
Tierärztliche Hochschule Hannover
Physiologisches Institut
Bedeutung von Kalium und Insulin für die Motilität der
Labmagenmuskulatur
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer
Doktorin der Veterinärmedizin
- Doctor medicinae veterinariae -
(Dr. med. vet.)
vorgelegt von
Gesche Türck, geb. Onken
aus Hamburg
Hannover 2009

Wissenschaftliche Betreuung: PD Dr. S. Leonhard-Marek
1. Gutachter: PD Dr. S. Leonhard-Marek
2. Gutachter: Univ.-Prof. Dr. J. Rehage
Tag der mündlichen Prüfung: 18.05.2009
Diese Arbeit wurde mit finanziellen Mitteln der H. Wilhelm Schaumann Stiftung (Gesche Türck) und
der Deutschen Forschungsgemeinschaft (LE 824/4 und 5) durchgeführt.

Für Nils, Matti und die Hühner


Inhaltsverzeichnis
Inhaltsverzeichnis
1 Einleitung........................................................................................................... 11
2 Literaturübersicht............................................................................................... 13
2.1 Der Labmagen ........................................................................................... 13
2.1.1 Anatomie des Labmagens .................................................................. 13
2.1.2 Enterisches Nervensystem ................................................................. 14
2.1.3 Glatte Muskelzellen (Textus muscularis nonstriatus) .......................... 14
2.1.4 Kontraktionsmechanismus .................................................................. 15
2.1.5 Labmagenmotilität............................................................................... 16
2.1.5.1 Physiologie der Labmagenmotilität .............................................. 16
2.1.5.2 Labmagenverlagerung ................................................................. 17
2.2 Kalium ........................................................................................................ 18
2.2.1 Kaliumhomöostase.............................................................................. 18
2.2.1.1 Aufnahme und Elimination ........................................................... 19
2.2.1.2 Verteilung zwischen Intra- und Extrazellulärraum........................ 20
2.2.1.3 Imbalancen im Kaliumhaushalt .................................................... 21
2.2.1.3.1 Hypokaliämie ............................................................................ 21
2.2.1.3.2 Hyperkaliämie ........................................................................... 22
2.2.2 Regulation des Kaliumhaushaltes....................................................... 23
2.3 Insulin......................................................................................................... 25
2.3.1 Insulinhomöostase .............................................................................. 25
2.3.1.1 Struktur und Biosynthese von Insulin........................................... 26
2.3.1.2 Sekretion und Abbau von Insulin ................................................. 27
2.3.1.3 Signaltransduktion am Erfolgsorgan ............................................ 29
2.3.2 Insulinresistenzphänomen .................................................................. 30
2.3.3 Beeinflussung der Labmagenmotilität durch Insulin ............................ 36
2.4 Hypothese .................................................................................................. 37

Inhaltsverzeichnis
3 Material und Methoden...................................................................................... 38
3.1 In-vitro-Motilitätsmessung der Labmagenmuskulatur ................................. 38
3.1.1 Versuchstiere ...................................................................................... 38
3.1.2 Entnahme der Muskelstreifen.............................................................. 38
3.1.3 Präparation der Zirkulär- und Längsmuskelstreifen............................. 38
3.1.4 Messung der mechanischen Aktivität der Muskelstreifen.................... 39
3.1.4.1 Versuchsvorrichtung .................................................................... 39
3.1.4.2 Versuchsschemata ...................................................................... 40
3.1.4.3 Motilitätsparameter ...................................................................... 41
3.2 In-vivo-Versuch: Bestimmung des Labmageneffluxes bei intravenöser
Applikation von Insulin und Kalium ....................................................................... 41
3.2.1 Versuchstiere ...................................................................................... 41
3.2.2 Versuchsaufbau .................................................................................. 43
3.2.2.1 Vorbereitung der Versuchstiere ................................................... 43
3.2.2.2 Vorbereitung der Infusionslösungen ............................................ 43
3.2.2.3 Ablauf des hyperinsulinämischen, euglykämischen Clamptests mit
gleichzeitiger Labmageneffluxmessung......................................................... 44
3.2.3 Analysen ............................................................................................. 45
3.2.3.1 Insulin........................................................................................... 45
3.2.3.2 Kalium.......................................................................................... 46
3.2.3.3 Chrom .......................................................................................... 46
3.2.3.4 Berechnung des Labmageneffluxes............................................. 47
3.3 Statistik....................................................................................................... 49
4 Ergebnisse ........................................................................................................ 50
4.1 In-vitro-Versuch.......................................................................................... 50
4.1.1 Aktivität der circulären Corpusmuskulatur........................................... 50
4.1.1.1 Basale Aktivität der circulären Corpusmuskulatur........................ 50
4.1.1.2 Wirkung steigender Kaliumgaben auf die circuläre
Corpusmuskulatur.......................................................................................... 53
4.1.1.3 Wirkung steigender Insulingaben auf die circuläre
Corpusmuskulatur.......................................................................................... 62

Inhaltsverzeichnis
4.1.1.4 Wirkung von Insulin bei steigenden Kaliumgaben auf die circuläre
Corpusmuskulatur.......................................................................................... 67
4.1.1.5 Wirkung von Insulin in Anwesenheit von Barium ......................... 71
4.1.1.6 Wirkung von Insulin in Anwesenheit von Glybenclamid ............... 74
4.1.1.7 Wirkung von Insulin in Anwesenheit von Ouabain ....................... 76
4.1.2 Aktivität der circulären Pylorusmuskulatur .......................................... 78
4.1.2.1 Basale Aktivität der circulären Pylorusmuskulatur ....................... 78
4.1.2.2 Wirkung steigender Kaliumgaben auf die circuläre
Pylorusmuskulatur ......................................................................................... 81
4.1.2.3 Wirkung steigender Insulingaben auf die circuläre
Pylorusmuskulatur ......................................................................................... 89
4.1.3 Aktivität der longitudinalen Corpusmuskulatur .................................... 97
4.1.3.1 Wirkung steigender Insulingaben auf die longitudinale
Corpusmuskulatur.......................................................................................... 97
4.2 In-vivo-Versuch .......................................................................................... 99
4.2.1 Insulinkonzentrationen ...................................................................... 100
4.2.2 Kaliumkonzentrationen...................................................................... 101
4.2.3 Labmagenefflux................................................................................. 104
5 Diskussion ....................................................................................................... 107
5.1 In-vitro-Versuch........................................................................................ 107
5.1.1 Reaktionen isolierter Labmagenmuskulatur auf Änderungen der
extrazellulären Kaliumkonzentration................................................................ 108
5.1.2 Reaktionen isolierter Labmagenmuskulatur auf Änderungen der
extrazellulären Insulinkonzentration ................................................................ 110
5.1.2.1 Keine Reaktion der Muskulatur auf Insulingaben....................... 111
5.1.2.2 Stimulation der Muskulatur durch Insulingaben ......................... 111
5.1.2.3 Hemmung der Muskulatur durch Insulingaben........................... 111
5.1.2.4 Mechanismen der Insulinhemmung ........................................... 113
5.1.2.4.1 Wirkung von Kalium in Anwesenheit von Insulin..................... 113
5.1.2.4.2 Wirkung von Insulin in Anwesenheit von Barium .................... 114
5.1.2.4.3 Wirkung von Insulin in Anwesenheit von Glybenclamid.......... 115

Inhaltsverzeichnis
5.1.2.4.4 Wirkung von Insulin in Anwesenheit von Ouabain .................. 116
5.1.2.4.5 Zusammenfassung der möglichen Mechanismen................... 116
5.2 In-vivo-Versuch ........................................................................................ 117
5.2.1 Insulinkonzentrationen ...................................................................... 118
5.2.2 Kaliumkonzentrationen...................................................................... 119
5.2.3 Efflux ................................................................................................. 121
5.2.3.1 Diskussion der Methode............................................................. 121
5.2.3.2 Diskussion der Ergebnisse......................................................... 121
6 Schlussfolgerungen......................................................................................... 124
7 Zusammenfassung.......................................................................................... 126
8 Summary ......................................................................................................... 129
9 Literaturverzeichnis ......................................................................................... 131
10 Anhang............................................................................................................ 168
10.1 Versuchsschemata................................................................................... 168
10.1.1 Kalium ............................................................................................... 168
10.1.2 Insulin................................................................................................ 169
10.1.3 Weiterführende Untersuchungen ...................................................... 170
10.2 Verlauf der Blutglukosekonzentration im In-vivo-Versuch ........................ 172
10.3 Verwendete Substanzen .......................................................................... 172
10.3.1 In-vitro-Versuch................................................................................. 172
10.3.2 In-vivo-Versuch ................................................................................. 173
11 Danksagung .................................................................................................... 175

Abkürzungsverzeichnis
Abkürzungsverzeichnis
°C Grad Celcius
µg Mikrogramm
µl Mikroliter
µU Mikrounits
a. p. ante partum
ATP Adenosin 5'-Triphosphat
C Kohlenstoff
Ca2+ Kalzium
CaCl2 Kalziumchlorid
Cl- Chlor
cm Zentimeter
CO2 Kohlenstoffdioxid
Cr3+ Chrom
CrCl3 Chromchlorid
d Tag
dest. destilliert
EDTA Ethylendiamintetraessigsäure
ENS Enterisches Nervensystem
g Gramm
GLUT Glucosetransporter
Glyb. Glybenclamid
ICC Interstitielle Zellen nach Cajal
Ins. Insulin
h Stunde
h2 Heritabilität
H2SO4 Schwefelsäure
HCl Salzsäure
HCO3- Bicarbonat
I. E. Internationale Einheiten

Abkürzungsverzeichnis
K+ Kalium
KCl Kaliumchlorid
kg Kilogramm
KGW Körpergewicht
l Liter
LMV Labmagenverlagerung
MgCl2 Magnesiumchlorid
min Minute
ml Milliliter
mM Millimolar
mmol Millimol
mN Millinewton
mU Milliunits
Na+ Natrium
NaCl Natriumchlorid
NaHCO3 Natriumbicarbonat
NaH2PO4 Natriumdihydrogenphosphat
NEFA Non esterified fatty acid
Ouab. Ouabain
O2 Sauerstoff
p.p. post partum
rpm rounds per minute
SGK 1 serum and glucocorticoid-inducible kinase
SSIK steady-state Insulinkonzentration
U Units
VIP Vasoaktives Intestinales Polypeptid
ZNS Zentralnervensystem

Einleitung 11
1 Einleitung
Die Labmagenverlagerung ist eine der häufigsten und bedeutsamsten Erkrankungen
hochleistender Milchkühe in der Frühlaktation. Aufgrund von Tierarztkosten,
Milcheinbußen, Fruchtbarkeitsstörungen und Tierverlusten entstehen zum Teil
erhebliche wirtschaftliche Schäden (GEISHAUSER et al. 2000). Die Pathogenese
dieser Erkrankung ist bislang nur unzureichend geklärt. Neben verschiedenen
anderen Faktoren wird eine Beteiligung von Imbalancen in der Kalium- und
Insulinhomöostase an der Entstehung der Labmagenverlagerung diskutiert.
Kalium wird mit der Nahrung aufgenommen und v. a. über die Niere, aber auch über
die Milch abgegeben. Es spielt eine wichtige Rolle in der Entstehung und
Aufrechterhaltung des Ruhemembranpotentials und ist somit maßgeblich an der
Erregbarkeit von Muskeln und Nerven beteiligt. Während hohe
Kaliumkonzentrationen schnell zum Tod des Tieres führen, kommt es durch lang
andauernde niedrige Kaliumwerte im Blut zu einer schlechten Entwicklung und einer
geringen Leistungsbereitschaft, ohne dass bereits die typischen Symptome einer
Hypokaliämie, wie z. B. Muskellähmungen auftreten (ROBERTS u. OMER 1965;
TELLE et al. 1964). Niedrige Blutkaliumspiegel stellen sich v. a. bei reduzierter oder
absolut eingestellter Futteraufnahme ein (RABINOWITZ et al. 1988), was u. a.
infolge von Labmagenverlagerungen beobachtet wird, treten aber auch im
Zusammenhang mit hohen Insulinkonzentrationen im Blut auf, da Insulin die
Aufnahme von Kalium in die Körperzellen fördert (VAN MEIRHAEGHE 1988).
Hyperinsulinämien werden sehr häufig im Zusammenhang mit
Labmagenverlagerungen beobachtet (OK et al. 2000). Ob die Insulinwerte allerdings
bereits vor der Labmagenverlagerung ansteigen oder dies eine Folge der Erkrankung
ist, ist noch nicht abschließend geklärt. Fest steht jedoch, dass Insulin nicht nur einen
hemmenden Einfluss auf den Labmagenefflux hat (HOLTENIUS et al. 2000),
sondern dass es auch bei Kühen mit anhaltend hohen Insulinspiegeln post
operationem zu verlängerten Motilitätsstörungen des Labmagens kommt
(PRAVETTONI et al. 2004).

12 Einleitung
Aufgrund der postpartalen und in besonderem Maße mit Labmagenverlagerungen
beobachteten Kalium- und Insulinimbalancen und der gegenseitigen Beeinflussung
dieser Faktoren ist es das Ziel dieser Arbeit, den Einfluss von Insulin und Kalium auf
die Motilität der Labmagenmuskulatur durch Untersuchungen in-vitro und in-vivo
darzustellen und zu klären, inwiefern eine mögliche hemmende Wirkung des Insulins
auf veränderte Kaliumgradienten an der glatten Muskulatur zurückzuführen ist.

Literaturübersicht 13
2 Literaturübersicht
2.1 Der Labmagen
2.1.1 Anatomie des Labmagens
Der rechts vom Pansen gelegene Labmagen besteht aus drei Teilen: Fundus,
Corpus und Pars pylorica. Während sich der dilatierte Fundus nahezu in der
Medianen direkt kaudal der Haube befindet, schiebt sich das eher konische Corpus
hinter und unter den Blättermagen und wendet sich nach rechts. Die Pars pylorica
zieht im Bereich des rechten Rippenbogens nach dorsal und geht schließlich am
Musculus sphincter pylori in das Duodenum über (VOLLMERHAUS u. ROOS 1999).
Über das Omentum minus ist der Labmagen mit der ventralen, über das Omentum
majus mit der dorsalen Bauchwand verbunden (KÖNIG et al. 1999). Die
Labmagenwand besteht aus fünf verschiedenen Schichten:
• Tunica serosa
• Tela subserosa
• Tunica muscularis
- Longitudinale Muskulatur
- Zirkuläre Muskulatur
• Tela submucosa
• Tunica mucosa (LIEBICH 1999).
Die Tunica muscularis besteht aus zwei Lagen glatter Muskulatur. Die äußere
Längsmuskulatur zieht vom Blättermagen her den gesamten Labmagen entlang, wird
in der Pars pylorica kräftiger und geht letztlich nahtlos ins Duodenum über. Auch die
innere Ringmuskulatur ist Bestandteil aller drei Labmagenabschnitte und bildet,
bevor auch sie in den Dünndarm übergeht, den Musculus sphincter pylori
(VOLLMERHAUS u. ROOS 1999). Die Innervation des Labmagens ist vegetativ. Der
Plexus coeliacus entlässt sympathische Fasern, die u. a. den Plexus gastricus
bilden, der die sympathische Versorgung des Labmagens übernimmt. Die
parasympathischen Äste entstammen dem Nervus vagus und zwar sowohl aus dem
dorsalen als auch aus dem ventralen Truncus vagalis (KÖNIG et al. 1999).

14 Literaturübersicht
2.1.2 Enterisches Nervensystem
Das in der Wand des Magen-Darmtrakts lokalisierte enterische Nervensystem
verfügt über eigene sensorische Neurone, Interneurone und Motoneurone, so dass
es in der Lage ist, Reflexe (wie z. B. den Transport des Chymus durch Kontraktion
oral gelegener Muskulatur bei gleichzeitiger aboraler Relaxation) unabhängig vom
ZNS oder dem vegetativen Nervensystem zu vermitteln (GERSHON et al. 1994).
Sensorische Neurone registrieren über Chemo- oder Mechanorezeptoren
verschiedene Stimuli wie Nährstoffgehalt, Osmolarität oder pH-Wert bzw.
Wandspannung, intraluminale Drücke, Scherreize oder Volumenänderungen (WOOD
1994). Vermutlich kodieren dabei afferente Nervenzellen lediglich die sensorischen
Stimuli, während die Aufgabe der eigentlichen Sensorfunktion anderen Zellen, z. B.
enterochromaffinen Zellen, obliegt (SCHEMANN 2000). Interneurone stellen die
Kommunikation innerhalb des enterischen Nervensystems sicher (WOOD 1994). Die
hemmenden und erregenden Motoneurone schließlich modulieren Effektoren, wie die
glatte Muskulatur (vor allem über ICC, interstitielle Zellen nach Cajal), sekretorische
Zellen oder Blutgefäße (WOOD 1994). Bis heute sind über 20 Neurotransmitter und
noch mehr Neuromodulatoren identifiziert, die an der Vermittlung der synaptischen
Vorgänge im enterischen Nervensystem beteiligt sind (MC CONALOGUE u.
FURNESS 1994).
2.1.3 Glatte Muskelzellen (Textus muscularis nonstriatus)
Die glatte Muskulatur erhielt ihren Namen durch das Fehlen einer erkennbaren
geordneten Myofibrillenstruktur. Bei den in der Labmagenwand vorkommenden
glatten Muskelzellen handelt es sich um den „Single-unit“-Muskeltyp, bei dem die
Zellen über sogenannte Nexus (gap junctions) in direkter Verbindung stehen
(LIEBICH 1999).
Der Zellkern befindet sich stets zentral im Sarkoplasma, das neben zahlreichen
Sarkosomen auch ein unterschiedlich stark ausgebildetes sarkoplasmatisches
Retikulum enthält, das als Kalziumspeicher dient (LIEBICH 1999). Nicht kontraktile
intermediäre Mikrofilamente (Desmin) bilden das Zytoskelett der Zelle. Die
Myofilamente (Aktin-, Myosin- und Intermediärfilamente) verlaufen netzartig ohne

Literaturübersicht 15
erkennbare regelmäßige Anordnung. Während die Aktin- und Intermediärfilamente
an sogenannten Haftplatten inserieren, bestehen die Myosinfilamente aus löslichen
Proteinen, die sich während der Kontraktion verdichten, so dass die Aktinfilamente
über sie hinweggleiten können (RÜEGG 2000). Auf der Oberfläche der glatten
Muskelzelle befindet sich ein Netz aus überwiegend Kollagen- und Retikulinfasern, in
dem auch Fasern des ENS verlaufen. Diese ziehen direkt an das Plasmalemm der
Muskelzelle, die auf der Zelloberfläche eine Vielzahl von adrenergen und cholinergen
Rezeptoren aufweist.
2.1.4 Kontraktionsmechanismus
Bei einer Erregung der glatten Muskelzelle erhöht sich die intrazelluläre
Kalziumkonzentration einerseits transient durch einen Einstrom von Kalzium über
rezeptorgesteuerte Kanäle und durch eine IP3-vermittelte Freisetzung von
Kalziumionen aus dem sarkoplasmatischen Retikulum und andererseits langfristig
durch Öffnen spannungsabhängiger Kalziumkanäle. Kalmodulin bindet das Kalzium
und kann nun zum einen die Myosin-Kinase, die ihrerseits durch Phosphorylierung
der Myosinfilamente die Myosin-ATPase aktiviert, und zum anderen eine
Proteinkinase aktivieren, die das Hemmprotein Caldesmon der Myosinbindungsstelle
des Aktins phosphoryliert und somit eine Bindung an den Kalzium-Kalmodulin-
Komplex ermöglicht (RÜEGG 2000). In der Folge kommt es zu einer Interaktion
zwischen Aktin und Myosin, wobei die so gebildeten Myosinquerbrücken unter ATP-
Spaltung zyklisch tätig sind. Die Trennung der Filamente erfolgt durch Abkopplung
der an das Myosin angehängten Phosphatgruppen durch die Myosinphosphatase.
Bei der Muskelrelaxation wird Kalzium in das sarkoplasmatische Retikulum
zurückgepumpt und durch ATP-getriebene Kalziumpumpen und Na+/Ca2+-
Austauscher in den Extrazellulärraum befördert (SOMLYO u. SOMLYO 1994).
Bei den glatten Muskelzellen vom single-unit-Typ entstehen die zur Kontraktion
führenden Aktionspotentiale nicht neurogen sondern myogen in sogenannten
Schrittmacherzellen, den interstitiellen Zellen nach Cajal (ICC) (SANDER 1996). Man
unterscheidet zwei ICC-Typen: ICC-MY, die zwischen Zirkulär- und Längsmuskulatur
anzutreffen sind, und die ICC-IM, die innerhalb der Muskelbündel liegen (BURNS et
al. 1995). ICC haben kein gleichbleibendes Ruhepotential sondern weisen langsame

16 Literaturübersicht
rhythmische Schwankungen auf (SANDER 1996). Verantwortlich für die Entstehung
dieser sogenannten slow waves ist die Erhöhung der intrazellulären
Kalziumkonzentration, die u. a. auf einer Freisetzung aus den Mitochondrien beruht,
da eine Hemmung mitochondrialer Aktivität die Frequenz der slow waves reduziert
(SUZUKI et al. 2006). Slow waves führen bei den ICC zu schnellen Depolarisationen
mit anschließender Plateauphase, aber nicht zwangsläufig zu Kontraktionen der
glatten Muskelzellen. Allerdings konnte eine positive Korrelation zwischen der
Amplitudenhöhe der slow waves und dem Auftreten von Kontraktionen der zirkulären
Muskelschicht festgestellt werden (THUNEBERG 1989). Kommt es jedoch
Acetylcholin-vermittelt zu einem extrazellulären Kalziumeintritt in die
Schrittmacherzelle, folgt eine Depolarisation mit einem weiteren Einstrom von
Kalzium durch potentialgesteuerte Kanäle, was zu einer Erhöhung und Verbreiterung
des Plateaus führt. Die Dauer des Plateaus bestimmt die Dauer der Kontraktion
(EHRLEIN 2000).
Über die gap junctions (Nexus) genannten niederohmigen Zellkontakte kann sich die
Depolarisation einer bereits erregten Zelle elektrotonisch auf benachbarte
Muskelzellen übertragen (KOMURO et al. 1999). Sobald deren Membran durch den
lokal über die gap junctions fließenden Strom bis zur Schwelle der
spannungsabhängigen Kalziumkanäle depolarisiert ist, erfolgt ein Aktionspotential,
das daraufhin weitere elektrotonisch gekoppelte Muskelzellen erregt (RÜEGG 2000).
2.1.5 Labmagenmotilität
2.1.5.1 Physiologie der Labmagenmotilität
Unter dem Aspekt der motorischen Funktion wird am Labmagen ein Magenspeicher
und eine Magenpumpe unterschieden. Die Motorik des Labmagens besteht aus
Spannungsänderungen des Magenspeichers und peristaltischen Wellen, die am
Magenkörper beginnen und zum Pylorus hin verlaufen. Im Bereich des
Magenspeichers sind sie jedoch nur schwach an der Oberfläche bemerkbar, an der
Magenpumpe hingegen werden sie zu stärkeren Einschnürungen, die sich in Ringen
auf den Pylorus zubewegen (PFEFFER 1987). Bei dem Entleerungsmechanismus
der Magenpumpe können drei Phasen unterschieden werden:

Literaturübersicht 17
1. Phase des Vorschubs: während des Ablaufs der peristaltischen Wellen über
die proximale Magenpumpe erschlafft der zuvor kontrahierte distale Abschnitt
(EHRLEIN 2000).
2. Phase der Entleerung und Durchmischung: Erreichen die Wellen die Mitte der
Magenpumpe, öffnet der M. sphincter pylori reflektorisch, während er sich bei
Ankunft der Kontraktionen am Sphincter bereits verschlossen hat (HILL 1969),
der Mageninhalt wird also nicht in den Darm gepresst, sondern gespült.
3. Phase der Rücktreibung und Zerkleinerung: durch den nicht vollständigen
Verschluss am mittleren Abschnitt der Magenpumpe, kommt es zu einem
Rückfluss von Chymus in den proximalen Teil (EHRLEIN 2000).
Diese Phasen treten aufgrund der den peristaltischen Wellen zu Grunde liegenden
slow waves zwar im Allgemeinen zyklisch ein, werden jedoch in erster Linie durch
vago-vagale Reflexschaltkreise moduliert (COTTRELL u. STANLEY 1992;
COTTRELL 1994). Die abomasale Effluxmenge ist durch Auslösung dieser Reflexe
u. a. abhängig von der Relaxation des Magenspeichers, der Einschnürungstiefe der
peristaltischen Wellen, der Weite des Pylorus, der rezeptiven Erschlaffung des
Duodenums, der Art der Duodenalkontraktionen und der Intensität der Fütterung
(MALBER u. RUCKEBUSCH 1991; LESTER u. BOLTON 1994). Zu einem
verzögerten Übertritt von Chymus ins Duodenum kommt es z. B. durch eine
langandauernde Erschlaffung des Magenspeichers, eine geringe Einschnürungstiefe
der Kontraktionen, eine ungenügende Relaxation von M. sphincter pylori oder
Duodenum und eine geringe Aktivität der duodenalen Muskelkontraktionen, die den
Chymus von oral nach aboral transportieren (EHRLEIN 2000). Neben der Aktivität
der Muskulatur des Magens selber ist also auch die des Dünndarms von Bedeutung
für die Entleerung des Magens (VAUPEL 2000).
2.1.5.2 Labmagenverlagerung
Das Krankheitsbild der Labmagenverlagerung wurde 1950 das erste Mal an einem
sechs Jahre alten Shorthorn-Rind (FORD 1950) beschrieben und tritt seither
zunehmend häufiger vor allem bei hochleistenden Milchrindern in den ersten
Wochen nach der Abkalbung auf (CONSTABLE et al. 1992). Als wesentliche, die
Entstehung der Labmagenverlagerung bedingende Voraussetzung gilt eine

18 Literaturübersicht
abomasale Gasansammlung in Kombination mit einer Atonie der
Labmagenmuskulatur bei ausreichendem abdominalen Platzangebot (z. B.
großrahmige Tiere) (GEISHAUSER 1995; STÖBER u. SARATSIS 1974). Die
ursächlichen Faktoren für die Entstehung von Gas und verminderter Motorik sind
jedoch bislang nur unzureichend geklärt, vermutet wird ein multifaktorielles
Geschehen. Als mögliche Auslöser der Atonien der Labmagenmuskulatur werden
neben hohen Plasmakonzentrationen an kurzkettigen Fettsäuren (LE BLANC et al.
2005; ZADNIK 2003), Ketonkörpern (ROHRBACH et al. 1999), Hypokalzämien
(DANIEL 1983), Endotoxinen (ROHRBACH et al. 1999; VLAMINCK et al. 1985),
Alkalosen (POULSEN 1976) und Hyperglykämien (HOLTENIUS et al. 2000) auch
hohe Insulinkonzentrationen im Zuge von Insulinresistenzen diskutiert (VAN
MEIRHAEGHE et al. 1988 a). Außerdem werden im Zusammenhang mit
Labmagenverlagerungen häufig niedrige Kaliumkonzentration im Blut nachgewiesen
(ESPERSEN u. SIMENSEN 1961), die durch reduzierte Nahrungsaufnahme
(RABINOWITZ et al. 1988), hohe Insulinkonzentrationen (VAN MEIRHAEGHE 1988)
und/oder abomasalen Reflux mit folgender Alkalose bedingt sein können (ROSSOW
1995).
Die Rollen von Kalium und Insulin im Stoffwechselgeschehen mit Hinblick auf eine
mögliche Beteiligung an der Pathogenese der Labmagenverlagerung werden im
Folgenden ausgeführt.
2.2 Kalium
2.2.1 Kaliumhomöostase
Kalium ist ein starker Elektrolyt, der in wässriger Lösung vollständig ionisiert vorliegt
und aufgrund seiner geringen Tendenz, Komplexe zu bilden, im Körper vorwiegend
dem Transport von Ladungen dient. Als monovalentes Kation spielt es eine
bedeutende Rolle in der Entstehung und Aufrechterhaltung des
Ruhemembranpotentials und somit in der neuromuskulären Erregbarkeit (PETRIDES
1997).
Die Referenzwerte der Plasma-Kaliumkonzentrationen beim Rind werden mit 3,5 bis
5,0 mmol/l (VETMEDLABOR 2007; SEJERSTED u. SJOGAARD 2000), bzw. mit 4,0

Literaturübersicht 19
bis 5,0 mmol/l (STÖBER u. GRÜNDER 1990) angegeben. Allerdings befinden sich
nur ca. zwei Prozent des Kaliums in der extrazellulären Flüssigkeit, so dass die
Kaliumwerte im Plasma nicht unbedingt die Gesamtsituation im
Stoffwechselgeschehen wiederspiegeln (BROBST 1986). Die große Mehrheit des
Kaliumgehaltes im Körper befindet sich im intrazellulären Bereich, vor allem in den
Skelettmuskelzellen, wo Kalium in Konzentrationen von ungefähr 120 – 140 mmol/l
vorliegt (SEJERSTED u. SJOGAARD 2000). Demzufolge können Diffusionsprozesse
den Kaliumgehalt im Plasma grundsätzlich beeinflussen. Die Konzentration von
Kalium im Plasma wird also durch zwei Komponenten bestimmt: die äußere (die
Differenz zwischen Aufnahme und Elimination) und die innere Kaliumbalance (die
Verlagerung der Ionen zwischen Intra- und Extrazellulärflüssigkeit) (BROBST 1986).
2.2.1.1 Aufnahme und Elimination
Kalium wird mit der Nahrung aufgenommen, wobei der Gehalt im Futter bei
laktierenden Kühen 0,8 % und bei güsten Tieren 0,5 % in der Trockenmasse
betragen sollte. Bei Nicht-Wiederkäuern wird über 85 % des mit der Nahrung
aufgenommenen Kaliums resorbiert, vorwiegend im Dünndarm und parazellulär
mittels solvent drag aufgrund der großen Durchlässigkeit der Zonulae occludentes
(FROMM u. HIERHOLZER 2000). Auch bei Kühen werden zwischen 72 und 97 %
des aufgenommenen Kaliums resorbiert (KHORASANI et al 1997; BANNINK et al.
1999), wobei bei hohem Kaliumangebot im Futter auch präduodenale Bereiche zur
Resorption beitragen können. Kalium wird bei laktierenden Kühen zu 75 % mit dem
Urin, zu 13 % mit dem Kot und zu 12 % mit der Milch ausgeschieden (WARD 1966),
bei nicht laktierenden Schafen werden 90 % des aufgenommenen Kaliums über die
Nieren ausgeschieden (DEWHURST et al. 1968). Die Höhe der Kaliumexkretion
steht in engem Zusammenhang mit der Nahrungsaufnahme. RABINOWITZ et
al. (1988) konnten zeigen, dass es bei Schafen nach der Fütterung zu einer
gesteigerten Kaliumausscheidung kam, wobei die Kaliumkonzentration im Urin umso
größer war, je mehr Kalium mit dem Futter aufgenommen wurde. An Tagen, an
denen die Tiere nicht gefüttert wurden, blieb der Peak in der Kaliumexkretion aus.
In der Niere wird Kalium in den Glomerula filtriert und der größte Teil im proximalen
Tubulus und der Henle-Schleife wieder resorbiert, um schließlich in die Lumina der

20 Literaturübersicht
distalen Tubuli und der Sammelgänge sezerniert zu werden. Die Exkretion ist ein
aktiver ATP-abhängiger Austausch gegen Natrium bzw. Wasserstoff (SWEENEY
1999).
2.2.1.2 Verteilung zwischen Intra- und Extrazellulärraum
Eine wichtige Voraussetzung für alle Lebensvorgänge ist die Aufrechterhaltung einer
hohen extrazellulären Natrium- sowie einer hohen intrazellulären
Kaliumkonzentration. Da die Zellmembran für geladene Ionen nur schwer permeabel
ist, wird diese Ionenverteilung durch die Aktivität der Na+/K+-ATPase erreicht, die den
aktiven Transport von drei Natrium- und zwei Kaliumionen durch die Zellmembran
katalysiert. Bei dieser Ionenpumpe handelt es sich um ein tetrameres Protein mit
zwei α- und zwei β-Untereinheiten. Erstere enthalten die Bindungsstelle für Natrium,
Kalium und ATP (STRYER 1999). Da sowohl die α- als auch die β-Untereinheiten in
drei Subtypen vorkommen, entsteht eine Familie von Isoenzymen, die sich
unterscheiden durch:
- ihre Affinität zu Natriumionen
- ihre hormonelle Regulierbarkeit (z. B. durch Schilddrüsenhormone, Insulin
oder Glucocorticoide) und
- ihre zelluläre Lokalisation (PETRIDES 1997).
Aus dem Zellinneren kann Kalium über verschiedene Kanäle in den
Extrazellularraum diffundieren. Kaliumkanäle stellen Homo- oder Heterotetramere
dar. Die Monomeruntereinheiten werden von mindestens zehn Genen kodiert, von
denen einzelne zusätzlich durch Spleißing unterschiedliche Domänenstrukruren
hervorrufen können. Durch vielfältige Kombinationen dieser unterschiedlichen
Genprodukte entsteht eine Vielzahl verschiedener Kaliumkanal-Isoformen (STRYER
1999). Damit ist die lokale Verteilung von Kaliumionen abhängig von der Anflutung
mit dem Blut, der Aktivität der Na+/K+-ATPase und der Offenwahrscheinlichkeit und
Leitfähigkeit der vorhandenen Kaliumkanäle.

Literaturübersicht 21
2.2.1.3 Imbalancen im Kaliumhaushalt
Störungen im Kalium-Gleichgewicht zwischen Aufnahme und Elimination und
zwischen Intra- und Extrazellulärraum können zu Hypo- und Hyperkaliämien führen,
ohne dass der absolute Kaliumgehalt im Körper stark verändert ist. Insbesondere
Störungen des Säure-Basen-Haushaltes und Hunger beeinflussen die
Kaliumhomöostase (BROBST 1986; LUNN u. MC GUIRK 1990). Abbildung 1 gibt
dazu eine Übersicht.
2.2.1.3.1 Hypokaliämie
Hypokaliämien durch Imbalancen im extrazellulären Raum können zum einen durch
eine verminderte Kaliumaufnahme und zum anderen durch eine forcierte
Ausscheidung entstehen. Eine zu geringe Kaliumkonzentration im Futter ist sehr
selten und kann lediglich bei hochleistenden Milchkühen vorkommen, die
überwiegend konzentratreiche Nahrung erhalten (WARD 1966). Weit häufiger sind
niedrige Blutkaliumwerte im Zusammenhang mit unzureichender Futteraufnahme
(z. B. durch Krankheit) zu beobachten. Insbesondere Tiere, die an kaliumreiche
Nahrung adaptiert sind, können bei plötzlich ausbleibender Kaliumaufnahme die
Ausscheidung nicht schnell und weit genug herunterregulieren, so dass es zur
Entstehung von Hypokaliämien kommt. Diese sind allerdings in der Regel nicht so
K+ K+
K+
Intracellular Potassium Extracellular Potassium Acidosis Excretion
Alkalosis
Dietary intake
Abbildung 1: Faktoren, die das intra- und extrazell uläre Kalium-Gleichgewicht
beeinflussen (SWEENEY 1999)

22 Literaturübersicht
stark ausgeprägt, dass sie zu klinischen Symptomen führen (SWEENEY 1999).
Passagestörungen im Magendarmtrakt, wie sie unter anderem bei
Labmagenverlagerungen auftreten, ziehen ebenfalls durch eine verminderte
Resorptionsrate im Dünndarm geringe Blutkaliumspiegel nach sich (DELGADO-
LECAROZ et al. 2000; TAGUCHI 1995).
Auch Gaben von Diuretika oder Niereninsuffizienzen können durch eine
unzureichende Rückresorption des in der Niere filtrierten Kaliums zu einer
Hypokaliämie führen (vermehrte Ausscheidung) (BROBST 1986; LUNN u. MC
GUIRK 1990). Im Zuge von metabolischen Alkalosen kommt es ebenfalls zu einem
Absinken der Kaliumspiegel im Blut (WARD et al. 1994) durch eine verstärkte
Aufnahme von Kalium in die Zelle. Wasserstoff wird dabei im Antiport gegen Natrium
aus der Zelle transportiert, welches wiederum über die Na+/K+-ATPase die Zellen
verlässt. Auf gleichem Wege entsteht im Nierenepithel der distalen Tubuli zusätzlich
ein größerer Diffusionsgradient für Kalium. Durch die zusätzlich verminderte Aktivität
des K+/H+-Antiporters, um die Sekretion der H+-Ionen zu reduzieren, kommt es
schließlich zu einer erhöhten Kaliumausscheidung (LUNN u. MC GUIRK 1990; MC
GUIRK u. BUTLER 1980).
Auf zellulärer Ebene haben geringe Kaliumspiegel im Plasma und der extrazellulären
Flüssigkeit ein vermehrtes Ausströmen von Kalium aus der Zelle über die
verschiedenen Kaliumkanäle zur Folge, so dass die Zelle hyperpolarisiert. Ein
Erreichen des Schwellenwertes zur Öffnung spannungsabhängiger Natrium-
(Nerven- und Skelettmuskelzelle) oder Kalziumkanäle (glatte Muskelzelle), das zur
Auslösung von Aktionspotentialen und Muskelkontraktionen führt, ist damit seltener
(PETRIDES 1997).
Klinisch äußert sich eine Hypokaliämie demzufolge in erster Linie in
Herzrhythmusstörungen und Skelettmuskelschwäche, die letztlich zu Festliegen in
autauskultatorischer Haltung führt (SIELMAN et al. 1997).
2.2.1.3.2 Hyperkaliämie
Bei ausreichender Wasseraufnahme und funktionstüchtigen Nieren ist der
Wiederkäuerstoffwechsel vor einer Hyperkaliämie durch eine schnell
anpassungsfähige renale Kaliumausscheidung gut geschützt (SWEENEY 1999).

