Embed Size (px)
Citation preview


Epitheliums• Tissus de revetement/glandulaire• Classification:Nombre de Couche/Simple,
plusieurs/stratification ; forme cellulaire pavimenteux,cylindrique,
• Pseudostratifie/cubique• Specialisation cellulaire/bordure en brosse• Echanges/compartiment intercellulair• Protection/keratine.• Regeneration• Repose membrane basale• Origine tissu embryonnaire/ectoderme

Epitheliums simples

• Toujours le même type de prélèvement, avec, en plus, une coloration nucléaire. Remarquez l'agencement étroit entre les cellules qui s'imbriquent parfaitement les unes entre les autres. Cette caractéristique est vraie pour tout épithélium.

• Voici la vue générale d'une coupe de rate. Il s'agit d'un organe très cellulaire, d'où son aspect relativement basophile et donc foncé, en 1. La flèche indique, en bordure de l'organe, une mince capsule conjonctive. Les fibres sont colorées ici par le safran. Cette capsule est recouverte par le péritoine, épithélium pavimenteux simple que nous allons observer à plus fort grossissement.

• En 1 : les diverses cellules de la rate; En 2 : la capsule conjonctive qui l'entoure; En 3 : l'épithélium pavimenteux simple péritonéal, appelé mésothélium. Les cellules de cet épithélium sont fortement aplaties. Leur noyau est allongé, lenticulaire. Le cytoplasme, peu abondant, est réduit à une mince couche pratiquement imperceptible, sauf entre les noyaux comme en 4.

• Au niveau de l'endocarde d'un coeur de vache, les cellules pavimenteuses de l'endothélium montrent un aspect caractéristique en forme "d'oeuf sur le plat". Les cellules sont moins étirées, ce qui fait que le noyau, en 1, est arrondi et fait protrusion dans la lumière. En 2, le liseré cytoplasmique reste mince.

• Voici une vue générale de la zone médullaire d'un rein, zone où se trouvent divers tubes rénaux, coupés ici transversalement. On voit notamment: En 1, des tubes limités par une paroi acidophile; En 2, d'autres bordés par une paroi plus épaisse et moins acidophile; En 3, des tubes à paroi fine.

• A fort grossissement, on voit ces divers tubes rénaux bordés par une seule assise de cellules: c'est un épithélium simple. En 1, un premier type de tube est bordé par un épithélium pavimenteux simple: noyau aplati et peu de cytoplasme. En 2, un épithéllium cubique simple: Le noyau, arrondi, occupe toute la hauteur cellulaire. En 3, les cellules sont plus hautes que larges, le cytoplasme plus abondant: c'est un épithélium cylindrique simple.

• Une belle image d'épithélium cubique simple nous est donnée ici, au niveau d'un canal excréteur d'une glande. En 1, se trouve la lumière du canal. En 2, un tissu conjonctif environnant. L'épithélium est séparé de ce conjonctif par une membrane basale, fléchée en 3. Les cellules épithéliales sont véritablement carrées, le noyau, arrondi, est entouré par très peu de cytoplasme.

• La paroi du gros intestin est bordée, du côté de sa lumière par un épithélium de revêtement simple, indiqué par la flèche. Il est constitué d'une seule assise cellulaire qui s'enfonce régulièrement pour former des glandes.

• Observons ces glandes du gros intestin à plus fort grossissement. En 1 est noté l'épithélium. En 2, un tissu conjonctif entoure les glandes. En 3, est fléché le noyau d'une cellule épithéliale. Il est situé dans la partie attenant au tissu conjonctif, c'est le pôle dit "basal". En 4, le cytoplasme occupe le pôle dit "apical", situé du côté de la lumière.

• Ces mêmes glandes, du gros intestin sont coupées ici transversalement. En 1, le pôle basal héberge le noyau. En 2, le cytoplasme occupe le pôle apical. Les cellules sont plus hautes que larges, donc cylindriques. Il n'y a qu'une assise de cellules. C'est donc un épithélium cylindrique simple.

• Un autre exemple d'épithélium cylindrique simple est illustré ici, dans la vésicule biliaire. On devine légèrement, au niveau des flèches, les limites cellulaires. Les cellules sont donc bien plus hautes que larges. On n'observe qu'une seule rangée de noyaux, situés ici plus au centre de la cellule.

Epitheliums stratifies

• Les cellules épithéliales peuvent se superposer en plusieurs couches pour former un épithélium stratifié. Le plus simple, comme ici, est formé de deux couches de cellules dont les noyaux sont fléchés en 1 et en 2. Ces cellules sont de forme cubique. On qualifie dès lors cet épithélium de bistratifié cubique.

• Nous observons ici un épithélium bistratifié cylindrique. Les cellules de la couche supérieure, fléchée, choisie par convention pour qualifier la forme de l'épithélium, sont en effet plus hautes, avec une quantité de cytoplasme plus importante. De tels épithéliums de revêtement bistratifiés se rencontrent généralement autour de canaux excréteurs de glandes.

• L'épithélium que nous observons sur cette image est constitué d'un grand nombre de couches cellulaires superposées. Entre l'assise basale, en 1, située contre le tissu conjonctif et l'assise superficielle en 2, faisant face à la lumière, on distingue de nombreuses couches de cellules. On qualifie cet épithélium de pluristratifié. Nous en distinguerons plusieurs types différents.

• Cet épithélium stratifié possède 2 caractéristiques bien propres. En 1, les cellules de son assise superficielle contiennent encore des noyaux. Ces cellules sont pavimenteuses. Ensuite, cet épithélium montre une coloration régulière sur toute sa hauteur, sans changement brusque de coloration car il n'est pas kératinisé. On le qualifie donc d'épithélium pavimenteux stratifié non kératinisé ou épidermoïde.

• En 1, les cellules de l'assise basale sont cylindriques simples. En 2, elles deviennent polyédriques puis s'aplatissent en 3 pour devenir pavimenteuses dans les couches superficielles. Ces dernières cellules gardent leur noyau. Progressivement, les cellules se chargent de glycogène qui apparait clair sur une coloration habituelle. En 4, sont fléchées des enclaves glycogéniques.

• La langue de poule est recouverte, en 1, par un épithélium épidermoïde très épais. On y reconnait en 2 une des caractéristiques de l'épidermoïde, à savoir, la persistance de noyaux dans les cellules superficielles. Toutefois, ces cellules superficielles se détachent en lambeaux donnant, en 3, cette image de desquamation filiforme très importante.

• En 1, la présence de noyaux dans les couches superficielles; en 2, les enclaves glycogéniques laissent penser à un épithélium épidermoïde. Mais en 3, la coloration plus rouge de la zone de surface montre qu'un début de kératinisation s'est produit. C'est un épithélium parakératosique.

• Nous décrivons à présent l'épithélium pavimenteux stratifié kératinisé de type A. On le trouve dans les zones où la protection doit être accrue. Ainsi, chez l'homme: au niveau de la plante des pieds et de la paume des mains. A faible grossissement, nous observons déjà, en 1, une rupture nette et brusque de coloration, surmontée, en 2, d'une zone épaisse qui est la partie kératinisée.

• On distingue: en 1 : l'assise basale ou germinative, couche unique de cellules cylindriques où se réalisent les mitoses. Elle est surmontée, en 2, par la couche épineuse de Malpighi, constituée de plusieurs assises superposées de cellules polygonales. En 3, la couche granuleuse produit un changement net de coloration. En 4, la couche cornée, épaisse, est kératinisée. La flèche montre un dépôt anormal de sang en surface.

• L'assise germinative ne repose pas à plat sur le conjonctif sous-jacent, mais on observe des soulèvements réguliers de ce conjonctif, appelés papilles dermiques. Ces papilles peuvent apparaitre sous diverses incidences de coupe. Elles permettent une meilleure adhésion de l'épithélium sur le conjonctif.

• A fort grossissement, on observe, en 1, de fins prolongements cytoplasmiques au pôle basal des cellules de l'assise germinative. Ils assurent un meilleur ancrage de l'épithélium sur le conjonctif. En 2, les cellules polyédriques de la couche épineuse semblent séparées par un espace intercellulaire où apparait une certaine striation.

• En 1, la couche épineuse de Malpighi. On rencontre ensuite la couche granuleuse, notée en 2. Le cytoplasme de ces cellules se remplit de grains denses, basophiles. Ce sont les grains de kératohyaline, précurseurs de la kératine. Au niveau de cette couche, les noyaux dégénèrent. Ils deviennent picnotiques, et donc très denses, comme en 3, puis disparaissent.

• Cet épithélium stratifié possède des noyaux au niveau de son assise superficielle, comme un épidermoïde. Mais la flèche indique un changement brusque de coloration dans cet épithélium. La bande plus rouge en surface est kératinisée. Cette kératinisation est incomplète car les noyaux subsistent dans les cellules. On parle d'épithélium parakératosique.

• La couche superficielle délimitée par les pointillés est la couche cornée, couche de cellules pavimenteuses mortes, anucléées et tout à fait kératinisées. Cette couche est très épaisse dans l'épithélium kératinisé de type A. La zone plus rouge, en 2, est un artéfact de tassement, de quelques couches cellulaires. L'ensemble ne forme qu'une seule et même couche

• Un plus fort grossissement de la couche cornée nous montre des cellules mortes, anucléées, de forme aplatie. Cette couche cornée est très épaisse dans le type A car ces cellules desquament très peu.

• Récapitulons les différentes couches d'un épithélium kératinisé : En 1 : l'assise germinative. En 2 : la couche épineuse de Malpighi. En 3 : la couche granuleuse. En 4 : la couche cornée, épaisse, dans le kératinisé de type A. Cet épithélium est traversé par les canaux sudorifères qui conduisent la sueur. On observe un sudorifère en 5 dans la portion conjonctive, en 6, dans l'épithélium.

• Rappelons également plusieurs artéfacts possibles. En 1 : un artéfact de tassement dû à un étirement trop faible lors de l'étalement de la coupe. Un étirement trop prononcé risquerait toutefois de produire un décollement de la couche cornée. En 2 : en surface du prélèvement, nous observons un faible dépôt de sang.

• A faible grossissement, l'épaisseur de la couche cornée, en 1, caractérise l'épithélium kératinisé de type A. On y rencontre, en 2, des bandes transversales plus foncées reparties régulièrement. Il s'agit de tassements de ces cellules kératinisées, au niveau des empreintes digitales. En 3, les images en "ronds de fumée" représentent un canal sudorifère traversant cette couche cornée.

• Nous observons ici une épine implantée dans un épithélium. En 1, les cellules de l'épine présentent un aspect très régulier, très géométrique, comme la plupart des cellules végétales. En 2, autour de ce corps étranger, se produit une réaction inflammatoire importante, afin de résorber l'épine.

• L'épithélium de type A que nous venons de décrire est rare. Le plus souvent, la peau est recouverte par un E.P.S.K. de type B, comme en 1. Cet épithélium s'invagine, en 2, et se différencie pour former des follicules pileux toujours annexés à ce type d'épithélium. En 3, un follicule pileux est coupé plus obliquement.

• L'E.P.S.K. de type B présente les mêmes couches que le type A, toutefois moins développées. En 1, est flèchée l'assise germinative. En 2, la couche épineuse de Malpighi, moins épaisse. En 3, la couche granuleuse. Bien que très mince, elle produit une rupture nette de coloration. En 4, la couche cornée est très fine car elle desquame rapidement en lamelles.

• Nous pouvons retrouver les mêmes couches sur cette image. Notons surtout: En 1 , la couche cornée, très fine. En 2, des traces de desquamation. L'épithélium repose sur le conjonctif qui forme également des papilles dermiques, en 3.

Epitheliums pseudostratifies

• En surface de l'épididyme, toutes les cellules épithéliales partent de la membrane basale, fléchée en 1. C'est donc un épithélium simple. Ces cellules sont de hauteur variable et se chevauchent. En 2, on ne voit qu'un seul noyau de haut tandis qu'en 3, plusieurs noyaux sont superposés. C'est un épithélium pseudostratifié. Les petites cellules, en 4, sont des cellules de remplacement.

• Un épithélium simple particulier, de type pseudostratifié, se rencontre aussi dans la trachée. Nous avons l'impression de voir plusieurs couches de cellules. Mais la superposition des noyaux n'est pas régulière. En réalité, toutes les cellules touchent la membrane basale.

Epithelium urinaire

• La lumière de la vessie que l'on voit en 1 est bordée, en 2 par un épithélium urinaire, ou urothélium. On le qualifie souvent d'épithélium mixte ou de transition.

