Embed Size (px)
Citation preview

UUNNIIVVEERRSSIIDDAADD PPOOLLIITTÉÉCCNNIICCAA DDEE MMAADDRRIIDD
EESSCCUUEELLAA TTÉÉCCNNIICCAA SSUUPPEERRIIOORR DDEE IINNGGEENNIIEERROOSS AAGGRRÓÓNNOOMMOOSS
TRABAJO FIN DE CARRERA
Estudio de la diversidad de poblaciones naturales de romero (Rosmarinus sp.) de la Península Ibérica mediante RAPDs
ÁÁllvvaarroo AAgguuaaddoo –– MMuuññoozz OOllmmeeddii ll llaa
MMaaddrr iidd ,, eenneerroo 22000077


Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs AAGGRRAADDEECCIIMMIIEENNTTOOSS
P á g i n a | I
AGRADECIMIENTOS
Quiero agradecer a todas las personas que han colaborado en este trabajo,
principalmente a mis tutoras Dña. Mª Carmen Martín Fernández y Dña. Elena
González Benito por la oportunidad que me han dado de realizar este estudio de
investigación, por su inestimable dirección y las horas que me han dedicado en este
trabajo.
Asimismo me gustaría dar las gracias a todos aquellos que a lo largo de este
tiempo han colaborado en mi formación, en especial a Beatriz Guerra que estuvo
ayudándome durante la fase experimental y a David Draper por la elaboración del
mapa donde quedaron reflejadas las poblaciones recolectadas.
Finalmente, deseo agradecer a mis padres, hermanos y a mi abuela, su apoyo
y confianza en mí.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs RREESSUUMMEENN
P á g i n a | II
RESUMEN
El presente trabajo se ha llevado a cabo en el contexto del proyecto de
investigación ‘Recolección, conservación y caracterización de germoplasma de
poblaciones españolas de varias especies de los géneros Rosmarinus y Origanum’
(Instituto Nacional de Investigaciones Agrarias), en el cual se plantea la
caracterización de la diversidad existente entre las poblaciones silvestres de romero
de España recolectadas y conservadas en el Banco de Semillas de la Universidad
Politécnica de Madrid (UPM).
Los objetivos del estudio han sido la puesta a punto de un protocolo para la
obtención de marcadores moleculares para caracterizar poblaciones de romero
(Rosmarinus officinalis L. y Rosmarinus eriocalyx Jordan & Fourr.), el estudio de la
diversidad interpoblacional y el análisis de factores (geográficos y ambientales) que
puedan explicar el grado de similitud entre las poblaciones estudiadas.
La metodología se basa en la obtención de marcadores moleculares RAPDs
(Random Amplified Polymorphic DNA) mediante el procesado de muestras de hoja
de trece poblaciones españolas de Rosmarinus. El nivel de similitud entre las
muestras estudiadas se analizó a partir de la matriz binaria de datos generada por
los RAPDs. Los análisis de clasificación de las muestras se realizaron por el método
de agrupamiento (UPGMA) y se representaron gráficamente mediante un
dendrograma. El test de correlación de Mantel se utilizó para comprobar el ajuste
de los dendrogramas en la representación del grado de similitud entre muestras, así
como en el análisis de la correlación entre matriz de distancias genéticas y las
matrices de distancias geográficas y ambientales. Finalmente, para estimar la
diversidad genética se utilizó la expresión propuesta por Nei de 1973 y la realización
de un análisis de varianza molecular (AMOVA).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs RREESSUUMMEENN
P á g i n a | III
Como conclusiones, la utilización de marcadores RAPDs ha permitido
establecer la singularidad de las poblaciones de las que proceden las muestras de
semillas conservadas en el banco de Germoplasma de la UPM. Existe una elevada
diversidad entre las poblaciones analizadas y también, en la mayoría de los casos,
entre las muestras de una misma población. Los resultados obtenidos también
permitieron detectar un posible error en la determinación de una de las poblaciones
estudiadas que había sido considerada como R. eriocalyx y que sin embargo los
análisis de los marcadores moleculares llevados a cabo parecen indicar que también
se trata de una población de R. officinalis. La falta de correlación observada entre la
matriz de distancias genéticas y las matrices de distancias geográficas y ambientales
refuerza la hipótesis de falta de diferenciación entre las distintas provincias
biogeográficas.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs AABBRREEVVIIAATTUURRAASS
P á g i n a | IV
Índice de ABREVIATURAS
- AFLP Amplified Fragment Lenght Polimorphism
- AMOVA Análisis de la Varianza Molecular
- d.e. desviación estándar
- dNTPs desoxinucleótidos
- PCR Polymerase Chain Reaction
- RAPD Random Amplified Polymorfic DNA
- RFLP Restriction Fragment Length Polimorphisms
- SSR Simple Sequence Repeats
- pb. pares de bases
- PVP polivinilpirrolidona

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs ÍÍNNDDIICCEE
P á g i n a | V
ÍNDICE
I. Introducción ..................................................................................................... pág.01
1.1. Descripción botánica y distribución de las poblaciones de romero ……………….. pág.01
1.1.1. Descripción geográfica y hábitat del género Rosmarinus …………….. pág.05
1.1.2. Especies objeto de estudio: R. officinalis y R. eriocalyx ………………... pág.06
1.2. Importancias de las labiadas: usos y cultivo ………………………………………... pág.09
1.2.1. Cultivo del romero ……………………………………….……………… pág.12
1.3. Importancia de la conservación ……………………………………………...…........ pág.14
1.4. Caracterización del material conservado …………………………………………… pág.20
1.4.1. La caracterización vegetal: diferentes enfoques ………………………. pág.22
1.4.2. La caracterización mediante marcadores moleculares ………………... pág.23
1.5. Técnicas moleculares usadas en caracterización de poblaciones …………………. pág.25
1.5.1. Tipos de marcadores moleculares …………………….………………... pág.25
1.5.2. Tipos de ensayos de marcadores moleculares ………………………… pág.26
1.6. Utilización de marcadores moleculares en estudios de poblaciones ……………… pág.33
1.7. Estudios de poblaciones de romero ………………………………………………… pág.34
II. Interés y objetivos …………………………………………………………… pág.35

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs ÍÍNNDDIICCEE
P á g i n a | VI
III. Material y métodos ……………………….…………………………………. pág.36
3.1. Material vegetal ……………………………………………………………………….. pág.36
3.1.1. Poblaciones estudiadas ………………………………………………………… pág.36
3.1.2. Recolección del material ……………………………………………………….. pág.38
3.1.3. Conservación del material ……………………………………………………… pág.39
3.2. Extracción de ADN ……………………………………………………………………. pág.40
3.2.1. Protocolo de extracción de ADN ………………………………………………. pág.40
3.2.2. Preparación de las soluciones de trabajo (ST) ………………………………… pág.41
3.3. Amplificación del ADN (RAPDs) ………………………………………………………. pág.41
3.3.1. Protocolo de amplificación mediante PCR …………………………………….. pág.42
3.4. Separación de los productos de amplificación por electroforesis ………………….. pág.43
3.4.1. Electroforesis en gel de agarosa al 1,5 % en TBE ………………………………. pág.43
3.4.2. Tinción y adquisición de imágenes …………………………………………….. pág.44
3.5. Análisis estadísticos …………………………………………………………………... pág.44
IV. Resultados y discusión ………………………………………………………. pág.47
4.1. Caracterización de las poblaciones de romero ………………………………………. pág.47
4.2. Análisis de agrupamiento …………………………………………………………….. pág.50
4.3. Distancias genéticas ….………………………………………………………………. pág.56

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs ÍÍNNDDIICCEE
P á g i n a | VII
4.4. Correlaciones entre distancias genéticas, geográficas y ambientales ……………... pág.59
4.4.1. Correlación entre las distancias geográficas y genéticas …………………….. pág.59
4.4.2. Correlación entre las distancias ambientales y genéticas ……………………. pág.60
4.5. Diversidad genética …………………………………………………………………... pág.62
4.5.1. Diversidad genética según Nei ………………………………………………… pág.63
4.5.2. Análisis de la varianza molecular (AMOVA) …………….…………………….. pág.66
4.6. Conclusiones ………………………………………………………………………….. pág.68
V. Bibliografía …….……………………………………………………………... pág.71
VI. Anejos ………………………………………………………………................ pág.78
Anejo I ……………………………………………………………………………………… pág.78
Anejo II ……………………………………………………………………………….......... pág.80
Anejo III ………………………………………………………………………………......... pág.83
Anejo IV ………………………………………………………………………………......... pág.90


Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 1
1. Introducción
El presente trabajo se ha realizado en el contexto de un proyecto de
investigación que lleva por título ‘Recolección, conservación y caracterización de
germoplasma de poblaciones españolas de varias especies de los géneros
Rosmarinus y Origanum’ financiado por el Instituto Nacional de Investigaciones
Agrarias (INIA).
Dentro de las tareas propuestas en dicho proyecto se encontraba la
caracterización mediante marcadores moleculares de las poblaciones silvestres de
romero recolectadas para su conservación en el Banco de Semillas de la Universidad
Politécnica de Madrid (UPM). La puesta a punto de la caracterización de esas
poblaciones y la obtención de unos resultados preliminares han dado lugar a este
trabajo.
1.1. Descripción botánica (clasificación taxonómica) y distribución de las
poblaciones de romero
El romero pertenece a la familia de las Labiadas (Lamiaceae), que se
encuentra dentro del orden Lamiales, subclase Asteriadae, clase Magnoliopsida,
División Magnoliophyta y reino Plantae (Cronquist, 1981). Todas o casi todas las
plantas de esta familia, consideradas como las plantas aromáticas por excelencia,
tienen el tallo cuadrado y las hojas opuestas, acopladas, una frente a la otra. En
general, las flores se forman a manera de una boca abierta, con ambas quijadas muy
separadas, llamadas labios de la corola: el labio superior, por lo común bilobulado y
algo más corto que el inferior, y éste trilobulado. El cáliz suele estar dividido en
cinco dientes o gajos (tanto iguales como desiguales). Casi todas poseen cuatro
estambres fértiles (una excepción es el romero), adheridos al tubo de la corola, y de
longitud casi siempre desigual: dos estambres largos y dos cortos. Las flores suelen

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 2
agruparse en inflorescencias de unas seis flores, entorno al tallo, y entre cada dos
hojas floríferas de las sumidades florales. El fruto se compone de cuatro
subunidades, que se ven en el fondo del cáliz, verdes y diminutas en la planta
florida; de color más o menos oscuro y endurecidas cuando llegan a la madurez. En
cada subunidad hay una semilla (Font Quer, 1981).
Las Labiadas agrupan a unas 3.000 especies de plantas herbáceas, de las
cuales, casi una décima parte vive en la Península Ibérica, ya que la mayoría de ellas
suelen crecer bien en suelos secos y soleados (Font Quer, 1981). En los países de la
región mediterránea viven unas 1.000 especies silvestres de labiadas
correspondientes a 48 géneros, aproximadamente la cuarta parte en número de
géneros y especies del total mundial (Morales, 2000). Según Flora Europaea existen
41 géneros de labiadas y 449 especies en toda Europa, de las cuales 290 se
encuentran en la Península Ibérica e Islas Baleares (corresponden a 36 géneros
diferentes, aproximadamente la tercer parte del total de la región Mediterránea)
(Morales, 2000). En la Península Ibérica viven 342 taxones de Labiadas (incluidas
subespecies), 11 especies viven en Baleares y 6 de ellas son endemismos de estas
islas. Centrándonos en España (incluidas las islas Baleares), las labiadas son en
general abundantes, pero escasean hasta desaparecer en las regiones húmedas del
norte y noroeste de la Península, desde Galicia al País Vasco.
Las propiedades aromáticas de estas plantas se encuentran en la esencia que
se halla alojada principalmente en las hojas, el cáliz y la corola, en el interior de unas
glándulas redondeadas que se suelen ver como diminutos puntitos brillantes a la luz
del sol, de color áureo o pálido. Cuando se hallan en las hojas, es frecuente que
mirándolas a contraluz aparezcan como otros tantos puntos traslúcidos (Font Quer,
1981).
Los géneros de la familia de las Labiadas se pueden agrupar de acuerdo al
número de especies que engloba cada uno, estableciéndose así tres grandes
grupos: el de los grandes, con más de 20 especies, el de los medianos de 5‐20, y los

