
Nanobiotechnologie 7Einführung
Sichtbarmachung im Nanomaßstab
Rastertunnelmikroskopie
Rasterkraftmikroskopie
Nachweis von Viren mittels Rasterkraftmikroskopie
Das Wiegen einzelner Bakterien und Viruspartikel
Nanopartikel und ihre Anwendung
Nanopartikel für die Markierung
Größenquantisierungs effekt und Farben von Nanokristallen
Nanopartikel für die Verabreichung von Arznei stoffen, DNA oder RNA
Nanopartikel in der Krebstherapie
Zusammenbau von Nanokristallen durch Mikroorganismen
Nanoröhrchen
Antibakterielle Nanoschichten
Nachweis von Viren durch Nanokabel
Ionenkanäle als Nanosensoren
Gentechnische Veränderung von DNA im Nanomaßstab
Mechanische DNA-Nanomaschinen
Kontrollierte Denaturierung von DNA durch Nanopartikel aus Gold
Kontrollierte Veränderung der Proteinstruktur durch DNA
Biomolekulare Motoren
Weiterführende Literatur

200 Nanobiotechnologie7
Einführung
Richard Feynman prognostizierte im Jahr 1959 als erster Wissenschaftler die Zukunft der Nanotechno-logie und äußerte die Vermutung, dass Maschinen und Materialien eines Tages auf atomarer Ebene kon-struiert würden: „Die physikalischen Gesetze spre-chen, soweit ich das beurteilen kann, nicht dagegen, Dinge Atom für Atom bewegen zu können.“
Die Molekularbiologie beschäftigt sich weitge-hend mit der Untersuchung von Mikroorganismen. Ein Mikrometer ist ein Millionstel eines Meters und in etwa die Länge einer Zelle von Escherichia coli, dem für den Genetiker wichtigsten Bakterium. Ein Nanometer ist ein Tausendstel eines Mikrometers (= 10–9 Meter; Abb. 7.1). Die Bezeichnungen mikro- und nano- stammen beide aus dem Griechischen. „mik-ros“ bedeutet „klein“, „nanos“ ist dagegen bildhafter und bedeutet „kleiner alter Mann“ oder „Zwerg“. „pico-“ stammt aus dem Spanischen, wo es eine kleine Menge bezeichnet. Vorsilben für noch gerin-gere Mengen sind in Tabelle 7.1 aufgeführt. Mit die-sen Begriffen lassen sich subatomare Dimensionen beschreiben. Handelt es sich jedoch um Massen und Volumina im Nanomaßstab, dann werden Begriffe wie Femtogramm und Zeptoliter verwendet.
Vor nicht allzu langer Zeit hat die Wissenschaft den Schritt in den Bereich der Nanotechnologie vollzogen. Wie der Name bereits andeutet, ist das Vordringen in diese Thematik eng verknüpft mit der Suche nach neuen praktischen Anwendungen, insbe-sondere in der Elektronik und der Materialforschung,
und weniger mit der Suche nach theoretischem Wis-sen. Zur Nanotechnologie gehört die individuelle Manipulation einzelner Moleküle oder Atome. Das ursprüngliche Ziel war möglicherweise, Materialien mit neuen oder stark verbesserten Eigenschaften zu finden, indem man sie Atom für Atom oder Molekül für Molekül aufbaut. Doch das Spektrum erweiterte sich und die Nanotechnologie beschäftigt sich mitt-lerweile mit Strukturen, die so klein sind, dass ihre Untersuchung und ihre Manipulation bis vor kur-zem nicht möglich waren. Im Nanomaßstab nehmen Quanteneffekte zu und die Materialien verhalten sich häufig ungewöhnlich, verglichen mit ihren Festkör-pereigenschaften.
Die Bestandteile von biologischen Zellen entspre-chen in ihrer Größe in etwa den Materialien, die von der Nanotechnologie untersucht werden. Nano-technologen suchen daher auch im Bereich der Zell-biologie nach nützlichen Strukturen, Prozessen und Informationen. Zelluläre Organellen wie Ribosomen lassen sich als programmierbare „Nanomaschinen“ betrachten. Die Nanotechnologie drängt somit auch in Richtung Molekularbiologie. Ein Großteil der „Nanobiotechnologie“ ist aus der Sicht der Materi-alforschung tatsächlich Molekularbiologie, und sie verwendet eine neue Terminologie.
Alle chemischen Reaktionen finden auf Mo-lekülebene statt. Was die echte Nanotechnologie allerdings davon abgrenzt, ist, dass hier einzelne Moleküle oder Nanostrukturen nach spezifischen Anleitungen zusammengesetzt werden. Ein Ribo-som polymerisiert nicht einfach Aminosäuren zu einer Kette. Es verwendet entsprechend der zur
7.1 GrößenvergleichDie Größe der Objekte reicht von einem Meter bis zu einem Picometer.
1 m0.1 m1 cm1 mm100 μm10 μm1 μm100 pm 1 nm 10 nm 100 nm10 pm1 pm }}}}
Kohlen-stoffatom
Protein T2-Phage
Wasserstoff-atom
Elektron
Elektron Mikroskop
Lichtmikroskop
unbewaffnetes Auge
Bakterien eukary-otische Zelle
kleinesInsekt
großerHund
kary-e Zelle
kleinesInsekt

Sichtbarmachung im Nanomaßstab 201 7
Verfügung stehenden Information spezielle Ami-nosäuren und verbindet sie in einer bestimmten Reihenfolge. Zu den entscheidenden Eigenschaften einer Nanomaschine gehört daher die Fähigkeit, Strukturen nicht nur auf Molekülebene zusammen-zusetzen, sondern dabei auch spezifisch und kon-trolliert vorzugehen.
Das wichtigste praktische Ziel für die Nano-biotechnologie ist die Verwendung von biologi-schen Komponenten für Anwendungen im Nano-maßstab. Einige dieser Anwendungen sind nichtbio-logischer Natur und stammen aus der Elektronik und der EDV, andere sind jedoch für die Biologie oder die Medizin von Bedeutung. In diesem Kapitel soll durch ausgewählte Beispiele gezeigt werden, wie biologi-sche Ansätze einen Beitrag zur Nanobiotechnologie leisten.
Viele Bestandteile biologischer Zellen liegen be-
züglich ihrer Größe im Nanobereich. Mit dem Fort-
schritt der Nanotechnologie ergeben sich viele Ver-
bindungen zur Biotechnologie und zur Gentechnik.
Sichtbarmachung im Nanomaßstab
Um Materialien auf atomarer Ebene manipulieren zu können, muss man einzelne Atome und Moleküle se-hen können. Obwohl sich Moleküle auch mit einem Elektronenmikroskop sichtbar machen lassen, war es die Entwicklung von Rastersondenmikroskopen (RSM) (engl. scanning probe microscopes, SPM), die ein neues Feld für die Nanotechnologie eröffneten. Diese Geräte besitzen eine Sonde, die die zu untersu-chende Oberfläche abtastet.
Alle Rastersondenmikroskope messen bestimmte Eigenschaften des Probenmaterials wie den elektri-schen Widerstand, den Magnetismus, die Tempera-tur oder die Lichtabsorption mithilfe einer sehr fei-nen Spitze, die sehr dicht über der Probenoberfläche positioniert wird. Das Mikroskop führt die Sonde in einem vorgegebenen Raster (Raster-Scan) über die Oberfläche (Abb. 7.2), wobei die gewünschte Eigen-schaft gemessen wird. Die Daten werden als geraster-tes Bild, ähnlich dem auf einem Fernsehbildschirm, dargestellt. Im Gegensatz zu herkömmlichen Mikros-kopen nutzen diese Geräte kein Linsensystem, sodass hier auch nicht die Lichtbrechung die Auflösung
Tabelle 7.1 Vorsilben und Größen
Längeneinheit Meter Beispiele
5,9 Terameter mittlerer Abstand zwi-schen Sonne und Pluto
Terameter 1012
150 Gigameter Abstand der Erde zur Sonne
Gigameter 109
380 Megameter Abstand des Mondes zur Erde
6,3 Megameter Radius der Erde
3,2 Megameter Länge der chinesischen Mauer
Megameter 106
Kilometer 103
30 Meter Länge eines Blauwals
Meter 1 Länge eines großen Hundes
Millimeter 10–3 kleines Insekt
Mikrometer 10–6 Bakterienzelle
500 Nanometer Wellenlänge sichtbaren Lichtes
100 Nanometer Größe eines typischen Virus
3,4 Nanometer eine Windung der DNA-Doppelhelix
Nanometer 10–9 Moleküle
350 Picometer Durchmesser eines Wassermoleküls
260 Picometer Atomabstand in festem Kupfer
77 Picometer Atomradius von Kohlen-stoff (= Auflösungsgrenze eines Rasterkraftmikros-kops aus dem Jahr 2004)
32 Picometer Atomradius von Wasser-stoff
Ångstrøm = 100 Picometer = 10–10 Meter
2,4 Picometer Wellenlänge eines Elektrons
Picometer 10–12
Femtometer 10–15 Radius eines Atomkerns
Attometer 10–18 Radius eines Protons
Zeptometer 10–21
Yoctometer 10–24 Radius eines Neutrinos

202 Nanobiotechnologie7
begrenzt, sondern die Größe der Sonde. Mit einigen dieser Mikroskope lassen sich die Proben nicht nur sichtbar machen, sondern auch verändern.
Das erste Rastersondenmikroskop war das Ras-tertunnelmikroskop (RTM) (engl. scanning tunne-ling microscope, STM), das Gerd Binnig und Heinrich Rohrer von IBM (s. folgender Abschnitt) entwickel-ten, wofür sie im Jahre 1986 den Nobelpreis erhiel-ten. Das RTM schickt Elektronen, also elektrischen Strom, durch die Probe und misst auf diese Weise den elektrischen Widerstand. Das Rasterkraftmikroskop (RKM) (engl. atomic force microscope, AFM) ist in der Biologie besonders nützlich und misst die Kräfte, die zwischen der Sondenspitze und der Probe wirken.
Die Sichtbarmachung einzelner Moleküle oder
Atome ist mithilfe von Rastersondenmikroskopen
möglich.
Rastertunnelmikroskopie
Wird eine Metallspitze sehr dicht an eine leitende Oberfläche herangeführt, dann fließen zwischen Spitze und Oberfläche Elektronen (der sog. Tun-
nelstrom). Die Stärke des Tunnelstromes ist expo-nentiell abhängig von dem Abstand zwischen Spitze und Oberfläche. Oberflächenkonturen lassen sich kartieren, indem ein Regelkreis angeschlossen wird, der dafür sorgt, dass der Tunnelstrom immer kon-stant bleibt, während die Spitze über der Oberflä-che schwebt. Durch Heben und Senken der Spitze beim Abtasten der Probe erhält man Informationen über die Topographie der Oberfläche im atomaren Maßstab (Abb. 7.3).
Mithilfe eines RTM lassen sich Atome auch be-wegen. Im Jahr 1989 führten D. M. Eigler und E. K. Schweizer das wahrscheinlich aufsehenerregendste Experiment der Nanotechnologie durch, indem sie 35 Xenonatome auf einer Nickeloberfläche zum IBM-Logo anordneten. Sie wählten Nickel, da hier die Täler zwischen den Reihen aus Nickelatomen ausreichend tief sind, um die Xenonatome an ihrem Platz zu halten. Die Täler sind also auch flach genug, sodass sich die Xenonatome über die Oberfläche zie-hen lassen. Um die Xenonatome zu bewegen, wurde die RTM-Spitze im imaging mode des Mikroskops über einem Xenonatom platziert. Als nächstes schal-tete man den scanning mode aus und die Spitze wurde abgesenkt, bis der Tunnelstrom um ein Vielfaches angestiegen war (fabrication mode). Das Xenonatom wurde von der RTM-Spitze angezogen und durch eine horizontale Bewegung an seinen neuen Platz
Rücklauf
Abtasten
7.2 Prinzip des Raster-ScansBeim Raster-Scan bewegt sich die
Sonde über der Zielregion hin und
her. Die Sonde tastet nur ab, wenn
sie sich in eine Richtung bewegt
(„Abtasten“). In die andere Richtung
bewegt sie sich schneller und ohne
Kontakt zur Probe („Rücklauf“).
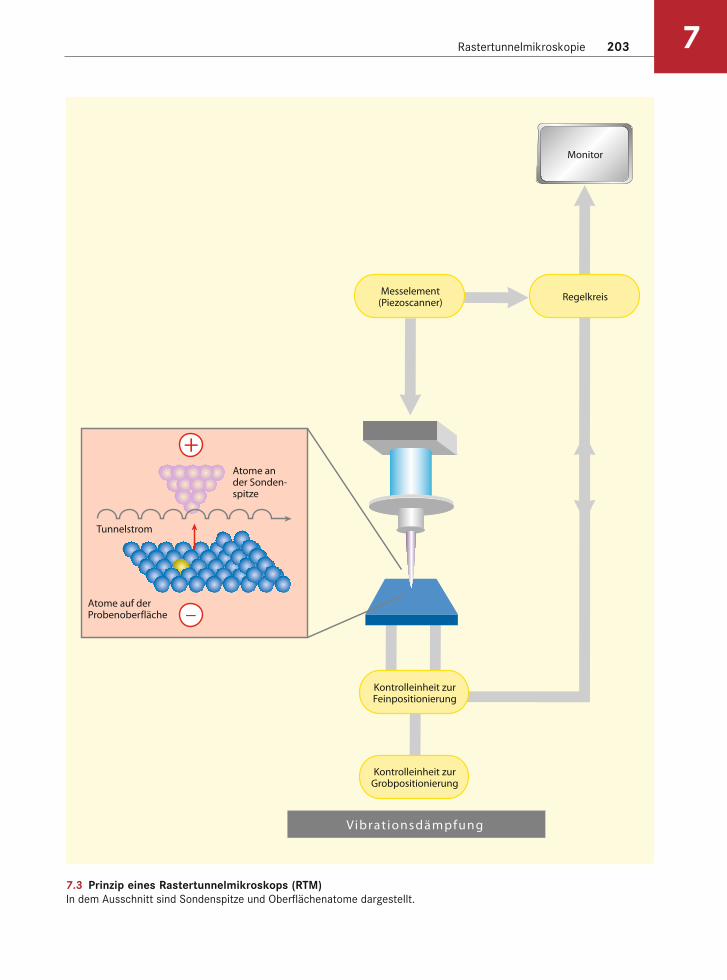
Rastertunnelmikroskopie 203 7
7.3 Prinzip eines Rastertunnelmikroskops (RTM)In dem Ausschnitt sind Sondenspitze und Oberflächenatome dargestellt.
Atome ander Sonden-spitze
Atome auf derProbenoberfläche
Tunnelstrom
+
−
Messelement(Piezoscanner)
Kontrolleinheit zurFeinpositionierung
Kontrolleinheit zurGrobpositionierung
V ibrat ionsdämpfung
Regelkreis
Monitor

