). Simultaneamente, foi
preparado o meio ágar Pipes contendo 750 ml de água deionizada, 15 g de ágar e
30,24 g de Pipes. O pH do meio foi ajustado para o pka do Pipes (6,8) com NaOH.
Ambos os meios foram autoclavados por 15 mina 121ºC/ 1 atm. Após o resfriamento
dos meios para aproximadamente 50ºC, a solução de CAS foi adicionada

27
cuidadosamente sobre o meio ágar-Pipes, com constante agitação, evitando-se desta
maneira a formação de espumá.
Os fungos foram cultivados em tubos de ensaio ou nas placas contendo o
meio de detecção de sideróforos, e a incubação foi conduzida a 28ºC e no caso do T
aurantiacus a 45ºC. A reação positiva para detecção de sideróforos foi visualizada com
a formação de um halo cor de rosa ou alaranjado em torno das colônias dos
respectivos fungos.
4.3.1 - Medida de crescimento fú,ngico e da velocidade da reação CAS em meio sólido
•
Placas de petri (10 cm de diâmetro) foram preparadas com 20 mi de ágar-
ME 2% e após solidificação, metade do meio foi retirado e substituído por ágar-CAS (10
mi). Os fungos foram inoculados com auxílio de uma alça na metade da placa contendo
ágar-ME 2%, o mais distante possível do limite entre os dois meios. As placas foram
incubadas em estufa, com temperatura controlada, durante três semanas, e o
crescimento das cepas foi monitorado diariamente e expressado como o número de
dias nec?ssários para o microrganismo cobrir metade da placa contendo o ágar-ME
2%,
A velocidade da reação CAS foi determinada medindo-se o avanço da
mudança de cor no ágar-CAS , a partir do limite entre os meios e expressada em
mm/dia. Placas controle, preparadas como descrito acima, mas sem inocular, foram .
incubadas nas mesmas condições.

28
4.4 - Produção de sideróforos em meio líquido
O cultivo dos fungos foi realizado em Erlenmeyers de 250 ml contendo
100 ml de extrato de malte, 2%. (Merck), por um período máximo de 30 dias. As
culturas foram incubadas sem agitação numa temperatura de 28ºC, exceto para o T
aurantiacus que foi cultivado a 45ºC. Amostras foram coletadas das culturas a cada 5
dias e centrífugadas antes de serem analisadas.
4.5 - Métodos químicos de detecção de sideróforos
4.5.1 - Método CAS
Para detecção da presença de sideróforos em cultura líquida foi
empregado o método universal de SCHWYN e NEILANDS (1987). Esse método
consiste na preparo de complexo indicador ternário contendo: O, 15 mM do reagente
CAS, 0,015 mM de uma solução de FeCb em HCI, 0,6 mM de HDTMA em solução . tampão de piperazina/ HCI pH 5,6. Todas as soluções aquosas foram preparadas
com água deionizada. O reagente CAS complexado com Fe(III) na solução possui uma
intensa cor azul com uma absortividade E = 100.000 M-1 cm" a 630 nm e pH 5,6. O
ensaio consistiu na adição de 0,6 ml da solução CAS para 0,6 ml do caldo de cultura
ou solução contendo sideróforo. A mistura foi incubada durante 1 hora à temperatura
ambiente e a leitura de absorbância foi determinada a 630 nm. Um controle foi
preparado utilizando-se uma alíquota do meio de cultura sem inocular.
As unidades percentuais de sideróforos foram definidas pela seguinte
fórmula (PAYNE, 1988):

29
Unidades % = (Ab - Aa) x 100
Ab
onde:
Ab = Absorbância do controle
Aa = Absorbância da amostra
4.5.2 - Método de Arnow
O método de Arnow foi utilizado para detecção de sideróforos do tipo
catecolato em sobrenadantes fúngicos (ARNOW, 1937). Esta metodologia consiste no
preparo das seguintes soluções: HCI 0,5 N, 1 O g de nitrito de sódio e 1 O g de molibdato
de sódio dissolvidos em 100 ml de água desionizada e NaOH 1 N.
Para 1 ml de sobrenadante ou a solução contendo sideróforo, adicionou-
se os reagentes na seguinte ordem: 1 ml de HCI, 1 ml da solução nitrito-molibdato
(o catecol neste ponto produz uma cor amarela), e 1 ml da NaOH (a cor muda para
vermelha). Esta cor é estável no mínimo 1 hora, e a absorbância foi medida a 51 O nm.
Para zerar o espectrofotõrnetro utilizou-se o meio de cultura não
inoculado, adicionado dos mesmos reagentes de Arnow. A concentração de sideróforos
nas culturas fúngicas foi determinada através de uma curva de calibração, preparada
com o ácido 2,3 dihidroxibenzóico {2,3 DHB) nas concentrações de 0,019; 0,038; 0,077;
0,23; 0,30; 0,38 e 0,46 mg/L, (Anexo 1 ).

30
• 4.5.3 - Método de Csáky
O ensaio de Csáky, foi utilizado para detecção de sideróforos do tipo
hidroxamato, e baseia-se na oxidação de hidroxilamina a nitrito, em meio ácido. O
nitrito formado foi estimado pela reação de cor com ácido sulfanílico e a-naftilatnina
(CSÁKY, 1948).
Para a utilização desse método 1 ml da amostra a- ser analisada foi
primeiramente hidrolisada com 1 ml de uma solução de H2S04 6 N, a uma temperatura
de 1 OOºC por 6 horas. Em seguida adicionou-se. 3 ml de uma solução de acetato de
sódio (35% p/v) e 1 ml de solução de ácido sulfanílico (1 g de ácido sulfanílico
dissolvido em 1 OOmL de uma solução aquecida de ácido acético 30% v/v) juntamente
com 0,5 ml da solução de iodo (1,3 g de iodo em 100 ml de ácido acético glacial).
Após 3-5 minutos, o excesso de iodo proveniente no meio reacional foi removido pela
adição de 1 ml da solução de arsenito de sódio (2% p/v). Adicionou-se posteriormente
1 rnl, da solução de a-naftilamina (3 g de a-naftilamina dissolvida em 1000 ml de ácido
acético, 30% v/v) e o volume foi completado para 1 O ml com água deionizada. Após
um período de 20 a 30 minutos, observou-se o aparecimento de cor púrpura,
· determinada a 526 nm.
O zero da absorbância do aparelho foi determinado através da leitura do
meio não inoculado adicionado dos mesmos reagentes de Csáky. Para determinar a
concentração de sideróforos nas culturas fúngicas foi feita uma curva de calibração,
preparada com cloreto de hidroxilamina nas concentrações de 0,34; 0,69; 1,38; 2,77;
4, 16; 5,55; 6,94 e 8,33 mg/L, (Anexo 2) .
••

31
4.5.4 - Método de Lowry modificado para detecção de ácidos hidroxâmicos
Ácidos hidroxâmicos, ferricromo e compostos relacionados interagem
com o reagente de Folin-Ciocalteau phenol produzindo um complexo azul sob
condições alcalinas que pode ser medido colorimetricamente, · sob as mesmas
condições descritas para estimar proteínas. Para o desenvolvimento dessa metodologia
foi utilizada uma solução contendo ·1 ml de sulfato de cobre 5 H20 1 %, 1 ml de
tartarato de sódio e potássio 4 H20 2%, 1 O ml de sulfato de dodecil e sódio 1 % e 88
ml carbonato de sódio em NaOH O, 1 N. A mistura da reação continha 2 ml dessa
solução e 200 µL do caldo fúngico. Após agitação a mistura foi mantida em repouso por
10 minutos, e em seguida adicionou-se 200 µL do reagente de Folin (diluído 1:1).
Novamente, a mistura foi agitada e após 30 minutos em repouso, a leitura de
absorbância foi realizadada a 660 nm. O espectrofotômetro foi zerado utilizando o meio
não inoculado seguido da adição dos mesmos reagentes do método de Lowry. A
concentração de sideróforos nas culturas fúngicas foi determinada utilizando uma curva
padrão construída com diferentes concentrações de desferal (desferrioxamina B-Ciba)
correspondentes a 0,016; 0,032; 0,065; 0,098; 0,131; O, 164; O, 196; 0,229 e 0,262 mg/L,
(Anexo 3).
4.6 -Teste da Atividade Oxidativa
A atividade oxidativa foi determinada utilizando-se dois substratos, o
,.. ABTS e a o-dianisidina. A oxidação do ABTS (E420 = 36.000 M-1cm-1) foi conduzida
. numa mistura de reação contendo 0,6 ml do caldo filtrado, 0,3 ml do tampão citrato-
fosfato 0,05 M (pH 3,0) e O, 1 ml dos respectivos substratos, num volume final de 1 ml.
A reação foi iniciada pela adição do ABTS e a velocidade de oxidação acompanhada
durante 1 O minutos a 420 nm (MACHUCA, 1995). Uma unidade de atividade oxidativa

32
· definida como a quantidade de enzima necessária para oxidar 1 µmol de substrato por
minuto e por ml de caldo de cultivo (UI/ mi).
A oxidação da o-dianisidina (t400 = 29.000 M-1cm-1) foi acompanhada por
uma reação similar à descrita para ABTS, apenas mudando o comprimento de onda
para 460 nm, (SZKLARZ et ai., 1989).
4.7 - Determinação de proteínas
O conteúdo de proteínas nas frações de baixa massa molar ( < 5 kDa) dos
fungos foi determinado de acordo com o método de BRADFORD, (1976). Esse método
consiste em adicionar 1 ml de amostra a 1 ml do reagente de Bradford, agitar e ler a
absorbância a. 595 nm. A concentração da amostra foi calculada pela curva de \
. calibração com albumina bovina, nas concentrações entre 2 e 4 µg/ ml.
4.8 - Separação dos sobrenadantes fúngicos de acordo com a massa molar
Os caldos de · cultivo de 30 dias P. cocos e T. versicolor foram
concentrados . 1 O vezes por liofilização e posteriormente transferidos para tubos de
· contendo uma membrana de corte 5 kDa (Ultrafree - 15 centrifugai Biomax - 3 KNMWL
- Millipore). Os caldos liofilizados foram centrifugados, (Centrífuga Jouan - Modelo MR
1812) por 30 minutos, a 25ºG e 2400 x g.
Nos caldos liofilizados e nas frações de baixa massa molar obtidas das
ultrafiltrações, realizaram-se as análises de detecção de sideróforos por. métodos
químicos específicos, não específicos. e de açúcares redutores, conforme o esquema 1
apresentado a seguir:

33
ESQUEMA 1
Cultura fúngica : (Extrato de malte 2 ºlo, 30 dias de
incubação)
Extrato Extracelular 1 ·· .. --- ----------------------------------------------- --------------··----.------------------------------------···········1
I Atividade Oxidativa I !
Liofilização
í"·~, .. ., .. _._ .. _ ....... -.E~~~-i~~ --d~- -- -·---~l : . 1
1 Arnow, Csáky, Lowry, 1 . ; 1
! Reação CAS - i Atividade Oxidativa I .L. !
[-·---.---·----····- - .. ·-· u1t~fil~~ção - 1
. 1
_ (Membranade __ corte_S_KDa)J
Fração de - , PM>5KDa 1 (Fraçãoll) [_ _-_ --. --- ------------------------------------------ ... ----------------------------------------------------
--- --·- · ·.·.~·.":.'~.)
'Fração de PM<SKDa (Fração 1)
_____ .,...,,...,...,.,_, ,,,.- - -.·-.·. ···.·--ii
Ens .. aios r : Arnow, Csaky, Lowry,, : Açúcares, Reação CASI I
Atividade Oxidativa I . . .. . ' .