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 3
pequeños, con menos de 5 especies. El grupo en el que nos vamos a centrar es el de
los pequeños, ya que en él se encuentra el género en estudio del presente trabajo.
Los 22 géneros pequeños, suman en total 41 especies, de ellas 2 endemismos
ibéricos y uno balear: Ancinos (3), Ajuga (4), Ballota (2), Calamintha (3), Cleonia (1),
Clinopodium (1), Dracocephalum (1), Glechoma (1), Horminum (1), Hyssopus (1),
Leonurus (1), Lycopus (1), Marrubium (3), Melissa (1), Melittis (1), Molucella (2),
Origanum (3), Prasium (1), Prunilla (4), Rosmarinus (3), Timbra (1) y Ziziphora (2)
(Morales, 2000).
La distribución del género Rosmarinus es entorno al mar Mediterráneo
(figura 1). Las tres especies incluidas en el género Rosmarinus son: R. officinalis L.
(figura 2), R. eriocalyx Jordan & Fourr (figura 3) y R. tomentosus Huber‐Morath &
Maire (figura 4). Esta última especie es endémica de una pequeña zona costera
entre las provincias de Granada y Málaga, con una distribución muy restringida y
cuyas poblaciones no se han incluido en este trabajo.
Fig. 1. Distribución del género Rosmarinus sp. Se aprecia la distribución entorno al Mediterráneo, destacando su alta presencia en el norte de África y la Península Ibérica (Fuente: de Bolòs y Vigo, 1995).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 4
Fig. 2. Rosmarinus officinalis L. Aspecto de las hojas y flores.
Fig. 3. Rosmarinus eriocalyx Jordan & Fourr. Detalles de las hojas y flores.
Fig. 4. Rosmarinus tomentosus Huber‐Morath & Maire. Detalle del arbusto. Presenta hojas dénsamente cubiertas con pelos blanquecinos.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 5
1.1.1. Distribución geográfica y hábitat del género Rosmarinus
Se distribuye por la cuenca del Mediterráneo, de donde es originario, tanto
en el sur de Europa como en el norte de África y suroeste de Asia, presentándose
tanto en forma silvestre como cultivada. Habita en laderas y collados de tierra baja,
encontrándose también en las proximidades del litoral (Morales, 2000).
El romero, junto con el espliego, son las plantas aromáticas más abundantes
en rodales o matas aisladas, frecuentemente asociado con el tomillo vulgar y
ajedrea, en altitudes desde el nivel del mar hasta los 1.500 m en las montañas más
cálidas, aunque el óptimo, en parte por su carácter de planta termófila, es en
altitudes inferiores a los 1.000 m, sobre todo en zonas bajas y medias, y con
precipitaciones entre 280‐300 y 600 mm anuales (Gónzalez López, 1982; Aberturas
Aguado, 1986).
Según la clasificación fito‐sociológica, dentro de los matorrales calcícolas
está el orden Rosmarinetalia, el cual engloba a romerales, tomillares, salviares, etc.
que se desarrollan sobre suelos bien drenados, formados a partir de sustratos
básicos, no salinos, como son las margas, calizas y dolomías principalmente
(Ferreras y Arozena, 1987; Sanchis Duato et al., 2003).
Aunque la planta del romero es muy plástica, los lugares en que se desarrolla
más favorablemente corresponden a zonas secas, áridas y soleadas, es decir, con
clima moderado, templado o templado‐cálido, más bien con poco agua,
principalmente ambiente de encinar (etapas degradadas por tala o quema, o laderas
pedregosas y erosionadas). El óptimo de estos suelos se produce en la región
mediterránea occidental y últimamente se han expandido notablemente debido a la
degradación de los bosques y de los suelos que en dicha región se daban. Con el
paso del tiempo, este tipo de matorrales se han constituido en comunidades
estables y permanentes (Sanchis Duato et al., 2003). Además, precisa de protección
contra el viento y heladas. Desaparece en las umbrías más frescas, llegando a

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 6
alcanzar las cotas más elevadas en las zonas de solana (López González, 1982;
Morales, 2000).
1.1.2. Especies objeto de estudio: R. officinalis y R. eriocalyx
Al romero se le conoce también como romaní, romeo, erromeru, alecrín,
alecrimzeiro. Según la etimología latina, romero procede de ‘ros’ (rocío) y ‘marinus‘
(mar), en alusión a su aroma y su hábitat próximo al litoral, pues ‘rocío de mar’
parecía indicar el hábitat de una especie típica de la cuenca mediterránea (Mendiola,
1989). La etimología griega le hace proceder de los vocablos ‘rhos’ (arbusto) y
‘myrinos‘ (aromático), característica de la planta, algo que es más acorde con la
realidad que la interpretación anterior, es decir, con su porte y aroma característico.
El nombre específico officinalis expresa o alude a sus aplicaciones farmacéuticas y
medicinales.
Rosmarinus officinalis L. es una planta de tipo subarbustiva o mata vivaz,
leñosa, rústica, erguida o ascendente, siempre verde, que puede alcanzar 2 m de
altura, muy ramificado (de 50‐80 cm de altura y de 1,5‐1,8 m de vuelo o anchura),
con ramas marrones y muchas hojas, con tallos cuadrangulares y algo vellosos
cuando son jóvenes, rodeándose y haciéndose quebradiza su corteza al envejecer,
al mismo tiempo que se pierde el vello. Es además una planta muy aromática y
melífera (Aberturas Aguado, 1986; Mendiola, 1989).
Las hojas son pequeñas, gruesas y estrechas (de 15 a 35‐40 mm x 1,2‐3,5 mm
de longitud), perennes, sentadas, casi lineales, enteras, opuestas, coriáceas,
lanceoladas, lampiñas, con los bordes enteros y vueltos hacia abajo, de color verde
(oscuro e intenso) brillante, ligeramente rugosas por el haz y suaves, con tomento
blanquecino, por su envés. Los pedúnculos y pedicelos son tomentosos‐estrellados.
Tanto las hojas como las flores tienen un olor o aroma intenso, algo picante y, en el
caso de las hojas, un sabor acre (Aberturas Aguado, 1986; Mendiola, 1989).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 7
Las flores (fig. 5a) son subsésiles, agrupadas en pequeños y cortos racimos
axilares (de las hojas) y terminales (por estar en las cimas de las ramas). Se
desprenden con gran facilidad. El cáliz es de 3‐4 mm de longitud, acampanado,
bilabiado, verde o pulverulento y tomentoso de joven, posteriormente subglabro,
con la garganta desnuda, donde el labio superior es oval, entero, y el inferior con
dos lóbulos lanceolados. La corola es de 10‐12 mm de longitud, de color azul o lila
pálido, a veces rosa y más rara vez blanca; bilabiada, con el tubo saliente, siendo el
labio superior en forma de casco, bífido, y el inferior con tres lóbulos, el central más
ancho y ligeramente cóncavo. Por último, el androceo está formado por dos
estambres únicamente, de filamentos salientes, insertos en la garganta de la corola
y provistos en su base de un pequeño diente, terminados por dos anteras lineares y
con un solo lóbulo (Aberturas Aguado, 1986; Mendiola, 1989).
El fruto es seco, en tetraquenio, con semillas menudas (fig. 5b). Los aquenios
son obovales, sésiles, de color marrón, con cuatro semillas menudas. En el interior
de cada aquenio hay un embrión desprovisto de albúmen, con dos cotiledones
convexos (Aberturas Aguado, 1986; Mendiola, 1989).
a) b)
Fig. 5. Rosmarinus officinalis L. Detalle de las flores (a) y el fruto en tetraquenio (b).
El romero está en flor casi todo el año, aunque la floración es mayor desde
febrero hasta noviembre. Especialmente es más intensa de marzo‐abril a mayo, si

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 8
bien se produce una renovación de la floración durante un amplio período
(Aberturas Aguado, 1986).
El Rosmarinus eriocalyx se diferencia fácilmente por sus inflorescencias
densamente cubiertas de largos pelos glandulares y sus hojas más cortas (5 a 15
mm), lampiñas y verdes; es de ramas grises y presenta mayor porte.
Rosmarinus officinalis L. es una especie de amplia distribución (figura 6), bajo
diversas formas, debido a su alta capacidad de adaptarse a áreas de condiciones
ecológicas muy variables. Esta especie habita principalmente en el contorno de la
región mediterránea. En lo que se refiere a España, se encuentra en las islas
Baleares y en la Península Ibérica. Sólo falta en algunos puntos del norte y noroeste,
siendo especialmente frecuente en las tierras bajas de clima cálido (López González,
2001).
Fig. 6. Distribución aproximada de R. officinalis L. (trazo de líneas continua) en el Mediterráneo occidental, y localidades en que se han encontrado distintos taxones: R. eriocalyx – R. tomensus – R. x mendizabalii – R. x lavandulaceus: 1. var. lavandulaceus; 2. var. noeanus; 3. var trogloditarum; 4. var. subtomentosus; 5. var clementei (Fuente: Rosua,1986).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 9
La distribución de Rosmarinus eriocalyx Jordan & Tour es mucho más escasa
comparada con R. officinalis L. Se distribuye por distintas regiones áridas y
semiáridas del Mediterráneo (ej. sur de Andalucía y norte de África), pero
principalmente en la zona subcostera de las provincias de Almería, Granada y
Málaga (López González, 2001). En muchas ocasiones se encuentra ligada a
formaciones de Tetraclinis articulata y en otras convive con R. officinalis L., con quien
hibrida con facilidad (Mendiola, 1989).
1.2. Importancia de las labiadas: usos y cultivo
La familia botánica Labiatae, plantas aromáticas por excelencia, ha tenido
siempre gran importancia para el hombre debido a los usos culinario y medicinal, tal
como indica Pío Font Quer en el compendio de El Dioscórides Renovado de plantas
medicinales, donde recopila un total de 62 plantas de esta familia (espliego,
alhucema, lavanda, salvia, toronjil, ajedrea, hisopo, mentas…) de las cerca de 3000
que contiene la publicación (Font Quer, 1981). Uno de los representantes más
conocidos es el romero y es en el que está centrado este trabajo.
Las aplicaciones del romero se basan en sus propiedades, derivadas de su
particular composición química, tanto de las partes enteras (hojas y sumidades
floridas) como de su extracto (aceite esencial) (Font Quer, 1981; Aberturas Aguado,
1986; Mendiola, 1989; Fernández‐Pola, 2001). Además de por su uso medicinal
(industria farmacéutica) y culinario (condimento), el romero es también utilizado en
jardinería (setos ornamentales y aromáticos) y, en el ámbito de la industria
alimentaria, como planta melífera (Font Quer, 1981; Aberturas Aguado, 1986;
Mendiola, 1989; Fernández‐Pola, 2001).
Las hojas y sumidades floridas se utilizan tanto para uso interno como
externo, dependiendo de la forma de aplicación. Las hojas en su uso interno, según
recopila Font Quer (1981) y los autores anteriormente mencionados, preparadas en

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 10
forma de infusión y extracto fluido, son consideradas como estomacales, en la
insuficiencia hepática, espasmolíticas, vulnerarias, tónicas y aperitivas, sedantes,
diaforéticas, emenagogas, antioxidantes y facilita la curación de la ictericia. Además,
las hojas frescas o desecadas a la sombra, cocidas en vino, se utilizan para aumentar
el apetito y favorecer la digestión, como diuréticas, también carminativas e incluso
febrífugas, coleréticas y colagogas (referido sobre todo a la secreción biliar),
pudiendo ser abortivo y tóxico en dosis elevadas (Font Quer, 1981; López González,
2001). En lo que se refiere a su uso externo, tanto hojas como sumidades floridas se
utilizan como cicatrizantes, desinfectantes y como antirreumáticas. En cuanto a la
cocina, por su sabor penetrante, el romero se emplea como condimento de asados
de carnes, pescados, parrilladas y caza, salsas, etc (Mendiola, 1989; Fernández‐Pola,
2001).
El aceite esencial es utilizado fundamentalmente por las industrias de
cosmética y de perfumería; en esta última, tanto para colonias (ya que mezcla muy
bien con otros aceites), como para aromatizar jabones, insecticidas domésticos,
desodorantes, ambientadores, detergentes, etc, no sólo por su olor fresco sino por
su facilidad de enmascarar los olores de alquitrán y fenoles. También es utilizado,
con carácter medicinal, como analgésico (en usos interno y externo),
antiespasmódico, detersivo, cicatrizante, y estimulante. Del romero se obtiene
aproximadamente un 2% de aceite esencial, cuya composición depende de la época
del año y de la zona en que se recolecte el romero (Mendiola, 1989; Fernández‐Pola,
2001).
En el aceite de romero se han descrito hasta 53 componentes diferentes,
siendo los compuestos más frecuentes: α‐pineno, canfor, 1,8‐cineol, canfeno (o
eucaliptol), borneol, β‐pineno, verbenona, β‐cariofileno, limoneno (hidrocarburo
aromático), α‐terpineol, mirceno, p‐cimeno, acetato de bronilo, linalool y terpinen‐4‐
ol (Salido, et al., 2003), componentes fenólicos (mentol, timol, pulegona, esclarol,
carvacrol, piperitona), pigmentos flavónicos, flavonglucósidos (apigenina y
luteolina), ácidos fenólicos (cafeico, clorogénico, neoclorogénico, rosmarínico,

