La replicazione del cromosoma di E. coli
Un unico replicone circolare di 4,6 milioni di bp
L’inizio della replicazione è un evento separato dalla replicazione del DNA e richiede uno specifico set di proteine. Essenzialmente si verifica la separazione dei filamenti complementari di DNA a un sito specifico, chiamato oriC, che permette l’ingresso e l’assemblaggio degli enzimi della replicazione.
oriC
DnaADnaBDnaCHuGirasiSSB
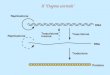
Le proteine richieste per l’inizio
La prima tappa nella formazione del complesso di inizio è il legame al sito oriC della proteina DnaA

Il sito oriC presenta sequenze ripetute di 9 e 13 basi all’interno di una regione di 245 bp che costituisce la sequenza minima necessaria per l’inizio della replicazione. Le due sequenze sono diverse ma comunque ricche in AT.
Si ritiene che DnaA riconosca inizialmente le sequenze di 9 bp e vi si leghi in maniera cooperativa fino a che 20-40 monomeri formino un nucleo centrale attorno al quale si avvolge il DNA di oriC Il risultato è l’apertura delle sequenze ripetute di 13 bp ricche in AT. A questo punto entrano DnaB, C ed altre proteine che formano ul complesso di 480.000 daltons.
DnaB è l’elicasi necessaria per separare la doppia elica. Altre proteine sono necessarie per l’inizio come pure per il prosieguo della replicazione e sono la DNA girasi che rimuove superavvolgimenti positivi e SSB che stabilizza il DNA a singola elica.

Si è venuta a formare così una forca replicativa da dove la sintesi di DNA potrà procedere bidirezionalmente non senza che sia prima stato sintetizzato un innesco di RNA che funge da primer per la DNA polimerasi III

Le varie fasi della replicazione possono essere seguite al ME. All’inizio si ha la struttura a “occhio” che si trasforma nella struttura intermedia a “teta” con l’avanzamento delle forche replicative
forma
terC
La replicazione termina al sito di terminazione indicato come terC dove si incontrano le due forche replicative. La regione contiene due sequenze di 23 bp che delimitano il punto di incontro.
23 bp
La completa replicazione del cromosoma richiede circa 40 minuti

La replicazione del cromosoma è seguita dalla segregazione: processo dove i cromosomi duplicati vengono disposti ai lati opposti della cellula in divisione. I batteri quali E. coli riescono a portare a termine questo processo in modo efficiente nonostante l’assenza di un apparato mitotico
I geni responsabili di questo processo non sono ancora del tutto noti ma sicuramente iplicato è l’operone mukFEB
Il gene B codifica per una proteina di 177 kDa che è la più grande proteina conosciuta in coli e ha una struttura simile alla miosina eucariotica. Ha una attività ATPasica e GTPasica e la parte globulare ha affinità per il DNA.
La segregazione del nucleoide
Mutanti in mukB portano alla formazione di cellule anucleate

Problematiche poste dalla replicazione
Quale è il segnale che dà inizio alla replicazione?
Quale meccanismo mette in relazione la fine della replicazione con la successiva divisione?
L’inizio della replicazione è determinato dal raggiungimento di una massa critica, quindi la frequenza di inizio è legata alla massa cellulare. Più un batterio cresce velocemente, cioè più velocemente può aumentare la sua massa, più frequentemente inizierà la replicazione. Deve pertanto esistere un meccanismo di controllo che correli la massa all’inizio della replicazione
Un modello per spiegare il fenomeno è che esista un iniziatore che venga sintetizzato costitutivamente e si accumuli proporzionalmente all’aumento della massa cellulare fino a raggiungere una concentrazione critica che avvii l’inizio della replicazione. Il modello non è stato ancora provato

Il ciclo cellulare di Escherichia coli
I C D
40 min 20 minvariabile
Fase I: periodo di interinizio. Osservabile in cellule che si dividono lentamente (T>60’); la cellula si prepara alla replicazione del DNA
Fase C: periodo necessario alla replicazione del DNA. Il limite è dovuto alla velocità di sintesi del DNA da parte della DNA polimerasi
Fase D: periodo della divisione. E’ il tempo che intercorre tra il completamento della replicazione del cromosoma e la divisione cellulare.

Il paradosso della divisione in E. coli
Come può un batterio dividersi ogni 20 minuti se il DNA impiega 40 minuti per duplicarsi?
forca replicativa

C D
40 min 20 min
T = 60’
0 10 20 30 40 50 60

D
T = 40’
0 10 20 30 40 50 60
replicazione iniziata 20’ prima

T = 20’
replicazione iniziata 20’ prima
replicazione iniziata 40’ prima
replicazione che inizia

1Lla distanza tra il centro dei due nuceoidi deve essere pari a una lunghezza cellulare per permettere la formazione del setto

La divisione nei batteri gram-negativi comporta la cresita interna e coordinata dei tre strati che delimitano il citoplasma sotto forma di un setto
La formazione del setto richiede una regolazione: 1) TEMPORALE2) SPAZIALE
La regolazione temporale fa in modo che la divisione avverrà sempre dopo la duplicazione del DNA (un blocco nella sintesi del DNA non sarà seguito dalla divisione). Un ruolo importante è svolto dalla proteina SulA (sistema sos) che inibisce in modo selettivo la principale proteina responsabile della formazione del setto (FtsZ).Altri fattori che attivano la formazione del setto devono ancora essere individuati.
La divisione in E. coli


La regolazione spaziale della divisione in E. coli
In E. coli come in altri batteri di forma bastoncellare, il setto si localizza esattamente nella posizione centrale della cellula.
Come viene scelta la corretta posizione del setto?
SI NO
Si ritiene che siano 2 i fattori coinvolti nel corretto posizionamento del setto:
1) OCCLUSIONE DEL NUCLEOIDE2) SISTEMA Min

Occlusione del nucleoide
Il nucleoide esercita un effetto negativo sul posizionamento del setto: in una regione occupata dal nucleoide non si formerà mai il setto.
Il volume cellulare è occupato dal nucleoide. Non ci sono siti liberi per la formazione del setto
In seguito alla segregazione del DNA, la regione mediana rimane libera dal nucleoide. Il setto può formarsi
Non è richiesta la completa separazione dei nucleoidi ma è sufficiente una diminuizione della concentrazione del DNA

