
UNIVERSIDADE FEDERAL DE PERNAMBUCO
DEPARTAMENTO DE ENERGIA NUCLEAR
COMISSÃO NACIONAL DE ENERGIA NUCLEAR
CENTRO REGIONAL DE CIÊNCIAS NUCLEARES DO NORDESTE
Programa de Pós-Graduação em Tecnologias Energéticas e Nucleares
ESTOQUES E DINÂMICA DO CARBONO E NITROGÊNIO
EM SOLOS SOB DIFERENTES COBERTURAS E USOS DE
TERRA EM PERNAMBUCO
KENNEDY NASCIMENTO DE JESUS
Orientador: Prof. Dr. Everardo V. S. B. Sampaio
Recife, PE
Fevereiro, 2017

KENNEDY NASCIMENTO DE JESUS
ESTOQUES E DINÂMICA DO CARBONO E NITROGÊNIO
EM SOLOS SOB DIFERENTES COBERTURAS E USOS DE
TERRA EM PERNAMBUCO
Tese submetida ao Programa de Pós-Graduação
em Tecnologias Energéticas e Nucleares para
obtenção do título de Doutor em Ciências, Área
de Concentração: Fontes Renováveis de Energia.
Orientador: Prof. Dr. Everardo V. S. B. Sampaio
Recife, PE
Fevereiro, 2017

Catalogação na fonte Bibliotecário Carlos Moura, CRB-4 / 1502
J58e Jesus, Kennedy Nascimento de.
Estoques e dinâmica do carbono e nitrogênio em solos sob
diferentes coberturas e usos de terra em Pernambuco. / Kennedy
Nascimento de Jesus. - Recife: O Autor, 2017.
112 f. : il., tabs.
Orientador: Prof. PhD. Everardo Valadares de Sá Barretto Sampaio.
Tese (doutorado) – Universidade Federal de Pernambuco. CTG.
Programa de Pós-Graduação em Tecnologias Energéticas e
Nucleares, 2017.
Inclui referências bibliográficas.
1. Classes de solo. 2. Carbono total. 3. Nitrogênio total. 4.
Argissolos. 5. Mudança de uso da terra. 6. Isótopos de 13C e 15N. I.
Sampaio, Everardo Valadares de Sá Barretto, orientador. II. Título.
UFPE
CDD 631.4 (21. ed.) BDEN/2017-08

Estoques e Dinâmica do Carbono e Nitrogênio
em Solos Sob Diferentes Coberturas e Usos de
Terra em Pernambuco
Kennedy Nascimento de Jesus
APROVADA EM: 02.02.2017
ORIENTADOR: Prof. Dr. Everardo Valadares de Sá Barreto Sampaio
COMISSÃO EXAMINADORA:
___________________________________________________________
Prof. Dr. Everardo Valadares de Sá Barreto Sampaio – DEN/UFPE
________________________________________________
Prof. Dr. Rômulo Simões Cezar Menezes – DEN/UFPE
_______________________________________________________
Profa. Dra. Maria do Socorro Bezerra de Araújo – DCG/UFPE
__________________________________________________________
Prof. Dr. Flávio Adriano Marques – EMBRAPA Solos-UEP Recife
_____________________________________________________________
Prof. Dr. Luís de França da Silva Neto – EMBRAPA Solos-UEP Recife
Visto e permitida a impressão
_____________________________________
Coordenador(a) do PROTEN/DEN/UFPE

Aos meus avós, José Eugênio e Genésia Vieira, pelos
melhores ensinamentos da vida.
“In memoriam”
Dedico

AGRADECIMENTOS
A DEUS, por manter inabalável a minha fé e por me mostrar, através da ciência, a
perfeição da natureza;
À minha mãe, Elcy Nascimento, pelo exemplo de garra e determinação, por ensinar-me
o sentido da família, do amor e da importância que a educação tem em nossas vidas;
Aos meus avós, meu pai e irmãos, meus tios e tias, primos e primas que, mesmo distante,
sempre estiveram torcendo pelo meu sucesso;
Ao Prof. Everardo Sampaio, agradeço profundamente pela orientação, pelo estímulo,
esclarecimentos, sugestões e correções oportunamente apresentadas, a permanente
disponibilidade e o voto de confiança e amizade durante a realização deste trabalho. Serei
eternamente grato;
À Universidade Federal de Pernambuco (UFPE), em especial ao Programa de Pós-
Graduação em Tecnologias Energéticas e Nucleares (PROTEN);
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela
concessão da bolsa de Doutorado no Brasil e pelo financiamento da pesquisa nos Estados
Unidos da América (EUA) na Texas A&M University (TAMU), através do projeto de cooperação
internacional CAPES/TAMU, “Influência do Uso da Terra na Ecohidrologia e Biogeoquímica da
Caatinga Brasileira” (« The Influence of Land Use on the Ecohydrology and Biogeochemistry of
the Brazilian Caatinga »), (Processo 10002-14-6 CAPES/TAMU, 2014.1 Doutorado Sanduíche);
À Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE),
a Fundação de Amparo à Ciência e Tecnologia do Estado de São Paulo (FAPESP) e ao
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pelo apoio
financeiro e logístico, principalmente através dos projetos “Estoques de Carbono na Vegetação
e no Solo em Áreas com Diferentes Usos da Terra em Pernambuco” (CNPq Universal, 14/2012,
Processo 473449/2012-9), “Impactos de Mudanças Climáticas sobre a Cobertura e Uso da
Terra em Pernambuco: Geração e Disponibilização de Informações para o Subsídio de
Políticas Públicas” (Processo APQ-0.077-5, 01/09, edital 05-2010) e “Estoque vertical de
carbono orgânico estável em diferentes solos, biomas e uso da terra no estado de
Pernambuco” (FACEPE/CAPES - Processo 0199-3.09/12);
Aos membros do Comitê de Acompanhamento do Doutorado, Prof. Rômulo Menezes,
Prof. Flávio Marques e o Pesq. Dário Primo, pelas inúmeras contribuições, sugestões e
correções durante a realização desse trabalho;

Aos professores Antônio Antonino (UFPE) e Jason West (TAMU), por todo o apoio nas
etapas que antecederam o intercâmbio e pela orientação durante o período do Doutorado
sanduíche nos Estados Unidos (EUA);
Aos pesquisadores da Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA),
especialmente à Vanderlise Giongo (EMBRAPA – Semiárido) e Luciano Accioly (EMBRAPA
– Solos, UEP Recife), pelo suporte nas análises laboratoriais e identificação das áreas de
amostragem do projeto;
Aos Pesquisadores e funcionários da Associação Plantas do Nordeste (APNE),
especialmente ao Eng. Ftal. Frans Pareyn, que contribuiu efetivamente nas atividades de campo
e nas discussões do projeto;
Ao Pedólogo Elmo Clark, pela fundamental ajuda na identificação das classes de solo e
aos Professores Gustavo Duda, da Unidade Acadêmica de Garanhuns (UAG – UFRPE) e Plínio
Camargo, do Centro de Energia Nuclear da Agricultura (CENA – USP), pelas contribuições
nas análises laboratoriais;
A todos do Departamento de Energia Nuclear (DEN – UFPE), professores, funcionários,
estagiários, colaboradores e colegas de pós-graduação pelo acolhimento, convivência e
amizade; em especial à Claudenice Maria, Gilberto Eloi, Pedro Antônio, Patryk Melo, Eliza
Albuquerque, Rodrigo Lustosa, Júlio Martins, Emmanuel Dutra, Taciana Santos, Mona
Vasconcelos, Monaliza Andrade, Diego Marcelino, Natache Ferrão, Nilson Medeiros, Mônica
Santana, Tânia Lúcia, Carol Gonzáles, Girleyde Lima, Wiliane Costa, Uemeson Santos,
Raphael Correia, Edilândia Dantas, Gabriela Santiago, Alice Sabrina, Fernando Pinto, Tiago
Diniz, Erick Xavier, Renata Souza, Andrea Avelino, Benaia Barros, Ana Dolores, Josenildo,
Zacarias, Nilvânia Monteiro e Kalidja;
As pessoas especiais que fazem parte das nossas vidas através de pequenos gestos de
carinho e ajuda, e que colaboraram de forma direta ou indireta para realização deste trabalho;
em particular à amiga Eline Lima, a quem tenho um enorme apreço;
Aos agricultores que permitiram as coletas das amostras de solo em suas propriedades
e forneceram informações importantes para essa pesquisa;
E a todos que de alguma forma contribuíram na elaboração deste trabalho.
A todos vocês, MUITO OBRIGADO!

“Aprender é a única coisa de que a mente nunca se
cansa, nunca tem medo e nunca se arrepende."
Leonardo da Vinci

RESUMO
A conversão de áreas de vegetação nativa em áreas destinadas à exploração agropecuária é
apontada como um dos principais fatores responsáveis pelas emissões de Gases de Efeito Estufa
para a atmosfera. Essas mudanças no uso da terra causam na paisagem um mosaico de áreas com diferentes usos e coberturas do solo. Porém, ainda é limitado o conhecimento da dinâmica
do carbono (C) e do Nitrogênio (N) do solo nesses sistemas, principalmente na região Nordeste do Brasil. Nesse sentido, o objetivo desse trabalho foi determinar as alterações dos estoques e
da dinâmica do C e N no solo devido às mudanças de uso da terra nas diferentes regiões fisiográficas e nas principais classes de solo de Pernambuco. Foram realizados três estudos, que
auxiliaram no alcance do objetivo proposto: 1) no primeiro, foi realizado um levantamento e compatibilização da literatura sobre os estoques de C na camada superficial (0-30 cm) dos solos
de Pernambuco. 2) no segundo estudo, foram quantificados os estoques de carbono (in situ) e
sua distribuição nas camadas ao longo do perfil do solo (0-100 cm) em áreas de vegetação nativa, agrícolas e pasto, nas principais regiões fisiográficas e classes de solo do Estado; e 3)
no terceiro, foram avaliados a dinâmica do C e do N, através da abundância natural de δ13C e
δ15N e das concentrações de C e N em perfis (0-100 cm) de Argissolos ao longo de um gradiente climático em Pernambuco em áreas representativas de floresta nativa, de pasto e agrícolas. Em
todos os estudos, as áreas de vegetação nativa foram tomadas como condição original do solo
de acordo com cada região estudada. Nos estudos 2 e 3 quantificou-se as concentrações de C e N por meio de analisador elementar CHN e no estudo 3 foram quantificadas a abundância
natural de δ 13C e δ 15N nos Argissolos do Estado, por meio de espectrômetro de massa.
Aproximadamente 368 Tg de C estão estocados nos solos de Pernambuco de 0-30 cm. De uma forma geral, os maiores estoques de carbono total ocorreram nas vegetações nativas densas e
decresceram da região úmida à semiárida Oeste, com grandes variações entre as classes de solos. A substituição da vegetação nativa para implantação de áreas de pastagens ou agrícolas
acarretou mudanças no sinal δ13C do solo, sendo maiores nas áreas úmidas e subúmidas do
estado em relação às áreas semiáridas. O enriquecimento de δ15N encontrado na superfície do
solo das regiões semiáridas em relação às áreas úmidas e subúmidas estão associados as menores concentrações de C e N presentes nos Argissolos dessa região, em decorrência das
maiores temperaturas e menores precipitações pluviais. Fica evidente que a intervenção humana nessas áreas através de práticas agropecuárias, reduz os estoques e modificam a dinâmica do C
e do N no solo, e para se contrapor a esta situação no Estado, são necessárias medidas mitigatórias, como a adoção de sistemas conservacionistas do solo e da água, através de
políticas públicas de incentivo às práticas conservacionistas em substituição à agricultura de
baixo C.
Palavras-chave: Carbono total. Nitrogênio total. Classes de solo. Argissolos. Mudança de uso
da terra.Isótopos de 13C e 15N.

ABSTRACT
The conversion of native vegetation areas into agricultural areas is pointed out as one of the
main factors responsible for the emissions of greenhouse gases to the atmosphere. Changes in
land use create a mosaic of areas in the landscape with different soil coverages and knowledge of the dynamics of C and N in these systems is limited. The objective of this work was to
determine the changes in soil carbon (C) and nitrogen (N) stocks and dynamics due to changes in land use in the different physiographic regions and in the main soil classes of Pernambuco
state. Three studies were carried out: 1) the first was a survey and compatibilization of the literature on C stocks in the surface layer (0-30 cm) of the soils of Pernambuco; 2) the second
study was a quantification of the C stocks along the different layers of the soil profile (0-100 cm) in areas under native vegetation, agriculture and pasture, in the main physiographic regions
and soil classes of Pernambuco; and 3) the third study evaluated soil C and N dynamics through
the natural abundance of δ13C and δ15N and the concentrations of C and N in profiles of Ultisols
(0-100 cm) along a climatic gradient in Pernambuco. In all studies, dense native vegetation areas were taken as the original, reference soil condition. In studies 2 and 3 the concentrations
of C and N were quantified using CHN elemental analyzer. In study 3 the natural abundance of
δ13C and δ15N were quantified by mass spectrometer. Approximately 368 Tg of carbon are
stored in the soils of Pernambuco in the 0-30 cm layer. The largest total carbon stocks occurred
under dense native vegetation and decreased from the humid region towards the semi-arid western state region, with large variations among soil classes. The replacement of the native
vegetation by agriculture or pasture caused changes in the soil δ13C signal, being greater in the
humid areas and subhumid regions than in the semi-arid region. The enrichment of δ15N in the
soil surface layer of the semi-arid regions in relation to the humid and subhumid regions is associated with the lower concentrations of C and N in these regions, due to their higher
temperatures and lower rainfall. It is evident that Human intervention in these areas through conventional farming practices reduces soil carbon stocks at levels lower than those found in
native areas, and modifies the C and N dynamics in the soil in these regions. To counteract this situation in the State, mitigating measures are necessary, such as the adoption of soil and water
conservation systems, through public policies to encourage conservation practices in place of
low carbon agriculture.
Keywords: Total carbon. Total nitrogen. Soil classes. Ultisols. Change of land use. Isotopes of 13C and 15N.

LISTA DE ILUSTRAÇÕES
Figura 1 - Localização geográfica do estado de Pernambuco e suas regiões fisiográficas ...... 36
Figura 2 - Características climáticas das regiões fisiográficas do Estado de Pernambuco. ..... 38
Figura 3 - Classes de solos mais representativas de Pernambuco. ........................................... 39
Figura 4 - Valores de densidade do solo coletados de 1997 a 2012 em trabalhos publicados em
áreas de Pernambuco na camada de 0-30 cm sob diversos usos e cobertura da terra. .... 43
Figura 5 - Mapa do Estado de Pernambuco com a localização dos perfis de solos utilizados para
a estimativa dos estoques de carbono.............................................................................. 44
Figura 6 - Distribuição dos pontos amostrais no estado de Pernambuco. ................................ 46
Figura 7 - Imagem com a demarcação dos diferentes usos da terra em um Latossolo. ........... 48
Figura 8 - Mapa de uso e cobertura da terra em Pernambuco. ................................................. 50
Figura 9 - Trincheira para coleta de amostras de solo. ............................................................. 52
Figura 10 - Análises de CT pelo método da combustão seca em um analisador elementar de
CHN. ............................................................................................................................... 53
Figura 11 - Estoques médios de C (Mg ha-1) na camada de 0-30 cm nos solos do Brasil e de
Pernambuco. .................................................................................................................... 64
Figura 12 – Médias dos estoques de C no solo de 0 a 30 cm, sob diferentes usos e coberturas da
terra no estado de Pernambuco. Barras representam o erro padrão da média. ................ 65
Figura 13 - Distribuição em profundidade do C do solo sob diferentes usos e coberturas da terra
na Zona da Mata de Pernambuco. ................................................................................... 67
Figura 14 - Distribuição em profundidade do C do solo sob diversos usos e coberturas da terra
na Zona do Agreste de Pernambuco................................................................................ 68
Figura 15 - Distribuição em profundidade do C do solo sob diversos usos e coberturas da terra
na Zona do Sertão leste de Pernambuco. ........................................................................ 69
Figura 16 - Distribuição em profundidade do C do solo sob diversos usos e coberturas da terra
na Zona do Sertão oeste de Pernambuco......................................................................... 70
Figura 17 - Redução nos estoques de CT em função das mudanças no uso da terra nas regiões
fisiográficas de Pernambuco. .......................................................................................... 76
Figura 18 - Teores de C (%) em perfis de Argissolos sob diferentes usos terra nas diferentes
regiões fisiográficas de Pernambuco. .............................................................................. 78
Figura 19 - Carbono e nitrogênio totais do solo (0-20 cm) em função da precipitação pluvial
média anual, em diferentes usos e coberturas da terra em Argissolos de Pernambuco. . 79

Figura 20 - Teores de N (%) em perfis de Argissolos sob diferentes usos e cobertura da terra
nas diferentes mesorregiões do estado de Pernambuco. ................................................. 82
Figura 21 - Nitrogênio total do solo (0-20 cm) em função do carbono orgânico total do solo sob
vários usos e coberturas em Argissolos de Pernambuco. ................................................ 83
Figura 22 - Composição isotópica de 13C (‰) em perfis de Argissolos sob diferentes usos e
cobertura da terra nas diferentes regiões do estado de Pernambuco. .............................. 86
Figura 23 - Variação dos valores de 15N (‰) em perfis de Argissolos sob diferentes usos e
cobertura da terra nas diferentes mesorregiões do estado de Pernambuco. .................... 88
Figura 24 - Abundância natural de 15N do solo (0-20 cm) em função da precipitação pluvial,
altitude, carbono total e nitrogênio total do solo sob diversos usos e coberturas em
Argissolos de Pernambuco. ............................................................................................. 89

LISTA DE TABELAS
Tabela 1 - Características das principais classes de solos presentes no estado de Pernambuco.
......................................................................................................................................... 40
Tabela 2 - Trabalhos utilizados para o levantamento dos estoques de C dos solos de
Pernambuco. .................................................................................................................... 42
Tabela 3 - Estratificação do estado de Pernambuco em regiões fisiográficas, classes de solos e
usos e cobertura da terra mais representativos. ............................................................... 47
Tabela 4 - Características dos principais sistemas de uso e cobertura da terra nas diferentes
regiões fisiográficas e classes de solos do estado de Pernambuco.................................. 49
Tabela 5 - Área dos diferentes tipos de uso e cobertura das terras de Pernambuco e proporção
em relação à área total do Estado. ................................................................................... 51
Tabela 6 - Abrangência da classe dos Argissolos no Estado de Pernambuco. ......................... 56
Tabela 7 - Principais usos e coberturas vegetais existentes em áreas de predominância de
Argissolos no estado de Pernambuco, com algumas de suas características. ................. 57
Tabela 8 - Localização dos pontos amostrais de Argissolos em Pernambuco. ........................ 58
Tabela 9 - Estoques de C (0 a 30 cm de profundidade) por unidade de área e para áreas cobertas
no território de Pernambuco, classe de solo, número de perfis (n) e erro padrão da média
(s(m)). .............................................................................................................................. 61
Tabela 10 - Estoques de carbono total (Mg ha-1) na camada de 1 m de profundidade, em solos
sob diferentes usos e coberturas da terra em Pernambuco. ............................................. 71
Tabela 11 - Estoques de CT (Mg ha-1) em camadas de solos sob diferentes usos e coberturas da
terra em Pernambuco....................................................................................................... 73
Tabela 12 - Relação C:N em perfis de Argissolos sob diferentes usos e coberturas da terra nas
diferentes regiões do estado de Pernambuco. ................................................................. 84

LISTA DE ABREVIATURAS E SIGLAS
12C Isótopo estável de cabono 12
13C Isótopo estável de cabono 13
13C/12C Razão isotópica 13C: 12C
14C Isótopo radioativo de carbono 14
14N Isótopo estável de nitrogênio 14
15N Isótopo estável de nitrogênio 15
C Carbono
C/N Relação carbono: nitrogênio
C3 Ciclo fotossintético C3
C4 Ciclo fotossintético C4
CAM Ciclo fotossintético CAM (Crassulacean Acid Metabolism)
CO Carbono orgânico
CO2 Dióxido de carbono
COS Carbono orgânico do solo
COT Carbono orgânico total
CT Carbono total
Ds Densidade do solo [g cm-3]
FBN Fixação biológica de nitrogênio
GEE Gases de efeito estufa
MOS Matéria orgânica do solo
N Nitrogênio
N2 Nitrogênio elementar (atmosférico)
N2O Óxido nitroso
NH4+ Nitrogênio em sua forma mineral amoniacal
NO Óxido nítrico
NO3- Nitrogênio em sua forma mineral nítrica
NT Nitrogênio total
PDB Pee Dee Belemnite (Padrão internacional do carbono)
PEPcase Enzima de carboxilação fosfenolpiruvato carboxilase (plantas C4)
Pg Petagrama (1 Pg = 1015g, ou um bilhão de toneladas)
ppm Partes por milhão

RuBisCo Enzima de carboxilação ribulose-1,5 bifosfato carboxilase (plantas C3)
Tg Teragrama (1 Tg = 1012g, ou 1 milhão de toneladas)
δ Desvio em relação ao padrão [ ‰ ]
δ13C Desvio em relação ao padrão de 13C [ ‰ ]
δ15N Desvio em relação ao padrão de 15N [ ‰ ]

Sumário
1 INTRODUÇÃO .............................................................................................. 17
2 REVISÃO DE LITERATURA ..................................................................... 20
2.1 Ciclos biogeoquímicos do carbono (C) e nitrogênio (N) ................................................ 20
2.1.1 Ciclo do C ........................................................................................................................ 20
2.1.2 Ciclo do N ........................................................................................................................ 21
2.2 A matéria orgânica do solo (MOS) ................................................................................. 23
2.2.1 Fatores determinantes no estoque de MOS ..................................................................... 24
2.2.1.1 Textura e mineralogia do solo ...................................................................................................24
2.2.1.2 Clima ..........................................................................................................................................25
2.2.1.3 Uso e cobertura da terra ............................................................................................................26
2.3 Isótopos estáveis de 13C e 15N no estudo da MOS .......................................................... 29
2.3.1 Abundância natural de 13C ............................................................................................... 29
2.3.1.1 Fracionamento isotópico de carbono pelas plantas ..................................................................30
2.3.1.2 Ciclos fotossintéticos C3, C4 e CAM ...........................................................................................31
2.3.2 Abundância natural de 15N .............................................................................................. 33
3 MATERIAL E MÉTODOS ........................................................................... 36
3.1 Caracterização das áreas de estudo ................................................................................ 36
3.1.1 Localização, limites e área............................................................................................... 36
3.1.2 Regiões fisiográficas e características edafoclimáticas ................................................... 37
3.1.3 Distribuição dos solos ...................................................................................................... 39
3.2 Estudo 1 - Levantamento dos estoques de carbono nos solos de Pernambuco. .......... 41
3.2.1 Estimativas dos estoques de C do solo ............................................................................ 41
3.3 Estudo 2 - Distribuição em profundidade e estoques de carbono em solos sob diferentes
coberturas e usos da terra em Pernambuco. ........................................................................ 45
3.3.1 Delineamento da área de estudo ...................................................................................... 45
3.3.2 Levantamento das classes de solos e coberturas vegetais existentes............................... 47
3.3.2.1 Usos e coberturas da terra .........................................................................................................49

3.3.3 Coleta das amostras de solo ............................................................................................. 51
3.3.4 Processamento e análise das amostras de solo ................................................................ 52
3.3.4.1 Cálculo dos estoques de C do solo .............................................................................................53
3.3.5. Análise dos dados ........................................................................................................... 55
3.4 Estudo 3 - Carbono, nitrogênio e abundância natural (δ13C e δ15N) do solo sob
diferentes usos ao longo de um gradiente climático em Pernambuco. .............................. 55
3.4.1 Levantamento dos Argissolos e das coberturas vegetais existentes ................................ 56
3.4.2 Coleta das amostras de solo ............................................................................................. 58
3.4.3 Processamento e análise das amostras de solo ................................................................ 59
3.4.4. Análise dos dados ........................................................................................................... 59
4 RESULTADOS E DISCUSSÃO ................................................................... 61
4.1 Estudo 1 - Levantamento dos estoques de carbono nos solos de Pernambuco. .......... 61
4.1.1 Estoques de C por classe de solo ..................................................................................... 61
4.1.2 Estoques de C no solo por uso e cobertura da terra ......................................................... 64
4.2 Estudo 2 - Distribuição em profundidade e estoques de carbono em solos sob diferentes
coberturas e usos da terra em Pernambuco. ........................................................................ 66
4.2.1 Distribuição do C no solo em profundidade .................................................................... 66
4.2.2 Estoques totais de C no solo ............................................................................................ 70
4.3 Estudo 3 - Carbono, nitrogênio e abundância natural (δ13C e δ15N) do solo sob
diferentes usos ao longo de um gradiente climático em Pernambuco. .............................. 77
4.3.1 Carbono total ................................................................................................................... 77
4.3.2 Nitrogênio total ................................................................................................................ 81
4.3.3 Relação C: N .................................................................................................................... 83
4.3.4 Abundância natural de δ13C ............................................................................................. 85
4.3.5 Abundância natural de δ15N............................................................................................. 88
5 CONCLUSÕES .............................................................................................. 92
REFERÊNCIAS ................................................................................................ 94

17
1 INTRODUÇÃO
A atual preocupação com as mudanças climáticas globais decorrentes do aumento nas
concentrações de gases de efeito estufa (GEE) na atmosfera, em especial do gás carbônico
(CO2), tem despertado a atenção da comunidade científica para o ciclo biogeoquímico do
carbono (C). O gás carbônico é responsável por cerca de 70% do potencial de elevação da
temperatura terrestre (IPCC, 2007). Nos últimos 250 anos, a concentração desse gás na
atmosfera aumentou 38%, alcançando os atuais 402 ppm, o mais alto nível observado nos
últimos 420 mil anos (NOAA, 2016).
A conversão de áreas de vegetação nativa em áreas destinadas à exploração agropecuária
é apontada como um dos principais fatores responsáveis pelas emissões de CO2 para a atmosfera
(LAL, 2001). A agricultura e a pecuária são as principais atividades relacionadas com o uso dos
solos, e mudanças decorrentes de práticas de manejo inadequadas, podem levar a um rápido
declínio dos estoques de C e nitrogênio (N) do solo, colaborando para o aumento das emissões
de GEE para a atmosfera e intensificando o efeito estufa sobre o globo terrestre (FREIXO et
al., 2002).
Nas últimas décadas, os ecossistemas terrestres vêm sendo considerados tão importantes
quanto os oceanos na retirada (sequestro) e no armazenamento (estoque) de C da atmosfera,
mitigando o efeito estufa. O maior compartimento de C dos ecossistemas terrestres são os solos,
contendo quantidades que superam em até quatro vezes as presentes na biomassa vegetal e em
mais de duas vezes as presentes na atmosfera do planeta (WATSON et al., 2000). Cerca de
2.050 Pg de carbono (1 Pg = 1015g, ou um bilhão de toneladas) estariam armazenados nos
ecossistemas terrestres, dos quais 70% estariam nos solos (WATSON et al., 1990). A
quantidade de C estocada no solo até um metro de profundidade estaria em torno de 1.576 Pg,
distribuídos em 12,8 bilhões de hectares, nos continentes (ESWARAN et al., 1993). Quando se
consideram apenas os 30 cm superficiais de solo, o estoque seria de 800 Pg (CERRI et al.,
2006), ou seja, quase a mesma quantidade armazenada no compartimento atmosférico.
Estima-se que a metade do C estocado no solo (0-100 cm), ou seja, cerca de 787 Pg,
esteja relacionada aos solos sob florestas (DIXON et al., 1994), enquanto as áreas sob pastagens
conteriam cerca de 500 Pg (SCHARPENSEEL, 1997) e as sob cultivo agrícola 170 Pg de C
(PAUSTIAN et al., 2000).
Estimativas para todo o território do Brasil (850 milhões de ha) mostraram estoques de
C em torno de 36,4±3,4 Pg na camada de 0–30 cm de solo (BERNOUX et al., 2002), o que

18
corresponde a aproximadamente 40% de todo o C armazenado nos solos da América Latina
(BERNOUX e VOLKOFF, 2006). Se for considerada a profundidade de 0 a 100 cm, os estoques
seriam da ordem de 65,9 a 67,5 Pg de C, dos quais 65% estariam na região amazônica brasileira
(BATJES, 2005). Na região semiárida do Nordeste (98 milhões de ha), Sampaio e Costa (2011)
estimaram os estoques de C na camada superficial do solo (0-20 cm) como sendo 2,8 Pg e na
camada de 20 aos 100 cm, 6,0 Pg; totalizando 8,8 Pg de 0 a 100 cm.
A principal via de entrada do C derivado do CO2 da atmosfera para o solo é a deposição
dos resíduos vegetais e animais, formados inicialmente pela fotossíntese. Parte deste carbono
acumula-se, formando a matéria orgânica do solo (MOS), que é o mais importante reservatório
de C nos ecossistemas terrestres (ROSCOE et al., 2006). Nos ecossistemas naturais, a fonte de
carbono orgânico (CO) do solo tem como principal origem, os resíduos da vegetação nativa,
enquanto nos agroecossistemas o carbono total (CT) pode ser derivado do material orgânico
remanescente da vegetação nativa, da decomposição do sistema radicular das culturas agrícolas,
da aplicação de calcário e da adubação orgânica (CARVALHO et al., 2009).
A importância do conhecimento da MOS, bem como sua relação com o manejo e uso
da terra, visa desenvolver estratégias para utilização sustentável dos solos, para reduzir o
impacto das atividades agrícolas sobre o ambiente. Isso se torna imprescindível na região do
presente estudo, visto que os cultivos agrícolas e as pastagens principalmente na região
semiárida do Estado são, na maioria das vezes, formados após a retirada da vegetação nativa
pelo uso do fogo e em áreas declivosas sem um manejo adequado dos solos (SAMPAIO e
COSTA, 2011). Neste cenário, a exploração desordenada e a retirada da cobertura vegetal
intensificam o processo erosivo, culminando com a diminuição da fertilidade dos solos, em
decorrência da redução da MOS. Consequentemente, o uso agropecuário é comumente seguido
pelo abandono das áreas, ocorrendo então a regeneração da vegetação nativa. Essa dinâmica
causa na paisagem um mosaico de áreas com diferentes usos e coberturas do solo, e é limitado
o conhecimento da dinâmica do C e N do solo nesses sistemas. Vários fatores podem influenciar
essa dinâmica, como o tipo de solo, o clima e o manejo agropecuário.
As dinâmicas de C e de N dos solos podem ser estudadas usando os isótopos estáveis
13C e 15N, especialmente úteis considerando as diferentes fontes e a complexidade de seus ciclos
biogeoquímicos (ALVES et al., 2005). As avaliações da dinâmica são baseadas na abundância
natural de δ13C e de δ15N da MOS (BUSTAMANTE et al., 2004; SISTI et al., 2004; ALVES
et al., 2005; JANTALIA et al., 2007). O δ13C e o δ15N quantificados através de técnicas
isotópicas têm sido bons indicadores do tipo de vegetação existente, e das modificações a que
uma área foi submetida. Em regiões tropicais, esta técnica tem sido utilizada nos estudos de

19
ciclagem da MOS em médio prazo, onde florestas (ciclo fotossintético C3) foram substituídas
por culturas do ciclo fotossintético C4 (JANTALIA et al., 2007; PINHEIRO et al., 2010,
COSTA JUNIOR et al., 2011a, b).
Nesse sentido, a presente pesquisa baseia-se na hipótese de que a mudança de uso da
terra, através da adoção de novos sistemas agropecuários, controlam a capacidade de acumular
C e N, a distribuição em profundidade e a taxa de sequestro das classes de solos em função das
diferentes regiões climáticas do estado de Pernambuco.
Sabendo-se que as mudanças no uso da terra causam na paisagem um mosaico de áreas
com diferentes usos e coberturas do solo, e que é limitado o conhecimento da dinâmica do C e
N do solo nesses sistemas e da importância desse tipo de conhecimento para mitigação da
mudança climática global, o objetivo desse trabalho foi determinar as alterações dos estoques e
da dinâmica do C e N no solo devido às mudanças de uso da terra nas diferentes regiões
fisiográficas e nas principais classes de solo do estado de Pernambuco.
Para tal, serão apresentados três trabalhos, que irão auxiliar na obtenção do objetivo
proposto.
Estudo 1 – Levantamento dos estoques de carbono nos solos de Pernambuco;
Estudo 2 – Distribuição em profundidade e estoques de C em solos sob diferentes
coberturas e usos da terra em Pernambuco;
Estudo 3 – Concentrações de C, N e abundância natural (δ13C e δ15N) em Argissolos
sob diferentes usos ao longo de um gradiente climático em Pernambuco.
O primeiro estudo refere-se ao levantamento e compatibilização da literatura sobre os
estoques de C na camada superficial (0-30 cm) dos solos de Pernambuco, realizado através de
uma ampla revisão de trabalhos publicados na literatura especializada, em boletins
exploratórios de levantamentos de solos e em periódicos científicos, buscando contribuir com
informações mais detalhadas sobre os estoques de C do Nordeste.
O Segundo estudo traz a quantificação dos estoques de carbono (in situ) e sua
distribuição nas camadas ao longo do perfil do solo (0-100 cm) em áreas de vegetação nativa,
cultivos agrícolas e áreas de pastagens, nas principais regiões fisiográficas e classes de solo do
estado de Pernambuco.
O terceiro e último estudo avalia a dinâmica do C e do N, através da abundância natural
de δ13C e δ15N, e das concentrações de C e N em perfis de Argissolos ao longo de um gradiente
climático em Pernambuco em áreas representativas de floresta nativa, de pasto e agrícolas.

20
2 REVISÃO DE LITERATURA
2.1 Ciclos biogeoquímicos do carbono (C) e nitrogênio (N)
A dinâmica dos ecossistemas depende da interação entre os ciclos biogeoquímicos, os
quais podem ser modificados pelas atividades humanas. As mudanças no uso da terra, por
exemplo, modificam profundamente os processos biogeoquímicos do solo, com reflexos no
estoque de C, na disponibilidade de N e na emissão de gases para a atmosfera (LIMA et al.,
2000). Para amortizar o impacto das atividades humanas sobre o ambiente é necessário um
entendimento da dinâmica global, regional e local de cada ciclo individualmente e das relações
entre eles (CANADELL et al., 2000).
2.1.1 Ciclo do C
Entende-se por ciclo biogeoquímico do C o conjunto de processos responsáveis por
retirar C de um determinado compartimento (reservatório), fazê-lo participar de compostos e
reações em outros compartimentos e após algum tempo devolvê-lo ao compartimento de origem
(PACHECO e HELENE, 1990). Na Terra, existem vários compartimentos de C e, dentre eles,
os principais são: os oceanos, as formações geológicas contendo carbono fóssil e mineral, os
ecossistemas terrestres (biota + solo) e a atmosfera (SCHLESINGER, 1997; GRACE, 2001).
Desses, o maior compartimento são os oceanos com 38.000 Pg C, seguido do compartimento
presente nas formações geológicas (5.000 Pg C). O carbono das formações geológicas consiste
de 4.000 Pg C presentes no carvão, 500 Pg C no petróleo e 500 Pg C no gás natural (o carbono
estocado nesses combustíveis fósseis não é permutável naturalmente, ocorrendo isto apenas
com a interferência humana). O compartimento de carbono do solo (2.500 Pg C) é o maior nos
ecossistemas terrestres (aproximadamente 4 vezes o compartimento de C da vegetação e 3,3
vezes o carbono da atmosfera) e é constituído pelo carbono orgânico (1.500 Pg C) e mineral
(1.000 Pg C).
Apesar da vegetação constituir um reservatório bastante limitado de carbono, é
responsável pelo fluxo mais intenso no ciclo biogeoquímico. Através da fotossíntese estima-se
que a produção primária sequestre anualmente cerca de 120 Pg C na forma de CO2 da atmosfera,
sendo que 60 Pg C retornam à atmosfera como respiração dos tecidos vivos. Acredita-se que os
outros 60 Pg C também retornem à atmosfera por meio da decomposição da MOS
(SCHLESINGER, 1997).