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 11
gálico, gentísico, siríngico) y una lactona diterpénica llamada picrosalvina o carnasol
(amarga, que en el proceso de destilación se transforma en el ácido carnosólico, no
amargo), una resina y ácidos orgánicos, junto con pequeñas cantidades de
rosmaricina (alcaloide), ácido ursólico, taninos, azúcares y elementos minerales (Na,
K, Ca, Mg, Fe, Cu, Zn y Mn) (Mendiola, 1989; Hidalgo et al., 1998; Fernández‐Pola,
2001; Del Bano et al., 2003; Almela et al., 2006; Moreno et al., 2006). Otros
componentes se encuentran en cantidad inferior al 2% (Atti‐Santos et al., 2005;
Carvalho et al., 2005; Ramírez et al., 2006).
El ácido rosmarínico extraído del romero posee una fuerte actividad
antioxidante que recientemente está siendo utilizada por la industria alimentaria. El
extracto de romero (extracto supercrítico) es uno de los componentes de la
patente Vidalim1 ® para el desarrollo de productos cárnicos funcionales, donde es
añadido por su capacidad antioxidante para preservar la relación de ácidos grasos
poliinsaturados omega‐6/omega‐3 introducidos en la composición de una conocida
marca de productos cárnicos española (Frial, S.A.).
En esta línea, la compañía eslovena Vitiva ha desarrollado un concentrado del
ácido rosmárinico en forma soluble en agua, dirigido al mercado de suplementos
dietéticos, cosméticos y productos alimenticios (una pureza del 5 al 70%). Este
ingrediente se considera de interés por su capacidad antioxidante, la cual puede
ejercerse tanto en alimentos evitando la oxidación de algunos de sus componentes
de interés nutricional, pero fácilmente oxidables, como para prevenir el daño celular
causado por radicales libres, que se generan de forma fisiológica en el metabolismo
aerobio de los seres vivos. Otra actividad de interés del ácido rosmarínico es su
capacidad antiinflamatoria, ya que procesos de este tipo están implicados en gran
número de enfermedades (asma, cardiovasculares, reumáticas) (Abramovic y
Abram, 2006).
1 Pertenece a un grupo de investigación de la Universidad Autónoma de Madrid y el grupo Frial (noviembre, 2005). Premios Mejor Empresa Alimentaria Española – Edición 2005. Dossier de prensa. Ministerio de Agricultura, Pesca y Alimentación (www.mapya.es).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 12
1.2.1. Cultivo del romero
La densidad de plantación óptima varía de 15.000 a 20.000 pies/ha. Respecto
a las labores culturales a realizar, son relativamente fáciles: araduras, riegos
moderados, escardas y acepta la poda sin ningún problema. En inviernos fríos se
debe proteger. Para impedir que germinen las semillas de las malas hierbas se
emplean diferentes herbicidas (ej. Linuron), que no deben usarse si la hoja va
destinada a herboristería (Muñoz, 1987; Mendiola, 1989; Fernández‐Pola, 2001). La
multiplicación – propagación puede hacerse por semilla y vegetativamente,
mediante esquejes, división de pies y acodos.
El romero es un cultivo poco exigente en cuanto a fertilización. En un cultivo
bien dirigido se impone un aporte bien fermentado en la labor preparatoria del
terreno de invierno, junto con otra labor a finales de primavera. Cabe reseñar que,
como planta poseedora de aceite esencial, requiere gran cantidad de azufre
(Muñoz, 1987; Mendiola, 1989; Fernández‐Pola, 2001).
La recolección puede realizarse mecánicamente. Mientras las hojas pueden
ser recolectadas en cualquier época del año (óptimo al final de verano) desde el
segundo o tercer año, las flores son recolectadas inmediatamente antes de la plena
floración. El secado se hace a la sombra, de forma extendida y sin exceder los 35ºC.
Las hojuelas se separarán de los tallos después del secado. Se suele destilar la planta
entera, salvo en las regiones muy secas, que sólo se hace con las ramas tiernas y
floridas. Las plantas recolectadas por la mañana temprano son más ricas en aceite
esencial que las cortadas por la tarde (Aberturas Aguado, 1986; Muñoz, 1987;
Mendiola, 1989; Fernández‐Pola, 2001).
Respecto al apartado de las enfermedades, se cree que el amarillamiento de
algunas ramas, seguido del desecamiento de la planta, es debido a la presencia de
nemátodos, aunque en la mayoría de los casos, el responsable es el frío. Otro
organismo que ataca a tallos y hojas, es un coleóptero parásito (Chrisolina americana

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 13
L.), el cual se combate eficazmente con Actellic 50‐E. El tiempo de duración de un
cultivo es de unos 10‐12 años, pero conviene renovarlo cada 5‐7 años. Los
rendimientos en hojas secas son del 20 al 25 % de la producción de planta fresca, de
la que se obtiene aceite esencial entre el 0,5 y 0,6 % (Muñoz, 1987; Mendiola, 1989;
Fernández‐Pola, 2001).
En cuanto a la comercialización, la demanda de hojas se satisface con plantas
de flora espontánea. En España, las principales provincias productoras son Murcia,
Granada y Jaén. La demanda de aceite esencial es baja y estabilizada, pero el aceite
esencial obtenido de cultivo no puede competir con el del silvestre ni con el del
Norte de África. A pesar de la gran importancia comercial del género, todavía hay
mucha necesidad de información en cuanto a su cultivo, recolección y manejo del
germoplasma. Consecuentemente, el grado de erosión no es bien conocido. Varias
especies de Rosmarinus están en la lista de especies raras, amenazadas y endémicas
de Europa. Varias instituciones a lo largo del mundo recogen material genético de
Rosmarinus, especialmente para fines de investigación.
El conocimiento de la amplia diversidad morfológica y química del género
Rosamarinus y la distribución nativa de sus diferentes taxa es esencial para mejorar
la explotación de este cultivo muy prometedor. El romero es un cultivo en el que la
mejora genética resulta interesante debido a su alta heterogeneidad química y
fisiológica. La mejora de cultivos es altamente recomendable teniendo en cuenta su
amplio rango de usos y las grandes dificultades que el hecho de que el material no
sea uniforme puede causar al sector. Teniendo en cuenta tanto a los productores
como a los consumidores, los esfuerzos de cualquier programa de mejora del
romero, deben estar dirigidos a mejorar los siguiente objetivos: parámetros
relacionados con el rendimiento (ej. hábito de crecimiento), desarrollo rápido de las
plantas jóvenes, ramificación, ratio hoja/tallo, tolerancia ante diversos factores
ambientales (salinidad y frío), resistencia a patógenos o enfermedades y
parámetros relacionados con la calidad (ej. mejores características aromáticas,
contendido y composición en aceite esencial, propiedades antioxidantes y

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 14
antimicrobianas). Para lograr estos objetivos las herramientas más adecuadas para
la mejora del cultivo son los métodos de selección e hibridación, combinados con
controles analíticos de la variabilidad encontrada en el material. Para poder ir más
allá, investigando en estas especies, se requiere de la aplicación de la biotecnología
en esta área, con la finalidad de obtener nuevas variedades. Para llevar a cabo esta
tarea y desarrollarla en profundidad, además de los caracteres de tipo fisiológico‐
agronómico que debe poseer cualquier nueva variedad para distinguirla de otras del
mismo cultivo ya existentes, es necesario utilizar otros caracteres morfológicos y
moleculares, que hagan más sencilla esta diferenciación, sin necesidad de recurrir a
su cultivo en campo, que es donde se muestran las características fisiológico‐
agronómicas. En todos estos procesos interesa la búsqueda de marcadores
(morfológicos o moleculares), es decir, caracteres que resulten ser fácilmente
observables y se encuentren estrechamente ligados, desde el punto de vista
genético, a una característica agronómica de interés (Muñoz, 1987; Mendiola, 1989;
Fernández‐Pola, 2001).
1.3. Importancia de la conservación
Desde el inicio de la agricultura, el hombre ha domesticado especies
vegetales para su aprovechamiento y ha seleccionado a lo largo de las generaciones
aquellos caracteres que mejor se adaptaban a sus necesidades. Simultáneamente
con el proceso de domesticación y diversificación, ha existido una constante
preocupación por la conservación de los recursos biológicos disponibles, sin
embargo, la conciencia de la erosión genética como un problema a escala mundial
no ha tenido lugar hasta bien entrado el siglo XX. A comienzos de dicho siglo, la
mayor parte de la población humana utilizaba técnicas de producción agraria que no
habían variado sustancialmente durante los últimos mil años, pero en unas pocas
décadas, la agricultura experimentó una profunda transformación que permitió
incrementar de forma espectacular la productividad alimentaria. La conocida
‘revolución verde’ fue consecuencia de la aplicación masiva de fertilizantes, la lucha

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 15
química contra plagas y enfermedades y la progresiva mecanización de las
actividades agrícolas, sin olvidarse, de que el principal factor fue la introducción de
cultivares vegetales cuidadosamente seleccionados por procedimientos de mejora
genética. De esta forma, se ha producido una doble erosión genética de las plantas
cultivadas: por un lado se ha reducido ostensiblemente su número y se han
conservado solamente aquellas en las que se han realizado programas de mejora, y
por otra parte, se ha reducido drásticamente el número de cultivares por especie
(Iriondo Alegría, 2001; Scarascia‐Mugnozza y Perrino, 2002).
Actualmente, los recursos fitogenéticos se encuentran en grave peligro de
extinción al ser sustituidas las variedades locales por variedades más productivas y
resistentes a diferentes tipos de estrés. Estos recursos contienen la variabilidad de
la información genética que las variedades productivas y resistentes modernas no
tienen, por lo que las variedades locales son necesarias para la creación hoy,
mañana y en los sucesivos años de nuevas variedades en un futuro. El concepto
clásico de recursos fitogenéticos incluye el material de reproducción o de
propagación vegetativa de variedades cultivadas, cultivares en desuso, cultivares
primitivos, especies vegetales silvestres emparentadas con cultivares, estirpes
genéticas especiales (entre ella las líneas y mutantes selectos de los mejoradores) y
especies de malas hierbas. Si se tiene en cuenta que con los conocimientos actuales
se permite hoy utilizar genes procedentes de cualquier ser vivo en los programas de
mejora de especies cultivadas, todas las especies vegetales, cultivadas o silvestres,
deben ser consideradas como recursos fitogenéticos. Por tanto, los recursos
fitogenéticos constituyen una auténtica garantía de la seguridad alimentaria
mundial ya que son fuente insustituible de características tales como la adaptación a
condiciones ambientales, la resistencia a enfermedades y plagas, los tratamientos
agronómicos, la calidad nutritiva y, principalmente, la productividad, aspecto
importante ante el crecimiento continuo de la población mundial. La reducción de la
base genética conlleva un aumento en la vulnerabilidad de los cultivos ante
inesperados cambios ambientales o a la aparición de nuevas plagas y enfermedades
(Nevo, 1998; Iriondo Alegría, 2001; García Olmedo et al., 2001).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 16
La toma de conciencia sobre la magnitud de los problemas mencionados ha
derivado en el desarrollo de iniciativas destinadas a contrarrestar la pérdida de
biodiversidad tanto en el ámbito de las plantas cultivadas como en el de las plantas
silvestres. La actual alarma global de crisis y extinción de la biodiversidad afecta
negativamente a la biosfera. La conservación de la biodiversidad es una de las
herramientas posibles para paliar el agotamiento de la biosfera por parte de los
humanos. La diversidad genética, la base de la evolución por selección natural, está
gravemente afectada en las especies emparentadas de las plantas cultivadas y su
exploración, evaluación, conservación in situ y ex situ es imprescindible para poder
garantizar un desarrollo sostenible (Nevo, 1998). En este contexto, la conservación
se define como la gestión de la utilización humana de la biosfera para que pueda
aportar el máximo beneficio sostenible a las generaciones presentes y futuras
(Tratado internacional sobre los recursos fitogenéticos para la alimentación y la
agricultura, www.fao.org).
La conservación puede realizarse, al menos teóricamente, a tres niveles de
organización: génica, organísmica y ecológica. Tradicionalmente, los métodos de
conservación se clasifican en dos grandes grupos: métodos de conservación in situ y
métodos de conservación ex situ. Los métodos de conservación in situ, permiten la
conservación de las especies en los entornos ecológicos naturales y culturales en los
que han desarrollado sus propiedades específicas. Por el contrario, la conservación
ex situ implica la conservación de las especies fuera de sus respectivos entornos,
llevándose esto a cabo, en el caso de las especies vegetales, bien a través de
colecciones de plantas (jardines botánicos y colecciones de plantas en campo) o de
bancos de germoplasma (Iriondo Alegría, 2001; Scarascia‐Mugnozza y Perrino,
2002).
Los jardines botánicos pueden considerarse las primeras instituciones
implicadas en la conservación ex situ de recursos fitogenéticos. En la actualidad hay
cerca de 1500 jardines botánicos por todo el mundo, de los cuales unos 800
desarrollan actividades de conservación. Las colecciones de plantas en campo,

