INDICE
1. IL MEDITERRANEO ............................................................................... 1
1.1 INTRODUZIONE........................................................................................................1 1.1.1 Caratteristiche idrologiche ............................................................................................................. 1 1.1.2 Caratteristiche biogeochimiche ..................................................................................................... 3 1.1.3 Il Mediterraneo orientale................................................................................................................ 4 1.1.4 Reti trofiche e flussi di carbonio ................................................................................................... 5 1.1.5 Il Mar Mediterraneo e la fosforo limitazione ............................................................................. 9 1.1.6 Il Microzooplancton....................................................................................................................... 10 1.1.7 Ruolo trofico del microzooplancton ........................................................................................... 11
1.2. SCOPO DEL LAVORO............................................................................................15
1.3. MATERIALI E METODI.........................................................................................16 1.3.1 Area di studio.................................................................................................................................. 16 1.3.2 Variabili ambientali....................................................................................................................... 17 1.3.3 Il metodo delle diluizioni............................................................................................................. 18 1.3.4 Campionamento ............................................................................................................................. 22 1.3.5 Distribuzione del microzooplancton.......................................................................................... 24 1.3.6 Analisi quali-quantitativa ............................................................................................................ 25
a) Picoplancton....................................................................................................................................... 25 b) Nanoplancton ..................................................................................................................................... 27 c) Microplancton .................................................................................................................................... 28
1.3.7 Produzione secondaria .................................................................................................................. 30
1.4 RISULTATI.................................................................................................................31 1.4.1 Dati ambientali............................................................................................................................... 31 1.4.2 Predazione sul picoplancton eterotrofo ..................................................................................... 31 1.4.3 Predazione sul picoplancton autotrofo ...................................................................................... 37 1.4.4 Predazione sul nanoplancton....................................................................................................... 41 1.4.5 Microzooplancton .......................................................................................................................... 48 1.4.6 Produzione secondaria .................................................................................................................. 57 1.4.7 Distribuzione .................................................................................................................................. 59
1.5. DISCUSSIONE E CONCLUSIONI.......................................................................63

2.OSTREOPSIS OVATA ............................................................................. 72
2.1 INTRODUZIONE......................................................................................................72
2.2 SCOPO DEL LAVORO.............................................................................................76
2.3 MATERIALI E METODI..........................................................................................77 2.3.1 CAMPIONAMENTO .................................................................................................................... 77
a) Esperimento di diluizione................................................................................................................ 77 b) Esperimento di grazing (giugno 2009)........................................................................................... 78 c) Esperimento di grazing (settembre 2009)...................................................................................... 79 d) Copepodi........................................................................................................................................... 79
2.3.2 ANALISI QUALI QUANTITATIVA.......................................................................................... 80
2.4 RISULTATI.................................................................................................................82 2.4.1 Caratterizzazione del popolamento microzooplanctonico (giugno 2009)............................ 82 2.4.2 Inizio dell’incubazione (T0).......................................................................................................... 83 2.4.3 Fine dell’incubazione (T24) – Crescita apparente...................................................................... 84 2.4.4 Esperimento di predazione del mesozooplancton (Grazing) – giugno 2009....................... 91 2.4.5 Caratterizzazione del popolamento microzooplanctonico (settembre 2009)....................... 92 2.4.6 Esperimento di predazione del mesozoo plancton (grazing) – settembre 2009 .................. 94
2.5 DISCUSSIONE E CONCLUSIONI........................................................................96

3. ANTARTIDE.......................................................................................... 100
3.1. Introduzione ............................................................................................................100 3.1.1 Ruolo del microzooplancton in Antartide............................................................................... 101 3.1.2 Caratterizzazione dell’area di studio........................................................................................ 102 3.1.3 Il Mare di Ross.............................................................................................................................. 105 3.1.4 Baia Terra Nova ............................................................................................................................ 107
3.2 MATERIALI E METODI........................................................................................108
3.3 RISULTATI...............................................................................................................109 Esperimento 1 ........................................................................................................................................ 109 Esperimento 2 ........................................................................................................................................ 110 Esperimento 3 ........................................................................................................................................ 111 Esperimento 4 ........................................................................................................................................ 114 Esperimento 5 ........................................................................................................................................ 115 Esperimento 6 ........................................................................................................................................ 117 Esperimento 7 ........................................................................................................................................ 117 Esperimento 8 ........................................................................................................................................ 119
3.4 DISCUSSIONE E CONCLUSIONI......................................................................121 Bibliografia
Allegati

Il Mediterraneo-Introduzione
1
1. IL MEDITERRANEO
1.1 INTRODUZIONE
Il Mar Mediterraneo è un mare chiuso che si estende tra 45°N e 30°N circa di
latitudine e tra 5°W e 36°E circa in longitudine, con una superficie di 2.560.0000 km2 e
una profondità massima di 5.020 m. Compreso fra le coste dell’Europa meridionale,
dell’Asia Occidentale e dell’Africa Settentrionale, il Mar Mediterraneo si sviluppa per una
lunghezza di 3.680 km, tra lo Stretto di Gibilterra e la costa orientale, e con una larghezza
media di 700 km, la cui massima estensione di 1.665 km è compresa tra il golfo della Sirte
e quello di Trieste.
Il Mediterraneo è in comunicazione con l’Oceano Atlantico tramite lo Stretto di
Gibilterra e con il Mar Nero tramite il Bosforo.
Il Mediterraneo è suddiviso in diversi sottobacini. Lo zoccolo siculo africano,
ampio sollevamento del fondo tra la Sicilia e il Capo di Bon in Algeria, divide questo mare
in un bacino orientale ed uno occidentale, che a loro volta sono divisi in mari e bacini
minori. Nel bacino occidentale, escludendo il sollevamento delle Baleari e qualche piccola
piattaforma, si denota che la scarpata continentale, che raggiunge i 2.400 m di profondità, è
molto ravvicinata alla costa e la piana abissale non presenta quasi rilievi.
1.1.1 Caratteristiche idrologiche
Il Mediterraneo è un bacino di evaporazione, in quanto le perdite di acqua dolce per
evaporazione sono maggiori degli apporti dovuti alle precipitazioni e ai fiumi. Il bilancio di
massa e di sale è però mantenuto in stato stazionario attraverso gli scambi con l’oceano
Atlantico che avvengono attraverso lo Stretto di Gibilterra. Il deficit di acqua dolce è
bilanciato dall’ingresso nel Mediterraneo di acqua superficiale atlantica relativamente poco
salata e dalla fuoriuscita di acqua a salinità elevata. La differenza di densità (Béthoux et al.,
1999) confina con il flusso entrante alla superficie (0-150 m), mentre il flusso in uscita
scorre sul fondo della soglia (150-300m) dello stretto di Gibilterra. Questa differenza di
densità è dovuta principalmente alla salinità (35.9 per le acque atlantiche e 37.9 per il
flusso uscente (Béthoux et al., 1999) ed è tanto significativa da mantenere riconoscibile

Il Mediterraneo-Introduzione
2
l’acqua atlantica presente nel Mediterraneo come Atlantic Water (AW). L’AW scorre poi
principalmente verso est fino alla parte orientale del Mediterraneo, confinata dalla forza di
Coriolis alla parte orientale del bacino. Nonostante l’influsso delle masse d’acqua che
attraversa e i parziali rimescolamenti che ne attenuano le particolarità in modo crescente
con il movimento verso est, l’AW resta identificabile in ogni parte del bacino.
Nel Mediterraneo sono inoltre presenti processi di formazione di acque profonde e
intermedie, rappresentati dallo sprofondamento di masse d’acqua superficiale. La
formazione delle acque è il risultato dell’aumento di densità provocato dall’aumento di
salinità dovuto alla forte evaporazione estiva e dal forte raffreddamento invernale che si
verifica in specifiche aree.
In particolare, si ha sprofondamento di acque dense nel Golfo del Leone e
nell’Adriatico Meridionale, mentre nel bacino levantino si ha formazione di acque
intermedie. Nel Golfo del Leone il raffreddamento invernale è causato dal maestrale, vento
freddo e secco incanalato dai Pirenei e dal Massiccio Centrale Francese in direzione nord-
ovest. Nell’Adriatico è solamente il raffreddamento invernale a guidare il flusso di densità
(Béthoux et al., 1999) poiché l’evaporazione e le precipitazioni si equivalgono. E’ quindi la
bora, vento freddo proveniente da nord-est, a raffreddare le masse d’acqua che poi
muovono verso sud lungo le coste italiane.
La Levantine Intermediate Water (LIW) è una massa d’acqua che si forma nel Bacino
levantino dalla trasformazione dell’AW. A causa dell’evaporazione la LIW acquista
elevata salinità e si estende per tutto il Mediterraneo orientale ad una profondità che varia
tra 100-150 m nella porzione più orientale del bacino e 250-300 m nella parte più
occidentale. La LIW poi oltrepassa lo Zoccolo siculo-africano e attraversa il Mediterraneo
occidentale ad una profondità di circa 400 m, costituendo una parte importante del flusso
uscente da Gibilterra. La LIW subisce un rimescolamento con le acque sottostanti e
soprastanti, entrambe a salinità inferiore, che attenua il caratteristico massimo di salinità a
profondità intermedia mano a mano che si procede da est verso ovest. AW e LIW, insieme
alla formazione di acque profonde, compongono il forzante termoalino del Mediterraneo.
Questa forza è l’origine della circolazione nel Mediterraneo, circolazione che riproduce in
scala ridotta quella degli oceani. La presenza del LIW infatti gioca un ruolo importante
nella formazione delle acque profonde, connettendo le celle di circolazione meridionali
delle acque profonde con le celle zonali formate da AW e LIW (fig.1.1)

Il Mediterraneo-Introduzione
3
Fig.1.1 Circolazione e formazione delle acque nel Mar Mediterraneo (Pinardi &
Masetti, 2000)
1.1.2 Caratteristiche biogeochimiche
La circolazione antiestuarina del Mediterraneo causa la fuoriuscita di acque
intermedie e profonde, arricchite in nutrienti, e l’ingresso di acque superficiali, povere di
nutrienti.
Il caso fosforo può servire a schematizzare il comportamento degli elementi con
lunghi tempi di residenza in ambiente marino. Per il fosforo si può assumere lo stato di
equilibrio (Béthoux, 1981) dato che la concentrazione di questo elemento non mostra
variazioni significative nel tempo. Béthoux (1991) stima la quantità di fosforo uscente da
Gibilterra (433 *106 kg P y -1) bilanciata dal fosforo trasportato dal flusso in entrata (86
*106 kg P y -1) e soprattutto dall’apporto terrestre (357*106 kg P y -1). L’utilizzo del fosforo
da parte dei processi biologici avviene principalmente negli strati superficiali, mentre negli
strati intermedi e profondi questo elemento può essere considerato conservativo al pari
della salinità.
Il fosforo inorganico disciolto, quindi nella forma utilizzata come nutriente per i
processi biologici, è altresì il maggiore fattore limitante per la produttività del
Mediterraneo. Il rapporto tra nitrati e fosfati (N : P) per l’Oceano Atlantico è di circa 16:1,

Il Mediterraneo-Introduzione
4
e corrisponde al tipico utilizzo di nutrienti da parte del fitoplancton (Redfield et al., 1963),
mentre nel Mediterraneo il valore caratteristico è 22:1. Questo ultimo valore, pur essendo
una schematizzazione del bacino nel suo complesso, indica da solo l’impoverimento dei
fosfati. Bisogna però sottolineare che nel tipico profilo verticale questo elemento presenta
un impoverimento superficiale, ovvero nella zona fotica, proprio dove è necessario per la
vita.
Anche l’azoto presenta un profilo simile, ma il nitroclino si posiziona ad una
profondità inferiore a quello del fosfoclino, portando il rapporto N : P nelle acque
superficiali a valori ben superiori a 22:1. In aggiunta a ciò recenti esperimenti (Diaz et al.,
2001) hanno evidenziato che l’arricchimento di fosfati in campioni d’acqua superficiale
aumenta la produttività e la richiesta fitoplanctonica di azoto, suggerendo quindi per il
fosforo il ruolo di fattore limitante.
1.1.3 Il Mediterraneo orientale
La circolazione nel bacino orientale può essere descritta come un sistema a tre strati
(Ribera d’Alcalà et al., 2003). L’acqua Atlantica entra attraverso lo Stretto di Sicilia (a
200 m). Quest’acqua fluisce verso il bacino orientale diventando più salata a causa delle
condizioni climatiche più calde e secche della regione orientale, in particolar modo durante
il periodo estivo. Le acque levantine intermedie (200-500 m di profondità) si generano
durante l’inverno vicino alla costa della Turchia e formano la maggior parte del flusso di
ritorno dell’acqua in uscita dal bacino attraverso lo stretto di Sicilia. Le acque profonde del
bacino orientale si formano nell’Adriatico meridionale e riempiono il bacino dagli 800 m
di profondità al fondo. Dal 1987 una nuova fonte di produzione di acque profonde si è
osservata nel bacino levantino e nel mar Ionio profondi (Roether et al., 2007). Quest’acqua
profonda formata nell’Egeo Meridionale viene scaricate nel bacino levantino attraverso lo
stretto di Kassos e questa sostituzione tra le sorgenti di acque dense di fondo è nota come il
transiente del Mediterraneo orientale.

Il Mediterraneo-Introduzione
5
1.1.4 Reti trofiche e flussi di carbonio
Il Mar Mediterraneo, per la sua configurazione di mare semi chiuso è caratterizzato
da un ricco complesso di dinamiche fisiche con tratti distintivi riguardanti in maniera
particolare la circolazione termoalina. La produzione primaria presenta un andamento
decrescente da ovest verso est. Il bacino del Mediterraneo è dominato da piccoli autotrofi,
microeterotrofi e piccoli copepodi. I microrganismi - fitoplancton, virus, batteri, flagellati,
ciliati e zooplancton in generale - rivelano una considerevole diversità e variabilità sia su
scala temporale che spaziale. Le maggiori differenze fra il bacino occidentale e quello
orientale sono state evidenziate studiando il fitoplancton ma anche la componente
microbica eterotrofa e le sue relazioni. In queste aree l’arricchimento intermittente di
nutrienti favorisce un’alternanza tra la comunità dominata da diatomee e la comunità
microbica. In questi casi la rete trofica classica sostituisce la rete microbica (fig.1.2)
Queste alternanze aumentano il flusso verso i livelli trofici superiori. Il sistema microbico
sembra subire contemporaneamente un controllo di tipo bottom-up e top-down. La rete
mistivora è dovuta alla grande varietà di modalità di assunzione del cibo e di preferenze di
prede da parte del mesozooplancton sul fitoplancton e sui ciliati.
Fig.1.2 Rete trofica classica
Il flusso di carbonio negli oceani dipende dall’efficienza della produzione primaria
e dai processi biogeochimici della zona fotica nonché dalla struttura delle reti trofiche. Fin

Il Mediterraneo-Introduzione
6
all’inizio degli anni ’80 si era creduto che il flusso di energia attraverso la rete trofica fosse
regolato da relazioni fra la produzione primaria microfitoplanctonica e i suoi predatori
rappresentati prevalentemente dal mesozooplancton (copepodi). Si credeva che la biomassa
fotosintetizzata fluisse attraverso la rete trofica classica ai livelli trofici superiori. In questo
schema gli organismi microfitoplanctonici erano i soli responsabili della produzione
primaria e non era stato del tutto considerato che la perdita di materia organica potesse
rappresentare il substrato per la degradazione batterica (Azam, 1998). In questo caso i
batteri non avevano un ruolo essenziale. Da quel momento si sono molto approfonditi gli
studi sui processi biogeochimici che avvengono negli oceani e questo ha portato ad una
nuova idea di rete trofica marina grazie alla scoperta dell’esistenza di un enorme numero di
microrganismi dalle dimensioni comprese fra 0.02 e 0.2 µm che includono virus e batteri
sia autotrofi che eterotrofi (Pomeroy, 1974; Azam et al., 1983; Rassoulzadegan, 1993;
Legendre & Rassoulzadegan, 1995; Fonda Umani, 2000).
La nuova tecnica di microscopia ad epifluorescenza proposta da Daley e Hobbies
nel 1975 ha notevolmente aumentato l’interesse per lo studio dei batteri marini. Questo
metodo di analisi permette di distinguere tra organismi autotrofi, sulla base della
fluorescenza naturale dei pigmenti fotosintetici (Waterbury et al., 1979) ed organismi
eterotrofi riconoscibili grazie ad una nuova tecnica di colorazione. La capacità di contare le
cellule microbiche e distinguerle in componente autotrofa ed eterotrofa ha permesso di fare
una stima generale delle abbondanze che va da 107-109 ind L-1 per la componente
eterotrofa e 105-108 ind L-1 per la componente autotrofa partendo da ambienti marini
eutrofici fino ad arrivare ad ambienti oligotrofici.
Si specula che una componente così abbondante possa avere un ruolo determinante
per il flusso di energia nell’ecosistema marino (Pomeroy, 1974). I procarioti sono infatti gli
unici organismi in grado di utilizzare la materia organica disciolta (DOM) e trasformarla in
biomassa utilizzabile dai predatori (Azam et al., 1983). I batteri non solo utilizzano il
DOM ma sono anche in grado di degradare la materia organica particellata (POM). Viene
così introdotto il concetto di circuito microbico o “microbical loop” (Azam et al., 1983)
che comprende le interazioni trofiche tra il pico-nano e microplancton partendo da
considerazioni quantitative: la più alta percentuale di carbonio organico in mare si trova in
fase disciolta.
L’origine del DOC può essere molto diversa: essudazione microalgale (William,
1990; Alledredge et al., 1993), perdita di materia cellulare durante i processi di predazione
(sloppy feeling) (Eppely et al., 1981), lisi cellulare spontanea o per infezione virale

Il Mediterraneo-Introduzione
7
(Fuhrman & Shuttle, 1993; Fuhrman & Noble, 1995), degradazione di pallottole fecali
“faecal pellets” prodotte dallo zooplancton (Honjio & Roman, 1978), e da processi di
escrezione.
Su questo substrato agiscono i batteri utilizzandolo e trasformandolo in propria
biomassa. Tale biomassa viene predata dal nanoplancton eterotrofo, il quale a sua volta è
predato dal microzooplancton. Tutti questi organismi restituiscono all’ambiente carbonio
inorganico sotto forma di CO2 attraverso la respirazione e materiale organico disciolto
(DOC) e quindi il “loop” si chiude. Il nanoplancton può predare sui batteri autotrofi ed
eterotrofi ma può predare anche piccole cellule eucarioti mentre il microzooplancton può
utilizzare come fonte di energia sia la frazione autotrofa che quella eterotrofa del
picoplancton e del nano plancton (fig.1.3).
Fig. 1.3 Rappresentazione delle tre reti trofiche: microbial loop, rete trofica
microbica, rete del pascolo.
Si può avere la percezione che una rete trofica (classica) escluda l’altra ( microbica)
e questa a sua volta escluda il microbial loop. In realtà a parte in situazioni ambientali
estreme, le tre reti sono presenti contemporaneamente in quella che viene definita
“mistivourus food chain“. Il prevalere di una sull’altra va a modificare il destino finale del
materiale fotosintetizzato. Nel microbial loop l’energia ricavata dal substrato viene

Il Mediterraneo-Introduzione
8
immediatamente respirata o trasformata in prodotti di escrezione quali faecal pellets
piccole e galleggianti utilizzate in superficie con ulteriore rilascio di CO2. Se prevale la
catena classica invece, i consumatori di ordine superiore immobilizzano nella loro
biomassa una parte del carbonio organico per tempi maggiori producendo faecal pellets più
pesanti che possono sedimentare al di sotto della zona fotica, fino ad arrivare sul fondo.
Fig.1.4 Ciclo del carbonio oceanico. La pompa biologica (sinistra) controllata
dalla rete trofica marina e la pompa di solubilità (destra) regolata da processi fisici e
chimici (Chisholm, 2000)
Il ruolo che i batteri giocano nell’ambiente marini è fondamentale sotto molteplici
aspetti: basti pensare alla loro importanza nel ciclo globale del carbonio e quindi allo
scambio di CO2 tra oceano e atmosfera nel contesto generale dell’incremento di tale gas e
dei suoi possibili effetti sul riscaldamento del pianeta. Nell’oceano sono state identificate
tre diversi tipi di “pompe” di CO2 (Volk and Hoffert, 1985): la pompa fisica legata alla
solubilità della CO2 all’interfaccia atmosfera-oceano, attiva nelle zone di formazione delle
acque dense; la pompa biologica dei carbonati, legata alla sedimentazione di organismi con
gusci calcarei e la pompa biologica dei tessuti molli (soft-tissue-pump) nota come la
pompa biologica della CO2 (fig.1.4).
Il carbonio biogenico negli oceani viene classificato in base al suo tempo di turn
over definito come il tempo che intercorre tra la fissazione del carbonio per via
fotosintetica e il suo rilascio in atmosfera (Legendre and LeFevre, 1992). Si distinguono 3
compartimenti di carbonio biogenico in base ai tempi di turn over: carbonio a vita breve

Il Mediterraneo-Introduzione
9
(short lived carbon >10-2 anni), costituito da organismi ad alto tasso di turn over e dal
carbonio organico disciolto labile. Il carbonio organico a vita lunga (long-lived carbon 10-
2-102 anni) e il carbonio biogenico sequestrato (>102 anni), che comprende i resti organici
seppelliti nei sedimenti, la sostanza organica refrattaria e la CO2 disciolta nelle acque
profonde derivata dai processi di ossidazione (respirazione) in situ dei composti organici.
La produzione primaria può, come abbiamo visto, essere respirata nella zona
eufotica, veicolata verso gli organismi di maggiori dimensioni o verso gli strati profondi
dell’oceano. La catena classica del pascolo è caratteristica di zone ad alta energia, sia in
termini idrodinamici che in termini di concentrazione di nutrienti come in aree costiere o di
upwelling dove si verificano fioriture di diatomee (Kiorbe, 1996). La catena microbica è
invece tipica di zone a bassa energia, con scarso apporto di nutrienti dove il carbonio viene
fotosintetizzato dai produttori primari ed essenzialmente utilizzato e respirato in zona
fotica (Kiorbe, 1996).
1.1.5 Il Mar Mediterraneo e la fosforo limitazione
Il Mediterraneo è da sempre considerato uno dei mari più oligotrofici del mondo e
dagli studi più recenti sembra essere quasi sempre in condizioni di fosforo limitazione. Per
questo ambiente Thingstad e Rassoulzadegan (1995) hanno messo a punto un semplice
modello basandosi su numerose osservazioni precedenti relative all’esistenza di una
fosforo limitazione sia per il fitoplancton che per i batteri; elevate concentrazioni
superficiali di organico disciolto (DOP e DOC) con gradienti decrescenti verso il fondo;
scarsa degradazione del DOC da parte dei batteri a causa della fosforo limitazione e della
predazione insieme; regolazione del ciclo di rigenerazione del fosforo da parte degli
organismi di taglia minore e particolarmente dei batteri.
Tre sono le potenziali classi di utilizzatori di fosforo inorganico: i batteri, i piccoli
autotrofi (flagellati) e le diatomee. A seconda della maggiore o minore disponibilità di
DOC (labile) da un lato e di silice organica dall’altro, l’incorporazione del fosforo
inorganico sarà più efficiente da parte di uno dei comparti di fissatori. Ad alte
concentrazioni di fosforo prevarrà la rete trofica classica che è più efficiente nel rispondere
ad improvvisi e brevi apporti di nutrienti tipici di zone costiere o di upwelling, aumentando
rapidamente il flusso di materiale sedimentario in colonna con accumuli al nutriclinio
(Kiorbe, 1996).

Il Mediterraneo-Introduzione
10
Scarsi apporti di fosforo inorganico e sufficiente disponibilità di DOC labile
favoriranno la catena microbica in zone più stabili ed oligotrofiche. La predazione giocherà
il suo ruolo in egual misura: abbondanti biomassa di eteronanoflagellati controlleranno
efficacemente le biomassa di batteri e favoriranno l’assunzione di fosforo da parte di
nanoflagellati autotrofi; consistenti biomassa di ciliati controlleranno efficacemente sia il
nanoplancton autotrofo che eterotrofo diminuendo la pressione di grazing di quest’ultimo
sui batteri, i quali saranno più efficienti nell’assunzione di fosforo: la presenza massiccia di
predatori di ordine superiore (copepodi) infine abbasserà via predazione il numero di
diatomee ma anche di ciliati spostando nuovamente l’equilibrio a favore del nanoplancton
((Fonda Umani, 2000).
1.1.6 Il Microzooplancton
Il termine zooplancton fu coniato da Hensen (1887) e comprende tutti quegli
organismi galleggianti nell’acqua le cui abilità locomotorie sono insufficienti a contrastare
l’andamento delle correnti.
Lo zooplancton è distinto dal fitoplancton in base alle modalità di nutrizione
eterotrofa o autotrofa. Lo zooplancton è definito come l’insieme degli organismi fagotrofi
in accordo con le loro preferenze alimentari che possono essere classificati in erbivori,
detritivori, onnivori o carnivori.
Il microzooplancton è composto da organismi di taglia compresa fra 10 – 20 (a
seconda della classificazione utilizzata) a 200 µm: molti protisti come ciliati, dinoflagellati,
foraminiferi, acantari ed i primi stadi larvali di molti metazoi chiamati solitamente
micrometazoi. Il plancton eterotrofo include anche i batteri osmotrofi. Comunemente tra i
flagellati si trovano anche forme mixotrofe, una combinazione di autotrofi ed eterotrofi, e
si possono trovare anche in alcuni phyla di metazoi (cnidaria e molluschi). Il plancton
marino comprende un’ampia varietà di organismi. Per distinguere le varie componenti del
plancton, Sieburth et al. (1978) hanno proposto un ordinamento basato sulla classe
dimensionale, al cui interno si identificano organismi con modalità trofiche diverse
(autotrofi, eterotrofi, mixotrofi). Il plancton viene così dimensionalmente distinto (tab.1.1);

Il Mediterraneo-Introduzione
11
Plancton Dimensione Organismi FEMTOPLANCTON 0.02-0.2 µm Virus,Batteri
PICOPLANCTON 0.2-2 µm Batteri, Cianobatteri, Proclorofite
NANOPLANCTON 2-10 µm Fitoflagellati, Coanoflagellati,Dinoflagellati, Ciliati,
MICROPLANCTON 10-200 µm Diatomee, Tintinnidi, Dinoflagellati, Radiolari, Ciliati, larve e uova di Metazoi
MESOPLANCTON 0.2-20 mm Crostacei (Copepodi, Eufasiacei, Cladoceri)
MACROPLANCTON 2-20 cm Meduse MEGAPLANCTON 20-200 cm Meduse, colonie di Tunicati
Tab.1.1 Suddivisione dimensionale del plancton
1.1.7 Ruolo trofico del microzooplancton
Negli ultimi decenni sono stati condotti studi sul microzooplancton relativi non solo
agli aspetti morfologici e sistematici, ma anche a quelli biochimici ed ecologici, allo scopo
di valutarne la rilevanza nella rete trofica marina. Più esattamente si è approfondito il ruolo
dei popolamenti microzooplanctonici nei trasferimenti energetici dai produttori primari ai
successivi anelli della rete trofica (Margalef, 1963; Montagnes et al., 1988; Epstein et al.,
1992, Stoeker & Capuzzo, 1990).
In particolare, l’interesse per questi organismi è aumentato dagli inizi degli anni
’80, da quando il concetto di rete trofica marina è cambiato passando dalla classica catena
alimentare della predazione (Azam 1998) ad una più complessa rete alimentare mistivora,
dove il micobial loop o rete microbica gioca un ruolo importante (Azam et al., 1983). La
rete trofica del pascolo prevedeva come produttori primari soltanto gli organismi
microfitoplanctonici, (diatomee, dinoflagellati autotrofi, coccolitoforidi, ecc). Il maggior
trasferitore di biomassa in queste reti era il mesozooplancton, gruppo di organismi tra cui
si trovano piccoli crostacei planctonici come i copepodi, che peraltro sono il gruppo di
metazoi più numeroso di tutto il pianeta. L’introduzione del concetto di “microbial loop”
ha modificato in parte il ruolo del microzooplancton che, in questo contesto, è il predatore
principale del nano e del picoplancton (Gast, 1985; Jonsson, 1986; Sherr et al., 1986;
Albright et al., 1987; Fenchel, 1982; Sherr & Sherr, 1987; Rassoulzadegan et al., 1988;
Bernard and Rassoulzadegan, 1990; Gonzales, 1996; Simek & Chrzanowski, 1992), e nella

Il Mediterraneo-Introduzione
12
catena del pascolo come predatore del fitoplancton di cui è capace di consumare anche il
100% della produzione giornaliera (Goldman,1984; Calbet & Landry, 2004). Inoltre,
costituisce il principale veicolo di trasferimento energetico dalla rete microbica a quella del
pascolo, in quanto a sua volta viene attivamente predato dal mesozooplancton (copepodi, e
larve di pesci) (Fonda Umani & Zanon 2000; Fonda Umani et al., 2005).
Nonostante l’ormai riconosciuta importanza di questa frazione planctonica non si è
ancora messo a punto un protocollo unico di campionamento, conseguentemente il
confronto tra dati di origine diversa deve essere affrontato con grande cautela considerando
le modalità di campionamento adottate.
L’abbondanza della comunità microplanctonica è regolata dalla presenza di risorse
ambientali come l’abbondanza delle prede (Heinbokel & Beers, 1979; Fenchel 1980b;
Verity 1985; Jonsson 1986; Rassoulzadegan et al., 1988; Hansen 1991; Hansen et al.,
1994) e dalla pressione di grazing derivante dai livelli trofici superiori esercitata dai
crostacei (Stoeker & Egloff 1987; Stoeker & Capuzzo, 1990; Gifford, 1991; Kivi et al.,
1996).
I ciliati loricati o tintinnidi sono uno dei gruppi più abbondanti che può
rappresentare anche il 50% dell’abbondanza microzooplanctonica (Dolan et al., 2009;
Fonda Umani et al., 2005) e sono peraltro gli organismi ideali per lo studio della struttura o
della composizione della comunità microzooplanctonica (Thompson et al., 1999). I ciliati
loricati sono caratterizzati dall’avere uno specifico rivestimento detto appunto lorica
(Dolan et al., 2005) sul quale è basata la classificazione. Sono facilmente identificabili al
microscopio grazie alla presenza della lorica che ha forme ben definite (Pierce & Turner
1993; Thompson & Adler, 2005; Dolan et al., 2005, 2007).
I ciliati planctonici sono considerati predatori di particellato sospeso catturato con il
movimento delle ciglia (Fenchel, 1980). I ciliati non predano in maniera indiscriminata, ma
selettivamente discriminando il tipo di preda in base una varietà di meccanismi tra i quali
anche l’uso di chemiosensori (Fenchel, 1980). I tintinnidi sono inoltre in grado di ingerire
prede di dimensioni superiori del 40-45% rispetto il diametro orale (Spittler, 1973).
I dinoflagellati rappresentano un gruppo piuttosto eterogeneo con più di 2000 specie
descritte. Una gran parte di queste specie sono mixotrofe, e sebbene dotati di cloroplasti
possono anche comportarsi da eterotrofi (Stoeker, 1999). I dinoflagellati presentano due
flagelli che permettono loro di muoversi attivamente, tanto che la loro velocità nel nuoto è
maggior di quella delle loro prede. Le forme eterotrofe presentano tre modalità di
predazione: ingestione diretta della preda, emissione di uno pseudopodio che cattura la

Il Mediterraneo-Introduzione
13
preda ed emissione di un imbuto che aspira il contenuto della preda (Hansen & Calado,
1999).
I mixotrofi sono prevalentemente organismi unicellulari (dinoflagellati,
primnesioficee, ciliati) che in primis hanno la capacità di fotosintetizzare ma al tempo
stesso possono nutrirsi in modo eterotrofo. Questa caratteristica viene usata per rinnovare
le riserve di carbonio cellulare, macronutrienti e amminoacidi (Stoecker & Gustafson
2003). Sono presumibilmente i fattori e le condizioni ambientali quali luce, nutrienti e
disponibilità di prede che modulano le varie modalità di predazione. Mentre per alcuni taxa
sembra che i mixotrofi siano associati ad ambienti oligotrofici, i protisti mixotrofi
sembrano essere abbondanti in ambienti eutrofici (Stoeker, 1998).
A partire dal concetto di rete microbica introdotto da Azam (Azam et al. 1983,
Pomeroy, 1974), il microzooplancton ha assunto un ruolo fondamentale nel trasferimento
dell’energia dai livelli trofici inferiori (rete microbica) alla rete del pascolo.
Il microzooplancton cresce alla stessa velocità delle cellule fitoplanctoniche e può
facilmente adattarsi alle variazioni delle disponibilità di cibo. Fa parte della dieta dei
grandi predatori come i copepodi (Kleppel, 1993; Roman & Gauzens, 1997; Calbet &
Landry, 1999 Roman et al., 2000; Rollwagen Bollens & Perny, 2003; Calbet & Saiz, 2005;
Irigoien et al., 2005; Liu et al., 2005), può allo stesso tempo essere preda e competere con i
consumatori di livello superiore nella rete mistivora (Rassoluzadegan, 1993). Quando
disponibile, il microzooplancton e specialmente i ciliati sono selettivamente predati dal
mesozooplancton (Wiadnyana & Rassoulzadegan, 1989; Stoecker & Capuzzo, 1990;
Verità & Paffenhofer, 1996; Roman et al., 2000; Rollwagen Bollens & Penry, 2003; Calbet
& Saiz, 2005; Liu et al., 2005).
Nonostante l’importanza di questa frazione non c’e’ ancora una modalità unica di
campionamento (retino o bottiglia), e non c’e’ accordo tra i ricercatori sul volume più
adeguato di campione, sul fissativo più idoneo (e le relative concentrazioni) (Choi &
Stoecker, 1989; Sherr & Sherr, 1993; Leakey et al., 1994; Stoecker et al., 1994 Gifford &
Caron, 2000; Zinabu & Bott, 2000; Karayanni et al., 2004; Modigh & Castaldo, 2005).
Conseguentemente ogni confronto quantitativo con altri dati deve essere considerato con
cautela in considerazione al protocollo utilizzato.
Nel Mar Mediterraneo le prime ricerche sul microzooplancton furono strettamente
tassonomiche (Entz, 1904, 1909, Laackmann, 1913, Jorgensen, 1924, Rampi, 1948, 1950);
solo a partire dagli anni ’80 le ricerche si sono indirizzate sulla distribuzione e

Il Mediterraneo-Introduzione
14
sull’importanza ecologica, ma bisogna arrivare ai primi anni ’90 perché i dinoflagellati
vengano presi in considerazione.

Il Mediterraneo-Introduzione
15
1.2. SCOPO DEL LAVORO
Lo scopo della tesi è stato quello di quantificare il flusso ci carbonio attraverso la
comunità microbica valutato attraverso la stima della predazione esercitata dal
microzooplancton sulla comunità microbica. Determinare sia le prede che i predatori dal
punto di vista arrivando al genere e specie (quando possibile).Lo scopo è stato anche
quello di verificare l’eventuale predazione selettiva esercitata dalla comunità eterotrofa,
l’analisi dei “black box” studiando gli effetti sinergici ed antagonisti della predazione
esercitata dal microzooplancton e dai nanoflagellati sul picoplancton eterotrofo.
Quantificare il tasso specifico di crescita dei predatori rappresentati dal
microzooplancton dopo un periodo di incubazione (produzione secondaria).
Comparare la composizione e la distribuzione spaziale del microzooplancton
raccolto e conservato con tre diverse tecniche di campionamento.
Questo lavoro di tesi si inserisce all’interno del progetto V.E.C.T.O.R.
(Vulnerability of the coasts and of the Italian marine ecosystems to climate change and
their role in the Mediterranean carbon cycles) che ha come scopo quello di approfondire le
conoscenze relative all’impatto esercitato dai cambiamenti climatici globali sull’ambiente
marino Mediterraneo, focalizzando l’attenzione sui processi fisici e biogeochimici delle
masse d’acqua. Si prefigge anche l’obiettivo di studiare il ruolo attivo esercitato dal bacino
del Mediterraneo nel ciclo globale del carbonio cercando di capire se il bacino del
Mediterraneo si comporta da produzione (source) o deposito (sink) di CO2.

Il Mediterraneo- Materiali e Metodi
16
1.3. MATERIALI E METODI
1.3.1 Area di studio
Fig.1.5 Stazioni di campionamento (V6, V7, V10, Viera)
I campioni di acqua di mare analizzati in questa tesi di dottorato sono stati raccolti
durante la “Transmediterranean cruise” all’interno del progetto V.E.C.T.O.R.
(VulnErabilità delle Coste e degli ecosisTemi marini italiani ai cambiamenti climatici e
loro ruOlo nei cicli del caRbonio mediterraneo) svoltasi dal 13 al 27 giugno 2007 nel
Mediterraneo orientale. La crociera oceanografica si è svolta a bordo della Nave
oceanografica N/O Universitas.
L’acqua di mare è stata campionata in 5 stazioni del Mediterraneo orientale
(fig.1.5). Condizioni meteorologiche avverse si sono presentate a sud di Creta tali da
bloccare ogni attività per più di 48 ore. Condizioni di mare non affrontabili hanno inoltre
impedito di raggiungere la stazione V9 che è stata sostituita con una stazione posta a 40
miglia a sud di Creta in corrispondenza del vortice di Ierapetra: la stazione è stata chiamata
Viera.
I campioni di acqua di mare superficiale sono stati raccolti in 4 stazioni (V6, V7,
V10, Viera) . Nella tabella 1.2 sono riportate le date e le coordinate di campionamento.

Il Mediterraneo- Materiali e Metodi
17
Tab. 1.2 Date, longitudine e latitudine, delle stazioni campionate
1.3.2 Variabili ambientali
Parametri idrologici : temperatura (°C) e salinità sono stati raccolti con una sonda
multiparametrica provvista di sensore per la temperatura, conducibilità ossigeno,
fluorescenza, transmittanza, pH, equipaggiata con 24 bottiglie Niskin da 12 litri ciascuna.
Parametri biogeochimici: ossigeno disciolto (AOU Wink.) nutrienti inorganici
(NO3, NO2 SiO2, PO4) così come il fosforo e l’azoto organico disciolti (TDP e TDN), usati
per la caratterizzazione della colonna d’acqua. I campioni per le analisi sono stati raccolti a
quote discrete mediante bottiglie Niskin montate sulla rosette.
L’ossigeno disciolto è stato analizzato a bordo con il metodo Winkler, mentre i
nutrienti inorganici disciolti (nitriti, nitrati, fosfati e silicati) sono stati campionati e
mantenuti a -20°C fino all’analisi condotta presso il laboratorio CNR-ISMAR di Trieste.
Produzione primaria: è stata misurata in situ lungo la colonna d’acqua utilizzando
radioisotopi in modo da stimare il tasso di produzione di carbonio. La clorofilla a è stata
campionata con un’analoga procedura utilizzata per la produzione primaria: sono stati
raccolti 5L di acqua allo scopo di calcolare i rapporti Produzione/Biomassa.
Data Stazione Longitudine Latitudine
14/06/2007 V6 17°59.980' E 38°29.720' N 10/06/2007 V7 20°52.500' E 35°08.120 N 24/06/2007 Viera 26°05.200' E 34°24.690 N 23/06/2007 V10 28°19.390 E 35°57.190 N

Il Mediterraneo- Materiali e Metodi
18
Stazioni V6 V7 V10 Viera 14/06/2007 16/06/2007 23/06/2007 24/06/2007
Temperatura °C 22.66 22.49 24.33 23.40 Salinità 38 38.73 39.36 39.27
Tot. Chl a(µg L-1) 0.059 0.046 0.057 0.035
PP (µgCL-1 h-1) 0.44 0.27 0.51 0.33
AOU Wink. µM-O2 -12.14 -4.81 -6.37 -12.53
NO3 µM-N 0.78 0.07 0.18 0.09
NO2 µM-N 0.13 0.03 0.02 0.02
SiO2 µM-Si 0.77 1.01 0.85 1.15
PO4 µM-P 0.12 0.01 0.02 0.01 Longitudine 17°59.980' E 20°52.500' E 26°05.200' E 28°19.390 E Latitudine 38°29.720' N 35°08.120 N 34°24.690 N 35° 57.190 N
Profondità (m) 5 5 5 5
Tab. 1.3 Variabili ambientali registrate durante il campionamento
1.3.3 Il metodo delle diluizioni
La maggior difficoltà riscontrata nel misurare il tasso di predazione del
microzooplancton in situ, risiede nel fatto che separare i predatori (microzooplancton) dalle
loro prede (microfitoplancton, nanoplancton e picoplancton) è impossibile, a causa delle
loro equiparabili dimensioni. Per valutare l’efficienza della predazione si utilizza ormai
come protocollo standard il metodo delle diluizioni proposto da Landry e Hassett (1982),
successivamente modificato da Landry et al. (1995) e Gallegos (1989) e ulteriormente
adattato per stimare anche la predazione del nanoplancton eterotrofo sul picoplancton. Il
metodo classico delle diluizioni si limita a stimare l’impatto del microzooplancton sulla
frazione autotrofa, valutando le differenze tra i valori di clorofilla. Altri ricercatori hanno
utilizzato analisi con l’HPLC per valutare la predazione in base a pigmenti specifici (Strom
et al., 1991; McManus et al., 1992; Verity et al., 1993; Waterhouse et al., 1995; Latasa et
al., 1997; Schlüter, 1998) così come è stata utilizzata la citofluorimetria (Reckermann et
al., 1997; Kuipers et al., 1999; Stelfox-Widdicombe et al., 2000; Aberle et al., 2007).
Per valutare l’efficienza della predazione a livello specifico si devono analizzare i
campioni al microscopio ottico rovesciato me se si vogliono avere informazioni
relativamente alla predazione sulla frazione eterotrofa (nano e picoplanctonica) i campioni
devono essere analizzati quali-quantitativamente al microscopio ad epifluorescenza

Il Mediterraneo- Materiali e Metodi
19
(Landry et al., 1984; Campbell et al., 1986; Caron et al., 1991; Landry et al., 1993; Verity
et al., 1993; 1996; Ayukai, 1996; Nejstgaard et al., 1997; James and Hall; 1998; Lessard &
Murrel, 1996; Caron et al., 2000; Fonda Umani & Zanon, 2000; Fonda Umani & Beran,
2003; Fonda Umani et al., 2004; 2005; Sakka Hlaili et al., 2007). A tutt’oggi in poche
ricerche si è tenuto nel dovuto conto lo studio della composizione dei predatori (microzoo-
e nanoplanctonici) e della loro possibile crescita durante l’esperimento (Gifford, 1995;
Paranjape, 1990; Verity et al., 1993; Froneman & Perissinotto, 1996, Froneman et al.,
1996; Strom & Strom, 1996; James & Hall, 1998; Dolan et al., 2000; Fonda Umani &
Beran, 2003), anche se è di fondamentale importanza controllare i possibili cambiamenti
quali e quantitativi durante l’esperimento.
A differenza di altri metodi proposti, quali ad esempio l’uso di prede fluorescenti,
particolarmente utilizzato per lo studio dell’impatto dei batterivori (Sherr et al., 1986), il
metodo delle diluizioni, estremamente semplice, non prevede alcuna manipolazione degli
organismi e consente di ottenere sia il tasso specifico di crescita delle prede
(microfitoplancton, nanoplancton, picoplancton) sia quello di mortalità indotta da
predazione degli organismi eterotrofi (Båmstedt et al., 2000). Successive diluizioni
dell’acqua di mare con la stessa acqua filtrata su 0.22 µm, allo scopo di eliminare ogni
organismo presente, riducono le probabilità d’incontro tra preda e predatore e consentono
di stimare il tasso apparente di crescita delle prede e il tasso mortalità dovuta a predazione.
Il tasso specifico di crescita delle prede si ottiene estrapolando la crescita apparente
al 100% di diluizione (cioè il tasso di crescita in mancanza di predatori); il tasso di
mortalità da predazione degli eterotrofi corrisponde al valore assoluto dell’angolo della
retta di regressione tra la crescita apparente delle prede e le frazioni di acqua non filtrata.
Il metodo si basa su tre presupposti:
1- Il tasso di predazione è costante, indipendentemente dalla concentrazione
delle prede e di conseguenza il popolamento dei consumatori non varia durante
l’incubazione (Evans & Paranjape, 1992);
2- Il tasso di crescita delle prede non varia in seguito alle diluizioni, cioè la
crescita di una cellula non dipende dalla densità del popolamento e segue una legge
esponenziale. Deve essere dunque garantita la non limitazione di nutrienti nel periodo di
incubazione previsto dal protocollo;

Il Mediterraneo- Materiali e Metodi
20
3- Il tasso di predazione è linearmente correlato con le diverse
concentrazioni delle prede.
Il tasso di crescita delle prede può essere descritto dalla seguente equazione
Ct= C0e(k-g)t
che può anche essere scritta:
(1/t)ln(Ct /C0)= k-g
dove:
Ct = numero di individui o biomassa totale al tempo t soggetti a crescita e
predazione al tempo t
C0= numero di individui o biomassa totale al t0
k= coefficiente istantaneo di crescita delle prede
g= coefficiente istantaneo di mortalità delle prede, dovuto alla predazione
t= tempo di incubazione, in genere 24 ore per comprendere il ciclo giornaliero
completo.
Il termine k, dato dal primo postulato non viene influenzato dalle diluizioni, ma
rimane costante. Il coefficiente g, in accordo con il secondo postulato, varia in modo
direttamente proporzionale alla densità dei predatori, risultando quindi indipendente dalle
variazioni della densità delle prede. k e g possono variare senza modificare il tasso di
crescita delle prede in condizioni naturali nelle differenti diluizioni.
Dal momento che k è costante e g è direttamente proporzionale alla diluizione, le
equazioni con le due incognite k e g possono essere risolte graficamente con una retta di
regressione relativa alla crescita apparente contro il fattore diluizione (fig.1.6).

Il Mediterraneo- Materiali e Metodi
21
Fig.1.6 Rappresentazione grafica del modello di regressione lineare
I coefficienti di crescita apparente, (1/t)ln(Ct /C0), sono riportati in ordinata, mentre
in ascissa vengono riportati i fattori di diluizione.
L’intercetta della retta con l’asse y, ossia il punto dove g=0, rappresenta il
coefficiente di crescita istantaneo k in assenza di predatori; la pendenza della retta
rappresenta invece il valore negativo del coefficiente istantaneo di mortalità dovuta alla
predazione , -g.
Conoscendo la concentrazione delle prede all’inizio dell’esperimento (C0), il
coefficiente istantaneo di crescita delle prede (k) ed il coefficiente di mortalità da
predazione (g) è possibile ricavare un altro utile parametro, il tasso di ingestione (I),
identificato con la quantità di prede eliminate dai predatori nell’unità di tempo (t) e di
volume.
Innanzitutto si calcola la concentrazione media delle prede <C> nel corso
dell’esperimento, mediante l’equazione:
<C>= (C0e(k-g)t- C0)/(k-g)
E poi si procede al calcolo del tasso di ingestione (µgCl-1g-1):
I= g*<C>

Il Mediterraneo- Materiali e Metodi
22
La produzione reale (Pr) rappresenta la quantità di biomassa (µgCl-1) prodotta dal
popolamento delle prede in presenza del predatore durante l’esperimento:
Pr = C0e(k-g)- C0
La produzione potenziale (Pp), invece è la quantità di biomassa (µgCl-1) che
avrebbe potuto essere prodotta dal popolamento in assenza del predatore:
Pp= C0 ek- C0
La produzione potenziale rimossa dalla predazione (PP%) rappresenta l’incidenza
della predazione sulla produzione potenziale:
PP% = [(Pp-Pr)/Pp] *100
Inoltre, è possibile calcolare la biomassa iniziale rimossa dalla predazione (SP%):
SP%= [(Pp-Pr)/(Pr+C0)] *100
1.3.4 Campionamento
Lo scopo principale di questo lavoro è stato quello studiare, applicando il metodo
delle diluizioni, la predazione da parte del microzooplancton sul comparto nano e
picoplanctonico. Si è voluto anche studiare la distribuzione del microzooplancton lungo il
transetto del mediterraneo orientale.

Il Mediterraneo- Materiali e Metodi
23
Fig.1.7 Rosette con bottiglie Niskin
Gli esperimenti di campionamento sono stati condotti a bordo della nave
oceanografica Universitas. Il campionamento di acqua di mare superficiale è stato condotto
utilizzando una rosette con 24 bottiglie Niskin da 12 L cad. (fig.1.7). Sono stati raccolti
circa 100 L d’acqua per ogni stazione campionata. Allo scopo di eliminare gli eventuali
predatori di taglia superiore, l’acqua è stata immediatamente filtrata con retino a maglia
200µm. Un’aliquota della stessa acqua è stata filtrata con pompa peristaltica (fig.1.8) e
filtro da 0.22 µm allo scopo di ottenere acqua pura, priva di qualsiasi organismo utilizzata
per l’allestimento delle diluizioni.
Fig.1.8 Pompa peristaltica per la filtrazione
Le operazioni di filtrazione sono state condotte con molta attenzione per evitare il
danneggiamento delle cellule con conseguente alterazione dei risultati dell’esperimento.
Sono state così allestite quattro diluizioni (100%, 80%, 50%, 20%) in tre repliche
ciascuna per il C0 e per il C24 per quattro parametri (micozooplancton, microfitoplancton,
nano- e picoplancton), per un totale di 24 bottiglie (12 bottiglie in policarbonato da

Il Mediterraneo- Materiali e Metodi
24
incubazione e 12 bottiglie in plastica per il C0) per il microplancton ed altre 24 bottiglie per
il nano ed il picoplancton (da 250 ml e da 50 ml ciascuna).
I campioni iniziali al C0 sono stati immediatamente fissati con formalina al 2% e
mantenuti al freddo (5°C) e al buio. Solo per la frazione picoplanctonica la formalina è
stata prefiltrata con un filtro a siringa Acrodisc per eliminare ogni possibile impurità. I
campioni di nanoplancton (C0 e C24) sono stati conservati in gluteraldeide all’1% (Sherr &
Sherr, 1993).
Considerata l’elevata oligotrofia del bacino del Mediterraneo orientale, ed allo
scopo di favorire la crescita del fitoplancton durante il periodo di incubazione, sono stati
aggiunti i seguenti nutrienti: 5 µM NaNO3 e 1 µM KH2PO4 rispettivamente. Allo scopo di
mantenere le condizioni ambientali costanti si è provveduto ad incubare i campioni in un
incubatore alle stesse condizioni di luce e di temperatura dell’ambiente naturale (fig.1.9).
Fig.1.9 Incubazione delle bottiglie in situ
1.3.5 Distribuzione del microzooplancton
E’ stata condotta anche un’analisi relativa alla distribuzione e composizione del
popolamento microzooplanctonico lungo il transetto ovest – est del bacino del
Mediterraneo, raccogliendo 5 L di acqua superficiale con le bottiglie Niskin. Di questi 300
ml venivano messi in bottiglie di vetro e fissate con Lugol al 2%, mentre l’acqua rimanente
veniva immediatamente filtrata con filtro a 10µm con filtrazione inversa. I campioni
concentrati sono stati messi in bottiglie da 250 ml, fissati con formalina al 2%, conservati
al buio e al freddo (5°C). Il Lugol e la formalina sono i fissativi più comunemente usati per

Il Mediterraneo- Materiali e Metodi
25
conservare al meglio i ciliati. Questa doppia fissazione è stata effettuata per comparare gli
effetti dei due fissativi, cercando di evidenziarne i vantaggi e gli svantaggi.
1.3.6 Analisi quali-quantitativa
L’analisi quali-quantitativa dei campioni è stata condotta nei laboratori del
Dipartimento di Scienza della Vita presso l’Università di Trieste con la supervisione della
prof.ssa Serena Fonda Umani.
a) Picoplancton
I campioni di picoplancton conservati in formalina prefiltrata al 2% sono stati
filtrati con pompa di filtrazione che produce una depressione tra 0.2-0.3 atm, utilizzando
un sottofiltro nero in policarbonato (NTG) da 0.2 µm sul quale è stato sovrapposto un
filtro in cellulosa da 0.4 µm (Millipore) (fig.1.10).
Fig.1.10 Rampa di filtrazione
L’analisi dei campioni è stata fatta seguendo la modifica del metodo di Porter e
Feig (1980). La componente eterotrofa è stata colorata al buio con DAPI (4’6 – diamidio -
2- phenylindole) per 15 minuti ad una concentrazione finale di 1µg mL-1. La molecola del
fluorocromo si lega in una conformazione quasi planare al solco del DNA, situato al centro
dell’elica formando un legame idrogeno. Il complesso DAPI-DNA, eccitato dai raggi UV,
fluoresce nel blu, mentre il fluorocromo legato in modo aspecifico fluoresce nel giallo
(Porter & Feig, 1980) (fig.1.11).