21
Os estoques de carbono nos solos sob florestas nativas estão em equilíbrio dinâmico
com o carbono na atmosfera, na vegetação e nos oceanos. Esses estoques são determinados
principalmente pelo clima, pelo relevo, pela vegetação e pela textura e composição
mineralógica dos solos, além das interrelações destes, como explícito pelos fatores de formação
dos solos (JENNY, 1994). A magnitude da variação destes estoques sob vegetação nativa é
bastante significativa: regiões de maior latitude, de climas úmidos e sob florestas apresentam
maiores estoques de C, sendo os menores estoques encontrados em regiões de clima árido e
semiárido (POST et al., 1982).
A queima de combustíveis fósseis e o desmatamento de florestas nativas pela queima
para uso agrícola, pastoril e para uso de madeiras, são apontados como os principais
responsáveis pelo crescente aumento de CO2 na atmosfera. As florestas contêm de 20 a 100
vezes mais carbono por unidade de área que as plantações ou pastos. Com o desmatamento o
carbono originalmente contido na vegetação e nos solos é liberado para a atmosfera em forma
de CO2. Somente uma quantidade relativamente pequena de carbono é redistribuída na Terra
ou levada pelos rios (IPCC, 2007).
Estima-se que a queima de combustíveis fósseis represente um incremento de CO2 para
a atmosfera da ordem de 6 Pg C ano-1. Já as emissões de carbono provenientes de mudanças de
uso da terra são de menor magnitude, cerca de 0,9 Pg C ano-1 (VITOUSEK, 1992), porém
sujeitas a consideráveis incertezas e divergências (KREILEMAN e BOUWMAN, 1994).
2.1.2 Ciclo do N
Assim como os demais ciclos biogeoquímicos, o ciclo do nitrogênio também tem seus
reservatórios e seus fluxos e processos pelos quais o nitrogênio troca de reservatório. A
disponibilidade de nitrogênio no ambiente controla o funcionamento de ecossistemas locais e
da biogeoquímica global (SCHLESINGER, 1997). A maior parte do N encontra-se na
atmosfera, na qual ocupa cerca de 78% do volume. Os estoques de nitrogênio orgânico no solo
e na biota terrestre são insignificantes quando comparados ao estoque da atmosfera. Pequenas
quantidades de N são encontradas também nos oceanos, rochas e sedimentos (CHAPIN et al.,
2002).
Embora 78% da atmosfera da Terra sejam compostas por nitrogênio molecular, esse
importante reservatório está indisponível diretamente para animais e plantas. A fixação do
nitrogênio é um processo de redução do nitrogênio atmosférico (N2) a amônia (NH3). A

22
capacidade de fixar nitrogênio é encontrada apenas em alguns seres procariontes (COSTA et
al., 2009).
Pode-se dividir a ciclagem biogeoquímica de N nos ecossistemas em função dos seus
processos, chamados de externos e internos. Os externos acrescentam ou retiram N dos
ecossistemas. Já os processos internos ciclam N entre os estoques dos ecossistemas (SCHIMEL
e BENNETT, 2004). O N pode ingressar no sistema solo-planta por deposições atmosféricas
(através de descargas elétricas), fixação biológica e adubações minerais ou orgânicas. Por outro
lado, pode sair por meio de remoção pelas culturas e variados mecanismos de perdas, que
incluem a lixiviação, a volatilização e a desnitrificação (COSTA et al., 2009). Descargas
elétricas na atmosfera incorporam N ao solo pela transformação do nitrogênio elementar (N2)
em óxidos que são convertidos em ácido nítrico, dissolvido na água das chuvas, resultando em
nitratos aproveitáveis pelas plantas (ODUM, 1983; MATSON et al.,1999). A fixação do N do
ar pode ser mediada por micro-organismos livres do solo e por micro-organismos que vivem
em simbiose com as plantas. Todo N acumulado na planta somente é incorporado ao solo à
medida em que os resíduos de raízes, folhas, galhos, cascas e troncos forem decompostos
(ODUM, 1983; CLEVELAND et al., 1999).
Do N existente no solo, cerca de 95 a 98% encontram-se na forma orgânica, que, para ser
aproveitada pelas plantas, necessita ser transformada para as formas inorgânicas (NH4+ e NO3
-
) pelo processo de mineralização. A formação de NH4+ acontece devido à mineralização da
matéria orgânica por microrganismos heterotróficos em condições redutoras (amonificação), já
o NO3- é formado pela oxidação do NH4
+ através das bactérias aeróbias do gênero
Nitrossomonas e Nitrobacter (nitrificação). O NO3- raramente é adsorvido às partículas do solo,
tem grande mobilidade na solução do solo e pode ser perdido via lixiviação e emissão gasosa.
A absorção pelas plantas depende da densidade radicular e suas reservas podem ser utilizadas
de forma eficiente. Já o NH4+ é adsorvido na argila e na matéria orgânica, possui baixa
mobilidade e célere empobrecimento nas proximidades das raízes. Logo, a utilização de
reservas de NH4+ depende mais do crescimento de densidade radicular que a do NO3
- (SPRENT,
1987).
Antes da interferência mais efetiva do homem no ciclo do N, provavelmente de 90 a 130
milhões de toneladas de N eram fixados biologicamente a cada ano (VITOUSEK et al., 1997).
No último século, as atividades humanas tem duplicado as quantidades de N ciclado entre os
ecossistemas terrestres e a atmosfera. Enquanto a queima de combustíveis fósseis libera para a
atmosfera quantidades de N superiores a 20 Tg a cada ano, a fixação de N2 atmosférico em
fertilizantes aumentou de 10 Tg ano-1, em 1950, para 80 Tg ano-1, em 1990, sendo esperado

23
para o ano 2030 uma fixação maior que 135 Tg ano-1 (VITOUSEK et al., 1997). Como
consequências da alteração no ciclo do N é possível observar: aumento na concentração global
de NO e N2O em decorrência da fertilização de solos; intensificação nos eventos de chuva ácida
e poluição fotoquímica; aumento na produtividade e nos estoques de C em regiões onde N é
limitante; favorecimento de invasão biológica e redução da diversidade; perdas por lixiviação
e eutrofização de corpos d’agua (HALL et al., 1996; RUSSOW et al., 2000).
2.2 A matéria orgânica do solo (MOS)
Dentre os constituintes do solo, a matéria orgânica do solo (MOS), principalmente o
carbono orgânico total (COT), exerce papel fundamental na preservação de sua qualidade e na
manutenção de suas funções ecológicas. A MOS consiste de uma mistura de compostos em
vários estágios de decomposição, que resultam da degradação biológica de resíduos de plantas
e animais, e da atividade sintética de microrganismos (STEVENSON, 1994). Os constituintes
principais da matéria orgânica são o carbono (52 - 58%), oxigênio (34 - 39%), hidrogênio (3,3
- 4,8%) e nitrogênio (3,7 - 4,2%) (SILVA et al., 2004). Devido às relações com outros
componentes do ambiente, a qualidade e a quantidade de MOS são definidas pelo ambiente de
formação, sendo sensível às alterações provocadas no solo. Dessa forma, a remoção da
vegetação nativa para uso agrícola e/ou pastoril da terra e determinadas práticas de manejo do
solo podem contribuir para o decréscimo ou acúmulo de MOS. A perda de quantidade e
qualidade de MOS contribui para a degradação do solo, e a sua recuperação, na maioria dos
casos, está relacionada com a promoção do aumento do conteúdo de MOS (STEVENSON,
1994).
O carbono orgânico é obtido pelas plantas através da fixação fotossintética do CO2 da
atmosfera. O oxigênio e o hidrogênio acompanham o metabolismo do carbono e são obtidos da
água também no processo de fotossíntese das plantas. Porém, o nitrogênio, essencial às plantas,
de modo geral, não faz parte do material de origem do solo. O conteúdo de nitrogênio mineral
encontrado em solução é produto ou da adição de fertilizantes ou de transformações do
nitrogênio orgânico em formas inorgânicas através da mineralização da matéria orgânica, sendo
encontrado em solução e absorvido pelas plantas principalmente na forma mineral, nítrica (NO3-
) ou amoniacal (NH4+) e, excepcionalmente em outras formas orgânicas como ácidos aminados
e vitaminas (MOREIRA e SIQUEIRA, 2003).
Dentro do conceito de qualidade do solo, a matéria orgânica do solo (MOS), tem sido
reconhecida como um dos mais importantes atributos (DORAN e PARKIN, 1994). Este

24
consenso em relação a MOS como indicador da qualidade do solo emana do fato de que seu
teor é muito sensível às práticas de manejo, principalmente nas regiões tropicais e subtropicais,
onde, nos primeiros anos de cultivo, mais de 50% da matéria orgânica previamente acumulada
podem ser perdidos por processos, como a decomposição microbiana e a erosão (PEIXOTO,
2008).
A matéria orgânica é componente imprescindível para a qualidade dos solos e ambientes
tropicais. Ela desempenha um papel significativo na produtividade dos solos, pois representa a
reserva dominante de N, enxofre (S) e fósforo (P) para as plantas. Além disso, ela influencia
diretamente os processos físicos, químicos e biológicos do solo. Fatores como por exemplo o
pH, a capacidade de troca iônica e estrutura do solo (ZECH et al., 1997), também podem
contribuir de forma indireta para o sequestro de C da atmosfera (BAYER et al., 2000;
GLATZELA et al., 2003), podendo ter efeito também na atividade microbiana e na estabilidade
de agregados (BALESDENT et al., 2000, KALBITZ et al., 2003).
2.2.1 Fatores determinantes no estoque de MOS
2.2.1.1 Textura e mineralogia do solo
Para condição semelhantes de clima, vegetação e manejo, solos argilosos apresentam
tendência a possuir maiores estoques de matéria orgânica que solos arenosos. Isso decorre das
maiores área superficial específica e quantidade de cargas nos solos argilosos, o que possibilita
maiores interações organominerais e menores taxas de mineralização (DICK et al., 2009). Em
muitos solos, o conteúdo de argila atua como importante determinante na estabilização da MOS,
e o conteúdo de carbono no solo geralmente está correlacionado com o conteúdo de argila. A
incorporação física da MOS dentro dos agregados de argila resulta na proteção dos polímeros
orgânicos ao ataque enzimático (ANDERSON, 1992). Além da estabilização física, as argilas
também promovem estabilização química da matéria orgânica coloidal à porção mineral do
solo. Como ambas possuem carga, cátions com mais de uma valência, como Ca++, Mg++, Al3+
e Fe3+, podem promover ligações eletrostáticas relativamente estáveis entre a matéria orgânica
e a porção mineral do solo (ANDERSON, 1992).
A proteção física da MOS pela microagregação, de certa forma, está relacionada com a
interação organomineral. Quanto mais intensa, maior é a possibilidade de formação de
microagregados (< 250 µm de diâmetro), cuja estabilidade protege fisicamente a MOS, tanto
por reduzir a difusão de oxigênio em seu interior, reduzindo assim, a atividade microbiana;

25
como por atuar como uma barreira física que dificulta a chegada do micro-organismo ao
substrato orgânico (BALESDENT et al., 2000). A proteção física por microagregação e a
interação organomineral são processos associados, de forma que é difícil separar a ação
individual de cada um na estabilização da MOS. A condição que favorece a interação
organomineral (solo argiloso, elevada área superficial específica e elevada densidade de grupos
funcionais) também favorece a formação dos microagregados. Além disso, a própria MOS
contribui para a estabilização desses microagregados e, em contrapartida, recebe a proteção
física, estabelecendo-se um efeito interativo (DICK et al., 2009).
2.2.1.2 Clima
O clima é um fator ativo de formação do solo, pois atua em processos que causam
mudanças nas características dos solos durante sua gênese (FANNING e FANNING, 1989). A
influência do clima nos processos pedogenéticos pode ser inferida através de características
como a mineralogia, teor e tipo de óxidos de ferro, acúmulo de matéria orgânica no solo e seu
grau de humificação (MARTIN NETO et al., 1998). As condições climáticas, tais como
temperatura e precipitação pluvial média anual, são os fatores mais importantes na
determinação dos níveis de matéria orgânica do solo (STEVENSON, 1994). Em climas frios e
úmidos ocorre maior acúmulo de matéria orgânica que em climas mais quentes e secos, devido
à menor atividade microbiana nestes sítios. Em climas tropicais a matéria orgânica acumula-se
mais no solo conforme aumenta a altitude.
Dalmolin et al. (2006) constataram que não apenas o teor de MOS aumentou com a
altitude, como também a proporção de grupos C-O alquil, que constituem as estruturas mais
lábeis da MOS. O efeito do clima foi mais pronunciado nos solos mais argilosos e ricos em
óxidos de Fe, evidenciando o papel dos complexos organominerais na estabilização da MOS.
Ao avaliar a influência do clima, expresso pela relação temperatura média
anual/precipitação anual, sobre o teor de carbono no solo, Tate (1992) observou um conteúdo
superior no estoque de carbono em ambientes úmidos sob temperaturas baixas e em altitudes
elevadas que em ambientes mais quentes e secos. Analogamente, Kämpf e Schwertmann (1983)
observaram numa climossequência no estado do Rio Grande do Sul, um aumento no teor de
carbono orgânico do solo com o aumento da umidade e a diminuição da temperatura.
A temperatura é o fator determinante na decomposição de resíduos, pois a taxa de perda
de C é bem maior nos períodos mais quentes do ano do que nos períodos mais frios. HOLTZ
(1995) estabelecem para regiões subtropicais a faixa de temperatura de 30 a 40º como a mais

26
favorável ao processo de decomposição. Também COSTA (1995) observou que a maior
decomposição de resíduos ocorreu em rampas que recebiam maior irradiação solar e portanto
tinham maiores temperaturas. Porém o efeito é extremamente complexo devido às interações
com outros fatores climáticos e com os tipos e características do solo e da matéria orgânica
(GREGORICH et al., 1994; HOLTZ, 1995).
Embora as condições climáticas de elevadas temperatura e precipitação possam
potencializar a mineralização da MOS nos trópicos, a interação organomineral pode
contrabalançar esses efeitos climáticos, principalmente quando associada ao maior potencial de
aporte de fitomassa pelos vegetais (SOLLINS et al., 1996).
2.2.1.3 Uso e cobertura da terra
O carbono e o nitrogênio são os principais componentes da matéria orgânica do solo e
os seus estoques variam em função das taxas de adição, por resíduos vegetais e, ou, animais, e
de perda, dentre elas, as decorrentes da erosão e da oxidação pelos microrganismos do solo. Em
solos sem ação antrópica, o teor e o estoque desses elementos são determinados basicamente
pela temperatura, pela umidade e pelo tipo de solo (BAYER e MIELNICZUK, 1997). Em
sistemas agropecuários, os estoques de C orgânico (CO) e de N total (NT) no solo são também
influenciados pelo manejo adotado. Em solos com intenso revolvimento, além das perdas por
erosão, ocorre aumento da atividade microbiana pela maior exposição dos resíduos aos
microrganismos e suas enzimas.
A exploração agropecuária geralmente promove aumento na taxa de oxidação da MOS,
favorecendo as emissões de CO2. Em solos de clima temperado, metade do conteúdo inicial de
C do solo é perdido em um intervalo de 50 a 100 anos de cultivo (FELLER e BEARE, 1997).
Em ambientes tropicais e subtropicais as perdas podem ser ainda maiores. Nos primeiros anos
de cultivo, mais de 50% da matéria orgânica previamente acumulada é perdida por diversos
processos, entre esses, a decomposição microbiana e a erosão (ANDREUX, 1996).
Silva et al. (1994), avaliando o efeito de cinco anos da monocultura de soja com preparo
intensivo do solo no Cerrado baiano, verificaram perdas de matéria orgânica, em relação ao
estoque inicial, na ordem de 80%, 76% e 41% sob Neossolo Quartzarênico, Latossolo de textura
média e Latossolo argiloso, respectivamente. Por outro lado, práticas conservacionistas, como
determinadas técnicas de plantio (plantio direto, cultivo mínimo, rotação de cultura e etc.) e
manejo das pastagens, florestas plantadas e sistemas agroflorestais podem reduzir

27
drasticamente estas perdas, mantendo-se os teores de MOS ou até mesmo aumentando-os
(CARTER, 2001).
A maioria das práticas agrícolas não provoca elevações significativas dos conteúdos de
carbono em pouco tempo, sendo necessário pelo menos entre 25 a 50 anos antes que um novo
equilíbrio seja atingido (BATJES, 2001). Campos (2003), no nordeste do Espírito Santo,
comparou o efeito da conversão de uma área desmatada em 1980 que foi mantida sob pastagem
por 10 anos e foi posteriormente cultivada com cana-de-açúcar por 12 anos, com o de outra área
que foi mantida sob pasto por 22 anos (após desmatamento). Todas as áreas do estudo (mata,
pastagem e cana-de-açúcar) eram adjacentes e com o mesmo tipo de solo (Latossolo, 80% de
areia). O estoque de carbono nos primeiros 100 cm de profundidade na área de mata foi de 62
Mg ha-1, na área de cana-de-açúcar de 50,9 Mg ha-1 e no pasto de 70,8 Mg ha-1. Embora a cana-
de-açúcar tenha sido fertilizada (a soqueira recebeu 80 kg N ha-1), a redução dos estoques de C
provavelmente foi associada ao cultivo intensivo usado no estabelecimento da cultura e nas
duas posteriores renovações em intervalos de cincos anos.
Fujisaka et al. (1998) avaliaram os efeitos da conversão de florestas nativas em áreas
com culturas agrícolas e pastagens nos estoques de carbono do solo em Rondônia. Constataram
redução da ordem de 80% nos estoques C no solo com a transformação, através do corte e
queima da vegetação, para lavouras de culturas anuais e posterior introdução de pastagem.
Silva et al. (2004) avaliaram áreas de pastagens quanto ao armazenamento de C na
profundidade de até 1 m em Latossolos argilosos na região do Cerrado e verificaram, após 4 a
8 anos, que a magnitude do estoque variou entre 97,1 a 113,0 Mg ha-1, concentrando-se nas
camadas superficiais, com cerca de 50% até 40 cm de profundidade.
A maior contribuição das pastagens para o acúmulo de C no solo advém do seu sistema
radicular. Nessas áreas, o teor de MOS é maior nas camadas superficiais e decresce com a
profundidade. Cerca de 40 a 50% do C do solo pode ser armazenado até 30 cm e amostragens
de até 60 cm capturam cerca de 90% da biomassa radicular e do C orgânico do solo de
ecossistemas com pastagens (LAL, 2002).
As informações disponíveis sobre o papel dos solos brasileiros no ciclo biogeoquímico
do carbono e do nitrogênio são ainda escassas, especialmente para os solos do semiárido. Isso
dificulta a realização de estimativas do estoque de C e N, bem como a simulação do impacto de
sistemas de manejo conservacionistas. A maioria dos relatos sobre estoques de C e N em solos
agrícolas brasileiros foram baseados em sistemas de cultivo de culturas anuais para produção
de grãos em condições de alta precipitação pluvial (FREIXO et al., 2002; MACHADO e
SILVA, 2001; SISTI et al., 2004).

28
Tiessen et al. (1998) apresentaram o estoque total de C de um Latossolo sob vegetação
natural na região Nordeste do Brasil como sendo próximo de 20 Mg ha-1, na camada superficial
(0-20 cm). Um Argissolo, na mesma região, apresentou aproximadamente 28 Mg ha-1, entre 0
e 20 cm (KAUFFMAN et al., 1993). Para um Luvissolo Crômico, sob caatinga hiperxerófila
no semiárido cearense, Maia et al. (2007) encontraram estoque de carbono de 48,4 Mg ha-1, na
mesma profundidade. Borges e Kiehl (1996) observaram, em um Latossolo Amarelo Álico na
Bahia, que o estoque total de carbono (100 t ha-1) sob vegetação natural até 100 cm de
profundidade, foi reduzido em torno de 20% com os cultivos de banana, manga e mandioca. O
cultivo de citros resultou numa menor redução do estoque, em torno de 10%.
Amorim (2009) avaliou a variação sazonal dos estoques médios de C em Argissolo sob
vegetação natural, no município de Petrolina, Pernambuco. No período da estação seca, os
valores foram de 16,5; 11,8 e 9,89 Mg ha-1 e após período de maior precipitação pluvial (estação
chuvosa) 14,2; 10,0 e 8,99 Mg ha-1 nas camadas de 0-10, 10-20 e 20-30cm, respectivamente.
Os estoques de carbono do solo diminuem exponencialmente com a profundidade
(MACHADO et al., 2001; SISTI et al., 2004). Em geral, 39-70% do carbono orgânico total na
camada de 0-100 cm de solo mineral estão presentes nos primeiros 30 cm; e 58-81% nos
primeiros 50 cm (BATJES, 1996). Perdas ou aportes de carbono do solo são maiores no
horizonte superficial (0-15 cm), que deve ser amostrado mais frequentemente (RICHTER, et
al., 1999). Entretanto, havendo mudanças no uso e manejo, a amostragem deve ser feita a
profundidades maiores, pois o acúmulo de carbono na superfície pode ser balanceado por perdas
em profundidade (WATSON et al., 2000). Não seria prático, todavia, definir uma profundidade
para todos os solos, onde o carbono seria analisado.
Os teores de N, nas diferentes profundidades do solo, apresentam o mesmo padrão de
distribuição dos teores de C, com os maiores valores observados em camadas mais superficiais,
onde há maior acúmulo de matéria orgânica, sendo, dessa forma, também influenciados pelos
sistemas de cultivos. Em solos agrícolas, os teores de nitrogênio variam, em geral, de 0,02 a
0,5% e, 98% do NT do solo encontram-se na forma orgânica (STEVENSON, 1994). A
relevância da inclusão do N nos estudos que avaliam a MOS reside no fato dos compostos
orgânicos representarem um importante reservatório de formas de N potencialmente
disponíveis para as culturas.
Segundo Stevenson (1994), os compostos nitrogenados representam em torno de 5% da
fração orgânica do solo, e cerca de 95 a 98% desse elemento estão ligados ao carbono dessa
fração. Assim, em virtude dessa estreita relação entre o C e o N no solo, a manutenção do
estoque de C e, por conseguinte, da MOS, é intimamente condicionada à quantidade de N do

29
solo. Portanto, o conteúdo de N orgânico do solo não pode ser acrescido de forma estável, sem
que haja acréscimo correspondente no total de C, e vice-versa.
A MOS apresenta em geral, uma relação C/N bastante estável, variando de 8 a 14,
quando ocorre a estabilização da MOS (húmus). Essa característica está diretamente
relacionada ao tipo de solo, clima e mesmo de vegetação. Áreas sob florestas e cultivos
agrícolas apresentam valores de relação C/N em torno de 12, enquanto em áreas sob pastagem
esse valor pode ser um pouco mais elevado (BATJES, 1996).
2.3 Isótopos estáveis de 13C e 15N no estudo da MOS
Isótopos são átomos do mesmo elemento químico (mesmo número de prótons), mas que
diferem em número de nêutrons, apresentando diferentes massas. O termo isótopo vem do grego
ISOS (mesmo ou igual) e TOPOS (lugar), referindo-se ao fato de que ocupam o mesmo lugar
na tabela periódica. Os isótopos podem ser estáveis, quando não emitem radiação; ou
radioativos, quando emitem algum tipo de radiação. Os isótopos estáveis, em relação aos
radioativos, possuem a vantagem de não apresentarem riscos à saúde humana e contaminação
ao ambiente (BOUTTON et al., 1988).
A substituição de florestas por cultivos agrícolas e/ou pastoris levam a importantes
mudanças na dinâmica do carbono e do nitrogênio do solo, e consequentemente na ciclagem
global desses elementos.
A utilização de traçadores isotópicos naturais de 13C e 15N são métodos bastante
utilizados para distinguir e quantificar a origem da matéria orgânica nos solos cultivados e a
dinâmica do nitrogênio (CADISCH et al., 1996).
2.3.1 Abundância natural de 13C
Na natureza, existem dois isótopos do C que são estáveis e se encontram em proporções
pouco variáveis ao longo do tempo. O 12C é o mais leve e representa a maior proporção em
relação ao total de C na natureza (98,89%); já o 13C conta com apenas 1,1% do total. A pequena
fração restante é do isótopo radioativo 14C. Essas proporções se mantêm relativamente estáveis
em qualquer resíduo orgânico. No entanto, a taxa desses dois isótopos pode variar como
resultado do fracionamento durante processos físicos, químicos e biológicos (BOUTTON,
1991). Quando o C passa de um estado físico-químico para outro, ocorre uma discriminação

30
entre os dois isótopos, ou seja, um isótopo muda de estado preferencialmente, ou mais
rapidamente, em relação ao outro. Isso faz com que ocorra uma variação na casa dos milésimos
na proporção 13C/12C (ALVES et al., 2006).
As análises da composição isotópica de C são feitas medindo-se a relação de 13C/12C
das amostras em relação a um padrão internacional, e o resultado é expresso em termos de
diferença de δ13C em relação ao padrão. O padrão internacional para o carbono é uma rocha
calcária encontrada em uma formação geológica denominada Pee Dee, na Carolina do Norte,
EUA, cuja abreviação é PDB (Pee Dee Belemnite), e tem uma relação molar de 13C/12C (R) de
0,01124. Os desvios em relação ao padrão são conhecidos como unidades δ (letra grega delta
minúscula), e como são valores muito pequenos convencionou-se expressá-los em partes por
mil (‰) (ALVES, et al., 2006).
O valor de δ do padrão é por definição 0‰. Esse padrão pode ser obtido na International
Atomic Energy Agency (IAEA) em Viena, Áustria e no National Institute of Standards and
Technology (NIST) nos Estados Unidos.
Os valores das amostras (vegetal e/ou solo) são expressos contra esse padrão quando as
razões isotópicas são determinadas usando-se um espectrômetro de massa. Porém, esses
padrões são muitas vezes de alto custo para o uso nas análises diárias, assim muitos laboratórios
utilizam seu “padrão interno de trabalho” (UNKOVICH et al., 2001).
A relação ¹³C/¹²C é calculada pela fórmula abaixo:
𝛿13𝐶𝑃𝐷𝐵(‰) = (𝑅𝑎𝑚𝑜𝑠𝑡𝑟𝑎 − 𝑅𝑃𝐷𝐵
𝑅𝑃𝐷𝐵) × 1.000
Onde: Ramostra é a razão isotópica 13C/12C da amostra e RPDB é a razão isotópica 13C/12C do
padrão.
Para usar estas pequenas variações isotópicas em estudos ambientais, a razão 13C/12C da
MOS deve ser medida com alta precisão. Usualmente são feitas análises em amostras gasosas
em espectrômetro de massa (BOUTTON, 1996).
2.3.1.1 Fracionamento isotópico de carbono pelos vegetais
Como resultado das propriedades bioquímicas das enzimas de fixação do CO2
atmosférico (RuBisCo e PEPcase) e das limitações à difusão do CO2 para o interior das folhas,

31
todos os vegetais discriminam contra o 13CO2 durante a fotossíntese, mas de forma diferente de
acordo com seus ciclos fotossintéticos (O’LEARY, 1988). Existem três diferentes tipos de
ciclos fotossintéticos entre os vegetais terrestres: C3, C4 e CAM ou metabolismo ácido das
crassuláceas. Cada um possui um modelo único de discriminação dos isótopos de carbono.
Essas diferenças na composição isotópica dos materiais orgânicos permitem, por exemplo,
traçar o carbono derivado de cada ciclo fotossintético na matéria orgânica acumulada no solo,
sendo este, um bom indicador do tipo de vegetação existente ou que ocorreu em determinada
área.
2.3.1.2 Ciclos fotossintéticos C3, C4 e CAM
O ciclo fotossintético C3 está presente em aproximadamente 85% das espécies de plantas
dominantes em muitos ecossistemas terrestres e nelas o δ13C varia de -30 a -24‰ (PDB), com
média de aproximadamente -27‰ (VOGEL, 1993). Vegetações naturais de florestas tropicais,
de regiões temperadas e de alta altitude são quase exclusivamente C3. Nas plantas C3, o
fracionamento do carbono ocorre nos dois passos do processo fotossintético: difusão/dissolução
e carboxilação. No segundo passo desse processo, a carboxilação pela enzima ribulose-1,5
bifosfato carboxilase (RuBisCo) causa um elevado fracionamento, pois essa enzima tem grande
afinidade por 12CO2, por ser mais leve que o 13C e demandar menos esforço na sua captura,
consequentemente a planta C3 é muito empobrecida em carbono pesado (GANNES et al.,
1998).
Já as plantas C4 usam a via fotossintética Hatch-Slack, em que a enzima de carboxilação
fosfenolpiruvato carboxilase (PEPcase) reduz o CO2 a ácido aspártico ou málico, discriminando
menos contra o 13C. Os valores isotópicos das plantas C4 demonstram que o seu δ13C permanece
aproximadamente em –14‰ (de –7 a –16‰) (LOPES e BENEDITO-CECILIO, 2002).
Espécies C4 representam menos que 5% nas florestas, mas são dominantes em ecossistemas
quentes ricos em gramíneas, como as savanas e as pastagens cultivadas nos trópicos em
plantações de gramíneas como o milho, a cana-de-açúcar e o sorgo (MENDONÇA et al., 2010).
As plantas que utilizam a via metabólica dos ácidos crassulásseos (CAM) possuem as
mesmas enzimas utilizadas na via C4, porém com atividade enzimática diferenciada. Essas
plantas fixam o CO2 durante a noite, usando a PEP-carboxilase com fracionamento,
comumente, encontrado em plantas C4, associado ao de C3, sendo sua variabilidade isotópica
registrada entre –10 e –28‰ (LOPES e BENEDITO-CECILIO, 2002). O ciclo fotossintético
CAM é encontrado em aproximadamente 10% das espécies vegetais e é dominante em

32
ecossistemas áridos ou semiáridos (MENDONÇA et al., 2010). Para fixar CO2, as plantas CAM
mudam do ciclo fotossintético C3 durante o dia para o ciclo fotossintético C4 durante a noite.
Valores de δ13C neste tipo se estendem em todo o alcance das plantas C3 e C4, mas usualmente
são intermediários.
O fracionamento de 13C ocorre na planta não somente pelas diferentes vias
fotossintéticas, mas também, em escala bem menor pelo fracionamento na formação de
moléculas recalcitrantes (lignina) com baixo 13C (PARK e EPSTEIN, 1961). O C mais leve
(12C) é usado preferencialmente pelos decompositores, enriquecendo a MOS em 13C ao longo
da decomposição (BLAIR et al., 1985). A não consideração dessas discriminações pode
conduzir a uma superestimação do C derivado de resíduos de plantas C4, consequentemente
subestimação da contribuição de plantas C3 e da estabilidade da MOS (CLAY et al., 2007).
O primeiro trabalho a utilizar o 13C para quantificar a contribuição de diferentes
vegetações ao carbono do solo no Brasil foi realizado por Cerri et al. (1985). Os autores
avaliaram a contribuição do carbono da cana-de-açúcar (planta C4) e a perda do carbono
derivado de mata, 12 e 50 anos após substituição da mata (ciclo C3). Utilizaram amostras em
fragmentos de mata preservadas como base para o cálculo da substituição de C do solo. O C
na MOS derivado da vegetação nativa, em áreas cultivadas, foi calculado pelo seguinte modelo:
%𝐶𝑂𝑆𝑓 = [(𝛿13𝐶𝑏 − 𝛿13𝐶𝑎)
(𝛿13𝐶𝑓 − 𝛿13𝐶𝑎)] × 100
Onde:
%COSf = o C derivado da floresta nativa (C3); δ 13Ca = a marcação na vegetação C4; δ13Cb = a
marcação atual do solo (C3 + C4); δ 13Cf = a marcação da floresta nativa (C3).
Trabalho semelhante publicado por Balesdent et al. (1987) mostrou uma taxa de
movimentação (“turnover”) de 22% de MOS derivada da mata nativa após 13 anos de
substituição da mata por cultivo de milho.
Balesdent et al. (1988) quantificaram a contribuição, no carbono do solo, do milho (C4)
ao longo de 40 anos de cultivo após retirada de pastagem C3 (pastagem cultivada por 61 anos
em substituição de vegetação nativa C4). Após mais de 100 anos de cultivo, mais de 50% do
COT era ainda derivado da vegetação nativa, correspondendo à fração menos lábil, associada
às frações mais finas do solo, com “turnover” superior a 1.000 anos. Amostras de solo foram
coletadas, em diferentes sucessões de cultivos, em 1915 (solo cultivado com C3 desde 1888),

33
1928, 1938, 1962 (após entrada de milho em 1950) e 1975 em parcelas com cultivos de plantas
C3 (trigo e pastagem) implantadas em 1915.
A aplicação da discriminação isotópica do 13C em estudos de “turnover” de MOS em
áreas que tiveram substituição de vegetação, com padrões fotossintéticos diferenciados, exige
duas situações básicas: tempo razoável de mudança de cultivos e testemunhas confiáveis ou
amostragem de solo antes da substituição do padrão de planta. Esse método, chamado de
mistura simples padrão, foi o primeiro a ser aplicado em estudos de MOS usando discriminação
isotópica. Esses dois trabalhos são até os dias de hoje a base para aplicação de 13C em estudos
de dinâmica de MOS. Ressalte-se que, mesmo sendo os primeiros, são muito recentes
comparando-se com outros temas dentro do estudo da ciência do solo. Quando não existe área
testemunha confiável para utilizar como base, é necessário que existam amostras de solo
coletadas em diferentes períodos, para que se possam acompanhar as alterações na abundância
natural e com isso quantificar a contribuição da nova vegetação para o C do solo.
Campos (2003) avaliou áreas adjacentes de mata e pasto por 22 anos e outras que foram
convertidas no plantio de cana-de-açúcar por 10 anos de pastagem. Após 22 anos de pastagem
houve um aumento significativo no estoque (0-100 cm) de C do solo (8,8 Mg C ha-1) em
comparação com a mata original. Na área que foi pastagem por 10 anos, seguida por 12 de cana-
de-açúcar, no entanto, o estoque de C do solo foi de 50,9 Mg C ha-1, sendo 18% menor do que
aquele sob a mata original. Com os dados de abundância de 13C foi possível avaliar a dinâmica
do C derivado da floresta, de pasto e da cana-de-açúcar nessa cronossequência. Sob a pastagem,
após 20 anos, o C derivado da floresta diminuiu em 9%; mas onde o solo foi intensivamente
revolvido sob a cultura da cana, a quantidade do C derivado da mata original diminuiu em 35%.
A redução no estoque de C da MOS sob a cana-de-açúcar, em comparação com o pasto, ocorreu
pela decomposição da MOS original da mata.
2.3.2 Abundância natural de 15N
Os isótopos estáveis podem prover medidas integradas a respeito da dinâmica de
nitrogênio (δ15N) ao longo do tempo (HANDLEY e HAVEN, 1992; HÖGBERG, 1997). Desta
forma, diminui-se a necessidade de medidas pontuais e repetitivas acerca dos processos que
regulam o ciclo do N. Determinar valores de δ15N em compartimentos chaves de ecossistemas,
como os solos e as plantas, pode permitir uma série de inferências acerca da dinâmica do N
(HÖGBERG, 1997).

34
Há dois isótopos estáveis de N (14N e 15N). A média natural de abundância no ar destes
isótopos é constante de 99,63% de 14N e 0,3663% de 15N. As fontes naturais de 15N variam de
-20 a +20‰. O nitrogênio presente na matéria orgânica do solo apresenta tendência de ser mais
rico em 15N que os tecidos das plantas, sugerindo uma preferência microbiana pelo 14N durante
o processo de decomposição (DAWSON et al., 2002).
Bustamante et al. (2004) entenderam que as grandes diferenças na variação de 15N foliar
encontradas entre as espécies vegetais do Cerrado indicavam que quando as plantas dividem o
mesmo ambiente demandam diferentes fontes de N. Martinelli et al. (1999) avaliaram médias
para o padrão de δ15N do solo em florestas tropicais e encontraram valores altos, assim como
Nardoto (2005) no Cerrado, mas com valores menores que nas florestas tropicais. Tais estudos
sinalizam que o ciclo de N é mais aberto em florestas que nas savanas tropicais.
Quando as leguminosas estão fixando N atmosférico, através da FBN, as suas
assinaturas isotópicas tendem a se aproximar de 0‰; logo, a conversão de N2 em formas
orgânicas de nitrogênio distinguem pouco contra o 15N (EHLERINGER e RUNDEL, 1989). Já
espécies não fixadoras de N apresentam uma grande diferença na razão isotópica, de acordo
com a taxa de mineralização de N no solo, (GARTEN e VAN MIEGROET, 1994). Portanto,
pode-se utilizar o δ15N foliar para determinar se a fonte de N tem maior participação atmosférica
ou do solo.
Apesar das diversas aplicações, a interpretação dos padrões de diferenças de 15N entre
lugares e tipos de solos, ainda estão sujeitas a controvérsias. Várias exceções às tendências
gerais já foram relatadas (AMUNDSON et al., 2003; SWAP et al., 2004; FREITAS et al., 2010)
e a relação entre o δ15N do solo e o clima ainda não é completamente clara (CRAINE et al.
2009). Os sinais isotópicos da maior parte das florestas tropicais ainda são desconhecidos e há
poucas pesquisas de sinais isotópicos ao longo de um gradiente de precipitação nos solos
existentes a curtas distâncias, principalmente, em regiões com temperaturas médias anuais
acima de 23° C (CRAINE et al., 2009; HANDLEY et al., 1999).
Vários estudos têm demostrado que os sinais de 15N do solo tendem a diminuir para zero
com o aumento da pluviosidade (AMUNDSON et al., 2003; NARDOTO et al., 2008; CRAINE
et al., 2009, FREITAS et al., 2015) e que esses sinais são mais altos em florestas tropicais do
que em florestas temperadas (MARTINELLI et al., 1999; AMUNDSON et al., 2003). As
diferenças nos sinais das plantas e do solo também diminuem com o aumento da temperatura e
com a diminuição da precipitação e da latitude (AMUNDSON et al., 2003). Isso ocorre porque
em lugares mais úmidos a abundâcia natural de 15N resulta principalmente da interação da
ciclagem interna do C entre os compartimentos orgânicos vivo e morto, com pequena perda de

35
N. Esse ciclo quase fechado é interrompido nos trópicos e, especialmente nos trópicos secos,
onde proporcionalmente, há maior fluxo de N orgânico para o compartimento inogânico, que é
mais sujeito a perdas, levando ao enriquecimento de 15N dos componentes do solo e da
vegetação (AUSTIN e VITOUSEK, 1988; HANDLEY e SCRIMGEOUR, 1997; HANDLEY
et al., 1999; MARTINELLI et al., 1999; FREITAS et al., 2010; ARANIBAR et al., 2004;
SWAP et al., 2004).

