
Exemples de contrôle de voies métaboliques
Contrôle: une seule enzyme ou plusieurs?
0 5 100
0.5
1
[SUBSTRAT] (mM)
VIT
ES
SE
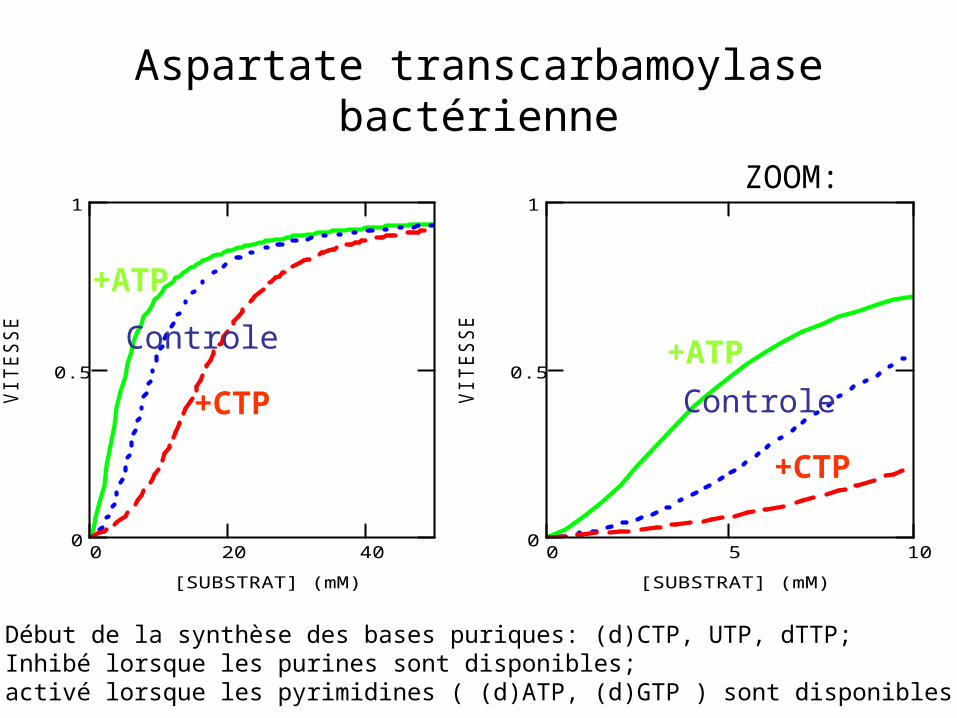
Aspartate transcarbamoylase bactérienne
Début de la synthèse des bases puriques: (d)CTP, UTP, dTTP; Inhibé lorsque les purines sont disponibles;activé lorsque les pyrimidines ( (d)ATP, (d)GTP ) sont disponibles
0 20 400
0.5
1
[SUBSTRAT] (mM)
VIT
ES
SE
ZOOM:
+ATP
+CTP
Controle +ATP
+CTP
Controle

