Embed Size (px)
Citation preview

Amino acids

AMINO ACIDS
They are molecules containing an amine group, a carboxylic acid group, and a side-chain that is specific to each amino acid.
The key elements of amino acid are carbon, hydrogen, oxygen, and nitrogen.
Amino acids are the basic structural building units of protein and other biomolecules; they are also utilized as an energy source.
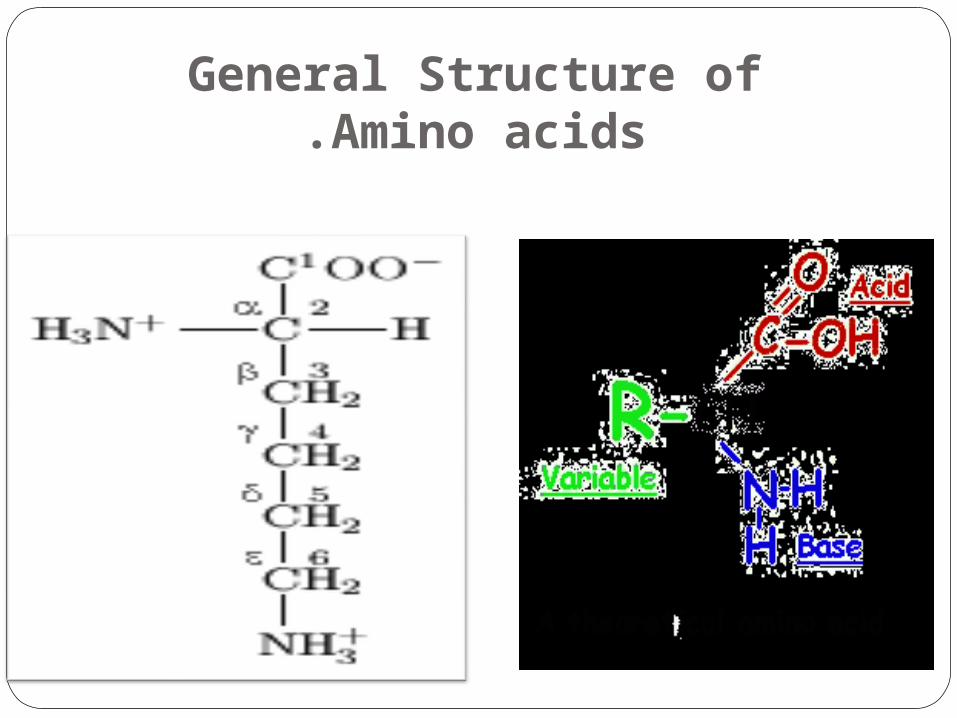
General Structure of Amino acids.

General Classification of Amino acids
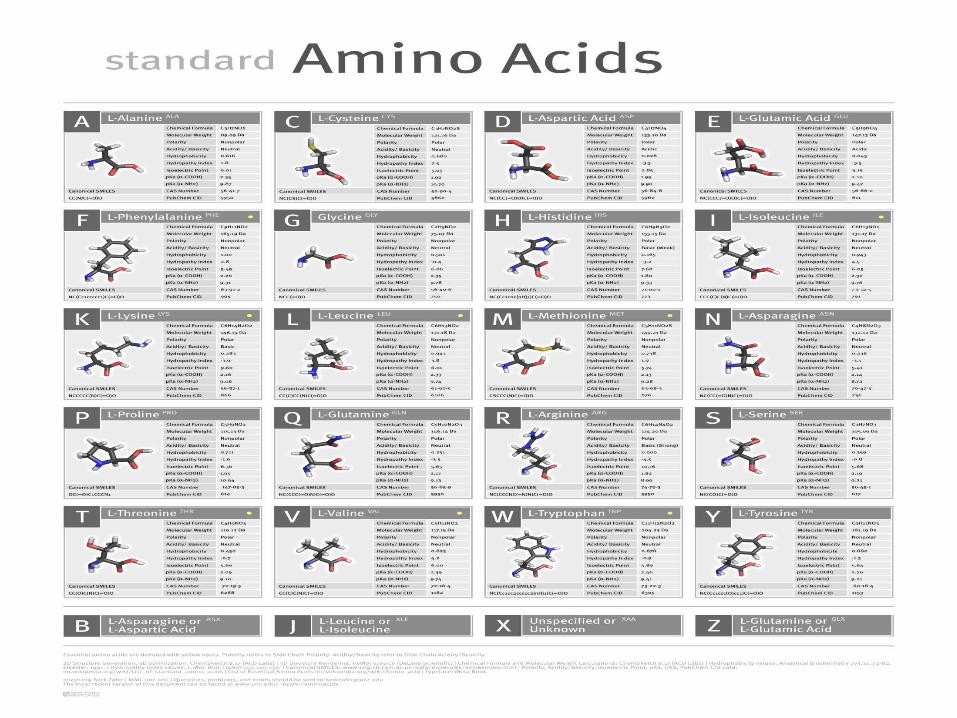
Standard amino acids
Non-standard amino acids

Standard Amino Acids
Amino acids join together to form short polymer chains called peptides or longer chains called either polypeptidesor proteins.
These polymers are linear and unbranched, with each amino acid within the chain attached to two neighboring amino acids.
Twenty-two amino acids are naturally incorporated into polypeptides and are called proteinogenic or natural amino acids. Of these, 20 are encoded by the universal genetic code. The remaining 2 are incorporated into proteins by unique synthetic mechanisms.

Non-standard amino acids
Aside from the 22 standard amino acids, there are many other amino acids that are called non-proteinogenic or non-standard.
They are either not found in proteins (for example carnitine, GABA), or are not produced directly.
Non-standard amino acids that are found in proteins are formed by post-translational modification, which is modification after translation during protein synthesis.
These modifications are often essential for the function or regulation of a protein; for example, the carboxylation of glutamate allows for better binding of calcium cations.

Some nonstandard amino acids are not found in proteins. Examples include lanthionine, 2-aminoisobutyric acid, dehydroalanine, and the neurotransmitter gamma-aminobutyric acid.
Nonstandard amino acids often occur as intermediates in the metabolic pathways for standard amino acids — for example, ornithine and citrulline occur in the urea cycle.

In human nutrition
When taken up into the human body from the diet, the 22 standard amino acids either are used to synthesize proteins and other biomolecules or are oxidized to urea and carbon dioxide as a source of energy.
The oxidation pathway starts with the removal of the amino group by a transaminase, the amino group is then fed into the urea cycle. The other product of transamidation is a keto acid that enters the citric acid cycle.
Glucogenic amino acids can also be converted into glucose, through gluconeogenesis.

Difference between essential and non-essential amino
acids
There are 20 different amino that make up all proteins in the human body.
These amino acids are needed to replenish tissue, red blood cells, enzymes, and other substances.
9 - 12 can be manufactured by the body-nonessential amino acids, not obtained from the diet.
The remaining 8 to 11 -essential amino acids, must be obtained from the diet.

Essential NonessentialHistidine AlanineIsoleucine Arginine*Leucine AsparagineLysine Aspartic acidMethionine Cysteine*Phenylalanine Glutamic acidThreonine Glutamine*Tryptophan GlycineValine Ornithine*
Proline*Selenocysteine*Serine*Taurine*Tyrosine*

Non-protein functions
Many amino acids are used to synthesize other molecules, for example:
Tryptophan is a precursor of the neurotransmitter serotonin.
Tyrosine is a precursor of the neurotransmitter dopamine.
Glycine is a precursor of porphyrins such as heme.
Arginine is a precursor of nitric oxide.Aspartate, glycine, and glutamine are precursors
of nucleotides.

Classification according to functions
Anabolic/Catabolic Responses and Tissue pH Regulation :- Glutamic AcidGlutamine
The Urea Cycle and Nitrogen ManagementArginineCitrullineOrnithineAspartic AcidAsparagine

Essential Amino Acids for Proteins and EnergyIsoleucineLeucineValineThreonineHistidineLysineAlpha-Aminoadipic Acid
Sulfur Containing Amino Acids for Methylation and GlutathioneMethionineCystineHomocysteineCystathionineTaurine

Neurotransmitters and PrecursorsPhenylalanineTyrosineTryptophanAlpha-Amino-N-Butyric AcidGamma-Aminobutyric Acid
Methylhistidines1-methylhistidine3-methylhistidine

Precursors to Heme, Nucleotides and Cell MembranesGlycineSerineSarcosineAlanineEthanolaminePhospethanolaminePhosphoserine
Bone Collagen Specific Amino AcidsProlineHydroxyprolineHydroxylysine
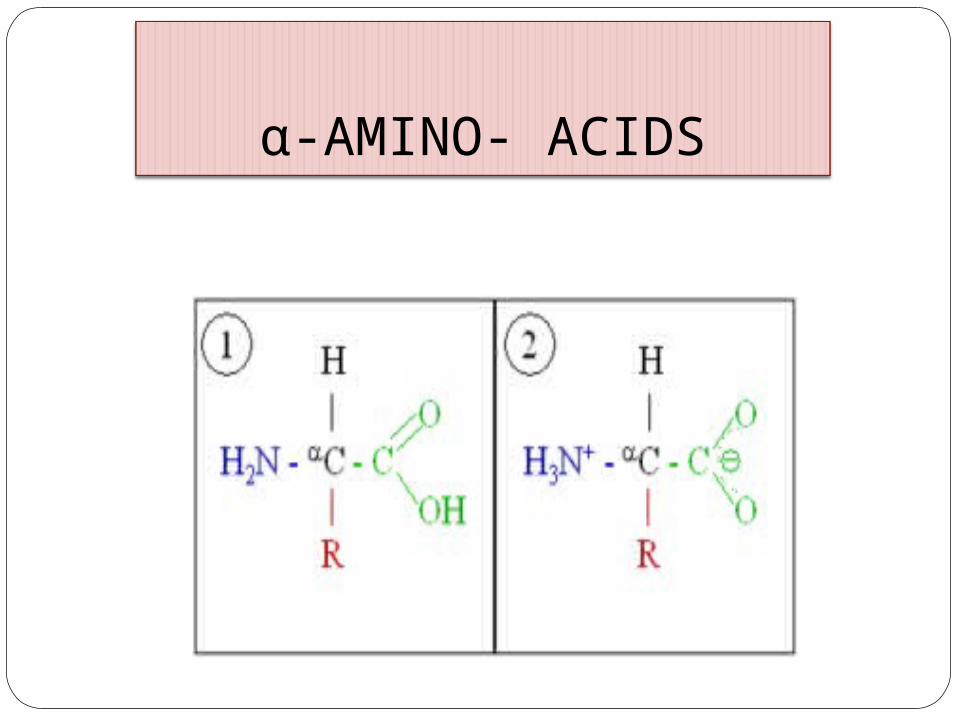
α-AMINO- ACIDS
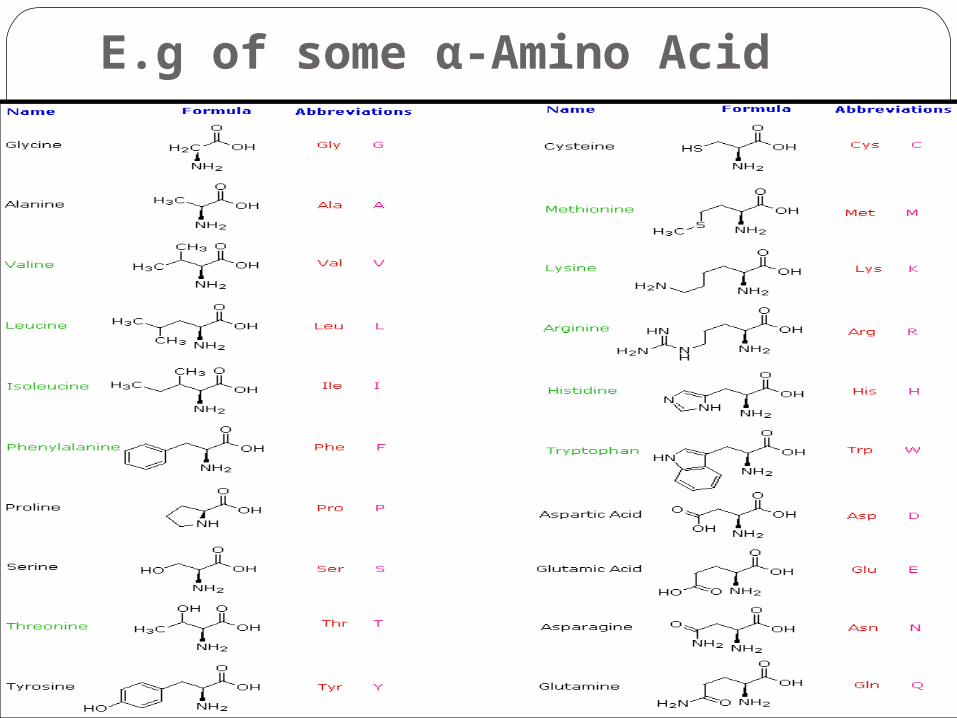
E.g of some α-Amino Acid
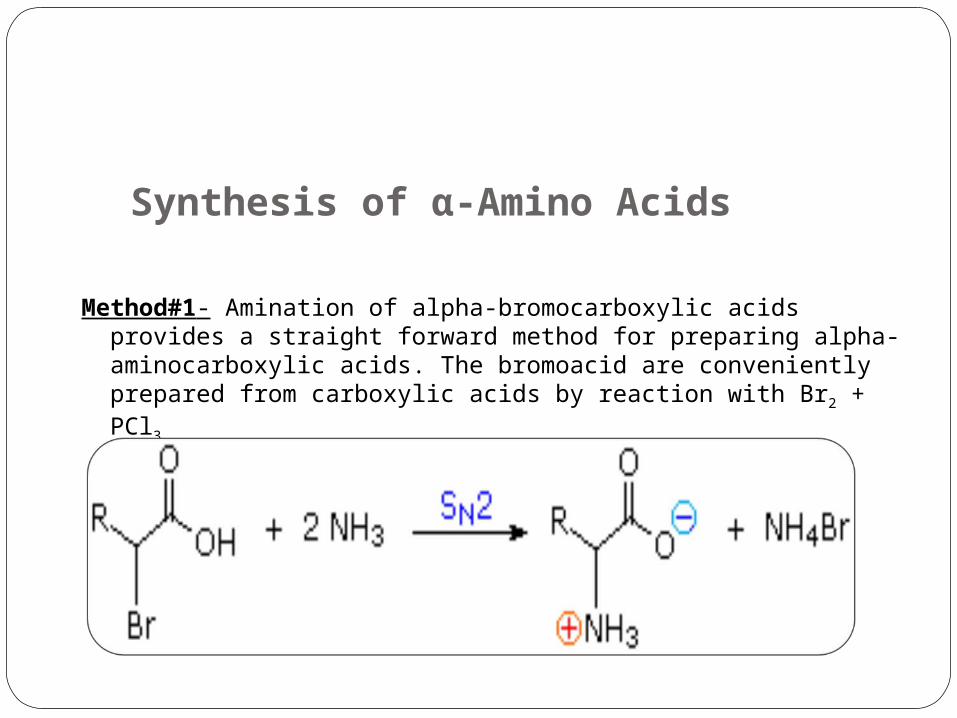
Synthesis of α-Amino Acids
Method#1- Amination of alpha-bromocarboxylic acids provides a straight forward method for preparing alpha- aminocarboxylic acids. The bromoacid are conveniently prepared from carboxylic acids by reaction with Br2 + PCl3.
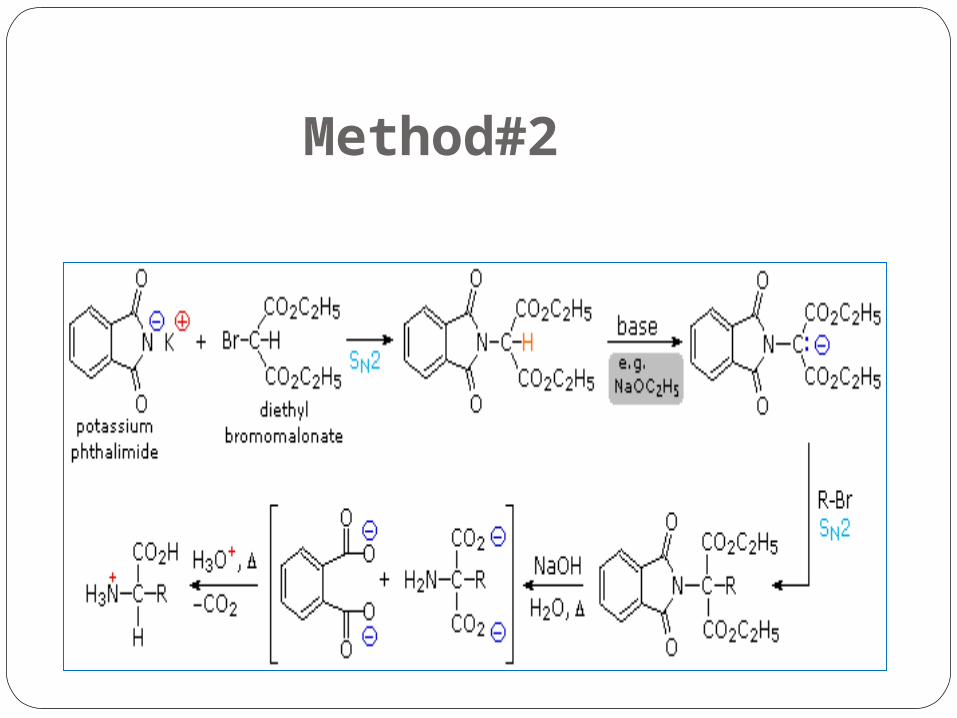
Method#2