36
3 MATERIAL E MÉTODOS
3.1 Caracterização das áreas de estudo
3.1.1 Localização, limites e área
Os estudos foram conduzido em Pernambuco. O estado está localizado no centro-leste
da região Nordeste do Brasil, entre os paralelos 7º 15’ e 9º 27’ S e os meridianos 34º 48’ e 41º
19’ W (CPRM, 2005) e tem como limites os estados da Paraíba (N), do Ceará (NW), de Alagoas
(SE), da Bahia (S) e do Piauí (W), além de ser banhado pelo oceano Atlântico (L). Pernambuco
ocupa (Figura 1) uma área de 98.311 km² e é constituído por 185 municípios, somando-se a
esses o arquipélago de Fernando de Noronha (IBGE, 2016).
O estado possui uma configuração espacial estreita no sentido norte-sul, apresentando
uma faixa marítima de apenas 187 km de extensão. No sentido leste-oeste alonga-se
Figura 1 - Localização geográfica do estado de Pernambuco e suas
regiões fisiográficas

37
consideravelmente, chegando a 784 km de extensão. Essa projeção para oeste faz com que cerca
de 80% de seu território esteja inserido na região de clima semiárido, onde as chuvas são poucas
e mal distribuídas, ocorrendo, periodicamente, o fenômeno das secas (CONDEPE/FIDEM,
2006).
3.1.2 Regiões fisiográficas e características edafoclimáticas
Pernambuco possui três regiões fisiográficas, Litoral-Mata ou Zona da Mata, Agreste e
Sertão (Figura 1). Estas três regiões são divididas em cinco Mesorregiões geográficas (Região
Metropolitana de Recife, Zona da Mata, Agreste, Sertão e Sertão do São Francisco) e 19
Microrregiões geográficas, onde estão inseridos os 185 municípios do estado (IBGE, 2001;
CONDEPE/FIDEM, 2006).
A Região da Zona da Mata (Litoral-Mata) corresponde à faixa de terra que vai da costa
atlântica aos primeiros contrafortes do Maciço da Borborema. É a menor das três regiões
fisiográficas, apresentando uma área de 8.738 km2 (IBGE, 2014). Apresenta clima tropical
quente e úmido, com temperaturas médias anuais em torno de 24ºC e precipitações pluviais
variando entre 1.200 mm a mais de 2.200 mm anuais. O relevo dessa região é modesto. De uma
planície litorânea, quase ao nível do mar, vai se elevando progressivamente, chegando a
altitudes em torno de 600 m, nas áreas próximas ao Maciço da Borborema. Os solos arenosos
da faixa litorânea dão lugar aos solos argilosos das encostas dos morros e colinas de formas
brandas e arredondadas, principalmente na parte sul. Na parte norte, os tabuleiros da Formação
Barreiras podem ter solos mais arenosos. A rede hidrográfica nessa região apresenta-se perene
e caudalosa drenando extensas áreas em consequência de maiores cotas pluviométricas. O
revestimento vegetal original era representado pela floresta tropical úmida, localmente chamada
de Mata Atlântica, que foi substituída em mais de 95% por cultivos agrícolas, principalmente
cana-de-açúcar (CONDEPE/FIDEM, 2006).
A Região do Agreste é uma área de transição entre a Zona da Mata e o Sertão, com uma
extensão de 24.396 km2 (IBGE, 2014). Essa região está localizada quase inteiramente sobre o
Maciço da Borborema, apresentando climas que vão do tropical úmido da Mata ao semiárido
do Sertão. Em áreas de maiores cotas altimétricas, principalmente nas vertentes expostas aos
ventos alísios de sudeste, surgem os brejos de altitude (porções mais úmidas em relação às áreas
circundantes), verdadeiros microclimas, cuja atividade agrícola é bastante diversificada e que,
como consequência, apresentam maior densidade demográfica. Nas áreas mais secas, a
vegetação predominante era de floresta tropical seca, chamada de caatinga arbórea, em grande
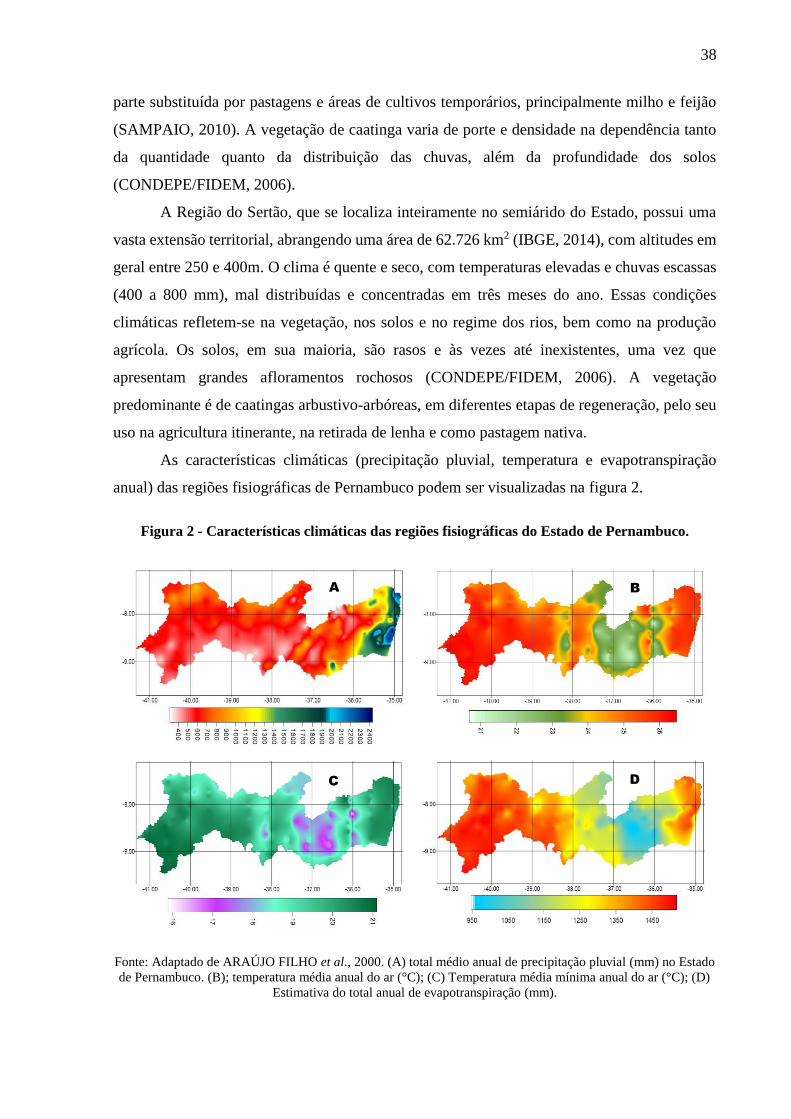
38
parte substituída por pastagens e áreas de cultivos temporários, principalmente milho e feijão
(SAMPAIO, 2010). A vegetação de caatinga varia de porte e densidade na dependência tanto
da quantidade quanto da distribuição das chuvas, além da profundidade dos solos
(CONDEPE/FIDEM, 2006).
A Região do Sertão, que se localiza inteiramente no semiárido do Estado, possui uma
vasta extensão territorial, abrangendo uma área de 62.726 km2 (IBGE, 2014), com altitudes em
geral entre 250 e 400m. O clima é quente e seco, com temperaturas elevadas e chuvas escassas
(400 a 800 mm), mal distribuídas e concentradas em três meses do ano. Essas condições
climáticas refletem-se na vegetação, nos solos e no regime dos rios, bem como na produção
agrícola. Os solos, em sua maioria, são rasos e às vezes até inexistentes, uma vez que
apresentam grandes afloramentos rochosos (CONDEPE/FIDEM, 2006). A vegetação
predominante é de caatingas arbustivo-arbóreas, em diferentes etapas de regeneração, pelo seu
uso na agricultura itinerante, na retirada de lenha e como pastagem nativa.
As características climáticas (precipitação pluvial, temperatura e evapotranspiração
anual) das regiões fisiográficas de Pernambuco podem ser visualizadas na figura 2.
Figura 2 - Características climáticas das regiões fisiográficas do Estado de Pernambuco.
Fonte: Adaptado de ARAÚJO FILHO et al., 2000. (A) total médio anual de precipitação pluvial (mm) no Estado
de Pernambuco. (B); temperatura média anual do ar (°C); (C) Temperatura média mínima anual do ar (°C); (D)
Estimativa do total anual de evapotranspiração (mm).
A B
C D
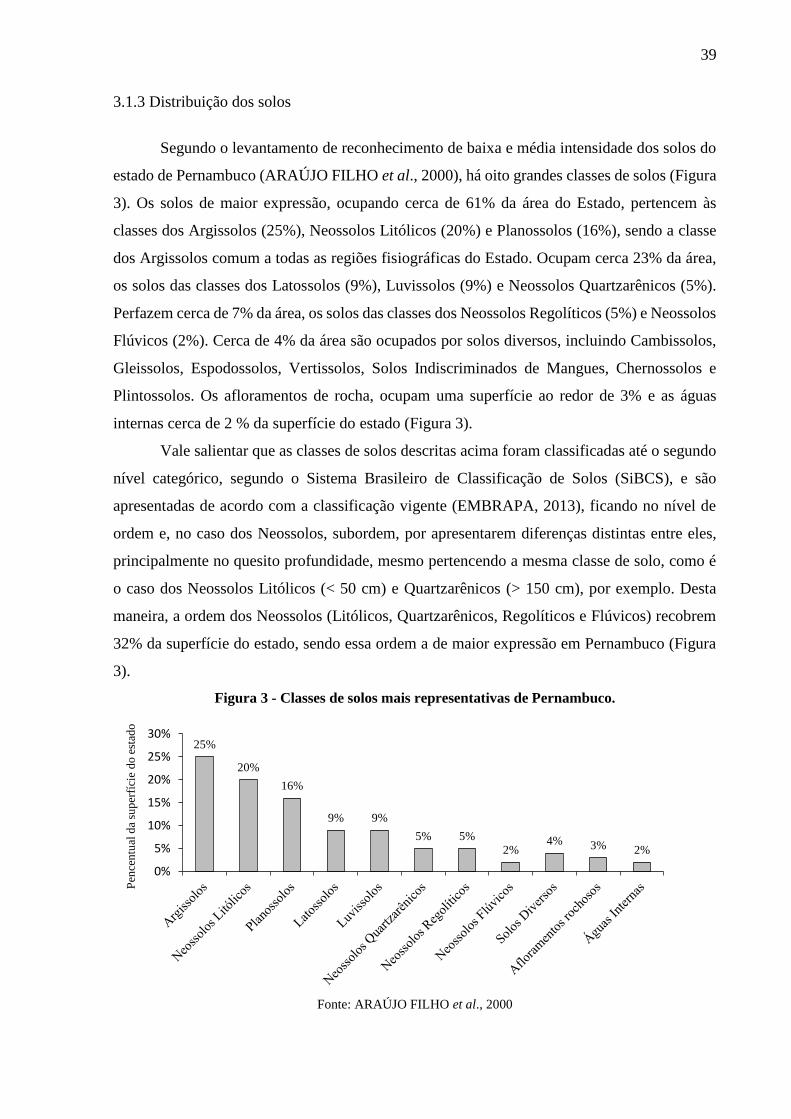
39
3.1.3 Distribuição dos solos
Segundo o levantamento de reconhecimento de baixa e média intensidade dos solos do
estado de Pernambuco (ARAÚJO FILHO et al., 2000), há oito grandes classes de solos (Figura
3). Os solos de maior expressão, ocupando cerca de 61% da área do Estado, pertencem às
classes dos Argissolos (25%), Neossolos Litólicos (20%) e Planossolos (16%), sendo a classe
dos Argissolos comum a todas as regiões fisiográficas do Estado. Ocupam cerca 23% da área,
os solos das classes dos Latossolos (9%), Luvissolos (9%) e Neossolos Quartzarênicos (5%).
Perfazem cerca de 7% da área, os solos das classes dos Neossolos Regolíticos (5%) e Neossolos
Flúvicos (2%). Cerca de 4% da área são ocupados por solos diversos, incluindo Cambissolos,
Gleissolos, Espodossolos, Vertissolos, Solos Indiscriminados de Mangues, Chernossolos e
Plintossolos. Os afloramentos de rocha, ocupam uma superfície ao redor de 3% e as águas
internas cerca de 2 % da superfície do estado (Figura 3).
Vale salientar que as classes de solos descritas acima foram classificadas até o segundo
nível categórico, segundo o Sistema Brasileiro de Classificação de Solos (SiBCS), e são
apresentadas de acordo com a classificação vigente (EMBRAPA, 2013), ficando no nível de
ordem e, no caso dos Neossolos, subordem, por apresentarem diferenças distintas entre eles,
principalmente no quesito profundidade, mesmo pertencendo a mesma classe de solo, como é
o caso dos Neossolos Litólicos (< 50 cm) e Quartzarênicos (> 150 cm), por exemplo. Desta
maneira, a ordem dos Neossolos (Litólicos, Quartzarênicos, Regolíticos e Flúvicos) recobrem
32% da superfície do estado, sendo essa ordem a de maior expressão em Pernambuco (Figura
3).
Figura 3 - Classes de solos mais representativas de Pernambuco.
Fonte: ARAÚJO FILHO et al., 2000
25%
20%
16%
9% 9%
5% 5%
2%4% 3% 2%
0%
5%
10%
15%
20%
25%
30%
Pen
cen
tual
da
sup
erfí
cie
do
est
ado

40
Na mesorregião da Zona da Mata predominam Argissolos (51%), Latossolos (19%) e
Gleissolos (9%); no Agreste, Neossolos Litólicos (25%), Planossolos (24%), Argissolos (20%)
e Neossolos Regolíticos (12 %); no Sertão, Neossolos Litólicos (22%), Planossolos (15%),
Neossolos Quartzarênicos (14%), Luvissolos (13%), Argissolos (12%) e Latossolos (9%). As
principais características desses solos estão descritas na tabela 1.
Tabela 1 - Características das principais classes de solos presentes no estado de Pernambuco.
Classes Características
Argissolos
Solos bem desenvolvidos, com presença de uma camada subsuperficial mais
argilosa, chamada de horizonte B textural. São pouco a muito profundos, com baixa
a média fertilidade. Ocorrem em áreas de relevo desde plano a muito acidentado.
Latossolos
Solos bem desenvolvidos, profundos, ácidos e pobres em nutrientes, com
estrutura granular, porosos, bem drenados, de textura variando de arenosa até muito
argilosa.
Luvissolos
Solos que variam de bem a imperfeitamente drenados, sendo normalmente
pouco profundos (60 a 120cm), com sequência de horizontes A, Bt e C, e nítida
diferenciação entre os horizontes A e Bt, devido ao contraste de textura, cor e/ou
estrutura entre eles. Podem ou não apresentar pedregosidade na parte superficial e
caráter solódico ou sódico, na parte subsuperficial.
Neossolos
Flúvicos
Solos pouco evoluídos, formados por sobreposição de camadas de
sedimentos aluviais recentes sem relações pedogenéticas entre os estratos. São solos
que apresentam um horizonte diagnóstico de superfície A e abaixo deste
normalmente havendo uma sucessão de camadas estratificadas C de natureza não
hidromórfica. Ocorrem normalmente nos ambientes de várzeas, planícies fluviais e
terraços aluvionares.
Neossolos
Litólicos
Solos muito pouco evoluídos, sendo muito rasos, com profundidades inferiores
a 50 cm. Apresentam grandes quantidades de minerais primários em relação às
outras classes, ou seja, possuem reserva de nutrientes, mas eles estão muito pouco
disponíveis às plantas.
Neossolos
Quartzarênicos
São solos pouco evoluídos, muito arenosos, excessivamente drenados, com
baixa capacidade de retenção de água e de nutrientes. São solos de baixa fertilidade
natural e ocorrem em relevo plano a suave ondulado.
Neossolos
Regolíticos
Solos pouco desenvolvidos, pouco profundos a profundos, de textura
normalmente arenosa (cascalhenta ou não). Às vezes apresentam fragipã, uma
camada endurecida, que dependendo da profundidade pode vir a ser uma limitação
para o uso agrícola. Possuem baixa fertilidade natural.
Planossolos
Solos medianamente desenvolvidos, rasos a pouco profundos, que possuem um
horizonte B bastante argiloso, com uma transição textural abrupta entre os horizontes
A e B. Essa transição abrupta resulta numa grande deficiência de drenagem. Bastante
susceptíveis à erosão. Possuem média fertilidade natural, sendo ácidos na superfície
e neutros na subsuperfície. Grande parte desses solos possui alta saturação por sódio,
conferindo-lhes característica de solos sódicos ou solódicos.
Fontes: ARAÚJO FILHO et al., 2000; ARAÚJO et al. (2008); EMBRAPA, 2013; IBGE, 2015.

41
3.2 Estudo 1 - Levantamento dos estoques de carbono nos solos de Pernambuco.
O estudo 1 refere-se ao levantamento dos estoques de C nos solos de Pernambuco,
realizado através de uma ampla revisão de trabalhos publicados na literatura especializada, em
boletins exploratórios de levantamentos de solos e em periódicos científicos.
3.2.1 Estimativas dos estoques de C do solo
Cálculos dos estoques de carbono do solo dependem da disponibilidade de dados sobre
concentrações de carbono (g C kg-1 solo) e da densidade do solo (Ds) das camadas ou horizontes
para as quais se deseja calcular os estoques:
Est. C = COT x Ds x Espessura
onde, Est. C é o estoque de C no solo em Mg ha-1, COT é o carbono orgânico total em
(g kg-1), e Espessura é espessura da camada ou horizonte em cm. Os valores de Ds são
necessários para converter as concentrações de carbono expressas em porcentagem do peso
seco, para peso de carbono por unidade de área (HOWARD et al., 1995). Entretanto, este dado
analítico está ausente em muitos trabalhos que contém dados de C orgânico, principalmente nos
boletins de levantamentos de solos (JACOMINE et al., 1973).
Um conjunto de dados de perfis de solo foi organizado a partir de trabalhos publicados,
nos quais as concentrações de C do solo foram quantificadas (Tabela 2). Esses dados
compreenderam 226 perfis (669 horizontes), coletados em 12 das 13 classes de solos existentes
no atual Sistema Brasileiro de Classificação de Solos (EMBRAPA, 2013), distribuídas por todo
território de Pernambuco, em áreas de vegetação natural e antropizada, áreas de pastagem e em
áreas de agricultura, entre os anos de 1973 e 2012.

42
Tabela 2 - Trabalhos utilizados para o levantamento dos estoques de C dos solos de
Pernambuco.
Conjunto de dados Perfis Fontes
Artigos científicos de 1997
a 2012 88
ANTUNES et al., 2010; ARAÚJO e SALCEDO,
1997; ARAÚJO et al., 2001; ARAÚJO et al., 2004;
BONFIM et al., 2004; GALINDO et al., 2008;
GIONGO et al., 2011; OLIVEIRA et al., 2003;
OLIVEIRA et al., 2009; SANTOS et al., 2012;
SIMÕES NETO et al., 2009; SIMÕES NETO et al.,
2012; SOUZA et al., 2010.
Solos de Referência de
Pernambuco 35 RIBEIRO et al., 1999
Levantamento exploratório
- Reconhecimento de solos
do Estado de Pernambuco
*103 JACOMINE et al., 1973
* Perfis sem valores de DS.
Os diferentes usos e coberturas da terra relatados nos trabalhos foram agrupados em
quatro tipos principais: Mata (Mata Atlântica), Caatinga, pastagem (áreas de pasto) e agricultura
(lavouras). As áreas de Mata e Caatinga foram ainda subdivididas em densa (vegetação natural)
e abertas (área antropizada). Para as áreas de pastagem, consideraram-se tanto as áreas de pastos
nativos quanto às áreas de pasto natural. É importante ressaltar que não há uma distinção clara
entre vegetação natural e pastos nativos, ambos correspondendo principalmente à vegetação de
Caatinga, quase toda ela ocupada eventualmente com rebanhos domésticos (SAMPAIO e
COSTA 2011). Admite-se que as Matas sejam vegetação de maior porte, mais alta, mais
fechada e mais arbórea, que os pastos nativos.
Os solos descritos nos artigos científicos e levantamentos foram classificados até o
segundo nível categórico, segundo o Sistema Brasileiro de Classificação de Solos, e são
apresentados de acordo com a classificação vigente (EMBRAPA, 2013), ficando no nível de
ordem e, no caso dos Neossolos, subordem, por apresentarem diferenças distintas entre eles. As
médias por uso e cobertura da terra foram obtidas identificando-se, para cada perfil, o uso e
cobertura em que ele ocorria.
De 669 horizontes (226 perfis contendo valores de carbono, quantificados
principalmente por combustão via úmida), apenas 361 horizontes (123 perfis) foram descritos
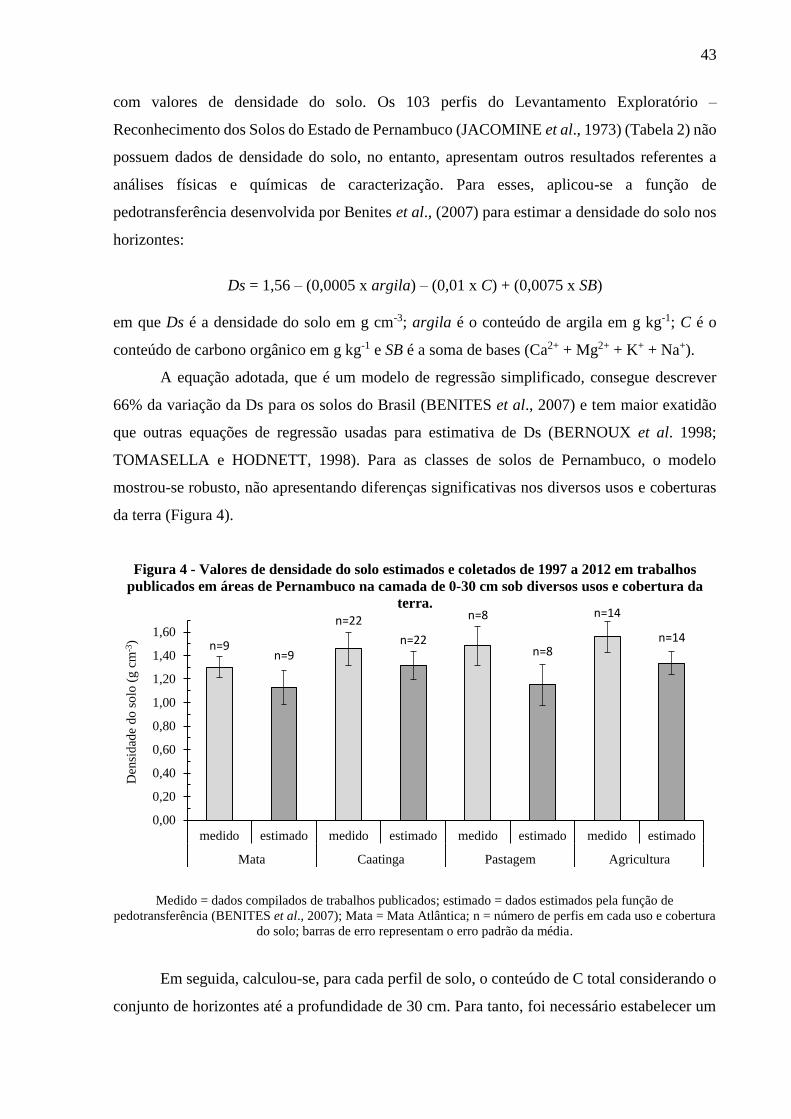
43
com valores de densidade do solo. Os 103 perfis do Levantamento Exploratório –
Reconhecimento dos Solos do Estado de Pernambuco (JACOMINE et al., 1973) (Tabela 2) não
possuem dados de densidade do solo, no entanto, apresentam outros resultados referentes a
análises físicas e químicas de caracterização. Para esses, aplicou-se a função de
pedotransferência desenvolvida por Benites et al., (2007) para estimar a densidade do solo nos
horizontes:
Ds = 1,56 – (0,0005 x argila) – (0,01 x C) + (0,0075 x SB)
em que Ds é a densidade do solo em g cm-3; argila é o conteúdo de argila em g kg-1; C é o
conteúdo de carbono orgânico em g kg-1 e SB é a soma de bases (Ca2+ + Mg2+ + K+ + Na+).
A equação adotada, que é um modelo de regressão simplificado, consegue descrever
66% da variação da Ds para os solos do Brasil (BENITES et al., 2007) e tem maior exatidão
que outras equações de regressão usadas para estimativa de Ds (BERNOUX et al. 1998;
TOMASELLA e HODNETT, 1998). Para as classes de solos de Pernambuco, o modelo
mostrou-se robusto, não apresentando diferenças significativas nos diversos usos e coberturas
da terra (Figura 4).
Medido = dados compilados de trabalhos publicados; estimado = dados estimados pela função de
pedotransferência (BENITES et al., 2007); Mata = Mata Atlântica; n = número de perfis em cada uso e cobertura
do solo; barras de erro representam o erro padrão da média.
Em seguida, calculou-se, para cada perfil de solo, o conteúdo de C total considerando o
conjunto de horizontes até a profundidade de 30 cm. Para tanto, foi necessário estabelecer um
n=9n=9
n=22
n=22
n=8
n=8
n=14
n=14
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
medido estimado medido estimado medido estimado medido estimado
Mata Caatinga Pastagem Agricultura
Den
sid
ade
do
so
lo (
g c
m-3
)
Figura 4 - Valores de densidade do solo estimados e coletados de 1997 a 2012 em trabalhos
publicados em áreas de Pernambuco na camada de 0-30 cm sob diversos usos e cobertura da
terra.
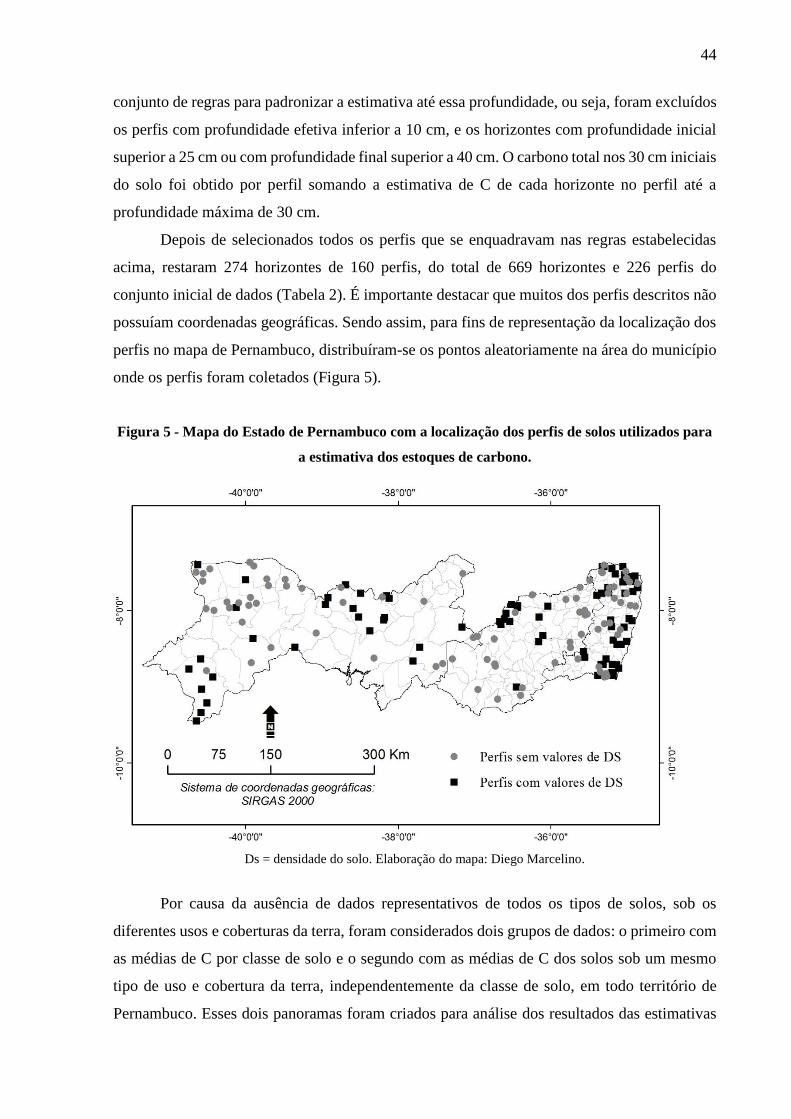
44
conjunto de regras para padronizar a estimativa até essa profundidade, ou seja, foram excluídos
os perfis com profundidade efetiva inferior a 10 cm, e os horizontes com profundidade inicial
superior a 25 cm ou com profundidade final superior a 40 cm. O carbono total nos 30 cm iniciais
do solo foi obtido por perfil somando a estimativa de C de cada horizonte no perfil até a
profundidade máxima de 30 cm.
Depois de selecionados todos os perfis que se enquadravam nas regras estabelecidas
acima, restaram 274 horizontes de 160 perfis, do total de 669 horizontes e 226 perfis do
conjunto inicial de dados (Tabela 2). É importante destacar que muitos dos perfis descritos não
possuíam coordenadas geográficas. Sendo assim, para fins de representação da localização dos
perfis no mapa de Pernambuco, distribuíram-se os pontos aleatoriamente na área do município
onde os perfis foram coletados (Figura 5).
Figura 5 - Mapa do Estado de Pernambuco com a localização dos perfis de solos utilizados para
a estimativa dos estoques de carbono.
Ds = densidade do solo. Elaboração do mapa: Diego Marcelino.
Por causa da ausência de dados representativos de todos os tipos de solos, sob os
diferentes usos e coberturas da terra, foram considerados dois grupos de dados: o primeiro com
as médias de C por classe de solo e o segundo com as médias de C dos solos sob um mesmo
tipo de uso e cobertura da terra, independentemente da classe de solo, em todo território de
Pernambuco. Esses dois panoramas foram criados para análise dos resultados das estimativas

45
da quantidade de C no solo, considerando-se alguns desses fatores isoladamente e, assim, obter
amostras com um maior número de perfis.
3.3 Estudo 2 - Distribuição em profundidade e estoques de carbono em solos sob diferentes
coberturas e usos da terra em Pernambuco.
Considerando a importância e a carência de estimativas dos estoques de carbono nos
solos no Nordeste do Brasil, e como ela varia com os usos da terra, as condições ambientais e
os tipos de solo, esse trabalho objetivou quantificar os estoques de carbono no solo de vegetação
nativa, cultivos e pastagens, in situ, nas principais regiões climáticas e classes de solo de
Pernambuco.
3.3.1 Delineamento da área de estudo
Para o levantamento dos estoques de C do solo, fez-se necessário à divisão do estado de
Pernambuco em regiões fisiográficas (Figura 1) (IBGE, 2001), levando-se em consideração as
principais classes de solo (EMBRAPA, 2006) presentes em cada região, bem como o tipo de
uso e cobertura vegetal existente. Assume-se que essa estratificação em três níveis hierárquicos
(região climática, tipo de solo e cobertura e uso da terra) garante que os principais fatores que
interferem nos estoques e na qualidade da MOS sejam avaliados.
Ao todo, foram amostrados 180 pontos (Figura 6), localizados em áreas representativas
distribuídas em função das três grandes regiões fisiográficas do Estado (Zona da Mata, úmida;
Agreste, subúmido e Sertão, semiárido), sendo esse último dividido em porção leste e porção
oeste, em função das diferenças nos tipos de solo, perfazendo ao todo, quatro regiões de coleta,
cobrindo todo o Estado de Pernambuco.

46
Em cada uma destas quatro regiões foram alocados pontos em função dos quatro tipos
principais de usos e coberturas da terra (vegetação nativa densa e aberta, áreas agrícolas e áreas
de pastagem) e das principais classes de solo, de forma que estas abrangessem mais de 90% e
70% da área de cada região, para os principais usos e solos, respectivamente. Em cada região,
para cada combinação de cobertura e solo foram escolhidos três pontos amostrais, tomados
como repetições.
Na região fisiográfica da Zona da Mata, foram realizadas coletas em dois tipos de solo
e em quatro tipos de uso e cobertura da terra, com três repetições cada, perfazendo um total de
24 situações ou extratos (Tabela 3). Para a região do Agreste foram 48 pontos amostrados (4
classes de solos x 4 tipos de uso e cobertura x 3 repetições); no Sertão Leste foram 60 pontos
(5 tipos de solo x 4 tipos de uso e cobertura x 3 repetições) e no Sertão Oeste 48 pontos amostrais
(4 tipos de solo x 4 tipos de uso e cobertura x 3 repetições).
Figura 6 - Distribuição dos pontos amostrais no estado de Pernambuco.
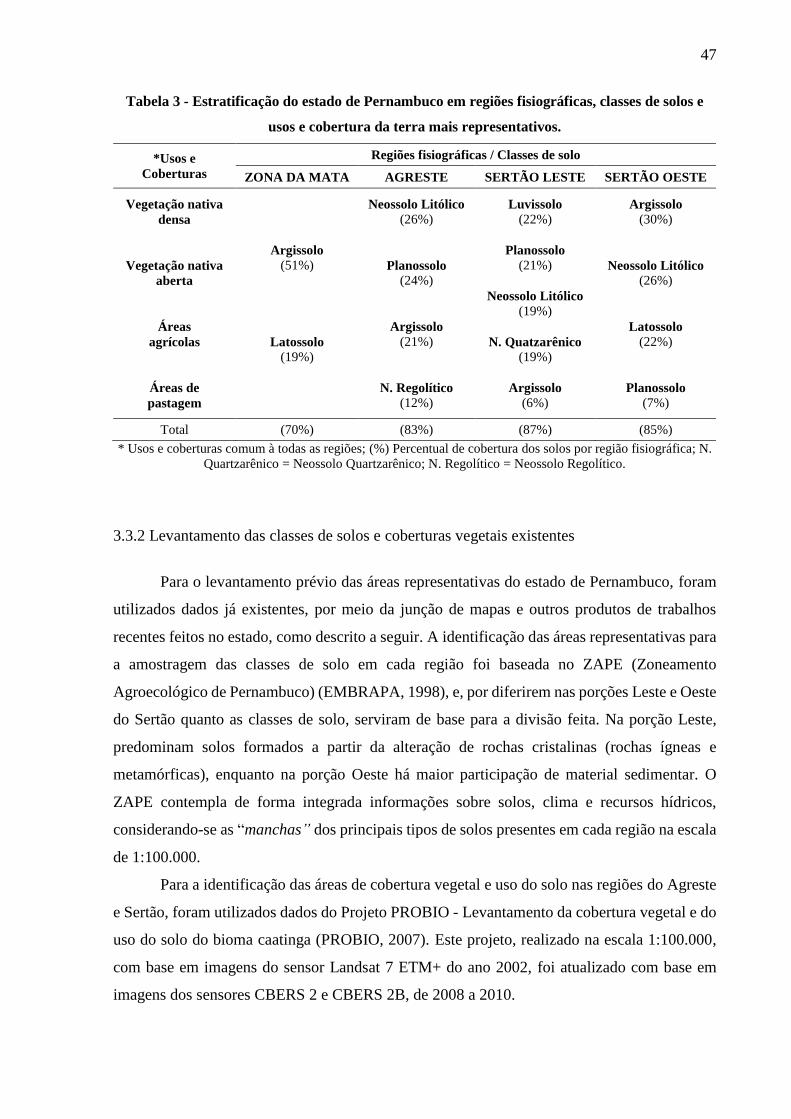
47
Tabela 3 - Estratificação do estado de Pernambuco em regiões fisiográficas, classes de solos e
usos e cobertura da terra mais representativos.
*Usos e
Coberturas
Regiões fisiográficas / Classes de solo
ZONA DA MATA AGRESTE SERTÃO LESTE SERTÃO OESTE
Vegetação nativa
densa
Vegetação nativa
aberta
Áreas
agrícolas
Áreas de
pastagem
Argissolo
(51%)
Latossolo
(19%)
Neossolo Litólico
(26%)
Planossolo
(24%)
Argissolo
(21%)
N. Regolítico (12%)
Luvissolo
(22%)
Planossolo
(21%)
Neossolo Litólico
(19%)
N. Quatzarênico (19%)
Argissolo
(6%)
Argissolo
(30%)
Neossolo Litólico
(26%)
Latossolo
(22%)
Planossolo
(7%)
Total (70%) (83%) (87%) (85%)
* Usos e coberturas comum à todas as regiões; (%) Percentual de cobertura dos solos por região fisiográfica; N.
Quartzarênico = Neossolo Quartzarênico; N. Regolítico = Neossolo Regolítico.
3.3.2 Levantamento das classes de solos e coberturas vegetais existentes
Para o levantamento prévio das áreas representativas do estado de Pernambuco, foram
utilizados dados já existentes, por meio da junção de mapas e outros produtos de trabalhos
recentes feitos no estado, como descrito a seguir. A identificação das áreas representativas para
a amostragem das classes de solo em cada região foi baseada no ZAPE (Zoneamento
Agroecológico de Pernambuco) (EMBRAPA, 1998), e, por diferirem nas porções Leste e Oeste
do Sertão quanto as classes de solo, serviram de base para a divisão feita. Na porção Leste,
predominam solos formados a partir da alteração de rochas cristalinas (rochas ígneas e
metamórficas), enquanto na porção Oeste há maior participação de material sedimentar. O
ZAPE contempla de forma integrada informações sobre solos, clima e recursos hídricos,
considerando-se as “manchas” dos principais tipos de solos presentes em cada região na escala
de 1:100.000.
Para a identificação das áreas de cobertura vegetal e uso do solo nas regiões do Agreste
e Sertão, foram utilizados dados do Projeto PROBIO - Levantamento da cobertura vegetal e do
uso do solo do bioma caatinga (PROBIO, 2007). Este projeto, realizado na escala 1:100.000,
com base em imagens do sensor Landsat 7 ETM+ do ano 2002, foi atualizado com base em
imagens dos sensores CBERS 2 e CBERS 2B, de 2008 a 2010.

48
O uso e a cobertura das terras do bioma Mata Atlântica (Zona da Mata), foi realizada
com base em dados primários utilizando-se, também, as imagens mais recentes dos sensores da
série CBERS. A geração das classes de uso (matas densas e abertas, caatingas densas e abertas,
agricultura e pastagem) foi feita pela interpretação visual considerando as seguintes
características: tonalidade, textura, cor e forma, com base em imagens da época chuvosa e da
época seca (ACCIOLY et al., 2017, no prelo), (Figura 7).
Fonte: equipe de Geoprocessamento da Embrapa Solos UEP Recife, PE.
Após o cruzamento das informações dos solos e dos usos e coberturas da terra, através
dos mapas e das imagens de satélite, fez-se necessária à validação dos pontos amostrais a nível
de campo. Com as coordenadas pré-estabelecidas, realizou-se então a validação, verificando-se
se os pontos pré-estabelecidos correspondiam à combinação prevista (solo x vegetação). Caso
não correspondessem, um novo ponto amostral era procurado. A escolha das áreas amostrais,
bem como a validação das áreas representativas, foram feitas com o apoio da equipe de
Geoprocessamento, da Embrapa Solos UEP Recife, como também com o apoio de pedólogos
da Embrapa Semiárido (CPTSA).
No reconhecimento das fitofisionomias abertas e densas (Tabelas 3 e 4), foram
utilizadas as cenas do sensor ccd1xs do satélite CBERS 2B, para a imagem da época seca e o
sensor LIS3 do satélite IRS P6 ou Resourcesat-1, para a época chuvosa. Para o sensor ccd1xs
(CBERS 2B) foram assinaladas às bandas multiespectral 2 (520 – 590 nm), 3 (630 – 690 nm) e
4 (770 – 890 nm). Para o sensor LIS3 foram assinaladas as bandas: 1 (520 – 590 nm), 2 (620 –
680 nm) e 3 (770 – 860 nm) (ACCIOLY et al., 2017, no prelo).
Figura 7 - Imagem com a demarcação dos diferentes usos da terra em um Latossolo.

49
Tabela 4 - Características dos principais sistemas de uso e cobertura da terra nas diferentes
regiões fisiográficas e classes de solos do estado de Pernambuco.
Usos e coberturas Características
VEGETAÇÃO
NATIVA
Densa
Zona úmida (Zona da Mata): Remanescente de mata nativa dos tipos
florestas perenifólias, subperenifólias e subcaducifólias, com vegetação densa,
sem qualquer exploração ou interferência antrópica visível.
Zona subúmida e semiárida (Agreste e Sertão): Remanescente de caatinga
nativa dos tipos hipoxerófilas e/ou hiperxerófilas densas, sem qualquer
exploração ou interferência antrópica visível.
Aberta
Zona úmida: Remanescente de mata nativa dos tipos florestas perenifólias,
subperenifólias e subcaducifólias, pouco densas e/ou abertas, devido as suas
características naturais e/ou interferência antrópica.
Zona subúmida e semiárida: Remanescente de caatinga nativa dos tipos
hipoxerófilas e hiperxerófilas pouco densas e/ou abertas, devido as suas
características naturais e/ou interferência antrópica.
ÁREAS
AGRÍCOLAS
Na região úmida predomina o cultivo da cana-de-açúcar em plantio
convencional, com o preparo do solo realizado com aração e gradagens. Nesse
sistema, o controle das plantas daninhas é feito com a aplicação de herbicidas
e/ou capina manual e adubação é realizada com fertilizantes “químicos”. Nas
regiões subúmida e semiárida, predominam a agricultura de subsistência com
cultivos de milho, feijão, mandioca e palma (para alimentação animal). A
adubação com fertilizantes “químicos” nesse sistema é praticamente inexistente
e as culturas podem ser adubadas com esterco de curral.
ÁREAS DE
PASTAGEM
Em todas as regiões fisiográficas, a grande maioria das pastagens é de
Brachiaria spp., exceto os sertões, e/ou pastagem natural, conduzidas sob pastejo
contínuo de bovinos e/ou caprinos em regime intensivo, semi-intensivo ou
extensivo e na maioria dos casos sem aplicação de fertilizantes.
Fontes: JACOMINE et al., 1973; CONDEPE/FIDEM, 2006
3.3.2.1 Usos e coberturas da terra
O mapa de uso e cobertura das terras na Escala 1:100.000 (Figura 8) foi elaborado em
parceria com a EMBRAPA Solos UEP Recife, através da classificação visual de imagens dos
sensores Landsat 5 TM e Landsat 8 LDCM pela composição colorida (RGB) das bandas 5, 4 e
3 para a imagem do sensor Landsat 8 LDCM e pelas bandas 4, 3 e 2 do sensor Landsat 5 TM
para o ano de 2013 (ACCIOLY et al., 2017, no prelo).
Na etapa de pré-processamento foram feitas as correções geométricas, a calibração
radiométrica com correção atmosférica e a segmentação das imagens. A imagem do sensor
Landsat 8 LDCM foi geometricamente corrigida (registrada) utilizando-se como referência
pontos de controle obtidos em imagens de alta resolução espacial, disponíveis no software
Google Earth. Após esse registro, a imagem do sensor Landsat 5 TM foi corregistrada à imagem
do Landsat 8 LDCM (ACCIOLY et al., 2017, no prelo).