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 17
conjunto de individuos de una misma especie seleccionados para que sean
representativos de la variabilidad genética existente, son particularmente
apropiadas para especies perennes que tardan años en producir semillas (ej.
especies forestales), especies que se reproducen exclusivamente de manera
vegetativa o para la conservación de plantas cuyas semillas no pueden ser
fácilmente almacenadas (ej. cultivos tropicales).
Los bancos de germoplasma son centros orientados al almacenamiento de
una gran parte de la variabilidad genética correspondiente a una determinada
especie o cultivar. Dentro de esta categoría podemos distinguir los bancos de
semillas, bancos de cultivo in vitro y bancos de genes o bancos de ADN. El
almacenamiento del material a conservar en forma de semillas – bancos de semillas
– constituye uno de los procedimientos de conservación ex situ más válidos y
extendidos en la actualidad, que permite mantener un gran número de semillas de
especies cultivadas durante largos períodos de tiempo y con un mínimo riesgo de
daños genéticos. Las semillas presentan una serie de características que hacen que
el almacenamiento sea el método más eficaz y económico para la conservación ex
situ de especies vegetales. Por un lado, las semillas, son unidades adapatadas a al
dispersión en el tiempo, y por tanto, capaces, en la mayoría de los casos, de
permanecer viables, de forma natural, durante largos períodos de tiempo; en
segundo lugar, su pequeño tamaño, unido a la posibilidad de que cada una de ellas
posea una constitución genética diferente, asegura la conservación de una gran
diversidad genética en un espacio reducido. Sin embargo, esta técnica no es
aplicable a todas las especies vegetales ya que las semillas de un número
considerable de especies no sobreviven a estas condiciones de almacenamiento
(semillas recalcitrantes). Si bien, la mayoría de las especies silvestres y de los
cultivos de las zonas templadas del planeta forman semillas ortodoxas, por lo que
pueden ser conservadas en bancos de semillas, con condiciones de temperatura y
humedad bajas (Iriondo Alegría, 2001).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 18
Una alternativa a los bancos de semillas, es la crioconservación, que permite
almacenar el material vivo a muy bajas temperaturas (en general a menos de ‐130
ºC). Las principales ventajas de la crioconservación sobre otras técnicas radican en la
ausencia de controles complicados de humedad y temperatura, la inexistencia de
daños de plagas y patógenos en las colecciones y, en teoría, en una viabilidad del
material vegetal sin límites temporales y sin daños genéticos, lo que permite, en
principio, reducir o eliminar la realización de controles periódicos de viabilidad.
Otra posibilidad para la conservación, es la aplicación de las técnicas de
cultivo in vitro, utilizada en los casos de especies con semillas recalcitrantes,
especies que no producen semillas o con producción reducida de semillas, especies
con ciclos de vida muy largos que no producen semillas hasta que la planta alcanza
su madurez al cabo de muchos años y clones con elevado grado de heterocigosis
que han sido seleccionados y mantenidos por el agricultor mediante propagación
vegetativa. Las ventajas de las técnicas de cultivo in vitro son principalmente la
obtención de elevadas tasas de multiplicación, el carácter aséptico del cultivo, el
reducido espacio ocupado por las colecciones, la reducción del riesgo de pérdidas
de material debido a catástrofes naturales y sus múltiples aplicaciones en
programas de mejora genética. Entre los inconvenientes que conlleva la utilización
de esta técnica se puede mencionar el elevado coste de infraestructura, la
necesidad de personal cualificado y los riesgos de alteraciones genéticas durante el
cultivo (variación somaclonal).
Con el avance de las técnicas de ingeniería genética que, en principio,
permiten la transferencia de genes entre especies totalmente distintas,
pertenecientes incluso a distintos reinos, se ofrece una nueva alternativa, la
instalación de bancos de ADN. En los bancos de ADN se procede a la extracción del
ADN procedente de individuos de una determinada especie o cultivar y
posteriormente, el ADN extraído se conserva a bajas temperaturas (cámaras
congeladoras a ‐80ºC o tanques de nitrógeno líquido). En la actualidad, esta
alternativa se utiliza en el caso de especies o cultivares cuyo genoma ha sido

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 19
estudiado en detalle y donde se conocen las secuencias de numerosos o
importantes genes (Iriondo Alegría, 2001).
La conservación in situ de especies cultivadas supone el mantenimiento de
variedades tradicionales por parte de los agricultores, bien porque existe una
subvención económica a las prácticas tradicionales, o porque determinadas
variedades tradicionales o de especies en desuso resultan competitivas en ámbitos
locales gracias al desarrollo y promoción de sus productos ligados a valores
culturales de la zona. En el caso de la conservación de especies silvestres, la solución
idónea consiste en la protección de los ecosistemas en donde habitan las especies
consideradas (conservación in situ), si bien, en muchas ocasiones, la degradación o
destrucción del hábitat natural hace que la conservación ex situ en bancos de
germoplasma constituya la única alternativa posible.
La conservación in situ implica una adecuada protección y gestión de los
ecosistemas naturales. Existen numerosas formas para proteger los espacios
naturales de la actividad humana, sin embargo, estas medidas no resultan siempre
suficientes para asegurar la supervivencia de las especies a conservar. En todo
momento, la conservación ex situ de especies silvestres constituye una herramienta
de apoyo fundamental para las operaciones de conservación in situ, al tiempo que
cumple una función de seguro ante la destrucción de las poblaciones naturales. Los
programas de conservación ex situ suplementan la conservación in situ
proporcionando almacenamiento a largo plazo, análisis, ensayos y métodos de
propagación de especies amenazadas y sus propágulos.
Para llevar a cabo una gestión adecuada de la conservación in situ resulta
necesario recavar previamente una gran cantidad de información básica sobre la
especie a proteger y su ecosistema. Una parte fundamental del proceso de
conservación son los procedimientos de documentación, caracterización y
evaluación del material conservado. La documentación hace referencia al conjunto
de datos que identifican cada muestra y describen la localización y características

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 20
del lugar de recolección así como la ubicación presente del material conservado. Los
descriptores juegan un papel importante en la caracterización y evaluación del
material conservado y se definen como características o propiedades de una
muestra que son identificables y medibles. En el proceso de caracterización se
utilizan descriptores que permiten una fácil y rápida discriminación entre fenotipos.
Se suelen utilizar caracteres que poseen un fuerte componente genético y que son
independientes de las condiciones ambientales. La utilización de marcadores
moleculares (ej. isoenzimas, RFLPs, RAPDs, AFLPs, SSRs) es muy recomendada en
estos casos (ver más adelante). En el proceso de evaluación los descriptores están
relacionados con caracteres que son útiles en la mejora del cultivo y que, a menudo,
dependen de las condiciones ambientales, tales como productividad y
susceptibilidad a diferentes tipos de estrés (Iriondo Alegría, 2001).
La conservación activa de los recursos fitogenéticos ha aportado ya
importantes beneficios a la agricultura. La conservación y evaluación de la
diversidad biológica vegetal no sólo aporta beneficios en el área de la producción
alimentaria, sino que puede contribuir sustancialmente a otras áreas de la actividad
humana, como la medicina o la industria. En la actualidad, el 25% de los
medicamentos de la industria farmacéutica tienen como principio activo
compuestos procedentes de plantas, mientras que en los países en desarrollo, el
80% de los fármacos son de origen vegetal. Muchos de los compuestos que se
emplean en investigación clínica proceden de material vegetal por lo que los
esfuerzos que se realicen en la conservación de recursos fitogenéticos serán
altamente rentables para la sociedad.
1.4. Caracterización del material conservado
La caracterización de las especies permite abordar el análisis de la diversidad
genética de una determinada especie, teniendo como objetivo el conocimiento de
la diversidad tanto a nivel interpoblacional como intrapoblacional. Ello resulta

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 21
fundamental a la hora de diseñar estrategias de conservación vegetal in situ y ex
situ. Para aumentar las probabilidades de éxito en el mantenimiento de especies a
través de programas de conservación, en la conservación in situ deberían
protegerse, de entre todas las áreas en las que habite la especie, aquellas que
presenten mayor diversidad genética, y en lo referente a la conservación ex situ, la
recogida de germoplasma vegetal deberá realizarse en las mismas zonas.
Sin embargo, este aspecto no solamente tiene interés desde el punto de
vista conservacionista, ya que la domesticación de especies silvestres con la
finalidad de poder ser utilizadas y/o aprovechadas por el ser humano, requiere
también un exhaustivo estudio de la biodiversidad existente en las poblaciones
naturales, con el objetivo de identificar aquellas que mejor se adaptan al cultivo.
Esta diversidad genética puede analizarse utilizando caracteres morfológicos,
generalmente cuantitativos, o bien utilizando marcadores moleculares (González‐
Andrés, 2001).
Otra finalidad de la caracterización es poder llevar a cabo una correcta
gestión de los bancos de germoplasma, para lo que es necesario evitar duplicados, e
identificar las sinonimias (el mismo material con diferentes nombres) y homonimias
(materiales diferentes con el mismo nombre). En este caso se trata de identificar
como igual o diferente, respectivamente, accesiones que pertenecen a la misma
especie, y con frecuencia a la misma variedad, como cuando se trabaja con clones
de cierta variedad. Por tanto es necesario buscar e identificar las características que
diferencien a materiales por otra parte tan similares. Los métodos de
caracterización que permiten evaluar estas diferencias con mayor fiabilidad están
basados en la utilización de marcadores moleculares (tanto isoenzimas como
marcadores de ácidos nucleicos), si bien en los últimos años las técnicas más
empleadas son las basadas en el estudio del ADN.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 22
1.4.1. La caracterización vegetal: diferentes enfoques
Los estudios de caracterización pueden estar basados en distintos tipos de
caracteres. Centrándose en las condiciones en las que se desarrollan los individuos,
los caracteres ecológicos analizan el hábitat, los parásitos, los alimentos, las
variaciones estacionales, etc., mientras que los geográficos hacen referencia entre
otros a la distribución de las especies, a las relaciones entre las poblaciones, tanto
de simpatría como de alopatría, etc (González‐Andrés, 2001).
Por otra parte, en el estudio directo de los organismos se pueden distinguir
los caracteres de tipo: morfológicos, citológicos, bioquímicos, embriológicos y
basados en el ADN. Los caracteres morfológicos son los más clásicos y estudian
cualquier órgano de la planta desde el punto de vista cualitativo (formas, colores,
presencia de estructuras singulares, etc) o desde el punto de vista cuantitativo.
Los caracteres citológicos se centran fundamentalmente en el estudio del
núcleo celular, en especial el número de cromosomas. Son los bioquímicos quienes
abarcan o incluyen el estudio de metabolitos del metabolismo principal y secundario
de las plantas, fundamentalmente los basados en compuestos de bajo peso
molecular (flavonoides, alcaloides, aminoácidos no proteicos, aceites esenciales) y
los basados en el estudio de proteínas (proteínas totales, isoenzimas).
Los embriológicos están basados en el desarrollo embrionario de los
individuos, siendo los basados en el estudio del ADN los que incluyen a todos los
marcadores moleculares basados en la secuencia primaria del ADN.
En el siglo XX se ha asistido a un espectacular crecimiento de los estudios
bioquímicos en plantas y a su aplicación en taxonomía, identificación y filogenia. En
la actualidad la caracterización con marcadores moleculares ha cobrado gran
predominio y protagonismo, aunque no por ello las técnicas basadas en la
morfología quedan obsoletas ni han sido desechadas. Existen posturas extremas a