Literaturübersicht 23
Daher liegen in den meisten Fällen hoher Kaliumkonzentrationen im Blut eine
eingeschränkte Nierentätigkeit oder eine verminderte Aufnahme von Kalium in die
Zellen vor. Azidosen z. B. werden im Körper unter anderem dadurch abgepuffert,
dass bei hohen H+-Konzentrationen im Blut die Aktivität der Na+/H+-Transporter und
im weiteren Verlauf auch die der Na+/K+-ATPasen reduziert ist, so dass weniger
Kalium in die Zellen aufgenommen wird (TREMBLAY 1990).
Generell ist bei hohen Kaliumkonzentrationen in der extrazellulären Flüssigkeit die
Diffusion von Kalium aus der Zelle durch die Kaliumkanäle vermindert. Das
Zellinnere und damit das Membranpotential wird aufgrund dessen positiver, so dass
das Schwellenpotential eher erreicht wird (PETRIDES 1997). Da jedoch die
spannungsabhängigen Natrium- bzw. Kalziumkanäle nach ihrer Öffnung und
zeitabhängigen Schließung zunächst refraktär sind, haben Hyperkaliämien im
Endeffekt ähnliche klinische Symptome zur Folge wie Hypokaliämien, bradykarde
Herzrhythmusstörungen stehen allerdings zunächst im Vordergrund (SWEENEY
1999). Bei Kühen führte eine experimentelle orale Applikation von 650 g KCl zum
Tod der Tiere (DENNIS u. HARBAUGH 1948).
2.2.2 Regulation des Kaliumhaushaltes
Zur Vermeidung starker, ggf. lebensbedrohlicher Schwankungen der Blutkaliumwerte
wird der (extrazelluläre) Kaliumhaushalt streng reguliert, insbesondere und primär
über die Niere. Dabei scheint es zusätzlich zum klassischen Feedback-System bei
der Regulation der Kaliumhomöostase einen weiteren Mechanismus zu geben, der
einen Anstieg des Blutkaliums nach einer Mahlzeit verhindert: Postprandial kommt es
zu einem Anstieg der renalen Kaliumexkretion, wobei genau die Menge
ausgeschieden wird, die zuvor mit der Nahrung aufgenommen wurde (YOUN u.
MCDONOUGH 2009). Wie dieses „Feedforward-System“ genau funktioniert, ist
ungeklärt. Denkbar wären Sensoren im Magendarmtrakt, die die resorbierte
Kaliummenge registrieren und in entsprechendem Maße die renale Exkretion
verstärken (YOUN u. MCDONOUGH 2009), nicht aber die Ausscheidungsmenge mit
dem Kot verändern (ANDERSON u. PICKERING 1962; DEWHURST et al. 1968).
Wahrscheinlich ist diese stimulierte Ausscheidung über die Niere nicht auf das
Mineralocorticoid Aldosteron zurückzuführen (GREEN u. RABINOWITZ 1979;

24 Literaturübersicht
RABINOWITZ et al. 1984 a; RABINOWITZ u. SARASON 1980; RABINOWITZ,
SARASON u. YAMAUCHI 1981). Zwar verstärkt Aldosteron im distalen Tubulus die
Kaliumsekretion durch Stimulation der Na+/K+-ATPase (SWEENEY 1999), und auch
RABINOWITZ et al. (1985) zeigten einen potenten kaliuretischen Effekt des
Aldosterons am Schaf bei hohen Plasmakaliumwerten. Allerdings waren diese Werte
nur durch KCl-Infusionen zu erzielen und konnten nicht im Zuge der
Nahrungsaufnahme beobachtet werden.
Auch eine Beteiligung von HCO3- und Glukagon an der postprandialen
Kaliumexkretion ist unwahrscheinlich, da beides zumindest bei Schafen nicht bzw.
erst in pharmakologischen Dosen zu einer vermehrten Ausscheidung von Kalium
über die Nieren führte (RABINOWITZ et al. 1984 b). Hingegen konnten
RABINOWITZ et al. (1984 b) mit intravenösen Infusionen von Propionat und Acetat
sowohl sinkende Kaliumspiegel im Plasma als auch eine verstärkte (Aldosteron-
unabhängige) Kaliumexkretion erzielen. Eine mögliche Erklärung sehen die Autoren
darin, dass die vermehrte Bereitstellung von Energie zu einer stärkeren (ATP-
abhängigen) Kaliumsekretion in der Niere führen könnte. Propionat könnte allerdings
auch über eine erhöhte Freisetzung von Insulin auf den Kaliumhaushalt wirken (DE
JONG 1982).
Darüber hinaus ist die Kaliumexkretion vom Urinvolumen abhängig: eine forcierte
Diurese erhöht die Kaliumausscheidung und umgekehrt wird sie durch eine geringe
Urinmenge gehemmt (BROBST 1986; LUNN u. MC GUIRK 1990).
Zwar ist auch Insulin vermutlich nicht die Ursache der vermehrten postprandialen
Kaliumausscheidung (RABINOWITZ et al. 1984 b), trägt aber dennoch maßgeblich
zur Senkung der Kaliumkonzentration im Blut bei, indem es dessen Aufnahme in die
Zellen verstärkt, so dass unter dem Einfluss von Insulin die intrazellulären
Kaliumgehalte in der Leber, im Fettgewebe und in der Muskulatur ansteigen (VAN
MEIRHAEGHE 1988; SWEENEY 1999). Es wurde z. B. gezeigt, dass Insulin den
intrazellulären Kaliumgehalt im Zwerchfell von Ratten (KAMMINGA et al. 1950;
CREESE u. NORTHOVER 1961; CREESE 1968) im Musculus soleus von Ratten
(CLAUSEN u. KOHN 1977; FLATMAN u. CLAUSEN 1965) und in der
Froschmuskulatur (MANERY et al. 1950, 1977; SMILLIE u. MANERY 1960;

Literaturübersicht 25
GOURLEY 1965) erhöht. Damit verhindert postprandial ausgeschüttetes Insulin
gleichfalls im Sinne einer Feedforward Reaktion das übermäßige Ansteigen der
Kaliumkonzentration im Plasma.
Diese durch Insulin bedingte erhöhte Kaliumaufnahme in die Muskelzellen konnte
durch Ouabain, einem Hemmer der Na+/K+-ATPase, aufgehoben werden (CLAUSEN
u. KOHN 1977; GOURLEY 1965; MANERY et al. 1977; ZEMKOVA et al. 1982).
Gleichzeitig reduziert Insulin den Natriumgehalt der Muskelzellen über eine Ouabain-
sensitive Steigerung des Natriumeffluxes (CLAUSEN u. KOHN 1977; CREESE u.
NORTHOVER 1961; CREESE et al. 1968; CHINET u. CLAUSEN 1984; MOORE
1973; ERLIJ u. GRINSTEIN 1976; KITASATO et al. 1980a, b, c; ERLIJ 1984).
Zudem zeigten CLAUSEN und HANSEN (1977), dass Insulin die Ouabain-
Bindungsrate steigerte, was, laut Autoren, für eine höhere Turnover-Rate der Na+/K+-
ATPase sprechen würde. Die Stimulation der Na+/K+-ATPase bietet also eine
mögliche Erklärung dafür, wie es zu einer vermehrten Aufnahme von Kalium in die
Zellen kommt.
Im Gegensatz dazu ist Insulin allerdings auch in der Lage, die Aktivität (BERWECK
et al. 1993) bzw. die Offenwahrscheinlichkeit (YASUI et al. 2007) von Kaliumkanälen
zu erhöhen, so dass es in einigen Geweben im Endeffekt in Anwesenheit von Insulin
zu einem Ausstrom von Kalium kommt.
2.3 Insulin
2.3.1 Insulinhomöostase
Für die physiologische Insulinkonzentration im Plasma von Rindern liegen keine
offiziellen Referenzwerte vor. In verschiedenen Untersuchungen konnte gezeigt
werden, dass die Insulinkonzentrationen im Plasma starken Schwankungen
unterliegen. Die Angaben der Autoren über die Insulinwerte gesunder,
trockenstehender, tragender Tiere variieren zwischen 8,9 (±2,4) µU/ml (KRÄFT
2004) und 27 (±3) µU/ml (SANO et al. 1993 b). Innerhalb eines Tages schwanken die
basalen Insulinwerte zudem abhängig von der Fütterung: 2 - 4 h nach der letzten
Mahlzeit steigen die Insulinwerte an und sinken dann wieder ab mit Erreichen des
Minimums 12 – 16 h nach der letzten Fütterung. Eine Kuh im Zeitraum 17 bis 11

26 Literaturübersicht
Wochen vor der Abkalbung weist deshalb an einem Tag Insulinwerte zwischen ca.
10 und 40 µU/ml auf (STAUFENBIEL et al. 1992). So erscheint nur ein Vergleich der
Insulinwerte innerhalb einer Studie und nicht zwischen den einzelnen
Untersuchungen sinnvoll.
Als anaboles Hormon stellt Insulin die Nährstoffversorgung (insulinabhängiger)
Gewebe sicher. Am Ende der Trächtigkeit kommt es im mütterlichen Metabolismus
jedoch allmählich zu einer Verschiebung der verschiedenen Metabolite zwischen
dem Eutergewebe und den übrigen Körpergeweben, die u. a. auf veränderte
Insulinspiegel im Blut zurückzuführen ist (MERSMANN 1987). Milchkühe in der
Spätträchtigkeit und Frühlaktation haben daher in der Regel niedrigere
Insulinkonzentrationen im Plasma als Tiere in der Mittel- und Spätlaktation und der
Trockenstehzeit (GRIZARD et al. 1986; MALVEN et al. 1987; BAUMANN u. CURRIE
1980; BLUM et al. 1972). Außerdem treten Hypoinsulinämien im Hungerzustand auf
(HOVE 1978 b; PETTERSON et al. 1994).
Hyperinsulinämien hingegen werden bei adipösen Tieren (MC CANN und REIMERS
1985; MC CANN et al. 1986), bei Tieren mit Laminitis, Mastitis oder Metritis
(HOLTENIUS u. HOLTENIUS 1996) und bei Kühen mit Labmagenverlagerung
beobachtet (OK et al. 2000; SEN et al. 2006).
2.3.1.1 Struktur und Biosynthese von Insulin
Das Proteohormon Insulin besteht aus zwei Peptidketten, einer sauren (A) und einer
basischen (B), die an verschiedenen Positionen über Disulphidbrücken miteinander
verbunden sind (siehe Abbildung 2).
S S
A-Kette
Gly-Ile-Val-Glu-Gln-Cys-Cys-Ala-Ser-Val-Cys-Ser-Leu-Tyr-Gln-Leu-Glu-Asn-Tyr-Cys-Asn
5 10 15 21
S
S
B-Kette
Phe-Val-Asn-Gln-His-Leu-Cys-Gly-Ser-His-Leu-Val-Glu-Ala-Leu-Tyr-Leu-Val-Cys-Gly-Glu-Arg-Gly-Phe-Phe-Tyr-Thr-Pro-Lys-Ala
5 10 15 20 25 30
Abbildung 2: Die Aminosäuresequenz von Rinderinsuli n (STRYER 1999)

Literaturübersicht 27
Insulin wird in den β-Zellen des Langerhansschen Inselapparates gebildet und
gespeichert (MARTIN u. CRUMP 2003): Nach Transkription der mRNA im Zellkern
und Translation im rauen endoplasmatischen Retikulum wird zunächst ein
Insulinvorläufer, das Präproinsulin, gebildet, bei dem die A- und B-Kette über ein
sogenanntes „connecting peptide“ (C-Peptid) miteinander verbunden sind. Dieses C-
Peptid ist in seiner Aminosäurenzusammensetzung, im Gegensatz zum restlichen
Protein, bei den verschiedenen Säugetieren höchst variabel. Es folgt die Spaltung
von Arginin-Arginin und Lysin-Arginin-Resten am C-Peptid durch Trypsin-like und
Carboxypeptidase-like Enzyme sowie die Bildung von Disulphidbrücken zwischen
der A- und B-Kette. Das daraus entstehende Proinsulin wird vom rauen
endoplasmatischen Retikulum in den Golgi-Apparat überführt. Es wird in
zytosolischen Granula bis zur Freisetzung gespeichert. Die definitive Überführung
des Proinsulins in Insulin erfolgt durch Abspaltung der die beiden Kettenenden
verbindenden Brücke (MARTIN u. CRUMP 2003).
2.3.1.2 Sekretion und Abbau von Insulin
Faktoren, die die Insulinsekretion stimulieren, sind Nährstoffe (z. B. Glukose,
Galaktose, Mannose, Arginin, Lysin, Leucin, Alanin, Fettsäuren, Kalium und
Kalzium), gastrointestinale Hormone (z. B. Glukagon, Sekretin und Cholezystokinin)
und parasympathische oder β-adrenerge Stimuli. Auslöser für eine Hemmung der
Insulinsekretion sind u. a. Fasten, Aufregung, Hypokalzämien, α-adrenerge Aktivität
und gastrointestinale Hormone (z. B. Galanin und Somatostatin) (MARTIN u.
CRUMP 2003).
Beim Monogastrier gilt ein Anstieg des Blutzuckerspiegels als wichtigster Auslöser
für den Mechanismus der Insulinfreisetzung. Mit der steigenden Konzentration im
Blut wird vermehrt Glukose in die β-Zellen aufgenommen. Durch eine unmittelbare
Metabolisierung der Glukose (durch eine hohe Glukokinaseaktivität), steigt die
zytosolische ATP-Konzentration, die eine Reduktion der Offenwahrscheinlichkeit
ATP-abhängiger Kaliumkanäle bedingt. Durch die verminderte Kaliumleitfähigkeit
kommt es zu einer Anhebung des Ruhemembranpotentials und damit zu einer
erhöhten Öffnungsrate spannungsabhängiger Kalziumkanäle. Der folgende

28 Literaturübersicht
Kalziumeinstrom führt zu einer gesteigerten Exozytose der Insulingranula und damit
zur Sekretion in die Blutbahn (LÖFFLER u. PETRIDES 1997).
Auch beim Wiederkäuer liegen verschiedene Untersuchungen über die Faktoren vor,
die auf die Insulinsekretion Einfluss nehmen:
TRENKLE (1971) konnte zeigen, dass sowohl die basalen Insulinspiegel als auch die
Insulinsekretionsrate bei gefütterten Schafen signifikant höher waren als die
fastender Tiere, wohingegen sich bei der Eliminationsrate keine Unterschiede
ergaben. Zumindest prepartum ist die Menge der Insulinsekretion nach
Glukoseinfusionen dabei abhängig von der Fütterungsintensität: Sie nahm mit der
Intensität der Fütterung zu (bei 71 MJ umsetzbarer Energie geringere
Insulinsekretion als bei 177 MJ) (HOLTENIUS et al. 2003). Verschiedene Autoren
konnten nachweisen, dass die Insulinsekretion vor allem von der
Propionatkonzentration im Portalblut und einem prandial erhöhten Vagotonus
abhängig ist (MANNS u. BODA 1967; MANNS et al. 1967; BLOOM u. EDWARDS
1981; SARTIN et al. 1985; MINEO et al. 1990; WEEKES 1991; GRIINARI et al.
1997). Darüber hinaus wurde in verschiedenen Studien die Insulinsekretion nach
Infusion unterschiedlicher Substanzen überprüft. So konnte durch intravenöse.
Glukoseapplikationen bei Milchkühen der Insulingehalt im Blut erhöht werden
(GIESECKE 1986), wobei SAKAI et al. (1996) eine stärkere Insulinantwort durch
Xylitolgaben erzielten (Xylitol ist ein intermediär vorkommendes Pentose-Derivat, das
keine Erhöhung des Blutzuckerspiegels bewirkt und daher auch „Diabetiker-Zucker“
genannt). Auch Fettsäureninfusionen, neben Propionat auch n-Butyrat und n-Valerat
(nicht hingegen Acetat), hatten ansteigende Insulinkonzentrationen im Blut zur Folge
(DE JONG 1982). Auch HORINO et al. (1968) zeigten einen Anstieg der
Plasmainsulinspiegel durch Infusion von Fettsäuren mit drei bis acht C-Atomen.
Interessant dabei war, dass es zu keiner Erhöhung der Glukosekonzentration im Blut
kam, die Fettsäuren also direkt und nicht etwa über eine Stimulierung der
Glukoneogese die Insulinsekretion verstärkte. Die Autoren ziehen aus ihrer Studie
den Schluss, dass Fettsäuren mit oben genannter Länge bei Wiederkäuern potentere
Stimulatoren der Insulinsekretion sind als Glukose. Im Gegensatz dazu zeigten
KNOWLTON et al. (1998) eine tendenziell stärkere Insulinantwort auf eine

Literaturübersicht 29
abomasale als auf eine ruminale Stärkeinfusion. Des Weiteren konnten ODA et al.
(1986) an Ziegen den Nachweis erbringen, dass durch β-adrenerge Substanzen eine
Insulinsekretion ausgelöst wird. Auch intravenöse Glukagonapplikationen erhöhen
den Insulingehalt im Plasma (HOLTENIUS u. TRAVEN 1990).
Nach Stimulation der Insulinsekretion kommt es zu einem biphasischen Anstieg der
Plasmakonzentration, der vermutlich darauf zurückzuführen ist, dass zunächst eine
Ausschüttung der Reserven erfolgt und dann eine Neuproduktion mit anschließender
Freisetzung; bei hungernden Kühen ist nur ein monophasischer Anstieg des
Insulingehaltes im Plasma festzustellen (HOVE 1978 b).
Neben reduzierten Insulinspiegeln im Hungerzustand (HOVE 1978 b) wurde nach
Gabe von α-Adrenergika (ODA et al. 1986) eine Hemmung der Insulinfreisetzung
beobachtet. Diese binden an α2-Rezeptoren der Plasmamembran der β-Zellen im
Pankreas und hemmen den Kalziumeinstrom in die Zelle und damit die Freisetzung
von Insulin (BROCKMAN 1986). In diesem Zusammenhang ist auch die mit
Hypokalzämien (GOFF 1999) einhergehende verminderte Insulinsekretion zu sehen.
Zudem konnte Somatostatin bei Ziegen eine Glukose-induzierte Insulinsekretion
kurzzeitig verhindern (MAGLAD et al. 1983), indem es Kaliumkanäle aktiviert und
damit eine Depolarisation der β-Zellen verhindert (BROCKMAN 1986).
Abgebaut wird das in der Blutbahn zirkulierende Insulin enzymatisch in der Leber. Es
hat im Serum eine Halbwertszeit von 10-15 Minuten. Bei der Bindung des Insulins an
die Insulinrezeptoren der Zielzellen kommt es im Zuge der Signaltransduktion zu
einer Internalisierung des Komplexes und zu einem anschließenden Abbau durch
lysosomale Enzyme. Dabei werden die Disulphidbrücken durch die Glutathion-
Insulin-Transhydrogenase gespalten und die entstandenen Ketten proteolytisch
abgebaut (LÖFFLER u. PETRIDES 1997).
2.3.1.3 Signaltransduktion am Erfolgsorgan
Bei der Signaltransduktion werden drei Stadien unterschieden: Phosphorylierung des
Insulinrezeptors, intrazelluläre Signalkaskade über second messenger und
Translokation z. B. der Glukosetransporter.

30 Literaturübersicht
Der Insulinrezeptor ist ein dimeres Molekül aus jeweils zwei gleichen α- und β-
Untereinheiten, die über eine Disulphidbrücke miteinander verknüpft sind. Während
die α-Untereinheiten auf der Zelloberfläche lokalisiert sind, befinden sich die β-
Anteile v. a. im Zellinneren, haben allerdings auch transmembranäre und
extrazelluläre Anteile (TORNQUIST u. AVRUCH 1988; TAYLOR 1991; LÖFFLER u.
PETRIDES 1997). Sie sind zudem mit einer Tyrosinkinase-Domäne ausgestattet
(KAHN 1994). Die Bindung von Insulin an die α-Untereinheit führt zu einer
Konformationsänderung der Untereinheiten und im Zuge dessen zu einem Wegfall
der Hemmwirkung der α-Untereinheit auf die Tyrosinkinase der β-Untereinheit. Im
weiteren Verlauf kommt es zu einer Autophosphorylierung einiger Tyrosylreste an
der β-Untereinheit und in der Folge zu einer Internalisierung des Insulin-Rezeptor-
Komplexes. Dieser Prozess löst durch sich intrazellulär ausbreitende second
messenger-Systeme eine Kaskade an Phosphorylierungen und
Dephosphorylierungen aus. Die entstehenden Botenstoffe sind in ihrer Wirkung auf
das Endergebnis spezifisch. Die Phosphorylierung des Insulinrezeptorsubstrates
stimuliert beispielsweise die Glykogensynthese, die des Guanosintriphosphats
fördert Wachstum und Genexpression und die der Phosphoinositolphosphat-Kinase
die Lipogenese, Proteinsynthese und die Translokation von Glukosetransportern
(KAHN 1994).
2.3.2 Insulinresistenzphänomen
KAHN (1978) beschreibt die Insulinresistenz als einen Zustand, in dem die
physiologische Insulinkonzentration eine verminderte Antwort erzeugt. BERSON und
YALOW (1970) sehen in ihr eine Situation, in der eine größere Insulinmenge benötigt
wird, um eine normale Reaktion hervorzurufen. Man spricht in diesem Fall von einer
verminderten Insulinsensitivität, bei der eine höhere Insulinkonzentration nötig ist, um
einen halbmaximalen biologischen Effekt (km-Wert) zu erzielen. Die Höhe des
maximalen biologischen Effektes bleibt unverändert. Bei einer reduzierten Insulin-
Response, die auch einer Insulinresistenz zugrunde liegen kann, ist hingegen der
maximale Effekt des Insulins bei unverändertem km-Wert vermindert (RIZZA et al.
1981; SANO et al. 1991).

Literaturübersicht 31
Auf molekularer Ebene können Abweichungen der Insulinwirkung im Körper an drei
verschiedenen Lokalisationen ihren Ursprung nehmen:
1. Im Prärezeptor-Level (Insulinsekretionsrate reduziert)
2. Im Rezeptor-Level (verminderte Dichte oder Ansprechbarkeit der Rezeptoren)
3. Im Postrezeptor-Level (intrazelluläre Signalkaskade unterbrochen) (VERNON
u. SASAKI 1991)
Ein Defekt im Prärezeptor-Level bedingt in der Regel eine Hypoinsulinämie. Eine
herabgesetzte Insulin-Response nimmt im Allgemeinen ihren Ursprung im Rezeptor-
Level, während eine fehlerhafte Signalübertragung im Postrezeptor-Level Ursache
einer reduzierten Insulin-Sensitivität ist.
HOLTENIUS und HOLTENIUS (1996) beschreiben bei der hochleistenden Milchkuh
zwei verschiedene Diabetesformen:
1. Typ-I-Diabetes: Hypoinsulinämische und –glykämische Tiere (in der
Frühlaktation bei negativer Energiebalance)
2. Typ-II-Diabetes: Hyperinsulinämische und –glykämische Tiere (häufig mit
Krankheit wie Metritis, Mastitis oder Laminitis einhergehend)
Zur Prüfung der Insulin-Resistenzlage werden euglykämische Insulinclamps oder
Glukosetoleranztests (GTT) durchgeführt. Im GTT, bei der Glukose intravenös
verabreicht wird, können z. B. basale und maximale Plasmainsulinkonzentrationen,
Glukose-Clearance, Verhältnis von Plasma-Glukose zu Plasma-Insulin und die Zeit
bis zum Wiedererreichen normaler Glukosewerte bestimmt werden (HAYIRLI et al.
2001). Beim euglykämischen Clamp wird eine bestimmte Menge an Insulin infundiert
und dabei der Glukosespiegel konstant gehalten. Aus verbrauchter Glukosemenge
und infundierter Insulinkonzentration kann der biologische Effekt abgeleitet werden
(SANO et al. 1991, 1993 a).
Die Entwicklung einer Insulinresistenz ist ein multikausales Geschehen. Im
Folgenden sind einige Faktoren aufgeführt.
Reproduktionsstadium:
Insulinresistenzen werden häufig im Zusammenhang mit fortgeschrittenen
Trächtigkeiten und einsetzender Laktation beobachtet (PRIOR u. CHRISTENSON
1978; DEBRASS et al. 1989; FAULKNER u. POLLOCK 1990; PETTERSON et al.

32 Literaturübersicht
1994). Der Nährstoffbedarf des wachsenden Fetus und später die einsetzende
Laktation fordern einen flexiblen mütterlichen Metabolismus. Bei Schafen beträgt die
fetale Glukoseaufnahme 42-50 % der mütterlichen Glukoseproduktion bzw. wird die
uterine Glukoseaufnahme mit 18 mg/kg Fetus/min angegeben (PRIOR u.
CHRISTENSON 1978). Der Glukosetransport in der Plazenta, die uterine
Glukoseaufnahme sowie die Glukosesekretion ins Euter sind dabei (anders als der
Transfer von Glukose in das mütterliche Muskel- oder Fettgewebe) weitestgehend
insulinunabhängig (HOVE 1978 a; HAY et al. 1984). Die Umverteilung dieser
Nährstoffe in Richtung Plazenta und Euter wird daher vor allem über eine geringere
Insulinsekretion und eine geringere Ansprechbarkeit des Körpergewebes auf Insulin
erzielt. STAUFENBIEL et al. (1992) haben die Insulin- und Glukoseregulation bei der
Milchkuh mittels Tagesprofilen im Zeitraum von 17 Wochen ante partum bis 52
Wochen post partum beurteilt. Dabei wurden die niedrigsten Insulinwerte im
Tagesprofil in der 23. Woche p.p. gemessen bei einer generellen Abnahme ab der 7.
Woche a. p.. Auch SANO et al. (1993 b) erbrachten den Nachweis, dass bei
laktierenden Kühen die Insulinsekretion nach Infusion von Glukose geringer ist als
bei Tockenstehern (23 vs. 102 µU/ml). Hingegen konnten sie im Glukoseverbrauch
bei hyperinsulinämischen Clamps keinen Unterschied zwischen laktierenden und
trockenstehenden Tieren feststellen. GIESECKE (1986) hat die Insulinantwort nach
einer Glukoseinfusion gemessen. Zu sehen war eine deutliche Abnahme der
Insulinspiegel 6 Wochen ante partum bis 6 Wochen post partum, was der Autor auf
eine verminderte Sekretion zurückführte. In einer weiteren Untersuchung zeigte er im
gleichen Zeitfenster zusätzlich an Erythrocyten eine Affinitätsabnahme für Insulin.
Vorausgesetzt, dass sich die an den Erythrocyten gewonnenen Erkenntnisse auf
andere Gewebe übertragen lassen, bedeutet das, dass neben reduzierten
Insulinspiegeln im Blut peripartal außerdem eine verminderte Ansprechbarkeit der
Gewebe auf Insulin vorliegen könnte.
KRÄFT (2004) hingegen demonstrierte, dass Milchkühe in der Frühlaktation im
Gegensatz zu trockenstehenden Kühen eine hohe Insulin-Sensitivität und -Response
aufweisen, was einer Unterversorgung extramammärer Gewebe bei niedrigen
Insulinspiegeln entgegen wirkt. So besteht eine grundsätzliche Übereinstimmung

Literaturübersicht 33
hinsichtlich der verminderten Insulinsekretion nach Glukoseapplikation in der
Spätträchtigkeit und Frühlaktation, wohingegen es gegenläufige Ergebnisse in Bezug
auf die Ansprechbarkeit der Gewebe auf Insulin in diesem Zeitraum gibt.
Der Effekt von Insulin auf den Fettstoffwechsel während der Trächtigkeit und der
Frühlaktation wurde von GUESNET et al. (1991) am Schaf untersucht. In den ersten
drei Monaten der Trächtigkeit konnten die Autoren eine verstärkte Lipogenese, eine
reduzierte Reaktion auf einen ß-lipolytischen Stimulus mittels Isoproterenol und eine
erhöhte Anzahl von Insulinrezeptoren mit hoher Affinität feststellen. Im letzten Drittel
der Trächtigkeit und in der Laktation kam es zu einem umgekehrten Bild. V. a. in der
Laktation war die Anzahl der Insulinrezeptoren vermindert und die Insulin-stimulierte
Lipogenese wurde unzureichend. Analysen von Dosiswirkungskurven im Zuge von
Insulininfusionen bei tragenden und nicht-tragenden Schafen zeigten, dass die
maximale Insulin-vermittelte Reduzierung von NEFAs und Glycerol im Plasma bei
tragenden Schafen signifikant geringer ist als bei den nicht-tragenden Tieren. Das
bedeutet, dass es bei Schafen während der Trächtigkeit zu einer verminderten
Ansprechbarkeit des Fettgewebes auf Insulin kommt (PETTERSON et al. 1994).
Während der späten Trächtigkeit sind u. a. die Konzentrationen von Progesteron,
Cortisol und Prolaktin im Plasma erhöht. RYAN und ENNS (1988) haben den Effekt
dieser Hormone auf die Insulinwirkung (maximale Insulinbindung und
Glukosetransportrate) an isolierten Fettgewebszellen von Ratten untersucht. Dabei
stellten sie fest, dass die Gabe von Prolactin den Glukosetransport hemmte und
Progesteron und Cortisol beides reduzierten. Das vor allem zu Beginn der Laktation
ebenfalls erhöhte Estradiol erzeugte hingegen eine gesteigerte maximale
Insulinbindung.
Ernährungszustand:
In einer Vielzahl von Untersuchungen an Wiederkäuern und Monogastriern konnte
gezeigt werden, dass mit Adipositas häufig Stoffwechselstörungen vergesellschaftet
sind. Erhöhte Konzentrationen an NEFAs im Blut korrelieren mit einer gestörten
intrazellulären Signalkaskade (ZIERATH et al. 1998; LE MARCHAND-BRUSTEL et
al. 1999), einer reduzierten Anzahl an GLUT-4-Transportern (VAN EPPS-FUNG et
al. 1997) und einer Hemmung der Insulin-stimulierten Glukoseaufnahme des

34 Literaturübersicht
peripheren Gewebes (KOOPMANS et al. 1996). GRIZARD und SZCZYGIEL (1983)
konnten an Schafen eine mit steigendem Körpergewicht abnehmende Insulinbindung
an Hepatocyten nachweisen. Es kommt im Zusammenhang mit Fettleibigkeit auch zu
Insulinresistenzen, die mit Hyperinsulinämien einhergehen. MC CANN und
REIMERS (1985) und MC CANN et al. (1986) konnten zeigen, dass es sowohl bei
adipösen Schafen als auch bei fettleibigen Färsen im Vergleich zu normal
konditionierten Tieren zu einer Reduktion der Glukoseregulation und zu erhöhten
Insulinspiegeln kam. Allerdings beschreiben z. B. HAYIRLI et al. (2002),
GARNSWORTHY und TOPPS (1982) und TREACHER et al. (1986) bei obesen
Wiederkäuern im Gegensatz zu monogastrischen Tieren einen verminderten Appetit.
HAYIRLI (2006) folgert daraus, dass Adipositas bei Nicht-Wiederkäuern mit
Hyperglykämie und –insulinämie und bei Wiederkäuern mit Hypoglykämie und –
insulinämie vergesellschaftet ist.
Eine moderate, kurzfristige Unterernährung hatte bei Schafen keine Auswirkungen
auf die Insulin-vermittelte Reduktion von NEFAs und Glycerol im Blut (PETTERSON
et al. 1994). Eine andere Untersuchung hingegen zeigt, dass bei gesunden Kühen
nach Infusion von Glukose höhere Insulinspiegel erzielt wurden als bei hungernden
Tieren (HOVE 1978 b). Die Autoren erklären dies zum einen mit einem durch HALSE
(1960) an Milchkühen dokumentierten Abfall der Plasma-Kalziumspiegel um 20 %
nach 48 Stunden Fasten. Zum anderen halten die Autoren den beim fastenden
Menschen nachgewiesenen Norepinephrin-Anstieg für einen möglichen hemmenden
Faktor. Zusätzlich bietet sich eine weitere Erklärungsmöglichkeit in dem durch die
reduzierte Futteraufnahme verminderten Propionatspiegel sowie der dadurch
bedingten Hypoglykämie (HERDT 2000; OSTERGAARD u. GRÖHN 1999).
Mineralstoffimbalancen:
In verschiedenen Untersuchungen konnte ein Zusammenhang zwischen
Hypokalzämien und verminderter Insulinsekretion nachgewiesen werden
(LITTLEDIKE et al. 1968; GOFF 1999). Beim Schaf ist bei bestehender
Hypokalzämie zusätzlich zur herabgesetzten Sekretion auch eine Hemmung der
Insulin-Clearance nachgewiesen (SCHLUMBOHM et al. 1997).

Literaturübersicht 35
Eine Hypokaliämie kann ähnlich hemmende Auswirkungen auf die Insulinsekretion
haben, da es bei niedrigen Kaliumspiegeln allgemein zu einem vermehrten Ausstrom
von Kalium aus der Zelle kommt. Dadurch wird deren Ruhemembranpotential
negativer und als Konsequenz kann die Zelle nicht mehr so leicht depolarisiert
werden. Dies ist allerdings die Voraussetzung dafür, dass die potentialabhängigen
Kalziumkanäle öffnen. Im Endeffekt wird also die Insulinsekretion gehemmt
(PETRIDES 1997). In die gleiche Richtung zielen Beobachtungen bei diabetischen
Menschen mit gleichzeitig bestehender Hypokaliämie, bei denen die Glukosetoleranz
durch Anhebung der Kaliumspiegel verbessert werden konnte (TOURNIAIRE et al.
1988).
Labmagenverlagerung und Ketose:
Auch verschiedene Erkrankungen bei Milchkühen können mit Insulinresistenzen
zusammen auftreten. Bei ketotischen Tieren kommt es bei unveränderten (VAN
MEIRHAEGHE et al. 1988 b) oder verminderten (BROCKMAN 1979; KRÄFT 2004)
basalen Insulinspiegeln zu einer geringeren Insulinsekretion als Antwort auf
Glukoseinfusionen verglichen mit gesunden Tieren im gleichen Laktationsstadium.
Auch HOVE (1978 b) erzielte in gesunden Kühen nach Infusion von Glukose höhere
Insulinspiegel als in ketotischen. Den geringen Anstieg der Insulinspiegel der
ketotischen Tiere erklärten die Autoren mit einer verminderten sekretorischen
Kapazität der ß-Zellen, herrührend von der Tage oder Wochen andauernden Phase
der die Ketose begleitenden Hypoglykämie. SAKAI et al. (1996) erzielten ähnliche
Ergebnisse: bei gesunden Kühen war nach Infusion von 500 ml einer 50 %igen
Glukoselösung die Insulinkonzentration im Plasma 7 mal, bei ketotischen Tieren nur
3 mal höher. Jedoch zeigte die Infusion von Xylitol ein anderes Bild: bei gesunden
Kühen war nach der Infusion von 1000 ml einer 25 %iger Xylitollösung die
Insulinkonzentration 9 mal, bei ketotischen Tieren hingegen 12 mal höher. Die
Autoren erklären diesen Insulinanstieg entweder mit einem verminderten Abbau von
Insulin, da die Diffusion von Xylitol ins periphere Gewebe Insulin unabhängig verläuft,
oder aber mit einer gesteigerten Synthese und Freisetzung. BECK et al. (1983)
erbrachten außerdem den Nachweis, dass ketotische Kühe neben einer
verminderten Insulinsekretion auch eine reduzierte Insulin-Sensitivität aufweisen.

36 Literaturübersicht
Tiere mit bestehender Labmagenverlagerung hingegen weisen signifikant höhere
Insulin- und Glukosewerte auf als gesunde Kühe. Auch auf eine Glukoseinfusion
reagieren erstere mit einer langanhaltenden erhöhten Insulinsekretion (VAN
MEIRHAEGHE et al. 1988 b; HOLTENIUS u. TRAVEN 1990). Tiere, die sowohl
ketotisch sind als auch an einer Labmagenverlagerung leiden, reagierten in dieser
Studie überwiegend mit einer starken Insulinantwort (VAN MEIRHAEGHE et al. 1988
b). OK et al. (2000) zeigten zudem, dass Kühe mit rechtsseitiger
Labmagenverlagerung noch höhere Insulinspiegel haben als solche mit linksseitiger.
2.3.3 Beeinflussung der Labmagenmotilität durch Insulin
Verschiedene Studien belegen, dass Insulin einen hemmenden Einfluss auf den
Labmagen hat. VAN MEIRHAEGE et al. (1988 a) demonstrierten eine Reduktion des
abomasalen Effluxes nach einer Insulin-Injektion von 0,2 U/kg KGW, obwohl der
hypoglykämische Effekt durch Gaben von Glukose oder Adrenalin abgepuffert
wurde. Insulin wirkt also unabhängig vom Blutglukosespiegel. Auch SUSTRONCK
(2000) und HOLTENIUS et al. (2000) konnten eine klare Korrelation zwischen
bestehender Hyperinsulinämie und verzögerter Labmagenentleerung zeigen, die
Glukose-unabhängig war.
Es blieb allerdings unklar, ob und inwiefern es sich bei der Hemmung um eine
Beeinflussung der Labmagenmuskulatur oder um einen sekundären Effekt
(beispielsweise durch verminderten Vorschub aus dem Pansen) handelte. Zu diesem
Zweck setzten PRAVETTONI et al. (2004, 2007) Kühen bei der Labmagenoperation
abomaso-duodenale Elektroden ein. Untersucht wurden Kühe mit
Labmagenverlagerung zu Beginn ihrer Laktation. Alle Tiere wiesen dabei erhöhte
Insulinspiegel auf. Nach der chirurgischen Korrektur sanken die Blutspiegel ab, was
eine langsame Erhöhung der abomaso-duodenalen myogenen Aktivität nach sich
zog. Aufgrund der unterschiedlich hohen Insulinspiegel nach der Operation wurden
die Tiere zudem in zwei gleich große Gruppen geteilt: Tiere mit hohen und solche mit
niedrigen Insulinspiegeln. Kühe mit hohen Plasmakonzentrationen zeigten sieben
Tage nach der Operation eine signifikant niedrigere myogene Aktivität als die Tiere
mit niedrigen Plasmainsulinwerten (PRAVETTONI et al. 2004).

Literaturübersicht 37
2.4 Hypothese
Ziel dieser Arbeit war es herauszufinden, ob und auf welche Weise Insulin und
Kalium einzeln oder in Kombination auf isolierte Labmagenmuskulatur wirken und
inwieweit sich die daraus gewonnenen Ergebnisse in In-vivo-Untersuchungen
bestätigen lassen.
Gesichert ist, dass experimentelle Insulininfusionen unter normoglykämischen
Bedingungen den Efflux von Ingesta aus dem Labmagen vermindern (HOLTENIUS
et al. 2004). Dies könnte bedeuten, dass Insulin direkt, also Glucose-unabhängig, auf
die Labmagenmotilität wirkt. Zudem wurde eine Korrelation zwischen verlängerter
postoperativer Labmagenatonie und hohen Insulinspiegeln festgestellt
(PRAVETTONI et al. 2004).
Ferner ist bewiesen, dass Insulin nach Futteraufnahme zu einer erhöhten Aufnahme
von Kalium in Zellen der Skelettmuskulatur führt. Insulin ist also in der Lage,
Kaliumgradienten über Zellmembranen zu beeinflussen. Ob die Insulinwirkung an der
Muskulatur des Labmagens über lokale Änderungen von Kaliumgradienten zustande
kommt, ist bisher nicht untersucht. Denkbar wäre eine Insulin-bedingte Stimulation
der Na+/K+-ATPase in Analogie zu den Vorgängen an der Skelettmuskelzelle oder
eine Aktivierung von Kaliumkanälen, wie es an Pericyten retinaler Kapillaren
(BERWECK et al. 1993) und glatten Muskelzellen der thorakalen Aorta von
Rattenembryonen (YASUI et al. 2007) nachgewiesen werden konnte.
Daraus ergeben sich folgende Fragestellungen:
1. Wie reagiert isolierte Labmagenmuskulatur auf Veränderungen der
extrazellulären Kalium-Konzentration?
2. Wie reagiert sie auf steigende Insulingaben?
3. Gibt es Hinweise für eine Beteiligung der Kalium-Leitfähigkeit oder der Na+/K+-
ATPase an dem Insulineffekt auf die Labmagenmuskulatur?
4. Lassen sich die Ergebnisse auf Untersuchungen in-vivo übertragen?

38 Material und Methoden
3 Material und Methoden
3.1 In-vitro-Motilitätsmessung der Labmagenmuskulat ur
3.1.1 Versuchstiere
Das Probenmaterial stammte einerseits von 22 gleichaltrigen, definiert gehaltenen
und gefütterten Ziegenböcken, die im Rahmen eines anderen Versuches am
hiesigen Institut geschlachtet wurden. Andererseits wurde Labmagenmaterial von 74
nichttragenden schwarzbunten Milchkühen aus dem Schlachthof in
Gleidingen/Laatzen gewonnen. Ihre Schlachtkörper und Innereien wiesen keinerlei
Veränderungen auf.
3.1.2 Entnahme der Muskelstreifen
Die Tiere wurden mittels Bolzenschuss betäubt und durch Entbluten getötet. Die
Entnahme des Probenmaterials erfolgte ca. 15 Minuten (Ziegen) bzw. 30 Minuten
(Kühe) nach der Tötung. Dabei wurden 5 x 5 cm große Präparate der
Labmagenwand aus dem proximalen Drittel der Magenpumpe (Corpus) und dem
Pylorus entnommen, mit erwärmter (37 °C) Pufferlösu ng gespült und schließlich
darin inkubiert.
Die Lösung hatte folgende Zusammensetzung:
NaCl: 117,0 mmol/l; KCl: 4,7 mmol/l; MgCl2: 1,2 mmol/l; NaH2PO4: 1,2 mmol/l; CaCl2:
2,5 mmol/l; NaHCO3: 25,0 mmol/l; Glukose: 11,0 mmol/l; pH-Wert 7,4 bei Begasung
mit O2/CO2 95/5 %.
3.1.3 Präparation der Zirkulär- und Längsmuskelstreifen
Die Präparation der Muskelstreifen erfolgte 10 Minuten (Ziegen) bzw. 60 Minuten
(Kühe) nach der Entnahme. Zu diesem Zweck wurden die Gewebe frisch
angeschnitten und mit der mukosalen Seite nach oben in einer mit Sylgard
beschichteten und mit Pufferlösung befüllten Petrischale mit Präpariernadeln
aufgespannt. Mit Hilfe von Pinzette und Präparierschere wurden Mukosa und
Submukosa unter dem Mikroskop (SZ40, Olympus, Hamburg) bei 10facher
Vergrößerung entfernt, 0,5 cm x 1,0 cm große Muskelstreifen parallel zur benötigten

Material und Methoden 39
Faserrichtung (zirkulär bzw. longitudinal) festgepinnt und geschnitten. Danach wurde
an beiden Enden der Muskelstreifen Nähgarn zur Befestigung an der
Messvorrichtung angeknotet.
Bei den Ziegen wurden neben zirkulärer Pylorus- und zirkulärer Corpusmuskulatur im
Gegensatz zu den Kühen auch longitudinale Corpusmuskulatur untersucht.
3.1.4 Messung der mechanischen Aktivität der Muskelstreifen
3.1.4.1 Versuchsvorrichtung
Die Apparatur zur Messung der mechanischen Aktivität der Muskelstreifen bestand
aus:
- 8 doppelwandigen Kammern, deren äußerer Bereich mit dem Wasserbad
verbunden war und deren innerer Bereich (in den die Einsätze mit den
fixierten Muskelstücken eingebracht werden) mit einem verschließbaren
Abfluss und der Möglichkeit zur Carbogenbegasung versehen war
- 8 isometrische Kraftaufnehmer, die zum Zwecke der Vordehnung
höhenverstellbar waren
- einer Messbrücke, die an einen analogen Digital-Wandler (Spider 8, Holtinger
Baldwin Messtechnik) und an ein computergestütztes Analysesystem
(catman®Easy Version 1.0, HBM Software) angeschlossen waren
Die präparierten Muskelstücke wurden mit Hilfe der Einsätze in die mit 10 ml 37 °C
warmer Pufferlösung befüllten Kammern verbracht und mit dem Faden des freien
Endes mit dem Haken des isometrischen Kraftaufnehmers verbunden. Zur Messung
der kontraktilen Aktivität wurden die eingespannten Muskelstreifen auf 20 mN
vorgedehnt.
Die Kraftaufnehmer dienten zur Umwandlung der mechanischen Aktivität in
elektrische Signale, die über die Messbrücke verstärkt und auf einem Monitor mit
Hilfe eines Analog-Digital-Wandlers und eines computergestützten Analysesystems
dargestellt wurden. Die Aktivität der Muskelstreifen wurde kontinuierlich mit 20 Hz
aufgezeichnet.
Nach einer Adaptationsphase von 90 Minuten wurden die Wirkungen verschiedener
Substanzen, die direkt in die Kammer gegeben wurden, getestet.