50
Fonte: EMBRAPA Solos UEP Recife e equipe de geoprocessamento do grupo de Pesquisa da Energia da
Biomassa – UFPE (ACCIOLY et al., 2017, no prelo).
Os resultados (Tabela 5) mostram que cerca de metade da área de Pernambuco é coberta
por caatingas densas e abertas, em proporções semelhantes (28 e 23%, respectivamente).
Naturalmente, estas caatingas estão na zona semiárida, principalmente nos Sertões Leste e
Oeste (Figuras 7 e 8) e, como a porção semiárida corresponde a cerca de 90% do estado, as
caatingas ocupam pouco menos de dois terços desta porção. Ainda na zona semiárida, há cerca
de 16% da área estadual ocupada com agricultura, principalmente com culturas de ciclo curto,
em roças de milho, feijão, algodão herbáceo, mandioca e outras intercaladas (jerimum,
melancia, maxixe, etc.). Grande parte desta área está na Zona do Agreste, onde resta menos
vegetação nativa. Apenas 1% é de agricultura irrigada, incluindo culturas permanentes (manga,
uva, coco, goiaba e outras), concentradas na área próxima do rio São Francisco, no polo de
Petrolina. As áreas de pastagem na região semiárida (separadas das da Zona da mata),
principalmente no Agreste, ocupam 16% da área estadual. Portanto, na região semiárida, os
usos e coberturas previstos para amostragem no planejamento da pesquisa mostraram-se
adequados, cobrindo mais de 90% do território regional (ACCIOLY et al., 2017, no prelo).
Figura 8 - Uso e cobertura da terra em Pernambuco.
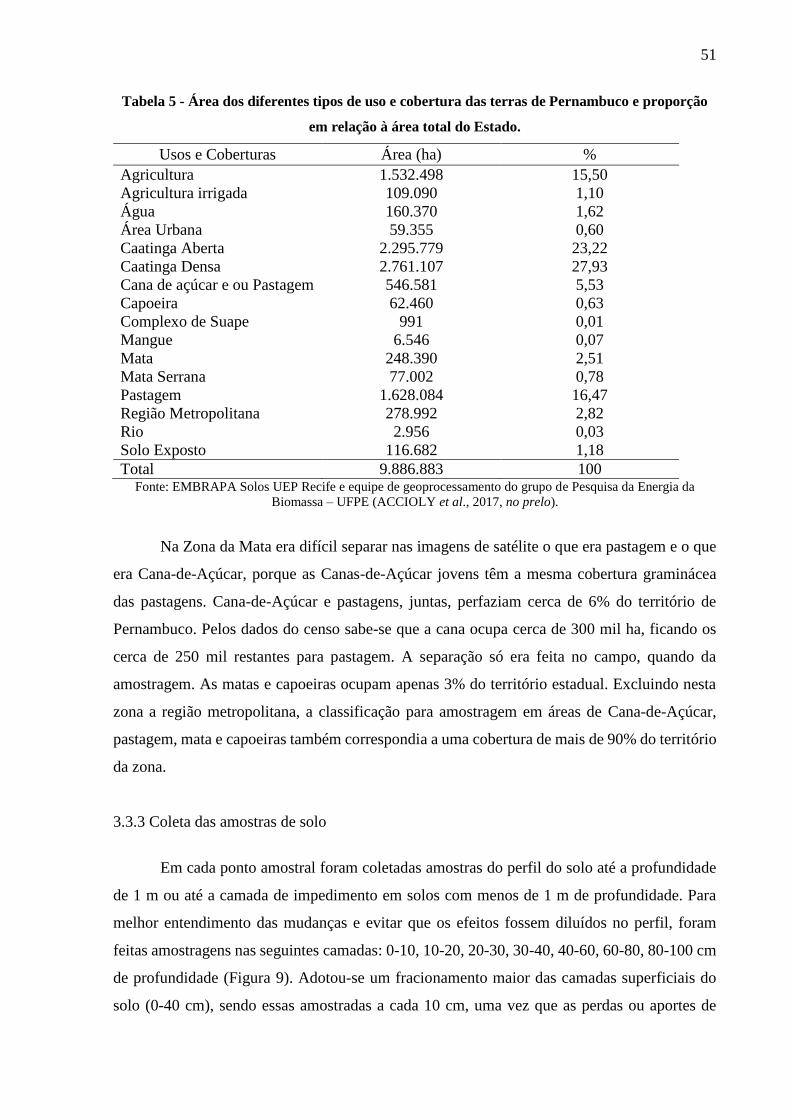
51
Tabela 5 - Área dos diferentes tipos de uso e cobertura das terras de Pernambuco e proporção
em relação à área total do Estado.
Usos e Coberturas Área (ha) %
Agricultura 1.532.498 15,50
Agricultura irrigada 109.090 1,10
Água 160.370 1,62
Área Urbana 59.355 0,60
Caatinga Aberta 2.295.779 23,22
Caatinga Densa 2.761.107 27,93
Cana de açúcar e ou Pastagem 546.581 5,53
Capoeira 62.460 0,63
Complexo de Suape 991 0,01
Mangue 6.546 0,07
Mata 248.390 2,51
Mata Serrana 77.002 0,78
Pastagem 1.628.084 16,47
Região Metropolitana 278.992 2,82
Rio 2.956 0,03
Solo Exposto 116.682 1,18
Total 9.886.883 100 Fonte: EMBRAPA Solos UEP Recife e equipe de geoprocessamento do grupo de Pesquisa da Energia da
Biomassa – UFPE (ACCIOLY et al., 2017, no prelo).
Na Zona da Mata era difícil separar nas imagens de satélite o que era pastagem e o que
era Cana-de-Açúcar, porque as Canas-de-Açúcar jovens têm a mesma cobertura graminácea
das pastagens. Cana-de-Açúcar e pastagens, juntas, perfaziam cerca de 6% do território de
Pernambuco. Pelos dados do censo sabe-se que a cana ocupa cerca de 300 mil ha, ficando os
cerca de 250 mil restantes para pastagem. A separação só era feita no campo, quando da
amostragem. As matas e capoeiras ocupam apenas 3% do território estadual. Excluindo nesta
zona a região metropolitana, a classificação para amostragem em áreas de Cana-de-Açúcar,
pastagem, mata e capoeiras também correspondia a uma cobertura de mais de 90% do território
da zona.
3.3.3 Coleta das amostras de solo
Em cada ponto amostral foram coletadas amostras do perfil do solo até a profundidade
de 1 m ou até a camada de impedimento em solos com menos de 1 m de profundidade. Para
melhor entendimento das mudanças e evitar que os efeitos fossem diluídos no perfil, foram
feitas amostragens nas seguintes camadas: 0-10, 10-20, 20-30, 30-40, 40-60, 60-80, 80-100 cm
de profundidade (Figura 9). Adotou-se um fracionamento maior das camadas superficiais do
solo (0-40 cm), sendo essas amostradas a cada 10 cm, uma vez que as perdas ou aportes de

52
carbono são maiores nas camadas superficiais do solo, enquanto que as camadas subsuperficiais
(40-100 cm) foram amostradas a cada 20 cm. Também foi determinada a densidade do solo de
cada camada amostrada com três repetições por sistema, utilizando o método do anel
volumétrico descrito em EMBRAPA (1997).
Para a retirada das amostras deformadas e indeformadas (para determinação da
densidade do solo) foi aberta uma trincheira em cada sítio com 0,7 x 0,7 x 1 m (Figura 9). Os
resíduos superficiais em cada ponto de coleta foram removidos antes da amostragem para evitar
contaminação. As amostras coletadas foram acondicionadas em sacos plásticos, etiquetadas e
encaminhadas ao laboratório de Fertilidade do Solo da Universidade Federal de Pernambuco,
UFPE.
(A) gabarito para marcação da trincheira. (B) trincheira de 0,7 x 0,7 x 1m. (C) detalhe da estratificação do perfil
do solo em camadas para coleta de amostras deformadas e indeformadas de solo.
3.3.4 Processamento e análise das amostras de solo
No laboratório, as amostras indeformadas de solo foram colocadas para secar em estufa
de ventilação forçada a 105ºC por 72 horas para posterior determinação da massa. Após
determinada a massa de solo seco, a densidade do solo foi então calculada, utilizando a
expressão: Ds = ms / v, onde Ds = a densidade do solo, ms = massa do solo seco em estufa à
105°C por 72 horas e v é igual ao volume do cilindro utilizado na amostragem do solo
indeformado (EMBRAPA, 1999).
A
Figura 9 - Trincheira para coleta de amostras de solo.
B C
00 - 10 cm
10 - 20 cm
20 - 30 cm
30 - 40 cm
40 - 60 cm
60 - 80 cm
80 - 100 cm

53
Já as amostras deformadas de solo foram secas ao ar e inicialmente passadas em peneira
com malha de 2 mm para a obtenção da terra fina seca ao ar (TFSA). Posteriormente, uma
alíquota de cada amostra foi macerada em almofariz e passada em peneira com malha 0,149
mm (100 mesh) para determinação dos teores de C.
As concentrações de CT foram determinadas pelo método da combustão seca, utilizando
um analisador elementar CHN (TruSpec CHN LECO® 2006, St. Joseph, EUA), nos
laboratórios de solos da Unidade Acadêmica de Garanhuns – UAG e da EMBRAPA CPATSA,
em Petrolina (Figura 10).
3.3.4.1 Cálculo dos estoques de C do solo
Os estoques totais de CT (Mg ha-1) para cada profundidade amostrada foram calculados
pelos métodos de camada e de massa de solo equivalentes, como descrito por Ellert et al. (2001).
O método da camada equivalente leva em consideração a espessura da camada e a densidade
do solo (BAYER et al., 2000), enquanto o método da massa equivalente de solo utiliza, como
referência, a massa de solo de um tratamento, que é tomada como base para o cálculo do estoque
em todos os demais tratamentos (ELLERT e BETTANY, 1995).
Figura 10 - Análises de CT pelo método da combustão seca em um analisador elementar de
CHN.

54
No presente estudo, foram utilizadas, como referência, as massas de solo das camadas
correspondentes das áreas sob Mata nativa (Mata Atlântica na Zona da Mata e Caatinga nas
mesorregiões do Agreste e Sertão), que representa a condição original do solo (DE BONA,
2004). Desta maneira, os estoques totais de CT foram calculados pela equação (1).
Estoque CT (Mg ha-1) = Cconc. * Ds * E * 10.000 m2 ha-1 * 0,001 Mg kg-1 * Fc (1)
onde,
Cconc. = concentração de C (kg Mg-1); Ds = densidade do solo (Mg m-3); E = espessura da
camada (m); Fc = fator de correção (1 – (cascalhos % + pedras %) / 100).
A massa de solo foi calculada usando a equação (2) e as espessuras das camadas de solo
foram ajustadas usando a equação (3).
Msolo = Ds * E * 10.000 m2 ha-1 (2)
onde,
Msolo = massa de solo por unidade de área (Mg ha-1)
Ead = (Msolo, equiv. - Msolo, camada) * 0.0001 ha m-2 / Ds subsuperficial (3)
onde,
Ead = Espessura adicional da camada subsuperfícial necessária para obter-se a massa de solo
equivalente (m); Msolo, equiv. = massa de solo equivalente; Msolo, camada = massa da camada
de solo mais pesada até 1 m de profundidade (Mg ha-1); Ds subsuperficial = densidade da
camada subsuperficial do solo (Mg m-3)

55
3.3.5. Análise dos dados
Os dados dos estoques de CT do solo das regiões úmida, subúmida e semiárida Leste
foram transformados em x = Log (x), e os obtidos na região semiárida Oeste em 𝑥 = √𝑥 , e
foram submetidos a análise de variância para avaliar a diferença entre os usos da terra e classes
de solo, em cada mesorregião. A comparação das médias foi feita pelo teste de Tukey a 5% (P
< 0,05) de significância. Para as análises e transformações dos dados foi utilizado o software
estatístico ASSISTAT® versão 7.7 beta (SILVA, 2016).
3.4 Estudo 3 - Carbono, nitrogênio e abundância natural (δ13C e δ15N) do solo sob
diferentes usos ao longo de um gradiente climático em Pernambuco.
Com o objetivo de identificar a origem da matéria orgânica no solo e a dinâmica do
carbono e nitrogênio, foram realizadas medidas isotópicas (13C e 15N) nas amostras coletadas
nos perfis de Argissolos de todas as regiões fisiográficas de Pernambuco, em áreas
representativas de floresta, áreas de pastagem e áreas agrícolas (identificados após avaliação
em campo).
Para o levantamento da abundância natural de δ13C e δ15N e das concentrações de C e
de N no solo sob os diferentes usos da terra, fez-se necessário o levantamento prévio das áreas
de predominância de Argissolos e dos usos e coberturas vegetais mais representativos do estado.
Os Argissolos são solos bem representativos em Pernambuco (25,23% da superfície) e
uma das poucas classes que entre as demais, está distribuída por todo o estado (Tabela 6). São
predominantemente encontrados na zona úmida costeira (51%) (EMBRAPA, 2006), ocorrem
sob vegetação de florestas subperenifólia e subcaducifólia, mas também estão presentes em
porções expressivas no agreste e sertão (20% e 12% respectivamente) (EMBRAPA, 2006). No
extremo oeste do estado, em áreas bastante secas, têm como vegetação primária as caatingas
hiperxerófilas. Em todos os casos constituem associações com outros solos, como os Latossolos
(ARAÚJO FILHO et al., 2000).

56
Tabela 6 - Abrangência da classe dos Argissolos no Estado de Pernambuco.
Símbolo Classe Área mapeada Superfície do Estado
km2 %
PAC Argissolos Acinzentados 436,05 0,44
PA Argissolos Amarelos 13.275,92 13,42
PVA Argissolos Vermelho-Amarelos 9.809,94 9,92
PV Argissolos Vermelhos 1.432,73 1,45
TOTAL 24.954,64 25,23
Fonte: Adaptado de ARAÚJO FILHO et al., 2000.
São solos bem desenvolvidos, variando de pouco a muito profundos e com fertilidade
oscilando de baixa a média (ARAÚJO FILHO et al., 2000). Nas áreas mais úmidas são
predominantemente cultivados com cana-de-açúcar e pastagens, podendo-se encontrar
mandioca, abacaxi, coco e citrus. No Oeste do estado, nas áreas próximas do rio são Francisco
estabeleceu-se o polo de fruticultura, utilizando a irrigação (ARAÚJO FILHO et al., 2000).
3.4.1 Levantamento dos Argissolos e das coberturas vegetais existentes
Para o levantamento foram utilizados dados existentes, por meio da junção de mapas e
outros trabalhos recentes feitos em Pernambuco, como descrito no item 3.3.2. Após o
levantamento prévio das manchas de Argissolos, observou-se a predominância de três usos e
coberturas: áreas de floresta (Mata Atlântica/ Caatinga), áreas de pastagem e áreas agrícolas
(cultivos de subsistência/ cana-de-açúcar) (Tabela 7).

57
Tabela 7 - Principais usos e coberturas vegetais existentes em áreas de predominância de
Argissolos no estado de Pernambuco, com algumas de suas características.
Usos e Coberturas Região Características
Vegetação
Nativa
Mata Atlântica Zona da Mata
Remanescente de Mata Atlântica do tipo
subperenifólia sem qualquer exploração ou
interferência antrópica visível.
Caatinga
Agreste
Sertão Leste
Sertão Oeste
Remanescente de caatinga nativa dos tipos
hipoxerófilas e/ou hiperxerófilas densas, sem qualquer
exploração ou interferência antrópica visível.
Áreas
Agrícolas
Cana-de-Açúcar Zona da Mata
Áreas cultivadas com cana-de-açúcar por longos
períodos no sistema convencional (aração, gradagem,
adubação e correção do solo).
Cultivos de
subsistência
Agreste
Sertão Leste
Sertão Oeste
Agricultura de subsistência com cultivos de milho,
feijão, mandioca e palma (para alimentação animal). A
adubação com fertilizantes químicos nessas áreas é
praticamente inexistente, havendo em algumas
adubação com esterco de curral.
Áreas de
Pastagem
Zona da Mata
Agreste
Sertão Leste
Sertão Oeste
Pastagens de Brachiaria spp. e/ou pastagem
natural, em sua grande maioria conduzidas sob pastejo
contínuo de bovinos e/ou caprinos em regime intensivo
ou semi-intensivo e na maioria dos casos extensivo sem
fertilização.
Fontes: JACOMINE et al., 1973; (CONDEPE/FIDEM, 2006)
Após o cruzamento das informações do solo e dos usos e coberturas da terra, por meio
dos mapas e das imagens de satélite, como descrito no item 3.3.2, foi feita a validação dos
pontos amostrais no campo. Com as coordenadas pré-estabelecidas, realizou-se então a
validação, verificando-se se os pontos pré-estabelecidos correspondiam à combinação prevista
(solo x vegetação). Caso não correspondessem, um novo ponto amostral era procurado.
Ao todo, foram amostrados 36 pontos, localizados em 13 municípios, em áreas
representativas das três grandes regiões fisiográficas de Pernambuco (Zona da Mata, Agreste,
e Sertão), sendo esse último dividido em porção Leste e porção Oeste, por diferirem quanto as
classes de solo, perfazendo ao todo, quatro regiões de coleta. Em cada região, para cada uso e
cobertura foram escolhidos três pontos amostrais, tomados como repetições, ficando assim
então: 4 regiões fisiográficas x 3 tipos de usos e coberturas x 3 repetições (Tabela 8).
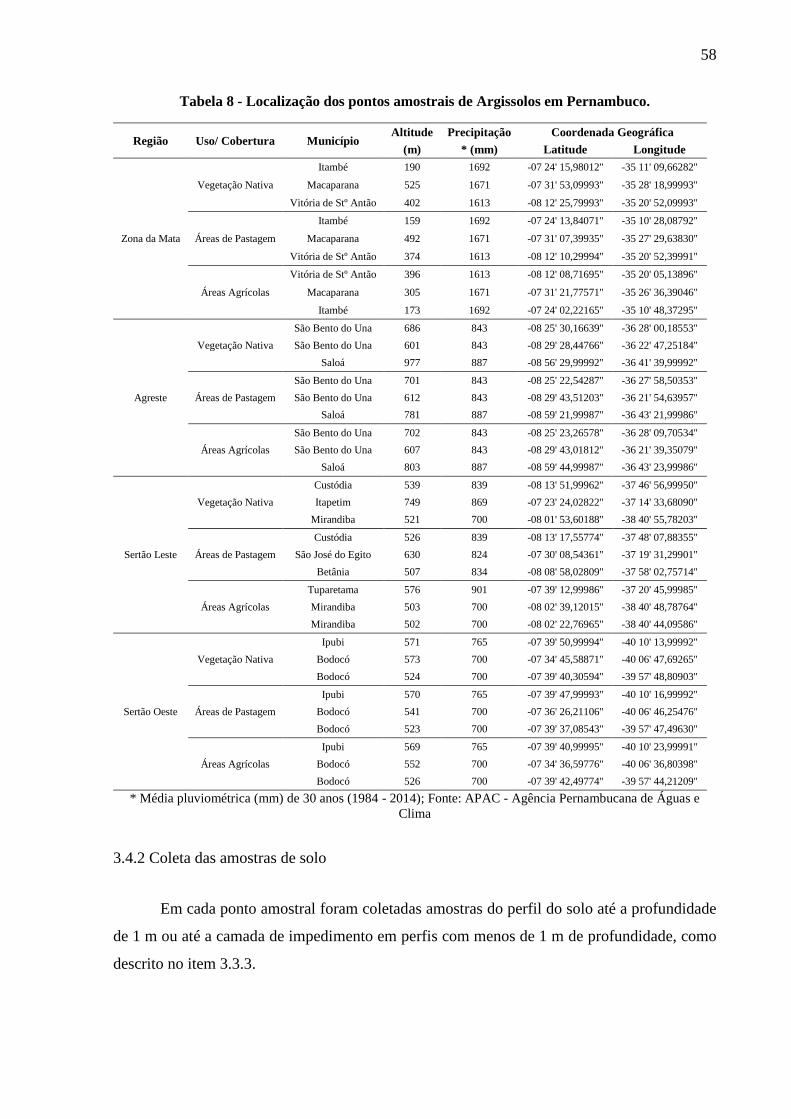
58
Tabela 8 - Localização dos pontos amostrais de Argissolos em Pernambuco.
Região Uso/ Cobertura Município Altitude Precipitação Coordenada Geográfica
(m) * (mm) Latitude Longitude
Zona da Mata
Vegetação Nativa
Itambé 190 1692 -07 24' 15,98012'' -35 11' 09,66282''
Macaparana 525 1671 -07 31' 53,09993'' -35 28' 18,99993''
Vitória de Stº Antão 402 1613 -08 12' 25,79993'' -35 20' 52,09993''
Áreas de Pastagem
Itambé 159 1692 -07 24' 13,84071'' -35 10' 28,08792''
Macaparana 492 1671 -07 31' 07,39935'' -35 27' 29,63830''
Vitória de Stº Antão 374 1613 -08 12' 10,29994'' -35 20' 52,39991''
Áreas Agrícolas
Vitória de Stº Antão 396 1613 -08 12' 08,71695'' -35 20' 05,13896''
Macaparana 305 1671 -07 31' 21,77571'' -35 26' 36,39046''
Itambé 173 1692 -07 24' 02,22165'' -35 10' 48,37295''
Agreste
Vegetação Nativa
São Bento do Una 686 843 -08 25' 30,16639'' -36 28' 00,18553''
São Bento do Una 601 843 -08 29' 28,44766'' -36 22' 47,25184''
Saloá 977 887 -08 56' 29,99992'' -36 41' 39,99992''
Áreas de Pastagem
São Bento do Una 701 843 -08 25' 22,54287'' -36 27' 58,50353''
São Bento do Una 612 843 -08 29' 43,51203'' -36 21' 54,63957''
Saloá 781 887 -08 59' 21,99987'' -36 43' 21,99986''
Áreas Agrícolas
São Bento do Una 702 843 -08 25' 23,26578'' -36 28' 09,70534''
São Bento do Una 607 843 -08 29' 43,01812'' -36 21' 39,35079''
Saloá 803 887 -08 59' 44,99987'' -36 43' 23,99986''
Sertão Leste
Vegetação Nativa
Custódia 539 839 -08 13' 51,99962'' -37 46' 56,99950''
Itapetim 749 869 -07 23' 24,02822'' -37 14' 33,68090''
Mirandiba 521 700 -08 01' 53,60188'' -38 40' 55,78203''
Áreas de Pastagem
Custódia 526 839 -08 13' 17,55774'' -37 48' 07,88355''
São José do Egito 630 824 -07 30' 08,54361'' -37 19' 31,29901''
Betânia 507 834 -08 08' 58,02809'' -37 58' 02,75714''
Áreas Agrícolas
Tuparetama 576 901 -07 39' 12,99986'' -37 20' 45,99985''
Mirandiba 503 700 -08 02' 39,12015'' -38 40' 48,78764''
Mirandiba 502 700 -08 02' 22,76965'' -38 40' 44,09586''
Sertão Oeste
Vegetação Nativa
Ipubi 571 765 -07 39' 50,99994'' -40 10' 13,99992''
Bodocó 573 700 -07 34' 45,58871'' -40 06' 47,69265''
Bodocó 524 700 -07 39' 40,30594'' -39 57' 48,80903''
Áreas de Pastagem
Ipubi 570 765 -07 39' 47,99993'' -40 10' 16,99992''
Bodocó 541 700 -07 36' 26,21106'' -40 06' 46,25476''
Bodocó 523 700 -07 39' 37,08543'' -39 57' 47,49630''
Áreas Agrícolas
Ipubi 569 765 -07 39' 40,99995'' -40 10' 23,99991''
Bodocó 552 700 -07 34' 36,59776'' -40 06' 36,80398''
Bodocó 526 700 -07 39' 42,49774'' -39 57' 44,21209''
* Média pluviométrica (mm) de 30 anos (1984 - 2014); Fonte: APAC - Agência Pernambucana de Águas e
Clima
3.4.2 Coleta das amostras de solo
Em cada ponto amostral foram coletadas amostras do perfil do solo até a profundidade
de 1 m ou até a camada de impedimento em perfis com menos de 1 m de profundidade, como
descrito no item 3.3.3.

59
3.4.3 Processamento e análise das amostras de solo
As amostras de solo foram secas ao ar e passadas em peneira com malha de 2 mm para
a obtenção da terra fina seca ao ar (TFSA). Posteriormente uma alíquota de cada amostra foi
macerada em almofariz e passada em peneira com malha de 0,149 mm (100 mesh) para
determinação dos valores da abundância natural de δ13C e de δ15N e dos teores de C e de N do
solo.
As concentrações de CT e de NT, bem como a abundância natural de δ13C e de δ15N,
foram determinadas por espectrometria de massa isotópica de fluxo contínuo (espectrômetro de
massa Finnigan Delta Plus acoplado em um auto-analisador de C e N total Carlo Erba EA 1108
– Finnigan MAT, Bremen, Alemanha), no Laboratório de Isótopos Estáveis do Centro de
Energia Nuclear na Agricultura, da USP, em Piracicaba, São Paulo. Os resultados foram
expressos na forma de δ13C e δ15N (‰), sendo o C em relação ao padrão internacional PDB e
o N em relação à composição atmosférica (N2 atmosférico), calculadas pelas equações 4 e 5,
respectivamente:
δ13C(‰) = [(R amostra – RPDB) / RPDB] x 1000 (4)
em que R amostra é a relação molar 13C/12C da amostra analisada, e RPDB, a relação
molar 13C/12C do padrão Pee Dee Belemnite (PDB).
δ15N(‰) = [(R amostra - R padrão) / R padrão] x 1000. (5)
onde R amostra é a relação molar 15N/14N da amostra analisada, e R padrão, a relação
molar 15N/14N da composição atmosférica (N2 atmosférico).
3.4.4. Análise dos dados
Os dados foram submetidos ao teste de hipóteses, utilizando-se o software estatístico
ASSISTAT® (SILVA, 2016), para verificar se apresentavam distribuição normal e atendiam
aos critérios necessários para aplicação de um teste paramétrico. Uma vez constatada a
normalidade, pelo teste de Shapiro-Wilk (SHAPIRO e WILK, 1965), os dados foram
submetidos à análise de variância (ANOVA), com aplicação do teste F e os valores médios,
quando significativos, foram comparados pelo teste de Scott-Knott (SCOTT e KNOTT, 1974)

60
ao nível de 5% (P < 0,05) de significância. No caso em que a normalidade dos dados não foi
verificada, foram transformados em Box-Cox (BOX e COX, 1964) e Yeo-Johnson (YEO e
JOHNSON, 2000), através do software estatístico MiniTab®.
Os usos da terra (floresta, pastagem e agricultura) foram comparados entre si em cada
mesorregião de Pernambuco e foram comparados isoladamente dentre as mesorregiões (zona
da Mata, Agreste, Sertão Leste e Sertão Oeste), para cada camada (0-10, 10-20, 20-30, 30-40,
40-60, 60-80 e 80-100 cm) de solo. Realizou-se também correlação de Pearson e regressão
linear entre as variáveis precipitação pluvial, altitude, CT, NT, δ13C e δ15N na camada de 0-20
cm. As análises estatísticas foram feitas usando-se os softwares Assistat versão 7.7 beta (2016)
e Microsoft Excel.

61
4 RESULTADOS E DISCUSSÃO
4.1 Estudo 1 - Levantamento dos estoques de carbono nos solos de Pernambuco.
4.1.1 Estoques de C por classe de solo
Os dados dos 160 perfis (274 horizontes) das diferentes classes de solos sob todos os
usos e coberturas da terra, resultaram numa estimativa de aproximadamente 353 Tg de C (1 Tg
= 1012g, ou 1 milhão de toneladas) para a camada de 0-30 cm nos solos de Pernambuco (Tabela
9). Vale ressaltar que o estoque total de C não incluiu as áreas com solos indiscriminados de
mangues, que correspondem a 0,19% do território do Estado (185,27 km²) (ARAÚJO FILHO
et al., 2000) e as classes dos Organossolos, para as quais não existem informações suficientes
de perfis e da área de abrangência no Estado.
Tabela 9 - Estoques de C (0 a 30 cm de profundidade) por unidade de área e para áreas cobertas
no território de Pernambuco, classe de solo, número de perfis (n) e erro padrão da média (s(m)).
Classes de solo *Área
n Estoque de C
s(m) C
Fontes (km2) (Mg ha-1) (Tg)
Chernossolo 39,50 4 74,02 12,91 0,29 8; 11
Nitossolo 10,10 3 57,55 13,21 0,06 8; 11
Vertissolo 357,09 6 48,10 6,45 1,72 8; 11
Gleissolo 1.234,14 8 46,72 7,17 5,77 2; 8; 11; 13; 14
N. Litólico 20.047,64 7 45,15 6,31 90,52 1; 8; 11
Espodossolo 363,71 5 42,77 6,43 1,56 1; 8; 13; 11; 14
Argissolo 24.954,64 49 40,56 3,25 101,21 2; 3; 4; 5; 7; 8; 9; 11;
13; 14; 15
Luvissolo 8.313,76 15 40,23 5,04 33,45 8; 10; 11
Plintossolo 57,70 2 37,75 0,32 0,22 9; 11
Latossolo 9.257,15 24 37,39 4,13 34,61 1; 2; 4; 8; 11; 13; 14
N. Flúvico 1.990,29 5 36,64 8,22 7,29 8; 11
Planossolo 15.174,57 19 31,50 3,45 47,79 6; 8; 10; 11
N. Regolítico 4.899,18 6 30,81 7,64 15,10 8; 11; 12
N. Quartz. 5.185,51 3 20,48 9,23 10,62 2; 8; 11
Cambissolo 1.622,34 4 15,24 7,86 2,47 8; 15
Totais 93.507,32 160 352,67 N. Flúvico = Neossolo Flúvico; N. Litólico = Neossolo Litólico; N. Quartz. = Neossolo Quartzarênico; N.
Regolít. = Neossolo Regolítico; Fontes: (1) ANTUNES et al., 2010; (2) ARAÚJO e SALCEDO, 1997; (3)
ARAÚJO et al., 2001; (4) ARAÚJO et al., 2004; (5) BONFIM et al., 2004; (6) GALINDO et al., 2008; (7)
GIONGO et al., 2011; (8) JACOMINE et al., 1972; (9) OLIVEIRA et al., 2003; (10) OLIVEIRA et al., 2009; (11)
RIBEIRO et al., 1999; (12) SANTOS et al., 2012; (13) SIMÕES NETO et al., 2009; (14) SIMÕES NETO et al.,
2012; (15) SOUZA et al., 2010. * Fonte: ARAÚJO FILHO et al., 2000.

62
Os Chernossolos, seguidos pelos Nitossolos, Vertissolos e Gleissolos possuem os
maiores estoques de carbono por classe, na camada de 0-30 cm, com 74, 58, 48 e 47 Mg ha-1
de C no solo, respectivamente, sob os diferentes usos e coberturas. Já os Cambissolos e os
Neossolos Quartzarênicos e Regolíticos apresentaram os menores estoques de C, com 15, 20 e
31 Mg ha-1, respectivamente (Tabela 9).
Os maiores estoques de C encontrados nas classes dos Chernossolos, Nitossolos,
Vertissolos e Gleissolos, estão associados ao grande acúmulo de argila presentes nesses solos
(ARAÚJO FILHO et al., 2000; EMBRAPA, 2013), conferindo-lhes maiores estoques de
matéria orgânica que os solos mais arenosos. Isso decorre das maiores área superficial
específica e quantidade de cargas presentes nos solos argilosos, o que possibilita maiores
interações organominerais e menores taxas de mineralização (DICK et al., 2009). Outros fatores
como a topografia e o hidromorfismo, também estão associados aos altos estoques de C, como
no caso dos Gleissolos, especificamente, que apresentam fortes limitações quanto a drenagem,
o que lhes conferem níveis mais altos de MOS, devido ao aporte de sedimentos e resíduos
orgânicos provenientes de áreas mais elevadas, e à lenta decomposição associada à baixa
atividade microbiana em condições anaeróbicas (RESCK et al., 2008).
Em contrapartida, os Neossolos Quartzarênicos que tem como principal característica a
textura muito arenosa (areia ou areia franca), apresentaram maiores estoques de C que os
Cambissolos, na camada de 0-30 cm (Tabela 9), onde a textura varia com maior frequência de
fraco-arenosa a franco-argiloarenosa (textura média) (ARAÚJO FILHO et al., 2000;
EMBRAPA, 2013).
Porém, não se pode desconsiderar que tais comparações merecem um certo cuidado,
principalmente em função da falta de padronização nos processos de amostragem realizados
nos trabalhos publicados. Além disso, o baixo número amostral, bem como o histórico de usos
das áreas e os fatores relacionados as diferenças texturais e mineralógicas existentes na própria
classe dos Cambissolos, em segundo e terceiro nível categórico, podem ter contribuído para os
baixos valores de C encontrados nesse levantamento, pois de acordo com Araújo Filho et al.
(2000), os Cambissolos apresentam características que variam muito de um local para outro,
apresentando uma grande heterogeneidade no material de origem, e outros fatores de formação
dos solos, como o clima e o relevo, por exemplo. Desta forma, a classe dos Cambissolos
comportam solos desde fortemente até imperfeitamente drenados, de rasos a profundos, de alta
a baixa saturação por bases, bem como, de alta a baixa atividade das argilas e ainda podem
ocorrer com e sem pedregosidade e em diversos relevos, desde o plano até montanhoso.

63
A média geral dos estoques de C na camada de 0-30 cm, ponderada pela proporção de
área das classes de solo, é da ordem de 38 Mg ha-1 e, como corresponde a todos os usos do solo,
pode ser extrapolada para 368 Tg de C, fazendo-se assim, a correção para a área total de solos
do Estado, incluindo as áreas para as quais não se tem informação (áreas indiscriminadas de
mangue e Organossolos).
O valor médio de 38 Mg ha-1 de carbono foi superior ao encontrado por Sampaio e Costa
(2011), que estimaram o estoque médio de C por área para todo o semiárido nordestino em 28
Mg ha-1. A explicação desse maior valor médio de C para os solos de Pernambuco deve-se
principalmente à incorporação dos perfis amostrados no bioma Mata Atlântica e na maior
profundidade utilizada neste trabalho (30 cm). Segundo Post et al. (1982), a magnitude da
variação dos estoques de C sob vegetação nativa é bastante significativa: regiões de maior
altitude, de climas úmidos e sob florestas apresentam maiores estoques de C, sendo os menores
estoques encontrados em regiões de clima árido e semiárido.
As estimativas dos estoques de C (Mg ha-1) das classes de solo em Pernambuco ficaram
próximas das calculadas por Fidalgo et al. (2007), que consideraram os diferentes tipos de solos
nos mais variados biomas (Amazônia, Caatinga, Cerrado, Mata Atlântica e Pantanal) em todo
Brasil, com exceção dos Cambissolos, Chernossolos, Gleissolos, Luvissolos e dos Neossolos
Litólicos e Regolíticos (Figura 11). É provável que as grandes variações dos Cambissolos,
Chernossolos, Gleissolos e Neossolos Regolíticos de Pernambuco estejam associadas ao
pequeno número de perfis amostrados. Nesta comparação, deve-se levar em conta também que
boa parte dos estoques de C estimados para os solos brasileiros (FIDALGO et al., 2007)
referem-se aos biomas Mata Atlântica e Amazônia, esse último, ainda predominantemente sob
vegetação natural.
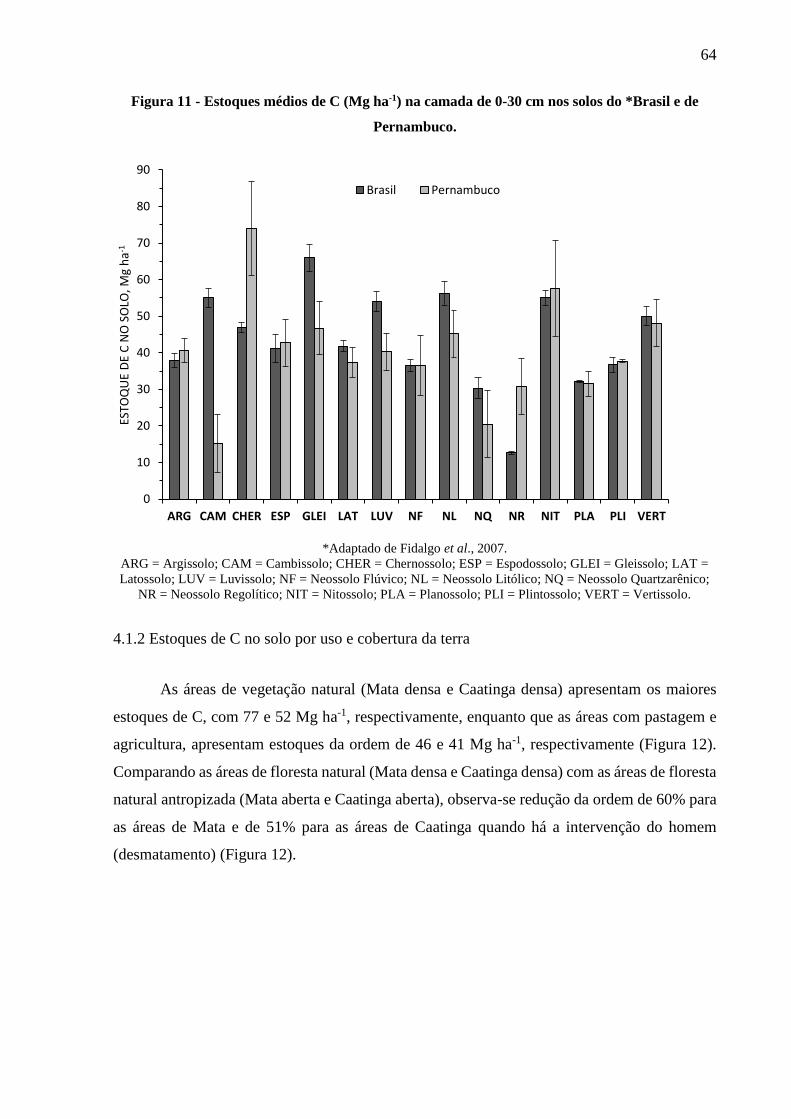
64
Figura 11 - Estoques médios de C (Mg ha-1) na camada de 0-30 cm nos solos do *Brasil e de
Pernambuco.
*Adaptado de Fidalgo et al., 2007.
ARG = Argissolo; CAM = Cambissolo; CHER = Chernossolo; ESP = Espodossolo; GLEI = Gleissolo; LAT =
Latossolo; LUV = Luvissolo; NF = Neossolo Flúvico; NL = Neossolo Litólico; NQ = Neossolo Quartzarênico;
NR = Neossolo Regolítico; NIT = Nitossolo; PLA = Planossolo; PLI = Plintossolo; VERT = Vertissolo.
4.1.2 Estoques de C no solo por uso e cobertura da terra
As áreas de vegetação natural (Mata densa e Caatinga densa) apresentam os maiores
estoques de C, com 77 e 52 Mg ha-1, respectivamente, enquanto que as áreas com pastagem e
agricultura, apresentam estoques da ordem de 46 e 41 Mg ha-1, respectivamente (Figura 12).
Comparando as áreas de floresta natural (Mata densa e Caatinga densa) com as áreas de floresta
natural antropizada (Mata aberta e Caatinga aberta), observa-se redução da ordem de 60% para
as áreas de Mata e de 51% para as áreas de Caatinga quando há a intervenção do homem
(desmatamento) (Figura 12).
0
10
20
30
40
50
60
70
80
90
ARG CAM CHER ESP GLEI LAT LUV NF NL NQ NR NIT PLA PLI VERT
ESTO
QU
E D
E C
NO
SO
LO, M
g h
a-1
Brasil Pernambuco
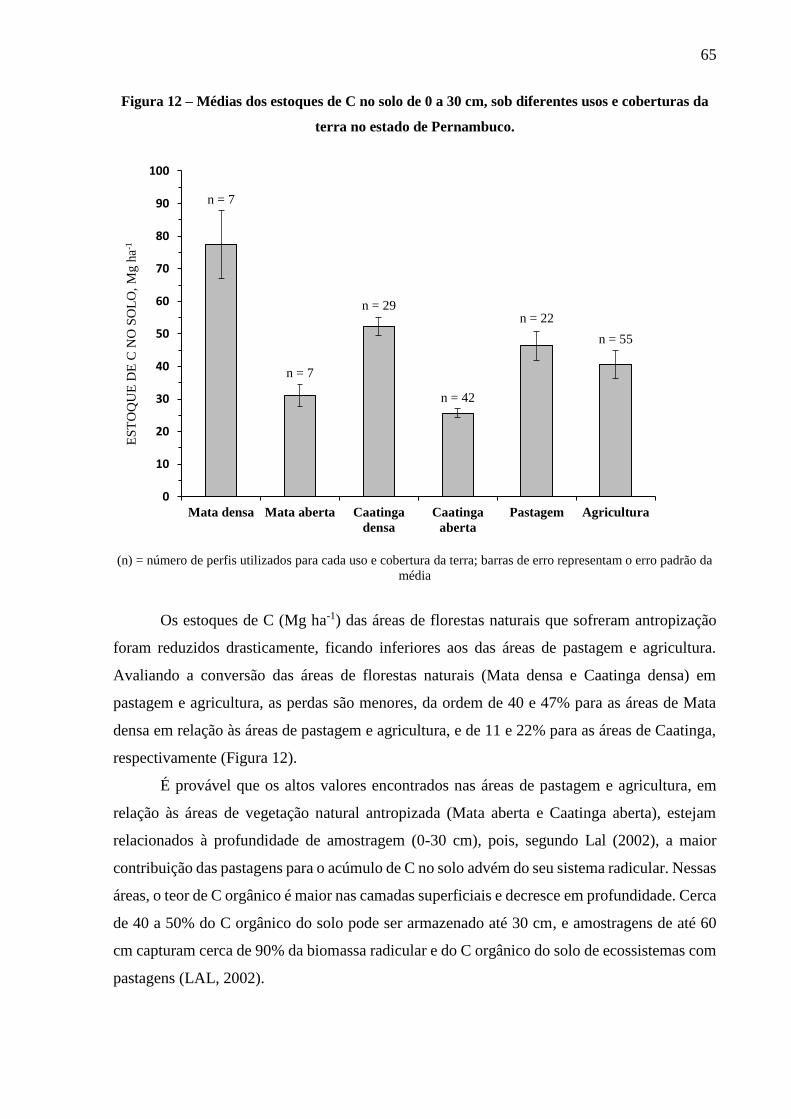
65
Figura 12 – Médias dos estoques de C no solo de 0 a 30 cm, sob diferentes usos e coberturas da
terra no estado de Pernambuco.
(n) = número de perfis utilizados para cada uso e cobertura da terra; barras de erro representam o erro padrão da
média
Os estoques de C (Mg ha-1) das áreas de florestas naturais que sofreram antropização
foram reduzidos drasticamente, ficando inferiores aos das áreas de pastagem e agricultura.
Avaliando a conversão das áreas de florestas naturais (Mata densa e Caatinga densa) em
pastagem e agricultura, as perdas são menores, da ordem de 40 e 47% para as áreas de Mata
densa em relação às áreas de pastagem e agricultura, e de 11 e 22% para as áreas de Caatinga,
respectivamente (Figura 12).
É provável que os altos valores encontrados nas áreas de pastagem e agricultura, em
relação às áreas de vegetação natural antropizada (Mata aberta e Caatinga aberta), estejam
relacionados à profundidade de amostragem (0-30 cm), pois, segundo Lal (2002), a maior
contribuição das pastagens para o acúmulo de C no solo advém do seu sistema radicular. Nessas
áreas, o teor de C orgânico é maior nas camadas superficiais e decresce em profundidade. Cerca
de 40 a 50% do C orgânico do solo pode ser armazenado até 30 cm, e amostragens de até 60
cm capturam cerca de 90% da biomassa radicular e do C orgânico do solo de ecossistemas com
pastagens (LAL, 2002).
n = 7
n = 7
n = 29
n = 42
n = 22
n = 55
0
10
20
30
40
50
60
70
80
90
100
Mata densa Mata aberta Caatinga
densa
Caatinga
aberta
Pastagem Agricultura
ES
TO
QU
E D
E C
NO
SO
LO
, M
g h
a-1