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 23
favor de un sistema u otro, pero en la actualidad hay gran cantidad de pruebas que
indican que ambos tipos de estudios son complementarios, y la integración de los
resultados obtenidos por ambos medios enriquece de manera notable las
conclusiones que puedan extraerse y contribuyen a una aproximación más exacta a
la realidad (González‐Andrés, 2001).
La clasificación de organismos y los estudios filogenéticos pueden ser
abordados desde dos perspectivas: la escuela cladística y el feneticismo. La escuela
cladística defiende que las relaciones existentes entre los organismos no pueden ser
deducidas a partir de cualquier similitud que presenten, sino que esas relaciones son
un reflejo de una serie de caracteres que sólo serán compartidos por dos o más taxa
que desciendan de un ancestro común.
El feneticismo entiende que la caracterización deber abordarse sobre
diferentes partes de la planta, en diferentes estados fisiológicos de las mismas, y
con diversos tipos de caracteres, debiéndose analizar todos ellos en conjunto,
otorgándoles a priori la misma importancia.
Con el desarrollo de las técnicas estadísticas de análisis multivariante, y de
los programas informáticos para llevarlos a cabo con rapidez y precisión, el
feneticismo ha encontrado el soporte técnico necesario para establecerse
sólidamente en el campo de la sistemática. Dicho soporte técnico permite analizar
cientos de caracteres simultáneamente y de una manera rápida.
1.4.2. La caracterización mediante marcadores moleculares
Se consideran marcadores moleculares todos aquellos basados en moléculas
biológicas, es decir todos los bioquímicos que engloban compuestos de bajo peso
molecular (flavonoides, alcaloides, terpenoides, etc.) y proteínas (proteínas totales,
isoenzimas), más los basados en la molécula de ADN.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 24
Frente a la metodología clásica de la caracterización basada en morfología,
fisiología y embriología, los marcadores moleculares presentan, entre otras las
siguientes claras ventajas: detectan mayor variabilidad, no son susceptibles de estar
afectados por el ambiente (sólo los basados en la molécula de ADN) y, valorados de
forma conjunta con los caracteres morfológicos (como resultado final de una serie
de rutas metabólicas complejas e interconectadas, controladas por un elevado
número de genes e influidas por factores externos), se pueden obtener mejores
resultados que con métodos independientes cuando se valora la herencia poligénica
(Torres Lamas, 2001).
El marcador molecular ideal, debe de tener las siguientes cualidades: 1) ser
polimórfico – presentar diferente expresión en las diferentes unidades taxonómicas
operativas a analizar. Así, por ejemplo, los isoenzimas son menos polimórficos que
los marcadores basados en ADN; 2) ser específico de un determinado locus – lo que
quiere decir que cada banda corresponda a un único locus, ya que con frecuencia en
determinados marcadores moleculares basados en ADN se da el problema de la “no
homología”, es decir, que bandas correspondientes a fragmentos de igual movilidad
electroforética, corresponden a diferentes loci. Este problema es tanto mayor
cuanto más alejados filogenéticamente se encuentren las unidades taxonómicas
operativas; 3) heredable y preferentemente con herencia codominante – es decir,
que cuando en un individuo se presentan dos alternativas alélicas de un locus,
ambas se manifiesten. Esto permite diferenciar homocigotos de heterocigotos, y
permite calcular las frecuencias de cada uno de los alelos en cada uno de los loci; 4)
estar ampliamente distribuido en el genoma – cuando lo que se pretende conocer
es la diversidad genética existente en un taxón, o bien establecer las relaciones
genéticas entre genotipos, interesa explotar la mayor parte del genoma, y en
consecuencia interesa que los marcadores estén distribuidos por todo el genoma; 5)
reproducible entre laboratorios y dentro de un mismo laboratorio – ser reproducible
y repetible; y 6) fácil, rápido y económico de detectar.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 25
1.5. Técnicas moleculares usadas en caracterización de poblaciones
Las técnicas y los marcadores moleculares aplicados al estudio de la
variación genética de las plantas se empezaron a utilizar a mediados del siglo
XX. Estos estudios son imprescindibles cuando se trata de conservar
eficazmente y utilizar los recursos fitogenéticos, ya que para ello hay que
ampliar los conocimientos acerca de la extensión y distribución de las
variaciones genéticas. Los marcadores más adecuados deben constituir
caracteres hereditarios, discriminar entre los individuos, poblaciones o taxa
examinados, ser fácilmente evaluables y aportar resultados que puedan ser
comparados con los de otros estudios (Westman y Kresovich, 1997).
1.5.1. Tipos de marcadores moleculares
Los marcadores moleculares pueden ser muy invariables o ser altamente
variables y pueden ser tanto proteínas como ácidos nucleicos. Existe una alta
variedad de fragmentos de ácidos nucleicos que son utilizados como
marcadores, entre los que se encuentran genomas completos, cromosomas
individuales, fragmentos de ADN o ARN y nucleótidos simples. Mientras
algunos se repiten una vez en el genoma, otros se repiten varias veces. Muchas
de las secuencias repetidas usadas como marcadores son no codificantes; otras
son elementos de familias multigenómicas. Algunas secuencias repetidas están
intercaladas por todo el genoma, bien distribuidas al azar o en grupo (Westman
y Kresovich, 1997).
Fue a partir del año 1970 cuando los marcadores moleculares basados en
el ácido nucleico ganaron popularidad, con la ventaja de la secuenciación del
ADN y el análisis de fragmentos de restricción. De esta forma, desde el siglo
pasado se han desarrollado numerosas técnicas que permiten detectar
variaciones en la secuencia de ADN al comparar distintos individuos. Gran parte

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 26
de la investigación actual se centra en la identificación de las secuencias
repetidas que suceden en tándem y existe una alta variedad de fragmentos de
ácidos nucleicos que son utilizados como marcadores. Las clases de
repeticiones en tándem se pueden diferenciar por la longitud de la unidad
central de repetición, el número de unidades repetidas por locus y la
abundancia y distribución de los loci (Westman y Kresovich, 1997).
1.5.2. Tipos de ensayos de marcadores moleculares
Generalmente, los ensayos con marcadores moleculares se clasifican en
función de si las moléculas estudiadas son proteínas o ácidos nucleicos y, en
este último, también por el tipo de material genético analizado en un ensayo de
marcador con ácido nucleicos: el genoma entero, un cromosoma, un fragmento
o un nucleótido. Alternativamente, los ensayos con marcadores pueden ser
clasificados por el tipo de carácter medido. Algunos métodos miden diferencias
cuantitativas, otros miden caracteres cualitativos, cada uno con dos o más
estados posibles. Los ensayos con marcadores también difieren en el número
de loci evaluados por análisis, bien si los múltiples loci son evaluados
simultáneamente o secuencialmente, y según el tipo y cantidad de información
que se necesita acerca del locus marcador antes de llevar a cabo el ensayo
(Westman y Kresovich, 1997).
Los tipos de ensayos con marcadores moleculares se pueden clasificar
en: ensayos con marcadores de proteínas, ensayos con marcadores de ácidos
nucleicos, que a su vez pueden utilizar marcadores de ADN genómico total y
cromosomas o bien marcadores de fragmentación. Estos últimos, los
marcadores de fragmentación, pueden ser: fragmentos que se producen, se
someten a electroforesis y se detectan, fragmentos producidos por enzimas de
restricción digestiva, fragmentos producidos por la reacción en cadena de la

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 27
polimerasa, fragmentos detectados directamente por sondas de hibridación o
por secuenciación de nucleótidos.
Cuando las proteínas se utilizan como marcadores genéticos, se supone
que cualquier variación entre proteínas refleja la variación hereditaria en la
secuencia de sus aminoácidos. Existe un tipo de ensayo de marcadores que
miden la distancia migrada por una proteína a través de un gel de almidón,
poliacrilamida o acetato de celulosa en respuesta a la aplicación de una
corriente eléctrica. Muchas de las proteínas usadas como marcadores son
isoenzimas, que son formas funcionalmente similares de una enzima,
codificada por loci diferentes o alelos diferentes de un locus, es decir, son
múltiples formas moleculares que catalizan una misma reacción. De esta forma
es posible separar enzimas codificadas por genes diferentes o productos de
diferentes alelos de un mismo gen (González‐Andrés, 2001). Con este tipo de
marcadores se llevan a cabo ensayos de medida de propiedades inmunológicas
de las proteínas, pero ha quedado en desuso porque pocas proteínas han
podido ser identificadas correctamente, pero no por ello han dejado de ser una
herramienta útil. Las isoenzimas son frecuentemente usadas para describir la
variación entre y dentro de poblaciones, especies y géneros, para mapas
genéticos y diagnóstico, y en algunos casos para identificación (Westman y
Kresovich, 1997). Uno de los usos más precisos de las isoenzimas es su
aplicación en estudios de análisis de variación intraespecífica, pero su uso está
limitado por el pequeño y limitado número de polimorfismos detectados
(Klaas, 1998).
Los ensayos con marcadores de ácidos nucleicos utilizan como
marcadores el ADN genómico total y los cromosomas. Aunque los marcadores
que utilizan proteínas son útiles como marcadores genéticos, el fenotipo que
cada proteína describe está determinado por el genotipo y por el tipo de tejido
muestreado, el estado fenológico de la planta, medio ambiente y proceso
postranscripcional. Por otra parte, el porcentaje del genoma muestreado por

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 28
los marcadores de proteínas es limitado, ya que estos marcadores sólo
describen las secuencias codificantes y no todas las proteínas pueden ser
separadas y detectadas por los métodos establecidos. En cambio, los ácidos
nucleicos están presentes en todos los organismos y son considerados material
genéticos comunes en la comparación virtual de cualquier taxa. La metilación
del ADN y la estructura secundaria pueden causar variaciones artificiales en
algunos ensayos, alterando la relación directa entre marcador y genotipo. Sin
embargo, estas circunstancias pueden ser detectadas a menudo y eliminadas.
Los marcadores de ácidos nucleicos describen genotipos y no fenotipos y
pueden muestrear tanto regiones codificantes como no‐codificantes de un
genoma. Los primeros ensayos con marcadores de ácidos nucleicos midieron
propiedades del ADN genómico total. Este tipo de marcadores han sido usados
para caracterizar y comparar secuencias repetidas entre especies de plantas e
híbridos (Westman y Kresovich, 1997).
Los ensayos con marcadores de ácidos nucleicos que utilizan fragmentos
de ADN o ARN son actualmente los marcadores genéticos más empleados. Este
tipo de marcadores requieren cantidades mínimas de material de partida y
aportan una información cualitativa de alta resolución acerca de la variación de
la secuencia (Westman y Kresovich, 1997).
Los fragmentos producidos, son separados por electroforesis en gel de
agarosa o poliacrilamida, y en ese momento son detectados. La longitud y
conformación de un fragmento determina su tasa de migración en el gel. Varios
enfoques diferentes se pueden tomar para detectar fragmentos. En algunos
ensayos todos los fragmentos son detectados y visualizados. En otros, los
fragmentos diana específicos son detectados y visualizados mediante
hibridación (Westman y Kresovich, 1997).
Los fragmentos producidos por digestión con enzimas de restricción son
muy utilizados en la genética de plantas. Estos fragmentos de restricción se

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 29
generan por medio del tratamiento de la doble cadena de ADN con
endonucleasas de restricción (que son enzimas generadas por bacterias).
Cuando se evalúan los fragmentos producidos, las variaciones en la
longitud de dichos fragmentos se denominan polimorfismos de longitud de
fragmentos de restricción (RFLPs).
El análisis de los fragmentos de ADN se comenzó a utilizar en la década
de los 70 del pasado siglo XX, con la introducción de los RFLPs, y creció
exponencialmente una década después con el desarrollo de la ‘reacción en
cadena de la polimerasa’ (PCR, de sus siglas en inglés). La PCR utiliza los
principios de reasociación del ADN y la acción de la ADN‐polimerasa para
amplificar los fragmentos de ácidos nucleicos in vitro. Cada una de las
soluciones de la reacción de la PCR contiene un ADN molde de doble cadena,
“primers” (cebador, oligonucleótidos cortos de cadena sencilla), ADN‐
polimerasa termoestable, cofactores enzimáticos y los cuatro
desoxinucleótidos (dNTPs: dATP, dCTP, dGTP, dTTP). El ADN molde se
desnaturaliza por el calor; la temperatura disminuye y los primers se unen a las
secuencias complementarias en el ADN molde. La solución se calienta de nuevo
y la polimerasa añade dNTPs en el extremo 3’ de cada uno de los primers
anillados. Se producen los fragmentos de doble cadena y sirven de molde para
el ciclo siguiente, el cual genera fragmentos con las secuencias del primer en
cada extremo. Durante los ciclos siguientes, estos fragmentos se amplifican en
progresión geométrica.
La especificidad y reproducibilidad de la PCR depende de varios
factores: la concentración y calidad de los ingredientes de la reacción, el diseño
y contenido de GC de los primers y la temperatura, duración y número de ciclos
llevados a cabo. Cuando se utilizan para describir variaciones entre y dentro de
las especies, los fragmentos amplificados se pueden detectar por el marcaje de

