
Dottorato in MEDICINA VETERINARIA
Trasmissione della malattia di Aujezsky nel cinghiale (Sus scrofa) e possibili conseguenze nella demografia
e gestione della specie in provincia di Pisa
Candidato Relatori Dr. Paolo Varuzza Prof. Alessandro Poli Mat. 431758 Dr. Antonio Felicioli
AA. 2007 – 2010 SSD
VET03 BIO 10

Pagina | 2
Indice …………………………………………………………………………………………………………... 2
Introduzione …………………..…………………………………………………………………………….. 3
Malattia di Aujeszky …………………………………………………………………………………….. 9
Il Cinghiale (cenni di biologia) …………………..……………………………………………….. 32
Area di studio ………………………………………………………………………………………………. 46
Materiali e metodi ………………….………………..……………………………………………….. 51
Risultati ……………………………………………..……..……………………………………………….. 64
Discussione ………………………………………..……..……………………………………………….. 83
Conclusioni ………………………………………..……..……………………………………………….. 91
Bibliografia ………………………………………..……..……………………………………………….. 93
Ringraziamenti ………………………………..……..……………………………………………….. 115

Pagina | 3
CAPITOLO 1 INTRODUZIONE

Pagina | 4
1. INTRODUZIONE A partire dagli anni ‘80 il cinghiale ha assunto in Italia un interesse sempre
maggiore per motivazioni di tipo economico, sociali e biologiche. Dal punto
di vista venatorio la specie è probabilmente, insieme al fagiano, la più
importante nel panorama italiano: la caccia al cinghiale, attira infatti
sempre più, un grande numero di cacciatori, ed oltre a costituire una
profonda eredità di tipo culturale, rappresenta una notevole fonte di
introiti a beneficio dell’economia locale e nazionali. La nascita del “mito”
cinghiale è stata supportata dall’incredibile espansione territoriale e
numerica della specie che nel giro di trent’anni ha quintuplicato l’areale di
distribuzione italiano, favorita da una straordinaria adattabilità e da vari
fattori, tra i quali le numerose immissioni a scopo venatorio operate in
varie parti del Paese. Di pari passo sono cresciuti i danni e le
problematiche connesse che fanno del cinghiale una delle specie
selvatiche più difficili da gestire nel panorama italiano. Per avere un’ idea
delle proporzioni raggiunte dalla caccia al cinghiale basti pensare che nella
sola Toscana durante l’ultima stagione venatoria sono stati abbattuti oltre
75.000 capi.
Alla luce di queste considerazioni è sempre più opportuno pensare ad una
sua gestione a 360°, che contempli il maggior numero di informazioni sulla

Pagina | 5
specie non solo comportamentali, biologiche, o di abbondanza, ma anche
sanitarie.
In gran parte d’Italia poco si conosce sulla demografia della specie,
soggetta a forti oscillazioni periodiche, probabilmente riconducibili solo in
parte alle disponibilità alimentari stagionali. Alcune malattie infettive da
sempre fungono da regolatrici delle popolazioni selvatiche. Studi condotti
per la malattia di Aujezsky in provincia di Pisa hanno fatto emergere nel
cinghiale valori alti di siero prevalenza che necessitano di essere
approfonditi per potenziali conseguenze sugli allevamenti suinicoli della
provincia ed per la demografia del cinghiale. La virosi infatti, di notevole
interesse nell’industria suinicola, dove causa ingenti danni economici legati
alla perdita di capi, potrebbe avere un impatto notevole anche sulla
fecondità del cinghiale. Il virus in tale specie potrebbe essere un ulteriore
elemento di regolazione demografica andando ad incidere in particolare
sul numero di feti concepiti e non sulla mortalità in soggetti post
svezzamento come avviene per il suino. Indagini condotte recentemente
presso il Dipartimento di Patologia Animale, Profilassi ed Igiene degli
Alimenti dell’Università di Pisa hanno consentito di acquisire informazioni
sulla presenza dell’infezione nelle popolazioni di cinghiali viventi in
Maremma e nella provincia di Pisa, dimostrando una sieropositività

Pagina | 6
superiore al 50% .È stato inoltre possibile dimostrare la presenza di
antigene virale nelle tonsille e nel tessuto linfoide di tali soggetti,
testimoniando una diffusione attiva dell’infezione nelle popolazioni di
cinghiale in Toscana.
Tale dato non esclude inoltre una potenziale trasmissione dell’infezione ai
suini di allevamento, specialmente di razze rustiche ed allevate al brado
che con facilità entrano in contatto con i cinghiali.
A questo scopo sono stati effettuati prelievi a campione sui cinghiali
abbattuti nell’arco di poche ore dalla morte grazie alla collaborazione delle
squadre di caccia al cinghiale. Per ogni animale abbattuto è stata
compilata una scheda riepilogativa con peso pieno e vuoto, età, sesso,
data e luogo di abbattimento. Si è proceduto poi alla raccolta di campioni
di siero, tonsille, linfonodi e apparati riproduttivi, tamponi nasali, vaginali,
prepuziali e faringei. Per le femmine, quando possibile è stato prelevato
tutto l’apparato riproduttivo ed i feti e successivamente analizzati e
studiati in laboratorio. Le indagini in precedenza svolte hanno dimostrato
la validità di questa metodica di raccolta. Per raggiungere inoltre
l’obiettivo ci proponiamo di allestire una metodica di identificazione dei
virus della malattia di Aujeszky basata su indagini virologiche su tamponi
nasali e genitali.

Pagina | 7
Gli obiettivi di questa ricerca si possono ricondurre a due tipologie: lo
studio delle demografia e la ricerca della malattia di Aujeszky con le
possibili relazioni tra produttività e malattie.
Nel primo caso avremo:
lo studio della produttività del cinghiale in provincia di Pisa;
confrontare la sex ratio al momento della nascita e quella desunta
dai dati di abbattimento;
stabilire la correlazione tra numero di feti e peso ed età delle
femmine;
Fig. 1.1 Obiettivi di studio

Pagina | 8
Per lo studio sanitario:
verificare la prevalenza della malattia di Aujeszky;
stabilire eventuali correlazioni con le varie classi di sesso ed età;
indagare sulle modalità di trasmissione della malattia in special
modo tra femmine e feti;
correlare la fertilità con la presenza dell’infezione nella popolazione
studiata;
valutare se e quanto la malattia di Aujeszky possa influire sulla
dinamica di popolazione del cinghiale.

Pagina | 9
CAPITOLO 2 MALATTIA DI AUJESKY

Pagina | 10
2. MALATTIA DI AUJESZKY o PSEUDORABBIA
2.1 Eziologia
Nel 1902 A. Aujeszky descrisse una nuova malattia che causava disturbi
nervosi distinta dalla rabbia. Questa entità morbosa fu definita in Europa
“Morbo di Aujeszky” mentre nel nuovo continente fu preferito il nome di
“Pseudorabbia” per distinguerla appunto dalla malattia causata dal
Rabdovirus agente infettivo della rabbia.
Nel 1910, Schmeidhofer, dimostrò tramite prove di filtrazione su lisati
cellulari di animali infetti che la patologia, così come la rabbia, era causata
da un agente di natura virale. Il virus fu classificato nel 1934 da Sabin e
Wright come appartenete al genere Herpesvirus. Il Comitato
Internazionale per la Tassonomia dei Virus classificò successivamente l’
Aujeszky Disease Virus (ADV) come appartenente alla sottofamiglia
alphaherpesvirinae.
Sebbene il suino sia l’ospite naturale del virus, definito anche PRV
(Pseudorabies Virus) molti altri animali possono fungere da ospiti
accidentali e venire infettati da questo, è nel suino comunque che le
manifestazioni morbose possono esplicarsi con una varietà di quadri
clinici. Il decorso clinico è prevalentemente determinato da una parte

Pagina | 11
dall’età del soggetto infettato e dal suo stato immunologico , dall’altra
dalla virulenza del ceppo e dalla dose infettante.
La malattia si esplica in modo grave in popolazioni che non sono mai
venute in contatto con PRV ed in particolar modo nei suinetti dove la
mortalità si attesta sul 100% nelle prime 2 settimane di vita, abbassandosi
però al 50 % nella terza e quarta settimana.
Nei suinetti inoltre si osservano diversi sintomi neurologici al contrario
degli animali adulti che mostrano ipertermia, mancanza di appetito, tosse,
e dispnea marcata.
Negli adulti la mortalità è generalmente inferiore al 5%.
Nei verri e nelle scrofe si possono osservare anche disturbi della fertilità,
solitamente correlati a periorchiti e metriti che possono portare ad
infertilità e ad aborto gli animali.
2.2 Classificazione
PRV ha la tipica architettura dei virioni appartenenti al genere
Herpesvirus. Le particelle virali mostrano una conformazione pressoché
sferica con un diametro di 150-180 nm con un core centrale circondato da
tre strati.

Pagina | 12
Il core del virus contiene acido desossi-ribo-nucleico organizzato in doppio
filamento con una lunghezza approssimativa di 140 Kb. Il capside ha
natura proteica ed è organizzato in simmetria icosaedrica.
Il virus presenta anche un envelope che riveste l’intera particella formato
da un doppio strato fosfolipidico nel quale sono incluse le proiezioni
esterne del virus di natura glicoproteica.
2.3 Epidemiologia
La Pseudorabbia è una affezione principalmente del suino, serbatoio e
fonte più importante di infezione naturale per un ampio gruppo di ospiti
secondari che comprendono bovini, pecore, capre, cani, gatti e molte
specie animali selvatiche (Thedford e Wright, 1980). A motivo
dell’evoluzione e dell’esito della malattia, tutte le specie animali diverse
dai suidi sono considerate ‘fondi cechi epidemiologici, che però
potrebbero contenere il virus e diffonderlo se mangiati da carnivori o altri
animali selvatici.
L’infezione da PRV diffonde lentamente e prevalentemente per contagio
diretto naso-naso o per via oro-fecale, ma può anche avvenire attraverso
alimenti contaminati, acqua, ingestione di tessuti infetti o attraverso

Pagina | 13
calzature, abiti e veicoli contaminati. Trasmissione indiretta avviene
solitamente per inalazione di virus in aerosol. Il virus può rimanere
infettante fino a 7 ore nell’aria con umidità relativa >55%. Esperienze
condotte in Gran Bretagna, Danimarca e USA indicherebbero che il virus
può viaggiare via aerosol fino a 2 km di distanza. Oltre che per via
respiratoria, la trasmissione può realizzarsi anche attraverso il latte delle
scrofe infette, il seme del verro (anche se impiegato per una sola
inseminazione artificiale) e per via diaplacentare.
Una fonte insidiosa d’infezione sono i suini convalescenti, malati cronici ed
infetti in forma latente, che si possono conservare a lungo eliminatori.
L’infezione latente da herpesvirus può essere definita come fase di
persistenza del virus nell’ospite non associata a replicazione né a reazione
immunitaria (Thedford e Wright, 1980). In suini con infezione latente, a
seguito di condizioni stressanti quali il parto, sovraffollamento o trasporti,
possono riprendere la moltiplicazione e dell’eliminazione virale, non
sempre associate alla comparsa della sintomatologia clinica.
Altri studi hanno dimostrato che il virus può sopravvivere fino a 7 ore in
acque di pozzo non clorate, per due giorni negli effluenti di vasconi di
decantazione così come nell’erba verde, nel terreno, nelle feci e nelle
granaglie, per tre giorni nei liquidi di lavaggio nasale, sulla plastica e sul

Pagina | 14
mangime pellettato, per quattro giorni nella lettiera di paglia. Il virus è
munito di envelope e per questo è inattivato dall’ essiccamento, la luce
solare diretta, dalle alte temperature e a tutti i solventi che sciolgono i
grassi.
La malattia, prima degli anni ’60, risultava localizzata nell’Europa orientale,
ma dal 1989 ha avuto una distribuzione mondiale raggiungendo 43 Paesi
(http:WWW.Defra.gov.uk/animalh/diseases/notifiable/disease/aujeszkys.1
989). Ci sono diverse ragioni per spiegare l’aumentata incidenza della
malattia: lo sviluppo, negli ultimi decenni, di ceppi nuovi e più virulenti; i
miglioramenti nelle tecniche diagnostiche con l’identificazione dei casi di
pseudorabbia precedentemente diagnosticati come peste suina; e
cambiamenti nella gestione dell’allevamento suino, con confinamento di
molti animali in ambienti chiusi e dove i parti si succedono a ciclo
continuo.
Ospiti a fondo cieco, come il cane, gatto, ma anche ratti e altri roditori
selvatici sopravvivono solo 2-3 giorni dall’infezione e possono trasmettere
il virus tra le aziende se mangiati da altri suini. Il ruolo degli insetti come
potenziali vettori è oggetto di indagine. Uccelli non sembrano ricoprire
alcun ruolo nella trasmissione.

Pagina | 15
Il virus della pseudorabbia è stato isolato da cinghiali in Texas e Florida e la
positività sierologia è stata dimostrata in numerosi paesi Europei, Italia
compresa. La diffusione del virus dal cinghiale al bestiame domestico è
stata riportata in numerose occasioni in Florida e Texas, dove è stata
sospettata la contaminazione da parte di cinghiali di foraggio distribuito a
bovini sul terreno. Questa trasmissione dal cinghiale a bestiame domestico
è vista come un serio ostacolo ai programmi di eradicazione della malattia
(http://texnat.tamu.edu/symposia/feral/feral-12.htm 1993).
L’unico metodo di controllo della pseudorabbia nei selvatici è a tutt’oggi il
controllo della popolazione.
2.4 Patogenesi
2.4.1 Ingresso del virus nella cellula
PRV si replica all’interno delle cellule suscettibili alla sua infezione tramite
l’aggancio del virione alla membrana cellulare e la conseguente fusione
della membrana stessa con quella dell’envelope virale.
Ad oggi si è arrivati alla caratterizzazione di 7 glicoproteine dell’envelope
virale (gB, gC, gD, gE, gH, gI e gL) che giocano un ruolo fondamentale
nell’adesione e nell’adsorbimento del virus alla cellula.

Pagina | 16
Il primo contatto del virus con la cellula ospite è mediato dalla
glicoproteina gC che si lega alle molecole di eparan-solfato (HS) presenti
sulla superficie cellulare (Mettenleiter et al., 1990). L’interazione gC-HS
viene inoltre modulata per mezzo della glicoproteina gD la quale converte
rapidamente la bassa affinità di legame HS-dipendente in un legame più
stabile HS-indipendente (Karger and Mettenleiter, 1993). La stabilità di
legame della particella virale alla cellula infettata è fondamentale e
necessaria per la successiva serie di processi che portano alla definitiva
fusione dell’envelope con la membrana cellulare. Le quattro glicoproteine
gB, gD, gH e gL formanti un complesso, ed in combinazione singolarmente
con gH e gL mostrano di essere attive durante il processo di penetrazione
nella cellula (Rauh and Mettenleiter, 1991; Peeters et al., 1992; Peeters et
al., 1992; Klupp et al., 1992; Klupp et al., 1994) anche se la precisa
sequenza delle cascata che porta alla formazione del complesso proteico
non è stata ancora chiarita. Nonostante ciò, si è arrivati alla conclusione
che gB, gD e gH sono indispensabili per il processo di ingresso nella cellula,
mentre gC non risulta fondamentale per questo.