66
Existem evidências de que práticas conservacionistas, como determinadas técnicas de
plantio (integração lavoura-pecuária-floresta), pastagens bem manejadas, florestas plantadas e
sistemas agroflorestais, também podem reduzir drasticamente estas perdas, mantendo os níveis
de C orgânico ou até mesmo aumentando-os (CARTER, 2001). A maioria das práticas agrícolas
não provoca elevações significativas dos níveis de carbono em pouco tempo, sendo necessários
pelo menos 25 a 50 anos antes que um novo equilíbrio seja atingido (BATJES, 2001).
Fujisaka et al. (1998) avaliaram os efeitos da conversão de florestas nativas em áreas
agrícolas e pastagens, na Amazônia, e constataram redução da ordem de 80% nos estoques C
no solo com a transformação, através do corte e queima da vegetação nativa. Ressaltaram que
a alta redução registrada deve ter ocorrido em curto prazo, pois ao longo do tempo, em
pastagens bem manejadas, esses estoques podem ser similares ou superiores ao de solos sob
floresta nativa. Já Borges e Kiehl (1996), observaram em um Latossolo na Bahia, que o estoque
total de carbono (100 Mg ha-1) sob vegetação natural até 100 cm de profundidade, foi reduzido
em torno de 20% com os cultivos de banana, manga e mandioca, enquanto o cultivo de citros
reduziu o estoque em torno de 10%. São valores que se aproximam bastante dos encontrados
nesse trabalho.
4.2 Estudo 2 - Distribuição em profundidade e estoques de carbono em solos sob diferentes
coberturas e usos da terra em Pernambuco.
4.2.1 Distribuição do CT no solo em profundidade
Os padrões de distribuição de CT nos perfis dos solos foram diferentes entre os usos e
coberturas da terra, em cada região fisiográfica, mas, de modo geral, correlacionaram-se
inversamente com a profundidade do solo, sendo as concentrações mais altas nas camadas
superficiais e decrescendo nas camadas subjacentes de forma exponencial ou potencial (Figuras
13, 14, 15 e 16). Entretanto, em alguns perfis de áreas agrícolas e de vegetação nativa aberta,
principalmente nas Zonas do Agreste e Sertão, as concentrações variaram pouco nas maiores
profundidades (Figuras 14 e 15), devido principalmente ao menor aporte de resíduos orgânicos
nos solos dessas regiões, em decorrência da menor produção de biomassa vegetal que é limitada
pela baixa precipitação pluvial.

67
Figura 13 - Distribuição em profundidade do C do solo sob diferentes usos e coberturas da terra
na Zona da Mata de Pernambuco.
As barras de erro representam o erro padrão da média (n = 3).
Reduções das concentrações de C com a maior profundidade dos solos confirmam os
resultados obtidos por diversos autores (ROSCOE e MACHADO 2002; ADUAN et al., 2003;
LAL, 2005; PAIVA e FARIA, 2007; PAIVA et al., 2011), assim como o fato do modelo
exponencial ser o que melhor explica a distribuição do C no perfil do solo (ARROUAYS e
PELISSIER, 1994; RICHTER et al., 1999; JOBBÁGY e JACKSON, 2000).
0
20
40
60
80
100
0 1 2 3 4 5
Pro
fun
did
ade
(cm
)
CT (%)
Mata nativa Densa
Argissolo
Latossolo
0 1 2 3 4 5
CT (%)
Mata nativa Aberta
0
20
40
60
80
100
Pro
fun
did
ade
(cm
)
Áreas Agrícolas Áreas de Pastagens
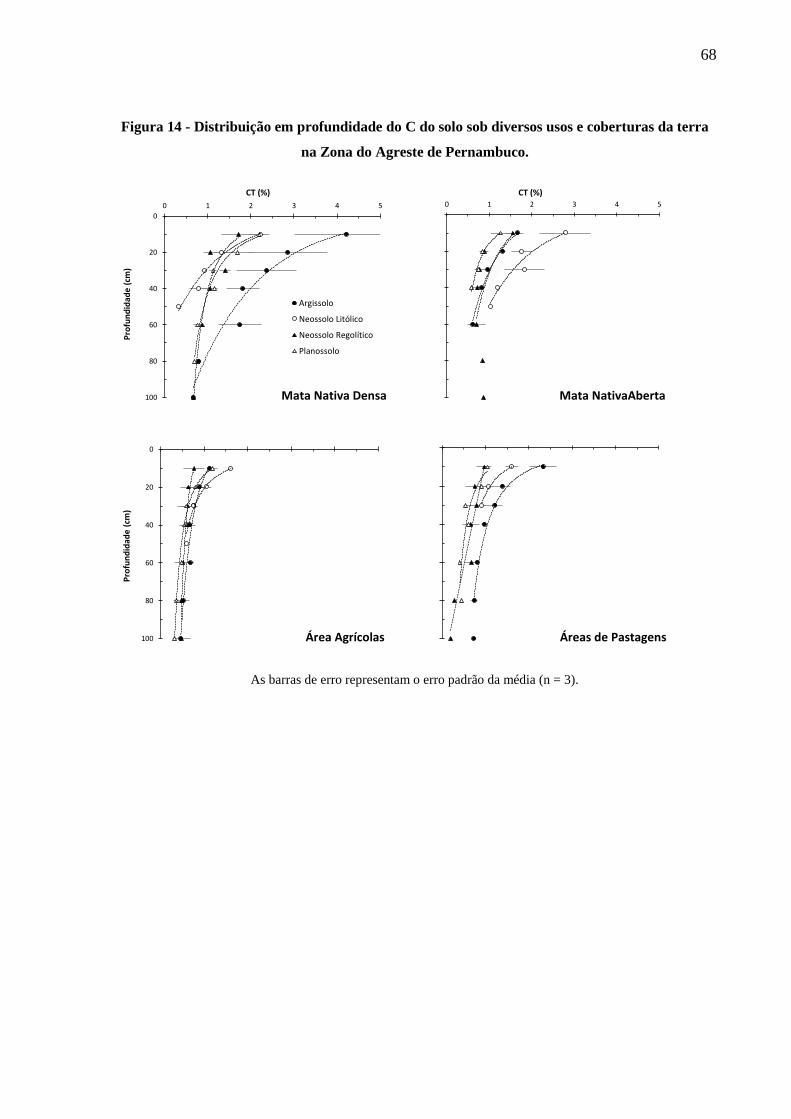
68
Figura 14 - Distribuição em profundidade do C do solo sob diversos usos e coberturas da terra
na Zona do Agreste de Pernambuco.
As barras de erro representam o erro padrão da média (n = 3).
0
20
40
60
80
100
0 1 2 3 4 5
Pro
fun
did
ade
(cm
)
CT (%)
Mata Nativa Densa
Argissolo
Neossolo Litólico
Neossolo Regolítico
Planossolo
0 1 2 3 4 5
CT (%)
Mata NativaAberta
0
20
40
60
80
100
Pro
fun
did
ade
(cm
)
Área Agrícolas Áreas de Pastagens
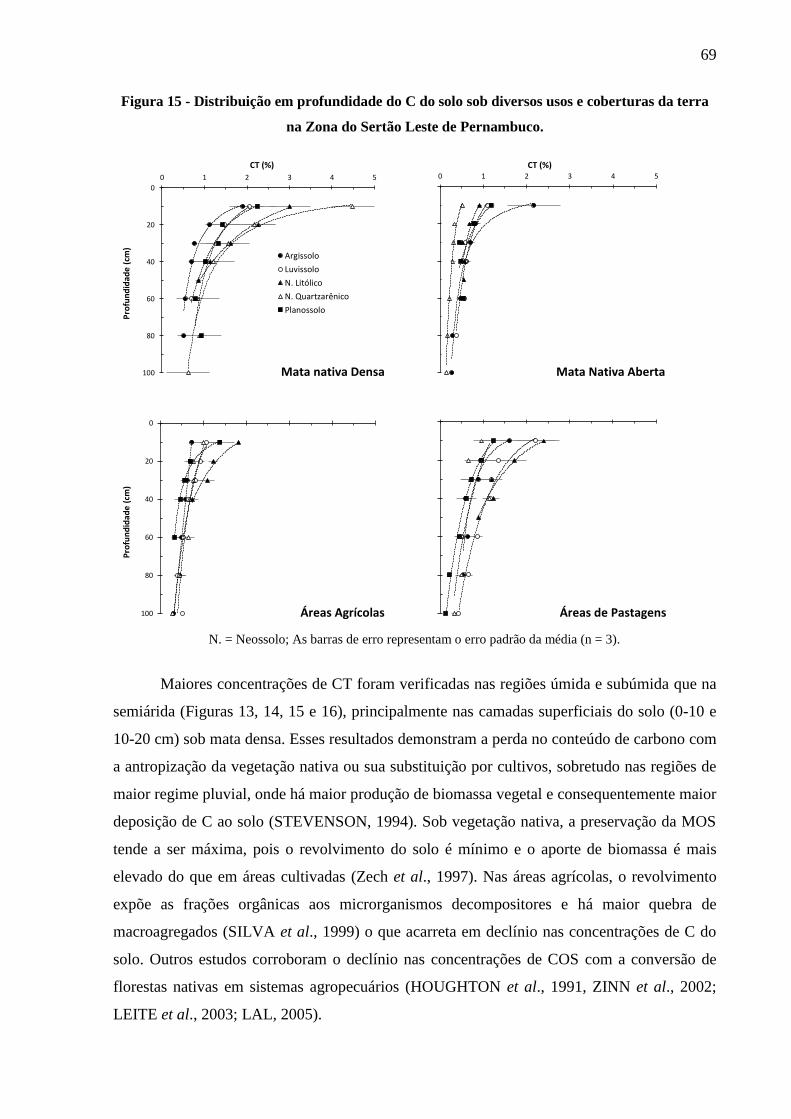
69
Figura 15 - Distribuição em profundidade do C do solo sob diversos usos e coberturas da terra
na Zona do Sertão Leste de Pernambuco.
N. = Neossolo; As barras de erro representam o erro padrão da média (n = 3).
Maiores concentrações de CT foram verificadas nas regiões úmida e subúmida que na
semiárida (Figuras 13, 14, 15 e 16), principalmente nas camadas superficiais do solo (0-10 e
10-20 cm) sob mata densa. Esses resultados demonstram a perda no conteúdo de carbono com
a antropização da vegetação nativa ou sua substituição por cultivos, sobretudo nas regiões de
maior regime pluvial, onde há maior produção de biomassa vegetal e consequentemente maior
deposição de C ao solo (STEVENSON, 1994). Sob vegetação nativa, a preservação da MOS
tende a ser máxima, pois o revolvimento do solo é mínimo e o aporte de biomassa é mais
elevado do que em áreas cultivadas (Zech et al., 1997). Nas áreas agrícolas, o revolvimento
expõe as frações orgânicas aos microrganismos decompositores e há maior quebra de
macroagregados (SILVA et al., 1999) o que acarreta em declínio nas concentrações de C do
solo. Outros estudos corroboram o declínio nas concentrações de COS com a conversão de
florestas nativas em sistemas agropecuários (HOUGHTON et al., 1991, ZINN et al., 2002;
LEITE et al., 2003; LAL, 2005).
0
20
40
60
80
100
0 1 2 3 4 5
Pro
fun
did
ade
(cm
)
CT (%)
Mata nativa Densa
Argissolo
Luvissolo
N. Litólico
N. Quartzarênico
Planossolo
0 1 2 3 4 5
CT (%)
Mata Nativa Aberta
0
20
40
60
80
100
Pro
fun
did
ade
(cm
)
Áreas Agrícolas Áreas de Pastagens

70
Figura 16 - Distribuição em profundidade do C do solo sob diversos usos e coberturas da terra
na Zona do Sertão Oeste de Pernambuco.
N. = Neossolo; As barras de erro representam o erro padrão da média (n = 3).
4.2.2 Estoques totais de C no solo
Os estoques totais de C (Mg ha-1) nos solos variaram bastante entre as mesorregiões do
estado e foram influenciados significativamente pelos usos e coberturas da terra, bem como
pelas concentrações e distribuição do C no perfil. Os maiores estoques de CT ocorreram, de
maneira geral, sob vegetações nativas densas e decresceram da região úmida à semiárida Oeste,
com grandes variações entre as classes solos (Tabela 10). Esse comportamento é típico de
situação sob vegetação nativa, como já mencionado anteriormente, pois o aporte de resíduos
vegetais na superfície do solo promove a decomposição lenta e gradual, que garante a constante
incorporação de material orgânico no solo, sobretudo nas regiões úmidas e subúmidas, onde há
maior produção de biomassa em função da maior disponibilidade de água.
0
20
40
60
80
100
0 1 2 3 4 5
Pro
fun
did
ade
(cm
)
CT (%)
Mata Nativa Densa
Argissolo
Latossolo
N. Litólico
Planossolo
0 1 2 3 4 5
CT (%)
Mata Nativa Aberta
0
20
40
60
80
100
Pro
fun
did
ade
(cm
)
Áreas Agrícolas Áreas de Pastagens
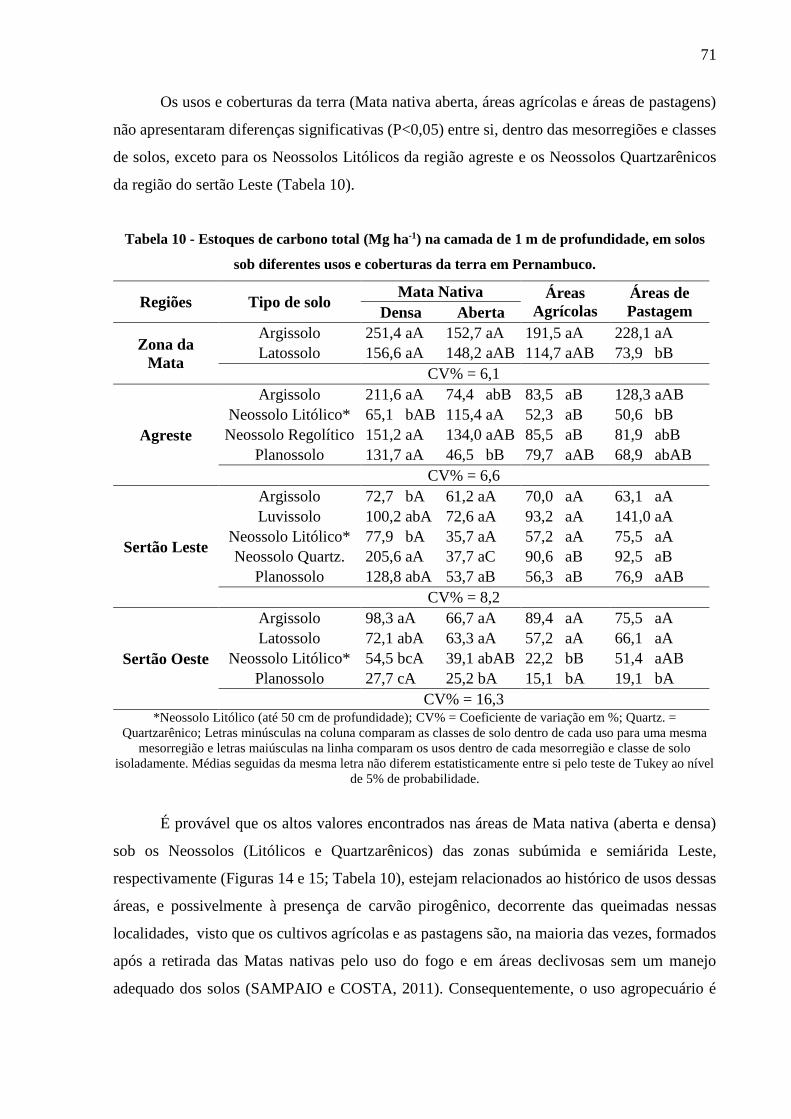
71
Os usos e coberturas da terra (Mata nativa aberta, áreas agrícolas e áreas de pastagens)
não apresentaram diferenças significativas (P<0,05) entre si, dentro das mesorregiões e classes
de solos, exceto para os Neossolos Litólicos da região agreste e os Neossolos Quartzarênicos
da região do sertão Leste (Tabela 10).
Tabela 10 - Estoques de carbono total (Mg ha-1) na camada de 1 m de profundidade, em solos
sob diferentes usos e coberturas da terra em Pernambuco.
Regiões Tipo de solo Mata Nativa Áreas
Agrícolas
Áreas de
Pastagem Densa Aberta
Zona da
Mata
Argissolo 251,4 aA 152,7 aA 191,5 aA 228,1 aA
Latossolo 156,6 aA 148,2 aAB 114,7 aAB 73,9 bB
CV% = 6,1
Agreste
Argissolo 211,6 aA 74,4 abB 83,5 aB 128,3 aAB
Neossolo Litólico* 65,1 bAB 115,4 aA 52,3 aB 50,6 bB
Neossolo Regolítico 151,2 aA 134,0 aAB 85,5 aB 81,9 abB
Planossolo 131,7 aA 46,5 bB 79,7 aAB 68,9 abAB
CV% = 6,6
Sertão Leste
Argissolo 72,7 bA 61,2 aA 70,0 aA 63,1 aA
Luvissolo 100,2 abA 72,6 aA 93,2 aA 141,0 aA
Neossolo Litólico* 77,9 bA 35,7 aA 57,2 aA 75,5 aA
Neossolo Quartz. 205,6 aA 37,7 aC 90,6 aB 92,5 aB
Planossolo 128,8 abA 53,7 aB 56,3 aB 76,9 aAB
CV% = 8,2
Sertão Oeste
Argissolo 98,3 aA 66,7 aA 89,4 aA 75,5 aA
Latossolo 72,1 abA 63,3 aA 57,2 aA 66,1 aA
Neossolo Litólico* 54,5 bcA 39,1 abAB 22,2 bB 51,4 aAB
Planossolo 27,7 cA 25,2 bA 15,1 bA 19,1 bA
CV% = 16,3 *Neossolo Litólico (até 50 cm de profundidade); CV% = Coeficiente de variação em %; Quartz. =
Quartzarênico; Letras minúsculas na coluna comparam as classes de solo dentro de cada uso para uma mesma
mesorregião e letras maiúsculas na linha comparam os usos dentro de cada mesorregião e classe de solo
isoladamente. Médias seguidas da mesma letra não diferem estatisticamente entre si pelo teste de Tukey ao nível
de 5% de probabilidade.
É provável que os altos valores encontrados nas áreas de Mata nativa (aberta e densa)
sob os Neossolos (Litólicos e Quartzarênicos) das zonas subúmida e semiárida Leste,
respectivamente (Figuras 14 e 15; Tabela 10), estejam relacionados ao histórico de usos dessas
áreas, e possivelmente à presença de carvão pirogênico, decorrente das queimadas nessas
localidades, visto que os cultivos agrícolas e as pastagens são, na maioria das vezes, formados
após a retirada das Matas nativas pelo uso do fogo e em áreas declivosas sem um manejo
adequado dos solos (SAMPAIO e COSTA, 2011). Consequentemente, o uso agropecuário é

72
comumente seguido pelo abandono das áreas, ocorrendo então a regeneração da vegetação
nativa.
Vale salientar também, que não há uma distinção clara entre mata nativa densa e mata
nativa aberta (ou pastos nativos), ambos correspondendo principalmente à vegetação de
caatinga, quase toda ela ocupada eventualmente com rebanhos domésticos. Admite-se que as
matas densas sejam vegetação de maior porte, mais alta, mais fechada e mais arbórea, que as
matas abertas (ou pastos nativos). Estes podem ir desde áreas com cobertura quase exclusiva
de herbáceas e subarbustos até capoeiras mais densas. Parte deles são áreas em regeneração
natural, correspondendo ao ciclo de pousio da agricultura itinerante. Parte mantém-se aberta
com retiradas periódicas de lenha e queimadas (SAMPAIO e COSTA, 2011).
De uma maneira geral, os estoques de CT nas camadas dos solos seguiram as mesmas
tendências dos teores de C em profundidade (Figuras 13, 14, 15 e 16), com os maiores estoques
nas camadas superficiais e decrescendo em direção as camadas mais profundas (Tabela 11). Em
geral, a camada superficial até 30 cm estocava cerca de 40 a 60% do carbono total do solo até
1 m de profundidade, mas podia chegar até 80% nas classes de solo com menos de 1 m de
profundidade, como alguns Planossolos e até quase 100% em Neossolos Litólicos, com
profundidade máxima de 50 cm (Tabela 11). Jobbagy & Jackson (2000) afirmaram que cerca
de 50% do carbono total do solo estão estocados em camadas mais profundas e diversos outros
autores também chamaram a atenção para a importância do subsolo no armazenamento de CT
(LORENZ e LAL, 2005; JANZEN, 2006; SCHÖNING et al., 2006)
Determinações de estoques de CT no solo até um metro de profundidade, sob diferentes
usos e coberturas da terra nas diferentes regiões do Nordeste são escassas, dificultando as
comparações. Porém, comparações limitadas às camadas mais superficiais dos solos na região
semiárida revelam resultados assemelhados aos aqui reportados. Em áreas de mata nativa sob
Argissolo no semiárido pernambucano, por exemplo, Kauffman et al. (1993) encontraram
estoques de C de 26,2 Mg ha-1 na camada de 0-20 cm, enquanto Fraga e Salcedo (2004)
reportaram valores de 28,6 Mg ha-1 para a camada de 0-15 cm. Esses valores são próximos aos
encontrados nesse estudo para as áreas de mata aberta, tanto no sertão Leste (22,6 Mg ha-1)
quanto Oeste (26,9 Mg ha-1), mas inferiores aos da mata densa (29,5 e 36,0 Mg ha-1,
respectivamente) considerando a camada de 0-20 cm (Tabela 4) em Argissolos. Já Maia et al.
(2007), trabalhando em área de caatinga hiperxerófila sobre Luvissolo no semiárido cearense,
encontraram valor na camada de 0-20 cm (44,4 Mg ha-1) bem próximo aos do presente estudo
em áreas de caatinga densa (46,9 Mg ha-1) sobre Luvissolo (Tabela 11).
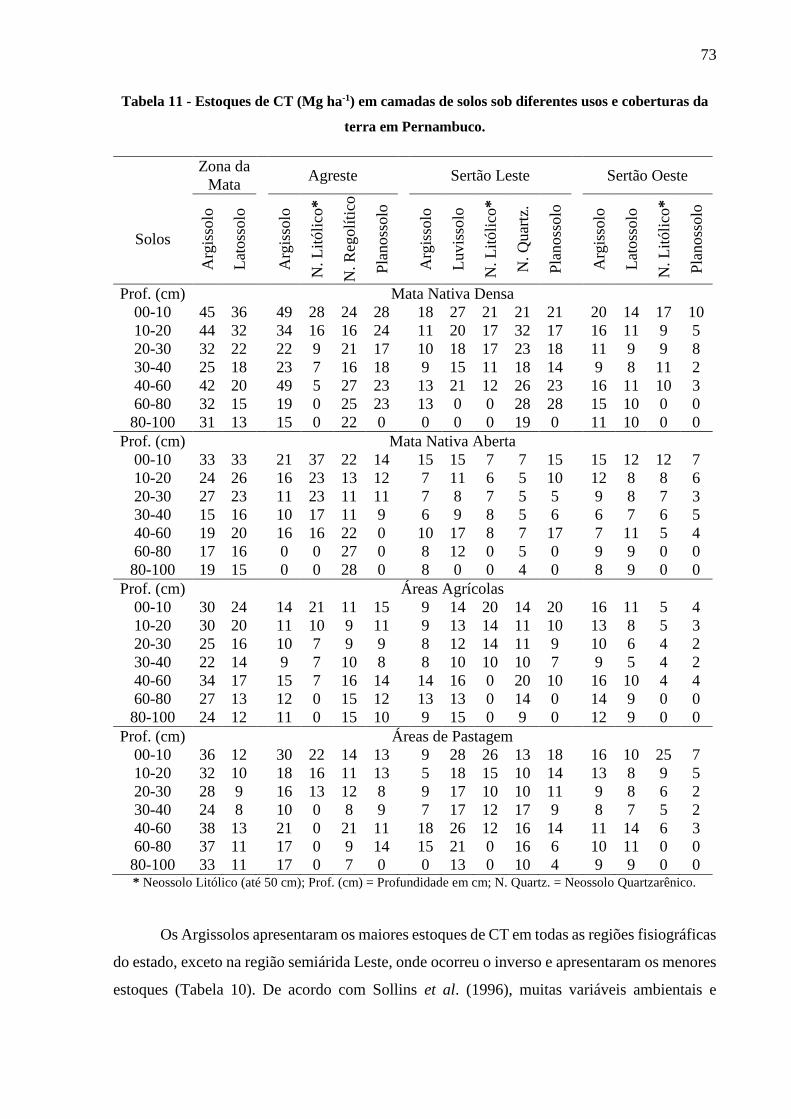
73
Tabela 11 - Estoques de CT (Mg ha-1) em camadas de solos sob diferentes usos e coberturas da
terra em Pernambuco.
Zona da
Mata Agreste Sertão Leste Sertão Oeste
Solos
Arg
isso
lo
Lat
oss
olo
A
rgis
solo
N. L
itóli
co*
N. R
egolí
tico
Pla
noss
olo
A
rgis
solo
Luvis
solo
N. L
itóli
co*
N. Q
uar
tz.
Pla
noss
olo
A
rgis
solo
Lat
oss
olo
N. L
itóli
co*
Pla
noss
olo
Prof. (cm) Mata Nativa Densa
00-10 45 36 49 28 24 28 18 27 21 21 21 20 14 17 10
10-20 44 32 34 16 16 24 11 20 17 32 17 16 11 9 5
20-30 32 22 22 9 21 17 10 18 17 23 18 11 9 9 8
30-40 25 18 23 7 16 18 9 15 11 18 14 9 8 11 2
40-60 42 20 49 5 27 23 13 21 12 26 23 16 11 10 3
60-80 32 15 19 0 25 23 13 0 0 28 28 15 10 0 0
80-100 31 13 15 0 22 0 0 0 0 19 0 11 10 0 0
Prof. (cm) Mata Nativa Aberta
00-10 33 33 21 37 22 14 15 15 7 7 15 15 12 12 7
10-20 24 26 16 23 13 12 7 11 6 5 10 12 8 8 6
20-30 27 23 11 23 11 11 7 8 7 5 5 9 8 7 3
30-40 15 16 10 17 11 9 6 9 8 5 6 6 7 6 5
40-60 19 20 16 16 22 0 10 17 8 7 17 7 11 5 4
60-80 17 16 0 0 27 0 8 12 0 5 0 9 9 0 0
80-100 19 15 0 0 28 0 8 0 0 4 0 8 9 0 0
Prof. (cm) Áreas Agrícolas
00-10 30 24 14 21 11 15 9 14 20 14 20 16 11 5 4
10-20 30 20 11 10 9 11 9 13 14 11 10 13 8 5 3
20-30 25 16 10 7 9 9 8 12 14 11 9 10 6 4 2
30-40 22 14 9 7 10 8 8 10 10 10 7 9 5 4 2
40-60 34 17 15 7 16 14 14 16 0 20 10 16 10 4 4
60-80 27 13 12 0 15 12 13 13 0 14 0 14 9 0 0
80-100 24 12 11 0 15 10 9 15 0 9 0 12 9 0 0
Prof. (cm) Áreas de Pastagem
00-10 36 12 30 22 14 13 9 28 26 13 18 16 10 25 7
10-20 32 10 18 16 11 13 5 18 15 10 14 13 8 9 5
20-30 28 9 16 13 12 8 9 17 10 10 11 9 8 6 2
30-40 24 8 10 0 8 9 7 17 12 17 9 8 7 5 2
40-60 38 13 21 0 21 11 18 26 12 16 14 11 14 6 3
60-80 37 11 17 0 9 14 15 21 0 16 6 10 11 0 0
80-100 33 11 17 0 7 0 0 13 0 10 4 9 9 0 0 * Neossolo Litólico (até 50 cm); Prof. (cm) = Profundidade em cm; N. Quartz. = Neossolo Quartzarênico.
Os Argissolos apresentaram os maiores estoques de CT em todas as regiões fisiográficas
do estado, exceto na região semiárida Leste, onde ocorreu o inverso e apresentaram os menores
estoques (Tabela 10). De acordo com Sollins et al. (1996), muitas variáveis ambientais e

74
antrópicas podem influenciar os estoques de C nas classes de solo, dentre elas composição e
quantidade do resíduo vegetal sobre o solo (serapilheira), fertilidade natural, temperatura,
precipitação pluvial, manejo e histórico da área, além da composição granulométrica do solo,
de modo que se torna difícil estabelecer um padrão definido para uma mesma classe de solo em
regiões fisiográficas e usos distintos.
Na região úmida, os Argissolos apresentaram maiores estoques de C que os Latossolos
em todos os quatro usos (Tabela 10), embora essa superioridade só tenha sido significativa
(P<0,05) nas áreas de pastagem, onde os Argissolos (228,1 Mg ha-1) estocaram três vezes mais
que os Latossolos (73,9 Mg ha-1).
A mudança de uso da terra, nessa região, resultou em decréscimo dos estoques de CT
do solo nos Latossolos, tendo as áreas de mata densa os maiores estoques (156,6 Mg ha-1),
chegando a ser duas vezes superiores aos das áreas de pastagem (73,9 Mg ha-1) (Tabela 10).
Dentre as regiões, a subúmida do Agreste foi onde os estoques de CT foram mais
influenciados pelos usos e coberturas da terra, em todas as classes de solo (Argissolo, Neossolo
Litólico, Neossolo Regolítico e Planossolo), com decréscimos nos usos quando comparados à
vegetação densa. Os maiores decréscimos ocorreram nos Argissolos, com reduções de mais de
60% nas áreas de mata aberta (74,4 Mg ha-1) e agricultura (83,5 Mg ha-1) em relação as áreas
de mata densa (211,6 Mg ha-1). Nas áreas de mata densa e pastagem os menores estoques foram
verificados nos Neossolos Litólicos, um fato influenciado pela reduzida profundidade destes
solos, que não ultrapassa os 50 cm (Tabela 10).
Na região semiárida Leste, as áreas de mata densa sobre Neossolos Quartzarênicos
(205,6 Mg ha-1) apresentaram os maiores estoques de C, significativamente superiores aos dos
Argissolos (72,7 Mg ha-1) e dos Neossolos Litólicos (77,9 Mg ha-1) que apresentaram estoques
mais de 60% inferiores (Tabela 10). Quanto à mudança de uso da terra, apenas nos Neossolos
Quartzarênicos e nos Planossolos houve reduções significativas.
Diferenças em relação à mudança do uso da terra na região semiárida Oeste só foram
verificadas nos Neossolos Litólicos, que sob mata densa (54,4 Mg ha-1) estocaram mais do
dobro (22,2 Mg ha-1) que sob cultivos agrícolas (Tabela 10). Sob matas densas os Argissolos
(98,3 Mg ha-1) apresentaram os maiores estoques, superando em 45% os Neossolos Litólicos
(54,5 Mg ha-1) e em 72% os Planossolos (27,7 Mg ha-1), mas não diferindo significativamente
dos Latossolos (72,1 Mg ha-1). Os Argissolos também foram superiores às outras classes
(Latossolos, Neossolos Litólicos e Planossolos) nas áreas de mata aberta (66,7 Mg ha-1),
agricultura (89,4 Mg ha-1) e pastagem (75,5 Mg ha-1), enquanto os menores estoques foram
verificados nos Planossolos, em todos os usos.