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 30
primers o de los dNTPs en la solución de reacción, por los productos marcados
de PCR, o por tinción de los geles después de la electroforesis.
Se han desarrollado y utilizado en la genética de plantas numerosos
ensayos de marcadores basados en PCR. Estos pueden ser clasificados según si
las secuencias dianas son conocidas previamente a la amplificación, o si las
secuencias de los primers están diseñadas o son arbitrarias, el número de
primers, y el tamaño del rango de productos amplificados.
Desde comienzos de los 90, se han desarrollado una serie de técnicas
basadas en la reacción en cadena de la polimerasa (PCR) que no requieren un
conocimiento previo de secuencias específicas, tanto de la región diana como de las
secuencias flanqueantes, o de genes en los organismos examinados (Klaas, 1998).
Entre ellas la RAPD (Random Amplified Polymorphic DNA = polimorfismo de ADN
amplificados al azar) es la más utilizada debido a su sencillez (Klaas, 1998; Torres
Lamas, 2001; Wang et al., 2004). Es en esta técnica en la que más nos detenemos
por ser la que se ha manejado para el presente trabajo.
En el proceso se utiliza un único primer de secuencia arbitraria y más corto
(normalmente de 10 nucleótidos) que los generalmente empleados para amplificar
fragmentos específicos, con las únicas limitaciones de poseer un contenido en G+C
comprendido entre el 50‐70% y carecer de secuencias palindrómicas de más de 6
bases. Distintos laboratorios ofrecen cebadores especialmente diseñados para el
ensayo RAPD, a priori utilizables en un amplio rango de especies que incluye tanto
organismos eucariotas como procariotas, y tanto para su uso con ADN nuclear
como ADN plastídico. Un programa típico de PCR / RAPD consta de unos 35‐45
ciclos, cada uno con las etapas de desnaturalización del ADN molde, unión y
elongación del primer. La temperatura de unión del cebador debe estar entre 35‐37
ºC. Al tratarse de un cebador corto, por encima de 38‐40 ºC no suele permanecer
unido al ADN molde, y por debajo de 35 ºC el porcentaje de uniones inespecíficas es
elevado (Torres Lamas, 2001).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 31
En cada reacción (fig. 7) el primer se anilla al patrón de las secuencias
complementarias en ambos sentidos (“sense” y “antisense”), haciendo posible
la amplificación de las regiones entre los primers en sentidos opuestos. El
número y longitud de fragmentos amplificados puede variar, de forma que, un
primer corto produce generalmente un amplio número de fragmentos
pequeños. En los análisis RAPDs, las muestras que son homocigotas para la
presencia de un fragmento no se pueden diferenciar de las heterocigotas, por
lo que la presencia de los fragmentos generalmente domina sobre la ausencia.
La reacción permite obtener millones de copias de un fragmento del ADN
molde mediante la repetición de un ciclo compuesto por una etapa de
desnaturalización, otra de unión de cebadores a las secuencias flanqueantes de
dicho fragmento y una final de elongación de los cebadores mediante una ADN
polimerasa termoestable.
Fig. 7. Pasos de la técnica PCR (Reacción en cadena de la polimerasa). Fases: 1) Desnaturalización; 2) Unión al cebador; 3) Elongación del cebador. * Los extremos 5` de las moléculas de ADN se representan mediante un círculo (Fuente: Torres Lamas, 2001).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 32
Esta técnica se ha utilizado ampliamente para realizar análisis entre
especies de plantas y también estudios dentro de una misma especie,
identificar entradas en los bancos de genes y cultivares y, en la construcción de
mapas genéticos (Westman y Kresovich, 1997).
La técnica de los RAPDs consta de las etapas siguientes: 1) extracción del
ADN; 2) amplificación de los fragmentos de ADN mediante PCR; 3) separación de los
fragmentos de ADN por electroforesis en geles de agarosa; 4) tinción de los
fragmentos de ADN con bromuro de etidio y 5) visualización con luz ultravioleta y
captación de imagen. El polimorfismo detectado es de tipo presencia/ausencia y se
debe a mutaciones en la secuencia del ADN molde a la que se une el cebador, o bien
a inserciones o deleciones entre las dos zonas de unión de los cebadores (Torres
Lamas, 2001).
Esta técnica ofrece varias ventajas: 1) no se precisa un conocimiento previo
del genoma; 2) se necesitan pequeñas cantidades de ADN (5‐50 ng de ADN por
reacción), lo cual es especialmente importante cuando se trabaja con especies
amenazadas; 3) los métodos de extracción de ADN son sencillos, ya que es
suficiente un ADN molde de calidad media para obtener buenas amplificaciones; 4)
en comparación con otras técnicas, el coste en infraestructura es medio
(básicamente se necesita un termociclador, un equipo de electroforesis y una
fuente de luz ultravioleta); 5) los ensayos son fáciles y rápidos; 6) los marcadores
son abundantes en el genoma y se encuentran distribuidos al azar; y 7) el número de
polimorfismos que se pueden encontrar es teóricamente ilimitado (Torres Lamas,
2001).
En contraposición, se producen algunos inconvenientes tales como: 1) la baja
reproducibilidad de los fragmentos amplificados, especialmente de los de alto peso
molecular (>1.600 pb); 2) la incertidumbre acerca de la homología de los fragmentos
amplificados de igual peso molecular (esto es, si tienen la misma secuencia); y 3)
son marcadores dominantes (Torres Lamas, 2001).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 33
Muchos otros ensayos con PCR son variaciones y combinaciones de los
primers de PCR ya diseñados y de los PCR arbitrarios. La mayoría se
desarrollaron para aumentar la reproducibilidad de los resultados y / o para
mejorar la discriminación entre genotipos altamente relacionados. Para realizar
el genotipo, diagnóstico y valorar la variación entre familias próximas, se
necesitan ensayos con marcadores basados en PCR con alta resolución.
1.6. Utilización de marcadores moleculares en estudios de poblaciones
Los marcadores moleculares pueden ser usados, si la técnica lo permite, para
analizar con una mayor o menor precisión las variaciones entre y dentro de
poblaciones, especies y géneros y, así poder conocer la estructura genética de un
determinado grupo de individuos. De esta forma es posible realizar estudios de
diagnosis, mapas genéticos y, en algunos casos, identificación.
Los estudios descriptivos que usan ensayos con marcadores moleculares
para examinar la variación entre y dentro de taxa específicos pueden generar una
información valiosa, pero algunas cuestiones sobre conservación de plantas como
son los recursos fitogenéticos, se concentran mejor mediante la realización de
varios estudios experimentales y complementarios (Westman y Kresovich, 1997).
El uso de técnicas moleculares permite, en labores de conservación de
poblaciones, comprobar si en varias entradas de un banco de germoplasma existen
duplicaciones, si las poblaciones son genéticamente distintas o si son subdivisiones
geográficas de una población, y si han ocurrido cambios genéticos en una entrada o
población al cabo del tiempo (Wetstman y Kresovich, 1997). En esta misma línea,
ayudan a evaluar las relaciones de similitud entre genotipos, la cantidad de variación
genética presente, y cómo la variación está distribuida entre los individuos,
poblaciones y taxa. La conclusión de estas valoraciones ayuda a decidir qué
genotipos son prioritarios para la conservación in situ, diseño de las estrategias de

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs II.. IInnttrroodduucccciióónn
P á g i n a | 34
muestreo de germoplasma y programas de regeneración, construcción de
colecciones base, estudios de flujo genético, y determinación del tamaño óptimo
para la conservación de poblaciones o entradas de bancos genéticos (Westman y
Kresovich, 1997; Bachmann, 2001).
1.7. Estudios de poblaciones de romero
Actualmente, Rosmarinus es un género poco estudiado desde el punto de
vista de la distribución y diversidad genética de las poblaciones existentes (Elamrani
et al., 2000; Herrera, 2004), sin embargo, sí ha sido estudiado en mayor medida para
la caracterización de los compuestos presentes en sus aceites esenciales (Almela et
al, 2006; Zaouali et al., 2005) debido a su alto interés por parte de la industria, en su
uso en diversos productos.
Hasta la fecha, sólo existe un trabajo publicado en el que se emplean
marcadores moleculares para abordar el estudio de poblaciones naturales de
romero (Martín y Hernández Bermejo, 2000). En dicho trabajo se emplean RAPDs
para el estudio de varias poblaciones de Rosmarinus tomentosus Huber‐Morath &
Maire (especie endémica en el sureste español). En este trabajo se llevó a cabo una
valoración inicial de la variación genética existente entre zonas muestreadas, entre
poblaciones y entre individuos. Los resultados obtenidos constituyen una
información de partida valiosa para la elaboración de estrategias de conservación
de la especie de romero citada que se encuentra en peligro de extinción.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIII.. IInntteerrééss yy oobbjjeettiivvooss
P á g i n a | 35
2. Interés y objetivos
Como ya se ha expuesto anteriormente en la Introducción, la caracterización
del material conservado resulta de gran valor en los diferentes ámbitos
relacionados con la conservación.
El objetivo de este trabajo fue poner a punto un protocolo para la obtención
de marcadores moleculares (RAPDs en este caso) que permitieran caracterizar la
diversidad existente entre las poblaciones silvestres de romero de España
recolectadas y conservadas en el Banco de Semillas de la Universidad Politécnica de
Madrid (UPM) en el marco del proyecto de investigación ‘Recolección, conservación
y caracterización de germoplasma de poblaciones españolas de varias especies de
los géneros Rosmarinus y Origanum’ financiado por el Instituto Nacional de
Investigaciones Agrarias (INIA).
Como objetivos parciales destacan:
Establecer un protocolo adecuado para la obtención de marcadores
moleculares tipo RAPD que permitan caracterizar las muestras analizadas.
Estudiar la diversidad interpoblacional detectada por los marcadores
empleados.
Analizar factores geográficos y ambientales que pudieran explicar el grado
de similitud existente entre las poblaciones estudiadas en base a los
marcadores moleculares analizados.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 36
3. Material y métodos
3.1. Material vegetal
3.1.1. Poblaciones estudiadas
Las muestras estudiadas proceden de poblaciones de romero silvestres
repartidas por toda la geografía española que han sido identificadas en el momento
de la recolección como (Rosmarinus officinalis L. y Rosmarinus eriocalyx Jordan &
Fourr). En la tabla 1 se muestra el número de la población, la especie y la localización
de las muestras tomadas.
Tabla 1. Listado de poblaciones de romero recolectadas.
Nº(Código) 1 Especie Término Provincia R01‐00 R. officinalis Turre Almería R04‐00 R. officinalis Valdeconcha Guadalajara R05‐00 R. officinalis Argamasilla de Alba Ciudad Real R06‐00 R. officinalis Alcaraz Albacete R07‐00 R. officinalis Villena Alicante R08‐00 R. officinalis Algimia de Almoacid Castellón R09‐00 R. officinalis San Martín de la Virgen del Moncayo Zaragoza R10‐00 R. officinalis Loporzano Huesca R11‐00 R. officinalis Leciñena Zaragoza R12‐00 R. officinalis Soto del Real Madrid R15‐01 R. officinalis Torrejón el Rubio Cáceres R19‐01 R. eriocalyx Tabernas Almería R21‐01 R. officinalis Conil de la Frontera / Chiclana Cádiz 1: Los números después del guión indican el año de recolección (2000 y 2001), que no se utilizarán en el resto del documento.
En la figura 8 se muestra la localización, en el mapa de la Península Ibérica, de
las poblaciones recolectadas.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 37
Fig. 8. Situación de las poblaciones de Rosmarinus officinalis [ ] y Rosmarinus eriocalyx [ ].
En la figura 9 se muestran las provincias biogeográficas en que está dividida
España. Seguido figura la tabla 2 en la que se indica a qué población biogeográfica
pertenece cada una de las muestras tomadas.
Fig. 9. Provincias biogeográficas (Rivas‐Martinez et al, 1987). Provincias biogeográficas de España y Portugal (Península Ibérica, Baleares y Canarias).‐ Región Eurosiberiana. I: Pirenaica. II: Cántabro‐atlántica. III: Orocantábrica. – Región Mediterránea. IV: Aragonesa. V: Catalana‐Valencia‐Provenzal. VI: Balear. VII: Castellano‐Maestrazgo‐Manchega. VIII: Murciano‐Almeriense. IX: Carpetano‐Ibérico‐Leonesa. X: Luso‐Extremadurense. XI: Gaditano‐Onubo‐Algarviense. XII: Bética. – Región Macaronésica. XIII: Canaria Occidental. XIV: Canaria Oriental.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 38
Tabla 2. Provincias biogeográficas a las que pertenecen las poblaciones de romero estudiadas. Nº Provincia biogeográfica Población I Pirenaica R10 IV Aragonesa R11 V Catalana – Valenciano – Provenzal R07, R08 VII Castellano – Mestrazgo – Manchega R04, R05 VIII Murciano – Almeriense R01, R19 IX Carpetano – Ibérico – Leonesa R09, R12 X Luso – Extremadurense R15 XI Gaditano – Onubo – Algarviense R21 XII Bética R06
Las poblaciones en las que se basa este trabajo sólo suponen una pequeña
muestra de todas las poblaciones de romero presentes en la Península Ibérica. Se ha
intentado tener al menos una población de cada una de las provincias
biogeográficas.
Estas poblaciones eran casi todas poblaciones con un alto número de
individuos, que no presentaban tamaños poblacionales limitantes. Sólo las
poblaciones R07, R15 y R21 mostraron un censo de menos de 1.000 individuos.
3.1.2. Recolección del material
En el proceso de recolección del material se tomaron semillas de por los
menos 100 individuos, además de muestras de hojas separadas de 6 individuos para
extracción de ADN y 5 ejemplares para preparar pliegos de herbario.
En la recolección de hojas, destinadas a la extracción del ADN y objeto de
este trabajo, se debe tener en cuenta que siempre se recogieron las hojas más
jóvenes posibles. Esta recogida de material se realizó al mismo tiempo que la
recolección de semillas.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 39
Se recogieron muestras de 6 individuos, una muestra (tallo) por individuo.
Cada muestra se envolvió en una toalla de papel, se humedeció (sin llegar a
encharcarla) y se guardó en una bolsa de plástico pequeña con cierre (siempre una
sola muestra por bolsa). Las bolsas fueron marcadas con el código de la población
(ej. R4) y otro número para indicar el individuo (ej. R4‐1). También se anotó la fecha
de recolección. Estas muestras se guardaron en nevera portátil, transcurriendo el
menor tiempo posible desde su recogida hasta su procesado de congelación.
Inicialmente, se recolectaron 24 poblaciones, pero durante el desarrollo de
los ensayos se descartaron algunas muestras por problemas en las amplificaciones,
pudiéndose estudiar sólo las mostradas en la tabla 1. Finalmente se incluyeron en el
análisis las muestras de 13 poblaciones.
3.1.3. Conservación del material
Al llegar al laboratorio, se seleccionaron las hojas más limpias y sanas,
prefiriéndose las hojas más jóvenes. Las hojas se limpiaron con una toalla de papel
humedecida con alcohol (arrastrando para quitar posibles depósitos). Si las hojas
resultaban ser demasiado grandes se partieron, empleando bisturí y pinzas.
Se almacenaron muestras de cada individuo recogido de forma individual y
muestras que contenían mezcla de tres individuos (mezcla de los individuos 1, 2 y 3 y
mezcla de los individuos 4, 5 y 6). La primera mezcla se la denonima “1” (por
ejemplo, de la población R01, R1.1) y a la segunda “2” (ej. R1.4). En todos los casos se
conservaron dos eppendorfs (previamente autoclavados) que contienen
aproximadamente 20 mg de hoja.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 40
3.2. Extracción de ADN
3.2.1. Protocolo de extracción de ADN
De cada población se utilizaron las dos muestras que contenínan hojas de 3
individuos cada una, excepto de las poblaciones R07 y R19, que sólo se pudieron
utilizar una de cada (R7.4 y R19.1).
Para la extracción del ADN de las hojas de las muestras tomadas se siguió el
método de Gawel & Jarret (1991) ligeramente modificado, quedando como se
especifica a continuación:
Se utilizaron aproximadamente 20 mg de material vegetal por muestra. Se
congeló con nitrógeno líquido en eppendorfs de 1,5 mL y se pulverizó el
material en el mismo tubo, utilizando un dispositivo manual (minimorteros
de plástico) hasta alcanzarse un grado de homogeneidad buena y suficiente.
Inmediatamente se añadieron 700 μL de tampón de extracción (mezcla de 2%
p/v CTAB (ver Anejo I) + 3,8 g∙L‐1 bisulfito de sodio + PVP 1%) al que se añadió
0,2% de β‐mercaptoetanol, sólo antes de su uso y precalentado a 65ºC. Se
mantuvieron las muestras en incubación a 65ºC durante, al menos, 1 hora.
Se añadió 600 μL de cloroformo:alcohol isoamílico (24:1 v/v) y se mezclaron
las muestras por inversión durante 5 minutos a temperatura ambiente.
Se centrifugó a 12.000 r.p.m. durante 5 minutos y el sobrenadante se
transfirió a un eppendorf limpio.
Se añadieron 500 μL de isopropanol frío (‐20ºC) a los 500 μL del
sobrenadante recuperado y se mantuvo en el congelador durante, al menos,
10 minutos. Después de mezclar por inversión se centrifugó nuevamente a
12.000 r.p.m. durante 5 minutos para precipitar el ADN.
Se eliminó el isopropanol completamente y se añadieron 300 μL de etanol al
70% a temperatura ambiente para lavar el precipitado de ADN.
Inmediatamente se centrifugó a 12.000 r.p.m. durante 5 minutos.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 41
Después de 1 ó 2 lavados consecutivos se dejaron los tubos abiertos e
invertidos durante al menos 12 horas para asegurar que el ADN centrifugado
quedara libre de alcohol;
Finalmente se resuspendieron las muestras en 100 μL de agua destilada y se
incubaron durante 2 horas a 37ºC.
El ADN extraído se conservó a ‐20ºC.
3.2.2. Preparación de las soluciones de trabajo
Se comprobó el estado y concentración del ADN extraído mediante
comparación con concentraciones conocidas de ADN en gel de agarosa.
Las extracciones originales de ADN se diluyeron en agua destilada
esterilizada hasta alcanzar una concentración de aproximadamente 2 μg∙μL‐1. Estas
diluciones son las soluciones de trabajo con las que se prepararon las
amplificaciones.
3.3. Amplificación del ADN (RAPDs)
Para las amplificaciones, inicialmente se probaron trece primers
decanucleótidos de secuencia arbitraria (se obtuvieron de Operon Technologies Inc,
Alameda, California. USA: OPF_03, OPF_08, OPF_13, OPO_04, OPO_05, OPO_06,
OPO_07, OPO_10, OPO_12, OPO_15, OPO_16, OPO_18, OPO_20). Fueron
seleccionados (OPF_13, OPO_07, OPO_10, OPO_15 y OPO_20) aquellos que
alcanzaron mayor nivel de polimorfismo (calidad de información aportada: bandas
consistentes y bien definidas en el mayor grado posible) y generaron un mayor
número de bandas, con el objetivo de hacer más eficaz el análisis de las muestras
recogidas.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 42
3.3.1. Protocolo de amplificación mediante PCR
Las reacciones de amplificación del ADN fueron realizadas en volúmenes de
25 μL que contenían los compuestos mostrados en la tabla 3.
Tabla 3. Componentes y cantidades de los compuestos usados en la PCR.
Componente Cantidad agua 14,85 μL
10 x buffer (1) 2,50 μL
Cl2Mg (50mM) 0,75 μL
dNTPs (50mM) 0,4 μL
primer (10 mM) 0,5 μL
Taq ADN polimerasa (2) 1,0 μL
ST muestra 5,0 μL
* 1 y 2 – tampón provisto por la casa comercial de la enzima (Biotools)
La mezcla de la reacción fue cubierta por una gota de aceite mineral. La
amplificación se realizó en un termociclador (DNA Thermal Cycler 480, Perkin‐Elmer
CETUR), que llevó a cabo la siguiente secuencia de ciclos programada (tabla 4).
Tabla 4. Ciclos, temperatura y tiempos de la PCR.
Num. ciclos Temperatura Tiempo 1 ciclo 94ºC 1 min.
35 ciclos 92ºC 45 s.
37ºC 1 min.
72ºC 2 min.
1 ciclo 72 ºC 3 min.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 43
3.4. Separación de los productos de amplificación por electroforesis
3.4.1. Electroforesis en gel de agarosa al 1,5 % en TBE
Una vez realizada la amplificación de las muestras, los productos de
amplificación se sometieron a una electroforesis en gel de agarosa al 1,5% (p/v). Para
la elaboración de los geles se siguió el siguiente protocolo:
Se añaden 2,25 g de agarosa a 150 mL de 1 x TBE (ver Anejo I) en un
erlenmeyer.
Se calienta en microondas a 600 w durante 2 minutos tapando el recipiente.
Si es necesario se vuelve a calentar durante medio minuto más a menor
potencia para conseguir que se disuelva por completo la agarosa.
Se prepara la bandeja de electroforesis, nivelando el soporte y regulando los
peines.
Se homogeniza la muestra mediante agitación y cuando la temperatura ha
descendido a 50‐60ºC se vierte la solución en la bandeja, comprobando que
no queden burbujas o cuerpos extraños.
El gel se deja enfriando hasta que gelifica completamente.
A continuación, se insertaron en los pocillos del gel alícuotas de 12 μL de los
productos de amplificación que contenían tampón de carga (ver Anejo I). En todos
los geles se incluyó “marcador en escalera” de 100 pb (Pharmacia) para determinar
el tamaño de los productos de amplificación (Anejo I). Todas las electroforesis se
llevaron a cabo inmersas en tampón 1 x TBE (Sambrook et al. 1989), con unos
tiempos medios de 2h y 30 min y una intensidad de corriente de 100 v.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 44
3.4.2. Tinción y adquisición de imágenes
Se llevó a cabo una tinción mediante inmersión en bromuro de etidio (0,5
μg∙mL‐1), durante 10‐15 minutos, seguido de un proceso de lavado en agua de 15‐20
minutos. Los geles se visualizaron y fotografiaron bajo luz ultravioleta con cámara
digital (Kodak DC 290) y fotos Polaroid.
Con la finalidad de comprobar la reproducibilidad, se realizaron al menos 2
reacciones por cada ‘primer’ con cada una de las muestras en estudio.
3.5. Análisis estadísticos
A partir del análisis de bandas generadas por los RAPDs (Random Amplified
Polymorphic DNA) se elaboró una matriz binaria de datos donde se han tenido en
cuenta las bandas de los RAPDs por su presencia (1) o ausencia (0). Los fragmentos
amplificados fueron nombrados primero por el primer usado (OPF_13, OPO_07,
OPO_10, OPO_15 y OPO_20) y seguido del tamaño de la banda expresado en pares
de bases (pb). Esta matriz permite cuantificar el número de bandas monomórficas y
polimórficas y constituye la herramienta base de todos los análisis llevados a cabo
en este trabajo.
La matriz de datos permitió el cálculo de las matrices de similitud usando los
índices o coeficientes de JACCARD (1908) y Dice (1945) (ver Anejo IV) mediante el
programa NTSYS (Numerical Taxonomy System for Cluster and Ordination Analysis). A
continuación, los datos de la matriz de similitud se sometieron a un análisis de
clasificación, usando el método de agrupamiento UPGMA (Unweighted Pair Group
Method with Arithmatic Mean) (Wilkie et al., 1993). La representación gráfica de los
resultados del análisis de clasificación se realizó mediante la obtención de un
dendrograma o árbol de similitud, usando para ello el método gráfico de SHAN
(Sequential Agglomerative Hierarical Nested Cluster Analysis). La obtención de las