Explaination of Method #2
By modifying the nitrogen as a phthalimide salt the propensity of amines to undergo multiple substitutions is removed, and a single clean substitution reaction of 1º- and many 2º-alkylhalides takes place.
This procedure is known as the Gabriel synthesis, it can be used to advantage in aminating bromomalonic esters, as shown in the upper equation of the following scheme.
Since the phthalimide substituted malonic ester has an acidic hydrogen (colored orange) activated by the two ester groups, this intermediate may be converted to an ambident anion and alkylated.
Finally, base catalyzed hydrolysis of the phthalimide moiety and the esters, followed by acidification and thermal decarboxylation, produces an amino acid and phthalic acid (not shown).
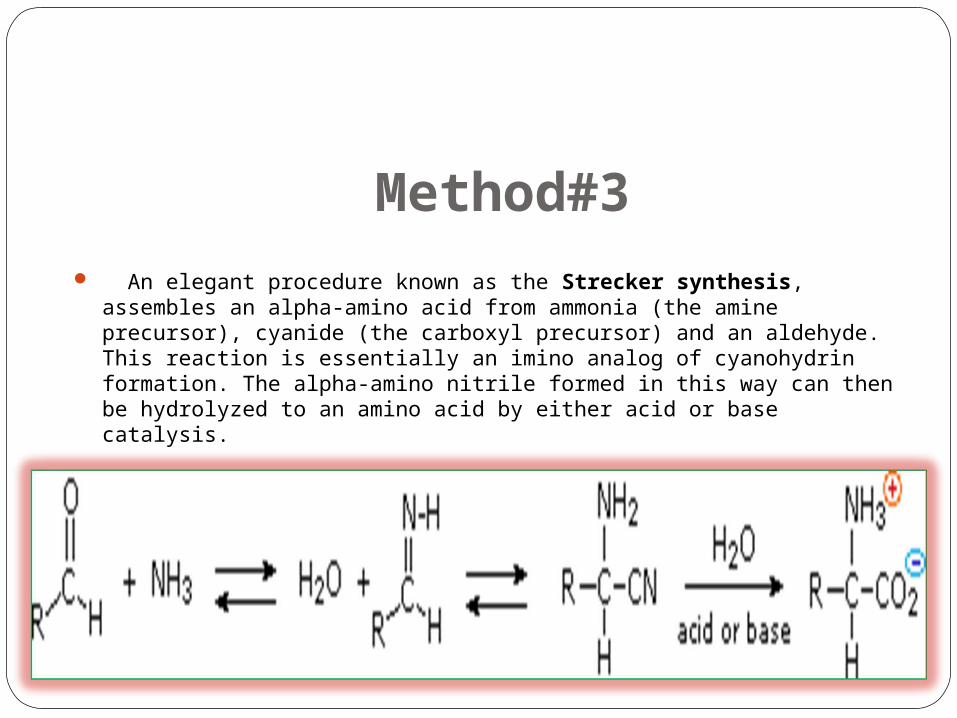
Method#3 An elegant procedure known as the Strecker synthesis, assembles an alpha-amino acid from
ammonia (the amine precursor), cyanide (the carboxyl precursor) and an aldehyde. This reaction is essentially an imino analog of cyanohydrin formation. The alpha-amino nitrile formed in this way can then be hydrolyzed to an amino acid by either acid or base catalysis.
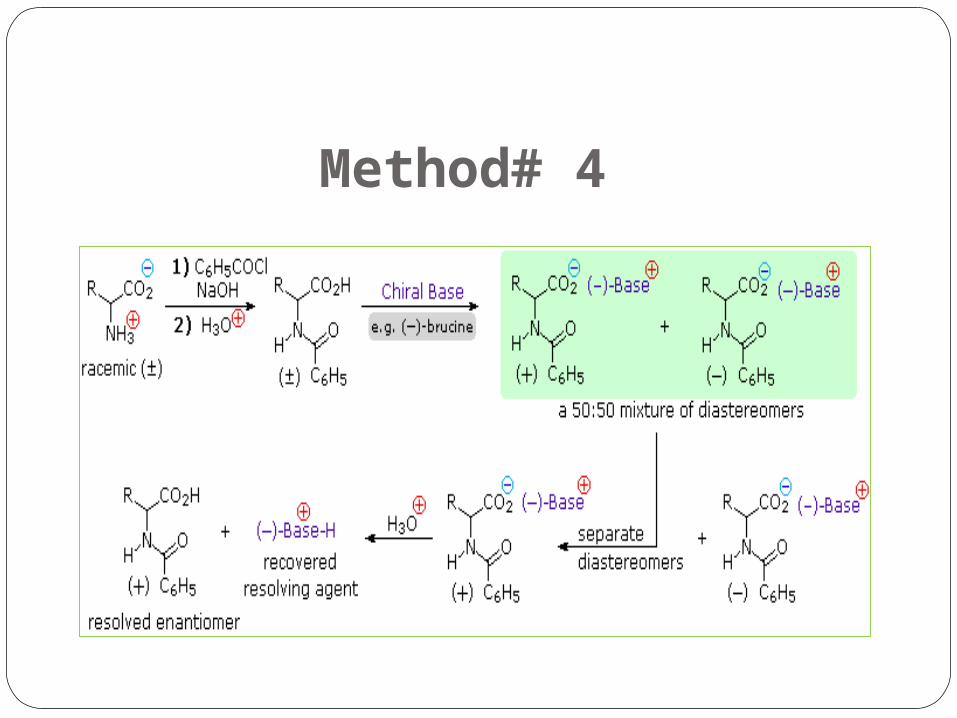
Method# 4

Explaination of Method # 4 Resolution The three synthetic procedures described above and many others that
can be conceived, give racemic amino acid products. If pure L or D enantiomers are desired, it is necessary to resolve these racemic mixtures.
A common method of resolving racemates is by diastereomeric salt formation with a pure chiral acid or base.
This is illustrated for a generic amino acid in the following diagram. Be careful to distinguish charge symbols shown in colored circles, from optical rotation signs shown in parenthesis.

In the initial display, the carboxylic acid function contributes to diastereomeric salt formation. The racemic amino acid is first converted to a benzamide derivative to remove the basic
character of the amino group. Next, an ammonium salt is formed by combining the carboxylic acid with an optically pure
amine, such as brucine (a relative of strychnine). The structure of this amine is not shown, because it is not a critical factor in the logical
progression of steps. Since the amino acid moiety is racemic and the base is a single enantiomer (levorotatory in this
example), an equimolar mixture of diastereomeric salts is formed (drawn in the green shaded box).
Diastereomers may be separated by crystallization, chromatography or other physical manipulation and in this way one of the isomers may be isolated for further treatment, in this illustration it is the (+):(-) diastereomer.
Finally the salt is broken by acid treatment, giving the resolved (+)-amino acid derivative together with the recovered resolving agent (the optically active amine). Of course, the same procedure could be used to obtain the (-)-enantiomer of the amino acid.