• Détaillons cet épithélium urinaire. On y classifie plusieurs variétés de cellules, selon leur forme, leur taille et la localisation de leur noyau. En 1, les cellules les plus basses constituent l'assise germinative. On y rencontre, en 2, de nombreuses mitoses. Les cellules, fléchées en 3, sont dites "cellules en raquette" en raison de leur forme. En 4, les cellules les plus hautes sont appelées "cellules en dôme".
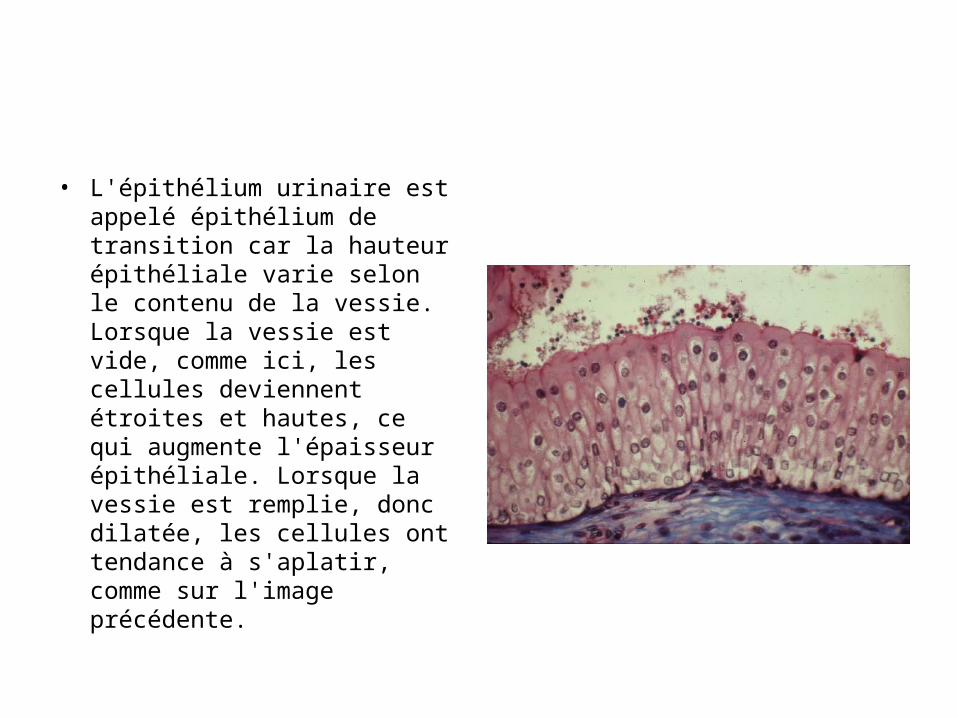
• L'épithélium urinaire est appelé épithélium de transition car la hauteur épithéliale varie selon le contenu de la vessie. Lorsque la vessie est vide, comme ici, les cellules deviennent étroites et hautes, ce qui augmente l'épaisseur épithéliale. Lorsque la vessie est remplie, donc dilatée, les cellules ont tendance à s'aplatir, comme sur l'image précédente.

• Toutes les cellules qui se divisent s'observent à la base de l'épithélium urinaire. Nous observons une cellule en prophase. En 1, le cytoplasme d'une cellule en mitose s'éclaircit fortement. En 2, les chromosomes se rassemblent en une masse très dense. La membrane nucléaire a disparu, ce qui explique l'aspect hérissé de ce noyau en prophase.

• La métaphase se caractérise par un fuseau mitotique bien formé, marqué par la flèche, et par le rassemblement des chromosomes au centre du fuseau, au niveau de la plaque équatoriale.

• Dans l'anaphase, les chromosomes-fils se sont séparés et migrent vers les deux pôles du fuseau mitotique.

• La télophase se marque, au niveau des flèches, par l'invagination de la membrane cellulaire. Il en résultera une séparation des deux cellules filles, qui ont reçu la même garniture chromosomique.

• Les cellules, fléchées en 1, sont appelées "cellules en raquette" en raison de leur forme. Leur cytoplasme, bourré de glycogène, apparait très clair en coloration ordinaire. En 2, les cellules en dôme sont de grandes cellules étalées, recouvrant généralement plusieurs cellules en raquettes. Elles sont souvent plurinucléées.

• Une coupe oblique de l'épithélium urinaire rase tangentiellement plusieurs cellules en dôme. Elles sont plus grandes que les cellules en raquette et possèdent généralement plusieurs noyaux.

• L'épithélium urinaire où l'on reconnait en 1 les cellules en raquette présente fréquemment, en 2, une condensation cytoplasmique au pôle apical des cellules en dôme. Cette condensation, appelée fausse-cuticule, se marque par une coloration à l'érythrosine plus prononcée. Ceci réalise une protection de l'épithélium contre la toxicité de l'urine présente dans la cavité vésicale, en 3.

• Le glycogène est un polysaccharide simple, et peut donc être mis en évidence par un P.A.S. Nous remarquons ici son accumulation dans les cellules en raquette.

Specialisations des epitheliums stratifies
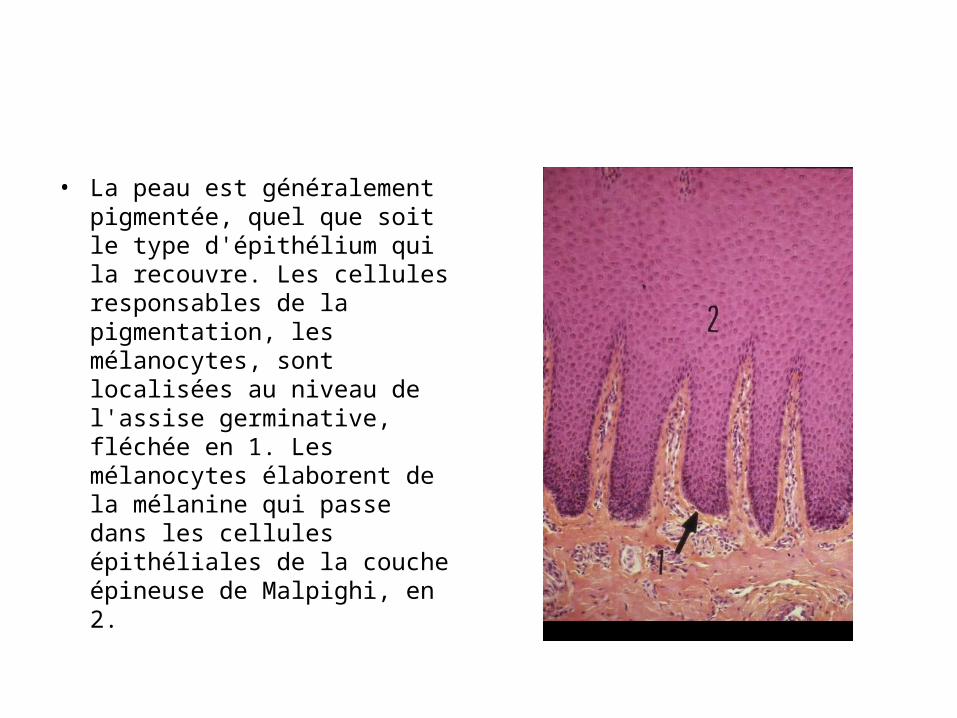
• La peau est généralement pigmentée, quel que soit le type d'épithélium qui la recouvre. Les cellules responsables de la pigmentation, les mélanocytes, sont localisées au niveau de l'assise germinative, fléchée en 1. Les mélanocytes élaborent de la mélanine qui passe dans les cellules épithéliales de la couche épineuse de Malpighi, en 2.

• En dehors de toute coloration particulière, les mélanocytes, apparaissent dans l'assise germinative, comme au niveau des flèches, comme des cellules arrondies à cytoplasme très clair, et à noyau dense. Sans coloration spéciale, il est toutefois impossible de les distinguer des cellules de Merckel, qui ont un rôle neuro-sensitif, ou des cellules de Langerhans, qui jouent un rôle immunitaire.

• Une coloration spéciale, appelée Fontana, permet de mettre en évidence la mélanine. On la voit ici au niveau d'un mélanocyte. Le Fontana est une réaction argentaffine. La mélanine contient en effet des groupements phénols capables de réduire eux-mêmes l'hydroxyde d'argent, ce qui se marque en noir.

• Le mélanocyte se situe au niveau de l'assise germinative. Il projette de nombreux prolongements cytoplasmiques, marqués par les flèches, remplis de mélanine. Ces bras mélanocytaires se ramifient entre les cellules épithéliales dans lesquelles ils déversent ce pigment.

• Le mufle de veau est un organe de choix pour l'étude de la pigmentation, car son épithélium est excessivement pigmenté. On a réalisé un Fontana sur cette coupe. Déjà à faible grossissement, on repère une coloration noire très importante à proximité de l'assise basale.

• En 1, est fléché un mélanocyte: cellule à cytoplasme clair, localisée dans l'assise germinative. L'épithélium étant trop pigmenté, on distingue mal ses bras. La mélanine est déversée dans les cellules épithéliales voisines, comme en 2. En montant dans les couches supérieures, les cellules entrainent avec elles la mélanine. On la remarque, en 3, coiffant les noyaux cellulaires.

• Nous observons la coupe d'un mufle de veau colorée à l'H.E.S. Sans coloration spéciale, la mélanine apparait en brun lorsqu'elle est abondante. On la trouve principalement: en 1, au niveau des cellules épithéliales de l'assise germinative. En 2, elle se concentre en calotte au-dessus du noyau des cellules de la couche épineuse.

• Nous décrivons à présent l'épithélium pavimenteux stratifié kératinisé de type A. On le trouve dans les zones où la protection doit être accrue. Ainsi, chez l'homme: au niveau de la plante des pieds et de la paume des mains. A faible grossissement, nous observons déjà, en 1, une rupture nette et brusque de coloration, surmontée, en 2, d'une zone épaisse qui est la partie kératinisée.

• On distingue: en 1 : l'assise basale ou germinative, couche unique de cellules cylindriques où se réalisent les mitoses. Elle est surmontée, en 2, par la couche épineuse de Malpighi, constituée de plusieurs assises superposées de cellules polygonales. En 3, la couche granuleuse produit un changement net de coloration. En 4, la couche cornée, épaisse, est kératinisée. La flèche montre un dépôt anormal de sang en
surface.

• L'assise germinative ne repose pas à plat sur le conjonctif sous-jacent, mais on observe des soulèvements réguliers de ce conjonctif, appelés papilles dermiques. Ces papilles peuvent apparaitre sous diverses incidences de coupe. Elles permettent une meilleure adhésion de l'épithélium sur le conjonctif.

• A fort grossissement, on observe, en 1, de fins prolongements cytoplasmiques au pôle basal des cellules de l'assise germinative. Ils assurent un meilleur ancrage de l'épithélium sur le conjonctif. En 2, les cellules polyédriques de la couche épineuse semblent séparées par un espace intercellulaire où apparait une certaine striation.

• En 1, la couche épineuse de Malpighi. On rencontre ensuite la couche granuleuse, notée en 2. Le cytoplasme de ces cellules se remplit de grains denses, basophiles. Ce sont les grains de kératohyaline, précurseurs de la kératine. Au niveau de cette couche, les noyaux dégénèrent. Ils deviennent picnotiques, et donc très denses, comme en 3, puis disparaissent.

• La couche superficielle délimitée par les pointillés est la couche cornée, couche de cellules pavimenteuses mortes, anucléées et tout à fait kératinisées. Cette couche est très épaisse dans l'épithélium kératinisé de type A. La zone plus rouge, en 2, est un artéfact de tassement, de quelques couches cellulaires. L'ensemble ne forme qu'une seule et même couche.
•

• Un plus fort grossissement de la couche cornée nous montre des cellules mortes, anucléées, de forme aplatie. Cette couche cornée est très épaisse dans le type A car ces cellules desquament très peu.

• Récapitulons les différentes couches d'un épithélium kératinisé : En 1 : l'assise germinative. En 2 : la couche épineuse de Malpighi. En 3 : la couche granuleuse. En 4 : la couche cornée, épaisse, dans le kératinisé de type A. Cet épithélium est traversé par les canaux sudorifères qui conduisent la sueur. On observe un sudorifère en 5 dans la portion conjonctive, en 6, dans l'épithélium.

• A faible grossissement, l'épaisseur de la couche cornée, en 1, caractérise l'épithélium kératinisé de type A. On y rencontre, en 2, des bandes transversales plus foncées reparties régulièrement. Il s'agit de tassements de ces cellules kératinisées, au niveau des empreintes digitales. En 3, les images en "ronds de fumée" représentent un canal sudorifère traversant cette couche cornée.

• Rappelons également plusieurs artéfacts possibles. En 1 : un artéfact de tassement dû à un étirement trop faible lors de l'étalement de la coupe. Un étirement trop prononcé risquerait toutefois de produire un décollement de la couche cornée. En 2 : en surface du prélèvement, nous observons un faible dépôt de sang.

• Nous observons ici une épine implantée dans un épithélium. En 1, les cellules de l'épine présentent un aspect très régulier, très géométrique, comme la plupart des cellules végétales. En 2, autour de ce corps étranger, se produit une réaction inflammatoire importante, afin de résorber l'épine.

• L'épithélium de type A que nous venons de décrire est rare. Le plus souvent, la peau est recouverte par un E.P.S.K. de type B, comme en 1. Cet épithélium s'invagine, en 2, et se différencie pour former des follicules pileux toujours annexés à ce type d'épithélium. En 3, un follicule pileux est coupé plus
obliquement.

• L'E.P.S.K. de type B présente les mêmes couches que le type A, toutefois moins développées. En 1, est flèchée l'assise germinative. En 2, la couche épineuse de Malpighi, moins épaisse. En 3, la couche granuleuse. Bien que très mince, elle produit une rupture nette de coloration. En 4, la couche cornée est très fine car elle desquame rapidement en lamelles.

• Nous pouvons retrouver les mêmes couches sur cette image. Notons surtout: En 1 , la couche cornée, très fine. En 2, des traces de desquamation. L'épithélium repose sur le conjonctif qui forme également des papilles dermiques, en 3.

• En microscopie électronique, on note, en 1, une cellule épithéliale, et en 2, un mélanocyte dont le cytoplasme est plus clair. La mélanine est synthétisée sous forme de grains denses aux électrons, fléchés en 3, appelés mélanosomes. On retrouve ces mélanosomes, en 4, déversés dans les cellules épithéliales.

• Une image en cryodécapage nous montre, en 1, un bras de mélanocyte infiltré entre deux cellules épithéliales. Le trait de fracture révèle les feuillets membranaires de plusieurs mélanosomes. En 2, les cellules épithéliales voisines sont remplies de tonofilaments (fléchés en 3). On y voit quelques mélanosomes.

• Intéressons-nous au problème des jonctions entre les cellules épithéliales. Dans la couche épineuse de Malpighi, les cellules semblent séparées par un petit espace clair. Une coloration Unna montre, dans ce pseudo-espace intercellulaire, de fins traits reliant deux cellules voisines. Ce sont les ponts d'unions avec, au centre, un épaississement ponctué appelé nodule de Bizzozero.