,34
4.9- Cromatografia em Sephadex G-25
Os extratos de baixa massa molar (200 µL) foram aplicadas à coluna de
Sephadex G-25.
A coluna foi previamente calibrada com as seguintes substâncias de
massa molar conhecida: Riboflavina (376,36 g/ mol), Cobalamina (1355,4 gl mol) e Azul
de dextrana (2000 kDa). Nas frações eluídas da coluria foram determinadas a absorção
à 280 nm e aquelas frações que apresentaram mais do que 0,2 unidades de
absorbância analisou-se a reação CAS, atividade oxidativa e açúcares redutores.
As frações correspondentes aos picos foram coletadas, reunidas e
liofilizadas e novamente realizadas as análises de CAS, atividade oxidativa e açúcares
redutores ( esquema 2).
4.1 O - Cromatografia em Sephadex G-1 O
•• Os ultrafiltrados de baixa massa molar ( < 5 kDa) de P. cocos e
T versicolor foram também aplicadas a uma coluna de filtração em gel Sephadex G-1 O.
Amostras (200 µL) foram aplicadas a coluna de Sephadex G-1 O
previamente equilibrada com substâncias de massa molar conhecida: Riboflavina
(376,36 g/ mol), Cobalamina (1355,4 g/ mol) e Azul de dextrana (2000 kDa).
· As frações· eluldas da coluna foram determinadas a 280 nm e aquelas 1
frações que apresentaram mais do que 0,2 unidades de absorbância analísou-se a a
reação CAS, atividade oxidativa e açúcares redutores.
35
ESQUEMA2
1 Extrato Extracelular .1
L :: Liofilização ]
U ltrafiltração (membrana de corte 5 KDa)
Fração de> Massa molar (Fração li)
Fração < Massa molar (Fração 1)
1 . Sephadex G-25
1 .
1 Liofilização
1
Reação CAS Atividade oxidativa Açúcares redutores

36
4.11 - Determinação de açúcares redutores
Foi feita a detecção de açúcares redutores de forma qualitativa, através
do método do ácido 3,5 - dinitrosalidlico (DNS) (MILLER, 1959). O procedimento
consistiu na adição de 0,5 mL das amostras de P. cocos e T. versicolor (PM < 5 kDa)
ou aquelas obtidas das colunas de filtração em gel em 3 mL de DNS. Em seguida esta .
solução foi aquecida em banho de água fervente por 5 minutos e· após resfriar,
adicionou-se 16,5 mL de água destilada nos tubos de ensaio com as respectivas
amostras.
As leituras de absorbância feitas a 540 nm, contra um branco, preparado
com água deionizada da mesma forma que foi feito para os extratos fúngicos.
4.12 - Estudo de Interferentes dos ensaios de determinação de sideróforos ·
'-. - Realizou-se o estudo do efeito de diferentes substâncias como
1nterferentes das reações de determinação de sideróforos. Para tal estudo, foram
· utilizados o ácido oxálico e a proteína soro albumina bovina (BSA).
Inicialmente analisou-se o efeito do ácido oxálico (3,9; 15,8; 35,5; 63,0;
98,5; 141,8 e 393,9 mg/L) na reação CAS. Após 1 hora e 6 horas da adição do
reagente CAS foram feitas leituras de absorbància a 630 nm . .. A partir das concentrações do ácido oxálico (1,2; 3,2; 6,3 mg/L) e o BSA
(0,4; 2 mg/L) foram realizados os testes referentes aos métodos de Arnow, Csáky e
Lowry para verificar a interferência no meio.

37
5. RESULTADOS E DISCUSSÃO
A produção de sideróforos por fungos decompositores de madeira foi
estudada em meio sólido e líquido. Diversas metodologias foram empregadas na
detecção destes compostos complexantes de ferro. Algumas metodologias se
baseiam na capacidade cornplexante dos sideróforos e outras na natureza estrutural
dos compostos.
5.1 - Detecção de sideróforos em meio sólido
•• Para estudar a produção de sideróforos em meio sólido a técnica de
trabalho mais utilizada é o cultivo de microrganismos em meio de cultura
suplementado com o reagente CAS (SCHWYN e NEILANDS, 1987):
Entretanto, estudos publicados revelaram um crescimento pobre por . parte de alguns microrganismos e a ausência completa de crescimento por parte de
outros no meio CAS (SCHWYN e NEILANDS, 1987). Este fato foi associf1dO à
presença' do detergente HDTMA, altamente tóxico, utilizado na preparação deste
meio. A fim de evitar a inibição do crescimento dos microrganismos no · meio
contendo o reagente CAS,. duas adaptações desse método foram propostas neste
trabalho.
Na primeira adaptação, o meio ágar-CAS foi recoberto por uma camada
de ágar-extrato de malte, 2% (ágar-ME), e das nove cepas de fungos repicadas em
tubos e em. placas, apenas P. coccineus, P. chrysosporium, P. pin! e T. versicolor

38
apresentaram crescimento micelial e a formação do halo cor de rosa em torno das
respectivas colônias após 15 dias de incubação. As outras espécies, P. cocos, L.
sulfureus, P. medula-panis, M. tremellosus e T. aurantiacus, ainda nestas condições,
apresentaram uma maior sensibilidade ao meio ágar-CAS, demonstrada pela
ausência de crescimento dessas espécies.
Apesar do ágar-CAS ser recoberto por uma camada de ágar-ME, pode
ter ocorrido a difusão de substâncias para a superfície do meio provocando a inibição
do crescimento de alguns fungos.
Em razão disso, foi proposta uma segunda adaptação da metodologia
original utilizando placas contendo metade de ágar-ME e metade de ágar-CAS,
sendo que . as cepas fúngicas foram inoculadas na metade contendo ágar-ME.
Quando os fungos foram inoculados nesses meios para detecção de sideróforos,
foram determinadas a velocidade de crescimento fúngico, a cor da reação CAS e a
velocidade de mudança de cor no meio ágar-CAS. Através desta adaptação foi
possível observar um bom crescimento de todas as espécies em estudo, sem
problemas de inibição (Tabela 1 ). Dependendo do fungo, o crescimento micelial foi
muito rápido e somente se deteve quando alcançou o limite entre os dois meios, na
metade da placa. Algumas placas foram escolhidas para fotografar e estão
mostradas na Figura 7.
A maioria dos fungos produziram mudança de cor no meio após ter
colonizado toda a metade da placa com o ágar-ME. Entretanto, P. chrysosporium e
T. versicolor alteraram a cor do ágar-CAS simultaneamente ao crescimento.
Todos os basidiomicetos produziram uma cor rosa no ágar-CAS e não
alaranjada, que é a cor tipicamente descrita na literatura para a remoção de ferro do ·
CAS por sideróforos (SCHWYN e NEILANDS 1987, PAYNE 1994). Esse resultado foi . . .
também observado para outros fungos pertencentes às classes deuteromiceto e
zigomiceto (MILAGRES et ai., 1999). Porém o ascomiceto T. aurantiacus mudou a
cor do ágar-CAS para alaranjado.

39
Tabela 1: Crescimento de fungos e detecção de sideróforos em placas
preparadas meio a meio
Espéciesª Crescimento (dias)" Reação CAS Reação completa no melo"
P. chrysosporium 4 Rosa +++
T. versicolor 7 Rosa ++
P. coccineus 8 Rosa ++
P. pini 18 Rosa +
P. medula-pànis 10 Rosa ++
M. tremellosus 15 Rosa +
P. cocos 15 Rosa +
• 15 Rosa L. su/fureus +
T. aurantiacus 15 Alaranjado +
Controle 18 Azul
ª Todas 'as espécies são Basidiomicetos, exceto o T. aurantiacus que ·é um
Ascomiceto
b Tempo requerido para o micélio fúngico cobrir a metade da placa ágar-ME
e Completa reação no meio ágar-CAS (-, sem mudança de cor, + 1,0 - 4,0; ++ 4, 1 -
8,0; +++ > 8,0 mm/dia de avanço da fronteira de mudança de cor do ágar-CAS)

40
Figura 7: Meio sólido contendo ãgar-CAS/ ãgar-ME inoculado com :
A- P. pini, B- P. chrysosporium, C- T. versicolor, D- M. tremellosus,
E- T. aurantiacus, F- L. su/fureus