Since amino acids are amphoteric, resolution could also be achieved by using the basic character of the amine function. For this approach we would need an enantiomerically pure chiral acid such as tartaric acid to use as the resolving agent.
Note that the carboxylic acid function is first esterified, so that it will not compete with the resolving acid.
Resolution of aminoacid derivatives may also be achieved by enzymatic discrimination in the hydrolysis of amides. For example, an aminoacylase enzyme from pig kidneys cleaves an amide derivative of a natural L-amino acid much faster than it does the D-enantiomer.
If the racemic mixture of amides shown in the green shaded box above is treated with this enzyme, the L-enantiomer (whatever its rotation) will be rapidly converted to its free zwitterionic form, whereas the D-enantiomer will remain largely unchanged.
the diastereomeric species are transition states rather than isolable intermediates. This separation of enantiomers, based on very different rates of reaction is called kinetic
resolution.
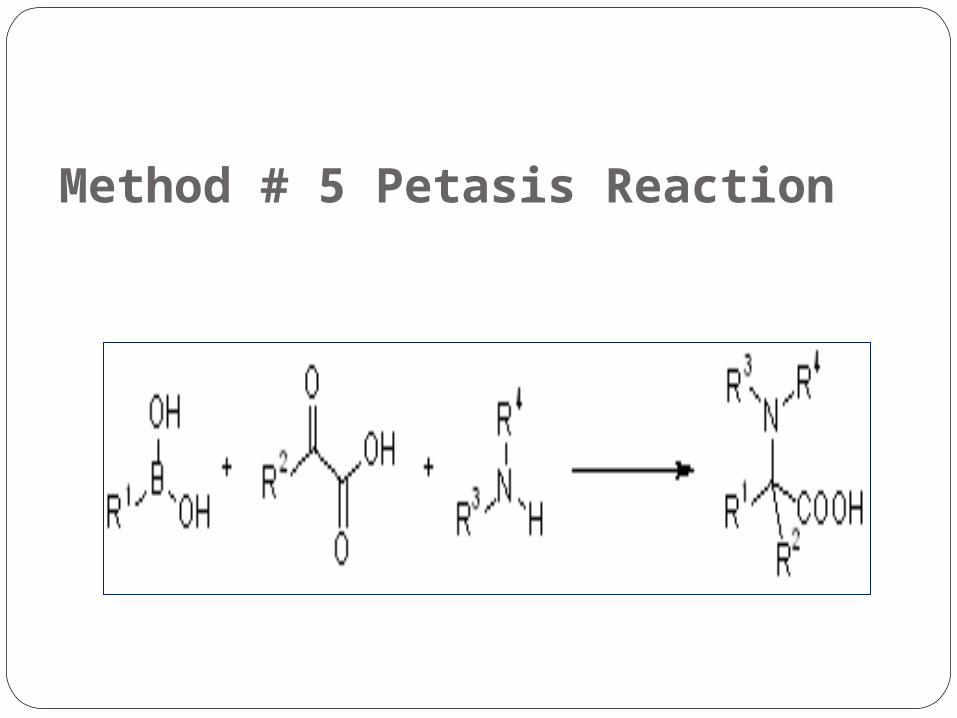
Method # 5 Petasis Reaction
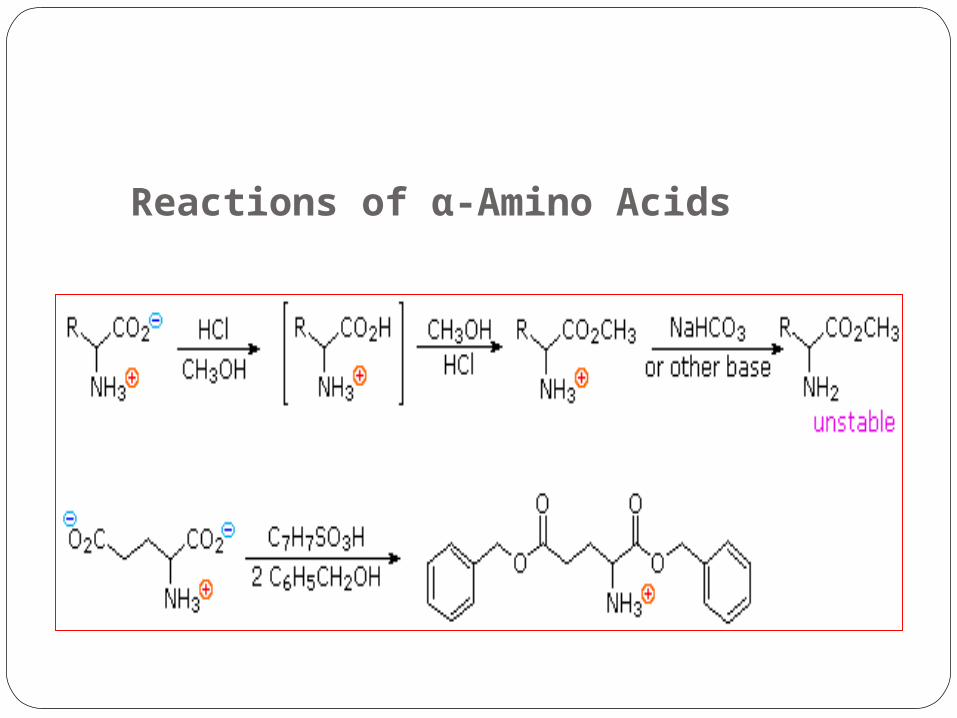
Reactions of α-Amino Acids
.1Carboxylic Acid Esterification.
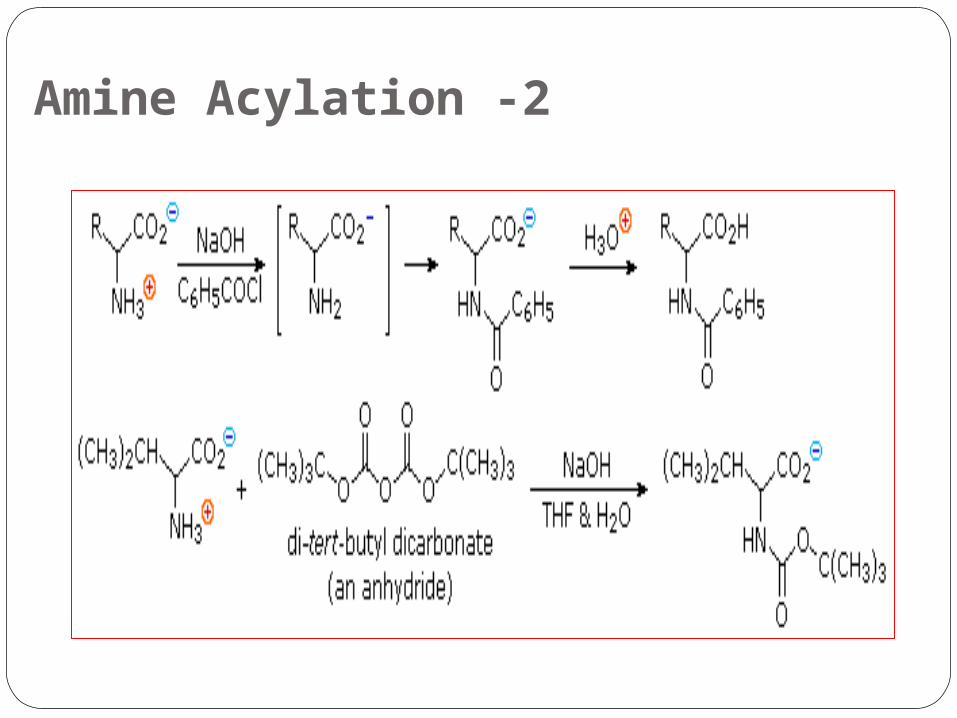
2 -Amine Acylation
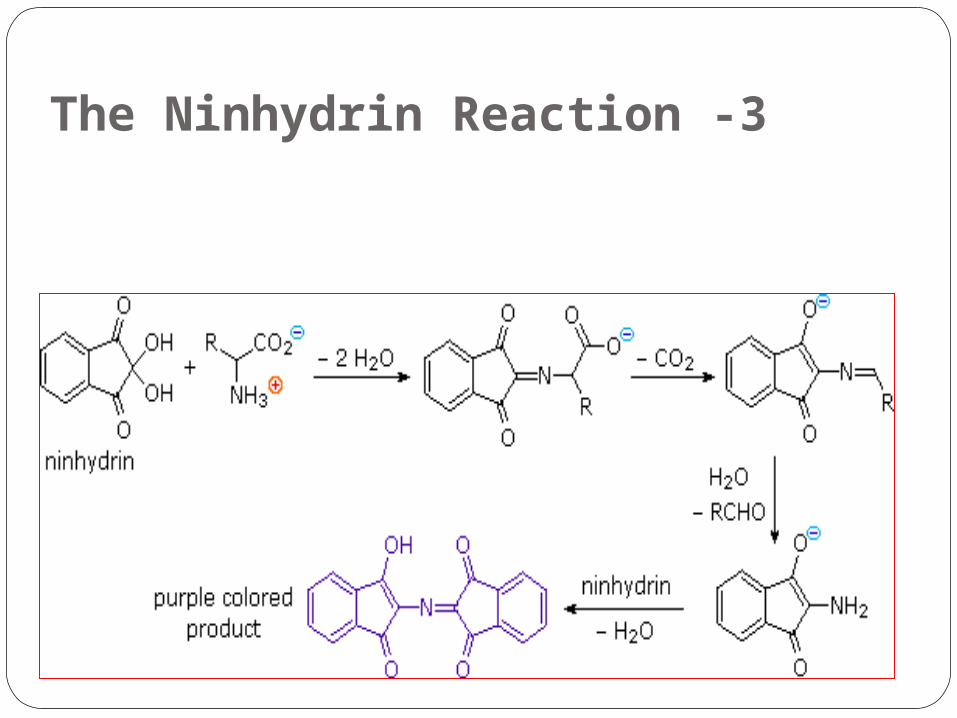
3 -The Ninhydrin Reaction
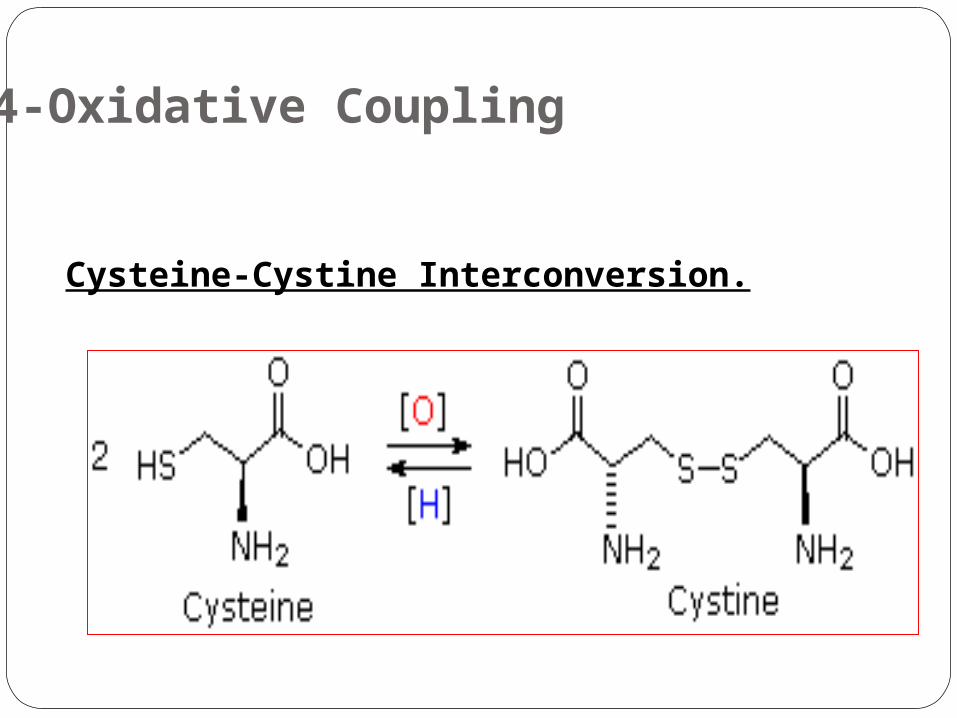
4-Oxidative Coupling
Cysteine-Cystine Interconversion.
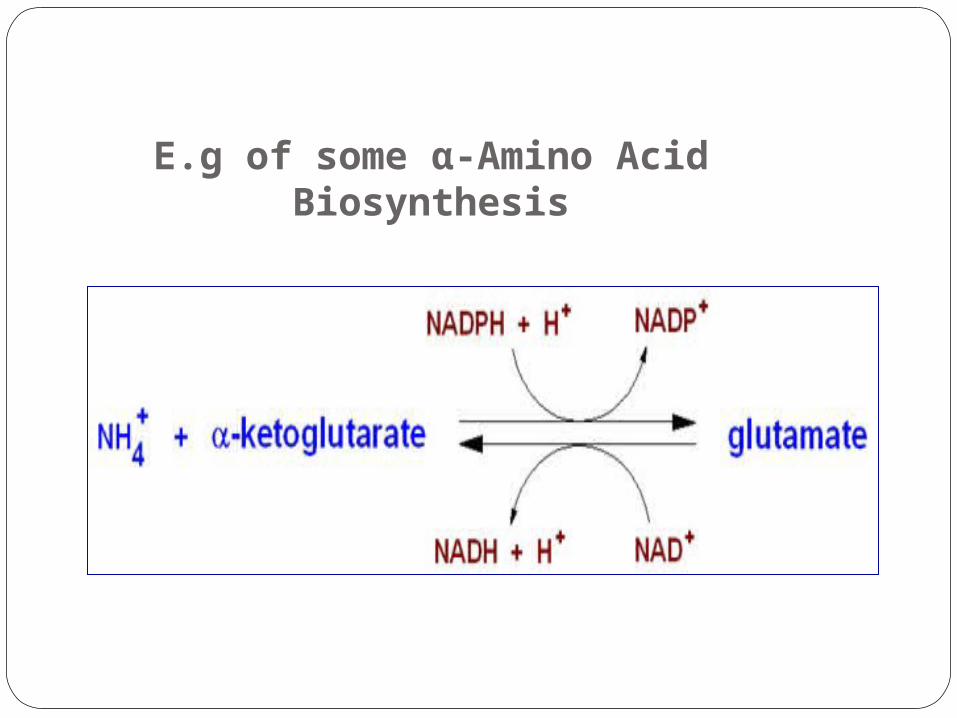
E.g of some α-Amino Acid Biosynthesis
Glutamate.
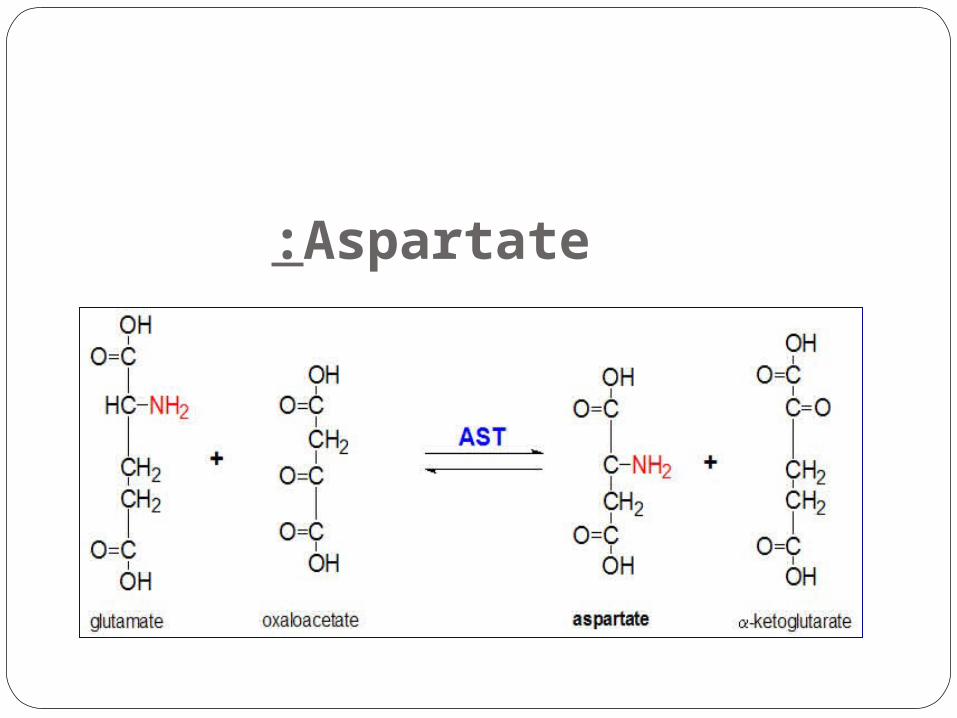
Aspartate:
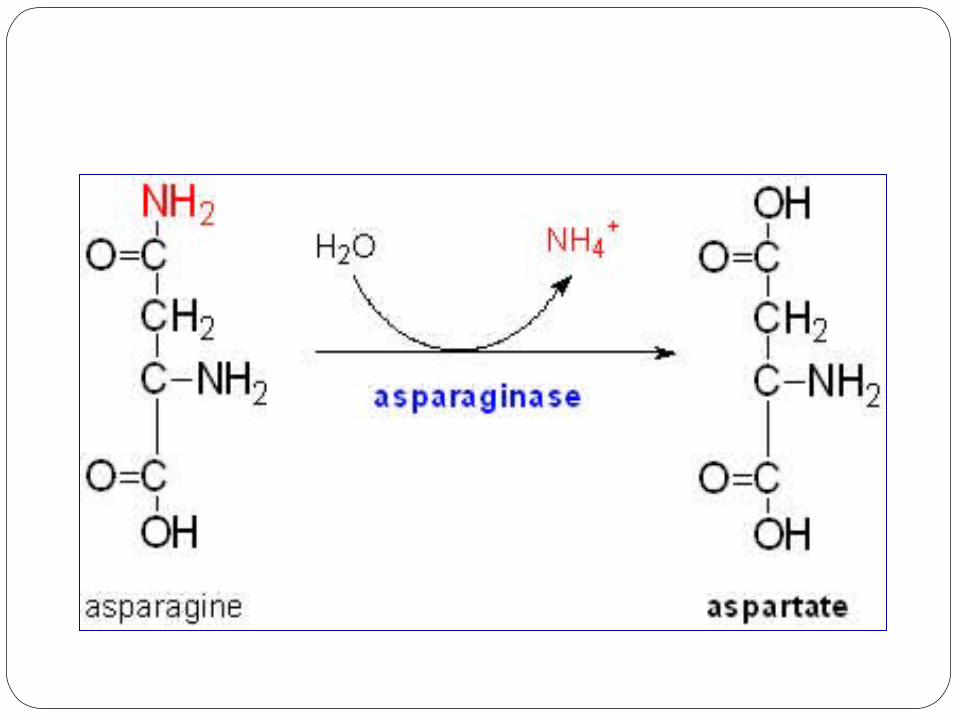
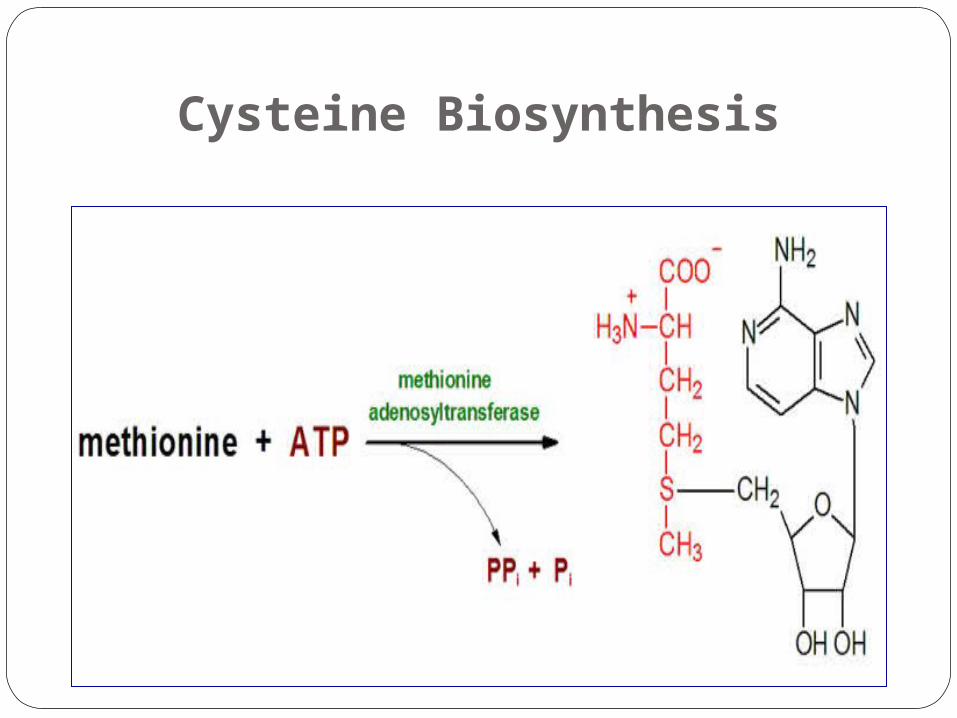
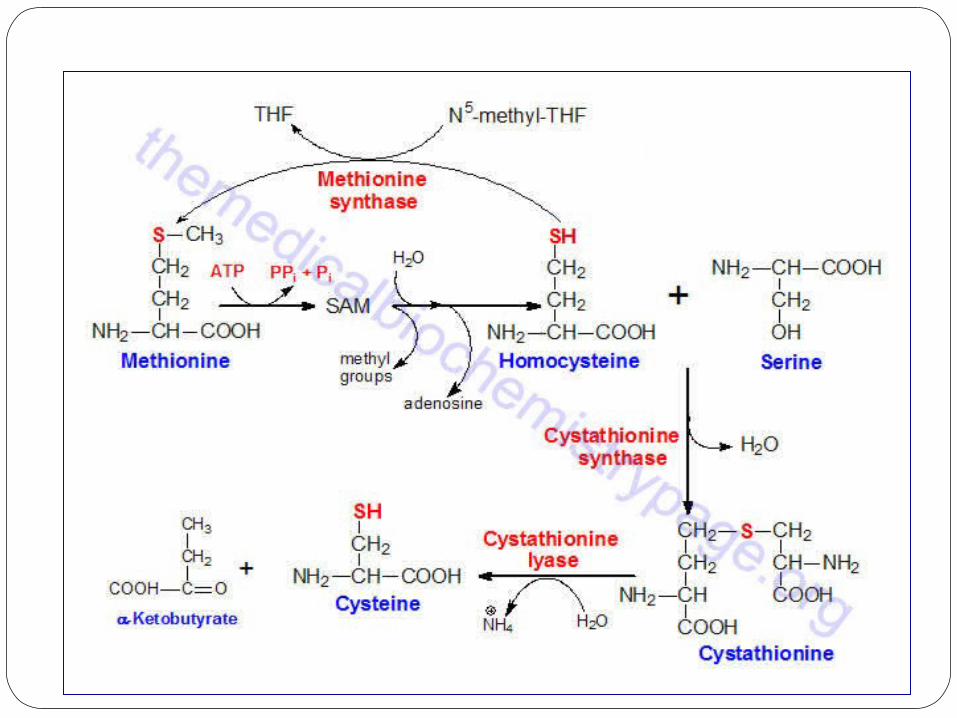
Cysteine Biosynthesis
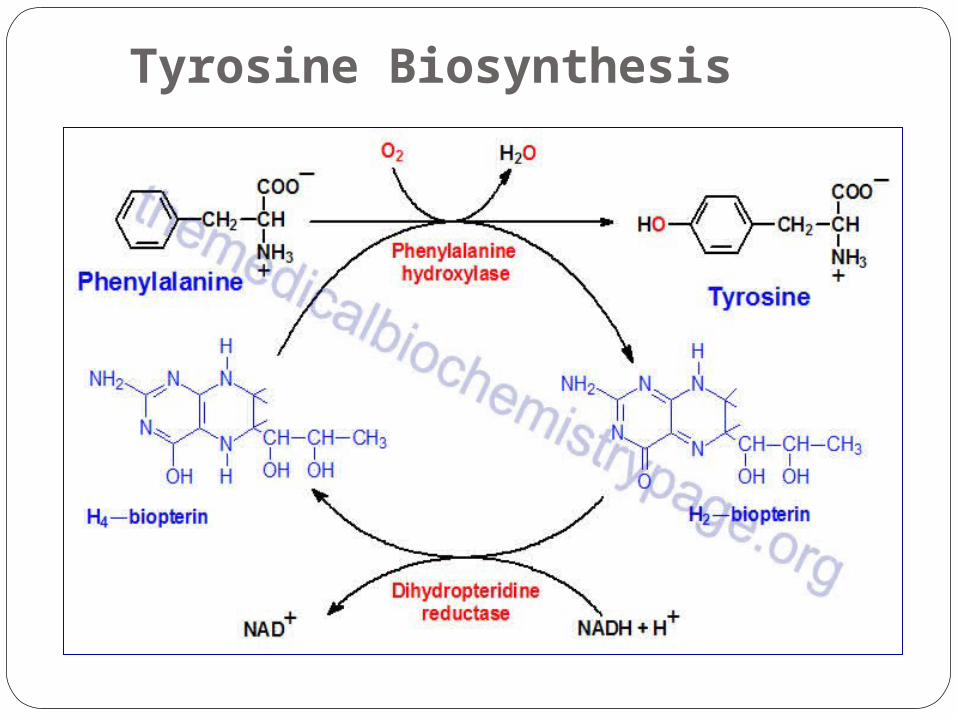
Tyrosine Biosynthesis
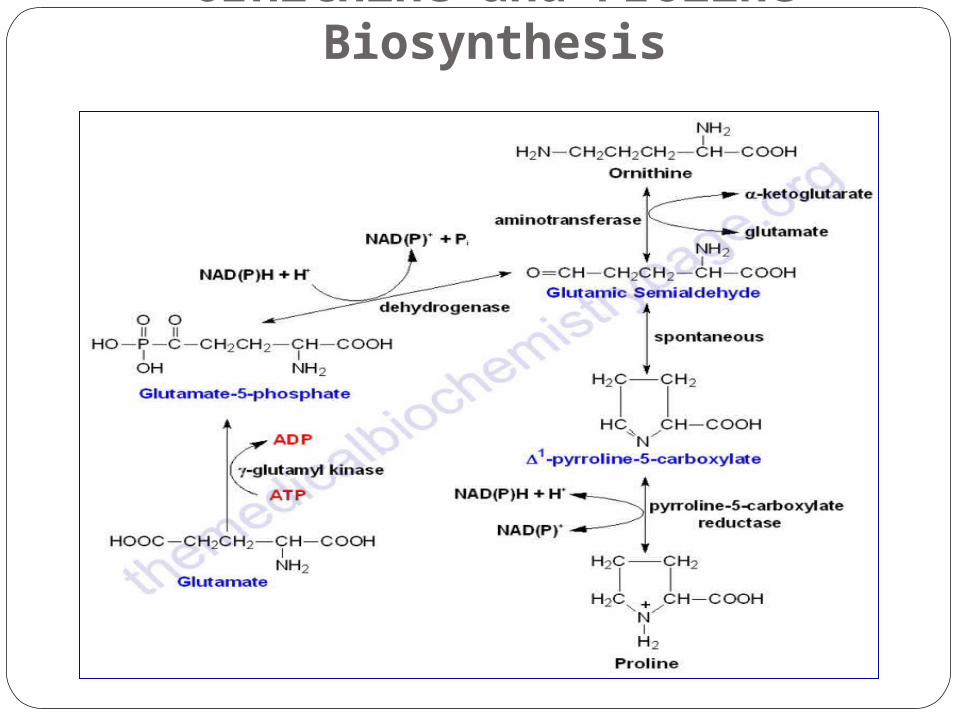
Ornithine and Proline Biosynthesis
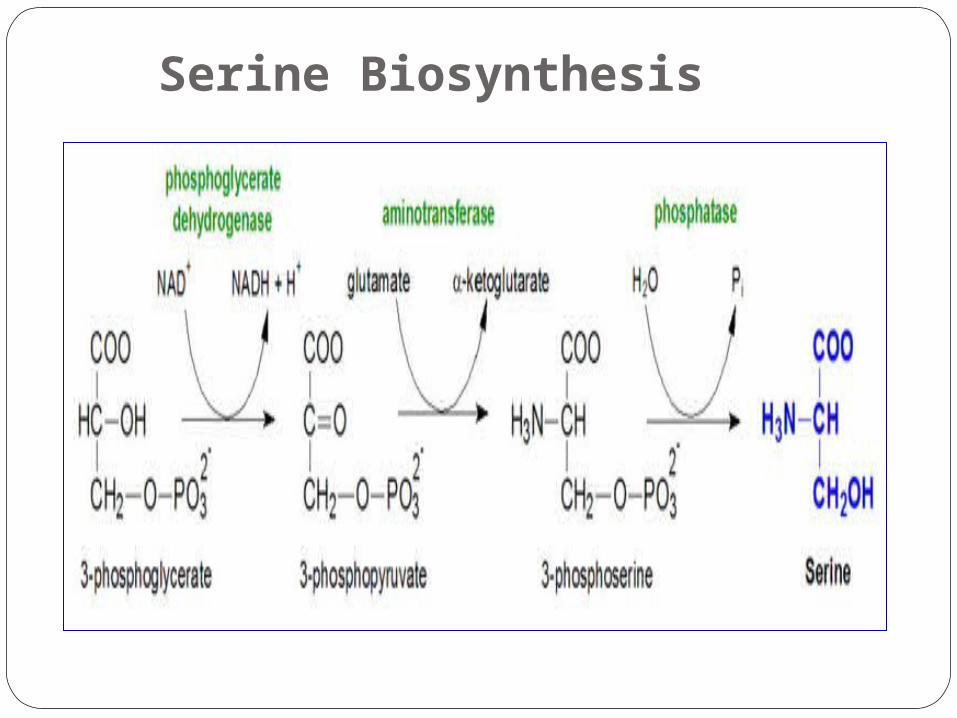
Serine Biosynthesis
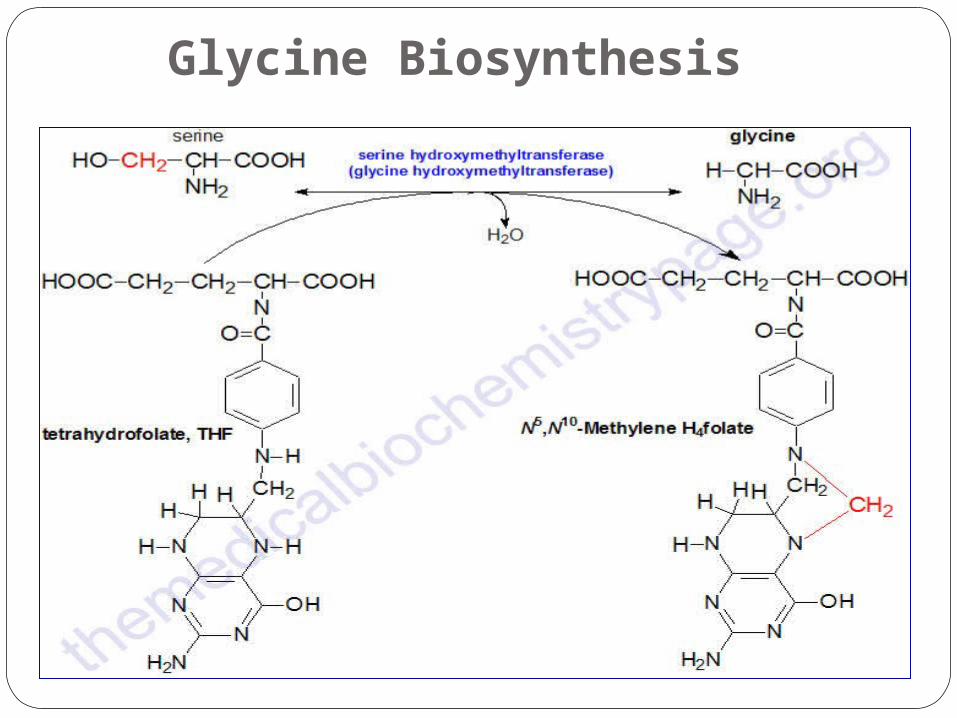
Glycine Biosynthesis