Pagina | 17
2.4.2 Replicazione virale
Le modalità per cui il capside si disintegra ed il materiale genomico
raggiunge il nucleo non sono ancora perfettamente comprese.
Una volta che il DNA di PRV viene a trovarsi all’interno del nucleo inizia la
sua trascrizione.
I primi geni ad essere trascritti vengono indicati con il nome di IE
(Immediate-Early). Fino ad oggi soltanto uno dei geni IE è stato
identificato. Questo gene, nominato IE180 (Cheung et al., 1990) codifica
per una proteina che a sua volta riconosce specifiche sequenze del
genoma virale e porta alla trascrizione di una altro gruppo di geni
denominati geni E (Early) (Workman et al., 1988).
Questi codificano per un gruppo di proteine che sono attive durante la
sintesi del DNA. L’ultimo gruppo di geni espressi, codifica principalmente
per glicoproteine strutturali. Una volta che sono stati prodotti tutti i
componenti strutturali, le nuove particelle virali vengono assemblate
tramite passaggio attraverso la membrana nucleare interna (Pol, 1990). I
neo-virioni passano quindi attraverso il reticolo endoplasmatico,
l’apparato del Golgi e quindi secreti all’esterno della cellula.
Il rilascio del virus all’esterno della cellula è influenzato dalle glicoproteine
gC, gE e gI (Schreurs et al., 1988 Zsak et al., 1989). La produzione del virus

Pagina | 18
è stata studiata in diverse linee cellulari dove la produzione di nuove
particelle virali appare evidente già 5 ore post-inoculazione (PI)
(Nauwynck, 1993).
I titoli del virus quindi crescono rapidamente 7 ore PI raggiungendo il
valore di 107.5 TCID50 per 106 cellule inoculate.
2.4.3 Diffusione diretta del virus da cellula a cellula
Un’importante ed alternativa via di infezione tramite il liquido
extracellulare è quella della trasmissione diretta del virus da cellula a
cellula dopo fusione delle membrane cellulari. Prima però che la fusione e
quindi il passaggio del virus possa avvenire, le cellule devono essere in
contatto l’una con l’altra. Sia l’adesione tra cellule che la fusione delle
membrane è mediata dalle glicoproteine dell’ envelope virale che
cominciano ad essere espresse sulla superficie delle cellule già 5 ore PI
raggiungendo un massimo 11 ore PI. Le principali glicoproteine coinvolte
nel processo di adesione cellulare sono gB, gC ed in grado minore gD
(Nauwynck, 1993). Una volta che le cellule sono in contatto può partire il
processo di fusione (Hanssens et al., 1993) in cui le glicoproteine coinvolte
sono principalmente gB e gH, in maniera minore gD e gE, mentra gC non

Pagina | 19
è coinvolta (Peeters et al., 1992a, Peeters et al., 1992b; Rauh and
Mettenleiter, 1991; Zsak et al., 1992).
2.4.4 Interazioni virus-ospite in assenza di immunità specifica
La virulenza di PRV è determinata dalla sua capacità di replicarsi ed
invadere le cellule del suino. Il sito primario di replicazione virale è senza
dubbio l’epitelio delle cavità nasali, le tonsille palatine, l’epitelio della
faringe e dei polmoni (Sabo et al., 1968; Sabo et al., 1969; Wittmann et al.,
1980; Miry and Pensaert, 1989; Pol, 1990; Kritas et al., 1994a; Kritas et
al.,1994b). Dopo la replicazione a livello delle cellule epiteliali dei suddetti
apparati, il virus diffonde nel connettivo sottostante in breve tempo. 24
ore PI, l’antigene virale può essere già ritrovato in gruppi di cellule
epiteliali, fibrociti e cellule nervose. 48 ore PI l’antigene virale viene
riscontrato in larghe placche che vanno incontro precocemente a necrosi e
che si estendono sopra gli epiteli interessati, il connettivo sottostante, le
cellule nervose e persino l’endotelio dei vasi sanguigni e linfatici della zona
sito di replicazione.

Pagina | 20
Il virus a questo punto può diffondersi dal sito primario di inoculazione a
tutti i livelli guadagnando il torrente ematico, la circolazione linfatica o
transitando passivamente lungo gli assoni dei nervi periferici.
La replicazione virale a livello dei siti secondari come linfonodi drenanti,
milza, bulbi olfattori, reni, ovaie ed utero inizia pressoché 48 ore PI.
Nonostante la presenza di immunità di tipo sia umorale che cellulo
mediata, il virus può essere isolato dalle tonsille dal 18° giorno PI (Sabo et
al., 1969) e dai secreti nasali dal 13° giorno PI (Pensaert et al., 1990
Vannier et al., 1990).
Il ruolo che giocano nel processo di replicazione le glicoproteine virali è
stato studiato ed interpretato tramite esperimenti di delezione sui geni
codificanti le suddette proteine creando dei ceppi mutati mancanti
appunto di una o delle altre proteine.
2.4.5 Interazione virus ospite in presenza di immunità specifica
Il grado in cui la replicazione virale può essere tenuta sotto controllo nel
suino dipende dalla situazione immunitaria dell’animale. Nonostante una
inibizione totale alla replicazione possa essere riscontrata a livello delle
cavità nasali e della faringe in suini reinfettati 1 o 2 mesi dopo la prima

Pagina | 21
inoculazione, l’antigene virale può essere ritrovato in un certo numero di
cellule alveolari nel polmone (Miry e Pensaert, 1989). Da queste dati è
stato ipotizzato che in prima istanza siano i macrofagi alveolari ad
infettarsi e quindi a trasmettere il virus all’epitelio alveolare e al
connettivo sottostante tramite contatto diretto tra cellula e cellula. In
suini immunizzati tramite vaccino la protezione immunitaria è meno
efficiente rispetto all’infezione naturale poiché l’antigene virale non solo
viene riscontrato a livello respiratorio profondo ma anche nell’epitelio
delle cavità nasali, tonsille e faringe (Miry e Pensaert, 1989; Wittmann et
al., 1980). Nonostante la presenza di immunità vaccinale, si può sempre
ritrovare una viremia cellulo-associata (Wittmann et al., 1980). Questo
permette a PRV di raggiungere organi profondi ed in caso di gravidanza
anche la placenta dove esplica azione lesiva che si può concludere in
interruzione della gestazione ed aborto ( Nauwynck e Pensaert. 1992).
Una volta infatti raggiunta la circolazione uterina, i leucociti veicolanti il
virus, aderiscono ai vasi sia tramite processi fisiologici sia tramite
l’espressione delle glicoproteine virali sulla superficie cellulare.
Dopo l’adesione dei leucociti ai vasi placentari, PRV è capace di
diffondere, sempre in associazione a queste cellule, attraverso l’ endotelio,
lo strato di fibrociti e quindi l’epitelio uterino(Hanssens et al., 1993).

Pagina | 22
Con questo meccanismo patogenetico definito a “cavallo di troia”, il
virus, anche in presenza di immunità vaccinale può essere veicolato per via
ematica e linfatica nell’organismo e quindi passare intatto, nelle scrofe,
attraverso la barriera placentare dove esplica la sua azione patogena sul
feto con conseguenze spesso fatali per questo.
2.5 Sintomatologia
Nella specie suina il quadro clinico dipende dallo stipite virale coinvolto,
dalla dose infettante e soprattutto dall’età dei colpiti. I suinetti sono molto
suscettibili, e le perdite possono raggiungere il 100% in quelli con meno di
7 giorni dove, in generale, sono osservati segni di malattia del sistema
nervoso centrale (tremori, pedalamento). La sintomatologia nervosa, pur
essendo occasionalmente segnalata in soggetti in fase di
magronaggio/ingrasso, è propria dei lattonzoli e dei suinetti appena
svezzati. Inizialmente, i suinetti mostrano astenia, anoressia e ipertermia
(41 °C). I sintomi neurologici possono già manifestarsi nelle prime 24 h con
tremori, scialorrea, atassia, nistagmo, opistotono, gravi crisi convulsive
epilettiformi. È anche possibile la paralisi del treno posteriore con
caratteristica posizione di cane seduto, o movimenti di rotazione su se

Pagina | 23
stessi con pedalage di tutti gli arti. Il prurito, costantemente presente nelle
altre specie sensibili, si manifesta molto raramente nel suino. Nella
maggior parte dei casi la morte sopraggiunge nell’arco di 24-36 ore.
Nei maialetti già svezzati i sintomi respiratori sono il principale problema
clinico, soprattutto per il sopraggiungere di complicazioni da germi di
irruzione secondaria. È stato riportato che il virus della pseudorabbia
inibisce la funzionalità dei macrofagi alveolari riducendo così la loro abilità
a spostarsi e distruggere i batteri. Nei suini magroni e nei grassi, la forma
respiratoria spesso poco pronunciata dal punto di vista clinico, talvolta
confusa con altre patologie, oppure comunemente definita ‘sindrome
influenzale, è in grado di condizionare altre infezioni respiratorie, con
perdite rilevanti in termini di incremento ponderale. È caratterizzata da
elevata morbilità (anche 100%) e, in assenza di complicazioni, bassa
mortalità (1-2%). La sintomatologia compare dopo un periodo di
incubazione di 3-6 giorni con ipertermia (41-42 °C), depressione, anoressia,
grave rinite con starnuti e secrezione nasale, che evolve in polmonite con
tosse secca e dispnea. Occasionali sono i casi di coinvolgimento del
sistema nervoso centrale. L’evoluzione della malattia in allevamento è
estremamente variabile con forme che diffondono rapidamente a tutti i

Pagina | 24
suini presenti indipendentemente dall’età e forme inapparenti,
diagnosticabili solo sierologicamente.
Nei riproduttori oltre alle manifestazioni respiratorie, compaiono disturbi
della sfera riproduttiva: nei verri è possibile osservare edema scrotale ed
azospermia totale o parziale; nella scrofa i sintomi variano a seconda del
momento dell’infezione, se si infetta nel primo terzo di gravidanza, può
riassorbire gli embrioni e ritornare in estro, mentre nel secondo e terzo
periodo della gestazione i sintomi vanno dall’aborto alla natimortalità.
2.6 Lesioni anatomo patologiche
Nei suini colpiti dalla forma nervosa, le lesioni macroscopiche sono spesso
assenti o poco significative, e si limitano alla iperemia delle meningi,
aumento del liquido cefalo-rachidiano, congestione della mucosa naso
faringea, tonsillite, necrosi puntiforme epatica e splenica. Sempre presenti,
anche in suini asintomatici, le lesioni microscopiche a carico del SNC. I
reperti istologici più frequenti sono la meningoencefalite non suppurativa
e la ganglioneurite, che possono coinvolgere tanto la sostanza grigia
quanto la bianca a seconda della via di ingresso del virus. È stata anche
osservata la presenza di manicotti perivascolari di monociti, necrosi
neuronale e ispessimento delle meningi per infiltrazione monocitica.

Pagina | 25
Tipica, oltre alla comparsa di focolai di necrosi cariorettica, la comparsa di
grossi inclusi nucleari.
Negli altri mammiferi, le sole lesioni macroscopicamente apprezzabili sono
quelle traumatiche indotte dal prurito; i reperti istologici non differiscono
da quelli che si riscontrano nel suino.
La forma respiratoria è caratterizzata dalla flogosi della mucosa delle
prime vie respiratorie e dalla comparsa di ampi focolai polmonari di
consolidazione, soprattutto a livello dei lobi craniali e del lobo cardiaco. Il
quadro istologico consiste in una bronchiolo-alveolite necrotizzante, con
essudazione alveolare fibrinosa ed emorragica. Nelle prime fasi
dell’infezione è anche possibile notare la presenza di tipici inclusi nucleari
all’interno delle cellule epiteliali. Nelle razze a cute chiara è frequente la
cheratocongiuntivite.
Nelle scrofe l’aborto è seguito da una leggera flogosi della mucosa uterina
che si presenta edematosa. I feti abortiti, se non mummificati, presentano
edema sottocutaneo, versamenti essudatizi nelle cavità splacniche, necrosi
puntiforme epatica e splenica, focolai necrotici polmonari e tonsillari.
Nei verri è possibile osservare lesioni degenerativo necrotiche a carico dei
tubuli seminiferi, della tunica albuginea del testicolo e la presenza di

Pagina | 26
spermatozoi anomali, che potrebbero anche essere una conseguenza
dell’ipertermia.
A livello epatico, PRV, come altri virus erpetici, è in grado di produrre
focolai necrotici (2-3 mm di diametro) sparsi su tutta la superficie. Queste
lesioni sono più tipicamente ritrovate in soggetti giovani (< 7 giorni). Nel
suino, ma anche nel bovino, nel cane e nel gatto, si possono inoltre
osservare fenomeni di miocardite non purulenta, di tipo interstiziale, con
focolai di necrosi. Altre lesioni possibili sono la gastrite catarrale acuta e
l’enterite emorragica.
Lesioni macroscopiche da virus della pseudorabbia sono spesso invisibili. Si
possono ritrovare rinite sierosa, tonsillite necrotica o emorragie ai
linfonodi polmonari e talvolta edema polmonare e lesioni polmonari
secondarie a batteri patogeni.
2.7 Diagnosi
La diagnosi clinica è relativamente semplice solo nei mammiferi diversi dal
suino, l’isolamento del virus consente una diagnosi nel caso di forme fatali
della malattia o forme cliniche dei maiali, mentre altre tecniche e test
sierologici sono richiesti per la diagnosi delle forme latenti.

Pagina | 27
Nella specie suina, il rilievo clinico deve necessariamente essere integrato
da una accurata indagine anamnestica, dall’esame necroscopico e da
specifici esami di laboratorio. L’esame istologico può essere molto utile,
ma solo le indagini di laboratorio, dirette e indirette, costituiscono
elementi di certezza.
Prove dirette cercano di mettere in evidenza il virus mediante isolamento
su substrati cellulari e/o dimostrazione dell’antigene presente nei
campioni. La natura dell’antigene virale può essere dimostrata con test
immunoenzimatici (ELISA, immunoperossidasi), prove di
immunofluorescenza diretta, siero neutralizzazione. Rientrano tra le prove
dirette anche quelle che, attraverso reazioni di amplificazione del genoma
(Polymerase Chain Reaction) o l’impiego di sonde a DNA (tecniche di
ibridazione in situ), si prefiggono di dimostrare la presenza del genoma
virale.
Cervello, milza, e polmoni sono gli organi di scelta per isolamento virale.
Tamponi nasali possono essere utilizzati per l’isolamento virale solamente
in caso di soggetti in fase di infezione acuta. Il test di immunoflurescenza si
può fare su tonsille o cervello.
Le prove indirette svelano la reazione immunitaria specifica dell’ospite nei
confronti dell’agente virale, tuttavia molti animali colpiti non vivono

Pagina | 28
abbastanza a lungo per produrre una marcata risposta anticorpale. Sono
quindi praticabili solo nella specie suina e vengono prevalentemente
utilizzate a fini epidemiologici, piuttosto che diagnostici. Ogni tecnica
sierologia usata dovrebbe essere sufficientemente sensibile per dare un
risultato positivo con il siero standard internazionale di referenza dell’OIE.
In caso di commercio internazionale, il test dovrebbe essere abbastanza
sensibile per reagire con il siero standard diluito ½.
Il test di sieroneutralizzazione è stato riconosciuto come metodo di
referenza per la sierologia, anche se, per motivi di diagnosi generale e per
comodità in caso di analisi su larga scala, è stato ampiamente rimpiazzato
dal test ELISA (http: www.oie.int/eng/normes 2000), che può tuttavia
avere scarsa specificità. Risultati falsi positivi sono riesaminati con un test
di sieroneutralizzazione. Anche la sieroagglutinazione in latex, nonostante
sia un test sensibile e rapido, ha poca specificità. Questo test permette di
ritrovare gli anticorpi specifici dopo 6-7 giorni dall’infezione, utilizzando l’
ELISA ne occorrono 7-8 e con la sieroneutralizzazione 8-10. Anticorpi
colostrali per la pseudorabbia possono essere presenti fino a 4 mesi di età.

Pagina | 29
2.8 Profilassi
2.8.1. Azione immuogena delle glicoproteine di PRV
Da quanto descritto fin ora risulta chiaro come le diverse glicoproteine
virali sono responsabili di importanti interazioni tra PRV e le cellule
dell’ospite. Pertanto non sorprende che l’immunità indotta da queste
specifiche glicoproteine è molto efficace nella protezione del suino, non
solo clinicamente ma anche virologicamente. L’immunità specifica verso le
glicoproteine dell’envelope consiste di due meccanismi: l’immunita’
umorale specifica e quella cellulare mediata da linfociti T citotossici. Per
quanto riguarda gli anticorpi, questi possono agire singolarmente (via
diretta) o mediare attività antivirali tramite l’attivazione del sistema
complemento o per mezzo dell’ interazione con cellule immunitarie come
i leucociti (via indiretta).
Gli effetti diretti vengono ottenuti tramite l’azione competitiva sui siti di
legame delle glicoproteine durante l’ingresso del virus, l’adesione cellulare
e la fusione delle cellule infettate. È stato inoltre dimostrato che alcuni
anticorpi sia di natura monoclonale che policlonale diretti contro gB, gC e
gD, bloccano PRV anche in assenza del sistema complemento (Wathen et
al ., 1985; Ben-Porat et al., 1986; Eloit et al., 1990; Marchioli et al., 1988;
Coe e Mengeling, 1990), che sieri monoclonali specifici per le glicoproteine

Pagina | 30
gB e gC , inibiscono l’adesione virale in cellule infettate (Hanssens et al.,
1993) ed infine che anticorpi monoclonali diretti verso gB, gD e gE,
riducono la larghezza delle placche di lisi in monostrati cellulari in coltura(
Nauwynck e Pensaert, 1995b). Gli effetti indiretti della reazione
immunitaria sono possibili non solo attraverso l’azione dei suddetti
anticorpi ma anche tramite l’azione di anticorpi diretti verso regioni
inattive delle glicoproteine dell’envelope. Inoltre il legame degli anticorpi
(IgG, IgM) può attivare il sistema complemento, causando un danno alla
membrana in cui sono ancorate le glicoproteine dell’envelope e questo
può a sua volta portare a lisi cellulare, liberazione del virus in ambiente
extracelluare con conseguente in attivazione di questo. Questo è il motivo
per cui anticorpi monoclonali non neutralizzanti e diretti verso gB, gC, gD e
gE , possono concorrere alla clearence virale in presenza delle frazioni del
sistema complemento (Wathen et al., 1985; Fuchs et al., 1990; Nakamura
et al., 1990). Gli anticorpi una volta legati alle frazioni delle glicoproteine
espresse sulla superficie delle cellule infette, possono causare la morte
cellulare anche tramite l’interazione con i recettori Fc espressi dai fagociti
(monociti, macrofagi e granulociti neutrofili). Nonostante sia provata
l’esistenza di anticorpi diretti verso altre proteine virali come gG o IEP180,
questi risultano immunologicamente di scarsa importanza. Il motivo di

Pagina | 31
questo può essere spiegato con il fatto che le proteine in questione
svolgono la loro attività e vengono espresse all’interno delle cellule infette
risultando quindi irraggiungibili dai corrispondenti anticorpi (Zuckerman
et al., 1990).