204 Nanobiotechnologie7
gebracht. Dort wurde der Tunnelstrom reduziert und das Atom abgelegt. Seitdem wurden viele Diagramme auf diese Weise erstellt. Der Kohlenmonoxidmann ist in Abbildung 7.4 dargestellt.
Aus biologischer Sicht hat das RTM den Nach-teil, dass sich nur elektrisch leitende Oberflächen untersuchen lassen, in der Praxis handelt es sich dabei in der Regel um Metallschichten. Ein Ras-terkraftmikroskop (s. folgender Abschnitt) hat den Vorteil, dass das Material nicht leitfähig sein muss. Es findet daher in der Biologie ein breites Anwen-dungsgebiet.
Das Rastertunnelmikroskop lässt sich einsetzen,
um einzelne Atome einer elektrisch leitenden
Oberfläche sichtbar zu machen oder sie zu be-
wegen.
Rasterkraftmikroskopie
Für die Sichtbarmachung im Nanomaßstab nutzt man häufig ein Rasterkraftmikroskop (RKM). Wie der Name schon sagt, beruht das Verfahren auf der Messung einer mechanischen Wechselwirkung und nicht auf der Messung fließender Partikel, seien es Photonen wie bei der Lichtmikroskopie oder Elektro-nen wie bei der Elektronenmikroskopie.
Physiker vergleichen die Arbeitsweise eines RKM häufig mit der eines altmodischen Plattenspielers, bei dem eine Nadel über die Oberfläche der Schall-platte kratzt. Für einen Biologen ist der Unterschied zwischen einem Lichtmikroskop und einem RKM ähnlich dem Unterschied zwischen dem Lesen eines Textes mit den Augen und dem Ertasten der Blinden-schrift mit den Fingerspitzen.
Das Rasterkraftmikroskop wurde im Jahr 1985 von Gerd Binnig, Calvin Quate und Christof Gerber entwickelt. Bei dem RKM gleitet eine feine Spitze, die an einem Träger angebracht ist, über die Ober-fläche einer Probe. Durch die zwischen Spitze und Probe auftretenden Kräfte biegt sich ein Federbalken (engl. cantilever) und diese Biegung wird dokumen-tiert. Die Spitze wird in einem vorgegebenen Raster über die Oberfläche geführt und das entstehende Bild der Oberflächentopographie wird auf einem Monitor dargestellt.
Während des Abtastens werden die Spitze oder die Probe durch ein sehr genau arbeitendes Stellele-ment aus piezoelektrischer Keramik bewegt. (Dieses Material verändert die Form als Reaktion auf eine angelegte Spannung.) Das Stellelement hat in der Regel die Form eines Röhrchens, das sich in alle drei räumlichen Dimensionen im Sub-Ångstrøm-Bereich zu bewegen vermag.
Die RKM-Sonde befindet sich am Ende eines biegsamen Federbalkens. Auf diesen Federbalken ist ein Laserstrahl gerichtet, der abgelenkt wird, wenn sich der Federbalken aufgrund der auf die Spitze wirkenden Kraft biegt. Die Biegung lässt sich dann anhand der Ablenkung des Laserstrahls messen, wie in Abbildung 7.5 dargestellt ist. Der Laserstrahl wird auf eine geteilte Photodiode gelenkt und die Unter-schiede zwischen dem A- und dem B-Signal sind ein Maß für die Biegung des Federbalkens. Kleine Verän-derungen sind dabei proportional zur aufgewendeten Kraft. Die Kraft zwischen Spitze und Probe lässt sich somit bestimmen.
Der Abstand zwischen Spitze und Probe wird so eingestellt, dass er im Bereich der Abstoßung
7.4 Der Kohlenmonoxidmann von ZeppenfeldDie Atome wurden mithilfe eines RTM angeordnet. Der
Mann besteht aus Kohlenmonoxid auf einer Platinober-
fläche. Mit freundlicher Genehmigung von International
Business Machines Corporation © 1995, IBM.

Nachweis von Viren mittels Rasterkraftmikroskopie 205 7
der intramolekularen Kraftkurve liegt. Das bedeutet, dass die RKM-Sonde durch ihre molekulare Wech-selwirkung von der zu untersuchenden Oberfläche abgestoßen wird. Durch die Abstoßung lässt sich die Oberflächentopographie messen und in einer Abbildung darstellen, in der Farben die relativen Höhen anzeigen. Es ist möglich, die Oberfläche nach ihren topographischen Gegebenheiten abzusuchen, anschließend die RKM-Sonde anzuheben und die Oberfläche ein weiteres Mal nach elektrostatischen oder magnetischen Kräften abzutasten. Diese Ergeb-nisse können ebenfalls graphisch dargestellt werden und lassen sich mit den topographischen Gegeben-heiten vergleichen.
Wie bei dem RTM ist es auch mit dem RKM möglich, einzelne Atome zu bewegen, obwohl dies nur einmal im Jahr 2003 gelang. Forscher der Universität von Osaka, Japan, entfernten ein ein-zelnes Siliziumatom aus einer Oberfläche und er-setzten es.
Durch den Einsatz eines RKM ist es möglich, polymere biologische Moleküle wie DNA oder Cel-lulose sichtbar zu machen und sogar einzelne Mo-nomere (bei hoher Auflösung sogar Atome) zu be-trachten.
Das Rasterkraftmikroskop kann Atome oder Mole-
küle detektieren, indem eine Oberfläche hinsicht-
lich ihrer Topographie oder ihrer elektromagneti-
schen Eigenschaften abgetastet wird.
Nachweis von Viren mittels Rasterkraftmikroskopie
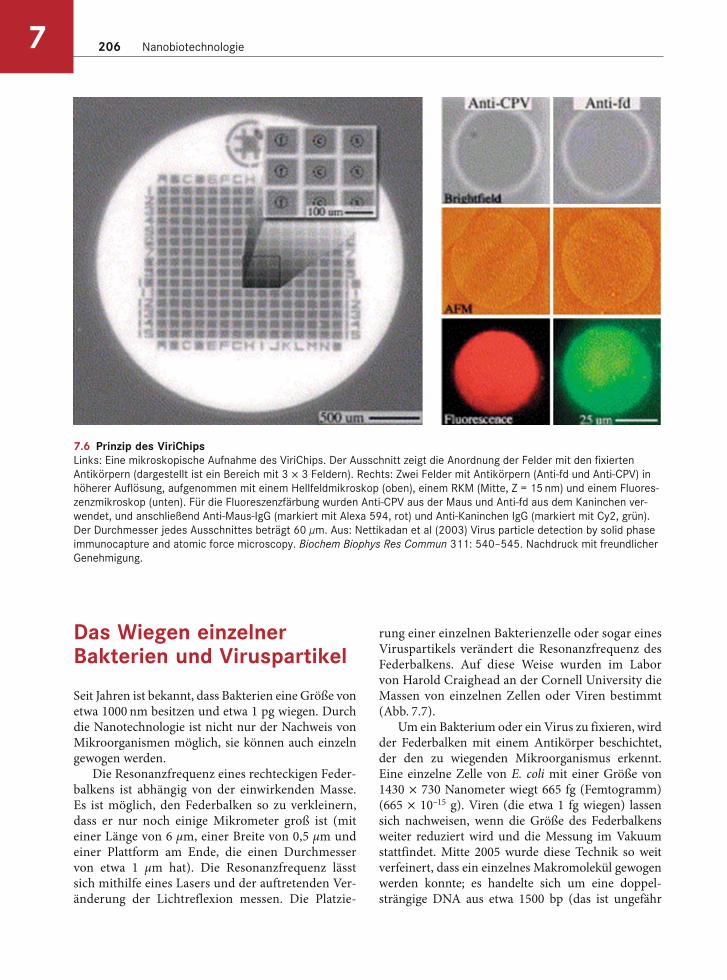
Ein Rasterkraftmikroskop (RKM) lässt sich einsetzen, um die Anwesenheit von Viruspartikeln nachzuwei-sen, dafür ist allerdings ein sogenannter „ViriChip“ notwendig. Dabei handelt es sich um einen Silizium-chip, der mit Antikörpern gegen das interessierende Virus beschichtet ist. In der Praxis ist der Chip in Be-reiche aufgeteilt, die mit verschiedenen Antikörpern beschichtet werden, damit viele Viren gleichzeitig nachgewiesen werden können. Von der zu untersu-chenden Probe wird ein Mikroliter auf den Chip auf-getragen und jedes Virus, das von den Antikörpern auf dem Chip erkannt wird, wird auf dem Chip ge-bunden. Die Chipoberfläche wird nun mithilfe eines RKM abgetastet, um die Anwesenheit von Virusparti-keln zu prüfen, die sich durch den Ort ihrer Bindung auf dem Chip identifizieren lassen (Abb. 7.6).
Der Chip wurde bereits erfolgreich mit einer Viel-zahl von Viren, einschließlich humanpathogener Vi-ren, getestet und wird zurzeit noch weiterentwickelt. Grundsätzlich sollte es möglich sein, Tausende von verschiedenen Viren mithilfe eines einzigen Chips nachzuweisen. Nur ein Tropfen Blut eines Patienten könnte dann auf viele Viren getestet werden.
Die Rasterkraftmikroskopie vermag einzelne Viren
nachzuweisen und zu identifizieren.
Laserstrahl Position desDetektors
Sondenspitze
7.5 Das Rasterkraftmikroskop (RKM)Die Auslenkung der Sondenspitze
durch die Probenoberfläche wird
mithilfe eines Lasers gemessen.
206 Nanobiotechnologie7
Das Wiegen einzelner Bakterien und Viruspartikel
Seit Jahren ist bekannt, dass Bakterien eine Größe von etwa 1000 nm besitzen und etwa 1 pg wiegen. Durch die Nanotechnologie ist nicht nur der Nachweis von Mikroorganismen möglich, sie können auch einzeln gewogen werden.
Die Resonanzfrequenz eines rechteckigen Feder-balkens ist abhängig von der einwirkenden Masse. Es ist möglich, den Federbalken so zu verkleinern, dass er nur noch einige Mikrometer groß ist (mit einer Länge von 6 μm, einer Breite von 0,5 μm und einer Plattform am Ende, die einen Durchmesser von etwa 1 μm hat). Die Resonanzfrequenz lässt sich mithilfe eines Lasers und der auftretenden Ver-änderung der Lichtreflexion messen. Die Platzie-
rung einer einzelnen Bakterienzelle oder sogar eines Viruspartikels verändert die Resonanzfrequenz des Federbalkens. Auf diese Weise wurden im Labor von Harold Craighead an der Cornell University die Massen von einzelnen Zellen oder Viren bestimmt (Abb. 7.7).
Um ein Bakterium oder ein Virus zu fixieren, wird der Federbalken mit einem Antikörper beschichtet, der den zu wiegenden Mikroorganismus erkennt. Eine einzelne Zelle von E. coli mit einer Größe von 1430 × 730 Nanometer wiegt 665 fg (Femtogramm) (665 × 10–15 g). Viren (die etwa 1 fg wiegen) lassen sich nachweisen, wenn die Größe des Federbalkens weiter reduziert wird und die Messung im Vakuum stattfindet. Mitte 2005 wurde diese Technik so weit verfeinert, dass ein einzelnes Makromolekül gewogen werden konnte; es handelte sich um eine doppel-strängige DNA aus etwa 1500 bp (das ist ungefähr
7.6 Prinzip des ViriChipsLinks: Eine mikroskopische Aufnahme des ViriChips. Der Ausschnitt zeigt die Anordnung der Felder mit den fixierten
Antikörpern (dargestellt ist ein Bereich mit 3 × 3 Feldern). Rechts: Zwei Felder mit Antikörpern (Anti-fd und Anti-CPV) in
höherer Auflösung, aufgenommen mit einem Hellfeldmikroskop (oben), einem RKM (Mitte, Z = 15 nm) und einem Fluores-
zenzmikroskop (unten). Für die Fluoreszenzfärbung wurden Anti-CPV aus der Maus und Anti-fd aus dem Kaninchen ver-
wendet, und anschließend Anti-Maus-IgG (markiert mit Alexa 594, rot) und Anti-Kaninchen IgG (markiert mit Cy2, grün).
Der Durchmesser jedes Ausschnittes beträgt 60 μm. Aus: Nettikadan et al (2003) Virus particle detection by solid phase
immunocapture and atomic force microscopy. Biochem Biophys Res Commun 311: 540–545. Nachdruck mit freundlicher
Genehmigung.