41
As diferenças nas respostas da reação CAS (rosa ou alaranjado)
. observada entre os microrganismos poderiam estar relacionadas as diferenças
estruturais dos sideróforos produzidos por cada cepa. Os dois grandes grupos de
sideróforos, hidroxamato e catecolato, produzem complexos fortemente coloridos na
presença de Fe(III). A pH neutro, a cor dos complexos monohidroxamatos. é vermelho-alaranjado e a dos trihidroxamatos é alaranjado, ao passo que a cor dos
complexos formados pelos catecóis é púrpura (NEILANDS 1984, PAYNE 1994). A
cor final no ágar-CAS deve ser o resultado de uma mistura entre a cor final do CAS
após reagir com um complexante e a cor do complexo que está sendo formado
(sideróforo-Fe(III)). Segundo PAYNE (1994), em altas concentrações de sideróforos,
a cor do complexo sideróforo-Fe(III) formado pode interferir na cor final da reação em
meio líquido e portanto nas leituras de absorbância. Dessa forma acredita-se que em
meio sólido possa acontecer o mesmo.
As diferentes cores (alaranjado ou rosa) produzidas no ágar-CAS
sugeriram a produção de sideróforos de naturezas diferentes, enquanto a
intensidade da cor poderia estar relacionada à concentraçãodo sideróforo.
A velocidade de mudança de cor, foi determinada pelo avanço da
mudança de cor azul do ágar-CAS. Os resultados mostraram que há diferenças entre
as cepas, e dessa forma pode-se determinar uma relação entre a velocidade da
mudança de cor e o crescimento microbiano. As cepas que cresceram mais
...., rapidamente no meio extrato de malte produziram uma rápida mudança de cor no
· ágar-CAS. A forma de expressar a velocidade da mudança de cor (mm/dia) não é exatamente uma quantificação da reação no meio sólido, uma vez que o inóculo não
foi padronizado.
À exceção do P. chrysosporium, T. versicolor e P. pini, que já haviam
sido estudadas por FEKETE et ai., (1989), os demais fungos estudados nesse
trabalho estão sendo descritos pela primeira vez como produtores de sideróforos em
meio sólido, usando o reagente CAS. DURÁN e MACHUCA (1995), verificaram que o
fungo termofílico T. aurantiacus produz um sideróforo, com estrutura de hidroxamato
em meio líquido, com propriedade de atuar na deslignificação de efluentes e· de
polpas celulóslcas. O interesse no estudo desses fungos decompositores de madeira

42
associados à produção de sideróforos vem aumentando nos últimos anos, desde que
foi sugerido que esses compostos apresentam um importante papel nos estágios
iniciais de decomposição da madeira. O desenvolvimento de novas metodologias de
detecção de sideróforos por tais fungos, que possibilite a ampliação da coleção de
cepas produtoras, pode viabilizar trabalhos futuros, dependendo da aplicação
desejada.
A técnica de separar o reagente CAS das células em crescimento foi
desenvolvida nesse trabalho para detecção de sideróforos em meio sólido, conforme
sugerido por SCHWYN e NEILANDS (1987). Com esse procedimento, foi possível
estudar a produção de sideróforos com todos os microrganismos selecionados para
esse trabalho, evitando o problema de inibição do crescimento das cepas, o qual é
causado pela toxicidade do reagente HDTMA, presente no ágar-CAS. Além disso, o
ensaio CAS modificado pode ser facilmente adaptado a todos os tipos de
,. microrganismos, visto que os meios de crescimento e detecção de sideróforos são
preparados separadamente. Isso possibilita a adição de suplementos (por exemplo,
vitaminas, aminoácidos, antibióticos, etc) ao meio de cultivo, estimulando o
crescimento microbiano, sem causar interferência na reação CAS.
5.2 - Produção de sideróforos em meio líquido
Após verificar que todos os fungos utilizados neste estudo
apresentaram reação positiva para a detecção de sideróforos em meio sólido
efetuou-se também o cultivo destes em meio líquido, constituído por extrato de malte,
2%. Amostras foram coletadas com intervalos de 5 dias para verificar a produção de
sideróforos, seguindo a metodologia descrita por SCHWYN e NEILANDS, (1987). A
maioria dos fungos estudados produziu uma intensa coloração rosa no meio sólido,
no entanto, em meio líquido esse resultado não se confirmou, houve apenas uma
redução da cor azul do reagente CAS, constatado pelo decréscimo na leitura de
absorbância das amostras.

43
P. chrysosporium, P. medula-panis e P. coccineus foram as cepas que
não alteraram a cor do CAS, e houve apenas uma redução de absorbância (Figura
8). Este fato pode estar associado a uma baixa produção dos compostos quelantes
ou a uma possível degradação do reagente CAS provocado por enzimas
liqninolfticas desses fungos.
P cocos, L. sulfureus e T. aurantiacus reagiram instantaneamente com
o CAS em meio líquido, alterando a cor de azul para rosa (Figura 9). P. cocos foi o
fungo que produziu a maior reação CAS atingindo a máxima porcentagem de
sideróforos no décimo dia de incubação e manteve-se praticamente constante, até o
•final do período de cultivo. Para o L. sulfureus, a cinética de produção de sideróforos
aumentou com o tempo de cultivo do fungo, atingindo o valor máximo após 20 dias.
No entanto, para o T. aurantiacus o pico máximo de reação com o CAS foi obtido no
quinto dia observando-se uma redução da cor da reação até o final de cultivo.

44
P. cnrysosporium P. medula-panis
, 100 100
~ 80 ~ 80 e.... e... li) 60 li) o o 60 · ... ... .2 40 · .E 40 ·O ·O ... ai & Ili 20 · "C "C 20 <Í> À <Í>
o ----- ,----,-·---- -----. o o 5 10 15 20 o 5 10 15 20
Período de Incubação (dias) Período de Incubação (dias) '\..
••
P. coccineus
5 10 15 20
100 ~ 80 - e.... li) 60 o ... .2 40 ·O ... Ili 20 "C
<Í> o o
Período de Incubação (dias)
Figura 8: Cinética da reação CAS produzida por extratos extracelulares de
P. chrysosporium, P. medula-panis e P. coccineus
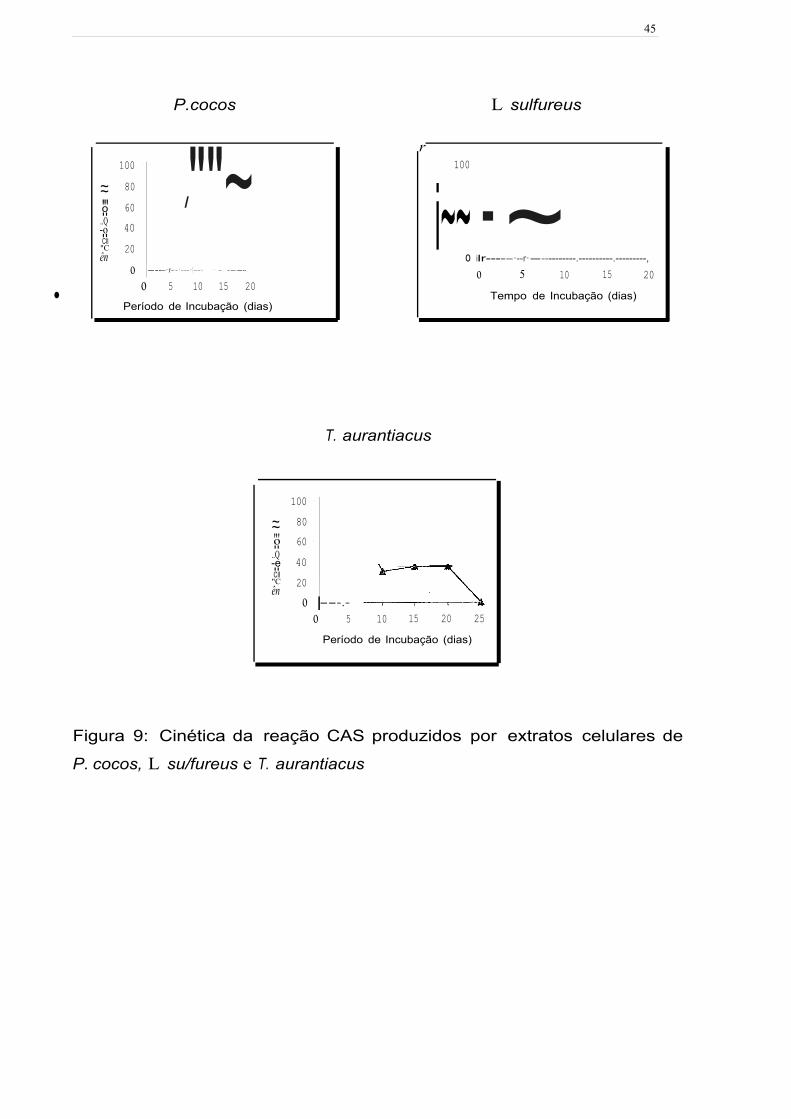
45
P.cocos
100 · ""~ ~ 80 ~ "' 60 I o ..
..Q 40 -o .. CII "C 20 ên
o -----·r-·-·---·1--·- ·-· -·. -----.
o 5 10 15 20 • Período de Incubação (dias)
L sulfureus
T. aurantiacus
r 100
i~~·~ o ilr-----·--r··-----------.----------.---------,
o 5 15
100 · ~ 80 ~ "' 60 · o ..
..Q 40 -e .. CII "C 20 ên
o ---.- o 5 10 15 20
10 Tempo de Incubação (dias)
20
Período de Incubação (dias)
25
Figura 9: Cinética da reação CAS produzidos por extratos celulares de
P. cocos, L su/fureus e T. aurantiacus

46
Os fungos M .tremellosus e T. versicolor não reagiram com o CAS
durante o período de incubação.
Dos fungos em estudo, somente o P. cocos, L. sulfureus e
T. aurantiacus reagiram positivamente com o CAS ( com mudança de cor)
independente do meio de cultura, sólido ou líquido.
Os outros fungos, embora tenham reagido positivamente em meio
sólido, quando cultivados em meio líquido produziram uma reação muito fraca com o
reagente CAS ( sem mudança de cor) e em alguns casos não houve reação. Essa
diferença entre a reação CAS observada em meio líquido e sólido podem ser
atribuídas a diferentes fatores associados à concentração de sideróforos. Em meio
.sólido, os micélios fúngicos crescem na superfície do meio e de forma diferente da
obervada em meio líquido. O crescimento de fungos em meio sólido pode aumentar
a excreção dos sideróforos, como observado para enzimas e outros metabólitos
(NIGANE e SINGH, 1994).
Em meio líquido, parece que os sideróforos necessitam ser produzidos
em altas concentrações para serem detectados através da reação CAS. Por outro
lado, a técnica de produção de sideróforos em meio líquido não permite detectar os
sideróforos no momento em que eles estão sendo produzidos, pois as amostras são
retiradas em intervalos de tempo e a reação CAS é realizada numa outra etapa. Isso
pode levar a uma menor reação CAS, visto que os sideróforos podem ser
degradados durante o período de incubação ou tornarem-se complexados com
outros metais. No final do período de cultivo, os micélios fúngicos foram coletados
para avaliação do crescimento através da determinação da massa _seca. A cinética
de produção de atividade oxidativa foi acompanhada simultaneamente à reação CAS
e as maiores atividades detectadas no final do cultivo, são mostradas na Tabela 2.