Il Mediterraneo- Materiali e Metodi
26
La componente autotrofa è stata filtrata separatamente con volumi maggiori ed
osservata al microscopio ad epifluorescenza.
Il picoplancton eterotrofo è stato filtrato in 9 repliche mentre per la componente
autotrofa sono state filtrate 3 repliche. I filtri sono stati mantenuti al buio alla temperatura
di -20° C fino all’analisi dei campioni. Al momento dell’analisi i filtri sono stati
posizionati sul vetrino portaoggetti tra due gocce di olio da immersione e coperti con un
vetrino coprioggetti.
Fig.1.11 Picoplancton eterotrofo (sx) ed aurotrofo visto all’epifluorescenza (dx)
Il conteggio del picoplancton è stato condotto con un microscopio ad
epifluoresenza Olympus BX60 F5 con lampada a mercurio 100W con obiettivo ad
immersione 100X (fig.1.12)
Fig.1.12 Microscopio ad epifluorescenza
L’abbondanza della frazione autotrofa è stata conteggiata usando la luce blu (λ=
450-490 nm) contando circa 150 cellule per filtro, mentre per la componente eterotrofa si è

Il Mediterraneo- Materiali e Metodi
27
utilizzata la luce UV (λ=365 nm) e sono state contate almeno 200 cellule per campo. Il
numero di cellule contate, sia per la componente eterotrofa che per quella autotrofa è stato
convertito in biomassa di carbonio usando un fattore di conversione di 20 fg C cell-1
(Ducklow & Carlson, 1992) e 200 fgC cell-1 (Caron et al., 1991) rispettivamente.
b) Nanoplancton
I campioni per la determinazione quantitativa del nanoplancton sono stati fissati
con gluteraldeide fino ad ottenere una concentrazione finale dell’1% (Verity et al., 1993) e
conservati in frigorifero. Successivamente in laboratorio l’acqua è stata filtrata in 3
repliche per ciascuna quota di diluizione (100%, 80%, 50%, 20%) utilizzando sottofiltri
Millipore da 1.2 µm di porosità per uniformare la filtrazione e filtri in policarbonato NTG
black 0.8 µm di porosità. Prima di procedere alla filtrazione i campioni sono stati colorati
con una soluzione di fluorocromo DAPI (4’-6-diamino-2-fenilindolo) con concentrazione
finale di 1µg mL-1 lasciata agire per circa 15’. I filtri sono stati mantenuti a – 20°C fino al
momento dell’analisi al microscopio.
Fig.1.13 Nanoplancton al microscopio ad epifluorescenza
Al momento dell’analisi i filtri sono stati posizionati sul vetrino portaoggetto tra
due gocce di olio da immersione e coperti con un vetrino portaoggetti. I conteggio sono
stati eseguiti al microscopio ad epifluorescenza Olympus BX 60 dotato di lampada a
vapori di mercurio (100 Watt), utilizzando un obiettivo 100X. Per distinguere le cellule dal
materiale detritico si osservano i campioni con raggi UV (λ=365nm). Il nanoplancton
eterotrofo osservato in luce blu-violetto (λ = 450-490 nm), appare di colore verde (Booth,

Il Mediterraneo- Materiali e Metodi
28
1993), mentre le cellule degli autotrofi si distinguono per la presenza al loro interno dei
cloroplasti di colore rosso-arancione dovuto all’autofluorescenza della clorofilla (fig.1.13).
La componente autotrofa non era distinguibile dalla componente eterotrofa, fatto questo
probabilmente dovuto alla perdita della fluorescenza della chl a quindi il nanoplancton è
stato conteggiato suddividendolo in 3 classi dimensionali: < 3 µm, 3-5- µm, >5 µm. La
frazione compresa tra 10-20µm che normalmente appartiene al nanoplancton, in questo
caso è stata analizzata al microscopio invertito utilizzando la tecnica di conteggio applicata
per il microplancton allo scopo di effettuare una corretta distinzione degli organismi con
dimensioni < di 20 µm. Per il nanoplancton sono state contate almeno 100 cellule per
filtro. Il numero di cellule litro è stato poi convertito in biomassa di carbonio utilizzando il
fattore di conversione di 183 fg C µm-3 (Caron et al., 1995).
c) Microplancton
I campioni per la determinazione del microzoo e microfitoplancton sono stati
conservati in formalina al 2% e mantenuti al freddo (5°C) e al buio fino al momento
dell’analisi in laboratorio. I campioni sono stati preconcentrati da un volume iniziale di 2 L
a circa 200 ml. Di questi, 100 ml sono stati messi a sedimentare per 72 ore (3 ore per cm di
altezza) in apposite colonnine di sedimentazione secondo il metodo Uthermöhl (1958)
(fig.1.14). L’analisi quali-quantitativa dei comparti microplanctonici è avvenuta al
microscopio
rovesciato Labovert utilizzando un obiettivo a 32 ingrandimenti. I campioni sono stati
conteggiati in 3 repliche al T0 e al T24. Il conteggio della componente eterotrofa è stato
effettuato sull’intera cameretta (fig.1.15).
Fig.1.14 Colonnina di sedimentazione

Il Mediterraneo- Materiali e Metodi
29
Fig.1.15 Cameretta di sedimentazione e microscopio rovesciato Labovert
I tintinnidi sono stati determinati in base alla forma e dimensione della lorica in
accordo con Kofoid e Campbell (1929, 1939) e Marshall (1973); i ciliati aloricati,
foraminiferi, radiolari e acantari, sono stati determinati seguendo le descrizioni di
Neudruck Asher et al., (1929). L’identificazione delle larve di metazoi e dei naupli di
copepodi è stata fatta in accordo con Trègouboff e Rose (1957). Gli altri piccoli flagellati
sono stati riconosciuti secondo Throndsen (1997) mentre i coccolitoforali secondo Heimdal
(1997). Il termine nanociliati descrive i ciliati < 20 µm (Pitta et al., 2001). Il numero di
individui conteggiato è stato poi convertito in cellule litro e biomassa di carbonio
determinando prima i biovolumi associando gli organismi a formule geometriche standard
(Edler, 1979) poi il biovolume è stato convertito in contenuto di carbonio usando specifici
fattori di conversione disponibili in letteratura (tab.1.4).
Microplancton Fattore di
conversione Bibliografia Ciliati aloricati 0.14 Putt & Stoeker, 1989 Nanociliati, Dinoflagellati < 20µm, Coccolitoforali, Foraminiferi < 50 µm 0.183 Caron et al., 1995a Ciliati loricati: Tintinnidi 444.5+(bv*0.053) Verity & Langdon, 1984 Dinoflagellati atecati 0.13 Lessard, non pubblicato Dinoflagellati tecati 0.14 Lessard, 1991 Foraminiferi > 50 µm 0.089 Michaels et al., 1995 Acantari 0.0026 Michaels et al., 1995 Larve mesoplanctoniche Naupli di copepodi 0.08 Beers & Stewart, 1970 Radiolari 0.14 Michaels et al., 1995 Diatomee 0.288*bv*0.811 Menden Deur & Lessard, 2000
Tab.1.4 Fattori di conversione per il microplancton

Il Mediterraneo- Materiali e Metodi
30
1.3.7 Produzione secondaria
Per stimare la crescita del microzooplancton dopo il tempo di incubazione previsto
(produzione secondaria) è stata calcolata la biomassa (µgCL-1) all’inizio dell’esperimento
(C0) e alla fine dell’esperimento (C48) per le tre repliche al 100% utilizzando il metodo
Uthermöhl (1958). 100ml di campione sono stati messi a sedimentare per 72 ore e l’analisi
dei campioni è stata condotta al microscopio invertito Labovert a 32 ingrandimenti
conteggiando gli organismi presenti nell’intera cameretta. Il numero di cellule conteggiato
è stato convertito prima in cell L-1 e poi in biomassa di carbonio attraverso fattori specifici
di conversione presenti in letteratura.
Il Mediterraneo -Risultati
31
1.4 RISULTATI
1.4.1 Dati ambientali
Le variabili ambientali per tutte le stazioni del Mediterraneo orientale sono
riassunte nella tabella 1.5. La temperatura aumenta passando dalla stazione V6 a quella
geograficamente più orientale V10. Lo stesso andamento si osserva per alla salinità che
aumenta allo stesso modo passando da valori pari a 38.00 a valori di 39.36 nella stazione
più orientale.
La Chl a è relativamente bassa in tutte le stazioni campionate in accordo con la
stagione di campionamento in cui si evidenziano scarse concentrazioni di nutrienti.
Tab1.5 Variabili ambientali. Temperatura, salinità, parametri biogeochimici,
produzione primaria e chl a sono stati gentilmente forniti da DISMAR (Università
di Ancona); CNR – ISMAR (Trieste); U.O. Saggiomo Stazione Zoologica A.
Dohrn (Napoli).
1.4.2 Predazione sul picoplancton eterotrofo
Inizio dell’incubazione (C0): l’analisi dei campioni all’inizio dell’esperimento (C0) serve
a dimostrare che ogni esperimento (V6, V7, Viera e V10) è stato correttamente allestito:
Stazioni V6 V7 Viera V10 Temperatura °C 22.66 22.49 23.4 24.33 Salinità (psu) 38.00 38.73 39.27 39.36
Tot chl (µg L-1) 0.059 0.046 0.035 0.057
PP (µgCm3 h-1) 0.44 0.27 0.33 0.51
NO3 µM-N 0.78 0.07 0.09 0.18
NO2 µM-N 0.13 0.03 0.02 0.02
SiO2 µM-N 0.77 1.01 1.15 0.85
PO4 µM-N 0.12 0.01 0.01 0.02

Il Mediterraneo -Risultati
32
all’aumentare del fattore diluizione c’è un’effettiva diminuzione nel numero di organismi
(fig.1.16,1.17,1.18,1.19).
St.V6 Picoplancton eterotrofo
y = 6.2622x + 1.2983r2 = 0.957
0
2
4
6
8
10
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
St.V7Picoplancton eterotrofo
y = 4.8972x + 0.7244r2 = 0.8234
0
2
4
6
8
10
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.16 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.97
Fig.1.17 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.90

Il Mediterraneo -Risultati
33
St. Viera Picoplancton eterotrofo
y = 4.1055x + 0.6736r2 = 0.9846
0
1
2
3
4
5
6
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
St. V10Picoplancton eterotrofo
y = 4.2841x + 1.6771r2 = 0.8742
0
2
4
6
8
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.19 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.93
Fig.1.18 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.99

Il Mediterraneo -Risultati
34
Fine dell’incubazione (C24): il tasso di crescita apparente è stato calcolato per ogni
diluizione in ogni stazione.
Nella stazione V6 (fig. 1.20) c’è una relazione significativa tra la crescita apparente
ed il fattore diluizione (r=0.71; p<0.001). Il coefficiente istantaneo di crescita k=1.408 ed il
coefficiente istantaneo di mortalità indotta da predazione g=-0.881 determina un tasso di
ingestione I= 9.51 µgC L-1 d-1.
St. V6 Picoplancton eterotrofo
y = -0,8817x + 1,4084r2 = 0,5147
0
0.5
1
1.5
2
2.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.20 Il tasso di crescita apparente presenta una correlazione significativa con p<0.001 r = 0.71
Nella stazione V7 (fig. 1.21) c’è una relazione significativa tra il tasso di crescita
apparente ed il fattore diluizione (r= 0.97 p<0.001). Il coefficiente istantaneo di crescita k=
1.66 ed il coefficiente istantaneo di mortalità indotta da predazione g=-1.405, determina
un tasso di ingestione I = 9.58 µg C L-1 d-1.

Il Mediterraneo -Risultati
35
St. V7Picoplancton eterotrofo
y = -1,4057x + 1,6661r2 = 0,9442
0
0.5
1
1.5
2
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Nella stazione Viera (fig. 1.22) c’è una relazione significativa tra il tasso di crescita
apparente ed il fattore diluizione (r=0.93 p< 0.001). Il coefficiente istantaneo di crescita
k= 2.09 ed il coefficiente istantaneo di mortalità indotta da predazione g=-1.838, determina
un tasso di ingestione I = 11.80 µgCL-1d-1
St. Viera Picoplancton eterotrofo
y = -1,838x + 2,0939r2 = 0,8693
0
0.5
1
1.5
2
2.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Nella stazione V10 fig. 23 c’è una relazione significativa tra il tasso di crescita
apparente ed il fattore diluizione (r=0.97 p< 0.001). Il coefficiente istantaneo di crescita
Fig 1.21 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.97
Fig.1.22 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.93

Il Mediterraneo -Risultati
36
k= 2.15 ed il coefficiente istantaneo di mortalità indotta da predazione g=-1.712, determina
un tasso di ingestione I = 13.06 µgC L-1 d-1 .
St. V10Picoplancton eterotrofo
y = -1,7124x + 2,1507r2 = 0,9526
0
0.5
1
1.5
2
2.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
I risultati ottenuti negli esperimenti di diluizione sono riportati nella tabella 1.6,
dove k = coefficiente istantaneo di crescita del popolamento picoplanctonico, g è il
coefficiente istantaneo di mortalità indotta da predazione; C0 è la concentrazione del
picoplancton eterotrofo all’inizio dell’esperimento; Cm è la concentrazione media delle
prede durante l’incubazione; I tasso di ingestione; Pr è la produzione reale in presenza dei
predatori durante l’esperimento; Pp è la produzione potenziale.
St. k g C0
µgCL -1 Cm
µgCL -1 I
µgCL -1 d-1 Pr
µgCL -1d-1 Pp
µgCL -1d-1 V6 1,40 0,88 7,48 10,80 9,51 3,94 15,21 V7 1,66 1,40 5,89 6,82 9,58 1,50 9,78 Viera 2,09 1,83 4,87 6,42 11,80 1,24 10,20 V10 2,15 1,71 6,14 7,63 13,06 2,68 13,20
Tab.1.6 Predazione del microzooplancton sul picoplancton eterotrofo
Fig.1.23 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.97

Il Mediterraneo -Risultati
37
1.4.3 Predazione sul picoplancton autotrofo Inizio dell’incubazione (C0):l’ analisi dei campioni all’inizio dell’esperimento (C0) serve
a dimostrare che ogni esperimento (V6, V7, Viera, V10) è stato correttamente allestito;
all’aumentare del fattore di diluizione c’è un’effettiva diminuzione nel numero di
organismi (fig. 1.24,1.25,1.26,1.27)
St. V6Picoplancton autotrofo
y = 0,4096x + 0,0223r2 = 0,8101
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
St. V7Picoplancton autotrofo
y = 0.2406x + 0.1486r2 = 0.4193
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.24 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.9
Fig.1.25 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.64

Il Mediterraneo -Risultati
38
St. Viera Picoplancton autotrofo
y = 0.3047x + 0.0878r2 = 0.7401
0
0.1
0.2
0.3
0.4
0.5
0.6
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.26 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.86
St. V 10 Picoplancton autotrofo
y = 0.2756x + 0.4313r2 = 0.4346
0
0.2
0.4
0.6
0.8
1
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.27 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.65
Fine dell’incubazione (C24): il tasso di crescita apparente è stato calcolato per ogni
diluizione in ogni stazione.
I grafici della crescita apparente verso il fattore diluizione non mostrano alcuna relazione
significativa in nessuna delle quattro stazioni campionate (fig.1.28, 1.29, 1.30,1.31).

Il Mediterraneo -Risultati
39
St. V6Picoplancton autotrofo
00.10.20.30.40.5
0.60.7
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C0)
Fig.1.28 Relazione tra crescita apparente e fattore diluizione
St. V7Picoplancton autotrofo
-1.6
-1.4
-1.2
-1
-0.8
-0.6
-0.4
-0.2
0
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C
0)
Fig.1.29 Relazione tra crescita apparente e fattore diluizione

Il Mediterraneo -Risultati
40
St. V10Picoplancton autotrofo
-1.2
-1
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C
0)
Fig.1.30 Relazione tra crescita apparente e fattore diluizione
St. VieraPicoplancton autotrofo
-0.4-0.3-0.2-0.1
00.10.20.3
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C0)
Fig.1.31 Relazione tra crescita apparente e fattore diluizione

Il Mediterraneo -Risultati
41
1.4.4 Predazione sul nanoplancton
L’analisi dei campioni è stata fatta suddividendo il nanoplancton in tre classi
dimensionali (< 3µm, 3-5 µm, >5 µm) senza distinzione fra autotrofi ed eterotrofi.
Inizio dell’incubazione (C0):l’analisi dei campioni all’inizio dell’esperimento (C0) ha
dimostrato il corretto allestimento dell’esperimento nelle stazioni campionate (V6, V7,
V10 e Viera). All’aumentare del fattore di diluizione corrisponde una riduzione nel numero
di organismi (fig.1.32, 1.33, 1.34, 1.35).
St. V6Nanoplancton
y = 2,6846x + 0,5882r2 = 0,6283
0
1
2
3
4
5
6
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.32 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.79
St.V7Nanoplancton
y = 5,7376x - 0,5588r2 = 0,7396
012345678
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.33 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.85

Il Mediterraneo -Risultati
42
St.V10Nanoplancton
y = 2,7023x + 0,3956r2 = 0,8793
00.5
1
1.52
2.53
3.54
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.34 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una correlazione significativa p<0.001; r = 0.93
St. VieraNanoplancton
y = 9,7576x - 2,4152r2 = 0,7661
-2.0
0.0
2.0
4.0
6.0
8.010.0
12.0
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.1.35 Abbondanza (µgC L-1) all’inizio dell’esperimento (C0) presenta una
correlazione significativa p<0.001; r = 0.87
Fine dell’incubazione (C24): il tasso di crescita apparente è stato calcolato per ogni
classe dimensionale (< 3µm, 3-5 µm > 5 µm). Nella stazione V6 c’è una correlazione
significativa tra il tasso di crescita apparente e il fattore diluizione relativamente alla
frazione < 3 µm (r=0.74; p< 0.01). Il coefficiente istantaneo di crescita delle prede k=0.878
ed il coefficiente di mortalità da predazione g= 0.721 determinano un tasso di ingestione I=
0.142 µgC L-1 d-1 (fig.1.36).

Il Mediterraneo -Risultati
43
St. V6Nanoplancton <3 µm
y = -0.7216x + 0.8789r2 = 0.5597
0
0.2
0.4
0.6
0.8
1
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.36 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.74
Nella stessa stazione c’è anche una correlazione significativa tra il tasso di crescita
apparente e il fattore diluizione relativamente alla frazione compresa 3-5 µm (r=0.78; p<
0.01). Il coefficiente istantaneo di crescita delle prede k=0.592 ed il coefficiente di
mortalità da predazione g= 0.398 determinano un tasso di ingestione I= 0.169 µgC L-1 d-1
(fig.1.37).
St. V6Nanoplancton 3-5 µm
y = -0.3983x + 0.5925r2 = 0.6191
0
0.2
0.4
0.6
0.8
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (C
t/C0)
Fig.1.37 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.78
Così come c’è una correlazione anche significativa tra il tasso di crescita apparente
e il fattore diluizione relativamente alla frazione >5 µm (r=0.64; p< 0.01). Il coefficiente
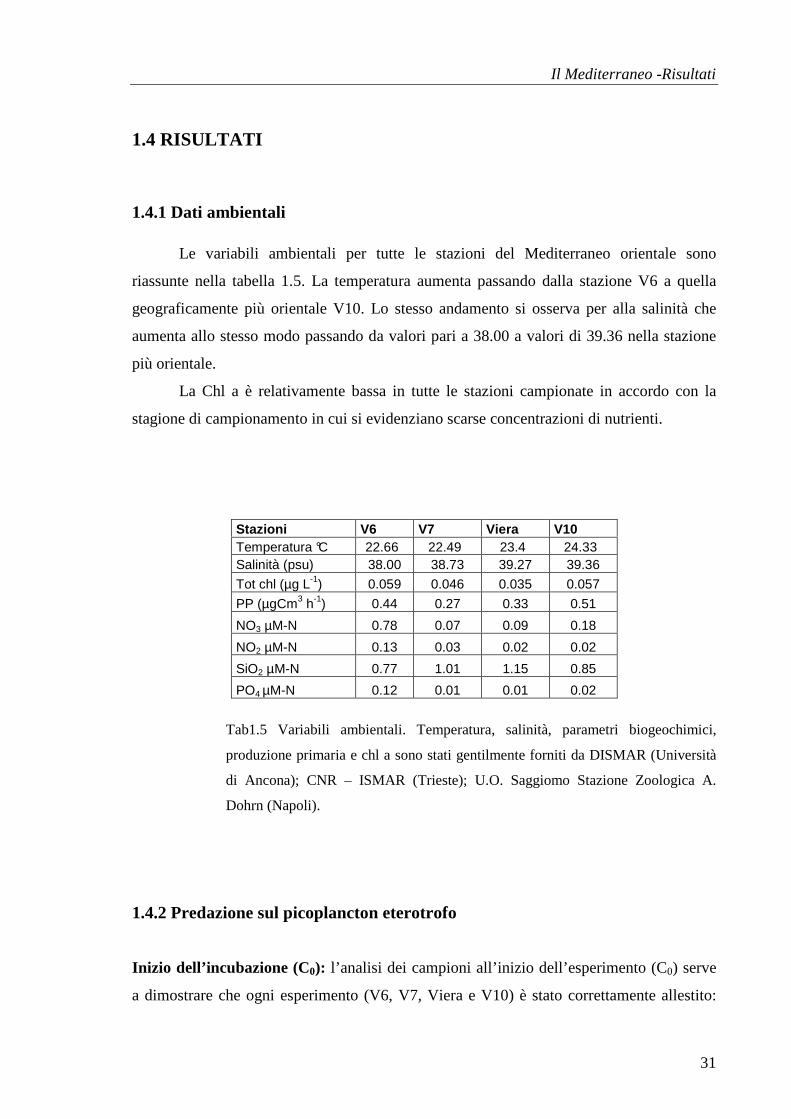
istantaneo di crescita delle prede k= 1.02 ed il coefficiente di mortalità da predazione g=
1.145 determinano un tasso di ingestione I= 0.829 µgC L-1 d-1 (fig.1.38).

Il Mediterraneo -Risultati
44
St. V6Nanoplancton >5 µm
y = -1.1452x + 1.0272r2 = 0.4279
-1-0.5
0
0.5
1
1.5
2
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (C
t/C0)
Fig.1.38 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.64
Nella stazione V7 c’è una correlazione significativa tra il tasso di crescita apparente
e il fattore diluizione relativamente alla frazione <3µm (r=0.64; p< 0.01). Il coefficiente
istantaneo di crescita delle prede k= 0.152 ed il coefficiente di mortalità da predazione g=
2.091 determinano un tasso di ingestione I= 0.273 µgC L-1 d-1 (fig.1.39)
St. V7Nanoplancton < 3 µm
y = -2.0914x + 1.5279r2 = 0.6909
-2-1.5
-1-0.5
00.5
11.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.39 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.83
Nella stessa stazione c’è una correlazione significativa tra il tasso di crescita
apparente e il fattore diluizione relativamente alla frazione compresa tra 3-5 µm (r=0.82;
p< 0.01). Il coefficiente istantaneo di crescita delle prede k= 2.163 ed il coefficiente di
mortalità da predazione g= 1.267 determinano un tasso di ingestione I= 0.541 µgC L-1 d-1
(fig.1.40).

Il Mediterraneo -Risultati
45
St. V7 Nanoplancton 3-5 µm
y = -1.2675x + 2.1638r2 = 0.6854
0
0.5
1
1.5
2
2.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.40 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.82
Al contrario non c’è alcuna correlazione significativa tra la crescita apparente ed il
fattore diluizione relativamente alla classe dimensionalmente > 5 µm (fig.1.41).
St. V7 Nanoplancton > 5 µm
0
0.5
1
1.5
2
2.5
3
0% 20% 40% 60% 80% 100%
Diluizioni
LN(C
t/C0)
Fig.1.41 Relazione fra crescita apparente e fattore diluizione
Nella stazione V10 non si verifica correlazione significativa per il nanoplancton di
dimensioni < 3 µm (fig.1.42).

Il Mediterraneo -Risultati
46
St. V10 Nanoplancton < 3 µm
-1
-0.5
0
0.5
1
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.42 Relazione fra crescita apparente e fattore diluizione
C’è una correlazione significativa tra il tasso di crescita apparente ed il fattore
diluizione relativamente alla frazione compresa tra 3-5 µm (r=0.82; p< 0.01). Il
coefficiente istantaneo di crescita delle prede k=0.830 ed il coefficiente di mortalità da
predazione g= 0.899 determinano un tasso di ingestione I= 0.249 µgC L-1 d-1 (fig.1.43)
St. V10Nanoplancton 3-5 µm
y = -0.8991x + 0.8304
r2 = 0.5577
-0.5
0
0.5
1
0% 20% 40% 60% 80% 100%
Diluizioni
LN(C
t/C0)
Fig.1.43 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.74
Così come c’è una correlazione significativa tra il tasso di crescita apparente ed il
fattore diluizione relativamente alla frazione >5 µm (r=; p< 0.01). Il coefficiente istantaneo
di crescita delle prede k=0.618 ed il coefficiente di mortalità da predazione g= 1.12
determinano un tasso di ingestione I= 0.688 µgC L-1 d-1 (fig.1.44)
Il Mediterraneo -Risultati
47
St. V10 Nanoplancton > 5 µm
y = -1.1227x + 0.618r2 = 0.5013
-1
-0.5
0
0.5
1
0% 20% 40% 60% 80% 100%
Diluizioni
LN(C
t/C0)
Fig.1.44 Il tasso di crescita apparente evidenzia una correlazione significativa con p<0.001; r = 0.70
Nella stazione Viera non c’è correlazione significativa tra il tasso di crescita
apparente ed il fattore diluizione per nessuna delle 3 classi dimensionali analizzate (fig.
1.45, 1.46, 1.47).
St. Viera Nanoplancton < 3 µm
-2
-1.5
-1
-0.5
0
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.45 Relazione fra crescita apparente e fattore diluizione

Il Mediterraneo -Risultati
48
St. Viera Nanoplancton 3-5 µm
-1-0.8-0.6-0.4-0.2
00.20.40.60.8
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.1.46 Relazione fra crescita apparente e fattore diluizione
St. Viera Nanoplancton > 5 µm
-2
-1.5-1
-0.5
0
0.51
1.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C0)
Fig.1.47 Relazione fra crescita apparente e fattore diluizione
1.4.5 Microzooplancton L’analisi dei campioni ha permesso di compilare una lista faunistica Sono stati
trovati ciliati aloricati, tintinnidi, dinoflagellati, foraminiferi, radiolari, acanatari, piccoli
metazoi quali nauplii di copepodi, piccoli copepoditi e piccole larve di metazoi.
Tra i ciliati aloricati sono stati identificati 6 generi per un totale di 7 specie: Laboea
strobila, Leegardiella spp. Lhomaniella spp. Strombidium conicum, S capitatum, S .
conicum, S. epidemum, S. neptuni, Strombilidium spp., Tontonia gracillima.
Tra i tintinnidi sono stati identificate 7 generi per un totale di 11 specie identificate:
Codonella brevicollis, Craterella torulata, Dadayiella ganymedes, Epiplocys blanda,

Il Mediterraneo -Risultati
49
Eutintinnus apertus, E. lusus-undae, E.macilentus, E. tubulosus, Rhabdonella cornucopia,
R. spiralis, Tintinnopsis compressa.
Tra i dinoflagellati sono stati identificati 4 generi con 10 specie: Gymnodinium spp.,
Gyrodinium fusiforme, G. impudicum, Ornithocercus magnificus, Protoperidinium
brevipes, P. crassipes, P. diabolus, P. divergens, P. oceanicum, P. steini, P. subinerme.
Tra i foraminiferi è stato identificato un solo genere: Globigerina spp.

Il Mediterraneo -Risultati
50
LISTA FAUNISTICA
PROTOZOA
CILIOPHORA
SPIROTRICHIA
OLIGOTRICHIDA
Strombidium acutum (Leegaard, 1915) (Kahl, 1932)
Strombidium capitatum (Leegaard, 1915) (Kahl, 1932)
Strombidium conicum (Lohmann, 1908) (Wulff, 1919)
Strombidium epidemum (Lynn Montagnes & Small, 1955)
Strombidium neptuni (Montagnes & taylor, 1904)
Strombilidium spp.
Tontonia gracillima (Fauré-fremiet, 1924)
Laboea strobila (Lhomann, 1908)
Leegaardiella spp.
Lhomanniella spp.
Oligotrichida indet.
PERITRICHIDA
Peritrichida indet.
HOLOTRICHIDA
Holotrichida indet.
TINTINNIDA
Acanthostomella conicoides (Kofoid & Campbell, 1939)
Acanthostomella minutissima (Kofoid & Campbell, 1929)
Amplectella occidentalis (Kofoid & Campbell, 1929)
Canthariella brevis (Kofoid & Campbell, 1929)
Codonella brevicollis (Kofoid & Campbell, 1929)
Coxliella laciniosa (Brandt, 1906)
Craterella armilla (Kofoid & Campbell, 1929)
Craterella torulata (Kofoid & Campbell, 1929)
Dadayiella ganymedes (Kofoid & Campbell, 1929)
Dictyocysta polygonata (Kofoid & Campbell, 1929)
Epiplocylis undella (Kofoid & Campbell, 1929)
Eutintinnus apertus (Kofoid & Campbell, 1929)

Il Mediterraneo -Risultati
51
Eutintinnus fraknoii (Daday, 1887)
Eutintinnus lusus-undae (Entz, 1885)
Eutintinnus macilentus ((Jørgensen, 1924)
Eutintinnus medium (Kofoid & Campbell, 1929)
Eutintinnus tubulosus (Ostenfeld, 1899)
Protorhabdonella curta (Cleve) (Jørgensen, 1924)
Rhabdonella spiralis (Fol) (Brandt)
Salpingella acuminata (Claparède & Lachmann, 1859)
Salpingella curta (Kofoid & Campbell, 1929)
Salpingella decurtata (Kofoid & Campbell, 1929)
Salpingella laminata (Kofoid & Campbell, 1939)
Salpingella rotundata (Kofoid & Campbell, 1939)
Steenstrupiella gracilis (Kofoid & Campbell, 1939)
Steenstrupiella steenstrupii (Claparède & Lachmann, 1858)
Stenosemella nivalis (Meunier, 1910)
Tintinnopsis beroidea (Stein, 1864)
Tintinnopsis minuta (Wailes, 1925)
Tintinnopsis nana (Lohmann, 1908)
Xystonella longicaudata (Brandt 1907)
SARCOMASTIGOPHORA
MASTIGOPHORA
DINOFLAGELLIDA
Gymnodinium spp. (Stein, 1878)
Gyrodinium fusiforme (Kofoid & Swezy, 1921)
Gyrodinium impudicum (Fraga et.al., 1995)
Ornithocercus magnificus (Stein, 1883)
Protoperidinium brevipes (Paulsen, 1908) (Balech, 1974)
Protoperidinium crassipes (Kofoid) (Balech, 1974)
Protoperidinium diabolus (Cleve) (Balech, 1974)
Protoperidinium divergens (Ehrenberg) (Balech 1974)
Protoperidinium oceanicum (Van Höffen) (Balech, 1974)
Protoperidinium steini (Jørgensen) (Balech, 1974)
Protoperidinium subinerme (Pausen) (Loeblich III, 1970)
Torodinium robustum (Kofoid & Swezy, 1921)

Il Mediterraneo -Risultati
52
FORAMINIFERIDA
Foraminiferida indet.
Globigerina spp.
RADIOLARIA
Radiolaria indet.
ACANTHARIA
Acantharia indet.
METAZOA
Metazoa ova
Metazoi larvae
ARTHROPODA
CRUSTACEA
COPEPODA
Copepoda nauplii

Il Mediterraneo -Risultati
53
a) Strombilidium spp.; b) Protoperidinium bipes; c) Ciliato loricato; d) Strombidium spp. e) Gymnodinium spp.; f) Gymnodinium spp., in forma coloniale
a b
c d
e f

Il Mediterraneo -Risultati
54

Il Mediterraneo -Risultati
55
L’abbondanza e la biomassa del microzooplancton sono state valutate all’inizio
dell’esperimento (C0) per le tre repliche (fig.1.48, 1.49). I grafici mostrano come nel
Mediterraneo orientale le abbondanze del microzooplancton rimangono piuttosto costanti.
Ci sono delle variazioni in termini di biomassa, dovute esclusivamente alla taglia degli
organismi presenti nelle varie stazioni. Nelle stazioni V6 e V10 sono presenti organismi di
taglia minore con conseguente minor contenuto di carbonio.
Abbondanza del microzooplancton
020406080
100120140160180
V6 V7 V10 Viera
Cel
l L-1
Fig.1.48 Abbondanza del microzooplancton (cell L-1)
Biomassa del microzooplancton
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
V6 V7 V10 Viera
µgC
L-1
Fig.1.49 Biomassa del microzooplancton (µgCL-1)
Nella fig. 1.50 sono stati messi a confronto in termini di abbondanza i 5 gruppi di
microzooplancton identificati nei campioni: Ciliati aloricati, tintinnidi, dinoflagellati, altri
protozoi (foraminiferi radiolari, acantari), micrometazoi (larve di metazoi, e nauplii di
copepodi). Dominano i ciliati aloricati con valori piuttosto costanti in tutte e quattro le

Il Mediterraneo -Risultati
56
stazioni campionate, seguiti immediatamente dai dinoflagellati con un’abbondanza
piuttosto omogenea lungo tutto il bacino del Mediterraneo orientale.
.
Abbondanza del microzooplancton
0
20
40
60
80
100
120
V6 V7 V10 Viera
Cel
l L-1
Ciliati aloricati
Tintinnidi
Dinoflagellati
Altri protozoi
Micrometazoi
Fig.1.50 Abbondanze dei gruppi di microzooplancton (Cell L-1)
In termini di biomassa, fig.1.51 dominano sicuramente i micrometazoi nelle
stazioni del Mediterraneo orientale, i ciliati aloricati sono particolarmente abbondanti nella
stazione più orientale di Viera. Mediamente i valori di biomassa degli altri gruppi si
mantengono al di sotto dei 0.10 µgCL-1 .
Biomassa del microzooplancton
-0.05
0.00
0.05
0.10
0.15
0.20
0.25
0.300.35
0.40
V6 V7 V10 Viera
µgC
L-1
Ciliati aloricati
Tintinnidi
Dinoflagellati
Altri protozoi
Micrometazoi
Fig.1.51 Biomassa dei gruppi di microzooplancton (µgC L-1)

Il Mediterraneo -Risultati
57
1.4.6 Produzione secondaria
L’abbondanza e la biomassa dei predatori è stata stimata per le tre repliche al 100%
all’inizio ed alla fine dell’incubazione allo scopo di verificare quali sono i gruppi che
crescono durante il periodo di incubazione.
Nella stazione V6 i tintinnidi, i dinoflagellati ed in misura minore gli altri protozoi
trovano prede sufficienti che permettono loro di crescere durante il periodo
d’incubazione. Gli incrementi di crescita corrispondono al 47% per i tintinnidi; il166% per
i dinoflagellati eterotrofi ed il 100% per gli altri protozoi (Fig.1.52)
Biomassa del microzooplancton St. V6
-0.05
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
Ciliati aloricati Tintinnidi Dinoflagellati Altri protozoi Micrometazoi
µgC
L-1
C0 C48
Fig.1.52 Biomassa del microzooplancton al C0 e al C48
Nella stazione V7 invece i ciliati aloricati, i tintinnidi, i protozoi ed i micrometazoi
non trovano prede sufficienti per crescere durante l’incubazione. Si verifica un incremento
di crescita solamente a carico dei dinoflagellati con una percentuale pari al 36% (fig.1.53).

Il Mediterraneo -Risultati
58
Biomassa del microzooplancton St. V7
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
Ciliati aloricati Tintinnidi Dinoflagellati Altri protozoi Micrometazoi
µgC
L-1
C0 C48
Fig.1.53 Biomassa del microzooplancton al C0 e al C48
Nella stazione V10 crescono in maniera significativa dopo il periodo di incubazione
i dinoflagellati (62%) e gli altri protozoi (86%) (fig 1.54).
Biomassa del microzooplancton St. V10
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
Ciliati aloricati Tintinnidi Dinoflagellati Altri protozoi Micrometazoi
µgC
L-1
C0 C48
Fig.1.54 Biomassa del microzooplancton al C0 e al C48
Nella stazione Viera la maggior parte dei gruppi presenti non trova cibo sufficiente
per crescere e si verifica dunque un’evidente riduzione della biomassa dopo l’incubazione.
Solamente gli altri protozoi crescono lievemente durante il periodo di incubazione
(fig.1.55).

Il Mediterraneo -Risultati
59
Biomassa del microzooplancton St. Viera
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
Ciliati aloricati Tintinnidi Dinoflagellati Altri protozoi Micrometazoi
µgC
L-1
C0 C48
Fig.1.55 Biomassa del microzooplancton al C0 e al C48
1.4.7 Distribuzione
Per studiare la distribuzione del microzooplancton nel bacino del Mediterraneo si
sono messi a confronto i risultati ottenuti con tre metodi di raccolta e conservazione dei
campioni d’acqua tal quale al C0. Lo scopo dell’elaborazione statistica dei dati è stato
quello di confrontare i risultati delle 3 repliche (2 L di campione fissato in formalina al
2%) con i 5 L di campione raccolto, fissato in formalina al 2% e poi concentrato a 250 ml e
i 300 ml di campione raccolto e fissato in soluzione di Lugol al 2%. I dati sono stati
elaborati statisticamente in collaborazione con il prof. Feoli. L’analisi dei risultati ha
evidenziato come non ci sono differenze statisticamente significative fra le 3 repliche
provenienti dai 2 L fissati in formalina al 2% (fig.1.56) come evidenziato dai bassi valori
di deviazione standard.

Il Mediterraneo -Risultati
60
Distribuzione del microzooplancton in superficie
0
50
100
150
200
250
300
V6 V7 V10 Viera
Stazioni
Cel
l L-1
Fig.1.56 Distribuzione del microzooplancton relativa ai 2 L di campione in 3 repliche
La distribuzione superficiale del microzooplancton è stata anche stimata sulla base
dei 5 L di campione raccolto, conservato in formalina al 2 % e poi concentrato fino a
250ml (fig.1.57). Questa modalità di raccolta e conservazione del campione determina
generalmente una sottostima del numero di organismi realmente presenti nell’ambiente di
campionamento. Si evidenzia una sovrastima, rispetto ai 2 L in tre repliche, solamente
nella stazione V7 dove sono stati conteggiati in totale 427 ind. L -1
Distribuzione del microzooplancton in superficie (5L )
0
50
100
150
200
250
300
350
400
450
V6 V7 V10 Viera
Stazioni
Cel
l L-1
Fig.1.57 Distribuzione del microzooplancton relativa ai 5 L di campione concentrato
a 250 ml
Analoghe analisi sono state condotte su 300 ml d’acqua raccolta e fissata con una
soluzione di lugol al 2% (fig.1.58 ). Con questa modalità di conservazione dei campioni si
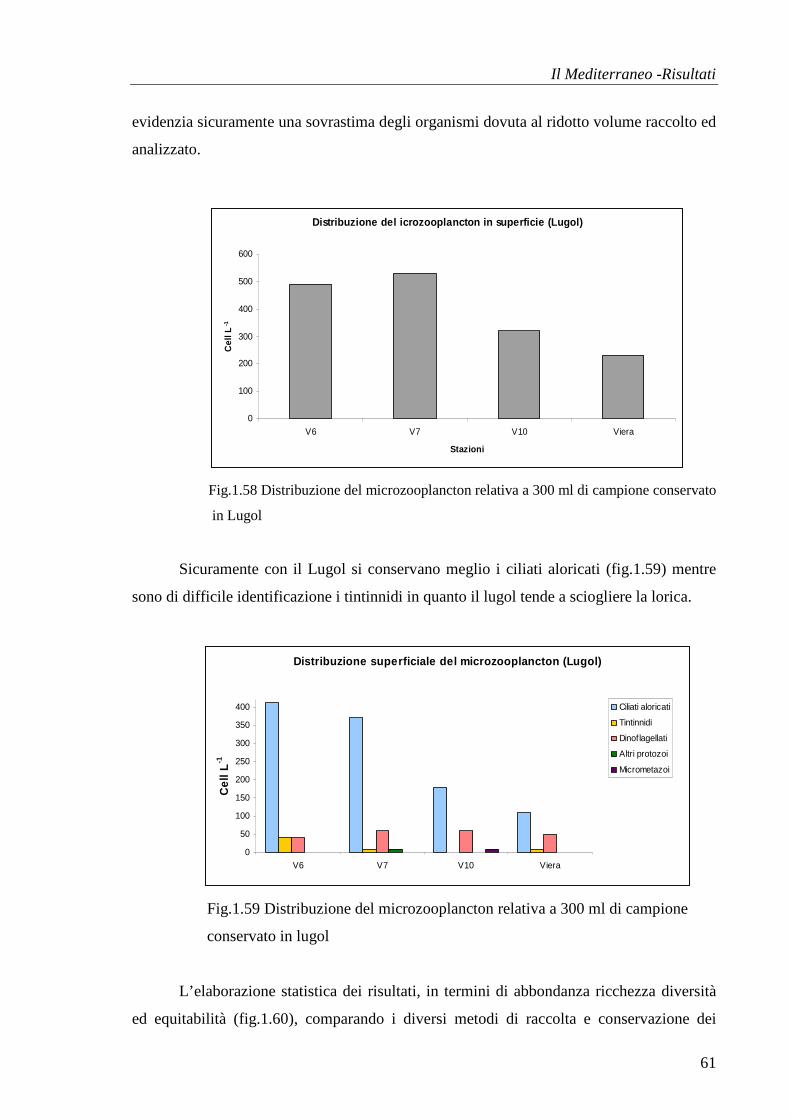
Il Mediterraneo -Risultati
61
evidenzia sicuramente una sovrastima degli organismi dovuta al ridotto volume raccolto ed
analizzato.
Distribuzione del icrozooplancton in superficie (Lu gol)
0
100
200
300
400
500
600
V6 V7 V10 Viera
Stazioni
Cel
l L-1
Fig.1.58 Distribuzione del microzooplancton relativa a 300 ml di campione conservato
in Lugol
Sicuramente con il Lugol si conservano meglio i ciliati aloricati (fig.1.59) mentre
sono di difficile identificazione i tintinnidi in quanto il lugol tende a sciogliere la lorica.
Distribuzione superficiale del microzooplancton (Lu gol)
0
50
100
150
200
250
300
350
400
V6 V7 V10 Viera
Cel
l L-1
Ciliati aloricati
Tintinnidi
Dinoflagellati
Altri protozoi
Micrometazoi
Fig.1.59 Distribuzione del microzooplancton relativa a 300 ml di campione
conservato in lugol
L’elaborazione statistica dei risultati, in termini di abbondanza ricchezza diversità
ed equitabilità (fig.1.60), comparando i diversi metodi di raccolta e conservazione dei

Il Mediterraneo -Risultati
62
campioni, ha confermato come i 2 litri in 3 repliche sono quelli che in termini di
abbondanza descrivono meglio il popolamento presente. I 5 litri concentrati sicuramente
sottostimano le abbondanze, mentre al contrario i 300ml conservati in lugol sovrastimano
il popolamento presente
0
1
2
3
4
5
6
7
1 replica 2L 2 replica 2L 3 replica 2 l 5 L lugol
0
50
100
150
200
250
300
350
ind. L1-
MARGALEF SHANNON BRILLOUIN J-pielou E Brillouin abbondanza
Fig.1.60 Abbondanza, ricchezza, diversità ed equitabilità del popolamento microzooplanctonico

Il Mediterraneo-Discussione e Conclusioni
63
1.5 DISCUSSIONE E CONCLUSIONI
L’area del Mediterraneo orientale dove è avvenuto il campionamento è una zona
caratterizzata da elevate oligotrofia estiva e fosforo limitazione (Thingstad &
Rassoulzadegan, 1995; Thingstad et al., 2005). Queste condizioni vanno sicuramente a
limitare la crescita batterica durante il periodo estivo (Dugdale & Wilkerson 1988;
Béthoux et al., 1992, Berland et al., 1980, Krom et al., 1991; Estrada et al., 1993;
Thingstad & Rassoulzadegan 1995, 1999). Le stesse condizioni ambientali sono state
riportate in letteratura nel bacino del Mediterraneo durante lo stesso periodo di
campionamento (Krom et al., 1991; Thingstad & Rassoulzadegan 1995; Dolan et al., 1999;
Crispi et al., 2001; Tanaka&e Rassoulzadegan, 2002). La marcata oligotrofia registrata nel
mio studio è presumibilmente dovuta al mite inverno precedente era con scarso
rimescolamento della colonna d’acqua, scarse precipitazioni e scarsi apporti di nutrienti di
origine antropica (Andersen et al., 2008). Il plancton di zone poco produttive è
caratterizzato solitamente da un’elevata biomassa eterotrofa spostando così il sistema verso
una più marcata eterotrofia dove prevale sicuramente la respirazione rispetto alla
fotosintesi (Gasol et al., 1997).
Durante questa campagna di campionamento invece dei classici approcci utilizzati
per valutare la predazione eterotrofa quali le variazioni della concentrazione della Chl a e
l’analisi dei pigmenti (Strom & Welschmeyer, 1991; McManus & Ederington-Cantrell,
1992; Verity et al., 1993; Waterhouse & Welschmeyer, 1995; Latasa et al., 1997;
Reckermann & Veldhuis, 1997; Kuiper & Witte, 1999; Stelfox-Widdicombe et al., 2000)
sono stati condotti direttamente al microscopio i conteggidel fitoplancton, del picoplancton
autotrofo ed eterotrofo e del nanoplancton, nonche’ dei predatori (microzooplancton).
Inoltre e’ stato possibile valutare la produzione secondaria ovvero la crescita dei predatori
dopo il periodo di incubazione. E’ stata anche analizzata la distribuzione del
microzooplancton nell’area del Mediterraneo orientale confrontandola con i dati ottenuti
dall’analisi di campioni conservati con tre metodi diversi, cercando di capire quale
modalità di conservazione è quella che meglio rappresenta la condizione presente in
natura.
L’analisi dei campioni preparati per verificare la predazione da parte del
microzooplancton sulla frazione picoplanctonica ha evidenziato valori di biomassa
batterica piuttosto costanti pari a 7.49 µgCL-1 partendo dalla stazione V6 con un leggero

Il Mediterraneo-Discussione e Conclusioni
64
decremento nella stazione più orientale Viera con un valore pari a 4.87 µgCL-1. Anche in
termini di abbondanza i valori rimangono piuttosto costanti ed in accordo con quelli trovati
in letteratura nel bacino del Mediterraneo (Ferrier Pagés & Rassoulzadegan 1991, 1994;
Tanaka & Rassoulzadegan 2002) con valori che vanno da 3.70x 108cell L-1 nella stazione
V6 a 2.50x108 cell L-1 nella stazione Viera (Siokou-Frangou et al., 2010).
I tassi di ingestione sono mediamente alti e si mantengono pressochè costanti in
tutte e quattro le stazioni campionate passando da valori ~ 9.5 µgCL-1 d-1 nelle stazioni V6
e V7, a valori pari a 13.06 µgCL-1 d-1 nella stazione più orientale V10. Si verifica dunque
predazione da parte del microzooplancton sul comparto picoplanctonico. Confrontando i
valori dei coefficienti di mortalità indotta da predazione (g) e di crescita (k) si evidenzia
che nelle stazioni V6, V7, V10, e Viera i valori di k sono sempre maggiori, quindi
nonostante ci sia predazione sul comparto picoplanctonico da parte del microzooplancton il
predatore non riesce a controllare la crescita della preda. Questo è in accordo con le
condizioni di spiccata oligotrofia nella quale riescono a ben svilupparsi i batteri che sono in
grado di ottimizzare al meglio le scarse risorse disponibili, non solo, ma sono anche capaci
di sviluppare strategie di difesa che permettono loro di sopravvivere alla predazione
(Pernthaler, 2005).
Le biomasse della frazione picoplanctonica autotrofa sono molto basse con valori <
a 0.5 µgCL-1 in tutte le stazioni. Non si verifica predazione sulla componente autotrofa in
nessuna della quattro stazioni del bacino orientale.
I risultati di questo esperimento sono stati messi a confronto con quelli derivati
dall’esperimento di predazione del solo nanoplancton sul comparto picoplanctonico,
condotto in superficie nelle stesse stazioni del Mediterraneo orientale sempre con lo stesso
protocollo (Landry & Hassett, 1982) in modo da capire meglio le dinamiche che si
stabiliscono nella rete trofica marina. La comprensione dei cosiddetti “black boxes” ci
permette di individuare le modalità di predazione presenti in ambienti particolarmente
oligotrofici come il Mediterraneo orientale. L’elaborazione dei dati ha permesso di
individuare 3 modelli:
- Modello 1: nella stazione V6 il tasso di ingestione dei soli nanoflagellati è pari a
20.56 µgCL-1 mentre in presenza del microzooplancton si dimezza a un valore pari a 9.52
µgCL-1. In questo caso il microzooplancton riduce la pressione di predazione della sola
frazione nano planctonica (fig.1.61)
- Modello 2: nelle stazioni V7 e V10 invece i tassi di ingestione relativi ai due
esperimenti hanno valori piuttosto simili: 10.35 µgCL-1e 9.59 µgCL-1 per il nanoplancton e

Il Mediterraneo-Discussione e Conclusioni
65
il microzooplancton rispettivamente. Si verifica inoltre predazione da parte del
microzooplancton sul comparto nanoplanctonico: si ha dunque una presunta mortalità
diretta sul comparto batterico (fig. 1,62).
- Modello 3: nella stazione Viera il nanoplancton non viene predato dal
microzooplancton, i due tassi di ingestione sul picoplancton si sommano (fig. 1.63).
St. V6
HNAN1.2 µgC L -1
Microzooplancton0.63 µgC L -1
Picoplancton eterotrofo7.48 µg C L -1
20.52 µgC L -1 g-1 9.52 µgC L -1 g -1
3.26 µgC L -1 g-1
Fig.1.62 Black box modello 2 con relative biomasse (µg C L-1) e tassi di ingestione (µg C L-1 g-1) di prede e predatori
Fig.1.61 Black box modello 1 con relative biomasse (µg C L-1) e tassi di ingestione (µg C L-1 g-1) di prede e predatori
St. V7 (V10)
HNAN2.04µgC L -1
Microzooplancton0.60 µgC L -1
Picoplancton eterotrofo5.89 µg C L -1
10.37 µgC L -1 g -1 9.59 µgC L -1 g-1
0.81 µgC L -1 g-1
Viera
HNAN1.4 µgC L -1
No grazing Microzooplancton0.63 µgC L -1
Picoplancton eterotrofo4.87 µgC L -1
8.79 µgC L -1 g-1 11.80 µgC L -1 g -1
Fig.1.63 Black box modello 3 con relative biomasse (µg C L-1) e tassi di ingestione (µg C L-1 g-1) di prede e predatori

Il Mediterraneo-Discussione e Conclusioni
66
Il nanoplancton è stato conteggiato suddividendolo in tre classi dimensionali (< 3
µm; 3-5 µm e >5µm), per verificare se esiste una predazione di tipo selettivo indirizzata
preferibilmente verso una taglia rispetto ad un’altra. In termini di biomassa i valori si
approssimano mediamente a 2µgCL-1, mentre in termini di abbondanza i valori sono
mediamente ~ 2.5 x 105 cell L-1 , in accordo con i valori trovati in letteratura nella stessa
area di campionamento (Tanaka & Rassoulzadegan, 2002; Ferrier-Pagès &
Rassoulzadegan, 1991; 1994). Si registra predazione da parte del microzooplancton sul
comparto nanoplanctonico in tre (V6, V7 e V10) delle quattro stazioni campionate. Nella
stazione V6 il microzooplancton preda su tutte e tre le frazioni dimensionali di HNF
conteggiate. Nella stazione V7 il microzooplancton preda unicamente gli organismi di
dimensioni < 3 µm e quelli fra 3-5 µm. Nella stazione V10 la predazione è rivolta verso gli
organismi presumibilmente più energetici di medie e grandi dimensioni. Dal confronto dei
valori assoluti dei coefficienti di crescita (k) e di mortalità (g) delle prede
nanoplanctoniche è possibile osservare come nelle stazioni V10, V6 per gli organismi di
taglia >5 µm e in V7 per quelli < 3 µm il valore di g>k evidenzi un efficace top down
control sulla crescita delle prede come già evidenziato da altri autori in ambienti
oligotrofici (Pernthaler, 2005).
Negli altri casi dove si verifica predazione k>g indica una crescita superiore alla
mortalità indotta da predazione: il microzooplancton non controlla la crescita dei NF.
Questa predazione selettiva indirizzata verso alcune classi dimensionali potrebbe derivare
dalla composizione della comunità microzooplanctonica. In linea generale in tutte le
stazioni del Mediterraneo orientale la comunità di predatori è dominata dai ciliati aloricati
e dai dinoflagellati eterotrofi di taglia medio piccola. I valori elevati dei tassi di ingestione
relativi al picoplancton pari ~11 µgCL-1 g-1 indicano una predazione indirizzata
prevalentemente verso il comparto dimensionalmente più piccolo.
Solamente nella stazione V10 il microzooplancton preda i NF di dimensioni
maggiori, mentre nella stazione più meridionale Viera non c’è alcuna predazione sul
nanoplancton.
Il microzooplancton ha due ruoli importanti nell’ecosistema marino: uno è quello di
intermediazione fra il picoplancton, il nanoplancton ed il mesozooplancton (Gifford, 1991;
Gifford et al., 1995) e l’altro è quello di essere un importante agente per la
rimineralizzazione del particolato organico a livello superficiale (Ducklow et al., 1986;). In
questo studio l’abbondanza (cell L-1) e la biomassa (µgCL-1) hanno un andamento piuttosto
omogeneo in tutte e 4 le stazioni campionate con valori ~

Il Mediterraneo-Discussione e Conclusioni
67
1.40x102 cell L-1 e ~0.60 µgCL-1. La composizione del popolamento di predatori è
costituita per la gran parte da ciliati aloricati seguiti dai dinoflagellati eterotrofi (Lessard e
Murrel, 1996). Nonostante il numero limitato di specie relative ai ciliati aloricati, questi
rappresentano i predatori dominanti del comparto microplanctonico (Dolan et al., 1999,
2002; Dolan, 2000; Pérez et al., 2000; Pitta et al., 2001; Modigh & Castaldo, 2002). I
tintinnidi, nell’area di studio rappresentano una frazione piuttosto limitata anche se sono
quelli che riescono a predare organismi di dimensioni fino alla metà del diametro della
apertura orale (Spittler, 1973; Heinbokel, 1978, b; Rassoluzadegan, 1978; Rassoulzadegan
& Etienne, 1981; Capriulo, 1982). I tintinnidi giocano un ruolo minore nei flussi relativi
alla rigenerazione dei nutrienti e la loro attività di predazione e di escrezione è sicuramente
meno importante dal punto di vista quantitativo rispetto ai ciliati aloricati. Le abbondanze
dei ciliati aloricati seguono un andamento decrescente dal Bacino del Mediterraneo
occidentale in cui troviamo valori ~ 3x103 cell L-1 fino ad arrivare a valori ~5.0 x 102 cell
L-1 nel Bacino del Mediterraneo orientale (Dolan et al., 2002; Di Poi, 2010).
I valori di abbondanza relativi ai tintinnidi e dei ciliati aloricati, oltre a seguire un
andamento decrescente, hanno valori mediamente inferiori rispetto a quelli trovati da
Dolan et al., (2002) derivanti da campionamenti effettuati nella stessa zona nel 1999.
I ciliati sono in grado di modulare le risorse di carbonio e sono caratterizzati da un
elevato tasso di crescita (Fenchel, 1990) e sono in grado utilizzare positivamente eventuali
surplus di risorse che si rendono disponibili durante i periodi di fioriture algali.
D’altra parte, quando le risorse energetiche sono scarse i ciliati mixotrofi possono
sia fotosintetizzare che fagocitare il cibo disponibile. L’importanza dei ciliati è
particolarmente rilevante negli ambienti oligotrofici dove la scarsa disponibilità di nutrienti
impone la necessità di disporre di modalità di assunzione di cibo diverse (Pitta &
Giannakourou, 2000). Nonostante la loro importanza, si hanno ancora poche informazioni
relative al ruolo dei ciliati nei sistemi oligotrofici (Strom et al., 1993; Stoeker et al., 1996).
I dinoflagellati eterotrofi, non hanno un andamento decrescente spostandosi verso
est, ma mantengono valori medi di abbondanza prossimi a 40 cell L-1 in tutte le stazioni
studiate.
Nella stazione V6, la più occidentale di quelle campionate i ciliati aloricati sono
presenti per il 66% contro il 7% dei tintinnidi. Nelle stazioni V7, V10 e Viera la
percentuale si approssima attorno al 55% mentre quella dei tintinnidi oscilla intorno
all’8%, in accordo con uno studio condotto nel Mediterraneo orientale nel 1997 (Pitta &
Giannakourou, 2000). Da uno studio condotto da Pitta et al. (2001), nel bacino del

Il Mediterraneo-Discussione e Conclusioni
68
Mediterraneo è emerso come in realtà circa l’88% dei ciliati aloricati di dimensioni < 30
µm contenesse nello stomaco prede <3µm e solamente l’8% dei piccoli ciliati contenesse
prede >3µm ed il 4% contenesse nello stomaco prede di entrambe le dimensioni. Anche nei
tintinnidi i risultati di quello studio evidenziano una predazione selettiva indirizzata
principalmente (80%) verso prede di piccole dimensioni.
Tra i ciliati sono presenti anche forme mixotrofe, quali Strombidium conicum e
Tontonia sp. che sono le forme che meglio riescono a sopravvivere in periodi di scarsa
disponibilità di cibo. Scarsa l’abbondanza dei tintinnidi pari a 4 specie identificate:
Dadayella ganymedes, Epiplocylis blanda, Eutintinnus tubulosus, Rhabdonella spiralis.
Nella stazione V7 sono state identificate 4 specie: Codonella sp., Eutintinnus
macilentus, E. tubulosus, Eutintinnus sp. Rhabdonella spiralis, Tintinnopsis sp.
Nella stazione V10, quella più orientale sono state identificate 6 specie: Codonella
sp., Dadayella ganymedes, Eutintinnus apertus, E. macilentus, E. tubulosus, Rhabdonella
spiralis, Tintinnopsis nana, Tintinnopsis sp.
Nella stazione più meridionale Viera, caratterizzata da una dinamica di correnti
tale da favorire il fenomeno di downwelling con il conseguente incremento dell’oligotrofia
già presente in quest’area, sono state identificate 4 specie: Craterella torulata, Epiplocys
blanda, Eutintinnus macilentus, E. tubulosus, Rhabdonella spiralis.
Confrontando la diversità ritrovata in questo lavoro con studi precedenti (Dolan et
al., 1999; Pitta et al., 2001; Pierce & Turner, 1993), emerge che la diversità dei tintinnidi è
piuttosto ridotta. Allo stesso tempo però l’ambiente è popolato per la maggior parte dai
ciliati non loricati che meglio ottimizzano le risorse pico e nanoplanctoniche disponibili
(Kivi e Setälä, 1995; Rassoulzadegan et al., 1988).
L’analisi dei campioni contenenti il microzooplancton dopo il periodo di
incubazione ha permesso di verificare la crescita degli organismi dopo il periodo di
incubazione (T48). Nella stazione V6 la predazione è indirizzata verso il comparto batterico
(I=9.51 µg C L-1 g-1 ); c’è anche una predazione, anche se in misura minore rivolta verso il
comparto nanoplanctonico. L’analisi dei campioni dopo il periodo di incubazione ha
permesso di evidenziare un incremento del popolamento dei dinoflagellati eterotrofi ed in
particolar modo di Protoperidinium steini, Gyrodinium fusiforme e Gyrodinium sp. Questo
incremento può essere spiegato considerando che le specie di dinoflagellati che crescono
sono di piccole dimensioni e quindi favorite dall’abbondanza della comunità microbica.
Anche i tintinnidi crescono in termini di biomassa anche se in realtà il loro incremento è
piuttosto ridotto poiché passano da 0.044 µgCL-1 al T0 a 0.065 µgCL-1 dopo l’incubazione.