40 Material und Methoden
3.1.4.2 Versuchsschemata
Es wurde sowohl die Wirkung von Kalium auf die glatte Labmagenmuskulatur
getestet als auch die von Insulin.
- Kalium: Der Einfluss von Kalium auf isolierte Muskelpräparate wurde anhand
einer Dosiswirkungskurve von 1 bis 10 mmol/l überprüft. Dazu wurde zu 10 ml
Pufferlösung mit einem Kaliumgehalt von 1 mmol/l entsprechende Mengen (10
bzw. 20 µl) einer 1 molaren Kaliumlösung gegeben. Equimolare NaCl-Gaben
dienten dabei als Zeit- und osmotische Kontrolle.
- Insulin: Auch der Einfluss von Insulin auf isolierte Labmagenmuskulatur wurde
mit Hilfe einer Dosiswirkungskurve (0 bis 120 mU Insulin/l) ermittelt. Dies
erfolgte zum Einen im Vergleich zu einer Zeitkontrolle und zum Anderen in
Anwesenheit verschiedener Kaliumkonzentrationen (3 und 5 mmol/l). Dazu
wurde eine Insulinlösung mit einer Konzentration von 7 U/l hergestellt, so dass
eine Zugabe von 10 µl zu 10 ml Pufferlösung zu einer
Konzentrationserhöhung um 7 mU Insulin/l führten. Bei der Kuh wurden
zusätzlich am Pylorus die Wirkung von verschiedenen Insulinkonzentrationen
(0, 40, 120, 200 mU/l) über die Zeit, d.h. ohne weitere Zugaben, gemessen.
- Weiterführende Untersuchungen an der zirkulären Corpusmuskulatur:
o Wirkung von 7 und 40 mU Insulin/l auf den Verlauf einer
Kaliumdosiswirkungskurve
o Wirkung von 80 mU Insulin/l in Anwesenheit des Kaliumkanalblockers
Barium (3 mmol/l)
o Wirkung von 80 mU Insulin/l in Anwesenheit des ATP-sensitive
Kaliumkanäle blockierenden Glybenclamids (0,05 mmol/l)
o Wirkung von 80 mU Insulin/l in Anwesenheit von 0,001 mmol Ouabain/l
(Hemmer der Na+/K+-ATPase)
Die genauen Versuchsschemata sind dem Anhang 10.1 zu entnehmen

Material und Methoden 41
3.1.4.3 Motilitätsparameter
Die Bestimmung der Motlitätsparameter erfolgte jeweils über 2 Minuten vor der
nächsten Zugabe und nach Wirkungseintritt der zugegebenen Substanz. Dabei
wurden folgende Parameter zur Analyse herangezogen:
Mittlere Kontraktionsamplituden [mN]: Die mittlere Kontraktionsamplitude wurde
durch die Summe der Delta zwischen den einzelnen Minima und Maxima geteilt
durch die Frequenz während der auszuwertenden Periode ermittelt.
Kontraktionsfrequenz [1/min]: Die Kontraktionsfrequenz entsprach der Anzahl der
Kontraktionen pro Minute.
Kontraktionsaktivität [mN/min]: Die Kontraktionsaktivität wurde durch Multiplikation
der mittleren Kontraktionsamplitude und der Kontraktionsfrequenz errechet.
Miteinander verglichen wurden gleiche Muskelpräparate einer Tierart.
3.2 In-vivo-Versuch: Bestimmung des Labmageneffluxe s bei intravenöser
Applikation von Insulin und Kalium
3.2.1 Versuchstiere
Bei den Versuchstieren handelte es sich um sechs Kühe der Rasse Holstein-Frisian
der Bundesforschungsanstalt für Landwirtschaft in Braunschweig, die sowohl im
Pansen wie auch im Duodenum fistuliert waren. Die Kühe waren klinisch gesund,
durchschnittlich 7 Jahre alt und in unterschiedlichen Laktationsstadien (4 in Laktion
-davon 2 tragend- und 2 trocken stehende Kühe –davon eine tragend). Alle Tiere
wurden in Anbindehaltung gehalten, morgens und nachmittags gemolken und mit
einer leistungsgerechten Menge an Maissilage und Kraftfutter versorgt.
Futtermittelproben ergaben folgende Zusammensetzung in der Trockensubstanz:

42 Material und Methoden
Tabelle 1: Futtermittelanalyse
Inhaltsstoffe in g/kg Maissilage Stroh Kraftfutterpellets
Rohasche 26,51 50,4 51,3
Rohprotein 43,4 39,1 175
Rohfett 13,94 381 30,7
Rohfaser 84,67 220 73,8
N-freie Extraktstoffe 217,71 544
Kalzium 1,94 2,87 6,16
Phosphor 1,28 0,63 5,02
Natrium 0,14 <0,11 2,47
Kalium 7,04 5,88 6,25
zusätzlich Minerallecksteine
Die genauen Daten der Tiere sind der Tabelle 2 zu entnehmen.
Tabelle 2: Leistungsdaten der Kühe an den Versuchst agen
Versuchstage Name Gewicht Letzte
Abkalbung
Trächtigkeit Milchleistung Ration
Silage
KF
16.-18.04.08 Reike 577 kg 28.09.07 tragend 16 l 26 kg
5 kg
21.-23.04.08 Stella 570 kg 17.12.06 tragend trocken 28 kg
28.-30.04.08 Wolke 560 kg 14.12.07 nicht
tragend
23 l 26 kg
5 kg
05.-07.05.08 Lotte 540 kg 13.11.07 tragend 19 l 30 kg
5 kg
07.-09.05.08 Inka 520 kg 31.03.08 nicht
tragend
13 l 20 kg + 2 kg
Stroh
13.-15.05.08 Ambra 705 kg 28.12.06 nicht
tragend
trocken 24 kg

Material und Methoden 43
Das Versuchsvorhaben wurde gemäß § 8 TSchG angemeldet und genehmigt.
3.2.2 Versuchsaufbau
3.2.2.1 Vorbereitung der Versuchstiere
Am Vortag des dreitägigen Versuches wurden die Tiere auf einer Viehwaage
gewogen und klinisch allgemein untersucht. Am Morgen der Versuchstage wurden
die Tiere gemolken und gefüttert, eventuell vorhandene minimale Futterreste wurden
aber vor Beginn des Versuches entfernt.
Am Morgen des ersten Versuchstages wurden zwei venöse Zugänge in beide Vv.
jugulares externae gelegt. Zu diesem Zweck wurde das obere Halsdrittel im Bereich
der Drosselrinne großflächig rasiert, mit Ethanol (96 %) entfettet und anschließend
jodiert (alkoholische Jodlösung, 10 % w/v). Ein Venenverweilkatheter (1,1 x 1,7 mm,
Länge 45 cm) wurde herzwärts geschoben und mittels zweier Hauthefte sowie mit
Klebeband fixiert. An beiden Kathetern wurde eine Heidelberger Verlängerung (140
cm) angebracht, die auf der einen Halsseite auf 15 cm gekürzt wurde und so einer
erleichterten Blutentnahme diente. Auf der anderen Seite wurde ein Dreiwegehahn
befestigt, an den einerseits die Insulininfusionspumpe und andererseits die
Glukoseinfusion (G40 % ad us. vet) angeschlossen wurde. Der Katheter wurde nach
jeder Blutentnahme mit physiologischer Kochsalzlösung (NaCl 0,9 %) gespült.
3.2.2.2 Vorbereitung der Infusionslösungen
Zunächst wurde eine Insulinstammlösung (7175 U Insulin/l) hergestellt, die dann,
individuell auf das Gewicht der Tiere abgestimmt, verdünnt werden konnte. Für die
Stammlösung wurde das Insulinpulver (Insulin from bovine pancreas) mit 500 ml
sterilem Aqua bidest. vermischt und durch Ansäuern mit 1 n HCl in Lösung gebracht.
Die Einstellung des pH-Wertes auf 4,5 erfolgte mittels eines Puffergemisches aus 0,1
n Kaliumhydrogenphosphat und 0,1 n Natronlauge (Mischverhältnis von 3,17:1, pH
6,5). Die Lösung wurde mit sterilem Aqua bidest. auf 1 l aufgefüllt, in 10 Portionen a
100 ml aufgeteilt und im Kühlschrank bei +2 °C in s terilen Glasflaschen aufbewahrt.
Entsprechend des Körpergewichtes der Tiere wurde am Abend des ersten
Versuchstages die Insulininfusionslösung in einer solchen Konzentration hergestellt,

44 Material und Methoden
dass den Tieren bei einer Infusionsmenge von 1 ml/min 1 mU/kgKGW/min infundiert
wurde. Der Lösung des dritten Versuchstages wurden 120 mmol Kaliumchlorid/l
zugesetzt, so dass die infundierte Kaliummenge 0,5 mmol/Tier/min betrug.
Zur Herstellung der Chrom-EDTA-Lösung wurden zunächst 143,88 g CrCl3 * 6H2O in
1l Aqua dest. unter Rühren mit einem Glasstab und 201 g Na2-EDTA * 2H2O in 1,5 l
Aqua dest. mit Zusatz von einigen Glasperlen unter Erwärmen gelöst. Anschließend
wurden beide Lösungen zusammengegossen und eine Stunde unter Rühren
gekocht. Nach Abkühlung und Lösen von 5,88 g CaCl2 * 2H2O wurde der pH-Wert
mit Hilfe von 10 n NaOH auf 6,0 eingestellt und die Lösung auf 3 l aufgefüllt.
Für die Herstellung der initialen Infusionslösung wurden 230 ml der Chrom-EDTA-
Lösung mit 770 ml Aqua dest. und für die folgenden Zugaben 90 ml Chrom-EDTA-
Lösung mit 510 ml Aqua dest. verdünnt.
3.2.2.3 Ablauf des hyperinsulinämischen, euglykämischen Clamptests mit
gleichzeitiger Labmageneffluxmessung
Ein Versuchsabschnitt dauerte drei Tage. Zu Beginn der Versuchstage wurde jeweils
eine klinische Allgemeinuntersuchung durchgeführt.
Tag 1: Um 6 Uhr wurde der erste Venenverweilkatheter für die Blutentnahme gelegt
und fixiert. Kurz vor 7 Uhr wurde eine erste Duodenalsaftprobe entnommen (100 ml)
und der pH-Wert gemessen (dieser lag stets zwischen 2,14 und 2,98), bevor um 7
Uhr die initiale Chrom-EDTA-Lösung und im Folgenden alle 10 Minuten 20 ml der
verdünnten Lösung in den Pansen gegeben wurde. Des weiteren wurden alle 20
Minuten bis 12 Uhr weitere Duodenalchymusproben genommen. Ab 7 Uhr 30
erfolgte zusätzlich alle 10 Minuten eine Blutentnahme mittels einer 2 ml Spritze zur
Bestimmung von Glucose im Vollblut bzw. mittels Serumröhrchen zur Bestimmung
von Kalium und Insulin im Serum. Nach Gerinnung wurden die Serumproben 10
Minuten bei 4000 U/min zentrifugiert und das Serum anschließend mit Hilfe von
Transferpipetten in Eppendorfgefäße (1 ml) überführt. Die Glukosewerte konnten
unmittelbar nach der Beprobung mit Hilfe eines Blutzuckermessgerätes bestimmt
werden. Nach Entnahme der Blutproben wurden die Katheter jeweils mit 10 ml
physiologischer Kochsalzlösung gespült und nach der letzten Probe mit dem
zugehörigen Inlet/Platzhalter versehen.

Material und Methoden 45
Um 12 Uhr 15 wurde der zweite Venenverweilkatheter gelegt und das Versuchstier
antibiotisch mit Procain-Penicillin (30.000 I.E./kg) versorgt. Gegen 15 Uhr wurden die
Chymusproben 15 Minuten bei 19000 U/min mit einem Sorvall SS-34-Rotor
zentrifugiert und der Überstand in Szintillationsgefäße pipettiert. Die Serum- und
zentrifugierten Chymusproben wurden bei –20 °C eing efroren und zu späteren
Zeitpunkten jeweils in kleinen Portionen für die weiteren Analysen aufgetaut.
Tag 2 und 3: Zusätzlich zum Ablauf des ersten Tages wurden von 8 Uhr bis 12 Uhr
die Insulinpumpe und die Glukoseinfusionsflasche mittels Infusionsbesteck am
Dreiwegehahn angeschlossen. Erstere wurde auf eine Infusionsrate von 1 ml/min
eingestellt, die Glukoseinfusion je nach Blutglukosewerten korrigiert. Um eine
genaue Infusionsmenge zu garantieren, wurde das Gewicht der Insulinlösung vor
und nach der Infusion bestimmt. Zusätzlich wurde eine Probe der
Insulininfusionslösung in Eppendorf-Cups pipettiert und bis zur Insulinbestimmung
zusammen mit den Chymus- und Serumproben bei –20 °C aufbewahrt.
Nach der vierstündigen Insulininfusionsperiode schloss sich eine eineinhalbstündige
Phase der Nachkontrolle an, in der zunächst Glukose über eine halbe Stunde weiter
infundiert und schließlich noch über eine weitere Stunde der Blutglukosespiegel
kontrolliert wurde.
Am letzten Versuchstag wurden gegen 14 Uhr die Katheter entfernt und die Tiere
abschließend untersucht.
3.2.3 Analysen
3.2.3.1 Insulin
Die Bestimmung der Insulinkonzentration im Serum erfolgte mittels eines für bovines
Insulin spezifischen ELISAs, einem „solid phase two-site enzyme immunoassay“.
Das bei Raumtemperatur aufgetaute Probandenserum (25 µl) wurde im
Doppelansatz auf eine mit monoklonalen, gegen Rinderinsulin gerichteten
Antikörpern beschichtete Mikrotiterplatte pipettiert und mit 100 µl einer Lösung
überschichtet, die monoklonale Peroxidase-konjugierte, gegen Rinderinsulin
gerichtete Antikörper enthielt. Nach einer Inkubationszeit von 2 h auf einem ELISA-
Schüttler (700 rpm) wurden in einem sechsfachen Waschvorgang ungebundene

46 Material und Methoden
Peroxidase-konjugierte Antikörper entfernt und die gebundenen mit Hilfe von 3,3´-
5,5´-tetramethylbenzidin (200 µl) dargestellt (Blaufärbung). Nach 15 Minuten wurde
die Reaktion mit 50 µl einer 0,5 molaren H2SO4-Lösung gestoppt (Gelbfärbung) und
innerhalb der nächsten halben Stunde spektrophotometrisch bei einer Wellenlänge
von 450 nm gemessen.
Mit den im Test enthaltenen Standards aus bovinem Insulin (0,05; 0,15; 0,5; 1,5 und
3 µg/l) und einem Nullstandard wurden an jedem Messtag Eichkurven erstellt.
Der Standardbereich des Tests lag bei 0,05-3,0 µg/l bei einer analytischen
Sensitivität von 0,025 µg/l. Für den Variationskoeffizienten der eigenen Proben
wurde 8,15 % ermittelt.
3.2.3.2 Kalium
Die Bestimmung von Kalium im Serum erfolgte gegen 15 Uhr des selben
Versuchstages. Das Probandenserum wurde im Labor der Klinik für Rinder,
Tierärztliche Hochschule, mittels einer ionensensitiven Elektrode (614 Na+/K+
Analyser) analysiert.
3.2.3.3 Chrom
Die Chromkonzentration im Duodenalchymus wurde mittels
Atomabsorptionsspektrometrie (UNICAM 969) im Institut für Tierernährung,
Tierärztliche Hochschule Hannover, bestimmt. Dazu wurden die Proben vor dem
Einfrieren und nach dem Auftauen mit 19000 rpm zentrifugiert.
Ein Flammen-Atomabsorptionsspektrometer besteht aus einer Lichtquelle
(Hohlkathodenlampe), der Absorptionszelle (Flamme), dem Monochromator (zur
Trennung der Resonanzlinie von anderen Spektrallinien) und dem Detektor
(Photomultiplier). Freie Metallatome werden durch Atomisierungsquellen erzeugt; die
Atomisierung kann dabei in der Flamme (Luft/Acetylen) erfolgen.
Das Prinzip der Atomabsorptionsspektrometrie beruht auf dem Lambert-Beer-
Gesetz:

Material und Methoden 47
Io
A= ----- = k * d * c Id
Io: Intensität des eingestrahlten Lichts, Id: Intensität des durchgestrahlten Lichts, k:
Konstante, c: Konzentration des Chroms, d: Schichtdicke
Die Absorption ist also proportional der durchstrahlten Schichtdicke und der
Konzentration des absorbierenden Stoffes, umgekehrt lässt die gemessene
Absorption Rückschlüsse auf die Konzentration des zu messenden Stoffes im
Untersuchungsmaterial zu.
3.2.3.4 Berechnung des Labmageneffluxes
Bei der im Folgenden beschriebenen Methode zur Berechnung des
Labmageneffluxes liegt die Annahme zugrunde, dass dieser mit dem Duodenalfluss
übereinstimmt, da die Berechnung des Labmageneffluxes mit Hilfe der gemessenen
„steady state“-Chromkonzentration im Duodenal- und nicht im Labmagenchymus
erfolgte. Dieser „steady state“ war dann erreicht, wenn sich das Chrom gleichmäßig
auf Vormägen und Labmagen verteilt hatte und die in den Pansen gegebene
Chrommenge der aus dem Labmagen abfließenden entsprach. Vorversuche, die an
drei Kühen zur Überprüfung der Methode zur Bestimmung des Effluxes durchgeführt
wurden, ergaben, dass dieser Zustand nach drei bis vier Stunden erreicht war
(Abbildung 3).

48 Material und Methoden
0
500
1000
1500
2000
120 180 240
min nach der ersten Chromzugabe
erre
chne
te E
fflux
men
ge in
m
l/min
Abbildung 3: Effluxmessungen während der Vorversuch e mit Erreichen des steady states nach
ca. drei bis vier Stunden
Der Efflux wurde dabei anhand der Chromwerte im Duodenalchymus und der pro
Minute zugegebenen Menge Chrom in den Pansen nach folgender Formel
berechnet:
Zugegebene Menge Chrom in mg/min
--------------------------------------------------------------- = l/min gemessene Chromkonzentration in mg/l Da allerdings an Tag 2 und 3 zum Zeitpunkt 0 die Chromwerte noch erhöht waren
von den Gaben am Vortag, mussten diese von den für die Effluxmessung genutzten
Werten subtrahiert werden. Jedoch ist in den verbleibenden vier Stunden vom
Zeitpunkt 0 bis zum steady state ein weiteres Abnehmen der verbliebenen
Chromkonzentration des Vortags anzunehmen. Diese Abnahme erfolgt exponentiell:
t = t0*e-yx
t0 = letzter Chromwert am Vortag; y = Chromeliminationsrate;
x = aktueller Messzeitpunkt

Material und Methoden 49
So konnte aus dem letzten Chromwert des Vortags und den ersten des aktuellen
Versuchstages, an denen das neu zugegebene Chrom das Duodenum noch nicht
erreicht hatte, der zu subtrahierende Chromwert kalkuliert werden.
3.3 Statistik
Die statistische Auswertung wurde unter Verwendung des Statistikprogrames SAS
Version 9.1 durchgeführt. Die Erstellung der Graphiken erfolgte mit dem Programm
Microsoft® EXCEL des Programmpaketes Office XP Professional 2003.
Zur Auswertung der Ergebnisse des In-vitro-Versuches wurden die Medianwerte der
Motilitätsparameter mit den jeweiligen Minima und Maxima errechnet, da in der
überwiegenden Anzahl die Daten nicht normal verteilt waren. Die Normalverteilung
wurde mit dem Test nach Kolmogorov-Smirnov überprüft.
Die angegebenen n-Zahlen entsprechen der Anzahl der Versuchstiere. Der Einfluss
der Testsubstanzen wurde zunächst mit der einfaktoriellen Varianzanalyse überprüft;
der Vergleich zweier Medianwerte sowohl zwischen den einzelnen Konzentrationen
als auch zwischen Versuchs- und Kontrollkurve erfolgte anschließend mittels signed-
rank-Test für verbundene Stichproben.
Im In-vivo-Versuch wurden die Mittelwerte für Kalium, Insulin und den
Labmagenefflux mit Hilfe einer einfaktoriellen Varianzanalyse für verbundene
Stichproben verglichen.
Alle Tests wurden auf einem Signifikanzniveau von p<0,05 durchgeführt.

50 Ergebnisse
4 Ergebnisse
4.1 In-vitro-Versuch
4.1.1 Aktivität der circulären Corpusmuskulatur
4.1.1.1 Basale Aktivität der circulären Corpusmuskulatur
Um den Einfluss der Zeit auf die Vorgänge an der isolierten Muskulatur unter Kalium-
oder Insulingaben einschätzen zu können, wurde zunächst das
Kontraktionsverhalten über einen Zeitraum von 2,5 h unter normokaliämischen
Bedingungen getestet. Präparate der circulären Corpusmuskulatur der Ziege zeigten
über die gemessene Zeit von 2,5 Stunden bei einer extrazellulären
Kaliumkonzentration von 4,7 mmol/l einen Anstieg der mittleren Amplitude bei in
etwa gleichbleibender Frequenz (Tabelle 3). Daraus ergab sich auch für die
Kontraktionsaktivität (Abbildung 4) eine signifikante Zunahme von 11,13 (3,97;58,27)
auf 48,49 (26,09;193,65) mN/min.
Tabelle 3: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf von Amplitude und Frequenz an der circuläre n Corpusmuskulatur der Ziege unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
Zeitpunkt
(h nach Versuchsbeginn)
Amplitude (mN) Frequenz (1/min)
0 2,6 (0,9;10,6)a 5 (3;7)
0,5 2,1 (0,8;13,7)a,b 6 (3;6)
1 3,1 (1,2;16,9)b,c 6 (3;8)
1,5 7,5 (1,1;15,5)b,c 7 (3;12)
2 9,0 (2,1;14,5)c 9 (3;9)
2,5 10,1 (3,7;27,7)c 7 (3;8)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte ein Einfluss
der Zeit auf die Amplitude circulärer Corpusmuskula tur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Zeitpunkten sind
mit unterschiedlichen Buchstaben gekennzeichnet.

Ergebnisse 51
0 0,5 1 1,5 2 2,50
10
20
30
40
50
60
Zeit (min)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 4: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur der Ziege unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte ein Einfluss
der Zeit auf die Kontraktionsaktivität circulärer C orpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Zeitpunkten sind
mit unterschiedlichen Buchstaben gekennzeichnet.
Im Gegensatz zur Ziege zeigten die Präparate der bovinen circulären
Corpusmuskulatur vom Rind bei 4,7 mmol Kalium/l über die Zeit eine Abnahme der
Amplitude bei zunächst zunehmender, nach einer Stunde dann aber abnehmender
Kontraktionsaktivität (von 24,72 (6,87;246,23) auf 39,14 (7,06;161,03) auf 23,54
(7,65;150,29) mN/min) und unveränderter Frequenz (Tabelle 4; Abbildung 5).
a a
a,b
a,b b b
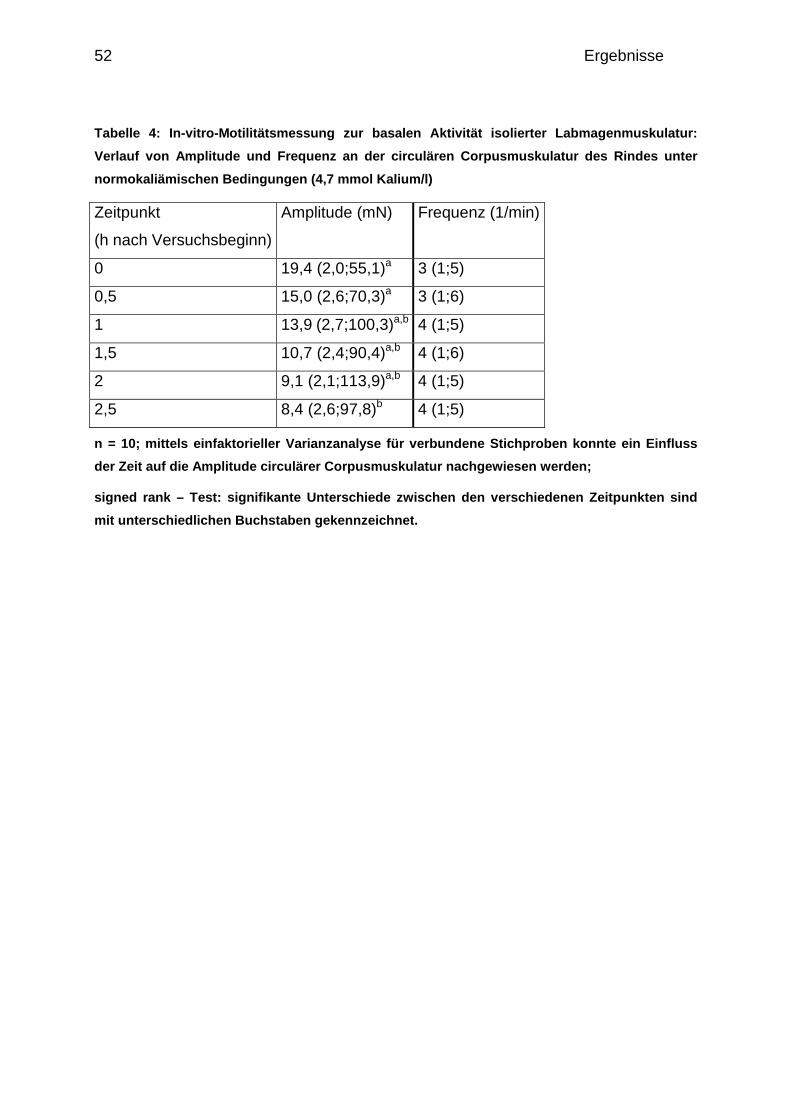
52 Ergebnisse
Tabelle 4: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf von Amplitude und Frequenz an der circuläre n Corpusmuskulatur des Rindes unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
Zeitpunkt
(h nach Versuchsbeginn)
Amplitude (mN) Frequenz (1/min)
0 19,4 (2,0;55,1)a 3 (1;5)
0,5 15,0 (2,6;70,3)a 3 (1;6)
1 13,9 (2,7;100,3)a,b 4 (1;5)
1,5 10,7 (2,4;90,4)a,b 4 (1;6)
2 9,1 (2,1;113,9)a,b 4 (1;5)
2,5 8,4 (2,6;97,8)b 4 (1;5)
n = 10; mittels einfaktorieller Varianzanalyse für verbunde ne Stichproben konnte ein Einfluss
der Zeit auf die Amplitude circulärer Corpusmuskula tur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Zeitpunkten sind
mit unterschiedlichen Buchstaben gekennzeichnet.

Ergebnisse 53
0 0,5 1 1,5 2 2,50
100
200
300
Zeit (h)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 5: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur des Rindes unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte kein Einfluss
der Zeit auf die Kontraktionsaktivität circulärer C orpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Zeitpunkten sind
mit unterschiedlichen Buchstaben gekennzeichnet.
4.1.1.2 Wirkung steigender Kaliumgaben auf die circuläre Corpusmuskulatur
Die Wirkung von Kalium auf die isolierte Labmagenmuskulatur wurde anhand einer
Kalium-Dosiswirkungskurve getestet. Equimolare Zugaben von NaCl zu
benachbarten Gewebestücken dienten dabei als Kontrolle für Zeit- und osmotische
Effekte (Tabelle 5). Zusätzlich wurden am selben Präparat NaCl- und KCl-Zugaben
nach zwischenzeitlichem Washout getestet.
a,b b
a,b a,b a,b
a

54 Ergebnisse
Tabelle 5: Versuchsschema zur Überprüfung der Kaliu mwirkung auf isolierte
Labmagenmuskulatur der Ziege
Kammer 1 Kammer 2
KCl (DWK-1) NaCl
Washout
Zeitvergleich
KCl (DWK-2)
Kammervergleich
DWK = Dosiswirkungskurve
Die circuläre Corpusmuskulatur der Ziege reagierte auf steigende Zugaben von KCl
mit einer Zunahme der Amplitude und auch der Kontraktionsaktivität (von 7,28
(0;54,25) auf 90,07 (33,87;228,08) mN/min) bei gleichbleibender Frequenz. In der
Kontrolle mit steigenden NaCl-Gaben kam es zu geringeren Zunahmen von
Amplitude und Kontraktionsaktivität 8,58 (0;58,86) auf 35,73 (10,84;167,60) mN/min)
bei ebenfalls unbeeinflusster Frequenz (Abbildung 6 und Tabelle 6). Beim Vergleich
der NaCl- und KCl-Effekte am selben Präparat zeigten sich deutliche Zeiteffekte, da
die später im Versuch applizierten Kaliummengen zu geringeren Reaktionen der
Muskelstreifen führte (Abbildung 7). Bei wiederum unveränderter Frequenz fiel die
Zunahme der Amplitude signifikant geringer aus. Daraus resultierte ein Anstieg der
Kontraktionsaktivität von lediglich 13,10 (0;62,54) auf 55,77 (7,50;105,16) mN/ min
bei 7 mmol/l bzw. 48,83 (10,30;85,45) mN/min bei 10 mmol Kalium/l. Beim Vergleich
der beiden Kalium-Dosiswirkungskurven, die um ca. 3 h zeitversetzt liegen, ist die
Abnahme der Parameter erkennbar (Abbildung 8).

Ergebnisse 55
Tabelle 6: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf von Frequenz und Amplitude an der circuläre n
Corpusmuskulatur der Ziege unter dem Einfluss steig ender Kaliumkonzentrationen
[K+] in
mmol/l
Amplitude (mN) Frequenz (1/min)
K+-DWK-1 Kontrolle
(NaCl)
K+-DWK-2 K+-
DWK-1
Kon-
trolle
K+-
DWK-2
4,7 3,4
(0;28,4)a,c,d
5,6
(1,5;18,7)a
5
(0;12)
6
(4;7)
1 1,1
(0;7,9)b
1,5
(0;7,4)b
2,5
(0;8,3)a
4
(0;7)
5
(0;8)
6
(0;8)
3 3,8
(0,35,0)a
2,6
(0;6,3)b
3,6
(0;9,9)a
5
(0;13)
4
(0;12)
4
(0;8)
4 6,2
(0;35,9)c
3,0
(0;6,7)b
3,8
(2,5;11,6)b
5
(0;14)
5
(0;13)
4
(2;8)
5 6,4
(0,9;41,2)d
3,6
(0;8,0)b
5,6
(2,2;14,7)b
6
(3;13)
6
(0;14)
5
(3;8)
6 9,5
(1,7;54,0)e * 4,4
(0,7;14,2)a,b * 8,1
(2,0;18,8)c
6
(3;12)
6
(4;10)
5
(3;9)
7 11,0
(1,7;54,0)e * 6,5
(0,8;15,7)a
8,4
(1,7;19,5)c
6
(3;12)
6
(4;9)
5
(3;8)
10 12,5
(3,7;76,5)e * 7,0
(1,7;19,7)a
8,3
(2,4;23,1)b,c
6
(3;16)
6
(5;9)
5
(3;7)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Amplitude circulärer Corpusmusku latur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert. Die Amp lituden von K +-DWK-1 und K +-DWK-2
unterscheiden sich bei einer Kalium - Konzentration von 10 mmol/l.
DWK = Kalium-Dosiswirkungskurve zu Beginn (K +-DWK-1) und im Anschluss an die NaCl-
Zugaben (K +-DWK-2)

56 Ergebnisse
4,7 1 3 4 5 6 7 100
50
100
150
200
250
K-DWK-1 Kontrolle
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 6: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
der Ziege unter dem Einfluss steigender KCl-Gaben ( Kammervergleich)
Bei Kontrollgeweben wurden gleichzeitig equimolare Mengen NaCl zugegeben.
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Corpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
a b a,c,d c d e e e
b b b b a,b a a a
* *

Ergebnisse 57
1 3 4 5 6 7 100
100
200
KontrolleK-DWK-2
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 7: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
der Ziege unter dem Einfluss steigender KCl-Gaben ( Zeitvergleich)
Bei Kontrollgeweben wurden gleichzeitig equimolare Mengen NaCl zugegeben.
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Corpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben ergaben sich nicht.
a a a a b a,b b
a a b b c c b,c

58 Ergebnisse
1 3 4 5 6 7 100
50
100
150
200
250
K-DWK-1 K-DWK-2
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 8: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
der Ziege unter dem Einfluss von Kalium-Dosiswirkun gskurven an unterschiedlichen
Präparaten und zu unterschiedlichen Zeitpunkten
n = 10; signed rank – Test: signifikante Unterschie de zwischen den beiden Kurven sind mit *
markiert.
Ähnliche Verhältnisse fanden sich an der circulären Corpusmuskulatur des Rindes
wieder: Auch hier äußerte sich der K+-Effekt in einer signifikanten Erhöhung der
Amplitude und demzufolge der Kontraktionsaktivität von 9,36 (0;78,38) auf 29,87
(0;157,73) mN/min (jeweils mit p<0,05) ohne Einflussnahme auf die Frequenz. In der
NaCl-Kontrolle sank sowohl die Frequenz als auch die Kontraktionsaktivität (von 7,85
(0;48,56) auf 2,89 (0;21,19) mN/min) bei unveränderter Amplitude ab (Abbildung 9).
Allerdings war, wie Abbildung 10 und Tabelle 8 zeigen, auch beim Rind eine
zusätzliche Abnahme der Parameter über die Zeit nachweisbar: Stiegen bei zwei
aufeinander folgenden Kalium- Dosiswirkungskurven bei gleichbleibender Frequenz
Amplitude und Kontraktionsaktivität zunächst noch von 9,25 (0;100,36) auf 34,43
(0;178,05) mN/min bei 6 mmol Kalium/l bzw. 29,72 (0;145,68) mN/min bei 10 mmol
Kalium/l an, so kam es bei der Wiederholung zu geringeren Veränderungen.
* *

Ergebnisse 59
Tabelle 7: Versuchsschema zur Überprüfung der Kaliu mwirkung auf isolierte
Labmagenmuskulatur vom Rind
DWK = Dosiswirkungskurve
Kammer 1 Kammer 2
Kontrolle (NaCl) KCl (K+-DWK-1)
Washout
Zeitvergleich
KCl (K+-DWK) KCl (K+-DWK-2)
Kammervergleich

60 Ergebnisse
Tabelle 8: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf von Frequenz und Amplitude an der circuläre n
Corpusmuskulatur des Rindes unter dem Einfluss stei gender Kaliumkonzentrationen
Amplitude (mN) Frequenz (1/min)
Kammervergleich Zeitvergleich Kammervergleich Zeitvergleich
[K+]
in
mM Kontrolle
(NaCl)
K+-DWK K+-
DWK-1
K+-
DWK-2
Kontrolle
(NaCl)
K+-
DWK
K+-
DWK-
1
K+-
DWK-
2
4,7 8,6
(0,7;24,6)a
9,0
(0;42,2)a
4
(1;9)a
4
(0;10)a
1 3,2
(0;7,6)b
2,6
(0,25,4)b
2,6
(0;18,2)a
3,0
(0;14,3)a
2
(0;9)a
3
(0;7)a
4
(0;7)
2
(0;6) 3 2,4
(0;7,6)b
3,1
(0;23,5)b
4,2
(0;35,7)b
2,8
(0;6,5)a
2
(0;9)a
4
(0;9)a
5
(0;6)
2
(0;7) 4 1,8
(0;5,3)b
4,2
(0;26,6)b,c
5,3
(0;42,3)b
2,6
(0;8)a
2
(0;10)a
4
(0;9)a
5
(0;7)
3
(0;6) 5 2,9
(0;7,3)b
4,5
(0;40,6)c
5,5
(0;42,2)b
* 2,7
(0;15,6)a
2
(0;10)a
3
(0;9)a
5
(0;8)
5
(0;6) 6 2,6
(0;6,0)b
* 5,0
(0;52,6)c
5,9
(0;35,6)b
4,0
(0;17,3)a
1
(0;4)b
3
(0;7)a
5
(0;7)
3
(0;7) 7 2,3
(0;8,9)b
* 6,1
(0;53,1)c
5,5
(0;37,9)b
4,8
(0;18,4)a
1
(0;3)b
3
(0;6)a
5
(0;7)
4
(0;6) 10 2,6
(0;10,6)b
* 6,3
(0;64,8)c
7,0
(0;32,4)b
5,0
(0;22,2)a
1
(0;2)b
2
(0;7)b
4
(0;6)
4
(0;9)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Amplitude circulärer Corpusmusku latur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.

Ergebnisse 61
4,7 1 3 4 5 6 7 100
100
200
K-DWK Kontrolle
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 9: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
des Rindes unter dem Einfluss steigender KCl-Gaben (Kammervergleich)
Bei Kontrollgeweben wurden gleichzeitig equimolare Mengen NaCl zugegeben.
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Corpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
a b b b c c c c
a b b b,c b,c c c c
* * * * *

62 Ergebnisse
1 3 4 5 6 7 100
100
200
300
K-DWK-1 K-DWK-2
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 10: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
des Rindes unter Einfluss von zwei nacheinander dur chgeführten Kalium-
Dosiswirkungskurven an demselben Präparat (Zeitverg leich)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Corpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
4.1.1.3 Wirkung steigender Insulingaben auf die circuläre Corpusmuskulatur
Die Wirkung von Insulin wurde ebenfalls in Form einer Dosiswirkungskurve
gemessen und zwar zum einen bei 4,7 mmol KCl/l mit einer Zeitkontrolle und zum
anderen im direkten Vergleich der Kurven bei 3 bzw. 5 mmol KCl/l. Es kam dabei
unter dem Einfluss von Insulin an der circulären Corpusmuskulatur der Ziege im
Vergleich zur Zeitkontrolle bei unveränderter Frequenz zu einer signifikant
geringeren Zunahme von Amplitude und Kontraktionsaktivität: Letztere stieg in
Anwesenheit von Insulin lediglich von 57,3 (0;35,61) auf 27,76 (0;68,87) mN/min, die
Zeitkontrolle hingegen von 11,13 (3,97;58,27) auf 48,49 (26,09;193,65) mN/min
a a a
a a,b a,b a,b b
b b b b b a
* *

Ergebnisse 63
(Abbildung 11). Durch abschließende Erhöhung der Kaliumkonzentration von 4,7 auf
10 mmol/l konnte sowohl mit als auch ohne Insulin nur eine nominelle Steigerung von
Amplitude und Kontraktionsaktivität (auf 29,48 (0;93,59) mN/min mit bzw. 55,67
(29,43;208,36) mN/min ohne Insulin) erzielt werden (Tabelle 9).
Zudem konnten bei Insulin-Dosiswirkungskurven in Anwesenheit unterschiedlicher
Kaliumkonzentrationen (3 und 5 mmol/l KCl) keine Unterschiede in Amplitude,
Frequenz und Kontraktionsaktivität zwischen den beiden Kaliumkonzentrationen
festgestellt werden (Tabelle 10).
Tabelle 9: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude und Frequenz an der circuläre n
Corpusmuskulatur der Ziege unter Einfluss einer Ins ulin-Dosiswirkungskurve mit Zeitkontrolle
(4,7 mmol Kalium/l)
[Insulin] in mU/l
bei 4,7 mmol KCl/l
Amplitude (mN) Frequenz (1/min)
Insulin-DWK Zeitkontrolle Insulin-
DWK
Zeitkontrolle
0 1,4 (0;6,5)a 2,6 (0,9;10,6)a 4 (0;6) 5 (3;7)
7 2,1 (0;7,7)a 2,1 (0,8;13,7)a 5 (0;7) 6 (3;6)
21 3,2 (0;10,4)a,b 3,1 (1,2;16,9)a,b 5 (0;7) 6 (3;8)
42 4,1 (0;13,8)b 7,5 (1,1;15,5)a,b 5 (0;10) 7 (3;12)
80 4,3 0;12,1)b * 9,0 (2,1;14,5)b 5 (0;9) 7 (3;9)
120 3,8 (0;19,8)b * 10,1 (3,7;27,7)b 5 (0;12) 7 (3;8)
120 bei 10 mmol KCl/l 5,9 (0,21,2)b * 11,8 (4,2;34,7)b 4 (0;6) 6 (3;8)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte ein Einfluss
von Insulin auf die Amplitude circulärer Corpusmusk ulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.