• En microscopie électronique, on voit deux cellules de la couche épineuse, en 1 et en 2, qui s'engrènent entre elles par des prolongements cytoplasmiques en forme de doigts. L'espace intercellulaire réel, est donc mince. Il est fléché en 3, entre les deux membranes plasmiques. En 4, les desmosomes sont des épaississements particuliers et localisés des membranes, d'où partent les tonofilamentst fléchés en 5.
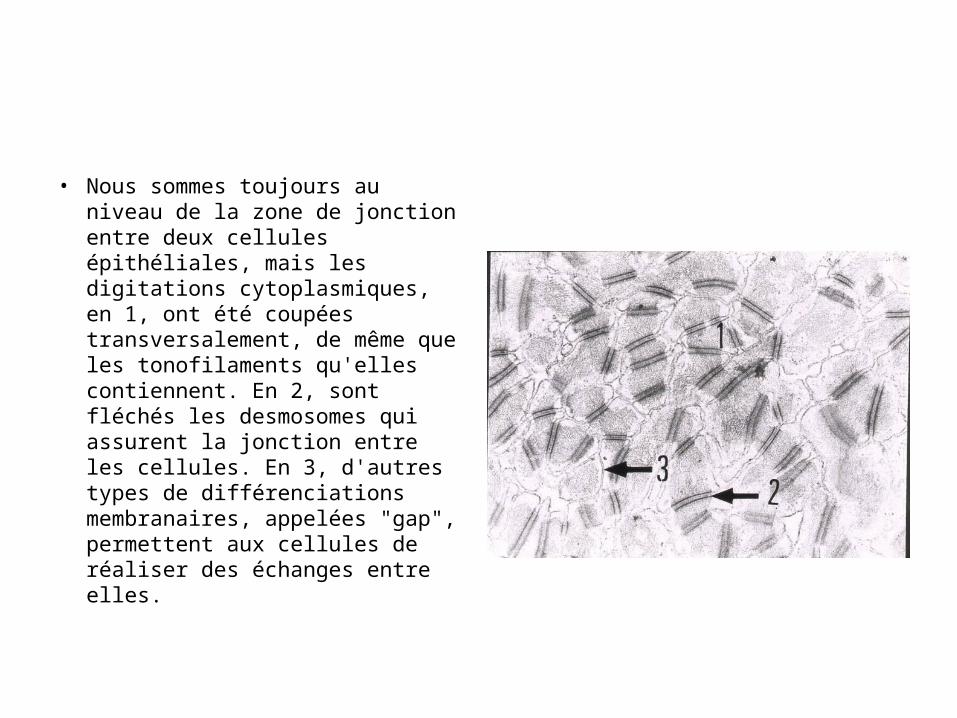
• Nous sommes toujours au niveau de la zone de jonction entre deux cellules épithéliales, mais les digitations cytoplasmiques, en 1, ont été coupées transversalement, de même que les tonofilaments qu'elles contiennent. En 2, sont fléchés les desmosomes qui assurent la jonction entre les cellules. En 3, d'autres types de différenciations membranaires, appelées "gap", permettent aux cellules de réaliser des échanges entre elles.

• A fort grossissement, un desmosome laisse voir cinq lignes denses aux électrons. En 1, les lignes les plus larges correspondent à des épaississements du feuillet protéique interne de la membrane de chaque cellule. En 2, sont fléchés les épaississements du feuillet protéique externe. L'espace intercellulaire montre, en 3, une ligne dense qui serait un épaississement du "cell coat".

• En 1, le cryodécapage révèle le feuillet P d'une membrane au niveau d'un desmosome. On y voit une forte agrégation de particules sur la face P, et en 2, une agrégation moins importante sur le feuillet E. En 3, est fléché l'espace intercellulaire. On situe en 4 un second desmosome dont les plans membranaires ne sont pas dévoilés. En 5, sont fléchés les tonofilaments dans le cytoplasme.

• En 1, on localise l'espace entre deux cellules voisines. Sur les différents feuillets membranaires, nous voyons, en 2, une agrégation importante de particules au niveau d'un desmosome. En 3, une agrégation de particules membranaires encore plus serrée et plus régulière caractérise le feuillet P d'une "gap". Son feuillet E, en 4, porte les empreintes de ces particules.

• La relation entre l'image optique et électronique de la zone de jonction entre deux cellules épithéliales s'explique bien. Le pseudo-espace intercellulaire est la zone d'interdigitation des cellules. Les ponts d'union représentent les tonofilaments et les nodules de Bizzozero sont à mettre en relation avec les desmosomes.

• Nous allons aborder l'étude des diverses différenciations des épithéliums stratifiés, en dehors de la kératinisation. Nous nous intéresserons tout d'abord au problème de la pigmentation épithéliale. Nous verrons ensuite quels sont les types de jonctions que l'on y rencontre entre les cellules.

Specialisation des epitheliums simples et pseudostratifies
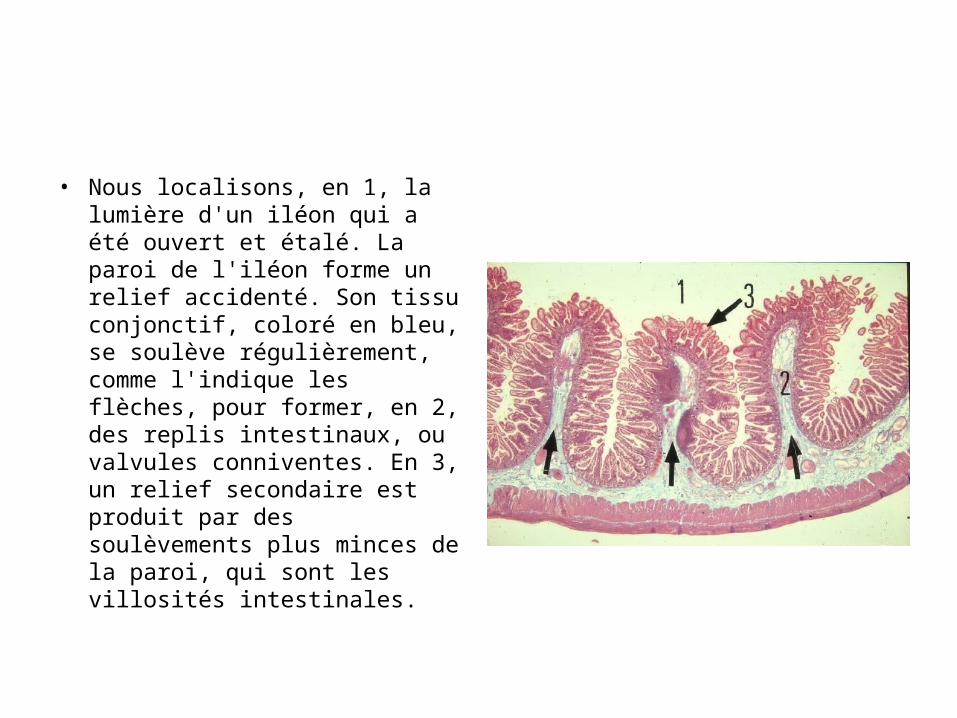
• Nous localisons, en 1, la lumière d'un iléon qui a été ouvert et étalé. La paroi de l'iléon forme un relief accidenté. Son tissu conjonctif, coloré en bleu, se soulève régulièrement, comme l'indique les flèches, pour former, en 2, des replis intestinaux, ou valvules conniventes. En 3, un relief secondaire est produit par des soulèvements plus minces de la paroi, qui sont les villosités intestinales.

• La muqueuse de l'iléon est hérissée d'un ensemble de structures en forme de doigts appelées villosités intestinales. En 1, le centre de la villosité est occupé par du tissu conjonctif recouvert, en 2, par un épithélium cylindrique simple. Notons en 3, que cet épithélium peut s'invaginer pour former des glandes appelées glandes de Lieberkühn. La lumière intestinale
se situe en 4.

• En 1, nous situons l'axe conjonctif d'une villosité intestinale. En 2, l'épithélium de revêtement, et en 3, la membrane basale qui sépare tout tissu épithélial du conjonctif sous-jacent. Dans les cellules épithéliales, on distingue en 4 le pôle basal qui, ici, contient les noyaux, et en 5, du côté de la lumière, le pôle apical.

• Deux types cellulaires constituent l'épithélium intestinal. En 1, les entérocytes, ou cellules à plateau strié, ont une différenciation apicale d'apparence floue, soulignée par une ligne plus colorée. En 2, les cellules caliciformes sont glandulaires et produisent du mucus. Ce mucus apparait clair en coloration ordinaire. On observe aussi en 3, des lymphocytes traversant l'épithélium.

• Détaillons les cellules à plateau strié. Le pôle basal est occupé par le noyau, très allongé. Face à la lumière, le pôle apical, possède une différenciation en plateau strié. Il est composé en 1 d'une zone externe épaisse et floue soulignée, en 2, par une ligne dense bien colorée. En 3, les épaississements ponctués sont les cadres obturants qui unissent les cellules voisines à leur pôle apical.

• En microscopie électronique à balayage, le plateau strié du pôle apical d'un entérocyte apparait formé, en 1, d'une série de petits prolongement fins et réguliers. Ces sortes de bâtonnets réguliers, serrés les uns contre les autres, sont appelés microvillosités. En 2, on distingue une zone sous-jacente d'aspect différent. Cette zone est appelée terminal web.

• En microscopie électronique à transmission, le pôle apical d'un entérocyte montre: En 1, les microvillosités : longues, grêles et régulières. C'est cette zone qui en optique apparait floue à l'apex des cellules. En 2, le terminal web est dépourvu d'organites cellulaires. Il contient, en 3, des microfilaments issus des microvillosités. En optique, le terminal web correspond au trait rouge plus souligné du plateau strié.

• A plus fort grossissement, on voit en 1 que les microvillosités représentent des soulèvements digitiformes de la membrane cellulaire. Elles sont recouvertes par une membrane plasmique unitaire. En 2, l'axe des microvillosités contient des microfilaments d'actine qui se prolongent en 3 dans la région du terminal web.

• Voici un ensemble de microvillosités coupées transversalement. Elles sont limitées, en 1, par une membrane plasmique unitaire dont on observe les deux feuillets protéiques denses. En 2, l'axe de la microvillosité contient les microfilaments. En 3, à l'extérieur de la membrane plasmique, on observe une sorte de chevelu: c'est une condensation plus prononcée du cell coat, fine couche de glycoprotéines normalement présente en surface des membranes plasmiques.

• A plus fort grossissement, on voit en 1 que les microvillosités représentent des soulèvements digitiformes de la membrane cellulaire. Elles sont recouvertes par une membrane plasmique unitaire. En 2, l'axe des microvillosités contient des microfilaments d'actine qui se prolongent en 3 dans la région du terminal web.
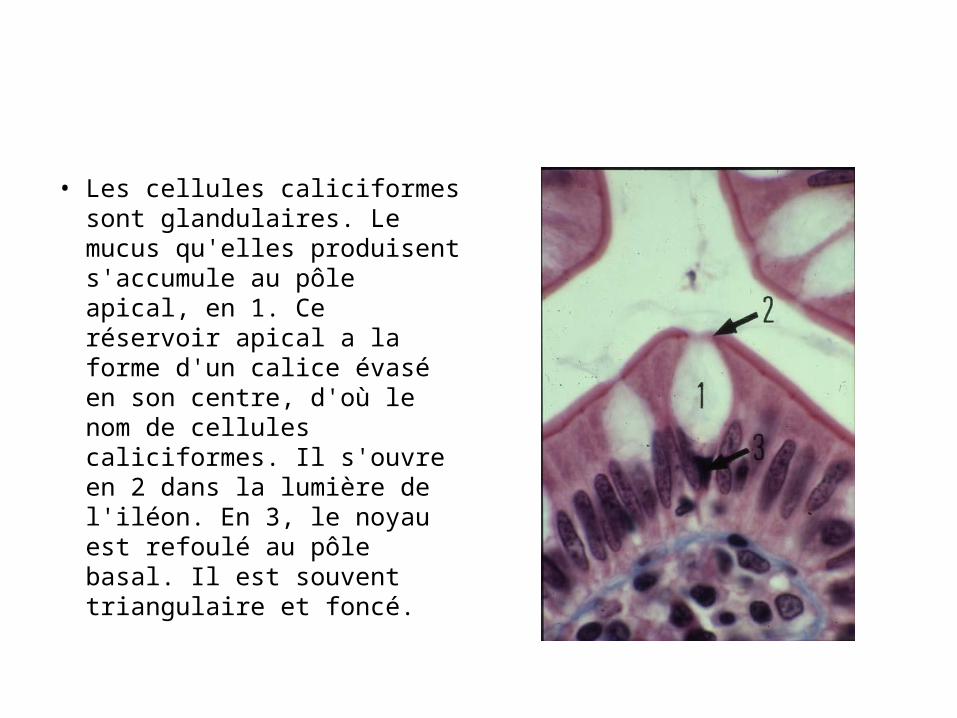
• Les cellules caliciformes sont glandulaires. Le mucus qu'elles produisent s'accumule au pôle apical, en 1. Ce réservoir apical a la forme d'un calice évasé en son centre, d'où le nom de cellules caliciformes. Il s'ouvre en 2 dans la lumière de l'iléon. En 3, le noyau est refoulé au pôle basal. Il est souvent triangulaire et foncé.

• Voici une cellule caliciforme en microscopie électronique à transmission. En 1, le noyau occupe le pôle basal riche en réticulum endoplasmique rugueux, fléché en 2. La sécrétion se fait sous forme de grains denses aux électrons, fléchés en 3, et appelés boules de mucigène. Ce mucigène est un précurseur du mucus proprement dit. Il s'accumule au pôle apical encore fermé de la cellule.