Nanopartikel und ihre Anwendung 207 7
die Größe einer typischen codierenden Sequenz). Die Weiterentwicklung des Verfahrens dürfte die Mes-sung kleiner Proteine oder anderer Moleküle mög-lich machen, deren Gewicht im Zeptogrammbereich (10–21 g) liegt.
Die Messung der Resonanzfrequenz eines Feder-
balkens mithilfe eines Lasers erlaubt es, einzelne
Bakterien oder Viren zu wiegen.
Nanopartikel und ihre AnwendungFortschritte der Wissenschaft, das unglaublich Kleine sichtbar zu machen und zu messen, waren der Ur-sprung der Nanotechnologie. Es folgte der Aufbau von Strukturen im Nanomaßstab. Mittlerweile wer-den einfache Nanostrukturen für eine Vielzahl analy-tischer Zwecke eingesetzt und eine zweite Generation solcher Strukturen für den Einsatz im klinischen Bereich befindet sich in der Entwicklung.
Wie ihr Name schon andeutet sind Nanopartikel kleiner als ein Mikrometer – in der Praxis zwischen 100 nm und 5 nm. Die Partikel sind in der Regel ku-gelförmig, doch auch Stäbchen, Platten und andere Formen werden eingesetzt. Sie können gefüllt oder auch hohl sein und aus einer Vielzahl von Materia-lien bestehen, die häufig in abgegrenzten Schichten mit unterschiedlicher Funktion übereinander gelagert sind. Typisch sind ein zentraler funktioneller Kern, eine Schutzschicht und eine äußere Schicht, die mit der biologischen Umgebung in Wechselwirkung tritt.
Der zentrale funktionelle Kern hat in der Regel nützliche optische oder magnetische Eigenschaften. Die bekannteste ist die Fluoreszenz. Die Schutz-schicht schützt die funktionelle Schicht vor den schädigenden Einflüssen von Luft, Wasser oder Zell-bestandteilen – sie schützt die Zelle auch vor allen toxischen Eigenschaften der funktionellen Schicht. Die äußere(n) Schicht(en) macht die Nanopartikel „biokompatibel“. Zu den erforderlichen Eigenschaf-ten gehören in der Regel die Wasserlöslichkeit und die spezifische Erkennung. Für biologische Zwecke werden Nanopartikel häufig durch eine hydrophile äußere Schicht wasserlöslich gemacht. Außerdem müssen auf der Außenseite chemische Gruppen für eine spezifische Anheftung an andere Moleküle oder Strukturen vorhanden sein (Abb. 7.8).
Die Verwendungsmöglichkeiten von Nanoparti-keln in der Biologie sind vielfältig:a) Fluoreszenzmarkierung und optische Codierung,b) Nachweis pathogener Mikroorganismen und/
oder spezifischer Proteine,c) Reinigung und Manipulation biologischer Mole-
küle,d) Gezielte Verabreichung von Arzneimitteln und/
oder Genen,e) Zerstörung von Tumoren auf chemischem oder
thermischem Weg,f) Verstärkung des Kontrastes bei der Magnetreso-
nanztomographie (MRT).
a
7.7 Das Wiegen eines einzelnen Bakteriumsa Rasterelektronenmikroskopische Aufnahme eines Feder-
balkens mit einer Länge von 6 μm, einer Breite von 0,5 μm
und einer Plattform mit einem Durchmesser von 1 × 1 μm.
Der Größenbalken entspricht 2 μm. Ilic B et al (2004) Virus
detection using nanoelectromechanical divices. Appl Phys
Lett 95: 2604–2607. © 2004. Nachdruck mit freundlicher
Genehmigung des American Institute of Physics. b Raster-
elektronenmikroskopische Aufnahme einer einzelnen E.
coli-Zelle, die durch einen Antikörper auf dem Federbalken
fixiert wird. Mit freundlicher Genehmigung der Graighead-
Gruppe, Cornell University. Ilic B et al (2001) Single cell
detection with micromechanical oscillators. J Vac Sci Tech-
nol B. 19: 2825–2828. © 2001. Nachdruck mit freundli-
cher Genehmigung des American Institute of Physics.
b

208 Nanobiotechnologie7
Nanopartikel werden mittlerweile bei vielen biologi-
schen Verfahren eingesetzt. Dazu gehören sowohl
analytische als auch klinische Anwendungen.
Nanopartikel für die Markierung
Betrachten wir das lumineszierende CdSe-Nano-stäbchen als Beispiel für ein Nanopartikel, das für Markierungen verwendet wird (Abb. 7.9). Diese Na-nostäbchen (stabförmige Nanokristalle) lassen sich in der Molekularbiologie als Fluoreszenzmarker ein-setzen, weil sie UV-Licht mit einer Wellenlänge von 550 nm absorbieren und Licht mit einer Wellenlänge von 590 nm emittieren. Sie wurden von Thomas Nann aus Freiberg, Deutschland, entwickelt.
Die Nanostäbchen sind etwa 3 nm breit und 10 bis 20 nm lang. Sie haben einen Kern aus lumines-zierendem Cadmiumselenid (CdSe), der von einer Schicht aus ZnS (Zinksulfid, Wurzit) umgeben ist, die den Kern vor Oxidation schützt. Außen befindet sich eine Schicht aus Siliziumdioxid (Silica), über die Phosphonate oder Amine an das Nanostäb-
chen gekoppelt werden können. Diese hydrophilen Gruppen sorgen für die Wasserlöslichkeit der Stäb-chen. Außerdem können die Nanostäbchen über diese äußeren chemischen Gruppen an Proteine binden.
Das Gerüst einer eukaryotischen Zelle ist aus zy-lindrischen Proteinstrukturen aufgebaut, die als Mi-krotubuli bezeichnet werden. Diese werden häufig in ihre Monomere (Tubulin) zerlegt und an einem anderen Ort wieder aufgebaut. Mithilfe von Nano-stäbchen lässt sich diese Umstrukturierung verfol-gen, indem man sie mit Tubulinmonomeren ver-knüpft. Die Zugabe von Guanosintriphosphat (GTP) stimuliert den Zusammenbau der Mikrotubuli, und durch die fluoreszierenden Nanostäbchen lässt sich beobachten, wie sie sich zu linearen Strukturen zu-sammenlagern.
Warum verwendet man hier eine komplexe viel-schichtige Nanostruktur statt eines einfachen Fluo-reszenzfarbstoffes?a) Obwohl die Nanokristalle einen engen Bereich
der Fluoreszenz haben, absorbieren sie doch ein relativ breites Spektrum an Licht (im Gegensatz zu dem engen Absorptionsspektrum eines typi-schen Fluoreszenzfarbstoffes). Somit bleichen sie während der Anregung nicht aus und lassen sich über einen langen Zeitraum bestrahlen und auch verfolgen.
physikalisch aktive, innereSchichten
Schutzschicht
hydrophile Schicht
Linker
anhaftende Moleküle
7.8 Typischer schichtweiser Auf-bau eines NanopartikelsDer physikalisch aktive Kern wird von
mehreren Schichten umgeben. Häu-
fig werden chemische Gruppen mit
der Außenseite verknüpft, damit das
Nanopartikel an andere biologische
Moleküle binden kann.

Nanopartikel für die Markierung 209 7
b) Das von Nanokristallen emittierte Licht ist sehr hell – es ist das Produkt aus dem molaren Ex-tinktionskoeffizienten und der Quantenausbeute. (Der molare Extinktionskoeffizient ist die Ab-sorption einer einmolaren Lösung eines reinen gelösten Stoffes bei gegebener Wellenlänge; je hö-her er ist, umso mehr Licht wird absorbiert. Die Quantenausbeute ist der Quotient aus absorbier-ten Photonen und den während der Fluoreszenz emittierten Photonen.)
c) Das Emissionsmaximum eines Nanokristalls ist abhängig von seiner Größe. Indem man einen Kristall einer bestimmten Größe herstellt, lässt sich jede beliebige Wellenlänge einstellen (s. un-ten).
Nanopartikel lassen sich ebenso spezifisch zu Gewe-ben wie Krebszellen dirigieren, indem man entspre-chende Antikörper oder Rezeptorproteine mit der Oberfläche der Partikel verknüpft. Fluoreszierende Nanopartikel werden häufig als Quantenpunkte (engl. quantum dots) bezeichnet und sind mittlerweile für viele Markierungen in der Biologie kommerziell er-hältlich. Auch Fluoreszenzfarbstoffe können mit an-deren Molekülen gekoppelt werden, doch sind Nano-partikel vielseitiger. Mit Quantenpunkten lassen sich DNA-Moleküle wie auch Proteine markieren. Die Kopplung von Quantenpunkten mit PCR-Primern ergibt fluoreszenzmarkierte PCR-Produkte – ein Ver-fahren, das als quantum dot-PCR bezeichnet wird.
Um den Kontrast bei der Magnetresonanztomo-graphie (MRT) zu verstärken, wurden viele Materi-alien eingesetzt. Zunehmend gehören dazu auch Na-
nopartikel, die verschiedene Materialien enthalten. So sind z.B. superparamagnetische Nanopartikel aus Eisenoxid, sog. SPIO-Nanopartikel (engl. superpa-ramagnetic iron oxide nanoparticles), ein gutes Kon-trastmittel für ein MRT. Ihre magnetischen Eigen-schaften variieren mit der Partikelgröße. Größere Partikel mit einer Größe über 300 nm werden für den Darm, die Leber und die Milz eingesetzt. Kleinere Partikel mit einer Größe zwischen 20 und 40 nm sind genauer und besser für die Detektion von frühen Tumorstadien der Lymphknoten geeignet als her-kömmliche Materialien.
Fluoreszierende Nanopartikel haben bei Markierun-
gen in der Biologie ein breites Anwendungsspek-
trum. Sie sind stabiler als herkömmliche fluoreszie-
rende Farbstoffe und häufig auch heller.
Amin AminPhos-
phonatPhos-
phonat
Siliziumdioxid
10–20 nm
3 nmZinksulfid
Cadmiumselenid
hydrophile Gruppen, die mit der Oberfläche verknüpft sind
7.9 CdSe-NanostäbchenLumineszierende CdSe-Nanostäb-
chen sind von Schutzschichten aus
Zinksulfid und Siliziumdioxid um-
geben. Über hydrophile chemische
Gruppen auf der Oberfläche lassen
sich die Stäbchen mit Proteinen
oder anderen biologischen Molekü-
len verknüpfen.
Terminologie im TrendNanopartikel werden mit einer Vielzahl von „Nano-
begriffen“ bezeichnet wie Nanostäbchen, Nanokris-
talle, Nanokapseln, Nanoröhrchen, Nanokabel usw.
abhängig von ihrer Form und Struktur. Hinzu kommen
die Quantenpunkte, hinter denen sich fluoreszierende
Nanokristalle verbergen, die ausreichend klein sind,
um einen Quanteneinschluss (engl. quantum confine-
ment) zu zeigen – sie können für Markierungen in der
Biologie verwendet werden.
Exkurs 7.1