Il Mediterraneo-Discussione e Conclusioni
69
Anche nella stazione V7 gli elevati tassi di ingestione calcolati (I=9.59 µg C L-1)
indicano predazione sui batteri. In misura minore vengono predati anche i nanoflagellati di
piccole e medie dimensioni (<3µm e 3-5µm). L’analisi dei campioni dopo il periodo di
incubazione indica una crescita dei soli dinoflagellati eterotrofi e in particolar modo
crescono come nella stazione V6 Protoperidinium steini, Gyrodinium fusiforme e
Gyrodinium sp.
Nella stazione V10 la predazione è indirizzata verso il comparto batterico (I=13.06
µg C L-1). Dopo l’incubazione crescono i dinoflagellati eterotrofi con un incremento pari a
0.05 µg C L-1. In questa stazione ed in Viera crescono anche i radiolari, con un incremento
dell’86% nella stazione V10 e del 59% nella stazione Viera.
Per individuare il metodo che meglio descrive la composizione del popolamento
microzooplanctonico presente nell’area di campionamento, i campioni raccolti per l’analisi
del microzooplancton sono stati conservati in tre modi diversi:
- 2 litri di acqua raccolta in 3 repliche e fissata con formalina al 2%,
prefilatrate su 200 µm;
- 5 litri di acqua fissata in formalina al 2% e concentrati con
filtrazione inversa a 250ml;
- 300 ml di acqua conservata in Lugol al 2%.
L’analisi dei campioni corrispondenti alle tre repliche iniziali dell’esperimento
di diluizione evidenzia una sostanziale omogeneità, verificata dai bassi valori di SD
riscontrati.
I campioni in 5 litri concentrati sicuramente descrivono meglio il popolamento dei
tintinnidi. Il Lugol serve per conservare meglio i ciliati non loricati; tende infatti a
sciogliere la lorica dei tintinnidi ed a provocare maggiori distorsioni nella forma degli
organismi oltre che a provocare a concentrazioni più elevate la formazione di
precipitati nei campioni che ne impediscono la corretta analisi (Stoecker et al., 1996;
Modigh & Castaldo, 2005). Il Lugol è stato utilizzato alla concentrazione del 2% che è
quella consigliat per i ciliati aloricate (Vaquè, 1997; Kuipers & Witte, 1999; Archer et
al., 2001; Bulit et al. 2003; Setälä e Kivi, 2003; Johansson et al., 2004)). La formalina
dall’altra parte provoca minori alterazioni della forma cellulare. In ogni caso ogni
modalità di fissazione presenta vantaggi e svantaggi; i 2 litri in tre repliche
rappresentano sicuramente il miglior compromesso per descrivere in modo attendibile
la comunità presente. I campioni di 5 litri sicuramente tendono ad amplificare i risultati

Il Mediterraneo-Discussione e Conclusioni
70
in termini di ricchezza mentre vanno a sottostimare l’abbondanza visti i volumi
analizzati.
I 300 ml fissati in Lugol invece permettono di contare meglio i piccoli ciliati,
danneggiando però la lorica dei tintinnidi; visto il ridotto volume analizzato questo ha
sicuramente portato a una sovrastima degli organismi in termini di abbondanza.
Per concludere, la marcata oligotrofia del Mediterraneo orientale
particolarmente accentuata durante il periodo di campionamento (maggio-giugno
2007), in aggiunta alle particolari caratteristiche idrologiche registrate prima dell’estate
quali: alte temperature; scarso rimescolamento della colonna d’acqua; scarse
precipitazioni e ridotti apporti antropici di nutrienti hanno creato un ambiente così
caratterizzato:
- Domina il popolamento batterico in termini di abbondanza e biomassa, grazie al
loro rapporto superficie - volume riescono ad ottimizzare l’assimilazione dei pochi
nutrienti presenti e la predazione è maggiormente indirizzata verso questo
comparto. In ogni caso in tutte le stazioni k>g e pertanto il microzooplancton non è
in grado di controllare la crescita batterica;
- Il picoplancton autotrofo è molto scarso (~0.1 µgCL-1) e non viene predato
- I tassi di ingestione relativi alla predazione sul nanoplancton sono piuttosto bassi e
indirizzati solamente verso alcune classi dimensionali. Il nanoplancton dunque,
insieme al picoplancton rappresentano la principale fonte di carbonio per il
microzooplancton in questo ambiente oligotrofico, c il quale esercita anche una
predazione selettiva;
- Non c’è predazione nei confronti del microfitoplancton presente con abbondanze
molto basse;
- L’unione dei miei risultati con quelli derivanti dall’esperimento di predazione del
solo nanoplancton sul picoplancton ha evidenziato 3 modelli di predazione presenti
nel Mediterraneo orientale:
1. dall’analisi dei valori dei tassi di ingestione emerge che quello ottenuto
dall’esperimento di predazione del solo nanoplancton è il doppio di quello ottenuto
dall’esperimento in cui c’è anche il microzooplancton; dunque il microzooplancton
riduce la pressione di predazione della frazione nanoplanctonica (V6)

Il Mediterraneo-Discussione e Conclusioni
71
2. i valori dei tassi di ingestione sono piuttosto simili; il microzooplancton preda
anche su HNAN, si suppone quindi una predazione diretta sul comparto batterico
(V7, V10)
3. il microzooplancton e HNAN predano in maniera indipendente; i tassi di
ingestione si sommano (Viera).
La comparazione di tre metodi di raccolta e conservazione dell’acqua di mare per
studiare la composizione del popolamento microzooplanctonico ha evidenziato che:
- a. i 2 litri in tre repliche sono quelli che meglio descrivono la comunità
microzooplanctonica nel bacino del Mediterraneo;
- b. i 5 litri conservati in formalina e concentrati a 250 ml sono quelli che
descrivono meglio la comunità in termini di ricchezza ma sicuramente vanno a
sottostimare l’abbondanza;
- c. i 300 ml conservati in Lugol, permettono una migliore identificazione dei
ciliati non loricati, causando però una perdita dei tintinnidi ed in termini di
abbondanza sovrastimano il popolamento.
Nel Mediterraneo orientale, oggetto di questo studio, si evidenzia la marcata
presenza del “microbial loop” riferito alle interazioni trofiche tra pico-nano e
microzooplancton. La maggior parte del carbonio in mare si trova in fase disciolta (DOC) e
deriva da processi di predazione, lisi cellulare, essudazione algale. Su questo substrato
agiscono i batteri trasformandolo in propria biomassa la quale viene a sua volta predata dal
nanoplancton eterotrofo che a sua volta viene predato dal microzooplancton. Con il
microbial loop coesiste la catena trofica classica che trasferisce l’energia agli organismi di
taglia superiore (copepodi, salpe ecc.) formando così la rete trofica mistivora.
Questo lavoro di tesi ha evidenziato l’elevata biomassa eterotrofa di piccola taglia
verso la quale è indirizzata la predazione. Nel Mediterraneo orientale prevale sicuramente
il “microbial loop” in cui l’energia ricavata dal DOC può venir restituita al substrato,
respirata immediatamente o trasformata in fecal pellets piccole e galleggianti con rilascio
di CO2 in atmosfera.
Questi risultati rappresentano un contributo significativo per la comprensione dei
flussi di carbonio nell’ecosistema marino del Mediterraneo orientale, che era uno degli
obiettivi della campagna “Transmediterranean cruise” all’interno del progetto
V.E.C.T.O.R.

Ostreopsis-Introduzione
72
2.OSTREOPSIS OVATA
IDENTIFICAZIONE DEI TRASFERIMENTI TROFICI DI OSTREO PSIS LUNGO LA RETE TROFICA PELAGICA.
2.1 INTRODUZIONE
Le fioriture algali sono massicce proliferazioni di alghe unicellulari che si
verificano in natura per azione di particolari condizioni ambientali (Sellner et al. 2003)
quali l’aumento di nutrienti in seguito ad afflussi fluviali in mare, incremento della
temperatura superficiale, stratificazione della colonna d’acqua, assenza di vento con basso
idrodinamismo, luminosità prolungata e fenomeni di downwelling (Brescianini et al.,
2006). L’impatto sull’ecosistema delle fioriture è grave: si manifestano alterazioni della
qualità e del colore dell’acqua, ipossia e/o anossia dei fondi con morie di invertebrati
bentonici come mollusci, celenterati ed echinodermi
Alcune specie di microalghe sono in grado di generare fioriture dannose per
l’ecosistema e per l’uomo e contribuiscono a causare il fenomeno denominato Harmful
Algal Blooms (HABs o fioriture algali pericolose)
Tra le specie introdotte recentemente nel Mediterraneo si annovera Ostreopsis
ovata, una microalga bentonica unicellulare (fig.2.1) descritta per la prima volta da Fukuyo
(1981) e successivamente da Besada et al., (1982) e Faust et al., (1996) appartenente alla
classe delle Dinophyceae. Al microscopio le cellule di Ostreopsis ovata presentano una
forma a goccia compressa in senso antero posteriore; le cellule sono ricoperte da placche di
cellulosa che formano due teche (epiteca e ipoteca) di uguali dimensioni che presentano
numerosi pori distribuiti casualmente sulla superficie. Le cellule presentano molti
cloroplasti e uno o due corpi rossi sul lato dorsale che probabilmente risultano essere il
residuo di cisti temporanee come accade in altri dinoflagellati (Aligizaki & Nikolaidis,
2006).

Ostreopsis-Introduzione
73
Fig.2.1 Ostreopsis ovata (A) cellula singola (B) cellule viste
al microscopio invertito (Cminiello, 2006)
Ostreopsis è un’alga epifita che vive preferibilmente sulle alghe rosse e brune e sugli
invertebrati bentonici (Ballatine et al., 1988; Faust et al., 1996; Fukuyo, 1981; Totti et al.,
2007; Besada et al., 1982; Chang et al., 2000; Hurbungs et al., 2001; Rhodes, 2010).
O. ovata ha una vasta distribuzione a livello globale specialmente nelle aree tropicali e
subtropicali nell’Oceano Pacifico, Atlantico e Indiano (Fukuyo, 1981; Besada et al., 1982;
Chang et al., 2000; Hurbungs et al., 2001).
Il primo rilevamento in Mediterraneo risale agli anni ’70 lungo la costa di
Villafranche-sur-Mer in Francia (Taylor, 1979; Guerrini et al., 2010) e la sua presenza
risulta essere molto ben documentata sebbene non fosse ancora collegata con gli
avvelenamenti da ciguatera (Aligizaki & Nikolaidis, 2006). Condizioni climatiche ottimali
hanno consentito a quest’alga di svilupparsi alle nostre latitudini. La prima segnalazione di
Ostreopsis lungo le coste italiane risale al 1994 quando venne osservata lungo la costa
tirrenica della regione Lazio (Tognetto et al., 1995). Monti et al., (2007) registrarono per la
prima volta la presenza di O. ovata nel Golfo di Trieste (Alto Adriatico) nel 2006. Fin dalla
fine degli anno ’90 sono state registrate massicce fioriture di Ostreopsis nel Mar Tirreno,
nel Mar Ligure e in Adriatico (Sansoni et al., 2003; Simoni et al., 2004; Gallitelli et al.,
2005; Totti et al., 2007; Mangialajo et al., 2008).
Questa diffusione di Ostreopsis nelle zone temperate è dovuta principalmente allo
sversamento delle acque di zavorra dalle navi cargo e da condizioni ambientali
particolarmente favorevoli quali acque poco profonde, riparate e con basso idrodinamismo
(Vila et al., 2001); temperatura intorno ai 20-25°C; mare calmo e disponibilità di substrato
(Simoni et al., 2004; Aligizaki & Nikolaidis 2006; Shears & Ross, 2009), periodi di alta
pressione atmosferica associati a condizione di mare calmo (Mangialajo et al., 2008).
Le fioriture si presentano come schiume superficiali (foaming) formate da migliaia di
cellule ricoperte da un substrato biotico o abiotico (fig.2.2). Sott’acqua si forma una

Ostreopsis-Introduzione
74
pellicola bruna che avvolge gli scogli e tutto ciò che si trova sul fondo. Questi aggregati
poi tendono a ritornare in sospensione grazie all’azione meccanica delle onde (Congestri
et al., 2006; Totti et al., 2007). Le fioriture si verificano durante i periodi caldi con picchi
in luglio e agosto (Simoni et al., 2004; Aligizaki & Nikolaidis 2006; Mangialajo et al.,
2008). Durante il periodo estivo l’incremento della densità algale da zero ad un valore
massimo è estremamente veloce mentre la riduzione della densità algale e la scomparsa di
O. ovata in autunno-inverno è piuttosto graduale. Questo andamento dimostra la loro
capacità di produrre forme di resistenza difficilmente individuabili nell’ambiente naturale
che si attivano quando la temperatura supera i 20-25°C (Simoni et al., 2004).
Fig.2. 2 Ostreopsis durante una fioritura: materiale gelatinoso in sospensione con
presenza di schiume superficiali
Ostreopsis è considerata una specie tossica, vista la produzione di una tossina
simile alla palitossina che provoca effetti dannosi anche a basse concentrazioni (Ciminiello
et al., 2006; 2008; Ramos & Vasconcelos 2010 Wang, 2008). Infatti durante la fase di
fioritura molte cellule si rompono rilasciando in mare elevate concentrazioni di tossina che
viene trasportata in aria attraverso l’areosol marino (Zingone, 2008; Guerrini et al., 2010).
La palitossina (PTX) è una potente tossina marina termostabile con una lunga
catena di atomi di carbonio (>100) che presenta simultaneamente regioni lipofile e idrofile
(Moore & Bartolini, 1981). In particolare, Ciminiello et al., (2008) analizzando campioni
di acqua di mare prelevati nella zona di Genova nel 2005 e 2006 dimostrarono che una
ovatossina-a prodotta da O. ovata è strettamente correlata alla palitossina poichè presenta
solamente 2 atomi di ossigeno in meno rispetto a quest’ultima che è una delle più potenti
tossine marine non proteiche conosciute (Ukena et al., 2001; Lenoir et al., 2004; Penna et
al., 2005; Guerrini et al., 2010).

Ostreopsis-Introduzione
75
Le fioriture di O. ovata provocano conseguenze gravi sugli ecosistemi e sulla salute
umana. In particolare si evidenziano segnali di sofferenza da parte di alcuni organismi
marini quali ricci e stelle di mare che perdono aculei e braccia (Ramos e Vasconcelos
2010) (fig.2.3).
Fig.2.3 Paracentrotus lividus e Coscinasterias tenuispina senza spine e senza braccia rispettivamente (Sansoni et al., 2003)
Nelle zone tropicali l’avvelenamento da palitossina, provocato dall’ingestione di
pesce contaminato provoca disturbi neurologici e gastrointestinali (Alcala et al., 1988;
Yasumoto, 1998; Onuma et al., 1999; Granéli et al., 2002). Casi di avvelenamento di
questo tipo, caratterizzati da un alto tasso di mortalità sono stati registrati nelle Filippine,
in Madagascar, in Giappone e alle Hawaii (Ramos & Vasconcelos, 2010).
Tali intossicazioni sono dovute al bioaccumulo delle tossine all’interno della rete
trofica: le microalghe vengono accumulate da organismi erbivori quali zooplancton, pesci
erbivori, molluschi filtratori e raschiatori, i quali possono giungere all’uomo tramite il loro
consumo diretto o attraverso successivi trasferimenti a livelli trofici superiori (Anderson &
White, 1992; Geraci et al., 1989; Mebs, 1998).
Fino ad ora in Italia non si è registrato alcun caso di mortalità conseguente
all’ingestione di pesce contaminato: si sono invece registrati, a seguito delle fioriture,
disturbi alle vie aeree, tosse, rinorrea, nausea e congiuntiviti nelle persone esposte
all’inalazione delle tossine prodotte dall’alga o da frammenti di Ostreopsis presenti
nell’aerosol marino (Brescianini et al., 2006; Ciminello et al., 2006; Durando et al., 2007).

Ostreopsis-Scopo del lavoro
76
2.2 SCOPO DEL LAVORO
Nell’ambito del Progetto di Ricerca “Ostreopsis ovata e Ostreopsis spp. nuovi
rischi di tossicità microalgale nei mari italiani” gli obiettivi della linea di ricerca relativa al
“Trasferimento dell’alga e/o delle tossine nella rete trofica” sono stati quelli di identificare
gli eventuali trasferimenti trofici di Ostreopsis lungo la rete trofica pelagica. Il
monitoraggio infatti delle acque costiere per verificare la presenza di alghe pericolose
potrebbe essere essenziale per accertare la potenziale formazione di fioriture. Per questo
tipo di monitoraggio si ricorre all’esame del plancton tramite microscopio ottico o ad
epifluorescenza (Sournia, 1978; Sellner et al., 2003), metodi questi che richiedono molto
tempo ed un elevato grado di esperienza tassonomica. Per questo motivo si stanno
mettendo a punto saggi basati su metodi molecolari (Scholin et al., 1994; Litaker &Tester,
2002; Popels et al., 2003) che consentono di individuare la presenza di minime quantità di
microalga su un elevato numero di campioni.
Per il nostro studio è stata allestita nel mese di giugno del 2009 una serie di
esperimenti di diluizione e di grazing con acqua di mare raccolta al largo della stazione
biologica di Pirano (Slo), arricchita con coltura di Ostreopsis per verificare l’eventuale
predazione da parte del microzooplancton e del mesozooplancton nei confronti dell’alga in
questione.
Un secondo esperimento di solo grazing è stato condotto nel mese di settembre
2009, campionando acqua di mare raccolta al largo della stazione biologica di Pirano (Slo).
Il microzooplancton è costituito da tutti gli organismi eterotrofi compresi tra i 10 -
20 e i 200 µm. E’ una delle componenti delle comunità microbiche ed è importante in
quanto riesce a predare organismi le cui dimensioni sono inferiori a quelle delle prede
tipiche del mesozooplancton. Molteplici studi hanno dimostrato che ad esso va attribuito
una notevole parte della predazione sul fitoplancton (Verity, 1986; Pierce & Turner, 1992,
Calbet & Saitz, 2005). Il microzooplancton costituisce inoltre il principale veicolo di
trasferimento energetico dalla rete microbica a quella del pascolo in quanto a sua volta
viene attivamente predato dal mesozooplancton (copepodi e larve) (Sherr et al., 1986;
Stoeker & Capuzzo, 1990; Gifford & Dag, 1990; Turner & Granéli, 1992). Nel mese di
aprile del 2010 si è allestito un esperimento nel quale organismi mesozooplanctonici sono
stati messi ad incubare in coltura pura di Ostreopsis allo scopo di verificare se in queste
condizioni estreme essi predino l’alga. Si è poi messo a punto un protocollo molecolare in
grado di identificare la presenza dell’alga all’interno dei copepodi.

Ostreopsis- Materiali e metodi
77
2.3 MATERIALI E METODI
Per valutare l’efficienza della predazione del microzooplancton sulla componente
microfitoplanctonica ed in particolare su Ostreopsis ovata si è utilizzato il metodo delle
diluizioni proposto da Landry e Hasset (1982) successivamente modificato da Landry et
al., (1995) e Gallegos (1989), che a differenza di altri metodi proposti (Heinbokel & Beers,
1979; Capriolo & Carpenter, 1980; Waterhouse & Welschemeyer, 1995) è estremamente
semplice e non prevede alcuna manipolazione degli organismi e permette di ottenere sia il
tasso specifico di crescita delle prede (microfitoplancton, nanoplancton, picoplancton) sia
quello specifico di predazione degli organismi eterotrofi. Il metodo delle diluizioni è stato
già esposto nel cap.1.3.3.
2.3.1 CAMPIONAMENTO a) Esperimento di diluizione
Nel mese di giugno del 2009 a Pirano, Slovenia, presso il laboratorio del NIB, sono
stati effettuati prelievi d’acqua di superficie in una stazione al largo nel Golfo di
Capodistria (Nord Adriatico) (fig.2.4).
Fig. 2.4 Area di campionamento (Alto Adriatico); Laboratorio NIB di Pirano (SLO)
L’acqua raccolta è stata filtrata su retino con vuoto di maglia pari a 200 µm per
eliminare tutti gli organismi di dimensioni maggiori alla frazione microplanctonica. Una
parte di quest’acqua prefiltrata è stata ulteriormente filtrata con pompa peristaltica su
membrana idrofila PFTE Millipore con porosità 0.22 µm.
L’acqua filtrata su 200 µm è stata diluita con quella filtrata su 0.22 µm allestendo il
seguente set di diluizioni seguendo questo schema:

Ostreopsis- Materiali e metodi
78
• 100% - solo acqua filtrata su 200µm
• 80% - di acqua filtrata su 200µm
• 50% - di acqua filtrata su 200 µm
• 10% - di acqua filtrata su 200 µm
Per ciascuna diluizione si sono allestite 3 repliche da 2 litri, poste in bottiglie da
incubazione in Nalgene. I campioni sono stati arricchiti con coltura di Ostreopsis ad una
concentrazione pari a 0.5 x 106 cell. L-1. Sono stati aggiunti 10 mL per litro di coltura al
100%; 8 mL per litro all’80%, 5 mL per litro al 50% e 1 mL per litro al 10%. I campioni
sono stati messi poi ad incubare in mare per 24 ore.
I campioni iniziali (T0) per ciascuna diluizione, sempre in 3 repliche, sono stati posti in
bottiglie di plastica da 2 litri, arricchiti con coltura di Ostreopsis con le stesse modalità dei
campioni precedenti e sono stati immediatamente fissati con formalina neutralizzata alla
concentrazione finale del 2%. Allo stesso modo sono stati fissati i campioni alla fine
dell’incubazione (T24).
b) Esperimento di grazing (giugno 2009)
In concomitanza con l’esperimento di diluizione è stato effettuato anche un
esperimento di grazing per valutare la predazione da parte del mesozooplancton nei
confronti di Ostreopsis.
Allo scopo si è raccolto il mesozooplancton con una rete WP da 200 µm di vuoto di
maglia mediante un campionamento obliquo al largo della stazione di Pirano. Gli
organismi vivi sono stati messi in un contenitore raffreddato da 5 L ed il campione è stato
immediatamente portato in laboratorio e sistemato in camera fredda per il mantenimento
degli organismi vitali. Si sono quindi selezionati gli organismi maggiormente
rappresentativi della comunità mesozooplanctonica al momento del prelievo a mare. Per
verificare la predazione del mesozooplancton nei confronti di O.ovata sono state allestite 3
bottiglie di Nalgene arricchite con 20ml di coltura di Ostreopsis a concentrazione nota
(500*103 cell L-1) nelle quali sono stati aggiunti circa 20 organismi planctonici vitali
rappresentativi della comunità naturale nel periodo di campionamento (Acartia clausi) in
stadi prevalentemente giovanili. Le bottiglie sono state messe ad incubare per 24 ore in
mare. 3 bottiglie di acqua tal quale (filtrata su 200 µm), arricchite allo stesso modo con
coltura di Ostreopsis sono state messe ad incubare alle stesse condizioni (bianco). Altre 3

Ostreopsis- Materiali e metodi
79
bottiglie della stessa acqua arricchita con Ostreopsis sono state immediatamente fissate
con formalina al 2% (T0). Alla fine dell’incubazione (T24) tutti i campioni sono stati fissati
come sopra.
c) Esperimento di grazing (settembre 2009)
Nel mese di settembre 2009 si è allestito un esperimento di grazing con acqua di
mare prelevata al largo della stazione biologica di Pirano (Slo).
L’acqua di mare raccolta in superficie è stata filtrata su retino con maglia di 200µm allo
scopo di eliminare tutti gli organismi di taglia superiore. Il mesozooplancton è stato
raccolto con rete WP2 da 200µm effettuando un campionamento obliquo. Gli organismi
vivi così raccolti sono stati immediatamente trasferiti in un contenitore freddo e portati in
laboratorio per la selezione.
L’esperimento è stato allestito come quello condotto nel mese di giugno. Il
popolamento di organismi planctonici vitali presenti nel mese di settembre era costituito
da Centropages.
d) Copepodi
Nel mese di maggio 2010, è stato condotto un esperimento nel quale si sono messi
un certo numero di organismi mesozooplanctonici, raccolti nel Golfo di Trieste con un
retino a maglia 200µm, in presenza di coltura di Ostreopsis pura. Le specie rappresentative
della comunità sono state selezionate allo stereomicroscopio Leica MZ6 a 40X. Sono state
allestite 12 vaschette ognuna contenente 50 ml di coltura di O.ovata (240 cell mL-1), alle
quali sono stati aggiunti i copepodi selezionati (Tab.1). I campioni sono stati messi ad
incubare mantenendoli in moderata agitazione per 3 ore. Alla fine dell’incubazione i
copepodi sono stati recuperati, lavati e trasferiti in minifalcon e conservati a -80° fino al
momento dell’analisi genetica.

Ostreopsis- Materiali e metodi
80
Campioni Copepodi Cop_1 Copepoditi (3) Cop_2 Centropages sp. (3)
Cop_3 Centropages sp. (3)
Cop_4 Acartia sp. (3)
Cop_5 Acartia sp. (4) + Centropages sp. (1)
Cop_6 Acartia sp (1) + Centropages sp. (1)
Cop_7 Acartia sp. (1) + Centropages sp. (1) + copepoditi (1)
Cop_8 Centropages sp. (1) + Oithona sp. (1) + copepoditi (1)
Cop_9 Centropages sp. (1) + copepoditi(2)
Cop_10 Acartia sp. (1) + Centropages sp. (1) + copepoditi (1)
Cop_11 Acartia sp. (1) + copepoditi (1)
Cop_12 Acartia sp. (1) + Corycaeus sp. (1) + copepoditi (1)
Tab.2.1 Specie di copepodi selezionate per valutare la predazione del mesozooplancton nei
confronti di O.ovata
2.3.2 ANALISI QUALI QUANTITATIVA
L’analisi quali-quantitativa dei popolamenti fitoplanctonici è stata condotta su
aliquote di 50 mL di subcampione preventivamente preconcentrato per 48 ore, per
garantire che tutti gli organismi possano sedimentare sul fondo delle bottiglie da dove per
aspirazione si elima il surnatante. Una frazione (100 ml) del campione così ridotto a 250
mL è stato messo a sedimentare in colonnina posta su cameretta di sedimentazione usando
il classico metodo Utermöhl (1958). Il tempo di sedimentazione per ogni cameretta è stato
di 30 ore. Il campione è stato conteggiato al microscopio rovesciato Olympus a 200x. Per il
calcolo del contenuto di carbonio del popolamento fitoplanctonico sono state effettuate le
misure lineari su almeno 100 organismi di ciascuna specie presente, quindi si sono
calcolati i relativi biovolumi, approssimando gli organismi a diverse forme geometriche
(Sieburth et al., 1978) ed infine si sono applicate le formule di trasformazione proposte da
Edler (1979).
I campioni di microzooplancton sono stati pre-concentrati allo stesso modo e quindi
analizzati con il metodo Utermöhl (1958). I campioni sono stati conteggiati con un
microscopio invertito Leitz Labovert con ingrandimento a 320X. La biomassa è stata
calcolata a partire dal biovolume mediante misurazione delle dimensioni lineari delle

Ostreopsis- Materiali e metodi
81
cellule che sono state associate a figure geometriche standard in accordo con Edler (1979)
e Hillebrand et. al. (1999).
I volumi risultanti sono stati trasformati in valori di carbonio organico usando
fattori di conversione presenti in letteratura (Edler, 1979; Strathmann, 1967; Putt &
Stoeker, 1989; Verity & Langdon, 1984; Beers & Stewart, 1970).

Ostreopsis- Risultati
82
2.4 RISULTATI 2.4.1 Caratterizzazione del popolamento microzooplanctonico (giugno 2009)
L’analisi del popolamento microzooplanctonico presente nel corso dell’esperimento
ha permesso di verificare anche l’eventuale crescita dello stesso dopo il periodo di
incubazione(tab. 2.2)
Microzooplancton T0
pgCL-1 x 105 T24
pgCL-1 x 105 T24 copepodi pgCL-1 x 105
Ciliati non loricati 1.57 0.92 1.9 Ciliati loricati 1.80 3.57 1.70 Dinoflagellati 7.43 5.97 8.96 Altri Protozoi 0.34 1.03 0.99 Micrometazoi 24.1 22.4 26.4
Tab.2.2Biomassa del microzooplancton al T0, T24 e T24 con mesozooplancton
Il popolamento iniziale è dominato da larve di micrometazoi e nauplii di copepodi
con una biomassa pari a 24.1x105 pgCL-1, da dinoflagellati eterotrofi (7.43x105 pgCL-1), da
ciliati loricati (1.80x105 pgCL-1) e da ciliati non loricati (1.57x105 pgCL-1).
Il confronto tra i valori di biomassa calcolati all’inizio (T0) e alla fine
dell’incubazione (T24) evidenzia una riduzione della biomassa dei ciliati non loricati che
passa a 9.25x104 pgCL-1, dei dinoflagellati (5.97x105 pgCL-1) e degli altri protozoi.
Aumenta sensibilmente la biomassa dei ciliati loricati che passa a 3.57x105 pgCL-1, mentre
rimane pressoché costante la biomassa dei micrometazoi.
L’analisi delle repliche contenenti anche il mesozooplancton alla fine del periodo
d’incubazione (T24) ha permesso di rilevare un aumento della biomassa dei ciliati non
loricati (1.93x105 pgCL-1) e dei dinoflagellati (8.96x105 pgCL-1). Si riduce debolmente la
biomassa degli altri protozoi ad un valore di 9.93x104 pgCL-1, mentre rimane pressoché
costante la biomassa dei micro metazoi (fig.2.5).

Ostreopsis- Risultati
83
Biomassa micozooplancton
0.00E+00
5.00E+00
1.00E+01
1.50E+01
2.00E+01
2.50E+01
3.00E+01
T0 T24 Bianco T24 meso
pgC
L-1*1
05
Ciliati non loricati
Ciliati loricati: Tintinnidi
Dinoflagellati eterotrofi
Altri protozoi
Micrometazoi
Fig.2.5 Biomassa dei diversi gruppi microzooplanctonici all’inizio e alla fine
dell’esperimento
2.4.2 Inizio dell’incubazione (T0)
Un’analisi complessiva dei campioni iniziali (T0) indica che l’esperimento è stato
preparato correttamente e pertanto alle rispettive diluizioni corrisponde un’effettiva
diminuzione degli organismi (fig.2.6).
Bontà delle diluizioni y = 240.16x + 47.756r2 = 0.9669
0.00E+00
5.00E+01
1.00E+02
1.50E+02
2.00E+02
2.50E+02
3.00E+02
3.50E+02
0% 20% 40% 60% 80% 100%
Diluizioni
pgC
L-1 *
106
Fig.2.6 Abbondanze in pgC L¹ del microfitoplancton al T0 (p<0.001)

Ostreopsis- Risultati
84
2.4.3 Fine dell’incubazione (T24) – Crescita apparente
Dopo aver effettuato l’analisi qualitativa e quantitativa dei campioni incubati per 24
ore è stato calcolato il coefficiente di crescita apparente in ogni diluizione, che è stato poi
riportato su un sistema di assi cartesiani nel quale in ascissa si trovano i fattori di diluizione
in ordinata la crescita espressa come ln (Ct/C0).
In fig.2.7 è riportata la crescita apparente di una Bacillariophycea, Chaetoceros
decipiens: si osserva una relazione significativa tra questa e i fattori di diluizione (r = 0.77,
p<0.01). Dai coefficienti istantanei di crescita k = 2.0977 e di predazione g = -1.5645 sono
stati ricavati il tasso di ingestione I = 84.812 µg C L̄ ¹d̄ ¹, la produzione potenziale Pp =
103.835 µg C L̄ ¹d̄ ¹ e la produzione reale Pr = 26.393 µg C L̄ ¹d̄ ¹.
Chaetoceros decipiens y = -1,5645x + 2,0977
r2 = 0,5962
-0.5
0
0.5
1
1.5
2
2.5
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
o)
Fig.2.7 Crescita apparente di Chaetoceros decipiens
In fig. 2.8 è riportata la crescita apparente di una seconda diatomea: Thalassionema
nitzschioides. In questo caso non si osserva alcuna relazione significativa tra questa e i
fattori di diluizione, il che significa che questa specie non è soggetta a predazione.

Ostreopsis- Risultati
85
Thalassionema nitzschioides
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C0)
Fig.2.8 Crescita apparente di Thalassionema nitzschioides
In fig. 2.9 è riportata la crescita apparente di una terza specie: Dactyliosolen
fragilissimus. Si osserva una relazione significativa tra questa e i fattori di diluizione (r =
0.61, p<0.01). Dai coefficienti istantanei di crescita k = 1.1021 e di predazione g = -0.6108
sono stati ricavati il tasso di ingestione I = 5.554 µg C L ¹d ¹, la produzione potenziale Pp
= 8.928 µg C L ¹d ¹ e la produzione reale Pr = 3.979 µg C L ¹d ¹.
Dactyliosolen fragilissimus y = -0.6108x + 1.1021r2 = 0.3704
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0% 20% 40% 60% 80% 100%
Diluizioni
Ln(C
t/C0)
Fig.2.9 Crescita apparente di Dactyliosolen fragilissimus

Ostreopsis- Risultati
86
In fig. 2.10 è riportata la crescita apparente di una quarta specie Cerataulina
pelagica: anche in questo caso si rileva una relazione significativa con i fattori di
diluizione e quindi si può asserire che la predazione è stata effettuata (r = 0.61, p<0.01).
Dai coefficienti istantanei di crescita k = 1.3633 e di predazione g = -0.7695 sono stati
ricavati il tasso di ingestione I = 12.675 µg C L̄ ¹d¯¹, la produzione potenziale Pp = 18.975
µg C L̄ ¹d̄ ¹ e la produzione reale Pr = 8.265 µg C L̄ ¹d̄ ¹.
Cerataulina pelagica y = -0,7695x + 1,3633r2 = 0,3741
00.20.40.60.8
11.21.41.61.8
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.2.10 Crescita apparente di Cerataulina pelagica
Le fig. 2.11 e 2.12 mostrano, invece, la crescita apparente di due specie di
Bacillariophyceae che non sono soggette a predazione. Si tratta di Cyclotella spp.e
Thalassiosira spp., entrambi organismi in grado di formare colonie organizzate in lunghe
catenelle. Si può notare dai grafici, infatti, l’assenza di relazione significativa con i fattori
di diluizioni.

Ostreopsis- Risultati
87
Cyclotella
0
0.2
0.4
0.6
0.8
1
1.2
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.2.11 Crescita apparente di Cyclotella spp.
Thalassiosira spp.
00.20.40.60.8
11.21.41.6
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.2.12 Crescita apparente di Thalassiosira spp.
Infine non si osserva alcuna relazione significativa nei confronti della microalga
Ostreopsis ovata (fig.2.13). La mortalità non è dovuta ad una predazione selettiva.
Ostreopsis ovata
-0.8
-0.6
-0.4
-0.2
0
0.2
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
0)
Fig.2.13 Crescita apparente di Ostreopsis ovata

Ostreopsis- Risultati
88
I risultati dell’esperimento di diluizione che riguardano le specie soggette a
predazione sono riassunti nella tabella 2.3, dove: k = coefficiente istantaneo di crescita, g =
coefficiente istantaneo di predazione, C0 = concentrazione iniziale (pg C L¯¹), <C> =
concentrazione media durante l’esperimento (pg C L¯¹), I = tasso di ingestione (pg C
L¯¹d̄ ¹), Pp = produzione potenziale (pg C L¯¹d̄ ¹), Pr = produzione reale (pg C L¯¹d̄ ¹).
Specie k g C₀
µgCL ¹
<C>
µgCL ¹ I
µgCL ¹d ¹ Pp
µgCL ¹d ¹ Pr
µgCL ¹d ¹
Chaetoceros 2,098 1,564 49,499 54,210 12,675 103,835 26,393 decipiens Dactyliosolen 1,102 0,611 8,101 9,094 5,554 8,928 3,979 fragilissimus Cerataulina 1,363 0,769 13,919 16,472 12,675 18,975 8,265 pelagica
Tabella 2.3 Risultati riguardanti le specie predate

Ostreopsis- Risultati
89
LISTA FLORISTICA Bacillariophyceae Dactyliosolen fragilissimus (Bergon) Hasle 1996 Navicula spp. Nitzschia longissima (Brébisson 1849) Ralfs 1861 Pleurosigma spp. W.Smith 1852 Proboscia alata (Brightwell) Sundstrom Thalassionema nitzschioides (Gruniw) Grunow ex Hustedt 1959 Chaetoceros danicus Cleve 1889 Chaetoceros decipiens Cleve 1873 Chaetoceros minimus (Levander) Marino,Giuffrè, Montresor et Zingone 1991Chaetoceros dadayi Pavillard 1913 Chaetoceros indet. Chaetoceros diversus Cleve 1873 Chaetoceros brevis Schutt 1895 Cerataulina pelagica (Cleve) Hendey 1937 Bacteriastrum furcatum Shadbolt 1854 Bacteriastrum delicatulum Cleve Thalassiosira indet. Cyclotella spp. Amphora spp. Cocconeis scutellum (Ehrenberg 1837) Boyer 1928 Licmophora gracilis (Ehrenberg) Grunow Plagiotropis spp. Diatomea Centrica>20 Pseudo-nitzschia pseudodelicatissima (Hasle) Hasle 1993 Pseudo-nitzschia indet. Pseudosolenia calcar-avis (Shultze) Sundstrom 1986 Leptocylindrus minimus Gran Thalassiosira rotula Meunier 1910 Hemiaulus hauckii Grunow in Van Heurck Dactyliosolen blavyanus (H. Peragallo) Hasle Dinophyceae Ceratium furca (Ehrenberg) Claparède et Lachmann 1859 Ceratium fusus (Ehrenberg) Dujardin 1841 Ceratium declinatum (Karsten) Jorgensen 1911 Dinophysis sacculus Stein 1883 Oxytoxum scolopax Stein 1883 Oxytoxum viride Schiller Prorocentrum minimum (Pavillard) Schiller 1933 Prorocentrum spp. Prorocentrum balticum (Lohmann) Loeblich III 1970 Prorocentrum gracile (Schutt) 1895 Ostreopsis ovata Fukuyo 1981 Lingulodinium polyedrum (Stein) Dodge 1989 Gymnodinium fusum Dinophysis caudata Saville-Kent 1881

Ostreopsis- Risultati
90
Tecati indet. Tecati > 20 µm Primnesiophyceae coccolitoforali indet. Incertae sedis Leucocryptos marina (Braaud) Butcher 1967

Ostreopsis- Risultati
91
2.4.4 Esperimento di predazione del mesozooplancton (Grazing) – giugno 2009
Nell’esperimento di grazing si è valutata la predazione da parte del
mesozooplancton sul popolamento di Ostreopsis ovata.
Il conteggio dei campioni in 3 repliche al T0, T24 e T24 con il mesozooplancton ha
permesso di compilare una tabella relativa alle concentrazioni iniziali e finali di Ostreopsis
ovata sia in termini di abbondanza (Tab.2.4), che in termini di biomassa (Tab.2.5).
Ostreopsis ovata Cell L-1*103 ±SD C0 46.29 9.68 C24 41.47 3.39 C24 copepodi 0.81 0.47
Tab.2.4 Abbondanza di Ostreopsis ovata
Ostreopsis ovata pgC L-1* 106 ±SD C0 191.63 39.05 C24 154.27 8.30 C24 copepodi 5.71 3.30
In termini di abbondanza la concentrazione iniziale e quella finale di Ostreopsis
ovata subiscono una leggera diminuzione passando da 4.63x104 cellule L-1 a 4.15x104
cellule L-1. L’abbondanza di O. ovata diminuisce sostanzialmente nei campioni contenenti
il mesozooplancton. Andamento analogo si rileva in termini di biomassa passando da
1.92x108 pgC L-1 e 1.54x108 pgC L-1 nei campioni al T0 e T24 a 5.71x106 pgC L-1 nei
campioni finali con il mesozooplancton (fig. 2.14, 2.15).
Tab.2.5 Biomassa di Ostreopsis ovata

Ostreopsis- Risultati
92
Grazing su Ostreopsis
0
10
20
30
40
50
60
C0 C24 C24 con Meso
Cel
l L-1 *
103
Grazing su Ostreopsis
0
50
100
150
200
250
C0 C24 C24 con Meso
pgC
L-1*1
06
2.4.5 Caratterizzazione del popolamento microzooplanctonico (settembre 2009)
La biomassa del microzooplanctonico all’inizio dell’esperimento (T0) è
caratterizzata prevalentemente da ciliati non loricati con valori pari a 2.18 x106 pgCL-1, e
dai micrometazoi (1.44x106 pgCL-1). I dinoflagellati eterotrofi sono presenti con valori pari
a 3.80x104 pgCL-1, seguiti dai ciliati loricati (9.11x104 pgCL-1). Gli ebridi, rappresentati
esclusivamente dalla specie Hermesium adriaticum, hanno una biomassa pari a 1.96x104
pgCL-1. Gli altri protozoi, peraltro presenti soltanto all’inizio dell’esperimento hanno una
biomassa pari a 3.82x103 pgCL-1 (Tab2.6).
Fig. 2.14 Abbondanza di Ostreopsis ovata
Fig. 2.15 Biomassa di Ostreopsis ovata

Ostreopsis- Risultati
93
Microzooplancton T0
pgC L-1*104 T24 Bianco pgC L-1*104
T24 copepodi pgC L-1*104
Ciliati non loricati 217.67 171.65 88.80 Ciliati loricati 2.40 9.77 0.00 Dinoflagellati 3.80 4.31 57.29 Ebriida 1.96 2.55 1.96 Micrometazoi 143.95 137.86 119.31 Protozoi 0.38
Tab.2.6 Biomassa microzooplancton al T0, T24 e T24 con copepodi
In fig.2.16 vengono riportate le variazione del popolamento durante l’incubazione,
in assenza ed in presenza del mesozooplancton aggiunto durante l’esperimento.La biomassa
dei ciliati non loricati si riduce dopo le 24 ore di incubazione ad un valore pari a 1.72x106
pgCL-1, diminuendo ulteriormente nelle repliche contenenti il mesozooplancton (0.88x106
pgCL-1). La biomassa dei ciliati loricati subisce un lievissimo incremento durante
l’incubazione, passando a 9.77x104 pgCL-1, mentre in presenza dei copepodi non si
ritrovano forme loricate.
I dinoflagellati aumentano leggermente durante l’incubazione e subiscono un
ulteriore incremento in termini di biomassa nei campioni con il mesozooplancton con
valori pari a 5.73x105 pgCL-1 .
Le biomasse degli ebridi e dei micrometazoi rimangono pressochè invariate nei
campioni al T0, in quelli di controllo e in quelli con il mesozooplancton.
Biomassa del microzooplancton
0
50
100
150
200
250
T0 T24 bianco T24 Copepodi
pgC
l-1*1
04
Ciliati non loricati
Ciliati loricati
Dinoflagellati
Ebriida
Micrometazoi
Protozoi
Fig.2.16 Biomassa microzooplancton all’inizio (T0) e alla fine dell’incubazione (T24)

Ostreopsis- Risultati
94
2.4.6 Esperimento di predazione del mesozoo plancton (grazing) – settembre 2009
Nell’esperimento di grazing si è voluta verificare la predazione da parte del
mesozooplancton sulla microalga Ostreopsis ovata.
Dal conteggio dei campioni (in 3 repliche) al T0, T24 bianco e T24 con i copepodi, si
è evidenziata una variazione in termini di biomassa di Ostreopsis (fig.2.17).
Grazing su Ostreopsis
0
1
2
3
4
5
6
T0 T24 bianco C24 + Centropages
pg C
L-1*1
08
Fig.2.17 Biomassa di Ostreopsis ovata
In termini di biomassa (Tab.2.6) si parte da una concentrazione iniziale pari a
1.92x108 pgCL-1 e si osserva un leggero decremento di biomassa nei campioni messi ad
incubare senza i copepodi.
La biomassa di Ostreopsis ovata, aumenta ad un valore di 3.65x108 pgCL-1 nei
campioni in cui si è aggiunto il mesozooplancton.
Ostreopsis ovata pgCL-1 *108 ± SD C0 1.92 0.39 C24 bianco 1.42 0.38 C24+ Centropages 3.65 1.85
Tab.2.6 Biomassa di Ostreopsis ovata

Ostreopsis- Risultati
95
In termini di abbondanza si verifica un andamento analogo in cui si verifica un
dimezzamento nel numero di cellule dopo l’incubazione senza i copepodi pari a 2.02* 104
cell L-1 a 5.19*104 cell L-1 in presenza di Centropages (Tab. 2.7)
Ostreopsis ovata Cell L-1 *104 ± SD C0 4.63 0.85 C24 bianco 2.02 0.53 C24+ Centropages 5.19 2.63
Tab.2.7 Abbondanza di Ostreopsis ovata

Ostreopsis ovata-Discussione e Conclusioni
96
2.5 DISCUSSIONE E CONCLUSIONI
Lo scopo principale del lavoro è stato quello di determinare la mortalità della
componente microfitoplanctonica ed in particolare di Ostreopsis ovata indotta dalla
predazione del microzooplancton per identificare l’eventuale trasferimento lungo la rete
trofica pelagica dell’alga e di conseguenza della sua tossina. Essendo uno dei primi studi
non si trovano in letteratura dati paragonabili a quelli ricavati in questo studio.
L’importanza che ha il microzooplancton all’interno della rete trofica marina è
accertata da tempo: Calbet e Landry (2004) affermano che la predazione da parte dei
micro-predatori è la maggiore causa della mortalità degli organismi fitoplanctonici negli
habitat oceanici. Inoltre, suggeriscono che ciò potrebbe essere vero anche nelle acque
costiere più produttive come il Nord Adriatico. In studi svolti precedentemente in questa
zona (Fonda Umani & Beran, 2003: Fonda Umani et al., 2005) è stato rilevato come il
microzooplancton predi effettivamente su un’ampia gamma di prede con una predazione
peraltro altamente selettiva.
La predazione è altamente selettiva e variabile in funzione alla composizione della
comunità delle prede che, come si può notare dalla lista floristica allegata, è molto
diversificata. In questo lavoro è stato possibile osservare l’influenza del predatore sul
microfitoplancton selezionando le specie più abbondanti, quindi ritenute significative, e
studiando il tasso di predazione su ciascuna. La fig.2.18 riporta le abbondanze di ognuna
delle sette specie considerate significative espresse in cellule L-1 al T0. Si può notare che di
queste sette specie sei sono Bacillariophyceae (diatomee).
Fig.2.18 Abbondanze delle specie significative al T0 (Cell. L-1)
Le specie maggiormente rappresentate sono: Cyclotella spp., con una percentuale
del 43%, Thalassiosira spp., in percentuale del 23%, e Leucocryptos marina per il 17%. Le
Abbondanze T 0 (cell L -1)
7%
3%
4%
3%
43%
23%
17%
Cyclotella spp
Thalassiosira spp
Leucocryptos marina
Chaetoceros decipiens
Thalassionema itzschioides
Dactyliosolen fragilissimus
Cerataulina pelagica

Ostreopsis ovata-Discussione e Conclusioni
97
quattro restanti, meno numerose, risultano essere: Chaetoceros decipiens per il 7%,
Dactyliosolen fragilissimus per il 4%, Thalassionema nitzschioides e Cerataulina pelagica
entrambe in percentuale del 3%. Di queste sette specie quelle che risultano essere predate
significativamente dal microzooplancton sono Chaetoceros decipiens, Cerataulina
pelagica e Dactyliosolen fragilissimus, quindi quelle presenti in minore concentrazione.
Ciò dimostra che la scelta del predatore non verte su queste tre specie per la loro
abbondanza. Nella fig. 2.19 è possibile osservare la abbondanze fitoplanctoniche dopo
l’incubazione (T24) sempre espresse in cellule L-1
Fig.2.19 Abbondanze delle specie significative al T24 (Cell L-1)
Si nota come le abbondanze percentuali relative ad ogni specie non vari
particolarmente al T24, quindi dopo l’attività di grazing, rispetto al T0. L’unica eccezione è
Thalassiosira spp. che supera, con una percentuale del 38 %, Cyclotella, 36%, mostrando
un maggior tasso di crescita. Le rimanenti cinque specie, comprese quelle predate,
risultano essere leggermente meno abbondanti. Pare quindi che l’impatto da predazione
venga controbilanciato dall’alto tasso di crescita delle diatomee, quindi che la mortalità
indotta dal microzooplancton non abbia effetto su una parte delle specie presenti che
rimangono disponibili per altri predatori. Questo conferma i dati ottenuti da Gallegos et al.
(1996) a febbraio durante la grande fioritura della diatomea Odontella sinensis in
Manukau Harbour, New Zealand. L’autore osserva come soltanto in primavera ci sia
predazione da parte del microzooplancton sulla componente fitoplanctonica, senza però
che ci sia un controllo consistente ma soltanto un contributo nello strutturare la comunità
fitoplanctonica.
Abbondanze T 24 (cell L -1)5%
3%
3%
2%
36%38%
13%
Thalassiosira spp Cyclotella
Leucocryptos marina
Thalassionema itzschioides
Dactyliosolen fragilissimus
Cerataulina pelagica
Chaetoceros decipiens

Ostreopsis ovata-Discussione e Conclusioni
98
Lo studio del popolamento microzooplanctonico ha evidenziato come questo sia
caratterizzato da micrometazoi e dinoflagellati eterotrofi che aumentano in termini di
biomassa durante l’incubazione anche nei campioni contenenti i copepodi. Diminuiscono
invece le abbondanze dei tintinnidi nei campioni con il mesozooplancton.
I risultati relativi all’esperimento di diluizione hanno evidenziato come non si
riscontri mortalità indotta da predazione nei confronti di Ostreopsis ovata da parte del
microzooplancton. La specie viene evidentemente selezionata in termini negativi, il
microzooplancton, costituito prevalentemente di piccole dinoflagellate, si nutre di altre
specie fitoplanctoniche di dimensioni più ridotte.
I dati ottenuti dall’analisi dei campioni contenenti il mesozooplancton hanno
evidenziato la diminuzione in termini di abbondanza e di biomassa di Ostreopsis ovata.
Poiché la biomassa del microzooplancton cresce durante il periodo di incubazione anche in
presenza dei copepodi, si può ipotizzare che i copepodi indirizzino la loro predazione verso
la microalga causando conseguentemente, il suo possibile trasferimento ai livelli trofici
superiori.
Lo studio del popolamento microzooplanctonico relativo all’esperimento di grazing
svoltosi nel mese di settembre ha evidenziato una netta dominanza di ciliati non loricati
all’inizio dell’esperimento che decrescono in termini di abbondanza e biomassa durante
l’incubazione ed ancor di più in presenza del mesozooplancton. I dinoflagellati invece
aumentano durante il periodo di incubazione anche nei campioni con i copepodi. I
micrometazoi rimangono invariati durante l’esperimento.
La biomassa di Ostreopsis ovata subisce un decremento passando dal T0 al T24,
questo potrebbe far pensare ad una mortalità naturale oppure ad una predazione da parte del
microzooplancton. Nei campioni con i copepodi la biomassa di Ostreopsis aumenta e
questo induce a ritenere che non ci sia stata predazione durante l’incubazione da parte di
Centropages spp.. Viceversa le differenze riscontrate tra l’inizio e la fine dell’incubazione a
carico dei ciliati non loricati che subiscono un marcato decremento indica una possibile
predazione da parte del mesozooplancton su questa frazione microzooplanctonica.
I due esperimenti di grazing hanno dato quindi risultati discordanti: nel primo caso
Ostreopsis viene predata, nel secondo caso non si osserva alcun decremento in presenza di
copepodi. Tali discordanze possono essere messe in relazione alle diverse specie di
predatori utilizzate nei due esperimenti: copepoditi di Acartia nel primo caso e di
Centropages nel secondo. Purtroppo quando si effettuano questi esperimenti si possono

Ostreopsis ovata-Discussione e Conclusioni
99
utilizzare soltanto i copepodi presenti in numero consistente nel campione di plancton
raccolto. Gli organismi devono essere vitali e catturati rapidamente per essere immessi nelle
bottiglie d’incubazione in numero tale da garantire che gli effetti della predazione sul
popolamento naturale, in questo caso arricchito con Ostreopsis, proveniente da coltura,
siano statisticamente significativi. Questo limita forzosamente la scelta degli organismi da
usare per l’esperimento.

Antartide-Introduzione
100
3. ANTARTIDE
3.1. Introduzione
Il termine “plancton” deriva da una parola greca che significa “vagabondo “ coniato
per la prima volta dallo scienziato tedesco Victor Hensen (1887). Il termine si riferisce
all’insieme degli organismi compresi fra 0.02 e >400 µm che sono trasportati dalle
correnti. Il plancton è stato poi organizzato dai biologi in classi e sottoclassi.
Si distingue in fitoplancton e zooplancton. Il fitoplancton è il termine generico che
indica quel gruppo di organismi microscopici che si trovano nella zona eufotica del mare.
Sono i responsabili della produzione primaria, incorporando C inorganico dall’ambiente
negli organismi viventi, processo questo che innesca la rete trofica. Si stima che il
fitoplancton contribuisce a circa la metà della produzione primaria globale mentre l’altra
metà deriva dalle piante.
La distribuzione del fitoplancton non è uniforme e questa dipende da molteplici
fattori. Poiché la fotosintesi rimuove dall’atmosfera CO2 rilasciando O2 la produzione
primaria globale dovuta al fitoplancton è una variabile importante nei modelli climatici. Il
processo fotosintetico arricchisce l’acqua di ossigeno. Esplosioni del popolamento
fitoplanctonico chiamate anche “bloom” possono avere effetti disastrosi specialmente nelle
regioni costiere. Alcune specie producono tossine che se presenti in alte concentrazioni
possono avvelenare pesci ed animali entrando nella rete trofica.
Il fitoplancton può sopravvivere e moltiplicarsi nella fascia superficiale dell’oceano
chiamata anche “zona eufotica”. La profondità della zona eufotica è variabile e dipende
dalla limpidezza dell’acqua, dalla latitudine e dalla stagione. Al largo degli oceani la zona
eufotica oscilla tra i 50 e i 100 metri. La produttività del fitoplancton è fortemente
influenzata da due fattori fondamentali quali la luce, disponibile solamente negli strati
superficiali e i nutrienti, disponibili prevalentemente nelle acque profonde. Mentre le
piante terrestri hanno sviluppato un apparato radicale in grado di risolvere i problemi di
trasporto ed assorbimento di nutrienti lungo il fusto, parte del fitoplancton è in grado di
spostarsi verticalmente per recuperare i nutrienti localizzati nelle acque più profonde. Ci
sono altri fattori che direttamente o indirettamente influenzano la produzione planctonica
quali la temperatura e la salinità dell’acqua. Alcune specie infatti si sono adattate a
sopravvivere a specifiche condizioni di temperatura e salinità. Questi parametri inoltre
regolano la stabilità della colonna d’acqua ed il conseguente grado di mescolamento,

Antartide-Introduzione
101
responsabile questo del trasporto di nutrienti disciolti nella zona eufotica ad utilizzo del
fitoplancton. Questi parametri fisici sono di fondamentale importanza per le dinamiche di
sviluppo del plancton. Le due classi principali di fitoplancton sono le diatomee e i
dinoflagellati. I dinoflagellati sono deboli nuotatori e nuotano grazie all’uso di flagelli
perlopiù contrastando l’effetto della gravità. Le diatomee invece non nuotano attivamente
ma hanno sviluppato meccanismi in grado di contrastare la velocità di precipitazione verso
il fondo.
3.1.1 Ruolo del microzooplancton in Antartide
I primi studi sul microzooplancton antartico risalgono alla della Deutsche Sudpolar
Expedition e sono datati 1901-1903 (Laakmann, 1907; Schroeder, 1909; Popofsky, 1909).
Fino a pochi anni orsono le interazioni trofiche nell’ambiente pelagico antartico
venivano si riferivano allo schema classico di rete alimentare, caratterizzato dalla
predazione da parte dello zooplancton di grandi dimensioni su diatomee di grandi
dimensioni. L’esempio classico di rete alimentare antartica è dato dallo schema:
«diatomee predate da krill, consumato da grandi carnivori
quali balene, pinguini, foche» (El-Sayed, 1971)
Negli ultimi anni l’attenzione si è rivolta verso un altro modello alimentare, che
prende in considerazione le comunità microbiche e che include batteri, fitoplancton di
piccole dimensioni e protozoi. Si ritiene che tali comunità rivestano un ruolo importante
nelle dinamiche dell’ecosistema pelagico antartico (Hewes et al., 1985); ci si è spostati
quindi dalla semplice rete alimentare classica verso una più complessa rete alimentare, che
potremmo definire “mistivora”.
Hewes et al. (1985) hanno dimostrato che il modello del “microbial loop”, proposto
da Azam et al. (1983), è applicabile anche alle acque antartiche. Numerosi studi hanno
identificato nei predatori microeterotrofi (microzooplancton) la chiave del sistema
microbico marino antartico (Buck e Garrison, 1983; Hewes et al., 1985; Garrison e Buck,
1989; Waters et al., 2000) Il ruolo dei processi microbici è ora argomento centrale di molti
studi nella regione antartica, con particolare attenzione rivolta alle interazioni autotrofi -
eterotrofi ed al ciclo del carbonio (Hewes et al., 1990; Davidson & Marchant, 1992;

Antartide-Introduzione
102
Kopczynska et al.; 1995; Archer et al.,1996a, 1996b; Leacky et al., 1996; Xiuren et al.,
1996; Arrigo et al., 1999; Fonda Umani et al., 1998, 2002).
Il microzooplancton è uno dei componenti delle comunità microbiche ed è
importante in quanto riesce a predare organismi le cui misure sono inferiori a quelle tipiche
delle prede del mesozooplancton; la predazione da parte del microzooplancton permette
quindi l’assimilazione nella rete alimentare di una percentuale maggiore della produzione
primaria. E’ una produzione significativa anche dal punto di vista quantitativo, molteplici
studi hanno dimostrato che ad esso va attribuito una notevole parte della predazione sul
fitoplancton negli oceani Atlantico (Burkill et al., 1993a; Verity et al., 1993), Indiano
(Burkill et al., 1993b) e Pacifico (Miller, 1993), arrivando a tassi di rimozione della
produzione primaria fitoplanctonica anche superiori al 100% (Landry et al., 1993).
Molti studi hanno indicato come il microbial loop assuma una maggiore importanza
alla fine della primavera australe, quando le comunità di grandi diatomee vengono
sostituite, nel ruolo di produttori primaripiu’ imporanti, dalle componenti pico e
nanoplanctoniche, e si ha un aumento della frazione eterotrofa dovuto all’abbondante
materiale organico particellato prodotto in precedenza (Alder e Boltovosky, 1993; Garrison
et al., 1998); di conseguenza i popolamenti microzooplanctonici raggiungono il loro
massimo di crescita a spese del microbial loop, riuscendo a “rimpacchettare” la materia
organica in dimensioni più adatte al consumo dei livelli trofici superiori arrivando fino a
sostenere il larga parte l’elevato standing stock di crostacei (krill) in Antartide.
3.1.2 Caratterizzazione dell’area di studio
Il continente antartico è circondato interamente dall’Oceano Meridionale, o
Antartico, che occupa un’area di circa 36 milioni di km2, rappresentando il 10% degli
oceani della Terra (fig. 1). Le estremità meridionali degli oceani Pacifico, Atlantico ed
Indiano sono collegate tra loro dalla Corrente Circumpolare Antartica, che fluisce a sud
della Convergenza Sub-Tropicale (40°S) guidata dai forti venti occidentali. A sud del 65°S
è presente un flusso d’acqua occidentale, adiacente al continente condizionato dai venti
prevalenti orientali: la Corrente Circumpolare Profonda (Circumpolar Deep Water -
CDW). La CDW è coinvolta nella formazione di tutte le masse d’acqua antartiche: le
acque meno dense, come l’acqua superficiale antartica (Antarctic Surface Water- ASW),
sono relegate allo strato superficiale, mentre le acque più dense, come l’acqua profonda

Antartide-Introduzione
103
antartica (Antarctic Bottom Water – ABW), sprofondano andando così a formare le acque
abissali.
Fig.3.1 Il continente Antartico
L’interesse dal punto di vista oceanografico e climatico per l’Oceano Meridionale è
legato proprio alla grande varietà di masse d’acqua prodotte ed alla loro capacità di
influenzare la circolazione generale degli oceani. In particolare, le acque di piattaforma,
prodotte lungo il margine del continente antartico, con temperature prossime al punto di
congelamento -1.9°C) contribuiscono alla ventilazione delle acque profonde, influenzando
il clima dell’intero oceano.
L’oceano meridionale gioca un ruolo molto importante anche nel ciclo globale del
carbonio, degli elementi biogenici, come azoto, fosforo e silice (Legendre & Michaud,
1998), e nel ciclo globale della materia (Sullivan et al., 1993). Il flusso del carbonio
biogenico verso i grandi metazoi (risorse rinnovabili) e verso le profondità marine
(carbonio sequestrato), gioca un ruolo centrale nella regolazione della concentrazione
atmosferica della CO2, ed è materia di grande interesse attuale (Hanson et al., 2000;
Trèguer et al, 2002). Volk e Hoffert (1985) hanno identificato tre tipi di meccanismi,
definiti pompe, che possono trasportare la CO2 dall’atmosfera all’oceano profondo.
La “solubility pump” (pompa di tipo fisico) è particolarmente attiva in aree di
formazione di acqua profonda che risulta da un incremento della densità prodotta o da un
abbassamento della temperatura (alle alte latitudini) o da un aumento della salinità (come
nella polynya di Baia Terra Nova).