64 Ergebnisse
0 7 21 42 80 1200
10
20
30
40
50
60
70
mit Insulin ohne Insulin
Insulinkonzentration (mU/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 11: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
der Ziege unter Einfluss einer Insulin-Dosiswirkung skurve mit Zeitkontrolle (4,7 mmol Kalium/l)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte ein Einfluss
von Insulin auf die Kontraktionsaktivität circuläre r Corpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
a a a,b b b b
a a,b b b ba
* *
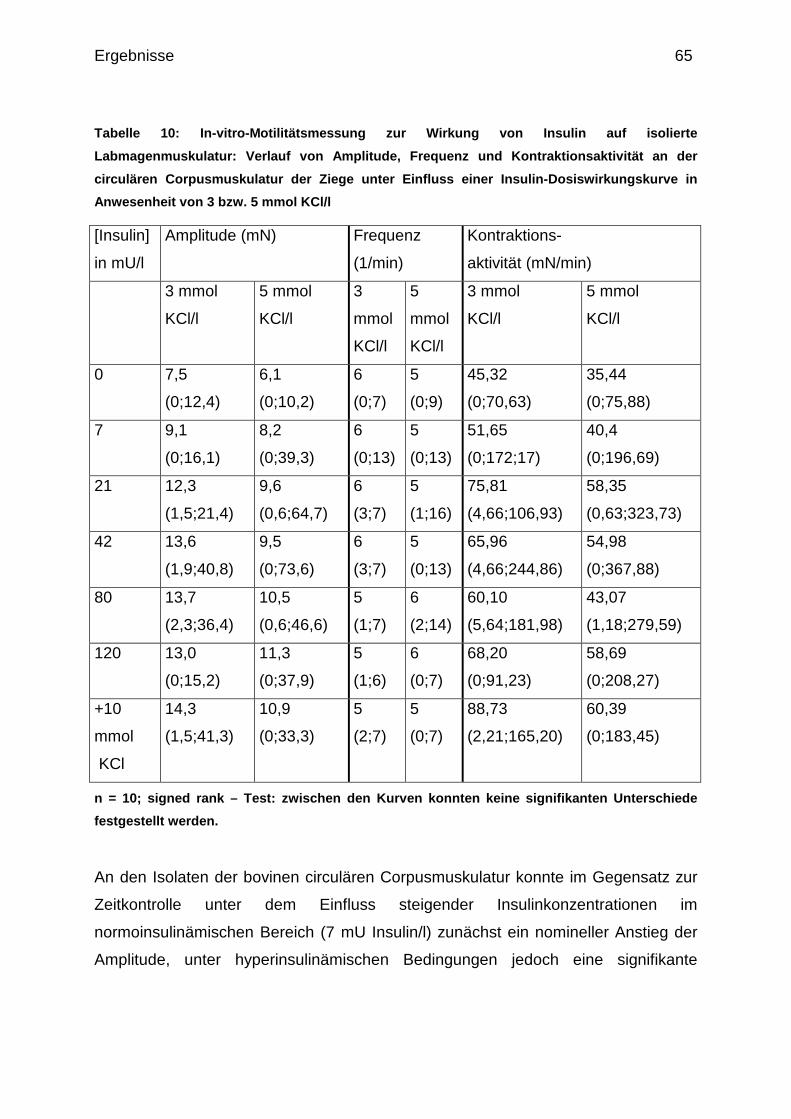
Ergebnisse 65
Tabelle 10: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude, Frequenz und Kontraktionsakt ivität an der
circulären Corpusmuskulatur der Ziege unter Einflus s einer Insulin-Dosiswirkungskurve in
Anwesenheit von 3 bzw. 5 mmol KCl/l
[Insulin]
in mU/l
Amplitude (mN) Frequenz
(1/min)
Kontraktions-
aktivität (mN/min)
3 mmol
KCl/l
5 mmol
KCl/l
3
mmol
KCl/l
5
mmol
KCl/l
3 mmol
KCl/l
5 mmol
KCl/l
0 7,5
(0;12,4)
6,1
(0;10,2)
6
(0;7)
5
(0;9)
45,32
(0;70,63)
35,44
(0;75,88)
7 9,1
(0;16,1)
8,2
(0;39,3)
6
(0;13)
5
(0;13)
51,65
(0;172;17)
40,4
(0;196,69)
21 12,3
(1,5;21,4)
9,6
(0,6;64,7)
6
(3;7)
5
(1;16)
75,81
(4,66;106,93)
58,35
(0,63;323,73)
42 13,6
(1,9;40,8)
9,5
(0;73,6)
6
(3;7)
5
(0;13)
65,96
(4,66;244,86)
54,98
(0;367,88)
80 13,7
(2,3;36,4)
10,5
(0,6;46,6)
5
(1;7)
6
(2;14)
60,10
(5,64;181,98)
43,07
(1,18;279,59)
120 13,0
(0;15,2)
11,3
(0;37,9)
5
(1;6)
6
(0;7)
68,20
(0;91,23)
58,69
(0;208,27)
+10
mmol
KCl
14,3
(1,5;41,3)
10,9
(0;33,3)
5
(2;7)
5
(0;7)
88,73
(2,21;165,20)
60,39
(0;183,45)
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
An den Isolaten der bovinen circulären Corpusmuskulatur konnte im Gegensatz zur
Zeitkontrolle unter dem Einfluss steigender Insulinkonzentrationen im
normoinsulinämischen Bereich (7 mU Insulin/l) zunächst ein nomineller Anstieg der
Amplitude, unter hyperinsulinämischen Bedingungen jedoch eine signifikante

66 Ergebnisse
Abnahme von Amplitude und Kontraktionsaktivität verzeichnet werden. Letztere
sank von 34,53 (3,48;186,39) auf 12,01 (4,41;78,97) mN/min bei 120 mU Insulin/l
(Abbildung 12 und Tabelle 11).
Tabelle 11: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude und Frequenz an der circuläre n
Corpusmuskulatur des Rindes unter Einfluss einer In sulin-Dosiswirkungskurve mit
Zeitkontrolle (4,7 mmol Kalium/l)
[Insulin] in mU/l
bei 4,7 mmol KCl/l
Amplitude (mN) Frequenz (1/min)
Insulin-DWK Zeitkontrolle Insulin-
DWK
Zeitkontrolle
0 22,2 (2,0;115,1)a 19,4 (2,0;55,1)a 3 (1;5) 3 (1;5)
7 25,1 (1,4;147,5)a,b 15,0 (2,6;70,3)a 3 (1;6) 3 (1;6)
21 13,3 (1,6;98,6)b,c 13,9 (2,7;100,3)a,b 3 (1;6) 4 (1;5)
42 14,1 (1,6;97,1)c 10,7 (2,4;90,4)a,b 3 (3;5) 4 (1;6)
80 5,4 (1,7;83,4)d * 9,1 (2,1;113,9)b 3 (1;5) 4 (1;5)
120 6,13 (1,8;65,7)d * 8,4 (2,6;97,8)b 3 (1;5) 4 (1;5)
n = 10; mittels einfaktorieller Varianzanalyse für verbunde ne Stichproben konnte ein Einfluss
von Insulin auf die Amplitude circulärer Corpusmusk ulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.

Ergebnisse 67
0 7 21 42 80 1200
100
200
300
mit Insulin ohne Insulin
Insulin (mU/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 12: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur
des Rindes unter Einfluss einer Insulin-Dosiswirkun gskurve mit Zeitkontrolle (4,7 mmol
Kalium/l)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Insulin auf die Kontraktionsaktivität circuläre r Corpusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
4.1.1.4 Wirkung von Insulin bei steigenden Kaliumgaben auf die circuläre
Corpusmuskulatur
In einem nächsten Schritt wurde geprüft, inwiefern normo- (7 mU Insulin) bzw.
hyperinsuliämische (42 mU Insulin) Bedingungen die Antwort der isolierten
Muskulatur auf eine Kalium-Dosiswirkungskurve beeinflussen. Getestet wurde dies
zeitgleich an drei Präparaten in Anwesenheit von 0, 7 und 40 mU Insulin/l. Dabei
ergaben sich keine Unterschiede zwischen den drei Gruppen (Abbildung 13 und
Tabelle 12).
a a,b a b,c c,d d
a a,b a,b a,b a,b b
* * *

68 Ergebnisse
Tabelle 12: In-vitro-Motilitätsmessung an isolierter Labmagenmu skulatur zur Einflussnahme
verschiedener Insulinkonzentrationen auf den Verlau f einer Kalium-Dosiswirkungskurve:
Verlauf von Amplitude und Frequenz an der circuläre n Corpusmuskulatur der Ziege unter
Einfluss von Kalium-Dosiswirkungskurven bei verschi edenen Insulinkonzentrationen
[Kalium]
in mmol/l
Amplitude (mN)
0 mU
Insulin/l
7 mU
Insulin/l
42 mU
Insulin/l
1 2,5 (0;8,3) 2,8 (0;15,5) 2,7 (0;12,1)
3 3,6 (0;9,9) 3,2 (1,5;23,5) 7,5 (0;11,5)
4 6,2 (2,5;11,6) 6,5 (2,1;20,6) 7,9 (0;12,2)
5 6,4 (2,2;14,7) 7,1 (1,3;22,5) 9,0 (1,2;14,8)
6 9,5 (2,0;18,8) 7,5 (1,7;24,0) 9,7 (1,1;16,4)
7 11,0 (1,7;19,5) 6,6 (1,3;25,6) 9,6 (0,7;18,1)
10 8,3 (2,4;23,1) 8,5 (1,4;16,9) 7,2 (1,0;21,4)
Frequenz (1/min)
1 6 (0;8) 5 (0;7) 6 (0;8)
3 4 (0;8) 6 (5;7) 5 (0;7)
4 4 (2;8) 5 (3;7) 5 (0;7)
5 5 (3;8) 6 (4;8) 6 (5;7)
6 5 (3;9) 6 (4;7) 5 (5;7)
7 5 (3;8) 6 (4;7) 5 (3;7)
10 5 (3;7) 5 (3;6) 5 (4;6)
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
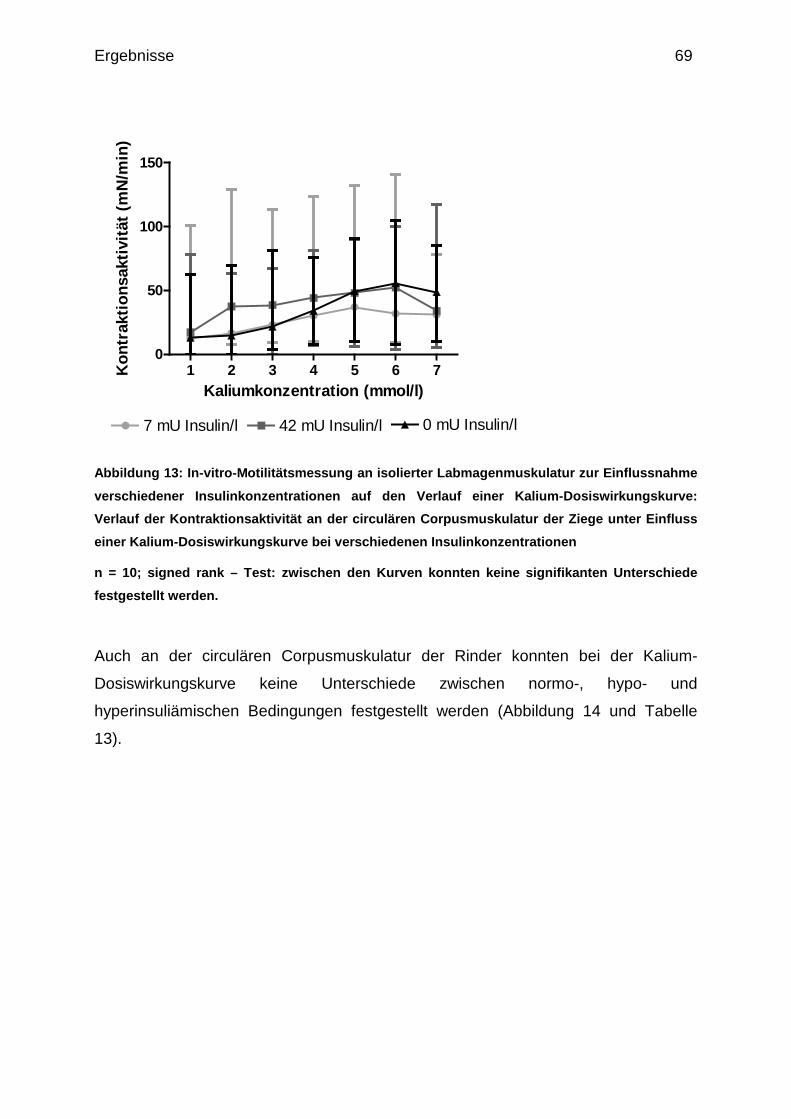
Ergebnisse 69
1 2 3 4 5 6 70
50
100
150
7 mU Insulin/l 42 mU Insulin/l 0 mU Insulin/l
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 13: In-vitro-Motilitätsmessung an isolierter Labmagenmu skulatur zur Einflussnahme
verschiedener Insulinkonzentrationen auf den Verlau f einer Kalium-Dosiswirkungskurve:
Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur der Ziege unter Einfluss
einer Kalium-Dosiswirkungskurve bei verschiedenen I nsulinkonzentrationen
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
Auch an der circulären Corpusmuskulatur der Rinder konnten bei der Kalium-
Dosiswirkungskurve keine Unterschiede zwischen normo-, hypo- und
hyperinsuliämischen Bedingungen festgestellt werden (Abbildung 14 und Tabelle
13).

70 Ergebnisse
Tabelle 13: In-vitro-Motilitätsmessung an isolierter Labmagenmu skulatur zur Einflussnahme
verschiedener Insulinkonzentrationen auf den Verlau f einer Kalium-Dosiswirkungskurve:
Verlauf von Amplitude und Frequenz an der circuläre n Corpusmuskulatur des Rindes unter
Einfluss von Kalium-Dosiswirkungskurven bei verschi edenen Insulinkonzentrationen
[Kalium] in
mmol/l
Amplitude (mN)
0 mU
Insulin/l
7 mU
Insulin/l
42 mU
Insulin/l
1 3,0 (0;14,3) 3,6 (0;6,6) 4,3 (0;21,7)
3 2,8 (0;6,5) 2,1 (0;6,3) 2,5 (0,5;22,6)
4 2,6 (0;8,0) 2,0 (0,5;5,6) 2,1 (0;18,0)
5 2,7 (0;15,6) 2,8 (0;5,1) 3,7 (0,5;16,9)
6 4,0 (0;17,3) 3,4 (0;5,8) 5,1 (0;17,7)
7 4,8 (0;18,4) 2,7 (0;5,2) 3,7 (0;16,8)
10 5,0 (0;22,2) 3,3 (2,7;5,4) 5,6 (0;20,0)
Frequenz (1/min)
1 2 (0;6) 2 (0;5) 2 (0;4)
3 2 (0;7) 2 (0;4) 2 (1;4)
4 3 (0;6) 3 (1;5) 3 (0;4)
5 5 (0;6) 3 (0;5) 3 (1;6)
6 3 (0;7) 3 (0;5) 3 (0;8)
7 4 (0;6) 4 (0;6) 3 (0;6)
10 4 (0;9) 4 (1;8) 3 (0;6)
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.

Ergebnisse 71
1 2 3 4 5 6 70
50
100
150
7 mU Insulin/l42 mU Insulin/l0 mU Insulin/l
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 14: In-vitro-Motilitätsmessung an isolierter Labmagenmu skulatur zur Einflussnahme
verschiedener Insulinkonzentrationen auf den Verlau f einer Kalium-Dosiswirkungskurve:
Verlauf der Kontraktionsaktivität an der circulären Corpusmuskulatur des Rindes unter
Einfluss einer Kalium-Dosiswirkungskurve bei versch iedenen Insulinkonzentrationen.
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
4.1.1.5 Wirkung von Insulin in Anwesenheit von Barium
Aufgrund der möglichen Wirkung von Insulin über eine veränderte
Membranleitfähigkeit von Kalium wurde in einem nächsten Schritt der Effekt von
Insulin in Anwesenheit des unspezifischen Kaliumkanalblockers Barium (3 mmol/l)
getestet. Nach Zugabe dieses Stoffes nahm die Amplitude bei unveränderter
Frequenz zu. Dadurch stieg die Kontraktionsaktivität von 29,03 (2,55;763,22) vor der
Zugabe auf 278,97 (1,57;1077,63) mN/min) 30 Minuten nach der Zugabe. Die Gabe
von 80 mU Insulin/l verringerte in diesen Versuchen Amplitude und
Kontraktionsaktivität (von 30,80 (5,89;583,60) auf 14,71 (2,94;212,19) mN/min) bei
ebenfalls unbeeinflusster Frequenz. In Anwesenheit von Insulin konnte die Wirkung
von Barium auf Amplitude und Kontraktionsaktivität nur nominell verstärkt werden.
Die Kontraktionsaktivität erhöhte sich durch Barium nach der Insulinhemmung von
14,71 (2,94;212,19) auf 238,97 (1,57;1077,63) mN/min. Umgekehrt hatte Insulin

72 Ergebnisse
nach vorheriger Blockade der Kaliumkanäle durch 3 mmol Barium/l keinen Einfluss
mehr auf die Muskulatur (Abbildung 15 und Tabelle 14).
Tabelle 14: In-vitro-Motilitätsmessung zur Einflussnahme von Ba rium auf die Wirkung von
Insulin auf isolierte Labmagenmuskulatur: Verlauf von Amplituden und Frequenzen an der
circulären Corpusmuskulatur des Rindes unter der Wi rkung von Insulin in Anwesenheit von
Barium
Amplitude (mN) Frequenz (1/min) t (min
nach
Ver-
suchs-
beginn)
Insulin (bei t =
2 min) + Ba++
(bei t = 32)
3 mmol Ba++/l
(bei t =
17 min)
Ba++ (bei t = 17
min) + Insulin
(bei t = 92)
Insulin+
Ba++
3 mmol
Ba++/l
Ba++ +
Insulin
0 7,7
(2,4;129,7) a
7,7
(1,3;152,6) a
9,6
(0,5;94,4) a
4 (3;5) 4 (2;5) 4 (2;5)
15 6,0
(2,4;83,6) a,b
13,0
(2,0;158,0) a
16,3
(0,8;96,0) a
3 (2;5) 4 (2;5) 3 (1;5)
30 5,3
(1,2;64,3) b
52,2
(1,7;190,4) b
15,1
(2,5;161,9) a
3 (2;4) 4 (2;5) 3 (1;4)
45 43,7
(3,8;242,5) c
73,5
(0,8;215,5) b
15,4
(2,3;135,4) a
3 (3;5) 4 (2;5) 3 (2;4)
60 74,0
(7,3;310,2) c
68,0
(1,0;223,6) b
19,5
(2,6;137,3) a
3 (2;4) 4 (2;5) 3 (2;4)
75 55,2
(4,7;323,0) c
59,7
(1,1;211,8) b
29,5
(10,3;241,3) a
3 (2;4) 4 (2;5) 3 (1;4)
90 52,7
(4,8;305,7) c
57,9
(1,2;218,4) b
32,0
(11,3;246,3) b
3 (1;5) 4 (2;5) 3 (2;5)
105 43,6
(6,4;308,5) c
57,3
(1,2;219,5) b
31,0
(10,1;239,4) b
3 (1;4) 4 (2;5) 3 (2;5)
120 39,5
(2,2;288,3) c
54,2
(0,9;210,9) b
27,8
(8,8;237,6) b
3 (1;5) 4 (2;5) 2 (2;4)

Ergebnisse 73
135 45,8
(0,9;298,3) c
56,7
(1,1;201,1) b
26,3
(8,1;247,7) b
3 (1;5) 4 (2;5) 2 (2;5)
150 57,0
(1,1;283,5) c
57,5
(1,2;218,2) b
28,8
(6,7;246,4) b
3 (1;5) 4 (2;5) 2 (2;4)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Barium auf die Amplitude circulärer Corpusmusku latur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
in der Wirkung von Barium auf den Verlauf von Ampli tude und Frequenz der drei Kurven
konnten nicht festgestellt werden.
60 1350
250
500
750
1000
1250 Insulin + Barium
Barium
Barium + Insulin
Zeit (min)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 15: In-vitro-Motilitätsmessung zur Einflussnahme von Ba rium auf die Wirkung von
Insulin auf isolierte Labmagenmuskulatur: Verlauf der Kontraktionsaktivitäten an der circulär en
Corpusmuskulatur des Rindes unter der Wirkung von I nsulin in Anwesenheit von Barium
1 = Zugabe von 80 mU Insulin/l (nach 2 min bei Ins. + Ba.; nach 92 min bei Ba. + Ins.)
2 = Zugabe von 3 mmol Barium/l (nach 17 min bei Ba. und Ba. + Ins.; nach 32 min bei Ins. + Ba.)
Signifikante Unterschiede (p<0,05) der Kontraktions aktivität entsprechen den in Tabelle 14 in
der Spalte „Amplitude“ dargestellten Signifikanzen. Signifikante Unterschiede in der Wirkung
von Barium auf den Verlauf der Kontraktionsaktivitä t der drei Kurven konnten nicht festgestellt
werden.
1 2 2
1

74 Ergebnisse
4.1.1.6 Wirkung von Insulin in Anwesenheit von Glybenclamid
Um die wahrscheinlich vorhandene Insulinwirkung auf die Kaliumkanäle zu
spezifizieren, wurden mittels 0,05 mmol Glybenclamid/l ATP-sensitive Kanäle
geblockt. Durch die Glybenclamidgabe nahmen bei unveränderter Frequenz sowohl
Amplitude als auch Kontraktionsaktivität (von 12,04 (1,96;32,62) auf 18,88 (121,55)
mN/min) unmittelbar nach der Zugabe zu. Die Gabe von 80 mU Insulin/l verringerte
bei ebenfalls unbeeinflusster Frequenz Amplitude und Kontraktionsaktivität.
In Anwesenheit von Insulin kam es nach der Zugabe von Glybenclamid zwar
ebenfalls zu einer Erhöhung von Amplitude und Kontraktionsaktivität (von 5,96
(0;12,65) auf 31,78 (1,77;60,33) mN/min, jedoch konnte kein signifikanter
Unterschied zu dem Anstieg der beiden Parameter ohne Insulin festgestellt werden.
Umgekehrt hatte Insulin nach vorheriger Glybenclamidgabe keinen Einfluss mehr auf
die Muskulatur (Abbildung 16 und Tabelle 15).

Ergebnisse 75
Tabelle 15: In-vitro-Motilitätsmessung zur Einflussnahme von Gl ybenclamid auf die Wirkung
von Insulin auf isolierte Labmagenmuskulatur: Verlauf von Amplituden und Frequenzen an der
circulären Corpusmuskulatur des Rindes unter der Wi rkung von Insulin in Anwesenheit von
Glybenclamid
Amplitude (mN) Frequenz (1/min) t
(min nach
Versuchs-
beginn)
Insulin (bei t = 2
min) + Glyb.
(bei t = 17 min)
0,05 mmol
Glyb./l (bei t
= 2 min)
Glyb. (bei t = 2
min) + Insulin
(bei t = 62 min)
Ins.+
Glyb.
0,05
mmol
Glyb./l
Gyb.
+ Ins.
0 4,3
(1,7;6,0) a
4,3
(1,0;13,0) a
2,8
(1,5;8,8) a
3 (2;4) 3 (2;4) 3 (1;3)
15 3,6
(1,2;6,1) b
5,4
(1,0;16,4) a
4,5
(2,0;11,0) a,b
2 (0;3) 3 (2;4) 3 (1;3)
30 5,2
(1,8;6,7) a
5,2
(2,1;22,3) a,b
4,3
(1,5;14,9) a,b
3 (2;4) 3 (1;4) 4 (1;4)
45 6,9
(1,0;10,2) c
5,4
(1,9;34,7) a,b
4,9
(1,4;21,2) a,b
3 (2;4) 3 (1;4) 3 (1;4)
60 7,5
(0,9;11,7) c
6,4
(1,8;39,7) b
6,1
(1,0;23,9) b
3 (2;4) 3 (1;4) 3 (1;4)
75 8,5
(0,7;13,9) c
7,6
(1,3;40,3) b
5,6
(1,3;24,5) b
3 (2;4) 3 (2;4) 3 (2;4)
90 9,0
(0,8;17,4) c,d
6,3
(1,5;46,3) b
6,6
(1,3;28,6) b
3 (2;4) 3 (2;4) 3 (2;4)
105 10,6
(1,2;20,1) d
7,8
(1,7;45,0) b
7,3
(1,3;30,0) b
3 (2;4) 3 (1;4) 3 (1;4)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Glybenclamid auf die Amplitude circulärer Corpu smuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
in der Wirkung von Glybenclamid auf den Verlauf von Amplitude und Frequenz der drei Kurven
konnten nicht festgestellt werden.

76 Ergebnisse
0 15 30 45 60 75 90 1050
100
200
Insulin + Glybenclamid
Glybenclamid
Insulin + Glybenclamid
Zeit (min)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 16: In-vitro-Motilitätsmessung zur Einflussnahme von Gl ybenclamid auf die Wirkung
von Insulin auf isolierte Labmagenmuskulatur: Verlauf der Kontraktionsaktivitäten an der
circulären Corpusmuskulatur des Rindes unter der Wi rkung von Insulin in Anwesenheit von
Glybenclamid
1 = Zugabe von 80 mU Insulin/l bzw. 0,05 mmol Glybe nclamid/l (nach 2 min)
2 = Zugabe von 0,05 mmol Glybenclamid/l (nach 17 mi n bei Insulin + Glybenclamid)
3 = Zugabe von 80 mU Insulin/l (nach 62 min bei Gly benclamid + Insulin)
Signifikante Unterschiede (p<0,05) der Kontraktions aktivität entsprechen den in Tabelle 15 in
der Spalte „Amplitude“ dargestellten Signifikanzen. Signifikante Unterschiede in der Wirkung
von Glybenclamid auf den Verlauf der Kontraktionsak tivität der drei Kurven konnten nicht
festgestellt werden.
4.1.1.7 Wirkung von Insulin in Anwesenheit von Ouabain
Aufgrund der in der Literatur beschriebenen Insulin-bedingten Stimulation der
Na+/K+-ATPase wurde in einem nächsten Schritt die Wirkung von Insulin in
Anwesenheit von 0,001 mmol Ouabain/l getestet.
Die Gabe dieses Hemmers der Na+/K+-ATPase führte bei unveränderter Frequenz
zunächst zu einem nominellen Anstieg von Amplitude und Kontraktionsaktivität (von
10,15 (2,55;47,82) auf 24,03 (0;127,53) mN/min) unmittelbar nach der Zugabe und
1
3
2

Ergebnisse 77
nach ca. 30 Minuten zu einem Abfall der beiden Parameter. Die Kontraktionsaktivität
verringerte sich im Zuge dessen auf 1,84 (0;28,25) mN/min. Die Gabe von 80 mU
Insulin/l verringerte bei ebenfalls unbeeinflusster Frequenz Amplitude und
Kontraktionsaktivität in diesem Versuch nur nominell.
Die Anwesenheit von Insulin hatte keinen signifikanten Einfluss auf die Wirkung des
Ouabains. Umgekehrt hatte Insulin nach vorheriger Ouabaingabe auch in diesem
Fall keinen Einfluss mehr auf die Muskulatur (Abbildung 17 und Tabelle 16).
Tabelle 16: In-vitro-Motilitätsmessung zur Einflussnahme von Ou abain auf die Wirkung von
Insulin auf isolierte Labmagenmuskulatur: Verlauf von Amplituden und Frequenzen an der
circulären Corpusmuskulatur des Rindes unter der Wi rkung von Insulin in Anwesenheit von
Ouabain
Amplitude (mN) Frequenz (1/min) t
(min
nach
Ver-
suchs-
beginn)
Ins. (bei t =
2 min) +
Ouabain (bei t
= 32 min)
Ouabain (bei t
= 2 min)
Ouabain (bei t
= 2 min) + Ins.
(bei t =
32 min)
Ins.+
Oua-
bain
Oua-
bain
Oua-
bain +
Ins.
0 3,4 (0,9;28,7) a 3,4 (1,1;19,1) a,b 2,4 (0;32,8) a,b 3 (2;5) 3 (2;5) 3 (0;4)
15 3,0 (1,8;28,9) a 4,4 (1,5;20,2) a,b 3,0 (1,0;36,2) a,b 3 (2;4) 3 (2;4) 3 (2;4)
30 2,7 (0;29,2) a 5,2 (0,9;22,3) a 3,1 (1,5;86,3) a 3 (0;3) 3 (2;8) 3 (2;4)
45 3,2 (0;24,8) a 7,2 (0;31,9) a 4,2 (1,3;75,5) a 3 (0;6) 3 (0;8) 3 (2;4)
60 5,3 (0,8;36,7) a 2,0 (0;7,2) b 1,4 (0;14,7) b 3 (2;7) 3 (0;8) 3 (0;4)
75 3,3 (0;12,8) a,b 1,7 (0;8,4) b 1,2 (0;23,6) b 3 (0;9) 3 (0;9) 3 (0;8)
90 1,3 (0;17,3) a,b 1,1 (0;4,3) b 0,7 (0;19,7) b 3 (0;9) 3 (0;8) 2 (0;9)
105 0,8 (0;9,5) b 0,7 (0;3,6) b 0,8 (0;19,9) b 2 (0;9) 3 (0;9) 2 (0;9)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Ouabain auf die Amplitude circulärer Corpusmusk ulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
in der Wirkung von Ouabain auf den Verlauf von Ampl itude und Frequenz der drei Kurven
konnten nicht festgestellt werden.

78 Ergebnisse
0 15 30 45 60 75 90 1050
50
100
150
200
250
Insulin + Ouabain
Ouabain
Ouabain + Insulin
Zeit (min)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 17: In-vitro-Motilitätsmessung zur Einflussnahme von Ou abain auf die Wirkung von
Insulin auf isolierte Labmagenmuskulatur: Verlauf der Kontraktionsaktivitäten an der circulär en
Corpusmuskulatur des Rindes unter der Wirkung von I nsulin in Anwesenheit von Ouabain
1 = 1. Zugabe von 80 mU Insulin/l bzw. 0,001 mmol O uabain/l (nach 2 min)
2 = 2. Zugabe von 80 mU Insulin/l bzw. 0,001 mmol O uabain/l (nach 32 min)
Signifikante Unterschiede (p<0,05) der Kontraktions aktivität entsprechen den in Tabelle 16 in
der Spalte „Amplitude“ dargestellten Signifikanzen. Signifikante Unterschiede in der Wirkung
von Ouabain auf den Verlauf der Kontraktionsaktivit ät der drei Kurven konnten nicht
festgestellt werden.
4.1.2 Aktivität der circulären Pylorusmuskulatur
4.1.2.1 Basale Aktivität der circulären Pylorusmuskulatur
Bei den Präparaten der circulären Pylorusmuskulatur der Ziege blieben über die
gemessene Zeit von 2,5 Stunden unter Standardbedingungen (bei 4,7 mmol KCl/l)
Frequenz, Amplitude und Kontraktionsaktivität weitestgehend konstant (Abbildung 18
und Tabelle 17).
1 2

Ergebnisse 79
Tabelle 17: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf von Amplitude und Frequenz an der circuläre n Pylorusmuskulatur der Ziege unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
Zeitpunkt (h nach Versuchsbeginn) Amplitude (mN) Frequenz (1/min)
0 0 (0;0,5) 0 (0;2)
0,5 0 (0;1,9) 0 (0;3)
1 0 (0;4,8) 0 (0;3)
1,5 0 (0;4,0) 0 (0;3)
2 0 (0;3,7) 0 (0;3)
2,5 0,2 (0;2,9) 2 (0;6)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte kein Einfluss
der Zeit auf Amplitude und Frequenz der circulären Pylorusmuskulatur nachgewiesen werden;
signed rank – Test: es konnten keine signifikanten Unterschiede zwischen den Zeitpunkten
festgestellt werden.
0 0,5 1 1,5 2 2,50
25
50
75
100
125
Zeit (min)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 18: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur der Ziege unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte kein Einfluss
der Zeit auf die Kontraktionsaktivität circulärer P ylorusmuskulatur nachgewiesen werden;
signed rank – Test: es konnten keine signifikanten Unterschiede zwischen den Zeitpunkten
festgestellt werden.
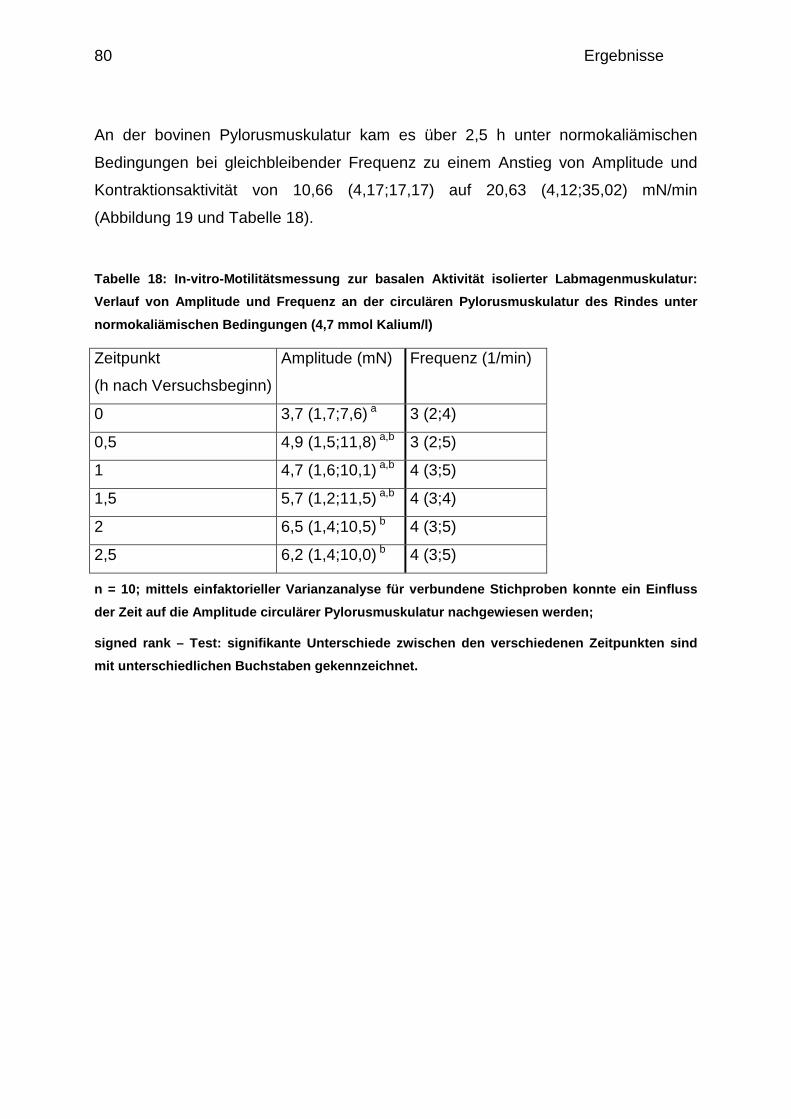
80 Ergebnisse
An der bovinen Pylorusmuskulatur kam es über 2,5 h unter normokaliämischen
Bedingungen bei gleichbleibender Frequenz zu einem Anstieg von Amplitude und
Kontraktionsaktivität von 10,66 (4,17;17,17) auf 20,63 (4,12;35,02) mN/min
(Abbildung 19 und Tabelle 18).
Tabelle 18: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf von Amplitude und Frequenz an der circuläre n Pylorusmuskulatur des Rindes unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
Zeitpunkt
(h nach Versuchsbeginn)
Amplitude (mN) Frequenz (1/min)
0 3,7 (1,7;7,6) a 3 (2;4)
0,5 4,9 (1,5;11,8) a,b 3 (2;5)
1 4,7 (1,6;10,1) a,b 4 (3;5)
1,5 5,7 (1,2;11,5) a,b 4 (3;4)
2 6,5 (1,4;10,5) b 4 (3;5)
2,5 6,2 (1,4;10,0) b 4 (3;5)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
der Zeit auf die Amplitude circulärer Pylorusmuskul atur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Zeitpunkten sind
mit unterschiedlichen Buchstaben gekennzeichnet.

Ergebnisse 81
0 0,5 1 1,5 2 2,505
1015202530354045
Zeit (h)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 19: In-vitro-Motilitätsmessung zur basalen Aktivität is olierter Labmagenmuskulatur:
Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur des Rindes unter
normokaliämischen Bedingungen (4,7 mmol Kalium/l)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
der Zeit auf die Kontraktionsaktivität circulärer P ylorusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Zeitpunkten sind
mit unterschiedlichen Buchstaben gekennzeichnet.
4.1.2.2 Wirkung steigender Kaliumgaben auf die circuläre Pylorusmuskulatur
Die circuläre Pylorusmuskulatur der Ziege reagierte unter steigenden Zugaben von
KCl zunächst mit einer Zunahme von Frequenz, Amplitude und demzufolge auch der
Kontraktionsaktivität. Bei einer Konzentration von 10 mmol Kalium/l kam es jedoch
zu einem Abfall aller drei Parameter, so dass die Kontraktionsaktivität nach dem
Anstieg von 0 (0;62,49) auf 14,47 (0;306,46) mN/min auf 3,75 (0;125,57) mN/min
absank. Allerdings konnten auch im Zuge steigender NaCl-Gaben ein ähnliches
Verhalten aller drei Parameter verzeichnet werden (Abbildung 20 und Tabelle 20). Im
Gegensatz zur circulären Corpusmuskulatur kam es allerdings nicht zu einer
Abnahme der Ansprechbarkeit des Muskels bei zwei hintereinander geschalteten
Dosiswirkungskurven am selben Präparat (Abbildungen 21 und 22).
a
a,b
a,b
b b
a,b

82 Ergebnisse
Tabelle 19: Versuchsschema zur Überprüfung der Kali umwirkung auf isolierte
Labmagenmuskulatur der Ziege
DWK = Dosis
wirkungskurve
Tabelle 20: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf von Frequenz und Amplitude an der circuläre n
Pylorusmuskulatur der Ziege unter dem Einfluss stei gender Kaliumkonzentrationen
[K+]
in
mM
Amplitude (mN) Frequenz (1/min)
K+-DWK 1 Kontrolle
(NaCl)
K+-DWK 2 K+-
DWK 1
Kon-
trolle
K+-
DWK 2
4,7 0,8 (0;31,9)a,b 0,6 (0;38,2)a,b 4 (0;8)a 2 (0;7)a
1 0 (0;18,0)a 0 (0;22,7)a 0,3 (0;7,2)a 0 (0;4)b 0 (0;5)b 1 (0;5)a
3 0,8 (0;26,0)a 0 (0;41,4)a 1,7 (0;116,3)c 2 (0;6)a 0 (0;6)a,b 2 (0;5)a
4 1,2 (0;25,0)a,b 0 (0;102,6)c 2,9 (0;102,5)c 3 (0;7)a 0 (0;6)a,b 2 (0;5)a
5 1,4 (0;33,2)a,b 0,7 (0;95,4)b * 5,4 (0;107,2)c 3 (0;7)a 1 (0;6)a,b 3 (0;6)a
6 1,5 (0;37,8)a,b 1,7 (0;100,2)b * 6,0 (0;117,9)c 4 (0;7)a 2 (0;6)a 3 (0;7)a
7 3,1 (0;38,2)b 1,7 (0;103,3)c 4,4 (0;120,7)c 4 (0;6)a 2 (0;6)a 3 (0;7)a
10 1,2 (0;39,0)a,b 1,0 (0;52,8)a,b 2,7 (0;52,0)a,c 2 (0;6)a 1 (0;5)a,b 2 (0;6)a
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Amplitude und die Frequenz circu lärer Pylorusmuskulatur nachgewiesen
werden; signed rank – Test: signifikante Unterschie de zwischen den verschiedenen
Konzentrationen einer Kurve sind mit unterschiedlic hen Buchstaben gekennzeichnet;
signifikante Unterschiede zu den Kontrollgeweben si nd mit * markiert. Die Amplituden von K +-
DWK-1 und K +-DWK-2 unterscheiden sich bei Kalium - Konzentratio nen von 5 und 6 mmol/l.
DWK = Kalium-Dosiswirkungskurve zu Beginn (K +-DWK-1) und im Anschluss an die NaCl-
Zugaben (K +-DWK-2)
Kammer 1 Kammer 2
KCl (DWK-1) NaCl
Washout
Zeitvergleich
KCl (DWK-2)
Kammervergleich

Ergebnisse 83
4,7 1 3 4 5 6 7 100
100
200
K-DWK-1 Kontrolle
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 20: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
der Ziege unter dem Einfluss steigender KCl-Gaben ( Kammervergleich)
Bei Kontrollgeweben wurden gleichzeitig equimolare Mengen NaCl zugegeben.
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Pylorusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
a a
a a a a a a a a a,b a,b a,b a,b b a,b
*

84 Ergebnisse
1 3 4 5 6 7 100
100
200
300
400
KontrolleK-DWK-2
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 21: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
der Ziege unter dem Einfluss steigender KCl-Gaben ( Zeitvergleich)
Bei Kontrollgeweben wurden gleichzeitig equimolare Mengen NaCl zugegeben.
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Pylorusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
a
a,b a a,b b b b a,b
a a a a a a
*

Ergebnisse 85
1 3 4 5 6 7 100
100
200
300
400
K-DWK-1 K-DWK-2
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 22: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
der Ziege unter Einfluss von Kalium-Dosiswirkungsku rven an unterschiedlichen Präparaten
und zu unterschiedlichen Zeitpunkten
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
An der circulären Pylorusmuskulatur des Rindes äußerte sich der K+-Effekt in einer
Erhöhung von Amplitude und Kontraktionsaktivität (von 3,53 (0;21,63) auf 33,55
(3,78;152,64) mN/min) bei gleichbleibender Frequenz. Dies unterschied sich deutlich
von den Kontrollgeweben, bei denen es unter NaCl-Gaben zu einer Abnahme der
Amplitude (Tabelle 22) und einer nominellen Reduktion der Kontraktionsaktivität (von
7,92 (0;38,06) auf 3,80 (0;10,89) mN/min) kam (Abbildung 23). Zudem war beim Rind
eine Abnahme der Parameter über die Zeit bei einer Hintereinanderschaltung von
zwei Kalium- Dosiswirkungskurven feststellbar, da eine Wiederholung nicht mehr zu
einem signifikanten Anstieg von Amplitude und Kontraktionsaktivität führte
(Abbildung 24).