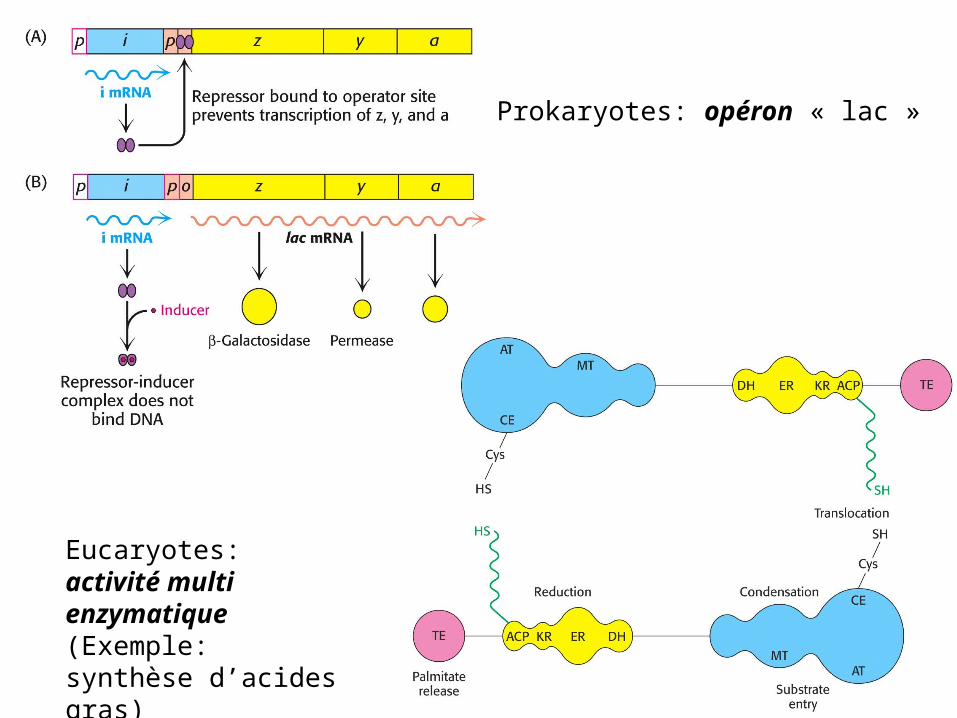
Prokaryotes: opéron « lac »
Eucaryotes: activité multi enzymatique (Exemple: synthèse d’acides gras)