Il sistema Min
Il sistema Min di E. coli consiste di 3 proteine: MinCMinDMinE
MinC è un inibitore della divisione ed è attivato da MinD. (complesso MinCD)
L’azione inibitrice di MinC è esercitata direttamente sulla proteina FtsZ
MinD è una proteina associata alla memrana MinE agisce su MinD ed è responsabile della localizzazione spaziale del complesso MinCD: la sua funzione è quella di far localizzare MinCD ai poli della cellula e lontano dal piano di divisione mediano.
Ma come agisce MinE?MinE provoca una rapida oscillazione di MinD tra un polo e l’altro della cellula nel giro di pochi secondi. In questo movimento è coinvolto anche MinC in quanto associato a MinD.

Comportamento dinamico di GFP-MinD
tratto da:The EMBO Journal, Vol. 20, No. 7 pp. 1563-1572, 2001

La rapida oscillazione fa in modo che l’inibitore del setto MinCD sia statisticamente più presente ai poli che al centro impedendo, quindi, la formazione del setto ai poli
I poli sono siti potenziali di divisione non occlusi dal nucleoide
La formazione di un setto a un polo porta alla formazione di minicellule (cellule di dimensioni più piccole e anucleate)
minicellula

Una fase precoce della divisione nei batteri è la formazione dello Z ring a livello del sito di divisione. La formazione dello Z ring è dovuta alla capacità della proteina FtsZ di polimerizzare
Formazione del setto e divisione in E. coli
FtsZ-GFP
Z-ring

La sintesi dello Z-ring parte da un sito di nucleazione localizzato a livello della membrana citoplasmatica e prosegue bidirezionalmente fino a formare un anello
FtsZGTP
GDP
Sito di nucleazione
Z-ring

Lo Z-ring va incontro a una successiva costrizione fornendo la forza che porta all’invaginazione della membrana citoplasmatica seguita dagli altri strati fino alla completa chiusura del setto e divisione della cellula

Oltre FtsZ altre proteine partecipano alla formazione del setto:ZipA, FtsA, FtsI, FtsL, FtsN, FtsQ, FtsK, FtsW
I geni che codificano per le proteine del setto sono essenziali
Per ogni gene sono stati isolati mutanti letali condizionali (termosensibili)
Questi mutanti, a temperatura non permissiva (42°C), hanno un fenotipo caratteristico (foto A): formano lunghi filamenti non settati o con il setto appena accennato
A)
B)
Fts = Filaments
Foto B) Mutanti Fts a temperatura permissiva (30°C)

FtsZ
E’ la proteina principale del setto e quella che interviene per prima
Proteina citoplasmatica di 40.000 D presente in circa 20.000 copie per cellula
E’ omologa alle tubuline eucariotiche
E’ una GTPasi. E’ capace di polimerizzare. La polimerizzazione richiede GTP
La polimerizzazione è inibita da SulA (controllo della replicazione) e da MinC (controllo spaziale della localizzazione del setto)
Tra le proteine Fts è quella più conservata tra i procarioti: è assente solo nelle clamidie. E’ presente in alcuni mitocondri e cloroplasti

FtsA
Proteina citoplasmatica presente in circa 200 copie per cellula
E’ omologa all’actina e ha una attività ATPasica
E’ una proteina di membrana.
Interagisce direttamente con FtsZ. Ancora lo Z-ring alla membrana
Sono proteine di membrana. Si associano allo Z-ring dopo FtsA e ZipA
Interagisce direttamente con FtsZ ed è la seconda a far parte del setto
ZipA
FtsI, L, N, K, Q, W
Il loro ruolo è ancora sconosciuto (FtsI è responsabile della sintesi del peptidoglicano a livello del setto)

I virus

I virus: caratteristiche generali
dimensioni estremamente piccole
parassitismo intracellulare obbligato
organizzazione strutturale semplice non cellulare
presenza di un solo tipo di acido nucleico (DNA o RNA)
I virus sono agenti infettivi con le seguenti caratteristiche distintive:
Spettro d’ospite. I virus si moltiplicano solo in particolari cellule ospiti e di conseguenza vengono suddivisi in tre classi principali: virus batterici (o batteriofagi)
virus animali
virus vegetali
si moltiplicano per sintesi separata e successiva riunione dei loro componenti

Origine dei virus
Evoluzione retrograda
I virus sarebbero forme degenerate di parassiti intracellulari: i loro genomi si sarebbero progressivamente ridotti in quanto la maggior parte delle funzioni vengono fornite dalla cellula ospite.
Questa ipotesi potrebbe spiegare l’origine di virus complessi quali i Poxvirus
Teoria della “evasione dei geni”
I virus si sarebbero evoluti da elementi genetici capaci di replicazione autonoma (plasmidi e trasposoni)

Struttura dei virus
Le particelle virali, o virioni, sono costituite da una o più molecole di DNA o RNA rivestite da un involucro proteico o capside.In alcuni virus il nucleocapside (acido nucleico + capside) può essere avvolto da un rivestimento membranoso (envelope)
Le dimensioni possono variare da 10 a 400 nm
In base alle caratteristiche morfologiche è possibile individuare 4 gruppi principali:
Virioni con capside icosaedrico
Virioni con capside elicoidale
Virioni dotati di envelope
Virioni complessi
Sia i capsidi elicoidali che icosaedrici sono strutture macromolecolari costruite da una o poche subunità proteiche, dette capsomeri, ripetute un gran numero di volte. Questa strategia costruttiva ha il vantaggio di utilizzare con la massima efficienza l’informazione immagazzinata nel genoma virale