86 Ergebnisse
Tabelle 21: Versuchsschema zur Überprüfung der Kali umwirkung auf isolierte
Labmagenmuskulatur vom Rind
DWK = Dosiswirkungskurve
Kammer 1 Kammer 2
Kontrolle (NaCl) KCl (K+-DWK-1)
Washout
Zeitvergleich
KCl (K+-DWK) KCl (K+-DWK-2)
Kammervergleich

Ergebnisse 87
Tabelle 22: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf von Frequenz und Amplitude an der circuläre n
Pylorusmuskulatur des Rindes unter dem Einfluss ste igender Kaliumkonzentrationen
Amplitude (mN) Frequenz (1/min) [K+]
in
mM
Kammervergleich Zeitvergleich Kammer-
vergleich
Zeit-vergleich
Kon-
trolle
K+-
DWK
K+-
DWK-1
K+-
DWK-2
Kon-
trolle
K+-
DWK
K+-
DWK-
1
K+-
DWK
-2
4,7 2,0
(0;6,2) a 1,8
(0;9,4) a
3
(0;9)
3
(0;9)
1 2,5
(0;9,5) a 1,5
(0;5,7) a
0,8
(0;3,5) a
1,3
(0;11,5)a
3
(0;9)
3
(0;9)
3
(0;4)
3
(0;4) 3 2,4
(0;7,6) a 1,7
(0;5,4) a
1,0
(0,4;3,3) a
1,2
(0;10,8) a
3
(0;9)
3
(0;9)
4
(3;7)
3
(0;4) 4 2,4
(0;4,3) a 2,1
(0;8,7) a
1,2
(0,4;3,9) a
1,5
(0;9,8) a
3
(0;9)
3
(0;9)
4
(3;5)
4
(0;4) 5 1,8
(0;3,6) a,b 2,8
(0,5;28,7) b
1,1
(0,5;8,8) a
2,4
(0;10,4) a
3
(0;9)
3
(2;9)
4
(3;7)
4
(0;6) 6 1,5
(0;3,0) b * 3,8
(1,1;36,0) b
1,7
(0,4;10,3) a
2,2
(0;11,7) a
3
(0;9)
4
(2;9)
4
(3;6)
4
(0;6) 7 1,2
(0;3,5) b * 4,5
(1,1;38,5) b
2,1
(0,7;13,9) a
2,6
(0;11,7) a
3
(0;9)
4
(3;9)
4
(3;5)
4
(0;7) 10 1,0
(0;3,0) b * 7,5
(1,1;38,2) b
4,2
(1,4;22,7) b
3,1
(0;19,5) a
4
(0;9)
4
(3;9)
4
(3;4)
4
(0;6)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Kalium auf die Amplitude circulärer Pylorusmusk ulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.

88 Ergebnisse
4,7 1 3 4 5 6 7 100
100
200
K-DWK Kontrolle
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 23: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
des Rindes unter dem Einfluss steigender KCl-Gaben (Kammervergleich)
Bei Kontrollgeweben wurden gleichzeitig equimolare Mengen NaCl zugegeben.
n = 10; mittels einfaktorieller Varianzanalyse für verbunde ne Stichproben konnte ein Einfluss
von Kalium auf die Kontraktionsaktivität circulärer Pylorusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
a a a a a,b b b b
a a a a a a a a
* * *

Ergebnisse 89
1 3 4 5 6 7 100
102030405060708090
K-DWK-2K-DWK-1
Kaliumkonzentration (mmol/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 24: In-vitro-Motilitätsmessung zur Wirkung von Kalium a uf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
des Rindes unter Einfluss von zwei nacheinander dur chgeführten Kalium-
Dosiswirkungskurven an demselben Präparat (Zeitverg leich)
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
4.1.2.3 Wirkung steigender Insulingaben auf die circuläre Pylorusmuskulatur
Bei den Ziegenpräparaten nahmen unter Einfluss steigender Insulingaben im
Gegensatz zur Zeitkontrolle sowohl Frequenz als auch Amplitude und
Kontraktionsaktivität zu. Letztere stieg dabei von 0 (0;33,11) auf 12,94 (0;105,56)
mN/min anstatt von 0 (0;0,88) auf 1,20 (0;10,30) mN/min bei der Zeitkontrolle. Die
Erhöhung der Kaliumkonzentration von 4,7 auf 10 mmol KCl/l hatte dabei keinen
Einfluss auf die Parameter weder bei der Zeitkontrolle noch in Anwesenheit von
Insulin (Abbildung 25 und Tabelle 23).

90 Ergebnisse
Tabelle 23: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude und Frequenz an der circuläre n
Pylorusmuskulatur der Ziege unter Einfluss einer In sulin-Dosiswirkungskurve mit Zeitkontrolle
(4,7 mmol Kalium/l)
[Insulin] in mU/l
bei 4,7 mmol KCl/l
Amplitude (mN) Frequenz (1/min)
Insulin-DWK Zeitkontrolle Insulin-DWK Zeitkontrolle
0 0 (0;7,4)a 0 (0;0,6)a 0 (0;5)a,b 0 (0;2)a
7 0 (0;9,9)a 0 (0;1,9)a 0 (0;10)a,b 0 (0;3)a
21 0,3 (0;22,9)a,b 0 (0;4,8)a 1 (0;5)a,b 0 (0;3)a
42 0,8 (0;18,7)a,b 0 (0;4,0)a 2 (0;5)b,c 0 (0;3)a
80 1,8 (0;43,8)b * 0 (0;3,7)a 3 (0;6)c 0 (0;3)a,b
120 3,9 (0;37,8)b * 0,2 (0;2,9)a 3 (0;6)c 2 (0;6)b
120 bei 10 mmol KCl/l 2,0 (0;42,0)b * 0,7 (0;1,9)a 2 (0;5)b,c 1 (0;6)b
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte ein Einfluss
von Insulin auf die Amplitude und die Frequenz der circulären Pylorusmuskulatur
nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.

Ergebnisse 91
0 7 21 42 80 1200
25
50
75
100
125
mit Insulinohne Insulin
Insulin (mU/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 25: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
der Ziege unter Einfluss einer Insulin-Dosiswirkung skurve mit Zeitkontrolle (4,7 mmol Kalium/l)
n = 6; mittels einfaktorieller Varianzanalyse für v erbundene Stichproben konnte ein Einfluss
von Insulin auf die Kontraktionsaktivität circuläre r Pylorusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben sind mit * markiert.
In Anwesenheit geringer Kaliumkonzentrationen (3 mmol KCl/l) fiel im Vergleich mit
höheren Konzentrationen (5 mmol KCl/l) der Anstieg der Parameter unter
Insulineinfluss ähnlich aus. Auch hier konnte keine Reaktion der Muskulatur auf eine
Erhöhung der Kaliumkonzentration auf 10 mmol KCl/l gezeigt werden (Abbildung 26
und Tabelle 24).
a a a a a a
a a a,b a,b b b
* *

92 Ergebnisse
Tabelle 24: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude und Frequenz an der circuläre n
Pylorusmuskulatur der Ziege unter Einfluss einer In sulin-Dosiswirkungskurve in Anwesenheit
von 3 bzw. 5 mmol KCl/l
[Insulin] in mU/l Amplitude (mN) Frequenz (1/min)
3 mmol KCl/l 5 mmol KCl/l 3 mmol KCl/l 5 mmol/l KCl
0 0 (0;1,8) 0 (0;23,4) 0 (0;5) 0 (0;6)
7 1,4 (0;4,9) 0,7 (0;38,7) 1 (0;5) 1 (0;6)
21 0 (0;5,1) 2,8 (0;19,4) 0 (0;4) 2 (0;6)
42 0,6 (0;6,2) 3,4 (0;25,5) 1 (0;5) 2 (0;6)
80 0,3 (0;8,4) 2,9 (0;30,7) 1 (0;5) 2 (0;6)
120 0,6 (0;16,5) 1,7 (0;34,9) 3 (0;5) 3 (0;5)
120 (10 mmol KCl/l) 0,8 (0;24,5) 3,3 (0;57,8) 2 (0;6) 3 (0;6)
n = 6; signed rank – Test: zwischen den Kurven konn ten keine signifikanten Unterschiede
festgestellt werden.

Ergebnisse 93
0 7 21 42 80 1200
50
100
150
5 mM KCL3 mM KCL
Insulin (mU/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 26: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
der Ziege unter Einfluss einer Insulin-Dosiswirkung skurve bei 3 und 5 mmol KCl/l
n = 6; signed rank – Test: zwischen den Kurven konn ten keine signifikanten Unterschiede
festgestellt werden.
An der bovinen Pylorusmuskulatur konnte eine ähnliche Zunahme von Amplitude und
Kontraktionsaktivität unter dem Einfluss einer Insulin-Dosiswirkungskurve
verzeichnet werden wie bei den Präparaten der Ziege, allerdings trat in diesem Fall
der gleiche Effekt auch bei der Zeitkontrolle auf (Abbildung 27 und Tabelle 25).

94 Ergebnisse
Tabelle 25: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude und Frequenz an der circuläre n
Pylorusmuskulatur des Rindes unter Einfluss einer I nsulin-Dosiswirkungskurve mit
Zeitkontrolle (4,7 mmol Kalium/l)
[Insulin] in mU/l
bei 4,7 mmol KCl/l
Amplitude (mN) Frequenz (1/min)
Insulin-DWK Zeitkontrolle Insulin-DWK Zeitkontrolle
0 3,1 (0,7;6,0) a 3,7 (1,7;7,6) a 3 (3;6) 3 (2;4)
7 3,0 (2,3;3,9) a,b 4,9 (1,5;11,8) a,b 3 (3;5) 3 (2;5)
21 4,8 (2,5;6,1) a,b 4,7 (1,6;10,1) a,b 4 (3;5) 4 (3;5)
42 5,7 (3,8;11,2) b 5,7 (1,2;11,5) a,b 4 (3;4) 4 (3;4)
80 5,4 (4,2;8,0) a,b 6,5 (1,4;10,0) b 4 (3;5) 4 (3;5)
120 6,2 (4,9;10,4) b 6,2 (1,4;10,0) b 4 (3;5) 4 (3;5)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte kein Einfluss
von Insulin auf die Amplitude und die Frequenz der circulären Pylorusmuskulatur
nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben ergaben sich nicht.

Ergebnisse 95
0 7 21 42 80 1200
10
20
30
40
50
mit Insulinohne Insulin
Insulin (mU/l)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 27: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
des Rindes unter Einfluss einer Insulin-Dosiswirkun gskurve mit Zeitkontrolle (4,7 mmol
Kalium/l)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte ein Einfluss
von Insulin auf die Kontraktionsaktivität circuläre r Pylorusmuskulatur nachgewiesen werden;
signed rank – Test: signifikante Unterschiede zwisc hen den verschiedenen Konzentrationen
einer Kurve sind mit unterschiedlichen Buchstaben g ekennzeichnet; signifikante Unterschiede
zu den Kontrollgeweben ergaben sich nicht.
Um einen etwaigen Insulineffekt deutlicher von einem reinen Zeiteffekt abgrenzen zu
können, folgte ein weiterer Versuch mit langfristigen Inkubationen in Pufferlösungen
mit verschiedenen Insulinkonzentrationen (40, 120 und 200 mU Insulin/l). Auch hier
konnten gegenüber der Zeitkontrolle ohne Insulin keine signifikanten Veränderungen
festgestellt werden (Abbildung 28 und Tabelle 26).
a a,b a,b b b b
a a,b a,b a,b b b

96 Ergebnisse
Tabelle 26: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude und Frequenz an der circuläre n
Pylorusmuskulatur des Rindes unter Einfluss verschi edener Insulinkonzentrationen mit
Zeitkontrolle (4,7 mmol Kalium/l)
Zeitpunkt (t = min
nach Versuchsbeginn)
Amplitude (mN)
Zeitkontrolle 40 mU
Insulin/l
(bei t = 5 min)
120 mU
Insulin/l
(bei t = 5 min)
200 mU
Insulin/l
(bei t = 5 min)
0 2,9 (1,3;15,2) 1,2 (0,7;24,0) 2,9 (0,5;47,7) 1,9 (0,5;8,5)
30 2,8 (1,3;14,8) 1,5 (0,7;26,8) 3,0 (0;52,4) 2,3 (0,5;12,2)
60 4,7 (0,6;18,1) 2,0 (0,6;24,6) 3,8 (0;59,7) 2,6 (0;17,8)
90 5,1 (0,6;13,7) 2,0 (0,5;23,4) 5,0 (0;60,0) 3,1 (0;27,9)
120 6,8 (0,6;14,2) 2,3 (0,5;19,8) 5,6 (0,9;60,4) 3,1 (0,7;27,5)
150 6,7 (0,7;16,2) 2,7 (0,5;17,0) 6,1 (0,8;63,0) 3,0 (1,1;31,7)
Frequenz (1/min)
0 3 (1;4) 3 (2;3) 3 (1;3) 3 (2;3)
30 3 (2;4) 3 (2;3) 1 (0;3) 3 (2;3)
60 3 (2;4) 3 (2;4) 1 (0;4) 3 (0;4)
90 3 (2;4) 3 (2;4) 1 (1;4) 3 (0;4)
120 3 (2;4) 3 (2;4) 1 (1;4) 3 (2;4)
150 3 (2;4) 3 (2;4) 1 (1;4) 3 (2;4)
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.

Ergebnisse 97
0 30 60 90 120 1500
100
200
ohne Insulin40mU
120mU 200mU
Zeit (min)
Kon
trak
tions
aktiv
ität
(mN
/min
)
Abbildung 28: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf der Kontraktionsaktivität an der circulären Pylorusmuskulatur
des Rindes unter Einfluss verschiedener Insulinkonz entrationen mit Zeitkontrolle (4,7 mmol
Kalium/l)
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
Insulinzugaben erfolgten nach 5 min.
4.1.3 Aktivität der longitudinalen Corpusmuskulatur
4.1.3.1 Wirkung steigender Insulingaben auf die longitudinale Corpusmuskulatur
An der longitudinalen Corpusmusksulatur konnten unter Einfluss einer Insulin-
Dosiswirkungskurve keine Veränderungen in Frequenz, Amplitude und
Kontraktionsaktivität nachgewiesen werden (Tabelle 27). Demzufolge zeigten sich
auch keine Unterschiede in den Reaktionen der isolierten Muskelstücke auf
steigende Insulingaben in Anwesenheit von unterschiedlichen Kaliumkonzentrationen
(3 und 5 mmol KCl/l) (Tabelle 28).

98 Ergebnisse
Tabelle 27: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude, Frequenz und Kontraktionsakt ivität an der
longitudinalen Corpusmuskulatur der Ziege unter Ein fluss einer Insulin-Dosiswirkungskurve
mit Zeitkontrolle (4,7 mmol Kalium/l)
[Insulin]
in mU/l
Amplitude (mN) Frequenz (1/min) Kontraktions-
Aktivität (mN/min)
Insulin-
DWK
Zeitkon-
tolle
Insulin-
DWK
Zeit-
kontrolle
Insulin-DWK Zeitkontolle
0 6,0
(0,7;27,9)
5,4
(0,5;17,1)
4 (2;6) 5 (2;7) 15,84
(2,40;139,30)
18,10
(4,71;131,11)
7 5,7
(0,6;30,1)
4,0
(0;20,0)
4 (2;7) 4 (2;7) 14,52
(1,77;135,53)
18,15
(2,55;99,66)
21 4,7
(0,5;28,4)
3,1
(0,5;16,6)
5 (2;7) 5 (1;7) 16,33
(1,96;155,93)
21,14
(2,35;101,24)
42 5,1
(0,7;26,7)
3,6
(0;28,6)
5 (2;6) 5 (2;7) 17,41
(2,40;146,76)
20,70
(2,35;114,58)
80 7,5
(0,7;25,6)
4,3
(0;37,9)
4 (2;7) 6 (2;7) 21,09
(2,06;128,02)
22,76
(2,75;91,18)
120 6,6
(0,5;22,7)
3,7
(0;24,9)
5 (2;7) 5 (2;7) 21,78
(2,45;101,97)
26,88
(2,16;100,06)
120 (10
mmol
KCl/l)
6,7
(0,7;31,3)
3,9
(0;29,1)
5 (2;6) 6 (2;6) 28,33
(3,43;187,76)
28,99
(1,57;102,42)
n = 10; mittels einfaktorieller Varianzanalyse für verbundene Stichproben konnte kein Einfluss
von Insulin auf die Amplitude, Frequenz und Kontrak tionsaktivität der longitudinalen
Corpusmuskulatur nachgewiesen werden;
signed rank – Test: zwischen den Kurven konnten kei ne signifikanten Unterschiede festgestellt
werden.
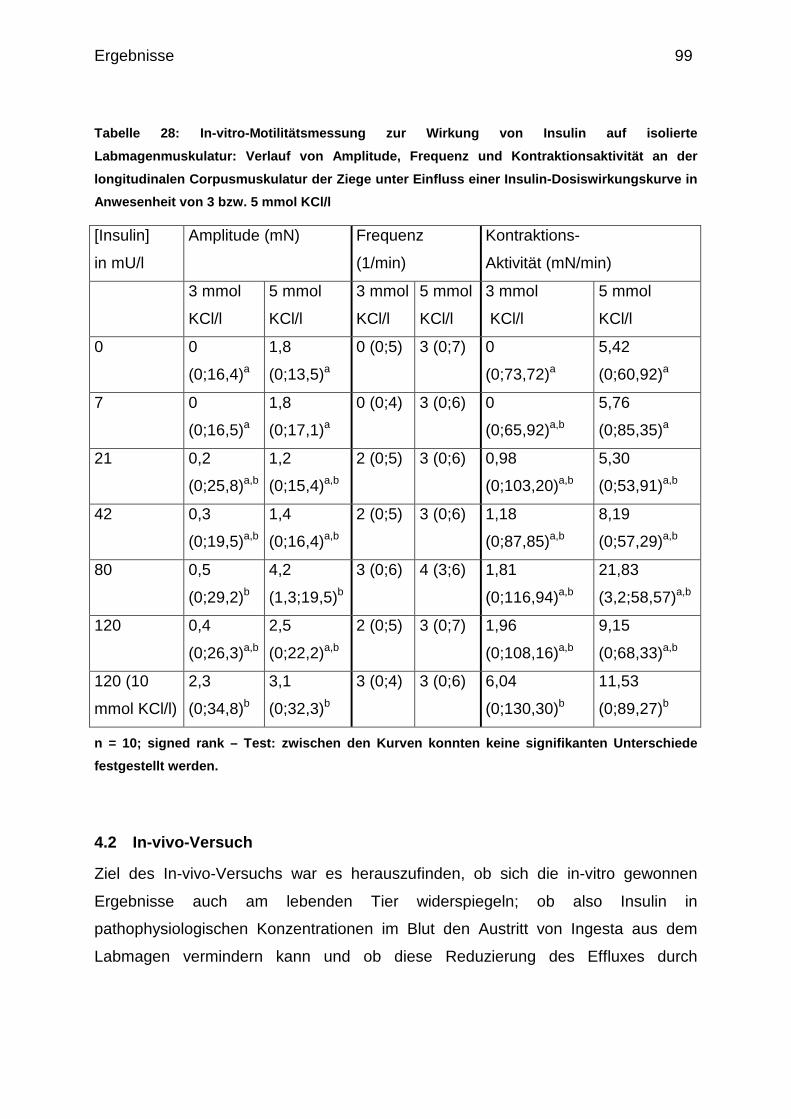
Ergebnisse 99
Tabelle 28: In-vitro-Motilitätsmessung zur Wirkung von Insulin auf isolierte
Labmagenmuskulatur: Verlauf von Amplitude, Frequenz und Kontraktionsakt ivität an der
longitudinalen Corpusmuskulatur der Ziege unter Ein fluss einer Insulin-Dosiswirkungskurve in
Anwesenheit von 3 bzw. 5 mmol KCl/l
[Insulin]
in mU/l
Amplitude (mN) Frequenz
(1/min)
Kontraktions-
Aktivität (mN/min)
3 mmol
KCl/l
5 mmol
KCl/l
3 mmol
KCl/l
5 mmol
KCl/l
3 mmol
KCl/l
5 mmol
KCl/l
0 0
(0;16,4)a
1,8
(0;13,5)a
0 (0;5) 3 (0;7) 0
(0;73,72)a
5,42
(0;60,92)a
7 0
(0;16,5)a
1,8
(0;17,1)a
0 (0;4) 3 (0;6) 0
(0;65,92)a,b
5,76
(0;85,35)a
21 0,2
(0;25,8)a,b
1,2
(0;15,4)a,b
2 (0;5) 3 (0;6) 0,98
(0;103,20)a,b
5,30
(0;53,91)a,b
42 0,3
(0;19,5)a,b
1,4
(0;16,4)a,b
2 (0;5) 3 (0;6) 1,18
(0;87,85)a,b
8,19
(0;57,29)a,b
80 0,5
(0;29,2)b
4,2
(1,3;19,5)b
3 (0;6) 4 (3;6) 1,81
(0;116,94)a,b
21,83
(3,2;58,57)a,b
120 0,4
(0;26,3)a,b
2,5
(0;22,2)a,b
2 (0;5) 3 (0;7) 1,96
(0;108,16)a,b
9,15
(0;68,33)a,b
120 (10
mmol KCl/l)
2,3
(0;34,8)b
3,1
(0;32,3)b
3 (0;4) 3 (0;6) 6,04
(0;130,30)b
11,53
(0;89,27)b
n = 10; signed rank – Test: zwischen den Kurven kon nten keine signifikanten Unterschiede
festgestellt werden.
4.2 In-vivo- Versuch
Ziel des In-vivo-Versuchs war es herauszufinden, ob sich die in-vitro gewonnen
Ergebnisse auch am lebenden Tier widerspiegeln; ob also Insulin in
pathophysiologischen Konzentrationen im Blut den Austritt von Ingesta aus dem
Labmagen vermindern kann und ob diese Reduzierung des Effluxes durch

100 Ergebnisse
gleichzeitige Kaliumgaben kompensiert werden kann. Der Versuch fand unter
normoglykämischen Bedingungen statt (10.2.1; Abbildung 33).
4.2.1 Insulinkonzentrationen
Wie die Abbildung 29 zeigt, konnte durch die Infusion von 1 mU Insulin/kg KGW/min
gegenüber den Basalwerten an Tag 1 im Mittel eine Steigerung der gemessenen
Insulinkonzentrationen im Blut von 0,68 (±0,59) µg/l auf 6,67 (±1,9) µg/l an Tag 2
bzw. auf 7,24 (±1,96) µg/l an Tag 3 erzielt werden. Der durch die Infusion bedingte
Anstieg der Insulinspiegel war bereits nach einer Stunde auf einem konstanten
Niveau. In Tabelle 29 sind die Einzeldaten der Versuchstiere an den drei
Versuchstagen aufgelistet. Wichtig ist, dass die vor Beginn der Infusionen an Tag 3
gemessenen Insulinwerte wieder alle im Normalbereich lagen.
0
2
4
6
8
10
keine Insulininfusion Insulininfusion von 1 mU/kgKGW/min
Insulininfusion von 1 mU/kgKGW/min mit
Kaliumsubstitution
Insu
linko
nz. i
n µg
/l
Abbildung 29: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Mittlere Insulinwerte an den drei Versuchstagen
n = 6; unterschiedliche Buchstaben (a,b) belegen st atistisch signifikante Unterschiede in der
einfaktoriellen Varianzanalyse mit einem Signifikan zniveau von p<0,05.
Bei den mittleren Insulinwerten handelt es sich an Tag 1 um den Mittelwert aller
Messzeitpunkte und an den Tagen 2 und 3 um die Mitt elwerte der Zeitpunkte von der 60. bis zur
240. Minute jeweils aller sechs Versuchstiere.
Die Standardabweichungen reflektieren die Streuung der jeweiligen Mittelwerte der 6
Versuchstiere.
a
b b

Ergebnisse 101
Tabelle 29: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Insulinwerte (in µµµµg/l) an den drei Versuchstagen zu unterschiedlichen
Zeitpunkten nach Beginn der Infusion
Tier 1 Tier 2 Tier 3 Tier 4 Tier 5 Tier 6
Tag 1, t = 0 min 0,21 1,03 0,51 1,61 0,02 0,80
Tag 1, t = 60 min 0,06 0,53 0,92 1,99 0,02 0,71
Tag 1, t = 120 min 0,02 0,25 0,61 1,75 0,01 0,85
Tag 1, t = 180 min 0,26 0,25 0,97 1,67 0,05 0,52
Tag 1, t = 240 min 0,29 0,46 0,78 1,64 0,05 0,52
Tag 2, t = 0 min 0,31 0,64 0,77 1,81 0,03 0,90
Tag 2, t = 60 min 6,16 7,01 4,89 8,98 10,13 4,81
Tag 2, t = 120 min 6,31 7,19 4,50 7,70 9,14 5,10
Tag 2, t = 180 min 4,42 8,56 4,28 7,74 6,54 5,05
Tag 2, t = 240 min 5,04 7,95 3,40 6,82 5,96 4,63
Tag 3, t = 0 min 0,69 0,83 1,04 2,11 0,04 0,63
Tag 3, t = 60 min 7,20 10,47 4,65 6,52 6,09 4,67
Tag 3, t = 120 min 9,42 8,20 4,31 9,49 7,61 5,23
Tag 3, t = 180 min 6,93 9,43 4,62 5,37 6,64 8,89
Tag 3, t = 240 min 5,83 9,64 4,66 8,59 8,00 9,45
n = 6; Beginn der Insulininfusion an Tag 2 und 3 je weils bei t = 2 min
4.2.2 Kaliumkonzentrationen
Durch die Insulininfusion konnten die Blutkaliumwerte im Mittel von 3,94 (±0,19)
mmol/l auf 3,51 (±0,25) mmol/l signifikant gesenkt werden. Erste Reaktionen traten
bereits innerhalb der ersten halben Stunde nach Beginn der Insulininfusionen auf.
Am dritten Versuchstag konnte die Substitution von 0,5 mmol KCl/min/Tier ein
signifikantes Absinken der Blutkaliumwerte verhindern: Im Mittel lagen diese an Tag
3 bei 3,83 (±0,19) mmol/l (Abbildungen 30 und 31). Die vor Beginn der Infusionen an

102 Ergebnisse
Tag 3 gemessenen Blutkaliumwerte lagen mit Ausnahme des zweiten Tieres alle im
normokaliämischen Bereich von 3,5 – 4,5 mmol/l (Tabelle 30).
3
3,5
4
4,5
keine Insulininfusion Insulininfusion von 1 mU/kgKGW/min
Insulininfusion von 1 mU/kgKGW/min mit
Kaliumsubstitution
[K] i
n m
mol
/l
Abbildung 30: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Mittlere Blutkaliumwerte an den drei Versuchstagen
n = 6; unterschiedliche Buchstaben (a,b) belegen st atistisch signifikante Unterschiede in der
zweifaktoriellen Varianzanalyse mit einem Signifika nzniveau von p<0,05.
Bei den mittleren Blutkaliumwerten handelt es sich an Tag 1 um den Mittelwert aller
Messzeitpunkte und an den Tagen 2 und 3 um die Mitt elwerte der Zeitpunkte von der 60. bis zur
240. Minute jeweils aller sechs Versuchstiere.
Die Standardabweichungen reflektieren die Streuung der jeweiligen Mittelwerte der 6
Versuchstiere.
a
b
a

Ergebnisse 103
3
3,2
3,4
3,6
3,8
4
4,2
4,4
4,6
0 60 120 180 240 0 60 120 180 240 300 30 90 150 210 270
Zeit in min
[K] i
n m
mol
/l
Abbildung 31: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Verlauf der Blutkaliumkonzentrationen aller Versuc hstiere an den drei
Versuchstagen
Die Pfeile markieren den Beginn der Insulininfusion .
Tabelle 30: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Blutkaliumwerte (in mmol/l) an den drei Versuchsta gen zu
unterschiedlichen Zeitpunkten nach Beginn der Infus ion
Tier 1 Tier 2 Tier 3 Tier 4 Tier 5 Tier 6
Tag 1, 0 min 3,83 3,81 3,56 3,77 3,84 3,89
Tag 1, 30 min 3,75 3,81 3,77 3,97 3,92 4,23
Tag 1, 60 min 3,68 3,81 3,71 3,86 3,82 3,99
Tag 1, 90 min 3,72 3,87 3,87 4,06 3,93 4,12
Tag 1, 120 min 3,94 4,04 3,91 4,16 3,95 3,90
Tag 1, 150 min 3,79 4,20 3,57 3,98 3,73 3,97
Tag 1, 180 min 3,81 4,05 3,94 4,35 3,90 4,08
Tag 1, 210 min 3,78 4,36 3,88 4,36 3,89 4,28
Tag 1, 240 min 3,99 4,26 3,76 4,32 3,92 4,11
Tag 1, 270 min 3,86 4,23 3,76 4,20 4,20 3,92
Tag 1 Tag 2 Tag 3
Kontrolle Insulin Insulin + Kalium

104 Ergebnisse
Tag 2, 0 min 3,77 4,20 3,86 4,10 3,99 3,91
Tag 2, 30 min 3,36 3,27 3,42 3,81 3,88 3,81
Tag 2, 60 min 3,35 3,37 3,34 3,84 3,58 3,75
Tag 2, 90 min 3,07 3,16 3,19 3,64 3,52 3,58
Tag 2, 120 min 3,11 3,30 3,76 3,76 3,34 3,62
Tag 2, 150 min 3,25 3,27 3,55 3,63 3,44 3,61
Tag 2, 180 min 3,19 3,31 3,56 3,82 3,37 3,61
Tag 2, 210 min 3,17 3,24 3,70 3,95 3,36 3,53
Tag 2, 240 min 3,27 3,20 3,43 3,71 3,49 3,89
Tag 2, 270 min 3,38 3,29 3,68 3,88 3,47 3,48
Tag 2, 300 min 3,32 3,22 3,6 3,99 3,81 3,74
Tag 3, 0 min 3,52 3,44 3,86 3,86 4,14 4,28
Tag 3, 30 min 3,72 3,42 2,88 3,93 4,31 4,10
Tag 3, 60 min 3,41 3,55 3,71 3,79 4,04 4,10
Tag 3, 90 min 3,56 3,79 3,82 3,88 4,11 3,85
Tag 3, 120 min 3,47 3,86 3,32 3,95 4,03 3,66
Tag 3, 150 min 3,63 3,82 3,34 3,98 4,11 3,89
Tag 3, 180 min 3,67 3,92 3,52 3,85 3,99 3,74
Tag 3, 210 min 3,64 4,36 3,51 3,88 4,01 3,78
Tag 3, 240 min 3,59 4,16 3,92 3,83 4,24 3,76
Tag 3, 270 min 3,48 4,26 3,57 3,88 4,40 3,69
Tag 3, 300 min 3,62 3,99 3,99 3,99 4,53 3,92
n = 6; Beginn der Insulininfusion an Tag 2 und 3 je weils bei t = 2 min
4.2.3 Labmagenefflux
Der mittlere Efflux aus dem Labmagen ins Duodenum unterschied sich an den drei
Versuchstagen nicht signifikant voneinander (Abbildungen 32 und 33). Bei den
einzelnen Versuchstieren traten unterschiedliche Reaktionen auf die Insulininfusion
auf: Bei vier von sechs Tieren kam es an Tag 2 zu einer Reduktion des Effluxes,
davon war nur bei zwei Tieren ein erneuter Anstieg am dritten Versuchstag

Ergebnisse 105
nachweisbar. Bei den zwei Tieren, bei denen es an Tag 2 zu einer Steigerung der
Effluxrate kam, war bei einem Tier an Tag 3 eine weitere Erhöhung, bei dem anderen
Tier eine Reduktion der Effluxmenge feststellbar (Tabelle 31).
0
50
100
150
200
250
300
keine Insulininfusion Insulininfusion von 1 mU/kgKGW/min
Insulininfusion von 1 mU/kgKGW/min mit
Kaliumsubstitution
Effl
uxm
enge
in m
l/min
Abbildung 32: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Mittlere Effluxmengen aus dem Labmagen ins Duodenu m an den drei
Versuchstagen
n = 6; die Standardabweichungen reflektieren die St reuung der jeweiligen Mittelwerte der 6
Versuchstiere.
Tabelle 31: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Effluxmengen in ml/min aus dem Labmagen ins Duoden um aller
Versuchstiere an den drei Versuchstagen
Tier 1 Tier 2 Tier 3 Tier 4 Tier 5 Tier 6
Tag 1 150 (±18) 149 (±24) 180 (±8) 165 (±10) 333 (±78) 146 (±16)
Tag 2 142 (±26) 133 (±5) 258 (±32) 194 (±11) 194 (±11) 113 (±9)
Tag 3 140 (±2) 180 (±42) 176 (±6) 192 (±6) 192 (±6) 168 (±15)
Die Standardabweichungen reflektieren die Streuung der Effluxmengen im steady state .