Antartide-Introduzione
104
La “carbonate pump” dipende dalla sedimentazione in profondità di organismi con
rivestimento calcareo.
La “soft-tissue pump” (pompa biologica della CO2) è attivata dall’incorporazione
fotosintetica di carbonio inorganico in molecole organiche da parte di alghe microscopiche,
seguita da un trasporto del fitodetrito verso le acque profonde.
Legendre e LeFévre (1992) hanno proposto di classificare i pools di carbonio
organico nell’oceano sulla base del loro tempo di turnover (cioè il tempo trascorso tra
assimilazione fotosintetica e ritorno come CO2 all’atmosfera) e hanno definito tre
principali compartimenti: carbonio organico a vita breve (<10-2 anni), carbonio organico a
vita lunga (10-2 – 102 anni) e carbonio biogenico sequestrato (>102 anni).
Il carbonio a vita breve viene incorporato in organismi con elevato tasso di turnover
e comprende anche il carbonio organico labile disciolto e viene trasportato attraverso la
rete trofica microbica (“micobial food web”: fitoplancton-batteri eterotrofi- protozoi).
Il carbonio a vita lunga include le risorse marine rinnovabile e transita attraverso la
rete alimentare classica (microalghe-crostacei-piccoli e grandi predatori) (Azam, 1998).
Il carbonio organico sequestrato comprende una gran varietà di forme: i resti
organici seppelliti nel sedimento, depositi inorganici di origine organica, materiale
organico disciolto refrattario e CO2 disciolta nelle acque profonde come risultato della
respirazione profonda (Legendre, 1996).
La produzione primaria (il carbonio fotosintetizzato) può essere metabolizzata dagli
organismi e restituita come CO2 all’atmosfera, attraverso la respirazione o può essere
incanalata tramite esportazione verticale di materiale in sedimentazione e/o attraverso la
biomassa dei grandi consumatori.
La struttura della rete trofica che sostiene l’ecosistema antartico risulta quindi più
complessa rispetto alla classica rete alimentare che si basava sul solo flusso di carbonio
dalle diatomee al krill ai predatori superiori. La rete trofica oggi si basa sulle componenti
più piccole, ma sicuramente più abbondanti dell’oceano: il picoplancton, il nanoplancton e
il microplancton.
Alla rete trofica classica sì è affiancata quella microbica (il microbial loop), che
sostiene la rete alimentare attraverso la produzione rigenerata.
La rete trofica microbica riveste un ruolo estremamente importante nell’ecosistema
antartico, dato che l’Oceano Meridionale presenta caratteristiche esclusive non
paragonabili con ambienti oligotrofici delle basse latitudini. che nelle acque antartiche vi è
L’efficienza ecologica della rete alimentare risulta condizionata e controllata dalla struttura

Antartide-Introduzione
105
trofica dell’ecosistema che è soggetta a variazioni stagionali estreme. L’estate australe
rappresenta il periodo di massima efficienza della produzione dei consumatori. Le
diatomee, che caratterizzano la fase primaverile, vengono soppiantate da organismi
autotrofi di dimensioni inferiori (<20µm); la produzione di sostanza organica disciolta è
elevata e le fonti principali sono l’essudazione, le perdite durante la predazione,
l’escrezione metabolica, la morte, l’autolisi o la lisi virale.
La sedimentazione del particellato organico è consistente e l’attività del micobial
loop garantisce il riciclaggio del carbonio organico disciolto liberato dai livelli trofici
superiori. Nella tarda estate, nel periodo che segue la fioritura algale, la biomassa
fitoplanctonica e la produzione primaria sono basse, prevale il nanoplancton autotrofo.
Per quanto riguarda la distribuzione dei diversi organismi nelle acque antartiche, si
osservano consistenti variazioni spazio-temporali (Smith et al., 1996). Nelle acque al largo,
tra la Circumpolare e la zona costiera, i livelli di biomassa del fitoplancton non
raggiungono mai valori molto alti e sembra che il microbial loop sostenga l’intero
ecosistema per tutto l’anno; nelle zone costiere ed in particolare in quelle di polynya del
Mare di Ross si registrano al contrario valori di produzione primaria e biomassa
fitoplanctonica molto più alti.
3.1.3 Il Mare di Ross
Dal punto di vista geografico l’Oceano Antartico è suddiviso in vari mari. I
principali sono: il Mare di Ross, il Mare di Weddell, il Mare di Scozia. Mari secondari di
minor estensione sono il Mare di Lazarev e il Mare di Riser-Larsen.
Il Mare di Ross occupa un’estesa baia che si incunea nel Continente Antartico ed è
limitato ad est dalla Terra Edward VII, ad ovest dalla Terra della Regina Vittoria, a sud
dalla barriera di ghiaccio (Ross Ice Shelf), mentre il limite settentrionale può identificarsi
nella congiungente Capo Adare - Capo Colbek. Le profondità di questo mare non sono
elevate e superano i 400-500 metri soltanto verso l’estremità settentrionale, a molta
distanza dalla costa.
Il Ross Ice Shelf è una piattaforma di ghiaccio permanente di grande importanza, in quanto
determina una circolazione che fa sprofondare acqua calda, la quale viene distinta in: acqua
di fondo a bassa salinità (Low Bottom Shelf Water – HBSW) e acqua di fondo ad alta
salinità (High Bottom Shelf Water – HBSW) (Jacobs et al., 1970; Gordon e Tchernia,
1972). Quest’ultima, chiamata anche Ross Sea Bottom Water, è formata dal mescolamento

Antartide-Introduzione
106
di acque profonde e di piattaforma. Tali acque si distribuiscono omogeneamente al di sotto
dei 400 m di profondità, ed alimentano le correnti di fondo che sono responsabili del
trasporto del freddo alle latitudini più basse, giocando un ruolo chiave nella circolazione
oceanica globale.
Un’altra caratteristica del Mare di Ross è la formazione stagionale della polynya,
un’area di ghiaccio sottile e di acque libere, circondata dal pack ice (fig. 3.2). Questo
fenomeno è determinato da forti venti catabatici provenienti dal plateau continentale (Bates
et al., 1998); ha origine a nord del Ross Ice Shelf ed, espandendosi rapidamente in
ampiezza nel mese di novembre, va a costituire la più ampia polynya della regione. Una
polynya di piccole dimensioni, che persiste per buona parte dell’inverno, si sviluppa nella
Baia Terra Nova a nord della lingua di ghiaccio del Drygalski (Arrigo et al., 1999). Le
zone di polynya sono caratterizzate da intense fioriture fitoplanctoniche e rappresentano le
aree più produttive dell’Oceano Antartico. Durante la primavera e l’estate, l’introduzione
di una gran quantità di acqua a bassa salinità e ad elevato tenore di nutrienti, derivante
dallo scioglimento dei ghiacci marini, e l’adeguata radiazione luminosa, garantiscono
un’ottimale crescita del fitoplancton (Smith e Gordon, 1997).
Fig.3.2 Polynya antartica

Antartide-Introduzione
107
3.1.4 Baia Terra Nova
Baia di Terra Nova è il sito di una polynya costiera con ricorrenza annuale nella
zona occidentale del Mare di Ross, la quale è stata descritta per la prima volta da
Bromwich e Kurtz (1984) e, più recentemente, in termini di distribuzione di masse d’acqua
e variabilità termoalina è stata analizzata da Budillon e Spezie (2000).
Studi precedenti in questa area (Innamorati et al., 1991, 1999) hanno dimostrato la
presenza di un massimo di clorofilla nel tardo dicembre, seguito da una temporanea
diminuzione della biomassa fitoplanctonica e poi da un altro massimo in febbraio. E’ stato
osservato che la stabilità della colonna d’acqua gioca un ruolo importante nel favorire e
mantenere la fioritura nella polynya. Una fioritura molto vasta e persistente di Phaeocystis
antarctica (nanoflagellati) è stata osservata in questa area da metà a fine gennaio (Arrigo &
McClain, 1994; Fonda Umani et al., 2002). Nella prima parte della fioritura primaverile
sono state osservate in proporzioni uguali P. antarctica e diatomee, mentre
successivamente, la composizione della comunità appare fortemente influenzata
dall’alternanza della dominanza nano/microfitoplancton.

Antartide-Materiali e Metodi
108
3.2 MATERIALI E METODI
In questo lavoro vengono riportati i
risultati di otto esperimenti di diluizione
effettuati durante diverse campagne antartiche
così come riassunto nella tabella 3.1 e in
figura 3.3
L’analisi dei campioni è stata effettuata
in Italia usufruendo delle attrezzature di
microscopia del Laboratorio di Biologia
Marina di Trieste.
Nella mia tesi ho ri-elaborato in modo
omogeneo i dati ottenuti dalle analisi condotte
da altri studenti.
Gli esperimenti sono stati realizzati per valutare l’effetto della predazione del
microzooplancton sul comparto fitoplanctonico utilizzando il metodo delle diluizioni già
descritto precedentemente.
Esp Anno Area campionamento Latitudine Longitudine 1 1997 Largo del M. di Ross 70°00' 77'' S 175°05' 5 6''W 2 1997 Polynya Baia T. Nova 75°13' 28'' S 164°56' 2 3'' W 3 2000/01 Polynya Baia T. Nova 74°54' 65'' S 164°21 ' 07'' W 4 2000/01 Largo del M. di Ross 70°00' 77'' S 175°05 ' 56''W 5 2000/01 Largo del M. di Ross 76°41' 67'' S 169°03 ' 29'' W 6 2003 Polynya Baia T. Nova 75°06' 08'' S 164°29' 0 7'' W 7 2003 Largo del M. di Ross 75°03' 43'' S 176°29' 1 7'' W 8 2005/06 Polynya Baia T. Nova 75°13' 28'' S 164°56 ' 23'' W
Tab.3 1 Campionamenti condotti durante diverse campagne antartiche
Fig.3.3 Siti di campionamento
nel Mare di Ross

Antartide-Risultati
109
3.3 RISULTATI
Esperimento 1
Questo esperimento è stato condotto nel dicembre del 1997 in Antartide. I
campionamenti sono stati fatti al Largo del Mare di Ross.
Cell L-1 T0 C0 <C> Fitoplancton totale 1.90E+03 1.464 1.89 Diatomee 1.43E+03 0.68 0.84 Dinophyceae 2.70E+02 0.78 1.05 Cryptophyceae 2.00E+02 0.50 1.00
Tab.3.2 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media durante l’esperimento
(µg C L-1)
I valori di abbondanza e biomassa riportati in tab.3.2 sono relativi alle specie
fitoplanctoniche dominanti durante il periodo di campionamento. I valori di biomassa
relativi alle specie trovate sono sempre <0.1 µgC L-1
L’analisi dei campioni alle varie diluizioni ha evidenziato un non corretto
allestimento dell’esperimento in quanto non si verifica una riduzione del numero di
organismi aumentando il fattore diluizione (fig.3.4).
Fitoplancton totale
0.00
0.50
1.00
1.50
2.00
2.50
0% 20% 40% 60% 80% 100%
diluizioni
µg C
L-1
Fig.3.4 Bontà delle diluizioni (C0) relative al fitoplancton totale

Antartide-Risultati
110
Esperimento 2
Questo esperimento è stato condotto nel dicembre del 1997 nella Polynya di Baia
Terra Nova. Le abbondanze relative al fitoplancton totale sono pari a 3.9x104 cell L-1 pari
ad una biomassa di 8.16 µg C L-1 (Tab.3.3)
Cell L-1 T0 C0 <C> Fitoplancton totale 3.90E+04 8.16 5.87 Diatomee 2.05E+04 4.57 2.84 Dinophyceae 7.22E+03 3.37 2.87 Cryptophyceae 1.04E+04 0.13 0.04
Tab. 3.3 Abbondanze e biomasse fitoplanctoniche C0=concentrazione
iniziale (µg C L-1); <C>= concentrazione media durante l’esperimento (µg C L-1)
L’analisi complessiva dei campioni iniziali (C0) alle varie diluizioni indica che
l’esperimento è stato correttamente allestito e quindi alle rispettive diluizioni di acqua
corrisponde un’effettiva diluizione degli organismi. La bontà delle diluizioni è verificata
anche per le specie più abbondanti (fig.3.5)
Fitoplancton totale y = 8.5337x + 0.4732r2 = 0.8942
0
2
4
6
8
10
0% 20% 40% 60% 80% 100%
diluizioni
µgC
L-1
Fig.3.5 Bontà delle diluizioni relative al fitoplancton totale al C0

Antartide-Risultati
111
Dopo l’analisi dei campioni all’inizio dell’esperimento, è stato calcolato il
coefficiente di crescita apparente in ogni diluizione che è stato riportato su un sistema di
assi cartesiani.
In fig.3.6 sono riportati i coefficienti di crescita apparente relativi al fitoplancton
totale e non si osserva una relazione significativa. La mortalità non è indotta da predazione
selettiva. Non c’è mortalità indotta da predazione neanche nei confronti dei gruppi più
abbondanti quali Diatomee, Dinophyceae e Cryptophyceae (Fig.3.6).
Fitoplancton totale
-10
-5
0
5
10
15
0% 20% 40% 60% 80% 100%
diluizioni
Ln(C
t/C0)
Fig.3.6 crescita apparente per il fitoplancton totale. Non si osserva una
relazione significativa
Esperimento 3
Questo esperimento è stato condotto durante l’estate australe del 2000/01. I
campionamenti sono stati condotti in Polynya di Baia Terra Nova
E’ stata condotta un’analisi del popolamento microzooplanctonico presente nel
corso dell’esperimento. Il popolamento iniziale è costituito da dinoflagellati eterotrofi
(56.21%), ciliati (38.33%), e fasi larvali di Metazoi (5.46%). Tra i dinoflagellati eterotrofi
sono stati distinti i generi Protoperidinium, diplaopsalis e la specie Gyrodinium lachima,
tra i ciliati i gruppi Strombilidiidae, Strombidiidae e Tintinnida. Il confronto tra i valori
misurati all’inizio (T0) e alla fine (T24) dell’incubazione, mette in evidenza un notevole
incremento della biomassa microzooplanctonica tranne per i Tintinnidi e i Metazoi
(Tab.3.4)

Antartide-Risultati
112
Microzooplancton T0 T24 Incremento µgCL-1 µgCL-1 % Strombilididi 0.79 2.05 159.5 Strombididi 3.83 11.39 197.4 Cymatocilis drigalski 0.73 1 38.1 Gyrodinium lachrima 0.66 1.39 110.6 Protoperidinium spp. 0.07 0.06 -13.2 Totale 6.09 15.84 159.9
Tab.3.4 Abbondanze dei diversi gruppi microzooplanctonici
all’inizio e alla fine dell’esperimento
La biomassa del fitoplancton totale è pari a 204.81 µg C L-1 (Tab 3.5).
C0 STD <C> k g r Pp Pr I
Fitoplancton totale 204.81 11.42 184.68 0.14 0.08 0.46 204.95 12.29 14.77 Nanoplancton totale 1.75 0.03 1.48 Fragilariopsis cylindrus 67.34 3.10 77.08 0.20 0.22 0.72 12.19 1.35 12.41 Fragilariopsis curta 123.69 8.17 141.68 0.18 0.16 0.53 21.77 1.48 23.24 Pseudonitzschia delicatissima 0.16 0.03 0.15 Pseudonitzschia heimii 1.55 0.33 1.63 Pseudonitzschia subcurvata 9.04 0.34 8.98 Phaeocystis antarctica 1.31 0.08 1.19
Tab. 3.5 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media, k= coefficiente di crescita, g=coefficiente di predazione, PP=produzione
potenziale (µg C L-1 g-1), Pr=produzione reale (µg C L-1 g-1), I=ingestione (µg C L-1 g-1)
L’analisi dei campioni all’inizio dell’esperimento (C0) ha permesso di verificare il
corretto allestimento dell’esperimento, sia relativamente al fitoplancton totale (fig. 3.7) sia
relativamente alle specie più abbondanti quali Fragilariopsis cylindrus, Fragilariopsis
curta, Pseudonitzschia delicatissima, Pseudonitzschia heimii, Pseudonitzchia subcurvata e
Phaeocystis antarctica (Tab.3.5)
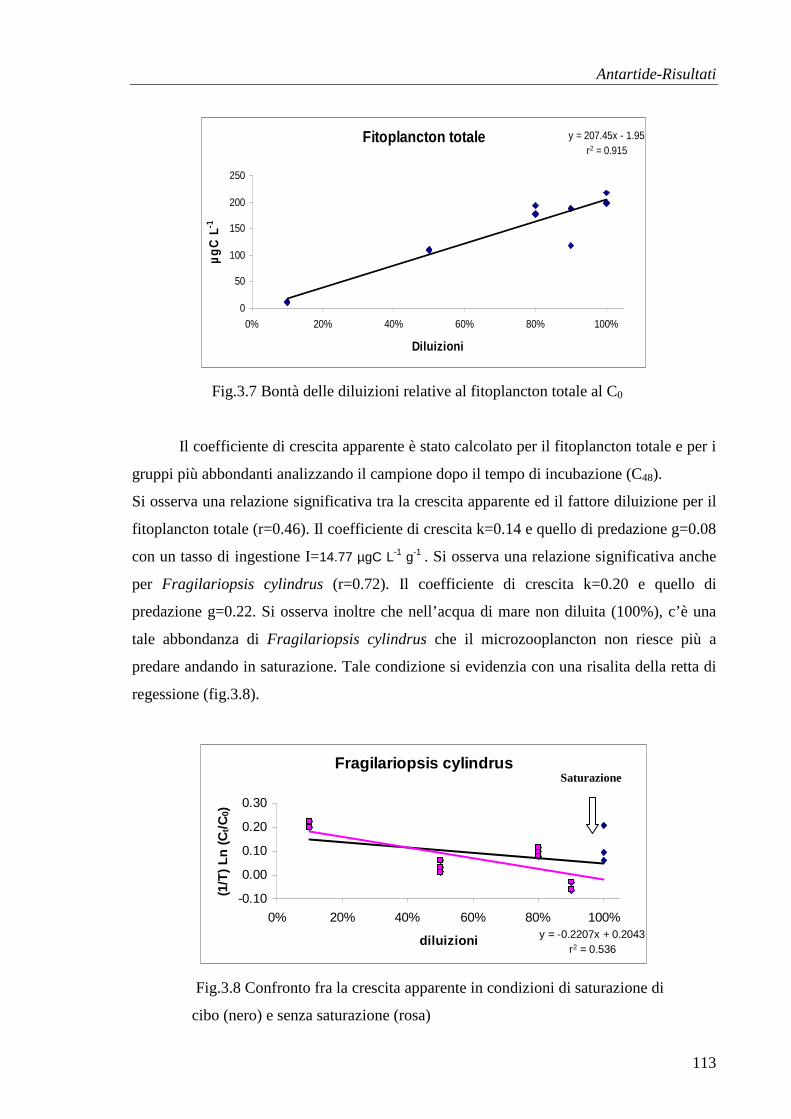
Antartide-Risultati
113
Fitoplancton totale y = 207.45x - 1.95r2 = 0.915
0
50
100
150
200
250
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.3.7 Bontà delle diluizioni relative al fitoplancton totale al C0
Il coefficiente di crescita apparente è stato calcolato per il fitoplancton totale e per i
gruppi più abbondanti analizzando il campione dopo il tempo di incubazione (C48).
Si osserva una relazione significativa tra la crescita apparente ed il fattore diluizione per il
fitoplancton totale (r=0.46). Il coefficiente di crescita k=0.14 e quello di predazione g=0.08
con un tasso di ingestione I=14.77 µgC L-1 g-1 . Si osserva una relazione significativa anche
per Fragilariopsis cylindrus (r=0.72). Il coefficiente di crescita k=0.20 e quello di
predazione g=0.22. Si osserva inoltre che nell’acqua di mare non diluita (100%), c’è una
tale abbondanza di Fragilariopsis cylindrus che il microzooplancton non riesce più a
predare andando in saturazione. Tale condizione si evidenzia con una risalita della retta di
regessione (fig.3.8).
Fragilariopsis cylindrus
y = -0.2207x + 0.2043r2 = 0.536
-0.10
0.00
0.10
0.20
0.30
0% 20% 40% 60% 80% 100%
diluizioni
(1/T
) Ln
(C
t/C0)
Fig.3.8 Confronto fra la crescita apparente in condizioni di saturazione di
cibo (nero) e senza saturazione (rosa)
Saturazione

Antartide-Risultati
114
Si verifica predazione anche nei confronti di Fragilariopsis curta (r=0.53), il coefficiente
di crescita k=0.18 e quello di predazione g=0.16.
Esperimento 4
I campioni relativi a questo esperimento sono stati raccolti in Antartide durante
l’estate australe 2000/01 al Largo del Mare di Ross.
Il popolamento iniziale è costituito da ciliati (87.57%) e dinoflagellati eterotrofi (12,43%).
Tra i ciliati sono stati distinti i gruppi Strobilidiidae, Strombidiidae e Tintinnida con la
specie Cimatocilis drigalskii tra i dinoflagellati i generi Protoperidinium e la specie
Gyrodinium lachrima (Tab 3.6)
Microzooplancton T0 T24 Incremento µgCL-1 µgCL-1 % Strombilididi 0.79 2.05 159.5 Strombididi 3.83 11.39 197.4 Cymatocilis drigalski 0.73 1.008 38.1 Gyrodinium lachrima 0.66 1.39 110.6 Protoperidinium spp. 0.076 0.066 -13.2 Totale 6.09 15.84 159.9
Tab. 3.6 Abbondanze dei diversi gruppi microzooplanctonici
all’inizio e alla fine dell’esperimento di diluizione
La biomassa fitoplanctonica all’inizio dell’esperimento (C0) è pari a 16.61 µg C L-1
(Tab.3.7)
C0 STD <C> k g r Pp Pr I Fitoplancton totale 16.61 1.80 14.45 0.11 0.50 0.681 1.78 -6.55 7.24 Corethron spp 5.30 0.70 5.74 0.82 0.86 0.799 4.36 -0.20 4.93 Fragilariopsis spp 0.54 0.07 0.34
Pseudonitzschia heimii 0.10 0.03 0.06 0.33 1.27 0.779 0.03 -
125.58 80.81 Pseudonitzschia subcurvata 5.89 0.97 4.17 0.35 0.53 0.731 2.05 -1.07 2.21 Phaeocystis antarctica 4.79 0.96 4.14 Tab. 3.7 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media, k= coefficiente di crescita, g=coefficiente di predazione, Pp=produzione potenziale (µg C L-1 g-1), Pr=produzione reale (µg C L-1 g-1), I=ingestione (µg C L-1 g-1)

Antartide-Risultati
115
L’analisi dei campioni all’inizio dell’esperimento (C0) ha permesso di verificare il
corretto allestimento dell’esperimento, sia per il fitoplancton totale (fig.3.9) che per le
specie più abbondanti quali Corethron sp., Fragilariopsis sp., Pseudonitzschia heimii,
Pseudonitzchia subcurvata e Phaeocystis antartica (Tab.3.7)
Fitoplancton totale y = 16.115x - 1.4157r2 = 0.9155
0.00
5.00
10.00
15.00
20.00
0% 20% 40% 60% 80% 100%
diluizioni
µgC
L-1
Fig.3.9 Bontà delle diluizioni relative al fitoplancton totale al C0
Si osserva una relazione significativa fra il coefficiente di crescita apparente ed il
fattore diluizione per il fitoplancton totale (r=0.80). Il coefficiente di crescita k=0.42 e
quello di mortalità da predazione g=0.36. Si osserva una relazione significativa anche nei
confronti di Corethron spp e di Navicula. Per le altre specie conteggiate nei campioni non
si verifica una correlazione significativa.
Esperimento 5
I campioni relativi a questo esperimento sono stati raccolti in Antartide durante
l’estate australe 2000/01 al largo del Mare di Ross. Il confronto delle biomassa del
popolamento microzooplanctonico all’inizio (C0) e alla fine dell’incubazione (C38)
evidenzia un notevole incremento della biomassa microzooplanctonica. Gruppi più
abbondanti risultano essere quelli dei dinoflagellati con la specie Protoperidinium spp e
Gyrodinium lachryma e quelli dei tintinnidi (Tab 3.8).

Antartide-Risultati
116
Microzooplancton T0 T24 Incremento µgCL-1 µgCL-1 % Diplopsalis sp. 0.85 1.06 25.1 Gyrodinium lachrima 5.01 8 59.71 Protoperidinium spp. 8.72 16.12 84.74 Ciliophora indet. 0.18 0.27 55.17 Strombidiidae indet. 1.55 1.39 -9.76 Tintinnidi 13.93 30.29 117.39 Metazoi (nauplii) 4.02 3.68 -8.33 Totale 34.26 60.83 77.54
Tab.3.8 Abbondanze dei diversi gruppi microzooplanctonici
all’inizio e alla fine dell’esperimento di diluizione
La biomassa fitoplanctonica all’inizio dell’esperimento (C0) è pari a 65.86 µg C L-1
(Tab.3.9)
C0 STD <C> k g r Pp Pr I Fitoplancton totale 65.86 2.53 67.69 0.42 0.36 0.80 27.59 3.95 24.30 Corethron spp 56.33 2.36 58.16 0.49 0.42 0.80 27.77 4.22 24.31 Fragilariopsis spp 6.00 0.65 5.66 Pseudonitzschia spp. 2.34 0.26 2.44 Navicula 0.04 0.01 0.04 0.54 0.49 0.84 0.02 0.00 0.02 Attheya 0.62 0.16 0.86 Nanoflagellati 0.14 0.03 0.16
Tab. 3.9 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media, k= coefficiente di crescita, g=coefficiente di predazione, Pp=produzione potenziale (µg C L-1 g-1), Pr=produzione reale (µg C L-1 g-1), I=ingestione (µg C L-1 g-1)
L’analisi dei campioni all’inizio dell’esperimento (C0) ha permesso di verificare il
corretto allestimento dell’esperimento.
Si verifica una correlazione significativa fra il coefficiente di crescita apparente ed il
fattore diluizione per il fitoplancton totale (r=0.80), il coefficiente di crescita k=0.49, e
quello di mortalità g=0.36. Tra le specie presenti vengono predate esclusivamente
Corethron sp.(r=0.80) e Navicula (r=0.84).
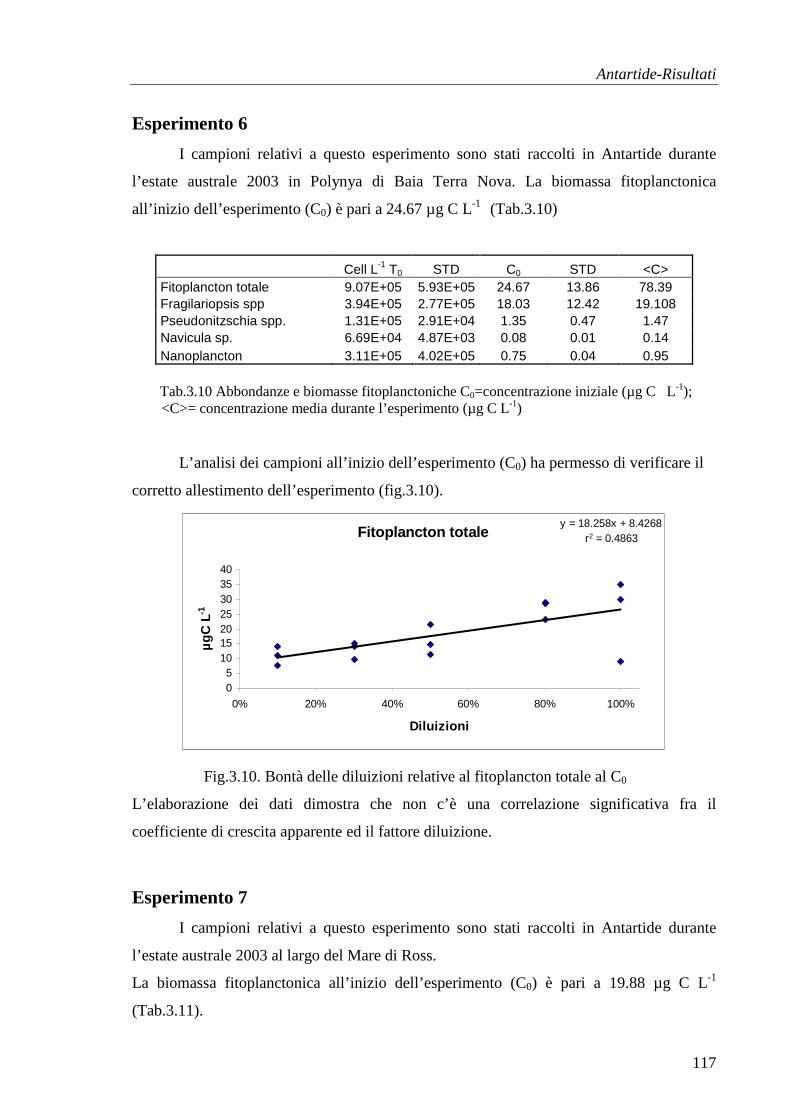
Antartide-Risultati
117
Esperimento 6
I campioni relativi a questo esperimento sono stati raccolti in Antartide durante
l’estate australe 2003 in Polynya di Baia Terra Nova. La biomassa fitoplanctonica
all’inizio dell’esperimento (C0) è pari a 24.67 µg C L-1 (Tab.3.10)
Cell L-1 T0 STD C0 STD <C> Fitoplancton totale 9.07E+05 5.93E+05 24.67 13.86 78.39 Fragilariopsis spp 3.94E+05 2.77E+05 18.03 12.42 19.108 Pseudonitzschia spp. 1.31E+05 2.91E+04 1.35 0.47 1.47 Navicula sp. 6.69E+04 4.87E+03 0.08 0.01 0.14 Nanoplancton 3.11E+05 4.02E+05 0.75 0.04 0.95
Tab.3.10 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media durante l’esperimento (µg C L-1)
L’analisi dei campioni all’inizio dell’esperimento (C0) ha permesso di verificare il
corretto allestimento dell’esperimento (fig.3.10).
Fitoplancton totaley = 18.258x + 8.4268
r2 = 0.4863
05
10152025303540
0% 20% 40% 60% 80% 100%
Diluizioni
µgC
L-1
Fig.3.10. Bontà delle diluizioni relative al fitoplancton totale al C0
L’elaborazione dei dati dimostra che non c’è una correlazione significativa fra il
coefficiente di crescita apparente ed il fattore diluizione.
Esperimento 7
I campioni relativi a questo esperimento sono stati raccolti in Antartide durante
l’estate australe 2003 al largo del Mare di Ross.
La biomassa fitoplanctonica all’inizio dell’esperimento (C0) è pari a 19.88 µg C L-1
(Tab.3.11).
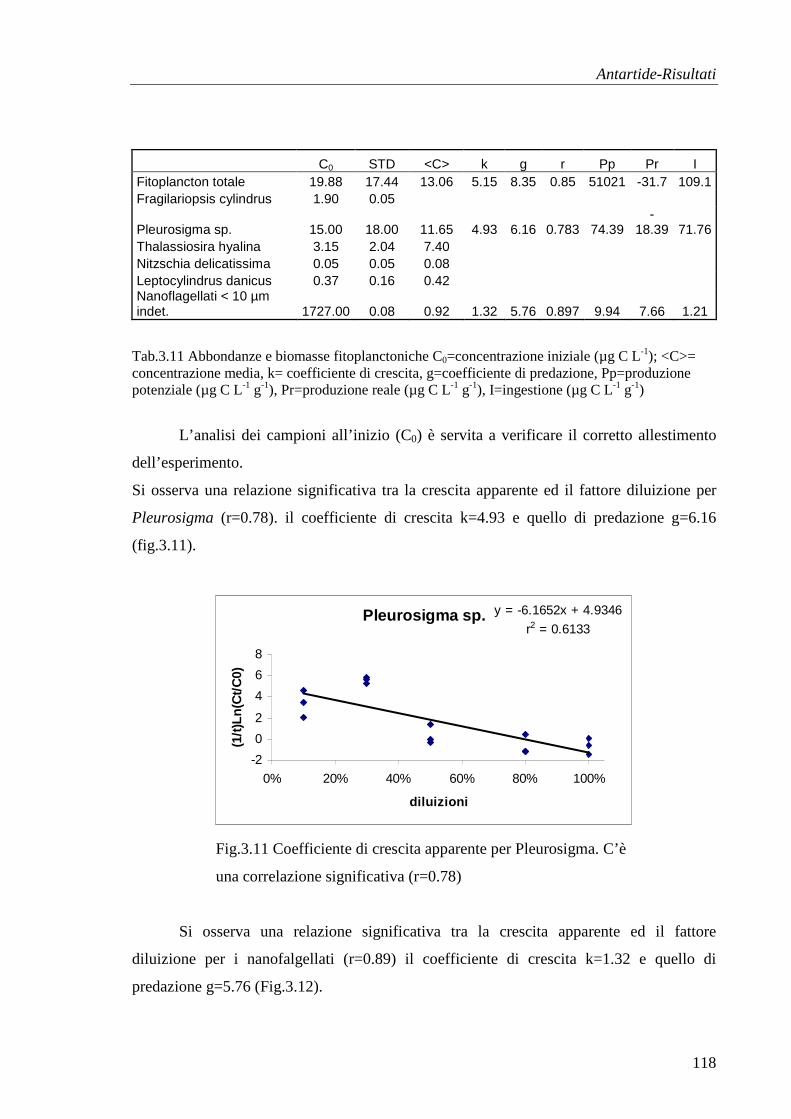
Antartide-Risultati
118
C0 STD <C> k g r Pp Pr I Fitoplancton totale 19.88 17.44 13.06 5.15 8.35 0.85 51021 -31.7 109.1 Fragilariopsis cylindrus 1.90 0.05
Pleurosigma sp. 15.00 18.00 11.65 4.93 6.16 0.783 74.39 -
18.39 71.76 Thalassiosira hyalina 3.15 2.04 7.40 Nitzschia delicatissima 0.05 0.05 0.08 Leptocylindrus danicus 0.37 0.16 0.42 Nanoflagellati < 10 µm indet. 1727.00 0.08 0.92 1.32 5.76 0.897 9.94 7.66 1.21
Tab.3.11 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media, k= coefficiente di crescita, g=coefficiente di predazione, Pp=produzione potenziale (µg C L-1 g-1), Pr=produzione reale (µg C L-1 g-1), I=ingestione (µg C L-1 g-1)
L’analisi dei campioni all’inizio (C0) è servita a verificare il corretto allestimento
dell’esperimento.
Si osserva una relazione significativa tra la crescita apparente ed il fattore diluizione per
Pleurosigma (r=0.78). il coefficiente di crescita k=4.93 e quello di predazione g=6.16
(fig.3.11).
Pleurosigma sp. y = -6.1652x + 4.9346
r2 = 0.6133
-2
0
2
4
6
8
0% 20% 40% 60% 80% 100%
diluizioni
(1/t)
Ln(C
t/C0)
Fig.3.11 Coefficiente di crescita apparente per Pleurosigma. C’è
una correlazione significativa (r=0.78)
Si osserva una relazione significativa tra la crescita apparente ed il fattore
diluizione per i nanofalgellati (r=0.89) il coefficiente di crescita k=1.32 e quello di
predazione g=5.76 (Fig.3.12).

Antartide-Risultati
119
Nanoflagellati y = -5.7631x - 1.3224
r2 = 0.8066
-8.00
-6.00
-4.00
-2.00
0.00
0% 20% 40% 60% 80% 100%
diluizioni
(1/t)
Ln
(Ct/C
0)
Fig.3.12 Coefficiente di crescita apparente per i nanoflagellati. Si osserva
una correlazione significativa (r=0.78)
Esperimento 8
I campioni relativi a questo esperimento sono stati raccolti in Antartide durante
l’estate australe 2005/06 in Polynya di Baia Terra Nova. La biomassa fitoplanctonica
all’inizio dell’esperimento (C0) è molto elevata e pari a 1.68E+04 µg C L-1 (Tab.3.12).
Cell L-1 T0 STD C0 STD <C> Fitoplancton totale 6.20E+08 6.16E+07 1.68E+04 8.59E+02 1.17E+04 Nitzschia longissima 4.18E+08 6.01E+07 8.15E+03 1.17E+03 7.64E+03 Fragilariopsis cylindrus 1.83E+08 5.03E+06 7.42E+03 3.92E+02 6.59E+03 Fragilariopsis curta 1.16E+07 1.34E+06 5.71E+02 6.62E+01 5.22E+02 Pleurosigma sp. 6.08E+06 1.07E+06 1.52E+02 2.69E+01 1.40E+02 Fragilariopsis kerguelensis 3.27E+05 1.89E+05 3.85E+01 2.22E+01 1.32E+02 Leptocylindrus danicus 4.00E+05 1.67E+05 2.42E+01 1.01E+01 3.96E+01 Nanoflagellati < 10 µm indet. 1.23E+05 6.11E+03 2.44E+04 1.21E+03 2.16E+01
Tab.3.12 Abbondanze e biomasse fitoplanctoniche C0=concentrazione iniziale (µg C L-1); <C>= concentrazione media durante l’esperimento (µg C L-1)
L’analisi dei campioni all’inizio dell’esperimento (C0) ha permesso di verificare il corretto
allestimento dell’esperimento (fig.3.13)
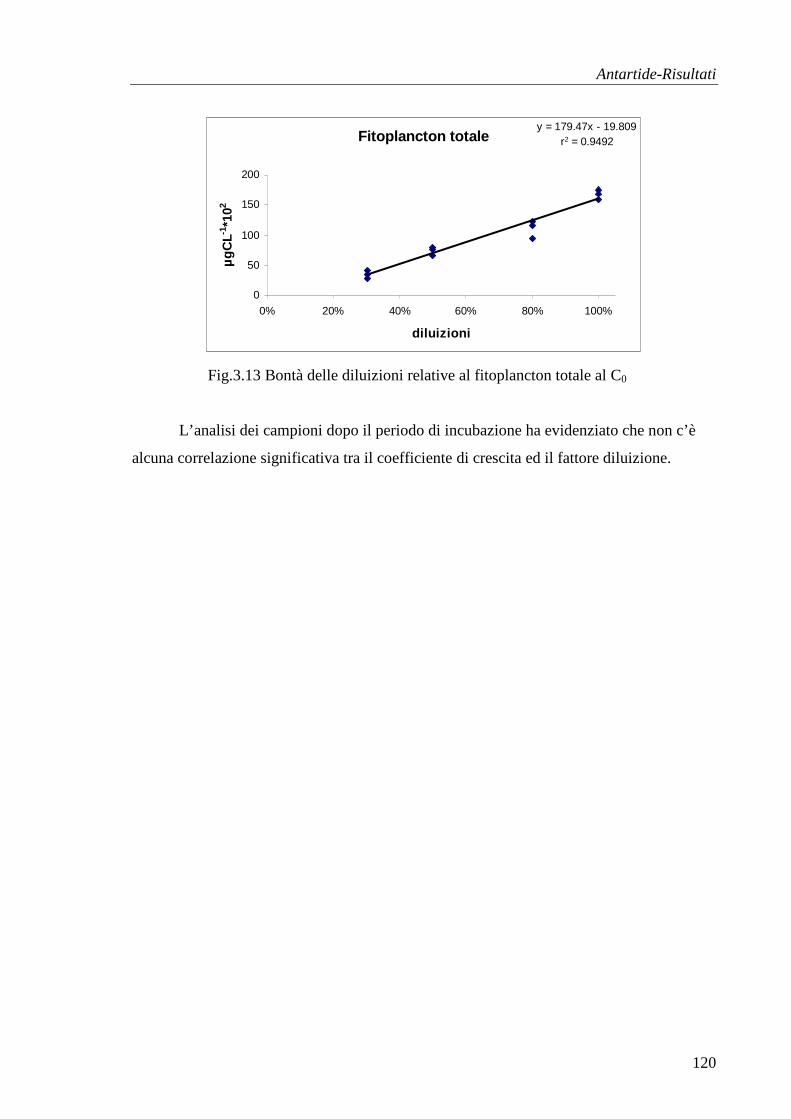
Antartide-Risultati
120
Fitoplancton totaley = 179.47x - 19.809
r2 = 0.9492
0
50
100
150
200
0% 20% 40% 60% 80% 100%
diluizioni
µgC
L-1
*102
Fig.3.13 Bontà delle diluizioni relative al fitoplancton totale al C0
L’analisi dei campioni dopo il periodo di incubazione ha evidenziato che non c’è
alcuna correlazione significativa tra il coefficiente di crescita ed il fattore diluizione.

Antartide-Discussione e Conclusioni
121
3.4 DISCUSSIONE E CONCLUSIONI
L’analisi dei dati relativi agli esperimenti condotti in Antartide nei diversi periodi
ha messo in evidenza risultati molto contrastanti. Negli esperimenti 1, 2, 6 e 8 non si
verifica predazione sul comparto fitoplanctonico. Esperimenti di diluizione sono stati
condotti da Caron et al., (2000) al centro del Mare di Ross utilizzando il metodo della
clorofilla: su 34 esperimenti eseguiti soltanto in 9 esperimenti si sono ottenuti risultati
significativi con concentrazioni di clorofilla < 1µg L-1. Solamente due esperimenti condotti
in gennaio-febbraio 1998 hanno dato un valore significativo di mortalità indotta da
predazione (g=0.26).
Negli esperimenti 1, 2, 6 e 8 il popolamento è costituito essenzialmente da
diatomee ed in particolar modo da Fragilariopsis cylindrus, Nitzschia longissima
Fragilariopsis spp. La costante presenza di Fragilariopsis nella comunità fitoplanctonica è
una caratteristica dell’estate australe nel Mare di Ross (Leventer & Dunbar, 1996; Cabrini
& Cataletto, 2000; Caron et al., 2000; Dennet et al., 2001; Fonda Umani et al., 2002,
2005). La netta dominanza di diatomee registrata è una caratteristica dell’area di
campionamento e la differente composizione floristica tra la polynya di Baia Terra Nova e
quella al largo del Mare di Ross è determinata secondo molti autori dalla profondità del
termoclino. Infatti dove la stratificazione è più superficiale, come nella zona prossima
all’area marginale per effetto dello scioglimento del pack-ice si osserva una dominanza di
diatomee. Nelle zone più al largo invece, lasciate libere dai ghiacci da più tempo la
stratificazione è più stabile ed il picnoclino più profondo sembra favorire Phaeocystis che
grazie all’aggregazione in fase palmelloide riesce a mantenersi nello strato fotico.
Le concentrazioni medie fitoplanctoniche sono molto variabili e vanno da 1.89 e
5.87 µgCL-1 registrati negli esperimenti condotti nel 1997 (esp. 1 e 2 ) a valori pari a 78.39
µgCL-1 registrati nell’esperimento 6. La concentrazione media fitoplanctonica registrata
nell’esperimento 8 è molto più alta delle altre (11.68x103 µgCL-1) e questo perché il
periodo di campionamento era caratterizzato da un’imponente fioritura fitoplanctonica.
Nonostante questa elevata concentrazione fitoplanctonica però non si verifica predazione
da parte del microzooplancton. E’ possibile che l’abbondante biomassa fitoplanctonica
riduca o come in questo caso sopprima la filtrazione da parte del microzooplancton durante
i periodi di fioritura (Caron et al., 2000).

Antartide-Discussione e Conclusioni
122
Negli esperimenti 3, 4, 5 e 7 si verifica predazione sul comparto fitoplanctonico. Il
popolamento fitoplanctonico è dominato da diatomee e più precisamente Fragilariopsis
curta, Fragilariopsis cylindrus e Pleurosigma. Le concentrazioni medie delle prede hanno
valori molto variabili, più bassi nelle stazioni al largo, pari a 13.06 µgC L-1 14.45 µgC L-1
e 78.39 µgC L-1.
Nell’esperimento 3 (Polynya di Baia Terra Nova) la concentrazione media del
fitoplancton totale è pari a 184.67 µgCL-1. L’analisi della mortalità indotta da predazione
riferita a ciascuna specie fitoplanctonica ha evidenziato un valore di g che va da 0.17 a
0.18 valori riconducibili a quelli in cui si è usata la clorofilla (Caron et al., 2000). Caron et
al., hanno condotto numerosi esperimenti durante fioriture a Phaeocystis e non hanno mai
osservato mortalità indotta da predazione microzooplanctonica. Nel nostro caso
Phaeocystis è presente con una concentrazione media pari a 1.19 µg C L-1 ma non risulta
preda gradita al microzooplancton, questo in accordo con studi precedenti (Smith et al.,
1998; Di Tullio 1998). In questo esperimento emerge che il microzooplancton, dominato
da dinoflagellati eterotrofi ed in particolar modo da Gyrodinium lachryma, da
Protoperidinium e Diplopsalis, preda preferenzialmente diatomee di piccole dimensioni
appartenenti al genere Fragilariopsis. In maniera particolare la predazione è indirizzata
verso la specie Fragilariopsis curta, molto abbondante al momento del campionamento
(<C> = 141.68 µgCL-1 ).
L’assunzione critica nell’approccio delle diluizioni è che i predatori consumano le
loro prede in proporzione diretta rispetto alla loro densità e che il livello naturale della
disponibilità di cibo è tale che i tassi d’ingestione di questi organismi non siano in
saturazione. A concentrazioni di cibo molto alte, come nel caso di Fragilariopsis curta,
questo non sarebbe vero per tutto o per parte del popolamento dei predatori. Nel caso
estremo ogni predatore mangerebbe un numero costante di prede indifferentemente
dall’effetto delle diluizioni sulla densità delle prede. Evans e Paranjape (1992) hanno
dimostrato che nel caso in cui i predatori si nutrono con tassi d’ingestione massimi saturi il
modello non lineare può adattarsi meglio ai dati ottenuti con l’esperimento delle
diluizioni. In questi grafici non-lineari, che mostrano una forma a L si osserva una
deviazione dei dati avendo una pendenza ripida vicino all’origine dell’asse delle ascisse e
nessuna pendenza man mano che ci si allontana dall’origine (fig.3.14).
Come descritto da Frost nel 1972, una concentrazione di prede può permanere nel
caso in cui il tasso di filtrazione dei predatori è massimo e il tasso di ingestione è funzione
lineare delle prede. Alle concentrazioni maggiori delle prede il tasso di ingestione può

Antartide-Discussione e Conclusioni
123
essere massimo raggiungendo il suo livello di saturazione e descrivendo un intervallo in
cui il tasso di filtrazione decresce all’aumentare delle prede (Frost, 1972). Inoltre alle basse
diluizioni, il tasso di filtrazione può essere uguale a zero descrivendo un intervallo in cui
non c’è relazione tra la risposta dei predatori e la concentrazione delle prede (Frost, 1975;
Rublee & Gallegos, 1989). Il modello non lineare sarà usato soltanto quando l’attività di
grazing alle densità maggiori delle prede raggiunge la saturazione (Moigis, 2006).
Crescita apparente
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0% 20% 40% 60% 80% 100%
Diluizioni
Ln (
Ct/C
O)
Fig.3.14 Il modello non lineare per rappresentare la condizione di saturazione di cibo
Nell’esperimento 5, condotto al Largo del Mare di Ross, la predazione è rivolta
principalmente nei confronti di Corethron spp. che è presente con una concentrazione
media pari a 5.74 µgCL-1 ed in misura minore verso Navicula, con un valore di g= 0.41.
In entrambe questi esperimenti i tassi di crescita sono sempre maggiori rispetto ai tassi di
predazione (k>g), in altre parole si verifica predazione ma questa non è comunque capace
di controllare la crescita nemmeno di quelle specie che risultano appetibili. Contrariamente
a quanto avviene in altri ambienti, dove il microzooplancton riesce a controllare lo
sviluppo quantomeno di determinati gruppi fitoplanctonici, predando qualche volta anche
più del 100% della produzione potenziale (Fonda & Beran, 2003) in ambiente antartico
questo non si verifica (Fronemann & Perissinotto, 1996). Caron et al., (2000) sostengono
che la scarsa efficienza esercitata dal microzooplancton è dovuta alle acque
particolarmente fredde quali quelle antartiche.
Nell’esperimento 4, condotto al largo del Mare di Ross, il popolamento utilizzato
per condurre gli esperimenti di diluizione è costituito da diatomee, in particolar modo da
Corethron spp. e da Phaeocystis antarctica. In quest’area di campionamento sono già state
riscontrate fioriture tardo primaverili caratterizzate dalla co-dominanza di Phaeocystis e
diatomee (Carlson et al., 1998; Caron et al., 2000; Dennett et al., 2001;). In questo

Antartide-Discussione e Conclusioni
124
esperimento il popolamento dei predatori è dominato da ciliati di grandi dimensioni quali
Strombidilidi e Strombidididi indeterminati, da Cymatocilis driglaski e dinoflagellati con
dominanza assoluta di Gyrodinium lachryma. E’ evidente che essendo il popolamento delle
prede costituito da organismi di dimensioni maggiori anche quello dei predatori subirà un
adattamento con una netta dominanza di organismi di taglia più grande. Si osserva infatti
come gli organismi di taglia più piccola quali Diplopsalis e Protoperidinium non abbiano
subito alcun aumento in biomassa dopo il periodo di incubazione. All’incremento
dimensionale del popolamento fitoplanctonico ne corrisponde uno analogo negli organismi
microzooplanctonici.
Nell’esperimento 7 condotto al largo del Mare di Ross la biomassa fitoplanctonica
media è pari a 13.06 µg C L-1 e la predazione è rivolta verso Pleurosigma (g=6.16),
diatomea di grandi dimensioni e verso Phaeocystis (g=5.76). In questo studio il
popolamento microzooplanctonico analizzato è piuttosto scarso se confrontato con studi
precedenti (Monti & Fonda Umani, 2000; Fonda Umani et al., 2000, 2005;). L’analisi dello
stesso dopo il periodo di incubazione indica che questo, costituito prevalentemente da
dinoflagellati quali Gyrodinium spp. e Gyrodinium lachryma, non è variato durante
l’esperimento. Questo sta ad indicare che i predatori hanno trovato cibo per sopravvivere
ma non a sufficienza da permettere loro di crescere durante l’incubazione. L’efficienza del
microzooplancton di predare sul comparto nanoplanctonico costituito essenzialmente da
Phaeocystis in forma libera, limita la formazione degli aggregati cellulari (forma
palmelloide) che è sicuramente una delle forme di adattamento dell’alga per sfuggire alla
pressione di predazione. Quando Phaeocystis passa in stadio palmelloide alimenta
esclusivamente il “microbial loop”, mentre il carbonio associato alle diatomee è in grado di
raggiungere i livelli trofici superiori favorendo così l’export di carbonio (Legendre &
Michaud 1998). La presenza di Phaeocystis in quest’area è confermata anche dai risultati
ottenuti nell’esperimento 4 condotto nella stessa zona di campionamento, al largo del Mare
di Ross nel 2001, confermando i risultati di lavori precedenti condotti nelle stessa area (Di
Tullio 1998; Smith & Gordon, 1997; Fonda Umani et al., 2002, 2005).
In questi due ultimi esperimenti analizzati i coefficienti di predazione sono sempre
maggiori rispetto a quelli di crescita delle prede (g>k) il che sta ad indicare un efficace top
down control esercitato dal microzooplancton sulla crescita delle prede.

Antartide-Discussione e Conclusioni
125
Lo scopo di questo lavoro è stato quello di elaborare i dati ottenuti dall’analisi di
campioni raccolti durante quattro campagne Antartiche, nella zona di Polynya di Baia
Terra Nova e nella Polynya del Mare di Ross, preparati attraverso il metodo delle
diluizioni. Questo ha permesso di verificare la mortalità indotta da predazione del
microzooplancton sul comparto fitoplanctonico arrivando alle seguenti conclusioni:
- L’analisi del popolamento microzooplanctonico ha evidenziato come le preferenze
alimentari del microzooplancton di grandi dimensioni sia indirizzato
prevalentemente verso prede (diatomee) di grandi dimensioni.
- Nelle stazioni in cui è presente Phaeocystis, anche se abbondante, questa non viene
predata, probabilmente perché poco appetibile e comunque soggetta a lisi batterica
con rimineralizzazione lungo la colonna d’acqua.
- La scarsa mortalità indotta da predazione rilevata negli esperimenti è in accordo
con studi precedenti condotti nella stessa area.
- Negli esperimenti 4 e 7 il tasso di predazione del microzooplancton supera sempre
quello di crescita delle prede (g>k) a testimonianza di un efficiente “top-down
control”: la maggior parte del carbonio organico fissato infatti viene consumato dai
protozoi eterotrofi, e successivamente respirato, da organismi le cui dimensioni
sono inferiori ai 200 µm piuttosto che da quelli appartenenti allo zooplancton di
maggiori dimensioni.
- Negli esperimenti 3 e 5 il tasso di predazione del microzooplancton non supera mai
quello di crescita delle prede (k>g) a testimoniare l’inefficienza del “top down
control”.
I risultati di questi esperimenti sono serviti a dare un contributo allo studio globale dei
flussi di carbonio all’interno del sistema pelagico in aree antartiche utilizzando il
protocollo delle diluzioni. Questo protocollo, ormai collaudato e di semplice allestimento,
non prevede alcuna manipolazione degli organismi e soprattutto, come dimostrato in
questo lavoro di tesi, può essere utilizzato con successo in ambienti diversi, da quelli
temperati a quelli più estremi come l’Antartide. Si è dimostrato inoltre valido strumento
per identificare i trasferimenti di alghe tossiche (Ostreopsis ovata) o delle sole tossine
lungo la rete trofica pelagica allo scopo di monitorare i rischi della tossicità algale nei mari
italiani.