Pagina | 32
CAPITOLO 3 IL CINGHIALE

Pagina | 33
3. Il CINGHIALE (cenni di biologia della specie)
3.1 Tassonomia
Unico rappresentante selvatico in Europa della famiglia dei Suidae, il
cinghiale rappresenta probabilmente la specie più importante nel
panorama faunistico venatorio in Italia. Da esso discendono i maiali
domestici originatosi indipendentemente in Asia ed in Europa circa 9000
anni fa. Le specie sono interfeconde tra di loro e in diverse parti dell’areale
del cinghiale, così come in Italia, si trovano popolazioni pure, ibride o solo
ibride. Numerose sono le sottospecie di cinghiale conosciuto in Eurasia,
alcune delle quali ritenute per molto tempo delle vere e proprie specie a
sé stanti. L’inquadramento sistematico delle varie sottospecie di cinghiale
risente della possibilità di incrociarsi sia con le razze domestiche di maiale
sia con varietà di cinghiale adattato a diverse aree geografiche della sua
distribuzione. I profondi spostamenti operati negli ultimi 50 anni dall’uomo
non hanno certo agevolato la situazione creando incroci tra cinghiali
centro-europei o dell’est Europa.
Attualmente sono riconosciute 16 sottospecie di cinghiale, sei di queste
interessano l’Europa.

Pagina | 34
3.2 Distribuzione
In Cinghiale è una specie cosmopolita, diffusa ampiamente in tutto il
Paleartico, e in alcune regioni dell’emisfero australe dove è stato
introdotta. Nel continente Eurasiatico, se pure con sottospecie differenti,
questo suide è riuscito a colonizzare ogni tipo di ambiente dalla macchia
mediterranea fino alle zone inospitali dell’Asia centrale. A nord rimane a
cavallo tra il 50° e 60° parallelo, mentre a sud l’areale della specie tocca
l’Africa mediterranea, su pure scomparsa dalla Libia e dall’Egitto, e arriva
fino all’India e alcune isole indonesiane a cavallo dell’equatore. In pratica è
presente dalla Penisola Iberica fino ad alcune isole del Giappone, ad
eccezione della regione Caucasica, dove si è estinto, e delle territori
desertici della Cina e della Mongolia.
Fig. 3.2.1 Distribuzione in Eurasia del cinghiale.

Pagina | 35
Nel Nuovo Mondo fa la sua comparsa in Florida nel 1532, attualmente è
presente in 19 stati degli Stati Uniti, e in Canada. L’azione umana ha
portato questa specie in Nuova Guinea e in molte isole limitrofe, in Nuova
Zelanda, e in Australia dove fu introdotta nel insieme a maiali, poi
rinselvatichiti e incrociati con il cinghiale.
In Europa il cinghiale è assente del tutto in Islanda, Danimarca, Norvegia e
Irlanda. Sparito nel 1650 in Gran Bretagna sta ricomparendo con
pochissimi nuclei isolati, frutto di immissioni illegali o di fughe da aziende
di caccia o allevamenti nel sud del Paese.
In Europa le dimensioni della specie crescono spostandosi dalla Spagna e
dall’Italia verso l’Ucraina. Non è un caso che i cinghiali dell’Est Europa
possano raggiungere i 350 Kg di peso, mentre in Sardegna o Spagna
toccano di rado gli 80-90 Kg. I fattori climatici giocano infatti, un ruolo
importante nella morfologia della specie, una taglia maggiore favorisce la
conservazione del calore in inverno e fa si che si possa muovere meglio.
Oggigiorno in Italia il cinghiale è l’ungulato selvatico con la più ampia
distribuzione e abbondanza. La specie è presente lungo tutto l’arco
appenninico, fino a raggiungere le Alpi occidentali senza soluzioni di
continuità. Nel resto dell’arco alpino la distribuzione risulta frammentaria,
probabilmente frutto di immissioni, e molto localizzata in alcune vallate

Pagina | 36
prealpine, ma sempre al di sotto del limite della vegetazione arborea.
Risulta assente nella Pianura Padana e lungo la fascia costiera adriatica,
fino in Puglia ad eccezione della Foresta Umbra. In Sardegna la specie è
presente in maniera diffusa, mentre in Sicilia è ritornato grazie a
immissioni operate negli ultimi decenni, così come in molte isole minori:
l’Elba, l’Asinara e Caprera.
Nel nostro Paese il cinghiale risulta presente in 90 province su 103 con un
areale complessivo di circa 170.000 Km2 corrispondenti a poco più della
metà del territorio italiano.
In quasi tutte le province dove è presente la specie è cacciata, ben 87 su
90, in altre è sottoposto a controllo numerico. Un calcolo grossolano,
basato sul numero di capi abbattuti, stima la popolazione italiana tra
300.000 e 500.000 cinghiali.

Pagina | 37
Fig. 3.2.2 Distribuzione in Italia del cinghiale.

Pagina | 38
3.3 Habitat e home range
Il cinghiale può vivere in quasi tutti gli ambienti naturali: dalla costa fino
limite del bosco in montagna, arrivando a frequentare aree a ridosso delle
grandi città o aree paludose.
Le condizioni richieste dal cinghiale si possono riassumere con poche
parole: un minimo di copertura vegetazionale e cibo ed acqua.
Quest’ultima risulta essere un elemento fondamentale non solo per
dissetarsi ma per quotidiani bagni per liberarsi dai parassiti e con funzione
termoregolatrice. Il bosco rimane l’ambiente prediletto del cinghiale. Qui
la specie trova rifugio durante il giorno, ci
L’Home range annuale è estremamente variabile da 3 km2 fino a 150 km2.
L’attività giornaliera si concentra in aree di circa 65 ha con distanze medie
espIorate per notte di 7 km. maschi solenghi hanno home range più ampi
rispetto alle femmine. Differenze in termini di estensioni degli home
ranges possono essere dovute all’influenza negativa determinata dalla
caccia in battuta.

Pagina | 39
3.4 Alimentazione
Dal punto di vista alimentare il cinghiale viene considerato una specie con
regime alimentare onnivoro a forte consumo vegetale e con costumi
opportunisti. La sua dieta annuale è composta quasi totalmente da
vegetali, di norma dal 98 al 100%, la rimanente parte può essere
rappresentata da invertebrati e, non di rado anche da vertebrati che
entrano nella dieta in maniera più o meno significativa. La composizione
del suo ampio spettro alimentare è molto variabile in funzione dell’annata,
della stagione e dell’area geografica. In pratica in cinghiale opera delle
scelte di volta in volta approfittando della disponibilità dei vari alimenti,
prediligendo sempre cibi al alto contenuto energetico. Si può affermare
che non esista un’alimentazione tipo, piuttosto una o due categorie
alimentari principali che variano periodicamente e che vengono integrate
a seconda della disponibilità. Gli alimenti di origine vegetale comprendono
erbe e parti sotterranei di piante, funghi, ma soprattutto frutti in
particolare castagne, faggiole e ghiande. Le castagne vengono quasi
sbucciate, mentre le ghiande e faggiole sono consumate per intero. La
parte animale è rappresentata da insetti, molluschi, lombrichi, rettili,
anfibi, uova e nidiacei, pesci morti, roditori, insettivori e piccoli di altri

Pagina | 40
mammiferi e carogne. Raramente arriva al 10%, ma è quasi sempre
presente per garantire proteine all’animale.
3.5 Organizzazione sociale
L’unità sociale di base è costituita dal gruppo familiare con una femmina
anziana dominante.
Spesso le femmine sono imparentate tra di loro. All’interno del gruppo
vige un ordine gerarchico ben preciso, ma non invariabile. I gruppi possono
esser anche molto numerosi 40-50 esemplari che raggruppano più unità
familiari.
La composizione dei gruppi può essere classificata in otto tipologie
differenti e può variare durante il corso dell’anno.:
Maschi solitari;
Gruppo di maschi giovani;
Femmine solitarie;
Gruppo di femmine senza piccoli;
Femmine adulte con piccoli (fino ad un anno) o giovani;
Gruppo familiare esteso ai giovani ed alle femmine dell’anno
precedente con o senza piccoli;

Pagina | 41
Gruppo familiare aggiunto, femmine con o senza piccoli
Gruppo multifamiliare composto da varie femmine anche non
imparentate, con piccoli.
3.6 Biologia riproduttiva
La biologia riproduttiva del cinghiale testimonia la sua capacità di
adattamento. Tra le varie specie di ungulati il genere Sus è quello con il più
alto tasso riproduttivo. Inoltre al contrario degli altri ungulati selvatici
italiani che hanno un periodo riproduttivo prestabilito e che dura poche
settimane il cinghiale non presenta un ciclo riproduttivo stagionale, ma gli
accoppiamenti avvengono tutto l’anno con un picco tra dicembre e
febbraio che coinvolge fino ed oltre 80% degli animali. Questa situazione è
stata amplificata nel corso degli ultimi decenni dagli incroci con le razze
domestiche che hanno portato all’aumento del numero di piccoli per parto
ed hanno sfatato il periodo dei parti.
Le femmine possono raggiungere la maturità sessuale prima dell’anno di
età, intorno agli 8-10 mesi, più in generale a 18 mesi. Per i maschi pur
essendo biologicamente maturi alla stessa età delle femmine, raggiungono
la piena maturità sociale solo intorno ai 4-5 anni.

Pagina | 42
La maturità sessuale delle femmine dipende da numerosi fattori, primi fra
tutti il peso e la disponibilità alimentare. Una buona percentuale di
femmine sotto l’anno si riproduce se raggiunge il 30kg di peso ma la
maggior parte ha bisogno di avere un peso tra i 30 ed i 40 Kg. Anche il
periodo di nascita, l’abilmente in cui vivendo da piccole e la quantità di
latte ricevuto influenzano l’età del parto. Un femmina nata alla fine
dell’inverno in un ambiente ricco dal punto di vista alimentare potrà avere
il suo primo calore prima del compimento dell’anno, mentre se nata a fine
primavera difficilmente all’inizio del periodo riproduttivo avrà raggiunto un
peso adeguato ad affrontare la riproduzione.
L’abbondanza di cibo determina sulle femmine diversi effetti:
permette l’accumulo di grasso e di riserve permettendogli di
affrontare al meglio l’inverno;
accelera la crescita nelle prime fasi di vita anticipando i tempi in cui
le giovani femmine raggiungono il peso soglia;
aumenta il tasso di fecondità, portando le femmine ad avere 3 parti
in due anni;
aumenta la prolificità e diminuisce il tasso di mortalità dei piccoli.
È dimostrata una correlazione positiva tra peso delle femmine e
percentuale di femmine gravide così come del n° di feti, mentre a

Pagina | 43
sua volta il numero di piccoli per parto risente dello stato di salute
generale delle femmina.
Le femmine anziane si accoppiano prima e di conseguenza anticipano il
parto rispetto alle primipare ed alle giovani. Il ciclo estrale dura 21-23
giorni e si ripete in caso di mancato concepimento, ma può essere anche
assente ad alte densità o con scarsezza di cibo. L’estro vero e proprio dura
50-60 ore. Le prime ad entrare in calore sono le femmine dominanti che
fungono da stimolo per le altre femmine del gruppo. Il ciclo estrale si
arresta durante l’estate fino ai primi dell’autunno. Per animali di 2-3 anni
di età si riproducono 80-90% delle femmine.
Le femmine in calore comunicano il loro stato alle altre femmine ed ai
maschi marcando la corteccia degli alberi con un secreto lacrimale. I
maschi adulti eccitati dall’odore delle femmine in estro marcano il
territorio con la saliva ed urina per scoraggiare gli altri maschi ad
avvicinarsi. La sincronizzazione dell’estro permette alle femmine del
gruppo di partorire ed allattare i piccoli nello stesso periodo determinando
una maggiore coesione del gruppo ed allo stesso tempo migliora
l’allevamento e la sicurezza.

Pagina | 44
L’ovulazione, fecondazione e sopravvivenza dei piccolo sono strettamente
legati al peso ed alle condizioni di alimentazione delle femmine.
I maschi individuano la femmina in estro dall’odore e la seguono
attivamente. Non sono rari gli scontri ed i combattimenti con altri maschi
che possono essere spettacolari con cariche, spinte e morsi. Il vincitore
avrà diritto ad accoppiarsi con la femmina. Il maschio accarezza la
femmina con il muso sui fianchi e sul ventre invitandola all’accoppiamento
che dura diversi minuti. Successivamente il maschio si allontana alla
ricerca di altre femmine.
La gestazione dura in media 115 giorni con un intervallo che oscilla tra 100
a 140 giorni.
Il numero dei piccoli varia da 2-3 a 5-6 per parto ed è proporzionato al
peso ed all’età delle femmine, di conseguenza l’aumento di un
popolazione dipende in gran parte dalla struttura di popolazione delle
femmine e dal tipo di ambiente.
Il cinghiale si riproduce una volta l’anno. Tuttavia se la disponibilità delle
risorse alimentari è alta, specie nel periodo invernale, aumenta il tasso di
ovulazione delle femmine, e in annate particolari i si possono avere due
parti nello stesso anno solare nella tarda primavera ed all’inizio

Pagina | 45
dell’autunno. Di norma il 50% dei piccoli in Italia, nasce tra aprile e maggio
con un picco ad aprile, e due minore a febbraio ed ottobre.
Fig. 3.6.1 Ciclo riproduttivo annuale del cinghiale. In alto annate normali in
basso annate eccezionali

Pagina | 46
CAPITOLO 4 AREA di STUDIO

Pagina | 47
4. AREA di STUDIO La ricerca è stata condotta in tutto il territorio della provincia di Pisa
suddividendolo in aree campione in base alla distribuzione del cinghiale,
delle caratteristiche ambientali e delle aree di caccia.
La provincia di Pisa si estende su una superficie di 244.587 ettari, occupati
da boschi per circa il 37 %.
Fig. 4.1 Aree boscate in provincia di Pisa

Pagina | 48
La distribuzione del cinghiale interessa gran parte del territorio provinciale,
se pure con densità differenti. Manca infatti solo in alcune parti
pianeggianti e con assenza copertura boschiva nella parte settentrionale
della provincia. Mentre in aree con caratteristiche ambientali simili, ma
situate a ridosso di corpi di bosco è considerato presente in quanto
utilizzate spesso come zone di spostamento o di foraggiamento.
Fig. 4.2 Distribuzione del cinghiale in provincia di Pisa.

Pagina | 49
Purtroppo mancano informazioni precise sulle densità raggiunte dalla
specie nei vari comprensori. Le uniche informazioni in nostro possesso
sono i dati degli abbattimenti.
Le zone dove il prelievo del cinghiale è organizzato in squadre di caccia
sono concentrate in sei aree principali.
Fig. 4.3 Distribuzione del cinghiale in provincia di Pisa e,in rosso, le aree di caccia
in battuta

Pagina | 50
Le stesse aree sono state utilizzate come base del campionamento. Per
poter confrontare i dati degli abbattimenti è stata divisa un’area nella
parte meridionale della provincia a cavallo tra i due ambiti territoriali di
caccia. Pertanto il campionamento si è svolto in sei aree che
raggruppavano più zone di caccia di diverse squadre.
Fig. 4.4 Aree di campionamento

Pagina | 51
CAPITOLO 5 MATERIALI e METODI

Pagina | 52
5. MATERIALI e METODI
5.1 Organizzazione del campionamento
Il periodo di campionamento è coinciso con l’arco temporale di caccia al
cinghiale in battuta previsto dal Calendario venatorio provinciale dal 1°
novembre al 31 gennaio.
Le squadre hanno cacciato tre giorni a settimana (mercoledì, sabato e
domenica).
I cinghiali abbattuti sono poi macellati un stanze attrezzate o in rari casi,
direttamente sul posto. Prima dell’eviscerazione degli animali, sono stati
prelevate le seguenti tipologie di campioni:
sangue;
tampone nasale;
tampone vaginale/prepuziale;
tonsille palatine;
linfonodi inguinali superficiali;
feti ed utero.