106 Ergebnisse
Tabelle 32 gibt eine Übersicht über die Einzeldaten aller Tiere an den drei
Versuchstagen.
Tabelle 32: In-vivo-Versuch zur Messung des Labmageneffluxes un ter basalen (Tag 1),
hyperinsulinämisch-hypokaliämischen (Tag 2) und hyp erinsulinämisch-normokaliämischen
(Tag 3) Bedingungen: Übersicht der Mittelwerte der Insulin- und Kaliumw erte im Blut und des
vorherrschenden Labmageneffluxes der einzelnen Tier e an den drei Versuchstagen
Tag 1 Tag 2 Tag 3
Insulin
in µg/l
Kalium
in
mmol/l
Efflux
in
ml/min
Insulin
in µg/l
Kalium
in
mmol/l
Efflux
in
ml/min
Insulin
in µg/l
Kalium
in
mmol/l
Efflux
in
ml/min
Tier
1 0,17 3,82 150 5,49 3,25 142 7,34 3,58 140
Tier
2 0,50 4,05 149 7,68 3,26 133 9,43 3,91 180
Tier
3 0,76 3,73 180 4,27 3,54 258 4,56 3,56 176
Tier
4 1,73 4,10 165 7,81 3,80 194 7,49 3,90 192
Tier
5 0,04 3,91 333 7,94 3,53 266 7,16 4,18 183
Tier
6 0,68 4,05 146 4,90 3,66 113 7,06 3,85 168

Diskussion 107
5 Diskussion
Die Ergebnisse des In-vitro- und des In-vivo-Versuchs sollten Aufschluss darüber
geben, inwieweit die insbesondere bei Kühen im peripartalen Zeitraum zu
beobachtenden Veränderungen in der Kalium- und Insulinhomöostase Einfluss auf
die Motilität der Labmagenmuskulatur und damit auf den abomasalen Efflux nehmen.
5.1 In-vitro -Versuch
In diesem Teil der Studie wurden Muskelpräparate aus dem Labmagen von Ziegen
und Rindern isoliert, in einer Pufferlösung inkubiert und in eine Vorrichtung zur
Messung der Motilität eingespannt. Es wurde circuläre Corpus- und
Pylorusmuskulatur (sowie bei der Ziege z. T. noch longitudinale Corpusmuskulatur)
hinsichtlich ihrer Reaktion auf Kalium- und Insulingaben untersucht. Dabei konnten
schon unter basalen Bedingungen (d. h. ohne jegliche Zugaben) sowohl
Unterschiede in der Aktivität zwischen Corpus- und Pylorusmuskulatur als auch bei
den Präparaten aus einer Region des Labmagens gefunden werden. Während die
Pylorusmuskulatur grundsätzlich eine geringe Spontanaktivität zeigte, wies die der
Corpusisolate erhebliche Unterschiede nicht nur zwischen den Einzeltieren, sondern
sogar bei aus direkter Nachbarschaft entnommenen Muskelstücken auf. Diese sehr
große Variation in der Motilität spiegelt sich in den z. T. erheblichen Unterschieden
der Minima und Maxima.
Die Differenzen zwischen Corpus- und Pylorusmuskulatur lassen sich durch ihre
jeweilige Funktion in vivo erklären. Während die Magenpumpe wellenförmige
Kontraktionen aufweist, die die Ingesta nach aboral befördern, erschlafft und
kontrahiert die Muskulatur am Pylorus, um den Chymus abfließen zu lassen (HILL
1969; MALBERT u. RUCKEBUSCH 1988). Letztere weist also im Gegensatz zur
Corpusmuskulatur nur in sehr viel geringerem Ausmaß rhythmische, spontane
Kontraktionen auf.
Der Grund für die großen Unterschiede der Spontanaktivität zwischen den
Präparaten eines Abschnitts ist unklar. Die Behandlung der Muskeln vor und
während der Präparation erfolgte stets im gleichen zeitlichen und technischen Ablauf.
Fett- und Bindegewebsauflagerungen wurden abpräpariert, ohne die Muskulatur zu

108 Diskussion
verletzen oder zu strapazieren. Diese Aspekte können zudem nicht die Unterschiede
zwischen Isolaten erklären, die direkt nebeneinander liegen. Auch das Gewicht der
Muskelstückchen korrelierte nicht mit ihrer Aktivität. Denkbar wäre hingegen, dass
der funktionelle Komplex zwischen ICC und Muskelzellen bzw. deren
Quervernetzung durch die Präparation in den einzelnen Muskelstückchen
unterschiedlich stark unterbrochen wird, so dass der „Taktgeber“ der Kontraktionen
in einigen Bereichen eines Muskelstückes entfällt und damit die gesamte
Kontraktionsaktivität geringer ausfällt (DAVIES 1989).
In Folge der unterschiedlichen Spontanmotilität war eine kontinuierlich durchgeführte
Kontrolle unerlässlich.
Da es sich bei den Isolaten der bovinen Labmagenwand um Material von
Schlachthoftieren handelte und die Kühe somit weder gleich alt noch definiert
gehalten und gefüttert wurden, diente Labmagenmuskulatur von gleichaltrigen
Ziegen, die im Rahmen eines anderen Versuches am hiesigen Institut gehalten und
schließlich geschlachtet wurden, der Kontrolle. Hinsichtlich der Variationen der
Motilität konnten keine Unterschiede zwischen den Kühen und Ziegen festgestellt
werden.
5.1.1 Reaktionen isolierter Labmagenmuskulatur auf Änderungen der
extrazellulären Kaliumkonzentration
Untersucht wurden die Antworten isolierter Labmagenmuskulatur auf
unterschiedliche Kaliumkonzentrationen an circulärer Corpus- und Pylorusmuskulatur
von Ziege und Rind. Als Kontrolle der in Form von KCl zugegebenen Mengen
dienten dabei equimolare NaCl-Gaben. Mit Ausnahme der circulären
Pylorusmuskulatur der Ziege kam es dabei über die Zeit zu starken Veränderungen
der Motilitätsparameter. Aus diesem Grund sind nur zeitgleich durchgeführte
Versuche aussagekräftig. An den Präparaten der Labmagenmuskulatur konnte in
unterschiedlicher Ausprägung eine durch steigende Kaliumgaben bedingte Zunahme
von Amplitude und Kontraktionsaktivität bei unbeeinflusster Frequenz festgestellt
werden. Das gilt auch für den für die Pathophysiologie relevanten Bereich zwischen
1 und 5 mmol Kalium/l. An der Pylorusmuskulatur der Ziege war nur ein Anstieg der
Parameter bis zu einer Kaliumkonzentration von 7 mmol/l feststellbar. Bei 10 mmol/l

Diskussion 109
kam es hingegen zu einem Abfall. Ein ähnlicher Verlauf insbesondere der Amplitude
war allerdings auch durch Gaben von NaCl zu beobachten, so dass hier die
Kaliumwirkung nicht eindeutig nachgewiesen werden konnte, da sich lediglich für
eine Konzentration von 7 mmol Kalium/l signifikante Unterschiede zwischen den
beiden Kurven ergaben.
Am Dünndarm von Hunden wiesen HARA und SZURSZEWSKI (1986) den Einfluss
extrazellulären Kaliums auf das Membranpotential glatter Muskelzellen nach: Je
höher die Kaliumkonzentrationen waren, desto weniger negativ wurde das
Membranpotential und umgekehrt. Im Zuge von Hypokaliämien wird somit das
Schwellenpotential zur Öffnung spannungsabhängiger Kalziumkanäle seltener
erreicht (KOBAYASHI et al. 1985), so dass es zu einer Abnahme der
Kontraktionsbereitschaft der Muskulatur bis hin zu Paralysen kommt (PETRIDES
1997; SIELMAN et al. 1997). Die bei hohen extrazellulären Kaliumkonzentrationen zu
beobachtende Steigerung der Muskelaktivität ist zwar auch auf die Beeinflussung
des Membranpotentials zurückzuführen, allerdings konnten KATO et al. (2007) am
Kaninchendarm zusätzlich bei Kaliumkonzentrationen von 9 bis 27 mmol/l eine
Modulation inhibitorischer Neurone feststellen, die durch Freisetzung eines Suramin-
sensitiven Transmitters die Stimulation glatter Muskelzellen verstärken. Ob die
Reaktionen der Labmagenpräparate dieser Untersuchung auf eine myogene oder
neurogene Beeinflussung zurückzuführen sind, ist nicht mit Sicherheit zu bestimmen.
Die veränderten Amplituden lassen jedoch die Vermutung zu, dass die ICC von den
äußeren Kaliumbedingungen beeinflusst werden, da die Potentialschwankungen
dieser Schrittmacherzellen mit Auslösen von Aktionspotentialen Grundvoraussetzung
für die Entstehung von Kontraktionen sind und die Frequenz dieser Aktionspotentiale
positiv mit der Kontraktionsstärke (und damit der Amplitudenhöhe) der Muskulatur
korreliert sind (DAVIES 1989). In vielen Studien konnte der Nachweis verschiedener
Kaliumkanäle in ICC erbracht werden (FLYNN et al. 1999; HATTON et al. 2001), was
zugleich eine Abhängigkeit dieser Zellen von der extrazellulären Kaliumkonzentration
bedeutet. Darüber hinaus stellten KITO und SUZUKI (2007) am Dünndarm von
Mäusen bei einer Erhöhung der extrazellulären Kaliumkonzentration von 5 auf 8,25
mmol/l eine Depolarisation der Zellmembran der ICC und eine Verringerung der

110 Diskussion
Amplitude des Plateaus der slow waves fest. Bei 15,3 mmol KCl/l war zusätzlich die
Frequenz letzterer reduziert. Die ICC waren in diesem Fall also abhängig vom
vorherrschenden Kaliummilieu.
Die Reaktionen der einzelnen Labmagenabschnitte auf veränderte extrazelluläre
Kaliumbedingungen waren in der vorliegenden Arbeit unterschiedlich ausgeprägt:
Die Kontraktionsaktivität der circulären Corpusmuskulatur der Ziege stieg von 7,28
(0;54,25) bei 1 mmol/l auf 90,07 (33,87;228,08) mN/min bei 10 mmol/l, während sich
die des Rindes nur von 9,36 (0;78,38) auf 29,87 (0;157,73) mN/min erhöhte. Eine
mögliche Erklärung hierfür liefert eine Studie an boviner Trachealmuskulatur:
GOSENS et al. (2003) konnten zeigen, dass die Stärke der durch Kalium induzierten
Muskelkontraktionen phenotypabhängig war. Auf Kalium- und Methacholingaben
hyperkontraktil reagierende Myozyten wiesen eine veränderte Kalziumsensitivität der
Muskelzellen auf. Ein ähnlicher Mechanismus könnte auch in dieser Arbeit die
Unterschiede in der Kontraktionsbereitschaft erklären.
Warum die circuläre Pylorusmuskulatur der Ziege auf NaCl-Gaben reagierte, bleibt
unklar. Eine Beeinflussung des Membranpotentials ist aufgrund der im Verhältnis
geringen Änderung der NaCl-Konzentration in der Pufferlösung von 117 auf 127
mmol/l eher unwahrscheinlich.
In vitro können also niedrige Kaliumspiegel die Labmagenmuskulatur hemmen und
sind damit grundsätzlich in der Lage, Atonien des Labmagens auszulösen, die in der
Folge zu Labmagenverlagerungen führen könnten.
5.1.2 Reaktionen isolierter Labmagenmuskulatur auf Änderungen der
extrazellulären Insulinkonzentration
Untersucht wurden die Antworten isolierter Labmagenmuskulatur auf
unterschiedliche Insulinkonzentrationen an longitudinaler Corpusmuskulatur von
Ziege und circulärer Corpus- und Pylorusmuskulatur von Ziege und Rind. Während
die circuläre Corpusmuskulatur unter Insulinzugaben im Gegensatz zur Kontrolle bei
unveränderter Frequenz mit einer Abnahme (Rind) bzw. einer geringeren Zunahme
(Ziege) von Amplitude und Kontraktionsaktivität reagierte, kam es bei der
Pylorusmuskulatur der Ziege zu einer Zunahme aller drei Parameter. Die übrigen
Gewebe reagierten nicht auf Insulingaben.

Diskussion 111
5.1.2.1 Keine Reaktion der Muskulatur auf Insulingaben
Nicht alle Abschnitte des Gastrointestinaltraktes reagieren auf Insulingaben mit
veränderter Motilität. BUENO und RUCKEBUSCH (1976) untersuchten beim Schaf
an verschiedenen Dünndarmabschnitten Veränderungen in der Motilität durch
Insulin. Während sie am Jejunum Reaktionen auf Insulingaben nachwiesen, konnten
sie sowohl am Duodenum als auch am Ileum keine Aktivitätsänderungen der
Muskulatur feststellen. Der Grund hierfür bleibt allerdings unklar.
5.1.2.2 Stimulation der Muskulatur durch Insulingaben
Schon PAVEL und MILCOU (1932) wiesen bei unterernährten Menschen eine durch
über längere Zeit verabreichtes Insulin hervorgerufene Hypermotilität des Darmes
nach, die sie auf einen direkt die Muskulatur stimulierenden Effekt zurückführten. In
vitro konnte am Jejunum von Schafen ebenfalls eine durch Insulin bedingte
Stimulation der Muskulatur gezeigt werden (BUENO u. RUCKEBUSCH 1976). Und
auch hyperinsulinämische Ratten weisen eine stärkere Motilität des Colons auf als
gesunde Kontrolltiere (FORREST u. PARSON 2003). Als Ursache hierfür konnten
die Autoren eine Ausschaltung inhibitorischer Neurone ausschließen. Sie vermuten
hingegen eine gesteigerte myogene Erregbarkeit als Auslöser. Ferner steht der
glatten Muskulatur bei hohen Insulinkonzentrationen und gleichzeitig unveränderter
Sensitivität der Gewebe vermehrt Glukose zur Verfügung (ARNQVIST 1975), die der
Grund für eine erhöhte Muskelaktivität sein könnte.
5.1.2.3 Hemmung der Muskulatur durch Insulingaben
Verschiedene Studien konnten in vivo einen hemmenden Einfluss von Insulin auf den
Labmagenefflux (HOLTENIUS et al. 2000; SUSTRONCK 2000; VAN MEIRHAEGE et
al. 1988 a) bzw. auf die abomasalen Muskelkontraktionen nachweisen
(PRAVETTONI et al. 2004, 2007). Die Ergebnisse der vorliegenden Arbeit belegen,
dass Insulin in der Lage ist, die circuläre Corpusmuskulatur zu hemmen. Da die
Motilität der Muskulatur aber entscheidend ist für die Aktivität der Magenpumpe,
kann dies den abomasalen Efflux vermindern.
Eine mögliche Ursache für die Hemmung der glatten Muskulatur ist in der
Beeinflussung des intrazellulären Kaliumhaushaltes zu sehen.

112 Diskussion
Sowohl an isolierter Frosch- (MOORE u. RABOVSKY 1979) als auch an
Säugetiermuskulatur (ZIERLER 1959) wurde unter Einfluss von Insulin eine
Hyperpolarisation der Zellmembran festgestellt. IANNACCONE et al. (1989) zeigten
an Skelettmuskulatur von Ratten, dass das negativere Membranpotential auf eine
Aktivierung der Na+/K+-ATPase zurückzuführen ist. Den gleichen Beweis lieferten
auch MOORE (1973) und GAVRYCK et al. (1974) an Froschmuskulatur und im
Rattenhirn. Ebenfalls an der Skelettmuskulatur des Frosches konnte gezeigt werden,
dass die Muskelzellen zweierlei Arten von Pumpen besitzen: aktive und inaktive.
Indem Insulin letztere stimuliert, kommt es zwar zu einem Kaliumeinstrom, aber zu
einem Netto-Kationenausstrom mit folgender Hyperpolarisation (ERLIJ u.
GRINSTEIN 1975). Zusätzlich stimuliert Insulin v. a. in der Leber den Na+/K+/Cl--
Symporter und den Na+/H+-Antiporter, so dass durch die Aufnahme von Na+ der
Na+/K+-ATPase Substrat geliefert wird (LANG 2000).
Die Stimulation der Na+/K+-ATPase führt zu einem Nettokationenausstrom, da nur
zwei Kalium- im Austausch gegen drei Natriumionen in die Zelle aufgenommen
werden. Zusätzlich kann Kalium aufgrund der erhöhten intrazellulären
Konzentrationen durch die Kaliumkanäle aus der Zelle diffundieren. So wird im
Endeffekt das Membranpotential der Muskelzelle stärker negativ, also
hyperpolarisiert. Das Schwellenpotential zur Öffnung spannungsabhängiger
Kalziumkanäle wird seltener erreicht und die myogene Aktivität somit gehemmt.
LANTZ et al. (1980) hingegen demonstrierten an Aggregaten von embryonalen
Herzzellen eine durch Insulin hervorgerufene Hyperpolarisation, die nicht durch die
Gabe des die Na+/K+-ATPase blockierenden Ouabains aufgehoben werden konnte,
also nicht auf einer Stimulation selbiger beruhte. ECKEL u. REINAUER (1990)
führten das durch Insulin bedingte negativere Membranpotential von Ratten-
Herzmuskelzellen auf eine erhöhte Kaliumleitfähigkeit der Zellmembran zurück. Eine
Untersuchung an Pericyten retinaler Kapillaren zeigte eine Aktivierung Apamin-
sensitiver Ca2+-abhängiger Kaliumkanäle durch Insulin (BERWECK et al. 1993) und
an glatten Muskelzellen der thorakalen Aorta von Rattenembryonen konnte eine
erhöhte Offenwahrscheinlichkeit ATP-sensitiver Kaliumkanäle in Anwesenheit von
Insulin nachgewiesen werden (YASUI et al. 2007). Da die Anwesenheit von Insulin

Diskussion 113
zudem grundsätzlich zu einer erhöhten Aufnahme von Kalium in Muskel-, Fett- und
Lebergewebe führt (VAN MEIRHAEGHE 1988; SWEENEY 1999), kommt es so
infolge lokaler oder auch genereller Hypokaliämien und einer erhöhten
Kaliumleitfähigkeit zu einem vermehrten Ausstrom von Kalium aus der Zelle und
ebenfalls zu einer Hyperpolarisation der Membran.
5.1.2.4 Mechanismen der Insulinhemmung
Um zu evaluieren, ob für den hemmenden Einfluss des Insulins auf die circuläre
Corpusmuskulatur des Labmagens ähnliche Mechanismen zugrunde liegen, wurden
weitere Untersuchungen durchgeführt. Einerseits wurde die Einflussnahme von
Kalium auf die Insulinwirkung untersucht und andererseits Kaliumkanäle mittels
Barium und Glybenclamid bzw. die Na+/K+-ATPase durch Ouabain blockiert.
5.1.2.4.1 Wirkung von Kalium in Anwesenheit von Insulin
Weder bei der Ziege noch beim Rind konnten signifikante Unterschiede zwischen
einer Kalium-Dosiswirkungskurve in An- oder Abwesenheit von Insulin an der
circulären Corpusmuskulatur festgestellt werden. Gleichzeitig reagierten alle
Präparate nur schwach auf die steigende extrazelluläre Kaliumkonzentration.
Letzteres ist auf die Konzeption des Versuchsprotokolls zurückzuführen (Anhang
10.1.1), da die Aktivität und Ansprechbarkeit der Präparate auf äußere Reize über
die Zeit abnahm (Ergebnisse 4.1.2.2). Die Gründe für das Fehlen von Unterschieden
zwischen den Kurven könnten auf einem Zusammenspiel von Na+/K+-ATPase und
Kaliumleitfähigkeit beruhen.
Wie oben beschrieben ist Insulin in der Lage, Muskelzellen durch Stimulierung von
Na+/K+-ATPasen zu hyperpolarisieren. Sowohl die durch Insulin stimulierte Aktivität
(MARUNAKA u. KITASATO 1985; MARUNAKA et al. 1986 b) als auch die basale
Aktivität der Na+/K+-Pumpe (MARUNAKA u. KITASATO 1985; MARUNAKA et al.
1986 a; MARUNAKA 1988) erhöht sich mit steigenden extrazellulären
Kaliumkonzentrationen. Dies würde für sich betrachtet bedeuten, dass Kaliumgaben
die Hyperpolarisation der Zellmembran in Anwesenheit von Insulin nicht verändern
oder verstärken. Allerdings ist das Ausmaß dieser Insulin-bedingten
Hyperpolarisation bei hohen Kaliumspiegeln vermindert (MARUNAKA 1987). Dies ist

114 Diskussion
in erster Linie darauf zurückzuführen, dass Kalium über eine Vielzahl von Kanälen in
Abhängigkeit des elektro-chemischen Gradienten über die Zellmembran diffundieren
kann. Der Beitrag der Kaliumleitfähigkeit zur Aktivität der Muskulatur überdeckt also
den hemmenden Einfluss der möglicherweise vorhandenen Insulin-bedingten
Stimulierung der Na+/K+-ATPase.
Eine durch Insulin hervorgerufene erhöhte Aktivität oder Offenwahrscheinlichkeit von
Kaliumkanälen müsste die Muskelzelle sensitiver werden lassen, insbesondere
gegenüber niedrigen extrazellulären Kaliumkonzentrationen. Bei einer Vielzahl von
geöffneten Kanälen ist die Wahrscheinlichkeit größer, dass lokale
Diffusionsgradienten für Kalium entstehen, es so vermehrt aus der Zelle austreten
kann und das Membranpotential (noch) negativer wird.
Dies spiegelt sich in den vorliegenden Ergebnissen nicht wider. Verschiedene
Gründe kommen dafür in Betracht. Zum Einen besteht die Möglichkeit, dass Insulin
weder die Na+/K+-ATPase noch die Kaliumkanäle beeinflusst, so dass keine
Unterschiede zwischen den Kurven festzustellen sind. Zum Anderen könnte es sein,
dass Insulin auf beide Komponenten einwirkt: Die Na+/K+-Pumpe erhöht und die
Aktivierung der Kanäle erniedrigt die intrazelluläre Kaliumkonzentration. Möglich ist
ferner, dass die Auswirkungen des Insulins auf die Muskulatur überlagert werden, da
auch die nervale Komponente und die ICC durch Kalium beeinflusst werden (s. 5.1).
5.1.2.4.2 Wirkung von Insulin in Anwesenheit von Barium
Durch die Gabe von Barium können die meisten Kaliumkanäle nicht-selektiv geblockt
werden (BERWECK et al. 1993). Im Zuge dessen kam es an der glatten Muskulatur
des Labmagens zu einer signifikanten Steigerung von Amplitude und
Kontraktionsaktivität, die nach ca. 30 Minuten ein neues Plateau erreichte. Diese
Aktivitätszunahme beruht auf einem Anstieg des Membranpotentials, das auf den
unterbundenen Kaliumausstrom zurückzuführen ist. Der Effekt auf die Muskulatur
war allerdings unterschiedlich stark ausgeprägt. Die Höhe des Plateaus der
einzelnen Präparate variierte dabei in etwa zwischen 2 und 1000 mN/min. Worauf
diese enormen Unterschiede in der Reaktionsstärke beruhen ist unklar. Denkbar

Diskussion 115
wären Unterschiede in der Ansprechbarkeit der Gewebe (s. 5.1) oder in der Anzahl
an Kaliumkanälen.
Nach Vorinkubation mit 80 mU Insulin/l, wodurch sich Amplitude und
Kontraktionsaktivität verringerten, kam es zu einer nominellen Verstärkung des
Bariumeffektes. Im Gegenzug hatte Insulin in Anwesenheit von Barium keinen
Einfluss mehr auf die Muskulatur. Diese Ergebnisse deuten auf eine Insulinwirkung
an den Kaliumkanälen hin. Auch BERWECK et al. (1993) konnten in ihrer Arbeit
zeigen, dass die durch Insulin induzierte Hyperpolarisation der Zellmembran boviner
retinaler Pericyten durch Bariumzugaben verhindert werden konnte und dass
umgekehrt Insulin in Anwesenheit des Kaliumkanalblockers keine Wirkung mehr
zeigte.
5.1.2.4.3 Wirkung von Insulin in Anwesenheit von Glybenclamid
Um die wahrscheinlich vorhandene Insulinwirkung auf die Kaliumkanäle zu
spezifizieren, wurden mittels 0,05 mmol Glybenclamid/l ATP-sensitive Kanäle
geblockt (BERWECK et al. 1993; TRICARICO et al. 1998; YASUI et al. 2007). Der
die myogene Kontraktion stimulierende Effekt war deutlich geringer ausgeprägt als
bei Barium, was auf die unvollständige Blockade der Kaliumkanalpopulation
zurückzuführen ist. Auch die Höhe der Plateaus variierte, vermutlich aufgrund
dessen, nicht so stark. Wie auch schon in Kombination mit Barium gezeigt, konnte
die Insulin-bedingte Hemmung von Amplitude und Kontraktionsaktivität durch
Glybenclamid aufgehoben werden. Insulin hatte zusätzlich in Anwesenheit von
Glybenclamid keinen Einfluss mehr auf die Muskulatur, was wiederum auf eine
mögliche Insulinwirkung an Glybenclamid-sensitiven Kaliumkanälen spricht.
Während BERWECK et al. (1993) die Insulin-induzierte Hyperpolarisation der Zellen
nicht durch Glybenclamid aufheben konnten, gelang dieses YASUI et al. (2007)
hingegen an Gefäßmuskelzellen von Rattenembryonen. Sie wiesen so erstmalig eine
Insulin-bedingte erhöhte Offenwahrscheinlichkeit ATP-abhängiger Kaliumkanäle
nach.

116 Diskussion
5.1.2.4.4 Wirkung von Insulin in Anwesenheit von Ouabain
In einem weiteren Ansatz wurde die Na+/K+-ATPase mittels 0,001 mmol Ouabain/l
gehemmt. Dabei kam es zunächst zu einem nominellen Anstieg, im weiteren Verlauf
jedoch zu einer Abnahme von Amplitude und Kontraktionsaktivität. VANHEEL und
BREYNE (2004) konnten am Endothel kleiner Magenarterien zeigen, dass es unter
Einwirkung von Ouabain initial durch Hemmung des Pumpenstroms zu einer
Depolarisation kam. Im Folgenden konnte jedoch eine Hyperpolarisation der
Muskelzellmembranen beobachtet werden. Diese könnte darauf zurückzuführen
sein, dass die bei der Blockade der Na+/K+-ATPase ansteigende intrazelluläre
Natriumkonzentration die Aktivität des Na+/Ca2+-Austauschers beeinflusst, so dass
sich Kalzium in der Zelle anreichert. Die Konsequenz daraus ist zum einen die
Kontraktion der Muskelzelle und zum anderen könnte es sein, dass Kalzium-
abhängige Kaliumkanäle öffnen (LEBRUN et al. 1993). Eine daraus entstehende
erhöhte Kaliumleitfähigkeit könnte die in der Arbeit von VANHEEL und BREYNE
(2004) gemessene Hyperpolarisation und die in der vorliegenden Arbeit gezeigte
sekundäre Abnahme der Motilitätsparameter erklären.
Durch die Gabe von 80 mU Insulin/l konnte in diesem Versuchsansatz keine
signifikante Abnahme von Amplitude und Kontraktionsaktivität festgestellt werden.
Die Gründe hierfür sind unklar, da die Insulinlösung aus derselben Charge stammte,
die auch in den vorherigen Versuchen eingesetzt wurde. Nach Vorinkubation mit
Insulin konnte zwar eine weniger ausgeprägte Ouabain-bedingte Stimulation der
Muskulatur nachgewiesen werden, die Unterschiede zu der Ouabainkontrolle waren
jedoch nicht signifikant. Nach Blockade der Na+/K+-ATPase hatte Insulin keine
Wirkung mehr auf die Labmagenpräparate. Die Insulinwirkung auf die circuläre
Corpusmuskulatur konnte somit durch Ouabain aufgehoben werden.
5.1.2.4.5 Zusammenfassung der möglichen Mechanismen
Die oben angeführten Ergebnisse weisen darauf hin, dass Insulin an der zirkulären
Corpusmuskulatur des Labmagens sowohl über eine erhöhte Kaliumleitfähigkeit als
auch über eine Stimulation der Aktivität der Na+/K+-ATPase wirken könnte. Da in der
vorliegenden Arbeit allerdings keine Membranpotentiale gemessen wurden, sondern

Diskussion 117
lediglich auf die Bewertung der Muskelreaktionen zurückgegriffen werden konnte, die
zudem erschwert wurde durch die starken Variationen der basalen und durch
Substratzugaben stimulierten Aktivität, könnte es sich bei den beobachteten
Änderungen auch um eine Überlagerung von Effekten und nicht um direkt
antagonistische Wirkungen handeln.
5.2 In-vivo -Versuch
In vitro konnte also gezeigt werden, dass Insulin Einfluss auf die Motilität der glatten
Labmagenmuskulatur nimmt und dass dieser Effekt unter Umständen auf veränderte
Kaliumtransportraten über die Zellmembran zurückzuführen ist. Ziel des In-vivo-
Versuchs war es deshalb zu untersuchen, ob sich diese Ergebnisse auch am
lebenden Tier widerspiegeln; ob also Insulin in pathophysiologischen
Konzentrationen im Blut den Austritt von Ingesta aus dem Labmagen vermindern
kann und ob diese Reduzierung des Effluxes durch gleichzeitige Kaliumgaben
kompensiert werden kann.
Bei den Versuchstieren handelte es sich um leistungsgerecht gefütterte, gesunde
Milchkühe der Rasse Holstein-Frisian in unterschiedlichen Reproduktionsstadien. Ein
Versuch dauerte drei Tage. Am ersten Tag wurden bei den Versuchstieren die
basalen Kalium-, Glukose- und Insulinwerte im Blut bestimmt und der normale, d.h.
nicht durch Kalium- oder Insulin beeinflusste Labmagenefflux gemessen. An den
folgenden Tagen wurde versucht, unter euglykämischen Bedingungen über eine
Infusion von Insulin (zweiter Versuchstag) die Kaliumkonzentration im Blut zu senken
bzw. diese durch zusätzliche Gabe von 0,5 mmol KCl/min/Tier konstant zu halten
(dritter Versuchstag) und dabei ggf. auftretende Änderungen des Effluxes zu
messen. Letzterer wurde mit Hilfe von kontinuierlichen intraruminalen Applikationen
von Chrom-EDTA ermittelt. Um das Erreichen des hierfür notwendigen steady states
nicht zu gefährden, wurde den Tieren die Futter- und Wasseraufnahme für die Dauer
des Versuchs (ca. fünf Stunden) verwehrt. Die Tiere haben die Versuchstage ohne
Komplikationen überstanden.

118 Diskussion
5.2.1 Insulinkonzentrationen
Durch die Infusion von 1 mU Insulin/kg KGW/min konnte gegenüber den Basalwerten
an Tag 1 im Mittel eine Steigerung der gemessenen Insulinwerte im Blut von 0,68
(±0,59) µg/l auf 6,67 (±1,9) µg/l an Tag 2 bzw. auf 7,24 (±1,96) µg/l an Tag 3 erzielt
werden. Die vor Beginn der Infusionen an Tag 3 gemessenen Insulinwerte
entsprachen alle den Basalwerten an Tag 1.
Insulininfusionen heben die basale Insulinkonzentration an, bis sich nach einer
gewissen Infusionsdauer ein Gleichgewicht zwischen Insulininfusion, pankreatischer
Insulinsekretion und Insulin-Clearance eingestellt hat und eine steady-state
Insulinkonzentration (SSIK) erreicht ist (BLUM et al. 1999). Dieser
Gleichgewichtszustand war in der vorliegenden Arbeit bereits nach einer Stunde
erreicht. Obwohl die Insulininfusion auf Basis der Körpergewichte für alle Tiere gleich
war, unterschieden sich die SSIKs bei den sechs Versuchstieren deutlich. Als
Ursache sind Unterschiede in der Plasma-Insulin-Clearance in den unterschiedlichen
Reproduktionsstadien zu vermuten (DEBRAS et al. 1989). Die Abnahme des
zirkulierenden Insulins ist z. B. bei laktierenden Schafen höher als bei
trockenstehenden. Dies sichert eine niedrige Insulinkonzentration im Plasma
während der Laktation, die für die Umverteilung der Nährstoffe hin zur Milchdrüse
wichtig ist (FAULKNER u. POLLOCK 1990). Vergleichbares wurde auch bei Ziegen
(GRIZARD et al. 1988), Kühen (HART et al. 1980) und Ratten (JONES et al. 1984)
beobachtet. Als weitere Möglichkeit wird die veränderte Insulinaufnahme durch die
Leber (LOMAX et al. 1979) während der Trächtigkeit und in der Laktation diskutiert
(SANO et al. 1991). So ist in der vorliegenden Studie bei den laktierenden Tieren
während der Insulininfusion zumindest an Tag 2 eine Abnahme der gemessenen
Plasmakonzentration trotz konstanter Infusionsrate zu verzeichnen.
Die Insulininfusionsrate von 1 mU/kg KGW/min wurde gewählt, da aus der Arbeit von
KRÄFT (2004) hervorging, dass mit dieser Menge Insulinkonzentrationen im Tier
erzielt werden, die auch pathophysiologischerweise noch vorkommen können. Sie
erzielte bei Infusionsraten von 0,5 mU Insulin/ kg KGW/ min im Plasma eine
Insulinkonzentration von ca. 27 mU/l, bei 2 mU Insulin/ kg KGW/ min hingegen schon
120-145 mU/l.

Diskussion 119
Die genaue Umrechnung der in µg/l gemessenen Insulinwerte in die sonst
verwendete Einheit mU/l ist nicht möglich, da Insulin mithilfe von Antikörpern
gemessen wurde und somit nicht bekannt ist, wie viel Prozent des Insulins
tatsächlich biologisch aktiv sind. In der Literatur wird diese Einheit jedoch auch z. T.
verwendet, so dass zumindest hinsichtlich der Basalwerte Vergleiche gezogen
werden können. HIPPEN et al. (1999) und BUTLER et al. (2004) geben hierfür
Konzentrationen zwischen 0,2 und 4,5 µg/l an, LEURY et al. (2003) zwischen 0,7 und
1,8 µg/l. Diese Angaben sind mit den Insulinspiegeln der vorliegenden Studie
vergleichbar. Nimmt man dann als Grundlage für die Bewertung anstelle absoluter
Werte die durch die Insulininfusion erzielte Steigerung der Plasmainsulinspiegel (in
dieser Arbeit im Schnitt um das Zehnfache), entspricht diese in etwa den
Konzentrationsunterschieden zwischen normo- und hyperinsulinämischen Kühen.
Erstere weisen je nach Laktationsstadium Werte zwischen 2,1 und 8,9 mU/l auf
(KRÄFT 2004) und letztere bis zu 42 mU/l (PRAVETTONI et al. 2007).
5.2.2 Kaliumkonzentrationen
Die Kaliumkonzentrationen im Plasma konnten durch die Insulininfusionen im Schnitt
signifikant von 3,94 (±0,19) mmol/l auf 3,51 (±0,25) mmol/l gesenkt werden. Die
Reduktion des Kaliumgehaltes erfolgte innerhalb der ersten halben Stunde nach
Beginn der Insulininfusion. Allerdings konnte nur bei 50 % der Tiere eine
Hypokaliämie mit Kaliumwerten von unter 3,5 mmol/l erzeugt werden. Durch die
Gabe von 0,5 mmol KCl/min/Tier konnte ein Absinken der Kaliumspiegel im Blut
verhindert werden. Im Mittel lagen die Werte dann bei 3,83 (±0,19) mmol/l und waren
damit signifikant höher als ohne Kaliumsubstitution und nicht mehr verschieden von
den Werten ohne Insulininfusion an Tag 1.
Entscheidend für die genaue Bestimmung des Kaliumgehaltes im Blut war die
unmittelbare Zentrifugation der Blutproben nach der Gerinnung und das Überführen
des Serums in Eppendorfgefäße, da durch die eintretende Hämolyse bei längerem
Stehen lassen der Proben der Kaliumgehalt des Plasmas steigt (VETMEDLABOR
2007).

120 Diskussion
Die Ergebnisse zeigen, dass Insulin in hyperinsulinämischen Konzentrationen beim
Rind in der Lage ist, den Kaliumgehalt der extrazellulären Flüssigkeit zu senken.
Dies steht mit den Aussagen von VAN MEIRHAEGHE (1988) und SWEENEY (1999)
im Einklang, die eine maßgebliche Beteiligung von Insulin an der Senkung der
Kaliumkonzentration im Blut durch eine verstärkte Aufnahme in Muskel-, Fettgewebs-
und Leberzellen feststellten. Allerdings treten starke Unterschiede zwischen den
Tieren auf: Während sich die Kaliumkonzentration im Plasma beispielsweise bei Tier
1 von 3,82 auf 3,25 mmol/l verringerte, reduzierte sie sich bei Tier 4 lediglich von
4,10 auf 3,80 mmol/l. Eine Korrelation zu der Höhe der im Tier erzielten
Insulinkonzentration bestand dabei nicht. Die während des Clamps infundierte
Glukosemenge zur Erhaltung euglykämischer Bedingungen unterschied sich bei den
Tieren nicht, so dass davon auszugehen ist, dass das verabreichte Insulin, das
zusätzlich aus einer Charge stammte, bei jedem Tier die gleiche biologische Aktivität
zeigte. Möglicherweise bestand bei einigen Versuchstieren eine Insulinresistenz bzw.
eine verminderte Insulin-Response hinsichtlich der Wirkung auf die
Kaliumhomöostase, wie es beim Menschen bereits gezeigt wurde (COHEN et al.
1991). Ebenfalls beim Menschen konnte z. B. auch eine selektive Insulinresistenz
des Proteinstoffwechsels (bei also unbeeinflusster Response des Glukose-, Kalium-
und Fettstoffwechsels) nachgewiesen werden (LUZI et al. 2008). In einer anderen
Studie traten hohe Insulinspiegel gemeinsam mit Hyperglykämien und
Hypokaliämien auf, so dass dort von einer verminderten Response des
Glukosestoffwechsels auszugehen ist (ZHAO et al. 2008). Bei Mäusen scheint
Insulin die SGK1 (serum and glucocorticoid-inducible kinase) in der Leber zu
aktivieren, die wiederum die Aktivität des Na+/H+-Austauschers, des Na+/K+/2Cl--Co-
Transporters und der Na+/K+-ATPase verstärkt und so letztlich den hypokaliämischen
Effekt von Insulin vermittelt (BOINI et al. 2008). Eine unterschiedliche Sensitivität
dieses Enzyms gegenüber Insulin in Geweben der Rinder könnte die von Tier zu Tier
unterschiedliche Abnahme der Kaliumkonzentrationen erklären.
Die Höhe der durch die Infusion erzielten Insulinkonzentration und die dadurch
bedingte Reduktion des Kaliumgehaltes im Blut erfüllen die Voraussetzung für eine
mögliche Übertragbarkeit der In-vitro-Ergebnisse auf das Tier.

Diskussion 121
5.2.3 Efflux
5.2.3.1 Diskussion der Methode
Der abomasale Efflux wurde mit Hilfe des Volumenmarkers Cr-EDTA in Anlehnung
an verschiedene am Pansen durchgeführte Untersuchungen gemessen (LOPEZ et
al. 1994; LOPEZ et al. 2003; VON ENGELHARDT et al. 2006). Cr-EDTA wurde zu
diesem Zweck über eine Fistel dem Pansen zugeführt und zwar in der Menge, die
dem aus dem Labmagen abfließenden Schätzwert entsprach. Die Berechnung des
Labmageneffluxes mithilfe der im Duodenalchymus gemessenen
Chromkonzentration erfolgte im steady state, also dann, wenn sich das Chrom
gleichmäßig auf Vormägen und Labmagen verteilt hatte. Vorversuche (n=3) ergaben,
dass dieser Zustand nach drei bis vier Stunden erreicht war (s. 3.2.3.4., Abbildung
3). Die mit dieser Methode bei den Vorversuchen gemessene basale Effluxmenge
aus dem Labmagen betrug im Mittel 150 ml/min. Dies entspricht auch den Werten
des ersten Versuchstages. Die hier verwandte Methode erweist sich also aufgrund
ihrer Reproduzierbarkeit für die Messung des abomasalen Effluxes als geeignet.
Allerdings ist es auch hier, ähnlich wie bei VAN MEIRHAEGE et al. (1988 a), nicht
möglich, den genauen Ursprung ggf. eintretender Veränderungen auszumachen, da
mit dieser Methode unklar bleibt, ob eine Beeinflussung des Effluxes auf einen
verminderten Vorschub aus dem Pansen oder auf einen direkten Effekt auf den
Labmagen zurückzuführen ist,
5.2.3.2 Diskussion der Ergebnisse
Im Schnitt war zwar bei den gemessenen Effluxmengen kein Unterschied zwischen
den Tagen feststellbar, allerdings ergaben sich zwischen den Versuchstieren
tendenzielle Abweichungen. Bei vier von sechs Versuchstieren kam es am zweiten
Versuchstag zu einer Abnahme, bei den übrigen zwei Tieren hingegen zu einem
Anstieg des Labmageneffluxes.
Dabei schien das Reproduktions- bzw. Trächtigkeitsstadium der Tiere keinen
Einfluss auf den Verlauf des Versuchs zu haben, und auch eine direkte Korrelation
zwischen der Höhe des Labmageneffluxes und der Höhe der Insulin- und
Kaliumkonzentrationen im Blut konnte nicht beobachtet werden. Allerdings fiel auf,

122 Diskussion
dass die Tiere, die am zweiten Versuchstag einen Anstieg des Effluxes zeigten, nur
geringgradige Unterschiede der Kaliumwerte zwischen den drei Versuchstagen
aufwiesen, die Beeinflussung des Kaliumhaushalts durch Insulin also geringer war,
als bei den übrigen Versuchstieren.
Reduktion des Effluxes unter Insulininfusion:
Der bei diesen Tieren nominell hemmende Einfluss des Insulins auf die
Labmagenentleerung entspricht früheren Untersuchungen anderer Autoren
(HOLTENIUS et al. 2000; PRAVETTONI et al. 2004, 2007; SUSTRONCK 2000; VAN
MEIRHAEGE et al. 1988 a), die allerdings z. T. Insulin in pharmakologischen Dosen
infundierten. Die vorliegende Studie zeigt mit Einschränkung, dass auch
pathophysiologische Konzentrationen den Efflux reduzieren könnten.
Der nominell hemmende Einfluss des Insulins war bei der Hälfte der Tiere (den
einzigen trockenstehenden Kühen) durch Kaliumgaben am dritten Versuchstag
kompensierbar, was zumindest bei diesen Kühen für eine mögliche Übertragbarkeit
der in vitro erzielten Ergebnisse auf Zustände in vivo spricht. Bei den Kühen, bei
denen der nominell hemmende Einfluss des Insulins auf den Labmagenefflux nicht
durch Kaliumgaben aufgehoben werden konnte, könnte Insulin nicht (nur) über
Änderungen der Kaliumhomöostase wirken. Denkbar wäre aber auch, dass die
lokalen Kaliumkonzentrationen an der Labmagenmuskulatur nicht den Plasmawerten
entsprechen und dass die substituierte Kaliummenge aus diesem Grund nicht
ausreichend ist, um einen Einfluss des Insulins zu kompensieren.
Die vorliegenden Verhältnisse könnten eine Begründung dafür liefern, weshalb es bei
nicht-laktierenden Kühen, bei denen Kalium die hemmende Insulinwirkung auf den
abomasalen Efflux offenbar kompensiert, nur in Ausnahmefällen zu
Labmagenverlagerungen kommt (DIRKSEN 1961; JUBB et al. 1991; DETILLEUX et
al. 1997).
Stimulation des Effluxes unter Insulininfusion:
Bei zwei der Tiere kam es unter Insulineinfluss zu einem nominell vermehrten
Ausfluss von Ingesta aus dem Labmagen. ARNQVIST (1975) geht davon aus, dass
Insulin der Labmagenmuskulatur über eine erhöhte Glukoseaufnahme in die Zellen
vermehrt Energie bereitstellt und auf diese Weise in der Lage ist, die Aktivität der

Diskussion 123
Myocyten indirekt zu steigern. Bei den beiden Kühen ist auffällig, dass der
Kaliumhaushalt von allen Versuchstieren am wenigsten durch die Insulininfusion
beeinflusst wurde: die Blutkaliumkonzentration sank lediglich um 0,2 bzw. 0,3 mmol/l,
bei den anderen vier Tieren um 0,4 bis 0,8 mmol/l. Damit war eine Insulin-bedingte
Änderung von Kaliumgradienten möglicherweise geringer ausgeprägt. Diese
abweichende Reaktion der Kühe könnte auf (genetischen) Unterschieden in der
Insulinsensitivität beruhen. Die Angaben zur Heritabilität von
Labmagenverlagerungen variieren zwar zwischen h2=0,04 (±0,017) (RICKEN 2003)
und h2=0,24 (GEISHAUSER et al. 1996), es besteht aber grundsätzlich
Übereinstimmung, dass genetische Unterschiede mehr oder weniger stark für die
unterschiedlichen Prädisposition der Kühe für eine Dislocatio abomasi verantwortlich
sein können.

