Embed Size (px)
Citation preview

Anny Loreni Díaz Gaitán 164002506Programa de Biología V Semestre
Mayo 2012

Propiedades de los gases en las fases
gaseosas y las soluciones acuosas.

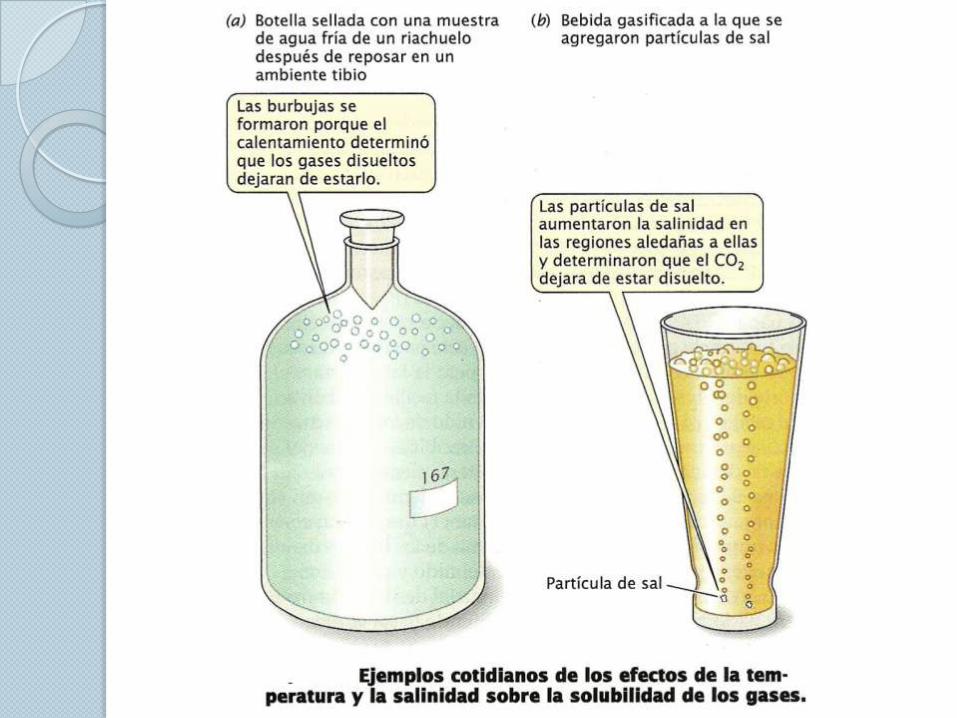
Difusión de los gases. Cuando los gases se difunden de un sitio a otro utilizan el mismo
mecanismo fundamental por medio del cual los solutos se difunden a
través de soluciones.
La ley fundamental de la difusión de los gases indica que los gases se
difunden en forma neta desde las áreas con presiones parciales
relativamente elevadas hacia las que presenta presiones parciales
relativamente bajas. Tasa de
movimiento
Factor de
proporcion
alidad
Diferencia de
presiones

Transporte de los gases por
convección.Este puede ser mucho más rápido que la difusión de los gases porque
moviliza moléculas del gas en una forma determinada y forzada, y no
dependiente de los movimiento aleatorios de las moléculas.


Respiración externa.En todos los animales los sistemas utilizados para el intercambio respiratorio
de gases con el medio ambiente utilizan una membrana respiratoria de
intercambio de gases o membrana respiratoria de intercambio; entonces la
respiración externa consiste en procesos por medio de los cuales se
transporta el O2 hacia la membrana respiratoria de intercambio de gases
desde el medio ambiente y el CO2 se elimina desde la membrana hacia el
medio ambiente exterior.

Las estructuras corporales especializadas típicas de la respiración externa
tienen membranas respiratorias de intercambio de gases que poseen
patrones de invaginaciones o evaginaciones que aumentan en forma
significativa la superficie de la membrana.
Las branquias son estructuras respiratorias que salen del cuerpo y están
rodeadas por el medio ambiente. En cambio, los pulmones son estructuras
respiratorias invaginadas dentro del cuerpo que contiene el medio ambiente.