Pagina | 53
Sangue- Il sangue è raccolto per gocciolamento durante le operazioni di
pulitura dell’animale o dalle cavità interne direttamente nelle provette di
vetro fornite. Da preferire i campioni non contaminati da altri liquidi
interni. Le provette, una volta tappate sono poste verticalmente in
frigorifero o all’aperto, se la temperatura risulta adeguata ( 4 - 10° C ),
facendo attenzione a non agitarle in qualsiasi momento per permettere al
sangue di sierare.
Tampone nasale- Tamponi sterili da passare nelle narici degli animali.
Preferibilmente in narici non imbrattate per la fuoriuscita di sangue. I
tamponi sono conservati nell’apposito porta tamponi con soluzione
fisiologica sterile, antibiotata e congelati in breve tempo in posizione
verticale in modo che il cotone rimanga immerso.
Tampone vaginale/prepuziale- Tamponi sterili da passare nella vulva fino
ad arrivare alla lunghezza totale del tampone o nel prepuzio cercando di
raccogliere materiale di desquamazione. I tamponi sono conservati
nell’apposito porta tamponi con soluzione fisiologica sterile, antibiotata e
congelati in breve tempo in posizione verticale in modo che il cotone
rimanga immerso.

Pagina | 54
Tonsille palatine- Il prelievo
viene effettuato tagliando con il
coltello od il bisturi tra le ossa
mandibolari e ribaltando la
lingua inferiormente in modo
da rendere visibile la faringe. Gli
organi si trovano alla base della
faringe e sono ben localizzabili Fig. 5.1.1 Tonsille
come due placche cribrate di circa 2x3 cm
o lievemente più piccole nei soggetti più giovani.
Linfonodi inguinali superficiali-
Il prelievo di questi organi viene
effettuato dopo lo scuoiamento
dell’animale. I linfonodi si
trovano a livello dell’inguine
lateralmente al pene nel
maschio ed immersi nel grasso
inguinale contiguo al tessuto Fig. 5.1.2 Linfonodi inguinali superficiali

Pagina | 55
mammario nella femmina. In alcuni animali questi risultano di dimensioni
notevoli (3-4 cmx2) in altri sono più piccoli e nascosti dal tessuto adiposo.
Al taglio mostrano una struttura caratteristica (diversa dai tessuti
circostanti e dal grasso), ben scapsulabili e di colorito tendente al grigio.
Feti e utero- Se la femmina risulta gravida è preferibile asportare
direttamente tutto l’utero lasciando i feti all’interno. Nell’eventualità che
la femmina non sia gravida o comunque che la gravidanza non sia
valutabile è comunque preferibile asportare direttamente l’utero senza
farne fuoriuscire il contenuto.
Fig. 5.1.3 Utero con feti

Pagina | 56
Ad ogni cinghiale è stato attribuito un codice univoco per località di
prelievo, e compilata una scheda riepilogativa dei dati del capo abbattuto
e sui campioni prelevati.
Per le modalità di caccia e di eviscerazione dei capi abbattuti non è stato
possibile avere le cinque tipologie di campioni per tutti i cinghiali
esaminati. Ad esempio è abitudine consolidata l’asportazione
immediatamente dopo l’abbattimento dell’apparato riproduttivo dei
maschi. Altrettanto spesso gli animali sono stati portati nel centro di
macellazione dissanguati .
La distribuzione del campionamento tra le varie aree di studio è stata
effettuata basandosi sugli abbattimenti delle stagione venatoria
precedente (2007/2008) considerando un 5% sul totale per area.
L’età degli animali abbattuti è stata stimata attraverso la valutazione del
grado di eruzione o di consumo dei denti. Gli animali campionati sono stati
divisi in tre classi di età: 0-1 anni, 1-2 anni, > 2 anni.

Pagina | 57
Squadra di caccia_____________________ ATC ________
Comune di
__________________________ Distretto___________
Resp. squadra Sig. _________________________Tel.____________
Resp. prelievo campioni Sig._____________________Tel. _________
Dati Abbattimenti Campioni prelevati (indicare sì o no)
Dat
a
CO
DIC
E
Sess
o
Età
Pe
so
San
gue
(2 p
rove
tte)
Ton
sille
Tam
po
ne
nas
ale
Tam
po
ne
pre
pu
zial
e/va
g.
linfo
no
do
ingu
inal
e
Ap
par
ato
rip
rod
utt
ivo
Ecto
par
assi
ti (
Zecc
he
ecc.
)
Alt
ro
Per informazioni si prega contattare il Sig.____________________________
Fig. 5.1.4 Scheda riepilogativa dei campioni prelevati

Pagina | 58
Fig. 5.1.5 Scheda riepilogativa di valutazione dell’età dei cinghiali. (Jagd und Fischerei modif.)

Pagina | 59
5.2 Feti
L’età dei feti campionati è stata valutata mediante il lavoro di Henry
(1968).
Giorni di età Morfologia del feto
30 appendici presenti, bocca aperta, occhi apparenti;
33 dita presenti, leggero sviluppo dell’orecchio che presenta l’apertura;
40 narici aperte, orecchio ancora piccolo e sottosviluppato, perdita delle sembianze di embrione;
50 zoccoli presenti, palpebre presenti, capezzoli visibili, orecchio molto piccolo in proporzione al corpo, ma tutte le altre strutture esterne si presentano nella stessa proporzione della nascita, scomparsa del sacco embrionale;
60 Orecchie proporzionate al corpo, zoccoli lunghi e orientati verso il ventre, follicoli piliferi visibili intono alle narici, sopracciglia e mento;
70 peli sul muso e sopracciglia lunghi 1-2 mm e di colore bianco, pigmentazione scura intorno al muso e alle zampe;
80 presenza di denti, peli sul mento, muso, sopracciglia, e sulle guance lunghi 3 mm o anche meno e di colore scuro, peli sull’estremità della coda, orecchie e linea mediana dorsale pigmentata di scuro;
90 peli sul mento, muso, sopracciglia e sulle guance lunghi 6-10 mm, follicoli piliferi evidenti sull’intero corpo, linea mediana dorsale pesantemente pigmentata , denti chiaramente visibili;
100 peli scarsi su tutto il corpo, striature longitudinali presenti sul corpo;
110 pelo sull’intero corpo, striature ben evidenti.

Pagina | 60
5.3 Analisi Sierologica
È stata ricercata la presenza di anticorpi specifici anti-PRV utilizzando un
test commerciale ELISA (CEDITEST PRV-gB, Cedi- Diagnostic B.V, Lelystad,
Olanda), seguendo le istruzioni della casa. In ogni test nelle piastre veniva
distribuito 100 μl di siero di riferimento PRV-1 nei pozzetti A1 e B1; 100 μl
di siero di riferimento PRV-2 nei pozzetti C1 e D1; 100 μl di siero di
riferimento PRV-3 nei pozzetti E1 e F1; e, nei pozzetti rimanenti i sieri da
testare. Dopo aver agitato delicatamente, le piastre sono state incubate a
37°C per 1 ora. Dopo aver effettuato 6 lavaggi con 200 μl del buffer di
lavaggio a pozzetto, nelle piastre sono stati distribuiti 100 μl a pozzetto del
coniugato diluito 1:30 con buffer di diluizione. Le piastre sono state
nuovamente agitate ed incubate per un ora a 37°C. Dopo altri 6 lavaggi
come riportato precedentemente, in ogni pozzetto sono stati distribuito
100 μl di cromogeno. Le piastre sono state incubate per 20’ a temperatura
ambiente e successivamente la reazione è stata arrestata con 100 μl a
pozzetto di stop solution. L’ intensità della reazione è stata letta mediante
uno spettrofotometro (Tirtek Multiskan Plus MK2, Made in Finland by
Eflab a Joint Venture Company Lab System and Flow Laboratories),
misurando la densità ottica dei pozzetti a 450 nm entro 15' e non oltre 60'.
Le densità ottenute sono state calcolate utilizzando la seguente formula:
PI= 100- [(OD450 test serum/OD450 maximun) x 100]

Pagina | 61
Il test è stato considerato valido quando: il valore massimo di densità
ottica (OD) non superava 1000; il valore della percentuale di inibizione del
siero di riferimento 1 era >50%; il valore della percentuale di inibizione del
siero di riferimento 2 era <50% ; quando questi criteri non erano rispettati
il test è stato considerato non valido. I campioni con una percentuale di
inibizione (PI) < 50% sono stati considerati negativi, mentre con la PI >
50%.
Per confermare i risultati della sierologia e per confrontare la presenza del
virus tra apparato nasale e genitale sono state effettuate analisi dei
tamponi attraverso nested-Pcr. Come controllo positivo è stato utilizzato
del tessuto nervoso di cane naturalmente infetto con herpesvirus suino e
confermato tramite IIC. Il bersaglio della nested-Pcr e delle successive real-
time Pcrs è stato il gene codificante per la glicoproteina virale di superficie
Gb ed un prodotto di 334bp e 196bp è stato ottenuto dal primo e dal
secondo passaggio rispettivamente.

Pagina | 62
5.4. Analisi virologica
Ulteriori analisi virologiche sono state effettuate per determinare la
localizzazione del virus nell'apparato nasale e/o genitale mediate tecnica
di SYBR Green PCR.
Il protocollo di PCR ha utilizzato la coppia di primer
Fwd2: 5’ - ACGGCACGGGCGTGATC-3’
Rev 2: 5’- GGTTCAGGGTACCCCGC-3
descritti da Ruiz-Fons et. al. 2007, che generano un prodotto di
amplificazione di 196bp. La specificità delle reazioni è stata valutata
tramite melt curve analysis.
Tutte le reazioni sono state effettuate in duplicato in un volume totale di
25μl utilizzando 5μl di templato costituito dal surnatante ottenuto da una
soluzione fisiologica sterile nella quale erano stati immersi i tamponi nasali
o genitali.
I campioni non idonei per eccessiva presenza di muco o sangue sono stati
scartati e per i casi dubbi le analisi sono state ripetute una seconda volta e

Pagina | 63
i prodotti di amplificazioni sono stati corsi su gel di agarosio come ulteriore
verifica della specificità del prodotto di amplificazione.
Come controllo positivo è stato utilizzato del tessuto nervoso di cane
naturalmente infetto con herpesvirus suino e confermato tramite IIC.
Controlli negativi sono stati effettuati utilizzando come templato 5
microlitri di soluzione fisiologica.
Le reazioni sono state effettuate utilizzando il ciclizzatore Rotorgene 6000
(Corbett research).
5.5 Produttività e dinamica di popolazione Nelle femmine che durante il campionamento erano palesemente gravide
è stato prelevato l’utero ed i feti e subito congelati a -40° C.
In una seconda fase i feti sono stati singolarmente pesati, misurati, sessati
e di ognuno è stata stimata l’età in giorni.
5.6. Statistica Le differenze tra i gruppi sono state verificate con il test del Chi quadro e
con il test di student.

Pagina | 64
CAPITOLO 6 RISULTATI

Pagina | 65
6. RISULTATI
6.1 Abbattimenti di cinghiale
Gli abbattimenti di cinghiale in provincia di Pisa sono soggetti ad
oscillazioni tipiche per questa specie.
Nella stagione venatoria 2009/2010 si è assistito ad un calo considerevole
spiegabile dagli abbattimenti in regime di controllo operati al di fuori del
periodo di caccia tradizionale.
Fig. 6.1.1 Andamento dei prelievi in battuta dei cinghiali in provincia di Pisa
Il campionamento è stato programmato per area sulla base dei capi
abbattuti nella stagione venatoria 2007/08.
0
1000
2000
3000
4000
5000
6000
7000
n°
cap
i pre
leva
ti
Stagione venatoria

Pagina | 66
6.2 Campionamento
Durante la stagione venatoria 2008/2009 sono stati campionati 281
cinghiali grazie alla collaborazione di 22 squadre di caccia al cinghiale
operanti in provincia di Pisa.
Un totale di 13 capi in più rispetto ai 268 previsti basandoci sul 5% del
prelievo della stagione venatoria 2007/08. La percentuale di
campionamento per la stagione venatoria 2008/2009, quindi realizzata, è
stata del 4,63%, a fronte di un aumento del prelievo di 712 capi.
Fig. 6.2.1 A sinistra n° di cinghiali abbattuti per area nella stagione
venatoria 2007/08 e a destra la ripartizione del campionamento

Pagina | 67
La distribuzione del campionamento in tutte le aree è stata superiore alla
soglia del 100% del previsto, ad eccezione di due dove si è campionata una
percentuale rispettivamente del 95 e del 96%.
Fig. 6.2.2 Percentuale di campionamento realizzato per area rispetto al
previsto
0,00
20,00
40,00
60,00
80,00
100,00
120,00
140,00
1 2 3 4 5 6
Per
cen
tual
e
Zone di campionamento
campionamento

Pagina | 68
Tra le tipologie di campione il tampone nasale è risultato quello più
abbondante (n. 260), mentre il tampone genitale (prepuziale o vaginale) è
stato quello meno campionato (n.139).
Zona sangue tonsille t. nasale t. gen. linfondi
1 22 20 22 22 20
2 10 19 19 12 18
3 51 46 49 30 21
4 38 45 46 32 46
5 33 73 75 35 36
6 43 33 49 8 23
totale 197 236 260 139 164
Tab. 6.2.1 Tipologie di campioni per area di campionamento
I motivi di tale differenza sono spiegati dal fatto che spesso si disponeva
solo della testa dell’animale abbattuto, quindi si potevano prelevare le
tonsille e si effettuava il tampone nasale. In altri casi il cinghiale era stato
eviscerato nel bosco e non era possibile effettuare il tampone
sull’apparato riproduttivo.
I campioni completi sono stati 80.
Per quanto riguarda i sessi sono stati campionati 114 maschi, 138 femmine
e 29 cinghiali di cui non è stato possibile determinarne il sesso. La sex ratio

Pagina | 69
è stata di 1:1,21- La ripartizione dei sessi campionati nelle varie zone è
riportata nella tabella successiva.
Zona Maschi Femmine Indeterminati
1 8 14 0
2 6 10 3
3 12 43 14
4 23 23 0
5 33 30 12
6 32 18 0
totale 114 138 29
Tab. 6.2.2 Campionamento per sesso e aree di studio
Il campionamento delle tre classi di età ha visto la classe giovanile meno
rappresentata con un numero di animali di circa la metà rispetto agli adulti
o alla classe dei piccoli. Di 28 cinghiali non è stato possibile stimare l’età, e
di tre non abbiamo l’età ed il sesso. Come era facile prevedere la maggior
parte degli animali indeterminati per sesso sono animali piccoli di solito
sotto i 10 Kg di peso, mentre per gli altri casi si è trattato di animali già
macellati, dei quali abbiamo avuto a disposizione solo la testa.