Capsidi a simmetria elicoidale
Di forma cilindrica, costituiti da un unico tipo di capsomero avvolto a spirale attorno all’asse centrale contenente l’acido nucleico
I capsidi possono essere rigidi (virus del mosaico del tabacco e il batteriofago M13) o flessibili (virus influenzali)
Esempio: virus del mosaico del tabacco (TMV)
Capside cilindrico rigido di 15-18 nm di diametro e circa 300 nm di lunghezza. L’unica proteina del capsomero è di 158 aa
Acido nucleico: RNA a singolo filamento di 6000 nucleotidi
Spesso la ripetizione di una sola proteina rappresenta l’unica possibilità di rivestire un acido nucleico di piccole dimensioni
Cristallizzato da Stanley nel 1935

Capsidi a simmetria icosaedrica
Icosaedro: solido regolare a 20 facce triangolari e 12 vertici
I capsomeri sono formati da 5 o 6 protomeri che possono essere anche tutti identici
I pentoni o pentameri possiedono 5 subunità e sono disposti ai vertici dell’icosaedro; gli esoni o esameri hanno 6 subunità e costituiscono le facce
Il numero totale di capsomeri per i diversi virus assume valori definiti da equazioni derivate dalla cristallografia; sono possibili solo i valori 12, 32, 42, 60, 72, 92, 162, 252 ecc
Esempi: i batteriofagi X174, MS2, Qparvovirus (12 capsomeri), poliovirus (32 capsomeri), papilloma virus (72 capsomeri), adenovirus (252)

Una volta sintetizzati ed esposti alle opportune condizioni, i protomeri interagiscono tra loro in modo specifico, associandosi spontaneamente a formare il capside
L’autoassemblaggio generalmente non richiede alcun intervento dall’esterno. Alcuni virus più complessi codificano per particolari fattori che, pur non essendo incorporati nel virione, risultano necessari per l’assemblaggio del virus
L’auto-assemblaggio dei virus
Protomeri e capsomeri sono legati tra loro mediante legami deboli non covalenti. Infatti, i capsidi (vuoti) sono facilmente dissociabili nei loro costituenti.

Virus dotati di envelopePresentano il nucleocapside rivestito da strutture membranarie complesse (involucro pericapsidico)
I virus con involucro sono comuni nel mondo animale, ma si conoscono anche virus battericiL’involucro pericapsidico è generalmente costituito da un doppio strato lipidico in cui sono inserite glicoproteine (spicole)
Il capside all’interno può avere simmetria elicoidale o icosaedricaP419a
La membrana deriva dalla cellula infettata in seguito al processo di liberazione dei virioni; le glicoproteine sono di origine virale

Virus con capsidi a simmetria complessa
Hanno una struttura complessa non assimilabile né a quella elicoidale né a quella icosaedrica
Rappresentanti: i poxvirus e i grandi batteriofagi
I poxvirus, di forma ovoidale o a mattone, sono i virus animali più grandi e possono essere osservati anche al microscopio ottico
Il genoma (DNA a doppio filamento) è associato a proteine e racchiuso in un nucleoide, a forma di disco biconcavo, delimitato da una membrana. Due corpi laterali, di forma ellittica, sono disposti tra il nucleoide e l’involucro esterno. L’involucro è di natura membranaria, rivestito da tubuli e fibre

I genomi virali
Per la maggior parte dei virus a RNA il materiale genetico è rappresentato da un singolo filamento di RNA (ss RNA)
Se la sequenza corrisponde a quella dell’mRNA virale il filamento è definito positivo o + Esempi: il virus polio, il virus del mosaico del tabacco, e il virus del sarcoma di Rous sono virus con genoma a ssRNA positivo
Se la sequenza è complementare a quella dell’mRNA virale il filamento è definito negativo o - Esempi: il virus della rabbia, il virus del morbillo, e il virus dell’influenza sono virus con genoma a ssRNA negativo
I virus animali con filamento positivo presentano modificazioni tipiche dei messaggeri eucariotici: 7-metilguanosina (cap) al 5’ e coda di poli-A al 3’Molti genomi a RNA risultano segmentati e si ritiene che ogni segmento codifichi per una proteina virale

colletto
guainaelicoidale
spine
fibrecaudali
asse tubulare(cavo)
placca basaleesagonale
testa
Virus con capsidi a simmetria complessa
I batteriofagi della serie T pari (T2, T4, T6)

Virus con capsidi a simmetria complessa
I batteriofagi T1, T5 e hanno una testa icosedrica, la coda è priva di guaina, e non termina con una base piatta ma con rudimentali fibre caudali
Il batteriofago lambda
testa icosaedrica
coda non contrattile
I batteriofagi T3 e T7 possiedono una testa icosedrica, la coda breve non contrattile e priva di fibre caudali
Tutti questi batteriofagi sono detti virus a simmetria binaria, poiché hanno sia la simmetria icosaedrica (testa) che elicoidale (coda)

I genomi virali
I virus contengono un unico tipo di acido nucleico in tutte e quattro le combinazioni: DNA a singola elica
DNA a doppia elicaRNA a singola elicaRNA a doppia elica
Anche la struttura del genoma è varia nei diversi virus. Il materiale genetico può essere organizzato in :
un’unica molecola lineare
un’unica molecola circolare (DNA)
segmentato in più molecole

I genomi virali
La capacità codificante va dalle 3-4 proteine dei virus più piccoli (MS2 e Qb) alle oltre 100 proteine dei batteriofagi della serie T-pari e dei Poxvirus
Nella maggior parte dei virus a DNA il genoma è costituito da un doppio filamento (ds DNA) che può essere lineare oppure circolare
GGGCGGCGACCT
CCCGCCGCTGGA
Circolarizzazione del DNA di
GGGCGGCGACCT
CCCGCCGCTGGA

La formazione dell’RNA messaggero virale
Virus a DNA
ds DNA+-
RNA pol usa come stampo il filamento -mRNA (+)
ss DNA+
DNA polmRNA (+)
+-
RNA pol
Affinché possano essere sintetizzate le nuove proteine virali, è prima necessario che siano sintetizzati nuovi messaggeri virali