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 45
matrices cofenéticas, a partir de las matrices de similitud, para cada índice de
similitud, permitió, a través de un análisis de correlación (Test de Mantel), estimar el
ajuste de los dendrogramas.
Paralelamente, se elaboró un diagrama de coordenadas principales,
mediante el uso del programa NTSYS, con el objeto de contrastar y complementar
los resultados de agrupamiento de muestras obtenidas por los dendrogramas a
partir de las matrices de similitud según Jaccard y Dice.
Por otra parte, se procedió al cálculo de la matriz de distancias genéticas de
acuerdo a la expresión de Nei (1978) mediante el uso del programa PopGene. Para
ello hubo que realizar una pequeña conversión previa de los valores de la matriz de
similitud en valores de dis‐similitud [D = (1‐S), siendo S el valor del coeficiente de
similitud]. A partir de ella se pudo generar un nuevo dendrograma, mediante el
método UPGMA, en el que las muestras aparecen agrupadas por poblaciones.
Para analizar la posible relación entre las distancias genéticas obtenidas y la
distribución geográfica de las poblaciones estudiadas, se construyó una matriz de
distancias geográficas, la cual recoge las distancias lineales entre los puntos de
localización de las poblaciones estudiadas.
Con el fin de analizar la posible correlación de la diversidad genética
observada con las condiciones ambientales de las localizaciones de las poblaciones
consideradas, se recogieron datos ambientales de estas poblaciones para construir
una matriz de distancias ambientales. Las variables consideradas fueron: altitud,
temperatura media anual y precipitación anual. La matriz obtenida fue comparada
con la matriz de distancias genéticas a través de un test de Mantel. Asimismo, cada
una de las tres variables ambientales consideradas fue comparada de manera
independiente con la misma matriz de distancias genéticas usando el mismo
método.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIIIII.. MMaatteerriiaall yy mmééttooddooss
P á g i n a | 46
Para cuantificar la diversidad genética observada se procedió a calcular el
estimador de la diversidad genética ‘h’ mediante la expresión de Nei (1973) usando
para ello el programa PopGene.
Dentro del mismo apartado y con el fin de establecer un reparto de la
variación observada a distintos niveles, centrándose fundamentalmente en las
diferencias entre las distintas provincias biogeográficas analizadas, se realizó un
análisis de variación molecular (AMOVA) a través del programa WinAmova.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 47
4. Resultados y discusión
4.1. Caracterización de las poblaciones de romero
Para la caracterización de las 13 poblaciones de romero en estudio, se
utilizaron los marcadores moleculares RAPDs (Random Amplified Polymorphic
DNAs). La técnica de los RAPDs es una técnica muy utilizada para caracterización,
tanto de especies cultivadas como en los estudios sobre la estructura genética de
poblaciones naturales (Martín et al, 1999; Nybon y Bartish, 2000; Ortíz‐Dorda et al.,
2005).
Se analizaron 24 muestras, 2 de cada población, excepto en las poblaciones
R07 y R19 en las que finalmente sólo se pudo estudiar una muestra. Cada una de
estas muestras representa una mezcla de 3 individuos de la población (ver apartado
3.1.3 de Material y Métodos). En la figura 10 se muestra uno de los geles obtenidos
de los productos amplificados con el primer OPO_07. Otras imágenes de los geles
obtenidos están recogidas en el Anejo II.
Fig. 10. Gel obtenido mostrando los productos amplificados con el primer OPO_07.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 48
El análisis de las muestras con los 5 primers seleccionados dio lugar a 51
bandas o marcadores (Anejo III, tabla A1). La nomenclatura de las bandas especifica
en primer lugar el primer que ha sido utilizado y en segundo lugar, el peso molecular
de la banda expresado en pares de bases (pb). La nomenclatura de las muestras,
está detallada en la tabla 1 (ver apartado de Material y Métodos).
El análisis de las bandas generadas, mostrado en la matriz de datos (Anejo III,
tabla A1), permite identificar 33 bandas polimórficas y 18 monomórficas. Si se
revisan las bandas de tipo polimórfico encontramos que no hay marcadores
específicos de población y que sólo en la población de San Martín de la Virgen del
Moncayo (R09) de Zaragoza, en lo que parecería ser un marcador monomórfico
(OPO 10_900), se constata la ausencia de banda para ambas muestras. Por otra
parte, tan sólo se encontraron bandas exclusivas de un individuo para una de las
muestras de las poblaciones de Argamasilla de Alba (R5.4) y Loporzano (R10.4) en
los marcadores OPO 07_325 y OPO 20_1600, respectivamente. También se observó
que algunos marcadores estaban presentes en prácticamente todas las muestras
estudiadas, excepto en una, siendo considerado entonces polimórfico: Valdeconcha
(R4.1) en las bandas OPF 13_1500 y OPO 07_1000; San Martín de la Virgen del
Moncayo (R9.4) en la banda OPO 15_950; Tabernas (R19.4) en la banda OPO
20_2000 y Conil de la Frontera / Chiclana (R21.10) en la banda OPO 10_1400, mientras
que para el resto de muestras hay presencia.
Los resultados que hemos obtenido con estas poblaciones de romero
pueden considerarse como una primera aproximación a la estimación de la
diversidad existente entre las poblaciones de romero muestreadas, ya que el
limitado número de primers que se pudo finalmente utilizar (13 primers ensayados,
pero sólo 5 utilizados), generaron 33 bandas polimórficas, cantidad relativamente
pequeña si se compara con otros estudios de este tipo donde se han podido evaluar
un mayor número de bandas polimórficas (Segarra‐Moragues et al., 2005). Sin
embargo, hay que tener presente que la frecuencia de bandas polimórficas
aportadas por los RAPDs varían de unas especies a otras; así por ejemplo, el maíz,