Bibliografia
I
Bibliografia
Aberle N., Lengfellner K., Sommer U. (2007) Spring bloom succession, grazing impact
and herbivore selectivity of ciliate communities in response to winter warming.
Oecologia 150:668-681
Albright L.J., Sherr E.B., Sherr B.F., Fallon R.D. (1987) Grazing of ciliated protozoa on
free particle attached bacteria. Mar. Ecol. Prog. Ser. 38:125-129
Alcala, A.C., Alcala, L.C., Garth, J.S., Yasumura, D., Yasumoto, T. (1988) Human fatality
due to ingestion of the crab Demanda reynaudii that contained a palytoxin-like
toxin. Toxicon 26(1) 105–107
Alder V.A., Boltovskoy D. (1993) The ecology of larger microzooplankton in the Weddel-
Scotia Confluence Area: horizontal and vertical distribution patterns. J.Mar.Res.,
51: 323-344
Aligizaki, K., Nikolaidis, G. (2006) The presence of the potentially toxic genera Ostreopsis
and Coolia (Dinophyceae) in the North Aegean Sea, Greece. Harmful Algae 5(6)
717-730
Alledredge A.L., Passow U., Logan B. (1993) The abundance and significance of a class of
large, transparent organic particles in the ocean. Deep Sea Res. 401:1131-1140
Andersen V., Goutx M., Prieur L., Dolan J.R. (2008) Short scale temporal variability of
physical, biological and biogeochemical processes in the NW Mediterranean Sea:
an introduction. Biogeosciences Discussions in the access reviewed discussion
forum of Bioscences 5: 5233-5251
Anderson, D.M., White, A.W. (1992) Marine biotoxins at the top of the food chain.
Oceanus 35(3) 55-61
Archer S.D., Stelfox-Widdicombe C.E., Burkill P.H (2001) A dilution approach to quantify
the production of dimethylsulphoniopropionate and dimethylsulphide due to
microzooplankton herbivory. Aquat. Microb. Ecol. 23: 131-145
Archer SD, Leakey R.J.G., Burkill P.H., Sleigh M.A. (1996a) Microbial dynamics in
coastal waters of East Antarctica: herbivory by heterotrophic dinoflagellates. Mar
Ecol Prog Ser 139:239-255

Bibliografia
II
Archer SD, Leakey R.J.G., Burkill P.H., Sleigh M.A. (1996b) Microbial ecology of the sea
ice at a coastal Antarctic site: community composition, biomass and temporal
change Mar Ecol Prog Ser 135:179-195
Arrigo K.R., McClain C.R. (1994) Spring phytoplanktonic production in the Western Ross
Sea. Science,266: 261-264
Arrigo K.R., Robinson D.H., Worthen D.L., Dunbar R.B., DiTullio G.R., VanWoert M.,
Lizotte M.P. (1999) Phytoplankton community structure and the drawdown of
nutrients and CO2 in the Sothern Ocean. Science , 283:365-367
Ayukai T. (1996) Possible limitation of the dilution technique for estimating growth and
grazing mortality rates of picoplanktonic cyanobacteria in oligotrophic tropical
waters. J. Exp. Mar. Biol. Ecol. 198: 102-111
Azam F. (1998) Microbial Control of Oceanic Carbon Flux: the Plot Thickness. Science
280: 694-696
Azam F., Fenchel T., Field J.G., Gray J.S., Mayer-Reil L.A., Thingstad F. (1983) The
ecological role of water column microbes in the sea. Mar. Ecol. Prog. Ser. 10: 257-
263
Ballatine, D.L., Tosteson, T.R., Bardales, A.T. (1988) Population dynamics and toxicity of
natural population of benthic dinoflagellates in southwestern Puerto Rico. J. Exp.
Mar. Biol. Ecol., 119, 201-212
Båmstedt U., Gifford D.J., Irigoien X., et al. (2000) Feeding. In Harris, Wiebe R., Lenz P.,
Skjoldal H.R., Huntley M. (eds), ICES Zooplankton Methodology Manual.
Academic Press, London. pp: 297-399
Bates N.R., Hansell D.A., Carlson C.A., Gordon L.I. (1998) Distribution of CO2 species,
estimates of net community production, and air-sea CO2 exchancge in the Ross Sa
polynya. J. Geoph. Res. Vol 103, C2:2883-2896
Beers J.R., Stewart G.L. (1970) The preservation of achantharians in fixed plankton
samples. Limnol. Oceanogr. 15: 825-827
Berland B.R., Bonin D.J., Maestrini S.Y.(1980) Auzote ou phoshore? Consideration sur la
“paradoxe nutrionnel” de la mer Méditerranée. Oceanol Acta 3:135-142

Bibliografia
III
Besada, E.G., Loeblich, L.A., Loeblich, A.R., (1982) Observations on tropical, benthic
dinoflagellates from ciguateric endemic areas: Coolia, Gambierdiscus, and
Ostreopsis. B. Mar. Sci. 32(3) 723-35.
Bèthoux J., Gentili B., Morin P., Nicolas E., Pierre C., and Ruiz-Pino D (1999) The
Mediterranean Sea: A miniature ocean for climatic and environmental studies and a
key for the climatic functioning of the North Atlantic. Prog. Oceanog. 44: 131-146
Bèthoux J.P. (1981) Le phosphore et l’azote en Mer Médirranée, bilans et fertilité
potentielle. Mar Chem. 10:141-158
Bèthoux J.P., Morin P., Madec C., Gentili B.(1992) Phosphorus and nitrogen behaviour in
the Mediterranean Sea. Deep-Sea res. 39 (9):1641-1654
Booth B.C. (1993) Estimate cell concentration and biomass of autotrophic plankton using
microscopy. In “Handbook of Methods in Acquatic Microbial Ecology”, Lewis
Publisher, p. 199-205
Brescianini, C., Grillo, C., Melchiorre, N., Bertolotto, R., Ferrari, A., Vivaldi, B., Icardi,
G., Gramaccioni, L., Funari, E., Scardala, S. (2006) Ostreopsis ovata algal blooms
affecting human health in Genoa, Italy, 2005 and 2006. Eurosurveillance 11(36)
Bromwich D.H., Kurtz D.D. (1984) Katabatic wind forcing of the Terra Nova Bay
Polynya. J Geophys Res 89:3561-3572
Buck K.R., Garrison D.L. (1983) Protists from the ice-edge region of the Weddel Sea.
Deep-Sea Res. 30:1261-1277
Budillon G., Spezie G. (2000) Thermohaline structure and variability in the Terra Nova
Bay polynya (Ross Sea) between 1995-98. Antarctic Sc. 12(4):501-516
Bulit C., Diaz-Avalos C., Signoret M. et al. (2003) Spatial structure of planktonic ciliate
patches in a tropical lagoon: an application of geostatistical methods. Aquat.
Microb. Ecol. 30: 185-196
Burkill P.H. Edwards E.S., John A.W.W., Sleigh M.A. (1993 a) Microzooplankton and
their herbivorous activity in the north east Atlantic Ocea. Deep Sea Res. II 40:1179-
1200

Bibliografia
IV
Burkill P.H. Leakey R.J.G., Owens N.J.P. Mantoura R.F.C. (1993b) Synechoccocus and its
importance to the microbial food web on the north west Indian Ocean. Deep Sea
Res. II 40:773-782
Cabrini M., Cataletto B. (1999) Fioritura di diatomee nel Mare di Ross nell’estate australe
1998. Atti A.I.O.L., XIII (2) :289-295
Calbet A., Landry M.R. (1999) Mesozooplankton influences on the microbial food web:
direct and indirect trophic interactions in the oligotrophic open ocean. Limnol.
Oceanogr. 44: 1370-1380
Calbet A., Landry M.R. (2004) Phytoplankton growth, microzooplankton grazing, and
carbon cycling in marine systems. Limnol. Oceanogr. 49: 51-57
Calbet A., Saiz E. (2005) The ciliate-copepod link in marine ecosystems. Aq. Microb.
Ecol. 38: 157-167
Campbell L., Carpenter E.J. (1986) Estimating the grazing pressure of heterotrophic
nanoplankton on Synechococcus spp. using the sea water dilution and selective
inhibitor techniques. Mar. Ecol. Prog. Ser. 33: 121-129
Capriulo G.M. (1982) Feeding of field collected tintinnid microzooplankton on natural
food. Mar. Biol. 71: 73-86
Capriulo, G.M., Carpenter, E.J., (1980) Grazing by 32 to 202 µm Micro-Zooplankton in
Long Island Sound. Mar Ecol., 56: 319-326
Carlson C.A., Ducklow H.W., Hansell D.A., Smith W.O.jr (1998) Organic carbon
partitioning during spring phytoplankton blooms in the Ross sea polynya and the
Sargasso sea. Limnol.Oceanograph. 43: 375-386
Caron D.A., Dam H.G., Kremer P., Lessard E.J., Madin L.P., Malone T.C., Napp J.M.,
Peele E.R., Roman M.R., Youngbluth M.J. (1995) The contribution of
microorganisms to particulate carbon and nitrogen in surface waters of the Sargasso
Sea near Bermuda. Deep-Sea Res. 42: 943-972
Caron D.A., Dennet M.R., Lonsdale D.J., Moran D.M., Shalapyonok L. (2000)
Microzooplankton herbivory in the Ross Sea,Antartica. Deep-sea Res.II 47:3249-
3272

Bibliografia
V
Caron D.A., Lim E.L., Miceli G., Waterbury J.B., Valois F.W. (1991) Grazing and
utilization of chroococcoid cyanobacteria and heterotrophic bacteria by protozoa in
laboratory cultures and a coastal plankton community. Mar. Ecol. Prog. Ser. 76:
205-217
Chang, F.H., Shimizu, Y., Hay, B., Stewart, R., Mackay, G., Tasker, R., (2000) Three
recently recorded Ostreopsis spp. (Dinophyceae) in New Zealand: temporal and
regional distribution in the upper North Island from 1995 to 1997. New Zeal. J. M.
Fresh. Res. 34(1) 29-39
Chisholm S.W. (2000) Stirring times in the Southern Ocean. Nature 47:685-687
Choi J.W., Stoecker D.K. (1989) Effects of fixative on cell volume of marine planktonic
protozoa. Appl. Environ. Microbiol. 55: 1761-1765
Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M., Magno, S.G., Tartaglione, L.,
Grillo, C., Melchiorre, N., (2006) The Genoa 2005 outbreak. Determination of
putative palytoxin in Mediterranean Ostreopsis ovata by a new liquid
chromatography tandem mass spectrometry method. Anal., Chem., 78, 6153-6159
Ciminiello, P., Dell’Aversano, C., Fattorusso, E., Forino, M., Tartaglione, L., Grillo, C.,
Melchiorre, N., (2008) Putative palytoxin and its new analogue, ovatoxin-a, in
Ostreopsis ovata collected along the Ligurian coast during the 2006 toxic outbreak.
Am. Soc. Mass Spectrom. 19(1) 111-120
Congestri, R., Penna, A., Zingone, A., (2006) BENTOX-NET, a research and management
initiative on Ostreopsis spp. and other benthic microalgal blooms on the Italian
coast. Harmful Algae News 32 11-12
Crispi G., Mosetti R., Solidoro C., Crise A. (2001) Nutrient cycling in the Mediterranean
basins: the role of the biological pump in the trophic regime. Ecol. Model. 138:
101-114
Daley, R. J., Hobbie J.E. (1975) Direct counts of aquatic bacteria by a modified epi-
fluorescent technique. Limnol. Oceanogr. 20:875-882
Davidson A.T., Marchant H.J. (1992) protest abundance and carbon concentration during a
Phaeocystis-dominated bloom at an Antarctic coastal site. Polar Biol. 12:387-395

Bibliografia
VI
Dennett M.R., Mathot S., Caron D.A., Smith W.O.Jr, Lonsdale D.J. (2001) Abundance and
distribution of heterotrophic nano- and microplankton in the southern Ross Sea.
Deep-sea Res. II 48: 4019-40
Di Poi E., (2010) Microzooplankton grazing impact along a trophic gradient from the
Atlantic Ocean to the western mediterranean sea. Tesi di Dottorato
Di Tullio G.R., (1998) Export of Phaeocystis antarctica to deep water and sediments of
Ross Sea. Ocean Science Meeting, American Geophysical Union, Vol.78, February
9-13 (Abstr.)
Diaz F., Raimbault P., Boudjellal B., Garcia N., Moutin T., (2001) Early spring
phosphorus limitation of primary productivity in a NW Mediterranean coastal zone
(Gulf of Lion) Mar. Ecol. Prog. Ser. 211:51-62
Dolan J., Gallegos C.L., Moigis A. (2000) Dilution effects on microzooplankton in dilution
grazing experiments. Mar. Ecol. Prog. Ser. 200: 127-139
Dolan J.R. .Ritchie M.E, Ras J. (2007) The "neutral" community structure of planktonic
herbivores, tintinnid ciliates of the microzooplankton, across the SE Tropical
Pacific Ocean. Biogeosciences Discussion 4 (1): 561-593
Dolan J.R., Claustre H., Carlotti F., Plounevez S., Moutin T. (2002) Microzooplankton
diversity: relationships of tintinnid ciliates with resources, competitors and
predators from the Atlantic Coast of Morocco to the Eastern Mediterranean. Deep
Sea Res. I 49: 1217-1232
Dolan J.R., Lemée R., Gasparini S., Mousseau L., Heyndrick C. (2005) Probing diversity
in the plankton: using patterns in Tintinnids (planktonic marine ciliates) to identify
mechanisms. Hydrobiologia 555 (1): 143-157
Dolan J.R., Vidussi F., Claustre H. (1999) Planktonic ciliates in the Mediterranean Sea:
longitudinal trends. Deep Sea Res. I 46: 2025-2039
Dolan J.R., Vidussi F., Claustre H. (1999) Planktonic ciliates in the Mediterranean Sea:
longitudinal trends. Deep Sea Res. I 46: 2025-2039
Ducklow H.W, Purdie D.A. Williams P.J.LeB, Davies J.M. (1986) Bacteriplankton: Asink
for carbon in a coastal marine plankton community. Science 232:871-873

Bibliografia
VII
Ducklow H.W., Carlson C.A. (1992) Oceanic bacterial production. In: Marshal KC (ed)
Advance in Microbial Ecology, Vol. 12. Plenum Press, New York. Pp: 113-181
Dugdale R.C. Wilkerson F.P. (1988) Nutrient sources and primary production in the
eastern Mediterranean. Oceanol. Acta Special Issue, 179-184
Durando, P., Ansaldi, F., Oreste, P., Moscatelli, P., Marensi, L., Grillo, C., Gasparini, R.,
Icardi, G.,(2007) Ostreopsis ovata and human health: epidemiological and clinical
features of respiratory syndrome outbreaks from a two-year syndromic surveillance,
2005-06, in north-west Italy. Eurosurveillance 12(23)
Edler L. (1979). Recommendations on methods for marine biological studies in the Baltic
Sea: phytoplankton and chlorophyll. Baltic Mar. Biol. Publ. 5: 1-38
El-Sayed SZ (1971) On the productivity of the Southern Ocean (Atlantic and Pacific
sectors). In: Holdgate MW (ed) Antarctic Ecology. Academic, NY, pp119-135
Entz G. (1904) Quarnero A. Tintinnidai. All. Kozl., 3:121-133
Entz G. (1909) Studien über Organisation und Biologie der Tintinniden. Arch. Protistenkd.
15: 94-226
Eppley R.W., Horrigan S.G., Fuhrman J.A., Brooks E.R., Price C.C., Sellner K. (1981)
Origin of dissolved organic matter in the Southern California waters: experiments
on the role of zooplankton. Mar. Ecol. Prog. Ser. 6: 149-159
Epstein S.S., Shiaris M. (1992) Size-selective grazing of coastal bacterioplankton by
natural assemblages of pigmented flagellates, and ciliates. Microb. Ecol. 23:211-
225
Estrada M., Marrasé C., Latasa M., Berdalet E., Delgado M., Riera T. (1993) Variability of
deep chlorophyll maximum characteristics in the Northwestern Mediterranean .
Mar Ecol Prog Ser 92:289-300
Evans G.T., Paranjape M.A. (1992) Precision of estimates of phytoplankton growth and
microzooplankton grazing when the functional response of grazers may be non
linear. Mar.Ecol.Prog.Ser.80:285-290
Faust, M.A., Morton, S.L., Quod, J.P. (1996) Further SEM study of marine dinoflagellates:
the genus Ostreopsis (Dinophyceae). J. Phycol 32, 1053-1065

Bibliografia
VIII
Fenchel T. (1980) Relationship between particle size selection and clearance in suspension
feeding ciliates. Limnol. Oceanogr. 25: 735-740
Fenchel T. (1982) Ecology of heterotrophuc microflagellates. Bioenergetiscs and growth.
Mar.Ecol.Prog.Ser.8:225-31
Fenchel T. (1990) The role of protozoa in nature in terms of functional properties related to
size. Zool. Sci. 7:51-58
Ferrier- Pagès C., Rassoulzaden F. (1991) Density-dependent effects of protozoans on
specific growth rates in pico- and nanoplanktonic assemblages. Limnol. Oceanogr.
36 (4): 657-669
Ferrier- Pagès C., Rassoulzaden F. (1994) Seasonal impact of the microzooplankton on
pico- and nanoplankton growth rates in the northwest Mediterranean Sea. Mar.
Ecol. Prog. Ser. 108: 283-294
Fonda Umani S. (2000) From inorganic solution to hydrogel. Biol. Mar. Medit. 7 (1): 140-
154
Fonda Umani S., Accornero A., Budillon G., Capello M., Tucci S., Cabrini M., Del Negro
P., Monti M., De Vittor C. (2002) Particulate matter and plankton dynamics in the
Ross Sea Polyny of Terra Nova Bay during the Austral Summer 1997/98 J. Mar
Sys 36: 29-49
Fonda Umani S., Beran A. (2003) Seasonal variations in the dynamics of microbial
plankton communities: first estimates from experiments in the Gulf of Trieste,
northern Adriatic Sea. Mar. Ecol. Prog. Ser. 247: 1-16
Fonda Umani S., Beran A., ParlatoS., Virgilio D., Zollet T., De Olazabal A., Lazzarini B.,
Cabrini M., (2004) Noctiluca scintillans Macartney in the Northern Adriatic Sea:
Long-term dynamics, relationships with temperature and eutrophication, and role in
the food web. J. Plank. Res. 26:545-561
Fonda Umani S., Cataletto B. (1996) Distribuzione del microzooplancton superficiale
durante la campagna oceanografica Eolie 1995. Caratterizzazione ambientale
marina del sistema Eolie e dei bacini limitrofi di Cefalù e Gioia (EOCUMM95)
Data Report: 297-304
Fonda Umani S., Monti M. (1993) Distribuzione dei popolamenti microzooplanctonici

Bibliografia
IX
nell'Arcipelago Toscano. In: Progetto Mare. Ricerca sullo stato biologico e fisico
dell'Alto Tirreno Toscano pp: 157-260
Fonda Umani S., Monti M.,Nuccio C. (1998) Microzooplankton biomass distribution in
Terra Nova Bay Ross Sea (Antarctica) J Mar Syst 17:289-303
Fonda Umani S., Shun Cheng Yong., Feoli E., Cataletto B., Cabrini M., Milani L. (1995)
Is it possible to identify any plankton succession in the Gulf of Trieste (Northern
Adriatic Sea) In: Biology and ecology of shallow coastal waters. (Eleftheriou A.,
Ansell A.D., Smith C.J., eds) Proceedings of 28th EMBS: 59-65
Fonda Umani S., Zanon V. (2000) Prime stime dell’efficienza della predazione del
microzooplancton nel Tirreno meridionale (isole Eolie) Atti dell’AIOL 13 (2): 133-
144
Fonda Umani, S., Tirelli, V., Beran, A., Guardiani, B. (2005) Relationships between
microzooplankton and mesozooplankton: competition versus predation on natural
assemblages of the Gulf of Trieste (northern Adriatic Sea) J. Plank. Res. 27: 973-
986
Froneman P.W., Perissinotto R. (1996a) Microzooplankton grazing and protozooplankton
community structure in the South Atlantic and in the Atlantic sector of the Southern
Ocean. Deep-sea Res.I 43:703-721
Froneman P.W., Perissinotto R. (1996b) Structure and grazing of the microzooplankton
comunities of the Subtropical Convergence and a warm-core eddy in the Atlantic
sector of the Southern Ocean. Mar.Ecol.Prog.Ser. 135:237-243
Frost B.W. (1972) Effects of size and concentration of food particles on the feeding
behaviour of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr.
17: 805-815
Frost B.W. (1975) A threshold feeding behaviour in Calanus pacificus. Limnol. Oceanog.
20:263-266
Fuhrman J.A., Noble R.T. (1995). Viruses and protists cause similar bacterial mortality in
coastal seawater. Limnol. Oceanogr. 40: 1236-1242
Fuhrman J.A., Suttle C.A. (1993) Viruses in marine planktonic system. Oceanography 6:
51-63

Bibliografia
X
Fukuyo, Y., 1981. Taxonomical study on benthic dinoflagellates collected in coral reefs. B.
JPN Soc. Sci. Fish. 47(8) 967-978
Gallegos C.L. (1989) Microzooplankton grazing on phytoplankton in the Rhode River,
Maryland: nonlinear feeding kinetics. Mar. Ecol. Prog. Ser. 57: 23-33
Gallitelli, M., Ungaro, N., Addante, L.M., Procacci, V., Silveri, N.G., Sabbà, C. (2005)
Respiratory illness as a reaction to tropical algal blooms occurring in a temperate
climate. J. Am. Med. Ass. 293(21) 2599-2600
Garrison D.L., Buck K.R. (1989) Protozooplankton in the Weddel Sea, Antarctica:
abundance and distribution in the ice edge zone. Polar Biol 9:341-351
Garrison D.L., Gowing M.M. Hughes M.P. (1998) Nano and microplankton assemblages
in the Northern Arabian Sea during the South West monsoon, august September
1995: a US-JGOFS study. Deep Sea Res. II 45:2269-2299
Gasol J.M., Del Giorgio P., Duarte C.M. (1997) Biomass distribution in marine planktonic
communities. Limnol. Oceanogr. 42 (6): 1353-1363
Gast V., (1985) bacteria as a food resource for microzooplancton in the Schlei fjord and
Baltic Sea with special references to ciliates. Mar. Ecol. Prog. Ser. 22:107-120
Geraci, J.R., Anderson, D.M., Timperi, R.J., Aubin, D. J. S., Early, G.A., Prescott, J.H.,
Mayo, C.A., (1989) Humpback whales (Megaptera novaeangliae) fatally poisoned
by dinoflagellate toxin. Can. J. Fish. Aquat. Sci. 46(11) 1895-1898
Gifford D.J. (1991) The protozoan-metazoan trophic link in pelagic ecosystems. J.
Protozool. 38: 81-86
Gifford D.J., Caron D. (2000) Sampling, preservation, enumerations and biomass of
marine protozooplankton. In: Harris R.P.; Wiebe P.H., Lenz J., Skjodeal H.R.,
Huntley M. (eds.). ICES Zooplankton method. Manual. Academic Press pp: 193-
221
Gifford D.J., Fessenden L.M., Garrahan P.R., Martin E. (1995) Grazing by
microzooplankton and mesozooplankton in the high-latitude North Atlantic Ocean:
spring versus summer dynamics. J. Geophys. Res. C4 100: 6665-6675

Bibliografia
XI
Gifford, D.J., Dagg, M.J., (1990) The microzooplancton – mesozooplancton link:
consumption of planktonic Protozoa by the calanoid copepods Acartia tonsa Dana
and Neocalanus pulchrus Murukawa. Mar. Microb. Food Webs, 5: 161-177
Goldman J.C. (1984) Oceanic nutrient cycles In: Fasham M.J.R. (ed). Flows of energy and
materials in marine ecosystems: theory and practice. Plenum Press, New York. pp:
137-170
González H.E., Sobarzo M., Figueroa D., Nothig E.M. (2000) Composition, biomass and
potential grazing impact of the crustacean and pelagic tunicates in the northern
Humboldt Current area off Chile: differences between El Niño and non-El Niño
years. Mar. Ecol. Prog. Ser. 195: 201-220
González J.M. (1996) Efficienti size-selective bacterivory by phagotrophic nanoflagellates
in aquatic system. Mar. Biol. 126:785-798
Granéli, E., Ferreira, C.E.L., Yasumoto, T., Rodrigues, E., Neves, M.H.B., (2002) Sea
urchins poisoning by the benthic dinoflagellate Ostreopsis ovata on the Brazilian
coast. Book of Abstracts, 10th International Conference on Harmful Algae, Florida,
USA, pp. 286
Guerrini, F., Pezzolesi, L., Feller, A., Riccardi, M., Ciminiello, P., Dell’Aversano., C.,
Tartaglione, L., Dello Iacovo, E., Fattorusso, E., Forino, M., Pistocchi, R., (2010)
Comparative growth and toxic profile of cultured Ostreopsis ovata from the
Tyrrhenian and Adriatic Seas. Toxicon 55(2-3) 1-10
Hansen B., Bjørnsen P.K., Hansen P.J. (1994) The size ratio between planktonic predators
and their prey. Limnol. Oceanogr. 39: 395-403
Hansen P.J. (1991) Quantitative importance and trophic role of heterotrophic
dinoflagellates in a coastal pelagic food web. Mar. Ecol. Prog. Ser. 73: 253-261
Hansen P.J., Calado A. J. (1999) Phagotrophic mechanisms and prey size selection in free-
living dinoflagellates. J. Eukaryot. Microbiol. 16: 382-389
Hanson R.B., Ducklow H.W., Field J.G (2000) The changes Ocean carbon cycle. A
midterm synthesis of the JGOFS. Cambridge Univ.Press, Cambridge,p 514
Heimdal B.R. (1997) Modern coccolithophorids. In: Tomas C.R. (ed.), Identifying Marine
Phytoplankton. Academic Press, San Diego, pp: 731-833

Bibliografia
XII
Heinbokel J.F. (1978a). Studies on the functional role of tintinnids in the Southern
California Bight. I. Grazing and growth rates in laboratory cultures. Mar. Biol. 47:
177-189
Heinbokel J.F. (1978b) Studies on the functional role of tintinnids in the southern
California Bight. II. Grazing rates of field populations. Mar. Biol. 47: 191-197
Heinbokel J.F., Beers J.R. (1979) Studies on the functional role of tintinnids in the
Southern California Bight. III. Grazing impact of natural assemblages. Mar. Biol.
52: 23-32
Hensen V. (1887) Uber die Bstimmung des Planktons oder des im Meere treibenden
Materials an Pflanzen und Tieren, V. Bericht der Commission zur
wissenschaftlichen untersuschung der deutschen Meere in Kiel, Paul Parey, Berlin
p.108
Hewes C.D. Sakshaug E., Reid F.M.H., Holm-Hansen O. (1990) Microbial autotrophic and
etherotrophic eukaryotes in Antarctic waters: relationship between biomass and
chlorophyll, adenosine triphosphate and particulate organic carbon. Mar. ecol Prog.
Ser., 63:27-35
Hewes C.D., Holm-Hansen O., Sakshaug E. (1985) Alternative pathways al lower trophic
levels in the Antarctic food web. In: Siegfried W.R., Condy P., Laws R.M., (eds)
Antarctic Nutrient Cycles and Food Webs, Proceedings of the 4th SCARSymposium
on Antarctic Biology. Springer, Berlin pp 283
Honjio S., Roman M.R. (1978) Marine copepod fecal pellets: production, preservation and
sedimentation. J. Mar. Res. 36: 45-57
Hurbungs, M.D., Jayabalan, N., Chineah, V. (2001) Seasonal distribution of potentially
toxic benthic dinoflagellates in the lagoon of Trou aux Biches, Mauritius. AMAS
Food and Agricultural Research Council, Réduit, Mauritius
Innamorati M., Lazzara L., Massi L., Mori C., Nuccio C., Saggiomo V. (1991)
Phitoplankton ecology. Nat. Sc. Com. Ant. Ocean Camp 1989-1990. Data Rep
1:141-252
Innamorati M., Mori C., Massi L., Lazzara L., Nuccio, C. (1999) Phytoplankton biomass
related to environmental factors in the Ross Sea. Springer-Verlag, Berlin, pp. 235-
252

Bibliografia
XIII
Irigoien X., Flynn K.J., Harris R.P. (2005) Phytoplankton blooms: a "loophole" in
microzooplankton grazing impact? J. Plank. Res. 27: 313-321
James M.R., Hall J.A. (1998) Microzooplankton grazing in different water masses
associated with the Subtropical Convergence round the South Island, New Zealand.
Deep-Sea Res. I 45: 1689-1707
Johansson M., Gorokhova E., Larsson U. (2004) Annual variability in ciliate community
structure, potential prey and predators in the open northern Baltic Sea proper. J.
Plank. Res. 26: 67-80
Jonsson P.R. (1986) Particle size selection, feeding rates and growth dynamics of marine
planktonic oligotrichous ciliates. Mar. Ecol. Prog. Ser. 33:265-277
Jørgensen E. (1924) Mediterranean Tintinnidae. Rep. Dan. oceanogr. Exped. Mediterr.
2 (J3): 1-110
Jørgensen E. (1927) Cialiata: Tintinnida. – In: Gimpe G., Remane A (eds). Die Tierwelt
der Nordund Ossee 2: 1-26 Gustav Fisher Verlag, Jena
Karayanni H., Christaki U., Van wambeke F., Dalby A.P. (2004) Evaluation of double
formalin Lugol’s fixation in assessing number and biomass of ciliates: an example
of estimations at mesoscale in NE Atlantic. J. Microbiol. Met. 56: 349-358
Kiørboe T. (1993) Turbulence, phytoplankton cell size, and the structure of pelagic food
webs. Advances in Marine Biology 29: 1-72
Kiørboe T. (1996) Material flux in the water column. In: Eutrophication in Coastal Marine
Ecosystem. Coastal and Estuarine Studies 52: 67-94
Kivi K. (1996) On the ecology of planktonic microprotozoans in the Gulf of Finland,
northern Baltic Sea. W.A. de Nottbeck Foundation Sci. Rep. 11: 1-36
Yliopistopaino, Helsinki
Kivi K., Setälä, O. (1995) Simultaneous measurement of food particle selection and
clearance rates of planktonic oligotrich ciliates (Ciliophora: Oligotrichina). Mar.
Ecol. Prog. Ser. 119: 125-137
Kleppel G.S. (1993) On the diets of calanoid copepods. Mar. Ecol. Prog. Ser. 99: 183-195
Kofoid C.A., Campbell A.S. (1929) A conspectus of the marine and freshwater ciliate
belonging to the suborder Tintinnoinea, with descriptions of new species

Bibliografia
XIV
principally from the Agassiz expedition to the eastern tropical Pacific 1904-1905.
Univ. Calif. Publ. Zool. 23: 1-403
Kofoid C.A., Campbell A.S. (1939) The ciliate: tintinnoinea. Bulletin of the Museum of
Comparative Zoology at Harvard college vol. LXXXIV. Cambridge, Mass, U.S.A.
pp: 1-471
Kopczynska E.E., Goeyens L., Semeneh M., Dehairs F. (1995) Phytoplankton composition
and cell distribution in Prydz Bay, Antarctica. J Plankton Res, 17 (4):685-707
Krom, M.D., Kress N., Brenner S., Gordon L.I., (1991) Phosphorus limitation of primary
productivity in the eastern Mediterranean Sea. Limnol. and Oceanogr. 36: 424-432
Kuipers B.R., Witte H.J. (1999) Grazing impact of microzooplankton on different size
classes of algae in the North Sea in early spring and mid-summer. Mar. Ecol. Prog.
Ser. 180: 93-104
Laackmann H. (1913) Adriatische Tintinnodeen. Sber. Akad. Wiss. Wwn. Kl. math. -
natunv. 122: 1-45
Laakmann H (1907) Antarktische Tintinnen. Zool Antarct 31:235-239
Landry M.R. (1993) Estimating rates of growth and grazing mortality of phytoplankton by
the dilution method. In: Kemp P.F., Sherr B., Sherr E., Cole J.J. (eds) Handbook of
methods in aquatic microbial ecology. Lewis Publishers, Ann. Arbor. MI. pp: 715-
772
Landry M.R., Hassett R.P. (1982). Estimating the grazing impact of marine
microzooplankton. Mar. Biol. 67: 283-288
Landry M.R., Kirsctein J., Constantinou J. (1995) A refined dilution technique for
measuring the community grazing impact of microzooplankton, with experimental
tests in the central equatorial Pacific. Mar. Ecol. Prog. Ser. 120: 53-63
Latasa M. Landry M.R, Schlüter L., Bidigare R.R. (1997) Pigment-specific growth and
grazing rates of phytoplankton in the Central Equatorial Pacific. Limnol. Oceanogr.
42 (2): 289-298
Leakey R.J.G., Burkill P.H., Sleigh M.A. (1994) A comparison of fixatives for the
estimation of abundances and biovolume of marine planktonic ciliate populations.
J. Plank. Res. 16 (4): 375 -389

Bibliografia
XV
Leakey R.J.G., Fenton N., Clarke A. (1996) The annual cycle of planktonic ciliates in the
nearshore waters at Signy Island, Antarctica J. Plan. Res. Vol 16, 7:841-856
Legendre L. (1996) The biological CO2 pump in seasonally ice-covered waters.Proc.NIPR
Symp.Polar Biol.9:61-74
Legendre L., Le Fèvre J. (1992) Interactions between hydrodynamics and pelagic
ecosystems: relevance to resource exploitation and climate change. IJ Mar. Sci
12:477- 486
Legendre L., Michaud (1998) Flux of biogenic carbon in oceans :size-dependent regulation
by pelagic food webs. Mar.Ecol.Prog.Ser. 164:1-11
Legendre L., Rassoulzadegan F. (1995) Plankton and nutrient dynamics in marine waters.
Ophelia 41: 153-172
Lenoir, S., Ten-Hage, L., Turquet, J., Quod, J.P., Bernard, C., Hennion, M.C. (2004) First
evidence of palytoxin analogues from an Ostreopsis mascarenensis (Dinophyceae)
benthic bloom in Southwestern Indian Ocean. J. Phycol., 40: 1042-1051
Lessard E.J. (1991) The trophic role of heterotrophic dinoflagellates in diverse marine
environments. Mar. Microb. Food Webs 2: 49-58
Lessard E.J., Murrell M.C. (1996) Distribution, abundance and size composition of
heterotrophic dinoflagellates and ciliates in the Sargasso Sea near Bermuda. Deep-
Sea Res. 43: 1045-1065
Lessard E.J., Murrell M.C. (1996). Distribution, abundance and size composition of
heterotrophic dinoflagellates and ciliates in the Sargasso Sea near Bermuda. Deep-
Sea Res. 43: 1045-1065
Leventer A., Dunbar R.B. (1996) Factoring influencing the distribution of diatoms and
other algae in the Ross Sea. J.Geogh Res. 101:18489-18500
Litaker, R.W., Tester, P.A (2002) Molecular methods for detecting and characterizing
harmful phytoplankton. In: Hurst, C.J., Crawford, R.L., Knudsen, G.R., McInerney,
M., Stetzenbach, L.D. (Eds), Environmental microbiology. Seconda edizione, ASM
Press, New York, pp. 342-353

Bibliografia
XVI
Liu H., Dagg M.J., Wu C.J., Chiang K.P. (2005) Mesozooplankton consumption of
microplankton in the Mississippi River plume, with special emphasis on planktonic
ciliates. Mar. Ecol. Prog. Ser. 286: 133-144
Mangialajo, L., Bertolotto, R., Cattaneo-Vietti, R., Chiantore, M., Grillo, C., Lemee, R.,
Margalef R. (1985) Introduction to the Mediterranean. Western Mediterranean,
Pergamon Press., chapter 1: 1-16
Margalef R. (1963) Role des Cilies dans le cycle de la vie pelagique en Mediterranee.
Rapp. Proc. Verb. Reun. CIESM, 17(2):511-512
Marshall, S.M. (1973) Respiration and feeding in copepods. Advances in Marine Biology
11: 57-120
McManus G.B., Ederington-Cantrell M.C. (1992) Phytoplankton pigments and growth
rates, and microzooplankton grazing in a large temperate estuary. Mar. Ecol. Prog.
Ser. 87: 77-85
Mebs, D., 1998. Occurrence and sequestration of toxins in food chain. Toxicon 36 (11):
1519-1522
Melchiorre, N., Moretto, P., Povero, P., Ruggieri, N. (2008) The toxic benthic
dinoflagellate Ostreopsis ovata: quantification of proliferation along the coastline
of Genoa, Italy. Mar. Pollut. Bull. 56(6) 1209–1214
Menden-Deuer S., Lessard E.J. (2000) Carbon to volume relationship for dinoflagellates,
diatoms and other protest plankton. Limnol. Oceanogr. 45: 596-579
Michaels A.F., Caron D.A., Swanberg N.R., Howse, F.A., Michaels C.M. (1995)
Planktonic sarcodines (Acantharia, Radiolaria, Foraminifera) in surface waters near
Bermuda: abundance, biomass and vertical flux. J Plank. Res. 17: 131-163
Miller C.B. (1993) Pelagic production processes in the Subartic Pacific. Prog. In Oceanog.
32:1-15
Modigh M- Castaldo S. (2002) Variability and persistence in tintinnids assemblages at a
Mediterranean coastal site. Aquat. Microb. Ecol. 28: 299-311
Modigh M., Castaldo S. (2005) Effect of fixatives on ciliates as related to cell size. J.
Plank. Res. 27: 845-849

Bibliografia
XVII
Moigis A.G. (2006) The clearance rate of microzooplankton as the key element for
describing estimated non-linear dilution plots demonstrated by a model. Mar.
Biol.149: 743–762
Montagnes D.J.S., Lynn D.H., Roff J.C., Taylor W.D. (1988) The annual cycle of
heterotrophic planktonic Ciliates in the waters surrounding the Isle of Shoals, Gulf
of Maine: an assessment of their trophic role. Mar. Biol. 99:21-30
Monti M., Fonda Umani S. (2000) Distbution of the main microzooplankton taxa in the
Ross Sea (Antarctica): Austral Summer 1994 In: Faranda F.M., Guglielmo L.,
Ianora A. (eds) Ross sea Ecology. Springer-Verlag, Berlin Heidelberg pp275-289
Monti, M., Minocci, M., Beran, A., Iveša, L., (2007) First record of Ostreopsis cfr. ovata
on macroalgae in the Northern Adriatic Sea. Mar. Pollut. Bull. 54(5) 598-601
Nejstgaard J.C., Gismervik I. Solberg P.T. (1997) Feeding and reproduction by Calanus
finmarchicus, and microzooplankton grazing during mesocosm blooms of diatoms
and the coccolithophore Emiliana huxleyi. Mar. Ecol. Prog. Ser. 147: 197-217
Nejstgaard J.C., Naustvoll L.J., Sazhim A. (2001a) Correcting for underestimation of
micrzooplankton grazing in incubation experiments with mesozooplankton. Mar.
Ecol. Prog. Ser. 221: 59-75
Neudruck Asher A. and Co. (1929) Nortisches plankton zoologischer tiel Protozoa. Pp: 1-
145
Paranjape M.A., (1990) Microzooplankton herbivory on the grand Bank (Newfoundland
Canada) A seasonal study. Mar. Biol. 107:321-328
Penna, A., Vila, M., Fraga, S., Giacobbe, M.G., Andreani, F., Riobò, P., Vernesi, C.,(2005)
Charaterization of Ostreopsis and Coolia (Dinophyceae) isolates in the Western
Mediterranean Sea based on morphology, toxicity and internal transcribed spacer
5.7s rDNA sequences. J. Phycol., 41: 212-225
Pérez M.T., Dolan J.R., Vidussi F., Fukai E.(2000) Dial vertical distribution of planktonic
ciliates within the surface layer of the NW Mediterranean (May 1995). Deep-Sea
Res. I 47: 479-503
Pernthaler J. (2005) Predation on prokaryotes in the water column and its ecological
implications. Nature reviews. 3:537-546

Bibliografia
XVIII
Pierce R.W., Turner J.T. (1993) Global biogeography of marine tintinnids. Mar. Ecol.
Prog. Series 94: 11-26
Pierce, R.W., Turner, J.T., (1992) Ecology of planktonic Ciliates in Marine Food Webs.
Rewieus in Aquatic Sciences. 6(2): 139-181
Pinardi N., Masetti E. (2000) “Variability of large scale general circulation of the
Mediterranean Sea from observations and modeling: a review.” Palaeogeography,
Palaeoclimatology, Palaeoecology 158: 153-173
Pitta P., Giannakourou A. (2000) Planktonic ciliates in the oligotrophic Eastern
Mediterranean: vertical, spatial distribution and mixotrophy. Mar. Ecol. Prog. Ser.
194: 269-282
Pitta P., Giannakourou A., Christaki U. (2001) Planktonic ciliates in the oligotrophic
Mediterranean Sea: longitudinal trends of standing stock, distributions and analysis
of food vacuole contents. Aquat. Microb. Ecol. 24: 297-311
Pomeroy L.R. (1974) The ocean’s food web, a changing paradigm. BioSci. 24 (9): 499-504
Popels, L.C., Cary, S.C., Hutchins, D.A., Forbes, R., Pustizzi, F., (2003) The use of
quantitative polymerase chain reaction for the detection and enumeration of the
harmful alga Aureococcus anophagefferens in environmental samples along the
United States East coast. Limnol. Ocean. Methods 1: 92-102
Popofsky A (1909) Die Radiolarien der Antarktis. Dtsch Sudpolar Exped 10:306
Porter K.G., Feig Y.S. (1980) The use of DAPI for identifying and counting aquatic
microflora. Limnol. Oceanogr. 25: 943-948
Putt M., Stoecker D.K. (1989) An experimentally determined carbon: volume ratio for
marine “oligotrichous” ciliates from estuarine and coastal waters. Limnol.
Oceanogr. 34 (6): 1097-1103
Ramos V., Vasconcelos, V., 2010. Palytoxin and analogs: biological and ecological effects.
Mar. Drugs 8(7) 2021-2037
Rampi L. (1948) I Tintinnoidi delle acque di San Remo. II. Osservazioni e conclusioni.
Boll. Pesca Piscic. Idrobiol. 3: 50-56
Rampi L. (1950) I Tintinnoidi delle acque di Monaco raccolti dall' Eider nell'anno 1913.
Bull. Inst. océanogr. Monaco 965: 1-7

Bibliografia
XIX
Rassoulzadegan F. (1978) Dimensions et taux d’ingestion des particules consommés par un
tintinnide: Favella ehrenbergii. Ann. Inst. Océanog. Paris 54: 17-24
Rassoulzadegan F. (1993) Protozoan patterns in the Azam-Ammerman’s bacteria-
phytoplankton mutualism. In: Guerrero R., Perdo-Aliò C. (eds). Trends in Microb.
Ecol. pp: 435-439
Rassoulzadegan F., Etienne M. (1981) Grazing rate of the tintinnid Stenosemella
ventricosa (Clap. & Lachm.) Jörg. on the spectrum of naturally occurring
particulate matter from a Mediterranean neritic area. Limnol. Oceanogr. 26: 258-
270
Rassoulzadegan F., Laval-Peuto M., Sheldon R.W. (1988) Partitioning of the food ration of
marine ciliates between pico- and nanoplankton. Hydrobiologia 159: 75-88
Reckermann M., Veldhuis M.J.W. (1997) Trophic interactions between picoplankton and
micro- and nanoplankton in the western Arabian Sea during the NE monsoon 1993.
Aquat. Microb. Ecol. 12: 263-273
Redfield A.C., Ketchum B.H., Richards F.A., (1963). The influence of organisms on the
composition of seawater. Hill, M.N.(Ed.), The Sea, vol. 2. Interscience, New York,
pp. 26– 77
Rhodes, L., 2010 (in press). World-wide occurrence of the toxic dinoflagellate genus
Ostreopsis Schmidt. Toxicon.
Ribera d’Alcalà M., Civitarese G., Conversano F., Cavezza R. (2003) Nutrien ratios and
fluxes hint in overlooked processes in the Mediterranean Sea. Jour. of Geoph.
Res.108 NO. C9 8106
Roether W., Klein B., Manca B.B., Theocharis A., Kioroglou (2007) Transient Eastern
Mediterranean deep waters in response to the massive dense-water output of the
Aegean Sea in the 1990s. Progr. In Ocean. 74:540-571
Rollwagen Bollens G.C., Penry D.L. (2003) Feeding dynamics of Acartia spp. copepods in
a large, temperate estuary (San Francisco Bay, CA). Mar. Ecol. Prog. Ser. 257: 139-
158
Roman M., Smith S., Wishner K., Zhang X., Gowing M. (2000) Mesozooplankton
production and grazing in the Arabian Sea. Deep-sea Res. II 47: 1423-1450

Bibliografia
XX
Roman M.R., Gauzens A.L. (1997) Copepod grazing in the equatorial Pacific. Limnol.
Oceanogr. 42: 623-634
Rublee P., Gallegos C. (1989) Use of fluorescently labelled algae (FLA) to estimate
microzooplancton grazing. Mar Ecol. Prog. Ser. 51:221-227
Sakka Hlaili A., Grami B., Hassine Hadj Mabronk, Gosselin M., Hamel D. (2007)
Phytoplankton growth and microzooplancton grazing rates in a restricted
Mediterranean lagoon (Bizerte Lagoon, Tunisia). Mar. Biol 151:767-783
Sansoni, G., Borghini, B., Camici, G., Casotti, M., Righini, P., Rustighi, C. (2003)
Fioriture algali di Ostreopsis ovata (Gonyaulacales: Dinophyceae): un problema
emergente. Biol. Amb. 17(1) 17-23
Schlüter L. (1998) The influence of nutrient addition on growth rates of phytoplankton
groups, and microzooplankton grazing rates in a mesocosm experiment. J. Exp.
Mar. Biol. Ecol. 228: 53-71
Scholin, C.A., Villac, M.C., Buck, K.R., Krupp, J.M., Powers, D.A., Fryxell, G.A. (1994)
Ribosomal DNA sequences discriminate among toxic and non-toxic Pseudo-
nitzschia species. Natural Toxins 2: 152-165
Schroder O (1909) Sticholonche zanclea und Wagnerella borealis. Deut Sudpolar Exp
10:317-322
Sellner, K.G., Doucette, G.J., Kirkpatrick, G.J. (2003) Harmful algal blooms: causes, impacts
and detection. J. Industrial Microbiol. Biotech., 30: 383-406
Setälä O., Kivi K. (2003) Planktonic ciliates in the Baltic Sea in summer: distribution,
species association and estimated grazing impact. Aquat. Microb. Ecol. 32: 287-297
Shears, N.T, Ross, P.M. (2009) Blooms of benthic dinoflagellates of the genus Ostreopsis;
an increasing and ecologically important phenomenon on temperate reefs in New
Zealand and worldwide. Harmful Algae 8(6) 916–925
Sherr E.B., Sherr B.F. (1987) High rates of consumption of bacteria by pelagic ciliates.
Nature 325:710-711
Sherr E.B., Sherr B.F. (1993) Preservation and storage of samples for enumerations of
heterotrophic protists. In : Kemp P.F., Sherr B.F., Sherr E.B., Cole J.J. (eds).

Bibliografia
XXI
Handbook of Method in Aquatic Microbial Ecology Lewis Publishers, New York
pp: 207-212
Sherr E.B., Sherr B.F., Paffenhöfer A. (1986) Phagotrophic protozoa as food for metazoans
a “missing” trophic link in marine pelagic food webs? Mar. Microb. Food Webs 1:
61-80
Sherr, E.B., Sherr, B.F., Fallon, R.D., Newell, S.Y. (1986) Small aloricate ciliates as a
major component of the marine heterotrophic nanoplancton. Limnol.Oceanog., 31:
177-183
Sieburth J.M., Smetacek V., Lenz J. (1978) Pelagic ecosystem structure: heterotrophic
compartments of the plankton and their relationship to plankton size fraction.
Limnol. Oceanogr. 23: 1256-1263
Simek K., Chrzanowski T. (1992) Direct and indirect evidence of size-selective grazing on
pelagic bacteria by freshwater nanoflagellates. Appl. Environ. Microbial 58:3715-
3720
Simoni, F., Di Paolo, C., Gori, L., Lepri, L. (2004) Further investigation on blooms of
Ostreopsis ovata, Coolia monotis, Prorocentrum lima, on the macroalgae of
artificial and natural reefs in the Northern Tyrrhenian Sea. Harmful Algae News 26
5-7
Siokou-Frangou I., Cristaki U., Mazzocchi M.G., Montresor M., Ribera d’Alcalà M.,
Vaquè D., Zingone A. (2010) Plankton in the open Mediterranean Sea: a review.
Biogeosc. 7:1543-1586
Smith Jr W.O., Nelson D.M., Di Tullio G.R., Levender A.R. (1996) Temporal and spatial
patterns in the Ross Sea : phytoplankton biomass, elemental composition
productivity and growth rates. Journal of Geophys.Res.101:18455-18466
Smith W.O., Craig A.C., Ducklow H., Hansell D.A. (1998) Growth dynamics of
Phaeocystis antartica dominated plankton assemblages from the Ross Sea.
Mar.Ecol.Prog.Ser. 168: 220-244
Smith W.O., Gordon L.I., (1997) Hyperproductivity of the Ross Sea (Antarctica) polynya
during austral spring. Geophysical Research Letters 24: 233-236
Sournia, A. (1978) Phytoplankton manual. Monographs on oceanographic methodology.
Voume 6. UNESCO, Parigi, pp 337

Bibliografia
XXII
Spittler P. (1973) Feeding experiments with tintinnids. Oikos Suppl. 15: 128-132
Stelfox-Widdicombe C.E., Edwards E.S., Burkill P.H., Sleigh M.A. (2000)
Microzooplankton grazing activity in the temperate and sub-tropical NE Atlantic:
summer 1996. Mar. Ecol. Prog. Ser. 208: 1-12
Stoecker D.K. (1998) Conceptual models of mixotrophy in planktonic protists and some
ecological and evolutionary implications. Europ. J. Protistol. 34: 281-290
Stoecker D.K., Capuzzo J.M. (1990) Predation on protozoa: its importance to zooplankton.
J. Plank. Res. 12: 891-908
Stoecker D.K., Egloff, D.A. (1987) Predation by Acartia tonsa Dana on planktonic ciliates
and rotifers. J. Exp. Mar. Biol. Ecol. 110: 53-68
Stoecker D.K., Gifford D.J., Putt M. (1994) Preservation of marine planktonic ciliates:
losses and cell shrinkage during fixation. Mar. Ecol. Prog. Ser. 110: 293-299
Stoecker D.K., Gustafson D.E. (2002) Predicting grazing mortality of an estuarine
dinoflagellate Pfiesteria piscicida. Mar. Ecol. Prog. Ser. 233: 31-38
Stoecker D.K., Gustafson D.E. (2003) Cell-surface proteolytic activity of photosynthetic
dinoflagellates. Acq. Micr. Ecol. 30:175-183
Stoecker, D.K. (1999) Mixotrophy among dinoflagellates. J. Eukaryot. Microbiol. 46: 397-
401
Stoeker D.K., Gustafson D.E., Verity P.G. (1996) Micro- and mesozooplankton at 140°W
in the equatorial Pacific: heterotrophs and mixotrophs. Aquat. Microb. Ecol. 10:
273-282
Strom S., Postel J.R., Booth B.C. (1993) Abundance, variability, and potential grazing
impact of planktonic ciliates in the open subartic Pacific Ocean. Prog. Oceanogr.
32:185-203
Strom S.L., Strom M.W. (1996) Microplankton growth, grazing, and community structure
in the northern gulf of Mexico. Mar. Ecol. Prog. Ser. 130:229-240
Strom S.L., Welschmeyer N.A. (1991) Pigment-specific rates of phytoplankton growth and
microzooplankton grazing in the open sub arctic Pacific Ocean. Limnol. Oceanogr.
36: 50-63

Bibliografia
XXIII
Sullivan C.W., Arrigo K.R., McClain C.R., Comiso J.C., Firestone J. (1993) Distributions
of phytoplankton blooms in the Southern Ocean. Science 262: 1832-1837
Tanaka T., Rassoulzadegan F. (2002) Full-depth profile (0-2000 m) of bacteria,
heterotrophic nanoflagellates and ciliates in the NW Mediterranean Sea: Vertical
portioning of microbial trophic structures. Deep-Sea Res. II 49: 2093-2107
Taylor, J.F.R. (1979) A description of the benthic dinoflagellate associated with maitotoxin
and ciguatoxin, including observations on Hawaiian material. In: Taylor, D.L.,
Seliger, H.H. (Eds.) Toxic dinoflagellate blooms. Elsevier North Holland, New
York, USA, pp. 71-76
Thingstad T.F. , Krom M.D. Mantoura R.F., Flaten G., Groom S., Herut B., Kress N., Law
C.S., Paternak A., Pitta P., Psarra S., Rassoulzadegan F., Tanaka T., Tselepides A.,
Wassmann P., Woodward E.M., Wexels-Riser C., Zodiatis G., Zohary T. (2005)
Nature of phosphorous limitation in the ultraoligotrophic Eastern Mediterranean
Science 309:1068-1071
Thingstad T.F., Rassoulzadegan F. (1995) Nutrient limitations, microbial food webs, and
'biological C-pumps': suggested interactions in a P-limited Mediterranean. Mar.
Ecol. Prog. Ser. 117: 299-306
Thingstad T.F., Rassoulzadegan F. (1999) Conceptual models for the biogeochemical role
of the photic zone microbial food web, with particular reference to the
Mediterranean Sea. Progr. Oceanogr. 44: 271-286
Thompson G.A., Adler V.A. (2005) Patterns in tintinnid species composition and
abundance in relation to hydrological conditions of the south western Atlantic
during austral spring. Aquat. Microb. Ecol. 40: 85-101
Thompson G.A., Alder V.A., Boltovskoy D., Brandini F. (1999) Abundance and
biogeography of tintinnids (Ciliophora) and associated microzooplankton in the
south western Atlantic Ocean. J. Plank. Res. 21: 1265-1298
Throndsen J. (1997) The planktonic marine flagellates. In: Tomas C.R. (ed). Identifying
Marine Phytoplankton. Academic Press, San Diego pp: 591-729
Tognetto L., Bellato, S., Moro, I., Andreoli, C. (1995) Occurence of Ostreopsis ovata
(Dinophyceae) in the Tyrrhenian Sea during summer 1994. Botanica Marina 38(1-
6) 291-295

Bibliografia
XXIV
Totti C., Cucchiari, E., Romagnoli, T., Penna, A. (2007) Bloom of Ostreopsis ovata on the
Conero Riviera (NW Adriatic Sea). Harmful Algal News 33 12-13
Trègouboff G., Rose M. (1957) Manuel de planctonologie mediterranéenne. Tome 1, 2.
CNRS, Paris
Tréguer P., Pondaven P., Boyd P.N., Anderson R.F., Nelson D.M. (2002) Climatic changes
and the cycle of carbon in the Southern Ocean. Deep-Sea Res.Part 2,
Trop.Stud.Oceanogr. 49 (16): 3103-3104
Turner, J.T., Granèli, E. (1992) Zooplankton feeding ecology: grazing during enclosure
studies of phytoplankton blooms from the west coast of Sweden. J. Exp. Mar. Biol.
Ecol., 157 (1): 19-31
Ukena, T., Satake, M., Usami, M., Oshima, Y., Naoki, H., Fujita, T., Kan Y., Yasumoto,
Y. (2001) Structure elucidation of ostreocin D, a palitoxin isolated from the
dinoflagellate Ostreopsis siamensis. Biosci. Biotechnol. Biochem., 65: 2585-2588
Utermöhl H. (1958) Zur Vervollkommnung der quantitativen Phytoplankton-Methodik.
Mitt. Int. Ver. Theor. Angew. Limnol. 9: 1-38
Vaqué D., Blough H.A., Duarde C.M. (1997) Dynamics of ciliate abundance, biomass and
community composition in an oligotrophic coastal environment (NW
Mediterranean). Aquat. Microb. Ecol. 12: 71-83
Verity P.G. (1985) Grazing, respiration, excretion, and growth rates of tintinnids. Limnol.
Oceanogr. 30: 1268-1282
Verity P.G., Langdon C. (1984) Relationships between lorica volume, carbon, nitrogen,
and ATP content of tintinnids in Narragansett Bay. J. Plank. Res. 6 (5): 859-868
Verity P.G., Paffenhöfer G.A. (1996) On the assessment of prey ingestion by copepods. J.
Plank. Res. 18: 1767-1779
Verity P.G., Stoecker D.K., Sieraki M.E. Nelson J.R. (1993) Grazing, growth and mortality
of microzooplankton during the 1989 North Atlantic spring bloom at 47°N, 18°W.
Deep Sea Res. I 40 (9): 1793-1814
Vila M., Garcés, E., Masò, M. (2001) Potentially toxic epiphytic dinoflagellate
assemblages on macroalgae in the NW Mediterranean. Aquat. Microb. Ecol., 26:
51-60

Bibliografia
XXV
Volk T., Hoffert M.I. (1985) Ocean carbon pumps: analysis of relative strengths and
efficiencies in ocean driven atmospheric CO2 change. In: ET. Sundquist, Broecker
W.S. (eds), The Carbon Cycle and Atmospheric CO2; Natural Variations Archean
to Present, Washington DC; American Geophysical Union, Geophysical
Monograph. 32: 99-110
Wang, D-Z. (2008) Neurotoxins from marine dinoflagellates: a breif review. Marine Drugs
6: 349-371
Waterbury J.B., Watson S.W., Guillard R.R.L., Brand L. (1979) Widespread occurrence of
a unicellular, marine, planktonic cyanobacterium. Nature 277: 293-294
Waterhouse T.Y, Welschmeyer N.A. (1995) Taxon specific analysis of microzooplankton
grazing rates and phytoplankton growth rates. Limnol. Oceanogr. 40 (4): 827-834
Waters R.L. van den enden R., Marchant H.J. (2000) Summer microbial ecology off East
Antarctica (80-150°E). Protistan community structure and bacterial abundance.
Deep-Sea Res II, 47:2401-2435
Wiadnyana N.N., Rassoulzadegan F. (1989) Selective feeding of Acartia clausi and
Centropages typicus on microzooplankton. Mar. Ecol. Prog. Ser. 53: 37-47
Williams P.J. Le B. (1990) The importance of losses during microbial growth:
Commentary on the physiology, measurement and ecology of the release of
dissolved organic material. Mar. Microb. Food Webs 4: 175-206
Xiuren N, Zilin L., Genhai Z., Junxian S., (1996) Size feactioned biomass and productivity
of phytoplankton and particulate organic carbon in the Southern Ocean. Polar Biol.
16:1-11
Zinabu G.M., Bott T.L. (2000) The effects of formaldehyde and Lugol’s iodine solution on
protozoan cell volume. Limnologica 30: 59-63

Allegati
Mediterraneo

PICOPLANCTON AUTOTROFO
V6 T0 µgC L-1 ± SD V6 T24 µgC L-1 ± SD V6 T0 Cell L-1 ± SD V6 T24 Cell L-1 ± SD 100% 0.485 0.089 100% 0.670 0.035 100% 2.43E+06 4.46E+05 100% 3.35E+06 1.77E+05
80% 0.295 0.009 80% 0.426 0.084 80% 1.48E+06 4.53E+04 80% 2.13E+06 4.18E+05
50% 0.195 0.031 50% 0.293 0.065 50% 9.73E+05 1.56E+05 50% 1.47E+06 3.23E+05
20% 0.138 0.025 20% 0.213 0.032 20% 6.91E+05 1.23E+05 20% 1.06E+06 1.59E+05
V7 T0 µgC L-1 ± SD V7 T24 µgC L-1 ± SD V7 T0 Cell L-1 ± SD V7 T24 Cell L-1 ± SD 100% 0.451 0.119 100% 0.375 0.066 100% 2.26E+06 5.93E+05 100% 1.88E+06 3.29E+05
80% 0.296 0.056 80% 0.209 0.017 80% 1.48E+06 2.81E+05 80% 1.04E+06 8.46E+04
50% 0.193 0.033 50% 0.145 0.021 50% 9.66E+05 1.63E+05 50% 7.23E+05 1.07E+05
20% 0.255 0.058 20% 0.070 0.009 20% 1.28E+06 2.90E+05 20% 3.49E+05 4.61E+04
Viera T0 µgC L-1 ± SD Viera T24 µgC L-1 ± SD Viera T0 Cell L-1 ± SD Viera T24 Cell L-1 ± SD 100% 0.417 0.104 100% 0.444227
37 0.103717
8 100% 2.09E+06 5.19E+05 100% 2.22E+06 5.19E+05
80% 0.320 0.037 80% 0.26606634
0.0282519
80% 1.60E+06 1.84E+05 80% 1.33E+06 1.41E+05
50% 0.197 0.023 50% 0.19461389
0.0434037
50% 9.87E+05 1.17E+05 50% 9.73E+05 2.17E+05
20% 0.178 0.010 20% 0.15935775
0.0296403
20% 8.92E+05 4.77E+04 20% 7.97E+05 1.48E+05
V10 T0 µgC L-1 ± SD V10 T24 µgC L-1 ± SD V10 T0 Cell L-1 ± SD V10 T24 Cell L-1 ± SD 100% 0.642 0.059 100% 0.611 0.074 100% 3.21E+06 2.97E+05 100% 3.05E+06 3.70E+05
80% 0.752 0.117 80% 0.804 0.168 80% 3.76E+06 5.87E+05 80% 4.02E+06 8.41E+05
50% 0.542 0.077 50% 0.608 0.152 50% 2.71E+06 3.87E+05 50% 3.04E+06 7.60E+05
20% 0.478 0.092 20% 0.185 0.015 20% 2.39E+06 4.61E+05 20% 9.27E+05 7.50E+04