47
Tabela 2: Reação CAS e atividade oxidativa de fungos em meio líquido
Cepas Massa seca (mg) Reação com CAS Atividade oxidativaª(Ul/mL)
em meio líquido
P. chrysosporium 83,4 o T versicolor 125,3 NR 7,6
P. coccineus 108,6 4,3
P. pini 46,1 NR o P. medula-panis 84,9 6,3
M. tremellosus 63,7 NR 9,4
P. cocos 85,6 + o L. su/fureus 91,5 + o
T aurantiacus 258,0 + 1,8
Os fungos foram incubados durante 20 dias, à exceção de T aurantiacus que foi
cultivado por 25 dias. ª Atividade oxidativa determinada pela oxidação de o-dianisidina
( + ) Diminuição da absorbância a 630 nm, com mudança de cor no CAS
( - ) Diminuição da absorbância a 630 nm
(NR) Não reagiu
•

48
Os fungos cresceram de maneira diferenciada no meio extrato de malte
2%, que pode ser constatado pela diferença nas massas secas obtidas no final do
período de cultivo. Dentre os fungos, o menor crescimento foi observado com o
P. pini, o qual pode ter sido responsável pela ausência de reação CAS e atividade
oxidativa. Essa ausência de atividade foi também observada para o
P. chrysosporium , P. cocos e L. sulfureus durante todo o período de cultivo. Nos
extratos dos fungos M. tremellosus, T versicolor, P. medula-panis e T. aurantiacus
foi detectada atividade através da oxidação de o-dianisidina. Em geral, as atividades
oxidativas foram baixas nesses extratos e tal fato pode estar associado à natureza
do substrato utilizado no ensaio de atividade. A especificidade pelo substrato das
oxidases varia grandemente dependendo da espécie fúngica, porém os substratos
mais eficientemente oxidados são a o-dianisidina, ácido 2,2' azino-bis (3 etíl-
benzotiazolina-6-sulfónico) (ABTS) e 2,6 dimetoxifenol (2,6 DMF) (ISHIGAMI et ai.,
1988; KOFUJITAetal., 1991; REHMAN eTHURSTON, 1992; THAKKEReta/., 1992;
GOSHADZE e ELISASHVIHI, 1994). Em um trabalho posterior a esse MACHUCA e
MILAGRES (1998), utilizaram ABTS, 2,6 DMF e a o-dianisidina em testes de
atividade oxidativa destas mesmas culturas fúngicas e constataram que o ABTS foi o
melhor substrato para a detecção de atividade oxidativa. Nesse estudo foi verificado
que essa atividade está relacionada à presença de compostos de baixa massa molar
( < 3 kDa) nos extratos do T versicolor e P. medula-panis.
Como um dos objetivos desse estudo foi caracterizar sideróforos
produzidos por um fungo de decomposição parda e outro de decomposição branca
foram escolhidos o P.cocos e o T versicolor para a continuidade do trabalho. O
fungo de decomposição parda, P. cocos foi escolhido pois produziu a maior reação
CAS, que pode estar associada a uma alta produção de compostos complexantes.
Entre os fungos de decomposição branca a escolha do T. versicolor baseou-se na
presença de atividade oxidativa e nos resultados anteriores que indicaram que essa
atividade oxidativa estaria relacionada a compostos complexantes de baixa massa
molar (MACHUCA e MILAGRES, 1998).

49
5.3 - Caracterização dos caldos de cultura de um fungo de decomposição parda e de um fungo de decomposição branca
P.cocos e T. versicolor foram cultivados por 30 dias em meio extrato de
malte 2% e alíquotas foram coletadas a cada 5 dias objetivando a realização de um
estudo mais amplo da cinética de produção de sideróforos, numa tentativa de
relacionar o método CAS com outros métodos de detecção dos sideróforos.
A determinação de sideróforos foi realizada através do método CAS, e
por métodos químicos específicos como o teste de Csáky , o teste de Arnow e o
teste de Lowry.
5.3.1 - Detecção pelo método CAS
O método CAS detecta a presença de sideróforos independentemente
do tipo de estrutura do composto. O reagente CAS não mede a concentração total de ~ sideróforos numa cultura, mas a quantidade de sideróforos não complexados,
presentes em um determinado momento. A condição para a reação ser positiva com
o reagente CAS é o sideróforo não estar complexado, caso contrário, o composto
não apresenta capacidade para captar o ferro do CAS.
A reação para o acompanhamento das cinéticas de produção de
sideróforos com os respectivos extratos fúngicos, foi realizada após 1 hora da adição
do reagente CAS. A reação CAS foi considerada positiva quando ocorreu a redução
da cor azul do reagente ou quando ocorreu mudança da cor azul da solução para
alaranjada ou púrpura ocasionada pela formação de complexos sideróforos-Fe.

50
Os resultados da cinética da reação CAS de P. cocos revelaram o
aparecimento da reação após 5 dias de cultivo e no décimo dia a reação ocorreu
acompanhada de mudança da cor azul do reagente. A máxima produção de
sideróforo ocorreu com 25 dias (68%) acompanhada da mudança na cor azul para
rosa. No período correspondente a 30 dias de cultivo observou-se uma redução na
porcentagem dos sideróforos produzidos pelo P.cocos (Figura1 O).
6 100
80 UI o ... .2 60 -e ... <li :E 40 U)
~ o 20
:e e.
4
- 2
o --~----+-----------------1------- o O 10 20 30
Período de Incubação (dias)
Figura 10: Cinética da reação CAS (-•-) e do pH (_.-) dos sobrenadantes de
cultivo do P. cocos
Paralelamente, o pH das culturas foi determinado e observou-se um
decréscimo do pH de 5,5 para 2,8, durante o período de incubação. Inicialmente,
atribuiu-se a mudança de cor da reação CAS aos baixos valores de pH da cultura,
uma vez que a diminuição do pH coincidiu com o aparecimento da reação CAS.
Os sideróforos ao complexar o Fedo CAS podem formar complexos de
cores variadas dependendo do tipo de sideróforo e do pH das culturas. Os ferri-

51
hidroxamatos exibem uma absorção máxima na faixa de 425 a 520 nm a qual pode
se alterar em função do pH. Os monohidroxamatos são os que apresentam esse
comportamento de maneira mais acentuada, alterando a cor da solução de amarelo
para púrpura em pH inferior a 5,0. Esse efeito de mudança de cor do complexo
férrico em função do pH pode interferir com a reação característica do CAS. Os
sideróforos do tipo catecol quando complexados com o ferro são de cor púrpura a
pHs neutros, mas o ferro se dissocia em pHs ácidos e a cor é perdida (PAYNE,
1994).
Tal efeito foi estudado com soluções de desferal preparadas em pH 2,0;
4,6 e 6,0 a fim verificar diferenças nos valores de absorbância das soluções. Os
resultados desses ensaios mostraram que em pHs mais ácidos (pH 2,0) houve maior
decréscimo na absorbância a 630 nrn. Essa diferença sugeriu-nos uma mudança no e
espectro de absorção do complexo Fe-hidroxâmico ou ainda que em pHs mais
ácidos haja uma maior facilidade de complexação.
Com a cultura de P. cocos, o pH foi ajustado para o valor
correspondente ao meio de extrato de malte 2% (pH 5,5), por adição de NaOH, e a
reação CAS foi novamente realizada. Os resultados da leitura de absorbância !.Í
obtidos com a cultura em pH 2,8 e em pH 5,5 foram os mesmos.
Segundo a literatura, P. · cocos quando cultivado por 6 dias em meio
mineral acrescido de glicose 1 %, produz elevadas concentrações de ácido oxálico
(2 mg/ml), o qual é responsável pela redução do pH do meio (ESPEJO e AGOSIN,
1991 ). Além de reduzir o pH, o ácido oxálico pode complexar o Fe3+ e também e-
reduzir o Fe3+ em Fe2+ (GOODELL et ai., 1997). Em razão disso, foi conduzido um
experimento adicionando-se ácido oxálico em concentrações variadas à solução
CAS e observou-se reação positiva, com mudança de cor do CAS para rosa em
concentrações de ácido oxálico acima de O, 39 mg/ml. Dessa forma, o ácido oxálico
se presente na cultura de P. cocos, poderia estar reagindo com o CAS da mesma
forma que um sideróforo.
A cinética da reação CAS foi acompanhada na cultura de
T. versicolor e verificou-se uma discreta redução na cor do reagente CAS com 5 dias
de cultivo. Nas determinações seguintes a reação CAS desapareceu, não se

52
observando mudança na cor azul do reagente CAS, ou ainda, um decréscimo no
valor da absorbâncía das amostras a 630 nm (Figura 11 ). Em meio sólido, esse
fungo cresceu rapidamente e mudou a cor do CAS para rosa. Uma explicação para a
ausência de reação CAS em meio líquido seria a baixa produção desses compostos
durante o período de cultivo.
Diferentemente do que ocorreu com P. cocos, o pH do meio de cultivo de
T. versicolor permaneceu praticamente constante durante todo o período de
incubação. Esse fungo produz ácido oxálico em baixas concentrações durante seu
metabolismo secundário, porém ele não se acumula no meio devido a presença da
enzima intracelular oxalato descarboxilase que o decompõe rapidamente ( DUTTON
et ai. , 1993).
20 /\ . \ o - -------- --~-~---k---~ o
- 6
4
100 -
80 - C/1 o .... .2 60 - -o ....
o (1)
:2 40 (/) ·-- 2 ~ o
o 10 20 30 Período de Incubação (dias)
Figura 11: Cinética da reação CAS (-~-) e do pH (-11-) dos sobrenadantes de
cultivo do T. versicolor