Pagina | 70
Età Maschi Femmine Indeterminati totale
0 - 1 40 37 21 99
1 - 2 11 33 3 47
> 2 57 49 2 108
Ind. 6 19 3 28
Tab. 6.2.3 Campionamento per sesso ed età ripartito nelle aree di studio
6.3 Feti
Le femmine risultate gravide in maniera evidente sono state 24 (17,39%)
suddivise in 5 aree. I feti identificati sono stati 151 con una media per
femmina di 6,29. Nelle diverse aree la media varia da un minimo di 5,5 feti
per femmina (area 2) ad un massimo di 7,5 (area 5). Il numero medio di
feti nelle femmine infette è risultato di 4,75.
Area N° femmine con feti
percentuale F feti /tot F
N° feti
Media feti/ femmine
1 12 85,71 78 6,5
2 2 20,00 15 7,5
3 4 9,30 22 5,5
4 2 8,70 13 6,5
5 4 13,33 23 5,7
totale 24 20,00 151 6,29
Tab. 6.3.1 Campionamento per sesso ed età ripartito nelle aree di studio

Pagina | 71
Di 117 feti è stato possibile stabilire il sesso, di questi 72 erano maschi e 56
femmine. Il rapporto sessi risultato di 1:0,77.
Il peso medio dei feti è stato di 317,05 grammi, non risultano differenze
significative tra i due sessi (t student P = N.S.).
Non sono emerse differenze significative tra il peso delle femmine ed il n°
di feti.
6.4 Risultati analisi sierologica
Per 127 campioni raccolti durante il campionamento è stato possibile
effettuare l’analisi sierologica. La loro distribuzione nelle 7 aree di indagine
è riportata nella tabella sottostante.
Area N° campioni
totali N° campioni
sierologia % campioni con
analisi sierologica
1 22 16 72,73
2 19 7 36,84
3 69 44 63,77
4 46 18 39,13
5 75 14 18,67
6 50 28 56,00
totale 281 127 45,20
Tab. 6.4.1 Ripartizione per area dei campioni utilizzati per l’analisi
sierologica

Pagina | 72
Tab. 6.4.2 Positività alla malattia dall’analisi sierologica per area (valori
assoluti)
La positività alla malattia è risultata variabile nelle diverse aree dal 50 al
85,71%
Tab. 6.4.3 Positività alla malattia dall’analisi sierologica per area espressa
in percentuale
Area Positivi (n°) Negativi (n°)
1 8 8
2 1 6
3 12 32
4 5 13
5 4 10
6 4 24
totale 34 93
Area Positivi (%) Negativi (%)
1 50,00 50,00
2 14,29 85,71
3 27,27 72,73
4 27,78 72,22
5 28,57 71,43
6 17,39 82,61
totale 26,77 73,23
Pagina | 73
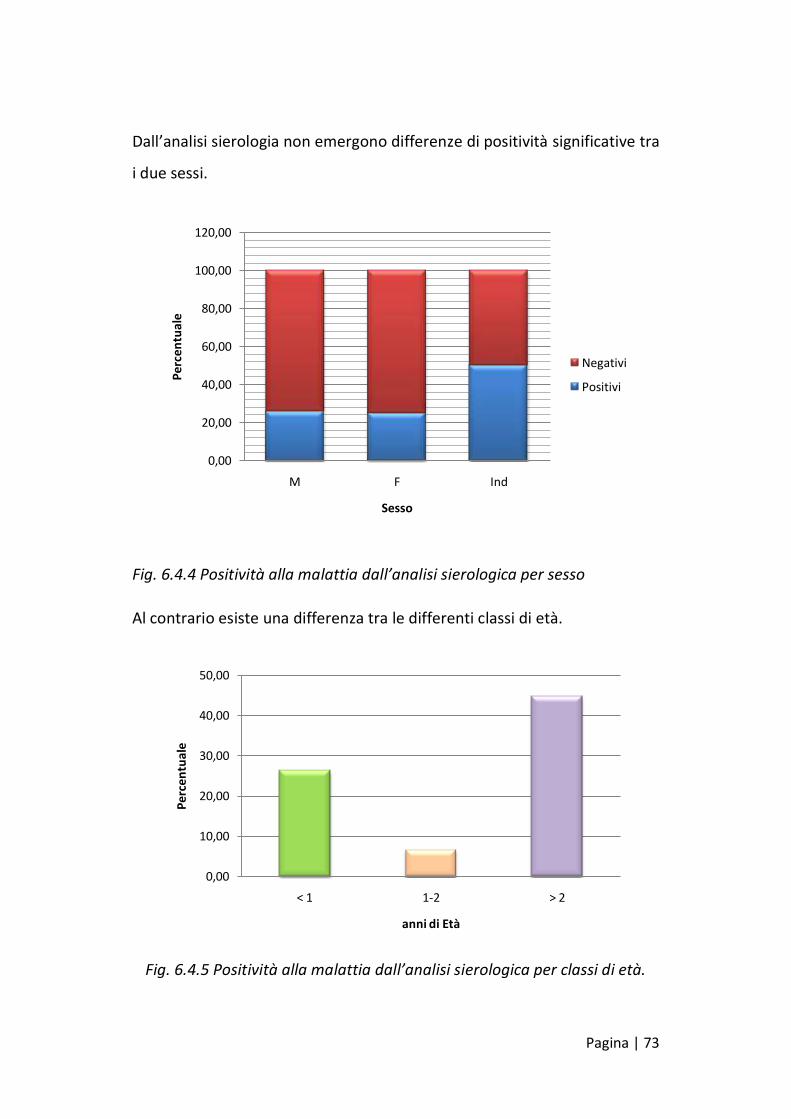
Dall’analisi sierologia non emergono differenze di positività significative tra
i due sessi.
Fig. 6.4.4 Positività alla malattia dall’analisi sierologica per sesso
Al contrario esiste una differenza tra le differenti classi di età.
Fig. 6.4.5 Positività alla malattia dall’analisi sierologica per classi di età.
0,00
20,00
40,00
60,00
80,00
100,00
120,00
M F Ind
Pe
rce
ntu
ale
Sesso
Negativi
Positivi
0,00
10,00
20,00
30,00
40,00
50,00
< 1 1-2 > 2
Pe
rcen
tual
e
anni di Età

Pagina | 74
Fig. 6.4.6 Positività alla malattia dall’analisi sierologica per classi di sesso e
ed età.
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
M F
Per
cen
tual
e
Sesso
< 1 1-2 > 2

Pagina | 75
6.5 Risultati analisi virologiche
Sono stati analizzati mediante Real PCR 399 tamponi nasali e genitali
rispettivamente 260 tamponi nasali e 139 genitali (dei quali 48 maschi, 88
femmine e 3 cinghiali di sesso indeterminato).
Fig. 6.5.1. Ripartizione dei campioni per tipologia e sesso
I tamponi nasali sono stati prelevati quasi in misura uguale (111 maschi,
127 femmine), al contrario i tamponi vaginali sono numericamente quasi il
doppio rispetto ai tamponi prepuziali rispettivamente: 90 e 49.
Gli animali di sesso indeterminato sono stati 22.
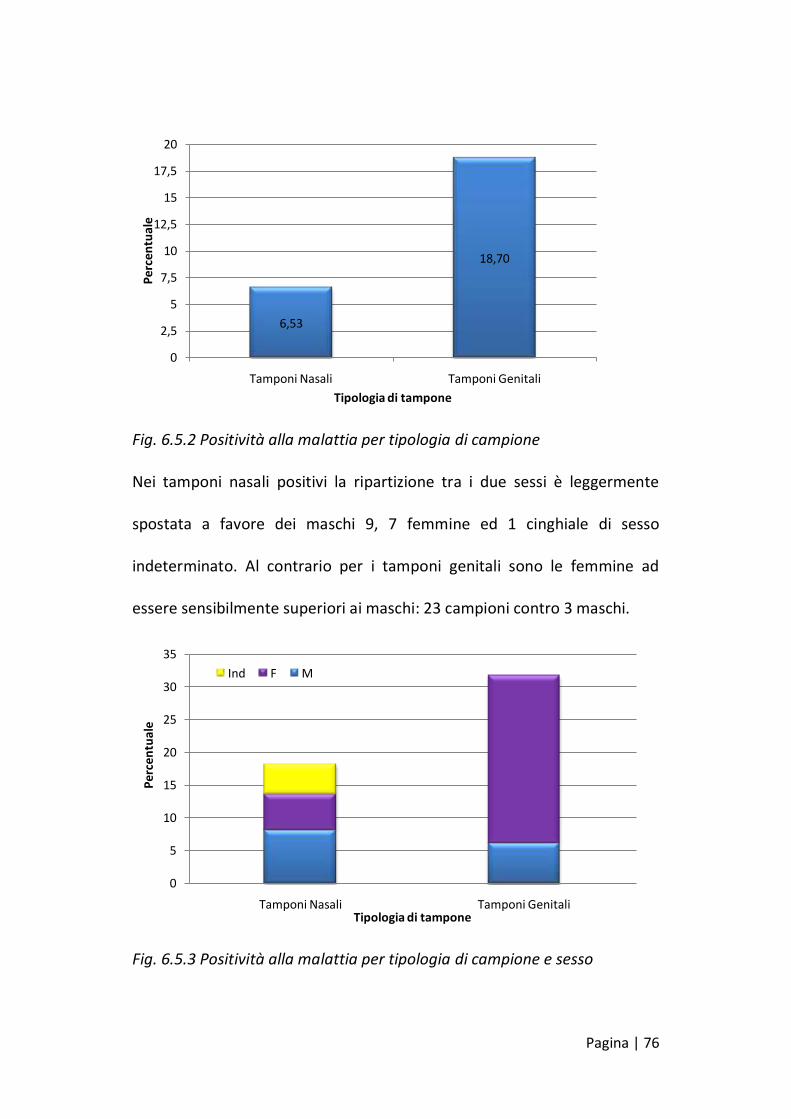
I campioni positivi alla PCR sono stati 43 dei quali 17 (6,54%) tamponi
nasali e 26 (18,71%) tamponi genitali.
0
20
40
60
80
100
120
140
M F Ind M F
Tamponi Nasali Tamponi Genitali
Per
cen
tual
e
Pagina | 76
Fig. 6.5.2 Positività alla malattia per tipologia di campione
Nei tamponi nasali positivi la ripartizione tra i due sessi è leggermente
spostata a favore dei maschi 9, 7 femmine ed 1 cinghiale di sesso
indeterminato. Al contrario per i tamponi genitali sono le femmine ad
essere sensibilmente superiori ai maschi: 23 campioni contro 3 maschi.
Fig. 6.5.3 Positività alla malattia per tipologia di campione e sesso
6,53
18,70
0
2,5
5
7,5
10
12,5
15
17,5
20
Tamponi Nasali Tamponi Genitali
Per
cen
tual
e
Tipologia di tampone
0
5
10
15
20
25
30
35
Tamponi Nasali Tamponi Genitali
Pe
rcen
tual
e
Tipologia di tampone
Ind F M

Pagina | 77
Due soli campioni, entrambi tamponi nasali di due femmine una giovane
ed una adulta, sono risultati positivi alla sierologia ed alla PCR.
Tamponi genitali e Tamponi nasali hanno mostrato una diversa
distribuzione della positività alla malattia tra le tre fasce di età
considerate.
La positività dei tamponi genitali è pressoché simile nei maschi piccoli
(meno di un anno) e negli animali adulti. Differentemente le femmine
registrano un valori differenti nelle tre classi con un massimo nella classe
piccola e poi valori crescenti nel passaggio da giovani a femmine adulte.
Fig. 6.5.4 Positività alla malattia dei tamponi genitali per sesso ed età
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
< 1 1-2 > 2 Ind
Pe
rcen
tual
e
Anni di Età
Maschi Femmine

Pagina | 78
Al contrario la positività nei tamponi nasali aumenta nei maschi con il
progredire dell’età, mentre le femmine non risultano positive sotto l’anno
e presentano valori simili nelle altre due classi di età. Il confronto è stato
ottenuto da un numero di campioni analizzato per le diverse classi e sessi è
molto simile.
Fig. 6.5.5 Positività alla malattia dei tamponi nasali per sesso ed età
0,00
2,00
4,00
6,00
8,00
10,00
12,00
< 1 1-2 > 2
Pe
rce
ntu
ale
Anni di Età
Maschi Femmine Ind.

Pagina | 79
6.6. Risultati analisi virologica sui feti Delle 24 femmine con feti, 4 sono risultate positive all’analisi tramite PCR
(16,67%), così come 6 dei 19 feti totali. La percentuale di feti positivi per
ogni femmina è variata dal 20 al 40% con una media di 31,58.
Fig. 6.6.1 Positività alla malattia nei feti di quatto femmine positive.
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
1 2 3 4
Per
cen
tual
e
Femmine

Pagina | 80
6.7 Effetti della malattia sulla popolazione In provincia di Pisa mancano dati di censimento della specie. Pertanto
abbiamo ipotizzato due diverse percentuali di prelievo pari al 60 e 70%
della popolazione, e basandoci sul dati del nostro campionamento
abbiamo stimato maschi e femmine.
Prelievo
popolazione
Popolazione
stimata
Femmine
stimate
60 % 10.123 5307
70 % 8677 4549
Tab. 6.7.1 Stima della popolazione di cinghiale e delle femmine in provincia
di Pisa.
Sulla base del n° di femmine abbiamo calcolato il n° di piccoli prodotti
ipotizzando che si riproducano il 50% delle femmine giovani e del 90%
femmine adulte, se pure con un n° medio di feti differente nelle due classi.
La ripartizione delle femmine nelle tre classi di età ed il n° di feti per le
femmine adulte sono stati basati sul nostro campionamento, mentre il n°
medio di feti per le femmine giovani è stato ipotizzato inferiore del 50%
rispetto alle adulte.
Sono state formulate due ipotesi a seconda della stima del n° di femmine
presenti nella popolazione di cinghiali della provincia di Pisa.

Pagina | 81
Ipotesi A n° femmine 4.549
Anni di età
Femmine (% che si riproducono)
Media feti Piccoli stimati
0-1 0 0 0
1-2 50 3,0 1.892
> 2 90 6,29 10.603
Tab. 6.7.3 Stima della n° di nuovi nati - ipotesi A.
Ipotesi B n° femmine 5.307
Anni di età
Femmine (% che si riproducono)
Media feti Piccoli stimati
0-1 0 0 0
1-2 50 3,0 1.892
> 2 90 6,29 10.603
Tab. 6.7.4 Stima della n° di nuovi nati - ipotesi B.
Nella prima ipotesi il totale del n° di piccoli potenziale è 12.495, nella
seconda ipotesi 14.577 piccoli, con una differenza di 2.082 capi.
Pur ipotizzando una mortalità dovuta a cause naturali ed alla malattia di
Aujezsky del 50% avremmo una popolazione in entrambi i casi in crescita

Pagina | 82
con valori che supererebbero abbondantemente il prelievo medio
annuale.
Dal 2005 al 2010 in provincia di Pisa sono stati abbattuti nei due ATC dalle
squadre di caccia in battuta, in media 4.500 cinghiali l’anno.
Stando al numero medio di feti registrato in questo studio e dalla
bibliografia (Massei et al., 2006; 1996) basterebbero un numero di
femmine tra 700 e 900 capi per produrre un numero sufficiente di piccoli
tale da mantenere stabile la popolazione.

Pagina | 83
CAPITOLO 7 DISCUSSIONE

Pagina | 84
7. DISCUSSIONE La sieroprevalenza della Malattia di Aujeszky nella popolazione di cinghiali
in provincia di Pisa mostra un’elevata variabilità negli anni e tra le diverse
aree. Analoga indagine nel 2006 aveva evidenziato una sieroprevalenza del
49% (Verin et al., 2006), passata a distanza di tre anni a circa il 27%.
Questo risultato può essere spiegato anche dalla maggiore dimensione del
campione rispetto alle precedenti indagini (Lari et al., 2006).
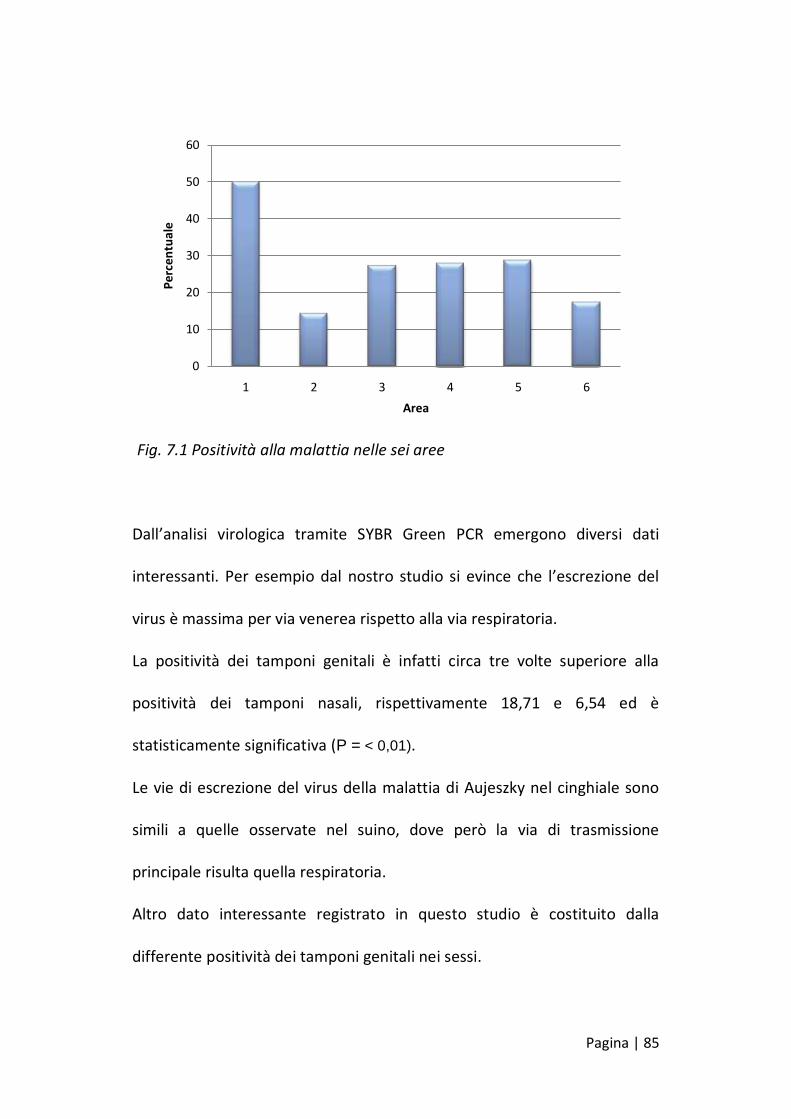
La differenza di positività alla malattia nelle sei aree oggetto di indagine è
statisticamente significativa (P < 0,05) e fa registrare valori estremi nelle
due aree dove vengono abbattuti meno cinghiali con una positività
massima nell’Area 1 (Monti Pisani) e di poco più del 14% nell’Area 2, a
cavallo dei comuni di S.Miniato, Palaia e Peccioli. Nel resto della provincia
la media si attesta a valori intorno al 25% di sieroprevalenza.
Le oscillazioni dei valori di sieroprevalenza sia negli anni che nelle aree
possono essere spiegate dagli alti tassi di prelievo cui la specie è
sottoposta dall’attività venatoria che possono raggiungere punte del 100%
della popolazione pre-parti.
Non esistono differenze significative di sieroprevalenza tra i sessi, mentre
questa risulta statisticamente rilevante nella classi di età in particolare nei
piccoli e soprattutto nei cinghiali adulti.
Pagina | 85
Fig. 7.1 Positività alla malattia nelle sei aree
Dall’analisi virologica tramite SYBR Green PCR emergono diversi dati
interessanti. Per esempio dal nostro studio si evince che l’escrezione del
virus è massima per via venerea rispetto alla via respiratoria.
La positività dei tamponi genitali è infatti circa tre volte superiore alla
positività dei tamponi nasali, rispettivamente 18,71 e 6,54 ed è
statisticamente significativa (P = < 0,01).
Le vie di escrezione del virus della malattia di Aujeszky nel cinghiale sono
simili a quelle osservate nel suino, dove però la via di trasmissione
principale risulta quella respiratoria.
Altro dato interessante registrato in questo studio è costituito dalla
differente positività dei tamponi genitali nei sessi.
0
10
20
30
40
50
60
1 2 3 4 5 6
Pe
rce
ntu
ale
Area