PICOPLANCTON ETEROTROFO
V6 T0 µgCL-1 ± SD V7 T0 µgCL-1 ± SD Viera T0 µgCL-1 ± SD V10 T0 µgCL-1 ± SD
100% 8.141 0.381 100% 6.668 1.041 100% 5.040 0.396 100% 6.140 0.704
100% 7.179 0.519 100% 5.520 1.047 100% 4.729 0.097 100% 6.156 0.349
100% 7.148 0.256 100% 5.489 0.406 100% 4.853 0.295 100% 6.125 0.352
80% 6.315 0.277 80% 3.826 0.198 80% 3.721 0.145 80% 4.594 0.474
80% 5.582 0.277 80% 4.524 0.351 80% 3.652 0.165 80% 4.617 0.344
80% 6.745 0.561 80% 3.791 0.232 80% 3.942 0.604 80% 4.629 0.291
50% 4.807 0.094 50% 3.171 0.165 50% 2.884 0.023 50% 5.202 0.543
50% 4.768 0.236 50% 2.861 0.545 50% 2.907 0.047 50% 3.900 0.272
50% 4.846 0.284 50% 4.892 0.896 50% 2.745 0.176 50% 3.869 0.107
20% 2.349 0.072 20% 1.530 0.137 20% 1.419 0.069 20% 2.163 0.266
20% 2.386 0.028 20% 1.698 0.056 20% 1.479 0.056 20% 2.493 0.093
20% 2.279 0.157 20% 1.451 0.161 20% 1.503 0.035 20% 2.368 0.082
V6 T24 µgCL-1 ± SD V7 T24 µgCL-1 ± SD Viera T24 µgCL-1 ± SD V10 T24 µgCL-1 ± SD
100% 19.104 1.142 100% 8.172 0.376 100% 8.606 0.524 100% 9.722 0.645
100% 11.118 1.587 100% 6.947 0.563 100% 6.807 0.457 100% 8.823 1.530
100% 12.126 0.839 100% 8.156 0.468 100% 8.513 1.070 100% 8.823 0.559
80% 9.536 0.838 80% 6.559 0.424 80% 6.210 0.393 80% 10.083 1.121
80% 10.734 0.693 80% 6.745 0.161 80% 5.873 0.222 80% 9.199 0.640
80% 10.129 0.773 80% 6.501 0.165 80% 5.024 0.665 80% 13.118 2.363
50% 12.924 1.815 50% 9.404 1.399 50% 7.482 0.635 50% 15.630 0.512
50% 18.759 1.661 50% 9.203 1.073 50% 7.683 0.295 50% 14.987 1.723
50% 17.142 1.548 50% 12.878 1.640 50% 8.071 0.524 50% 18.003 1.299
20% 5.182 0.162 20% 5.996 0.622 20% 8.597 0.535 20% 11.820 0.672
20% 10.099 3.371 20% 5.652 0.157 20% 10.834 0.234 20% 13.997 1.441
20% 8.169 3.155 20% 6.447 0.105 20% 9.922 1.571 20% 16.337 0.778

PICOPLANCTON ETEROTROFO
V6 T0 Cell L-1 ± SD V7 T0 Cell L-1 ± SD Viera T0 Cell L-1 ± SD V10 T0 Cell L-1 ± SD
100% 4.07E+08
1.90E+07
100% 3.33E+08
5.20E+07
100% 2.52E+08 1.98E+07 100% 3.07E+08 3.52E+07
100% 3.71E+08
2.59E+07
100% 2.76E+08
5.23E+07
100% 2.36E+08 2.28E+07 100% 3.08E+08 1.75E+07
100% 3.57E+08
1.28E+07
100% 2.74E+08
2.03E+07
100% 2.43E+08 5.35E+07 100% 3.06E+08 1.76E+07
80% 3.16E+08
1.38E+07
80% 1.91E+08
9.92E+06
80% 1.83E+08 7.26E+06 80% 2.30E+08 2.37E+07
80% 2.79E+08
1.38E+07
80% 2.26E+08
1.76E+07
80% 1.83E+08 8.24E+06 80% 2.31E+08 1.72E+07
80% 3.37E+08
2.80E+07
80% 1.90E+08
1.16E+07
80% 1.97E+08 3.02E+07 80% 2.31E+08 1.45E+07
50% 2.40E+08
4.70E+06
50% 1.59E+08
8.25E+06
50% 1.44E+08 1.16E+06 50% 2.60E+08 2.72E+07
50% 2.38E+08
1.18E+07
50% 1.43E+08
2.72E+07
50% 1.45E+08 2.33E+06 50% 1.95E+08 1.36E+07
50% 2.42E+08
1.42E+07
50% 2.45E+08
4.48E+07
50% 1.37E+08 8.78E+06 50% 1.93E+08 5.37E+06
20% 1.17E+08
3.58E+06
20% 7.65E+07
6.85E+06
20% 7.09E+07 3.44E+06 20% 1.08E+08 1.33E+07
20% 1.19E+08
1.40E+06
20% 8.49E+07
2.82E+06
20% 7.40E+07 2.79E+06 20% 1.25E+08 4.65E+06
20% 1.14E+08
7.84E+06
20% 7.26E+07
8.05E+06
20% 7.51E+07 1.76E+06 20% 1.18E+08 4.09E+06
V6 T24 Cell L-1 ± SD V7 T24 Cell L-1 ± SD Viera T24 Cell L-1 ± SD V10 T24 Cell L-1 ± SD
100% 9.55E+08
5.71E+07
100% 4.09E+08
1.88E+07
100% 4.30E+08 2.62E+07 100% 4.86E+08 3.22E+07
100% 5.56E+08
3.46E+07
100% 3.47E+08
2.81E+07
100% 3.40E+08 2.28E+07 100% 4.41E+08 7.65E+07
100% 6.06E+08
3.86E+07
100% 4.08E+08
2.34E+07
100% 4.26E+08 5.35E+07 100% 4.41E+08 2.79E+07
80% 4.77E+08
4.19E+07
80% 3.28E+08
2.12E+07
80% 3.11E+08 1.97E+07 80% 5.04E+08 5.61E+07
80% 5.37E+08
3.46E+07
80% 3.29E+08
1.31E+07
80% 2.90E+08 9.61E+06 80% 4.60E+08 3.20E+07
80% 5.06E+08
3.86E+07
80% 3.42E+08
1.91E+07
80% 2.61E+08 3.39E+07 80% 6.56E+08 1.18E+08
50% 6.46E+08
9.08E+07
50% 4.70E+08
6.99E+07
50% 3.74E+08 3.17E+07 50% 7.82E+08 2.56E+07
50% 9.38E+08
8.31E+07
50% 4.60E+08
5.37E+07
50% 4.04E+08 2.91E+07 50% 7.49E+08 8.61E+07
50% 8.57E+08
7.74E+07
50% 6.44E+08
8.20E+07
50% 4.19E+08 2.68E+07 50% 9.00E+08 6.49E+07
20% 2.59E+08
8.09E+06
20% 3.00E+08
3.11E+07
20% 4.30E+08 2.68E+07 20% 5.91E+08 3.36E+07
20% 5.05E+08
1.69E+08
20% 2.83E+08
7.86E+06
20% 5.55E+08 2.98E+07 20% 7.00E+08 7.21E+07
20% 4.08E+08
1.58E+08
20% 3.22E+08
5.27E+06
20% 5.23E+08 6.26E+07 20% 8.17E+08 3.89E+07

NANOPLANCTON - µgC L -1
A B C Media ± SD A B C Media ± SD 100% 0.168 0.174 0.207 0.183 0.021 100% 0.274 0.104 0.178 0.185 0.085
80% 0.116 0.140 0.119 0.125 0.013 80% 0.093 0.077 0.104 0.091 0.014
50% 0.081 0.059 0.048 0.063 0.017 50% 0.065 0.053 0.051 0.056 0.007 V6 T0 < 3µm
20% 0.058 0.061 0.146 0.088 0.050
V7 T0 < 3µm
20% 0.042 0.044 0.039 0.042 0.002
100% 0.488 0.301 0.352 0.380 0.097 100% 0.664 0.378 0.352 0.465 0.173
80% 0.234 0.203 0.166 0.201 0.034 80% 0.221 0.300 0.339 0.287 0.060
50% 0.273 0.233 0.314 0.273 0.040 50% 0.164 0.172 0.188 0.175 0.012 V6 T0 3-5 µm
20% 0.148 0.133 0.173 0.152 0.020
V7 T0 3-5 µm
20% 0.055 0.102 0.117 0.091 0.033
100% 0.995 0.448 0.721 0.721 0.274 100% 1.447 1.628 1.085 1.387 0.276
80% 0.543 0.464 0.336 0.448 0.104 80% 0.965 0.301 0.482 0.583 0.343
50% 0.407 0.259 0.382 0.349 0.079 50% 0.398 0.470 0.579 0.482 0.091 V6 T0 >5 µm
20% 0.253 0.213 0.213 0.227 0.023
V7 T0 >5 µm
20% 0.217 0.181 0.217 0.205 0.021
100% 0.185 0.187 0.141 0.171 0.026 100% 0.112 0.064 0.076 0.084 0.025
80% 0.098 0.083 0.069 0.083 0.015 80% 0.072 0.051 0.046 0.056 0.014
50% 0.089 0.083 0.080 0.084 0.004 50% 0.078 0.044 0.038 0.053 0.022 Viera T0 < 3µm
20% 0.051 0.059 0.059 0.056 0.005
V10 T0 < 3µm
20% 0.056 0.056 0.060 0.057 0.003
100% 0.625 0.515 0.395 0.512 0.115 100% 0.313 0.245 0.316 0.291 0.040
80% 0.273 0.311 0.311 0.299 0.022 80% 0.234 0.159 0.169 0.188 0.041
50% 0.091 0.077 0.075 0.081 0.009 50% 0.244 0.166 0.186 0.199 0.041 Viera T0 3-5 µm
20% 0.078 0.074 0.078 0.077 0.002
V10 T0 3-5 µm
20% 0.164 0.177 0.151 0.164 0.013
100% 0.995 0.663 0.516 0.725 0.245 100% 0.724 0.543 0.780 0.682 0.124
80% 0.362 0.310 0.207 0.293 0.079 80% 0.678 0.536 0.536 0.583 0.082
50% 0.241 0.241 0.254 0.245 0.007 50% 0.407 0.351 0.334 0.364 0.038 Viera T0 >5 µm
20% 0.136 0.060 0.090 0.095 0.038
V10 T0 >5 µm
20% 0.253 0.158 0.285 0.232 0.066
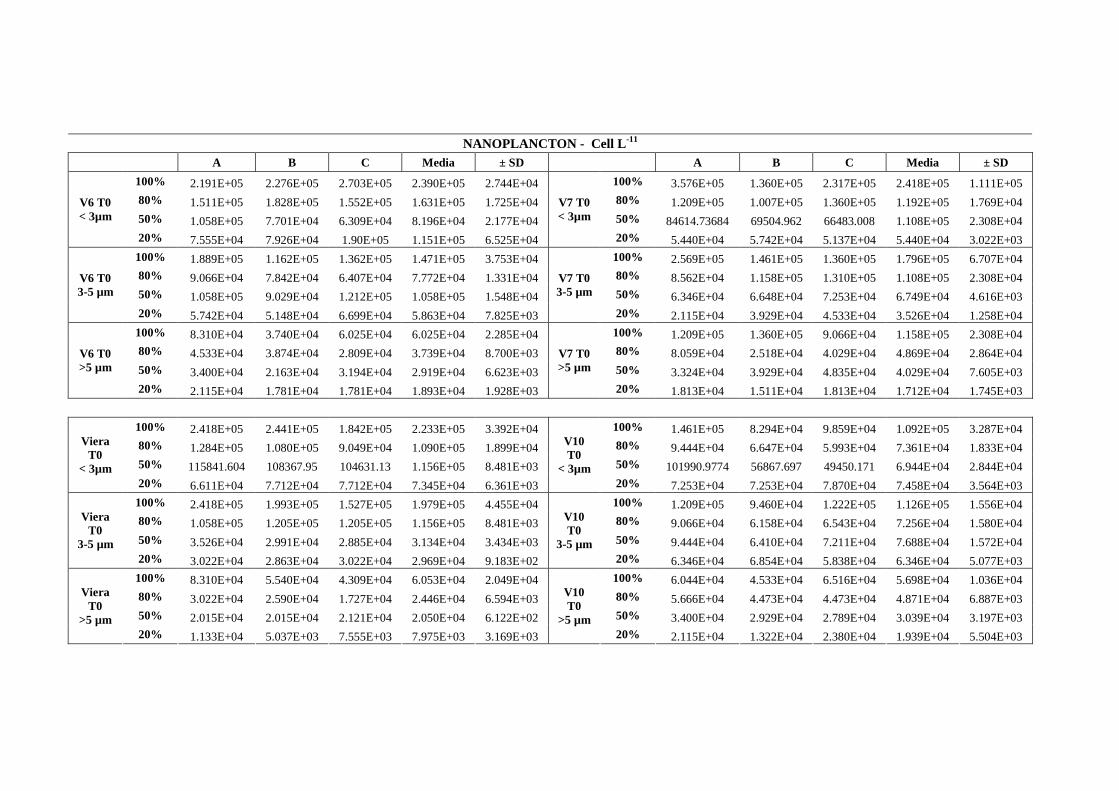
NANOPLANCTON - Cell L -11 A B C Media ± SD A B C Media ± SD
100% 2.191E+05 2.276E+05 2.703E+05 2.390E+05 2.744E+04 100% 3.576E+05 1.360E+05 2.317E+05 2.418E+05 1.111E+05
80% 1.511E+05 1.828E+05 1.552E+05 1.631E+05 1.725E+04 80% 1.209E+05 1.007E+05 1.360E+05 1.192E+05 1.769E+04 50% 1.058E+05 7.701E+04 6.309E+04 8.196E+04 2.177E+04 50% 84614.73684 69504.962 66483.008 1.108E+05 2.308E+04
V6 T0 < 3µm
20% 7.555E+04 7.926E+04 1.90E+05 1.151E+05 6.525E+04
V7 T0 < 3µm
20% 5.440E+04 5.742E+04 5.137E+04 5.440E+04 3.022E+03
100% 1.889E+05 1.162E+05 1.362E+05 1.471E+05 3.753E+04 100% 2.569E+05 1.461E+05 1.360E+05 1.796E+05 6.707E+04 80% 9.066E+04 7.842E+04 6.407E+04 7.772E+04 1.331E+04 80% 8.562E+04 1.158E+05 1.310E+05 1.108E+05 2.308E+04 50% 1.058E+05 9.029E+04 1.212E+05 1.058E+05 1.548E+04 50% 6.346E+04 6.648E+04 7.253E+04 6.749E+04 4.616E+03
V6 T0 3-5 µm
20% 5.742E+04 5.148E+04 6.699E+04 5.863E+04 7.825E+03
V7 T0 3-5 µm
20% 2.115E+04 3.929E+04 4.533E+04 3.526E+04 1.258E+04
100% 8.310E+04 3.740E+04 6.025E+04 6.025E+04 2.285E+04 100% 1.209E+05 1.360E+05 9.066E+04 1.158E+05 2.308E+04 80% 4.533E+04 3.874E+04 2.809E+04 3.739E+04 8.700E+03 80% 8.059E+04 2.518E+04 4.029E+04 4.869E+04 2.864E+04
50% 3.400E+04 2.163E+04 3.194E+04 2.919E+04 6.623E+03 50% 3.324E+04 3.929E+04 4.835E+04 4.029E+04 7.605E+03
V6 T0 >5 µm
20% 2.115E+04 1.781E+04 1.781E+04 1.893E+04 1.928E+03
V7 T0 >5 µm
20% 1.813E+04 1.511E+04 1.813E+04 1.712E+04 1.745E+03
100% 2.418E+05 2.441E+05 1.842E+05 2.233E+05 3.392E+04 100% 1.461E+05 8.294E+04 9.859E+04 1.092E+05 3.287E+04 80% 1.284E+05 1.080E+05 9.049E+04 1.090E+05 1.899E+04 80% 9.444E+04 6.647E+04 5.993E+04 7.361E+04 1.833E+04 50% 115841.604 108367.95 104631.13 1.156E+05 8.481E+03 50% 101990.9774 56867.697 49450.171 6.944E+04 2.844E+04
Viera T0
< 3µm 20% 6.611E+04 7.712E+04 7.712E+04 7.345E+04 6.361E+03
V10 T0
< 3µm 20% 7.253E+04 7.253E+04 7.870E+04 7.458E+04 3.564E+03
100% 2.418E+05 1.993E+05 1.527E+05 1.979E+05 4.455E+04 100% 1.209E+05 9.460E+04 1.222E+05 1.126E+05 1.556E+04 80% 1.058E+05 1.205E+05 1.205E+05 1.156E+05 8.481E+03 80% 9.066E+04 6.158E+04 6.543E+04 7.256E+04 1.580E+04
50% 3.526E+04 2.991E+04 2.885E+04 3.134E+04 3.434E+03 50% 9.444E+04 6.410E+04 7.211E+04 7.688E+04 1.572E+04
Viera T0
3-5 µm 20% 3.022E+04 2.863E+04 3.022E+04 2.969E+04 9.183E+02
V10 T0
3-5 µm 20% 6.346E+04 6.854E+04 5.838E+04 6.346E+04 5.077E+03
100% 8.310E+04 5.540E+04 4.309E+04 6.053E+04 2.049E+04 100% 6.044E+04 4.533E+04 6.516E+04 5.698E+04 1.036E+04
80% 3.022E+04 2.590E+04 1.727E+04 2.446E+04 6.594E+03 80% 5.666E+04 4.473E+04 4.473E+04 4.871E+04 6.887E+03 50% 2.015E+04 2.015E+04 2.121E+04 2.050E+04 6.122E+02 50% 3.400E+04 2.929E+04 2.789E+04 3.039E+04 3.197E+03
Viera T0
>5 µm 20% 1.133E+04 5.037E+03 7.555E+03 7.975E+03 3.169E+03
V10 T0
>5 µm 20% 2.115E+04 1.322E+04 2.380E+04 1.939E+04 5.504E+03

NANOPLANCTON - µgC L -1
A B C Media ± SD A B C Media ± SD 100% 0.255 0.192 0.186 0.211 0.038 100% 0.084 0.104 0.043 0.077 0.031 80% 0.162 0.195 0.130 0.162 0.032 80% 0.110 0.122 0.107 0.113 0.008 50% 0.135 0.142 0.105 0.127 0.020 50% 0.147 0.143 0.087 0.125 0.033
V6 T24 < 3µm
20% 0.089 0.082 0.110 0.093 0.014
V7 T24 < 3µm
20% 0.066 0.098 0.139 0.101 0.037 100% 0.488 0.481 0.447 0.472 0.022 100% 0.440 0.420 0.332 0.397 0.057 80% 0.313 0.229 0.250 0.264 0.044 80% 0.215 0.322 0.459 0.332 0.122 50% 0.365 0.252 0.233 0.283 0.071 50% 0.371 0.260 0.319 0.317 0.055
V6 T24 3-5 µm
20% 0.300 0.250 0.234 0.261 0.034
V7 T24 3-5 µm
20% 0.195 0.221 0.339 0.252 0.076 100% 0.995 0.758 0.455 0.736 0.271 100% 1.266 1.131 1.221 1.206 0.069 80% 0.814 0.398 0.470 0.561 0.222 80% 1.176 1.085 0.633 0.965 0.291 50% 0.482 0.402 0.287 0.390 0.098 50% 0.543 0.603 0.814 0.653 0.142
V6 T24 >5 µm
20% 0.362 0.603 1.105 0.690 0.379
V7 T24 >5 µm
20% 0.392 0.301 0.301 0.332 0.052
100% 0.330 0.238 0.307 0.292 0.048 100% 0.061 0.055 0.044 0.054 0.008 80% 0.087 0.083 0.103 0.091 0.011 80% 0.127 0.120 0.135 0.127 0.007 50% 0.068 0.073 0.076 0.072 0.004 50% 0.127 0.107 0.129 0.121 0.012
Viera T24 < 3µm
20% 0.082 0.181 0.105 0.123 0.051
V10 T24 < 3µm
20% 0.071 0.066 0.066 0.068 0.003 100% 0.918 1.109 1.262 1.097 0.173 100% 0.313 0.208 0.288 0.270 0.054 80% 0.186 0.241 0.309 0.245 0.062 80% 0.205 0.205 0.171 0.194 0.020 50% 0.163 0.293 0.342 0.266 0.093 50% 0.247 0.427 0.457 0.377 0.114
Viera T24 3-5 µm
20% 0.127 0.192 0.195 0.171 0.039
V10 T24 3-5 µm
20% 0.210 0.286 0.362 0.286 0.076 100% 2.261 1.515 1.439 1.738 0.454 100% 1.131 0.565 0.391 0.696 0.386 80% 0.633 0.844 1.055 0.844 0.211 80% 0.452 0.517 0.452 0.474 0.037 50% 0.241 0.201 0.040 0.161 0.106 50% 0.211 0.362 0.241 0.271 0.080
Viera T24 >5 µm
20% 0.023 0.028 0.028 0.026 0.003
V10 T24 >5 µm
20% 0.136 0.407 0.560 0.367 0.215

NANOPLANCTON - Cell L -1
A B C Media ± SD A B C Media ± SD 100% 3.324E+05 2.505E+05 2.428E+05 2.752E+05 4.966E+04 100% 1.095E+05 1.360E+05 5.666E+04 1.007E+05 4.039E+04 80% 2.115E+05 2.541E+05 1.692E+05 2.116E+05 4.242E+04 80% 1.435E+05 1.587E+05 1.398E+05 1.473E+05 9.994E+03 50% 1.763E+05 1.856E+05 1.366E+05 1.661E+05 2.601E+04 50% 1.914E+05 1.864E+05 1.133E+05 1.637E+05 4.369E+04
V6 T24 < 3µm
20% 1.158E+05 1.069E+05 1.431E+05 1.219E+05 1.885E+04
V7 T24 < 3µm
20% 8.562E+04 1.284E+05 1.813E+05 1.318E+05 4.794E+04 100% 1.889E+05 1.862E+05 1.727E+05 1.826E+05 8.673E+03 100% 1.700E+05 1.624E+05 1.284E+05 1.536E+05 2.213E+04 80% 1.209E+05 8.851E+04 9.670E+04 1.020E+05 1.683E+04 80% 8.310E+04 1.247E+05 1.775E+05 1.284E+05 4.733E+04 50% 1.410E+05 9.732E+04 8.998E+04 1.094E+05 2.760E+04 50% 1.435E+05 1.007E+05 1.234E+05 1.226E+05 2.142E+04
V6 T24 3-5 µm
20% 1.158E+05 9.653E+04 9.037E+04 1.009E+05 1.329E+04
V7 T24 3-5 µm
20% 7.555E+04 8.562E+04 1.310E+05 9.737E+04 2.951E+04 100% 8.310E+04 6.332E+04 3.799E+04 6.147E+04 2.261E+04 100% 1.058E+05 9.444E+04 1.020E+05 1.007E+05 5.770E+03 80% 6.799E+04 3.324E+04 3.929E+04 4.684E+04 1.857E+04 80% 9.821E+04 9.066E+04 5.288E+04 8.059E+04 2.429E+04 50% 4.029E+04 3.358E+04 2.398E+04 3.262E+04 8.197E+03 50% 4.533E+04 5.037E+04 6.799E+04 5.456E+04 1.190E+04
V6 T24 >5 µm
20% 3.022E+04 5.037E+04 9.234E+04 5.764E+04 3.169E+04
V7 T24 >5 µm
20% 3.274E+04 2.518E+04 2.518E+04 2.770E+04 4.362E+03
100% 4.306E+05 3.105E+05 4.006E+05 3.806E+05 6.254E+04 100% 7.933E+04 7.224E+04 5.808E+04 6.988E+04 1.082E+04 80% 1.133E+05 1.079E+05 1.349E+05 1.187E+05 1.428E+04 80% 1.662E+05 1.564E+05 1.760E+05 1.662E+05 9.777E+03 50% 8.814E+04 9.475E+04 9.916E+04 9.402E+04 5.545E+03 50% 1.662E+05 1.392E+05 1.683E+05 1.579E+05 1.623E+04
Viera T24
< 3µm 20% 1.077E+05 2.359E+05 1.374E+05 1.603E+05 6.713E+04
V10 T24
< 3µm 20% 9.255E+04 8.653E+04 8.653E+04 8.853E+04 3.477E+03
100% 3.551E+05 4.291E+05 4.882E+05 4.241E+05 6.671E+04 100% 1.209E+05 8.059E+04 1.114E+05 1.043E+05 2.107E+04 80% 7.177E+04 9.304E+04 1.196E+05 9.481E+04 2.397E+04 80% 7.933E+04 7.933E+04 6.611E+04 7.492E+04 7.633E+03 50% 6.296E+04 1.133E+05 1.322E+05 1.028E+05 3.580E+04 50% 9.570E+04 1.653E+05 1.769E+05 1.460E+05 4.391E+04
Viera T24
3-5 µm 20% 4.911E+04 7.429E+04 7.555E+04 6.632E+04 1.492E+04
V10 T24
3-5 µm 20% 8.122E+04 1.105E+05 1.399E+05 1.105E+05 2.933E+04
100% 1.889E+05 1.265E+05 1.202E+05 1.452E+05 3.796E+04 100% 9.444E+04 4.722E+04 3.269E+04 5.811E+04 3.228E+04 80% 5.288E+04 7.051E+04 8.814E+04 7.051E+04 1.763E+04 80% 3.777E+04 4.317E+04 3.777E+04 3.957E+04 3.116E+03 50% 2.015E+04 1.679E+04 3.358E+03 1.343E+04 8.884E+03 50% 1.763E+04 3.022E+04 2.015E+04 2.266E+04 6.663E+03
Viera T24
>5 µm 20% 1.889E+03 2.361E+03 2.361E+03 2.204E+03 2.726E+02
V10 T24
>5 µm 20% 1.133E+04 3.400E+04 4.675E+04 3.069E+04 1.794E+04

MICROZOOPLANCTON
Stazione V6 ABC T0 Cell L-1 µgC L-1 Cell L-1 µgC L-1 Cell L-1 µgC L-1 OLIGOTRICHIDA Laboea strobila 0 0 0 0 1 0.007 Legardiella sp 8 0.006 0 0 0 0
Lohmanniella sp. 12 0.003 6 0.002 8 0.002
Nanociliati 17 0.004 58 0.015 39 0.010
Oligotricha indet. 12 0.051 10 0.045 12 0.051
Strombidium acutum 8 0.037 6 0.030 3 0.012
Strombidium capitatum 0 0 0 0 3 0
Strombidium conicum 0 0 0 0 5 0.012
Strombidium sp. 6 0.003 6 0.003 8 0.004
Tontonia sp. 0 0 0 0 1 0.007 PERITRICHIDA
Peritrichida indet. 0 0 8 0.037 9 0.042 HOLOTRICHIA
Holotrichia indet. 5 0.011 10 0.021 16 0.034 TINTINNIDA
Dadayiella ganymedes 0 0 3 0.002 4 0.004
Epiplocylis blanda 0 0 3 0.014 0
Eutintinnus tubulosus 6 0.013 0 0 3 0.005
Rhabdonella spiralis 0 0 4 0.037 4 0.037
Tintinnide indet. 6 0.021 0 0 0 0 DINOFLAGELLIDA
Gymnodinium sp. 1 0 5 0.001 5 0.001
Gyrodinium fusiforme 10 0.007 13 0.009 0 0
Gyrodinium lachryma. 3 0.011 0 0 0 0
Ornythocercus magnificus 0 0 0 0 1 0.004
Protoperidinium brevipes 0 0 0 0 1 0.003
Protoperidinium crassipes 0 0 3 0.032 0 0
Protoperidinium sp. 3 0.006 5 0.010 4 0.008
Protoperidinium divergens 1 0.014 0 0 3 0.031
Protoperidinium steini 8 0.012 5 0.005 3 0.003
Protoperidinium subinerme 0 0 4 0.009 0 0
FORAMINIFERIDA
Globigerina 3 0.002 1 0.001 0 0
RADIOLARIA
Radiolaria indet. 8 0.009 0 0 3 0.027
COPEPODA
Copepoda nauplii 5 0.257 4 0.193 7 0.369
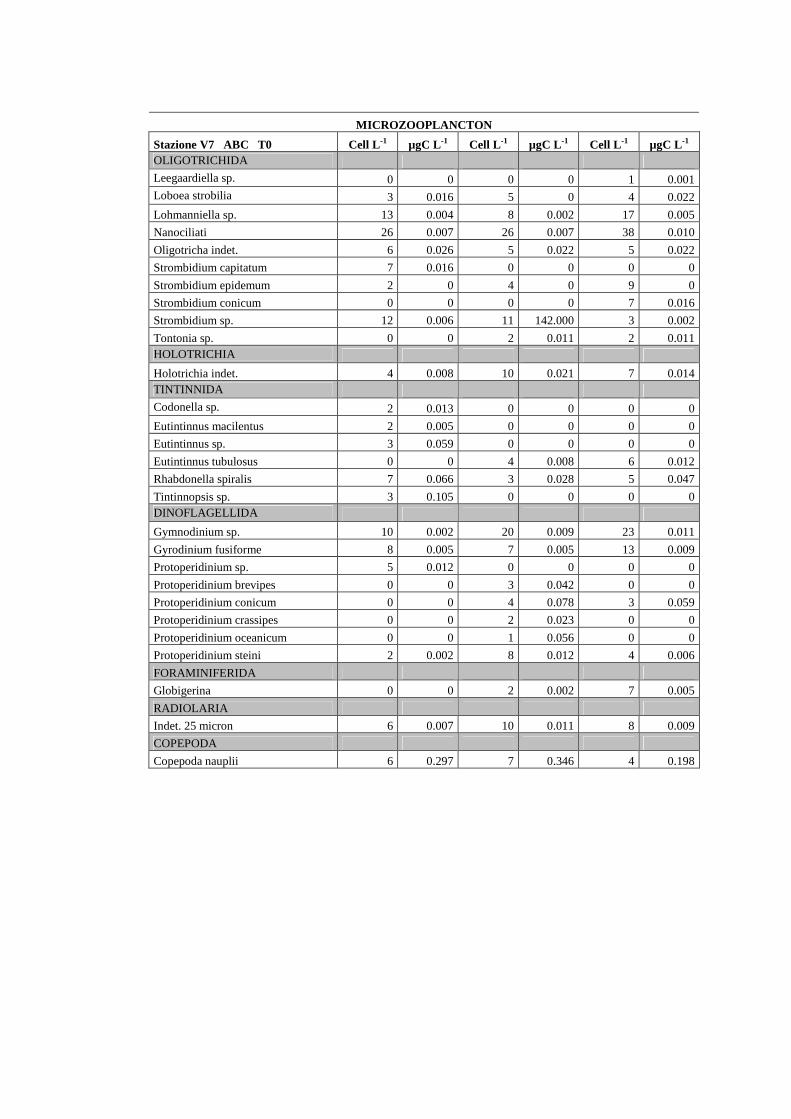
MICROZOOPLANCTON
Stazione V7 ABC T0 Cell L-1 µgC L-1 Cell L-1 µgC L-1 Cell L-1 µgC L-1 OLIGOTRICHIDA Leegaardiella sp. 0 0 0 0 1 0.001 Loboea strobilia 3 0.016 5 0 4 0.022
Lohmanniella sp. 13 0.004 8 0.002 17 0.005
Nanociliati 26 0.007 26 0.007 38 0.010
Oligotricha indet. 6 0.026 5 0.022 5 0.022
Strombidium capitatum 7 0.016 0 0 0 0
Strombidium epidemum 2 0 4 0 9 0
Strombidium conicum 0 0 0 0 7 0.016
Strombidium sp. 12 0.006 11 142.000 3 0.002
Tontonia sp. 0 0 2 0.011 2 0.011 HOLOTRICHIA
Holotrichia indet. 4 0.008 10 0.021 7 0.014 TINTINNIDA Codonella sp. 2 0.013 0 0 0 0
Eutintinnus macilentus 2 0.005 0 0 0 0
Eutintinnus sp. 3 0.059 0 0 0 0
Eutintinnus tubulosus 0 0 4 0.008 6 0.012
Rhabdonella spiralis 7 0.066 3 0.028 5 0.047
Tintinnopsis sp. 3 0.105 0 0 0 0 DINOFLAGELLIDA
Gymnodinium sp. 10 0.002 20 0.009 23 0.011
Gyrodinium fusiforme 8 0.005 7 0.005 13 0.009
Protoperidinium sp. 5 0.012 0 0 0 0
Protoperidinium brevipes 0 0 3 0.042 0 0
Protoperidinium conicum 0 0 4 0.078 3 0.059
Protoperidinium crassipes 0 0 2 0.023 0 0
Protoperidinium oceanicum 0 0 1 0.056 0 0
Protoperidinium steini 2 0.002 8 0.012 4 0.006
FORAMINIFERIDA
Globigerina 0 0 2 0.002 7 0.005
RADIOLARIA
Indet. 25 micron 6 0.007 10 0.011 8 0.009
COPEPODA
Copepoda nauplii 6 0.297 7 0.346 4 0.198

MICROZOOPLANCTON
Stazione Viera ABC T0 Cell L-1 µgC L-1 Cell L-1 µgC L-1 Cell L-1 µgC L-1 OLIGOTRICHIDA Leegardiella spp. 12 0.066 0 0 0 0 Loboea strobilia 4 0.022 4 0.020 3 0.016
Lohmanniella sp. 0 0 5 0.001 7 0.002
Nanociliati 10-20 micron 30 0.008 14 0.004 35 0.009
Oligotricha indet. 0 0 4 0.016 9 0.039
Strombidium conicum 5 0.012 0 0 6 0.014
Strombidium epideum 4 0 4 0 0 0
Strombidium sp. 18 0.009 17 0.008 15 0.008 HOLOTRICHIA
Holotrichia indet. 16 0.033 6 0.011 6 0.012 TINTINNIDA
Craterella torulata 2 0.001 0 0 0 0
Epiplocys blanda 0 0 0 0 2 0
Eutintinnus macilentus 4 0.011 3 0.007 2 0.005
Eutintinnus tubulosus 3 0.006 2 0.004 0 0
Rhabdonella spiralis 4 0.038 4 0.035 5 0.047 DINOFLAGELLIDA
Gymnodinium sp. 7 0.002 14 0.003 17 0.004
Gyrodinium fusiforme 18 0.012 9 0.006 13 0.009
Protoperidinium crassipes 5 0.057 0 0 0 0
Protoperidinium steini 10 0.009 8 0.008 4 0.004
FORAMINIFERIDA
Globigerina 5 0.004 3 0.002 5 0.004
RADIOLARIA
indet. 25 micron 0 0 2 0.002 5 0.006
METAZOA
Larve 0 0 3 0.006 3 0.007
COPEPODA
Copepoda nauplii 5 0.247 7 0.322 4 0.198

MICROZOOPLANCTON
Stazione V 10 ABC T0 Cell L-1 µgC L-1 Cell L-1 µgC L-1 Cell L-1 µgC L-1 OLIGOTRICHIDA Loboea strobilia 5 0.027 3 0.016 5 0.027
Lohmanniella sp. 12 0.003 6 0.002 18 0.005
Nanociliati 13 0.003 23 0.006 26 0.007
Oligotricha indet. 3 0.013 1 0.004 4 0.017
Strombidium capitatum 0 0 2 0.005 4 0.009
Strombidium conicum 4 0.009 3 0.007 2 0.005
Strombidium epidemum 3 0 0 0 3 0
Strombidium neptuni 0 0 1 0.007 0 0
Strombidium sp. 15 0.008 6 0.003 14 0.028
Tontonia sp. 0 0 0 0 3 0.016 HOLOTRICHIA
Holotrichia indet. 8 0.016 6 0.012 12 0.025 TINTINNIDA Codonella sp. 0 0 2 0.013 0 0
Dadayella ganymedes 2 0.002 2 0.002 0 0
Eutintinnus apertus 0 0 0 0 0 0
Eutintinnus macilentus 0 0 5 0.013 5 0
Eutintinnus tubulosus 4 0.008 2 0.004 1 0.002
Rhabdonella spiralis 3 0.028 3 0.028 4 0.038
Tintinnopsis nana 0 0 0 0 2 0.001
Tintinnopsis sp. 0 0 0 0 2 0.070 DINOFLAGELLIDA
Gymnodinium sp. 11 0.002 17 0.003 16 0.002
Gyrodinium fusiforme 9 0.006 8 0.005 14 0.010
Ornythocercus magnificus 0 0 0 0 3 0.010
Protoperidinium sp. 7 0.017 0 0 0 0
Protoperidinium crassipes 0 0 4 0.046 0 0
protoperidinium diabolus 0 0 0 0 4 0.065
Protoperidinium divergens 3 0.033 0 0 0 0
Protoperidinium steini 9 0.008 17 0.015 10 0.009
FORAMINIFERIDA
Globigerina 3 0.002 3 0.002 0 0
ACANTHARIA
Indet. 25 micron 3 0.003 0 0 0 0
RADIOLARIA
Indet. 25 micron 9 0.010 7 0.008 0 0
METAZOA
Larve micrometazoi 9 0.021 2 0.005 2 0.005
COPEPODA
Copepoda nauplii 6 0.297 4 0.198 4 0.221

MICROZOOPLANCTON
Stazione V6 ABC T48 Cell L-1 µgC L-1 Cell L-1 µgC L-1 Cell L-1 µgC L-1 OLIGOTRICHIDA Laboea strobila 3 0.016 0 0 3 0.016
Lohmanniella sp. 5 0.001 5 0.001 7 0.002
Nanociliati 9 0.002 24 0.006 29 0.008
Oligotricha indet. 12 0.052 6 0.026 5 0.021
Strombidium conicum 3 0.007 5 0.012 4 0.009
Strombidium emergens 0 0 2 0 0 0
Strombidium sp. 9 0.005 6 0.003 11 0.005 HOLOTRICHIA
Holotrichia indet. 5 0.010 5 0.010 4 0.008 TINTINNIDA
Eutintinnus lusus undae 0 0 0 0 2 0.010
Eutintinnus sp 0 0 0 0 3 0.057
Eutintinnus macilentus 0 0 3 0.008 0 0
Rhabdonella cornucopia 2 0.005 0 0 0 0
Rhabdonella spiralis 0 0 4 0.038 0 0
Rhabdonella sp. 0 0 2 0.006 0 0
Tintinnopsis minuta 0 0 0 0 3 0.002
Tintinnopsis sp 0 0 0 0 2 0.069
Undella sp 1 0 0 0 0 0 DINOFLAGELLIDA
Gymnodinium sp. 25 0.011 40 0.015 38 0.018
Gyrodinium fusiforme 32 0.011 26 0.021 18 0.058
Ornythocercus magnificus 0 3 0.021 0 0
Protoperidinium sp. 0 0 0 0 2 0.004
Protoperidinium crassipes 0 3 0.037 0 0
Protoperidinium diabolus 2 0.035 0 0 2 0.034
Protoperidinium divergens 4 0.048 3 0.036 3 0.035
Protoperidinium steini 24 0.023 13 0.013 6 0.006
Protoperidinium subinerme 6 0.137 0 0 2 0.004
FORAMINIFERIDA
Globigerina 1 0.001 3 0.003 5 0.005
RADIOLARIA
Indet 25 micron 5 0 5 0.006 4 0.004
METAZOA
Larve micrometazoi 2 0.005 2 0.005 2 0.002
COPEPODA
Copepoda nauplii 2 0.099 4 0.198 5 0.241

MICROZOOPLANCTON
Stazione V7 ABC T48 Cell L-1 µgC L-1 Cell L-1 µgC L-1 Cell L-1 µgC L-1 OLIGOTRICHIDA
Laboea strobila 4 0.021 3 0.015 3 0.016
Lohmanniella sp. 8 0.002 3 0.001 5 0.001
Nanociliati 30 0.008 22 0.006 31 0.008
Oligotricha indet. 6 0.025 5 0.020 5 0.030
Strombidium acutum 0 3 0.013 0 0
Strombidium conicum 3 0.007 0 0 2 0.005
Strombidium sp. 16 0.008 11 0.006 10 0.005 HOLOTRICHIA
Holotrichia indet. 10 0.020 6 0.004 5 0.010 TINTINNIDA
Codonella aspera 2 0.007 0 0 0 0 Eutintinnus apertus 0 0 0 0 1 0.002
Eutintinnus tubulosus 3 0.006 4 0 3 0.006
Rhabdonella spiralis 3 0.027 3 0.026 2 0.018
Rhabdonella sp. 2 0.006 0 0 2 0.006
Tintinnopsis compressa 2 0.005 0 0 0 0
Tintinnopsis sp 2 0.068 2 0.066 0 0
Xystonella sp. 0 0 1 0.008 0 0 DINOFLAGELLIDA
Gymnodinium sp. 23 0.023 13 0.012 13 0.016
Gyrodinium fusiforme 27 0.008 9 0.011 0 0
Ornythocercus magnificus 0 0 0 0 0 0
Protoperidinium sp. 3 0.006 4 0.007 0 0
Protoperidinium brevipes 4 0.007 2 0.003 0
Protoperidinium crassipes 0 0 3 0.035 5 0
Protoperidinium diabolus 2 0.034 3 0.049 3 0.050
Protoperidinium divergens 3 0.034 0 0 0 0
Protoperidinium oceanicum 2 0.107 0 0 0
Protoperidinium steini 18 0.018 18 0.017 11 0.010
Protoperidinium subinerme 3 0.007 0 0 0 0
FORAMINIFERIDA
Globigerina 3 0.003 2 0.002 2 0.002
RADIOLARIA
Indet. 25 micron 4 0.004 3 0.003 2 0.002
METAZOA
Larve micrometazoi 2 0.004 1 0.002 2 0.002
COPEPODA
Copepoda nauplii 4 0.198 3 0.139 4 0.190

MICROZOOPLANCTON
Stazione Viera A T72 Cell L-1 µgC L-1 Stazione V 10 A T72 Cell L-1 µgC L-1
OLIGOTRICHIDA OLIGOTRICHIDA
Laboea strobila 1 0.006 Loboea strobilia 3 0.016
Lohmanniella sp. 5 0.001 Lohmanniella sp. 4 0.001
Nanociliati 13 0.003 Nanociliati 15 0.004
Oligotricha indet. 5 0.023 Oligotricha indet. 3 0.013
Strombidium sp. 8 0.004 Strombidium acutum 0 0
HOLOTRICHIA Strombidium capitatum 0 0
Holotrichia indet. 0 0 Strombidium conicum 0 0
TINTINNIDA Strombidium epidemum 0 0
Eutintinnus lusus undae 0 0 Strombidium neptuni 0 0
Eutintinnus sp 0 0 Strombidium sp. 3 0.001
Eutintinnus macilentus 0 0 HOLOTRICHIA
Rhabdonella cornucopia 0 0 Holotrichia indet. 3 0.006
Rhabdonella spiralis 3 0.031 TINTINNIDA
Rhabdonella sp. 0 0 Craterella torulata 0 0
Tintinnopsis minuta 0 0 Epiplocylis sp 0 0
Tintinnopsis sp 0 0 Eutintinnus tubulosus 0 0
Undella sp 0 0 Rhabdonella spiralis 2 0.018
DINOFLAGELLIDA DINOFLAGELLIDA
Gymnodinium sp. 3 0.001 Gymnodinium sp. 14 0.013
Gyrodinium fusiforme 7 0.002 Gyrodinium fusiforme 17 0.015
Gyrodinium sp. 8 0.002 Protoperidinium diabolus 1 0.016
Ornythocercus magnificus 0 0 Protoperidinium oceanicum 2 0.108
Protoperidinium sp. 0 0 Protoperidinium steini 23 0.021
Protoperidinium crassipes 0 0 FORAMINIFERIDA
Protoperidinium diabolus 0 0 Globigerina 0 0
Protoperidinium divergens 0 0 RADIOLARIA
Protoperidinium steini 3 0.003 Indet. 40 micron 3 0.014
Protoperidinium subinerme 0 0 Indet. 60 micron 2 0.031
FORAMINIFERIDA METAZOA
Globigerina 4 0.004 Larve micrometazoi 0 0
RADIOLARIA COPEPODA
Indet. 25 micron 7 0.013 Copepoda nauplii 3 0.144
METAZOA
Larve micrometazoi 2 0.003
COPEPODA
Copepoda nauplii 2 0.061
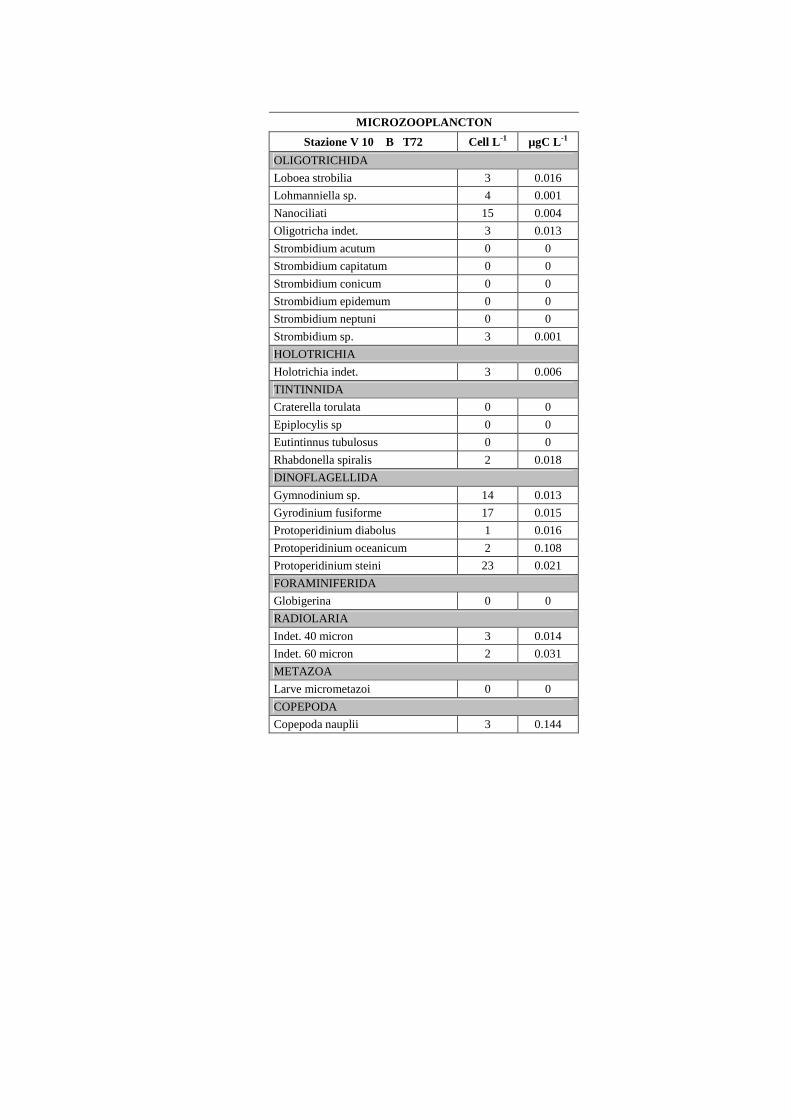
MICROZOOPLANCTON
Stazione V 10 B T72 Cell L-1 µgC L-1
OLIGOTRICHIDA
Loboea strobilia 3 0.016
Lohmanniella sp. 4 0.001
Nanociliati 15 0.004
Oligotricha indet. 3 0.013
Strombidium acutum 0 0
Strombidium capitatum 0 0
Strombidium conicum 0 0
Strombidium epidemum 0 0
Strombidium neptuni 0 0
Strombidium sp. 3 0.001
HOLOTRICHIA
Holotrichia indet. 3 0.006
TINTINNIDA
Craterella torulata 0 0
Epiplocylis sp 0 0
Eutintinnus tubulosus 0 0
Rhabdonella spiralis 2 0.018
DINOFLAGELLIDA
Gymnodinium sp. 14 0.013
Gyrodinium fusiforme 17 0.015
Protoperidinium diabolus 1 0.016
Protoperidinium oceanicum 2 0.108
Protoperidinium steini 23 0.021
FORAMINIFERIDA
Globigerina 0 0
RADIOLARIA
Indet. 40 micron 3 0.014
Indet. 60 micron 2 0.031
METAZOA
Larve micrometazoi 0 0
COPEPODA
Copepoda nauplii 3 0.144

Allegati
Ostreopsis ovata

FITOPLANCTON
T0 100% A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 38697,698 27785140,67 26015,3 37358172,32 38697,698 55570281,33
Thalassionema nitzschioides 12192,0161 172638,9473 18796 266151,7105 12635,983 178925,5197
Dactyliosolen fragilissimus 19348,849 7403643,585 19348,8 7403643,585 24815,6667 9495466,696
Cerataulina pelagica 11566,7845 11095669,33 13535,8 12984504,31 18427,4752 17676924,18
Cyclotella 197542,535 8930897,992 199017 8997546,484 197542,535 8930897,992
Thalassiosira indet. 114987,446 17616076,66 117199 17954847,36 97297,0693 14905911,02
Dinophyceae
Ostreopsis piccolo 19348,849 31010561,56 30073,6 48199272,83 24569,967 39378490,86
Ostreopsis medio 17690,3762 124427030,3 26275,4 184810736,2 20906,8083 147050126,7
Leucocryptos marina 4422,59406 167051,7127 116093 4385107,457 114987,446 4343344,529
T24 100% A B Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 1,24E+04 35282718,3 21522 82561560,84
Thalassionema nitzschioides 8,38E+03 234839,7446 5936,17 403141,5616
Dactyliosolen fragilissimus 1,04E+04 7845939,177 6508,79 12329332,99
Cerataulina pelagica 9,29E+03 17648641,1 5177,45 20404221,05
Cyclotella 1,45E+05 12996456,03 65885,6 13396346,99
Thalassiosira indet. 8,56E+04 25972420,71 32544 66760010,29
Dinophyceae
Ostreopsis piccolo 13794,5912 43779616,33 10253,4 37305938,71
Ostreopsis medio 8354,60778 116362315,4 8847,74 111095562,8
Leucocryptos marina 32105,2688 2021151,711 124078 4686728,605
T0 80% A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 41411,5626 59467417,94 42617,7 61199478,67 65707,1117 94356069,53
Thalassionema nitzschioides 8134,41407 115183,3033 11754,8 166447,8189 9786,16558 138572,1046
Dactyliosolen fragilissimus 19420,9565 7431234,803 12888,7 4931733,196 29023,2735 11105465,38
Cerataulina pelagica 14551,7611 13959067,87 10252,4 9834797,818 16871,3773 16184206,14
Cyclotella 88451,8812 3998909,549 130467 5898391,584 134889,119 6098337,061
Thalassiosira indet. 75921,198 11631127,54 64443,5 9872746,26 92137,3762 14115446,04
Dinophyceae
Ostreopsis piccolo 19420,9565 31126128,87 20303,7 32540952,9 19348,849 31010561,56
Ostreopsis medio 14409,0968 101347823,1 17520,3 123230616,5 9614,33491 67623386,03
Leucocryptos marina 66338,9109 2505775,69 76658,3 2895563,019 95528,0317 3608316,993

FITOPLANCTON
T24 80% A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 65327,2425 185763511,9 165110 237099867 129360,876 185763511,9
Thalassionema nitzschioides 10253,3823 287500,7781 15064,5 213312,768 20303,7273 287500,7781
Dactyliosolen fragilissimus 20303,7273 15384194,46 43488,8 16640570,34 40205,4005 15384194,46
Cerataulina pelagica 15038,294 28565909,48 27055,9 25953883,98 29778,8 28565909,48
Cyclotella 185373,03 16595474,63 247665 11196946,74 367075,307 16595474,63
Thalassiosira indet. 85614,05 25972420,71 185749 28456739,22 169532,772 25972420,71
Dinophyceae
Ostreopsis piccolo 6381,17143 20251795,31 18611,8 29829206,84 12635,983 20251795,31
Ostreopsis medio 4423,86981 61615308,28 13007,6 91490463,46 8760,13823 61615308,28
Leucocryptos marina 58627,0125 4385107,457 110565 4176292,816 116093,094 4385107,457
T0 50 % A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 19348,849 13892570,33 13007,6 18679086,16 19348,849 27785140,67
Thalassionema nitzschioides 6096,00803 86319,47367 9398,01 133075,8552 6317,99151 89462,75984
Dactyliosolen fragilissimus 9674,4245 3701821,793 9674,42 3701821,793 12407,8333 4747733,348
Cerataulina pelagica 5783,39223 5547834,666 6767,91 6492252,154 9213,73762 8838462,09
Cyclotella 98771,2673 4465448,996 99508,4 4498773,242 98771,2673 4465448,996
Thalassiosira indet. 57493,7228 8808038,329 58599,4 8977423,681 48648,5347 7452955,509
Dinophyceae
Ostreopsis piccolo 9674,4245 15505280,78 15036,8 24099636,41 12284,9835 19689245,43
Ostreopsis medio 8845,18812 62213515,15 13137,7 92405368,09 10453,4041 73525063,35
Leucocryptos marina 2211,29703 83525,85633 58046,5 2192553,729 57493,7228 2171672,264
T24 50 % A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 106142,257 152420977,1 124938 179412184,7 33485,3465 48085177,03
Thalassionema nitzschioides 41813,6238 15999705,48 12776,4 180913,4875 12192,0198 172639,0004
Dactyliosolen fragilissimus 11002,5545 155796,1711 50122,7 19178962,43 42617,7228 16307245,44
Cerataulina pelagica 37592,0594 36060934,83 40607,4 38953504,29 35040,5545 33613352,67
Cyclotella 210810,317 9530677,225 182801 8264363,467 232186,188 10497074,57
Thalassiosira indet. 141523,01 21681325,12 135995 20834399,11 147419,802 22584713,66
Dinophyceae
Ostreopsis piccolo 9503,88119 15231949,59 8504,99 13631018,51 11339,9802 18174680,77
Ostreopsis medio 9296,47525 65387688,3 8511,41 59865824,82 10529,9802 74063668,72
Leucocryptos marina 68866,099 2601233,491 20504,8 774512,4179 32221,7426 1217090,515

FITOPLANCTON
T0 10 % A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 9674,4245 6946285,167 6503,81 9339543,08 9674,4245 13892570,33
Thalassionema nitzschioides 3048,00401 43159,73684 4699,01 66537,92762 3158,99576 44731,37992
Dactyliosolen fragilissimus 4837,21225 1850910,896 4837,21 1850910,896 6203,91667 2373866,674
Cerataulina pelagica 2891,69612 2773917,333 3383,95 3246126,077 4606,86881 4419231,045
Cyclotella 49385,6337 2232724,498 49754,2 2249386,621 49385,6337 2232724,498
Thalassiosira indet. 28746,8614 4404019,164 29299,7 4488711,841 24324,2673 3726477,754
Dinophyceae
Ostreopsis piccolo 4837,21225 7752640,39 7518,41 12049818,21 6142,49175 9844622,715
Ostreopsis medio 4422,59406 31106757,58 6568,85 46202684,04 5226,70207 36762531,67
Leucocryptos marina 1105,64851 41762,92816 29023,3 1096276,864 28746,8614 1085836,132
T24 10 % A B C Cell L-1 pgC L-1 Cell L-1 pgC L-1 Cell L-1 pgC L-1
Bacillariophyceae
Chaetoceros decipiens 38329,1485 55040908,41 37592,1 53982443,62 23509,5842 33759916,89
Thalassionema nitzschioides 10593,6436 150005,9929 12285 4700728,611 10786,8119 4127465,698
Dactyliosolen fragilissimus 14373,4257 5499847,626 10530 149104,5196 11566,7921 163785,7758
Cerataulina pelagica 15794,9703 15151641,16 11167 10712215,58 11952,9505 11466106,82
Cyclotella 73709,901 3332404,624 120516 5448481,336 75921,2079 3432377,21
Thalassiosira indet. 55282,4356 8469269,141 58599,4 8977422,923 37592 5759094,4
Dinophyceae
Ostreopsis piccolo 6229,0099 9983286,081 5947,62 9532306,171 7250,15842 11619889,32
Ostreopsis medio 6030,81188 42418318,45 4914 34563110,4 5749,36634 40438743,07
Leucocryptos marina 23509,5842 888011,9325 27641,2 1044073,017 22997,4851 868668,7562

MICROZOOPLANCTON GIUGNO 2009 Pirano 100%
A T0 Cell L-1 pg C L-1
Pirano 100%
B T0 Cell L-1 pg C L-1
Pirano 100%
C T0 Cell L-1 pg C L-1
OLIGOTRICHIDA OLIGOTRICHIDA OLIGOTRICHIDA
Laboea strobila 3 16645.45 Laboea strobila 4 22420.41 Laboea strobila 13 72130.30
Legardiella sp 0 0.00 Legardiella sp. 4 3163.27 Legardiella sp. 0 0.00
Lohmanniella sp. 6 1654.55 Lohmanniella sp. 12 3342.86 Lohmanniella sp. 26 7169.70
Nanociliati 38 10094.95 Nanociliati 24 6440.82 Nanociliati 24 6375.76
Oligotricha indet. 4 17559.60 Oligotricha indet. 6 26608.16 Oligotricha indet. 10 43898.99
Strombidium conicum 21 49933.33 Strombidium conicum 18 43236.73 Strombidium conicum 25 59444.44
Strombidium sp. 16 18505.05 Strombidium sp. 16 8179.59 Strombidium sp. 28 14169.70
Tontonia sp. 2 10878.79 Tontonia sp. 0 0.00 Tontonia sp. 3 16318.18
HOLOTRICHIA HOLOTRICHIA HOLOTRICHIA
Holotrichia indet. 11 22822.22 Holotrichia indet. 9 18863.27 Holotrichia indet. 24 49050.75
TINTINNIDA TINTINNIDA TINTINNIDA
Canthariella brevis 2 1404.04 Canthariella brevis 0 0.00 Canthariella brevis 0 0.00
Eutintinnus tubulosus 3 6048.48 Eutintinnus tubulosus 0 0.00 Eutintinnus tubulosus 2 4032.28
Eutintinnus apertus 33 78533.33 Eutintinnus apertus 27 62506.12 Eutintinnus apertus 35 83292.93
Eutintinnus fraknoi 14 167490.91 Eutintinnus fraknoi 11 132942.86 Eutintinnus fraknoi 7 83745.45
Eutintinnus lusus-undae 1 5052.53 Eutintinnus lusus-undae 0 0.00 Eutintinnus lusus-undae 0 0.00
Rhabdonella cuspidata 1 3012.12 Rhabdonella cuspidata 3 9128.57 Rhabdonella cuspidata 0 0.00
Stentrupiella gracilis 1 1226.26 Stentrupiella gracilis 2 2477.55 Stentrupiella gracilis 0 0.00
Stenosemella nivalis 1 1062.63 Stenosemella nivalis 0 0.00 Stenosemella nivalis 0 0.00
Tintinnopsis minuta 0 0.00 Tintinnopsis minuta 7 3792.86 Tintinnopsis minuta 4 2145.45
Eutintinnus spp. 1 19748.48 Eutintinnus spp. 2 39900.00 Eutintinnus spp. 0 0.00