La ventilación de los pulmones, las branquias u otras membranas
respiratorias de intercambio de gases puede ser activa o pasiva.
La ventilación es activa si el animal produce las corrientes ventilatorias de
aire o de agua que fluyen desde y hacia la membrana por medio de
fuerzas de succión o de presión que requiere el uso de energía
metabólica.
La ventilación es pasiva si las corrientes de aire o de agua ambiental
inducen el flujo desde y hacia la membrana respiratoria de gases en
forma directa o indirecta.

Principios del intercambio de gases por
ventilación activa.

Es característico que en las estructuras que desarrollan ventilación corriente
la presión parcial de O2 en la sangre que sale del órgano respiratorio sea
menor que la presión parcial de O2 en el medio exhalado.
Cuando la ventilación es unidireccional, las dos relaciones más evidentes que
se desarrollan entre el flujo del medio y el de la sangre son concurrente y a
contracorriente.

Un tercer modo de intercambio es el intercambio respiratorio de gases de
corriente cruzada, que consiste en la descomposición del flujo en varias
corrientes, cada una de las cuales desarrolla el intercambio con el medio en
una parte de su trayectoria.

La propiedad del aire y el agua que es importante para comprender los
cambios relativos en las presiones parciales de O2 y CO2 es el coeficiente de
capacitancia, que se define como el cambio en la concentración total del gas
por unidad de cambio en la presión parcial del gas.

Respiración en los vertebrados.En las especies que forman cualquier grupo de vertebrados relacionados
desde el punto de vista filogenético la superficie total de la membrana
respiratoria de intercambio de gases en una especie es una función
alométrica del tamaño corporal.
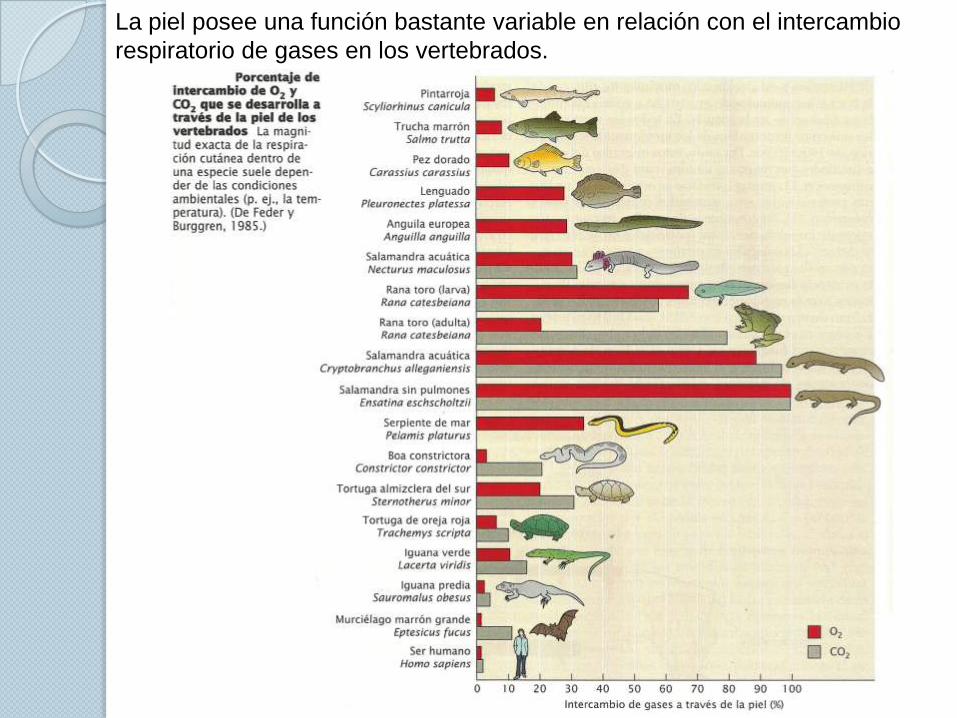
La piel posee una función bastante variable en relación con el intercambio
respiratorio de gases en los vertebrados.