75
Comparando os estoques totais de CT (0-100 cm) sob vegetação nativa densa, em todas
as classes de solo por região fisiográfica, levando em conta que a principal característica dessas
áreas é não ter sofrido antropização recente e de maior monta, a sequência decrescente nos
estoques foi: na região úmida, Argissolo > Latossolo; na região subúmida, Argissolo >
Neossolo Regolítico > Planossolo > Neossolo Litólico; região semiárida Leste: Neossolo
Quartzarênico > Planossolo > Luvissolo > Neossolo Litólico > Argissolo; e na região semiárida
Oeste: Argissolo > Latossolo > Neossolo Litólico > Planossolo.
As médias gerais de todas as classes de solo para os usos e coberturas da terra em cada
região fisiográfica indicam que as maiores reduções nos estoques de CT do solo em relação as
áreas de referência, sob mata densa, ocorrem nas regiões subúmida e semiárida Leste, com
reduções que variam de 34 a 55%, e de 16 a 27%, respectivamente (Figura 17).
Sampaio e Costa (2011) estimaram os estoques de carbono em diferentes usos da terra
no Nordeste, considerando reduções de 25 e 33%, com a mudança de caatinga para agricultura,
nas camada de 0-20 e 20-100 cm, respectivamente. São valores inferiores aos encontrados nesse
estudo para a região semiárida Leste (27%) e Oeste (37%) (Figura 17).
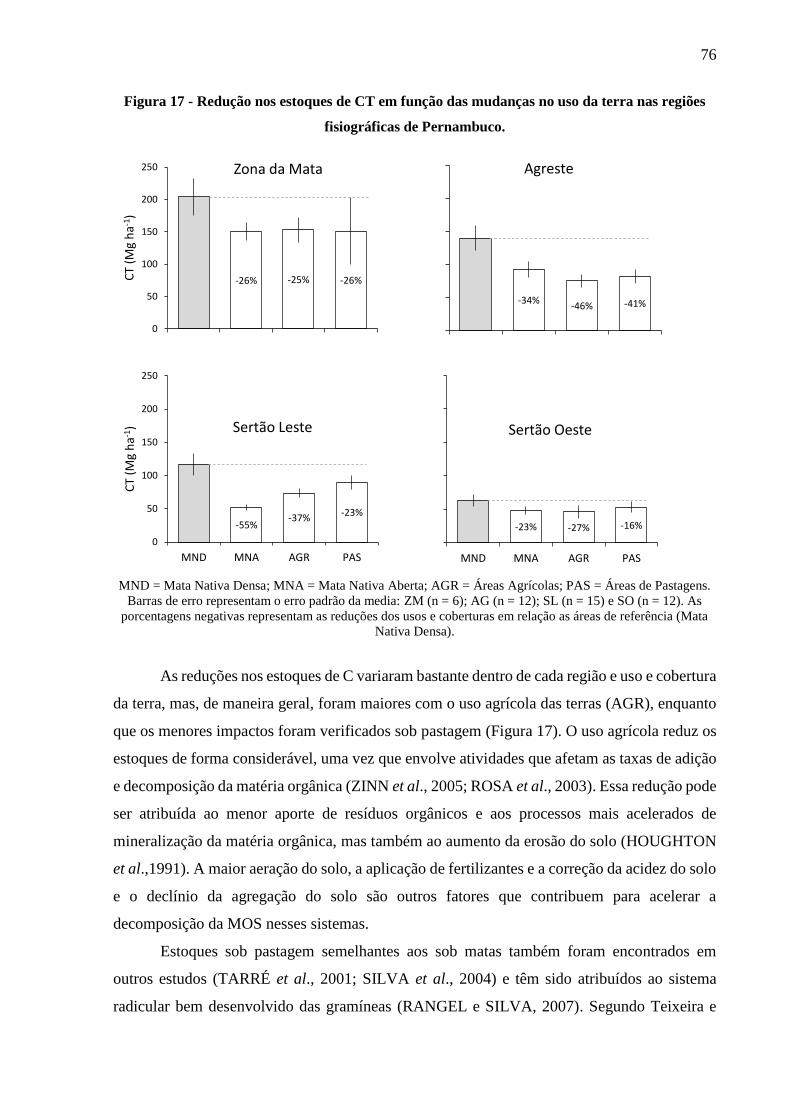
76
Figura 17 - Redução nos estoques de CT em função das mudanças no uso da terra nas regiões
fisiográficas de Pernambuco.
MND = Mata Nativa Densa; MNA = Mata Nativa Aberta; AGR = Áreas Agrícolas; PAS = Áreas de Pastagens.
Barras de erro representam o erro padrão da media: ZM (n = 6); AG (n = 12); SL (n = 15) e SO (n = 12). As
porcentagens negativas representam as reduções dos usos e coberturas em relação as áreas de referência (Mata
Nativa Densa).
As reduções nos estoques de C variaram bastante dentro de cada região e uso e cobertura
da terra, mas, de maneira geral, foram maiores com o uso agrícola das terras (AGR), enquanto
que os menores impactos foram verificados sob pastagem (Figura 17). O uso agrícola reduz os
estoques de forma considerável, uma vez que envolve atividades que afetam as taxas de adição
e decomposição da matéria orgânica (ZINN et al., 2005; ROSA et al., 2003). Essa redução pode
ser atribuída ao menor aporte de resíduos orgânicos e aos processos mais acelerados de
mineralização da matéria orgânica, mas também ao aumento da erosão do solo (HOUGHTON
et al.,1991). A maior aeração do solo, a aplicação de fertilizantes e a correção da acidez do solo
e o declínio da agregação do solo são outros fatores que contribuem para acelerar a
decomposição da MOS nesses sistemas.
Estoques sob pastagem semelhantes aos sob matas também foram encontrados em
outros estudos (TARRÉ et al., 2001; SILVA et al., 2004) e têm sido atribuídos ao sistema
radicular bem desenvolvido das gramíneas (RANGEL e SILVA, 2007). Segundo Teixeira e
-26% -25% -26%
0
50
100
150
200
250
CT
(Mg
ha-1
)
Zona da Mata
-34%-46% -41%
Agreste
-55%-37%
-23%
0
50
100
150
200
250
MND MNA AGR PAS
CT
(Mg
ha-1
) Sertão Leste
-23% -27% -16%
MND MNA AGR PAS
Sertão Oeste

77
Bastos (1989), as pastagens apresentam distribuição regular do sistema radicular até 1 metro de
profundidade, com distribuição de 46% das raízes na camada superficial do solo (0-10 cm). Os
estoques de C no solo dependem, dentre outros fatores, do clima, do conteúdo e natureza das
argilas e do uso e do sistema de manejo adotado (EBELING et al., 2013). Em áreas de matas
nativas de regiões semiáridas, os estoques de CT são menores que os encontrados sob matas
nativas de áreas úmidas (POST et al., 1982). Devido à baixa pluviosidade típica dessas regiões,
o desenvolvimento da vegetação, seja ela de caatinga, áreas agrícolas ou pastos é limitado.
Outros estudos corroboram os resultados aqui encontrados, demostrando reduções
drásticas de CT no solo com a mudança de uso da terra em regiões semiáridas. Liu et al. (2010)
concluíram que pastagens nativas e áreas com arbustos nativos na região semiárida da China
apresentam estoques de C aproximadamente 35% maiores que solos com cultivos
convencionais. Já Fracetto et al. (2012) observaram perdas de 50% de C em áreas agrícolas
quando comparadas a de vegetação nativa no semiárido brasileiro. Enquanto Traoré et al.
(2015) encontraram reduções de C de 49 e 54% em áreas degradadas sob Cambissolos e
Luvissolos quando comparadas as áreas preservadas no semiárido da África.
4.3 Estudo 3 - Carbono, nitrogênio e abundância natural (δ13C e δ15N) do solo sob
diferentes usos ao longo de um gradiente climático em Pernambuco.
4.3.1 Carbono total
As concentrações de CT nos Argissolos variaram entre as mesorregiões e entre os
diferentes usos da terra, em todas as camadas. As maiores concentrações foram encontradas nas
regiões da Zona da Mata, úmida e do Agreste, subúmido, que diferiram (p < 0,05) das demais
regiões semiáridas (sertão Leste e Oeste), (Figura 18).
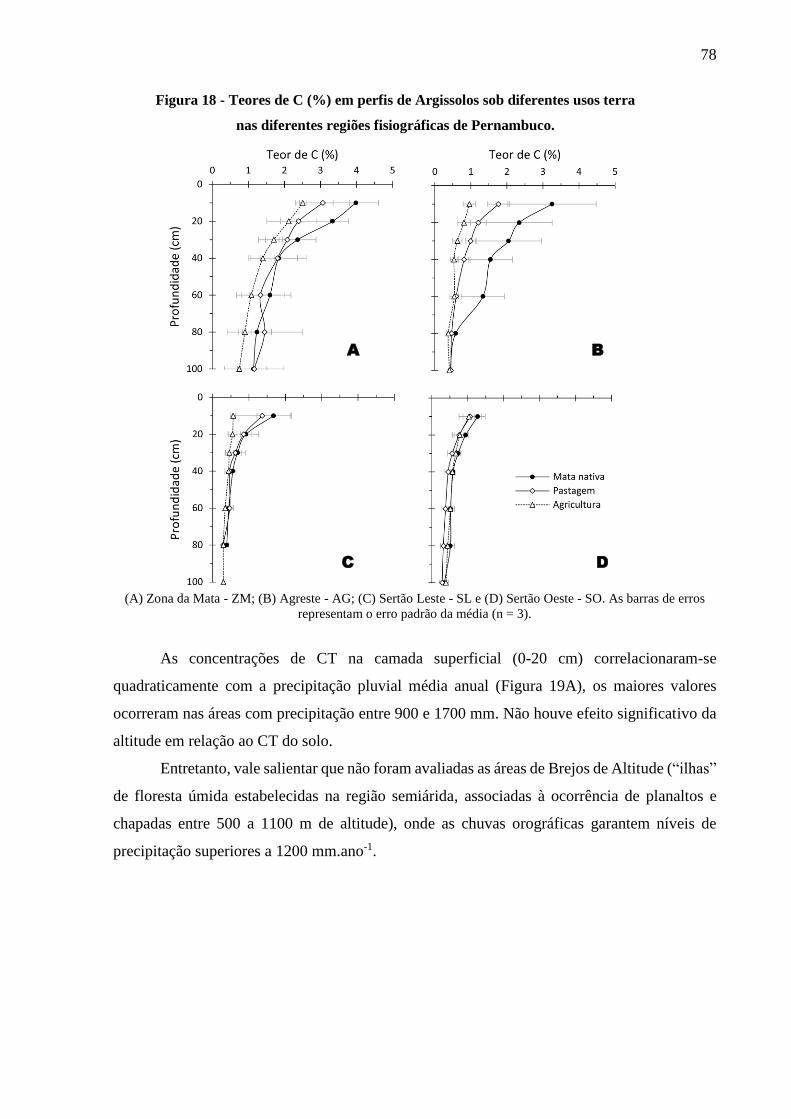
78
(A) Zona da Mata - ZM; (B) Agreste - AG; (C) Sertão Leste - SL e (D) Sertão Oeste - SO. As barras de erros
representam o erro padrão da média (n = 3).
As concentrações de CT na camada superficial (0-20 cm) correlacionaram-se
quadraticamente com a precipitação pluvial média anual (Figura 19A), os maiores valores
ocorreram nas áreas com precipitação entre 900 e 1700 mm. Não houve efeito significativo da
altitude em relação ao CT do solo.
Entretanto, vale salientar que não foram avaliadas as áreas de Brejos de Altitude (“ilhas”
de floresta úmida estabelecidas na região semiárida, associadas à ocorrência de planaltos e
chapadas entre 500 a 1100 m de altitude), onde as chuvas orográficas garantem níveis de
precipitação superiores a 1200 mm.ano-1.
A B
C D
Figura 18 - Teores de C (%) em perfis de Argissolos sob diferentes usos terra
nas diferentes regiões fisiográficas de Pernambuco.
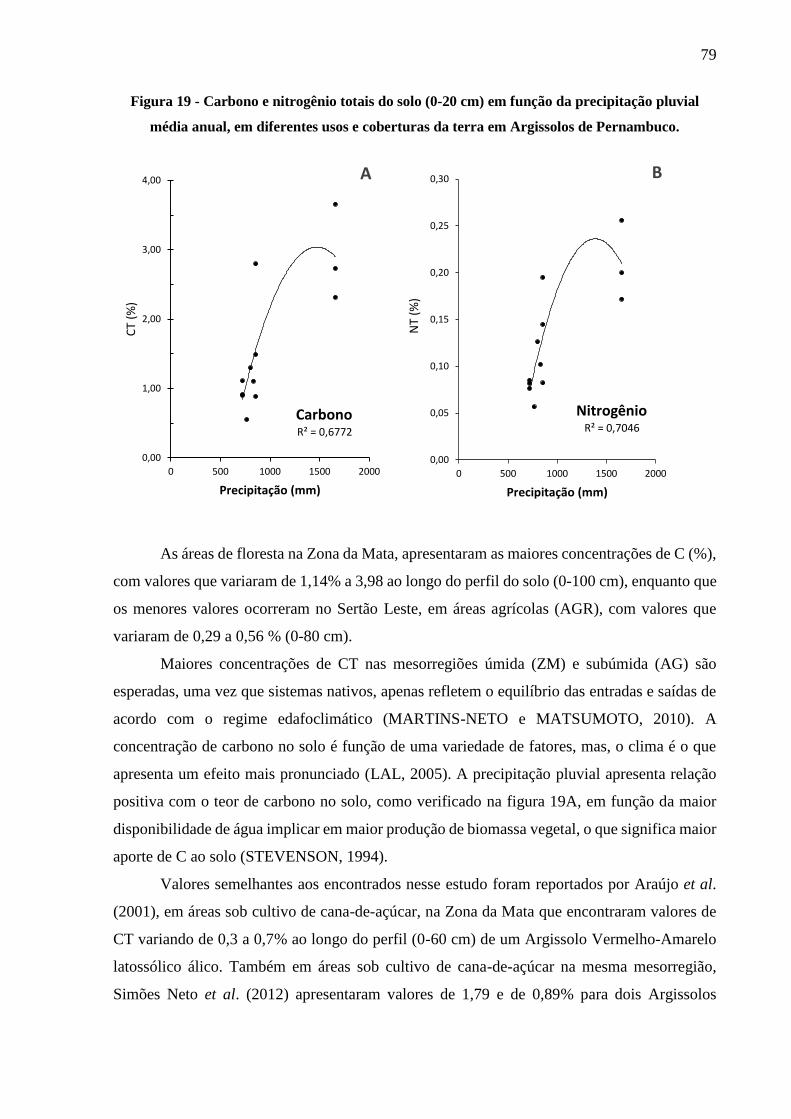
79
Figura 19 - Carbono e nitrogênio totais do solo (0-20 cm) em função da precipitação pluvial
média anual, em diferentes usos e coberturas da terra em Argissolos de Pernambuco.
As áreas de floresta na Zona da Mata, apresentaram as maiores concentrações de C (%),
com valores que variaram de 1,14% a 3,98 ao longo do perfil do solo (0-100 cm), enquanto que
os menores valores ocorreram no Sertão Leste, em áreas agrícolas (AGR), com valores que
variaram de 0,29 a 0,56 % (0-80 cm).
Maiores concentrações de CT nas mesorregiões úmida (ZM) e subúmida (AG) são
esperadas, uma vez que sistemas nativos, apenas refletem o equilíbrio das entradas e saídas de
acordo com o regime edafoclimático (MARTINS-NETO e MATSUMOTO, 2010). A
concentração de carbono no solo é função de uma variedade de fatores, mas, o clima é o que
apresenta um efeito mais pronunciado (LAL, 2005). A precipitação pluvial apresenta relação
positiva com o teor de carbono no solo, como verificado na figura 19A, em função da maior
disponibilidade de água implicar em maior produção de biomassa vegetal, o que significa maior
aporte de C ao solo (STEVENSON, 1994).
Valores semelhantes aos encontrados nesse estudo foram reportados por Araújo et al.
(2001), em áreas sob cultivo de cana-de-açúcar, na Zona da Mata que encontraram valores de
CT variando de 0,3 a 0,7% ao longo do perfil (0-60 cm) de um Argissolo Vermelho-Amarelo
latossólico álico. Também em áreas sob cultivo de cana-de-açúcar na mesma mesorregião,
Simões Neto et al. (2012) apresentaram valores de 1,79 e de 0,89% para dois Argissolos
A
R² = 0,6772
0,00
1,00
2,00
3,00
4,00
0 500 1000 1500 2000
CT
(%)
Precipitação (mm)
Carbono
B
R² = 0,7046
0,00
0,05
0,10
0,15
0,20
0,25
0,30
0 500 1000 1500 2000
NT
(%)
Precipitação (mm)
Nitrogênio

80
Amarelos na camada de 0-20 cm. Já Nascimento et al. (2004), reportaram valores mais baixos
(0,6%) para a mesma camada de um Argissolo Vermelho-Amarelo Distrófico de textura média
cultivado com milho e feijoeiro na Zona da Mata. Para a região semiárida do Sertão Oeste, em
áreas de caatinga degradada, Souza et al. (2010) reportaram valores de CT de 0,2 a 0,5 (0-105
cm) em um Argissolo Vermelho-Amarelo Distrófico abrúptico cambissólico e Oliveira et al.
(2003), apresentaram valores de 0,3 a 0,8% (0-100 cm) em dois Argissolos Amarelos
Eutróficos.
Os teores de CT tenderam a diminuir em profundidade (Figura 18), um padrão
observado em vários trabalhos, decorrente dos maiores aportes de matéria orgânica nas camadas
superficiais do solo (ARROUAYS e PELISSIER, 1994; RICHTER et al., 1999; JOBBÁGY e
JACKSON, 2000; ZHOU et al., 2006; BAYER et al., 2006; SALTON et al., 2011). Em geral,
não foram encontradas diferenças significativas (p < 0,05) entre os usos e coberturas da terra
(floresta, áreas de pastagem e áreas agrícolas), embora, as áreas de floresta e pastagem tenham
apresentado maiores concentrações de C ao longo do perfil do solo (0-100 cm) que as áreas de
cultivo agrícola, sobretudo nas camadas mais superficiais nas mesorregiões úmidas e
subúmidas de Pernambuco (Figura 18).
Os resultados corroboram o trabalho de Barreto et al. (2008), que avaliaram os teores
de CT sob três diferentes usos e coberturas da terra (Mata Atlântica, cultivo de cacau e
pastagem) no sul do Bahia e concluíram que a substituição da mata nativa pelo cultivo de cacau
ou pastagem não alteraram os teores de CT e de suas frações nas camadas superficiais do solo.
Em estudo semelhante, Tarré et al. (2001), compararam os níveis de carbono no solo de áreas
de pastagem (Brachiaria humidicola) com nove anos de uso e consorciada com leguminosa
(Desmodium ovalifolium), com uma área de Mata Atlântica adjacente, e também não
encontraram diferenças significativas entre os sistemas avaliados. Em contrapartida, os
resultados divergem de outros estudos, que apontam diferenças entre os usos e coberturas da
terra, após a retirada da vegetação nativa para a introdução de áreas agrícolas e/ou pastoris
(SILVA et al., 2004; MAIA et al., 2009; SIQUEIRA NETO et al., 2009).
Os trabalhos encontrados na literatura muitas vezes são contraditórios em relação às
diferenças entre os teores de C encontrados em solos de mata nativa e pastagens. Muitos
mostram que, pelo maior aporte de matéria orgânica proporcionado pelas raízes, os solos sob
pastagens apresentam teores iguais ou superiores aos encontrados em ambientes de mata
(GERALDES et al., 1995; CERRI et al., 1996; NEILL et al., 1996; FERNANDES et al., 1997;
MARCHIORI JÚNIOR e MELO, 1999; TARRÉ et al., 2001), enquanto outros mostram valores
mais altos nos solos de mata (DANTAS, 1979; DADALTO et al., 1986; NOORDWIJK et al.,

81
1997; ZECH et al., 1997 ZINN et al., 2002; LEITE et al., 2003; LAL, 2005), proporcionado
pelo maior aporte global de matéria orgânica.
É provável, que o baixo número amostral, bem como o histórico de usos das áreas
agrícolas e os fatores relacionados as diferenças texturais e mineralógicas existentes na própria
classe dos Argissolos, em segundo e terceiro nível categórico, tenham contribuído para a alta
variabilidade dos dados, proporcionando dessa maneira uma ausência de diferenças estatísticas
significativas entre os usos da terra nas mesorregiões do estudo, sobretudo entre as áreas de
cultivos agrícolas e as áreas de vegetação nativa. Sendo assim, torna-se difícil estabelecer
padrões quanto à mudança da vegetação nativa para outros usos da terra, devido à complexidade
da dinâmica do C no solo, e de outros fatores como a variabilidade espacial, tipos de clima,
diferenças texturais e mineralógicas e histórico de manejo das áreas, como já reportado por
diversos autores na literatura (FREITAS et al., 2000; BAYER et al., 2006; MARCHÃO et al.,
2009; MAIA et al., 2009) e também constatadas nesse estudo.
4.3.2 Nitrogênio total
As concentrações de N seguiram a mesma tendência das de C sendo superiores (p >
0,05) nas regiões da Mata e Agreste que aos dos Sertões, correlacionando-se quadraticamente
com a precipitação pluvial média anual na camada superficial do solo (Figura 19B) e sem efeito
significativo para a altitude. Também não foram verificadas diferenças significativas entre os
usos e coberturas do solo nas camadas estudadas, exceto para as áreas agrícolas, onde o
Argissolo da mesorregião do AG é significativamente inferior ao da ZM (Figura 20).
Os usos e coberturas da terra também não apresentaram diferenças significativas, assim
como para as concentrações de C nas regiões da ZM, SL e SO, apenas nas camadas
subsuperficiais (20-30 e 30-40 cm) nos Argissolos da mesorregião AG. Para essas camadas do
solo, as concentrações foram maiores nas áreas de floresta e pastagem do que nas de agricultura
(Figura 20), tendência essa, também verificada nas demais camadas do perfil, embora sem
diferirem estatisticamente.
As concentrações de N nas áreas de floresta na região úmida (ZM) e subúmida (AG),
variaram de 0,07 a 0,28% e 0,04 a 0,22% ao longo do perfil do solo (0-100 cm),
respectivamente. Já as menores concentrações, foram encontradas nas áreas de agricultura na
mesorregião do Sertão Leste, com valores que variaram de 0,06% na camada de 0-10 cm e
0,04% na camada de 80-100 cm (Figura 19).
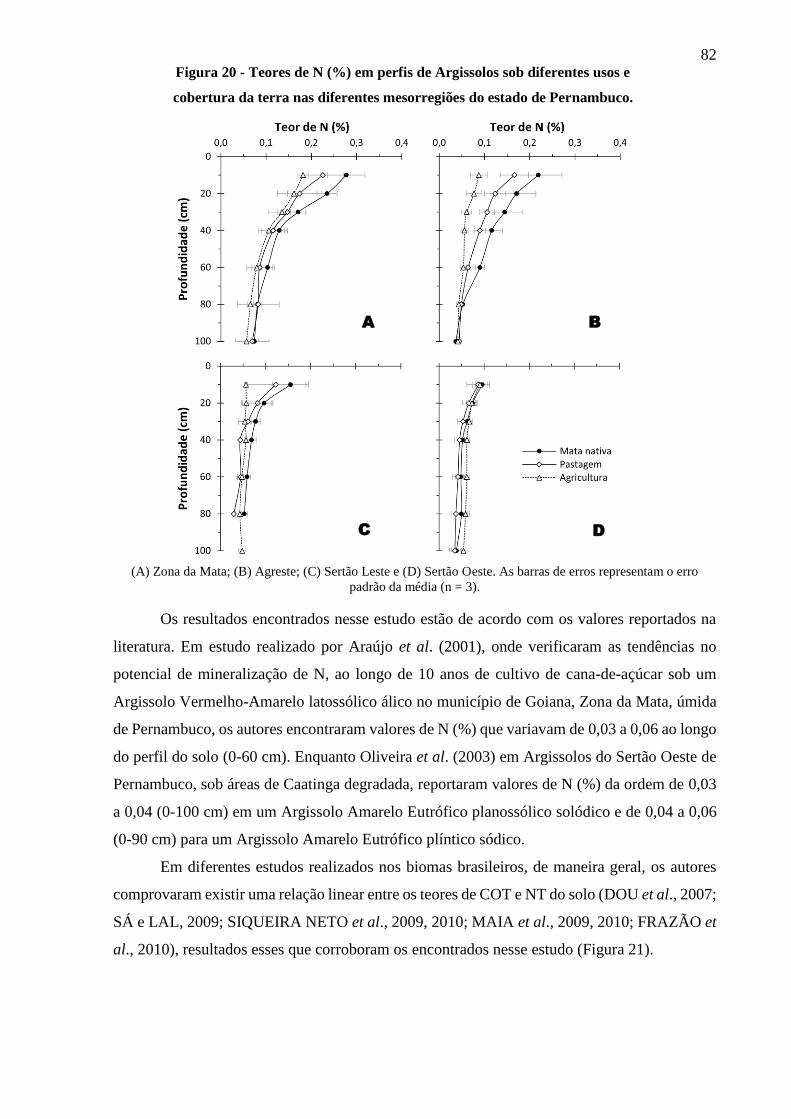
82
(A) Zona da Mata; (B) Agreste; (C) Sertão Leste e (D) Sertão Oeste. As barras de erros representam o erro
padrão da média (n = 3).
Os resultados encontrados nesse estudo estão de acordo com os valores reportados na
literatura. Em estudo realizado por Araújo et al. (2001), onde verificaram as tendências no
potencial de mineralização de N, ao longo de 10 anos de cultivo de cana-de-açúcar sob um
Argissolo Vermelho-Amarelo latossólico álico no município de Goiana, Zona da Mata, úmida
de Pernambuco, os autores encontraram valores de N (%) que variavam de 0,03 a 0,06 ao longo
do perfil do solo (0-60 cm). Enquanto Oliveira et al. (2003) em Argissolos do Sertão Oeste de
Pernambuco, sob áreas de Caatinga degradada, reportaram valores de N (%) da ordem de 0,03
a 0,04 (0-100 cm) em um Argissolo Amarelo Eutrófico planossólico solódico e de 0,04 a 0,06
(0-90 cm) para um Argissolo Amarelo Eutrófico plíntico sódico.
Em diferentes estudos realizados nos biomas brasileiros, de maneira geral, os autores
comprovaram existir uma relação linear entre os teores de COT e NT do solo (DOU et al., 2007;
SÁ e LAL, 2009; SIQUEIRA NETO et al., 2009, 2010; MAIA et al., 2009, 2010; FRAZÃO et
al., 2010), resultados esses que corroboram os encontrados nesse estudo (Figura 21).
A B
C D
Figura 20 - Teores de N (%) em perfis de Argissolos sob diferentes usos e
cobertura da terra nas diferentes mesorregiões do estado de Pernambuco.
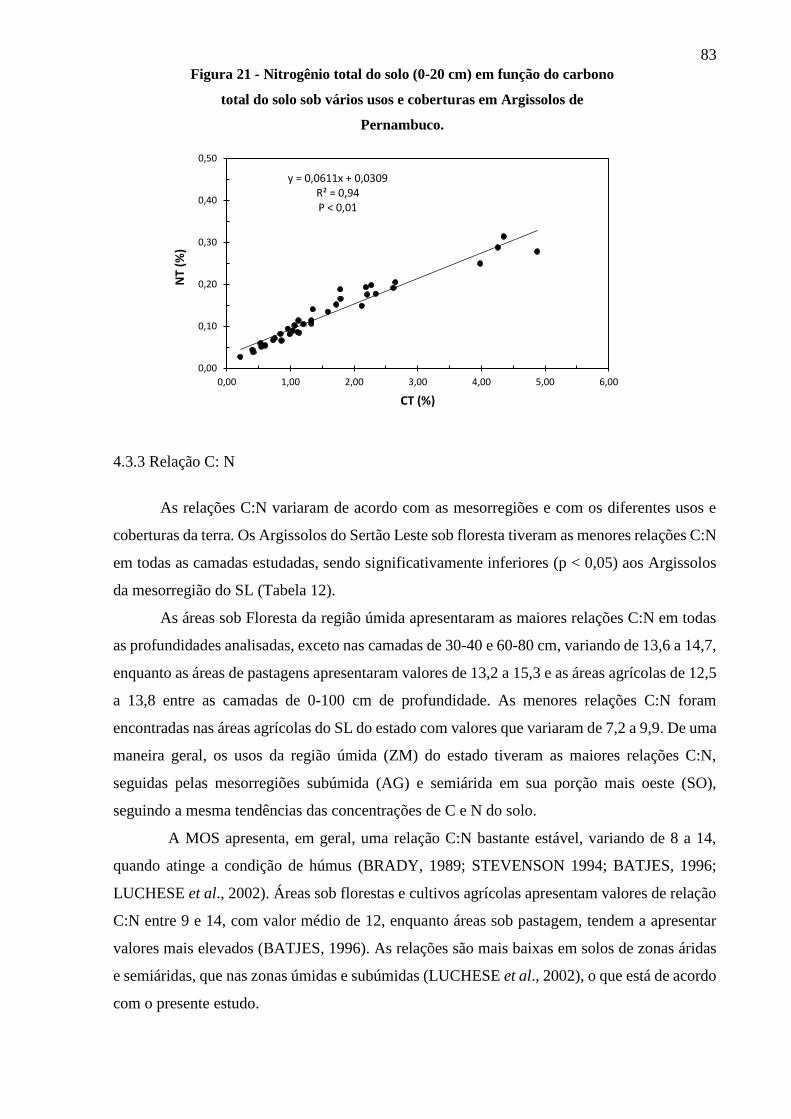
83
4.3.3 Relação C: N
As relações C:N variaram de acordo com as mesorregiões e com os diferentes usos e
coberturas da terra. Os Argissolos do Sertão Leste sob floresta tiveram as menores relações C:N
em todas as camadas estudadas, sendo significativamente inferiores (p < 0,05) aos Argissolos
da mesorregião do SL (Tabela 12).
As áreas sob Floresta da região úmida apresentaram as maiores relações C:N em todas
as profundidades analisadas, exceto nas camadas de 30-40 e 60-80 cm, variando de 13,6 a 14,7,
enquanto as áreas de pastagens apresentaram valores de 13,2 a 15,3 e as áreas agrícolas de 12,5
a 13,8 entre as camadas de 0-100 cm de profundidade. As menores relações C:N foram
encontradas nas áreas agrícolas do SL do estado com valores que variaram de 7,2 a 9,9. De uma
maneira geral, os usos da região úmida (ZM) do estado tiveram as maiores relações C:N,
seguidas pelas mesorregiões subúmida (AG) e semiárida em sua porção mais oeste (SO),
seguindo a mesma tendências das concentrações de C e N do solo.
A MOS apresenta, em geral, uma relação C:N bastante estável, variando de 8 a 14,
quando atinge a condição de húmus (BRADY, 1989; STEVENSON 1994; BATJES, 1996;
LUCHESE et al., 2002). Áreas sob florestas e cultivos agrícolas apresentam valores de relação
C:N entre 9 e 14, com valor médio de 12, enquanto áreas sob pastagem, tendem a apresentar
valores mais elevados (BATJES, 1996). As relações são mais baixas em solos de zonas áridas
e semiáridas, que nas zonas úmidas e subúmidas (LUCHESE et al., 2002), o que está de acordo
com o presente estudo.
y = 0,0611x + 0,0309R² = 0,94P < 0,01
0,00
0,10
0,20
0,30
0,40
0,50
0,00 1,00 2,00 3,00 4,00 5,00 6,00
NT
(%)
CT (%)
Figura 21 - Nitrogênio total do solo (0-20 cm) em função do carbono
total do solo sob vários usos e coberturas em Argissolos de
Pernambuco.
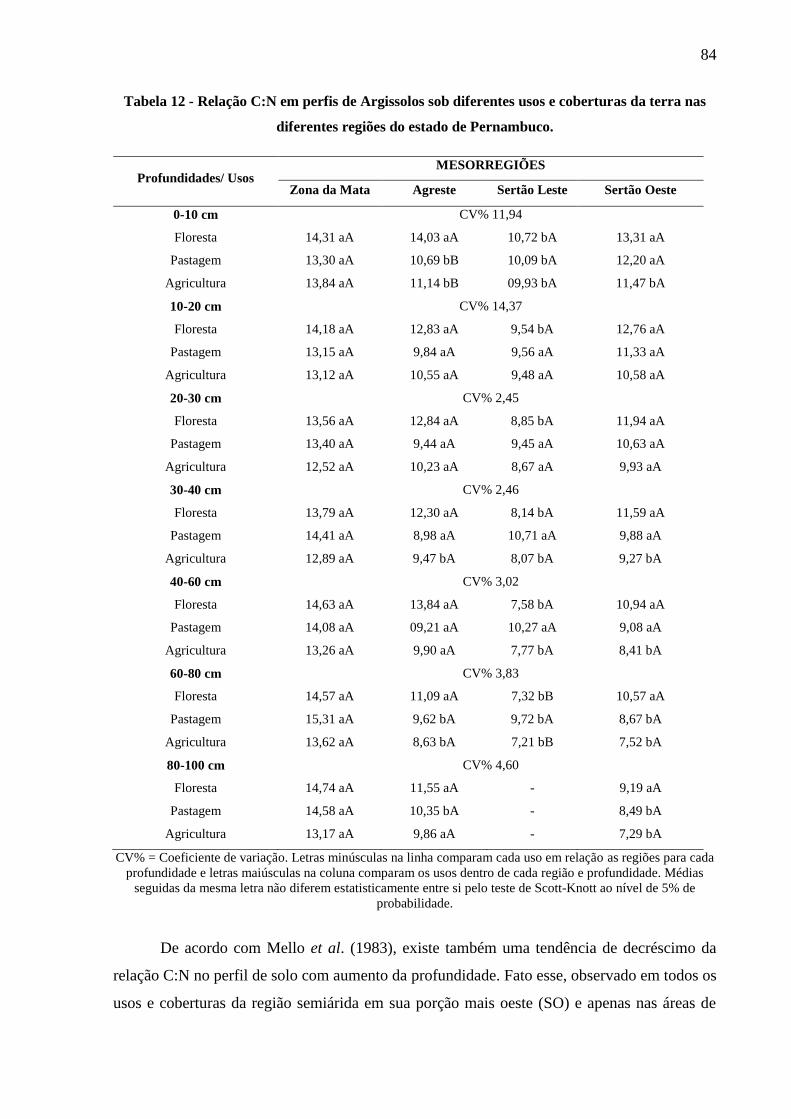
84
Tabela 12 - Relação C:N em perfis de Argissolos sob diferentes usos e coberturas da terra nas
diferentes regiões do estado de Pernambuco.
Profundidades/ Usos MESORREGIÕES
Zona da Mata Agreste Sertão Leste Sertão Oeste
0-10 cm CV% 11,94
Floresta 14,31 aA 14,03 aA 10,72 bA 13,31 aA
Pastagem 13,30 aA 10,69 bB 10,09 bA 12,20 aA
Agricultura 13,84 aA 11,14 bB 09,93 bA 11,47 bA
10-20 cm CV% 14,37
Floresta 14,18 aA 12,83 aA 9,54 bA 12,76 aA
Pastagem 13,15 aA 9,84 aA 9,56 aA 11,33 aA
Agricultura 13,12 aA 10,55 aA 9,48 aA 10,58 aA
20-30 cm CV% 2,45
Floresta 13,56 aA 12,84 aA 8,85 bA 11,94 aA
Pastagem 13,40 aA 9,44 aA 9,45 aA 10,63 aA
Agricultura 12,52 aA 10,23 aA 8,67 aA 9,93 aA
30-40 cm CV% 2,46
Floresta 13,79 aA 12,30 aA 8,14 bA 11,59 aA
Pastagem 14,41 aA 8,98 aA 10,71 aA 9,88 aA
Agricultura 12,89 aA 9,47 bA 8,07 bA 9,27 bA
40-60 cm CV% 3,02
Floresta 14,63 aA 13,84 aA 7,58 bA 10,94 aA
Pastagem 14,08 aA 09,21 aA 10,27 aA 9,08 aA
Agricultura 13,26 aA 9,90 aA 7,77 bA 8,41 bA
60-80 cm CV% 3,83
Floresta 14,57 aA 11,09 aA 7,32 bB 10,57 aA
Pastagem 15,31 aA 9,62 bA 9,72 bA 8,67 bA
Agricultura 13,62 aA 8,63 bA 7,21 bB 7,52 bA
80-100 cm CV% 4,60
Floresta 14,74 aA 11,55 aA - 9,19 aA
Pastagem 14,58 aA 10,35 bA - 8,49 bA
Agricultura 13,17 aA 9,86 aA - 7,29 bA
CV% = Coeficiente de variação. Letras minúsculas na linha comparam cada uso em relação as regiões para cada
profundidade e letras maiúsculas na coluna comparam os usos dentro de cada região e profundidade. Médias
seguidas da mesma letra não diferem estatisticamente entre si pelo teste de Scott-Knott ao nível de 5% de
probabilidade.
De acordo com Mello et al. (1983), existe também uma tendência de decréscimo da
relação C:N no perfil de solo com aumento da profundidade. Fato esse, observado em todos os
usos e coberturas da região semiárida em sua porção mais oeste (SO) e apenas nas áreas de

85
Floresta da porção leste (SL). Os demais usos e coberturas das mesorregiões úmida (ZM) e
subúmida (AG) não apresentaram tal característica (Tabela 12).
Os valores de relação C:N em áreas agrícolas encontrados nesse estudo são maiores aos
encontrados por Pedra et al. (2012) que reportaram valores de relação C:N da ordem de 11 a 13
em áreas sob cultivo convencional com uso de aração e gradagem na camada de 0-20 cm em
um Argissolo Vermelho-Amarelo da região úmida de Sergipe e aos encontrados por Fracetto et
al. (2012), que avaliaram a camada superficial (0-30 cm) de áreas agrícolas cultivadas com
mamona e também de áreas de caatinga preservada em um Latossolo Vermelho Eutrófico de
textura argilosa no semiárido da Bahia. Os autores reportaram valores de relação C:N da ordem
de 8 a 9, e de 5 a 12, paras as áreas agrícolas cultivadas com mamona e caatinga preservada,
respectivamente.
Os usos e coberturas da terra avaliados nesse estudo (floresta, pastagem e agricultura)
não diferiram entre si nas mesorregiões, exceto na camada de 0-10 cm na zona subúmida (AG),
onde as áreas de floresta foram superiores as demais, e na camada 80-100 cm no SL, onde as
áreas sob pastagem foram superiores (Tabela 12).
4.3.4 Abundância natural de δ13C
As abundâncias naturais de 13C (‰) praticamente não variaram entre as regiões em todas
as camadas de solo, com exceção das camadas de 0-10 cm, onde foram maiores nas áreas de
pastagem e agricultura e na camada de 80-100 cm nas áreas de Floresta. Não houve efeito
significativo da precipitação ou altitude ou CT com a abundância natural de 13C na camada
superficial do solo (0-20 cm). Já os usos e coberturas da terra (Floresta, Pastagem e
Agricultura), apresentaram diferenças significativas entre si dentro das mesorregiões ZM, AG
e SO, exclusivamente nas camadas superficiais do solo (até 40 cm) (Figura 22).
As áreas sob Pastagem das regiões do SL (-22,85) e SO (-24,81) apresentaram os
menores valores de δ13C ‰ na camada de 0-10 cm, diferindo estatisticamente (p < 0,05) das
mesorregiões úmida (-20,13) e subúmida (-19,80). Nas áreas de Floresta, não foram observadas
diferenças significativas entre as mesorregiões estudadas para a camada de 0-10 cm. Entretanto,
tais diferenças foram observadas na última camada do perfil (80-100 cm), onde as áreas de
Floresta da zona úmida (ZM) apresentaram os menores valores (-24,98) diferindo das demais
mesorregiões para esse uso (Figura 22).
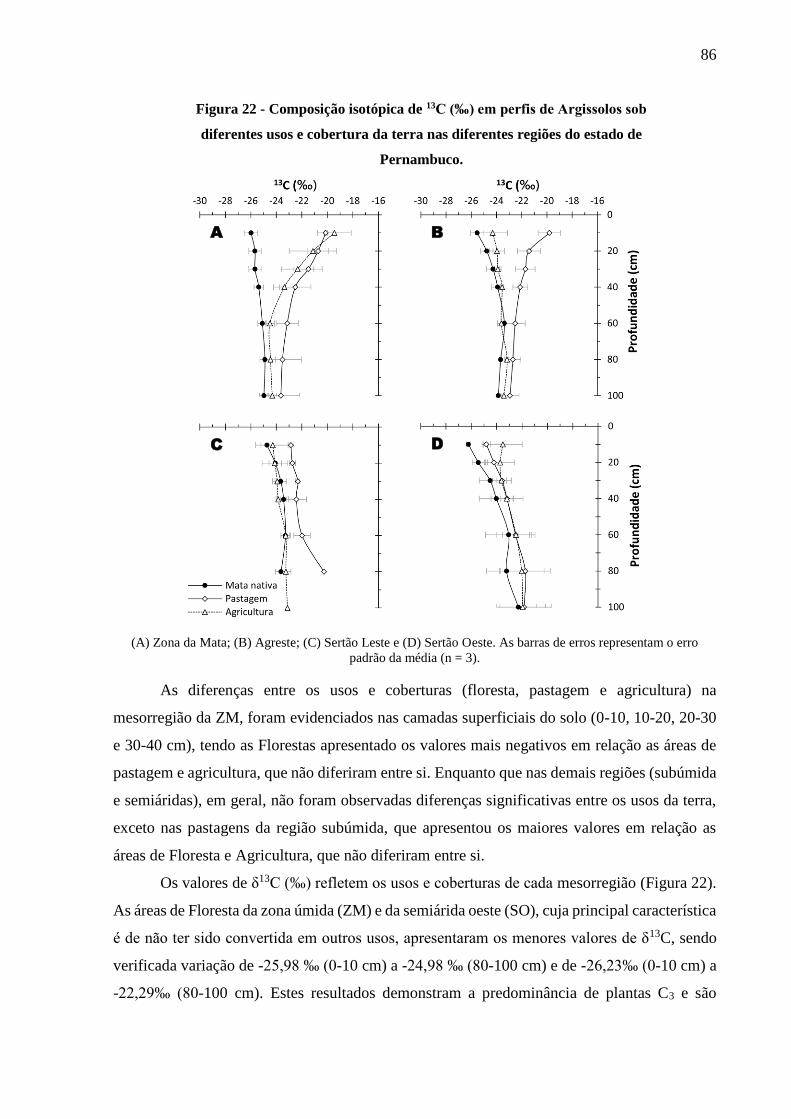
86
(A) Zona da Mata; (B) Agreste; (C) Sertão Leste e (D) Sertão Oeste. As barras de erros representam o erro
padrão da média (n = 3).
As diferenças entre os usos e coberturas (floresta, pastagem e agricultura) na
mesorregião da ZM, foram evidenciados nas camadas superficiais do solo (0-10, 10-20, 20-30
e 30-40 cm), tendo as Florestas apresentado os valores mais negativos em relação as áreas de
pastagem e agricultura, que não diferiram entre si. Enquanto que nas demais regiões (subúmida
e semiáridas), em geral, não foram observadas diferenças significativas entre os usos da terra,
exceto nas pastagens da região subúmida, que apresentou os maiores valores em relação as
áreas de Floresta e Agricultura, que não diferiram entre si.
Os valores de δ13C (‰) refletem os usos e coberturas de cada mesorregião (Figura 22).
As áreas de Floresta da zona úmida (ZM) e da semiárida oeste (SO), cuja principal característica
é de não ter sido convertida em outros usos, apresentaram os menores valores de δ13C, sendo
verificada variação de -25,98 ‰ (0-10 cm) a -24,98 ‰ (80-100 cm) e de -26,23‰ (0-10 cm) a
-22,29‰ (80-100 cm). Estes resultados demonstram a predominância de plantas C3 e são
A B
D C
Figura 22 - Composição isotópica de 13C (‰) em perfis de Argissolos sob
diferentes usos e cobertura da terra nas diferentes regiões do estado de
Pernambuco.