Virus a RNA positivo
ss RNA+mRNA (+)funziona direttamente da mRNA
-Replicasi
(RNA polimerasi RNA dipendente)
+
La formazione dell’RNA messaggero e replicazione virale
N.B. Sia nelle cellule eucariotiche che procariotiche sono presenti solo RNA polimerasi DNA dipendenti (non possono usare RNA come stampo)
Il filamento + funge direttamente da mRNA che codifica, oltre alle altre proteine necessarie, anche la Replicasi (Rna pol RNA dipendente). Questa inizialmente sintetizza RNA negativi usando come stampo il filamento positivo del genoma, e poi li usa come stampi per produrre ulteriori molecole positive che costituiranno i genomi della progenie virale

Virus a RNA negativo
- ss RNA mRNA (+)
RNA polimerasi RNA dipendenteassociata al virus
-
La formazione dell’RNA messaggero e replicazione virale
L’RNA genomico non può funzionare come mRNA né le cellule ospiti hanno polimerasi capaci di trascrivere mRNA usando come stampo RNA.
Come può essere sintetizzato RNA messaggero?
Per superare il problema questi virus portano, nel loro capside, alcune molecole di Trascrittasi (Rna pol RNA dipendente) che inizialmente sintetizza RNA positivo che funge da messaggero, e da questo, in seguito sintetizzera gli RNA negativi che costituiranno il genoma della progenie virale

La formazione dell’RNA messaggero e replicazione virale
Virus a RNA a doppio filamento
ds RNA+mRNA (+)
RNA polimerasi RNA dipendenteassociata al virus
-
Replicasi(RNA polimerasi RNA dipendente)
-usa come stampo il filamento
+-
Anche in questo caso l’RNA genomico non può funzionare come RNA messaggero
Alla sintesi del messaggero provvede una Trascrittasi associata al virus e inettata insieme al genoma virale. Il messaggero virale codifica per un’altra RNA pol RNA dipendente che svolge la funzione di Replicasi e sintetizzerà RNA a doppio filamento per le nuove particelle virali.
Trascrittasi

La formazione dell’RNA messaggero e replicazione virale
+ ss RNA
mRNA (+)
+-
Trascrittasi inversa
DNA polimerasi RNA dipendenteassociata al virus
+ ss RNA
(virione)
Retrovirus
Sono virus a singolo filamento di RNA + ma si differiscono da tutti gli altri perché sintetizzano i propri mRNA e replicano il proprio genoma passando attraverso un intermedio a DNA
Sono caratteristici anche per il fatto di essere “diploidi”. Infatti, nel loro capside sono presenti sempre due filamenti di RNA. Associati al genoma sono anche la Trascrittasi inversa (DNA polimerasi RNA dipendente) e tRNA di origine virale che svolge la funzione di primer per la sinstesi del DNA virale

tRNA virale
5’ 3’
attività RNAsi H
La trascrizione inversa
degradazione RNA
ibrido RNA/DNA
5’3’
sintesi del filamento complementare di DNA

Coltivazione dei virus
I virus animali vengono coltivati inoculando animali sensibili o uova embrionate di pollo (uova fecondate e incubate per 6-8 giorni)
Di recente i virus vengono coltivati su monostrati di cellule animali; questa tecnica mette in evidenza aree localizzate di lisi cellulare chiamate placche, o foci proliferativi (virus oncogeni)
I virus vegetali possono essere coltivati impiegando colture di tessuti vegetali, oppure piante intere. Le foglie si inoculano meccanicamente spalmandole con una miscela di virus e una sostanza abrasiva. Nelle aree infette si sviluppa una lesione necrotica localizzata
I batteriofagi sono i virus più facilmente coltivabili. L’infezione di una coltura di batteri sensibili porta nel giro di circa un’ora alla lisi dei batteri con produzione di una notevole quantità di virus (lisato)

Metodi di saggio dei virus
È possibile determinare la quantità di virus presente in un campione:1) contando il numero di particelle virali2) misurando la concentrazione di unità infettanti
Il numero delle particelle virali può essere anche 1 milione di volte superiore al numero delle unità infettanti
Conteggio del numero di particelle viralidiretto: microscopio elettronicoindiretto: saggio di emoagglutinazione
Conteggio delle unità infettantisaggio delle placchedose infettante (ID50) e dose letale (LD50)

Conteggio al ME
Il campione virale viene mescolato a microscopiche sferette di latex a concentrazione nota. Per una determinata superficie si fa il rapporto tra il numero di virus e il numero di sfere e si risale alla concentrazione virale

Saggio di emoagglutinazione
eritrociti virus
diluizione 10 20 40 80 160 320 640 1280 2560 5120
Diluizioni seriali di sospensione virale vengono aggiunte ai globuli rossi. Il titolo emoagglutinante è dato dalla diluizione più alta che dà ancora agglutinazione

Saggio delle placche
Ogni placca ha avuto origine da una singola particella virale, quindi, il numero delle placche prodotte equivale al numero di virioni infettanti o di unità formanti placca (PFU).
Dal numero delle placche e considerando la diluizione effettuata, si risale alla concentrazione di particelle infettanti.
placche di lisi su tappeto batterico
4-5 ml di top agarinfezione
0,2 ml di batteri
ultima diluizione virale
dopo 24h

Il saggio delle placche può essere applicato anche a virus animali e vegetali quando si dispone di cellule sensibili all’infezione che crescano in vitroIn alternativa, molti virus animali possono essere inoculati sulla membrana corionallantoidea degli embrioni di pollo dove producono lesioni localizzate dette pustole
Per i virus vegetali si spalma una sospensione di virus con una sostanza abrasiva su foglie sensibili. I virus daranno delle lesioni necroticheIn tutti i casi, moltiplicando il fattore di diluizione per il numero di pustole o lesioni necrotiche si ottiene la concentrazione di unità infettanti di una sospensione virale
Quando è impossibile ricorrere al saggio delle placche si usa il metodo della diluizione finale. Organismi viventi o colture cellulari sono inoculati con diluizioni seriali di una sospensione virale per determinare la diluizione che uccide o danneggia il 50% delle cellule o degli organismi impiegati.
LD50 = dose letale per il 50% del campione
ID50 = dose infettante per il 50 % del campione