Beta Amino Acid
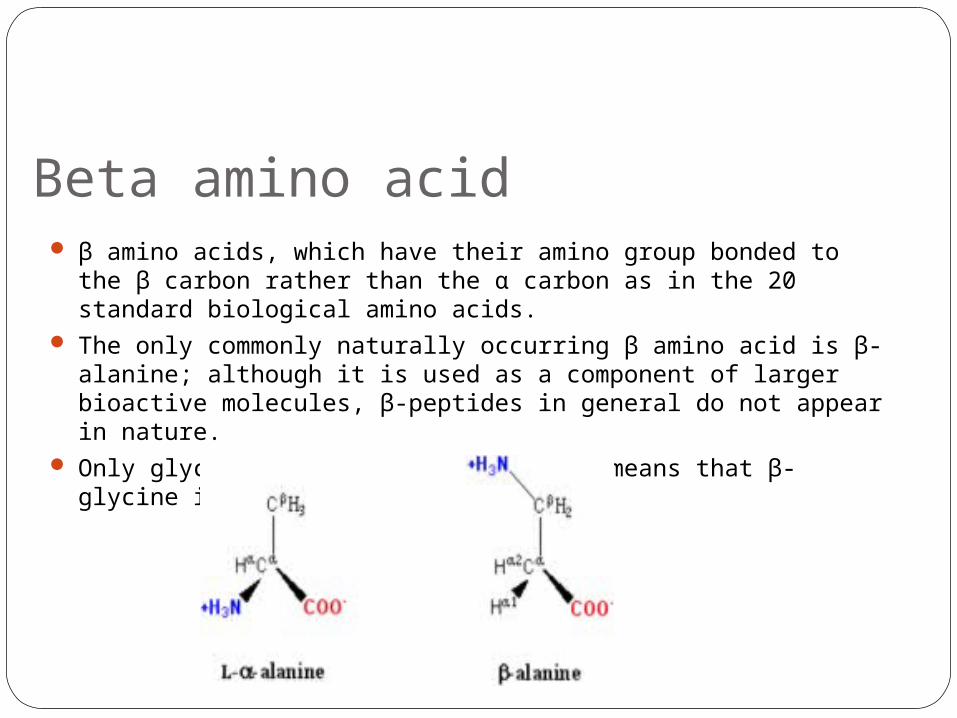
Beta amino acid β amino acids, which have their amino group bonded to the β
carbon rather than the α carbon as in the 20 standard biological amino acids.
The only commonly naturally occurring β amino acid is β-alanine; although it is used as a component of larger bioactive molecules, β-peptides in general do not appear in nature.
Only glycine lacks a β carbon, which means that β-glycine is not possible.