Régulation coordonnée de la synthèse et de la dégradation du glycogène par l’AMPc:
GlycogèneGlucose P
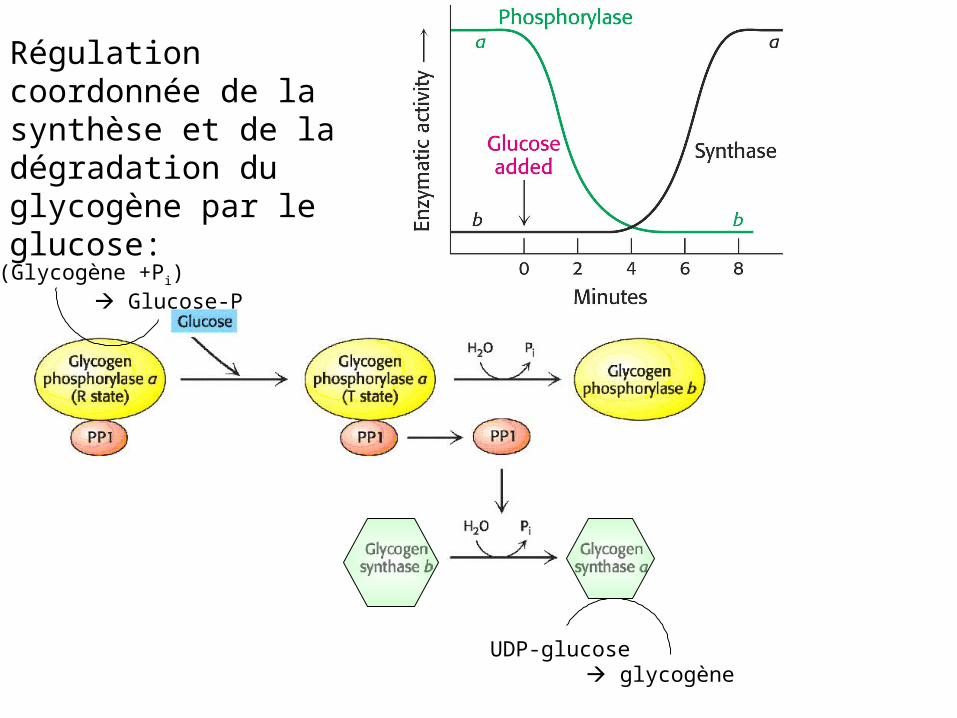
(Glycogène +Pi) Glucose-P
UDP-glucose glycogène
Régulation coordonnée de la synthèse et de la dégradation du glycogène par le glucose:
Glucose
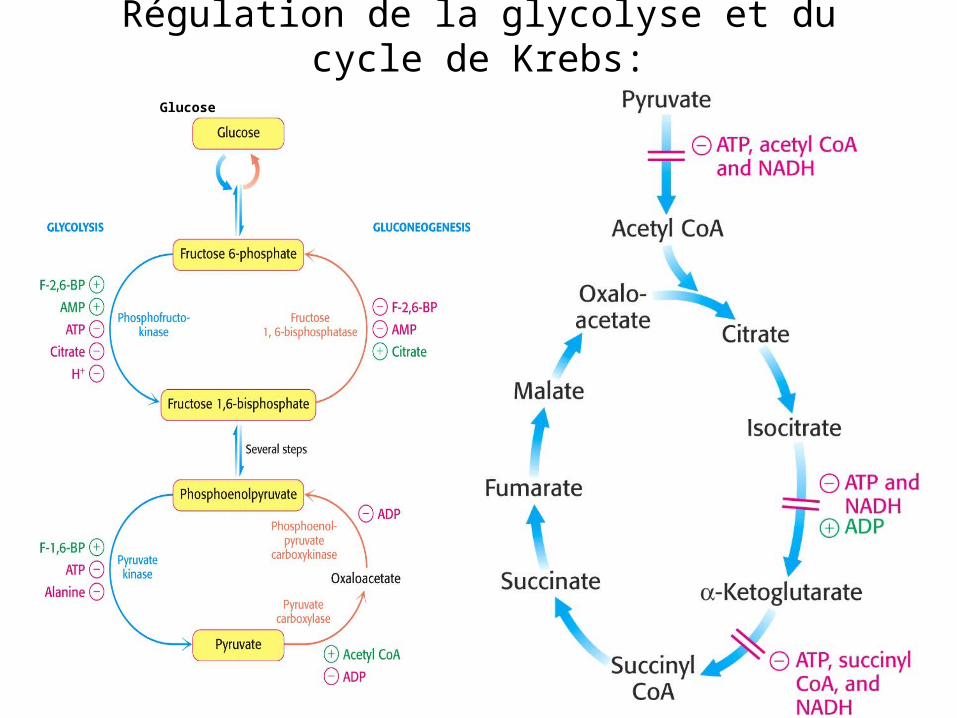
Régulation de la glycolyse et du cycle de Krebs:
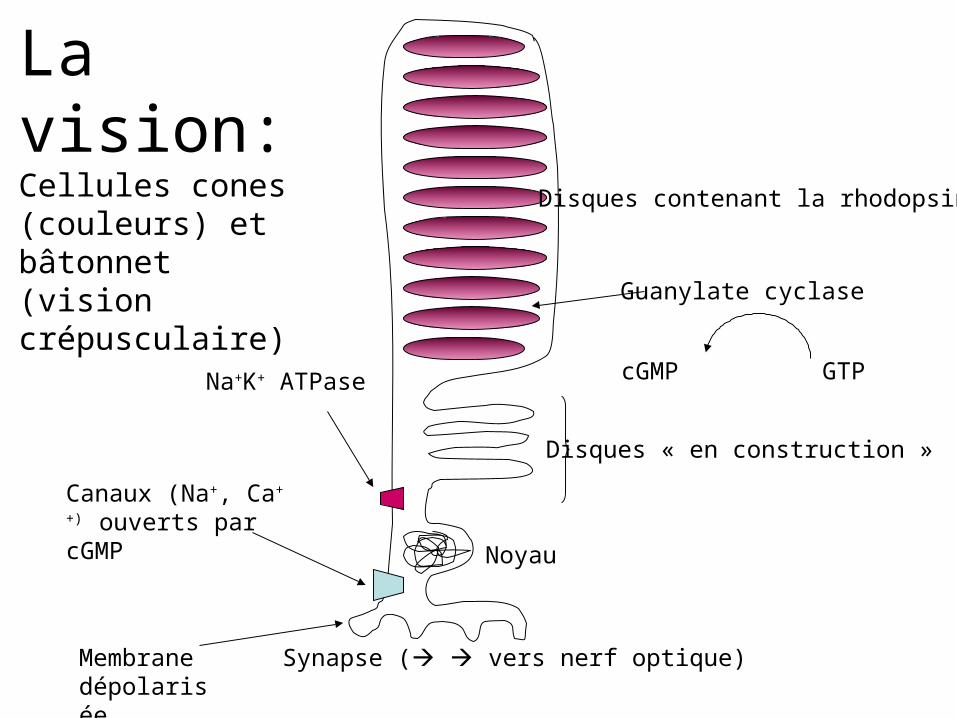
Disques contenant la rhodopsine
Disques « en construction »
Noyau
Synapse ( vers nerf optique)
Canaux (Na+, Ca++)
ouverts par cGMP
Guanylate cyclase
cGMP GTP
Membrane dépolarisée
Na+K+ ATPase
La vision:Cellules cones (couleurs) et bâtonnet (vision crépusculaire)