En todos los vertebrados los músculos responsables de los movimientos
respiratorios son esqueléticos. Por tanto, estos músculos deben ser
estimulados por neuronas motoras para desarrollar cada contracción.
El ritmo respiratorio se origina en las neuronas pertenecientes a un
generador central de patrones; estas neuronas desarrollan respuestas
rítmicas de impulsos nerviosos que se dirigen a los músculos respiratorios
y los estimulan para desarrollar patrones rítmicos de contracción.
Casi toda evidencia sugiere que los generadores centrales de patrones
para la respiración en todos los vertebrados se localizan dentro del tronco
encefálico: en el bulbo raquídeo y a veces en otras partes asociadas del
cerebro.

Respiración en los peces.

Respiración en los anfibios.De todos los grupos de vertebrados los anfibios lograron la mejor
combinación entre respiración en el agua y la respiración en el aire. Muchos
de estos animales se mueven desde un ambiente acuático hacia uno terrestre
durante su desarrollo y muchos respiran en ambos ambientes durante su
adultez.

Respiración en los reptiles.

Respiración en los mamíferos.

Respiración en las aves.

Respiración de invertebrados acuáticos y
grupos afines.

Respiración en los insectos y otros
artrópodos traqueados.


Pigmentos
respiratorios. La hemoglobina es uno de los varios tipos de pigmentos respiratorios o
pigmentos transportadores de O2 que desarrollaron los animales.
Todos los tipos de pigmentos respiratorios son metaloproteínas.
Todas estas sustancias poseen colores intensos por lo menos en parte
del tiempo, lo que explica la razón por la cual se denominan pigmentos.
Las hemoglobinas de la sangre desempeñan papeles importantes como
amortiguadores y participan en transporte de CO2 además de dedicarse al
transporte de O2.

Propiedades químicas y distribución de los
pigmentos respiratorios.Se conocen cuatro categorías químicas de pigmentos respiratorios:
hemoglobinas, hemocianinas, hemeritrinas y clorocruorinas.

Varios artrópodos y moluscos poseen hemocianinas a base de cobre.
Las hemocianinas solo se observan en dos filos: los artrópodos y los
moluscos pero representa la segunda clase más frecuente de pigmentos
respiratorios.
Las hemocianinas no contienen hemo, hierro ni porfirinas. El metal que
se encuentra en la molécula es el cobre y está unido en forma directa con
la proteína.
Cada sitio fijador de O2 en una hemocianina contiene dos átomos de
cobre; por tanto, la relación de fijación es de una molécula de O2 cada dos
moléculas de Cu.Las clorocruorinas se asemejan a las hemoglobinas y aparecen en algunos
anélidos.
Las clorocruorinas, también denominadas “hemoglobinas verdes”, se
observan en solo cuatro familias de anémonas de mar y de poliquetos
sedentarios.
Las clorocruorinas siempre se encuentran disueltas en el plasma fuera de
las células y tienen similitudes químicas con las hemoglobinas
extracelulares disueltas en el plasma de muchos anélidos.
Las clorocruorinas difieren de las hemoglobinas en el tipo de hierro y
porfirina que poseen.Las hemeritrinas a base de hierro no contienen hemo.
Las hemeritrinas se desarrollan en todos los gusanos sipuncúlidos, en
muchos branquiópodos, en ambos géneros del filo priapúlido y en una de
las familias de anélidos marinos (magelónidos).
Las hemeritrinas no contienen hemo y están compuestas de Fe unido en
forma directa con una proteína.

Características de la fijación del O2 a los
pigmentosrespiratorios.

Efecto Bohr: la afinidad por el O2 depende de la presión parcial de CO2 y del
pH.
La disminución del pH o el aumento de la presión parcial de CO2 en un líquido
o en un tejido corporal que contienen un pigmento respiratorio suele producir
una disminución de la afinidad del pigmento respiratorio por el O2, este efecto
se denomina efecto Bohr o desviación Bohr.

El efecto Root: en rara ocasiones el CO2 y el pH afectan la capacidad de
transporte de O2 en los pigmentos respiratorios.
En algunos tipos de animales debido a las propiedades distintivas de sus
pigmentos respiratorios el aumento de la presión parcial de CO2 o la
disminución del pH de la sangre no solo induce un efecto Bonr sino que
además reduce la cantidad de O2 que se une al pigmento respiratorio cuando
se satura. La disminución de la cantidad de O2 ligado al pigmento cuando
está saturado se denomina efecto Root.