53
5.3.2 - Detecção de sideróforos por métodos baseados na estrutura dos compostos
Os métodos específicos de Csáky, Arnow e Lowry usados na w determinação de sideróforos não produziram reação durante os diferentes períodos
de incubação nos quais os sideróforos foram detectados pelo método CAS. Somente
no final da incubação (30 dias) os caldos de P. cocos e T versicolor reagiram
positivamente ao método de Csáky, para detecção de estruturas de sideróforo do
tipo hidroxamato. Os caldos de P. cocos também reagiram com o método de Lowry o
utilizado na determinação de proteínas e hidroxamatos (Tabelas 3 e 4).
5.4 - Obtenção e análise de uma fração de baixa massa molar dos extratos de P. cocos e T. versicolor para detecção de sideróforos
Com 30 dias, os caldos foram coletados e liofilizados e os testes de
detecção de sideróforos novamente conduzidos com os extratos fúnqicos
concentrados (1 O vezes). Após esta etapa de concentração observou-se um
aumento das reações com CAS, Csáky e Lowry e o aparecimento de reação com o
método de Arnow, para sideróforo do tipo catecol (Tabelas 3 e 4).
Depois de concentrar os caldos por liofilização estes foram ultrafiltrados
através de uma membrana de 5 kDa com o objetivo de obter uma fração de baixa
massa molar livre de proteínas extracelulares e outros peptídeos fúngicos de alta
massa molar. Com essa fração ( < 5 kOa) foram novamente conduzidos os métodos
de detecção de sideróforos.
A reação CAS na fração de baixa massa molar de P. cocos , da mesma
forma como observada com o caldo liofilizado, foi instantânea e acompanhada de
mudança de cor do reagente de azul para rosa. Verificou-se que a reação CAS

54
manteve-se alta, mesmo após a ultrafiltração, indicando que compostos
responsáveis pela reação CAS positiya devem possuir massa molar abaixo de 5 kDa.
Os sideróforos do tipo catecolato foram principalmente detectados no
caldo concentrado por liofilização. Observou-se o aparecimento da cor alaranjada e
não vermelha como indicado pelo método. O ultrafiltrado obtido dessa amostra
(< 5 KDa) foi analisado e verificou-se um decréscimo de 15 vezes na intensidade
. da reação. Esse resultado mostra que compostos que reagem com o método de -~
Arnow se encontram também na fração de alta massa molar. Não deve ser
descartada a possibilidade de haver compostos não catecóis, interferindo no método
de Arnow.
Tabela 3: Determinação de sideróforos nos extratos de P. cocos
P.cocos CAS líquidoª ARNOW CSÁKY LOWRY
Caldo de cultura
de 30 dias de 18,6% ND 0,50 mg/L 27,2 mg/L
incubação
Caldo
liofilizado 87,8% 487 mg/L 2,8 mg/L 423,4 mg/L
Ultrafiltrado
(massamolar 89,2% 31 mg/L 2,8 mg/L 196,5 mg/L
< 5 kDa)
NO - não detectado
ª Os sideróforos foram expressos em porcentagem de reação CAS

55
Com o teste Csáky obteve-se reações positivas tanto no caldo
concentrado como no ultrafiltrado, indicando que os compostos que reagem a esse
método apresentam massa molar inferior a 5 kDa.
Os resultados obtidos da reação de Lowry com o ultrafiltrado ( <5 kDa)
foram positivos confirmando a produção de estruturas do tipo hidroxamato de baixa
massa molar. Parte dos compostos que reagiram com esse reagente não
apareceram na fração de baixa massa molar, e são provavelmente proteínas
excretadas pelo fungo durante seu metabolismo. O método de Lowry usado nesse
teste é o mesmo que se utiliza na determinação de proteínas, mas também é
·~ indicado para determinação da concentração de sideróforos do tipo hidroxamato
(SUBRAMANIAN et ai., 1965). A validade do método foi confirmada pela utilização
do sideróforo comercial desferrioxamina 8 (desferal), com estrutura de hidroxamato.
Foi feito um ensaio utilizando uma concentração de desferal de 130 mg/L e obteve-
se uma leitura de absorbância de 0,301. O desferal não absorve em 280 nm
podendo-se dessa forma afirmar que compostos de natureza não protéica reagem
com o método de Lowry se apresentarem ácidos hidroxâmicos. O meio extrato de
malte, utilizado no cultivo dos fungos reage com o método de Lowry (Abs2so = O, 17 4)
provavelmente devido a presença de aminoácidos, tirosina e triptofano, em sua
constituição.
Para o caldo do T. versicolor as mesmas análises foram realizadas, e
estão apresentadas na Tabela 4.
Os testes realizados com o T. versicolor para detecção de sideróforos
utilizando o método CAS foram negativos, mesmo após o caldo ser concentrado por
liofilização. Esse resultado já havia sido observado nos estudos anteriores, porém
nesse experimento pode-se verificar que a ausência de reação CAS em
T. versicolor parece não ser devido à sua concentração. Assim, outros fatores
ligados à velocidade de complexação de ferro pelos compostos produzidos por
T. versicolor podem estar ocorrendo (SCHWYN e NEILANDS, 1987).
Com o teste de Csáky obteve-se reações positivas tanto no caldo
concentrado como no ultrafiltrado, indicando que os compostos que reagem a esse
método apresentam massa molar inferior a 5 kDa.

56
O teste de Lowry produziu uma reação positiva no caldo de cultura
concentrado por liofilização e na fração de baixa massa molar ( < 5 KDa),
confirmando a produção de estruturas do tipo hidroxamato por T. versicolor. Da
mesma forma que para os extratos de P. cocos , o método de Lowry pode detectar
ácidos hidroxâmicos e proteínas presentes no meio de cultura de T. versicolor.
Porém, quando a cultura extracelular é ultrafiltrada por uma membrana de 5 kDa, é
·~ esperado que na fração menor que 5 kDa a presença de ácidos hidroxâmicos seja o
principal responsável pela reação com Lowry.
Tabela 4: Determinação de sideróforos nos extratos de T. versicolor
T. versicolor CAS líquido ARNOW CSÁKY LOWRY
Caldo de cultura
de 30 dias de NO NO 0,56 mg/L ..
incubação - - Caldo
liofilizado NO 391 mg/L 3,11 mg/L 318,8 mg/L
Ultrafiltrado
(massa molar NO NO 2,47 mg/L 397,6 mg/L
< 5 kOa)
NO - Não detectado

57
A possibilidade de existirem proteínas na fração menor que 5 kDa foi verificada
através do método de Bradford, o qual é mais. sensível que o método de Lowry e
apresenta menos interferências. A determinação proteínas através do método de
Bradford foi negativa na fração menor que 5 kDa. Assim, a reação positiva com o
método de Lowry na fração de baixa massa molar somente pode ser devida à presença de ácidos hidroxâmicos ou de aminoácidos como tirosina e triptofano.
O ensaio de Arnow foi positivo somente na fração concentrada por
liofilização, ao passo que no meio de cultura com 30 dias de incubação e na fração
de baixa massa molar a reação foi negativa. Isso sugere a presença de compostos
no meio de cultura que podem reagir através do ensaio de Arnow, para detecção de
catecóis, que possuem uma massa molar acima de 5 kDa.
Comparando as reações produzidas nos ultrafiltrados dos dois fungos
observa-se que ambos deram reação positiva para compostos com estrutura de
hidroxamato (Tabelas 3 e 4). No entanto, apenas com o P. cocos foi detectada
reação CAS e reação para catecóis. Esses resultados nos levam a sugerir que os
catecóis detectados nesse meio sejam os principais responsáveis pela reação CAS.
Conforme anteriormente mencionado, a reação CAS foi obtida instantaneamente e
essa é uma característica descrita para os sideróforos do tipo catecolato (SCHWYN
e NEILANDS, 1987). A velocidade de troca de ferro entre os compostos quelantes do
tipo hidroxamato e o CAS é mais lenta, podendo levar horas para ocorrer.
Uma vez que a literatura cita que fungos decompositores de madeira
produzem ácido oxálico (DUTTON et ai, 1993), e em nossos resultados foi, verificado
que o ácido oxálico reage com o 'CAS, foi realizado um ensaio para verificar sua
interferência ao método Arnow, Csáky e Lowry. Observou-se que o ácido oxálico não
reage com nenhum dos métodos, não sendo detectado a formação de um complexo
colorido no meio reacional dos métodos de caracterização de sideróforos. Esse
resultado reforça a especificidade do método pela estrutura do composto.

58
5.5 - Cromatografia em Sephadex G-25
Os caldos liofilizados e as frações resultantes da ultrafiltração foram
submetidas à cromatografia de filtração em gel (Sephadex G-25).
O perfil cromatográfico obtido no caldo liofilizado de P. cocos
apresentou três picos distintos de absorbância a 280 nm. O primeiro pico, eluiu
próximo ao volume morto da coluna (Vo), constituindo fração de massa molar
superior a 5 kDa. Essa fração não reagiu com o CAS ou apresentou atividade
oxidativa. Os outros picos eluíram na região de fracionamento da coluna e após
análise detectou-se reação CAS no pico correspondente a 25 ml (Figura 12 A).
A fração de baixa massa molar de P. cocos, obtida após a ultrafiltração
dos extratos liofilizados, foi aplicado à coluna de Sephadex G-25 e um único pico foi
evidenciado. As frações foram reunidas e uma análise qualitativa mostrou que as
frações apresentaram a reação CAS e grande quantidade de açúcares redutores
(Figura 12 B).
A massa molar da fração que reagiu com o CAS não pode ser
determinada através da filtração em gel por falta de padrões de baixa massa molar.
No entanto, estima-se que esteja abaixo de 1300 Da, visto que foi eluída após a
cobalamina (massa molar de 1355,4 g/mol). Embora a cromatografia em gel seja
bastante utilizada na determinação da massa molar de sideróforos, a metodologia
recomendada para este tipo de composto é a técnica de espectrometria de massa,
uma vez que a utilização de cromatografia geralmente conduz a valores
subestimados (MACHUCA, 1995).
O extrato de T. versicolor foi também aplicado à coluna de Sephadex G-
25 e dois picos eluíram desta coluna, sendo um bastante definido, com uma alta
absorção e outro menor (Figura 13 A). Foi feita análise CAS e de atividade oxidativa
nas frações correspondentes aos picos e foi detectada uma pequena atividade
oxidativa em ambas frações. A atividade correspondente ao primeiro pico deve-se
provavelmente a enzimas lignocelulolíticas que possam estar presentes no meio. No
entanto, a atividade oxidativa do segundo pico sugere a presença de um composto

59
(A)
1 Vo V1 V2
0,8
j 1 1 o 0,6 CIO N ui .a 0,4 <(
0,2
o -----· o 20 40 60
Vol. Acumulado (ml)
(B)
1 -,
i 1 0,8 1 1
o 1
CX) 0,6 j N .,;
' .o 0,4 j <! I l l 02Í ~ • 1
OJ ...... ~•HUUH --- 0 20 40
Vol. Acumulado (ml)
60
Figura 12: Cromatografia em Sephadex G-25 das frações do caldo extracelular
do P. cocos, caldo concentrado por liofilização (A), fração de baixa massa
molar (PM < 5 KDa) (B). A coluna foi eluída com água deionizada, com um fluxo
·~ de 0,8 mUmin e frações de 1,5 ml foram coletadas. A coluna foi calibrada com
blue dextran PM = 2000 KDa (Vo) = 15 ml, cobalamina PM = 1355,4 g/mol
(V1) = 24 ml e riboflavina PM = 376,37 g/mol (V2) = 39 ml.