Efficienza di piastramento
I conteggi ottenuti con il saggio delle placche sono sempre inferiori a quelli ottenuti con il microscopio elettronico. L’efficienza con cui i virus infettano le cellule ospiti raramente raggiunge il 100% e spesso può essere considerevolmente inferiore.
Una efficienza inferiore al 100% può essere in parte dovuta alla presenza di particelle virali difettive, ma più spesso è dovuta al fatto che, nelle condizioni usate, l’infezione da parte di queste particelle non ha avuto successo.Mentre per i virus batterici l’efficienza di piastramento supera spesso il 50%, nel caso di molti virus animali può essere molto bassa, tra lo 0,1 e 1%.
Il rapporto quantitativo tra il numero delle unità infettanti e il numero delle particelle virali (determinato mediante conteggio al microscopio elettronico) è definita efficienza di infezione.
E.I =Unità Infettanti
Conteggio al ME
Anche se sono necessarie molti virus per infettare una cellula, l’infezione è iniziata sempre da un solo virus

Le fasi della moltiplicazione dei batteriofagi
1. Adsorbimento
2. Iniezione dell’acido nucleico
3. Fase replicativa precoce
4. Replicazione del genoma virale
5. Sintesi delle proteine capsidiche
6. Assemblaggio del capside e impacchettamento del genoma virale
7. Rilascio dei virus maturi (lisi)
1. Adsorbimento
2. Iniezione dell’acido nucleico
3. Fase replicativa precoce
4. Replicazione del genoma virale
5. Sintesi delle proteine capsidiche
6. Assemblaggio del capside e impacchettamento del genoma virale
7. Rilascio dei virus maturi (lisi)

Esperimento di crescita a ciclo unico (one-step)
L’ esperimento one-step permette di seguire le varie fasi della moltiplicazione virale partendo da un’infezione sincrona di una coltura batterica
Delbruck e Ellis, 1939
1) infezione di una coltura batterica (T=0 dell’esperimento). Tutti i batteriofagi inizieranno il ciclo di moltiplicazione
contemporaneamente
2) lavaggio della coltura per eliminare i batteriofagi non ancora adsorbiti
3) ad intervalli di tempo viene determinato il numero di batteriofagi
liberati mediante il saggio delle placche

Curva di crescita a ciclo unico
periodo di latenza
periodo di eclisse
periodo di crescita
volume di scoppio
Tempo (minuti)
Num
ero
dei b
att
eri
ofa
gi
periodo di eclisse: periodo durante il quale non è possibile rilevare la presenza di particelle virali infettive neanche all’interno dei batteri
periodo di latenza: periodo di sviluppo dei fagi nel batterio infettato. I fagi non sono ancora liberati all’esterno. Nella fase finale sono presenti particelle mature nel batterio.periodo di crescita: le cellule vanno incontro a lisi liberando i fagi infettanti. Quando tutti i batteri infettati saranno lisati si raggiunge il plateau
plateau
N. fagi al plateau
N. fagi latenza=

L’adsorbimento dei batteriofagi
I batteriofagi non aderiscono a un punto qualunque della superficie batterica ma riconoscono recettori specifici
I recettori sono normali componenti della superficie dell’ospite quali proteine della parete, polisaccaridi, lipopolisaccaridi, acidi teicoici, flagelli e pili
In assenza di siti recettoriali il virus non può adsorbirsi
Se un recettore è alterato, l’ospite diventa resistente alla infezione da parte del virus che usa quel recettore. Tuttavia, anche i virus possono mutare per la struttura che riconosce il recettore tornando, così, capaci di infettare un ospite resistente
Fago PBSI di B. subtilis

Penetrazione dell’acido nucleico
Nella gran maggioranza dei batteriofagi è solo l’acido nucleico virale a entrare nel batterio.
I meccanismi di penetrazione differiscono notevolmente tra i diversi fagi finora studiati e sono in gran parte ancora oscuri.
Nei fagi T-pari l’adsorbimento è dovuto al contatto tra le fibre caudali e il recettore (proteina della membrana esterna). La penetrazione del DNA si verifica dopo che la placca basale si è adagiata sulla superficie cellulare e si verificano cambiamenti conformazionali sia nella placca che nella guaina. Quest’ultima si contrae permettendo all’asse tubulare di penetrare attraverso la parete. Infine, con un meccanismo ancora sconosciuto, il DNA dalla testa, passando attraverso l’asse tubulare, entra nella cellula.
E. coli infettato da T4

I geni con funzioni correlate sono generalmente raggruppati per essere espressi contemporaneamente in un preciso istante del ciclo di replicazioneI geni introdotti nella cellula ospite vengono espressi secondo una ordinata sequenza temporale che consente una efficiente coordinazione degli eventi che portano alla produzione della progenie virale
Regolazione dell’espressione dei geni fagici
Geni precoci immediati: trascritti subito dopo l’ingresso, codificano per prodotti che partecipano alla replicazione del DNA virale e che bloccano le attività del batterio. La loro trascrizione si arresta nel giro di pochi minuti.
Geni intermedi o precoci ritardati: partecipano alla replicazione e alla ricombinazione del DNA. Possono essere trascritti per l’intero ciclo replicativo
Geni tardivi: codificano per proteine capsidiche, fattori che partecipano all’assemblaggio e proteine litiche
Il passaggio da una fase all’altra è geneticamente controllato attraverso la sintesi di nuova RNA polimerasi (T7) o fattori che alterano la specificità della RNA polimerasi batterica

geni precoci immediati
blocca il fattore sigmageni precoci ritardati
Una delle proteine precoci immediate ha la funzione di bloccare il fattore sigma e quando raggiunge una certa concentrazione, i geni precoci immediati sono bloccati
Il controllo della trascrizione in T4 coinvolge la sintesi di proteine che modificano la specificità della RNA polimerasi dell’ospite in modo da farle riconoscere i diversi promotori fagici. I promotori precoci immediati sono letti direttamente dall RNA polimerasi attraverso il fattore sigma dell’ospite
fattori che modificano l’RNA polimerasi
geni ritardati
Regolazione della trascrizione in T4