MICROZOOPLANCTON GIUGNO 2009
Pirano 100% A T0
Cell L-1 pg C L-1 Pirano 100% B T0
Cell L-1 pg C L-1 Pirano 100% C T0
Cell L-1 pg C L-1
DINOFLAGELLIDA DINOFLAGELLIDA DINOFLAGELLIDA
Gymnodinium sp. 27 6000.00 Gymnodinium sp. 41 8979.59 Gymnodinium sp. 30 6666.67
Gyrodinium fusiforme 58 127210.39 Gyrodinium fusiforme 95 64530.61 Gyrodinium fusiforme 85 57696.97
Protoperidinium bipes 32 565171.72
Protoperidinium bipes 20 356836.73
Protoperidinium bipes 29 512186.87
Protoperidinium crassipes 3 37375.76 Protoperidinium crassipes 0 0.00 Protoperidinium crassipes 0 0.00
Protoperidinium sp. 4 9850.51 Protoperidinium sp. 9 18165.31 Protoperidinium sp. 20 39959.60
Protoperidinium diabolus 3 52984.85 Protoperidinium diabolus 0 0.00 Protoperidinium diabolus 0 0.00
Protoperidinium depressum 3 181245.45 Protoperidinium depressum 0 0.00 Protoperidinium depressum 0 0.00
Protoperidinium divergens 5 55914.14 Protoperidinium divergens 4 45187.76 Protoperidinium divergens 4 48173.74
Protoperidinium steini 11 10733.33 Protoperidinium steini 6 5914.29 Protoperidinium steini 6 5854.55
ACANTHARIA Protoperidinium subinerme 2 4651.02 Protoperidinium subinerme 3 6906.06
indet 25 µm 4 4626.26 indet 25 µm 0 0.00 ACANTHARIA
FORAMINIFERIDA FORAMINIFERIDA indet 25 µm 2 2313.13
Globigerina 12 9236.36 Globigerina 9 6997.96 FORAMINIFERIDA
RADIOLARIA RADIOLARIA Globigerina 19 19115.15
Radiolaria indet. 7 8095.96 Radiolaria indet. 5 5841.84 RADIOLARIA
METAZOA METAZOA Radiolaria indet. 10 47363.64
Larve micrometazoi 2 2642.42 Larve micrometazoi 6 8008.16 METAZOA
COPEPODA COPEPODA Larve micrometazoi 7 9248.48
Copepoda nauplii 34 1698901.01 Copepoda nauplii 57 2826742.86 COPEPODA
Copepoda nauplii 55 2698254.55

MICROZOOPLANCTON GIUGNO 2009
Pirano 100% A T24
Cell L-1 pg C L-1
Pirano 100% B T24
Cell L-1 pg C L-1
Pirano 100% C T24
Cell L-1 pg C L-1
OLIGOTRICHIDA OLIGOTRICHIDA OLIGOTRICHIDA
Laboea strobila 3 19023.38 Laboea strobila 2 10666.02 Laboea strobila 5 27742.42
Lohmanniella sp. 16 4254.55 Lohmanniella sp. 6 1590.29 Lohmanniella sp. 22 6066.67
Nanociliati 7 1821.65 Nanociliati 14 3574.76 Nanociliati 19 5047.47
Oligotricha indet. 6 26339.39 Oligotricha indet. 8 33755.34 Oligotricha indet. 3 13169.70
Strombidium conicum 8 18342.86 Strombidium conicum 4 9141.75 Strombidium conicum 18 42800.00
Strombidium sp. 29 14314.29 Strombidium sp. 42 20915.53 Strombidium sp. 16 8096.97
HOLOTRICHIA HOLOTRICHIA HOLOTRICHIA
Holotrichia indet. 3 7113.42 Holotrichia indet. 0 0.00 Holotrichia indet. 7 14523.23
TINTINNIDA TINTINNIDA TINTINNIDA
Eutintinnus apertus 23 38561.04 Eutintinnus apertus 18 31599.03 Eutintinnus apertus 40 69212.12
Eutintinnus fraknoi 25 297381.82 Eutintinnus fraknoi 14 160986.41 Eutintinnus fraknoi 28 334981.82
Eutintinnus minuta 2 33854.55 Eutintinnus minuta 2 37963.11 Eutintinnus minuta 0 0.00
Rhabdonella spiralis 3 32561.04 Rhabdonella spiralis 3 27384.47 Rhabdonella spiralis 0 0.00
Tintinnopsis minuta 3 1379.22 Tintinnopsis minuta 0 0.00 Tintinnopsis minuta 9 4827.27
Salpingella rotundata 0 0.00 Salpingella rotundata 0 0.00 Salpingella rotundata 2 6676.77
DINOFLAGELLIDA DINOFLAGELLIDA DINOFLAGELLIDA
Gymnodinium sp. 58 26625.97 Gymnodinium sp. 38 17379.61 Gymnodinium sp. 36 16690.91
Gyrodinium fusiforme 55 37679.65 Gyrodinium fusiforme 74 50174.76 Gyrodinium fusiforme 55 37090.91
Protoperidinium bipes 19 333047.62 Protoperidinium bipes 22 390441.75 Protoperidinium bipes 25 441540.40
Protoperidinium crassipes 4 49580.09 Protoperidinium crassipes 0 0.00 Protoperidinium crassipes 6 69412.12
Protoperidinium divergens 7 82583.55 Protoperidinium divergens 10 115757.28 Protoperidinium divergens 3 36130.30
Protoperidinium steini 30 29272.73 Protoperidinium steini 24 23446.60 Protoperidinium steini 34 33175.76
Protoperidinium subinerme 0 0.00 Protoperidinium subinerme 3 6163.11 Protoperidinium diabolus 4 65600.00
Protoperidnium diabolus 0 0.00 Protoperidinium diabolus 1 15763.11 Protoperidinium depressum 6 5630.30
FORAMINIFERIDA FORAMINIFERIDA FORAMINIFERIDA
Globigerina 3 2587.01 Globigerina 4 3867.96 Globigerina 9 9054.55
RADIOLARIA RADIOLARIA RADIOLARIA
Indet. 25 micron 3 Indet 25 micron 2 2217.48 Indet 25 micron 7 8074.75

MICROZOOPLANCTON GIUGNO 2009
Pirano 100% A T24 Cell L-1 pg C L-1
Pirano 100% B T24 Cell L-1 pg C L-1
Pirano 100% C T24 Cell L-1 pg C L-1
METAZOA 3 7930.74 METAZOA 4 8893.20 METAZOA 4 3151.52
Larve micrometazoi 3 7930.74 Larve micrometazoi 4 8893.20 Larve micrometazoi 4 3151.52
COPEPODA 42 2055812.99 COPEPODA 45 2209250.49 COPEPODA 49 2448416.16
Copepoda nauplii 42 2055812.99 Copepoda nauplii 45 2209250.49 Copepoda nauplii 49 2448416.16
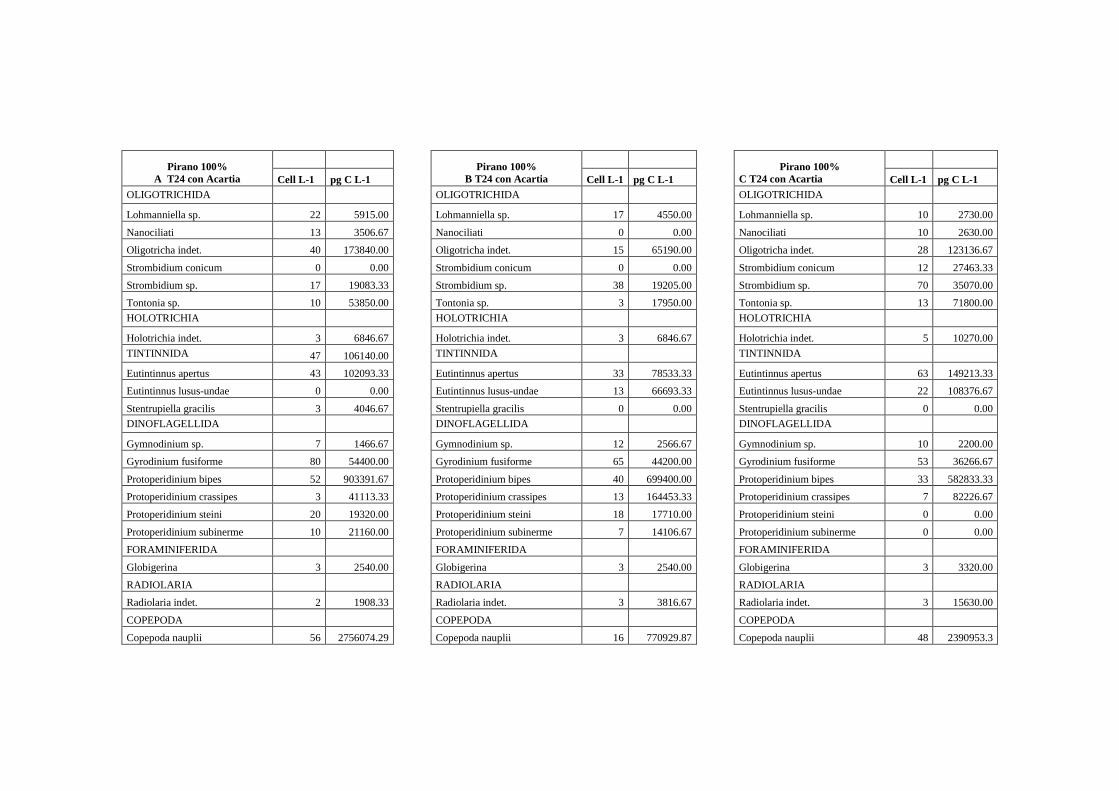
Pirano 100%
A T24 con Acartia Cell L-1 pg C L-1 Pirano 100%
B T24 con Acartia Cell L-1 pg C L-1 Pirano 100%
C T24 con Acartia Cell L-1 pg C L-1 OLIGOTRICHIDA OLIGOTRICHIDA OLIGOTRICHIDA
Lohmanniella sp. 22 5915.00 Lohmanniella sp. 17 4550.00 Lohmanniella sp. 10 2730.00
Nanociliati 13 3506.67 Nanociliati 0 0.00 Nanociliati 10 2630.00
Oligotricha indet. 40 173840.00 Oligotricha indet. 15 65190.00 Oligotricha indet. 28 123136.67
Strombidium conicum 0 0.00 Strombidium conicum 0 0.00 Strombidium conicum 12 27463.33
Strombidium sp. 17 19083.33 Strombidium sp. 38 19205.00 Strombidium sp. 70 35070.00
Tontonia sp. 10 53850.00 Tontonia sp. 3 17950.00 Tontonia sp. 13 71800.00
HOLOTRICHIA HOLOTRICHIA HOLOTRICHIA
Holotrichia indet. 3 6846.67 Holotrichia indet. 3 6846.67 Holotrichia indet. 5 10270.00
TINTINNIDA 47 106140.00 TINTINNIDA TINTINNIDA
Eutintinnus apertus 43 102093.33 Eutintinnus apertus 33 78533.33 Eutintinnus apertus 63 149213.33
Eutintinnus lusus-undae 0 0.00 Eutintinnus lusus-undae 13 66693.33 Eutintinnus lusus-undae 22 108376.67
Stentrupiella gracilis 3 4046.67 Stentrupiella gracilis 0 0.00 Stentrupiella gracilis 0 0.00
DINOFLAGELLIDA DINOFLAGELLIDA DINOFLAGELLIDA
Gymnodinium sp. 7 1466.67 Gymnodinium sp. 12 2566.67 Gymnodinium sp. 10 2200.00
Gyrodinium fusiforme 80 54400.00 Gyrodinium fusiforme 65 44200.00 Gyrodinium fusiforme 53 36266.67
Protoperidinium bipes 52 903391.67 Protoperidinium bipes 40 699400.00 Protoperidinium bipes 33 582833.33
Protoperidinium crassipes 3 41113.33 Protoperidinium crassipes 13 164453.33 Protoperidinium crassipes 7 82226.67
Protoperidinium steini 20 19320.00 Protoperidinium steini 18 17710.00 Protoperidinium steini 0 0.00
Protoperidinium subinerme 10 21160.00 Protoperidinium subinerme 7 14106.67 Protoperidinium subinerme 0 0.00
FORAMINIFERIDA FORAMINIFERIDA FORAMINIFERIDA
Globigerina 3 2540.00 Globigerina 3 2540.00 Globigerina 3 3320.00
RADIOLARIA RADIOLARIA RADIOLARIA
Radiolaria indet. 2 1908.33 Radiolaria indet. 3 3816.67 Radiolaria indet. 3 15630.00
COPEPODA COPEPODA COPEPODA
Copepoda nauplii 56 2756074.29 Copepoda nauplii 16 770929.87 Copepoda nauplii 48 2390953.3

MICROZOOPLANCTON SETTEMBRE 2009
Pirano 100% A T0 CellL-1 pg C L-1
Pirano 100% B T0 CellL-1 pg C L-1
Pirano 100% C T0 CellL-1 pg C L-1
OLIGOTRICHIDA OLIGOTRICHIDA OLIGOTRICHIDA
Nanociliati 10-20 micron 9 1722.86 Nanociliati 10-20 micron 10 2010.00 Nanociliati 10-20 micron 7 1435.71
Oligotricha indet. 94 409765.71 Oligotricha indet. 71 310428.57 Oligotricha indet. 83 360097.14
Strombidium conicum 21 360263.57 Strombidium conicum 36 600439.29 Strombidium conicum 50 840615.00
Strombidium sp. 140 70140.00 Strombidium sp. 113 56541.43 Strombidium sp. 86 42942.86
Strombidium neptuni 14 44875.80 Strombidium neptuni 17 53850.96 Strombidium neptuni 14 44875.80
Tontonia spp. 31 1208900.00 Tontonia spp. 27 1044050.00 Tontonia spp. 26 989100.00
HOLOTRICHIA HOLOTRICHIA HOLOTRICHIA
Holotrichia indet. 6 11737.14 Holotrichia indet. 17 35211.43 Holotrichia indet. 20 41080.00
TINTINNIDA TINTINNIDA TINTINNIDA
Codonellopsis sp. 9 30857.14 Codonellopsis sp. 6 20571.43 Codonellopsis sp. 6 20571.43
DINOFLAGELLIDA DINOFLAGELLIDA DINOFLAGELLIDA
Gymnodinium sp. 20 3120.00 Gymnodinium sp. 17 2674.29 Gymnodinium sp. 23 3565.71
Gyrodinium fusiforme 54 36914.29 Gyrodinium fusiforme 46 31085.71 Gyrodinium fusiforme 40 27200.00
Protoperidinium sp. 4 5674.29 Protoperidinium sp. 3 3782.86 METAZOA
RADIOLARIA RADIOLARIA Metazoa ova 11 8914.29
Radiolari indet. 10 11450.00 Radiolari indet 0 0.00 EBRIDA
ACANTHARIA ACANTHARIA Hermesinum adriaticum 27 16638.57
indet. 0 0.00 indet. 0 0.00 COPEPODA
METAZOA METAZOA Copepoda nauplii 16 791488.00
Metazoa ova 9 6685.71 Metazoa ova 6 4457.14
EBRIDA EBRIDA
Hermesinum adriaticum 29 17514.29 Hermesinum adriaticum 40 24520.00
COPEPODA COPEPODA
Copepoda nauplii 56 2756074.29 Copepoda nauplii 16 770929.87

MICROZOOPLANCTON SETTEMBRE 2009
Pirano 100% A T24 Bianco
CellL-1
pg C L-1
Pirano 100% B T24 Bianco
CellL-1
pg C L-1
Pirano 100% C T24 Bianco
CellL-1
pg C L-1
OLIGOTRICHIDA OLIGOTRICHIDA OLIGOTRICHIDA
Laboea strobila 0 0.00 Laboea strobila 0 0.00 Laboea strobila 4 23541.43
Lohmanniella sp. 83 22620.00 Lohmanniella sp. 51 13889.47 Lohmanniella sp. 21 5850.00
Nanociliati 21 5635.71 Nanociliati 39 10150.88 Nanociliati 51 13525.71
Oligotricha indet. 229 993371.43 Oligotricha indet. 239 1036940.35 Oligotricha indet. 231 1005788.57
Strombidium conicum 57 134514.29 Strombidium conicum 102 239529.82 Strombidium conicum 140 329560.00
Strombidium sp. 169 193014.29 Strombidium sp. 165 188824.56 Strombidium sp. 246 281342.86
Strombidium neptuni 46 143602.56 Strombidium neptuni 35 110221.26 Strombidium neptuni 51 161552.88
Tontonia sp. 20 107700.00 Tontonia sp. 14 75578.95 HOLOTRICHIA
HOLOTRICHIA HOLOTRICHIA Holotrichia indet. 6 11737.14
Holotrichia indet. 9 17605.71 Holotrichia indet. 11 21621.05 TINTINNIDA
TINTINNIDA TINTINNIDA Codonellopsis sp. 10 36000.00
Codonellopsis sp. 9 30857.14 Codonellopsis sp. 12 44210.53 Eutintinnus macilentus 0 0.00
Tintinnopsis sp. 4 150527.14 Eutintinnus macilentus 5 20157.89 Salpingella decurtata 7 11364.29
Salpingella decurtata 0 0.00 Salpingella decurtata 0 0.00 DINOFLAGELLIDA
DINOFLAGELLIDA DINOFLAGELLIDA Gymnodinium sp. 19 4085.71
Gymnodinium sp. 20 4400.00 Gymnodinium sp. 18 3859.65 Gyrodinium fusiforme 19 12628.57
Gyrodinium fusiforme 11 7771.43 Gyrodinium fusiforme 9 5964.91 Protoperidinium divergens 0 0.00
Protoperidinium divergens 0 0.00 Protoperidinium divergens 0 0.00 Protoperidinium diabolus 1 24978.57
Protoperidinium diabolus 1 24978.57 Protoperidinium diabolus 2 30675.44 Protoperidinium steini 4 4140.00
Protoperidinium steini 4 4140.00 Protoperidinium steini 2 1694.74 EBRIDA
EBRIDA EBRIDA Hermesinum adriaticum 34 21017.14
Hermesinum adriaticum 49 29774.29 Hermesinum adriaticum 42 25810.53 COPEPODA
COPEPODA COPEPODA Copepoda nauplii 28 1399094.95
Copepoda nauplii 37 1837382.86 Copepoda nauplii 18 899418.18

Pirano 100%
A T24 + Centropages CellL-1 pg C L-1 Pirano 100%
A T24 + Centropages CellL-1 pg C L-1 Pirano 100%
A T24 + Centropages CellL-1 pg C L-1 OLIGOTRICHIDA OLIGOTRICHIDA OLIGOTRICHIDA
Nanociliati 46 12203.20 Nanociliati 24 6312.00 Nanociliati 22 5891.20
Oligotricha indet. 182 792710.40 Oligotricha indet. 147 639731.20 Oligotricha indet. 214 931782.40
Strombidium sp. 26 29312.00 Strombidium sp. 19 9619.20 Strombidium sp. 30 15230.40
Strombilidium neptuni 32 100521.79 Strombilidium neptuni 38 120626.15 Strombilidium neptuni 34 105547.88
DINOFLAGELLIDA DINOFLAGELLIDA DINOFLAGELLIDA
Gymnodinium sp. 5 1056.00 Gymnodinium sp. 3 704.00 Gymnodinium sp. 6 1408.00
Gyrodinium fusiforme 10 6528.00 Gyrodinium fusiforme 8 5440.00 Gyrodinium fusiforme 5 3264.00
Protoperidinium steini 5 4636.80 Protoperidinium steini 0 0.00 Protoperidinium steini 3 3091.20
EBRIIDA EBRIIDA EBRIIDA
Hermesinum adriaticum 42 25484.61 Hermesinum adriaticum 26 15682.83 Hermesinum adriaticum 29 17643.19
METAZOA METAZOA METAZOA
Larve micrometazoi 4 5284.85 Larve micrometazoi 3 4185.60 Larve micrometazoi 3 4185.60
COPEPODA COPEPODA COPEPODA
Copepoda nauplii 8 399741.41 Copepoda nauplii 27 1345529.60 Copepoda nauplii 37 1820422.40

Allegati
Antartide

FITOPLANCTON T0
A FITOPLANCTON T0
B FITOPLANCTON T0
C Rapp. Diluiz 1:0 cell/litro �gC/litro cell/litro µgrC/litro cell/litro �gC/litro
Bacillariophyceae Actinocyclus actinochilus 10 0.010 Chaetoceros cf. dichaeta 60 0.015 Actinocyclus actinochilus 70 0.073 Chaetoceros sp. 90 0.013 Cylindrotheca closterium 270 0.004 Chaetoceros sp. 20 0.003 Coscinodiscus sp. 50 0.192 Fragilariopsis cylindrus 1110 0.138 Coscinodiscus sp. 30 0.115 Cylindrotheca closterium 430 0.007 Fragilariopsis kerguelensis 1860 0.180 Cylindrotheca closterium 460 0.007 Fragilariopsis cylindrus 520 0.064 Fragilariopsis sp. 960 0.106 Fragilariopsis cf. curta 270 0.016 Fragilariopsis kerguelensis 2200 0.213 Navicula sp 20 0.033 Fragilariopsis cylindrus 2000 0.248 Fragilariopsis sp. 550 0.061 Pseudo-nitzschia cf seriata 110 0.019 Fragilariopsis kerguelensis 1730 0.167 Pseudo-nitzschia cf. seriata 110 0.014 Pseudo-nitzschia sp. 100 0.002 Fragilariopsis sp. 810 0.089 Pseudo-nitzschia sp. 30 0.003 Stellarima microtrias 30 0.061 Navicula sp 20 0.033 Stellarima microtrias 40 0.004 Thalassiosira sp. 20 0.008 Nitschia sp. 40 0.170
Thalassiosira sp. 130 0.499 Forme centriche indet. (> 10 µm) 10 0.020 Pseudo-nitzschia cf. pseudodelicatissima
20 0.000
Forme centriche indet. (> 10 µm) 30 0.004 Pseudo-nitzschia cf seriata 280 0.049 Pseudo-nitzschia sp. 20 0.000 Stellarima microtrias 20 0.041 Thalassiosira sp. 240 0.101
Forme centriche indet. (> 10 µm)
100 0.197
Dinophyceae Amphidinium sp. 10 0.002 Gyrodinium spp. 20 0.028 Amphidinium sp. 20 0.004 Gymnodinium sp. 280 0.304 Prorocentrum micans 10 0.023 Gymnodinium sp. 90 0.098 Gyrodinium sp. 160 0.227 Prorocentrum sp. 50 0.145 Gyrodinium sp. 30 0.042 Prorocentrum cf. antarcticum 60 0.206 Forme indeterminate > 10 µm 50 0.301 Prorocentrum cf. antarcticum 50 0.172 Forme indeterminate (20-100 µm) 250 1.503 Prorocentrum sp. 10 0.029 Forme indet. (20-100 µm) 140 0.842
Cryptophyceae Forme indeterminate 90 0.001 Forme indeterminate 70 0.001 Forme indeterminate 140 0.002
ANTARTIDE ESPERIMENTO 1

FITOPLANCTON T0 A
FITOPLANCTON T0 B
FITOPLANCTON T0 C
Rapp. diluiz 1:1 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Bacillariophyceae
Cylindrotheca closterium 300 0.005 Cylindrotheca closterium 110 0.002 Actinocyclus actinochilus 10 0.010 Fragilariopsis cylindrus 110 0.014 Fragilariopsis cylindrus 140 0.017 Cylindrotheca closterium 40 0.001 Fragilariopsis kerguelensis 380 0.037 Fragilariopsis kerguelensis 730 0.071 Fragilariopsis cylindrus 290 0.036 Fragilariopsis sp. 320 0.035 Fragilariopsis sp. 250 0.028 Fragilariopsis kerguelensis 730 0.071 Navicula sp 50 0.083 Pleurosigma spp. 10 0.018 Fragilariopsis sp. 90 0.010 Peudo-nitzschia pseudodelicatissima 10 0.000
Pseudo-nitzschia pseudodelicatissima 20 0.000 Nitzschia sp. 10 0.043
Peudo-nitzschia seriata 40 0.007 Pseudo-nitzschia cf seriata 150 0.026 Pseudo-nitzschia pseudodelicatissima 10 0.000
Peudo-nitzschia sp. 40 0.001 Stellarima microtrias 30 0.061 Pseudo-nitzschia cf seriata 80 0.014 Stellarima microtrias 10 0.020 Thalassiosira sp. 10 0.004 Pseudo-nitzschia sp. 20 0.000
Thalassiosira sp. 30 0.013 Forme centriche indeterminate > 10 µm 50 0.098 Stellarima microtrias 20 0.041
Forme centriche indeterminate > 10 um 300 0.590
Thalassiosira sp. 180 0.076
Forme pennate indeterminate 30 0.059
Forme centriche indeterminate > 10 µm 80 0.157
Forme indeterminate > 10 µm 10 0.020 Dinophyceae
Prorocentrum micans 60 0.140 Amphidinium sp. 10 0.002 Gymnodinium spp. 20 0.022 Gymnodinium sp. 20 0.022 Prorocentrum cf. antarcticum 10 0.034 Prorocentrum micans 10 0.023 Forme indeterminate > 10 µm 70 0.421 Forme indeterminate > 10 µm 40 0.240
Cryptophyceae Forme indeterminate 220 0.003 Forme indeterminate 130 0.002
ANTARTIDE ESPERIMENTO 1

FITOPLANCTON T0 A
FITOPLANCTON T0 B
FITOPLANCTON T0 C
Rapp. diluiz. 1:3 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Bacillariophyceae
Cylindrotheca closterium 150 0,002 Cylindrotheca closterium 40 0,001 Actinocyclus actinochilus 10 0,010 Fragilariopsis cylindrus 220 0,027 Fragilariopsis cf. curta 20 0,001 Cylindrotheca closterium 80 0,001 Fragilariopsis kerguelensis 1310 0,127 Fragilariopsis cylindrus 900 0,112 Fragilariopsis cylindrus 430 0,053 Fragilariopsis sp. 990 0,109 Fragilariopsis kerguelensis 1490 0,144 Fragilariopsis kerguelensis 310 0,030 Navicula sp 30 0,050 Fragilariopsis sp. 380 0,042 Fragilariopsis sp. 40 0,004 Pseudo-nitzschia cf. seriata 160 0,028 Navicula sp 20 0,033 Nitzschia sp. 20 0,085 Pseudo-nitzschia spp. Nitzschia sp. 30 0,128 Pseudo-nitzschia seriata 80 0,010 Stellarima microtrias 40 0,081 Pseudo-nitzschia cf. seriata 70 0,009 Thalassiosira sp. 30 0,115
Thalassiosira spp. 10 0,004 Thalassiosira sp. 30 0,115 Forme centriche indeterminate > 10 µm 20 0,002
Forme centriche indeterminate > 10 um 180 0,354
Forme centriche indeterminate > 10 µm 50 0,006
Dinophyceae Gymnodinium spp. 20 0,022 Gymnodinium spp. 10 0,011 Gymnodinium spp. 20 0,022 Forme indeterminate 10 0,060 Prorocentrum sp. 10 0,029 Prorocentrum cf. antarcticum 10 0,034 Forme indeterminate > 10 µm 30 0,180 Forme indeterminate 30 0,180
Cryptophyceae Forme indeterminate 40 0.001 Forme indeterminate 130 0,002 Forme indeterminate 70 0,001
ANTARTIDE ESPERIMENTO 1

FITOPLANCTON T0 A
FITOPLANCTON T0 B
FITOPLANCTON T0 C
Rapp. Diluiz. 3:1 Cell L-1 �gCL-1 Cell L-1 �gCL-1 Cell L-1 �gCL-1 Bacillariophyceae
Cylindrotheca closterium 80 0,001 Cylindrotheca closterium 260 0,004 Cylindrotheca closterium 90 0,001 Fragilariopsis cylindrus 720 0,089 Fragilariopsis cf. curta 30 0,002 Fragilariopsis cylindrus 720 0,089 Fragilariopsis kerguelensis 540 0,052 Fragilariopsis cylindrus 900 0,112 Fragilariopsis kerguelensis 1160 0,112 Fragilariopsis sp. 770 0,085 Fragilariopsis kerguelensis 1200 0,116 Fragilariopsis sp. 170 0,019 Navicula sp 10 0,017 Fragilariopsis sp. 210 0,023 Nitzschia sp. 40 0,170 Pleurosigma sp. 20 0,036 Navicula sp 20 0,033 Pleurosigma sp. 10 0,018 Pseudo-nitzschia pseudodelicatissima 10 0,000 Nitzschia sp. 20 0,085 Pseudo-nitzschia cf. seriata 70 0,009 Pseudo-nitzschia cf seriata 140 0,017 Pseudo-nitzschia cf. seriata 230 0,029 Pseudo-nitzschia sp. 40 0,004 Pseudo-nitzschia sp. 30 0,003 Pseudo-nitzschia sp. 50 0,005 Stellarima microtrias 40 0,004 Thalassiosira sp. 40 0,154 Stellarima microtrias 20 0,002 Thalassiosira sp. 160 0,614 Forme centriche indeterminate > 10 µm 90 0,011 Thalassiosira sp. 90 0,345
Forme centriche indeterminate > 10 µm 30 0,004
Forme centriche indeterminate > 10 µm 90 0,011
Dinophyceae Gymnodinium spp. (10 - 20 µm) 10 0,011 Gymnodinium spp. 50 0,054 Gymnodinium spp. 50 0,054 Prorocentrum micans 70 0,164 Gyrodinium sp. 10 0,014 Gyrodinium spp. 30 0,042 Forme indeterminate > 10 µm 10 0,060 Prorocentrum cf. antarcticum 40 0,138 Prorocentrum cf. antarcticum 40 0,138 Forme indeterminate (20-100 µm) 90 0,541 Prorocentrum sp. 10 0,029 Forme indet (20-100µm) 40 0,240
Cryptophyceae Forme indeterminate 110 0.001 Forme indeterminate 120 0.002 Forme indeterminate 70 0.001
ANTARTIDE ESPERIMENTO 1

FITOPLANCTON T0
A FITOPLANCTON T0
B FITOPLANCTON T0
C Rapp. diluiz. 1:0 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Cell L-1 �gC L-1
Bacillariophyceae Actinocyclus actinochilus 20 0,025 Actinocyclus actinochilus 10 0,012 Actinocyclus actinochilus 20 0,025 Chaetoceros cf. dichaeta 10 0,002 Chaetoceros cf convolutus 10 0,000 Chaetoceros sp. 60 0,008 Chaetoceros sp. 100 0,014 Chaetoceros cf dichaeta 20 0,005 Corethron criophylum 80 0,495 Coscinodiscus sp. 10 0,002 Chaetoceros sp. 200 0,028 Coscinodiscus spp. 10 0,002 Cylindrotheca closterium 9320 0,144 Corethron criophylum 50 0,310 Cylindrotheca closterium 880 0,014 Fragilariopsis cylindrus 1210 0,229 Cylindrotheca closterium 1430 0,022 Fragilariopsis cf. curta 30 0,002 Fragilariopsis kerguelensis 740 0,102 Fragilariopsis cylindrus 560 0,106 Fragilariopsis cylindrus 190 0,036 Fragilariopsis sp. 540 0,147 Fragilariopsis kerguelensis 420 0,058 Fragilariopsis kerguelensis 230 0,032 Nitschia sp. 60 0,256 Fragilariopsis sp. 290 0,079 Fragilariopsis sp. 200 0,055 Pseudo-nitzschia cf. pseudodelicatissima
180 0,003 Navicula sp. 60 0,099 Navicula sp. 40 0,066
Pseudo-nitzschia cf seriata 360 0,053 Nitschia sp. 30 0,128 Nitzschia sp. 20 0,085
Pseudo-nitzschia sp. 40 0,001 Pseudo-nitzschia cf. pseudodelicatissima 90 0,002
Pseudo-nitzschia cf. pseudodelicatissima 30 0,001
Stellarima microtrias 10 0,016 Pseudo-nitzschia cf seriata 410 0,060 Pseudo-nitzschia cf seriata 230 0,034 Thalassiosira sp. 120 0,041 Pseudo-nitzschia sp. 100 0,002 Pseudo-nitzschia sp. 90 0,002
60 0,118 Stellarima microtrias 10 0,016 Thalassiosira sp. 180 0,062 Forme centriche indet > 10 µm Thalassiosira sp. 240 0,082 10 0,020
Forme centriche indet > 10 µm 30 0,059
Forme centriche indet > 10 µm
Dinophyceae Amphidinium spp. (10 - 50 µm) 850 0,181 Amphidinium sp. 10 - 50 µm 20 0,004 Amphidinium sp. 20 0,004 Gymnodinium sp. 610 0,522 Gymnodinium cf. baccatum 30 0,016 Gymnodinium cf. diploconus 10 0,254 Gyrodinium sp. 1290 0,279 Gymnodinium sp. 10 - 50 µm 2330 1,996 Gymnodinium sp. 10 -50 µm 1330 1,139 Prorocentrum cf. antarcticum 30 0,115 Gymnodinium sp. > 50 µm 90 0,675 Gymnodinium sp. > 50 µm 50 0,375 Prorocentrum sp. 50 0,192 Gyrodinium cf. fusiforme 30 0,133 Gyrodinium sp. 10 - 50 µm 6540 1,413 Forme indeterminate > 10 µm 1720 1,034 Gyrodinium sp. 10 - 50 µm 2630 0,568 Gyrodinium sp. > 50 µm 30 0,184 Forme indeterminate > 10 µm 1350 0,811 Prorocentrum cf. antarcticum 40 0,153

FITOPLANCTON T0
A FITOPLANCTON T0
B FITOPLANCTON T0
C Rapp. diluiz. 1:0 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Cell L-1 �gC L-1
Cryptophyceae Forme indeterminate 1360 0,018 Forme indeterminate 4600 0,060 Forme indeterminate 14600 0,191
Fitoflagellati indeterminati > 10 µm Fitoflagellati indeterminati 630 0,057 Fitoflagellati indeterminati 590 0,053 Fitoflagellati indeterminati 610 0,055
ANTARTIDE ESPERIMENTO 2

FITOPLANCTON T0 A
FITOPLANCTON T0 B
FITOPLANCTON T0 C
Rapp. Diluiz. 1:1 Cell L-1 �gCL-1 Cell L-1 µgrC L-1 Cell L-1 µgC L-1 Bacillariophyceae
Actinocyclus actinochilus 20 0,025 Actinocyclus actinochilus 30 0,037 Actinocyclus actinochilus 30 0,037 Chaetoceros dichaeta 20 0,005 Chaetoceros sp. 130 0,018 Chaetoceros sp. 130 0,018 Chaetoceros sp. 20 0,003 Corethron criophylum 40 0,248 Corethron criophylum 40 0,248 Cylindrotheca closterium 650 0,010 Cylindrotheca closterium 1770 0,027 Cylindrotheca closterium 1770 0,027 Fragilariopsis cylindrus 490 0,093 Fragilariopsis cylindrus 180 0,034 Fragilariopsis cylindrus 180 0,034 Fragilariopsis kerguelensis 460 0,063 Fragilariopsis kerguelensis 40 0,005 Fragilariopsis kerguelensis 40 0,005 Fragilariopsis sp. 90 0,025 Fragilariopsis sp. 100 0,027 Fragilariopsis sp. 100 0,027 Nitschia sp. 40 0,170 Navicula sp. 20 0,033 Navicula sp. 20 0,033 Pseudo-nitzschia cf. pseudodelicatissima 90 0,002
Pseudo-nitzschia cf. pseudodelicatissima 90 0,002
Pseudo-nitzschia cf. pseudodelicatissima 90 0,002
Pseudo-nitzschia cf seriata 230 0,034 Pseudo-nitzschia cf seriata 170 0,025 Pseudo-nitzschia cf seriata 170 0,025 Pseudo-nitzschia cf subcurvata 10 0,008 Pseudo-nitzschia sp. 80 0,002 Pseudo-nitzschia sp. 80 0,002 Pseudo-nitzschia sp. 70 0,002 Stellarima microtrias 20 0,031 Stellarima microtrias 20 0,031 Stellarima microtrias 10 0,016 Thalassiosira sp. 80 0,027 Thalassiosira sp. 80 0,027
Thalassiosira sp. 20 0,007 Forme centriche indeterminate > 10 µm 30 0,059
Forme centriche indeterminate > 10 µm 30 0,059
Forme centriche indeterminate > 10 µm 20 0,039
Dinophyceae Amphidinium sp. 10 - 50 µm 30 0,006 Gymnodinium sp. 10 - 50 µm 1030 0,882 Gymnodinium cf. baccatum 10 0,005 Gymnodinium sp. 10 - 50 µm 80 0,069 Gyrodinium sp. 10 - 50 µm 2290 0,495 Gymnodinium cf. diploconus 10 0,254 Gymnodinium sp. > 50 µm 130 0,975 Gyrodinium sp. > 50 µm 30 0,184 Gymnodinium sp. 10 - 50 µm 420 0,360 Gyrodinium sp. 10 - 50 µm 130 0,028 Forme indeterminate > 10 µm 400 0,240 Gymnodinium sp. > 50 µm 10 0,075 Gyrodinium sp. > 50 µm 60 0,368 Gyrodinium sp. 10 - 50 µm 3540 0,765 Forme indeterminate > 10 µm 590 0,355 Prorocentrum cf. antarcticum 10 0,038
Cryptophyceae Forme indeterminate 590 0,008 Forme indeterminate 4420 0.058 Forme indeterminate 4440 0,058
ANTARTIDE ESPERIMENTO 2

FITOPLANCTON T0 A
FITOPLANCTON T0 B
FITOPLANCTON T0 C
Rapp. diluiz.1:3 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Bacillariophyceae
Actinocyclus actinochilus 10 0,012 Actinocyclus actinochilus 10 0,012 Actinocyclus actinochilus 20 0,025 Chaetoceros dichaeta 10 0,002 Asteromphalus hookery 10 0,025 Corethron criophylum 20 0,124 Chaetoceros sp. 60 0,008 Chaetoceros sp. 80 0,011 Corethron inerme 20 0,012 Corethron criophilum 110 0,681 Corethron criophylum 60 0,371 Cylindrotheca closterium 1030 0,016 Cylindrotheca closterium 910 0,014 Cylindrotheca closterium 1100 0,017 Fragilariopsis cylindrus 3410 0,644 Fragilariopsis cf. curta 30 0,002 Fragilariopsis cf. curta 120 0,007 Fragilariopsis kerguelensis 760 0,104 Fragilariopsis cylindrus 560 0,106 Fragilariopsis cylindrus 510 0,096 Fragilariopsis sp. 950 0,259 Fragilariopsis kerguelensis 360 0,049 Fragilariopsis kerguelensis 300 0,041 Navicula sp. 240 0,396 Fragilariopsis sp. 500 0,136 Fragilariopsis sp. 200 0,055 Nitzschia sp. 30 0,128
Navicula sp. 80 0,132 Navicula sp. 170 0,281 Pseudo-nitzschia cf. pseudodelicatissima 20 0,000
Nitschia sp. 60 0,256 Nitzschia sp. 50 0,213 Pseudo-nitzschia cf seriata 320 0,047 Pseudo-nitzschia cf. pseudodelicatissima 50 0,001
Pseudo-nitzschia cf. pseudodelicatissima 70 0,001 Pseudo-nitzschia sp. 80 0,002
Pseudo-nitzschia cf seriata 200 0,029 Pseudo-nitzschia cf seriata 150 0,022 Thalassiosira sp. 150 0,051
Pseudo-nitzschia sp. 10 0,000 Pseudo-nitzschia sp. 200 0,004 Forme centriche indeterminate > 10 µm 30 0,059
Stellarima microtrias 20 0,031 Thalassiosira sp. 170 0,058 Forme pennate indet. > 10 µm 110 0,017 Thalassiosira sp. 80 0,027 Forme centriche indet. > 10 µm 70 0,138
Dinophyceae Amphidinium sp. 10 - 50 µm 20 0,004 Gymnodinium sp. 10 - 50 µm 800 0,685 Amphidinium sp. 10 - 50 µm 10 0,002 Gymnodinium cf. guttula 10 0,014 Gyrodinium sp. 10 - 50 µm 1430 0,309 Gymnodinium sp. 10 - 50 µm 220 0,188 Gymnodinium sp. 10 - 50 µm 230 0,197 Forme indeterminate > 10 µm 270 0,162 Gymnodinium sp. > 50 µm 30 0,225 Gyrodinium sp. 10 - 50 µm 280 0,060 Gyrodinium sp. 10 - 50 µm 1310 0,283 Gyrodinium sp. > 50 µm 30 0,184 Gyrodinium sp. > 50 µm 20 0,123 Prorocentrum cf. antarcticum 20 0,077 Prorocentrum cf. antarcticum 20 0,077
Forme indeterminate > 10 µm Forme indeterminate 870 0,011 Forme indeterminate 4380 0,057 Forme indeterminate 3940 0,052
ANTARTIDE ESPERIMENTO 2

FITOPLANCTON T0 A
FITOPLANCTON T0 B
FITOPLANCTON T0 C
Rapp. diluiz. 3:1 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Cell L-1 �gC L-1 Bacillariophyceae
Chaetoceros sp. 100 0,014 Actinocyclus actinochilus 50 0,062 Actinocyclus actinochilus 40 0,050 Coscinodiscus oculus-iridis 10 0,154 Chaetoceros sp. 20 0,003 Chaetoceros sp. 420 0,059 Coscinodiscus sp. 30 0,006 Corethron criophilum 30 0,186 Corethron criophylum 100 0,619 Cylindrotheca closterium 1250 0,019 Cylindrotheca closterium 440 0,007 Cylindrotheca closterium 600 0,009 Fragilariopsis cylindrus 320 0,060 Eucampia cf. antarctica 20 0,025 Fragilariopsis cylindrus 40 0,008 Fragilariopsis kerguelensis 340 0,047 Fragilariopsis cylindrus 1060 0,200 Fragilariopsis kerguelensis 180 0,025 Fragilariopsis sp. 350 0,095 Fragilariopsis kerguelensis 1460 0,200 Navicula sp. 20 0,033
Nitschia sp. 30 0,128 Fragilariopsis sp. 460 0,125 Pseudo-nitzschia cf. pseudodelicatissima 20 0,000
Pseudo-nitzschia cf. pseudodelicatissima 110 0,002 Navicula sp. 80 0,132 Pseudo-nitzschia cf seriata 200 0,029 Pseudo-nitzschia cf seriata 200 0,029 Nitschia sp. 40 0,170 Pseudo-nitzschia sp. 30 0,001 Pseudo-nitzschia cf subcurvata 10 0,008 Pleurosigma sp. 10 0,018 Thalassiosira sp. 40 0,014
Pseudo-nitzschia sp. 110 0,002 Pseudo-nitzschia cf. pseudodelicatissima 40 0,001
Forme centriche indeterminate > 10 µm 70 0,138
Stellarima microtrias 10 0,016 Pseudo-nitzschia cf seriata 210 0,031 Forme pennate indet.> 10 µm 180 0,027 Thalassiosira sp. 180 0,062 Pseudo-nitzschia sp. 80 0,002
110 0,216 Thalassiosira sp. 90 0,031 Forme centriche indet. > 10 µm Forme pennate indeterminate 300 0,046
Dinophyceae Amphidinium sp. 10 - 50 µm 250 0,053 Amphidinium sp. 10 - 50 µm 50 0,011 Gymnodinium cf. diploconus 20 0,508 Gymnodinium sp. 10 - 50 µm 600 0,514 Gymnodinium sp. 10 - 50 µm 930 0,797 Gymnodinium sp. 10 - 50 µm 1560 1,336 Gyrodinium sp. 10 - 50 µm 1390 0,300 Gymnodinium sp. > 50 µm 40 0,300 Gymnodinium sp. > 50 µm 30 0,225 Prorocentrum cf. antarcticum 60 0,230 Gyrodinium cf. modestum 30 0,019 Gyrodinium sp. 10 - 50 µm 4140 0,894 Forme indeterminate > 10 µm 1550 0,932 Gyrodinium sp. 10 - 50 µm 199 0,430 Gyrodinium sp. > 50 µm 10 0,061 Gyrodinium sp. > 50 µm 40 0,245 Prorocentrum cf. antarcticum 10 0,038
Cryptophyceae Forme indeterminate 1430 0,019 Forme indeterminate 5760 0,075 Forme indeterminate 5660 0,074
ANTARTIDE ESPERIMENTO 2

100% A 100%B 100%C 90%A 80%A 50%A 10%A 100%A 100%B 100%C 90%A 80%A 50%A 10%A Microzooplancton T0
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Dinoflagellati Gyrodinium lachrima 400 464 392 378 286 202 46 2.656 3.081 2.603 2.510 1.899 1.341 0.305 Protoperidinium antarcticum 4 2 2 0.244 0.122 0.122 Protoperidinium applanatum 34 34 12 20 18 12 4 0.146 0.146 0.051 0.086 0.077 0.051 0.017 Protoperidinium archiovatum 6 28 16 18 16 6 4 0.015 0.072 0.041 0.046 0.041 0.015 0.010 Protoperidinium defectum 12 14 6 8 16 10 4 0.009 0,011 0.005 0.006 0.012 0.008 0.003 Protoperidinium incertum 4 2 4 8 4 2 0.017 0.009 0.017 0.035 0.017 0.009 Protoperidinium mediocre 14 12 12 8 10 4 2 0.099 0.085 0.085 0.057 0.071 0.028 0.014 Protoperidinium melo 6 6 4 2 0.018 0.018 0.012 0.006 P. psudoantarcticum 8 8 16 8 2 2 0.121 0.121 0.242 0.121 0.030 0.030 Protoperidinium raphanum 26 18 34 34 22 8 4 0.124 0.086 0.162 0.162 0.105 0.038 0.019 Protoperidinium sp.1 Protoperidinium sp.2 Diplopsalis spp. 258 260 188 254 234 152 28 0.364 0.367 0.265 0.359 0.330 0.215 0.040 Strobilididi Strobilididae indet.1 34 22 20 30 40 22 8 0.067 0.044 0.040 0.059 0.079 0.044 0.016 Strobilididae indet.2 114 74 110 42 48 26 4 1.044 0.678 1.007 0.385 0.440 0.238 0.037 Strobilididae indet.3 36 36 30 36 22 6 0.905 0.905 0.754 0.905 0.553 0.151 Strobilididae indet.4 20 24 10 2 1.256 1.508 0.628 0.126 Strobilididae indet.5 10 10 6 1.431 1.431 0.859 Strombididi Strombidiidae indet.3 14 32 48 68 30 4 0.078 0.179 0.268 0.380 0.168 0.022 Strombidiidae indet.4 Strombidiidae indet.5 14 6 14 12 8 4 0.445 0.191 0.445 0.382 0.254 0.127 Strombidiidae indet.6 14 0.934 Tintinnidi Cymatocylis drigalskii 12 4 12 16 6 0.263 0.088 0.263 0.351 0.131 Laackmaniella naviculifera 6 12 4 0.055 0.110 0.037 Laackmaniella prolongata 20% 4 10 6 0.335 0.067 0.168 0.101 Metazoi Nauplius indet. 1 6 6 0.018 0.018 Nauplius indet. 2 14 14 18 8 14 4 0.268 0.268 0.345 0.153 0.268 0.077 Larvae indet. 14 14 2 6 0.193 0.193 0.028 0.083
ANTARTIDE ESPERIMENTO 3

100% A 100% B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T48
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 Dinoflagellati Gyrodinium lachrima 496 600 640 524 552 544 364 352 640 232 232 272 32 16 28 Protoperidinium antarcticum 4 32 4 4 Protoperidinium applanatum 20 12 40 16 28 48 20 28 32 16 8 24 8 Protoperidinium archiovatum 32 16 8 24 24 12 20 28 4 12 8 4 Protoperidinium defectum 32 88 40 44 64 48 24 48 52 8 20 16 4 4 Protoperidinium incertum 16 44 32 24 20 12 8 12 16 4 4 Protoperidinium mediocre 12 16 8 8 4 12 16 20 24 16 4 8 4 Protoperidinium melo 12 12 16 8 8 8 16 4 P. psudoantarcticum 12 12 8 12 8 4 12 12 8 8 4 Protoperidinium raphanum 20 56 24 32 28 24 28 16 24 12 4 16 4 Protoperidinium sp.1 4 4 8 16 8 8 Protoperidinium sp.2 8 8 4 8 16 4 12 12 4 4 Diplopsalis spp. 340 468 480 448 420 492 312 284 540 152 192 212 40 24 16 Strobilididi Strobilididae indet.1 72 84 80 68 72 32 20 28 40 24 12 24 4 Strobilididae indet.2 84 84 32 60 56 32 20 28 40 20 8 24 4 4 Strobilididae indet.3 20 76 32 64 32 60 36 40 32 24 16 24 4 Strobilididae indet.4 16 28 16 32 20 16 20 28 24 12 8 12 4 Strobilididae indet.5 16 8 16 4 4 12 4 16 Strombididi Strombidiidae indet.3 96 68 64 88 132 56 36 56 98 24 16 44 8 4 8 Strombidiidae indet.4 8 20 16 12 8 12 8 8 24 4 8 4 Strombidiidae indet.5 16 8 12 12 8 12 12 16 4 12 4 Strombidiidae indet.6 4 Tintinnidi Codonellopsis gaussi 8 8 4 4 Cymatocylis drigalskii 16 16 12 4 16 4 4 12 12 8 4 Laackmaniella naviculifera 4 4 20 4 4 4 8 4 8 4 Laackmaniella prolongata 8 8 8 4 4 4 4 4 12 Metazoi Nauplius indet. 1 4 8 4 12 4 Nauplius indet. 2 8 12 8 24 20 16 20 12 16 12 4 4 Larvae indet. 12 8 12 12 16 16 4 4 4 4 4
ANTARTIDE ESPERIMENTO 3

100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T48
µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Dinoflagellati
Gyrodinium lachrima 3.293 3.984 4.250 3.479 3.665 3.612 2.417 2.337 4.250 1.540 1.540 1.806 0.212 0.106 0.186
Protoperidinium antarcticum 0.244 1.950 0.244 0.244
Protoperidinium applanatum 0.086 0.051 0.171 0.069 0.120 0.206 0.086 0.120 0.137 0.069 0.034 0.103 0.034
Protoperidinium archiovatum 0.083 0.041 0.021 0.062 0.062 0.031 0.052 0.072 0.010 0.031 0.021 0.010
Protoperidinium defectum 0.024 0.067 0.031 0.034 0.049 0.037 0.018 0.037 0.040 0.006 0.015 0.012 0.003 0.003
Protoperidinium incertum 0.070 0.191 0.139 0.104 0.087 0.052 0.035 0.052 0.070 0.017 0.017
Protoperidinium mediocre 0.085 0.113 0.057 0.057 0.028 0.085 0.113 0.141 0.170 0.113 0.028 0.057 0.028
Protoperidinium melo 0.035 0.035 0.047 0.023 0.023 0.023 0.047 0.012
P. psudoantarcticum 0.181 0.181 0.121 0.181 0.121 0.060 0.181 0.181 0.121 0.121 0.060
Protoperidinium raphanum 0.095 0.266 0.114 0.152 0.133 0.114 0.133 0.076 0.114 0.057 0.019 0.076 0.019
Protoperidinium sp.1 0.007 0.007 0.015 0.029 0.015 0.015
Protoperidinium sp.2 0.068 0.068 0.034 0.068 0.136 0.034 0.102 0.120 0.034 0.034
Diplopsalis spp. 0.480 0.661 0.678 0.633 0.593 0.695 0.441 0.401 0.762 0.215 0.271 0.299 0.056 0.034 0.023
Strobilididi
Strobilididae indet.1 0.142 0.168 0.158 0.135 0.142 0.063 0.040 0.055 0.079 0.047 0.024 0.047 0.008
Strobilididae indet.2 0.769 0.769 0.293 0.549 0.513 0.293 0.183 0.256 0.366 0.183 0.073 0.220 0.037 0.037
Strobilididae indet.3 0.503 1.910 0.804 1.608 0.804 1.508 0.905 1.005 0.804 0.603 0.402 0.603 0.101
Strobilididae indet.4 1.005 1.759 1.005 2.010 1.256 1.005 1.256 1.759 1.508 0.754 0.503 0.754 0.251
Strobilididae indet.5 2.290 1.145 2.290 0.572 0.572 1.717 0.572 2.290
Strombididi
Strombidiidae indet.3 0.536 0.380 0.357 0.491 0.737 0.313 0.201 0.313 0.547 0.134 0.089 0.246 0.045 0.022 0.045
Strombidiidae indet.4 0.112 0.279 0.223 0.168 0.112 0.168 0.112 0.112 0.335 0.056 0.112 0.056
Strombidiidae indet.5 0.509 0.254 0.382 0.382 0.254 0.382 0.382 0.509 0.127 0.382 0.127
Strombidiidae indet.6 0.267
Tintinnidi
Codonellopsis gaussi 0.091 0.091 0.046 0.046
Cymatocylis drigalskii 0.351 0.351 0.263 0.088 0.351 0.088 0.088 0.263 0.263 0.175 0.088
Laackmaniella naviculifera 0.037 0.037 0.183 0.037 0.037 0.037 0.073 0.037 0.073 0.037
Laackmaniella prolongata 0.134 0.134 0.134 0.067 0.067 0.067 0.067 0.067 0.201
Metazoi
Nauplius indet. 1 0.012 0.024 0.012 0.036 0.012
Nauplius indet. 2 0.153 0.230 0.153 0.459 0.383 0.306 0.383 0.230 0.306 0.230 0.077 0.077
Larvae indet. 0.165 0.110 0.165 0.165 0.220 0.220 0.055 0.055 0.055 0.055 0.055
ANTARTIDE ESPERIMENTO 3

100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C Fitoplancton T0
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Diatomee
Fragilariopsis curta 1.97E+06 1.90E+06 2.15E+06 1.80E+06 1.80E+06 1.10E+06 1.89E+06 1.70E+06 1.76E+06 1.01E+06 1.09E+06 1.02E+06 1.13E+05 1.01E+05 1.07E+05
Fragilariopsis cylindrus 2.69E+06 2.71E+06 2.92E+06 2.69E+06 2.68E+06 1.61E+06 2.73E+06 2.45E+06 2.40E+06 1.69E+06 1.51E+06 1.59E+06 1.63E+05 1.72E+05 1.77E+05
Pseudonitzchia subcurvata 8.86E+05 9.28E+05 9.54E+05 8.00E+05 8.92E+05 7.06E+05 6.94E+05 8.42E+05 7.66E+05 4.92E+05 5.02E+05 5.32E+05 3.38E+04 7.30E+04 8.80E+04
Pseudonitzchia hemii 6.60E+03 1.01E+04 9.44E+03 6.32E+03 6.40E+03 5.88E+03 6.68E+03 1.04E+04 7.36E+03 7.08E+03 6.60E+03 6.08E+03 9.80E+02 6.20E+02 8.00E+02
P. pseudodelicatissima 8.20E+03 9.08E+03 1.02E+04 5.44E+03 5.44E+03 5.88E+03 7.04E+03 1.22E+04 8.20E+03 7.58E+03 8.24E+03 7.80E+03 5.80E+02 6.80E+02 1.02E+03
Nanoflagellati <10µm 1.43E+05 1.38E+05 1.42E+05 1.02E+05 8.84E+04 1.28E+05 1.11E+05 9.00E+04 9.44E+04 8.29E+04 6.20E+04 7.63E+04 1.62E+04 1.43E+04 1.69E+04
Primnesioficee
Phaeocystis antarctica 1.13E+05 1.02E+05 1.03E+05 9.63E+04 6.63E+04 1.02E+05 7.91E+04 7.60E+04 8.29E+04 5.15E+04 5.40E+04 5.92E+04 9.80E+03 1.02E+04 1.10E+04
Diatomee µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Fragilariopsis curta 121.300 116.980 132.780 111.060 110.810 67.990 116.860 105.010 108.470 62.070 67.130 63.180 7.002 6.232 6.620
Fragilariopsis cylindrus 65.270 65.850 70.910 65.420 65.030 39.030 66.390 59.490 58.470 41.020 36.640 38.540 3.969 4.172 4.296
Pseudonitzchia subcurvata 8.680 9.090 9.350 7.840 8.740 6.920 6.800 8.250 7.510 4.820 4.920 5.210 0.331 0.715 0.863
Pseudonitzchia hemii 1.170 1.790 1.680 1.120 1.140 1.050 1.190 1.860 1.310 1.260 1.170 1.080 0.174 0.110 0.142
P. pseudodelicatissima 0.120 0.140 0.150 0.080 0.080 0.090 0.110 0.180 0.120 0.110 0.120 0.120 0.009 0.010 0.015
Nanoflagellati <10µm 1.770 1.710 1.760 1.270 1.100 1.590 1.370 1.120 1.170 1.030 0.770 0.950 0.200 0.178 0.209
Primnesioficee
Phaeocystis antarctica 1.400 1.260 1.280 1.190 0.820 1.260 0.980 0.940 1.030 0.640 0.670 0.730 0.111 0.126 0.136
ANTARTIDE ESPERIMENTO 3