60
capaz de catalisar reações do tipo redox. Resultado semelhante foi encontrado para
T. aurantiacus e sugeriu-se um mecanismo de ação em que o ferro presente no
composto seria o responsável pelo potencial de oxidação de diversos substratos
(MACHUCA, 1995).
A fração de baixa massa molar de T. versicolor, obtida da ultrafiltração,
foi aplicada à coluna Sephadex G-25 e foi identificado um pico largo, mas com baixa
absorbância a 280 nm, porém com atividade oxidativa (Figura 13 .B). Em todas as
frações foi detectada a presença de açúcares redutores, sendo que a maior
concentração de açúcar coincidiu com o pico de absorbância máxima.
A massa molar da fração com atividade oxidativa foi estimada através
dos resultados dessa cromatografia e obteve-se um valor próximo de 1300 Da.
5.6 - Cromatografia em Sephadex G-1 O
A fim de melhorar a resolução da separação das frações de baixa
massa molar dos extratos fúngicos, as amostras foram aplicadas a uma coluna
Sephadex G-1 O. Essa coluna por apresentar um limite de exclusão menor que a
Sephadex G-25, esperava-se poder determinar com maior precisão a amssa molar
dos compostos e obter mais picos de A28o-
Quando o ultrafiltrado .de P. cocos foi aplicada a essa coluna, foi obtido
somente um pico de A280 que eluiu no volume morto da coluna, e onde foi detectada
a reação CAS. Esse resultado impossibilitou a determinação da amssa molar do
composto, mais indicou que ela seja superior a 700 Da (Figura 14 ).
Foi feita a determinação de açúcares redutores em todas as fra'ções
eluídas dessa cromatografia e verificou-se que eles começaram a ser eluídos na ·~
fração correspondente ao volume morto de 19 ml, porém o maior pico eluiu com 21
ml.
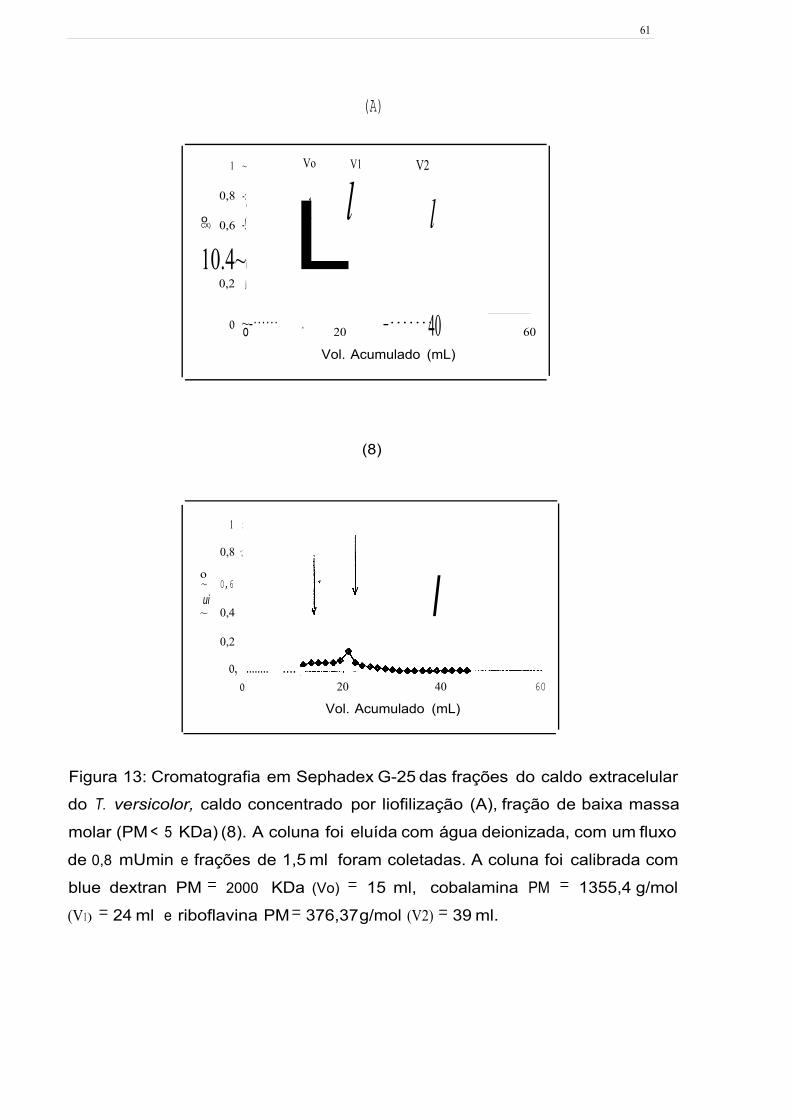
61
(A)
1 ~ Vo V1 V2
0,8 -: 1 l l ' '
0,6 -! o CX)
10.4~ L i 0,2 j
o ~-······ . -······· o 40 60 20
Vol. Acumulado (mL)
(8)
1 :
0,8 ·;
l o ~ 0,6 ui ~ 0,4
0,2
o, ........ ....., o 40 60 20
Vol. Acumulado (mL)
Figura 13: Cromatografia em Sephadex G-25 das frações do caldo extracelular
do T. versicolor, caldo concentrado por liofilização (A), fração de baixa massa
molar (PM < 5 KDa) (8). A coluna foi eluída com água deionizada, com um fluxo
de 0,8 mUmin e frações de 1,5 ml foram coletadas. A coluna foi calibrada com
blue dextran PM = 2000 KDa (Vo) = 15 ml, cobalamina PM = 1355,4 g/mol
(V1) = 24 ml e riboflavina PM = 376,37 g/mol (V2) = 39 ml.

62
A utilização de uma coluna com limite de exclusão menor possibilitou a separação
dos açúcares da fração que reagiu com o CAS. A partir desse resultado, ficou
demonstrado que a reação CAS não sofre interferências de açúcares presentes nos
meios de cultura, mesmo em altas concentrações.
Os extratos de baixa massa molar ( < 5 KDa) do T. versicolor, quando
aplicados à coluna Sephadex G-1 O, também eluíram com um único pico onde foi
detectada uma pequena atividade oxidativa. A massa molar dessa fração foi
estimada como sendo superior a 700 Da (Figura 15). As características apresentadas
por essa fração poderiam corresponder a um peptídeo, porém, não se encontra
informação na literatura que peptídeos possuam características de catalisar a
oxidação de substratos. Algumas das propriedades da fração com atividade
oxidativa, parcialmente purificada de T. versicolor, foram semelhantes ao encontrado
por MACHUCA (1995), para o ascomiceto Thermoascus aurantiacus.
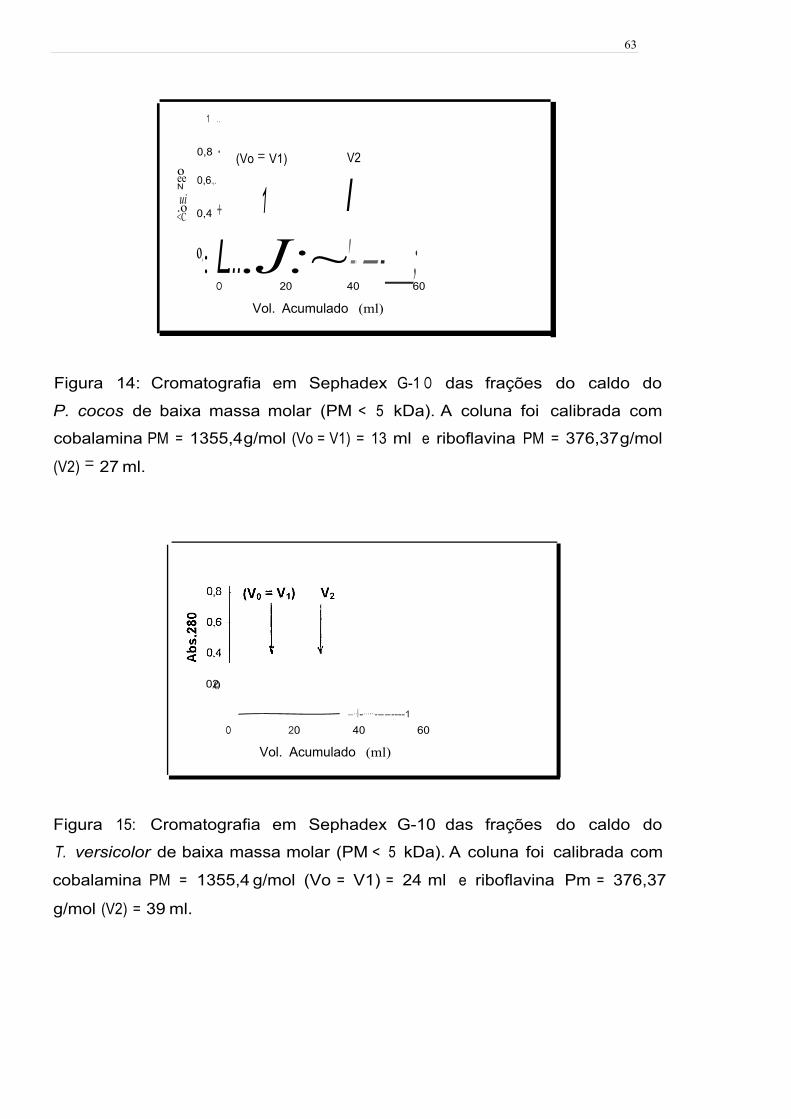
63
1 ..
0,8 • (Vo = V1) V2 o
1 l ee 0,6.,. N
ui 0,4 + .o
<C
º·: L ... .J:~'-- --____; O 20 40 60
Vol. Acumulado (ml)
Figura 14: Cromatografia em Sephadex G-1 O das frações do caldo do
P. cocos de baixa massa molar (PM < 5 kDa). A coluna foi calibrada com
cobalamina PM = 1355,4 g/mol (Vo = V1) = 13 ml e riboflavina PM = 376,37 g/mol
(V2) = 27 ml.
02 00
~--··Í·-······-------1
O 20 40 60
Vol. Acumulado (ml)
Figura 15: Cromatografia em Sephadex G-10 das frações do caldo do
T. versicolor de baixa massa molar (PM < 5 kDa). A coluna foi calibrada com
cobalamina PM = 1355,4 g/mol (Vo = V1) = 24 ml e riboflavina Pm = 376,37
g/mol (V2) = 39 ml.