Efectos térmicos: la afinidad por el oxígeno depende de la temperatura del
tejido.
Los cambios de la afinidad asociados con la temperatura aumenta el aporte
de O2 a los músculos en una forma análoga a la descrita para el efecto Bohr.
Por tanto, el desarrollo de efectos relacionados con la temperatura y de
efectos Bohr en forma simultanea promueve la descarga de O2 en los
músculos activos y ambos efectos tienden a reducir la afinidad del pigmento
respiratorio por el O2 en forma independiente cuando la sangre atraviesa los
músculos.

Transporte de CO2.

La cantidad de bicarbonato formado depende de los amortiguadores de la
sangre.
El bicarbonato es la forma dominante típica en la sangre de los animales.
El grado de formación del bicarbonato no solo depende de los principios de
la solubilidad sino que además de la acción de los compuestos que actúan
como amortiguadores del pH.
Las reacciones de los amortiguadores está representada en la siguiente
ecuación general:


El transporte del CO2 se interpreta con las curvas de equilibrio de este gas.
Las curvas de equilibrio del CO2 en la sangre tienen un valor interpretativo
similar a las del O2
Efecto Haldane: la curva de
equilibrio del CO2 depende de
la oxigenación de la sangre.
La curva de equilibrio del CO2
en la sangre de un animal
suele cambiar de acuerdo con
la oxigenación del pigmento
respiratorio en la sangre,
fenómeno que se denomina
efecto Haldane.
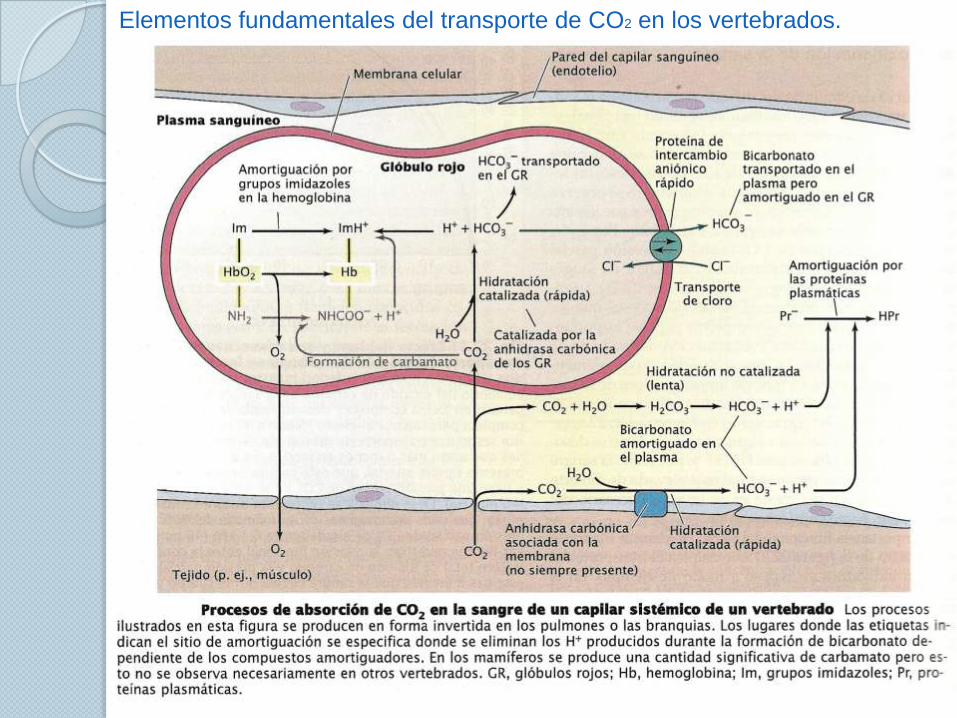
Elementos fundamentales del transporte de CO2 en los vertebrados.

Bibliografí
a. Eckert. (2004). Fisiología Animal: Mecanismos y Adaptaciones. España:
McGraw-Hill.
Richard W. Hill, G. A. (2005). Fisiología Animal. España: Médeca
Panamericana.