87
característicos de solos sob florestas nativas no Brasil. Valores semelhantes aos encontrados
nesse estudo foram obtidos por Tarré et al. (2001), Balbinot (2009) e Pinheiro et al. (2010), em
estudos também realizados na Mata Atlântica, assim como os realizados em outros biomas
brasileiros (ROSCOE et al., 2000; JANTALIA et al., 2007; MARTINELLI et al., 2009;
COSTA JÚNIOR et al., 2011; ARAÚJO et al., 2011).
As mudanças mais significativas nos valores de 13C ocorreram nos Argissolos sob
pastagem e agricultura na região úmida (ZM), e sob pastagem na região subúmida (AG) até a
profundidade de 40 cm (Figura 22), indicando alterações na cobertura vegetal que, em um
passado recente, passou por transformação antrópica, mudando de vegetação do ciclo C3 para
vegetação do ciclo C4. Estes resultados indicam que a contribuição de plantas C4 na matéria
orgânica do solo nessas regiões para esses usos é de vários anos atrás (± 50 anos) porque já
atingiu camadas mais profundas do solo, como também observado por Smith e Epstein (1971).
Nas áreas sob Mata nativa de uma maneira geral, os perfis de δ13C (0-100 cm) não
apresentaram alterações bruscas, indicando ocupação contínua por vegetação do ciclo C3
(Figura 22). Verificou-se nessas mesmas áreas um enriquecimento isotópico em profundidade,
com diferenças de 1,00; 1,67; 1,11 e 3,94 ‰ menos negativo, da superfície até o intervalo entre
80-100 cm paras as mesorregiões úmida (ZM), subúmida (AG), semiárida Leste (SL) e
semiárida Oeste (SO), respectivamente. Esse padrão de enriquecimento de δ13C em
profundidade, também tem sido reportado por outros autores (SALIMON, 2003; TELLES et
al., 2003) e pode estar relacionado tanto com a migração e redeposição de partículas de argila
que normalmente carregam maior valor de δ13C, como as diferenças que ocorrem nas taxas de
decomposição de resíduos orgânicos com diferentes assinaturas de δ13C, como celulose e
hemicelulose, que são frequentemente 1-2 ‰ mais enriquecidos em δ13C, enquanto a lignina é
2-6 ‰ inferior em relação ao valor δ13C do tecido total da planta (LIU et al., 2014).
De acordo com Martinelli et al. (2009), quando a vegetação predominante é do tipo C3
e não existem vestígios de mudança no tipo de vegetação em tempos longínquos, o valor de
δ13C da MOS aumenta em direção as camadas mais profundas do solo. Geralmente, o
incremento nos valores de δ13C da MOS é cerca de 3 a 4‰ entre a camada superficial e as
camadas mais profundas. Padrão esse, também observado nas áreas de Floresta do semiárido
Oeste (SO), indicando que aquela vegetação sempre foi composta por plantas C3 e menos
evidente nas demais mesorregiões do estado.

88
4.3.5 Abundância natural de δ15N
Os valores referentes a abundância natural de δ15N (‰), apresentaram diferenças
estatísticas entre as mesorregiões para cada uso e cobertura da terra. Embora, não tenham
apresentado diferenças entre os usos dentro de cada mesorregião isoladamente (Figura 23).
(A) Zona da Mata; (B) Agreste; (C) Sertão Leste e (D) Sertão Oeste. As barras de erros representam o erro
padrão da média (n = 3).
Nas áreas sob floresta, os maiores valores foram encontrados nas regiões semiáridas do
sertão (SL e SO), que apresentaram-se significativamente superiores aos Argissolos da regiões
úmida (ZM) e subúmida (AG), embora essas diferenças só tenham sido evidenciadas nas
camadas superficiais de 0-10 cm e 10-20 cm do solo (Figura 23).
A abundância natural de δ15N da camada superficial do solo (0-20 cm) variou
significativamente entre as mesorregiões, correlacionando-se negativamente de forma potencial
com a precipitação pluvial (Figura 24 A) e linearmente com os teores de CT e NT do solo
A B
D C
Figura 23 - Variação dos valores de 15N (‰) em perfis de Argissolos sob diferentes
usos e cobertura da terra nas diferentes mesorregiões do estado de Pernambuco.

89
(Figura 24 C, D) e de forma quadrática com a altitude, sendo os maiores valores encontrados
com altitudes intermediárias (entre 500 e 700 m), figura 24 B.
Figura 24 - Abundância natural de 15N do solo (0-20 cm) em função da precipitação pluvial,
altitude, carbono total e nitrogênio total do solo sob diversos usos e coberturas em Argissolos de
Pernambuco.
Os resultados encontrados nesse estudo, demostram haver um enriquecimento isotópico
de δ15N das mesorregiões semiáridas onde as temperaturas são mais altas e as chuvas mais
escassas em relação as áreas úmidas e subúmidas do estado. A explicação é que, em locais
úmidos, a reciclagem interna de N em forma orgânica é grande e as perdas de N mineral são
pequenas, causando dessa maneira um enriquecimento em 15N. Em locais mais secos,
proporcionalmente mais N pode permanecer em formas inorgânicas e ser perdido por
volatilização ou lixiviação. Em escalas menores, no nível de paisagem, locais com maior
disponibilidade de água, como depressões topográficas, em geral possuem vegetação com
y = 372,36x-0,514
R² = 0,7315P < 0,01
0
2
4
6
8
10
12
14
16
0 500 1000 1500 2000
δ1
5 N (‰)
Precipitação (mm)
A
y = -6E-05x2 + 0,0661x - 7,3506R² = 0,6356
P < 0,05
200 400 600 800
Altitude (m)
B
y = -1,8802x + 14,283R² = 0,7341
P < 0,01
0
2
4
6
8
10
12
14
16
0 1 2 3 4
δ1
5 N (‰
)
CT (%)
C
y = -28,25x + 14,903R² = 0,6665
P < 0,01
0,00 0,10 0,20 0,30
NT (%)
D

90
folhas mais enriquecidas que locais mais secos, o que é relacionado a maiores taxas de
mineralização e desnitrificação (GARTEN, 1993; HANDLEY e SCRIMGEOUR, 1997).
Outros trabalhos publicados na literatura, também encontraram fortes correlações entre
a composição isotópica do N do solo disponível para as plantas e a disponibilidade de água
(Precipitação). Esse padrão de enriquecimento ou empobrecimento é diferente para
comparações em diferentes escalas. Em escalas globais (HANDLEY e SCRIMGEOUR, 1997;
HANDLEY, 1999), grandes transectos geográficos (ARANIBAR et al., 2004; SWAP et al.,
2004) e em escalas regionais (OLIVEIRA, 2014; FREITAS et al., 2015), a tendência é sempre
de diminuição do sinal de 15N das folhas à medida que a precipitação pluvial média anual
aumenta. Dessa forma, estando também associada a quantidade MOS, uma vez que regiões de
maior regime pluvial resultam em maiores aportes de C no solo.
Os resultados também corroboram o trabalho de Mendonça et al. (2010) que ao avaliar
o teor de δ15N de áreas de Cerradão e de Caatinga em solos da Chapada do Araripe, CE,
observaram que os menores valores de δ15N estavam associados à maior quantidade de CT e
foram encontrados no solo de Cerradão. Na Caatinga, onde a superfície do solo apresentou
menor conteúdo de CT, encontraram-se os maiores valores de δ15N, o que é um indício de
mineralização. Resultados também reportados por Loss et al. (2011 e 2014).
Nas áreas sob Pastagem, as diferenças foram verificadas nas camadas de 10-20, 20-30,
60-80 e 80-100 cm. Em todas essas profundidades observou-se superioridade dos Argissolos
das regiões do AG e do SO sobre os Argissolos das regiões da ZM e do SL (Figura 24). Não
correspondendo dessa forma, a tendência observada nas áreas de floresta, de enriquecimento
isotópico ao longo do gradiente climático (precipitação) à medida que se afasta das áreas mais
úmidas do estado em direção as regiões semiáridas.
Os valores de δ15N do solo encontrados nas áreas de pastagem e de floresta da ZM foram
os menores entre todos os usos e mesorregiões do estado. Os valores variaram de 6,72 a 7,90
‰ e de 8,02 a 9,75 ‰ de 0-100 cm para as áreas de pastagem e floresta, respectivamente. Os
menores valores paras as áreas de PAS foram encontrados nas duas camadas mais profundas
do perfil (6,95 ‰ de 60-80 cm e 6,72 ‰ de 80-100 cm), enquanto que nas áreas de floresta os
menores valores foram verificados nas camadas superficiais (8,02 ‰ de 0-10 cm e 8,40 de 10-
20 cm).
Os resultados encontrados nessas áreas, diferem dos resultados encontrados por Freitas
et al., 2015 em áreas de floresta da zona úmida da região Nordeste do Brasil. Os autores
reportaram valores médios de δ15N do solo variando entre 3 e 6 ‰ na camada de 0-20 cm e
sugeriram portanto, que essas áreas com baixos valores de δ15N podem ter ciclos fechados com

91
imobilização rápida ou absorção de N mineralizado ou, alternativamente, elevadas perdas
compensadas pela alta fixação e baixa deposição. Por outro lado, os resultados desse estudo
corroboram o trabalho de Mendonça et al., 2010, que reportaram valores médios de δ15N do
solo (0-140 cm) entre 5 e 9 ‰ para áreas de floresta úmida, transição floresta úmida/cerradão,
e 3 tipos de cerradão/gramíneas, e de 13 a 15‰ para áreas de caatinga no extremo sul do estado
do Ceará, no nordeste do Brasil.

92
5 CONCLUSÕES
Estudo 1 - Levantamento dos estoques de carbono nos solos de Pernambuco.
1. As estimativas dos estoques de carbono nos solos de Pernambuco evidenciaram o
elevado potencial das classes dos Chernossolos, Nitossolos, Vertissolos e Gleissolos em estocar
C, principalmente nos biomas Mata Atlântica e Caatinga sem intervenção antrópica.
2. Ainda que, as classes dos Argissolos, Neossolos Litólicos, Planossolos e Latossolos
não estoquem as maiores quantidades de C por área, são essas classes, que recobrem as maiores
extensões territoriais do Estado e consequentemente, apresentam os maiores estoques totais de
C no solo para a camada de 0-30 cm.
3. Estão estocados em Pernambuco, aproximadamente 368 Tg de C (0-30 cm) e a média
geral dos estoques de C, ponderada pela proporção de área das classes de solo, é da ordem de
38 Mg ha-1 na camada de 0-30 cm.
Estudo 2 - Distribuição em profundidade e estoques de carbono em solos sob diferentes
coberturas e usos da terra em Pernambuco.
1. Os teores de CT correlacionam-se negativamente com a profundidade do solo de
forma quadrática (exponencial ou potencial);
2. Os maiores estoques de CT ocorreram nas vegetações nativas densas e decresceram
da região úmida à semiárida Oeste, com grandes variações entre as classes solos. As camadas
superficiais até 30 cm de profundidade estocam cerca de 40 a 60% do CT total do solo até 1 m;
3. As maiores reduções nos estoques de CT do solo com a mudança de uso da terra
(agricultura e pastagem) em relação as áreas de referência (vegetação densa) foram verificadas
nas regiões subúmidas e semiárida Leste, com reduções que variam de 34 a 55% e de 16 a 27%,
respectivamente.

93
Estudo 3 - Carbono, nitrogênio e abundância natural (δ13C e δ15N) do solo sob diferentes
usos ao longo de um gradiente climático em Pernambuco.
1. Não foram encontradas diferenças nas concentrações de C e N nos Argissolos entre
as áreas antropizadas (áreas agrícolas e áreas de pastagem) e as áreas de Floresta.
2. As maiores concentrações de CT e NT foram encontradas nas áreas de floresta das
regiões úmidas e subúmidas com precipitações entre 900 e 1700 mm;
3. A substituição da vegetação de floresta por pastagens ou cultivos agrícolas levou a
aumentos do sinal de δ13C do solo, pela incorporação de C derivado de plantas C4, sendo
maiores nas áreas úmidas e subúmidas do estado em relação às áreas semiáridas;
4. Os valores de δ15N do solo são inversamente relacionados às concentrações de C e de
N. O enriquecimento de δ15N encontrados na superfície das mesorregiões semiáridas em relação
às áreas úmidas e subúmidas estão associados as menores concentrações de C e N presentes nos
Argissolos dessa região, em decorrência das maiores temperaturas e menores índices
pluviométricos.
Conclusões gerais
O carbono e o nitrogênio nos solos de Pernambuco são regulados por fatores
edafoclimáticos, como a precipitação pluvial, altitude/temperatura, textura e pela profundidade
do solo. Em função do maior armazenamento em superfície, é comum o modelo exponencial
ou potencial se ajustarem aos teores de C e N no perfil de solo. Os maiores estoques de C nos
solos do Estado estão alocados em áreas de florestas preservadas das regiões fisiográficas com
maiores precipitações pluviais e sobretudo nas camadas superficiais das classes de solo com
maior conteúdo de argila. A intervenção humana nessas áreas através de práticas agropecuárias
convencionais, reduz os estoques de carbono no solo a patamares inferiores ao encontrados nas
áreas nativas, e modificam a dinâmica do C e do N no solo nessas regiões, contribuindo dessa
maneira, para a emissão GEE para a atmosfera. Para se contrapor a esta situação no Estado, são
necessárias medidas mitigatórias, como a adoção de sistemas conservacionistas do solo e da
água, a exemplo da rotação de culturas (gramíneas + leguminosas), integração Lavora-Pecuária-
Floresta (iLPF) Silviculturais, dentre outras, através de políticas públicas de incentivo às
práticas conservacionistas em substituição à agricultura de baixo carbono.

94
REFERÊNCIAS
ACCIOLY, L. J. O.; SILVA, E. A; CAVALCANTI JUNIOR, E. A.; SILVA, A.; ALVES, E.
S.; PEREIRA, A. G. S.; SILVA, R. S.; RAMOS, R. R. D. Mapeamento do uso e da cobertura
das terras do semiárido pernambucano (escala 1:100.000). Boletim de Pesquisa da Embrapa
Solos, 2017. No prelo.
ADDSCOTT, T. M. Entropy and sustainability. European Jornal of Soil Science, v.46, n.2,
p.161-168, 1995.
ADUAN, E. A.; VILELA, M. F.; KLINK, C. A. Ciclagem de carbono em ecossistemas
terrestres: o caso do cerrado brasileiro. Planaltina: Embrapa Cerrados, 2003. 30 p.
ALVES, B. J. R.; CAMPOS, D. V.; SISTI, C. P. J.; URQUIAGA, S.; BODDEY, R. M.
Emprego do isótopo estável 13C para o estudo da dinâmica da matéria orgânica do solo. In:
ROSCOE, R.; MERCANTE, F. M.; SALTON, J. C. (Eds.). Dinâmica da matéria orgânica
do solo em sistemas conservacionistas: modelagem matemática e métodos auxiliares. Dourados: Embrapa Agropecuária Oeste, p. 133-161, 2006.
ALVES, B. J. R.; ZOTARELLI, L.; JANTALIA, C. P.; BODDEY, R. M.; URQUIAGA, S.
Emprego de isótopos estáveis para o estudo do carbono e do nitrogênio no sistema solo-planta.
In: AQUINO, A. M.; ASSIS, R. L. (Eds). Processos biológicos no sistema solo-planta:
Ferramentas para uma agricultura sustentável. Brasília, Embrapa-SCT. p. 343-350. 2005.
AMORIM, L. B. Caracterização da serapilheira em caatinga preservada e mudanças no carbono
do solo após desmatamento sem queima. Dissertação de Mestrado (Mestrado em Ciência do
Solo) – Recife: Departamento de Agronomia, Universidade Federal Rural de Pernambuco,
2009, 66 p.
AMUNDSON, R.; AUSTIN, A. T.; SCHUUR, E. A. G.; YOO, K.; MATZEK, V.; KENDALL,
C.; UEBERSAX, A.; BRENNER, D.; BAISDEN, W. T. Global patterns of the isotopic
composition of soil and plant nitrogen. Global Biogeochemical Cycles, v. 17, n. 1, p. 31.1-
31.10, 2003.
ANDERSON, J. M. Responses of soils to climate change. Advances in Ecological Research,
v. 22, p. 63-91, 1992.
ANDREUX, F. Humus in word soils. In PICCOLO, A. (Ed.) Humic substances in terrestrial
ecosystems. Amsterdam: Elsevier, p. 45-100, 1996.
ANTUNES, P. D.; SAMPAIO, E. V. S. B.; FERREIRA JÚNIOR, A. L. G.; GALINDO, I. C.
L.; SALCEDO, I. H. Distribuição de 137Cs em três solos representativos do estado de
Pernambuco. Revista Brasileira de Ciência do Solo, Viçosa-MG, v. 34, p. 935-943, 2010.
ARANIBAR, J. N.; OTTER, L.; MACKO, S. A.; FERAL, C. J. W.; EPSTEIN, H.E.; DOWTY,
P. R.; ECKARDT, F.; SHUGART, H. H.; SWAP, R. J. Nitrogen cycling in the soil-plant system
along a precipitation gradient in the Kalahari sands. Global Change Biology, v. 10, p. 359-373,
2004.

95
ARAÚJO FILHO, J. C; BURGOS, N.; LOPES, O. F.; SILVA, F. H. B. B; MEDEIROS, L. A.
R; MELO FILHO, H. F. R; PARAHYBA, R. B. V; CAVALCANTI, A. C.; OLIVEIRA NETO,
M. B.; SILVA, F. B. R.; LEITE, A. P.; SANTOS, J. C. P.; SOUSA NETO, N. C.; SILVA, A.
B.; LUZ, L. R. Q. P.; LIMA, P. C.; REIS, R. M. G.; BARROS, A. H. Levantamento de
reconhecimento de baixa e média intensidade dos solos do estado de Pernambuco. Rio de
Janeiro: Embrapa solos, Boletim de Pesquisa, n. 11, 382 p., 2000.
ARAÚJO, A. M. S.; SAMPAIO, E. V. S. B.; SALCEDO, I. H. Mineralização do C e do N em
amostras de solo cultivado com Cana-de-açúcar, ao longo de dez anos com e sem fertilização
nitrogenada. Revista Brasileira de Ciência do Solo, v. 25, p. 43-53, 2001.
ARAÚJO, M. S. B.; SALCEDO, I. H. Formas preferenciais de acumulação de fósforo em solos
cultivados com Cana-de-açúcar na região Nordeste. Revista Brasileira de Ciência do Solo,
Viçosa-MG, v. 21, p. 643-650, 1997.
ARAÚJO, M. S. B.; SAMPAIO, E. V. S. B.; SILVEIRA, M. M. L. Estoques de fósforo em
solos do semiárido de Pernambuco e da Paraíba. In: MENEZES, R. S. C.; SAMPAIO, E. V. S.
B.; SALCEDO, I. H. Fertilidade do solo e produção de biomassa no semiárido. Recife:
Editora Universitária da UFPE, p. 47-61, 2008.
ARAÚJO, M. S. B.; SCHAEFER, C. E. G. R.; SAMPAIO, E. V. S. B. Frações de fósforo após
extrações sucessivas com resina e incubação, em Latossolos e Luvissolos do semiárido de
Pernambuco. Revista Brasileira de Ciência do Solo, v. 28, p. 259-268, 2004.
ARROUAYS, D.; PELISSIER, P. Modeling carbon storage profiles in temperate forest humic
loamy soils of France. Soil Science, v. 157, n. 157, p. 185-192, 1994.
ASSIS, C.P.; OLIVEIRA, T.S.; DANTAS, J.A.N. & MENDONÇA, E.S. Organic matter and
phosphorus fractions in irrigated agroecosystems in a semi-arid region of Northeastern Brazil.
Agric. Ecosystem Environment, v. 138, p. 74-82, 2010.
AUSTIN, A. T.; VITOUSEK, P. M. Nutrients dynamics on a precipitation gradient in Hawaii.
Oecologia, v. 113, p. 519-529, 1998.
BALBINOT, R. Carbono, nitrogênio e razões isotópicas δ13C e δ15N no solo e vegetação de
estágios sucessionais de Floresta Ombrófila Densa Submontana. Tese de Doutorado, Curitiba:
Universidade Federal do Paraná, 2009.
BALESDENT, J.; CHENU, C., BALABANE, M. Relationship of soil organic matter dynamics
to physical protection and tillage. Soil and Tillage Research, Amsterdam, v. 53, p. 215-230,
2000.
BALESDENT, J.; WAGNER, G. H.; MARIOTTI, A. Soil organic matter turnover in long-term
field experiments as revealed by 13C natural abundance. Soil Science Society of America
Journal, v. 52, p. 118–124, 1988.
BALESDENT, J; MARIOTTI, A; GUILLET, B. Natural 13C abundance as a tracer for studies
of soil organic matter dynamics. Soil Biology and Biochemistry, v. 19, p. 25-30, 1987.

96
BARRETO, A. C.; FREIRE, M. B. G. S.; NACIF, P. G. S.; ARAÚJO, Q. R.; FERIRE, F. J.;
INÁCIO, E. S. B. Fracionamento químico e físico do carbono orgânico total em um solo de
mata submetido a diferentes usos. Revista Brasileira de Ciência do Solo, v. 32, p. 1471-1478,
2008.
BATJES, N. H. Options for increasing carbon sequestration in West African soils an
exploratory study with special focus on Senegal. Land Degradation Development, v. 12,
p.131-142, 2001.
BATJES, N. H. Organic carbon stocks in the soils of Brazil. Soil Use and Management,
Oxford, v. 21, p. 22-24, 2005.
BATJES, N. Total carbon and nitrogen in the soils of the world. European Journal of Soil
Science, v. 47, p.151-163, 1996.
BAYER, C. e MIELNICZUK, J. Características químicas do solo afetadas por métodos de
preparo e sistemas de cultura. Revista Brasileira de Ciência do Solo, v. 21, p. 105-112, 1997.
BAYER, C.; MARTIN-NETO, L.; MIELNICZUK, J.; CERETTA, C. A. Effect of no-till
cropping systems on soil organic matter in a sandy clay loam Acrisol from Southern Brazil
monitored by electron spin resonance and nuclear magnetic resonance. Soil and Tillage
Research, v. 53, p. 95-104, 2000.
BAYER, C.; MARTIN-NETO, L.; MIELNICZUK, J.; PAVINATO, A.; DIECKOW, J. Carbon
sequestration in two Brazilian Cerrado soils under no-till. Soil and Tillage Research, v.86,
p.237-245, 2006.
BAYER, C.; MIELNICZUK, J.; AMADO, T. J. C.; MARTINNETO, L.; FERNANDES, S. A.
Organic matter storage in a sandy clay loam Acrisol affected by tillage and cropping systems
in southern Brazil. Soil and Tillage Research, v. 54, p. 101-109, 2000.
BENITES, V. M.; MACHADO, P. L. O. A.; FIDALGO, E. C. C.; COELHO, M. R.; MADARI,
B. E. Pedotransfer functions for estimating bulk density of Brazilian soils. Geoderma,
Amsterdam, NL, v. 139, p. 90-97, 2007.
BERNOUX, M. and VOLKOFF, B. Soil carbon stock in soil ecoregions of Latin America. In:
LAL, R.; CERRI, C.C.; BERNOUX, M.; ETCHEVERS, J.; CERRI, C. E. P. Carbon
sequestration in soils of Latin America. New York, Haworth, p. 65-75, 2006.
BERNOUX, M.; ARROUAYS, D.; CERRI, C.; VOLKOFF, B.; JOLIVET, C. Bulk densities
of Brazilian Amazon soils related to other soil properties. Soil Science Society of America
Journal, Madison, v. 62, p. 743-749, 1998.
BERNOUX, M.; CARVALHO, M. C. S.; VOLKOFF, B.; CERRI, C. C. Brazil’s soil carbono
stocks. Soil Science Society of America Journal, Madson, v. 66, p. 888-896, 2002.

97
BERNOUX, M.; CERRI, C. C.; CERRI, C. E. P.; SIQUEIRA NETO, M.; METAY, A.;
PERRIN, A.; SCOPEL, E.; BLAVET, D.; PICCOLO, M. C. Influence du semis direct avec
couverture végétale sur la séquestration du carbone et érosion au Brésil. In: ROOSE, E.; DE
NONI, G.; PRAT C.; GANRY, F.; BOURGEON, G. (Eds.). Gestion de la biomasse, érosion
et séquestration du carbone. Bulletin Du Reseau Erosion, v. 23, p. 323-337. 2004.
BLAIR N.; LEU A.; MUNOZ, E.; OLSEN, J.; KWONG, E.; D. DESMARAIS. Carbon isotopic
fractionation in heterotrophic microbial metabolism. Applied and Environmental
Microbiology, v. 50, p. 996–1001, 1985.
BONFIM, E. M. S.; FREIRE, F. J.; SANTOS, M. V. F.; SILVA, T. J. A.; FREIRE, M. B. G.
S. Níveis críticos de fósforo para Brachiaria brizantha e suas relações com características
físicas e químicas em solos de Pernambuco. Revista Brasileira de Ciência do Solo, Viçosa-
MG, v. 28, p. 281-288, 2004.
BORGES, A. L.; KIEHL, J. C. Alterações da matéria orgânica de um Latossolo amarelo Álico
de Cruz das Almas (BA), pelo cultivo com frutíferas perenes e mandioca. Revista Brasileira
de Ciência do Solo, v. 20, p. 313-318, 1996.
BOUTTON, T. W. Stable carbon isotope ratios of natural materials: I. Sample preparation and
mass spectrometric analysis. In: COLEMAN, D. C.; FRY, B. Carbon isotope techniques.
Academic Press, Inc. 1991.
BOUTTON, T. W. Stable carbon isotope ratios of soil organic matter and their use as indicators
of vegetation and climate change. In BOUTTON, T. W.; YAMASAKI, S. I. (Ed.). Mass
spectrometry of soils. Marcel Dekker, p. 47-82, 1996.
BOUTTON, T. W.; TYRRELL, H. F.; PATTERSON, B. W; VARGA, G. A.; KLEIN, P. D.
Carbon kinetics of milk formation in holstein cows in late lactation. Journal of Animal
Science, v. 66, p. 2336-2645, 1988.
BOX, G. E. P. and COX, D. R. An Analysis of Transformations. Journal of Royal Statistical
Society. B, 39, 211-252, 1964.
BRADY, N. C. Natureza e propriedades dos solos. Rio de Janeiro: Freitas Bastos, 1989. 898p.
BUSTAMANTE, M. M. C.; MARTINELLI, L. A.; SILVA, D. A.; CAMARGO, P. B.; KLINK.
C. A.; DOMINGUES T. F.; SANTOS R. V. 15N natural abundance in woody plants and soils
of central brazilian savannas (cerrado). Ecological Applications, Nova York, v. 14, p. 200-
213. 2004.
CADISCH, G.; IMHOF, H.; URQUIAGA, S.; BODDEY, R. M.; GILLER, K. E. Carbon
turnover (delta-c-13) and nitrogen mineralization potential of particulate light soil organic
matter after rainforest clearing. Soil Biology Biochemistry, v.28, p. 1555–1567, 1996.
CAMPOS, D. V. B. Uso da técnica 13C e fracionamento físico da matéria orgânica em solos
sob cobertura de pastagens e cana-de-açúcar na região da mata atlântica. Tese de Doutorado,
Universidade Federal Rural do Rio de Janeiro, Seropédica, 2003, 220 p.

98
CANADELL, J. G.; MOONEY, H. A.; BALDOCCHI, D. D.; BERRY, J. A.; EHLERINGER,
J. R.; FIELD, C. B.; GOWER, S. T.; HOLLINGER, D. Y.; HUNT, J. E.; JACKSON, R.B.;
RUNNIG, S. W.; SHAVER, G. R.; STEFFEN, W.; TRUMBORE, S. E.; VALENTINI, R.;
BOND, Y. Carbon metabolism of the terrestrial biosphere: a multitechnique approach for
improved understandin. Ecosystems. v. 3, p. 115-130. 2000.
CARTER, M. R. Organic matter and sustainability. In: REES, R. M., BALL, B. C.,
CAMPBELL, C. D.; WATSON, C. A. (Ed.). Sustainable management of soil organic matter.
CABI Publishing, p. 9-22, 2001.
CARVALHO J. L. N., CERRI C. E. P., FEIGL B. J., PICOLLO M. C., GODINHO V. P.,
CERRI C. C. Carbon sequestration in agricultural soils in the Cerrado region of the Brazilian
Amazon. Soil and Tillage Research, v.103, n. 2, p. 342-349. 2009.
CERRI, C. C.; BERNOUX, M.; CERRI, C. E. P.; LAL, R. Challenges and opportunities of soil
carbon sequestration in Latin America. In: LAL, R.; CERRI, C. C.; BERNOUX, M.;
ETCHEVERS, J.; CERRI, C. E. P. Carbon sequestration in soils of Latin America. New
York, Haworth, p.41-47, 2006.
CERRI, C. C.; BERNOUX, M.; VOLKOFF, B.; MORAES, J. L. Dinâmica do carbono nos
solos da Amazônia. In: ALVAREZ V., V. H.; FONTES, L. E. F.; FONTES, M. P. F. Os solos
nos grandes domínios morfoclimáticos do Brasil e o desenvolvimento sustentável. Sociedade
Brasileira de Ciência do Solo, p. 61-69, 1996.
CERRI, C.C.; BALESDENT, J.; FELLER, C.; VICTORIA, R.; PLENECASSAGNE, A.
Application du tracage isotopique naturel en 13C, à l’étude de la dynamique de la matière
organique dans les sols. Comptes Rendus de l’Academie dês Sciences de Paris, t. 300, Série
II, nº 9, p. 423-428, 1985.
CHAPIN F. S.; MOONEY, H. A.; CHAPIN, M C.; MATSON, P. Principles of terrestrial
ecosystem ecology. Springer. 2002.
CLAY, D. E.; CLAPP, C. E.; RESSE, C.; LIU, Z.; CARLSON, C. G. WOODARD, H; BLY,
A. Carbon-13 fractionation of relic soil organic carbon during mineralization and effects
calculated half-lives. Soil Science Society of America Journal, v. 71, n. 3, p. 1003-1009, 2007.
CONDEPE/FIDEM - AGÊNCIA ESTADUAL DE PLANEJAMENTO E PESQUISAS DE
PERNAMBUCO. Aspectos geoambientais e socioeconômicos da bacia hidrográfica do Rio
Una, grupos de bacias hidrográficas de pequenos rios litorâneos GL 4 e GL 5. Recife, PE: Série
Bacias Hidrográficas de Pernambuco, v. 3, 85 p. 2006.
CORAZZA, E. J.; SILVA, J. E.; RESCK, D. V. S.; GOMES, A. C. Comportamento de
diferentes sistemas de manejo como fonte ou depósito de carbono em relação à vegetação de
cerrado. Revista Brasileira de Ciência do Solo, Campinas, n. 23, p. 425-432, 1999.
COSTA JUNIOR, C.; PICCOLO, M. C.; SIQUEIRA NETO, M.; CAMARGO, P. B.; CERRI,
C. C.; BERNOUX, M. Carbono total e 13C em agregados do solo sob vegetação nativa e
pastagem no bioma cerrado. Revista Brasileira de Ciência do Solo, Viçosa, v. 35, n. 4, p.
1241-1252. 2011a.

99
COSTA JUNIOR, C.; SIQUEIRA NETO, M.; PICCOLO, M. C.; CAMARGO, P. B.
Nitrogênio e abundância natural de 15N em agregados do solo sob diferentes usos da terra no
Bioma Cerrado. Ensaios e Ciência: Ciências Biológicas, Agrárias e da Saúde, v. 15, p. 47-
66, 2011b.
COSTA, A. R.; MADARI, B. E.; CARVALHO, M. T. M.; MACHADO, P. L. O. A.
BERNARDES, T. G.; SILVEIRA, P. M. Uso do nitrogênio na agricultura e suas implicações
na emissão do gás de efeito estufa óxido nitroso (N2O). Santo Antônio de Goiás, GO: Embrapa
Arroz e Feijão. Documentos, v. 249, 47 p. 2009.
COSTA, J. B. Caracterização e Constituição do Solo. 7ª ed. Lisboa, Ed. Fundação Calouste,
2004.
COSTA, L. A. M. Radiação solar global: Algumas considerações sobre seus efeitos no solo em
que se incorporou efluente de biodigestor. Dissertação de Mestrado (Mestrado em produção
vegetal), São Paulo: Departamento de Agronomia, Universidade Estadual Paulista "Júlio de
Mesquita Filho" - UNESP, 1995, 133p.
CPRM. Projeto cadastro de fontes de abastecimento por água subterrânea no Estado de
Pernambuco, Diagnósticos dos Municípios, CPRM, 2005.
CRAINE, J. M.; ELMORE, A. J.; AIDAR, M. P. M.; BUSTAMANTE, M.; DAWSON, T. E.;
HOBBIE, E. A.; KAHMEN, A.; MACK, M. C.; McLAUCHLAN, K. K.; MICHELSEN, A.;
NARDOTO, G. B.; PARDO, L. H.; PEÑUELAS, J.; REICH, P. B.; SCHUUR, E. A. G.;
STOCK, W. D.; TEMPLER, P. H.; VIRGINIA, R. A.; WELKER J. M.; WRIGHT, I. J. Global
patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar
nutrient concentrations, and nitrogen availability. New Phytologist Journal, v. 183, p. 980–
992, 2009.
DADALTO, G. G.; COSTA, L. M.; MOURA FILHO, W. Alterações em características
químicas de solos cultivados com pastagens. Revista Ceres, v. 3, p. 395-403, 1986.
DALMOLIN, R. S. D.; GONÇALVES, C. N.; DICK, D. P.; KLAMT, E.; KNICKER, H.;
KÖGEL-KNABNER, I. Organic matter characteristics and distribution in Ferralsols profiles of
a climosequence in Southern Brazil. European Journal of Soil Science, v. 57, p. 644-654,
2006.
DANTAS, M. Pastagens da Amazônia Central: Ecologia e fauna do solo. Acta Amazônica, v.
9, p. 5-51, 1979.
DAWSON, T. E.; MAMBELLI, S.; PLAMBOECK, A. H.; TEMPLER, P. H.; TU, K. P. Stable
isotopes in plant ecology. Annual Review of Ecology and Systematics, v.33, p.507-559, 2002.
DE BONA, F. D. Dinâmica da matéria orgânica do solo em sistemas irrigados por aspersão sob
plantio direto e preparo convencional. Dissertação de Mestrado, Porto Alegre, Universidade
Federal do Rio Grande do Sul, 2005. 130p.
DICK, D. P.; NOVOTNY, E. H.; DIECKOW, J.; BAYER, C. IX - Química da matéria orgânica
do solo. In: MELO, V. F. e ALLEONI, L. R. F. Química e mineralogia do solo: Parte II –
Aplicações. Sociedade Brasileira de Ciência do Solo – SBCS, v. 2, 685 p., 2009.

100
DIXON, R. K.; BROWN, S.; HOUGHTON, R. A.; SOLOMON, A.M.; TREXLER, M. C.;
WISNIEWSKI, J. Carbon pools and flux of global forest ecosystems. Science, 263:185- 190,
1994.
DON, A.; SCHUMACHER, J.; FREIBAUER, A. Impact of tropical land-use change on soil
organic carbono stocks – a meta-analysis. Global Change Biology, v.17, n.4, p.1658-1670,
2011.
DORAN, J. W. e PARKIN, T. B. Defining and assessing soil quality. In: DORAN, J. W.;
COLEMAN, D. C.; BEZDICEK, D. F. e STEWART, B. A. eds. Defining soil quality for a
sustainable environment. Madison, Soil Science Society of America, n.35, p. 3-22, 1994.
DOU, F.; WRIGTH, A. L.; HONS, F. M. Depth distribution of soil organic C and N after long-
term soybean cropping in Texas. Soil and Tillage Research, Amsterdam, v. 94, n. 2, p. 530-
536, 2007.
EBELING, A. G.; ANJOS, L. H. C.; PEREIRA, M. G.; VALLADARES, G. S.; PÉREZ, D. V.
Substâncias húmicas e suas relações com o grau de subsidência em Organossolos de diferentes
ambientes de formação no Brasil. Revista Ciência Agronômica, v. 44, n. 2, 2013.
EHLERINGER, J. R., RUNDEL, P. W., Stable isotopes: history, units, and instrumentation. In:
RUNDEL, P. W., EGLERINGER, J. R., NAGY, K. A. (Eds.). Stable isotopes in ecological
research. Ecological Studies, 68. Springer-Verlag, p.1-15, 1989.
ELLERT, B. H. e BETTANY, J. R. Calculation of organic matter and nutrients stored in soils
under contrasting management regimes. Canadian Journal of Soil Science, v. 75, p. 529-538,
1995.
ELLERT, B. H.; JANZEN H. H.; McCONKEY, B. G. (2001). Measuring and comparing soil
carbon storage. In: LAL, R.; KIMBLE, J. M.; FOLLET, R. F.; STEWART, B. A. (eds)
Assessment methods for soil carbon. Lewis imprint of the CRC Press, Boca Raton, FL, p.
131–146.
EMBRAPA – EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema
Brasileiro de Classificação de Solos. Brasília, DF: Embrapa Solos, 3ª ed. revisada e ampliada,
353 p., 2013.
EMBRAPA – EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Zoneamento
Agrícola de Pernambuco. CD-Rom, 1998.
EMBRAPA – EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Serviço
Nacional de Levantamento e Conservação de Solo. 1997. Manual de Métodos de Análises de
Solos. Rio de Janeiro, 212 p., 1997.
EMBRAPA - EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Sistema
Brasileiro de Classificação de Solos. Brasília, DF, 353 p, 2013.
EMBRAPA - EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Manual de
métodos de análise de solo. Centro Nacional de Pesquisa de Solos. 2. edição revisada – Rio de
Janeiro, 1999.