Chemical synthesis
The chemical synthesis of β amino acids can be challenging, especially given the diversity of functional groups bonded to the β carbon and the necessity of maintaining chirality.
In the alanine molecule shown, the β carbon is achiral; however, most larger amino acids have a chiral atom.
A number of synthesis mechanisms have been introduced to efficiently form β amino acids and their derivatives notably those based on the Arndt-Eistert synthesis (A method of increasing the length of an aliphatic acid by one carbon by reacting diazomethane with acid chloride).
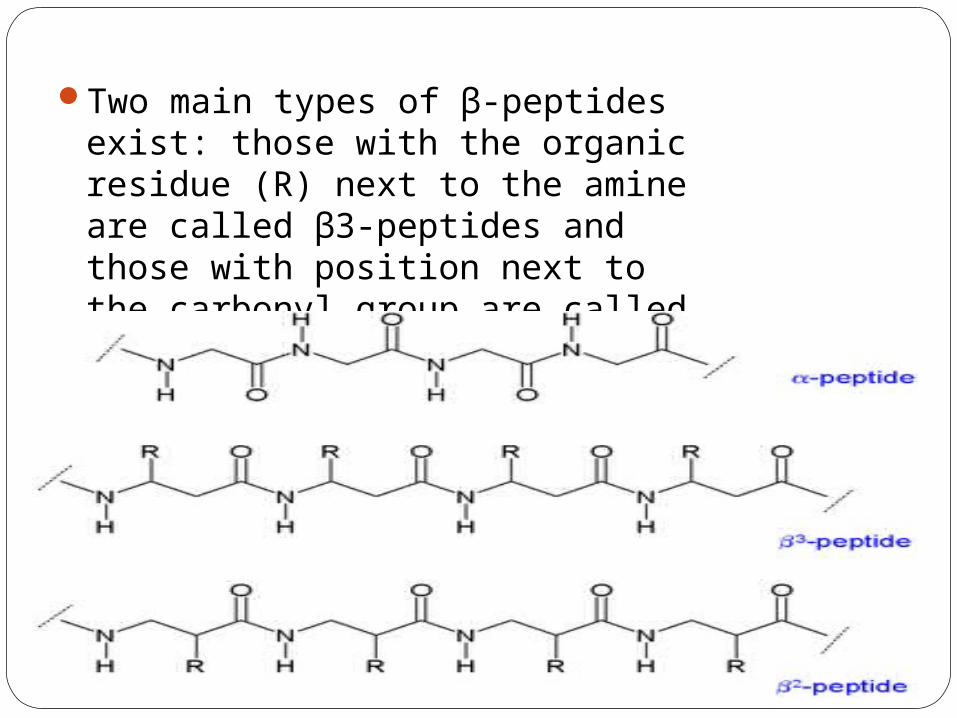
Two main types of β-peptides exist: those with the organic residue (R) next to the amine are called β3-peptides and those with position next to the carbonyl group are called β2-peptides.

Secondary structureBecause the backbones of β-peptides are longer than those of peptides that consist of α-amino acids, β-peptides form different secondary structures. The alkyl substituents at both the α and β positions in a β amino acid favor a gauche conformation about the bond between the α-carbon and β-carbon. Many types of helix structures consisting of β-peptides have been reported. These conformation types are distinguished by the number of atoms in the hydrogen-bonded ring that is formed in solution; 8-helix, 10-helix, 12-helix, 14-helix, and 10/12-helix have been reported. Generally speaking, β-peptides form a more stable helix than α-peptides.

Arndt–Eistert reactionThe Arndt-Eistert synthesis is a series of
chemical reactions designed to convert a carboxylic acid to a higher carboxylic acid homologue (i.e. contains one additional carbon atom) and is considered a homologation process.
Arndt-Eistert synthesis is a popular method of producing β-amino acids from α-amino acids. Acid chlorides react with diazomethane to give diazoketones. In the presence of a nucleophile (water) and a metal catalyst (Ag2O), diazoketones will form the desired acid homologue.

While the classic Arndt-Eistert synthesis uses thionyl chloride to convert the starting acid to an acid chloride, any procedure can be used that will generate an acid chloride.3 RCOOH + PCl3(phosphorus trichloride) → 3 RCOCl +
H3PO3RCOOH + PCl5(phosphorus pentachloride) → RCOCl +
POCl3 + HCl
Diazoketones are typically generated as described here, but other methods such as diazo-group transfer can also apply.
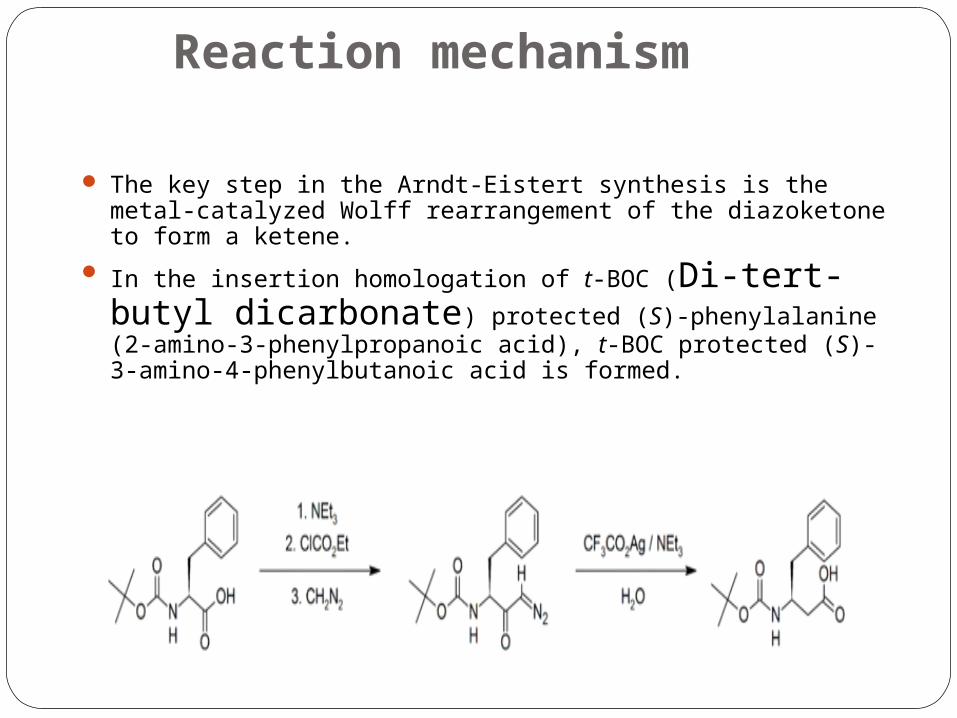
Reaction mechanism
The key step in the Arndt-Eistert synthesis is the metal-catalyzed Wolff rearrangement of the diazoketone to form a ketene.
In the insertion homologation of t-BOC (Di-tert-butyl dicarbonate) protected (S)-phenylalanine (2-amino-3-phenylpropanoic acid), t-BOC protected (S)-3-amino-4-phenylbutanoic acid is formed.

Wolff rearrangement of the α-diazoketone intermediate forms a ketene via a 1,2-rearrangement, which is subsequently hydrolysed to form the carboxylic acid. The consequence of the 1,2-rearrangement is that the methylene group α- to the carboxyl group in the product is the methylene group from the diazomethane reagant.
It has been demonstrated that the rearrangement preserves the stereochemistry of the chiral centre as the product formed from t-BOC protected (S)-phenylalanine retains the (S) stereochemistry
Heat, light, platinum, silver, and copper salts will also catalyze the Wolff rearrangement to produce the desired acid homologue.
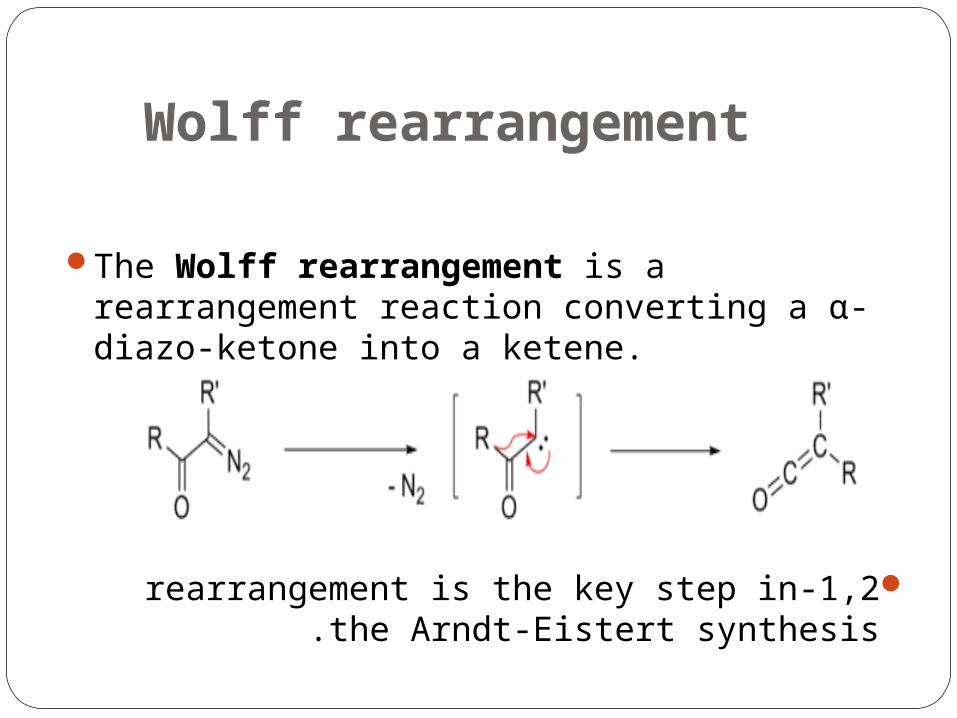
Wolff rearrangement
The Wolff rearrangement is a rearrangement reaction converting a α-diazo-ketone into a ketene.
1,2-rearrangement is the key step in the Arndt-Eistert synthesis .