• Par la suite, le mucigène s'hydrate et se transforme en mucus, beaucoup plus clair en microscopie électronique. Une incidence oblique montre, en 1, le pôle apical d'une cellule caliciforme à ce stade. Ce pôle apical est ouvert dans la lumière de l'iléon et on voit, en 2, le mucus qui recouvre le plateau strié des entérocytes voisins. Ce mucus protège la paroi intestinale.

• Outre ces deux types cellulaires, on observe à la base de l'épithélium, en 1, des petites cellules à noyau dense et arrondi logées dans un petit espace clair. Il s'agit de lymphocytes situés dans un espace de Grunhagen. En 2, on retrouve ces lymphocytes à différentes hauteurs entre les cellules épithéliales.

• En 1, une cellule caliciforme dont on voit le pôle apical ouvert. En 2, une incidence plus oblique ne nous montre pas cette communication avec la lumière. En 3, les cellules à plateau strié ou entérocytes. Au niveau de la ligne dense, on observe les cadres obturants qui marquent les jonctions intercellulaires qui existent au pôle apical des cellules.

• Une coupe tangentielle de l'épithélium intestinal observée à très fort grossissement nous montre, en 1, le pôle apical ouvert des cellules caliciformes, en 2, les cadres obturants. Ces jonctions apicales existent sur tout le pourtour cellulaire, enserrant complètement chaque cellule. Suite au tassement, les cellules sont déformées, deviennent hexagonales, d'où l'image en "nid d'abeille" de ces cadres obturants.

• Les cadres obturants vus en microscopie optique représentent une succession de 3 types de jonctions cellulaires. En 1 : une tight junction ou zonula occludens. En 2 : une zonula adhaerens. En 3, un desmosome ou macula adhaerens. Les jonctions 1 et 2, de type zonula, s'étendent sur tout le pourtour cellulaire, ce qui explique en optique les images en "nid d'abeille", en incidence tangentielle.

• Le même complexe de jonctions, se retrouve entre, en 1, une cellule caliciforme et, en 2, une cellule à plateau strié. On observe toujours la même succession, à savoir : En 3, une tight junction, en 4, une zonula adhaerens, en 5, un desmosome.

• Des colorations histochimiques permettent de mettre en évidence certains constituants. Ici, un mucicarmin est un colorant signalétique qui colore en rouge les mucopolysaccharides acides que contient le mucus. Ce mucus est synthétisé, en 1, au niveau des cellules caliciformes et est déversé, en 2, dans la lumière intestinale où il joue un rôle de protection.

• Voici deux cellules caliciformes à fort grossissement. Les mucopolysaccharides acides du mucus sont colorés par le mucicarmin. En 1, on voit le mucus s'écouler dans la lumière par le pôle apical ouvert. Remarquez, en 2, le noyau triangulaire de ces cellules, contre-coloré à l'hémalun. En 3, le plateau strié des entérocytes ne contient pas de mucopolysaccharides acides.

• Le PAS, qui utilise le réactif de Schiff, met en évidence les glucides. Il colore en rouge-violet, en 1, le mucus des cellules caliciformes et, en 2, le plateau strié. Nous avons vu en microscopie électronique que les microvillosités du plateau strié sont recouvertes par le "cell coat". Les mucopolysaccharides et les glycoprotéines du "cell coat" oxydent le réactif de Schiff.

• Le bleu alcian est un colorant signalétique mettant uniquement en évidence les mucopolysaccharides acides. Seul donc le mucus des cellules caliciformes et le mucus présent dans la lumière est coloré en bleu turquoise.

• Sur ces deux villosités intestinales, on voit: en 1, les cellules à plateau strié; en 2, les cellules caliciformes. Signalons un artéfact fréquent: en 3, l'espace ménagé entre l'épithélium et le tissu conjonctif. L'axe conjonctif contient des cellules musculaires lisses qui, lors de la fixation, se sont contractées et ont produit ce décollement.

• Voici une vue, à faible grossissement, de la trompe utérine coupée transversalement. Sa lumière, en 1, est encombrée par de nombreux soulèvements du tissu conjonctif, comme en 2. Ce conjonctif se ramifie abondamment, formant une véritable arborisation recouverte, en 3, par le tissu épithélial que nous allons observer.

• Nous observons un de ces soulèvements. Au centre, en bleu se trouve le tissus conjonctif. Il est recouvert par un épithélium où nous observons déjà deux variétés cellulaires: en 1, des cellules à noyau relativement arrondi, montrant une différenciation apicale. Il s'agit de cils vibratils; en 2, des cellules à
noyau très allongé, non ciliées.

• Sur une incidence parfaite de l'épithélium, nous observons une seule rangée de cellules côte à côte. Il s'agit d'un épithélium cylindrique simple. A la base, en 1, se situent quelques cellules de remplacement. En 2, les cils vibratils des cellules ciliées apparaissent comme de longs prolongements réguliers au pôle apical. Ils sont soulignés, en 3, par un trait plus coloré appelé plateau blépharoblastique.

Glandes exocrines simples

• Sur une coupe de lèvre observée du côté menton, nous étudierons les caractéristiques des glandes sébacées, flèchées en 1. A faible grossissement, nous voyons des massifs cellulaires très clairs, disposés en rosettes autour d'un follicule pileux, noté en 2. Ce follicule dérive de l'épithelium de surface, flèché en 3, qui s'est invaginé.

• En 1, l'épithélium de revêtement de la lèvre côté menton est de type kératinisé B. En 2, nous trouvons des follicules pileux. En 3, les glandes sébacées apparaissent comme des amas très clairs, en coloration ordinaire. Il s'agit de glandes exocrines simples, n'ayant même pas de canal excréteur propre; Elles utilisent le follicule pileux auquel elles sont annexées en guise de canal excréteur.
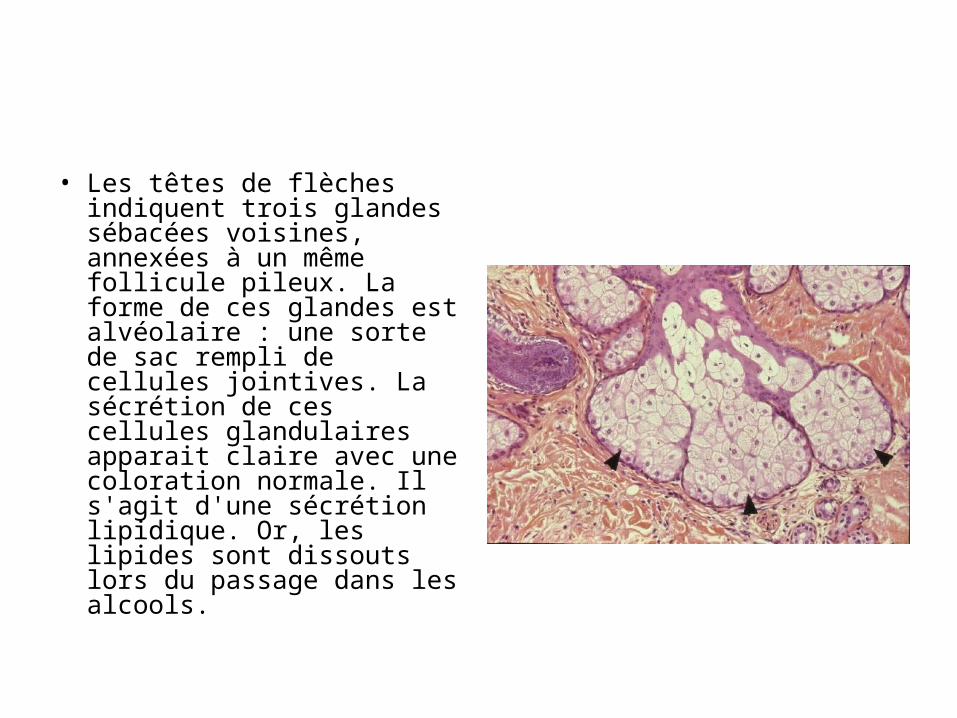
• Les têtes de flèches indiquent trois glandes sébacées voisines, annexées à un même follicule pileux. La forme de ces glandes est alvéolaire : une sorte de sac rempli de cellules jointives. La sécrétion de ces cellules glandulaires apparait claire avec une coloration normale. Il s'agit d'une sécrétion lipidique. Or, les lipides sont dissouts lors du passage dans les alcools.

• Sur une coloration H.E.S., nous observons une glande sébacée bien coupée longitudinalement. Cette glande alvéolaire est limitée en 1 par une couche de cellules basales. Cette assise cellulaire donnera naissance à deux types de cellules: en 2, les cellules glandulaires et en 3, des formations cloisonnantes constituées de cellules non glandulaires. On observe en 4 la paroi du follicule pileux. En 5, une seconde glande sébacée est coupée tangentiellement.

• Un fort grossissement dans la portion basale de la glande nous montre: en 1, les cellules glandulaires où les lipides s'accumulent sous forme de gouttelettes. Nous ne voyons que l'image négative de ces lipides dissouts par les alcools et le toluol. Vu cet aspect réticulé, on donne à ces cel1ules le nom de spongiocytes. En 2, les cellules des formations cloisonnantes, non glandulaires, ont un cytoplasme rosé et une forme étoilée.

• De la base vers le sommet de la glande sébacée, les cellules subissent une maturation. En 1, à la base, les spongiocytes, remplis de fines gouttelettes lipidiques, ont encore une trame cytoplasmique rosée, bien visible. Progressivement, en 2, les noyaux deviennent picnotiques et les gouttelettes lipidiques plus volumineuses. Finalement, les cellules superficielles, en 3, perdent leur noyau. Le cytoplasme est bourré de lipides, d'où son aspect très clair.

• La même charge lipidique progressive s'observe sur cette image. Les cellules superficielles, en 1, bourrées de graisses vont se détacher et tomber, en 2, dans la cavité du follicule pileux qui sert de canal excréteur. C'est un mode de sécrétion holocrine. En 3, sont fléchées les formations cloisonnantes, constituées de cellules non glandulaires.

• La maturation cellulaire et donc l'image d'holocrinie que nous venons de décrire, ne s'observe que sur une coupe bien longitudinale de la glande alvéolaire, comme en 1. Diverses incidences de coupes peuvent se rencontrer. Ainsi, en 2, une section transversale dans la portion basale ne montre que les spongiocytes. Nous ne voyons pas l'annexion de cette glande au follicule pileux, noté en 3.

• Une coupe à congélation permet la conservation des graisses, que révèle ici une coloration spécifique: le soudan rouge. En 1, à la base de l'alvéole glandulaire, les cellules ne contiennent que quelques gouttes lipidiques, tandis qu'à l'apex de la glande, en 2, les lipides remplissent complètement les cellules.

• La paroi de l'utérus comprend: en 1, le myomètre, couche essentiellement musculaire, et face à la lumière, l'endomètre. A sa base, en 2 une région plus foncée (ou zone résiduelle) sert à régénérer, en 3, la partie superficielle (ou zone fonctionnelle) qui desquame à chaque menstruation. En 4, l'endomètre contient des glandes tuhuleuses rectilignes qui deviennent sinueuses dans la zone résiduelle. Elles sont formées par l'invagination de l'épithélium de surface.

• A fort grossissement, voici une glande de l'endomètre. Sa lumière, en 1, est bordée, en 2, par un épithelium cylindrique simple formé d'un seul type cellulaire: des cellules cylindriques à noyau basal et à pôle apical rosé. Lorsqu'elles entreront en phase sécrétoire, ces cellules synthétiseront du glycogène qui passera dans la lumière. Observons, en 3, que des lymphocytes peuvent traverser cet épithélium.

• Nous observons une biopsie de l'endomètre. Il s'agit d'un curetage de la portion superficielle de la zone fonctionnelle. Les glandes y apparaissent en coupe transversale.

• A fort grossissement, nous observons ces glandes tubuleuses de l'endomètre coupées transversalement. Leur lumière arrondie est bordée par un épithélium cylindrique simple où l'on ne distingue qu'un seul type cellulaire. La zone fonctionnelle de l'endomètre régénère après chaque menstruation. Il est donc normal d'y rencontrer de nombreuses mitoses. Ainsi, en 1, au niveau de l'épithelium glandulaire; en 2, dans
le conjonctif très cellulaire.

• La peau face palmaire est recouverte en surface par un épiderme, fléché en 1, qui est de type kératinisé. Dans le derme, on distingue: en 2, le derme moyen, riche en fibres conjonctives et en 3, l'hypoderme qui apparait beaucoup plus clair, car il est riche en lobules graisseux. A la limite derme-hypoderme, se localisent, en 4, de nombreux amas rougeâtres: ce sont les glandes sudoripares que nous allons détailler.

• La muqueuse du duodénum possède, comme l'iléon, en 1 des villosités , en 2 des glandes de Lieberkühn, tubuleuses rectilignes. On trouve ensuite, en 3, des amas glandulaires très nombreux et très clairs en coloration habituelle. Ce sont les glandes de Brunner.

• Les glandes de Brünner du duodénum ont une sécrétion mucoïde qui réagit positivement au P.A.S. Une magnifique incidence longitudinale nous montre, en 1, une lumière tubulaire et, en 2, de nombreuses ramifications latérales. Cette glande est qualifiée de tubuleuse ramifiée. Elle est exocrine simple.

La prostate contient une partie glandulaire. Chez l'homme, il s'agit de glandes tubulo-alvéolaires : une lumière centrale tubulaire, en 1, présente, en 2, de nombreuses petites boursoufflures latérales, de forme alvéolaire. Diverses incidences peuvent s'observer au niveau de ces glandes.

Glandes exocrines composées

• La parotide est une des principales glandes salivaires. Elle est exocrine. A faible grossissement, on distingue une lobulation de la portion glandulaire, notée en 1. Ces lobes sont séparés, en 2, par une faible composante conjonctive. A petit grossissement, la portion glandulaire apparait relativement foncée. Ceci est propre à la nature de la sécrétion: la parotide a une sécrétion séreuse qui est sombre.