64
6 - CONCLUSÕES
1~ • O cultivo em meio sólido, realizado em placas contendo ágar-CAS/ ágar-ME
possibilita selecionar um maior número de cepas produtoras de sideróforos, mesmo
aquelas descritas como sensíveis ao reagente CAS, ou com alguma exigência
nutricional específica.
• Todas .as cepas produziram reação CAS positiva em meio sólido, mas somente 3
cepas fúngicas apresentaram um comportamento semelhante no meio líquido. Em
meio líquido parece que os sideróforos necessitam ser produzidos em altas
concentrações para serem detectados através da reação CAS. A velocidade de
complexação de ferro dos compostos pode diferir entre as cepas fúngicas, podendo ser
visualizada pela intensidade de mudança de cor da reação CAS
- -~
• Os métodos de detecção de sideróforos mostraram que as reações ocorreram nas
frações de baixa massa molar dos extratos fúngicos.
• P. cocos reduziu o pH do meio de crescimento e reagiu positivamente com o CAS, ao
método de Arnow, Csáky e Lowry. A partir desses resultados, conclui-se que o fungo
produza compostos de baixa massa molar do tipo catecolato e hidroxamato. No entanto
não se pode excluir que a reação CAS bastante intensiva produzida por esse fungo
seja também devido à presença de ácidos orgânicos excretados no meio.
Os testes realizados com T. versicolor para detecção de sideróforos indicaram a
presença de composto com estrutura do tipo hidroxamato

65
• A fração de baixa massa molar ( < 5 KDa) de P. cocos apresentou máxima reação
CAS e não apresentou atividade oxidativa, porém, na fração de baixa massa molar do
T. versicolor detectou-se atividade oxidativa e ausência da reação CAS.
O estudo da produção de sideróforos por fungos de decompositores de madeira
permitiu-nos concluir que há uma correlação entre a máxima atividade oxidativa e a
mínima reação CAS.

66
7 - REVISÃO BIBLIOGRÁFICA
ARNOW, L. E. Colorimetric determination of the components of 3,4-
dihydroxyphenylalanine-tyrosine mixtures. Journal of Biological Chemistry, v. 118,
p. 531-537, 1937.
ATKIN, C. L., NEILANDS, J. B. Rhodotorulic acida diketopiperazine dihydroxamine
acid with growth fator activity. 1. lsolation and characterization. Biochemistry, v. 7,
p. 3734-3739, 1968.
BACKA, S., GIERER, J., REITBERGER, T., NILSSON, T. Hidroxil radical activity in
brown-rot fungi studied by a new chemiluminescence method. Holzforschung, v.
46, p. 61-67, 1992.
BAGG, A., NEILANDS, J. B. Molecular mechanism of regulation of siderophore-
mediated iron assimilation. Microbiological Reviews, v. 51, p. 509-518, 1987.
BARRASA, J. M., GONZÁLES, A. E., MARTÍNEZ, A. T. Ultrastructural Aspects of
Fungai Delignification of Chilean Woods by Ganoderma australe and Phebia
chrysocrea. Holzforschung, v. 16, p. 1-8, 1992.

67
BLANCHETTE, R. A, BURNES, T. A, EERDMANS, M. M., AKHTAR, M. Evaluating
isolates of Phanerochaete chrysosporium and Ceriporiopsis subvermisporafor
use in biological pulping process. Holzforschung, v. 96, p. 109-115, 1992. •'
BLANCHETTE, R. A Delignification by wood-decay fungi. Annual Reviews of
Phytopathology, v. 29, p. 381-398, 1991.
BRADFORD, M.M. A rapid and sensitive methods for the quantitation of microgram
quantities of protein utilizing the principie of protein-dye binding, Analytical
Biochemistry, v. 72, p. 248-254, 1976.
CSÁKY, T. Z. On the estimation of bound hydroxylamine in biological rnaterials. Acta
Chemica Scandinavica, p. 450-454, 1948.
CHIMIAK, A, HIDER, R. C., LUI, A, NEILANDS, J. B., NOMOTO, K., SUGIURA, Y.
Siderophores from microrganisms and plants. Berlin: Springer - Verlag, 1984.
COWLING, E. B. Physical and chemical constraints in the hydrolysis of cellulose and
lignocellulosic material. Biotechnology and Bioengineering Symposium, v. 5, p.
163-181, 1975.
DANIEL, G. Use of electron microscopy for aiding our understanding of wood
biodegradation. FEMS Microbiology Reviews, v. 13, p. 199-234, 1994.

68
DANIEL, G., NILSSON, T., PETTERSON, B. Intra and extracellular localization of
ü lignin peroxidase during the degradation of solid wood fragments by
Phanerochaete chrysosporium by using transmission electron microscopy and
imuno-gold labeling. Applied and Environmental Microbiology, v. 55, p. 871-881,
1989.
DÚRAN, N., MACHUCA, A Kraft pulp bleaching by na extracellular low molecular
weight metabolite from the ascomycete Thermoascus aurantiacus. Hold ais Koh
und Werkstott, v. 53, p. 346, 1995.
DUTTON, M. V., EVANS, C. S., ATKEY, P. T., WOOD, D. A Oxalate production by
basidiomycetes, including the white-rot species Coriolus versicolor and
Phanerochaete chrysosporium. Applied Microbiology and Biotechnology, v. 39, p.
5-10, 1993.
- ·- .... > ERIKSSON, K. E., BLANCHETTE, R. , ANDER, P. Microbial and enzimatic
degradation of wood and wood components. Berlin: Springer Verlag, 1990. p
407 .
. \ERIKSSON, K., KIRK, T. K. Biopulping, biobleaching and treatment of kraft bleaching
effluents with white-rot fungi. ln: MOO-YOUNG, M. ed. Comprehensive
Biotechnology: The principies, aplications and regulations of
Biotechnology in lndustry, Agriculture and Medicine. New York: Pergamon Press,
1985. V. 3, p.271-294

69
ERIKSSON, K. Microbial degradation of celulose and lignin. ln: INTER. SYMP. ,,
WOOD PULP CHEMISTRY, Stockholm, 1981. v. 3, p. 60-65
ESPEJO, E., AGOSIN, E. Production and degradation of acid oxalic by brown-rot
fungi. Applied and Environmental Microbiology, v. 57, n. 7, p. 1980-1986, 1991.
EVANS, C. S., DUTTON, M. V., GUILLÉN, F., VENESS, R. G. Enzymes and small
molecular mass agents involved with lignocellulose degradation. FEMS
Microbiology Reviews, v. 13, p. 235-239, 1994.
EVANS, C. S., GALLAGHER, 1. M., ATKEY, P. T., WOOD, D. A. Localization of
degradative enzymes in white-rot decay of lignocellulose. Biodegradation, v. 2, p.
93-106, 1991.
FEKETE, F. A., CHANDHOKE, V., JELLISON, J. lron-binding compounds produced
by wood-decaying Basidiomycetes. Applied and Environmental Microbiology, v.
55, p. 2720-2722, 1989.
FEKETE, F. A., SPENCE, T., EMERY, T. A rapid and sensitive paper electrophoresis
assays for the detection of microbial siderophores elicited in solid-plating culture.
Analytical Biochemistry, v. 131, p. 516-519, 1983.
FENGEL, D., WEGENER, G. Wood: chemistry, ultraestruture, reactions.Berlin: W. de
Gruyter, 1984. p. 613

70
FLUORNOY, D. S., KIRK, T. K., HIGHLEY, T. L. Wood decay by brown-rot fungi:
changes in pare structure and cell wall volume. Holzforschung, v. 45, p. 383-388,
1991.
GOODELL, 8., JELLISON, J., LIU, J., DANIEL, G., PASZCZYNSKI, A, FEKETE, F.,
KRISHNAMURTHY, S., JUN, L., XU, G. Low molecular weight chelators and
phenolic compounds isolated from wood fungi and their role in the fungai
biodegradation of wood. Journal of 8iotechnology, v. 53, p. 133-162, 1997.
GOODELL, 8., LIU, J., JELLISON, J., LU, J., PASZCZYNSKI, A, FEKETE, F., A
Chelation activity and hydroxyl radical production mediated by low molecular
weight phenolate compounds isolated from Gloeophyl/um trabeum. ln: INT.
CONFERENCE 810TECHNOL. PULP ANO PAPER IND., 6, Viena, Austria, O-
F1-233, V. 45, 1995.
GOSHADZE, M. K., ÉLISASHVILI, V. 1. 8iosynthesis and some properties of laccase
from Lentinus tigrimus IBR - 101. Biochemical (Biokhimiya), v. 58, p. 1448-1452,
1994.
HAMMEL, K. E., KAWAI, S., JENSEN Jr., K. A, BAO, W., SREBOTNIK, E. Can fungi
degrade lignin without lignin peroxidase? ln: INT. CONFERENCE
810TECHNOL. PULP ANO PAPER IND., 6, Viena, Austria, O-F1-233, v. 45, 1995. ,.