Clinical potential
β-peptides are stable against proteolytic degradation in vitro and in vivo, an important advantage over natural peptides in the preparation of peptide-based drugs.
β-Peptides have been used to mimic natural peptide-based antibiotics such as magainins (A family of peptides with broad-spectrum antimicrobial activity has been isolated from the skin of the African clawed frog Xenopus laevis), which are highly potent but difficult to use as drugs because they are degraded by proteolytic enzymes in the body.

BETA ALANINEAlso known as 3-aminopropanoic acid,It is a non-essential amino acid and is the
only naturally occurring beta-amino acid. Not to be confused with alanine, beta- alanine
is classified as a non-proteinogenic amino acid as it is not used in the building of proteins.

β-Alanine is not used in the biosynthesis of any major proteins or enzymes. It is formed in vivo by the degradation of dihydrouracil and carnosine. It is a component of the naturally occurring peptides carnosine and anserine and also of pantothenic acid (vitamin B5), which itself is a component of coenzyme A. Under normal conditions, β-alanine is metabolized into acetic acid.
β-Alanine is the rate-limiting precursor of carnosine, which is to say carnosine levels are limited by the amount of available β-alanine.
Supplementation with β-alanine has been shown to increase the concentration of carnosine in muscles, decrease fatigue in athletes and increase total muscular work done.
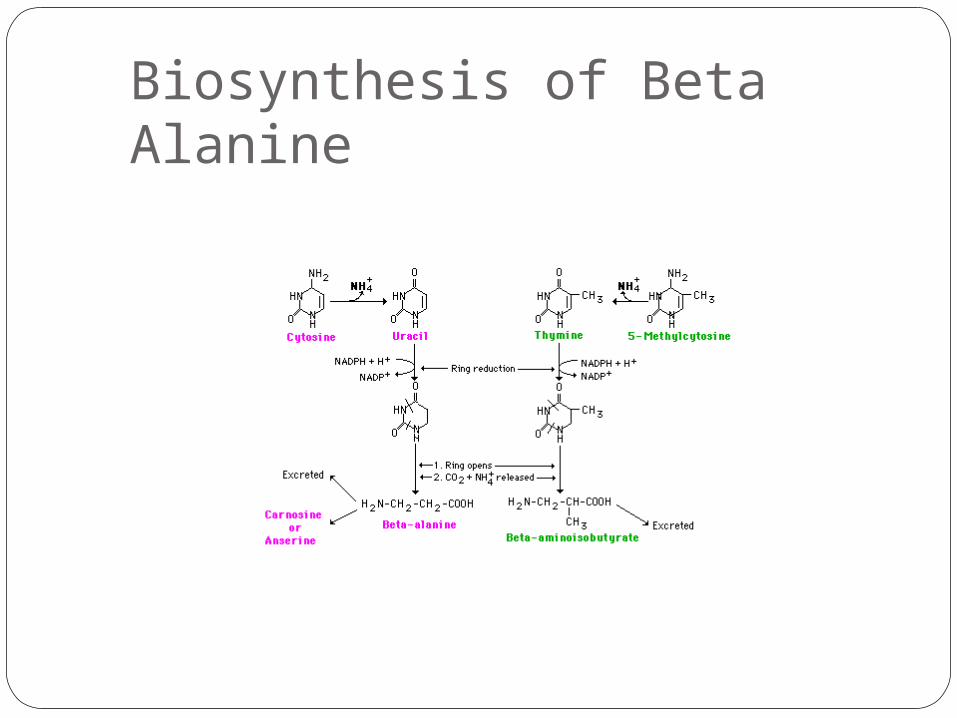
Biosynthesis of Beta Alanine

Carnosine (beta-alanyl-L-histidine) is a dipeptide of the amino acids beta-alanine and histidine. It is highly concentrated in muscle and brain tissues.
Anserine (beta-alanyl-N-methylhistidine) is a dipeptide found in the skeletal muscle and brain of mammals, and birds.
It is an antioxidant (about 5 times that of carnosine) and helps reduce fatigue


γ-Aminobutyric acid (GABA)
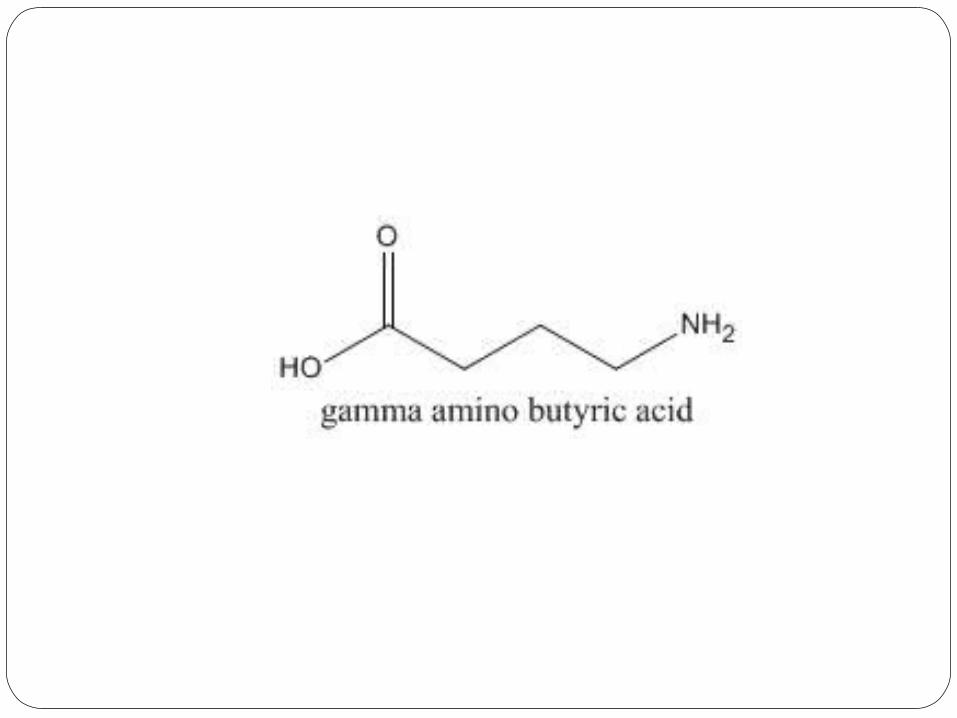

FunctionsIt plays a role in regulating neuronal excitability
throughout the nervous system. In humans, GABA is also directly responsible for the regulation of muscle tone.
GABA acts at inhibitory synapses in the brain by binding to specific transmembrane receptors in the plasma membrane of both pre- and postsynaptic neuronal processes.
This binding causes the opening of ion channels to allow the flow of either negatively charged chloride ions into the cell or positively charged potassium ions out of the cell.
In both cases, the membrane potential is decreased. This action results in a negative change in the transmembrane potential, usually causing hyperpolarization

Synthesis
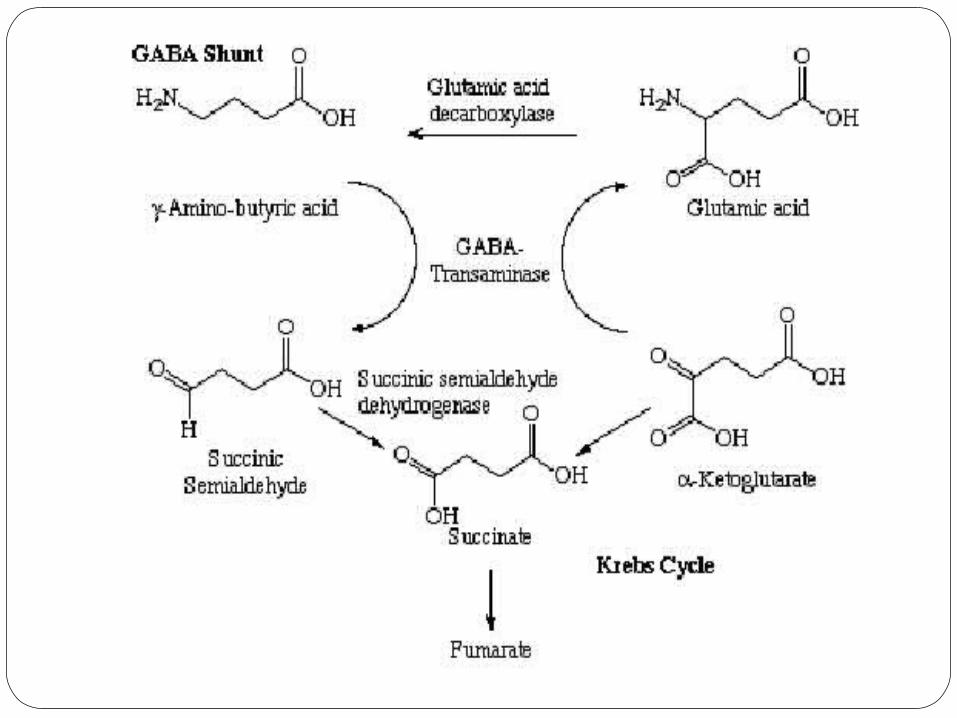
GABA does not penetrate the blood-brain barrier; it is synthesized in the brain.
It is synthesized from glutamate using the enzyme L-glutamic acid decarboxylase ( GAD) and pyridoxal phosphate (which is the active form of vitamin B6) as a cofactor via a metabolic pathway called the GABA shunt.
The synthesis of GABA is linked to the Kreb's cycle.

This process converts glutamate, the principal excitatory neurotransmitter, into the principal inhibitory neurotransmitter (GABA).
GABA is destroyed by a transamination reaction, in which the amino group is transferred to alpha-oxoglutaric acid (to yield glutamate), with the production of succinic semialdehyde, and then succinic acid. The reaction is catalysed by GABA transaminase.
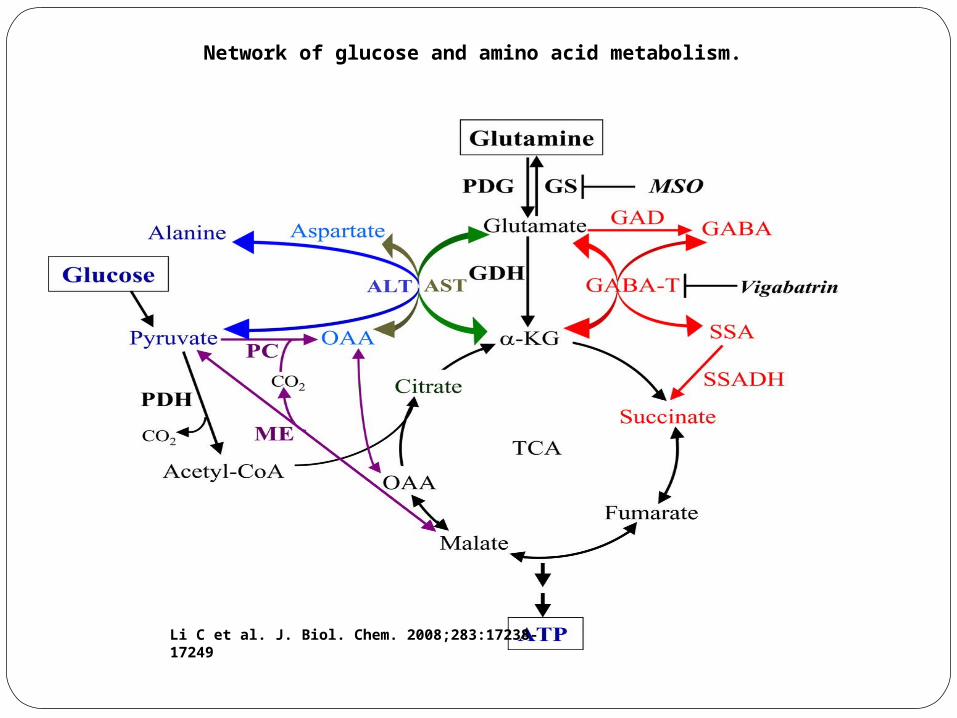
Network of glucose and amino acid metabolism.
Li C et al. J. Biol. Chem. 2008;283:17238-17249