124 Schlussfolgerungen
6 Schlussfolgerungen
In der vorliegenden Arbeit konnte gezeigt werden, dass Insulin neben möglichen
sekundären Effekten auch einen direkten Einfluss auf die Motilität der glatten
Muskulatur des Labmagens hat.
Die in vitro gezeigte Hemmung der circulären Corpusmuskulatur könnte zu einem
verminderten Vorschub führen, der, bei gleichzeitig unbeeinflusster oder gar
stimulierter Pylorusmuskulatur, in einem reduzierten Labmagenefflux resultieren
würde. Dieser deutete sich in vivo bei vier von sechs Kühen an. Die Insulin-bedingten
Störungen der Labmagenmotilität könnten demnach ursächlich an der Entstehung
der Atonien, die im Zusammenhang mit Labmagenverlagerungen beobachtet
werden, beteiligt sein.
Aus den In-vitro-Versuche mit Barium, Glybenclamid und Ouabain ergeben sich
Hinweise auf eine mögliche Beteiligung von Kaliumkanälen und der Na+/K+-ATPase
an der Insulinwirkung auf die Labmagenmuskulatur des Rindes.
Um diese Hinweise zu bestätigen, könnten in weiteren Versuchen Messungen von
Membranpotentialen insbesondere an der circulären Corpusmuskulatur durchgeführt
werden. Auf diese Art und Weise könnte genauer geprüft werden, in welchem
Umfang Insulin über eine veränderte Kaliumleitfähigkeit und/oder eine Stimulation
der Na+/K+-ATPase das Membranpotential erniedrigt und damit das Öffnen von
spannungsabhängigen Kalziumkanälen mit nachfolgender Muskelkontraktion
erschwert.
Aufgrund der großen individuellen Unterschiede im In-vivo-Teil dieser Arbeit, wäre
eine Fortführung dieses Versuches an einer größeren Tierzahl sinnvoll, um
Folgendes zu evaluieren:
1. Bei wie vielen Kühen kommt es infolge euglykämischen hyperinsulinämischen
Clamps tatsächlich zu einer Reduktion des Labmageneffluxes?
2. Bei wie vielen Tieren ist dieser durch Ausgleich des Kaliumhaushaltes zu
kompensieren?

Schlussfolgerungen 125
3. Bestätigt sich der Verdacht, dass insbesondere trocken stehende Kühe
tatsächlich mit einem Wiederanstieg der Effluxmenge auf ausgleichende
Kaliumgaben reagieren?
4. Wie viele Kühe reagieren bei einer Insulininfusion von 1 mU/kg KGW/min mit
einer starken Abnahme der Kaliumkonzentration im Blut? Gibt es genetische
Unterschiede?
5. Bestätigt sich der Verdacht, dass die Höhe der Reduktion der Kaliumspiegel
entscheidend ist für den Verlauf der Effluxmessungen?

126 Zusammenfassung
7 Zusammenfassung
Gesche Türck: Bedeutung von Kalium und Insulin für die Motilität der
Labmagenmuskulatur
Die Labmagenverlagerung ist eine häufige Erkrankung hochleistender Milchkühe,
deren Pathogenese bislang nur teilweise geklärt ist. Hypokaliämien und
Insulinimbalancen werden oft im Zusammenhang mit gestörter Labmagenmotilität
beobachtet und könnten damit an der Entstehung der Verlagerung beteiligt sein. Ziel
dieser Arbeit war es herauszufinden, wie isolierte Labmagenmuskulatur einerseits
auf Veränderungen der extrazellulären Kalium-Konzentration und andererseits auf
steigende Insulingaben reagiert. Im Falle eines auftretenden Insulineffektes sollte
geklärt werden, inwieweit eine Insulin-bedingte Hemmung der Muskelaktivität auf
eine erhöhte Kalium-Leitfähigkeit und/oder eine Stimulation der Na+/K+-ATPase
zurückgeführt werden kann, wie es bereits in anderen Geweben nachgewiesen
werden konnte. In einem letzten Schritt wurde überprüft, inwieweit in vivo der
Labmagenefflux durch Insulininfusion reduziert werden kann und ob diese Reduktion
durch Kaliumsubstitution kompensierbar ist.
Im Zuge des In-vitro-Versuchs wurde von 22 Ziegenböcken und 74 nicht tragenden
Milchkühen der Rasse Holstein-Frisian bei der Schlachtung aus der Labmagenwand
zirkuläre Muskulatur aus Pylorus und Corpus isoliert und in einer Pufferlösung
inkubiert. Bei der Ziege wurde zusätzlich auch longitudinale Corpusmuskulatur zur
Überprüfung eines möglichen Insulineffektes untersucht. Mit Hilfe von
Kraftaufnehmern wurde die spontane isometrische Kraftentwicklung dieser
Muskelstreifen bei verschiedenen Zugaben von Kalium (1 bis 10 mmol/l) und Insulin
(0 bis 120 mU/l) gemessen. Ausgewertet wurden Frequenz (1/min) und Amplitude
(mN) der Muskelkontraktionen sowie die Kontraktionsaktivität (mN/min). Im Rahmen
des In-vivo-Versuchs wurde der Labmagenefflux von sechs fistulierten Kühen mithilfe
von Chrom-EDTA gemessen. Diese Versuche erfolgten unter euglykämischen
Bedingungen ohne Infusion (Tag 1), unter Infusion von Insulin (1 mU/kg KGW/min;

Zusammenfassung 127
(Tag 2) und unter Insulininfusion mit einer zusätzlichen Kaliumsupplementierung von
0,5 mmol/Tier/min (Tag 3).
In vitro konnte an allen Präparaten mit Ausnahme der Pylorusmuskulatur der Ziege in
unterschiedlicher Ausprägung eine durch steigende Kaliumgaben bedingte Zunahme
von Amplitude und Kontraktionsaktivität bei unbeeinflusster Frequenz festgestellt
werden.
Hinsichtlich der Wirkung von Insulin auf die isolierte Labmagenmuskulatur gab es
unterschiedliche Reaktionen. Während die circuläre Corpusmuskulatur unter
Insulinzugaben im Gegensatz zur Kontrolle bei unveränderter Frequenz mit einer
Abnahme (Rind) bzw. einer geringeren Zunahme (Ziege) von Amplitude und
Kontraktionsaktivität reagierte, kam es bei der Pylorusmuskulatur der Ziege zu einer
Zunahme aller drei Parameter. Die übrigen Gewebe reagierten nicht auf Insulin. Die
hemmende Wirkung des Insulins auf die zirkuläre Corpusmuskulatur des Rindes
konnte dabei durch Gaben von Kalium, Barium (3 mmol/l; Kaliumkanalblocker),
Glybenclamid (0,05 mmol/l; Blocker ATP-sensitiver Kaliumkanäle) und Ouabain
(0,001 mmol/l; Hemmer der Na+/K+-ATPase) aufgehoben werden.
In vivo konnte im Zuge der Insulininfusion zwar eine Absenkung der
Kaliumkonzentration im Blut an Tag 2 und durch die Kaliumsupplementierung ein
Wiederanstieg der Werte an Tag 3 erzielt werden, jedoch kam es nur bei vier von
sechs Tieren zu einem nominellen Abfall des Labmageneffluxes am zweiten und
davon nur bei zwei Tieren zu einem Wiederanstieg am dritten Versuchstag.
In vitro konnte also zum einen gezeigt werden, dass die extrazelluläre
Kaliumkonzentration in direktem Zusammenhang mit der Motilität der glatten
Muskulatur des Labmagens steht und damit in der Lage wäre, die Aktivität der
Magenpumpe und die Effluxmenge zu beeinflussen. Zum anderen konnten
Amplitude und Kontraktionsaktivität der zirkulären Corpusmuskulatur durch Insulin
gehemmt werden. Da diese Hemmung durch Gabe von Kalium, Barium,
Glybenclamid sowie Ouabain kompensierbar war, weisen die Ergebnisse der
Motilitätsmessung darauf hin, dass Insulin seine Wirkung an der zirkulären
Labmagenmuskulatur über eine Erhöhung der Kaliumleitfähigkeit und/oder eine
Erhöhung der Aktivität der Na+/K+-ATPase entfalten könnte.

128 Zusammenfassung
Bezüglich der Übertragbarkeit der in vitro gewonnenen Erkenntnisse auf
Bedingungen in vivo kann aufgrund der großen individuellen Unterschiede in der
Reaktion des Effluxes auf die Insulin- und Kaliuminfusionen keine abschließende
Aussage getroffen werden.

Summary 129
8 Summary
Gesche Türck: Effect of potassium and insulin on the abomasal smooth muscles
Abomasal displacement is a common disease of high-yielding dairy cows of which
the pathogenesis is only partly understood. Hypokalemia and insulin imbalances are
often observed in connection with disturbed abomasal motility, and might be involved
in the development of abomasal displacement.
Aim of the present study was to test the reaction of isolated abomasal smooth
muscle stripes to changes in the extracellular potassium-concentration and to
increasing doses of insulin. In case of an insulin-effect the contribution of an
enhanced potassium-conductance and a stimulation of the Na+/K+-pump to this effect
had to be tested, in analogy to insulin effects known from other tissues. At least we
investigated in vivo, if the abomasal outflow could be reduced by infusions of insulin
and if potassium –substitutions might be able to compensate a potential reduction.
Motility measurements in vitro were performed with circular smooth muscle stripes
isolated from abomasal pylorus and corpus of 22 male goats and 74 non-pregnant
Holstein-Frisian cows immediately after slaughter and incubated in buffer solution. A
potential insulin-effect was also tested with longitudinal smooth muscles from caprine
corpus abomasi. Spontaneous isometric contractions were measured in the presence
of increasing concentrations of potassium (1 to 10 mmol/l) and insulin (0 to 120
mU/l). Contractions were analysed for frequency (1/min), amplitude (mN) and
contraction-activity (mN/min). For the in-vivo part of the study, the abomasal outflow
of six fistulated cows was measures with Cr-EDTA. These experiments were
conducted under euglycemic conditions without infusions (day 1), with infusion of
insulin (1 mU/kg BW/min; day 2) and with infusion of insulin together with an
additional potassium-supplement of 0.5 mmol/cow/min (day 3).
All tissues with the exception of goat pylorus reacted to increasing potassium
concentrations with a distinct increase in amplitude and contraction-activity while the
frequency was not influenced. The effect of insulin on the isolated smooth muscle
stripes differed between the localisations. While circular muscles of the corpus

130 Summary
reacted with a decrease of amplitude and contraction-activity (in cows) or a less
pronounced increase of these parameters compared with control (in goats), the
caprine pylorus showed an increase in all three parameters. The remaining tissues
didn’t react to insulin. The inhibitory effect of insulin on the circular smooth muscles
of the corpus could be antagonized by potassium, barium (3 mmol/l; inhibitor of K+-
channels), glibenclamide (0.05 mmol/l; inhibitor of ATP-sensitive K+-channels) and
ouabain (0.001 mmol/l; inhibitor of the Na+/K+-pump).
The insulin infusion in the in vivo experiment induced a decrease in the blood
potassium-concentration (day 2) which could be overcome by the potassium-
supplements (on day 3). However only four of the six cows showed a nominally
reduction in abomasal outflow on day 2 and only two of them showed a re-increase in
the outflow on day 3.
In conclusion, the results of the in vitro experiments showed a direct effect of
extracellular potassium concentration on the motility of the abomasal smooth muscle.
Potassium may thus influence the activity of the stomach-pump and the abomasal
outflow. Insulin inhibited amplitude and contraction-activity of circular muscles from
abomasal corpus. Since this effect could be antagonized by the addition of
potassium, barium, glibenclamide or ouabain, these results point to an increase in
potassium-conductance and/or an increase in Na+/K+-pump activity as possible
mechanisms of insulin.
Due to the individual differences between cows concerning the changes in abomasal
outflow after the insulin- and potassium-infusions, it is not possible to draw a definite
conclusion to which extent these in-vitro-results would be applicable to the
circumstances in vivo.

Literaturverzeichnis 131
9 Literaturverzeichnis
ANDERSON, R. S. u. E. C. PICKERING (1962):
Effects of intravenous infusion of potassium chloride on potassium and sodium
excretion and on the rate of urine formation in the cow.
J. Physiol. 164, 180-188
ARNQVIST, H. J. (1975):
Effect of alloxan-diabetes on the metabolism of rabbit colon smooth muscle.
Acta Physiol. Scand. 93, 500-504
BANNINK, A., VALK, H. u. A. M. VANVUUREN (1999):
Intake and excretion of sodium, potassium, and nitrogen and the effects on urine
production by lactating dairy cows.
J. Dairy Sci. 82, 1008-1018
BAUMANN, D. E. u. W. B. CURRIE (1980):
Partitioning of nutrients during pregnancy and lactation: a review of mechanisms
involving homeostasis and homeorhesis.
J. Dairy. Sci. 63, 1514-1529
BECK, U., STRANGASSINGER, M., GIESECKE, D. u. J. MEYER (1983):
The testing of insulin receptor activity in ruminants.
In: 5. Int. Congr. On Production Disease in Farm Animals,
Uppsala, Schweden, 1983, Proc. S. 175-178
BERSON, S. A. u. R. S. YALOW (1970):
Insulin antagonist and insulin resistance.
In: ELLENBERG, M. u. H. RIFKINS (Hrsg.)
Diabetes Mellitus: Theory and practice
McGraw-Hill, New York, S. 388-412

132 Literaturverzeichnis
BERWECK,S., THIEME, H., LEPPLE-WIENHUES, A., HELBIG, H. u. M.
WIEDERHOLT (1993):
Insulin-induced hyperpolarization in retinal capillary pericytes.
Investig. Ophthalm. Vis. Sci. 34(12), 3402-3407
BLOOM, S. R. u. A. V. EDWARDS (1981):
Pancreatic endocrine responses to stimulation of the peripheral ends of the vagus
nerves in conscious calves.
J. Physiol. 315, 31-41
BLUM, J. W., R. M. BRUCKMAIER u. P.-Y. VACHER (1999):
Insulin-dependent whole-body glucose utilization and insulin-responses to glucose in
week 9 and week 19 of lactation in dairy cows fed rumen-protected crystalline fat or
free fatty acids.
Domest. Anim. Endocrinol. 16(2), 123 - 134.
BÖCK, P. u. H.-G. LIEBICH (1999):
Endokrines System (Systema endocrinum).
In: Funktionelle Histologie der Haussäugetiere: Lehrbuch und Farbatlas für Studium
und Praxis (3rd edn)
F. K. Schattauer Verlagsgesellschaft mbH, Stuttgart, S. 157-175
BOINI, K. M., GRAF, D., KUHL, D., HÄUSSINGER, D. u. F. LANG (2008):
SGK1 dependence of insulin induced hypokalemia.
Pflügers Arch. Jul 30. [Epub ahead of print]
BROBST, D. (1986):
Review of the pathophysiology of alterations in potassium homeostasis.
J. Am. Vet. Med. Assoc.188(9),1019-25.

Literaturverzeichnis 133
BROCKMAN, R. P. (1978):
Roles of glucagon and insulin in the regulation of metabolism in ruminants - a review.
Can. Vet. J. 19(3), 55-62
BROCKMAN, R. P. (1979):
Roles for insulin and glucagons in the development of ruminant ketosis – a review.
Can. Vet. J. 20(5), 121-126
BROCKMAN, R. P. (1986):
Pancreatic and adrenal hormonal regulation of metabolism.
In: MILLIGAN, L. P., GROVUM, W. L. u. A. DOBSON (Hrsg.)
Control of digestion and metabolism in ruminants
A Reston Book, Prentice-Hall, Englewood Cliffs, New Jersey, S. 405-419
BUENO, L. u. M. RUCKEBUSCH (1976):
Insulin and jejunal electrical activity in dogs and sheep.
Am. J. Physiol. 230 (6), 1538-1544
BUTLER, S. T., PELTON, S. H. u. W. R. BUTLER (2004):
Insulin increases 17β-estradiol production by the dominant follicle of the first
postpartum follicle wave in dairy cows.
Reprod. 127, 537-545
CHINET, A. u. T. CLAUSEN (1984) :
Energetics of active sodium-potassium transport following stimulation with insulin,
adrenaline or sulbutamol in rat soleus muscle.
Pflügers Arch. 401, 160-166

134 Literaturverzeichnis
CLAUSEN, T. u. O. HANSEN (1977):
Active Na-K transport and the rate of ouabain binding. The effect of insulin and other
stimuli on skeletal muscle and adipocytes.
J. Physiol. 270, 415-430
CLAUSEN, T. u. P. G. KOHN (1977):
The effect of insulin on the transport of sodium and potassium in rat soleus muscle.
J. Physiol. 265, 19-42
COHEN, P., BARZILAI, N., LERMAN, A., HAREL, H., SZYLMAN, P. U. E. KARNIELI
(1991):
Insulin effects on glucose and potassium metabolism in vivo: evidence for selective
insulin resistance in humans.
J. Clin. Endocrinol. Metab. 73(3),564-568
CONSTABLE, P. D., MILLER, G. Y., HOFFSIS, G. F., HULL, B. L. u. D. M. RINGS
(1992):
Risk factors for abomasal volvolus and left abomasal displacement in cattle.
Am. J. Vet. Rec. 53(7), 1184-1192
COTTRELL, D. F. (1994):
Vagal reflex inhibition of motility in the abomasal body of sheep by antral and
duodenal tension receptors.
Vet. Res. Commun. 18(4), 319-330
COTTRELL, D. F. u. H. G. STANLEY (1992):
An excitatory body-antral reflex in the sheep abomasums.
Exp. Physiol. 77(4), 565-574

Literaturverzeichnis 135
CREESE, R. (1968):
Sodium fluxes in diaphragm muscle and the effects of insulin and serum proteins.
J. Physiol. 197, 255-278
CREESE, R. u. J. NORTHOVER (1961):
Maintenance of isolated diaphragm with normal sodium content.
J. Physiol. 155, 343-357
DANIEL, R. C. W. (1983):
Motility of the rumen and abomasums during hypocalcaemia.
Can. J. Comp. Med. 47, 276-280
DAVIES, J.V. (1989):
Normal intestinal motility.
Vet. Clin. North Am. Equine. Pract. 5, 271-281.
DEBRASS, E., GRIZARD, J., AINA, E., TESSERAUD, S., CHAMPREDON, C. u. M.
ARNAL (1989):
Insulin sensitivity and responsiveness during Lactation and dry period in goats.
Am. J. Physiol. 256, E295-E302
DE JONG, A. (1982):
Patterns of plasma concentrations of insulin and glucagons after intravascular and
intraruminal administration of volatile fatty acids in the goat.
J. Endocrinol. 92(3), 357-370
DELGADO-LECAROZ, R., WARNICK, L. D., GUARD, C. L., SMITH, M. C. u. D. A.
BARRY (2000):
Cross-sectional study of the association of abomasal displacement or volvolus with
serum electrolyte and mineral concentrations in dairy cows.
Can. Vet. J. 41, 301-305

136 Literaturverzeichnis
DENNIS, J. u. F. G. HARBAUGH (1948):
The experimental alteration of blood potassium and calcium levels in cattle.
Am. J. Vet. Res, 9, 20-25
DETILLEUX, J.C., Y.T. GRÖHN, S.W. EICKER u. R.L. QUAAS (1997):
Effects of left displaced abomasum on test day milk yields of Holstein cows.
J. Dairy Sci. 80, 121-126.
DEWHURST, J. K., HARRISON, F. A. u. R. D. KEYNES (1968):
Renal excretion of potassium in the sheep.
J. Physiol. 195, 609-621
DIRKSEN, G. (1961):
Vorkommen, Ursachen und Entwicklung der linksseitigen Labmagenverlagerung
(Dislocatio abomasi sinistra) des Rindes.
Dtsch. Tierärztl. Wschr. 68, 8-12.
DRAZNIN, B., SUSSMAN, K., MAO, M., LEWIS, D. u. N. SHERMAN (1987):
The existence of an optimal range of cytosolic free calcium for insulin-stimulated
glucose transport in rat adipocytes.
J. Biol. Chem. 262(30:25), 14385-14388
DUDEL, J. (2000):
Informationsvermittlung durch elektrische Erregung.
In: SCHMIDT, R. F., THEWS, G. u. F. LANG
Physiologie des Menschen
Springer-Verlag, Berlin, Heidelberg, New York, S. 20-42

Literaturverzeichnis 137
ECKEL, J. u. H. REINAUER (1990):
Modulation of transmembrane potential of isolated cardiac myocytes by insulin and
isoproterenol.
Am. J. Physiol. 259, H554-H559
EHRLEIN, H. J. (2000):
Muskelphysiologie: glatte Muskulatur.
In: VON ENGELHARDT, W. u. G. BREVES (Hrsg.)
Physiologie der Haustiere
Enke im Hippokrates Verlag, Stuttgart, S. 126-130
EHRLEIN, H. J. (2000):
Motorik des einhöhligen Magens und des Labmagens.
In: VON ENGELHARDT, W. u. G. BREVES (Hrsg.)
Physiologie der Haustiere
Enke im Hippokrates Verlag, Stuttgart, S. 317-323
ERLIJ, D. (1984):
Vanadate stimulates the pumped movements of Na in skeletal muscle.
Pfügers Arch. 400, 413-417
ERLIJ, D. u. S. GRINSTEIN (1975):
The number of sodium ion pumping sites in skeletal muscle and its modification by
insulin.
J. Physiol. 259, 13-31
ESPERSEN, A. G. u. M. G. SIMENSEN (1961):
Alkalose ved hojresidig lobedilatation.
Nord. Vet.-Med. 18, 147-159

138 Literaturverzeichnis
FAULKNER, A. u. H. T. POLLOCK (1990):
Metabolic responses to euglycaemic hyperinsulinaemia in lactating and non-lactating
sheep in vivo.
J. Endocrinol. 124, 59-66
FICKOVA, M., ZORAD, S. u. L. MACHO (1997):
The effect of in vivo thyroxin treatment on insulin receptors, glucose transport and
GLUT 4 in rat adipocytes.
Horm. Metabol. Res. 29, 16-19
FLATMAN, J. A. u. T. CLAUSEN (1979):
Combined effects of adrenalin and insulin on active eletrogenic Na+-K+ transport in
rat soleus muscle.
Nature 281, 590-581
FLYNN, E. R. M., MCMANUS, C. A., BRADLEY, K. K., KOH, S. D., HEGARTY, T.
M., HOROWITZ, B. u. K. M. SANDERS (1999):
Inward rectifier potassium conductance regulates membrane potential of canine
colonic smooth muscle.
J. Physiol. 518, 247-256
FORD, E. J. H. (1950):
A case of displacement of the bovine abomasums
Vet. Rec. 62, 763-764
FORREST, A. u. M. PARSONS (2003) :
The enhanced spontaneous activity of the diabetic colon is not the consequence of
impaired inhibitory control mechanisms.
Auton. Autacoid. Pharmacol. 23(3),149-158

Literaturverzeichnis 139
FROMM, M. u. K. HIERHOLZER (2000):
Niere.
In: VON ENGELHARDT, W. u. G. BREVES (Hrsg.)
Physiologie der Haustiere
Enke im Hippokrates Verlag, Stuttgart, S. 254-284
GARNSWORTHY, P. C. u. J. H. TOPPS (1982):
The effect of body condition of dairy cows at calving on their food intake and
performance when given complete diets.
Anim. Prod. 35, 113-119
GAVRYCK, W. A., MOORE, R. D. u. R. C. THOMPSON (1974):
Effect of insulin upon membrane-bound (Na++K+)-ATPase extracted from frog
skeletal muscle.
J. Physiol. 252, 43-58
GEISHAUSER, T. (1995):
Abomasal displacement in the bovine – an review on character, occurrence,
aetiology and pathogenesis.
Zentralbl. Vet Med. A. 42(4), 229-251
GEISHAUSER, T., M. DIEDERICHS u. R. BEUING (1996):
Schätzung der Erblichkeit von Labmagenverlagerung bei Deutsch-Schwarzbunten
Rindern in Hessen.
J. Vet. Med. A. 43, 87-92.
GEISHAUSER, T., LESLIE, K. E. u. T. F. DUFFIELD (2000):
Metabolic aspects in the etiology of displaced abomasums.
Vet. Clin. North Am. Food Anim. Pract. 16, 255-265

140 Literaturverzeichnis
GERSHON, M., KIRCHGESSNER, A. L. u. P. R. WADE (1994):
Functional anatomy of the enteric nervous system.
In: L. R. JOHNSON (Hrsg.)
Physiology of the gastrointestinal tract (3rd edn.)
Raven Press, New York, S. 381-422
GIESECKE, D. (1986):
Insulin deficiency and metabolic disorders in high-yielding dairy cows.
J. South Afr. Vet. Assoc. 57(1), 67-70
GOFF, J. P. (1999):
Physiologic factors to consider in order to prevent periparturient disease in the dairy
cow, with special emphasis on milk fever.
Western Dairy management conference, 8. – 10. April 1999, Las Vegas, Nevada, S.
99-112
GOSENS, R., NELEMANS, S. A., HIEMSTRA, M., GROOTTE BROMHAAR, M. M.,
MEURS, H. u. J. ZAAGSMA (2003):
Insulin induces a hypercontractile airway smooth muscle phenotype.
Europ. J. Pharmacol. 481, 125-131
GOURLEY, D. R. H. (1965) :
Effect of insulin on potassium exchange in normal and ouabain-treated skeletal
muscle.
J. Pharmacol. Exp. Ther. 148, 339-347
GRATHAM, B. D. u. V. A. ZAMMIT (1988):
Role of carnitine palmitoyltransferase I in the regulation of hepatic ketogenesis during
the onset and reversal of chronic diabetes.
J. Biochem. 249, 409-414

Literaturverzeichnis 141
GREEN, D. M. u. L. RABINOWITZ (1979):
Renal components of feed dependent K excretion in sheep (Absract).
Kidney Int. 16, 833
GRIINARI, J. M., MC GUIRE, M. A., DWYER, D. A., BAUMAN, D. E. u. D. L.
PALMQUIST (1997):
The role of insulin in the regulation of milk fat synthesis in dairy cows.
J. Dairy Sci. 80, 1076-1084
GRIZARD, J., BALAGE, M. u. M. MANIN (1986):
Controle hormonal du metabolisme hepatique chez les ruminants.
Reprod. Nutr. Develop. 26(1B), 245-257
GRIZARD, J., C. CHAMPREDON, E. AINA, C. SORNET u. E. DEBRAS (1988):
Metabolism and action of insulin and glucagon in goat during lactation and dry
period.
Horm. Metabol. Res. 20, 71 - 76.
GRIZARD, J. u. M. SZCZYGIEL (1983):
Insulin binding to liver plasma membranes from growing ruminating sheep.
Maintained on different dieats.
Horm. Metabol. Res. 15, 475-481
GUESNET, P. M., MASSOUD, M. J. u. Y. DEMARNE (1991) :
Regulation of adipose tissue metabolism during pregnancy and lactation in the ewe:
the role of insulin.
J. Anim. Sci. 69, 2057-2065

142 Literaturverzeichnis
HALSE, K. (1960):
Changes in serum calcium and magnesium in cows subjected to short periods of
fasting.
Proc. 8th Int. Grassland Congr. 553
HARA, Y. u. J. H. SZURSZEWSKI (1986):
Effect of potassium and acetylcholine on canine intestinal smooth muscle.
J. Physiol. 372, 521-537
HART, I. C., J. A. BINES u. S. V. MORANT (1980):
The secretion and metabolic clearance rates of growth hormone, insulin and prolactin
in high and low-yielding cattle at four stages of lactation.
Life Sci. 27, 1839 - 1847
HATTON, W. J., MASON, H. S., CARL, A., DOHERTY, P., LATTEN, M. J., KENYON,
J. L., SANDERS, K. M. u. B. HOROWITZ (2001):
Functional and molecular expression of a voltage-dependent K+ channel (KV1.1) in
interstitial cells of Cajal.
J. Physiol. 533, 315-327
HAY, W. W., SPARKS, J. W., GILBERT, M., BATTAGLIA, F. C. u. G. MESCHIA
(1984):
Effect of insulin on glucose uptake by the maternal hindlimb and uterus, and by the
fetus in conscious pregnant sheep.
J. Endocrinol. 100(1), 119-124
HAYIRLI, A. (2006):
The role of exogenous insulin in the complex of hepatic lipidosis and ketosis
associated with insulin resistance phenomenon in postpartum dairy cattle.
Vet. Res. Com. 30, 749-774

Literaturverzeichnis 143
HAYIRLI, A., BREMMER, D. R., BERTICS, S. J., SOCHA, M. T. u. R. R. GRUMMER
(2001):
Effect of chromium supplementation on production and metabolic parameters in
periparturient dairy cows.
J. Dairy Sci. 84, 1218-1230
HAYIRLI, A., BERTICS, S. J. u. R. R. GRUMMER (2002):
Effects of slow-release insulin on production, liver, triglyceride, and metabolic profiles
of Holsteins in early lactation.
J. Dairy Sci. 85, 2180-2191
HERDT, T. (2000) :
Ruminant adaptation to negative energy balance: Influences on the Etiology of
ketosis and fatty liver.
Vet. Clin. North Am. Food. Anim. Pract. 16(2), 215-230
HILL, H. (1969):
Pathophysiologie der Verduung.
In: SPÖRRI, H. u. H. STÜNZI
Pathophysiologie der Haustiere.
Verlag Paul Parey, Berlin und Hamburg, S. 215-251
HOLTENIUS, P. u. M. TRAVEN (1990):
Impaired glucose tolerance and heterogeneity of insulin responses in cows with
abomasal displacement.
J. Vet. Med. A. 37, 445-451
HIPPEN, A. R., SHE, P., YOUNG, J. W., BEITZ, D. C., LINDBERG, G. L.,
RICHARDSON, L. F. u. R. W. TUCKER (1999):
Alleviation of fatty liver in dairy cows with 14-Da intravenous infusions of glucagons.
J. Dairy Sci. 82, 1139-1152

144 Literaturverzeichnis
HOLTENIUS, K., STERNBAUER, K. u. P. HOLTENIUS (2000):
The effect of the plasma glucose level on the abomasal function in dairy cows.
J. Anim. Sci. 78, 1930-1935
HOLTENIUS, K., AGENÄS, S., DELAVAUD, C. u. Y. CHILLIARD (2003):
Effects of feeding intensity during the dry period. 2. metabolic and hormonal
responses.
J. Dairy Sci. 86, 883-891
HOLTENIUS, P. und K. HOLTENIUS (1996):
New aspects of ketone bodies in energy metabolism of dairy cows: a review.
J. Vet. Med. A. 43, 579-587
HORINO, M., MACHLIN, L. J., HERTELENDY, F. u. D. M. KIPNIS (1968):
Effect of short-chain fatty acids on plasma insulin in ruminant and nonruminant
species.
Endocrinol. 83, 118-128
HOVE, K. (1978 a):
Maintenance of lactose secretion during acute insulin deficiency in lactating goats.
Acta Physiol. Scan. 103, 173-179
HOVE, K. (1978 b):
Insulin secretion in lactating cows: Responses to glucose infused intravenously in
normal, ketonemic, and starved animals.
J. Dairy Sci. 61(10), 1407-1413
IANNACCONE, S. T., LI, K. X., SPERELAKIS, N. u. D. A. LATHROP (1989):
Insulin-induced hyperpolarization in mammalian skeletal muscle.
Am. J. Physiol. Cell Physiol. 256, C368-C374

Literaturverzeichnis 145
JONES, R. G., V. ILIC u. D. H. WILLIAMSON (1984):
Regulation of lactating-rat mammary-gland lipogenesis by insulin and glucagon in
vivo. The role and site of action of insulin in the transition to the starved state.
Biochem. J. 223, 345 - 351
JUBB, T.F., J. MALMO, G.M. DAVIS u. A.S. VAWSER (1991):
Left-side displacement of the abomasum in dairy cows at pasture.
Aust. Vet. J. 68, 140-142.
KAHN, C. R. (1978):
Insulin resistance, insulin sensitivity, and insulin unresponsiveness: a necessary
distinction.
Metabol. 27, 1893-1902
KAHN, C. R. (1994)
Insulin action, diabetogenes, and the cause of type II diabetes.
Diab. 43, 1066-1084
KAMMINGA, C. E., WILLEBRANDS, A. F., GREON, J. u. J. R. BLICKMAN (1950):
Effect of insulin on the potassium and inorganic phosphate content of the medium in
experiments with isolated rat diaphragms.
Sci. 111, 30-31
KATZUNG, B. G. (1995):
Pancreatic hormones and antidiabetic drugs.
In: Basic and clinical pharmacology (6th edn.)
Appleton and Lange, Norwalk, Connecticut, S. 637-654

146 Literaturverzeichnis
KATO, K., KITO, Y. u. H. SUZUKI (2007):
Mechanical and electrical responses modulated by excitation of inhibitory nerves
during stimulation with high-potassium solutions in circular smooth muscle of the
rabbit rectum.
J. Smooth Muscle Res. 43(6), 229-246
KHORASANI, G. R., JANZEN, R. A., MCGILL, W. B. u. J. J. KENNELLY (1997):
Site and extent of mineral absorption in lactating cows fed whole-crop cereal grain
silage of alfalfa silage.
J. Anim. Sci. 75(1), 239-248
KITASATO, H., SATO, S., MARUNAKA, Y. MURAYAMA, K. U. K. NISHIO (1980a):
Effects of ouabain on the Na efflux in high internal Na and insulin-preincubated
muscles.
Jpn. J. Physiol. 30, 591-602
KITASATO, H., SATO, S., MARUNAKA, Y. MURAYAMA, K. U. K. NISHIO (1980b):
Apparent affinity changes induced by insulin of Na-K transport system in frog skeletal
muscles.
Jpn. J. Physiol. 30, 603-616
KITASATO, H., SATO, S., MURAYAMA, K. U. K. NISHIO (1980c):
The interaction between the effects of insulin and ouabain on the activity of Na
transport system in frog skeletal muscle.
Jpn. J. Physiol. 30, 115-130
KITO, Y. u. H. SUZUKI (2007):
Role of K+ channels in the regulation of electrical spontaneous activity of the mouse
small intestine.
Pflügers Arch. 455, 505-514

Literaturverzeichnis 147
KNOWLTON, K. F., DAWSON, T. E., GLENN, B. P., HUNTINGTON, G. B. u. R. A.
ERDMAN (1998):
Glucose metabolism and milk yield of cows infused abomasally or ruminally with
starch.
J. Dairy. Sci. 81, 3248-3258
KOBAYASHI, S., KANAIDE, H. u. M. NAKAMURA (1985):
K+-depolarization induces a direct release of Ca2+ from intracellular storage sites in
cultured vascular smooth muscle cells from rat aorta.
Biochem. Biophysic. Res. Commun. 129 (3), 877-884
KÖNIG, H. E., SAUTET, J. u. H.-G. LIEBICH (1999):
Verdauungsapparat (Apparatus digestorius)
In: KÖNIG, H. E. u. H.-G. LIEBICH (Hrsg.)
Anatomie der Haussäugetiere: Lehrbuch und Farbatlas für Studium und Praxis, Band
II, Organe, Kreislauf- und Nervensystem
F. K. Schattauer Verlagsgesellschaft mbH, Stuttgart, S. 15-80
KOMURO, T., SEKI, K. U. K. HORIGUCHI (1999):
Ultrastructural characterization of the interstitial cells of Cajal.
Histol. Histopathol. 11, 769-786
KOOPMANS, S. J., SIPS, H. C. u. H. M. KRANS (1996):
Pulsatile intravenous insulin replacement in streptozotocin diabetic rats is more
efficient than continuous delivery: effect of glycemic control, insulin-mediated glucose
metabolism and lipolysis.
Diabetolog. 39, 391-400

148 Literaturverzeichnis
KRÄFT, S. (2004):
Charakterisierung der peripheren Insulin-Response und Insulin-Sensitivität bei
trockenstehenden, laktierenden und leberverfetteten Milchkühen ohne und mit
Ketose mittels hyperinsulinämischer, euglycämischer Clamps.
Hannover, Tierärztl. Hochsch., Diss.
LANG, F. (2000):
Wasser- und Elektrolythaushalt.
In: SCHMIDT, R. F., THEWS, G. u. F. LANG
Physiologie des Menschen
Springer-Verlag, Berlin, Heidelberg, New York, S. 772-792
LANTZ, R. C., ELSAS, L. J. u. R. L. DE HAAN (1980):
Ouabain-resistant hyperpolarization induced by insulin in aggregates of embryonic
heart cells.
Proc. Natl. Acad. Sci. USA 77(5), 3062-3066
LE BLANC, S. J., LESLIE, K. E. u. T. F. DUFFIELD (2005):
Metabolic predictors of displaced abomasum in dairy cattle.
J. Dairy Sci. 88, 159-170
LEBRUN, P., MALAISSE, W. J. u. A. HERCHUELZ (1993) :
Na+-K+ pump activity and the glucose-stimulated Ca2+-sensitive K permeability in the
pancreatic B-cell.
J. Membr. Biol. 74, 67-73
LE-MARCHAND-BRUSTEL, Y., TANTI, J. F., CORMONT, M., RICORT, J. M.,
GREMEAUX, T. u. S. GRILLO (1999) :
From insulin receptor signalling to GLUT-4 translocation abnormalities in obesity and
insulin resistance.
J. Receptor Signal Transduct. Res. 19, 217-228

Literaturverzeichnis 149
LESTER, G. D. u. J. R. BOLTON (1994):
Effect of dietary composition on abomasal and duodenal myoelectrical activity.
Res. Vet. Sci. 57(3), 270-276
LEURY, B. J., BAUMGARD, L. H., BLOCK, S. S., SEGOALE, N., EHRHARDT, R. A.,
RHOADS, R. P., BAUMAN, D. E., BELL, A. W. u. BOISCLAIR, Y. R. (2003):
Effect of insulin and growth hormone on plasma leptin in periparturient dairy cows.
Am. J. Physiol. Regul. Integr. Comp. Physiol. 285, R1107-R1115
LIEBICH, H.-G. (1999):
Muskelgewebe (Textus muscularis)
In: Funktionelle Histologie der Haussäugetiere: Lehrbuch und Farbatlas für Studium
und Praxis (3rd edn)
F. K. Schattauer Verlagsgesellschaft mbH, Stuttgart, S. 85-96
LITTLEDIKE, E. T., WITZEL, D. A. u. S. C. WHIPP (1968):
Insulin: Evidence for inhibition of release in spontaneous hypocalcemia.
Proc. Soc. Exp. Biol. Med. 129, 135-141
LÖFFLER u. PETRIDES (1997):
Stoffwechsel der Kohlenhydrate.
In: LÖFFLER u. PETRIDES (Hrsg.)
Biochemie und Pathobiochemie, 5. Aufl.
Springer Verlag, Berlin, Heidelberg, New York, S. 378-423
LOMAX, M. A., G. D. BAIRD, C. B. MALLINSON u. H. W. SYMONDS (1979):
Differences between lactating and non-lactating dairy cows in concentration and
secretion rate of insulin.
Biochem. J. 180, 281 – 289

150 Literaturverzeichnis
LOPEZ, S., HOVELL, F.D. u. N. A. MACLEOD (1994)
Osmotic pressure, water kinetics and volatile fatty acid absorption in the rumen of
sheep sustained by intragastric infusions.
Br. J. Nutr. 71(2), 153-168
López, S., Hovell, F. D. D., Dijkstra, J. u. J. France (2003):
Effects of volatile fatty acid supply on their absorption and on water kinetics in the
rumen of sheep sustained by intragastric infusions.
J. Anim. Sci. 81, 2609-2616
LUNN, D. P. u. S. M. MC GUIRK (1990):
Renal regulation of electrolyte and acid-base balance in ruminants.
Vet. Clin. North Am. Food Anim. Pract. 6(1), 1-28
LUZI, L., ZOPPINI, G., TARGHER, G., BATTEZZATI, A., MUGGEO, M. u. E.
BONORA (2008):
Insulin effect on serum potassium and auto-inhibition of insulin secretion is intact in a
patient with leprechaunism despite severe impairment of substrates metabolism.
Diab. Metab. Res. Rev. 24(3), 205-210
MAGLAD, M. A., WASFI, I. A. u. S. E. ADAM (1983):
Inhibition of glucose-induced insulin secretion by somatostatin in goats.
Res. Vet. Sci. 35(1), 19-21
MANERY, J. F., FISHER, K. C., GOURLY, D. R. H., WILSON, D. L. u. M. I. TAYLOR
(1950):
The influence of carbohydrate and insulin on the potassium content of leucocytes
and muscle.
Biol. Bull. 99, 312