Replicazione e sintesi delle proteine del fago T4
T4 ha uno dei più grandi genomi virali (1,7 x 105 bp) costiutito da DNA a doppio filamento lineare
Vengono interrotte le attività del batterio (DNA, RNA e proteine batteriche non vengono più sintetizzati).
Immediatamente dopo l’ingresso del DNA vengono sintetizzate le proteine precoci: sono gli enzimi coinvolti nella trascrizione e replicazione del DNAvirale e enzimi che degradano il DNA batterico per fornire nucleotidi
Dopo 5 minuti ha inizio la replicazione del DNA virale
La sintesi del DNA di T4 richiede una notevole preparazione perché contiene idrossimetilcitosina al posto della citosina, che successivamente viene anche glucosilata. Questi resudui glucosilati proteggono il DNA di T4 dall’attacco di alcune endonucleasi di E. coli, dette anche enzimi di restrizione, che altrimenti distruggerebbero il DNA virale effettuando tagli in siti specifici. Questo meccanismo di difesa batterica è detto restrizione. Il DNA batterico è protetto dall’azione di questi enzimi grazie alla presenza di enzimi di modificazione che effettuano modificazioni nei siti specifici riconosciuti dagli enzimi di restrizione

1 2 3 4 5 6 7 8 9 1
2 3 4 5 6 7 8 9 1 2
3 4 5 6 7 8 9 1 2 3
4 5 6 7 8 9 1 2 3 4
Il genoma di T4 è un dsDNA lineare e presenta il fenomeno della permutazione circolare e della ridondanza terminale (una stessa sequenza ripetuta alle estremità del DNA)
Organizzazione del genoma del fago T4

La replicazione del DNA di T4
Dopo diversi cicli di replicazione si vengono a formare concatenameri di lunghezza considerevole (fino a 20 copie di DNA)
concatenamero

Assemblaggio delle particelle fagiche
I geni tardivi dirigono la sintesi di tre tipi diversi di proteine:
1) proteine strutturali del fago2) proteine che intervengono nell’assemblaggio ma non faranno parte del fago3) proteine coinvolte nel processo di lisi batterica e liberazione dei fagi
Le proteine necessarie all’assemblaggio vengono sintetizzate contemporameamente e sono utilizzate in “catene di montaggio” quasi indipendenti fino ad avere il fago maturoL’impacchettamento del DNA avviene quando la testa è matura: circa 500 m di DNA devono essere inseriti in una cavità di appena 0,1 m.
L’impacchettamento avviene secondo il meccanismo detto “a testa piena” a partire dai concatenameri sintetizzati. Quando il volume della testa è totalmente riempito il concatenamero viene tagliato. In questo modo entra il 102% del genoma virale.
Questo processo è responsabile della ridondanza terminale e della permutazione circolare del genoma di T4

1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
102% 102% 102% 102%
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4
Ridondanza terminale e permutazione circolare del genoma di T4

X174 e il fenomeno dei geni sovrapposti
X174 è un piccolo batteriofago icosaedrico con genoma a DNA circolare a singolo filamento
È un esempio di come virus contenenti genomi di lunghezza limitata sfruttino al massimo l’informazione genetica attraverso il fenomeno dei geni sovrapposti
5386
I 5386 nucleotidi del genoma di X174 non sarebbero sufficienti a codificare tutte le proteine virali identificate. Si è scoperto, quindi, che una sequenza può codificare anche per due proteine: i geni sono sovrapposti.
Il gene E è contenuto all’interno di D su un altro schema di lettura. La proteina A* è parte della proteina A ed è ottenuta tramite un re-inizio della traduzione.
A sintesi della forma replicativaA* inibizione della sintesi di DNA dell’ospiteE lisi della cellula ospite

polarità +
FR forma replicativa a doppia elica
+
-
enzima che taglia la singola elica
elica nascente
filamento spiazzato
punto di crescita
una rotazione completa
filamento +
taglio e circolarizzazione di un genoma completo ad opera della proteina virale A
+
La replicazione a “cerchio rotante” di X174

Caratteristiche di un batterio lisogeno
I batteri lisogeni hanno il DNA virale (profago) integrato nel loro cromosoma (nel caso del fago P1 il DNA provirale e libero nel citoplasma).Il DNA del profago viene duplicato insieme a quello dell’ospite e trasmesso da una generazione di batteri a quella successiva.
All’interno di una popolazione di batteri lisogeni una piccola frazione (tra 10-3 e 10-6) delle cellule produce virus e muore.
Un batterio lisogeno per un fago è immune all’infezione con lo stesso fago. Rimane sensibile all’infezioni di fagi differenti. L’immunità è dovuta agli stessi sistemi regolativi virali che bloccano la via litica.
In particolari condizioni quasi tutta la popolazione di batteri lisogeni può andare incontro a lisi e produrre nuovi fagi. Il fenomeno va sotto il nome di induzione.
Un fago temperato può indurre cambiamenti fenotipici nel batterio. Tale cambiamento è detto conversione lisogena. Es.: il fago temperato di Corynebacterium diphteriae produce la tossina responsabile della difterite

Batteriofagi temperati e lisogenia
ciclo litico ciclo lisogenico
induzioneprofago
batterio lisogeno

Il batteriofago lambda
testa icosaedrica
coda non contrattile
64 nm
150 nm
GGGCGGCGACCT
5’5’
CCCGCCGCTGGA
sito cos
sito cos
dsDNA 48.502 bp

Fase precoce immediata
Solo due geni vengono espressi: cro e N.
Sono gli unici due geni i cui promotori sono riconosciuti dalla RNA polimerasi batterica.
Il prodotto del gene N (antiterminatore) permette il passaggio alla fase successiva.
Regione d’immunità

Fase precoce ritardata
La trascrizione procede verso destra per i geni della replicazione e verso sinistra per i geni della ricombinazione.
Il prodotto del gene Q (antiterminatore) permette il passaggio alla fase successiva.
A questo punto viene effettuata la scelta tra il ciclo litico e il ciclo lisogenico

Fase tardiva
È stata scelta la via litica. I geni precoci non vengono più trascritti. Vengono sintetizzate le proteine strutturali della coda e della testa e gli enzimi responsabili della lisi.
Se viene scelta la via litica il cromosoma di non si integra nel cromosoma batterico