Delta Amino Acids
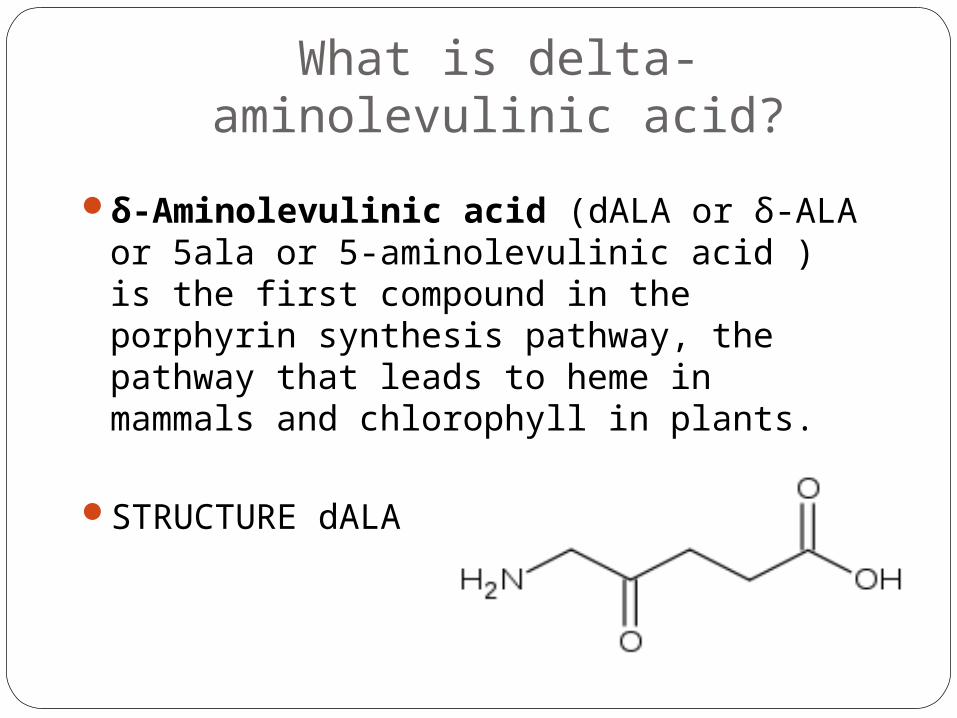
What is delta-aminolevulinic acid?
δ-Aminolevulinic acid (dALA or δ-ALA or 5ala or 5-aminolevulinic acid ) is the first compound in the porphyrin synthesis pathway, the pathway that leads to heme in mammals and chlorophyll in plants.
STRUCTURE dALA

Heme SynthesisHeme synthesis occurs partly in the
mitochondria and partly in the cytoplasm. The process begins in the mitochondria because
one of the precursors is found only there. Since this reaction is regulated in part by the concentration of heme, the final step (which produces the heme) is also mitochondrial.
Many of the intermediate steps are cytoplasmic. Notice in the diagram of the pathway that there is a branch with no apparent useful endproduct.
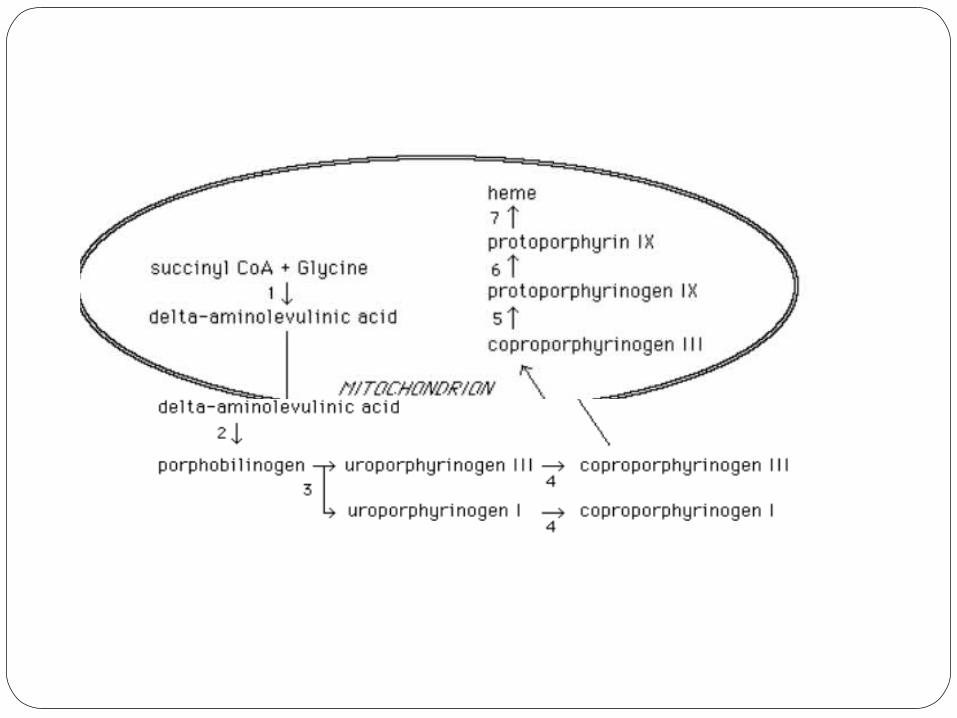

Outline of the porphyrin synthesis pathway.
1) delta-aminolevulinic acid synthase (ALA synthase).
2) delta-aminolevulinic acid dehydratase (ALA dehydratase)
3) uroporphyrinogen I synthase and uroporphyrinogen III cosynthase
4) uroporphyrinogen decarboxylase 5) coproporphyrinogen III oxidase 6) protoporphyrinogen IX oxidase 7) ferrochelatase

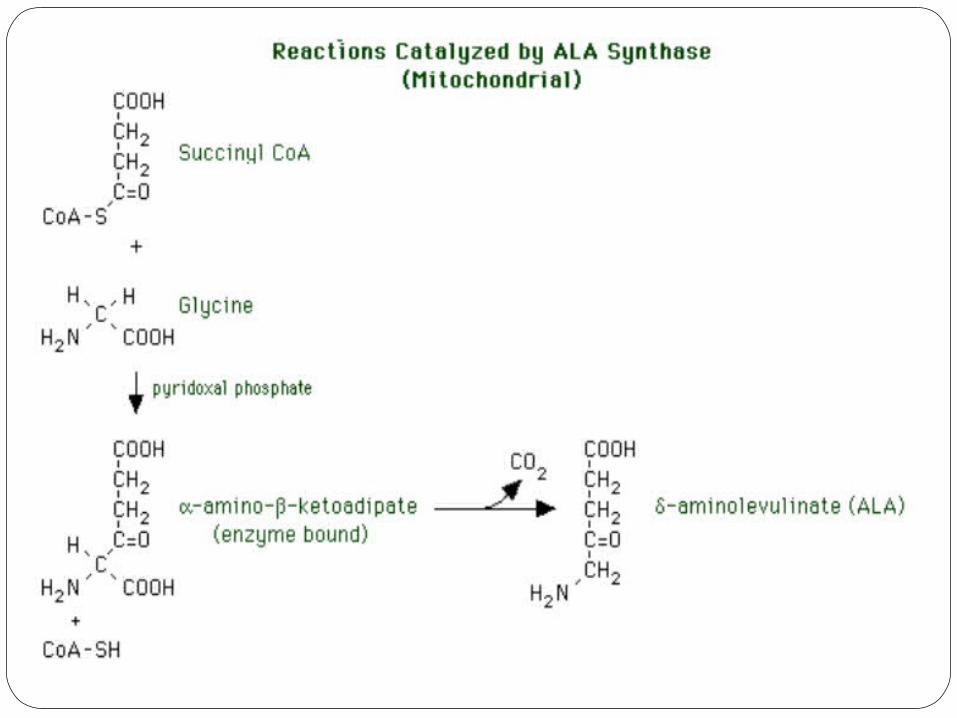
1) Delta-aminolevulinic acid synthase (ALA
synthase)The delta-aminolevulinic acid synthase (ALA
synthase) reaction occurs in the mitochondria.
The substrates aresuccinyl CoA (from the tricarboxylic acid
cycle) glycine (from the general amino acid pool)

An essential cofactor is pyridoxal phosphate (vitamin B-6).
The reaction is sensitive to nutritional deficiency of this vitamin.
Drugs which are antagonistic to pyridoxal phosphate will inhibit it. Such drugs include
1. penicillamine, is used as a form of immunosuppression to treat rheumatoid arthritis. It works by reducing numbers of T-lymphocytes, inhibiting macrophage function, decreasing IL-1, decreasing rheumatoid factor, and preventing collagen from cross-linking.

It is used as a chelating agent: Wilson's disease and cystinuria, Penicillamine is the second line treatment for arsenic poisoning, after
dimercaprol (BAL)
2. isoniazid also known as isonicotinylhydrazine (INH), is an organic compound that is the first-line anti tuberculosis medication in prevention and treatment.
The reaction occurs in two steps. 1. Condensation of succinyl CoA and glycine to form enzyme-bound alpha-
amino-beta-ketoadipate. 2. Decarboxylation of alpha-amino-beta-ketoadipate to form delta-
aminolevulinate. This is the rate-limiting reaction of heme synthesis in all tissues, and it is
therefore tightly regulated.

There are two major means of regulating the activity of the enzyme.
1. The first is by regulating the synthesis of the enzyme protein. This is important because its half life is only about one hour.
Enzyme synthesis is repressed by heme and hematin. It is stimulated by barbiturates (as a result, these drugs exacerbate
certain porphyrias). steroids with a 4,5 double bond, such as testosterone and certain oral
contraceptives. This double bond can be reduced by two different reductases to form either a 5-alpha or a 5-beta product. Only the 5-beta product affects synthesis of ALA synthase. Since the 5-beta reductase appears at puberty, some porphyrias are not manifested until this age.
2. The second control is feedback inhibition by heme and hematin, presumably by an allosteric mechanism. Hence, heme has a dual role in decreasing its own rate of synthesis.
The product of the reaction, ALA, diffuses into the cytoplasm, where the next several steps of heme synthesis occur.

2) delta-aminolevulinic acid dehydratase (ALA
dehydratase)The ALA dehydratase reaction occurs in the cytoplasm; the
product is porphobilinogenThe substrates are two molecules of ALA.The reaction is a condensation to form
porphobilinogen, the first pyrrole.Two molecules of water are released. The asymmetry of the
reaction relative to the two molecules of substrate results in the pyrrole ring having two different substituent groups:
1. acetic acid 2.propionic acid.
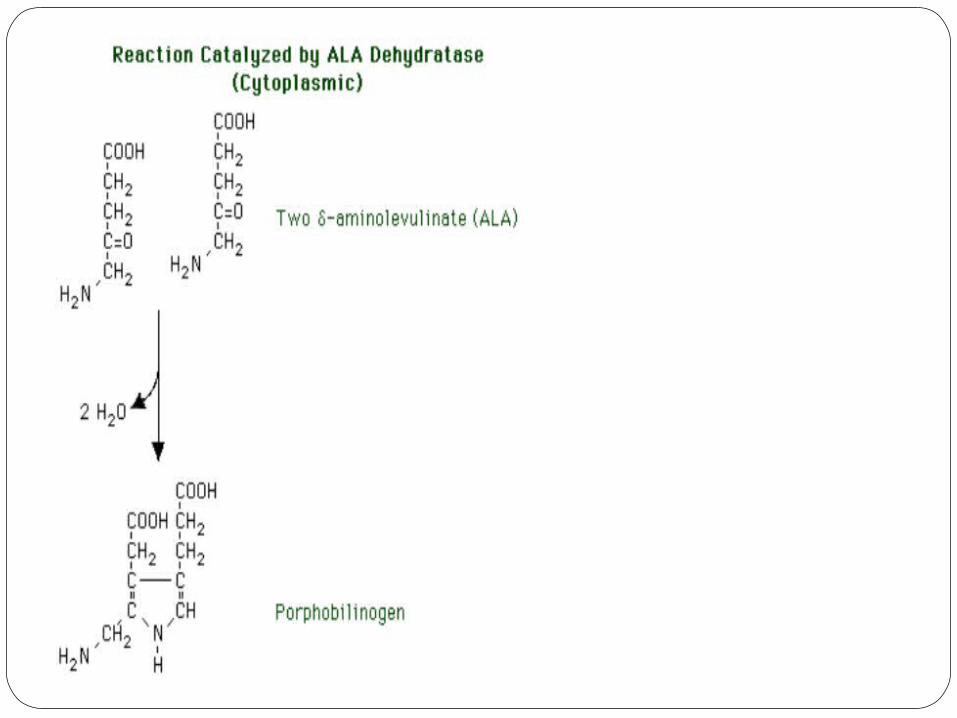

ALA dehydratase is a -SH containing enzyme.It is very susceptible to inhibition by heavy
metals, especially lead.increased urinary excretion of its substrate is a
good indicator of lead poisoning. This is because when ALA dehydratase is
inhibited its substrate, delta-aminolevulinate (ALA), accumulates. This is in part simply because it is no longer being used, and in part because it can no longer continue down the pathway to form heme, which would serve as a feedback inhibitor of further ALA synthesis. ALA continues to be made, and the excess is excreted in the urine.