Literaturverzeichnis 151
MANERY, J. F., DRYDEN, E. E., STILL, J. S. u. G. MADAPALLIMATTAM (1977):
Enhancement (by ATP, insulin, and lack of divalent cations) of ouabain inhibition of
cations transport and ouabain binding in frog skeletal muscle: effect of insulin and
ouabain on sarcolemmal (Na+K) Mg ATPase.
Can. J. Physiol. Pharmacol. 55, 21-33
MALBERT, C. H. u. Y. RUCKEBUSCH (1988):
Gastroduodenal motor activity associated with gastric emptying rate in sheep.
J. Physiol. 401, 227-239
MALBER, C. H. u. Y. RUCKEBUSCH (1991):
Emptying of the abomasum in adult ruminants.
Reprod. Nutr. Dev. 31(1), 1-25
MALVEN, P. V., HAED, H. H., COLLIER, R. J. u. F. C. BUONOMO (1987):
Periparturient changes in secretion and mammary uptake of insulin and in
concentrations of insulin and insulin-like growth factors in milk of dairy cows.
J. Dairy Sci. 70, 2254-2265
MANNS, J. G. u. J. M. BODA (1967):
Insulin release by acetate, propionate, butyrate, and glucose in lambs and adult
sheep.
Am. J. Physiol. 212(4), 747-755
MANNS, J. G., BODA, J. M. u. R. F. WILLES (1967):
Probable role of propionate and butyrate in control of insulin secretion in sheep.
Am. J. Physiol. 212(4), 756-764

152 Literaturverzeichnis
MARTIN, P. A. u. M. H. CRUMP (2003):
The endocrine pancreas.
In: MC DONALD, L. E. (Hrsg.)
Veterinary endocrinology and reproduction (6th edn.)
Lea and Febiger, Philadelphia, S. 141-163
MARUNAKA, Y. (1986 a):
The stimulatory action of [Na]0 reduction on insulin-stimulated 3-o-methyl-D-glucose
efflux from frog sartorius muscle; a change in the membrane potential as a mediator
of insulin action.
IRCS Med. Sci. 14, 453-454
MARUNAKA, Y. (1986 b):
Effects of internal Na concentration on ouabain-sensitive Na efflux and membrane
potential.
IRCS Med. Sci. 14, 601-602
MARUNAKA, Y. (1987):
The effect of the internal Na concentration on the electrogenicity of the insulin-
stimulated Na,K-pump in frog skeletal muscles.
Comp. Biochem. Physiol. 86A, 133-136
MARUNAKA, Y. u. H. KITASATO (1985):
The inhibitory action of insulin-induced hyperpolarization 3-o-methyl-D-glucose efflux
from frog sartorius muscles.
IRCS Med. Sci. 13, 637-638
MC CANN, J. P. u. T. J. REIMERS (1985):
Glucose response to exogenous insulin and kinetics of insulin metabolism in obese
and lean heifer.
J. Anim. Sci. 61, 612-618

Literaturverzeichnis 153
MC CANN, J. P., ULLMANN, M. B., TEMPLE, M. R., REIMERS T. J. u. E. N.
BERGMAN (1986):
Insulin and glucose response to glucose injection in fed and fastedobese and lean
sheep.
J. Nutr. 116, 1287-1297
MC CONALOGUE, K. u. J. B. FURNESS (1994) :
Gastrointestinal neurotransmitters.
Bailleres. Clin. Endocrinol. Metabol. 8, 51-76
MC GUIRK, S. M. u. D. G. BUTLER (1980):
Metabolic alkalosis with paradoxic aciduria in cattle.
J. Am. Vet. Med. Assoc. 177(6), 551-554
MERSMANN, H. J. (1987):
Nutritional and endocrinological influences on the composition of animal growth.
Prog. Food Nutr. Sci. 11(2), 175-201
MINEO, H., KANAI, M., KATO, S. u. J. I. USHIJIMA (1990):
Effects of intravenous injection of butyrate, valerate and their isomers on endocrine
pancreatic responses in conscious sheep (ovis aries).
Comp. Biochem. Physiol. A. 95(3), 411-416
MOORE, R. D. (1973):
Effect of insulin upon the sodium pump in frog skeletal muscle.
J. Physiol. 232, 23-45
MOORE, R. D. u. J. L. RABOVSKY (1979):
Mechanism of insulin action on resting membrane potential of frog skeletal muscle.
Am. J. Physiol. Cell Physiol. 236, C249-C254

154 Literaturverzeichnis
NELSON, D. R., PETERSEN, G. C., HUHN, J. C. u. W. A. OLSON (1995):
Electromyography of the reticulum, abomasums and duodenum in dairy cows with
left displacement of the abomasums.
J. Vet. Med. A. 42, 325-337
O`BRIEN, R. M. u. D. K. GRANNER (1990):
PEPCK gene as a model of inhibitory effects of insulin on gene transcription.
Diab. Care, 13, 327-334
ODA, S., OHTOMO, Y., OHNEDA, A., SASAKI, Y. u. T. TSUDA (1986):
Adrenergic modulation of pancreatic glucagons and insulin secretion in goats.
Comp. Biochem. Physiol. A. 84(4) 723-728
OK, M., SEN, I., BIRDANE, F. M., SEVINC, M., ASLAN, V. u. F. ALKAN (2000):
Concentration of insulin and glucose in dairy cows with abomasal displacement.
Indian Vet. J. 77, 961-962
OSTERGAARD, S. u. Y. T. GRÖHN (1999):
Effects of diseases on test day milk yield and body weight of dairy cows from Danish
research herds.
J. Dairy Sci. 82, 1188-1201
PAVEL, I. U. S. M. MILCOU (1932) :
Action de l’insuline sur l’intestin.
Compt. Rend. Soc. Biol. 109, 776-778
PETRIDES (1997):
Wasser- und Elektrolythaushalt.
In: LÖFFLER u. PETRIDES (Hrsg.)
Biochemie und Pathobiochemie, 5. Aufl.
Springer Verlag, Berlin, Heidelberg, New York, S. 679-706

Literaturverzeichnis 155
PETTERSON, J. A., SLEPETIS, R., EHRHARDT, R. A., DUNSHEA, F. R. u. A. W.
BELL (1994) :
Pregnancy but not moderate undernutrition attenuates insulin suppression of fat
mobilization in sheep.
J. Nutr., 124, 2431-2436
PFEFFER, E. (1987):
Stoff- und Energiewechsel: Verduung.
In: SCHEUNERT, A. , TRAUTMANN, A. u. G. WITTKE
Lehrbuch der Veterinär-Physiologie
Verlag Paul Parey, Berlin und Hamburg, S. 29-57
POULSEN, J. S. D. (1976):
Aetiology and pathogenesis of abomasal displacement in dairy cattle.
Nord. Vet. Med. 28, 299-303
PRAVETTONI, D., DOLL, K., HUMMEL, M., CAVALLONE, E., RE, M. u. A. G.
BELLONI (2004):
Insulin resistance and abomasal motility disorders in cows detected by use of
abomasoduodenal electromyography after surgical correction of left displaced
abomasums.
Am. J. Vet. Res. 65(10),1319-24
PRAVETTONI, D., DOLL, K., HUMMEL, M., CAVALLONE, E., MORANDI, N. u. A. G.
BELLOLI (2007):
Glucose tolerance test during abomaso-duodenal electromyography in three cows
operated for left displaced abomasums.
Dtsch. Tierärztl. Wochenschr. 114, 58-63

156 Literaturverzeichnis
PRIOR, R. L. u. R. K. CHRISTENSON (1978):
Insulin and glucose effects on glucose metabolism in pregnant and nonpregnant
ewes.
J. Anim. Sci. 46(1), 201-210
RABINOWITZ, L. u. R. L. SARASON (1980):
The concurrent kaliuresis and the delayed kaliuresis produced by KCl infusion in
sheep (Abstract).
Physiologist. 23, 14
RABINOWITZ, L., SARASON, R. L. u. H. YAMAUCHI (1981):
Aldosterone and the control of potassium excretion in sheep (Abstract).
Kidney Int. 21, 285
RABINOWITZ, L., SARASON, R. L. u. H. YAMAUCHI (1984 a):
Sheep renal potassium excretion: efferent kaliuretic regulatory factors.
Am. J. Physiol. Renal Physiol. 247, F520-F526
RABINOWITZ, L., SARASON, R. L, TANASOVICH, C., MENDEL, V. E. u, R. P.
BROCKMAN (1984 b):
Effects of glucagons, insulin, propionate, acetate, and HCO3 on K excretion in sheep.
Am. J. Physiol. Regul. Integr. Comp. Physiol. 246, R197-R204
RABINOWITZ, L., SARASON, R. L. u. H. YAMAUCHI (1985):
Effect of aldosterone on potassium excretion during potassium chloride infusion in
sheep.
Am. J. Physiol. 249 (4 Pt 2), R455-R461
RABINOWITZ, L., GREEN, D. M., SARASON, R. L. u. H. YAMAUCHI (1988):
Homeostatic potassium excretion in fed and fasted sheep.
Am. J. Physiol. 254(2 Pt 2), R357-380

Literaturverzeichnis 157
RICHARD, A. L., MC CUTCHEON, S. N. u. D. E. BAUMAN (1985):
Responses of dairy cows to exogenous bovine growth hormone administered during
early lactation.
J. Dairy Sci. 68, 2385-2389
RICKEN, M. W. (2003):
Labmagenverlagerung beim Rind: Analyse von genetischen Faktoren und
ökonomischen Auswirkungen auf die Milchproduktion.
Hannover, Tierärztl. Hochsch., Diss.
RIZZA, R. A., MANDARINO, L. J. u. J. E. GERICH (1981):
Mechanisms of insulin resistance in man.
Am. J. Med. 70, 169-176
ROBERTS, W. K. u. V. V. S. OMER (1965):
Dietary potassium requirement of fattening steers (Abstract).
J. Anim. Sci. 24, 902
ROHRBACH, B. W., CANNEDY, A. L., FREEMAN, K. u. B. D. SLENNING (1999):
Risk factors for abomasal displacement in dairy cows.
J. Am. Vet. Ned. Assoc. 214(11), 1660-1663
ROSSOW, N. (1995):
Erkrankungen der Vormägen und des Labmagens.
In: Innere Medizin für Tierärzte
Eugen Ulmer Verlag GmbH u. Co, Stuttgart (Hohenheim), S. 155-173

158 Literaturverzeichnis
RÜEGG, J. C. (2000):
Muskel.
In: SCHMIDT, R. F., THEWS, G. u. F. LANG
Physiologie des Menschen
Springer-Verlag, Berlin, Heidelberg, New York, S. 67-91
RYAN, E. A. u. L. ENNS (1988):
Role of gestational hormones in the induction of insulin resistance.
J. Clin. Endocrinol. Metabol. 67, 341-347
SAKAI, T., HAMAKAWA, M. u. S. KUBO (1996):
Glucose and xylitol tolerance tests for ketotic and healthy dairy cows.
J. Dairy Sci. 79, 372-377
SANDER, K. M. (1996):
A case for intestinal cells of Cajal as pacemakers and mediators of
neurotransmission in the gastrointestinal tract.
Gastroenterol. 111, 492-515
SANO, H., NAKAI, M., KONDO, T. u. Y. TERASHIMA (1991):
Insulin responsiveness to glucose and tissue responsiveness to insulin in lactating,
pregnant, and nonpregnant, nonlactating beef cows.
J. Anim. Sci. 69, 1122-1127
SANO, H., HATTORI, N., TODOME, Y., TSURUOKA, J., TAKAHASHI, H. u. Y.
TERASHIMA (1993 a):
Plasma insulin and glucagon responses to intravenous infusion of propionate and
their autonomic control in sheep.
J. Anim. Sci. 71, 3414-3422

Literaturverzeichnis 159
SANO, H., NARAHARA, S., KONDO, T., TAKAHASHI, A. u. Y. TERASHIMA
(1993 b):
Insulin responsiveness to glucose and tissue responsiveness to insulin during
lactation in dairy cows.
Dom. Anim. Endocrinol. 10(3), 191-197
SARTIN, J. L., CUMMINS, K. A., KEMPPAINEN, R. J., CARNES, R., MC CLARY, D.
G. u. J. C. WILLIAMS (1985):
Effect of propionate infusion on plasma glucagons, insulin and growth hormone
concentrations in lactating dairy cows.
Acta Andocrinol. (Copenh) 109(3), 348-354
SCHEMANN, M. (2000):
Enterisches Nervensystem und Innervation des Magen-Darm-Traktes.
In: VON ENGELHARDT, W. u. G. BREVES (Hrsg.)
Physiologie der Haustiere
Enke im Hippokrates Verlag, Stuttgart, S. 308-316
SCHLUMBOHM, C., SPORLEDER, H. P., BURTLER, H. u. J. HARMEYER (1997):
Effect of insulin on glucose and fat metabolism in ewes during various reproductive
states in normal and hypcalcaemia.
Dtsch. Tierärztl. Wochenschr. 104, 359-365
SEJERSTED, O. M. u. G. SJOOGAARD (2000):
Dynamics and consequences of potassium shifts in skeletal muscle and heart during
exercise.
Physiol. Rev. 80, 1411-81

160 Literaturverzeichnis
SEN, I., OK, M. u. A. COSKUN (2006):
The level of serum ionised calcium, aspartate aminotransferase, insulin, glucose,
betahydroxybutyrate concentrations and blood gas parameters in cows with left
displacement of abomasums.
Pol. J. Vet. Sci. 9(4), 227-232
SIELMAN, E. S., SWEENEY, R. W., WHITLOCK, R. H. u. R. Y. REAMS (1997):
Hypokalemia syndrome in dairy cows: 10 cases (1992-1996).
J. Am. Vet. Med. Assoc. 210(2), 240-243
SMITH, T. R., ELMENDORF, J. S., DAVID, T. S. u. J. TURISKY (1997):
Growth hormone-induced insulin resistance: role of the insulin receptor, IRS-1,
GLUT-1. and GLUT-4.
Am. J. Physiol. 272, 1071-1079
SOMLYO, A. P. u. A. V. SOMLYO (1994):
Signal transduction and regulation in smooth muscle.
Nature 372, 231-236
STAUFENBIEL, R., RISCHK, U., SCHUMACHER, B. u. W. BECKER (1992):
Beurteilung der Insulin- und Glukoseregulation bei der Milchkuh mittels
Tagesprofilen, dem einfachen und modifizierten Glukosetoleranztest.
Dtsch. Tierärztl. Wochenschr. 99, 69-75
STÖBER, M. u. H.-D. GRÜNDER (1990):
Kreislauf.
In: ROSENBERGER, G., DIRKSEN, G., GRÜNDER, H.-D. u. M. STÖBER (Hrsg.)
Die klinische Untersuchung des Rindes (3. Aufl.)
Verlag Paul Parey, Berlin und Hamburg, S. 194-242

Literaturverzeichnis 161
STÖBER, M. u. P. SARATSIS (1974):
Vergleichende Messungen am Rumpf von schwarzbunten Kühen mit und ohne
linksseitiger Labmagenverlagerung.
Dtsch. tierärztl. Wochenschr. 81, 564-565
STRYER, L. (1999):
Der molekulare Bauplan des Lebens.
In: Biochemie (4th edn.)
Spektrum Akademischer Verlag Heidelberg, Berlin, Oxford, S. 1-152
STRYER, L. (1999):
Membrankanäle und Pumpen.
In: Biochemie (4th edn.)
Spektrum Akademischer Verlag Heidelberg, Berlin, Oxford, S. 309-342
SUSTRONCK, B. (2000):
Studies on abomasal emptying in cattle.
In: Aetiologie, Pathogenese, Diagnostik, Prognose, Therapie und Prophylaxe der
Dislocatio abomasi.
Leipziger Universitätsverlag, S. 113-126
SUZUKI, H., KITO, Y., HASHITANI, H. u. E. NAKAMURA (2006):
Factors modifying the frequency of spontaneous activity in gastric muscle.
J. Physiol. 576.3, 667-674
SWEENEY, R. W. (1999):
Treatment of potassium balance disorders.
Vet. Clin. North Am. Food anim. Prac. 15(3), 609-617

162 Literaturverzeichnis
TAGUCHI, K. (1995):
Relationship between degree of dehydration and serum electrolytes and acid-base
status in cows with various abomasal disorders.
J. Vet. Med. Sci. 57(2), 257-260
TAYLOR, R (1991):
Review: Insulin action 1991.
Clin. Endocrinol. 34, 159-171
TELLE, P. P., PRESTON, R. L., KINTNER, L. D. u. W. H. PFANDER (1964):
Definition of ovine potassium requirement.
J. Anim. Sci. 23, 59-66
THUNEBERG, L. (1989):
Interstitial cells of Cajal.
In: SCHULTZ, S. G., WOOD, J. D. u. B. B. RAUNER (Hrsg.)
Handbook of physiology: The gastrointestinal system I (6th edn.)
Waverly Press, Baltimore, Maryland, USA, S. 349-386
TORNQUIST, H. E. u. J. AVRUCH (1988):
Relationship of site-specific beta subunit tyrosine autophosphorylation to insulin
activation of the insulin receptor (tyrosine) protein kinase activity.
J. Biol. Chem. 263(10), 4593-4601
TOURNIAIRE, J., BAJARD, L., HARFOUCH, M., REBATTU, B. u. D. GARREL
(1988):
Restoration of insulin sensitivity after correction of hypokalemia due to chronic
tubulopathy in a diabetic patient.
Diab. Metab. 14(6), 717-720

Literaturverzeichnis 163
TREACHER, R. J., REID, I. M. u. C. J. ROBERTS (1986):
Effect of body condition at calving on the health and performance of dairy cows.
Anim. Prod. 43, 1-6
TREMBLAY, R. R. (1990) :
Intravenous fluid therapy in calves.
Vet. Clin. North Am. Food Anim. Pract. 6(1), 77-101
TRENKLE, A. (1971):
Postprandial changes in insulin secretion rates in sheep.
J. Nutr. 101, 1099-1104
TRICARICO, D., PIERNO, S., MALLAMACI, R., BRIGIANI, G. S., CAPRIULO, R.,
SANTORO, G. u. D. C. CAMERINO (1998):
The biophysical and pharmacological characteristics of skeletal muscle ATP-sensive
K+ channels are modified in K+-depleted rat, an animal model of hypokalemic periodic
paralysis.
Mol. Pharm. 54, 197-206
VAN EPPS-FUNG, M., WILLIFORD, J., WELLS, A. u. R. W. HARDY (1997):
Fatty-acid induced insulin resistance adipocytes.
Endocrinol. 138, 4338-4345
VANHEEN, B. u. J. BREYNE (2004):
Endothelium-dependent hyperpolarization in small gastric arteries.
Cardiovasc. Res. 63, 331-337
VAN MEIRHAEGHE, H. (1988):
Insulin activity in cattle and development of insulin resistance in the postpartum
period.
Vlaams Diergeneeskundig Tijdschrift. 57(1), 1-10

164 Literaturverzeichnis
VAN MEIRHAEGHE, H., DEPREZ, P., VAN DEN HENDE, C. u. E. MUYLLE
(1988 a):
The influence of insulin on abomasal emptying in cattle.
J. Vet. Med. A. 35, 213-220
VAN MEIRHAEGHE, H., DEPREZ, P., VAN DEN HENDE, C. u. E. MUYLLE
(1988 b):
Plasma glucose clearance and insulin response in cows with abomasal
displacement.
J. Vet. Med. A. 35, 221-228
VAUPEL, P. (2000):
Funktionen des Magen-Darm-Trakts.
In: SCHMIDT, R. F., THEWS, G. u. F. LANG
Physiologie des Menschen
Springer-Verlag, Berlin, Heidelberg, New York, S. 806-848
VERNON, R. G. u. S. SASAKI (1991):
Control of responsiveness of tissues to hormones.
In: TSUDA, T., SASAKI, Y. U. R. KAWASHIMA (Hrsg.)
Physiological aspects of digestion and metabolism in ruminants.
Academie Press New York, Boston, London, S. 155-182
VETMEDLABOR (2007):
www.vetmedlabor.de/downloads/VetMedLabor_LVZ_02-2007.pdf
VLAMINCK, K., VAN MEIRHAEGHE, H., VAN DEN HENDE, C., OYAERT, W. u. E.
MUYLLE (1985):
Einfluß von Endotoxinen auf die Labmagenentleerung beim Rind.
Dtsch. Tierärztl. Wochenschr. 92, 392-395

Literaturverzeichnis 165
VOLLMERHAUS, B. u. H. ROOS (1999):
Speiseröhre, Magen, Darm, Darmanhangsdrüsen.
In: NICKEL, R., SCHUMMER, A. u. E. SEIFERLE
Hrsg.: FREWEIN, J., GASSE, H., LEISER, R., ROOS, H., THOME, H.,
VOLLMERHAUS, B. u. H. WAIBL
Lehrbuch der Anatomie der Haustiere, Band II, Eingeweide (8th edn.)
Parey Buchverlag Berlin, S. 103-214
VON ENGELHARDT, W., HAARMEYER, P. u. M. LECHNER-DOLL (2006):
Feed intake, forestomach fluid volume, dilution rate and mean retention of fluid in the
forestomach during water deprivation and rehydration in camels (Camelus sp.).
Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 143(4),504-507
WARD, G. M. (1966):
Potassium metabolism of domestic ruminants-a review.
J. Dairy Sci. 49(3), 268-276
WARD, J. L., SMITH, D. F., FUBINI, S. L. u. D. M. DEUEL-AROMANDO (1994):
Evaluation of abomasal outflow diversion as an experimental model of
hypochloremic, hypokalemic metabolic alkalosis in lactating cows.
Can. J. Vet. Res. 58, 13-19
WEEKES, T. E. C. (1991):
Hormonal control of glucose metabolism.
In: TSUDA, T., SASAKI, Y. U. R. KAWASHIMA (Hrsg.)
Physiological aspects of digestion and metabolism in ruminants.
Academie Press New York, Boston, London, S. 183-200

166 Literaturverzeichnis
WOOD, J. D. (1994):
Physiology of the enteric nervous system.
In: L. R. JOHNSON (Hrsg.)
Physiology of the gastrointestinal tract (3rd edn.)
Raven Press, New York, S. 423-482
YASUI, S., MAWATARI, K., KAWANO, T., MORIZUMI, R., HAMAMOTO, A.,
FURUKAWA, H., KOYAMA, K., NAKAMURA, A., HATTORI, A., NAKANO, M.,
HARADA, N., HOSAKA, T., TAKAHASHI, A., OSHITA, S. u. Y. NAKAYA (2007):
Insulin Activates ATP-Sensitive Potassium Channels via Phosphatidylinositol 3-
Kinase in Cultured Vascular Smooth Muscle Cells.
J. Vasc. Res. 45, 233-243
YOUN; J. H. u. A. A. MC DONOUGH (2009):
Recent advances in understanding integrative control of potassium homeostasis.
Annu. Rev. Physiol. 71, 2.1-2.21
ZADNIK, T. (2003):
A comparative study of the hemato-biochemical parameters between clinically
healthy cows and cows with displacement of the abomasums.
Acta Vet. 53(5-6), 297-309
ZAMMIT, V. A. (1996):
Review article: Role of insulin in hepatic fatty acid partitioning: emerging concepts.
J. Biochem. 314, 1-14
ZHAO, L., GU, L. Q., WANG, W. Q., NING, G. u. J. M. LIU (2008):
Extreme hyperglycaemia and hypokalaemia in a 96-year-old diabetic patient: case
report and discussion of management issues.
Br. J. Biomed. Sci. 65(1), 30-33

Literaturverzeichnis 167
ZEMKOVA, H., TEISINGER, J. U. F. VYSKOCIL (1982):
The comparison of vanadyl (IV) and insulin-induced hyperpolarization of the
mammalian muscle cell.
Biochem. Biophys. Acta. 720, 405-410
ZIERATH, J. R., LIVINGSTON, J. N., THORNE, A., BOLINDER, J., REYNISDOTTIR,
S., LORNQVIST, F. u. P. ARNER (1998):
Regional difference in insulin inhibition of non-esterified fatty acid release from
human adipocytes: relation to insulin receptor phosphorylation and intracellular
signalling through the insulin receptor substrate-1 pathway.
Diabetol. 41, 1343-1354
ZIERLER, K. L. (1959):
Hyperpolarization of muscle by insulin in a glucose-free environment.
Am. J. Physiol. 197, 524-526
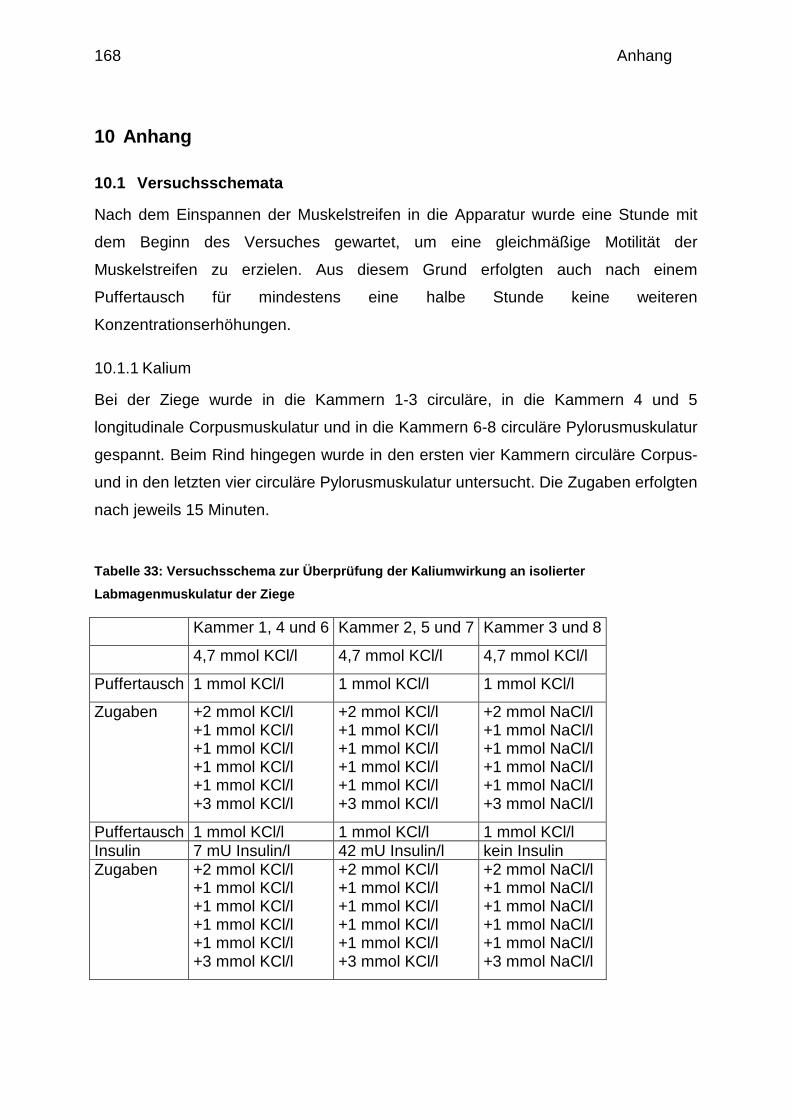
168 Anhang
10 Anhang
10.1 Versuchsschemata
Nach dem Einspannen der Muskelstreifen in die Apparatur wurde eine Stunde mit
dem Beginn des Versuches gewartet, um eine gleichmäßige Motilität der
Muskelstreifen zu erzielen. Aus diesem Grund erfolgten auch nach einem
Puffertausch für mindestens eine halbe Stunde keine weiteren
Konzentrationserhöhungen.
10.1.1 Kalium
Bei der Ziege wurde in die Kammern 1-3 circuläre, in die Kammern 4 und 5
longitudinale Corpusmuskulatur und in die Kammern 6-8 circuläre Pylorusmuskulatur
gespannt. Beim Rind hingegen wurde in den ersten vier Kammern circuläre Corpus-
und in den letzten vier circuläre Pylorusmuskulatur untersucht. Die Zugaben erfolgten
nach jeweils 15 Minuten.
Tabelle 33: Versuchsschema zur Überprüfung der Kali umwirkung an isolierter
Labmagenmuskulatur der Ziege
Kammer 1, 4 und 6 Kammer 2, 5 und 7 Kammer 3 und 8
4,7 mmol KCl/l 4,7 mmol KCl/l 4,7 mmol KCl/l
Puffertausch 1 mmol KCl/l 1 mmol KCl/l 1 mmol KCl/l
Zugaben +2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +3 mmol NaCl/l
Puffertausch 1 mmol KCl/l 1 mmol KCl/l 1 mmol KCl/l Insulin 7 mU Insulin/l 42 mU Insulin/l kein Insulin Zugaben +2 mmol KCl/l
+1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +3 mmol NaCl/l

Anhang 169
Tabelle 34: Versuchsschema zur Überprüfung der Kali umwirkung an isolierter
Labmagenmuskulatur des Rindes
Kammer 1 und 5 Kammer 2 und 6 Kammer 3 und 7 Kammer 4 und 8
4,7 mmol KCl/l 4,7 mmol KCl/l 4,7 mM KCl 4,7 mM KCl
Puffertausch 1 mmol KCl/l 1 mmol KCl/l 1 mmol KCl/l 1 mmol KCl/l
Zugaben
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +3 mmol NaCl/l
Puffertausch 1 mmol KCl/l 1 mmol KCl/l 1 mmol KCl/l 1 mmol KCl/l
Insulin 7 mU Insulin/l 42 mU Insulin/l kein Insulin kein Insulin
Zugaben +2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +1 mmol KCl/l +3 mmol KCl/l
+2 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +1 mmol NaCl/l +3 mmol NaCl/l
10.1.2 Insulin
Die Aufteilung der Präparate auf die zur Verfügung stehenden Kammern entspricht
der unter 10.1.1 aufgeführten. Beim Rind wurde nur der Versuch mit der Zeitkontrolle
durchgeführt. Die Zugaben erfolgten nach jeweils 30 Minuten.

170 Anhang
Tabelle 35: Versuchsschema zur Überprüfung der Insu linwirkung an isolierter
Labmagenmuskulatur
Ziege:
Rind:
Kammer 1, 2, 4, 6 und 7
Kammer 1, 3, 5 und 7
Kammer 3, 5 und 8
Kammer 2, 4, 6 und 8
5 mmol KCl/l (nicht beim Rind);
4,7 mmol KCl/l
3 mmol KCl/l (nicht beim Rind);
4,7 mmol KCl/l ohne Insulinzugaben
Zugaben + 7 mU Insulin/l + 7 mU Insulin/l
+ 14 mU Insulin/l + 14 mU Insulin/l
+ 21 mU Insulin/l + 21 mU Insulin/l
+ 38 mU Insulin/l + 38 mU Insulin/l
+ 40 mU Insulin/l + 40 mU Insulin/l
Nur Ziege! auf 10 mM KCl auf 10 mM KCl
Aufgrund der fehlenden Insulinwirkung an der circulären Pylorusmuskulatur des
Rindes wurde ein weiterer Versuch durchgeführt.
Tabelle 36: Versuchsschema zur Überprüfung der Insu linwirkung an isolierter
Pylorusmuskulatur des Rindes
Kammer 1 und 5 Kammer 2 und 6 Kammer 3 und 7 Kammer 4 und 8
Zeit 4,7 mmol KCl/l 4,7 mmol KCl/l 4,7 mmol KCl/l 4,7 mmol KCl/l
0 h
0,5-2,5 h kein Insulin 40 mU Insulin/l 120 mU Insulin/l 200 mU Insulin/l
10.1.3 Weiterführende Untersuchungen
An der circulären Corpusmuskulatur des Rindes wurden weitergehende
Untersuchungen einerseits mit den Kaliumkanalblockern Barium und Glybenclamid
und andererseits mit Ouabain, einem Hemmer der Na+/K+-ATPase, vorgenommen.

Anhang 171
Tabelle 37: Versuchsschema zur Überprüfung der Insu linwirkung in Anwesenheit von Barium;
Gesamtversuchsdauer: 150 min
Zeitpunkt
(min nach Versuchsbeginn)
Kammer
2 und 6
Kammer
3 und 7
Kammer
4 und 8
0 80 mU
Insulin/l
3 mmol Barium/l 3 mmol Barium/l
30 3 mmol Barium/l
60
90 80 mU Insulin/l
Tabelle 38: Versuchsschema zur Überprüfung der Insu linwirkung in Anwesenheit von
Glybenclamid; Gesamtversuchsdauer: 105 min
Zeitpunkt
(min nach Versuchsbeginn)
Kammer
2 und 6
Kammer
3 und 7
Kammer
4 und 8
0 80 mU
Insulin/l
0,05 mmol
Glybenclamid/l
0,05 mmol
Glybenclamid/l
15 0,05 mmol
Glybenclamid/l
30
60 80 mU Insulin/l
Tabelle 39: Versuchsschema zur Überprüfung der Insu linwirkung in Anwesenheit von Ouabain;
Gesamtversuchsdauer: 105 min
Zeitpunkt
(min nach
Versuchsbeginn)
Kammer
2 und 6
Kammer
3 und 7
Kammer
4 und 8
0 80 mU Insulin/l 0,001 mmol Ouab./l 0,001 mmol Ouab./l
15
30 0,001 mmol Ouab./l 80 mU Insulin/l

172 Anhang
10.2 Verlauf der Blutglukosekonzentration im In-vivo- Versuch
Die Effluxmessungen fanden unter normoglykämischen Bedingungen statt. Durch
Intravenöse Gaben von Glukose konnten die Blutkonzentrationen während der
Insulininfusionen weitestgehend konstant gehalten werden (Abbildung 33).
0
20
40
60
80
100
120
140
160
0 90 180 270 0 90 180 270 300 60 150 240 300
Zeit in min
[Glu
kose
] in
mg/
dl
Abbildung 33: Verlauf der Blutglukosekonzentratione n aller Versuchstiere an den drei
Versuchstagen
Die Pfeile markieren den Beginn der Insulininfusion .
10.3 Verwendete Substanzen
10.3.1 In-vitro-Versuch
Für die Pufferlösung:
NaCl: 117,0 mmol/l (Art. Nr. 1.06404.1000, Merck, Darmstadt)
KCl: 4,7 mmol/l (Art. Nr. 1.04936.0500, Merck, Darmstadt)
MgCl2: 1,2 mmol/l (Art. Nr. 1.05833.0250, Merck, Darmstadt)
NaH2PO4: 1,2 mmol/l (Art. Nr. S-5011, Sigma Aldrich, Steinheim)
CaCl2: 2,5 mmol/l (Art. Nr. 2381, Merck, Darmstadt)
NaHCO3: 25,0 mmol/l (Art. Nr. S-6014-1 KG, Sigma Aldrich, Steinheim)
Glukose: 11,0 mmol/l (Art. Nr. 1.08337.1000, Merck, Darmstadt)
Kontrolle Insulin + Kalium Insulin
Tag 2 Tag 3 Tag 1
Anhang 173
Als Zugaben:
Tabelle 40: Liste der für die in-vitro-Motilitätsme ssungen verwendeten Substanzen
Substanz Stammlösung Endkonzentration
im Organbad
Hersteller
NaCl 1 mol/l Aqua dest. 1/2/3/4/5/6/9 mmol/l Art. Nr. 1.06404.1000,
Merck, Darmstadt
KCl 1 mol/l Aqua dest. 1/3/4/5/6/7/10
mmol/l
Art. Nr. 1.04936.0500,
Merck, Darmstadt
Insulin 7/40 U/l Aqua dest. 7/21/40/80/120/200
mU/l
Art. Nr. I5500-250 MG,
Sigma-Aldrich, Taufkirchen
Ouabain 0,001 mol/l Aqua
dest.
0,001 mmol/l Art. Nr. 0-3125, Sigma, St.
Louis, USA
Glybenclamid 0,05 mol/l Aqua
dest.
0,05 mmol/l Art. Nr. 233-570-6, Sigma,
St. Louis, USA
Bariumchlorid-
Dihydrat
3 mol/ l Aqua dest. 3 mmol/l Art. Nr. 1719, Merck,
Darmstadt
10.3.2 In-vivo-Versuch
- Venenverweilkatheter (1,1 x 1,7 mm, Länge 45 cm, Cavafix Certo, Braun,
Melsungen) Heidelberger Verlängerung (140 cm, Art. Nr. 97583, WDT,
Hannover)
- Dreiwegehahn (Art. Nr. 96819, WDT, Hannover)
- Insulininfusionspumpe (InfusomatfmS, Braun, Melsingen)
- Glukoseinfusion (G40 % ad us. vet., Braun, Melsungen)
- physiologische Kochsalzlösung (NaCl 0,9 %, Braun, Tuttlingen)
- Insulinpulver (Insulin from bovine pancreas, I5500-250 MG; Sigma-Aldrich,
Taufheim)
- 143,88 g CrCl3 * 6H2O (Merck-Artikel-Nr.: 2487/250 g)

174 Anhang
- 201 g Na2-EDTA * 2H2O (Merck-Artikel-Nr.: 8421/1 kg)
- 5,88 g CaCl2 * 2H2O (Merck-Artikel-Nr.: 2381/250 g)
- 1 n NaOH (Merck-Artikel-Nr.: 6498)
- Tischzentrifuge (Labofuge 200, Heraeus Sepatech)
- Eppendorfgefäße (1 ml; Eppendorf, Hamburg)
- Blutzuckermessgerät (SensoCard, Art. Nr. 0197, WDT, Hannover)
- Messstreifen zur Blutzuckermessung
- Procain-Penicillin (30.000 I.E./kg; Art. Nr. 21762, WDT, Hannover)
- Zentrifuge (Sorvall RC-5C und Sorvall RC-5B, Du Pont, Willington)
- einen für bovines Insulin spezifischen ELISA (Mercodia Bovine Insulin ELISA,
Mercodia AB, Uppsala)
- 614 Na+/K+-Analyser (CIBA-CORNING)
- Atomabsorptionsspektrometer (UNICAM 969)

Danksagung 175
11 Danksagung
Frau PD Dr. Sabine Leonhard-Marek, Physiologisches Institut der Tierärztlichen
Hochschule Hannover, danke ich für die Überlassung des interessanten Themas, die
jederzeit gebotene freundliche Betreuung und Unterstützung sowie die kritische
Durchsicht meiner Dissertation.
Der Schaumann-Stiftung und der DFG danke ich für die großzügige finanzielle
Unterstützung.
Den wunderbaren Mitarbeiterinnen aus den verschiedenen Laboren (insbesondere
Marion Loh, Marion Burmester und Susanne Hoppe) danke ich für ihre Ideen und
ihre Hilfe bei anfallenden Reparaturen, beim Anrühren bunter Lösungen, beim
Finden verloren geglaubter Sachen und bei der Überwindung unlösbarer
Herausforderungen. Ihr hattet es wirklich nicht immer leicht.
Den Mitarbeitern der FAL Braunschweig (insbesondere Herrn Dr. Lebzien, Herrn
Badke und Herrn Mundstock) danke ich für ihre ausgesprochen freundliche und
hilfreiche Unterstützung bei meinen Versuchen.
Herrn Dr. Martin Beyerbach aus dem Institut für Biometrie, Epidemiologie und
Informationsverarbeitung der Tierärztlichen Hochschule Hannover danke ich für die
biometrische Beratung im Rahmen meiner Dissertation.
Herrn Prof. Dr. Kamphues und Frau Meyer danke ich für die Bestimmung der
Chromkonzentration und die Futtermittelanalyse.
Herrn Prof. Dr. Markus Pröpsting und seiner tollen Arbeitsgruppe danke ich für die
großartige Hilfe beim Messen der Insulinproben. Ihr Engagement hat mich nachhaltig
beeindruckt.

176 Danksagung
Herrn Prof. Dr. Jürgen Rehage und Marian Kusenda danke ich für ihre
Hilfsbereitschaft bei der Planung des In-vivo-Versuchs, der ohne ihre Kenntnisse und
Tipps sicher nicht so reibungslos abgelaufen wäre.
Herrn Dr. Martin Höltershinken und seinen Mitarbeitern danke ich für die Bestimmung
der Kaliumblutwerte und die Beratung hinsichtlich des Umgangs mit nicht
funktionierenden Kaliumelektroden.
Ein riesig großer Dank gilt natürlich auch den einzigartigen Mitstreitern:
Meike und Manu: Ihr habt mir in Leipzig das Leben gerettet
Margit: BS bleibt unvergessen, auch wenn Du noch immer auf versprochene
Ergebnisse wartest
Nina, Katha, Diana und die Schmuddelkinder: das lustigste Büro der Welt, das noch
immer keinen Anrufbeantworter für die Betriebsferien hat
Susi, Melanie und Jenny: die lustigsten Bürobesucher der Welt
1000 Dank auch an Sophie, Yasmin und Jan, die mir im ganzen letzten Jahr immer
wieder Unterschlupf, Verpflegung und Bespaßung boten – was hätte ich nur ohne
Euch gemacht?
Meinem wunderbaren Mann Nils und dem Rest unser kleinen Familie danke ich für
all das tägliche Chaos, ohne das mein Leben so langweilig wäre. Ich bin froh, dass
ich Euch habe.
Recommended

