ANTARTIDE ESPERIMENTO 4
Microzooplancton 100% A 90%A 80%A 50%A 10%A 100%A 90%A 80%A 50%A 10%A T0 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Dinoflagellati Gyrodinium lachrima 130 80 60 32 10 0.656 0.403 0.302 0.161 0.050 Diplopsalis 20 14 4 2 2 0.028 0.020 0.006 0.003 0.003 Protoperidinium applanatum 30 10 4 4 4 0.067 0.022 0.009 0.009 0.009 Protoperidinium defectum 12 8 4 4 0 0.009 0.006 0.003 0.003 Tintinnid i Cymatocyclis drigalskii 32 24 16 14 2 0.733 0.549 0.366 0.320 0.046 Strobilididi Strobilididae indet. 2 26 20 12 4 2 0.238 0.183 0.109 0.036 0.018 Strobilididae indet. 3 22 16 12 8 2 0.552 0.402 0.301 0.201 0.050 Strombididi Strombididae indet. 3 42 40 36 24 2 0.233 0.222 0.200 0.133 0.011 Strombididae indet. 6 54 48 36 12 4 3.301 3.201 2.400 0.800 0.266

100% A 100%B !00%C 90%A 90%B 90%C 80%A 80%B 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T24 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Dinoflagellati Gyrodinium lachrima 240 320 268 96 104 100 48 40 28 20 32 4 4 8 Diplopsalis 28 0 16 0 4 24 8 0 8 12 20 4 0 4 Protoperidinium applanatum 0 28 20 56 40 32 16 20 12 16 12 4 0 0 Protoperidinium defectum 0 16 16 4 0 8 0 0 0 0 4 0 0 0 Tintinnid i Cymatocyclis drigalskii 44 40 48 24 28 32 20 20 16 12 16 0 4 0 Strobilididi Strobilididae A 2 60 44 52 16 20 28 12 16 12 16 8 0 4 4 Strobilididae A 3 84 60 44 20 28 24 16 20 12 16 16 0 4 0 Strombididi Strombididae B 3 60 72 64 56 40 44 32 20 24 20 16 4 4 0 Strombididae B 6 160 200 136 92 104 116 80 84 20 40 44 0 4 8
100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T24 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Dinoflagellati Gyrodinium lachrima 1.212 1.616 1.353 0.485 0.525 0.505 0.242 0.202 0.141 0.100 0.162 0.020 0.020 0.004 Diplopsalis 0.039 0 0.022 0 0.006 0.003 0.011 0 0.011 0.017 0.028 0.006 0 0.006 Protoperidinium applanatum 0 0.063 0.045 0.125 0.090 0.072 0.036 0.045 0.027 0.036 0.027 0.009 0 0 Protoperidinium defectum 0 0.012 0.012 0 0 0.006 0 0 0 0 0.003 0 0 0 Tintinnid i Cymatocyclis drigalskii 1.008 0.916 1.100 0.550 0.642 0.733 0.458 0.458 0.367 0.275 0.367 0 0.092 0 Strobilididi Strobilididae A 2 0.549 0.403 0.476 0.147 0.183 0.256 0.110 0.147 0.110 0.147 0.073 0 0.037 0.037 Strobilididae A 3 2.110 1.507 1.105 0.503 0.704 0.603 0.402 0.503 0.302 0.402 0.402 0 0.101 0 Strombididi Strombididae B 3 0.334 0.401 0.356 0.312 0.223 0.245 0.178 0.111 0.134 0.111 0.089 0.022 0.022 0 Strombididae B 6 10.670 13.338 9.070 6.135 6.936 7.736 5.335 5.602 1.334 2.668 2.934 0 0.267 0.534
ANTARTIDE ESPERIMENTO 4

100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%B 50 %A 50%B 50%C 10%A 10%B 10%C Fitoplancton T0
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L -1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cel l L-1 cell L-1 cell L-1 cell L-1 Bacillariophyceae Corethron criophilum 4.60E+02 4.00E+02 3.80E+02 3.00E+02 2.40E+02 2.20E+02 3.20E+02 2.00E+02 2.20E+02 8.00E+01 1.00E+02 2.00E+02 4.00E+01 6.00E+01 2.00E+01 Corethron inerme 8.00E+02 6.00E+02 7.00E+02 5.00E+02 8.80E+02 6.60E+02 5.60E+02 4.80E+02 5.20E+02 3.00E+02 5.60E+02 5.00E+02 4.00E+01 8.00E+01 4.00E+01 Fragilariopsis curta 7.00E+03 8.00E+03 9.00E+03 2.00E+03 3.08E+03 3.30E+03 4.60E+03 2.00E+03 8.00E+03 3.66E+03 4.60E+03 2.80E+03 1.10E+03 1.06E+03 1.04E+03 Fragilariopsis cylindrus 1.40E+03 1.20E+03 2.00E+03 1.28E+03 4.20E+02 8.00E+02 2.00E+03 1.40E+03 2.44E+03 7.20E+02 1.00E+03 1.40E+03 3.60E+02 4.00E+02 2.00E+02 Pseudonitzschia hemii 2.30E+03 1.30E+03 1.80E+03 6.00E+02 8.40E+02 9.00E+02 8.00E+02 1.06E+03 1.00E+03 5.00E+02 3.80E+02 4.00E+02 1.00E+02 1.20E+02 8.00E+01 Pseudonitzschia subcurvata 6.00E+05 7.00E+05 5.02E+05 3.40E+05 4.00E+05 3.52E+05 1.32E+05 1.80E+05 1.00E+05 1.20E+05 9.00E+04 7.00E+04 6.00E+04 4.00E+04 5.00E+04 Primnesioficee Phaeocystis antarctica 6.00E+05 5.00E+05 4.00E+05 5.00E+05 4.10E+05 3.60E+05 4.38E+05 3.60E+05 3.40E+05 1.50E+05 9.50E+04 1.00E+05 3.00E+04 2.00E+04 1.80E+04 Bacillariophyceae µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Corethron criophilum 0.963 0.837 0.795 0.628 0.502 0.460 0.670 0.418 0.460 0.167 0.209 0.418 0.080 0.125 0.041 Corethron inerme 5.072 3.804 4.438 3.170 5.579 4.184 3.550 3.043 3.296 1.902 3.550 3.170 0.253 0.507 0.253 Fragilariopsis curta 0.433 0.495 0.557 0.123 0.190 0.204 0.284 0.123 0.495 0.226 0.284 0.173 0.068 0.065 0.064 Fragilariopsis cylindrus 0.038 0.032 0.053 0.034 0.011 0.021 0.053 0.037 0.065 0.019 0.026 0.037 0.009 0.010 0.005 Pseudonitzschia hemii 0.126 0.072 0.099 0.033 0.046 0.049 0.044 0.058 0.055 0.027 0.020 0.022 0.006 0.006 0.004 Pseudonitzschia subcurvata 5.880 6.860 4.919 3.332 3.920 3.449 1.293 1.764 0.980 1.176 0.882 0.686 0.588 0.392 0.490 Primnesioficee Phaeocystis antarctica 5.748 4.790 3.832 4.790 3.920 3.448 4.196 3.448 3.257 1.437 0.910 0.958 0.287 0.191 0.172 Fitoplancton T24 Bacillariophyceae Corethron criophilum 6.E+02 6.E+02 5.E+02 4.E+02 3.E+02 3.E+02 2.E+02 2.E+02 2.E+02 2.E+02 1.E+02 3.E+02 2.E+01 4.E+01 2.E+01 Corethron inerme 8.E+02 6.E+02 9.E+02 8.E+02 9.E+02 7.E+02 4.E+02 5.E+02 4.E+02 3.E+02 3.E+02 3.E+02 1.E+02 1.E+02 2.E+02 Fragilariopsis curta 2.E+03 2.E+03 2.E+03 2.E+03 3.E+03 2.E+03 7.E+02 9.E+02 7.E+02 6.E+02 7.E+02 6.E+02 3.E+02 4.E+02 5.E+02 Fragilariopsis cylindrus 4.E+02 2.E+03 6.E+02 2.E+03 2.E+03 1.E+03 1.E+02 3.E+02 1.E+02 1.E+02 1.E+02 9.E+01 4.E+01 6.E+01 8.E+01 Pseudonitzschia hemii 6.E+02 4.E+02 5.E+02 6.E+02 8.E+02 5.E+02 4.E+02 5.E+02 4.E+02 2.E+02 3.E+02 3.E+02 1.E+02 1.E+02 2.E+02 Pseudonitzschia subcurvata 3.E+05 2.E+05 2.E+05 3.E+05 2.E+05 3.E+05 7.E+04 8.E+04 7.E+04 5.E+04 4.E+04 4.E+04 4.E+04 3.E+04 4.E+04 Primnesioficee Phaeocystis antarctica 4.E+05 4.E+05 3.E+05 2.E+05 3.E+05 2.E+05 2.E+05 2.E+05 2.E+05 1.E+05 1.E+05 1.E+05 2.E+04 1.E+04 1.E+04 Bacillariophyceae µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Corethron criophilum 1.256 1.298 1.005 0.837 0.682 0.670 0.418 0.335 0.418 0.460 0.293 0.586 0.042 0.083 0.041 Corethron inerme 5.325 3.930 5.706 4.945 5.579 4.311 2.536 2.916 2.536 1.902 1.775 1.775 0.887 0.760 1.014 Fragilariopsis curta 0.123 0.128 0.108 0.123 0.156 0.104 0.042 0.054 0.044 0.037 0.040 0.035 0.018 0.024 0.030 Fragilariopsis cylindrus 0.010 0.054 0.017 0.043 0.045 0.025 0.003 0.008 0.003 0.002 0.002 0.002 0.001 0.001 0.002 Pseudonitzschia hemii 0.033 0.024 0.028 0.030 0.041 0.028 0.024 0.028 0.022 0.011 0.014 0.015 0.006 0.007 0.008 Pseudonitzschia subcurvata 2.940 2.250 2.156 2.548 2.156 2.940 0.686 0.764 0.666 0.470 0.352 0.392 0.392 0.313 0.372

100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C
Fitoplancton T0
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 Fragilariopsis curta 42288 43440 49344 40464 39584 50400 48400 41408 44544 18704 12288 19120 2928 10032 3856 Fragilariopsis cylindrus 50432 56416 55536 38656 44144 57264 55296 49856 45712 18528 13520 19792 3072 13952 6816 Fragilariopsis kerguelensis 9424 14672 15424 5008 9344 12144 13376 8336 12128 3616 3104 5472 336 720 848 Pseudonitzschia subcurvata 275200 340800 332800 153600 241600 214400 280000 230400 176000 268800 156800 217600 83200 59200 137600 Pseudonitzschia hemii 1328 1056 1472 1392 976 720 992 928 1168 352 736 912 176 128 240 Corethron criophilum 13216 11120 15568 6256 8672 10944 5264 7648 6496 4752 4992 1712 368 896 992 Corethron inerme 4384 5632 3456 1424 2624 3488 1632 2448 3152 1936 2304 480 112 560 384 Navicula sp. 30928 30080 39008 26160 32976 42928 34080 29216 27104 11472 4288 7760 4752 10736 2384 Attheya sp. 14800 11696 19584 9904 14752 9024 13552 17152 13376 4128 3440 6528 656 2976 560 Dictyocha speculum 64 32 16 16 48 0 0 0 0 16 0 0 0 0 0 Diatomea centrica indet. 432 512 656 320 864 400 240 272 144 96 16 192 0 64 0 Diatomea pennata indet. 96 80 0 0 32 0 0 0 0 0 0 0 0 0 0 Prorocentrum sp. 400 320 544 464 416 208 288 176 384 160 352 128 64 112 80 Nanoflagellati < 10 µm indet. 13568 18496 12672 16832 10160 7984 5184 12192 6192 8256 3024 3600 3376 3680 3136 100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Fragilariopsis curta 2.088 2.145 2.436 1.998 1.954 2.488 2.390 2.044 2.199 0.923 0.607 0.944 0.145 0.495 0.190 Fragilariopsis cylindrus 2.076 2.323 2.286 1.591 1.817 2.358 2.277 2.053 1.882 0.763 0.557 0.815 0.126 0.574 0.281 Fragilariopsis kerguelensis 1.107 1.723 1.812 0.588 1.097 1.426 1.571 0.979 1.424 0.425 0.365 0.643 0.039 0.085 0.100 Pseudonitzschia subcurvata 1.998 2.474 2.416 1.115 1.754 1.557 2.033 1.673 1.278 1.951 1.138 1.580 0.604 0.430 0.999 Pseudonitzschia hemii 0.048 0.038 0.053 0.050 0.035 0.026 0.036 0.033 0.042 0.013 0.026 0.033 0.006 0.005 0.009 Corethron criophilum 27.680 23.290 32.607 13.103 18.163 22.922 11.025 16.018 13.606 9.953 10.456 3.586 0.771 1.877 2.078 Corethron inerme 27.797 35.710 21.913 9.029 16.638 22.116 10.348 15.552 19.986 12.275 14.609 3.043 0.710 3.551 2.435 Navicula sp. 0.035 0.034 0.044 0.030 0.038 0.049 0.039 0.033 0.031 0.013 0.005 0.009 0.005 0.012 0.003 Attheya sp. 0.601 0.475 0.795 0.402 0.599 0.366 0.550 0.696 0.534 0.168 0.140 0.265 0.027 0.121 0.023 Dictyocha speculum 0.049 0.024 0.012 0.012 0.036 0.000 0.000 0.000 0.000 0.012 0.000 0.000 0.000 0.000 0.000 Diatomea centrica indet. 0.089 0.105 0.135 0.066 0.178 0.082 0.049 0.056 0.030 0.020 0.003 0.039 0.000 0.013 0.000 Diatomea pennata indet. 0.082 0.068 0.000 0.000 0.027 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Prorocentrum sp. 0.184 0.147 0.250 0.213 0.191 0.096 0.132 0.081 0.176 0.073 0.162 0.059 0.029 0.051 0.037 Nanoflagellati < 10 µm indet. 0.130 0.177 0.121 0.161 1.097 0.076 0.050 0.117 0.059 0.079 0.029 0.034 0.032 0.035 0.030
ANTARTIDE ESPERIMENTO 5

100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C
Fitoplancton T38
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 Fragilariopsis curta 37552 38784 45056 37024 36144 45760 44000 39536 40848 19472 14096 20096 3280 10928 5712 Fragilariopsis cylindrus 48736 49184 48832 39888 40256 52112 51648 44160 42048 18336 10752 22000 4272 14032 6048 Fragilariopsis kerguelensis 8288 12768 12304 3968 6672 11056 14224 7664 12608 3744 4240 5712 288 816 896 Pseudonitzschia subcurvata 337600 379200 310400 147200 380800 225600 206400 260800 266400 211200 171200 182400 97600 107200 115200 Pseudonitzschia hemii 864 1280 1792 496 752 1152 1296 992 784 832 688 736 432 256 496 Corethron criophilum 15776 11536 14304 7440 10752 14112 6720 9712 8432 6480 6912 2464 576 1168 1568 Corethron inerme 4864 5984 3792 3168 1856 4416 2816 3248 3664 2544 3136 784 304 832 544 Navicula sp. 37424 34976 42224 31536 39488 45872 41968 32800 32608 15008 6768 12368 6080 16560 3504 Attheya sp. 38480 12672 30480 12224 16768 11568 24288 15040 22384 5488 6224 4752 2144 3216 1296 Dictyocha speculum 32 0 96 0 32 0 0 0 0 0 0 0 16 0 0 Diatomea centrica indet. 480 624 448 672 272 544 192 112 224 80 144 64 32 16 48 Diatomea pennata indet. 48 16 32 16 0 0 0 32 0 0 0 0 0 0 0 Prorocentrum sp. 288 512 432 240 384 352 400 336 144 256 288 464 128 144 48 Nanoflagellati < 10 µm indet. 22160 13776 16992 18992 18352 11440 7664 11152 12864 9392 6016 6272 6640 3360 5408 100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Fragilariopsis curta 1.854 1.915 2.224 1.828 1.784 2.259 2.172 1.952 2.017 0.961 0.696 0.992 0.162 0.54 0.282 Fragilariopsis cylindrus 2.006 2.025 2.01 1.642 1.657 2.145 2.126 1.818 1.731 0.755 0.443 0.906 0.176 0.578 0.249 Fragilariopsis kerguelensis 0.973 1.5 1.445 0.466 0.784 1.299 1.6471 0.9 1.481 0.44 0.498 0.671 0.034 0.096 0.105 Pseudonitzschia subcurvata 2.451 2.753 2.254 1.069 2.765 1.638 1.498 1.893 1.789 1.533 1.243 1.324 0.709 0.778 0.836 Pseudonitzschia hemii 0.031 0.046 0.064 0.018 0.027 0.041 0.046 0.036 0.028 0.03 0.025 0.026 0.015 0.009 0.018 Corethron criophilum 33.042 24.162 29.959 15.583 22.52 29.557 14.075 20.341 17.66 13.572 14.477 5.161 1.206 2.446 3.284 Corethron inerme 30.841 37.942 24.044 20.087 11.768 28 17.855 20.594 23.232 16.131 19.884 4.971 1.928 5.275 3.449 Navicula sp. 0.043 0.04 0.048 0.036 0.045 0.052 0.048 0.037 0.037 0.017 0.008 0.014 0.007 0.019 0.004 Attheya sp. 1.562 0.514 1.237 0.496 0.681 0.47 0.986 0.61 0.909 0.223 0.253 0.193 0.087 0.131 0.053 Dictyocha speculum 0.024 0 0.073 0 0.024 0 0 0 0 0 0 0 0.012 0 0 Diatomea centrica indet. 0.099 0.128 0.092 0.138 0.056 0.112 0.039 0.023 0.046 0.016 0.03 0.013 0.007 0.003 0.01 Diatomea pennata indet. 0.041 0.014 0.027 0.014 0 0 0 0.027 0 0 0 0 0 0 0 Prorocentrum sp. 0.132 0.235 0.198 0.11 0.176 0.162 0.184 0.154 0.066 0.118 0.132 0.213 0.059 0.066 0.022 Nanoflagellati < 10 µm indet. 0.212 0.132 0.163 0.182 0.176 0.11 0.073 0.107 0.123 0.09 0.058 0.06 0.064 0.032 0.052
ANTARTIDE ESPERIMENTO 5

100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T0 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Diplopsalis sp. 568 752 656 504 592 472 528 544 272 360 312 512 16 152 56 Gyrodinium lachryma 1088 896 776 848 992 584 680 608 872 448 416 392 128 40 120 Protoperidinium applanatum 56 80 120 72 40 96 56 88 48 72 24 64 16 0 48 Protoperidinium incertum 408 432 496 384 328 448 344 384 216 248 192 264 88 16 0 Ptotoperidinium pseudoantaecticum 384 344 448 296 336 432 240 288 336 184 168 136 0 56 24 Ciliophora indet. 48 112 72 144 160 16 112 40 56 96 0 48 104 0 40 Strombidiidae indet. 256 232 168 240 144 296 248 192 176 144 120 72 24 0 16 Cymatocylis drigalskii 304 336 376 264 272 184 216 152 328 168 128 112 40 80 0 Codonellopsis gaussi 152 128 112 96 72 128 64 96 48 152 56 40 0 32 8 Laackmanniella prolongata 200 176 216 112 168 240 152 208 192 24 80 144 72 0 0 Metazoi (nauplii) 64 128 96 72 80 48 112 40 64 8 56 24 48 0 8 100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Diplopsalis sp. 0.734 0.972 0.848 0.651 0.765 0.610 0.682 0.703 0.351 0.465 0.403 0.662 0.021 0.196 0.072 Gyrodinium lachryma 5.927 4.881 4.228 4.620 5.404 3.182 3.705 3.312 4.751 2.441 2.266 2.136 0.697 0.218 0.654 Protoperidinium applanatum 0.119 0.170 0.255 0.153 0.085 0.204 0.119 0.187 0.102 0.153 0.051 0.136 0.034 0.000 0.102 Protoperidinium incertum 2.109 2.233 2.564 1.985 1.696 2.316 1.778 1.985 1.117 1.282 0.993 1.365 0.455 0.083 0.000 Ptotoperidinium pseudoantarcticum 6.113 5.476 7.132 4.712 5.349 6.877 3.821 4.585 5.349 2.929 2.675 2.165 0.000 0.892 0.382 Ciliophora indet. 0.110 0.256 0.165 0.330 0.367 0.037 0.256 0.092 0.128 0.220 0.000 0.110 0.238 0.000 0.092 Strombidiidae indet. 1.809 1.640 1.187 1.696 1.018 2.092 1.753 1.357 1.244 1.018 0.848 0.509 0.170 0.000 0.113 Cymatocylis drigalskii 7.307 8.076 9.037 6.345 6.538 4.422 5.192 3.653 7.883 4.038 3.076 2.692 0.961 1.923 0.000 Codonellopsis gaussi 1.861 1.567 1.371 1.175 0.881 1.567 0.783 1.175 0.588 1.861 0.686 0.490 0.000 0.392 0.098 Laackmanniella prolongata 4.249 3.739 4.589 2.380 3.569 5.099 3.230 4.419 4.079 0.510 1.700 3.060 1.530 0.000 0.000 Metazoi (nauplii) 2.679 5.359 4.019 3.014 3.349 2.010 4.689 1.675 2.679 0.335 2.345 1.005 2.010 0.000 0.335
ANTARTIDE ESPERIMENTO 5
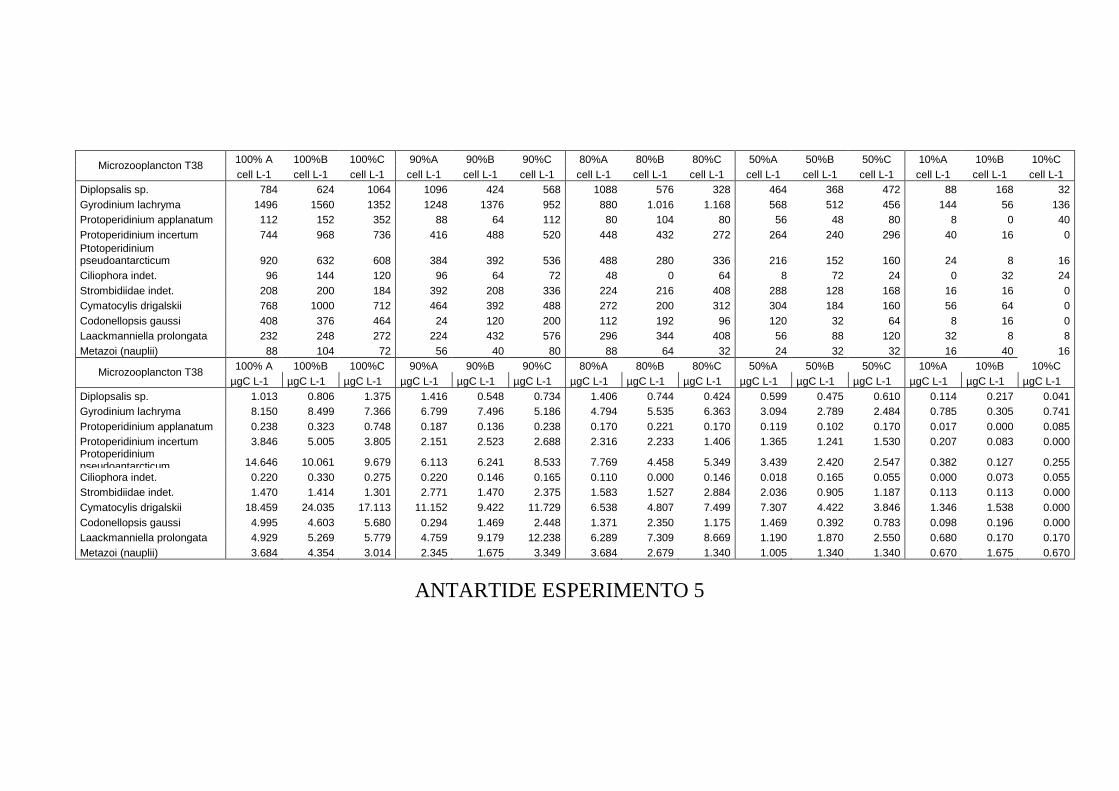
100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T38 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Diplopsalis sp. 784 624 1064 1096 424 568 1088 576 328 464 368 472 88 168 32 Gyrodinium lachryma 1496 1560 1352 1248 1376 952 880 1.016 1.168 568 512 456 144 56 136 Protoperidinium applanatum 112 152 352 88 64 112 80 104 80 56 48 80 8 0 40 Protoperidinium incertum 744 968 736 416 488 520 448 432 272 264 240 296 40 16 0 Ptotoperidinium pseudoantarcticum 920 632 608 384 392 536 488 280 336 216 152 160 24 8 16 Ciliophora indet. 96 144 120 96 64 72 48 0 64 8 72 24 0 32 24 Strombidiidae indet. 208 200 184 392 208 336 224 216 408 288 128 168 16 16 0 Cymatocylis drigalskii 768 1000 712 464 392 488 272 200 312 304 184 160 56 64 0 Codonellopsis gaussi 408 376 464 24 120 200 112 192 96 120 32 64 8 16 0 Laackmanniella prolongata 232 248 272 224 432 576 296 344 408 56 88 120 32 8 8 Metazoi (nauplii) 88 104 72 56 40 80 88 64 32 24 32 32 16 40 16
100% A 100%B 100%C 90%A 90%B 90%C 80%A 80%B 80%C 50%A 50%B 50%C 10%A 10%B 10%C Microzooplancton T38 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Diplopsalis sp. 1.013 0.806 1.375 1.416 0.548 0.734 1.406 0.744 0.424 0.599 0.475 0.610 0.114 0.217 0.041 Gyrodinium lachryma 8.150 8.499 7.366 6.799 7.496 5.186 4.794 5.535 6.363 3.094 2.789 2.484 0.785 0.305 0.741 Protoperidinium applanatum 0.238 0.323 0.748 0.187 0.136 0.238 0.170 0.221 0.170 0.119 0.102 0.170 0.017 0.000 0.085 Protoperidinium incertum 3.846 5.005 3.805 2.151 2.523 2.688 2.316 2.233 1.406 1.365 1.241 1.530 0.207 0.083 0.000 Protoperidinium pseudoantarcticum 14.646 10.061 9.679 6.113 6.241 8.533 7.769 4.458 5.349 3.439 2.420 2.547 0.382 0.127 0.255 Ciliophora indet. 0.220 0.330 0.275 0.220 0.146 0.165 0.110 0.000 0.146 0.018 0.165 0.055 0.000 0.073 0.055 Strombidiidae indet. 1.470 1.414 1.301 2.771 1.470 2.375 1.583 1.527 2.884 2.036 0.905 1.187 0.113 0.113 0.000 Cymatocylis drigalskii 18.459 24.035 17.113 11.152 9.422 11.729 6.538 4.807 7.499 7.307 4.422 3.846 1.346 1.538 0.000 Codonellopsis gaussi 4.995 4.603 5.680 0.294 1.469 2.448 1.371 2.350 1.175 1.469 0.392 0.783 0.098 0.196 0.000 Laackmanniella prolongata 4.929 5.269 5.779 4.759 9.179 12.238 6.289 7.309 8.669 1.190 1.870 2.550 0.680 0.170 0.170 Metazoi (nauplii) 3.684 4.354 3.014 2.345 1.675 3.349 3.684 2.679 1.340 1.005 1.340 1.340 0.670 1.675 0.670
ANTARTIDE ESPERIMENTO 5
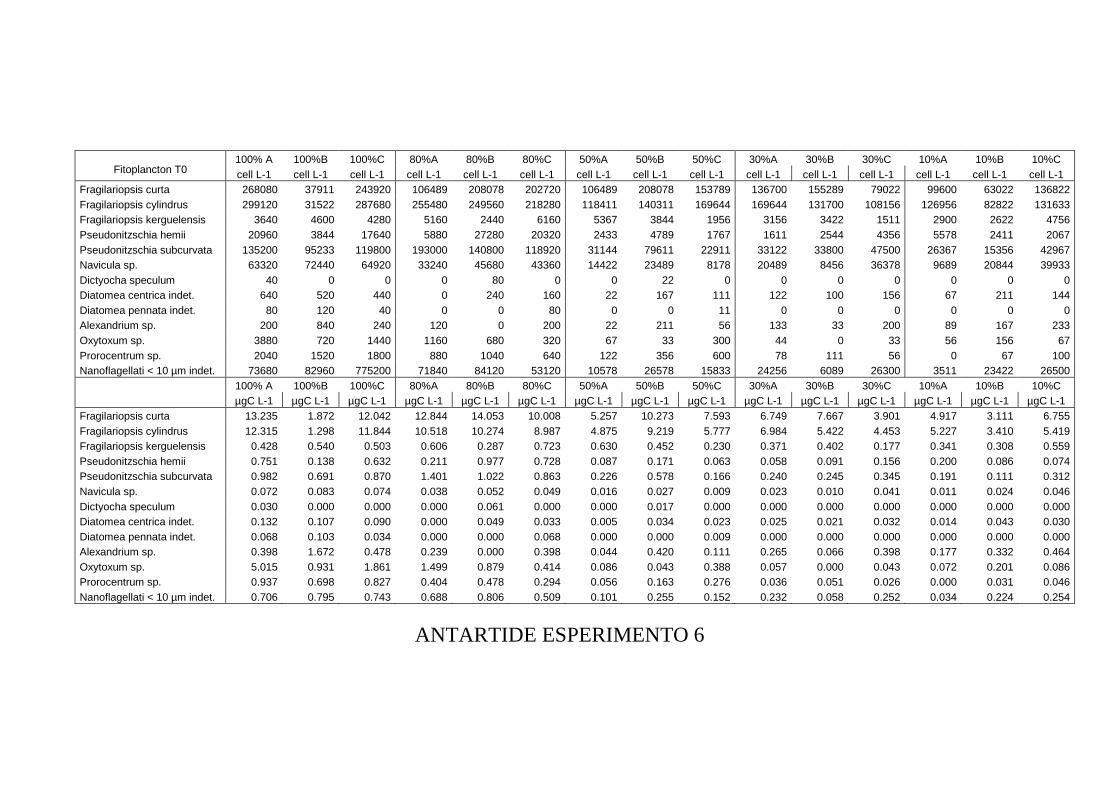
100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C
Fitoplancton T0
cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 Fragilariopsis curta 268080 37911 243920 106489 208078 202720 106489 208078 153789 136700 155289 79022 99600 63022 136822 Fragilariopsis cylindrus 299120 31522 287680 255480 249560 218280 118411 140311 169644 169644 131700 108156 126956 82822 131633 Fragilariopsis kerguelensis 3640 4600 4280 5160 2440 6160 5367 3844 1956 3156 3422 1511 2900 2622 4756 Pseudonitzschia hemii 20960 3844 17640 5880 27280 20320 2433 4789 1767 1611 2544 4356 5578 2411 2067 Pseudonitzschia subcurvata 135200 95233 119800 193000 140800 118920 31144 79611 22911 33122 33800 47500 26367 15356 42967 Navicula sp. 63320 72440 64920 33240 45680 43360 14422 23489 8178 20489 8456 36378 9689 20844 39933 Dictyocha speculum 40 0 0 0 80 0 0 22 0 0 0 0 0 0 0 Diatomea centrica indet. 640 520 440 0 240 160 22 167 111 122 100 156 67 211 144 Diatomea pennata indet. 80 120 40 0 0 80 0 0 11 0 0 0 0 0 0 Alexandrium sp. 200 840 240 120 0 200 22 211 56 133 33 200 89 167 233 Oxytoxum sp. 3880 720 1440 1160 680 320 67 33 300 44 0 33 56 156 67 Prorocentrum sp. 2040 1520 1800 880 1040 640 122 356 600 78 111 56 0 67 100 Nanoflagellati < 10 µm indet. 73680 82960 775200 71840 84120 53120 10578 26578 15833 24256 6089 26300 3511 23422 26500 100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Fragilariopsis curta 13.235 1.872 12.042 12.844 14.053 10.008 5.257 10.273 7.593 6.749 7.667 3.901 4.917 3.111 6.755 Fragilariopsis cylindrus 12.315 1.298 11.844 10.518 10.274 8.987 4.875 9.219 5.777 6.984 5.422 4.453 5.227 3.410 5.419 Fragilariopsis kerguelensis 0.428 0.540 0.503 0.606 0.287 0.723 0.630 0.452 0.230 0.371 0.402 0.177 0.341 0.308 0.559 Pseudonitzschia hemii 0.751 0.138 0.632 0.211 0.977 0.728 0.087 0.171 0.063 0.058 0.091 0.156 0.200 0.086 0.074 Pseudonitzschia subcurvata 0.982 0.691 0.870 1.401 1.022 0.863 0.226 0.578 0.166 0.240 0.245 0.345 0.191 0.111 0.312 Navicula sp. 0.072 0.083 0.074 0.038 0.052 0.049 0.016 0.027 0.009 0.023 0.010 0.041 0.011 0.024 0.046 Dictyocha speculum 0.030 0.000 0.000 0.000 0.061 0.000 0.000 0.017 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Diatomea centrica indet. 0.132 0.107 0.090 0.000 0.049 0.033 0.005 0.034 0.023 0.025 0.021 0.032 0.014 0.043 0.030 Diatomea pennata indet. 0.068 0.103 0.034 0.000 0.000 0.068 0.000 0.000 0.009 0.000 0.000 0.000 0.000 0.000 0.000 Alexandrium sp. 0.398 1.672 0.478 0.239 0.000 0.398 0.044 0.420 0.111 0.265 0.066 0.398 0.177 0.332 0.464 Oxytoxum sp. 5.015 0.931 1.861 1.499 0.879 0.414 0.086 0.043 0.388 0.057 0.000 0.043 0.072 0.201 0.086 Prorocentrum sp. 0.937 0.698 0.827 0.404 0.478 0.294 0.056 0.163 0.276 0.036 0.051 0.026 0.000 0.031 0.046 Nanoflagellati < 10 µm indet. 0.706 0.795 0.743 0.688 0.806 0.509 0.101 0.255 0.152 0.232 0.058 0.252 0.034 0.224 0.254
ANTARTIDE ESPERIMENTO 6

100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C Fitoplancton T36 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Fragilariopsis curta 365200 426800 347600 425360 303520 473880 143067 171389 144978 153567 178400 162156 54844 126578 140633 Fragilariopsis cylindrus 506000 532400 378400 396720 251000 485440 132822 165467 138711 138511 152533 176356 57978 120456 124956 Fragilariopsis kerguelensis 6440 1720 4960 17040 11840 15680 5033 9156 3678 2111 4189 1822 1289 6978 5367 Pseudonitzschia hemii 18640 12680 15280 5280 15360 11920 2300 4867 4144 2033 1311 2633 2767 1689 4744 Pseudonitzschia subcurvata 85600 183560 157280 69080 58240 135640 47711 45856 18956 43656 38733 30833 25122 30322 40933 Navicula sp. 128640 187680 193880 46560 41280 18160 15022 10178 6989 31933 35700 18133 20367 5356 10822 Dictyocha speculum 0 120 0 0 0 0 0 0 0 0 0 0 0 0 0 Diatomea centrica indet. 560 400 280 880 320 640 122 178 78 67 133 100 111 22 122 Diatomea pennata indet. 160 40 0 0 0 0 0 0 0 0 0 0 0 0 0 Alexandrium sp. 480 80 760 600 360 320 56 89 11 67 0 100 33 0 0 Oxytoxum sp. 1800 2360 2720 440 760 120 200 44 144 44 178 11 100 11 89 Prorocentrum sp. 2280 1760 2400 360 960 1440 222 300 133 100 33 22 0 33 22 Nanoflagellati < 10 µm indet. 132240 101520 127640 50920 24640 35080 8189 9367 6311 37356 31589 27233 49478 21867 36167 100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 Fragilariopsis curta 18.030 21.071 17.161 21.000 14.985 23.395 7.063 8.461 7.158 7.582 8.808 8.006 2.708 6.249 6.943 Fragilariopsis cylindrus 20.832 21.919 15.579 16.333 10.334 19.986 5.468 6.812 5.711 5.703 6.280 7.261 2.387 4.959 5.144 Fragilariopsis kerguelensis 0.756 0.202 0.583 2.001 1.391 1.842 0.591 1.075 0.432 0.248 0.492 0.214 0.151 0.82 0.630 Pseudonitzschia hemii 0.667 0.454 0.547 0.189 0.55 0.427 0.082 0.174 0.148 0.073 0.047 0.094 0.099 0.06 0.170 Pseudonitzschia subcurvata 0.621 1.333 1.142 0.502 0.423 0.985 0.346 0.333 0.138 0.317 0.281 0.224 0.182 0.22 0.297 Navicula sp. 0.147 0.214 0.221 0.053 0.047 0.021 0.017 0.012 0.008 0.036 0.041 0.021 0.023 0.006 0.012 Dictyocha speculum 0 0.091 0 0 0 0 0 0 0 0 0 0 0 0 0 Diatomea centrica indet. 0.115 0.082 0.058 0.181 0.066 0.132 0.025 0.037 0.016 0.014 0.027 0.021 0.023 0.005 0.025 Diatomea pennata indet. 0.137 0.034 0 0 0 0 0 0 0 0 0 0 0 0 0 Alexandrium sp. 0.955 0.159 1.512 1.194 0.716 0.637 0.111 0.177 0.022 0.133 0 0.199 0.066 0 0 Oxytoxum sp. 2.327 3.051 3.516 0.569 0.982 0.155 0.259 0.057 0.187 0.057 0.230 0.014 0.129 0.014 0.115 Prorocentrum sp. 1.047 0.808 1.102 0.165 0.441 0.661 0.102 0.138 0.061 0.046 0.015 0.010 0 0.015 0.010 Nanoflagellati < 10 µm indet. 1.267 0.973 1.223 0.488 0.236 0.336 0.078 0.09 0.06 0.358 0.303 0.261 0.474 0.209 0.346
ANTARTIDE ESPERIMENTO 6

100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C Microzooplancton T0 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Diplopsalis sp. 433 1267 744 244 644 344 256 378 167 78 22 33 56 89 122 Gyrodinium lachryma 1156 567 1011 1067 944 1.233 411 233 189 267 133 200 156 344 100 Protoperidinium sp. 800 2056 1056 956 1467 711 1433 622 922 656 389 800 1067 544 567 Protoperidinium defectum 3756 7367 4378 4644 4167 5244 2867 4122 2156 2078 3656 2156 2533 1922 956 Ciliophora indet. 478 122 167 156 100 67 22 0 33 0 0 0 0 11 33 Oligotrichida indet. 211 144 133 56 78 0 0 11 0 0 0 0 0 0 0 Strombidiidae indet. 44 0 33 0 11 0 0 0 0 0 0 0 0 0 0 Laackmanniella prolongata 156 133 89 111 100 133 44 22 56 56 22 22 44 44 33 Metazoi (nauplii) 22 11 33 22 33 22 11 11 22 0 0 11 22 11 0
100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 90%A 90%B 90%C 10%A 10%B 10%C Microzooplancton T0 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Diplopsalis sp. 3.184 9.307 5.470 1.796 4.735 2.531 1.878 2.776 1.225 0.571 0.163 0.245 0.408 0.653 0.898 Gyrodinium lachryma 17.274 8.472 15.115 15.946 14.118 18.437 6.146 3.488 2.824 3.986 1.993 2.990 2.325 5.149 1.495 Protoperidinium sp. 6.803 17.480 8.976 8.126 12.473 6.047 12.189 5.291 7.843 5.575 3.307 6.803 9.071 4.630 4.819 Protoperidinium defectum 6.899 13.532 8.042 8.532 7.654 9.634 5.266 7.573 3.960 3.817 6.715 3.960 4.654 3.531 1.755 Ciliophora indet. 1.094 0.280 0.382 0.352 0.229 0.153 0.051 0.000 0.076 0.000 0.000 0.000 0.000 0.025 0.076 Oligotrichida indet. 15.467 10.580 9.766 4.066 5.700 0.000 0.000 0.814 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Strombidiidae indet. 0.703 0.000 0.527 0.000 0.176 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 Laackmanniella prolongata 3.304 2.832 1.889 2.361 2.125 2.832 0.944 0.472 1.180 1.180 0.472 0.472 0.944 0.944 0.708 Metazoi (nauplii) 1.627 0.813 2.440 1.627 2.440 1.627 0.814 0.814 1.628 0.000 0.000 0.814 1.628 0.814 0.000
ANTARTIDE ESPERIMENTO 6

100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C Microzooplancton T36 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Diplopsalis sp. 2044 2767 1911 944 411 156 156 122 289 67 100 44 56 11 44 Gyrodinium lachryma 1033 1689 1833 800 1011 1367 311 400 211 244 178 344 111 133 256 Protoperidinium sp. 2178 2522 1433 678 1156 967 433 367 467 411 457 622 844 411 989 Protoperidinium defectum 18411 13478 14889 8533 12811 4878 3633 5156 4833 2811 1956 3189 1644 2167 789 Ciliophora indet. 244 100 278 22 11 0 11 0 22 0 22 0 0 0 0 Oligotrichida indet. 56 0 44 11 0 0 0 0 0 0 0 0 0 0 0 Strombidiidae indet. 33 11 0 0 0 0 0 0 0 0 0 0 0 0 0 Laackmanniella prolongata 100 0 78 67 22 33 67 56 89 22 44 33 0 11 33 Metazoi (nauplii) 0 22 22 11 0 22 11 0 11 11 0 0 0 0 11
100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C Microzooplancton T36 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Diplopsalis sp. 15.021 20.329 14.042 6.939 3.021 1.143 1.143 0.898 2.123 0.490 0.735 0.327 0.408 0.082 0.327 Gyrodinium lachryma 15.447 25.247 27.406 11.959 15.115 20.430 4.651 5.980 3.156 3.654 2.658 5.149 1.661 1.993 3.820 Protoperidinium sp. 18.520 21.449 12.189 5.764 9.827 8.221 3.685 3.118 3.969 3.496 3.969 5.291 7.181 3.496 8.410 Protoperidinium defectum 33.821 24.759 27.351 15.676 23.534 8.960 6.674 9.471 8.879 5.164 3.592 5.858 3.021 3.980 1.449 Ciliophora indet. 0.560 0.229 0.636 0.051 0.025 0 0.025 0 0.051 0 0.051 0 0 0 0 Oligotrichida indet. 4.066 0 3.253 0.814 0 0 0 0 0 0 0 0 0 0 0 Strombidiidae indet. 0.527 0.176 0 0 0 0 0 0 0 0 0 0 0 0 0 Laackmanniella prolongata 2.125 0 1.653 1.416 0.472 0.708 1.416 1.180 1.889 0.472 0.944 0.708 0 0.236 0.708 Metazoi (nauplii) 0 1.627 1.627 0.814 0 1.628 0.814 0 0.184 0.814 0 0 0 0 0.814
ANTARTIDE ESPERIMENTO 6

100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C Fitoplancton T0
cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 Fragilariopsis cylindrus 61067 3733 33067 13867 5333 15867 9600 11200 9600 11333 18000 9067 11067 2267 2133 Pleurosigma sp. 10400 1067 3867 14800 1333 1200 2533 1600 21067 2533 3200 1467 267 800 4444 Thalassiosira hyalina 11733 1200 4267 1867 667 2800 1067 1467 2267 1733 533 333 Nitzschia delicatissima 11600 2133 1467 3333 1200 8533 3200 5067 2933 800 1867 1200 133 400 933 Leptocylindrus danicus 4667 1333 267 1333 533 400 533 667 400 267 400 Nanoflagellati < 10 µm indet. 3.97E+06 1.03E+07 1.03E+07 7.40E+06 7.67E+06 8.59E+06 5.82E+06 5.69E+06 6.08E+06 4.36E+06 4.76E+06 5.42E+06 3.31E+06 3.97E+06 3.04E+06 100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 Fragilariopsis cylindrus 0.636 0.583 0.682 0.736 0.084 0.138 0.161 0.061 0.146 0.161 0.061 0.146 0.084 0.061 0.015 Pleurosigma sp. 7.359 36.578 1.082 32.682 0.433 1.948 11.471 0.216 1.082 0.866 0.866 0.216 14.934 Thalassiosira hyalina 5.479 1.619 2.366 1.993 0.872 1.245 0.125 0.125 0.498 0.498 0.311 Nitzschia delicatissima 0.023 0.119 0.012 0.087 0.064 0.075 0.026 0.017 0.006 0.006 0.009 0.038 0.003 0.020 Leptocylindrus danicus 0.209 0.358 0.538 0.179 0.090 0.030 0.149 0.03 Nanoflagellati < 10 µm indet. 1.720 1.655 1.806 1.355 1.335 1.335 0.981 1.061 1.119 0.948 0.948 0.943 0.686 0.765 0.746
100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C Fitoplancton T36 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1 cell -1
Fragilariopsis cylindrus 11067 10133 11867 2 2 2 2800 1067 2533 2800 1067 2533 1467 1067 267 Pleurosigma sp. 4533 22533 667 20133 267 1200 7067 133 667 267 533 133 9200 Thalassiosira hyalina 5867 533 2533 2133 933 1333 133 133 533 Nitzschia delicatissima 1067 5467 533 4000 2933 3467 1200 800 2000 267 400 1733 133 933 Leptocylindrus danicus 933 1600 2400 800 400 667 133 133 Nanoflagellati < 10 µm indet. 9.92E+06 9.52E+06 1.02E+07 9.12E+06 8.99E+06 8.99E+06 6.61E+06 7.14E+06 7.54E+06 5.95E+06 5.55E+06 5.42E+06 4.63E+06 3.85E+06 5.02E+06 100% A 100%B 100%C 80%A 80%B 80%C 50%A 50%B 50%C 30%A 30%B 30%C 10%A 10%B 10%C µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 µgC L -1 Fragilariopsis cylindrus 3.511 0.215 1.901 0.797 0.307 0.912 0.552 0.644 0.552 0.652 1.035 0.521 0.636 0.13 0.123 Pleurosigma sp. 16.882 1.732 6.277 24.025 2.164 1.948 4.112 2.597 34.198 4.112 5.195 2.381 0.433 1.299 7.215 Thalassiosira hyalina 10.959 1.121 3.985 1.743 0.623 2.615 2.117 0.125 1.619 0.498 0.311 0.623 Nitzschia delicatissima 0.252 0.046 0.032 0.072 0.026 0.185 0.069 0.11 0.064 0.017 0.041 0.026 0.003 0.009 0.02 Leptocylindrus danicus 1.045 0.299 0.06 0.299 0.119 0.09 0.119 0.149 0.09 0.06 0.09 Nanoflagellati < 10 µm indet. 0.056 0.145 0.145 0.103 0.107 0.12 0.082 0.08 0.085 0.061 0.067 0.076 0.046 0.055 0.042
ANTARTIDE ESPERIMENTO 7

cell L-1 T0 T36
Microzooplancton A B C A B C
Gyrodinium lachryma 4266 6133 4533 1867 4667 4400
Laboea strobila 800 667 533 267 1067 533
Gyrodinium sp 8800 9687 9141 17333 16133 17200
Protoperidinium diabulum 409 682 341 205
Dinoflagellati
Prorocentrum sp.p 533 133 0 133 400 0%
ANTARTIDE ESPERIMENTO 7

100% A 100% B 100% C 80% A 80% B 80% C 50% A 50% B 50% C 30% A 30% B 30% C Fitoplancton T0 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Nitzschia longissima 4.50E+08 4.56E+08 3.49E+08 2.78E+08 2.01E+08 2.71E+08 1.57E+08 1.42E+08 1.63E+08 7.90E+08 4.06E+08 9.40E+08 Fragilariopsis cylindrus 1.88E+08 1.78E+08 1.82E+08 1.41E+08 1.14E+08 1.33E+08 9.96E+07 8.45E+07 1.01E+08 3.99E+07 3.93E+07 5.02E+07 Fragilariopsis curta 1.31E+07 1.10E+07 1.06E+07 8.95E+06 6.55E+06 8.08E+06 3.16E+06 4.91E+06 6.55E+06 2.73E+06 3.49E+06 4.04E+06 Fragilariopsis kerguelensis 1.09E+05 4.36E+05 4.36E+05 8.73E+05 1.42E+06 3.27E+05 1.20E+06 0.00E+00 0.00E+00 5.46E+05 0.00E+00 2.18E+05 Pleurosigma sp. 7.31E+06 5.35E+06 5.57E+06 9.06E+06 4.58E+06 4.15E+06 3.82E+06 3.06E+06 3.71E+06 2.95E+06 2.51E+06 2.40E+06 Leptocylindrus danicus 2.18E+05 4.36E+05 5.46E+05 3.27E+05 1.09E+05 2.18E+05 4.36E+05 2.18E+05 4.36E+05 1.09E+05 1.09E+05 2.18E+05 Navicula sp. 0.00E+00 1.09E+05 2.18E+05 0.00E+00 2.18E+05 3.27E+05 0.00E+00 1.09E+05 0.00E+00 1.09E+05 0.00E+00 1.09E+05 Pseudonitzscha heimii 2.18E+05 3.27E+05 2.18E+05 2.18E+05 1.09E+05 3.27E+05 1.09E+05 1.09E+05 2.18E+05 0.00E+00 2.18E+05 1.09E+05 Thalassiosira gravida 0.00E+00 0.00E+00 4.36E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 Thalassiosira antarctica 1.09E+05 0.00E+00 0.00E+00 1.09E+05 1.09E+05 3.27E+05 1.09E+05 0.00E+00 1.09E+05 0.00E+00 1.09E+05 0.00E+00 Chaetoceros sp. 0.00E+00 2.18E+05 0.00E+00 4.36E+05 4.36E+05 1.09E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 Primnesiophyceae Phaeocystis antarctica 5.46E+04 3.82E+05 1.09E+05 1.64E+05 5.46E+04 0.00E+00 0.00E+00 1.09E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00
100% A 100% B 100% C 80% A 80% B 80% C 50% A 50% B 50% C 30% A 30% B 30% C Fitoplancton T0 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Nitzschia longissima 8768.32 8878.8 6800.92 5409.29 3915.68 5275.44 3057.33 2764.13 3180.56 1538.23 790.36 1829.3 Fragilariopsis cylindrus 7758.7 6990.47 7511.61 5799.93 4699.25 5480.96 4101.73 3477.26 4137.68 1644.29 1617.33 2066.59 Fragilariopsis curta 646.49 544.13 522.58 441.77 323.24 398.67 156.23 242.43 323.24 134.68 172.4 199.33 Fragilariopsis kerguelensis 12.82 51.27 51.27 102.53 166.61 38.45 140.98 0 0 64.08 0 25.63 Pleurosigma sp. 182.78 133.68 139.13 226.43 114.58 103.67 95.48 76.39 92.75 73.66 62.75 60.02 Leptocylindrus danicus 13.2 26.4 33 19.8 6.6 13.2 26.4 13.2 26.4 6.6 6.6 13.2 Navicula sp. 0 0.12 0.25 0 0.25 0.37 0 0.12 0 0.12 0 0.12 Pseudonitzscha heimii 7.82 11.72 7.82 7.82 3.91 11.72 3.91 3.91 7.82 0 7.82 3.91 Thalassiosira gravida 0 0 692.93 0 0 0 0 0 0 0 0 0 Thalassiosira antarctica 101.92 0 0 101.92 101.92 305.76 101.92 0 101.92 0 101.92 0 Chaetoceros sp. 0 30.71 0 61.41 61.41 15.35 0 0 0 0 0 0 Primnesiophyceae Phaeocystis antarctica 33.46 234.23 66.92 100.38 33.46 0 0 66.92 0 0 0 0
ANTARTIDE ESPERIMENTO 8

100% A 100% B 100% C 80% A 80% B 80% C 50% A 50% B 50% C 30% A 30% B 30% C Fitoplancton T36 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1 cell L-1
Nitzschia longissima 3.73E+08 3.37E+08 3.89E+08 1.99E+08 1.72E+08 1.73E+08 9.97E+07 1.23E+08 7.91E+07 3.83E+07 4.06E+07 3.28E+07 Fragilariopsis cylindrus 1.41E+08 1.18E+08 1.61E+08 5.49E+07 1.01E+08 1.08E+08 5.66E+07 6.83E+07 6.53E+07 2.88E+07 2.97E+07 2.31E+07 Fragilariopsis curta 1.10E+07 9.71E+06 7.97E+06 5.57E+06 6.44E+06 6.33E+06 3.82E+06 3.06E+06 3.06E+06 2.07E+07 2.40E+06 1.64E+06 Fragilariopsis kerguelensis 2.62E+06 1.31E+06 1.86E+06 8.73E+05 5.46E+05 1.31E+06 7.64E+05 0.00E+00 5.46E+05 2.18E+05 1.09E+06 0.00E+00 Pleurosigma sp. 4.36E+06 4.80E+06 6.11E+06 4.58E+06 5.02E+06 3.82E+06 2.07E+06 2.07E+06 2.40E+06 1.42E+06 1.64E+06 1.53E+06 Leptocylindrus danicus 7.64E+05 7.64E+05 4.36E+05 3.27E+05 4.36E+05 6.55E+05 2.18E+05 2.18E+05 1.64E+05 1.09E+05 2.73E+05 0.00E+00 Navicula sp. 1.09E+05 9.82E+05 3.27E+05 1.09E+05 0.00E+00 3.27E+05 0.00E+00 3.27E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00 Pseudonitzscha heimii 0.00E+00 3.27E+05 1.64E+05 7.64E+05 1.09E+05 3.27E+05 0.00E+00 1.09E+05 1.64E+05 0.00E+00 5.46E+05 1.09E+05 Thalassiosira gravida 0.00E+00 1.09E+05 6.55E+05 0.00E+00 6.55E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 Thalassiosira antarctica 0.00E+00 3.27E+05 4.36E+05 2.18E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 Chaetoceros sp. 2.18E+05 0.00E+00 0.00E+00 3.27E+05 0.00E+00 2.18E+05 3.27E+05 1.09E+05 0.00E+00 0.00E+00 1.09E+05 2.18E+05 Primnesiophyceae Phaeocystis antarctica 0.00E+00 1.64E+05 1.64E+05 0.00E+00 1.64E+05 1.09E+05 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00 0.00E+00
100% A 100% B 100% C 80% A 80% B 80% C 50% A 50% B 50% C 30% A 30% B 30% C Fitoplancton T36 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1 µgC L-1
Nitzschia longissima 7268.33 6552.34 7566.84 3875.31 3352.65 3376.03 1941.91 2388.08 1540.35 745.74 790.36 639.51 Fragilariopsis cylindrus 5795.44 4843.01 6626.57 2259.77 4137.68 4429.69 2331.65 2812.36 2686.57 1186.04 1224.23 952.43 Fragilariopsis curta 544.13 479.48 393.28 274.76 317.86 312.47 188.56 150.85 150.85 102.36 118.52 80.81 Fragilariopsis kerguelensis 307.6 153.8 217.88 102.53 64.08 153.8 89.72 0 64.08 25.63 128.16 0 Pleurosigma sp. 109.12 120.04 152.77 114.58 125.49 95.48 51.83 51.83 60.02 35.46 40.92 38.19 Leptocylindrus danicus 46.2 46.2 26.4 19.8 26.4 39.6 13.2 13.2 9.9 6.6 16.5 0 Navicula sp. 0.12 0.12 0.37 0.12 0 0.37 0 0.37 0 0 0 0 Pseudonitzscha heimii 0 11.72 5.86 27.35 3.91 11.72 0 3.91 5.86 0 1.95 3.91 Thalassiosira gravida 0 173.23 1039.39 0 1039.39 0 0 0 0 0 0 0 Thalassiosira antarctica 0 305.76 407.68 203.84 0 0 0 0 0 0 0 0 Chaetoceros sp. 30.71 0 0 46.06 0 30.71 46.06 15.35 0 0 15.35 30.71 Primnesiophyceae Phaeocystis antarctica 0 100.38 100.38 0 100.38 66.92 0 0 0 0 0 0
ANTARTIDE ESPERIMENTO 8

Microzooplancton T0 100% A 100% B 100% C 100% A 100% B 100% C Cell L-1 Cell L-1 Cell L-1 Cell L-1 Cell L-1 Cell L-1 Dinoflagellida Gyrodinium sp. 3.84E+04 4.40E+04 4.80E+04 5.44E+04 4.96E+04 4.80E+04 Gyrodynium lachryma 4.00E+04 4.96E+04 3.68E+04 5.04E+04 4.24E+04 4.16E+04 Protoperidinium applanatum 5.60E+04 4.64E+04 4.16E+04 6.24E+04 4.96E+04 4.08E+04 Oligotrichida Laboea strobila 4.80E+03 6.40E+03 3.20E+03 4.80E+03 8.00E+03 3.20E+03 Strobilididae indet. 2.08E+04 1.28E+04 6.40E+03 1.28E+04 8.80E+03 1.20E+04 Tintinnida Cymatocylis drigalskii 2.40E+03 1.60E+03 1.60E+03 1.60E+03 1.60E+03 1.60E+03
Microzooplancton T36 100% A 100% B 100% C 100% A 100% B 100% C µg CL-1 µg CL-1 µg CL-1 µg CL-1 µg CL-1 µg CL-1 Dinoflagellida Gyrodinium sp. 2.09E+01 2.40E+01 2.62E+01 2.97E+01 2.70E+01 2.62E+01 Gyrodynium lachryma 2.18E+02 2.70E+02 2.00E+02 2.75E+02 2.31E+02 2.27E+02 Protoperidinium applanatum 1.19E+02 9.87E+01 8.84E+01 1.33E+02 1.05E+02 8.67E+01 Oligotrichida Laboea strobila 2.68E+01 3.57E+01 1.79E+01 2.68E+01 4.47E+01 1.79E+01 Strobilididae indet. 1.90E+02 1.17E+02 5.86E+01 1.17E+02 8.06E+01 1.10E+02 Tintinnida Cymatocylis drigalskii 4.81E+01 3.21E+01 3.21E+01 3.21E+01 3.21E+01 3.21E+01
ANTARTIDE ESPERIMENTO 8
Recommended