• De plus près, on observe en 1, la portion glandulaire, sécrétrice. Il s'agit d'un ensemble d'unités sécrétantes de forme arrondie, serrées les unes contre les autres. L'unite sécrétante de base porte le nom d'acinus. En 2, la portion excrétrice est formée de canaux excréteurs de différents types. C'est une glande exocrine composée. La parotide est riche en cellules adipeuses, notées en 3.

• La parotide est qualifiée de glande acineuse, en raison de son type d'unité secrétante, l'acinus, qui possède une forme ballonnée. La lumière étroite, fléchée sur l'image est bordée par un ensemble de cellules pyramidales: le pôle basal est plus large que l'apex cellulaire. Le noyau, arrondi occupe ce pôle basal. Le pôle apical contient les grains de sécrétion. Toutefois, cette sécrétion est très labile et l'on n'en voit que l'image en négatif.

• Cette parotide de cheval a pu être fixée très rapidement après son prélèvement. Les grains de sécrétion ont été conservés, c'est pourquoi la portion glandulaire, en 1, est fortement colorée. La sécrétion séreuse est foncée. C'est une sécrétion enzymatique. En 2, est fléché un canal excréteur.

• Voici un acinus observé dans cette parotide de cheval. En 1, est fléchée sa lumière. La sécrétion séreuse en 2 se fait sous forme de grains: les grains de zymogène, relativement foncés, qui occupent le pôle apical de la cellule. Le noyau est refoulé dans le tiers basal. Ce pôle basal, en 3, est basophile, car il est riche en reticulum rugueux. En 4, une membrane basale entoure chaque acinus.

• En 1, la coupe transversale d'un acinus nous montre sa lumière, étroite, bordée par les cellules sécrétantes pyramidales. Vu l'étroitesse de cette lumière, la plupart des sections au niveau des acini passent comme en 2, en dehors de la lumière, dans les cellules sécrétantes.

• En 1, la lumière étroite d'un acinus se prolonge en 2, par de fins conduits radiaires pénétrant entre deux cellules sécrétantes voisines. Ce sont les canalicules intercellulaires, simples dépressions sans paroi propre, limitées de part et d'autre par les membranes des deux cellules adjacentes. Ces canalicules augmentent la surface d'évacuation de la sécrétion qui peut être excrétée latéralement, avant de passer dans la lumière de l'acinus.

Tissus conjonctif
• Dense, lache• Adipeux/cartilagineux, osseux• Soutien/• Mesoderme• Cellules :
fibroblastes,adipocytes,mastocytes,osteocytes, macrophages, chondocytes…
• Matrice extracellulaire/ substance fondamentale• Fibres reticulaires, collagesnes et elastiques

Colorations spécifiques.• Pour visualiser les autres types de
fibres conjonctives, il faut recourir à d'autres colorations spécifiques. Une orcéine colore les fibres élastiques en brun. On voit en 1 que ces fibres élastiques sont fines et possèdent généralement un trajet sinueux. Les autres fibres conjonctives, c'est-à-dire collagènes et réticuliniques, sont marquées en 2 par une contre-coloration au safran.

Colorations spécifiques.• Les fibres réticuliniques sont
spécifiquement mises en évidence par un Wilder qui est une imprégnation argentique contre-colorée ici par un vert lumière. L'imprégnation argentique marque les fibres réticuliniques en noir. Elles sont disséminées dans l'ensemble du conjonctif, mais concentrées en 1 au niveau de la membrane basale qui sépare tout tissu épithélial en 2, de tout tissu conjonctif, en 3.

Colorations spécifiques.
• Nous observons toujours un Wilder. Seule la contre-coloration diffère. Au niveau de la membrane basale, en 1, les fibres réticuliniques, marquée en noir, présentent un aspect grillagé très caractéristique.

Membrane basale• La membrane basale sépare
tout tissu épithélial de tout tissu conjonctif. Ainsi, en 1, un Wilder révèle la présence de fibres réticuliniques autour d'un épithelium glandulaire. Cet épithélium, en 2, est cylindrique simple. Il s'agit d'une glande sudoripare.

Fibres de reticuliniques/membrane basale
• Voici la lumière d'un vaisseau sanguin bordée en 1 par un épithélium pavimenteux simple. Un Wilder montre l'existence de fibres réticuliniques, au niveau de la membrane basale située juste sous le mince endothélium, en 2. Mais de plus, en 3, les fibres réticuliniques forment une sorte de grillage dans la paroi de certains vaisseaux

• Les fibres réticuliniques de la membrane basale sont séparées par une substance fondamentale particulièrement riche en mucopolysaccharides qui sont mis en évidence par la méthode de P.A.S., en 1. En 2, la substance fondamentale du tissu conjonctif sous-jacent contient nettement moins de mucopolysaccharides.
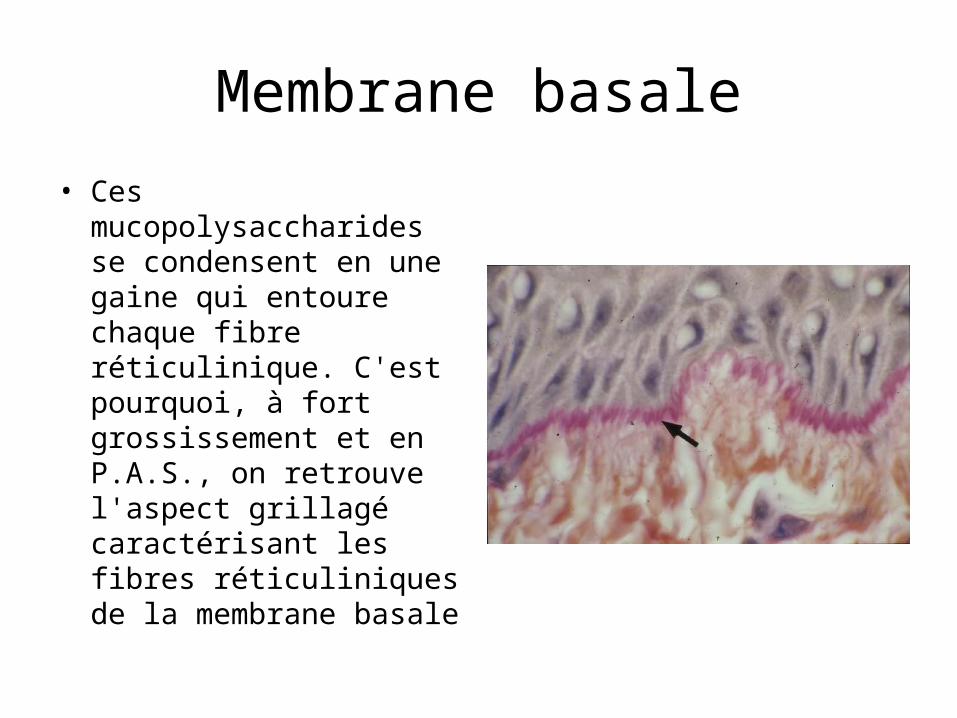
Membrane basale• Ces mucopolysaccharides se
condensent en une gaine qui entoure chaque fibre réticulinique. C'est pourquoi, à fort grossissement et en P.A.S., on retrouve l'aspect grillagé caractérisant les fibres réticuliniques de la membrane basale

• Sur un P.A.S., on repère, en 1, une membrane basale qui entoure ici une portion sudoripare. L'incidence est oblique, donc l'épithélium est difficile à observer. On voit en 2, un noyau entouré par une masse cytoplasmique importante. C'est un cylindrique simple

Fibroblastes• Après avoir détaillé les différents
types de fibres conjonctives et la composition de la substance fondamentale, il nous reste à décrire les types de cellules rencontrées dans le tissu conjonctif. Nous observons ici plusieurs fibroblastes. Ce sont des cellules à noyau généralement fusiforme et à cytoplasme très effilé si bien qu'on ne le perçoit généralement pas sur les préparations optiques. Les fibroblastes interviennent dans la synthèse des fibres conjonctives.

• En microscopie électronique, nous observons un fibroblaste. Son noyau est allongé, ovoïde. Le cytoplasme qui l'entoure en 1 est mince, étiré en un fin liseré périnucléaire. Il émet en 2 de nombreux prolongements très fins qui se dirigent entre les faisceaux de fibres collagènes.

Abces• Nous aurons l'occasion
d'étudier les différents types de cellules à caractère histiocytaire dans une coupe de foie pathologique où l'on remarque, en 1, la présence de petits abcès autour desquels se produit, en 2, une réaction histiocytaire. En 3, subsiste du parenchyme hépatique tout à fait normal.

Histiocyte• L'histiocyte, fléché sur cette
image, constitue la première forme de ces cellules. Il s'agit d'un monocyte qui vient de quitter les vaisseaux sanguins pour passer dans le conjonctif. Il se caractérise par un noyau volumineux, à chromatine fine et dispersée. Ce noyau est encoché et donc réniforme.

Macrophages• Rapidement l'histiocyte se
transforme en macrophage fléché en 1, que nous observons ici dans une zone bourrée de lymphocytes, en 2. Le noyau s'est arrondi. Il est volumineux, clair, et unique. Ce macrophage a phagocyté ici des débris de cellules voisines lysées. On voit, dans son cytoplasme, des déchets nucléaires sous forme d'amas denses. En 3, nous voyons le cytoplasme d'un macrophage voisin également bourré de phagosomes.
•

• Diverses cellules sanguines jouent un rôle dans les processus inflammatoires et se retrouvent dans le conjonctif. En 1, des cellules à noyau arrondi et dense, borde d'un fin liseré cytoplasmique sont des lymphocytes. En 2, toutes les cellules bourrées de granulations acidophiles et à noyau plurilobé souvent même bilobé sont des éosinophiles. En 3, un neutrophile a un noyau plurilobé, mais un cytoplasme plus clair.

• Des réactions inflammatoires se produisent autour de tout corps étranger dans un tissu. Ainsi, dans une coupe de peau, nous trouvons en 1 une épine qui s'est implantée. Ses cellules sont très géométriques et régulières. Tout autour en 2 apparaît une réaction inflammatoire où l'on peut trouver diverses cellules à
caractère histiocytaire.

• En polarisant on peut mieux voir la partie de l'épine qui est anisotrope. On retrouve au niveau des flèches des morceaux d'épine dans les cellules qui forment un manchon autour de ce corps étranger. Ceci prouve donc bien le caractère macrophagique de ces cellules.

• Nous observons ici une foule de macrophages qui ont résorbé un exsudat sanguin dans un tissu conjonctif. Les dépôts jaunâtres dans le cytoplasme sont des pigments d'hémosidérine qui résulte de la dégradation des globules rouges.

• Voici, en microscopie électronique à balayage un macrophage, fléché en 1, occupé à phagocyter un globule rouge, fléché en 2. Cette photo a été prise dans la rate qui est le siège de destruction des vieux globules rouges.

• Nous rechercherons des plasmocytes dans une glande mammaire où on les rencontre en grand nombre. En effet, les plasmocytes synthétisent des anticorps qui passeront dans le lait maternel. Les portions glandulaires de la glande mammaire, en 1, sont groupées en lobules. En 2, le tissu conjonctif intralobulaire est très cellulaire. C'est là que se trouvent les nombreux plasmocytes. Par contre, en 3, le conjonctif entourant les lobules est plus riche en fibres.

• Dans le conjonctif intralobulaire de la glande mammaire, nous voyons plusieurs plasmocytes en 1. Ils se caractérisent par un noyau en position tout à fait excentrique, parfois double ; une chromatine condensée le plus souvent en damier ; un cytoplasme basophile. Souvent, en 2, on distingue une zone moins basophile, en croissant clair au-dessus du noyau: c'est le centre cellulaire. En 3, se trouve la portion glandulaire.

• Voici un plasmocyte observé en microscopie électronique. En 1, son noyau excentrique possède une hétérochromatine abondante, amassée en damier ; en 2, son cytoplasme est très riche en R.E.R. qui intervient dans la synthèse des anticorps, ceci explique la basophilie de ce cytoplasme en optique ; en 3, le centre cellulaire, moins basophile, est la zone du Golgi que l'on voit mal ici.

Mastocyte• Les mastocytes sont des
cellules rondes ou ovoïdes. Leur noyau arrondi est unique et central. Le cytoplasme est bourré de grosses granulations rondes très acidophiles.Attention toutefois de ne pas le confondre avec un éosinophile qui contient, lui aussi de grosses granulations rouges, mais dont le noyau est plurilobé.

• Les mastocytes, en 1, se rencontrent le plus souvent près des petits vaisseaux sanguins. On voit en 2, l'endothélium (épithélium pavimenteux simple) qui délimite un vaisseau. Les granulations des mastocytes contiennent en effet de l'héparine, puissant anticoagulant. de l'histamine qui agit sur la vasodilatation.

• Voici deux mastocytes observés dans une coupe de langue de rat colorée au bleu de toluidine. Toute la coupe est bleu pâle, à l'exception des grains des mastocytes, fléchés en 1, qui ont fait virer le bleu de toluidine en violet. Ces grains ont donc la propriété de produire un virage métachromatique. Le noyau central, fléché en 2, n'a pas produit de métachromasie.

• Nous retrouvons les caractéristiques du mastocyte en microscopie électronique : cellule ovoïde, avec un noyau unique et central et un cytoplasme bourré de granulations. Ces grains sont de densité variable. Dans le conjonctif avoisinant, on reconnaît : en 1, un fibroblaste avec son noyau et peu de cytoplasme ; en 2, les prolongements cytoplasmiques de fibroblastes ; en 3, des faisceaux de fibres collagènes.

• Les cellules graisseuses ou adipocytes se groupent souvent en lobules et forment le tissu adipeux. Ces cellules sont polygonales, vu leur tassement. Dans le tissu adipeux secondaire, le cytoplasme apparaît vide. En 1, une grosse goutte lipidique a été dissoute lors des techniques. Le fin liseré cytoplasmique est refoulé en périphérie de même que le noyau en 2. Ceci donne à la cellule un aspect en "bague à chaton".