210 Nanobiotechnologie7
Größenquantisierungs-effekt und Farben von Nanokristallen
Werden Materialien in ausreichend kleine Fragmente zerteilt, beginnen Quanteneffekte ihre physikalischen Eigenschaften zu beeinflussen. Die fluoreszierenden Nanopartikel sind Halbleiter, die ausreichend klein sind, um solche Quanteneffekte zu zeigen.
Halbleiter sind Substanzen, die nur unter be-stimmten Bedingungen elektrischen Strom leiten. Bei Halbleitern des N-Typs besteht der Strom (wie bei normalen elektrischen Kabeln) aus negativ geladenen Elektronen. Bei Halbleitern des P-Typs besteht der Strom jedoch aus Löchern. Ein Loch entsteht, wenn einem Atom ein Elektron fehlt. Obwohl es im phy-sikalischen Sinne keine Partikel sind, vermögen die Löcher von einem Atom zum nächsten zu wandern und dienen als Ladungsträger. Elektronen und Lö-cher können rekombinieren und sich so auslöschen, ein Prozess, bei dem Energie frei wird. Umgekehrt kann die Energie, die von verschiedenen Halbleitern absorbiert wird, ein Elektron von einem Atom lö-sen; gleichzeitig entsteht ein Loch, und Elektron und Loch können sich anschließend in unterschiedliche Richtungen bewegen.
Markierungen mit Nanopartikeln lassen sich un-ter Verwendung verschiedener Emissionswellenlän-gen herstellen, die den UV-Bereich, das sichtbare Spektrum und den Nah-Infrarot-Bereich abdecken. Die Emissionswellenlängen variieren abhängig vom Material des Halbleiters. Durch den Größenquanti-sierungseffekt (engl. quantum size effect, Abb. 7.10) ist es möglich, dass derselbe Halbleiter, abhängig von der Größe des Nanopartikels, verschiedene Wellen-längen emittiert. Je kleiner die Nanopartikel, umso geringer ist die abgestrahlte Wellenlänge (d.h. umso höher ist die Energie).
Fluoreszierende Nanopartikel sind eine Art mini-aturisierte, lichtemittierende Dioden (LEDs). Dabei handelt es sich um Halbleiter, die Energie absorbie-ren (entweder elektrisch oder in Form von Licht), wodurch Paare aus Elektronen und Löchern ent-stehen. Rekombinieren die Elektronen wieder mit den Löchern, wird Licht emittiert. Bei Festkörpern hängen die Energie und damit die Wellenlänge des emittierten Lichts von der chemischen Zusammen-setzung des Halbleiters ab. Im Nanomaßstab kom-men allerdings Quanteneffekte zum Tragen. Ist die physikalische Größe des Halbleiters geringer als der natürliche Radius (der Bohr-Radius) des Elektron-Loch-Paares, dann muss zusätzlich Energie aufge-bracht werden, um das Paar einzuschließen. Dieser Vorgang wird als Quanteneinschluss (engl. quantum
7.10 GrößenquantisierungseffektNanokristalle unterschiedlicher
Größe absorbieren UV-Licht und
emittieren Energie. Die Wellenlänge
des emittierten Lichtes hängt von
der Größe des Nanokristalls ab. Je
kleiner der Kristall, umso energie-
reicher ist das emittierte Licht. Aus:
Riegler und Nann (2004) Application
of luminescent nanocrystals as la-
bels for biological molecules. Anal
Bioanal Chem 379 (7–8): 913–919.

Nanopartikel in der Krebstherapie 211 7
confinement) bezeichnet und tritt bei Nanokristallen mit einer Größe von 20 nm oder weniger auf. Je klei-ner der Halbleiterkristall, umso mehr Energie ist er-forderlich und umso energiereicher ist das emittierte Licht (d.h. umso kürzer ist seine Wellenlänge).
Die emittierte Wellenlänge eines fluoreszierenden
Nanopartikels hängt von seiner Größe ab und lässt
sich daher leicht manipulieren.
Nanopartikel für die Verabreichung von Arznei-stoffen, DNA oder RNA
Da sich Nanopartikel spezifisch zu Geweben diri-gieren lassen, können sie eingesetzt werden, um bio-logisch aktive Moleküle wie Arzneistoffe und gen-technische Konstrukte zu ihrem Bestimmungsort zu bringen.
Große polymere Moleküle wie DNA können selbst zu Nanopartikeln mit einer Größe von 50 bis 200 nm verdichtet werden. Hinzu kommt die Ad-dition eines positiv geladenen Moleküls (z.B. eines kationischen Lipids oder Polylysin), um die negative Ladung der Phosphatgruppen des Zucker-Phosphat-Rückgrats zu neutralisieren. Andere Moleküle kön-nen hinzugefügt werden und erhöhen die Selektivität für bestimmte Zellen oder Gewebe.
Alternativ kann man hohle Nanopartikel (Nano-kapseln) einsetzen, um kleinere Moleküle zu trans-portieren. Solche Nanokapseln müssen aus biokom-patiblen Materialien wie Polyethylenimin (PEI) oder Chitosan bestehen. Letzteres wird zurzeit häufig ein-gesetzt, es ist natürlicher Herkunft und biologisch abbaubar. Chitin ist ein β-1,4-verknüpftes Polymer aus N-Acetyl-D-glucosamin, das in Zellwänden von Insekten und Pilzen vorkommt. Es ist nach der Cel-lulose das am häufigsten vorkommende Biopolymer. Aus Chitin lässt sich Chitosan herstellen, indem man durch Alkalibehandlung alle Acetylgruppen des Chi-tins entfernt.
Ein interessanter Ansatz verbindet zwei zurzeit aktuelle Technologien: die Nutzung von Nanokapseln für den Transport von siRNA (kurze interferierende RNA). Die gezielte Verabreichung von siRNA löst RNA-Interferenz aus, die zur Zerstörung einer Ziel-mRNA führt (s. Kap. 5). Die siRNA kann mRNA von
Genen zum Ziel haben, die hauptsächlich in Krebs-zellen exprimiert werden oder die für bestimmte Viren charakteristisch sind.
Hohle Nanopartikel lassen sich einsetzen, um DNA,
RNA oder Protein gezielt an ihren Bestimmungsort
zu bringen.
Nanopartikel in der Krebstherapie
Tumorzellen können durch toxische Chemikalien oder durch lokale Hitzeeinwirkung zerstört werden. In beiden Fällen ist das Ziel, die wirksamen Agenzien gezielt zu den Krebszellen zu bringen, um gesundes Gewebe zu schützen. Werden toxische Substanzen eingesetzt, muss das Reagens nicht nur spezifisch zum Bestimmungsort transportiert werden, sondern darf auch nicht in das umliegende Gewebe diffundie-ren. Beides lässt sich realisieren, indem man hohle Nanopartikel für den Transport einsetzt. Nanoparti-kel können Tumore zum Ziel haben, indem man sie mit spezifischen Rezeptoren oder reaktiven Gruppen koppelt. Diese werden so gewählt, dass sie Proteine erkennen, die ausschließlich oder hauptsächlich auf der Oberfläche der Krebszellen vorkommen. Ziel ist eine orale Verabreichung dieser Nanopartikel.
Problematisch ist die Diffusion, doch sie lässt sich auf ein gewisses Maß beschränken, indem Na-nopartikel konstruiert werden, die den Wirkstoff nur langsam freisetzen. Eine sinnvolle Alternative ist die Herstellung eines toxischen Wirkstoffes in-nerhalb des Nanopartikels, nachdem dieser in die Krebszelle geschleust wurde. Bei der photodyna-mischen Krebstherapie entsteht Singulettsauerstoff, indem ein Laser einen photosensitiven Farbstoff an-geregt hat. Der Singulettsauerstoff ist hoch reaktiv und zerstört durch die Oxidation von Lipiden vor allem biologische Membranen. Nach der Diffusion aus dem Nanopartikel heraus, reagiert der toxische Sauerstoff so schnell, dass er die Krebszelle nicht verlässt (Abb. 7.11).
Nanopartikel lassen sich auch einsetzen, um Krebszellen durch lokale Hitzeeinwirkung abzutöten. Eine Möglichkeit ist die Verwendung eines magneti-schen Kerns. Ein wechselndes Magnetfeld liefert die Energie und heizt die Nanopartikel auf eine Tempe-

212 Nanobiotechnologie7
ratur auf, die für Säugerzellen tödlich ist. Eine andere Möglichkeit ist der Einsatz von Nanokapseln, deren Kern (häufig aus Siliziumdioxid) von einer dünnen Metallschicht (z.B. aus Gold) umgeben ist. Durch Veränderung des Kerndurchmessers und der Dicke der Metallschicht ist es möglich, solche Nanopartikel so einzustellen, dass sie aus jedem Bereich des Spek-trums, angefangen bei UV-Licht über den sichtbaren Bereich bis hin zum Infrarot, Licht absorbieren. Da lebende Gewebe am wenigsten im Nah-Infrarot-Be-reich absorbieren, werden die Nanopartikel so konst-ruiert, dass sie Strahlungsenergie aus diesem Bereich des Spektrums absorbieren. Eine Bestrahlung mit den entsprechenden Wellenlängen heizt auf diese Weise das umgebende Gewebe auf.
Nanopartikel lassen sich einsetzen, um Krebszellen
durch lokale Hitzeeinwirkung oder Freisetzung eines
toxischen Produkts abzutöten.
Zusammenbau von Nanokristallen durch Mikroorganismen
Seit vielen Jahren ist bekannt, dass Bakterien eine Vielzahl von metallischen Elementen anreichern und sie, in der Regel durch Oxidation oder Reduktion, chemisch modifizieren. So können Bakterien z.B. Se-
len- oder Telluranionen anreichern und zu elemen-tarem Selen oder Tellur reduzieren, die entweder als Präzipitat auf der Zelloberfläche oder im Zellinneren abgelagert werden. Bestimmte Arten des Bakteriums Pseudomonas, die in metallkontaminierten Böden vorkommen, und auch der Pilz Verticillium können Nanokristalle aus Silber bilden.
Wie man erst kürzlich entdeckte, fallen Partikel aus Cadmiumsulfid mit einer Größe von 2 bis 5 nm aus, wenn Escherichia coli in einem cadmiumchlorid- und natriumsulfidhaltigen Medium kultiviert wird. Mit anderen Worten: Bakterien „synthetisieren“ auf biologischem Wege Halbleiternanokristalle.
Noch raffinierter ist die Anwendung eines Pha-gen-Displays für die Selektion von Peptiden, die Halbleiternanokabel aufbauen können. Wie in Ka-pitel 9 beschrieben, ist das Phagen-Display eine Methode, mit deren Hilfe sich Peptide selektieren lassen, die an bestimmte, ausgewählte Zielmoleküle binden. DNA-Abschnitte, die eine Bibliothek aus Peptidsequenzen codieren, werden mit einem Gen für ein Hüllprotein eines Bakteriophagen fusioniert. Die zusätzliche Sequenz wird entweder an den C- oder N-Terminus des Proteins geheftet, wo sie die normale Funktion des Hüllproteins nicht stört. Wird das Hybridprotein in das Phagencapsid eingebaut, dann werden die entsprechenden Peptide auf der Außenseite des Phagenpartikels präsentiert. Die Pha-genbibliothek wird anschließend auf die Bindung an ein Zielmolekül hin untersucht und die bindenden Phagen weiter analysiert.
Phagen-Display-Bibliotheken wurden auch mit dem Ziel durchsucht, Peptide zu finden, die an ZnS-
angeregter Farbstoffaggregiert
organisch veränderteNanopartikel
aus Siliziumdioxid
FRET zurlichtempfindlichen
Substanz
Zerstörungder Membran
3O2
1O2
Laser im Nah-Infrarot-Bereich7.11 Nanopartikel für die Frei-setzung von SingulettsauerstoffDer Laser, der Licht im Nah-
Infrarot-Bereich abstrahlt, regt den
Farbstoff an, der Teil des Nanopar-
tikels ist. Die Energieübertragung
auf die leichtempfindliche Subs-
tanz mittels FRET (Fluoreszenz-Re-
sonanz-Energie-Transfer) wandelt
normalen (Triplett-)Sauerstoff in
Singulettsauerstoff um.