Pagina | 86
Questo dato molto più alto nelle femmine (P < 0,01) può trovare
spiegazione nella biologia riproduttiva della specie. Nel cinghiale se pure
entrambi i sessi raggiungono la maturità fisiologica ad un anno di età, i
maschi difficilmente prendono parte agli accoppiamenti prima del 4° -5°
anno di età, quando conquistano la maturità sociale. La specie è poliginica
pertanto aumenta la possibilità dei maschi di contrarre la malattia man
mano prendono parte agli accoppiamenti.
Diversamente nei tamponi nasali non vi sono differenze significative tra i
sessi, mentre emerge un aumento della positività nei tamponi nasali con il
progredire dell’età dei maschi ( anche se non significativo).
I valori di positività alla malattia registrati attraverso le tre metodologie
evidenziano come una percentuale importante della popolazione di
cinghiali sia venuta a contatto con il virus di Aujeszky e come una gran
parte di essa funga da escretrice del virus per via venerea o aerea.
26,77
6,54
18,71
0
5
10
15
20
25
30
sierologia Tamponi nasali Tamponi genitali
Per
cen
tual
e
Fig. 7.2 Percentuali di positività alla malattia registrati con le tre
diverse metodologie

Pagina | 87
Per quanto riguarda l’analisi della produttività e la ricerca della malattia di
Aujeszky nei feti, le femmine trovate gravide sono state in percentuale
bassa probabilmente a causa del periodo di campionamento legato
necessariamente al prelievo in battuta e di conseguenza al calendario
venatorio.
Nell’ambito degli ungulati italiani il cinghiale è sicuramente la specie quella
con il più alto Incremento utile annuo. Tuttavia le condizioni ambientali
sono determinanti sulla percentuale di femmine che prende parte alla
riproduzione e sul numero di piccoli prodotti.
Nel nostro studio il rapporto tra i sessi è notevolmente differente prima
della nascita rispetto a quanto campionato e ai dati degli abbattimenti
realizzati in regime di controllo da febbraio a settembre (Abbattimenti art.
37 L.R.3/94).
Feti
Campionamento Abbattimenti in controllo
Rapporto M: F
1 : 0,77 1 : 1,21 1 : 1,24
Tab. 7.1 Rapporto sessi nel cinghiale in provincia di Pisa registrato con tre
metodologie differenti

Pagina | 88
Pedone et al., 1995 hanno registrato un rapporto sessi uguale tra maschi e
femmine in quattro diverse aree della Toscana tra cui l’Alta Val di Cecina
compresa in tre delle nostre aree di studio.
È probabile che anche per il cinghiale vi sia una diversa mortalità in fase
giovanile come registrato per altre specie di ungulati ad es. il capriolo
(Varuzza, 2005).
Per quanto riguarda l’influenza della malattia di Aujeszky, dal nostro studio
è stata rilevata una differenza di n° medio di feti tra femmine sane (6,6
feti/femmina) e malate (4,74 feti/femmina). Nonostante questo i dati in
nostro possesso si riferiscono a poche femmine probabilmente
insufficiente per trarre delle conclusioni.
A prima vista la differenza è di poco superiore al 10% in termini di piccoli
prodotti.
Ipotesi A Ipotesi B
Femmine sane
12.495 14.577
Femmine infette
11.025 12.864
Differenza N° feti
1.470 1.713
Tab. 7.1 Differenza del n° di feti prodotti da femmine sane e femmine
infette per la provincia di Pisa sulla base dei risultati di questo studio.

Pagina | 89
È ragionevole ipotizzare che la popolazione sia regolata soprattutto dal
numero di femmine che si riproducono, condizione fortemente influenzata
dall’ambiente e dal peso per le primipare. Purtroppo il dato non è oggetto
di verifica da parte delle squadre di caccia.

Pagina | 90
CAPITOLO 8 CONCLUSIONI

Pagina | 91
8. CONCLUSIONI La malattia di Aujeszky nel cinghiale rimane ancora poco conosciuta in
particolare la specie sembra aver raggiunto una certa resistenza al virus
tale da non subirne i gravi effetti che si registrano nel suino domestico.
Dai nostri dati emerge che l’escrezione della malattia avviene
prevalentemente per via sessuale, e interessa in misura maggiore animali
adulti.
Probabilmente gli alti tassi di prelievo sulla specie condizionano
pesantemente non solo la diffusione della malattia, ma anche il suo
campionamento e lo studio dell’incidenza nei sessi e nelle diverse età.
Tra le metodologie di indagine sia l’analisi sierologica che l’analisi
virologica con SYBR Green PCR possono essere impiegate con ottimi
risultati anche su larga scala, fornendo un buon rapporto risultati-
sforzo/metodologia campionamento e costi. Il campionamento attraverso
la raccolta del sangue e dei tamponi nasali e o genitali può essere
effettuato con l’aiuto dei referenti delle squadre di caccia al cinghiale
previa formazione sulle modalità di campionamento e di conservazione dei
campioni.

Pagina | 92
Rimangono da approfondire le relazioni di positività tra feti e femmine, in
particolare in aree chiuse tali da permettere e verificare una adeguata
sperimentazione.
Per quanto riguarda la dinamica di popolazione la gestione di una specie di
così grande importanza nel panorama faunistico italiano non può
prescindere da una raccolta di dati biologici indispensabili per pianificarne i
piani di prelievo.
Parametri come peso delle femmine, stato riproduttivo, n° di feti e classi di
età degli animali abbattuti devono entrare nella norma dei dati raccolti su
tutto il territorio provinciale solo così nel giro di pochi anni potremmo
sopperire alla mancanza di informazioni sull’entità della popolazione di
cinghiali su base provinciale, informazione essenziale per definire la
gestione della specie.

Pagina | 93
CAPITOLO 9 BIBLIOGRAFIA

Pagina | 94
8. BIBLIOGRAFIA
AA.VV. 1989.
http://www.defra.gov.uk/animalh/diseases/notifiable/disease/auje
szkys. htm 1989; Aujeszky’s disease in pigs – Notifiable Diseases
(DEFRA, UK).
Albinia E., Cabriolet R., Eveno E et al. 1996.
Premiers résultats du CNEVA sur le dépérissement fetal du porcelet
en fin de post-sevrage. Suppl Sem Vet. 834 : 1-2.
Abaigar T. 1992.
Paramètres de la reproduction chez le sanglier (Sus scrofa) dans le
sud- est de la péninsule ibérique. Mammalia 56: 245-250.
Abaigar T. 1993.
Regimen alimentario del Jabalí (Sus scrofa, L. 1758) en el sureste
Iberico. Doñana, Acta vertebrata 20: 35-47.
Acevedo P, Escudero M. A., Munoz R., Gortázar C. 2006.
Factors affecting wild boar abundance across an environmental
gradient in Spain. Acta Theriologica 51: 327-336.
Ahmad E., Brooks J E, Hussain I, Khan M. H. 1995.
Reproduction in Eurasian wild boar in central Punjab, Pakistan. Acta
Theriologica 40: 163-173.
Albinia E., Mespedale A., Chenut G. et al. 2000.
A serological survey on classical swine fever (CSF), Aujeszky’s
disease (AD) and porcine reproductive and respiratory syndrome
(PRRS) virus infection in French wild boar from 1991 to 1998. Vet
Microbiol. 77: 43-57.
Allan G.M., Kennedy S., McNeilly F. et al. 1999.
Experimental reproduction of wasting disease and death by co-
infection of pigs with porcine circovirus and porcine parvovirus. J of
Comp Pathol. 121: 1-11.
Allan G.M., Mackie D.P., McNair J et al. 1994.
Production preliminary characterization and application of
monoclonal antibodies to porcine circovirus. Vet Immunol
Immunopathol.; 43: 357- 371.

Pagina | 95
Allan G.M., Mcneilly F., Cassidy J.P. et al. 1995.
Pahogenesis of porcine circovirus; experimental infections of
colostrums deprived piglets and examination of pig foetal
material. Vet Microbiol. 44: 49-64
Andrzejewski R., Jezierski W. 1978.
Management of a wild boar population and its effects on
commercial land. Acta Theriologica 23: 309-339.
Artois M., Depner K.R., Guberti V., Hars J., Rossi S., Rutili D. 2002.
Classical swine fever (hog cholera) in wild boar in Europe. Rev. sci.
tech. Off. Int. Epiz. 21: 287-303.
Aumaître A, Queré J. P., Peiniau J. 1984.
Influence du milieu sur la reproduction hivernale et la prolificité de
la laie. Symposium international sur le sanglier, F. Spitz, D. Pépin,
eds. Colloques del´I.N.R.A., Toulouse (France): 69-78.
Baber D W, Coblentz B. E. 1987.
Diet, nutrition, and conception in feral pigs on Santa Catalina Island.
Journal of Wildlife Management 51: 306-317.
Badia J., Spitz F., Valet G. 1991.
Estimate of the size of a hunted population. Ecological Modelling
55: 113-122.
Ben-Porat T., DeMarchi J.M., Lomniczi. B., Kaplan AS. 1986.
Role of glycoproteins of pseudorabies virus in eliciting neutralizing
antibodies. Virology. 154: 325-334.
Boitani L., Mattei L., Nonis D., Corsi F. 1994.
Spatial and activity patterns of wild boars in Tuscany, Italy.
Journal of Mammalogy 75: 600-612.
Boitani L., Trapanese P., Mattei L . 1995.
Demographic patterns of a wild boar (Sus scrofa L.) population in
Tuscany, Italy. IBEX - Journal of Mountain Ecology 3: 197-201.
Boitani L., Trapanese P., Mattei L., Nonis D. 1995.
Demography of a wild boar (Sus scrofa, L.) population in Tuscany,
Italy. Gibier Faune Sauvage 12: 109-132.
Bouldoire J. L. 1984.

Pagina | 96
Le statut du sanglier en France. Evolution entre 1975 et 1982.
Symposium international sur le sanglier, Toulouse (France): 173-
186.
Briedermann L. 1971.
Zur Reproduktion des Schwarzwildes in der Deutschen
Demokratischen Republik. Tagungsbericht der deutschen
Akademie für Landwirtschaftswissenschaften Berlin 113: 169-186.
Briedermann L. 1981.
Das Schwarzwild. in: Buch der Hege. Berlin, DLV. 1: 163-195.
Briedermann L. 1986.
Schwarzwild. Berlin, Neumann-Neudamm.
Brown T.T., Kwang K.O., Fuller. F.J. 1995.
Detection of pseudorabies viral DNA in tonsillar epithelial cells of
latently infected pigs. Am. J. Vet. Res. 56: 587-594.
Buhk H.J., Tisher I., Koch M.A. 1985.
Cloning and sequencing of porcine circovirus (PCV) genome.
Zentralbe Bakteriol Org A. 260: 465.
Cahill S., Llimona F. 2002.
Demographics of a wild boar Sus scrofa Linnaeus, 1758 population
in a metropolitan park in Barcelona. Wild boar research 2002. A
selection and edited papers from the "4th international wild boar
symposium", Lousa, Portugal: 37-52.
Capua I., Fico R., Banks M., Tamba M., Calzetta G. 1997
Isolation and characterization of an Aujeszky’s disease virus
naturally infecting a wild boar (Sus scrofa). Vet. Microbiol. 55: 141-
6.
Capua I., Casaccia C., Calzetta G., Caporale V. 1997.
Characterisation of Aujeszky’s disease viruses isolated from
domestic animals and from a wild boar (Sus scrofa) in Italy between
1972 and 1995. Vet. Microbiol. 51: 143-149
Cheung A., Vlcek C., Paces V., Schwyzer M. 1990.
Update and comparison of the immediate-early gene DNA
sequences of two pseudorabies virus isolates. Virus Genes. 4: 261-
265.

Pagina | 97
Clarke C. M. H., Dzieciolowski R. M., Batcheler D., Frampton C. M. 1992.
A comparison of tooth eruption and wear and dental cementum
techniques in age determination of New Zealand feral pigs. Wildlife
research 19: 769-777.
Cordioli P., Callegari S., Berlinzani A., Foni E., Candotti P., Barigazzi G. 1993.
Indagine sierologia su cinghiali dell’appennino Parmense. Atti Soc.
It. Sci. Vet. 47: 1065-1068.
Cugnasse J. M., Teillaud P., Bon R. 1987.
Résultats préliminaires sur l'activité diurne et sur la structure des
groupes de sangliers (Sus scrofa L.) dans les Monts de l'Espinouse.
Gibier Faune Sauvage 4: 267-277.
Dardaillon M. 1984.
Organisation sociale et reproduction chez le sanglier en Camargue.
Symposium international sur le sanglier, Colloques de l´I.N.R.A.,
Toulouse (France), INRA: 159-165.
Dardaillon M. 1987.
Seasonal feeding habits of the wild boar in a Mediterranean
wetland, the Camargue (Southern France). Acta Theriologica 32:
389-401.
Delcroix I., Mauget R., Signoret J P. 1990.
Existence of synchronization of reproduction at the level of the
social group of the European wild boar (Sus scrofa). Journals of
reproduction and fertility 89: 613-617.
Dulac G.C., Afshar A. 1989.
Porcine circovirus antigens in PK/15 cell line (ATCC CCL-33) and
evidence of antibodies to circovirus in Canadian pigs. Can J Vet Res.
53: 431-433.
Durio P., Gallo Orsi U., Macchi E., Perrone A. 1995.
Structure and monthly birth distribution of a wild boar population
living in mountainous environment. IBEX - Journal of Mountain
Ecology 3: 202-203.
Edwards S., Sands J. J. 1994.
Evidence of circovirus infection in British pigs. Vet Rec. 134: 680-
681. 87
Ellis J., Bratanich A., Clark E.G. et al. 2000.

Pagina | 98
Co-infection by porcine circoviruses and porcine parvovirus in pigs
with naturally acquired post-weaning multisistemic wasting
syndrome. J Vet Dian Invest 12.
Elbers A.R., Dekkers L.J., van der Giessen J.W. 2000.
Sero-surveillance of wild boar in The Netherlands, 1996-1999. Rev
Sci Tech.19: 848-54.
Ellis J., Hassard L., Clark E.G. et al. 1998.
Isolation of circovirus from lesions of pigs with post-weaning
multisistemic wasting syndrome. Can Vet J. 39: 44-51.
Ellis J., Krakowka S., Allan G. et al. 1999.
The clinical scope of PRRSV infection has expanded since 1987; an
alternative explanation. Vet Pathol. 36: 262-265.
Fernández-Llario P., Carranza J. 2000.
Reproductive performance of the wild boar in a Mediterranean
ecosystem under drought conditions. Ethology Ecology & Evolution
12: 335-343.
Fernández-Llario P., Carranza J. & Mateos-Quesada P. 1999.
Sex allocation in a polygynous mammal with large litters: the wild
boar. Animal Behaviour 58: 1079-1084.
Fernández-Llario P & Mateos-Quesada P. 1998.
Body size and reproductive parameters in the wild boar Sus scrofa.
Acta Theriologica 43: 439-444.
Fernández-Llario P. & Mateos-Quesada P. 2003.
Population structure of the wild boar (Sus scrofa) in two
Mediterranean habitats in the western Iberian Peninsula. Folia
Zoologica 52: 143-148.
Fernández-Llario P., Mateos-Quesada P. 2005.
Influence of rainfall on the breeding biology of wild boar (Sus scrofa)
in a Mediterranean ecosystem. Folia Zoologica 54: 240-248.
Fernández-Llario P., Mateos-Quesada P. 2005.
Udder preference in wild boar piglets. Acta Ethologica 8: 51-55.
Fernández-Llario P., Mateos-Quesada P, Silverio A., Santos P. 2003.
Habitat effects and shooting techniques on two wild boar (Sus
scrofa) populations in Spain and Portugal. Zeitschrift für J
agdwissenschaft 49: 120-129.