• Une coupe à congélation permet la conservation des graisses puisqu'il n'y a pas de passage dans les alcools. Diverses colorations propres aux graisses, ici le noir soudan, révèlent le gros globule graisseux qui emplit pratiquement toute la cellule, dans le cas d'un tissu adipeux secondaire.

• Le tissu adipeux primaire ou graisse brune possède des cellules plus petites, polygonales. Le noyau, en 1, est central, parfois légèrement excentrique. Les graisses dissoutes sont dispersées dans le cytoplasme en 2, sous forme de petites gouttelettes ; seul reste un fin réseau cytoplasmique, d'où l'aspect spongieux et le nom de spongiocyte. La graisse brune, peu fréquente chez l'homme est très développée chez les animaux hibernants.

• Classifions les différents conjonctifs localisés dans la peau, face palmaire. Dans le derme papillaire, à la base de l'épithélium, les trois éléments du tissu conjonctif, c'est-à-dire cellules, fibres et substance fondamentale sont en proportion quasi équivalentes. On le qualifie de tissu conjonctif lâche.

• Dans le derme moyen, les fibres prédominent. Les plus nombreuses sont les fibres collagènes qui forment des faisceaux denses, sans orientation spéciale. C'est un tissu conjonctif dense irrégulier.

• Dans l'hypoderme, se localisent des corpuscules de Vater Pacini. En 1, des terminaisons nerveuses sont entourées d'un conjonctif particulier. Des fibres collagènes, fléchées en 2, alternent régulièrement avec des fibroblastes, fléchés en 3 et se disposent concentriquement d'une façon régulière. C'est un exemple de tissu conjonctif dense régulier, qualifié ici de tissu conjonctif lamellaire.

Tendon• Le tendon est constitué d'un tissu
conjonctif dense. Les fibres collagènes prédominent. Elles se disposent, en 1, en faisceaux parallèles et denses. Les fibroblastes, en 2, s'agencent en files parallèles entre les fibres tendineuses. Il s'agit d'un conjonctif dense régulier. Remarquons que ces fibres collagènes gardent parfois la coloration rosée de l'érythrosine à cause de leur densité.

• Un autre exemple de tissu conjonctif dense régulier est le tissu conjonctif propre de la cornée. Nous observons en 1 des faisceaux collagènes bien orientés, séparés par des cellules (marquées en 2) qui portent le nom de kératocytes. Les fibres collagènes d'un faisceau sont parallèles entre elles, mais chaque faisceau est disposé perpendiculairement à son voisin. Cette alternance ne se voit pas ici.

Tissu conjoncyif muqueux: predominsnce de la substance fondamentale
• Dans le cordon ombilical, la gelée de Wharton est le seul exemple de tissu conjonctif où la substance fondamentale prédomine. Etant dissoute par la plupart des fixateurs, on doit la situer dans tous les espaces clairs, en 1 . On rencontre en outre en 2 quelques fibres conjonctives et en 3 des cellules mésenchymateuses particulières émettant des prolongements anastomosés.
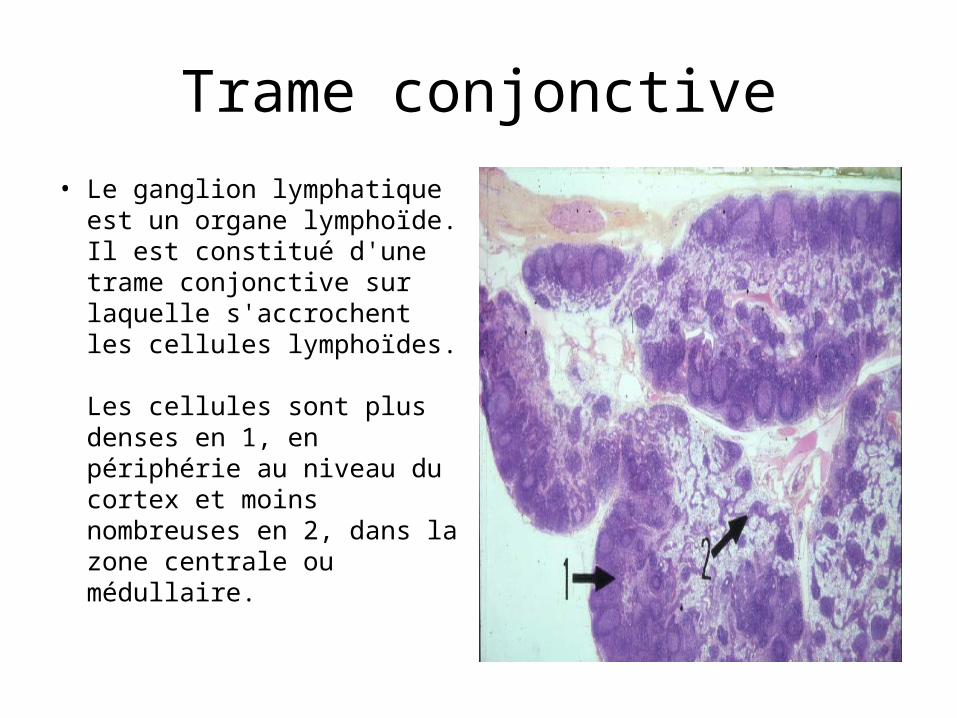
Trame conjonctive• Le ganglion lymphatique est
un organe lymphoïde. Il est constitué d'une trame conjonctive sur laquelle s'accrochent les cellules lymphoïdes. Les cellules sont plus denses en 1, en périphérie au niveau du cortex et moins nombreuses en 2, dans la zone centrale ou médullaire.

• La trame de base du ganglion lymphatique est un tissu conjonctif réticulaire. On l'observe bien dans la zone centrale ou médullaire du ganglion, là où la densité des cellules lymphoïdes est moindre. Ce tissu est riche en cellules fléchées en 1 qui forment un premier réseau. Elles ont en effet de nombreux prolongements qui s'anastomosent avec les cellules voisines. Sur ce réseau, s'accrochent, en 2, les cellules lymphoïdes.

• Le réseau cellulaire est doublé d'un second réseau de fibres réticuliniques ainsi que le révèle ici l'imprégnation argentique d'un Wilder.

Tissu cartilagineux
• Fibres elastiques,collagenes,• Matrice solidifiee• Vascularise, non innerve,• Chondrocytes,Fibroblastes,

• Cette coupe longitudinale de la trachée va nous permettre d'étudier le tissu cartilagineux. La paroi trachéale contient en effet de nombreux nodules cartilagineux marqués par les flèches. Le cartilage est un tissu conjonctif particulier, avec des cellules, des fibres et une substance fondamentale qui a la particularité d'être solide. Il se colore donc en jaune sur un H.E.S.
•

Trachee• La lumière de la trachée est
directement bordée, en 1, par un épithélium respiratoire sous lequel se localisent, en 2, des unités sécrétantes tubulo-acineuses. Le tissu cartilagineux, en 3, se présente sous forme de nodules. Il s'est formé par métaplasie du tissu conjonctif avoisinant, fléché en 4, appelé périchondre.

Perichondre• En périphérie du nodule
cartilagineux, nous trouvons le périchondre. Il possède, en 1, un feuillet stérile externe, riche en fibres conjonctives et, en 2, un feuillet fertile interne, beaucoup plus cellulaire. Ce périchondre se transforme pour donner du tissu cartilagineux. C'est un phénomène de métaplasie.

Périchondre• En 1, le feuillet stérile du
périchondre contient des fibres conjonctives très denses et des fibroblastes aux noyaux allongés, marqués par la tête de flèche. En 2, dans le feuillet fertile, les cellules s'arrondissent et élaborent la substance cartilagineuse. Ce sont les chondroblastes. En 3, la cellule complètement entourée de cette substance est le chondrocyte, logé dans son chondroplaste. C'est la croissance appositionnelle du cartilage.

Croissance cartilage• Le cartilage peut aussi s'accroître
de façon interstitielle, c'est-à-dire par mitose des chondrocytes. Si les mitoses se succèdent dans des directions diverses, on aboutit à un groupe de chondrocytes disposés circulairement en 1 : ce sont les groupes isogéniques coronaires. Les chondrocytes, logés dans leur chondroplaste, sont séparés en 2 par une jeune substance cartigineuse, plus pâle.

Croissance du cartilage• Au contraire, les mitoses peuvent
se faire successivement suivant une seule direction de l'axe du fuseau mitotique. On aboutit alors à un groupe de chondrocytes alignés, comme l'indiquent les flèches. Ce sont les groupements isogéniques axiaux. Dans ces groupements, les chondrocytes apparaissent
"empilés".
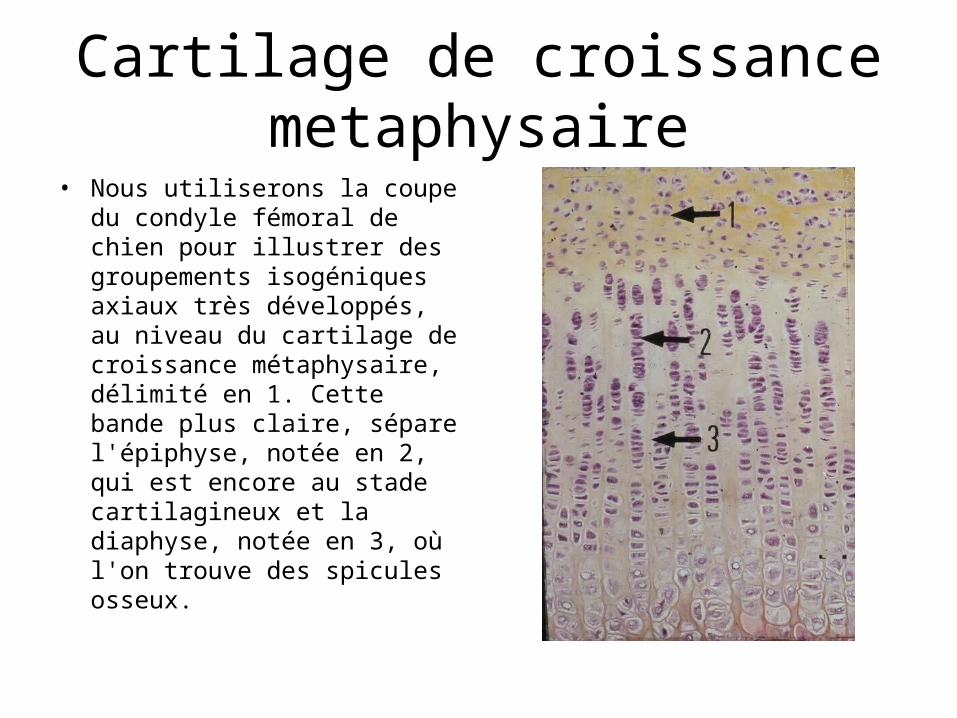
Cartilage de croissance metaphysaire
• Nous utiliserons la coupe du condyle fémoral de chien pour illustrer des groupements isogéniques axiaux très développés, au niveau du cartilage de croissance métaphysaire, délimité en 1. Cette bande plus claire, sépare l'épiphyse, notée en 2, qui est encore au stade cartilagineux et la diaphyse, notée en 3, où l'on trouve des spicules osseux.

Cartilage de croissance metaphysaire
• Nous retrouvons ici ce même cartilage de croissance, en métachromasie car la substance cartilagineuse est métachromatique. Les chondrocytes dégénèrent progressivement. En 1, ils se superposent en "piles d'assiettes". En 2, ils s'hypertrophient, de même que leur logette ou chondroplaste. En 3, ils se vacuolisent puis dégénèrent, tandis que la substance cartilagineuse se calcifie.

Groupements isogéniques coronaires, groupements isogéniques axiaux, ligne d'érosion
• En 1, le cartilage épiphysaire, avec ses nombreux groupements isogéniques coronaires. En 2, le cartilage de croissance constitué de groupements isogéniques axiaux excessivement développés. Nous avons vu que ces chondrocytes dégénèrent. A leur base, en 3, ils sont détruits par de grandes cellules, appelées chondroclastes. Cette ligne d'attaque des chondroclastes constitue la ligne d'érosion.

Ligne d'érosion • Nous situons en 1 les
chondrocytes hypertrophiés du cartilage de croissance. En 2, au niveau de la ligne d'érosion, les chondroclastes détruisent ce cartilage, dégénéré. Il s'agit de cellules géantes, plurinucléées. Attention toutefois : plus bas, en 3, apparaît un tissu plus coloré, qui est de l'os. La cellule géante fléchée en 4 est donc un ostéoclaste et non pas un chondroclaste.

Cartilage articulaire• Au niveau des articulations,
subsiste une zone de cartilage. C'est le cartilage articulaire. On y reconnaît en 1, des groupements isogéniques coronaires et, en 2, axiaux, résultant de la croissance interstitielle. Il n'existe plus ici de croissance appositionnelle, donc plus de périchondre. Les cellules fléchées en 3 sont donc des chondrocytes tassés et non des chondroblastes.

Cartilage articulaire • Avec une réaction de
métachromasie, nous observons ces groupements isogéniques du cartilage articulaire. On voit bien le chondroplaste, jeune substance cartilagineuse, qui entoure chaque chondrocyte.

Substance fondamentale du cartilage
• On reconnaît un tissu cartilagineux à la présence, en 1, de petits groupements isogéniques. Un PAS, met en évidence, en 2, la substance fondamentale de ce cartilage. Elle contient des mucopolysaccharides acides, plus particulièrement l'acide chondroitine sulfurique. Les chondrocytes contiennent du glycogène, PAS positif. Il fuit devant le fixateur, ceci donne, en 3, l'artéfact classique en "nid d'hirondelle".