Nanoröhrchen 213 7
oder CdS-Nanokristalle binden. Für die Fusion mit dem Peptid verwendete man das Protein VIII aus dem Bakteriophagen M13. ZnS wurde durch das Pep-tid VISNHAGSSRRL gebunden und CdS von dem Peptid SLTPLTTSHLRS. Da das Capsid des Bakte-riophagen mehrere Kopien des Hüllproteins enthält, werden auch viele Kopien des Peptids präsentiert. Auf der Phagenoberfläche ordnen sich also mehrere Nanokristalle an und da M13 ein filamentöser Phage ist, ist das Ergebnis ein Halbleiterkabel (Abb. 7.12).
Nanokristalle und Nanokabel können auch von
nichtmodifizierten Bakterien oder durch den Ein-
satz raffinierter Phagen-Display-Verfahren zusam-
mengesetzt werden.
Nanoröhrchen
Nanoröhrchen aus Kohlenstoff sind Zylinder, die aus reinem Kohlenstoff bestehen und einen Durchmes-ser zwischen 1 bis 50 nm und eine Länge von bis zu 10 μm haben. Reiner, elementarer Kohlenstoff liegt in Diamanten oder als Graphit vor. Im Diamanten ist jedes C-Atom kovalent mit vier anderen Kohlen-stoffatomen verbunden, sodass ein dreidimensiona-les, extrem stabiles, tetraedrisches Kristallgitter ent-steht. Graphit besteht dagegen aus flachen Lagen von hexagonal angeordneten C-Atomen. Innerhalb der Graphitschichten ist jedes C-Atom kovalent mit drei Nachbaratomen verbunden. Die Lagen verschieben sich seitwärts übereinander, weil sich zwischen den Atomen der einzelnen Schichten keine Bindungen ausbilden.
Um ein Nanoröhrchen herzustellen, wird eine einzelne Graphitschicht zu einem Zylinder aufge-rollt. Die Schichten können gerade oder in einem bestimmten Winkel zum Kohlenstoffgitter gerollt sein und Röhrchen mit unterschiedlichen Durch-messern bilden. Abhängig vom Durchmesser und der angelegten Spannung, können die Nanoröhrchen als metallische Leiter oder Halbleiter fungieren. Es überrascht nicht, dass die Röhrchen in der Elektronik reichlich Anwendung finden, ein Thema, das den Rahmen dieses Buches allerdings sprengen würde.
In der Biotechnologie steht der Einsatz der Nano-röhrchen noch am Anfang, doch zukünftig sollen andere Biomoleküle wie Enzyme, Hormonrezeptoren oder Antikörper mit der Oberfläche der Röhrchen verknüpft werden. Ein Ziel ist, Biosensoren zu kon-struieren, bei denen z.B. die Wechselwirkung eines Antikörpers mit seinem Antigen das elektrische Ver-halten des Nanoröhrchens ändert. Die Bindung von Hormonen, Pathogenen, Verunreinigungen usw. soll ein elektrisches Signal generieren.
Eines der wichtigsten Probleme bei der Kopplung der Röhrchen an Proteine ist die Hydrophobizität der Nanoröhrchenoberfläche. Eine Lösung ist, zunächst die Oberfläche durch die Zugabe von nichtionischen Detergenzien wie Triton-X100 zu verändern. Der hy-drophobe Bereich des Detergens bindet an die Röhr-chenoberfläche und die hydrophile Region bindet die Proteine. Alternativ lassen sich auch chemische Reagenzien einsetzen, die mit der Kohlenstoffober-fläche reagieren und Seitenketten ausbilden, die re-aktive funktionelle Gruppen tragen. Proteine können dann durch eine Reaktion mit diesen Gruppen an-gehängt werden (Abb. 7.13). Die Konstruktion von
7.12 Zusammenbau eines Nanokabels durch einen BakteriophagenDas Phagen-Display liefert gentechnisch veränderte Vari-
anten des M13-Hüllproteins (Protein VIII), mit eingebauten
Peptiden. Einige dieser Peptide vermögen CdS zu binden.
In Anwesenheit von CdS-Kristallen bildet sich auf der
Oberfläche des Bakteriophagen ein Nanokabel.
Protein VIII
Protein VIII, dasSLTPLTTSHLRSpräsentiert
Screening der Phagen-bibliothek nach einem CdS-bindenden Peptid
Zugabe von CdS
CdS CdS CdS CdS CdS CdS CdS
CdS CdS CdS CdS CdS CdS CdS
Nanokabel auf dem Bakteriophagen M13
veränderter Bakteriophage M13
Wildtyp des Bakteriophagen M13

214 Nanobiotechnologie7
Maschinen durch die Kombination von biologischen Molekülen mit Nanoröhrchen steckt noch in den Kinderschuhen, doch der Fortschritt auf diesem Ge-biet ist rasant.
7.13 Die Verknüpfung von Nanoröhrchen mit funktionellen organischen Gruppena Nanoröhrchen aus Kohlenstoff werden mit Säure behandelt, um sie zu reinigen und um Carboxylgruppen herzustellen.
b Alternativ können sie mit Aminosäurederivaten und Aldehyden reagieren, um komplexere hydrophile Gruppen mit der
äußeren Oberfläche zu verknüpfen. Aus: Bianco, Kostarelos und Prato (2005) Applications of carbon nanotubes in drug
delivery. Curr Opin Chem Biol 9: 674–679. Nachdruck mit freundlicher Genehmigung.
HOOC
H 2SO 4/
HNO 3
1) R-NHCH2 CO
2 H
(CH2 O)n /DMF2) HCI/DCM
HOOC
HOOC
O
N
N
NH
O
OOO
R =
O
O
O
a
b
NH3+
NH3+
COOH COOH
COOHCOOH
COOH
HOOC
HOOC
Magnetosomen: natürliche, magnetische Nanopartikel aus BakterienNatürlich vorkommende, magnetische Nanopartikel wer-
den von magnetotaktischen Bakterien wie Magnetospiril-
lum gebildet. Diese Mikroorganismen können Magnetfel-
der wahrnehmen und orientieren sich an ihnen. Die Orga-
nismen enthalten Magnetosomen, die aus Kristallen aus
magnetischem Eisenoxid (Magnetit, Fe3O4) in Nanogröße
oder, was seltener der Fall ist, aus Eisensulfid (Greigit,
Fe3S4) bestehen, eingehüllt in Protein. Die Magnetoso-
men ordnen sich entlang der Zellachse in einer Reihe an.
Die Synthese der Proteinhülle und die Mineralisierung
des magnetischen Kerns werden genetisch kontrolliert.
Zumindest in einigen Fällen bilden Gene, die für die Bil-
dung von Magnetosomen verantworlich sind, auf dem
bakteriellen Chromosom Cluster.
Es ist möglich, andere Moleküle mit der Außenseite
eines Magnetosoms zu verbinden, indem man die Pro-
teinhülle der Magnetosomen gentechnisch verändert.
Im Labor von Dr. Tadashi Matsunaga der University of
Agriculture and Technology in Tokyo wurde das Gen für
das Mms16-Protein von Magnetospirillum magneticum
mit den Genen für Luciferase und den Dopaminrezeptor
fusioniert. Die fusionierten Proteine wurden auf der Ober-
fläche des Magnetosoms präsentiert. Nach der Zerstö-
rung der Bakterienzelle konnte man die Magnetosomen,
die die Fremdproteine trugen, durch magnetische Wech-
selwirkungen isolieren. Dadurch wird die Analyse mem-
brangebundener Rezeptoren wie denen des menschli-
chen Nervensystems erleichtert.
Exkurs 7.2
Es lassen sich hohle Nanoröhrchen herstellen, die
verschiedene, biologisch nützliche Seitenketten
tragen.