71
HARVEY, P. J., SCHOEMAKER, H. E., PALMER, J. M. Veratryl alcohol as a
mediator and the role of radical cation in lignin biodegradation by Phanerachaete
chrysosporium Febs Letters, v. 195, p. 242-246, 1986.
HELM, D. van der, WINKELMANN, G. Hydroxamates and polycarboxylates as iron
transport agents (siderophores) in fungi. ln: WINKELMANN, G., WINGE, D. R. eds. Metal ions in fungi. New York : Marcel Dekker, 1994. cap. 2, p. 39-98.
HIDER, R. C. Siderophore mediated absorption of iron. Estructure Bonding, v, 58, p.
25-87, 1984.
HIGHLEY, T. Degradation of cellulose by Poria placenta in the presence of
compounds that affect hydrogen peroxide. Material und Organismen, v. 15, p. 81-
90, 1980.
HIRANO, T., TANAKA, H., ENOKI, A. Extracellular substance from the brown-rot
basidiomycete Tyromyces palustris that reduces molecular oxygen to ràdicals
and ferric to ferrous iron. Mokuzai Gakkaishi, v. 41, p. 334-341, 1995.
HOLZBERG, M., ARTIS, W. M. Hydroxamate siderophore production by opportunistic
and systemic fungai pathogens, lnfection and lmmunity, v. 40, n. 3, p, 1134-1139,
1983.
ISHIGAMI, T., HIROSE, Y., YAMADA, Y. Characterization of polyphenol oxidase from
Chaetomuim thermophile, a thermophilic fungus. Journal of General and Applied
Microbiology, v. 34, p. 401-407, 1988.

72
JELLISON, F., CHANDHOKE, V., GOODELL, B., FEKETE, A The isolation and
immunolocalization of iron-binding compounds produced by Gloeophyllum
trebeum. Applied Microbiology and Biotechnology, v. 35, p. 805-809, 1991.
JELLISON, J., GOODELL, B., FEKETE, F., CHANDHOKE, V. Fungai siderophores
and their role in wood biodegradation. ln: ANNUAL MEETING, 21, Rotorua, New
Zealand, 13-18 maio 1990.
JOSELEAU, J. P., GHARIBIAN, S., COMTAT, J., LEFEBVRE, A:, ruel, K. lndirect
involvement of ligninolytic enzyme systems in cell wall degradation. FEMS
Microbiology Reviews, v. 13, p. 255-264, 1994.
KIRK, T. K., FARREL, R. L. Enzimatic combustion: The microbial degradation of
lignin. Annual Reviews of Microbiology, v. 41, p. 465-505, 1987.
KOENIGS, J. W. Hidrogen peroxide and iron: A proposed system for decomposition
of wood by brown-rot basidiomycetes. Wood Fiber, v. 6, p. 66-80, 197 4. - r
KOFUJITA, H., OHTA, T., ASADA, Y., KUWAHARA, M. Purification and
characterization of laccase from Lentinus edodes. Mokuzai Gakkaishi, v. 37, p.
562-569, 1991.
LEONG, S. A, NEILANDS, J. B. Siderophore production by phytopathogenic
microbial species. Archives of Biochemistry and Biophysics, v. 218, p. 351-359,
1982.

73
MACHUCA. A., MILAGRES, A. M. F. Oxidative activity of iron-binding chelators
produced by wood-rot fungi. ln: INTERNACIONAL CONFERENCE ON
BIOTECHNOLOGY IN THE PULP ANO PAPER INDUSTRY, 7., Vancouver-
Canadá, 16-19 jun. 1998. p 8-245-8-248.
MACHUCA, A. Produção e caracterização parcial de um composto de baixa massa
molécular, com atividade fenoloxidásica, de Thermoascus aurantiacus.
Campinas: UNICAMP, 1995 (Tese de Doutorado)
MARTÍNEZ, J. L., DELAGDO-IRRIBARREN, A., BAQUERO, F. Mechanisms of iron
acquisition and bacterial virulence. FEMS Microbiology Reviews, v. 75, p. 45-56,
1990.
MESSENGER, A. J. M., RATLEDGE, C. Siderophores. ln: MOO-YOUNG, M. ed.
Comprehensive Biotechnology: The Principies, Applications and Regulations of
Biotechnology in lndustry, Agriculture and Medicine. Oxford: Pergamon Press,
1985. v. 3, cap. 13, p. 275-295
MILAGRES, A. M. F., MACHUCA, A., NAPOLEÃO, D. AS., Detection of siderophore
production from severa! fungi and bacteria by a modification of chromo azurol S
(CAS) agar plate assay. Journal of Microbiological Methods, 1999.
MILLER, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar.
Analytical Chemistry, v. 31, p. 426-428, 1959.

74
NEILANDS, J. 8. Methodology of siderophores. Struct. Bonding, v. 58, p. 1-24, 1984.
NIGANE, P., SINGH, D. Solid-state (substrate) fermentation systems and their
applications in biotechnology. Journal of Basic Microbiology, v. 34, n 6, p. 405-
423, 1994.
PAYNE, S. M. Detection, isolation and characterization of siderophores. ln: lron
assays and deferration of media. Methods in Enzymology, v. 235, p. 329-344,
1994.
PAYNE, S. M. lron and virulence in the family Enterobacteriaceae. CRC Criticai
Reviews in Microbiology, v. 16, p. 81-111, 1988.
RAASKA, L., VIIKARI, L., MATTILA-SANDHOLM, T. Detection of siderophore in
growing culture of Pseudomonas spp. J. lnd. Microbial, v. 11, p. 181-186, 1993.
REHMAN, A. U., THURSTON, C. F. Purification of laccase I from Armillaria mellea.
Journal of General Microbiology, v. 138, p. 1251-1257, 1992.
RIQUELME,M. Fungai siderophores in plant-microbe interactions. Microbiologia SEM,
1996.

75
RUEL, K., AMBERT, K., JOSELAU, J. P lnfluence of enzyme equipment of white-rot
fungi on the patterns of wood degradation. FEMS Microbiology Reviews, v. 13, p.
241-254, 1994.
SAVORY, J. G. Breakdown of timer by sacomycetes and fungi imperfect. Annals of
Applied Biology, v. 41, p. 336-347, 1954.
SCHWYN, B., NEILANDS, J. B. Universal chemical assay for the detection and
determination of siderophore. Analytical Biochemistry, v. 160, p. 47-56, 1987.
SHAH, S., KARKHANIS, V., DESAI, A lsolation and characterization of siderophore,
with antimicrobial activity, from Azospirillum lipoferum M. Current Microbiology, v.
25, p. 347-351, 1993.
SHIMADA, M., MA,D-B, AKAMATSU, Y., HATTORI, T. A proposed role of oxalic acid
in wood decay systems of wood-rotting Basidiomycets. FEMS Microbiology
Reviews, v. 13, p. 285-296, 1994.
SREBOTNIK, E., MESSNER, K. A simple method that uses differential staining and
light microscopy to asses the selectivity of wood delignification by white rot fungi.
Applied and Environmental Microbiology, v. 55, p. 871-881, 1994.
SREBOTNIK, E., MESSNER, K. lrnrnunoelectron microscopial study of the porosity of
brown-rot degraded pine wood. Holzforschung, v. 45, p. 95-101, 1991.

76
SUBRAMANIAN, K. N., PADMANABAN, G., SARMA, P. S. Folin-ciocalteu reagent for
the estimation of siderochromes. Analytical Biochemistry, v. 12, p. 106-112, 1965.
SZKLARZ, G. D., ANTIBUS, R. K., SINSA BAUGH, R. L., LINKINS, A. E. Production
of phenoloxidases and peroxidases by wood-rotting fungi. Mycologia, v. 81, p.
234-240, 1989.
TANAKA, H., HIRANO,T., ENOKI, A. Extracellular substance from the white-rot
basidiomycete lrpes lacteus for production and reduction of H202 during wood
degradatíon. Mokuzai Gakkaishi, v. 39, p. 493-499, 1993.
THAKKER, G. D., EVANS, C. S., RAO, K. K. Purification and characterization of
laccase from Monocillium indicum saxena. Applied Microbiology and
Biotechnology, v. 37, p. 321-323, 1992.
. - :._..,.,
TSCHIERSKE, M. DRECHSEL, H., JUNG, G., ZAHNER, H. Production of rhizoferrin
and new anoulogues obtained by directed fermatatiom. Applied Microbiology and
Biotechnology, v. 45, p. 664-670, 1996.
WOOLDRIDGE, K. G., WILLIANS, P. H. lron uptake mechanisms of pathogenic
bacteria. FEMS Microbiology Reviews, v. 12, p. 325-348, 1993.
WOOD, P. M. Pathways production of Fenton's reagent by wood rotting fungi. FEMS
Microbiology Reviews, v. 13, p. 313-320, 1994.

77
ANEXO 1
1 ' 1
0,8 1
1 ' ! i
0,6 1 1
0,5
o ..... U)
ui .e <t 0,4
i ! !
0,2 I 1 o +-----------,---------- · -- -T------,-----------------,---------------1
y= 1,2443x + 0,0037 R2= 0,9952
0,2 0,3 0,4
FIGURA 1 A: Curva de calibração preparada com o ácido 2,3 dihidroxibenzóico
utilizada para detecção de sideróforos do tipo catecolato através do método de
Arnow
, . ...
o 0,1
Cone. do ácido 2,3 dihidroxibenzóico (mg/ml)

78
ANEXO 2
2,5 1 1
2 J
U) 1,5 N IO ui .e <(
1 1 0,5 -1
o _J_:_ , __
y = 0,239x + 0,0019 R2 = 0,9961
--·--·· · t":" ··---~----.------·,---------,
6 8 10 o 2 4 Cone. üe cloreto de hidroxilamina (mg/ml)
FIGURA 2 B: Curva de calibração preparada com o cloreto de hidroxilamina
utilizada para detecção de sideróforos do tipo hidroxamato através do método de ~~
Csáky

79
ANEX03
1
0,8
o 0,6 .. <D <D
clÍ .a <! 0,4 ·
y = 3,5291x + 0,0062 R2 = 0,9946
0,2 ·
o .. -···-·-··-, ··-·.····--······--·r-------.
0,3 0,4 o 0,1 0,2 Cone.de desferal (mg/ml)
FIGURA 3 C: Curva de calibração preparada com o desferal utilizada para ,
detecção de sideróforos do tipo hidroxamato através do método de Lowry
__ _.,.,
Recommended