Cartilage élastique • Le cartilage du pavillon de
l'oreille, de l'aile du nez et de l'épiglotte chez l'homme, est un cartilage dit "élastique", car les fibres élastiques y sont très nombreuses. Sur un H.E.S. comme ici, elles apparaissent (en 1) assez rosées. On reconnaît toujours facilement un tissu cartilagineux au fait que les chondrocytes, forment, en 2, des groupements isogéniques.

Cartilage élastique
• Une coloration à l'orcéine met en évidence, en brun, les nombreuses fibres élastiques qui entourent les chondrocytes d'un cartilage élastique.

Cartilage élastique
• Une autre coloration spécifique des fibres élastiques est la fuchsine paraldéhyde qui révèle, en 1, ce réseau dense de fibres élastiques entourant les groupements isogéniques, fléchés en 2.

Tissux osseux
• Substance mineralisee/ osteoide( calcium, phosphate, fluor, cristaux d’hydroxyapatite)
• Cellules: osteoblastes, osteocytes, osteoclastes• Fibres collagenes• Systeme de Havers• Os long, os courts• Perioste• Moelle osseuse, Travees osseuses

Vue générale du pariétal d'un embryon et du cuir chevelu
• Voici une coupe dans le pariétal d'un embryon où nous pourrons étudier la formation du tissu osseux. La zone superficielle de la peau contient des ébauches embryonnaires de follicules pileux que l'on voit, en 1, en coupe transversale. En 2, un décollement réel permet un glissement de la peau sur le pariétal sous-jacent, constitué, en 3, de spicules osseux.

Spicules osseux • C'est au niveau de ces
spicules osseux que nous étudierons la formation du tissu osseux par métaplasie à partir du tissu conjonctif environnant. Sur coupe, ces spicules d'os semblent fragmentés. En réalité, sur une vue à trois dimensions, ces différents spicules s'anastomosent les uns aux autres.

Métaplasie du tissu conjonctif en tissu osseux
• Voici, en 1, une ébauche de formation de tissu osseux. Le conjonctif environnant est riche en fibroblastes, fléchés en 2. Ces cellules très effilées et aplaties vont se métaplasier en se transformant tout d'abord en ostéoblastes (fléchés en 3), cellules plus volumineuses, plus géométrique et à noyau plus arrondi.

Métaplasie du tissu conjonctif en tissu osseux
• La métaplasie du tissu conjonctif en tissu osseux s'observe nettement ici ; fléchés en 1, les fibroblastes du tissu conjonctif se transforment en ostéoblastes, fléchés en 2. Ces cellules, géométriques, se disposent étroitement les unes à côté des autres, à la manière d'un épithélium. Ces ostéoblastes en disposition épithélioide modifient la substance fondamentale. Une fois emprisonnés dans cette substance fondamentale modifiée, ils deviennent des ostéocytes (fléchés en 3).

Métaplasie du tissu conjonctif en tissu osseux
• Nous voyons: en 1, les fibroblastes du conjonctif avoisinant le spicule osseux ; fléchée en 2, la disposition épithélioide des ostéoblastes ; indiqués en 3, les ostéocytes, souvent rétractés par la fixation à l'intérieur de leur logette ou ostéoplaste. Les ostéocytes ne se divisent plus. Il n'existe donc pas de groupements isogéniques, différence majeure avec le tissu cartilagineux.

Résorption osseuse• Le tissu osseux est aisément
identifiable par, en 1, la disposition épithélioïde des ostéoblastes et par l'absence de groupes isogéniques. Les ostéocytes en 2, sont largement isolés par la substance osseuse. Au niveau du pariétal, pour permettre l'expansion du cerveau, ont lieu simultanément en 1 formation du tissu osseux à la face externe de l'os et résorption osseuse à la face interne par des cellules géantes, les ostéoclastes fléchés en 3.

Résorption osseuse • En 1, du tissu osseux. En 2,
une cellule plurinucléée ou ostéoclaste occupée à phagocyter un ostéocyte (fléché en 3). En 4, une zone floue apicale témoigne de la présence d'une bordure en brosse qui augmente la surface de contact cellulaire. En 5, on donne le nom de lacune de Howship à cette zone blanche, zone osseuse qui vient d'être détruite.

Calcification de la substance osseuse
• La méthode de Von Kossa substitue de l'argent aux ions calcium et colore donc en brun la substance osseuse calcifiée, comme en 1. Cette méthode révèle également en 2, la présence d'une jeune substance osseuse non calcifiée et donc non colorée en brun: c'est le liseré pré-osseux coloré ici par le safran situé sous l'assise épithélioide des ostéoblastes (fléchée en 3).

Calcification de la substance osseuse
• Est fléché en 1, un spicule osseux bien formé, où l'on retrouve en brun la substance osseuse calcifiée colorée par le Kossa et en 2, coloré en jaune pâle, le liseré pré-osseux. En 3, on assiste au début de la formation d'un autre spicule osseux qui n'en est encore pratiquement qu'au stade de substance pré-osseuse, non calcifiée.

Résorption osseuse
• Nous voyons en 1 le reste d'une travée osseuse calcifiée, marquée par la méthode Von Kossa. Elle est presque entièrement résorbée par les ostéoclastes, fléchés en 2.

Comparaison du tissu osseux et du tissu cartilagineux
• Voici une vue générale des fosses nasales où l'on rencontre à la fois une armature de tissu cartilagineux, en 1 et du tissu osseux en 2. Déjà à faible grossissement on les distingue tous deux par leur différence de coloration: le tissu osseux apparaît plus foncé que le tissu cartilagineux.

Comparaison du tissu osseux et du tissu cartilagineux
• Sur un détail, on distingue plus facilement encore ces deux types de tissu. Dans le tissu cartilagineux, les cellules ou chondrocytes se disposent toujours, en 1 en petits groupements isogéniques. Les ostéocytes, en 2, restent bien isolés et individualisés. D'autre part, autour du tissu osseux, en 3, nous retrouvons la disposition épithélioïde des ostéoblastes.

Comparaison du tissu osseux et du tissu cartilagineux
• Au niveau de l'articulation d'une patte de veau, on rencontre également os et cartilage. En surface, se trouve le cartilage articulaire, indiqué en 1, qui apparaît plus clair et d'où se détachent, en 2, un ensemble de lames anastomosées de tissu osseux, plus fortement coloré. Fléché en 3, du tissu conjonctif sépare ces travées osseuses.

Cartilage articulaire• Voici une vue du cartilage
articulaire où l'on voit apparaître progressivement depuis la périphérie vers la base du cartilage des groupes isogéniques de plus en plus importants, que ce soit, en 1, des groupes isogéniques coronaires ou, en 2 des groupements isogéniques axiaux.

Cartilage articulaire et os sous-jacent
• De ce cartilage articulaire, noté en 1, partent, en 2, de nombreuses travées osseuses anastomosées entre elles. A moyen grossissement, la différence de coloration permet de les reconnaître facilement.

Comparaison du tissu osseux et du tissu cartilagineux
• A la limite os/cartilage, on rencontre des imbrications de ces deux tissus l'un dans l'autre. On voit encore très bien ici, en 1, les groupes isogéniques caractéristiques du tissu cartilagineux. Il est encerclé en 2 par un tissu osseux dont les critères ont éte signalés précédemment.

Tissus musculaires
• Cellules allongees: fibre musculaire• Sarcoplasme : organites, fialaments d’actine
et myosine• M. strie : unite contractile= sarcomere/strie
z…,noyaux en peripherie• M. lisse: cellule allongee, forme en
banane/faisceaux, noyau central• M.cardiaque: cellules anastomosees strie
scalariforme, noyau central

Tissu musculaire• Pour l'étude des
caractéristiques du tissu musculaire lisse, nous observerons la paroi de la vessie. Nous devinons en 1 l'épithélium urinaire qui borde la lumière. La paroi de la vessie contient en 2 de nombreux faisceaux musculaires lisses que nous pourrons détailler sous diverses incidences.

Cellule musculaire fusiforme• En section longitudinale, la cellule
musculaire lisse est fusiforme, très allongée. En 1, son noyau occupe le centre cellulaire, là où le diamètre est le plus large. Il est allongé dans le sens de la cellule et est unique. Le cytoplasme contient les myofibrilles, éléments contractiles, qui sont disposées selon le grand axe de la cellule tout en laissant libre, en 2, un cône cytoplasmique périnucléaire qui contient les organites cellulaires.

Cellule musculaire lisse contractee
• Quand la cellule est contractée, le noyau peut se plisser, prenant un aspect dit en "tire-bouchon".

Noyau cellules musculaires• Cette image en "tire-
bouchon" du noyau se retrouve ici, dans des cellules musculaires lisses qui occupent la paroi d'un vaisseau sanguin. Ce noyau tire-bouchonné est propre aux cellules musculaires lisses

Faisceau de cellules• Le plus souvent, les cellules
musculaires lisses se disposent en faisceaux, s'imbriquant étroitement les unes entre les autres en quinconce. Du tissu conjonctif, surtout constitué de fibres réticuliniques et collagènes, assure la cohésion de l'ensemble: on le voit, marqué par la flèche, sur une coupe tangentielle des cellules.

Reseau de fibres reticuliniques
• Un Wilder met en évidence le fin réseau de fibres réticuliniques qui entoure chaque cellule musculaire lisse. On a donné le nom de manchon pellucide à ce réseau.

Faisceau de cellules musculaires lisses en coupe transversale
• En coupe transversale, les cellules musculaires lisses se caractérisent par une inégalité de diamètre des plans de section, selon que la coupe passe, comme en 1, dans l'extrémité effilée de la cellule, ou, comme en 2, plus au centre. Ces sections ont un aspect polygonal vu le tassement des cellules imbriquées entre les autres.

Faisceau de cellules musculaires lisses en coupe transversale
• L'irrégularité de diamètre des cellules musculaires lisses en coupe transversale se voit mieux à fort grossissement. En 1, les sections très minces sont celles qui passent par l'extrémité effilée d'une cellule. En 2, les sections les plus larges passent par le noyau. Remarquez le peu de cytoplasme autour de ce noyau. En 3, des sections larges sans noyau ont été faites dans la
zone juxtanucléaire.
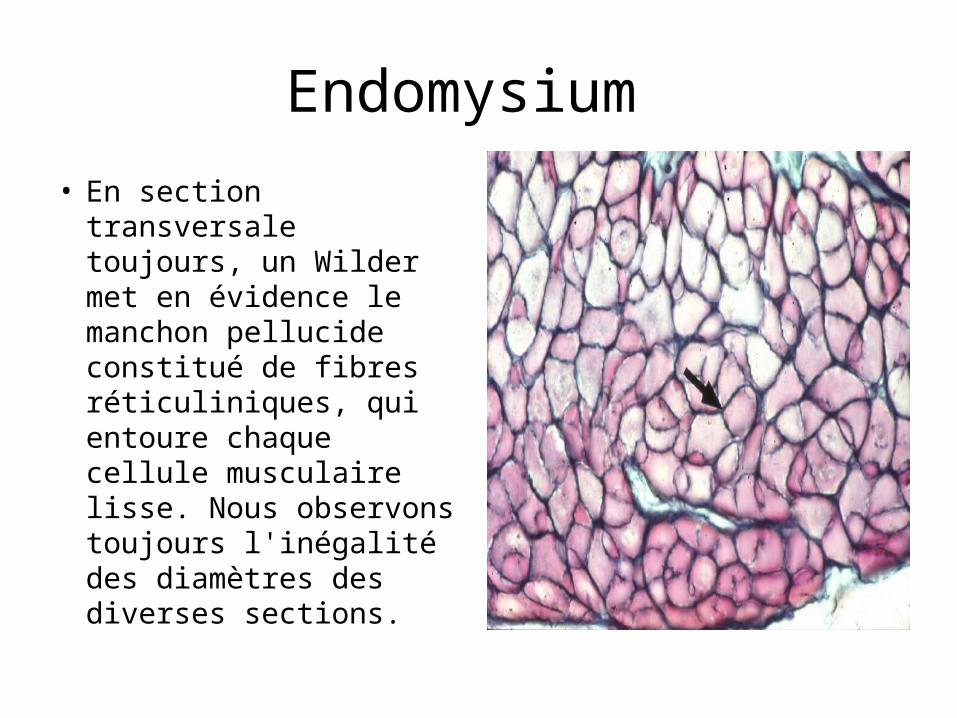
Endomysium
• En section transversale toujours, un Wilder met en évidence le manchon pellucide constitué de fibres réticuliniques, qui entoure chaque cellule musculaire lisse. Nous observons toujours l'inégalité des diamètres des diverses sections.

Tissu musculaire squelettique

Vue générale d'une langue • C'est dans une coupe de
langue que nous étudierons le tissu musculaire strié squelettique. La langue est un organe essentiellement musculaire. On voit déjà, à faible grossissement, les nombreux amas rosés dans le conjonctif.

Diverses incidences de coupe; périmysium
• Les cellules musculaires striées se regroupent en faisceaux dont l'orientation peut varier. C'est ainsi que nous voyons côte à côte en 1 des faisceaux coupés longitudinalement et, en 2, coupés transversalement. Le tissu conjonctif qui entoure chaque faisceau, fléché en 3 par exemple, porte le nom de périmysium.

Incidences de coupe:longitudinale et transversale
• En 1, en coupe longitudinale, la cellule musculaire striée squelettique a la forme d'un long cylindre de diamètre régulier. Les noyaux, marqués par les flèches sont multiples et rejetés en périphérie tout au long de la cellule. Dès lors, en coupe transversale (en 2), les diamètres des surfaces de section sont de taille beaucoup plus homogène. Ces sections sont de forme polygonale.

Striations• Le cytoplasme de la cellule
musculaire striée squelettique est rempli de myofibrilles fléchées en 1 allongées dans le sens de la cellule, ce qui donne une légère striation longitudinale. Mais de plus, en 2, apparaît une striation transversale avec alternance de bandes claires et de bandes sombres. C'est la raison pour laquelle on appelle ce muscle: muscle strié.