Ionenkanäle als Nanosensoren 215 7
Antibakterielle Nanoschichten
Nanoschichten werden durch viele Nanoröhrchen gebildet, die so nebeneinander ausgerichtet sind, dass die Achsen ihrer Zylinder parallel zueinander stehen. Nanoschichten, die Farben verändern und Bakterien abtöten, wurden aus speziell konstruier-ten Lipiden geschaffen, die sich in Abhängigkeit von den Bedingungen zu einer Vielzahl von Nano-strukturen zusammenlagern. In Wasser entstehen Nanoröhrchen. Durch partielle Rehydrierung ge-trockneter Nanoröhrchen lagern sich die Röhrchen Seite an Seite nebeneinander, und es bildet sich eine Nanoschicht.
Das Lipid besteht aus einer langen Kohlenwas-serstoffkette (C25) mit einer Diacetylengruppe in der Mitte. Ein einzelnes Nanoröhrchen hat einen Durchmesser von etwa 100 nm und ist etwa 1000 nm lang. Die Wände der Nanoröhrchen bestehen aus fünf Lipiddoppelschichten. Beide, sowohl die einzel-nen Lipidmoleküle als auch die zusammengesetzte Schicht, töten Bakterien ab. Wie andere langkettige Aminoverbindungen auch, wirken sie als Detergen-zien und zerstören die Zellmembran. Somit stellt die Nanoschicht eine Oberfläche dar, die für Bakterien tödlich ist. Diese Eigenschaft könnte sehr nützlich sein für die Anwendung der Nanoschichten in der Biomedizin.
Diacetylenverbindungen haben die interessante Eigenschaft, ihre Farbe zu verändern. Zu Beginn ist die Nanoschicht weiß, trifft jedoch UV-Licht auf, färbt sich die Schicht in tiefes Blau. Die UV-Strahlung führt zu Quervernetzungen zwischen den Acetylengruppen und benachbarten Molekülen und die Polymerisierung stabilisiert die Schicht. Blaue Nanoschichten verändern ihre Farbe als Re-aktion auf eine Vielzahl von Substanzen. Deter-genzien und Säuren färben sie von blau nach rot oder gelb, und die Anwesenheit von Bakterien wie E. coli ergibt Rot- oder Rosatöne. Solche Materi-alien ließen sich sowohl als Biosensoren als auch zum Schutz gegen bakterielle Kontamination ein-setzen.
Nanoröhrchen lassen sich zu Schichten arrangie-
ren, und es entstehen Oberflächen, die antibakteri-
ell wirken oder als Biosensoren eingesetzt werden
können.
Nachweis von Viren durch Nanokabel
Nanokabel haben einen Durchmesser im Nanobe-reich, doch sie können mehrere Mikrometer lang sein. Sie können metallisch sein und als elektrische Leiter dienen, oder sie bestehen aus halbleitenden Materialien.
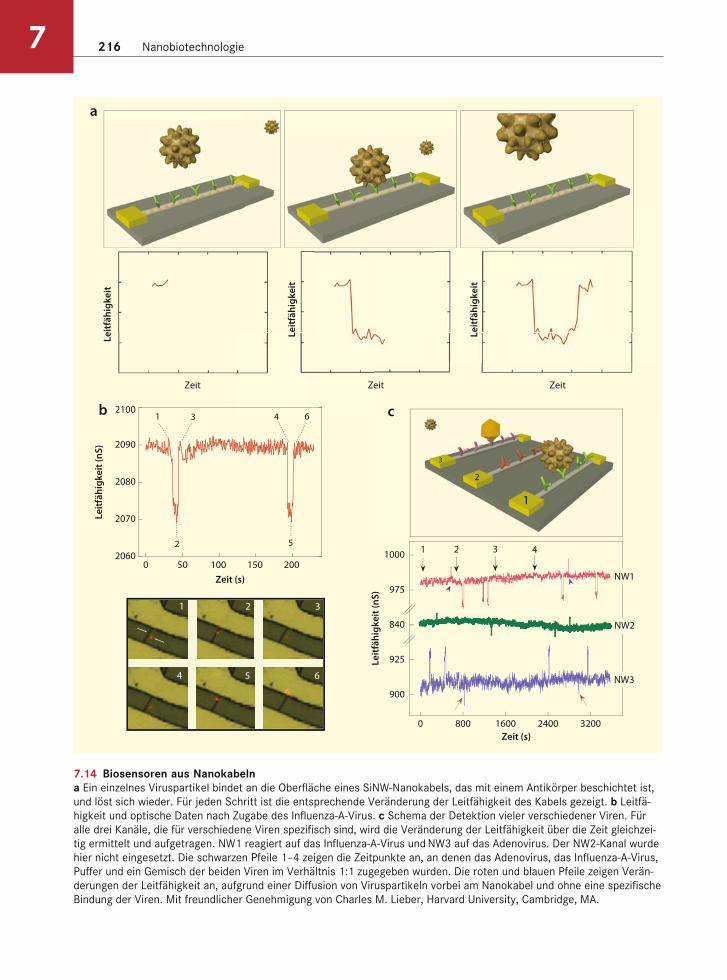
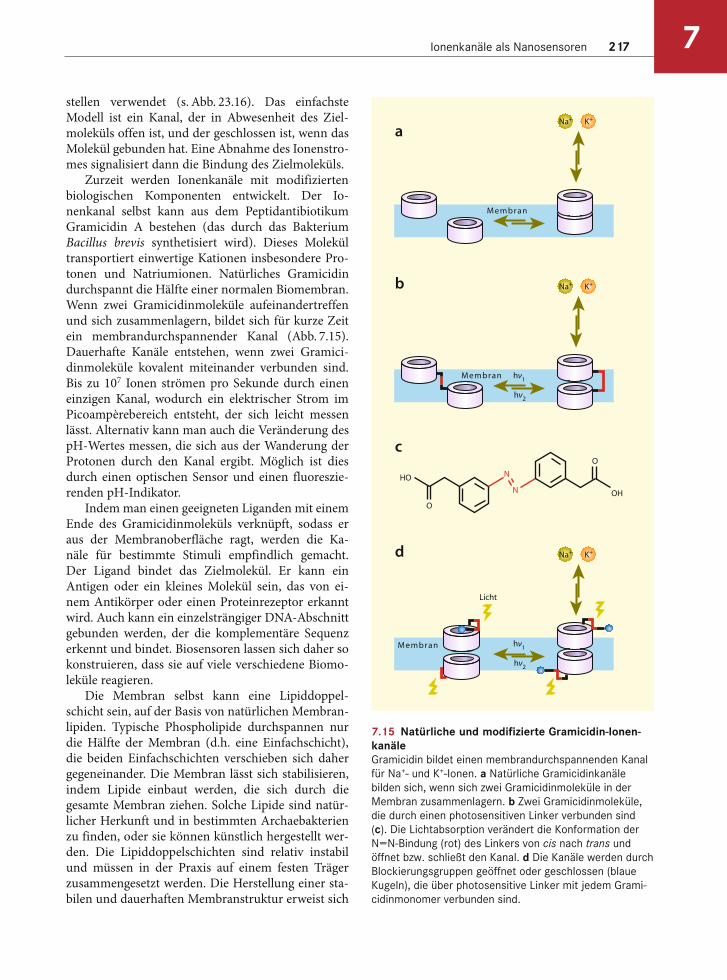
Aus Silizium-Halbleiternanokabeln lassen sich Biosensoren herstellen. Die Kabel können mit An-tikörpern beschichtet werden, die spezifisch an ein Virus binden. Die Bindung verändert die Leitfähig-keit des Nanokabels. Bei einem Siliziumnanokabel vom P-Typ nimmt die Leitfähigkeit ab, wenn die Oberflächenladung auf dem Viruspartikel positiv ist. Umgekehrt nimmt sie zu, wenn die Virenoberfläche negativ geladen ist. Durch diesen Ansatz lassen sich einzelne Viren nachweisen (Abb. 7.14). Außerdem ist es möglich, eine einzelsträngige DNA an dem Nanokabel zu befestigen. In diesem Fall wird die Leitfähigkeit durch die Bindung des komplementä-ren Stranges verändert. Zu den zukünftigen Anwen-dungen gehören sowohl klinische Tests als auch die Verwendung als Sensoren für die Überwachung von Lebensmitteln, Wasser und der Atmosphäre, um die Gesundheit der Bevölkerung zu sichern.
Sensoren aus Nanokabeln vermögen einzelne Viren
spezifisch nachzuweisen. Die Bindung eines Virus-
partikels verändert die Leitfähigkeit des Kabels.
Ionenkanäle als Nanosensoren
Komplexer als Nanoröhrchen und Nanokabel sind Ionenkanäle im Nanomaßstab, die in Membranen eingebaut werden. Diese Kanäle lassen sich so regu-lieren, dass Ionen sie nur unter bestimmten Bedin-gungen passieren können. Der Ionenstrom erzeugt einen elektrischen Strom, der von einer entspre-chenden Apparatur registriert, verstärkt und darge-stellt wird.
Ionenkanäle lassen sich als Biosensoren einsetzen, indem man eine Bindungsstelle für ein Zielmolekül an den Kanaleingang heftet. Wie in Kapitel 23 be-schrieben, werden häufig Antikörper als Bindungs-
216 Nanobiotechnologie7
7.14 Biosensoren aus Nanokabelna Ein einzelnes Viruspartikel bindet an die Oberfläche eines SiNW-Nanokabels, das mit einem Antikörper beschichtet ist,
und löst sich wieder. Für jeden Schritt ist die entsprechende Veränderung der Leitfähigkeit des Kabels gezeigt. b Leitfä-
higkeit und optische Daten nach Zugabe des Influenza-A-Virus. c Schema der Detektion vieler verschiedener Viren. Für
alle drei Kanäle, die für verschiedene Viren spezifisch sind, wird die Veränderung der Leitfähigkeit über die Zeit gleichzei-
tig ermittelt und aufgetragen. NW1 reagiert auf das Influenza-A-Virus und NW3 auf das Adenovirus. Der NW2-Kanal wurde
hier nicht eingesetzt. Die schwarzen Pfeile 1–4 zeigen die Zeitpunkte an, an denen das Adenovirus, das Influenza-A-Virus,
Puffer und ein Gemisch der beiden Viren im Verhältnis 1:1 zugegeben wurden. Die roten und blauen Pfeile zeigen Verän-
derungen der Leitfähigkeit an, aufgrund einer Diffusion von Viruspartikeln vorbei am Nanokabel und ohne eine spezifische
Bindung der Viren. Mit freundlicher Genehmigung von Charles M. Lieber, Harvard University, Cambridge, MA.
2060
2070
2080
2090
2100
0 50 100 150 200
Leitfä
hig
keit
(nS)
Zeit (s)
1 2 3
4 5 6
1
2
3 4
5
6
Zeit
Leitfä
hig
keit
a
b
Zeit
Leitfä
hig
keit
Zeit
Leitfä
hig
keit
900
925
840
975
1000
0 800 1600 2400 3200
Leitfä
hig
keit
(nS)
Zeit (s)
NW1
NW3
1 2 3 4
1
3
2
NW2
c
Ionenkanäle als Nanosensoren 217 7
stellen verwendet (s. Abb. 23.16). Das einfachste Modell ist ein Kanal, der in Abwesenheit des Ziel-moleküls offen ist, und der geschlossen ist, wenn das Molekül gebunden hat. Eine Abnahme des Ionenstro-mes signalisiert dann die Bindung des Zielmoleküls.
Zurzeit werden Ionenkanäle mit modifizierten biologischen Komponenten entwickelt. Der Io-nenkanal selbst kann aus dem Peptidantibiotikum Gramicidin A bestehen (das durch das Bakterium Bacillus brevis synthetisiert wird). Dieses Molekül transportiert einwertige Kationen insbesondere Pro-tonen und Natriumionen. Natürliches Gramicidin durchspannt die Hälfte einer normalen Biomembran. Wenn zwei Gramicidinmoleküle aufeinandertreffen und sich zusammenlagern, bildet sich für kurze Zeit ein membrandurchspannender Kanal (Abb. 7.15). Dauerhafte Kanäle entstehen, wenn zwei Gramici-dinmoleküle kovalent miteinander verbunden sind. Bis zu 107 Ionen strömen pro Sekunde durch einen einzigen Kanal, wodurch ein elektrischer Strom im Picoampèrebereich entsteht, der sich leicht messen lässt. Alternativ kann man auch die Veränderung des pH-Wertes messen, die sich aus der Wanderung der Protonen durch den Kanal ergibt. Möglich ist dies durch einen optischen Sensor und einen fluoreszie-renden pH-Indikator.
Indem man einen geeigneten Liganden mit einem Ende des Gramicidinmoleküls verknüpft, sodass er aus der Membranoberfläche ragt, werden die Ka-näle für bestimmte Stimuli empfindlich gemacht. Der Ligand bindet das Zielmolekül. Er kann ein Antigen oder ein kleines Molekül sein, das von ei-nem Antikörper oder einen Proteinrezeptor erkannt wird. Auch kann ein einzelsträngiger DNA-Abschnitt gebunden werden, der die komplementäre Sequenz erkennt und bindet. Biosensoren lassen sich daher so konstruieren, dass sie auf viele verschiedene Biomo-leküle reagieren.
Die Membran selbst kann eine Lipiddoppel-schicht sein, auf der Basis von natürlichen Membran-lipiden. Typische Phospholipide durchspannen nur die Hälfte der Membran (d.h. eine Einfachschicht), die beiden Einfachschichten verschieben sich daher gegeneinander. Die Membran lässt sich stabilisieren, indem Lipide einbaut werden, die sich durch die gesamte Membran ziehen. Solche Lipide sind natür-licher Herkunft und in bestimmten Archaebakterien zu finden, oder sie können künstlich hergestellt wer-den. Die Lipiddoppelschichten sind relativ instabil und müssen in der Praxis auf einem festen Träger zusammengesetzt werden. Die Herstellung einer sta-bilen und dauerhaften Membranstruktur erweist sich
7.15 Natürliche und modifizierte Gramicidin-Ionen-kanäleGramicidin bildet einen membrandurchspannenden Kanal
für Na+- und K+-Ionen. a Natürliche Gramicidinkanäle
bilden sich, wenn sich zwei Gramicidinmoleküle in der
Membran zusammenlagern. b Zwei Gramicidinmoleküle,
die durch einen photosensitiven Linker verbunden sind
(c). Die Lichtabsorption verändert die Konformation der
N=N-Bindung (rot) des Linkers von cis nach trans und
öffnet bzw. schließt den Kanal. d Die Kanäle werden durch
Blockierungsgruppen geöffnet oder geschlossen (blaue
Kugeln), die über photosensitive Linker mit jedem Grami-
cidinmonomer verbunden sind.
d
c
Na+ K+
a
Na+ K+b
hν1
HO
OH
O
O
N
N
Licht
Membran
Membran
Membran
hν2
Na+ K+
hν1
hν2

218 Nanobiotechnologie7
als schwierig und Ionenkanalsensoren befinden sich noch im Entwicklungsstadium.
Ionenkanalsensoren öffnen oder schließen sich als
Reaktion auf die spezifische Bindung eines Mole-
küls. Sie lassen sich für den Nachweis von Zielmo-
lekülen einsetzen.
Gentechnische Veränderung von DNA im Nanomaßstab
In der klassischen Gentechnik wird die DNA-Se-quenz gezielt verändert, um neue Kombinationen genetischer Information zu schaffen. Und auch wenn die DNA im größeren Maßstab umorganisiert wird, so bleibt sie doch eine lineare, doppelsträngige Helix mit Basenpaaren.
Die gentechnische Veränderung im Nanomaßstab hat die Herstellung von Strukturen zum Ziel, bei denen DNA als Strukturelement dient, und nicht die Veränderung der genetischen Information selbst. DNA ist in dieser Hinsicht sehr attraktiv, weil die Doppelhelix ein leicht handhabbares Modul darstellt. Außerdem lässt sich die natürliche Eigenschaft der Basenpaarung nutzen, um zwei DNA-Moleküle mit-einander zu verbinden. Um dreidimensionale Struk-turen zu schaffen, ist allerdings eine Verzweigung der DNA notwendig. Verzweigte Strukturen entste-hen zwar auch in der Natur (z.B. bei der Holliday-Struktur, die sich während der Rekombination durch Crossing-over bildet), doch sind sie nicht stabil und dauerhaft.
Kreuzförmige DNA lässt sich herstellen, indem man vier sorgfältig konstruierte Einzelstränge mit unterschiedlichen Sequenzen mischt. Jeder Strang bil-det über die Hälfte seiner Länge Basenpaare mit zwei anderen Strängen aus (Abb. 7.16). Besitzen die beiden ursprünglichen Stränge zwei kohäsive Enden, können die beiden Kreuze zu einer zweidimensionalen Matrix verbunden werden. Die Einzelstrangbrüche können, falls erwünscht, mit DNA-Ligase geschlossen werden. Die Prinzipien für die Herstellung der Verzweigung lassen sich auch auf die dritte Dimension übertragen, wodurch kubische DNA-Gitter entstehen.
Die DNA-Doppelhelix hat einen Durchmesser von etwa 2 nm und eine Windung umfasst etwa 3,5 nm. DNA lässt sich daher für die Herstellung von Gerüsten im Nanomaßstab einsetzen. Diese können
wiederum verwendet werden, um andere Bestand-teile wie metallische Nanokabel oder Nanoschalt-kreise zusammenzusetzen. Während die DNA über lange Strecken flexibel ist, ist sie im Nanomaßstab (bis zu ca. 50 nm) jedoch relativ starr.
Solche kreuzförmigen DNA-Moleküle (und die dreidimensionalen Produkte) haben den Nachteil, dass die Kreuzungen flexibel sind und keinen festen 90°-Winkel haben. Um hier Abhilfe zu schaffen, wur-den steifere DNA-Moleküle mit doppeltem Crossing-over (DX-DNA-Moleküle) konstruiert. Grundlage
7.16 Verzweigte DNA aus vier einzelnen Strängena Ein verzweigtes DNA-Molekül mit vier Armen. Vier ver-
schiedene Stränge (mit vier Farben dargestellt) werden
miteinander kombiniert und es entsteht ein Molekül mit
vier Armen (I, II, III und IV). Der Verzweigungspunkt dieses
Moleküls ist stabil. b Die Bildung eines zweidimensionalen
Gitters aus vierarmigen Molekülen mit kohäsiven Enden.
X und Y sind kohäsive Enden und X’ und Y’ sind die ih-
nen entsprechenden Enden. Vier Monomere lagern sich
parallel aneinander und es entsteht eine Gitterstruktur.
DNA-Ligase schließt die Einzelstrangbrüche im Gitter. Aus
Seeman (1999) DNA engineering and its application to na-
notechnology. Trends Biotechnol 17: 437. Nachdruck mit
freundlicher Genehmigung.
GCGTTAGG
C G T G C T C AG C A C G A G T T G A T A C C G
A C T A T G G C
CGCAATCC
CCGAATGC
GGCTTACG
I
II
a
b
III
IV
Y ’
Y ’
YY
YX
X
XX’
X’
X’
Y ’