Pagina | 99
Fonseca C., Santos P., Monzón A., Bento P., Alves da Silva A., Alves J.,
Silvério A., Soares A. M.V. M., Petrucci-Fonseca F. 2002.
Reproduction in the wild boar (Sus scrofa Linnaeus, 1758)
populations of Portugal. Wild boar research 2002. A selection and
edited papers from the "4th international wild boar symposium",
Lousa, Portugal: 53-65.
Fournier-Chambrillon C., Maillard D. & Fournier P. 1996.
Variabilité du régime alimentaire du sanglier (Sus scrofa L.) dans les
garrigues de Montpellier (Hérault). Gibier Faune Sauvage, Game
Wildlife 13: 1457-1476.
Fruzinski B. 1995.
Situation of wild boar population in western Poland. IBEX - Journal
of Mountain Ecology 3: 186-187.
Gaffuri A., Tranquillo V.M., Pellicioli L., Lanfranchi P. 2002.
Monitoraggio sanitario della fauna selvatica in provincia di B
ergamo. Rivista dell'OEVR (Osservatorio Epidemiologico IZS
Brescia) Anno 5 - n.5.
Gaillard J-M, Brandt S & Jullien J-M 1993.
Body weight effect on reproduction of young wild boar (Sus scrofa)
females: a comparative analysis. Folia Zoologica 42: 204-212.
Gaillard J-M, Pontier D, Brandt S, Jullien J-M & Allainé D. 1992.
Sex differentiation in postnatal growth rate: a test in a wild boar
population. Oecologia 90: 167-171.
Gaillard J-M., Vassant J., Klein F. 1987.
Quelques caractéristiques de la dynamique des populations de s
angliers (Sus scrofa scrofa) en milieu chassé. Gibier Faune Sauvage
4: 31-47.
Galeota-Wheeler J., Osorio F.A. 1991.
Investigation of sites of pseudorabies virus latency, using
polimerase chain reaction. Am. J. Vet. Res. 52: 1799-1803
Gallo Orsi U., Macchi E., Perrone A. & Durio P.1995.
Biometric data and growth rates of wild boar population living in
the Italian Alps. IBEX - Journal of Mountain Ecology 3: 60-63.
Geisser H., Reyer H-U. 2005.

Pagina | 100
The influence of food and temperature on population density of
wild boar Sus scrofa in the Thurgau (Switzerland). Journal of Zoology
267: 89-96.
Giovannini A., Cancellotti F.M., Turilli C., Randi E. 1988.
Serological investigations for some bacterial and viral pathogens in
fallow deer (Cervus dama) and wild boar (Sus scrofa) of the San Rossore
Preserve, Tuscany, Italy. J Wildl Dis. 1988. 24: 127-32.
Gortazar C., Vincente J., Fierro Y., Leon L., Cubero M.J., Gonzales M. 2002.
Natural Aujeszky’s disease in a Spanish wild boar population. Ann
N Y Acad Sci 969: 210-212.
Goulding M.J. 2001.
Possible genetic sources of free-living wild boar (Sus scrofa) in
southern England. Mammal Review 31: 245-248.
Green A.J. 2001.
Mass/length residuals: measures of body condition or generators of
spurious results? Ecology 82: 1473-1483.
Gresham A., Jackson G., Giles N. 2000. PMWS and porcine nephropathy
syndrome in Great Britain. Vet Rec. 146: 143.
Groot Bruinderink G.W., Hazebroek E. 1996.
Wild boar (Sus scrofa scrofa L.) rooting and forest regeneration on
podzolic soils in the Netherlands. Forest Ecology and Management
88: 71-80.
Groot Bruinderink G. W, Hazebroek E., Van der Voot A. 1994.
Diet and condition of wild boar, Sus scrofa scrofa, without
supplementary feeding. Journal of Zoology 233: 631-648.
Grieco V., Gelmetti D. Finazzi G., Brocchi E., Finazzi M. 1997.
Immunohistologic diagnosis of psuedorabies (Aujeszky’s disease)
using monoclonal antibodies. J. Vet. Diagn. Invest. 9: 326-8.
Guberti V., Ferrari G., Fenati M., De Marco M.A., Pasquali T. 2002.
Pseudorabies in wild boar. Proceeding 4th Meeting European
Association of Zoo- and Wildlife Veterinarians (EAZWV) 8-12 May,
Heidelberg, Germany.
Hahn E.C., Page G.R., Hahn P.S., Gillis K.D., Romero C., Annelli J.A., Gibbs
E.P.J. 1997.

Pagina | 101
Mechanisms of transmission of Aujeszky’s disease virus originating
from feral swine in the USA. Vet. Microbiol. 55: 123-130.
Henry V.G. 1966.
European wild hog hunting season recommendations based on
reproductive data. Proceedings of the annual conference
Southeastern association of game and fish commission 20: 139-145.
Henry V.G. 1968.
Fetal development in European wild hogs. Journal of Wildlife
Management 32: 966-970.
Henry V.G. 1969.
Estimating whole weights from dressed weights for European wild
hogs. Journal of Wildlife Management 33: 222-225.
Herrero J., García-Serrano A., Couto S., Ortuño V.M., García-González R.
2006.
Diet of wild boar Sus scrofa L. and crop damage in an intensive
agroecosystem. European Journal of Wildlife Research 52: 6.
Hines R.K., Lukert P.D. 1994.
Porcine circovirus as a cause of congenital tremors in newborn pigs.
Proc Am Assoc Swine Pract. 344-345.
Hinrichs U., Ohlinger V.F., Pesh S. et al. 1999.
First report of porcine circovirus type-2 infection in Germany.
Tieraerztl Umsch.; 54: 255-258.
Höfle U., Vicente J., Fernández-de-Mera I.G., Villanúa D., Acevedo P., Ruiz-
Fons F., Gortázar C. 2002.
Health risks in game production: the wild boar. Wild boar research
2002. A selection and edited papers from the "4th international wild boar
symposium", Lousa, Portugal: 197-206.
Henry V. G. 1968.
Foetal development in European wild hogs. J. Wildlife Manage., 32
(4): 966 -971.
Horner G. 1991.
Pig circovirus antibodies present in New Zealand pigs. Surveil
Wellington. 18: 23.
Jakob E. M., Marshall S.D., Uetz G.W. 1996.

Pagina | 102
Estimating fitness: a comparison of body condition indices. Oikos
77: 61-67.
Jacobs L., Mulder W.A.M., Van Oirschot J.T., Gielkens A.L.J., Kimman T.G.
1993. Deleting two amino acids in glycoprotein gI of pseudorabies virus
decreases virulence and neurotropism for pigs, but does not affect
immunogenicity. J. Gen. Viral. 74: 2201-2206.
James W. H., 1998. Porcine sex ratios and sex combinations within litters: comment on Meikle et al. (1997) and Mendl et al. (1997). Animal Behaviour 55: 767- 769. Janeau G., Spitz F. 1984. L'espace chez le sanglier (Sus scrofa scrofa L.): occupation et mode d'utilisation. Gibier Faune Sauvage 1: 73-89. Jullien J-M., Brandt S., Vassant J., Chantecaille S. 1991. Des sangliers chassés en battue en forêt domaniale de Châteauvillain/Arc-en-Barrois: leurs "stratégies" pour échapper aux chasseurs et aux chiens. Bulletin Mensuel del'Office National de la Chasse 162: 29-36. Kaberghs J. 2004. Sanglier - Impact du nourrissage artificiel. Chasse & Nature Mai 2004: 29- 32. Kabudi P., Fetter S., Libois R.M., Schumacker R., Ruwet J.C. 1987. Etude du régime alimentaire du sanglier (Sus scrofa L.) dans les Ardennes belges. Cahiers d'Ethologie appliquée 7: 223-246. Kaden V., Lange B. 2001. Oral immunisation against classical swine fever (CSF): onset and duration of immunity. Veterinary Microbiology 82: 301-310. Kimman T.G., de Wind N., Oei-Lie N., Pal J.M.A., Berm A.J.M., Gielkens
A.L.J. 1992.
Contribution of single genes within the unique short region of
Aujesaky’s disease virus (suid herpes virus type 1) to virulence,
pathogenesis and immunogenicity. J. Gen. Viral. 73: 243-25 1.
Kimman T.G. 1994.
Immunological protection against pseudorabies virus. Proc. O.I.E.
Symp. on Aujesrky’s Disease, Bangkok, Thailand, pp. 1 l-22.
Kit S., Kit M., Pirtle CC. 1985.
Attenuated properties of thymidine kinase-negative deletion
mutant of pseudorabies virus. Am. J. Vet. Res. 46: 1359-1367.

Pagina | 103
Klupp B., Baumeister J., Karger A., Visser N., Mettenleiter T.C. 1994.
Identification and characterization of a novel structural glycoprotein
in pseudorabies virus, gL. J. Virol. 68: 3868-3878.
Klupp B., Visser N., Mettenleiter T.C. 1992.
Identification and characterization of pseudorabies virus
glycoprotein H. J. Virol. 66: 3048-3055.
Kodera Y., Kanzaki N. 2001. Food habits and nutritional condition of Japanese wild boar in
Iwami district, Shimane prefecture, western Japan. Wildlife
Conservation Japan 6: 109-117.
Kozdrowski R., Dubiel A. 2004. The effect of season on the properties of wild boar (Sus scrofa L.) semen. Animal Reproduction Science 80: 281-289. Krakowa S., Ellis J.A., McNeilly F. et al. 2001.
Activation of the immune system is the pivotal event in the
production of wasting disease in pigs infected with porcine
circovirus-2 (PCV-2). Vet Pat. 38: 31-42.
Kritas S.K., Nauwynck H.J., Pensaert M.B. 1995.
Dissemination of wild-type and gC-, gE- and gI-deleted mutants of
Aujeszky’s disease virus in the maxillary nerve and trigeminal
ganglion of pigs after intranasal inoculation. J. Gen. Viral. 76: 2063-
2066.
Kritas S.K., Pensaert M.B., Mettenleiter T.C. 1994.
Role of envelope glycoproteins gI, gp63 and gII1 in the invasion and
spread of Aujeszky’s disease virus in the olfactory nervous pathways
of the pig. J. Gen. Viral. 75: 2319-2327.
Kritas S.K., Pensaert M.B., Mettenleiter T.C. 1994.
Invasion and spread of single glycoprotein deleted mutants of
Aujeszky’s disease virus in the trigeminal nervous pathway of pigs after I
ntranasal inoculation. Vet. Microbial. 40: 323-334.
Kruuk L. E. B., Clutton-Brock T. H., Albon S. D., Pemberton J. M., Guinness F E. 1999. Population density affects sex ratio variation in red deer. Nature
399: 459-461.
Kuipel M., Stevenson G.W., Mittal S.K., Clark E.G. 1998.

Pagina | 104
Circovirus-like disease in weaned pigs in Indiana. Vet Pathol. 35:
303- 307.
Lari A., Lorenzi D., Nigrelli D., Brocchi E., Faccini S., Poli A. 2006.
Pseudorabies virus in European wild boar from central Italy. J Wildl
Dis. 42:319-24.
Larochelle R., Morin M ., Antaya M. et al. 1999.
Identification and incidence of porcine circovirus in routine field
cases in Quebec as determined by PCR. Vet Rec. 145: 140-142.
LeCann P., Albina E., Madec F. 1997.
Piglet wasting disease. Vet Rec. 141: 660.
Lemel J, Truvé J & Söderberg B. 2003.
Variation in ranging and activity behaviour of European wild boar
Sus scrofa in Sweden. Wildlife Biology 9: 29-36.
Lucas X., Martínez E. A., Roca. J., Vázquez J. M., Gil M. A., Pastor L. M.,
Alabart J. L. 2002.
Relationship between antral follicle size, oocyte diameters and
nuclear maturation of immature oocytes in pigs. Theriogenology 58:
871-885.
Maillard D & Fournier P. 2002. Timing and synchrony of births in the wild boar (Sus scrofa Linnaeus, 1758) in a mediterranean habitat: the effect of food availability. Wild boar research 2002. A selection and edited papers from the "4th international wild boar symposium", Lousa, Portugal: 67-74. Markina F. A., Sáez-Royuela C., De Garnica R. 2002- Physical development of wild boar in the Cantabric Mountains, Álava, Northern Spain. Wild boar research 2002. A selection and edited papers from the "4th international wild boar symposium", Lousa, Portugal: 25-34. Marchioli C.C., Yancey R.J., Timmins .J.G., Post L.E., Young B.R. and
Povendo L.A. 1988.
Protection of mice and swine from pseudorabies virus-specific
mouse monoclonal antibodies. Am. J. Vet. Res. 49: 860-864.
Massei G, Cowan D. P., Miller L. 2006. Fertility control to manage wild boar: a viable option? Sixth international symposium on wild boar (Sus scrofa) and on sub-order

Pagina | 105
Suiformes, Holy Monastery of Kykkos, Cyprus, Ministry of the Interior, Cyprus: 45. Massei G, Genov P., Staines B. W. 1996. Diet, food availability and reproduction of wild boar in a Mediterranean coastal area. Acta Theriologica 41: 307-320. Massei G., Genov P.V. 2002. The environmental impact of wild boar. Wild boar research 2002. A selection and edited papers from the "4th international wild boar symposium", Lousa, Portugal: 135-145. Mattioli S., Pedone P. 1995. Dressed versus undressed weight relationship in wild boars (Sus scrofa) from Italy. IBEX - Journal of Mountain Ecology 3: 72-73. Matschke G.H. 1964. The influence of oak mast on European wild hog reproduction. Annual Conference Southeastern Association Game Fish Commission: 35-39. Matschke G.H. 1967. Aging European wild hogs by dentition. Journal of Wildlife Management 31: 109-113. Mauget R. 1972. Observations sur la reproduction du sanglier (Sus scrofa L.) à l'état sauvage. Annales de Biologie Animale, Biochimie et Biophysique 12: 195- 202. Mauget R, Aumaître A., Peignier Y. 1988. Caractérisation de l'état trophique du sanglier, en milieu forestier, à partir du gras mésentérique et des paramètres sanguins. Reproduction Nutrition Development 28:969-978. Mauget R., Pépin D. 1987 Energy intake, growth and timing of puberty in the European wild boar Sus scrofa L. 18th IUGB Congress, Krakow, Swiat Press: 205-209. Meehan B.M., McNeilly F. 1998.
Todd D. et al. Characterization of novel circoviruses associated with
wasting syndromes in pigs. Gen Virol. 79: 2171-2179.
Meikle D.B., Drickamer L.C., Vessey S.H., Rosenthal T.L., Fitzgerald K.S. 1993. Maternal dominance rank and secondary sex ratio in domestic swine. Animal Behaviour 46: 79-85. Meikle D.B., Vessey S.H., Drickamer L.C. 1998. Mechanisms of sex-ratio adjustment in domestic swine: reply to James. Animal Behaviour 55: 770-772.

Pagina | 106
Mendl M., Broom D. M., Zanella A. J. 1998. Multiple mechanisms may affect birth sex ratio in domestic pigs. Animal Behaviour 55: 773-776. Mendl M, Zanella A. J., Broom D. M., Whittemore C. T. 1995. Maternal social status and birth sex ratio in domestic pigs: an analysis of mechanisms. Animal Behaviour 50: 1361-1370. Mettenleiter T.C., Lomniczi B., Zsak L., Medveczky 1., Ben-Porat T., Kaplan
AS. 1989: J.T. Van Oirschot (Editor)
Analysis of the factors that affect virulence of pseudorabies virus.
Proc. CEC Seminar onVaccination and Control of Aujeszky’s
Disease. pp. 3- 11.
Miry C., Pensaert M. 1989.
Aujeszky’s disease virus replication in tonsils and respiratory tract of
non-immune and immune pigs. In: J.T. van Oirschot (Editor),
Vaccination and Control of Aujeszky’s Disease, pp. I63- 173.
Morandi F., Verin R., Sarli G., Canetti N., Scacco M., Panarese S., Poli A. 2010. Porcine circovirus type 2 (PCV2) antigen localisation and post- weaning multisystemic wasting syndrome (PMWS) in free- ranging wild boar (Sus scrofa spp scrofa) in Italy., European Journal of Wildlife Research, ISSN: 1612-4642 doi: 10.1007/s10344-010-0365-1 2010 Morandi F., Verin R., Sarli G., Canetti N., Scacco M., Panarese S., Poli A. 2010. Immunolocalizzazione degli antigeni di circovirus suino tipo 2 (PCV2): diffusione dell'infezione e diagnosi di PMWS in popolazioni di cinghiali in Italia. Large Animal Review, vol. 16; p. 77-84, ISSN: 1124-4593 2010 Moretti M. 1995. Biometric data and growth rates of mountain population of wild boar (Sus scrofa L.), Ticino, Switzerland. IBEX - Journal of Mountain Ecology 3: 56-59. Mörner T., Obendorf D.L., Artois M., Woodford M.H. 2002.
Surveillance and monitoring of wildlife T diseases. Rev. sci. tech. Off.
int. Epiz. 21: 67-76.
Morozov I., Sirinaurumiter T., Sorden S.D. et al. 1998.