Striations• Sur une coloration particulière,
on voit en 1, les disques sombres encore appelés disques A car ils sont anisotropes en polarisation ; en 2, les disques clairs sont isotropes et donc appelés disques I. En leur centre, en 3, est fléchée une ligne plus dense: la raie Z. L'espace délimité par 2 raies Z successives porte le nom de sarcomère. Au centre de la bande A, en 4, apparaît une zone plus claire: c'est la bande H.

Striations• Sur cette image, la striation
transversale est particulièrement évidente. En 1, la bande A est bien marquée, contrastant ainsi d'autant plus avec la bande l fléchée en 2. Remarquez toutefois, en 3, que les myofibrilles d'une même cellules striée squelettique peuvent être à des états de contraction différents.

Coupe transversale dans un faisceau de fibres musculaires striées squelettiques; endomysium et périmysium
• En coupe transversale, les sections sont polygonales et de calibre assez régulier. Les noyaux étant multiples, la plupart des sections montrent un ou plusieurs noyau refoulés en périphérie de la cellule, comme en 1. Entre les cellules, se trouve du tissu conjonctif, fléché en 2 et appelé endomysium. Il contient de petits capillaires sanguins, fléchés en 3. Autour des faisceaux musculaires, le conjonctif, fléché en 4, constitue le périmysium.

Disposition des myofibrilles • En section transversale, les
myofibrilles apparaissent sous forme de petits points. Elles sont soit réparties de façon assez homogène dans la cellule, comme en 1, soit regroupées en petits amas constituant les champs de Conheim, fléchés en 2.

Champs de Conheim • Une autre image, pour nous
montrer en 1 la répartition en champs de Conheim des myofibrilles. Remarquez toujours la localisation périphérique des noyaux, lorsque la section les révèle. En 2, est fléché l'endomysium.

Myofibrilles parallèles.• En microsocpie électronique, on
distingue les nombreuses myofibrilles parallèles. En 1, est notée l'épaisseur d'une myofibrille. Elles sont séparées les unes des autres par une fine bande de cytoplasme appelé "sarcoplasme" ; Il renferme : en 2, du glycogène relativement abondant ; en 3, des mitochondries encore appelées "sarcosomes" ; en 4, des vésicules de réticulum lisse,souvent dilatées qui forment le réticulum sarcoplasmique.

Coupe longitudinale dans une fibre musculaire striée squelettique
• A plus fort grossissement, nous voyons trois myofibrilles séparées par du sarcoplasme. En 1, est fléchée la raie Z. L'espace compris entre deux raies Z consécutives forme le sarcomère, unité de base de la contraction musculaire. En 2, la bande claire ou bande l est formée de fins filaments d'actine. En 3, la bande A contient des filaments épais de myosine entre lesquels coulissent les fins filaments d'actine.

Coupe longitudinale dans une fibre musculaire striée squelettique
• Les bandes I, indiquées en 1, alternent régulièrement avec les bandes A, délimitées en 2.En 3, est fléchée la raie Z, zone d'attache des filaments d'actine de deux sacomères voisins. Au centre de la bande A, en 4, se localise la bande H qui ne contient que des filaments épais. En son sein, en 5, apparaît une ligne plus dense ou ligne M.

Muscle strié squelettique
• A un grossissement plus important, nous observons une triade de Porter et Palade en coupe longitudinale. En 1, est fléché le système T, invagination de la membrane plasmique. Il est flanqué de chaque côté, en 2, des deux citernes terminales du reticulum sarcoplasmique. Entre les deux membranes, on observe des pointillés denses qui sont des structures de couplage.

Deux parties de fibres musculaires striées squelettiques séparées par de l'endomysium
• En 1 est noté un des nombreux noyaux qui occupent le cytoplasme périphérique. On y observe en outre des nombreuses mitochondries, en 2. Chaque cellule musculaire est délimitée par le sarcolemme formé : en 3, de la membrane plasmique recouverte en 4 d'une lame basale. Les deux cellules sont séparées par l'endomysium, tissu conjonctif où nous voyons : en 5, des fibres conjonctives, en 6, le prolongement cytoplasmique d'un fibroblaste.

Divers niveaux de section des myofibrilles en coupe transversale• Nous voyons, en 1, une
myofibrille sectionnée au niveau de sa bande H puisque seuls les filaments épais de myosine y sont présents. En 2, des petites expansions latérales de ces filaments épais laissent penser que nous pourrions être au niveau de la ligne M.

Muscle lisse

Comparaison du tissu musculaire strié squelettique et du lisse
• Dans la tunique musculaire du passage oesophago-gastrique, nous trouvons côte à côte : en 1, des cellules musculaires striées squelettiques ; en 2, des cellules musculaires lisses. Nous pourrons les différencier en coupe longitudinales et en coupe
transversale.

Comparaison du tissu musculaire strié squelettique et du lisse
• En coupe longitudinale, la striation des cellules musculaires striées squelettiques est évidente. Les noyaux, fléchés en 1, sont nombreux et logés en périphérie de la cellule. En 2, les cellules musculaires lisses sont de diamètre beaucoup plus petits. Elles sont fusiformes et le noyau est unique et central.
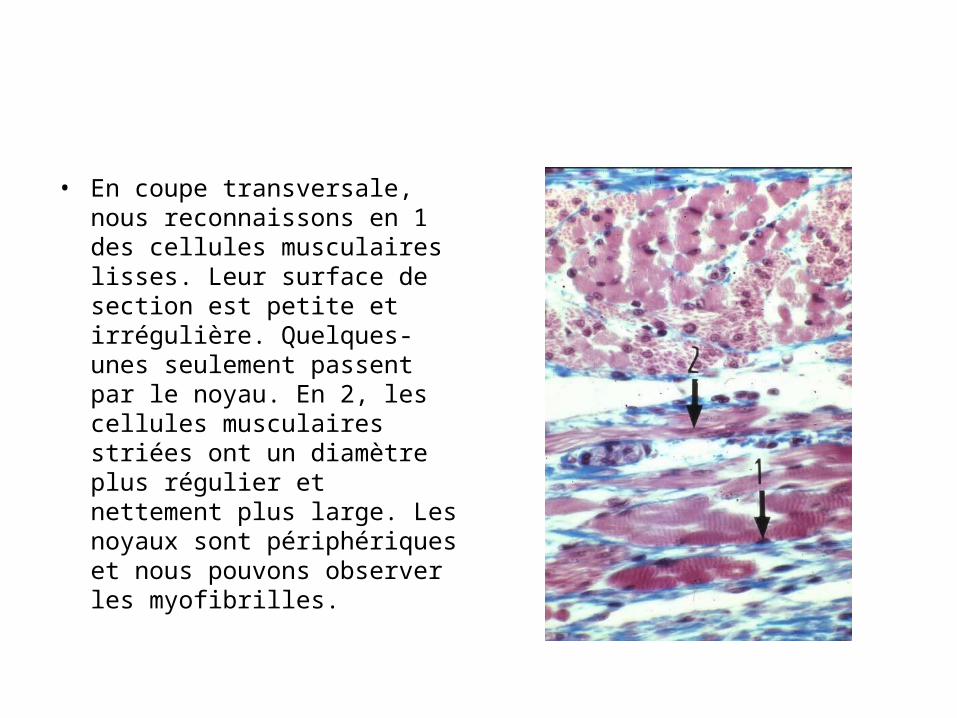
• En coupe transversale, nous reconnaissons en 1 des cellules musculaires lisses. Leur surface de section est petite et irrégulière. Quelques-unes seulement passent par le noyau. En 2, les cellules musculaires striées ont un diamètre plus régulier et nettement plus large. Les noyaux sont périphériques et nous pouvons observer les myofibrilles.

Muscle striee cardiaque

Vue générale du myocarde
• La paroi du coeur, encore appelée myocarde, est composée de cellules musculaires striées cardiaques ou cellules myocardiques que nous allons détailler.

Fibres musculaires striées cardiaques en coupe longitudinale• En coupe longitudinale, les
cellules musculaires striées cardiaques ont la forme de longs cylindres qui présentent, au niveau des flèches, de nombreuses ramifications anastomosées avec les cellules voisines. Elles forment ainsi un vaste réseau tridimensionnel complexe. Toutefois, chaque cellule reste bien individualisée.

Sang

Frottis sanguin• Voici un frottis sanguin coloré par
le May-Grumwald Giemsa. Les éléments figurés du sang sont d'une part, en 1, les globules rouges, et d'autre part les globules blancs. On y distingue, en 2, des monomorphonucléaires agranuleux (à noyau simple et sans granules cytoplasmiques) et en 3 des polymorphonucléaires granuleux (à noyau plurilobulé et dont 1e cytoplasme contient des grains.
•

Erythrocytes • Observons les globules rouges.
Ils sont anucléés chez l'homme et les mammifères, ont un diamètre d'environ 7 microns et présentent, en 1, une zone centrale plus claire, entourée d'un halo périphérique plus coloré. Ceci témoigne d'une dépression centrale du globule rouge. En 2, à cause de cette dépression, ils peuvent se
superposer.

• En microscopie électronique à balayage, on voit bien cette dépression centrale et l'empilement des globules rouges. Une dépression similaire existe sur la face opposée que l'on ne peut observer ici. Les globules rouges sont donc des cellules biconcaves.
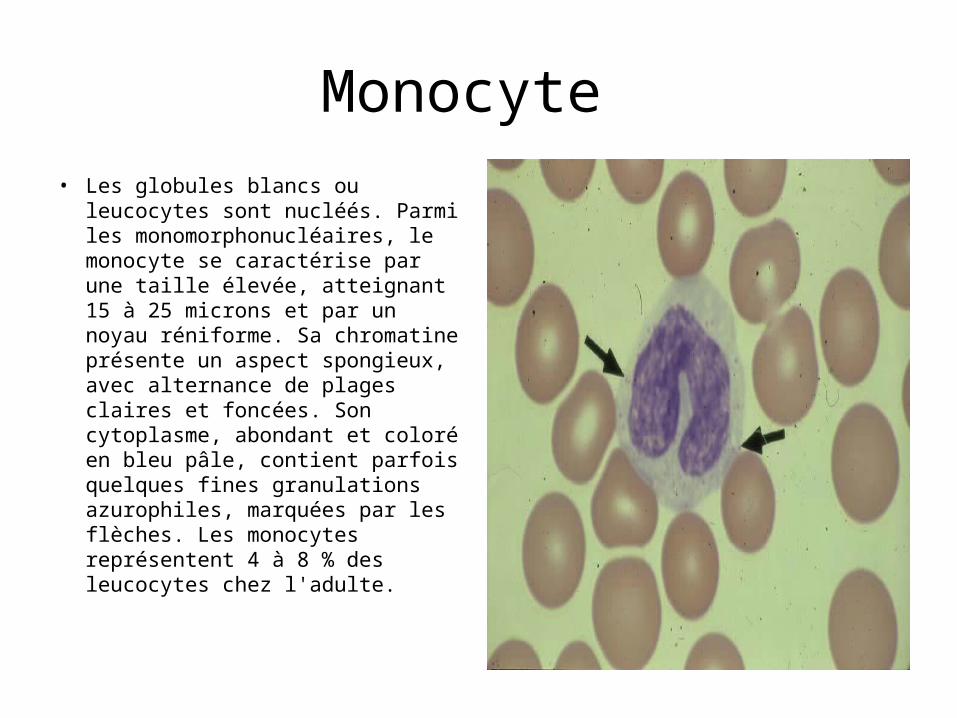
Monocyte • Les globules blancs ou leucocytes
sont nucléés. Parmi les monomorphonucléaires, le monocyte se caractérise par une taille élevée, atteignant 15 à 25 microns et par un noyau réniforme. Sa chromatine présente un aspect spongieux, avec alternance de plages claires et foncées. Son cytoplasme, abondant et coloré en bleu pâle, contient parfois quelques fines granulations azurophiles, marquées par les flèches. Les monocytes représentent 4 à 8 % des leucocytes chez l'adulte.

Monocyte • En microscopie électronique, sur
une bonne incidence, on observe le noyau réniforme déprimé en son centre. Dans l'encoche cytoplasmique se localise, en 1, un Golgi, généralement bien développé. Les granulations azurophiles se présentent comme en 2 : ce sont de petits lysosomes. Ils ne sont pas nombreux. Leur nombre et leur contenu enzymatique augmenteront lorsque la cellule se transformera en histiocyte .

Polymorphonucléaire neutrophile
• En microscopie electronique, l'aspect des granulations peut varier selon les espèces. Souvent, les granules sont subdivisés en fines granulations ponctuées emballées dans une membrane. La taille de ces fines particules peut varier d'un granule à l'autre, mais est généralement homogène au sein d'un même granule.

Polymorphonucléaire éosinophile• Les polymorphonucléaires
éosinophiles présentent généralement deux lobes dans leur noyau, plus rarement trois ou plus, toujours reliées par un petit pont nucléaire, marqué par la flèche. Ils sont caracterisés par la présence de grosses granulations très acidophiles, colorées en rouge vif et tassées dans leur cytoplasme. Les éosinophiles représentent 2 % des globules blancs.

Polymorphonucléaire éosinophile• En microscopie électronique,
nous observons le cytoplasme d'un éosinophile à un fort grossissement. Les granulations éosinophiles sont très particulières. Le plus souvent allongées, en forme de grains de riz, elles se caractérisent, en 1, par un cristalloïde dense aux électrons, au sein d'une matrice homogène ou granulaire plus claire. En 2, nous voyons un fragment des lobes nucléaires.

Thrombocytes• On observe ici un agrégat de
plusieurs plaquettes sanguines. Sur coloration au Giemsa, on distingue dans chaque plaquette, en 1, un point central bien coloré en violet: c'est le chromatomère. Il est entouré d'une zone plus pâle et homogène, en 2, appelée hyalomère. Les plaquettes sanguines interviennent dans les phénomènes de coagulation.