La via lisogenica
attP
attB
Il cromosoma di si integra nel cromosoma batterico.Una volta integrato un solo gene di è responsabile del mantenimento dello stato lisogenico: il gene cI

PR/ORPL/OL
crocIN cIIcIII
La regione di immunità di
Fase precoce immediata
antiterminatore
Cro
responsabile della via litica
RNA polimerasi batterica
gene trascritto
gene spento
I promotori PR/OR e PL/OL sono riconosciuti dalla RNA polimerasi batterica

PR/ORPL/OL
crocIN cIIcIII
La regione di immunità di
Fase precoce ritardata
protegge cII
cII
regolatore positivo di cI
RNA polimerasi batterica
gene trascritto
gene spento
cIII

PR/ORPL/OL
crocIN cIIcIII
La regione di immunità di
Fase precoce ritardata
cII + cIII
Repressore cI
PE
responsabile del mantenimento dello stato lisogenico

1
92
132
236
N
C
Il dominio N-terminale costituisce il sito di legame all’operatore
Il dominio C-terminale è responsabile della formazione del dimero
Il repressore è capace di legarsi al DNA solamente come dimero
N
C
N
C
Il repressore cI di lambda

PR/ORPL/OL
crocIN cIIcIII
La regione di immunità di
PE
OR1OR2OR3
trascrizione di cI
trascrizione di cro
PRM
PR

L’interruttore genetico di
OR1OR2OR3
trascrizione di cI
trascrizione di cro
PRM
PR
Il legame di Cro in OR3 blocca la sintesi del repressore cI. E’ stata scelta la via litica

L’interruttore genetico di
OR1OR2OR3
trascrizione di cI
trascrizione di cro
PRM
PR
Il legame del repressore cI in OR1 blocca la sintesi di Cro.
Il repressore cI ha una maggiore affinità per il sito OR1

L’interruttore genetico di
OR1OR2OR3
trascrizione di cI
trascrizione di cro
PRM
PR
Il legame del dimero in posizione OR1 favorisce il legame di un secondo dimero in posizione OR2 (legame cooperativo)
L’affinità per OR2 è inferiore all’affinità per OR1

L’interruttore genetico di
OR1OR2OR3
trascrizione di cI
trascrizione di cro
PRM
PR
Il legame del dimero in posizione OR2 regola positivamente la trascrizione del repressore stesso (regolazione autogena )
controllonegativo
controllopositivo
RNApolimerasi

PR/ORPL/OL
crocIN cIIcIII
Fase precoce immediata
antiterminatore
Cro
responsabile della via litica
I promotori PR/OR e PL/OL sono riconosciuti dalla RNA polimerasi batterica

PR/ORPL/OL
crocIN cIIcIII
Fase precoce ritardata
protegge cII
cII
regolatore positivo di cI
cIII

PR/ORPL/OL
crocIN cIIcIII
Fase precoce ritardata
cII + cIII
Repressore cI
PE

Ciclo litico
OR1OR2OR3
PRM
PR
PR/ORPL/OL
crocIN cIIcIII
PE
Verso la fine della fase precoce ritardata Cro si lega anche a OR2 e OR3 come pure a PL/OL bloccando la trascrizione dei geni precoci

Ciclo lisogenico
OR1OR2OR3
PRM
PR
PR/ORPL/OL
crocIN cIIcIII
PE
La continua trascrizione di cI assicura il mantenimento dello stato lisogenico.
Un eccesso di cI viene ricondotto alla norma grazie al legame in posizione OR3

L’immunità alla superinfezione
Batterio lisogeno per risulata immune all’infezione di
cI
Il repressore codificato dal profago blocca la trascrizione dei geni precoci del DNA infettante legandosi a PR/OR e PL/OL

Induzione del ciclo litico
L’induzione rappresenta una risposta a fattori ambientali (luce UV) o mutageni chimici che danneggiano il DNA dell’ospite
Come parte della risposta SOS, aumenta la concentrazione della proteina RecA che normalmente è responsabile della ricombinazione genetica
Ad alta concentrazione RecA interagisce con cI
N
C
RecA
attività endoproteolitica dell’estremità C-terminale del repressore
C
N

Induzione del ciclo litico
trascrizione dei geni litici

Mutazioni a carico dei geni cI, cII e cIII danno placche dal fenotipo “chiaro”
Mutazioni a carico dei siti OR e OL possono essere “virulente”


Per la classificazione vengono prese in considerazione le caratteristiche morfologiche e la composizione in acido nucleico

Adsorbimento dei virus animali
Adsorbimento avviene mediante l’incontro casuale tra il virus e la cellula ospite. Generalmente una glicoproteina sulla membrana plasmatica della cellula ospite costituisce il recettore
La capacità di un virus di infettare un determinato tipo di cellula o di tessuto risiede in gran parte nella distribuzione dei recettori
Il sito di legame sulla superficie del virus può essere rappresentato semplicemente da una proteina strutturale del capside come ad esempio le fibre che si estendono dai vertici degli icosaedri negli adenovirus, o le spicole dei virus dotati di envelope

Penetrazione e spoliazione
La penetrazione consiste nel passaggio del virus attraverso la membrana plasmatica.
La spoliazione è l’allontanamento del capside e la liberazione dell’acido nucleico virale.
In alcuni virus nudi, come il poliovirus, dopo l’adsorbimento alla membrana cellulare si verificano cambiamenti strutturali nel capside che portano alla liberazione nel citoplasma del solo acido nucleico.

In virus rivestiti come i paramyxovirus, l’envelope virale si fonde con la membrana citoplasmatica e il nucleocapside entra nella cellula.

Tuttavia la maggior parte dei virus con envelope entra nella cellula attraverso un processo di endocitosi mediato dall’interazione tra il virus e i recettori localizzati in porzioni specializzate della membrana citoplasmatica rivestite, sul lato citoplasmatico, di una proteina detta clatrina
In seguito al contatto la membrana si introflette formando delle vescicole contenenti le particelle virali. Le vescicole si fondono con i lisosomi e l’acidificazione sembra favorire l’espoliazione. La membrana della vescicola e l’envelope si fondono e il nucleocapside viene liberato nel citoplasma.
La spoliazione termina con la disgregazione del capside e la liberazione dell’acido nucleico.