Mechanische DNA-Nanomaschinen 219 7
sind hier zwei Isomere der antiparallelen DX-DNA, mit einer ungeraden (DAO) oder geraden (DAE) Zahl an halben Drehungen zwischen den Crossing-over-Stellen (Abb. 7.17). (Es gibt auch Moleküle mit doppeltem Crossing-over und parallelen Strängen, doch sie sind aus struktureller Sicht nicht so gut geeig-net.) DAE und DAO-Einheiten lassen sich zu einem starren Netzwerk zusammenfügen, wenn geeignete kohäsive Enden vorhanden sind. Und es ist möglich, den zentralen, kurzen DNA-Strang der DAE-Struktur durch einen längeren Strang zu ersetzen, der aus dem Netzwerk herausragt (DAE + J). Dadurch können verzweigte Strukturen geschaffen werden.
Welches Ziel verfolgt man mit der Herstellung von Netzwerken und dreidimensionalen Strukturen aus DNA? Der plausibelste Grund ist die mögliche Ver-wendung der DNA als Gerüst für den Zusammenbau
elektronischer Schaltkreise im Nanomaßstab. Bislang wurde nichtverzweigte DNA als Gerüst eingesetzt, um lineare Nanokabel aus Metall herzustellen. Verschie-dene Metalle (Gold, Silber, Kupfer, Palladium, Platin) bildeten eine Hülle um die DNA, und die Durch-messer variierten von 3 bis 100 nm. Es sollte letztlich möglich sein, diese beiden Ansätze miteinander zu kombinieren und Schaltkreise aus metallumhüllten, dreidimensionalen DNA-Strukturen zu konstruieren.
DNA lässt sich auch als strukturgebendes Molekül
betrachten. Es ist möglich, dreidimensionale Netz-
werke aus DNA herzustellen, deren Sequenz so ge-
zielt synthetisiert wurde, dass sich diese verzweigten
Strukturen bilden. Solche DNA-Strukturen lassen
sich als Gerüst verwenden, um metallische Kabel
und Schaltkreise im Nanomaßstab herzustellen.
Mechanische DNA-Nanomaschinen
Eine weitere, eher futuristische Anwendung für drei-dimensionale DNA-Strukturen ist die eines Gerüs-tes für mechanische Nanomaschinen. Notwendige Bestandteile einer solchen Maschine sind sich be-wegende Teile. Etliche Prototypen solcher „DNA-Maschinen“ wurden entworfen oder konstruiert und verdeutlichen das Konzept. Alle beruhen auf reversib-len Veränderungen in der Konformation einer DNA-Struktur, die durch Veränderungen der Basenpaa-rung angetrieben wird. Verursacht werden können sie durch eine Modifikation der physikalischen Be-dingungen (Hitze, Salz usw.) oder durch die Addition von einzelsträngigen DNA-Abschnitten (Abb. 7.18). Wird anfangs ssDNA eingesetzt, dann gibt man ei-nen weiteren Einzelstrang zu, der komplementär zu dem ersten ist, sodass sich die Maschine wieder in die Ausgangsposition zurück bewegt. Das Ergebnis ist ein mechanischer Kreislauf, der sich im Grunde genommen einsetzen lässt, um eine bestimmte Auf-gabe zu erfüllen. Die beiden ssDNA-Moleküle sind eine Art „Treibstoff “ und das Abfallprodukt ist eine doppelsträngige DNA, die aus den beiden ssDNA-Treibstoff-Elementen besteht. (Dieses Schema beruht nicht auf der Spaltung von kovalenten Bindungen. Es ist daher keine enzymatische Reaktion und un-terscheidet sich von der Verwendung von DNA als Desoxyribozym, wie in Kapitel 5 beschrieben.)
7.17 Starre DNA-NanomoduleAus Molekülen mit doppeltem Crossing-over (DX-Mole-
külen) lassen sich Netzwerke herstellen. a DAE und DAO
sind zwei antiparallele DX-Isomere. DAE + J ist ein DAE-
Molekül, bei dem eine zusätzliche Kreuzung den Einzel-
strangbruch im grünen Strang von DAE ersetzt. b Zweidi-
mensionales Netzwerk aus DX-Molekülen. Komplementäre
kohäsive Enden sind durch komplementäre geometrische
Formen dargestellt. A ist ein herkömmliches DX-Molekül,
B* ist dagegen ein DX + J-Molekül mit einer vertikal her-
ausragenden DNA-Haarnadelstruktur (schwarzer Punkt).
Aus Seeman (1999) DNA engineering and its application
to nanotechnology. Trends Biotechnol 17: 437. Nachdruck
mit freundlicher Genehmigung.
a
b
DAE
A B*
DAO DAE + J

220 Nanobiotechnologie7
DNA wurde auch für den Einsatz als Gerüst für
Nanomaschinen vorgeschlagen. Prototypen wurden
bereits hergestellt, anhand derer man das Konzept
überprüft hat.
Kontrollierte Denaturierung von DNA durch Nanopartikel aus Gold
Die DNA-Hybridisierung wird sowohl im Labor als auch in der klinischen Diagnose häufig für den Nachweis von Zielsequenzen eingesetzt. Bevor eine Hybridisierung stattfinden kann, muss die DNA-Doppelhelix zu Einzelsträngen denaturiert werden.
Dieses geschieht durch das Erhitzen der gesamten DNA. Durch die Fortschritte in der Nanotechnologie ist es zukünftig vielleicht möglich, einzelne DNA-Moleküle gezielt dissoziieren zu lassen.
Dazu lagern sich Nanopartikel mit einer Größe von etwa 1,4 nm und bestehend aus weniger als 100 Goldatomen an die doppelsträngige DNA. Wird diese Struktur Radiowellen ausgesetzt (die durch ein wechselndes Magnetfeld entstehen), wirkt das Gold als eine Art Antenne. Es absorbiert Energie und heizt die DNA-Moleküle auf, mit denen es ver-bunden ist. Die Doppelhelix wird zu Einzelsträngen aufgeschmolzen. Die Hitze breitet sich über einen Bereich von etwa 10 nm aus, sodass die umgeben-den Moleküle nicht beeinflusst werden. Sie klingt innerhalb von weniger als 50 Picosekunden ab, so-dass die DNA schnell zwischen einem einzelsträngi-gen und einem doppelsträngigen Zustand wechseln kann, wenn das Magnetfeld an- und ausgeschaltet wird. Das Verfahren lässt sich auf dsDNA anwen-den, die aus zwei getrennten Einzelsträngen besteht (Abb. 7.19), oder auf Stamm-Schleife-Strukturen, die durch die Faltung eines einzelnen Stranges ent-stehen.
Praktische Anwendungen werden sich erst in der Zukunft ergeben. Da Radiowellen jedoch lebendes Gewebe sehr effizient durchdringen, könnte es mög-lich sein, das Verhalten einzelner DNA-Moleküle von außen zu steuern. Metallantennen unterschiedlicher Materialien oder Größen könnten eingesetzt werden, um verschiedene DNA-Moleküle auf Radiowellen unterschiedlicher Frequenzen einzustellen.
Durch die Anlagerung von metallischen Antennen
lässt sich DNA mithilfe von Radiowellen schmelzen.
Es könnte zukünftig möglich sein, das Verhalten
von DNA von außen zu steuern.
Kontrollierte Veränderung der Proteinstruktur durch DNA
Allosterische Proteine verändern ihre Struktur als Reaktion auf die Bindung von Signalmolekülen (al-losterischen Effektoren) an einer spezifischen Stelle im Molekül. Wesentlich bei der allosterischen Kon-trolle ist, dass die Veränderung der Struktur im
7.18 Prototyp einer DNA-MaschineEin DNA-Nanomotor, entworfen von J. J. Li und W. Tan.
Die aufeinanderfolgende Addition von komplementären
DNA-Strängen (markiert mit α und β) verursachte eine
Konformationsänderung. Der DNA-Nanomotor wechselt
zwischen einer gefalteten Quadruplexstruktur und einer
doppelsträngigen Struktur. Der Nanomotor dehnt sich aus
und kontrahiert in einer wurmähnlichen Bewegung. Aus Ito
und Fukusaki (2004) DNA as a nanomaterial. J Mol Cataly-
sis B: Enzymatic 28: 155–166. Nachdruck mit freundlicher
Genehmigung.
α
α β
β