101
ESWARAN, H.; VAN DEN BERG, E. e REICH, P. Organic carbon in soils of the word. Soil
Science Society of America Journal, 57:192- 194, 1993.
FANNING, D.S.; FANNING, M.C.B. Soil: morphology, genesis and classification. John
Wiley e Sons, 395 p., 1989.
FELLER, C.; BEARE, M. Physical control of soil organic matter dynamics in the tropics.
Geoderma, v. 79, p. 69-116, 1997.
FERNANDES, E. C. M.; MONTAVALLI, P. P.; CASTILLA, C.; MUKURUMBIRA, L.
Management control of soli organic matter dynamics in tropical land-use systems. Geoderma,
v. 79, p. 49-67, 1997.
FIDALGO, E. C. C.; BENITES, V. M.; MACHADO, P. L. O. A.; MADARI, B. E.; COELHO,
M. R.; MOURA, I. B.; LIMA, C. X. Estoque de carbono nos solos do Brasil. Rio de janeiro:
Embrapa Solos, 2007, 26 p. (Embrapa solos. Boletim de pesquisa e desenvolvimento, 121).
FRACETTO, F. J. C.; FRACETTO, G. G. M.; CERRI, C. C.; FEIGL, B. J.; SIQUEIRA NETO,
M. S. Estoques de carbono e nitrogênio no solo cultivado com mamona na caatinga. Revista
Brasileira de Ciência do Solo, v. 36, p. 1545-1552, 2012.
FRAZÃO, L. A.; SANTANA, I. K. S.; CAMPOS, D. V. B.; FEIGL, B. J.; CERRI, C. C.
Estoques de carbono e nitrogênio e fração leve da matéria orgânica em Neossolo Quartzarênico
sob uso agrícola. Pesquisa Agropecuária Brasileira, v. 45, p. 1198-1204, 2010.
FREITAS, A. D. S.; SAMPAIO, E. V. S. B.; FERNANDES, A. R.; SANTOS, C. E. R. S.
Biological nitrogen fixation in legume trees of the Brazilian caatinga. Journal of Arid
Environment, v. 74, p. 344–349, 2010.
FREITAS, A. D. S.; SAMPAIO, E. V. S. B.; RAMOS, A. P. S. R.; BARBOSA, M. R. V.;
LYRA, R. P.; ARAÚJO, E. L. Nitrogen isotopic patterns in tropical forest along a rainfall
gradient in Northeast Brazil. Plant and Soil, n. 391, p. 109-122, 2015.
FREITAS, P. L.; BLANCANEAUX, P.; GAVINELLI, E.; LARRÉ-LARROUY, M. C.;
FELLER, C. Nível e natureza do estoque orgânico de latossolos sob diferentes sistemas de uso
e manejo. Pesquisa Agropecuária Brasileira. v. 35, p. 157-170, 2000.
FREIXO, A. A.; MACHADO, P. L. O. A.; GUIMARÃES, C. M.; SILVA, C. A. FADIGAS, F.
S. Estoques de carbono e nitrogênio e distribuição de frações orgânicas de Latossolo do Cerrado
sob diferentes sistemas de cultivo. Revista Brasileira de Ciência do Solo, v. 26, p. 425-434,
2002.
FUJISAKA, S.; CASTILLA, C.; ESCOBAR, G.; RODRIGUES, V.; VENEKLASS, E. J.;
THOMAS, R.; FISHER, M. The effects of forest conversion on annual crops and pastures:
Estimates of carbon emissions and plant species loss in a Brazilian Amazon colony.
Agriculture, Ecosystems and Environment, v. 69, p. 17-26, 1998.

102
GALINDO, I. C. L.; RIBEIRO, M. R.; SANTOS, M. F. A. V.; LIMA, J. F. W. F.; FERREIRA,
R. F. A. L. Relações solo-vegetação em áreas sob processo de desertificação no município de
Jataúba-PE. Revista Brasileira de Ciência do Solo, v. 32, p. 1283-1296, 2008.
GANNES, L. Z.; DEL RIO, C. M.; KOCK, P. Natural abundance variations in stable isotopes
and their potential uses in animal physiological ecology. Comparative Biochemistry and
Physiology, v. 119a, n. 3, p. 725-737, 1998.
GARTEN, C. T. e VAN MIEGROET, H. Relationships between soil nitrogen dynamics and
natural 15N abundance in plant foliage from Great Smoky Mountain National Park. Canadian
Journal of Forest Research, v. 24, n.8, p. 1636-1645, 1994.
GARTEN, C. T. Variation in foliar 15N abundance and the availability of soil nitrogen on
Walker Branch watershed. Ecology, v. 74, p. 2098-2113, 1993.
GERALDES, A. P. A.; CERRI, C. C.; FEIGL, B. J. Biomassa microbiana de solo sob pastagens
na Amazônia. Revista Brasileira de Ciência do Solo, v. 19, p. 55-60, 1995.
GIONGO, V.; GALVÃO, S. R. S.; MENDES, A. M. S.; GAVA, C. A. T.; CUNHA, T. J. F.
Soil Organic Carbon in the Brazilian Semi-arid Tropics. Dynamic Soil, Dynamic Plant, v. 5,
p. 12-20, 2011.
GLATZELA, S.; KALBITZ, K.; DALVAC, M.; MOOREC, T. Dissolved organic matter
properties and their relationship to carbon dioxide efflux from restored peat bogs. Geoderma,
Amsterdam, v.113, p.397-411, 2003.
GRACE, J. Carbon cycle. Encyclopedia of Biodiversity, v. 1, p. 609-629, 2001.
GREGORICH, E. G.; CARTER, M. R; ANGERS, D. A; MONREAL C. M.; ELLERT, B.H.
Towards a minimum data set to assess soil organic matter quality in agricultural soils.
Canadian Journal of Soil Science, v. 74 p. 367-385, 1994.
GRIFFITHS, H.; BORLAND, A.; GILLON, J.; HARWOOD, K.; MAXWELL, K.; WILSON,
J. Stable isotopes reveal exchanges between soil, plants and the atmosphere. In: PRESS, M. C.;
SCHOLES, J. D.; BARKER, M. G. (Ed.). Physiological plant ecology. Blackwell Science, p.
415-441, 1999.
HALL, S. J.; MATSON, P. A.; ROTH, P. M. NOx emissions from soil: implications for air
quality modeling in agricultural regions. Annual Review Energy and the Environment. v.
21, p. 311-46, 1996.
HANDLEY, L. L. e RAVEN, J. A. The uses of natural abundance of nitrogen isotopes in plant
physiology and ecology. Plant Cell and Environment, v. 15, p. 965-985, 1992.
HANDLEY, L. L.; AUSTIN, A. T.; ROBINSON, D.; SCRIMGEOUR, C. M.; RAVEN, J. A.;
HEATON, T. H. E.; SCHMIDT, S.; STEWART, G. R. The 15N natural abundance (δ15N) of
ecosystem samples reflects measures of water availability. Australian Journal Plant
Physiology, v. 26, p. 185–199, 1999.

103
HANDLEY, L. L.; SCRIMGEOUR, C. M. Terrestrial plant ecology and 15N natural abundance:
the present limits to interpretation for uncultived systems with original data from a Scottish old
field. Advances in Ecological Research, v. 27, p. 133-212, 1997.
HÖGBERG, P. Transley review No 95 – N-15 natural abundance in soil-plant system. New
Phytologist, v. 137, n.2, p. 179-203, 1997.
HOLTZ, G. P., Dinâmica da decomposição da palhada e da distribuição do carbono, nitrogênio
e fósforo numa rotação de culturas sob plantio direto na região de Carambeí-PR. Dissertação
de Mestrado - Universidade Federal do Paraná, 129 p.,1995.
HOUGHTON, R. A.; SKOLE, D. L.; LEFKOWITZ, D. S. Changes in the landscape of Latin
America between 1850 and 1985: II net release of CO2 to the atmosphere. Forest Ecology
Management, Amsterdam, v. 38, n. 3/4, p. 173- 199, Feb. 1991.
HOWARD, P. J. A.; LOVELAND, P. J.; BRADLEY, R. I.; DRY, F. T. HOWARD, D. M.;
HOWARD, D. C. The carbon content of soil and its geographical distribution in Great Britain.
Soil Use and Management, v. 11, p. 9-15, 1995.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE. Disponível em:
<http://cidades.ibge.gov.br/xtras/uf.php?lang=&coduf=26&search=pernambuco>.
Acesso em setembro, 2016.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE. Censo
Demográfico de Pernambuco, 2000. Rio de Janeiro, 2001.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA – IBGE. Manual técnico de
pedologia. Rio de Janeiro, IBGE: Manuais técnicos em geociências, n. 4, 3ª ed., 430 p. 2015.
INTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE, 2014. Disponível em:
<http://www.ibge.gov.br/home/geociencias/cartogramas/estadual.html> (acesso em:
novembro, 2014).
IPCC - Intergovernmental Panel on Climate Change. Climate change 2007. Climate change
impacts, adaptation and vulnerability. Working Group II. Geneva, Switzerland: IPCC, 2007.
JACOMINE, P. K. T.; CAVALCANTI, A. C.; BURGOS, N.; PESSOA, S. C. P.; SILVEIRA,
C. O. Levantamento exploratório – reconhecimento de solos do Estado de Pernambuco.
Recife, SUDENE, 1973. v.1. (Boletim Técnico, 26. Série Pedologia, 14).
JANTALIA, C. P.; RESCK, D. V. S.; ALVES, B. J. R.; ZOTARELLI, L.; URQUIAGA, S.;
BODDEY, R. M. Tillage effect on C stocks of a clayey oxisol under a soybean-based crop
rotation in the Brazilian Cerrado. Soil and Tillage Research, v. 95, n. 1-2, p. 97-109. 2007.
JANZEN, H. H. The soil carbon dilemma: shall we hoard it or use it? Soil Biology and
Chemistry, Elmsford, v. 38, n. 3, p. 419-424, 2006.
JENNY, H. Factors of soil formation. A system of quantitative pedology. Dover, 281p.,
1994. (Reimpresso do original de 1941).

104
JOBBÁGY, E. G. and JACKSON, R. B. The vertical distribution of soil organic carbono and
its relation to climate and vegetation. Ecological Applications, v. 10, n. 2, p. 423-436, 2000.
KALBITZ, K.; SCHMERWITZ, J.; SCHWESIG, D.; MATZNER, E. Biodegradation of soil
derived dissolved organic matter as related to its properties. Geoderma, Amsterdam, v.113, p.
273-291, 2003.
KÄMPF, N.; SCHWERTMANN, U. Goethite and hematite in a climosequence in Southern
Brazil and their application in classification of kaolinitic soils. Geoderma, v.29, p.27-39, 1983.
KAUFFMAN, J. B.; HUGHES, R. F.; HEIDER, C. Carbon pool and biomass dynamics
associated with deforestation, land use, and agricultural abandonment in the neotropics.
Ecological Applications, v. 19, n. 5, p. 1211-1222, 2009.
KAUFFMAN, J. B.; SANFORD JR., R. L.; CUMMINGS, D. L.; SALCEDO, I. H.; SAMPAIO,
E. V. S. B. Biomass and nutrient dynamics associated with slash fires in neotropical dry forests.
Ecology, v. 74, n.1, p. 140-151, 1993.
KREILEMAN, G. J. J.; BOUWMAN, A. F. Computing land use emissions of greenhouse
gases. Water, Air and Soil Pollution, v. 76, p.231-250, 1994.
LAL, R. Forest soils and carbon sequestration. Forest Ecology and Management, v. 220, n.
1/3, p. 242-258, 2005.
LAL, R. Potential of desertification control to sequester carbon and mitigate the greenhouse
effect. Climatic Change, v. 51, p. 35-72, 2001.
LAL, R. Soil carbon dynamics in cropland and rangeland. Environmental Pollution, v.116, p.
353-362, 2002.
LAURENCE, W.F. et al. Relationship between soils and Amazon forest biomass: a landscape-
scale study. Forest Ecology and Management, v.118, n. 1-3, p.127-138, 1999.
LEITE, L. F. C.; MENDONÇA, E. S.; NEVES, J. C. L.; MACHADO, P. L. O. A.; GALVÃO,
J. C. C. Estoques totais de carbono orgânico e seus compartimentos em Argissolo sob floresta
e sob milho cultivado com adubação mineral e orgânica. Revista Brasileira de Ciência do
Solo, v. 27, n. 5, p. 821-832, 2003.
LIMA, D. A.; Estudos Fitogeográficos de Pernambuco. Anais da Academia Pernambucana
de Ciência Agronômica, v. 4, p. 243-274, 2007.
LIMA, M. A. Emissão de gases de efeito estufa provenientes de sistemas agrícolas no Brasil.
Biotecnologia, Ciência e Desenvolvimento, v. 3, p. 38-43, 2000.
LIU, N.; GALLINO, R.; BISTERZO, S.; DAVIS A. M.; SAVINA, M. R.; PELLIN, M. J. The 13C-pocket structure in AGB models: Constraints from zirconium isotope abundances in single
mainstream SiC grains. The Astrophysical Journal, v. 788, n. 2, p. 1-7, 2014.

105
LIU, X.; FENG-MIN, L.; DA-QIAN, L.; GUO-JUN, S. Soil Organic Carbon, Carbon Fractions
and Nutrients as Affected by Land Use in Semi-Arid Region of Loess Plateau of China.
Pedosphere, v. 20, n. 2, p. 146-152, 2010.
LOPES, C. A. e BENEDITO-CECILIO, E. Variabilidade isotópica (δ13C e δ15N) em produtores
primários de ambientes terrestres e de água doce. Acta Scientiarum, v. 24, n. 2, p. 303-312,
2002.
LORENZ, K.; LAL, R. The depth distribution of soil organic carbon in relation to land use and
management and the potential of carbon sequestration in subsoil horizons. Advanced in
Agronomy, San Diego, v. 88, n. 8, p. 35-66, 2005.
LOSS, A.; PEREIRA, M. G.; GIÁCOMO, S. G.; PERIN, A.; DOS ANJOS, L. H. C. Agregação,
carbono e nitrogênio em agregados do solo sob plantio direto com integração lavoura-pecuária.
Pesquisa Agropecuária Brasileira, v. 46, p. 1269-1276, 2011.
LOSS, A.; PEREIRA, M. G.; PERIN, A.; DOS ANJOS, L. H. C. Abundância natural de δ13C
e δ15N em sistemas de manejo conservacionista no cerrado. Bioscience Journal, v. 30, p. 604-
615, 2014.
LUCHESE, E. B.; FAVERO, L. O. B.; LENZI, E. Fundamentos da Química do Solo Teoria
e Prática. 2ª ed. Freitas Bastos, Rio de Janeiro, 2002.
MACHADO, P. L. O. A.; SILVA, C. A. Soil management under no-tillage systems in the
tropics with special reference to Brazil. Nutrient Cycling in Agroecosystems, v. 61, n. 1-2, p.
119-130, 2001.
MAIA, S. M. F.; OGLE, S. M.; CERRI, C. C.; CERRI, C. E. P. Changes in soil organic carbon
storage under different agricultural management systems in the Southwest Amazon Region of
Brazil. Soil and Tillage Research, v. 106, p. 177-184, 2010.
MAIA, S. M. F.; OGLE, S. M.; CERRI, C. E. P.; CERRI, C. C. Effect of grassland management
on soil carbono sequestration in Rondônia and Mato Grosso states, Brazil. Geoderma, v. 149,
p. 84-91, 2009.
MAIA, S. M. F.; XAVIER, F. A. S.; OLIVEIRA, T. S.; MENDONÇA, E. S.; ARAUJO FILHO,
J. A. Organic carbon pools in a Luvisol under agroforestry and conventional farming systems
in the semi-arid region of Ceará, Brazil. Agroforesty System, v. 71, p. 127-138, 2007.
MAIA, S.M.F.; XAVIER, F.A.S.; OLIVEIRA, T.S.; MENDONÇA, E.S. & ARAUJO FILHO,
J.A. Impactos de sistemas agroflorestais e convencional sobre a qualidade do solo no semi-
árido cearense. Revista Árvore, v. 30, p. 837-848, 2006.
MARCHÃO, R. L.; BECQUER, T.; BRUNET, D.; BALBINO, L. C.; VILELA, L.;
ROSSARD, M. Carbon and nitrogen stocks in a Brazilian clayey Oxisol: 13-year effects of
integrated crop-livestock management systems. Soil and Tillage Research, v. 103, p. 442-450,
2009.

106
MARCHIORI JÚNIOR, M. e MELO, W. J. Carbono da biomassa microbiana, e atividade
enzimática em um solo sob mata natural, pastagem e cultura do algodoeiro. Revista Brasileira
de Ciência do Solo, v. 23, p. 257-263, 1999.
MARTIN NETO, L.; ROSELL, R.; SPOSITO, G. Correlation of spectroscopy indicators of
humification with mean annual rainfall along a temperate grassland climosequence.
Geoderma, v. 81, p. 305-311, 1998.
MARTINELLI, L. A.; PICCOLO, M. C.; TOWNSEND, A. R.; VITOUSEK, P. M.; CUEVAS,
E.; McDOWELL, W. H.; ROBERTSON, G. P.; SANTOS, O. C.; TRESEDER, K. Nitrogen
stable isotopic composition of leaves and soil: Tropical versus temperate forests.
Biogeochemistsry, v. 46, p. 45-65, 1999.
MARTINS, C. M.; GALINDO, I. C. L;/ SOUZA, E. R.; POROCA, H. A. Atributos químicos
e microbianos do solo de áreas em processo de desertificação no semiárido de Pernambuco.
Revista Brasileira de Ciência do Solo, v. 34, p. 1883-1890, 2010.
MARTINS-NETO F. L.; MATSUMOTO S. N. Qualidade do solo e nutrição de plantas em
sistemas de produção de café (Coffea arabica L.). Coffee Science, v. 5, p. 206-213, 2010.
MATSON, P.A.; DOWELL, M. C.; TOWNSEND, A.R.; VITOUSEK, P. M. The globalization
of N deposition: ecosystem consequences in tropical environments. Biogeochemistry, v. 46, p.
67-83, 1999.
MELLO, F. A. F.; SOBRINHO, M. O. C. B.; ARZOLLA, S.; SILVEIRA, R. I.; NETTO, A.
C.; KIELH, J. C. Fertilidade do Solo. NOBEL, São Paulo, 1983.
MENDONÇA, L. A. R.; FRISCHKORN, H. SANTIAGO, M. F.; CAMARGO, P. B.; LIMA,
J. O. G.; MENDES FILHO, J. Identificação de mudanças florestais por 13C e 15N dos solos da
Chapada do Araripe, Ceará. Revista Brasileira de Engenharia Agrícola e Ambiental, v.14,
n.3, p.314-319, 2010.
MENEZES, R. C. S.; GARRIDO, M. S.; PEREZ M., A. M. Fertilidade dos solos no semi-
árido. In: CONGRESSO BRASILEIRO DE CIÊNCIA DO SOLO, 30. Recife, 2005.
Palestras... Recife, UFRPE/SBCS, 2005. CD-ROM.
MENEZES, R. S. C. e SAMPAIO, E. V. S. B. Simulação dos fluxos e balanços de fósforo em
uma unidade de produção agrícola familiar no semi-árido paraibano. In: SILVEIRA, L.M.;
PETERSEN, P. e SABOURIN, E., orgs. Agricultura familiar e agroecologia no semi-árido:
Avanços a partir do Agreste da Paraíba. Rio de Janeiro, AS-PTA, 2002. p.249-260.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo. UFLA, 626p.,
2003.
NARDOTO, G. B. Abundância natural de 15N na Amazônia e Cerrado – implicações para a
ciclagem de nitrogênio. (Tese de Doutorado) – Escola Superior de Agricultura Luiz de Queiroz
– ESALQ/USP, 100 p., 2005.

107
NARDOTO, G. B.; OMETTO, J. P. H. B.; EHLERINGER, J. R.; HIGUCHI, N.;
BUSTAMANTE, M. M. C.; MARTINELLI, L. M. Understanding the influences of spatial
patterns on N availability within the Brazilian Amazon Forest. Ecosystems, v. 11, p. 1234–
1246, 2008.
NASCIMENTO, C. W. A.; BARROS, D. A. S.; MELO, E. E. C.; OLIVEIRA, A. B. Alterações
químicas em solos e crescimento de milho e feijoeiro após aplicação de lodo de esgoto. Revista
Brasileira de Ciências do Solo, v. 28, p. 385-392, 2004.
NEILL, C.; FRY, B.; MELILLO, J.; STEUDLER, P.; MORAES, F. L. & CERRI, C. C. Forest-
and pasture-derived carbon contributions to carbon stocks and microbial respiration of tropical
pasture soils. Oecologia, v. 107, p. 113-119, 1996.
NOAA (National Oceanic and Atmospheric Administration). 2016. Trends in Atmospheric
Carbon Dioxide. Disponível em: <http://www.esrl.noaa.gov/gmd/ccgg/trends/weekly.html>
Accesso em Setembro, 2016.
NOORDWIJK, M. V.; CERRI, C. C.; WOOMER, P. L.; NUGROHO, K. e BERNOUX, M.
Soil carbon dynamics in the humid tropical forest zone. Geoderma, v. 79, p. 187-225, 1997.
O’LEARY, M. H. Carbon isotopes in photosynthesis. Bioscience, v. 38, p. 328-336, 1988.
ODUM, E. P. Ecologia. Editora Guanabara Koogan S. A. 434 p. 1983.
OLIVEIRA, L. B.; FONTES, M. P. F.; RIBEIRO, M. R.; KER, J. C. Morfologia e classificação
de Luvissolos e Planossolos desenvolvidos de rochas metamórficas do semiárido do Nordeste
brasileiro. Revista Brasileira de Ciência do Solo, Viçosa-MG, v. 33, p. 1333-1345, 2009.
OLIVEIRA, L. B.; RIBEIRO, M. R.; FERRAZ, F. B.; JACOMINE, P. K. T. Classificação de
solos planossólicos do Sertão do Araripe (PE). Revista Brasileira de Ciência do Solo, v. 27,
p. 685-693, 2003.
OLIVEIRA, T. E. G. Estudo da bolota Quercus rotundifolia Lam. como indicador precoce dos
stress hídrico ao longo de um gradiente climático no Alentejo (SE, Portugal). Dissertação de
Mestrado (Mestrado em Biologia da Conservação) - Lisboa: Departamento de Biologia
Animal, Universidade de Lisboa, 2014, 62 p.
PACHECO, M. R. P. S. e HELENE, M. E. M. Transcrições e notas: Atmosfera, fluxos de
carbono e fertilização por CO2. Estudos Avançados, v.4, n.9, p. 204–220, 1990.
PAIVA, A. O.; FARIA, G. E. Estoque de carbono do solo sob Cerrado Sensu Stricto no Distrito
Federal, Brasil. Revista Tropica: Ciências Agrárias e Biológicas, v. 1, p. 59-65, 2007.
PAIVA, A. O.; REZENDE, A. V.; PEREIRA, R. S. Estoque de carbono em cerrado sensu
stricto do Distrito Federal. Revista Árvore, v. 35, p. 527-538, 2011.
PARK, R e EPSTEIN, S. Metabolic fractionation of 13C and 12C in plants. Plant Physiology,
v. 36, p. 133-138, 1961.
PAUSTIAN, K.; SIX, J.; ELLIOTT, E.T.; HUNT, H. W. Management options for reducing
CO2 emissions from agricultural soils. Biogeochemistry, v. 48, p. 147-163, 2000.

108
PEDRA, W. N.; PEDROTTI, A.; SILVA, T. O.; MACEDO, F. L.; GONZAGA, M. I. S.
Estoques de carbono e nitrogênio sob diferentes condições de manejo de um Argissolo
Vermelho Amarelo, cultivado com milho doce nos tabuleiros costeiros de Sergipe. Semina:
Ciências Agrárias, v. 33, p. 2075-2090, 2012.
PEIXOTO, M. F. S.; Atributos físicos, químicos e biológicos como indicadores da qualidade
do solo. Bahia: UFRB, 2008. 22 p. (Documento 01).
PINHEIRO, E. F. M; LIMA, E; CEDDIA, M. B.; URQUIAGA, S.; ALVES, B. J. R.;
BODDEY, R. M. Impact of pre-harvest burning versus trash conservation on soil carbon and
nitrogen stocks on a sugarcane plantation in the Brazilian Atlantic forest region. Plant and Soil,
v. 333, p. 71-80, 2010.
POST, W. M.; EMANUEL, W. R.; ZINKE, P. J.;STANGENBERGER, A. Soil carbon pools
and world life zones. Nature, v. 298, p.156- 158, 1982.
PROBIO - Projeto de Conservação e Utilização Sustentável da Diversidade Biológica
Brasileira. Subprojeto – Levantamento da Cobertura Vegetal e do Uso do solo do Bioma
Caatinga. 2007.
http://mapas.mma.gov.br/geodados/brasil/vegetacao/vegetacao2002/caatinga/documentos/rela
torio_final.pdf
RANGEL, O. J. P.; SILVA, C. A. Estoques de carbono e nitrogênio e frações orgânicas de
Latossolo submetido a diferentes sistemas de uso e manejo. Revista Brasileira de Ciência do
Solo, v. 31, n. 6, p. 1609-1623, dez. 2007.
RESCK, D. V. S.; FERREIRA, E. A. B.; FIGUEIREDO, C. C.; ZINN, Y. L. Dinâmica da
Matéria Orgânica no Cerrado. In: SANTOS, G. A.; SILVA, L. S.; CANELLAS, L. P.;
CAMARGO, F. A. O. Fundamentos da matéria orgânica do solo: ecossistemas tropicais e
subtropicais. 2. Ed. rev. e atual. – Porto Alegre: Metropole, 2008. 654 p.
RIBEIRO, M. R.; JACOMINE, P. K. T.; LIMA, J. F. W. F. Caracterização e classificação de
solos de referência do Estado de Pernambuco. Recife, Universidade Federal Rural de
Pernambuco, 1999. 140p.
RICHTER, D. D.; MARLEWITZ, D.; TRUMBORE, S. E.; WELLS, C. G. Rapid accumulation
and turnover of soil carbon in an aggrading forest. Nature, v. 400, p. 56-58, 1999.
ROSCOE, R.; MACHADO, P. L. O. de A. Fracionamento físico do solo em estudos da
matéria orgânica. Dourados: Embrapa Agropecuária Oeste; Rio de Janeiro: Embrapa Solos,
2002. 86 p.
ROSCOE, R.; MERCANTE, F. M.; MENDES, I. C.; REIS-JÚNIOR, F. B.; SANTOS, J. C. F.;
HUNGRIA, M. Biomassa microbiana do solo: fração mais ativa da matéria orgânica. In:
Dinâmica da Matéria Orgânica do Solo em Sistemas Conservacionistas - Modelagem
Matemática e Métodos Auxiliares, Embrapa, Dourados-MS, p. 163-198, 2006.

109
RUSSOW, R.; SICH, I.; NEUE, H. U. The formation of the trace gases NO and N2O in soils
by the coupled processes of nitrification and denitrification: results of kinetic 15N tracer
investigations. Chemosphere – Global Change Science. n. 2, p. 359-366, 2000.
SÁ, J. C. M.; LAL, R. Stratification ratio of soil organic matter pools as na indicator of carbon
sequestration in a tillage chronosequence on a Brazilian Oxisol. Soil and Tillage Research, v.
103, n. 1, p. 46-56, 2009.
SALIMON, C. I. Respiração do solo sob florestas e pastagens na Amazônia Sul-Ocidental,
Acre. Tese de Doutorado, Piracicaba: Escola Superior de Agricultura Luiz de Queiroz, 2003.
SALTON, J. C.; MIELNICZUK, J.; BAYER, C.; FABRICIO, A. C.; MACEDO, M. C. M.;
BROCH, D. L. Teor e dinâmica do carbono no solo em sistemas de integração lavoura-pecuária.
Pesquisa Agropecuária Brasileira, v. 46, p. 1349-1356, 2011.
SAMPAIO, E. V. S. B. Caracterização do bioma caatinga: características e potencialidades, in:
GARIGLIO, M. A., KAGEYAMA, P. Y. (Eds.), Uso Sustentável e Conservação dos
Recursos Florestais da Caatinga. GARIGLIO, M. A. SAMPAIO, E. V. S. B.; CESTARO, L.
A.; KAGEYAMA, P. Y. (Orgs.), Brasília: Serviço florestal brasileiro, p. 29-48. 2010.
SAMPAIO, E. V. S. B.; COSTA, T. L. Estoques e Fluxos de Carbono no Semiárido Nordestino:
Estimativas Preliminares. Revista Brasileira de Geografia Física, v. 06, p. 1275-1291, 2011.
SANTOS, J. C. B.; SOUZA JÚNIOR, V. S.; CORRÊA, M. M.; RIBEIRO, M. R.; ALMEIDA,
M. C.; BORGES, L. E. P. Caracterização de Neossolos Regolíticos da região semiárida do
estado de Pernambuco. Revista Brasileira de Ciência do Solo, v. 36, p. 683-695, 2012.
SCHARPENSEEL, H.W. Preface to workshop ‘Management of carbon in tropical soils under
global change: Science, practice and policy’. Geoderma, v. 79, p. 1-8, 1997.
SCHIMEL, J. P. e BENNETT, J. Nitrogen mineralization: challenges of a changing paradigma.
Ecology, v. 85, n. 3, p. 591-602, 2004.
SCHLESINGER, W. H. Biogeochemistry: an analysis of global change. 2nd. ed. Academic
Press, 588 p., 1997.
SCHÖNING, I.; TOTSCHE, K. U.; KÖGEL-KNABNER, I. Small scale spatial variability of
organic carbon stocks in litter and solum of a forested Luvisol. Geoderma, v. 136, n. 3/4, p.
631-642, 2006.
SCOTT, A. J.; KNOTT, M. A Cluster analysis method for grouping means in the analysis of
variance. Biometrics, v.30, p.507-512, 1974.
SHAPIRO, S. S.; WILK, M. B. An analysis of variance teste for normality (complete samples).
Biometryka, v. 52, p. 591-611, 1965.
SILVA, C. A.; ANDERSON, S. J.; VALE, F. R. Carbono, nitrogênio e enxofre em frações
granulométricas de dois Latossolos submetidos a calagem e adubação fosfatada. Revista
Brasileira de Ciência do Solo, v. 23, p. 593-602, 1999.

110
SILVA, F. A. S.; ASSISTAT Versão 7.7 beta (2016) - Homepage http://www.assistat.com;
DEAG - CTRN-UFCG – Atualizado em 01/03/2016.
SILVA, J. E. RESCK, D.V.S.; CORAZZA, E. J.; VIVALDI L. Carbon storage in clayey Oxisol
cultivated pastures in the “Cerrado” region, Brazil. Agriculture, Ecosystems and
Environment, v. 103, p. 357-363, 2004.
SILVA, J. E.; LEMAINSKI, J.; RESCK, D. V. S. Perdas de matéria orgânica e suas relações
com a capacidade de troca catiônica em solos da região de cerrado do oeste baiano. Revista
Brasileira de Ciência do Solo, v. 18, p. 541-547, 1994.
SILVA, J. E.; RESCK, D. V. S.; CORAZZA, E. J.; VIVALDI, L. Carbon storage in clayey
Oxisol cultivated pastures in the “Cerrado” region, Brazil. Agriculture, Ecosystems &
Environment, v. 103, p. 357-363, 2004.
SIMÕES NETO, D. E.; OLIVEIRA, A. C.; FREIRE, F. J.; FREIRE, M. B. G. S.;
NASCIMENTO, C. W. A.; ROCHA, A. T. Extração de fósforo em solos cultivados com Cana-
de-açúcar e suas relações com a capacidade tampão. Revista Brasileira de Engenharia
Agrícola e Ambiental, v. 13, p. 840-848, 2009.
SIMÕES NETO, D. E.; OLIVEIRA, A. C.; ROCHA, A. T.; FREIRE, F. F.; FREIRE, M. B. G.;
NASCIMENTO, C. W. A. Características agroindustriais da Cana-de-açúcar em função da
adubação fosfatada, em solos de Pernambuco. Revista Brasileira de Engenharia Agrícola e
Ambiental, v. 16, p. 347-354, 2012.
SIQUEIRA NETO, M.; PICCOLO, M. C.; SCOPEL, E.; COSTA JUNIOR, C.; CERRI, C.C.;
BERNOUX, M. Carbono total e atributos químicos com diferentes usos do solo no Cerrado.
Acta Scientiarum Agronomy, v. 31, n. 4, p. 709-717, 2009.
SISTI, C. P. J.; SANTOS, H. P.; KOHHANN, R.; ALVES, B. J. R.; URQUIAGA, S.;
BODDEY, R. M. Change in carbon and nitrogen stocks in soil under 13 years of conventional
or zero tillage in southern Brazil. Soil Tillage Research, Amsterdam, v. 76, n. 1, p. 39-58.
2004.
SMITH, B.N.; EPSTEIN, S. Two categories of 13C/12C ratios for higher plants. Plant
Physiology, v. 47, p. 380-384, 1971.
SOLLINS, P.; HOMANN, P.; CALDWELL, B. Stabilization and destabilization of soil organic
matter: mechanisms and controls. Geoderma, v. 74, p. 65-105, 1996.
SOUSA, F. P.; FERREIRA, T. O.; MENDONÇA, E.S.; ROMERO, R.E. & OLIVEIRA, J.G.B.
Carbon and nitrogen in degraded Brazilian semi-arid soils undergoing desertification. Agric.
Ecosystem Environment, v. 148, p. 11-21, 2012.
SOUZA, R. V. C. C.; RIBEIRO, M. R.; SOUZA JUNIOR, V. S.; CORRÊA, M. M.;
ALMEIDA, M. C.; CAMPOS, M. C. C.; RIBEIRO FILHO, M. R.; SCHULZE, M. B. B.
Caracterização de solos em uma topoclimossequência no maçico de Triunfo – Sertão de
Pernambuco. Revista Brasileira de Ciência do Solo, v. 34, p. 1259-1270, 2010.

111
SPRENT, J. I. The ecology of the nitrogen cycle. Cambridge studies in ecology, Cambridge
University Press, 1987.
STEVENSON, F. J. Humus chemistry: genesis, composition, reactions. New York: John
Wiley & Sons, 1994. 496 p.
SWAP, R.J.; ARANIBAR, J.N.; DOWTY, P.R.; GILHOOLY III, W.P.; MACKO, S.A. Natural
abundance of 13C and 15N in C3 and C4 vegetation of southern Africa: patterns and implications.
Global Change Biology, v. 10, p. 350-358, 2004.
TARRÉ, R.; MACEDO, R; CANTARUTTI, R. B.; REZENDE, C. P.; PEREIRA, J. M.;
FERREIRA, E.; ALVES, B. J. R.; URQUIAGA, S. & BODDEY, R. M. The effect of the
presence of a forage legume on nitrogen and carbon levels in soils under Brachiaria pastures
in the Atlantic forest region of the South of Bahia, Brazil. Plant and Soil, v. 234, p. 15-26,
2001.
TATE, K. R. Assessment, based on a climosequence of soils in tussock grasslands, of soil
carbon storage and release in response to global warming. Journal of Soil Science, v. 43, p.
697-707, 1992.
TEIXEIRA, L. B.; BASTOS, J. B. Matéria orgânica nos ecossistemas de floresta primária
e pastagem na Amazônia central. Belém: EMBRAPACPATU, 1989. 26 p. (Boletim de
pesquisa, 99).
TELLES, E. D. C.; CAMARGO, P. B.; MARTINELLI, L. A.; TRUMBORE, S. E.; COSTA,
E. S.; SANTOS, J.; HIGUCHI, N.; OLIVEIRA JÚNIOR, R. C. Influence of soil texture on
carbon dynamics and storage potential in tropical forest soils of Amazonia. Global
Biogeochemical Cycles, v. 17, n. 2, p. 1-12, 2003.
TIESSEN, H.; FELLER, C.; SAMPAIO, E. V. S. B.; GARIN, P. Carbon sequestration and
turnover in semiarid savannas and dry forest. Climatic Change, v. 40, p. 105-117, 1998.
TOMASELLA, J.; HODNETT, M. G. Estimating soil water retention characteristics from
limited data in Brazilian Amazonia. Soil Science, v. 163, p. 190-202, 1998.
TRAORÉ, S.; OUATTARA, K.; ILSTEDT, U.; SCHMIDT, M.; THIOMBIANO, A.;
MALMER, A.; NYBERG, G. Effect of land degradation on carbon and nitrogen pools in two
soil types of a semi-arid landscape in West Africa. Geoderma, v. 241-242, 2015.
TREVISAN, R.; MATTOS, M. L. T.; HERTER, F. G. Atividade microbiana em Argissolo
Vermelho-Amarelo distrófico coberto com aveia preta (Avena sp.) no outono, em um pomar de
pessegeiro. Ciência Rural, v. 7, n.2, p. 83-89, 2002.
UNKOVICH, M.; PATE, J.; MCNEILL, A.; GIBBS, D. J. Stable isotope techniques in the
study of biological processes and functioning of ecosystems. Dordrecht: Kluwer Academic
Publisher, 291 p., 2001.
VITOUSEK, P. M. Global environmental change: an introduction. Annual Review of Ecology
and Systematics, v. 23, p. 1-14, 1992.

112
VITOUSEK, P. M.; MOONEY, H. A.; LUBCHENCO, J.; MELILLO, J. Human domination
of Earth’s Ecosystems. Science. n. 277, p. 494-499, 1997.
VOGEL, J. C. Variability of carbon isotope fractionation during photosynthesis. In: Ehleringer,
J. R.; Hall, A. E.; Farquhar, G. D. (ed.). Stable isotopes and plant carbon – Water relations.
Academic Press, p. 29-38, 1993.
WATSON, R. T.; NOBLE, I. R.; BOLIN, B.; RAVINDRANATH, N. H.; VERARDO, D. J.;
DOKKEN, D. J.; Land use, land-use change and forestry: A special report of the IPCC,
Cambridge University Press, 2000.
WATSON, R. T.; RODHE, H.; OESCHGER, H.; SIEGENTHALER, U. I. Greenhouse gases
and aerosols. In: HOUGHTON, J. T.; JENKINS, G. J.; EPHRAUMS, J. J. (Eds.), Climate
Change. Cambridge University Press, p. 1-40, 1990.
YEO, I. K., JOHNSON, R. A. A New Family of Power Transformation to Improve Normality
or Symmetry. Biometrika, v. 87, 954-959, 2000.
ZECH, W.; SENESI, N.; GUGGENBERGER, G.; KAISER, K.; LEHMANN, J.; MIANO, T.
M.; MILTNER, A.; SCHROTH, G. Factors controlling humification and mineralization of soil
organic matter in the tropics. Geoderma, v. 79, n. 1/4, p. 117-161, 1997.
ZHOU, G.; LIU, S.; LI, Z.; ZHANG, D.; TANG, X.; ZHOU, C.; YAN, J.; MO, J. Old-growth
forest can accumulate carbon in soils. Science, v. 314, p. 1417-1418, 2006.
ZINN, Y. L.; RESK, D.; SILVA, J. E. Soil organic as affected by afforestation with Eucalyptus
and Pinus in the Cerrado region of Brazil. Forest Ecology e Management, v. 166, n. 1/3, p.
285-294, 2002.
Recommended











![Bioquimica UFP [Exercicios] Prof Dr Pedro Silva](https://img.pdfslide.tips/doc/110x75/5572001f49795991699ed4ff/bioquimica-ufp-exercicios-prof-dr-pedro-silva.jpg)