Trascrizione e replicazione
L’espressione dei geni precoci serve a ottenere il controllo della cellula ospite e a replicare il genoma virale.
Alcuni virus animali inibiscono la sintesi di DNA, RNA e proteine dell’ospite ma generalmente non degradano il DNA cellulare.
Virus a DNA
Per quanto riguarda i virus a DNA, la replicazione del DNA virale avviene nel nucleo cellulare con l’eccezione dei poxvirus: la loro replicazione avviene nel citoplasma grazie alla sintesi di una DNA polimerasi specifica.
Spesso la trascrizione dei virus a DNA avviene ad opera della RNA polimerasi cellulare ma alcuni virus possono anche portarsi una RNA polimerasi specifica (poxvirus).

Trascrizione e replicazione
Virus a RNA
Nella maggior parte dei virus a RNA la replicazione avviene nel citoplasma.
Virus a RNA positivo
ss RNA+mRNA (+)funziona direttamente da mRNA
-Replicasi
(RNA polimerasi RNA dipendente)
+
Il filamento + funge direttamente da mRNA che codifica, oltre alle altre proteine necessarie, anche la Replicasi (Rna pol RNA dipendente). Questa inizialmente sintetizza RNA negativi usando come stampo il filamento positivo del genoma, e poi li usa come stampi per produrre ulteriori molecole positive che costituiranno i genomi della progenie virale

Virus a RNA negativo
- ss RNA mRNA (+)
RNA polimerasi RNA dipendenteassociata al virus
-
L’RNA genomico non può funzionare come mRNA né le cellule ospiti hanno polimerasi capaci di trascrivere mRNA usando come stampo RNA.
Come può essere sintetizzato RNA messaggero?
Per superare il problema questi virus portano, nel loro capside, alcune molecole di Trascrittasi (Rna pol RNA dipendente) che inizialmente sintetizza RNA positivo che funge da messaggero, e da questo, in seguito sintetizzera gli RNA negativi che costituiranno il genoma della progenie virale
Trascrizione e replicazione

Virus a RNA a doppio filamento
ds RNA+mRNA (+)
RNA polimerasi RNA dipendenteassociata al virus
- -usa come stampo il filamento
+-
Anche in questo caso l’RNA genomico non può funzionare come RNA messaggero
Alla sintesi del messaggero provvede una Trascrittasi associata al virus e inettata insieme al genoma virale. Il messaggero virale codifica per un’altra RNA pol RNA dipendente che svolge la funzione di Replicasi e sintetizzerà RNA a doppio filamento per le nuove particelle virali.
Trascrittasi
Trascrizione e replicazione

+ ss RNA
mRNA (+)
+-
Trascrittasi inversa
DNA polimerasi RNA dipendenteassociata al virus
+ ss RNA
(virione)
Retrovirus
Sono virus a singolo filamento di RNA + ma si differiscono da tutti gli altri perché sintetizzano i propri mRNA e replicano il proprio genoma passando attraverso un intermedio a DNA
Sono caratteristici anche per il fatto di essere “diploidi”. Infatti, nel loro capside sono presenti sempre due filamenti di RNA. Associati al genoma sono anche la Trascrittasi inversa (DNA polimerasi RNA dipendente) e tRNA di origine virale che svolge la funzione di primer per la sinstesi del DNA virale
Trascrizione e replicazione

I geni tardivi dirigono la sintesi delle proteine capsidiche, che si assemblano spontaneamente come avviene per la morfogenesi dei batteriofagi. L’assemblaggio di virus con envelope è generalmente simile a quello dei virus nudi.
Sintesi e assemblaggio
Liberazione dei virioni
I meccanismi di liberazione di virus nudi differiscono da quelli dei virus dotati di envelope.
I virus nudi in genere vengono liberati mediante lisi della cellula ospite.
Nei virus con envelope la formazione dell’envelope è contemporanea alla liberazione e la cellula ospite può continuare a riversare virus all’esterno per un certo periodo di tempo


Inizialmente le proteine codificate dal virus vengono incorporate nella membrana citoplasmatica, poi per gemmazione del nucleocapside si forma l’envelope.

Infezioni acute: sono caratterizzate da una comparsa abbastanza rapida e da una durata ridotta; le cellule infettate generalmente vanno incontro a morte (effetto citopatico) e vengono liberati numerosi virioni. Le infezioni acute sono tipiche dei picornavirus, herpesvirus e adenovirus.
Effetti citopatici 1) inibizione della sintesi di DNA, RNA e proteine2) danneggiamento dei lisosomi con liberazione di
enzimi idrolitici3) alterazione della membrana plasmatica per
inserimento delle proteine virali (attacco del sistema immunitario)
4) alta concentrazione delle proteine virali5) rotture cromosomiche

Infezioni persistenti o croniche: possono durare anche molti anni. I virus possono replicarsi più lentamente senza causare una sintomatologia clinica. Sono presenti anticorpi diretti contro il virus infettante.Il virus dell’epatite B, il virus di Epstein-Barr e il virus del morbillo possono dare infezioni persistenti.
Infezioni latenti: dovute a virus che, dopo una iniziale fase moltiplicativa, arrestano il proprio ciclo di sviluppo e diventano quiescenti. La fase di latenza può durare un certo periodo dopo il quale può verificarsi una recudescenza dell’infezione con una manifestazione spesso diversa da quella iniziale.I virus erpetici possono dare infezioni latenti.
L’ herpes simplex di tipo 1 infetta in genere i bambini e rimane quiescente per anni a livello dei gangli del sistema nervoso; periodicamente può attivarsi e dare le caratteristiche lesioni erpetiche (herpes labiale)Il virus della varicella zoster (herpes zoster) provoca la varicella come prima manifestazione; non viene eliminato dal sistema immunitario ma diventa quiescente nei gangli dei nervi spinali. La manifesatzione ricorrente è il “fuoco di Sant’Antonio” per la riattivazione del virus a livello dei nervi intercostali
Recommended