Pagina | 107
Detection of a novel strain of porcine circovirus in pis with post-
weaning multisistemic wasting syndrome. J Clin Microbiol. 36:
2535-2541.
Mulder W.A.M., Pol J.M.A., Kimman T.G., Kok G.L., Priem J., Peeters B.P.H.
1995. gD-negative pseudorabies virus can spread transneuronally via
direct cell-to-cell transmission in its natural host, Abstract No. 52.
20th bit. Herpesvirus Workshop, Groningen. The Netherlands.
Muller T., Klupp B., Zellmer R., Teuffert J., Ziedler K., Possardt C., Mewes L.,
Dresenkamp B., Conraths F.J., Mettenleiter TC. 1998.
Characterisation of pseudorabies virus isolated from wild boar (Sus
scrofa). Vet Rec. 143: 337-40.
Muller T., Teuffert J., Ziedler K., Possardt C., Kramer M., Staubach C.,
Conraths FJ. 1998.
Pseudorabies in the European wild boar from eastern Germany. J.
Wildlife Dis. 34 : 251-258.
Náhlik A., Sándor G. 2003. Birth rate and offspring survival in a free ranging wild boar Sus scrofa population. Wildlife Biology 9: 37-42. Nakamura T., Ihara T., Nagata T., Ishihama A., Ueda S. 1990.
A complement-dependent neutralizing monoclonal antibody against
glycoprotein II of pseudorabies virus. Vet. Microbial. 24: 193-198.
Nauwynck H.J., Pensaert M.B. 1992.
Abortion induced by cell-associated pseudorabies virus in
vaccinated sows. Am. J. Vet. Res. 53: 489-493.
Nauwynck H.J., Pensaert M.B. 1995.
Effect of specific antibodies on the cell-associated spread of
pseudorabies virus in monolayers of different cell types. Arch. Virol.
140: 1137- 1146.
Nauwynck H.J. 1993.
The cell-associated spread of Aujeszky’s disease virus and its
possible role in the pathogenesis of abortion in vaccinated sows.
Ph.D. thesis, Gent, Belgium.
Nayar G.P.S., Hamel A., Lin L. 1997.

Pagina | 108
Detection andcharacterization of porcine circovirus associated with
postweaning multisistemic wasting syndrome in pigs. Can Vet . 38:
385-386.
Niagro F.D., Forsthoefel A.N., Lawthr R.P. et al. 1998.
Beak and feather disease virus and porcine circovirus genomes;
intermediates between the geminivirus and plant circovirus. Arch
Virol. 143: 1723-1744.
Oggiano A., Patta C., Laddomada A., Caccia A. 1991.
Indagine epidemiologica sulla diffusione della malattia di Aujeszky
nei cinghiali della Sardegna. Atti Soc. It. Sci. Vet. : 1157-61.
Pallares F., Halbur P.G., Opriessnig T. et al. 2002.
Porcine circovirus type 2 (PCV-2) coinfections in US field cases of
postweaning multisystemic wasting syindrome (PMWS). J.Vet
Diagnostic Investigation. 14: 515-519.
Pedone P, Mattioli L, Mattioli S, Siemoni N, Lovari C & Mazzarone V (1991) Body growth and fertility in wild boars of Tuscany, Central Italy. XXth congress of the international union of game biologists, Gödöllö, Hungary: 604-609. Pedone P, Mattioli S & Mattioli L. 1995. Body size and growth patterns in wild boars of Tuscany, Central Italy. IBEX – Journal of Mountain Ecology 3: 66-68. Peeters B., De Wind N., Broer R., Gielkens A. and Moormann R. 1992.
Glycoprotein H of pseudorabies is essential for entry and cell-to-cell
spread of the virus. J. Virol. 66: 3888-3892.
Peeters B., De Wind N., Hooisma M., Wagenaar F., Gielkens A., Moormann
R. 1992:
Pseudorabies virus envelope glycoproteins gp50 and gI1 are
essential for virus penetration, but only gI1 is involved in
membrane fusion. J. Viral. 66: 894-905.
Pensaert M.B., De Smet K., De Waele K. 1990.
Extent and duration of virulent virus excretion upon challenge of
pigs vaccinated with different glycoprotein-deleted Aujeskzy’s
disease vaccines. Vet. Micro-biol. 22: 107-117.
Pépin D. 1991.

Pagina | 109
Alimentation, croissance et reproduction chez la laie: études en conditions naturelles et en captivité. Institut National de la Recherche Agronomique Productions Animales 4: 183-189. Pépin D & Mauget R. 1989. The effect of planes of nutrition on growth and attainment of puberty in female wild boars raised in captivity. Animal Reproduction Science 20: 71-77. Pépin D., Spitz F., Janeau G., Valet G. 1987. Dynamics of reproduction and development of weight in the wild boar (Sus scrofa) in South-west France. Zeitschrift für Säugetierkunde 52: 21-30. Pol J.M.A. 1990.
Interferons affect the morphogenesis and virulence of pseudorabies
virus. Ph.D. thesis, Utrecht, The Netherlands.
Poli A., Guarda F., Mazzei M. et al. 2002.
Osservazioni preliminari sulla prevalenza ed il tipo di
glomerulonefriti nel suino. XXVIII Meeting annuale Soc. It. Pat. All.
Suino.
Randi E. 2005. Management of wild ungulate populations in Italy: Captive- breeding, hybridization and genetic consequences of translocations. Veterinary Research Communications 29:71-75. Rodrigue-Arroja G.M., Segalès J., Rossel C. et al. 1999.
Aujeszky’s disease virus infection concurrent with postweaning
multisistemic wasting syndrome in pigs. Vet Rec. 144: 152-153.
Romeo G., Mattii M-, Cimino L., Massolo A. 2001. Produttività del cinghiale in una popolaziona soggetta a prelievo venatorio. III Congresso Italiano di Teriologia - Biologia e gestione dei Mammiferi, San Remo, Italy: 113. Romero C.H., Meade P.N., Shultz J.E., Chung H.Y., Gibbs E.P., Hahn E.C.,
Lollis G. 2001.
Venereal transmission of pseudorabies virusesindigenous to feral
swine. J. Wildl. Dis. 37: 289-96.
Romero C.H., Meade P.N., Homer B.L., Shultz J.E., Lollis G. 2003.
Potential sites of virus latency associated with indigenous
pseudorabies viruses in feral swine. J. Wildlife Dis. 39: 567-575.

Pagina | 110
Romero C.H., Meade P.N., Santagata J., Gillis K., Lollis G., Hahn E.C., Gibbs
E.P.J. 1997.
Genital infection and transmission of pseudorabies virus in feral
swine in Florida, USA. Vet. Microbiol. 55: 131-139.
Ros Bascuñana C., Biörnerot L., Ballagi-Pordány A., Robertsson J- Å., Belák
S. 1997.
Detection of pseudorabies virus genomic sequences in apparently
uninfected “single reactor” pigs. Veterinary Microbiology. 55: 37-
47.
Rossel C., Segalès J., Plana-Duran J. et al. 1999.
Pathological, immunohistochemical and in-situ hybridization studies
of natural cases of post-weaning multisistemic wasting syndrome
(PMWS) in pigs. J Comp Pathol. 120: 59-78.
Ruiz-Fons F., Vicente J., Vidal D., Hofle U., Villanúa D., Gauss C., Segalés J., Almería S., Montoro V., Gortázar C. 2006. Seroprevalence of six reproductive pathogens in European wild boar (Sus scrofa) from Spain: The effect on wild boar female reproductive performance. Theriogenology 65:731-743. Ruiz-Fons F., Vidal D., Hofle U.,Vicente J., Gortazar C. 2007.
Aujeszky's disease virus infection patterns in European wild boar.
Vet Microbiol. 120:241-50.
Sabo A., Rajcani J., Blaskovic D. 1968.
Studies on the pathogenesis of Aujeszky’s disease virus. I.
Distribution of the virulent virus in piglets after peroral infection.
Acta Viral., 12: 214- 221.
Sabo A., Rajcani I., Blaskovic D. 1969.
Studies on the pathogenesis of Aujeszky’s disease virus. III. The
distribution of virulent virus in piglets after intranasal infection. Acta
Virol. 13: 407-414.
Sáez-Royuela C., Tellería J. L. 1986. The increased population of the wild boar (Sus scrofa L.) in Europe. Mammal Review 16: 97-101. Sáez-Royuela C & Tellería J. L. 1987. Reproductive trends of the wild boar (Sus scrofa) in Spain. Folia
Zoologica36: 21-25.
Sanchez R., Nauwynck H., Pensaert M. 2001.

Pagina | 111
Serological survey of porcine circovirus 2 antibodies in domestic and
feral pig population in Belgium. Proceedings of the European
Society for Veterinary Virology.
Santos P., Fernández-Llario P., Fonseca C., Monzón A., Bento P., Soares A. M. V. M., Mateos-Quesada P., Petrucci-Fonseca F. 2006. Habitat and reproductive phenology of wild boar (Sus scrofa) in the western Iberian Peninsula. European Journal of Wildlife Research 52: 207-212. Saunders G., Giles J. 1995. Ecological comparison of two wild pig populations in semi-arid and s ub-alpine Australia. IBEX - Journal of Mountain Ecology 3: 152-155. Schulze C., Segalés J., Neumann G. et al. 2004.
Identification of postweaning multisystemic wasting syndrome in
European wild boar (Sus scrofa). Vet Rec. 154: 694-696.
Segalés J., Marco I., Calsamiglia M. 2002.
Evidence of porcine circovirus type 2 infection in wild boar (Sus
scrofa) in Spain. Congress Ames, Iowa USA.
Servanty S. 2003. Variations des paramètres de la reproduction et de la croissance post- natale: cas d'un ongulé à fécondité élevée, le sanglier (Sus scrofa scrofa). Analyse et modélisation des systèmes biologiques: 30. Servanty S, Gaillard J M, Allaine D, Brandt S & Baubet E (2007) Litter size and fetal sex ratio adjustment in a highly polytocous species:the wild boar. Behavioral Ecology 18: 427-432. Singer F J, Otto D K, Tipton A R & Hable C P. 1981. Home ranges, movements, and habitat use of European wild boar in Tennessee. Journal of Wildlife Management 45: 343-353. Singer F. J., Swank W. T., Clebsch E. E. C. 1984. Effects of wild pig rooting in a deciduous forest. Journal of Wildlife Management 48:464-473. Spillane P., Kennedy S., Meehan B., et al. 1998.
Porcine circovirus infection in the Republic of Ireland. Vet Rec. 143:
511-512.
Spencer P. B. S., Hampton J O. 2005. Illegal translocation and genetic structure of feral pigs in Western Australia. Journal of Wildlife Management 69: 377-384. Spitz F, Gleize J-C., Duncan P. 1990.

Pagina | 112
Particularités de la croissance pondérale du sanglier: cas des populations de Camargue (sud de la France). Mammalia 54: 405- 414. Spitz F, Janeau G., Valet G. 1984. Eléments de démographie du sanglier (Sus scrofa) dans la région de Grésigne. Acta Oecologica, Oecologia Applicata 5: 43-59. Spitz F, Valet G., Brisbin I. L. J. 1998. Variation in body mass of wild boars from Southern France. Journal of Mammalogy 79: 251-259. Sweeney J. M., Probost E. E., Sweeney J. R. 1979. Reproductive biology of a feral hog population. Journal of Wildlife Management 43:555-559. Teillaud P. 1986. La vie sociale chez le sanglier: Co-émergence collectivité - individualité. Cahiers d'Ethologie appliquée 6: 157-184. Tellería J. L., Sáez-Royuela C.1985. L'évolution démographique du sanglier (Sus scrofa) en Espagne: Gestion du sanglier. Mammalia 49: 195-202. Toro M. A., Fernandez A., Garcia-Cortes L. A., Rodriganez J., Silio L. 2006. Sex ratio variation in Iberian pigs. Genetics 173: 911-917.
Tozzini F., Poli A., Della Croce G. 1982.
Experimental infection of european wild swine (Sus scrofa) with
pseudorabies virus. J. Wildlife Dis. 18: 425-428
Uzal A & Nores C. 2002. Endogenous and exogenous constraints in the population changes of wild boar (Sus scrofa Linnaeus, 1758). Wild boar research 2002. A selection and edited papers from the "4th international wild boar symposium", Lousa, Portugal: 83-98. Varuzza P. 2005. Il Capriolo: biologia e gestione. Edt. Geographica,308 pp Verin R., Ressel L. , Nigrelli D.Brocchi E. , Faccini S. , Varuzza P. , Poli A. 2006. Pseudorabies virus in wild boar (Sus scrofa) from central Italy. Atti VII Conference of European Wildlife Disease Association. p. 17. Saint.Vincent 27-30 Settembre 2006. Verin R. , Ricci M. , Taccini E. , Poli A. 2007. Diffusione di PCV-2 nella popolazione di cinghiali (Sus Scrofa) delle Provincie di Grosseto e Pisa. Atti IV CongressoNazionale AIPvet, Alberese(GR) 24-25 Maggio 2007

Pagina | 113
Vicente J., Ruiz-Fons F., Vidal D., Höfle U., Acevedo P., Villanúa D., Fernández-de-Mera I. G., Martin M. P., Gortázar C. 2005. Serosurvey of Aujezky's disease virus infection in European wild boar in Spain. Veterinary record 156: 408-412. Warwick B.L., 1928.
Prenatal growth ofswine. Journal of morphology and physiology 46:
59-84.
Wittmann G., Jakubik J., Ahl R. 1980.
Multiplication and distribution of Aujeszky’s disease (pseudo-rabies)
virus in vaccinated and non-vaccinated pigs after intranasal
infection. Arch. Virol. 66: 227-240.
Wittmann G., Ohlinger V., Rziha H.J. 1983.
Occurrence and reactivation of latent Aujeszky’s disease virus
following challenge in previously vaccinated pigs. Arch Virol.
75: 29-41.
Workman J., Abmayr S., Cromlish W., Roeder. R. 1988.
Transcriptional regulation by the immediate-early protein of
pseudorabies virus during in vitro nucleosome assembly. Cell. 55: 21
l-219.
Zhang G., Stevens R., Leader D.P. 1990.
The protein kinase encoded in the short unique region of
pseudorabies virus: Description of the gene and identification
of its product in virions and in infected cells. J. Gen. Viral. 71: 1757-
1765.
Zsak L., Zuckermann F., Sugg N., Ben-Porat T. 1992.
Glycoprotein g1 of pseudorabies virus promotes cell fusion and
virus spread via direct cell-to-cell transmission. J. Virol. 66: 2316-
2325.
Zsak L., Mettenleiter T.C., Sugg N., Ben-Porat T. 1989.
Release of pseudorabies virus from infected cells is controlled by
several viral functions and is modulated by cellular components. J.
Virol. 63:5475-5477.
Zuckerman F.A., Zsak L., Mettenleiter T.C., Ben-Porat T. 1990.

Pagina | 114
Pseudorabies virus glycoprotein gII1 is a major target for murine
and swine virus-specific cytotoxic T-lymphocytes. J. Virol. 64: 802-
812.

Pagina | 115
CAPITOLO 10 RINGRAZIAMENTI

Pagina | 116
10. RINGRAZIAMENTI Un grosso aiuto al campionamento è stato dato dal Dott. Ranieri VERIN e
dalla Dott.ssa Daniela GIUSTINI, le analisi sierologiche e virologiche sono
frutto del prezioso aiuto dei Dott. Ranieri Verin e Maurizio Mazzei.
Si ringraziano, gli Ambiti Territoriali di Caccia della provincia di Pisa: ATC 14
e 15, il Dr. Vito Mazzarone Responsabile dell’Ufficio Fauna della Provincia,
e le squadre di caccia al cinghiale della provincia Pisa, in particolare i Sigg.
Silvio Azara,
Ivanio Berti,
Lorenzo Biagiotti
Giorgio Bocelli,
Paolo Buselli,
Egiziano Cheli,
Giulio Cheli,
Riccardo Donati,
Sergio Fabbri,
Massimo Giannelli,
Ercoli Maurizio,
Ivo Giusti,
Alfiero Manetti,
Marco Martini,
Maurizio Mariotti,
Giorgio Nerli,
Carlo Panizzo,
Francesco Prato
Marco Rossi,
Michele Ruotolo,
Francesco Salutini,
Gabriele Stefanucci,
Andrea Vaghetti,
Alessandro Valori,
Andrea Vivarelli,
Un grazie particolare va ad Antonio Felicioli per avermi coinvolto e
incoraggiato in questo progetto ed al prof. Poli per aver supportato dal
primo momento questo studio fornendo spunti e preziosi suggerimenti.
Recommended

