Kontrollierte Veränderung der Proteinstruktur durch DNA 221 7
Protein übertragen wird und sich an einer anderen Stelle des Moleküls auswirkt. Bei allosterischen En-zymen verändert die Bindung eines allosterischen Effektors an einer entfernt liegenden Stelle die Kon-formation des aktiven Zentrums und kann dessen Affinität für das Substrat beeinflussen. Auf diese Weise werden einige Enzyme als Reaktion auf ein Signalmolekül aktiviert oder inaktiviert. So wird die Phosphofructokinase z.B. durch hohe Konzentrati-onen von AMP aktiviert, da AMP einen Mangel an Energie signalisiert. Durch die Aktivierung erhöht sich der Fluss von Metaboliten durch die Glykolyse. In ähnlicher Weise verändern viele DNA-bindende Proteine wie Repressoren und Aktivatoren ihre Ge-stalt, nachdem sie kleine Signalmoleküle gebunden haben.
Es ist möglich, die Struktur eines Proteins auch künstlich durch die Anwendung einer mechanischen Kraft zu verändern. Zeigen konnte man dies durch die Verknüpfung eines einzelsträngigen, 60 Basen langen DNA-Abschnittes mit den beiden Enden eines Proteins. Für diesen Ansatz waren chemische „Hand-griffe“ im Zielprotein erforderlich. Diese wurden in das Zielprotein eingesetzt, indem man die Amino-
säuren an geeigneten Stellen durch Cystein ersetzte. An diese reaktiven SH-Gruppen wurde anschließend die DNA geknüpft.
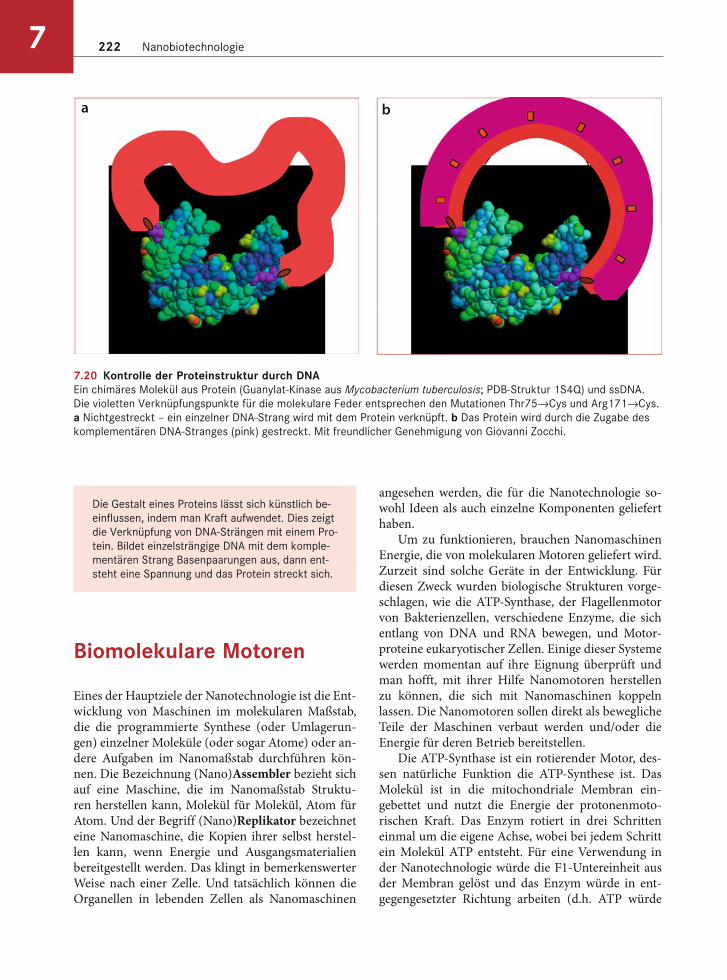
Doppelhelikale DNA ist viel stabiler als ssDNA. Die Addition bzw. Bindung des komplementären Stranges, bei der Bildung der Doppelhelix, führt zu einer Spannung. Im Labor von Giovanni Zocchi an der UCLA führte man diesen Ansatz mit einem maltosebindenden Protein und einem Enzym, der Guanylat-Kinase, durch. Wurde das maltosebin-dende Protein gestreckt, dann weitete sich die Bin-dungsstelle für die Maltose über das optimale Maß hinaus und die Affinität für den Zucker sank. Bei der Guanylat-Kinase (Abb. 7.20) verringerte die Span-nung die Enzymaktivität, da die Affinität für das Substrat reduziert wurde. In diesem Fall ließ sich das Enzym durch die Zugabe einer DNase aktivieren, die die DNA spaltete.
Möglicherweise ergeben sich zukünftig Anwen-dungen für dieses Verfahren. So sind Biosensoren vorstellbar, die DNA-Sequenzen nachweisen können. Außerdem könnte es möglich sein, Enzyme oder andere Proteine von außen zu kontrollieren, indem man eine geeignete ssDNA (oder RNA) zugibt.
Streptavidin
Au
F
Au
F
Au
F
Au
F
B
B B
B
Agarose-kügelchen
7.19 Steuerung der DNA-Dena-turierung durch Nanopartikel aus GoldAn das Ende eines DNA-Stranges
sind Nanopartikel aus Gold (Au)
gebunden, das andere Ende wurde
mit einem Fluoreszenzfarbstoff
(F) verknüpft. Der komplementäre
Strang trägt an einem Ende ein Bio-
tin-Tag (B). Das Biotin wird durch
Streptavidin gebunden und koppelt
den DNA-Strang dadurch an ein
Agarosekügelchen. Absorbiert das
Gold Energie, dann schmelzen die
beiden Stränge. Der Strang mit
dem Fluoreszenzfarbstoff wird in
den Überstand freigesetzt und kann
anhand der Fluoreszenz nachgewie-
sen werden.
222 Nanobiotechnologie7
Die Gestalt eines Proteins lässt sich künstlich be-
einflussen, indem man Kraft aufwendet. Dies zeigt
die Verknüpfung von DNA-Strängen mit einem Pro-
tein. Bildet einzelsträngige DNA mit dem komple-
mentären Strang Basenpaarungen aus, dann ent-
steht eine Spannung und das Protein streckt sich.
Biomolekulare Motoren
Eines der Hauptziele der Nanotechnologie ist die Ent-wicklung von Maschinen im molekularen Maßstab, die die programmierte Synthese (oder Umlagerun-gen) einzelner Moleküle (oder sogar Atome) oder an-dere Aufgaben im Nanomaßstab durchführen kön-nen. Die Bezeichnung (Nano)Assembler bezieht sich auf eine Maschine, die im Nanomaßstab Struktu-ren herstellen kann, Molekül für Molekül, Atom für Atom. Und der Begriff (Nano)Replikator bezeichnet eine Nanomaschine, die Kopien ihrer selbst herstel-len kann, wenn Energie und Ausgangsmaterialien bereitgestellt werden. Das klingt in bemerkenswerter Weise nach einer Zelle. Und tatsächlich können die Organellen in lebenden Zellen als Nanomaschinen
angesehen werden, die für die Nanotechnologie so-wohl Ideen als auch einzelne Komponenten geliefert haben.
Um zu funktionieren, brauchen Nanomaschinen Energie, die von molekularen Motoren geliefert wird. Zurzeit sind solche Geräte in der Entwicklung. Für diesen Zweck wurden biologische Strukturen vorge-schlagen, wie die ATP-Synthase, der Flagellenmotor von Bakterienzellen, verschiedene Enzyme, die sich entlang von DNA und RNA bewegen, und Motor-proteine eukaryotischer Zellen. Einige dieser Systeme werden momentan auf ihre Eignung überprüft und man hofft, mit ihrer Hilfe Nanomotoren herstellen zu können, die sich mit Nanomaschinen koppeln lassen. Die Nanomotoren sollen direkt als bewegliche Teile der Maschinen verbaut werden und/oder die Energie für deren Betrieb bereitstellen.
Die ATP-Synthase ist ein rotierender Motor, des-sen natürliche Funktion die ATP-Synthese ist. Das Molekül ist in die mitochondriale Membran ein-gebettet und nutzt die Energie der protonenmoto-rischen Kraft. Das Enzym rotiert in drei Schritten einmal um die eigene Achse, wobei bei jedem Schritt ein Molekül ATP entsteht. Für eine Verwendung in der Nanotechnologie würde die F1-Untereinheit aus der Membran gelöst und das Enzym würde in ent-gegengesetzter Richtung arbeiten (d.h. ATP würde
7.20 Kontrolle der Proteinstruktur durch DNAEin chimäres Molekül aus Protein (Guanylat-Kinase aus Mycobacterium tuberculosis; PDB-Struktur 1S4Q) und ssDNA.
Die violetten Verknüpfungspunkte für die molekulare Feder entsprechen den Mutationen Thr75→Cys und Arg171→Cys.
a Nichtgestreckt – ein einzelner DNA-Strang wird mit dem Protein verknüpft. b Das Protein wird durch die Zugabe des
komplementären DNA-Stranges (pink) gestreckt. Mit freundlicher Genehmigung von Giovanni Zocchi.
a b
Biomolekulare Motoren 223 7
Fracht
Motordomäne
spiralisierte Domäne
C-terminales Ende
Mikrotubulus
Kinesinrezeptor leichte Kette des Kinesins
schwereKette desKinesins
+
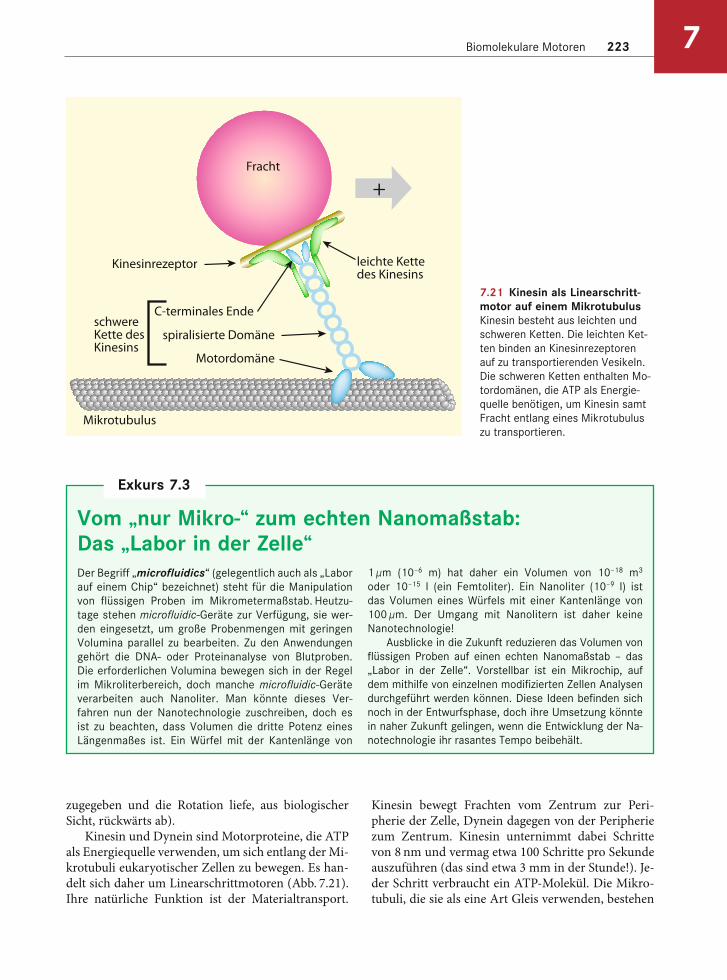
7.21 Kinesin als Linearschritt-motor auf einem MikrotubulusKinesin besteht aus leichten und
schweren Ketten. Die leichten Ket-
ten binden an Kinesinrezeptoren
auf zu transportierenden Vesikeln.
Die schweren Ketten enthalten Mo-
tordomänen, die ATP als Energie-
quelle benötigen, um Kinesin samt
Fracht entlang eines Mikrotubulus
zu transportieren.
Vom „nur Mikro-“ zum echten Nanomaßstab: Das „Labor in der Zelle“Der Begriff „microfluidics“ (gelegentlich auch als „Labor
auf einem Chip“ bezeichnet) steht für die Manipulation
von flüssigen Proben im Mikrometermaßstab. Heutzu-
tage stehen microfluidic-Geräte zur Verfügung, sie wer-
den eingesetzt, um große Probenmengen mit geringen
Volumina parallel zu bearbeiten. Zu den Anwendungen
gehört die DNA- oder Proteinanalyse von Blutproben.
Die erforderlichen Volumina bewegen sich in der Regel
im Mikroliterbereich, doch manche microfluidic-Geräte
verarbeiten auch Nanoliter. Man könnte dieses Ver-
fahren nun der Nanotechnologie zuschreiben, doch es
ist zu beachten, dass Volumen die dritte Potenz eines
Längenmaßes ist. Ein Würfel mit der Kantenlänge von
1 μm (10–6 m) hat daher ein Volumen von 10–18 m3
oder 10–15 l (ein Femtoliter). Ein Nanoliter (10–9 l) ist
das Volumen eines Würfels mit einer Kantenlänge von
100 μm. Der Umgang mit Nanolitern ist daher keine
Nanotechnologie!
Ausblicke in die Zukunft reduzieren das Volumen von
flüssigen Proben auf einen echten Nanomaßstab – das
„Labor in der Zelle“. Vorstellbar ist ein Mikrochip, auf
dem mithilfe von einzelnen modifizierten Zellen Analysen
durchgeführt werden können. Diese Ideen befinden sich
noch in der Entwurfsphase, doch ihre Umsetzung könnte
in naher Zukunft gelingen, wenn die Entwicklung der Na-
notechnologie ihr rasantes Tempo beibehält.
Exkurs 7.3
zugegeben und die Rotation liefe, aus biologischer Sicht, rückwärts ab).
Kinesin und Dynein sind Motorproteine, die ATP als Energiequelle verwenden, um sich entlang der Mi-krotubuli eukaryotischer Zellen zu bewegen. Es han-delt sich daher um Linearschrittmotoren (Abb. 7.21). Ihre natürliche Funktion ist der Materialtransport.
Kinesin bewegt Frachten vom Zentrum zur Peri-pherie der Zelle, Dynein dagegen von der Peripherie zum Zentrum. Kinesin unternimmt dabei Schritte von 8 nm und vermag etwa 100 Schritte pro Sekunde auszuführen (das sind etwa 3 mm in der Stunde!). Je-der Schritt verbraucht ein ATP-Molekül. Die Mikro-tubuli, die sie als eine Art Gleis verwenden, bestehen

224 Nanobiotechnologie7
aus Proteinzylindern mit einem äußeren Durchmes-ser von 30 nm.
Proteine, die chemische und mechanische Energie
ineinander umwandeln, werden auf einen Einsatz
als molekulare Motoren für den Antrieb zukünftiger
Nanomaschinen getestet.
� Weiterführende Literatur
Bogunia-Kubik K, Sugisaka M (2002) From molecular bio-
logy to nanotechnology and nanomedicine. BioSystems
65: 123–138
Bianco A, Kostarelos K, Prato M (2005) Applications of
carbon nanotubes in drug delivery. Curr Opin Chem
Biol 9: 674–679
Choi B, Zocchi G, Wu Y, Chan S, Jeanne Perry L (2005) Al-
losteric control through mechanical tension. Phys Rev
Lett 95: 78–102
Hamad-Schifferli K, Schwartz JJ, Santos AT, Zhang S, Ja-
cobson JM (2002) Remote electronic control of DNA
hybridization through inductive coupling to an attached
metal nanocrystal antenna. Nature 415 152–155
Hartmann U (2006) Nanotechnologie. Spektrum Akademi-
scher Verlag, Heidelberg
Hess H, Bachand GD, Vogel V (2004) Powering nanodevices
with biomolecular motors. Chem Eur J 10: 2110–2116
Ilic B, Yang Y, Craighead HG (2004) Virus detection using
nanoelectromechanical devices. Appl Phys Lett 85: 27
Li JJ, Tan W (2002) A single DNA molecule nanomotor.
Nano Lett 2: 315–318
Liu D, Park SH, Reif JH, LaBean TH (2004) DNA nanotubes
self-assembled from triple-crossover tiles as templates
for conductive nanowires. Proc Natl Acad Sci USA 101:
717–722
Matsunaga T, Okamura Y (2003) Genes and proteins in-
volved in bacterial magnetic particle formation. Trends
Microbiol 11: 563–541
Niemeyer CM, Mirkin CA (2004) Nanobiotechnology: Con-
cepts, Applications and Perspectives. Wiley-VCH
Papazoglou ES, Parthasarathy A (2007) BioNanotechno-
logy (Synthesis Lectures on Biomedical Engineering).
Morgan and Claypool Publishers, San Rafael, CA
Patolsky F, Zheng G, Hayden O, Lakadamyali M, Zhuang X,
Lieber CM (2004) Electrical detection of single viruses.
Proc Natl Acad Sci USA 101: 14 017–14 022
Riegler J, Nann T (2004) Application of luminescent nanoc-
rystals as labels for biological molecules. Anal Bioanal
Chem 379: 913–919
Zhang S (2003) Fabrication of novel biomaterials through
molecular self-assembly. Nat Biotechnol 21: 1171–1178
Recommended

