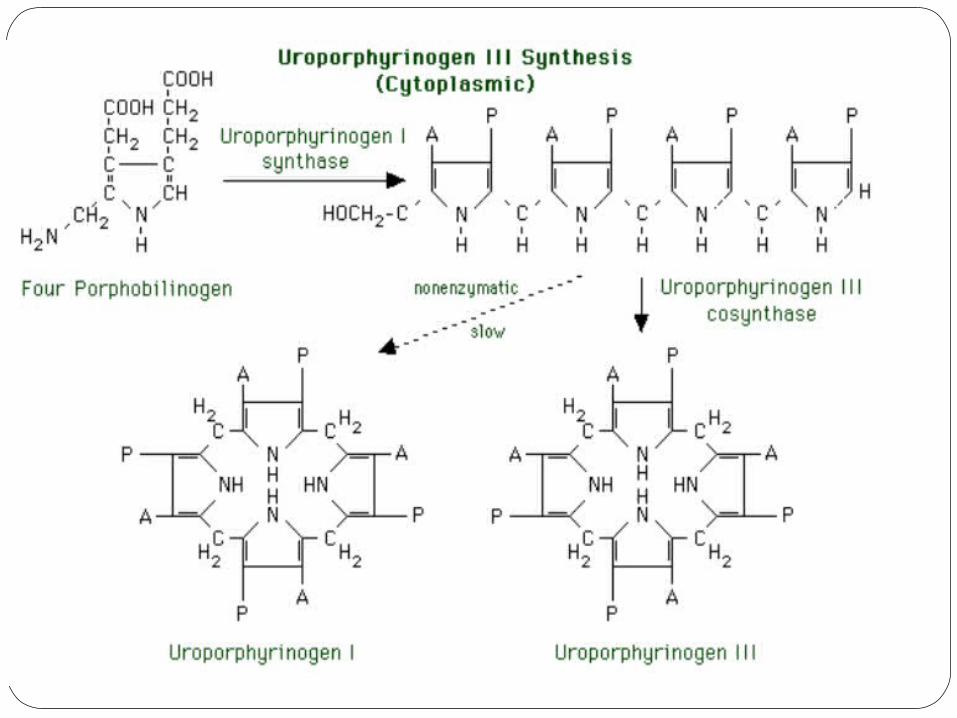
3) Uroporphyrinogen I synthase and
uroporphyrinogen III cosynthase
Production of uroporphyrin III requires two enzymes. The substrates are four molecules of porphobilinogen. 1. The first reaction is catalyzed by uroporphyrinogen I
synthase. The porphobilinogen molecules lose their amino groups. A linear tetrapyrrole with alternating acetic acid and
propionic acid groups is produced. This linear molecule cyclizes slowly (nonenzymatically) to yield uroporphyrinogen I. Without the second reaction (below), the heme synthesis pathway would end with porphyrinogens of the I series, which have no known function.

2. The second reaction is catalyzed by uroporphyrinogen III cosynthase. This enzyme rapidly converts the alternating linear tetrapyrrole to the cyclic uroporphyrinogen III, which has the substituents of its IV ring reversed: AP AP AP PA. This is the physiologically useful product.

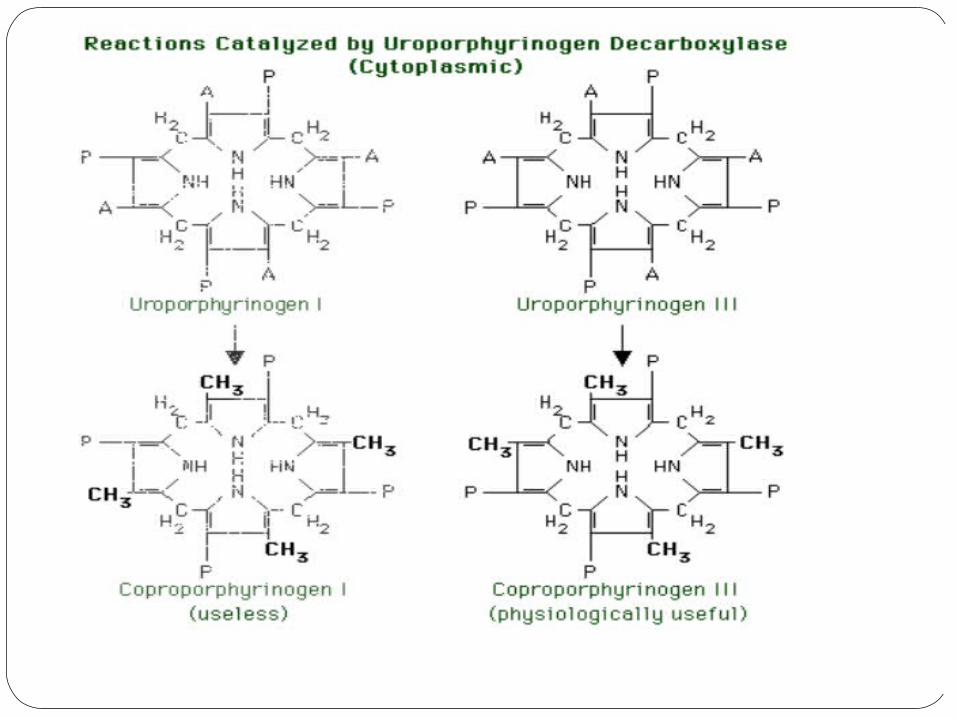
4) Uroporphyrinogen decarboxylase
Uroporphyrinogen decarboxylase decarboxylates the acetic acid groups, converting them to methyl groups.
The physiologically significant substrate is uroporphyrinogen III. The product is coproporphyrinogen III, which is transported
back to the mitochondria, where the remainder of heme synthesis occurs.
The substituent pattern in the coproporphyrinogen is MP MP MP PM.
Uroporphyrinogen decarboxylase also acts on uroporphyrinogen I, yielding coproporphyrinogen I. Coproporphyrinogen I has no known function, and its formation is thought to be a blind pathway.

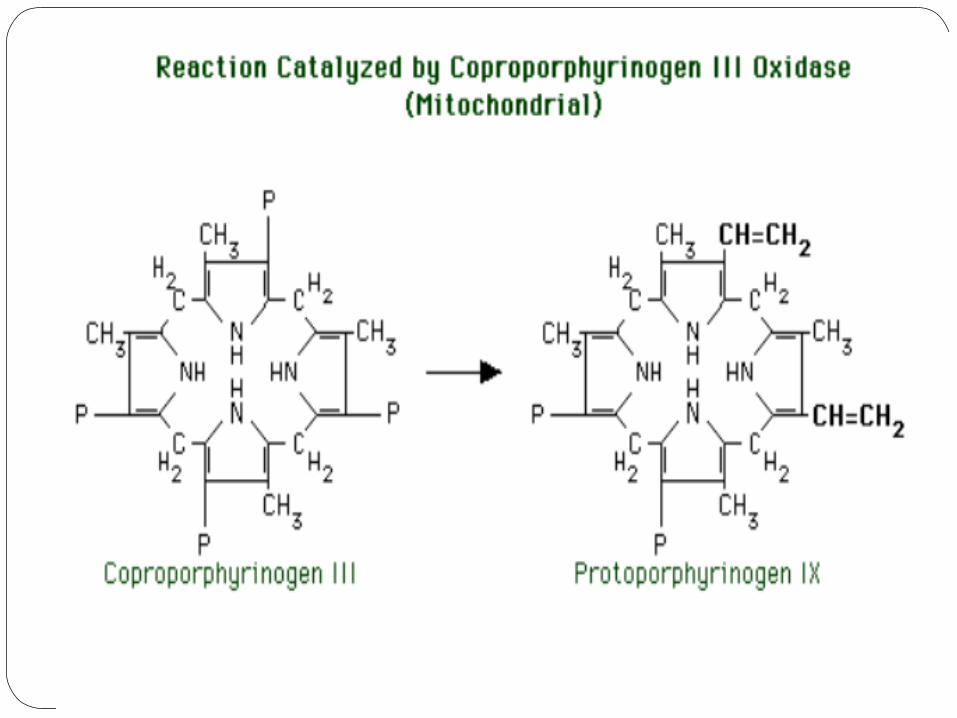
5) Coproporphyrinogen III oxidase
The mitochondrial enzyme, coproporphyrinogen III oxidase, catalyzes the next reaction.
The substrate is coproporphyrinogen III. The reaction is conversion of the propionic acid groups of
rings I and III to vinyl groups. We now have the final substituent pattern of MV MV MP PM (note that "Petrarchan" pattern of the last four substituent groups).
The product is protoporphyrinogen IX. (Some naming systems would call this protoporphyrinogen III to preserve the logic of the nomenclature, but "protoporphyrinogen III" is a departure from a time-honored tradition of referring to this and subsequent compounds by the number "IX.")

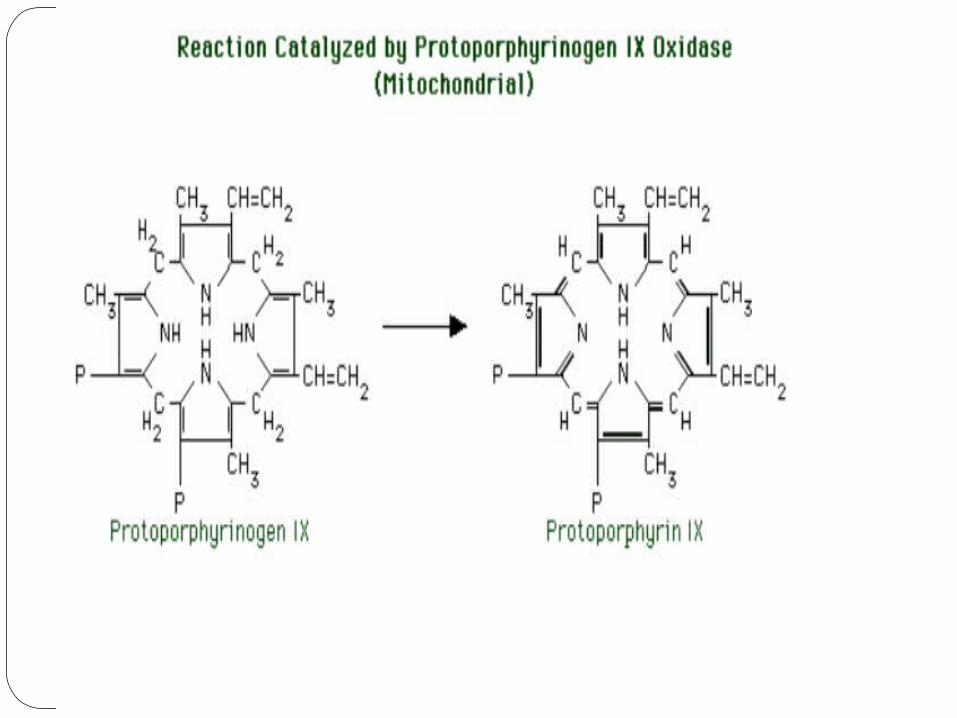
6) Protoporphyrinogen IX oxidase
Protoporphyrinogen IX oxidase converts the methylene bridges between the pyrrole rings to methenyl bridges. Resonance of double bonds around the entire great ring, with its resulting stabilization, is now possible.

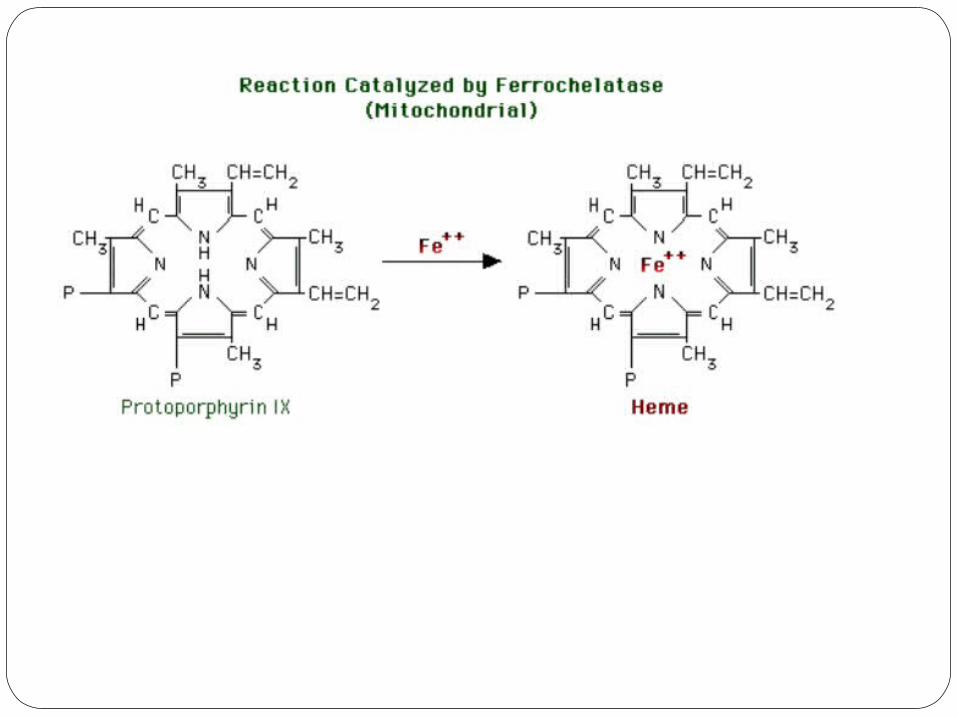
7) Ferrochelatase
Ferrochelatase adds iron (II) to protoporphyrin IX, forming heme
The enzyme requires iron (II), ascorbic acid and cysteine (reducing agents).
Ferrochelatase is inhibited by lead.

Physiological regulation of heme synthesis
Substrate availability: iron (II) must be available for ferrochelatase.
Feedback regulation: heme is a feedback inhibitor of ALA synthase.
Subcellular localization: ALA synthase is in the mitochondria, where the
substrate, succinyl CoA, is produced. ALA synthase is synthesized in the
cytoplasm, and is transported into the mitochondria (like many other
mitochondrial proteins). Its transport across the mitochondrial membrane
may be regulated.

In erythropoietic cells, heme synthesis is coordinated with globin synthesis. If heme is available, globin synthesis proceeds. If heme is absent:
a cAMP independent protein kinase is active. the kinase phosphorylates and thereby
inactivates, the eukaryotic initiation factor, eIF-2. This prevents further globin synthesis
Effects of drugs and steroids: Remember, certain drugs and steroids can increase heme synthesis via increased production of the rate-limiting enzyme, ALA synthase
![Diagnostic usefulness of an amino acid tracer, a-[N … · ORIGINAL ARTICLE Diagnostic usefulness of an amino acid tracer, a-[N-methyl-11C]-methylaminoisobutyric acid (11C-MeAIB),in](https://img.pdfslide.tips/doc/110x75/5b85588c7f8b9a317e8ded7e/diagnostic-usefulness-of-an-amino-acid-tracer-a-n-original-article-diagnostic.jpg)




