La rhodopsine, un récepteur de la lumière, couplé aux protéines G…

A l’obscurité:
• La rhodopsine est inactive. • La transducine est donc inactive.• La guanylate cyclase synthétise du cGMP.• Le cGMP ouvre des canaux Ca++ et maintient le taux de
Ca++ élevé, donc la cellule dépolarisée.• Le Ca++ active la GCAP (Guanylate Cyclase Activating
Protein), qui active la Guanylate cyclase. Boucle de rétroaction positive!
• Le Ca++ active la recoverine, qui inactive partiellement la rhodopsine kinase et augmente sa spécificité pour la métarhodopsine II (active)
• Le Ca++ active l’adénylate cyclase. L’AMPc active la PKA, qui phosphoryle et inactive la phosducine.

Contrôles: récupération à faible luminosité
• La Métarhodopsine II est instable: pert son rétinal, qui est rapidement réduit en « tout trans rétinol ». Il doit être capté par une des cellules de la base de la rétine pour être converti en 11-cis rétinol, puis en 11-cis rétinal, puis retransporté vers la cellule en batonnet (ou cone) pour régénérer la rhodopsine.
• La métarhodopsine II active une kinase spécialisée, la « rhodopsine kinase », qui la phosphoryle. La métarhodopsine II phosphorylée active puis reconnaît l’arrestine (un inhibiteur compétitif) plutôt que la transducine…
• La transducine TαGTP est une GTPase: elle hydrolyse le GTP en
GDP, puis interagit avec avec Tβγ plutôt qu’avec la phosphodiestérase… Cette interaction permet à la fois d’inactiver la transducine, et de l’amener à proximité de la rhodopsine pour un éventuel nouveau cycle d’activation-désactivation.
• Des protéines spécialisées (« RGS ») augmentent l’activité GTPase de la transducine et facilitent son inactivation

Après un « flash » lumineux:• Le taux de Ca++ diminue suite à la fermeture des canaux. Suite à
cette diminution:– L’adénylate cyclase est inactive. La phosducine est alors
déphosphorylée par des phosphatases. Elle reconnaît la sous-unité Tβγ et la dissocie de la membrane. Le complexe TαGDP ne peut plus se
réassocier à la membrane, et ne peut donc plus être activé par la rhodopsine.
– La recoverine est inhibée, et ne reconnaît plus la rhodopsine kinase. Cette dernière devient peu sélective, et phosphoryle aussi bien la rhodopsine (non encore activée) que la métarhodopsine II (active).
– La CGAP est activée, et l’activité guanylate cyclase augmente (transitoirement)
– En cas d’éblouissement prolongé: le Na+ chute également. La CGAP est alors inactivée, et la guanylate cyclase est inhibée par la sous-unité Tβγ. Ceci permet d’éviter un cycle futile synthèse-dégradation du cGMP.

Quelques exemples de contrôle :
• « Opérons » bactériens• Synthèse d’acides gras• Synthèse et utilisation du glycogène• Glycolyse et néoglucogénèse• Cycle Krebs• Vision• Etc…
La règle: contrôle « partagé », réparti à plusieurs niveaux. Pourquoi?
Recommended

