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 49
especie de polinización cruzada, es altamente polimórfica, mientras que el tomate,
especie con autopolización, es menos polimórfica (Wilkie et al., 1993; Hayward y
Sackville Hamilton, 1997).
En la tabla 5 se recoge a modo de resumen la cantidad de bandas totales
generadas por cada primer, diferenciando entre polimórficas y monomórficas.
Estos datos van acompañados del grado de polimorfismo alcanzado y el tamaño de
bandas obtenidas para cada uno de ellos.
Tabla 5. Tipos de bandas, porcentaje de polimorfismo y tamaño de bandas generado (expresado en pb).
PRIMER BANDAS
AMPLIFICADAS
% POLIMORFISMO Tamaño de bandas (pb)
TOTAL P M OPF – 13 7 2 5 28,57 650 – 1800 OPO – 07 13 8 5 61,53 325 – 1300 OPO – 10 9 3 6 50,00 600 – 2300 OPO – 15 7 5 2 71,42 800 – 2000 OPO – 20 15 15 0 100,00 450 – 2000
TOTAL de bandas encontradas
51 33 18 64,70
Los resultados mostrados en la tabla 5 reflejan grandes diferencias en el
porcentaje de polimorfismos detectados por cada uno de los primers empleados.
Así, mientras en el primer OPO‐20 todos los marcadores analizados resultaron ser
polimórficos, en el primer OPF‐13 sólo el 28,57% eran polimórficos. El valor medio de
marcadores polimórficos detectados, considerando conjuntamente los 5 primers
empleados, resulta ser del 64,70%. Comparado este valor con el obtenido en otros
trabajos similares resulta un tanto bajo. En el trabajo de estudio de diversidad
genética de Rosmarinus tomentosus se obtuvo un valor del 86% de marcadores
polimórficos de las 109 bandas estudiadas (Martín y Hernández Bermejo, 2000),
igualmente se pueden ver valores similares en otros estudios de otras especies (p.e.
Renau‐Morata et al., 2005). Sin embargo, hay que tener en cuenta que la obtención

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 50
en este caso de valores tan bajos está en parte justificado por el tipo de muestras
analizadas, que son mezcla de varios individuos, mientras que en los trabajos
mencionados analizaban cada individuo separadamente.
4.2. Análisis de agrupamiento
A partir de la matriz de datos (matriz de presencias y ausencias, 0 y 1, de los
marcadores analizados para las 24 muestras estudiadas, Anejo III, tabla A1), se
calcularon los índices de similitud entre todas las muestras. La matriz de similitud
obtenida mediante el coeficiente de Jaccard (Anejo III, tabla A2) presenta valores
que van de 0,568 (entre las muestras R8.4 y R15.4) a 0,971, el valor más alto,
observado entre las dos muestras analizadas de la población R11.
Excepto en las poblaciones R07 y R19, en las que sólo se pudo analizar una
muestra, en las otras poblaciones se estudiaron 2 muestras (mezcla de 3 individuos
de la población cada una de ellas), por lo que es de esperar que los índices de
similitud entre muestras de la misma población presenten valores más altos. El valor
más alto detectado mediante el índice de Jaccard se corresponde de hecho con uno
de estos valores, el correspondiente a las muestras de la población R11 como ya se
ha mencionado. Sin embargo, aunque en general todos ellos presentan valores de
similitud bastante altos (entre 0,971 – población R21 – y 0,681 – población R09) no
son los valores más altos encontrados, habiendo índices de igual valor o incluso
mayor entre muestras de distintas poblaciones.
Los resultados del análisis de similitud se muestran gráficamente en el
dendrograma obtenido mediante el método UPGMA (figura 11). En este
dendrograma se observa que el nivel de similitud mínimo es de 0,68, donde se
separan las muestras de la población de San Martín de la Virgen del Moncayo (R09)
del resto; las muestras de dicha población muestran entre ellas un coeficiente de
similitud de 0,86.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 51
Al nivel de similitud del 69% se separa el grupo que contiene a las muestras
de la población de Alcaraz (R06) y la muestra R8.4 del resto de muestras. Las
muestras de la población R06 presentan entre sí una similitud del 84%, y se separan
de la de R8.4 al nivel de similitud 0,76.
El grupo que contiene las 19 muestran restantes se divide en otros 2
subgrupos al nivel 0,73 de similitud, separándose a su vez el mayor de ellos en otros
2 grupos a un nivel muy cercano al anterior (aproximadamente 0,74).
El primer subgrupo mencionado (grupo A en el dendrograma), contiene las
muestras de las poblaciones de Valdeconcha (R04), Argamasilla de Alba (R05) y
Leciñena (R11). Las muestras de la población R11 se separan del resto a un nivel del
0,75, presentando entre ellas una similitud del 0,97, la más alta de las observadas en
este trabajo. Sin embargo, las muestras de las poblaciones R04 y R05 no se agrupan
por población, presentando subgrupos con muestras mezcladas de ambas.
Para interpretar el otro subgrugrupo se han denominado como subgrupos B
y C (ver dendrograma de la figura 11) los 2 grupos en que se divide. El grupo B
contiene las muestras de las poblaciones del Soto del Real (R12), Torrejón el Rubio
(R15), Conil de la Frontera / Chiclana (R21) y la muestra R19.4 de la población de
Tabernas. Al igual que ocurría en el grupo A, se observa que sólo una de las
poblaciones que forman este grupo muestra un subgrupo formado únicamente con
las muestras de la población (R21 en este caso), apareciendo las muestras de las
otras dos poblaciones del grupo (R12 y R15) mezcladas entre sí. En esta
interpretación no se ha considerado la muestra R19.4 por ser la única de su
población.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 52
Fig. 11. Dendrograma UPGMA, índice de JACCARD [NTSYS 2.20] de las muetras de cada población de romero en estudio.
El grupo C contiene las muestras de las poblaciones de Turre (R1), Loporzano
(R10) y las muestras R7.1 y R8.1, de las poblaciones de Villena y Algimia de Almoacid,
respectivamente. Estas muestras, al nivel de 0,77 de similitud se escinden en dos
nuevos grupos. Por un parte, se encuentran mezcladas las muestras R7.1, R8.1 y
R10.1, y por otro las dos muestras de la población R01, agrupadas entre sí a un nivel
del 84%, junto con la muestra R10.4.
El dendrograma obtenido a partir de la matriz de similitud usando el índice de
Jaccard refleja una gran diversidad entre las muestras analizadas, tanto entre
poblaciones distintas como incluso entre las muestras de una misma población,
habiendo casos en que las dos muestras de una población aparecen agrupadas en
grupos distintos, como es el caso de las muestras de la población R08. Sólo dos
poblaciones, R11 y R21 presentan un alto grado de similitud entre sus muestras.
Grupo C
Grupo B
Grupo A

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 53
Otro de los resultados más significativos que muestra el dendrograma es el
hecho de que la muestra R19.4 correspondiente a la población de Tabernas
aparezca agrupada en el grupo B junto con las muestras de las poblaciones R12, R15
y R21, ya que esta población fue determinada en el momento de la recolección
como Rosmarinus eriocalyx. Los resultados obtenidos a partir de los marcadores
moleculares de esta muestra en este trabajo, no respaldan dicha determinación, ya
que la muestra R19.4 aparece agrupada junto con muestras de R. officinalis a un
nivel de similitud bastante alto. Existe la posibilidad de que la posición de esta
muestra dentro del dendrograma se deba a un error en la determinación durante la
recolección o a la existencia de procesos de hibridación en esta población. Sin
embargo, tampoco se puede asegurar que se trate de un híbrido entre R. officinalis y
R. eriocalyx ya que no se han incluido en el análisis muestras de otras poblaciones de
R. eriocalyx que pudieran confirmar este hecho.
Las conclusiones a las que se llega al emplear para la obtención de la matriz
de similitud el índice de Dice (Anejo III, tabla A3) y el dendrograma obtenido a partir
de esta matriz (figura 12) son similares a las obtenidas utilizando el índice de
Jaccard, aunque hay que señalar que los valores obtenidos en los coeficientes de
similitud con el primer índice son algo mayores. Esto puede deberse en parte a que
en la expresión del índice de Dice se da un peso mayor a las bandas comunes de las
muestras comparadas (Anejo IV).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 54
Fig. 12. Dendrograma UPGMA, índice de DICE [NTSYS 2.20] de las muestras de cada población de romero en estudio.
El análisis a través del test de Mantel (1967), se llevó a cabo para comprobar
el ajuste de los dendrogramas, comparando la matriz cofenética obtenida a partir
de la matriz de similitud y esta última. Los niveles de correlación que se obtuvieron
para los índices de similitud de Jaccard y Dice, fueron de 0,75396 (P < 0,05) y de
0,73405 (P < 0,05), respectivamente. Estos valores, de acuerdo con la clasificación
hecha por Rohlf (2005), están incluidos dentro del rango de correlaciones
consideradas como pobres estadísticamente (0,7 ≤ r < 0,8), pero sí son
estadísticamente significativos.
Por otra parte, para poder contrastar y complementar la información
extraída a partir de ambos dendrogramas, en lo referente a la evaluación del grado
de diferencia entre poblaciones y a su posible agrupación, se llevó a cabo la
elaboración de un diagrama de coordenadas principales (NTSYS) (figura 13). Los tres
primeros ejes explican el 18,0%, 12,1% y 10,4% de la varianza total.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 55
Fig. 13. Diagrama de coordenadas principales, índice de JACCARD [NTSYS 2.20] – Vistas 2D (a) y 3D (b) de las muestras de cada población de romero en estudio.
a
b

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 56
Como se puede apreciar en las vistas 2D (figura 13.a) y 3D (figura 13.b), las
muestras de las poblaciones R09 y R21 siguen permaneciendo juntas; en menor
grado lo hacen las de las poblaciones R04, R05, R06, R10, R11, R12 y R15. A la vista de
esta disposición de las muestras se observa que éstas se hallan muy dispersas, sin
terminarse de constituir algún subgrupo bien definido, hasta el punto de que
algunas de ellas se separan mucho una de la otra (casos de las poblaciones R01 y
R08).
La distribución 2D de las muestras se asemeja a la obtenida en el
dendrograma, pero hay que reseñar que ciertas muestras o poblaciones aparecen
conjuntamente con otras que antes no lo hacían. Respecto a la vista 3D, aparecen
ciertas diferencias en cuando al grado de similitud presentado por las muestras de
una misma población, que en la vista 2D pudieran aparentar ser más cercanas. Todo
ello refuerza los resultados obtenidos inicialmente en el dendrograma, sobre la alta
diversidad de las muestras estudiadas.
4.3. Distancias genéticas
El cálculo de las distancias genéticas fue hecho de acuerdo a la expresión de
Nei (1978) mediante el programa PopGene .
De este análisis se obtiene como resultado la matriz de distancias genéticas
para el total de poblaciones (tabla 6), la cual recoge las frecuencias de alelos de
acuerdo con Nei (1978).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 57
Tabla 6. Identidad genética de Nei (por encima de la diagonal) y distancia genética de Nei
(por debajo de la diagonal).
pop ID R1 R4 R5 R6 R7 R8 R9 R10 R11 R12 R15 R19 R21
R1 **** 0.8710 0.8885 0.7902 0.8719 0.8537 0.8070 0.9475 0.8215 0.9172 0.9181 0.8482 0.8384
R4 0.1381 **** 0.9459 0.7974 0.8303 0.8397 0.8092 0.8901 0.8033 0.8906 0.8795 0.7606 0.8323
R5 0.1183 0.0556 **** 0.8311 0.8676 0.8234 0.8772 0.9029 0.8613 0.8783 0.8502 0.7571 0.8118
R6 0.2354 0.2264 0.1850 **** 0.8014 0.8651 0.7796 0.8139 0.7298 0.8194 0.7880 0.8064 0.7673
R7 0.1370 0.1860 0.1421 0.2214 **** 0.8929 0.8325 0.9423 0.7631 0.8503 0.7998 0.7647 0.7549
R8 0.1581 0.1747 0.1943 0.1450 0.1133 **** 0.7999 0.9334 0.8081 0.8191 0.7661 0.8098 0.7577
R9 0.2144 0.2117 0.1310 0.2490 0.1833 0.2233 **** 0.8403 0.7214 0.7893 0.7464 0.7115 0.7416
R10 0.0539 0.1164 0.1021 0.2059 0.0594 0.0689 0.1739 **** 0.8728 0.9027 0.8611 0.8285 0.8006
R11 0.1966 0.2190 0.1493 0.3150 0.2704 0.2131 0.3266 0.1361 **** 0.8323 0.7817 0.8025 0.7533
R12 0.0864 0.1159 0.1297 0.1992 0.1622 0.1995 0.2366 0.1024 0.1835 **** 0.9674 0.8738 0.8810
R15 0.0854 0.1284 0.1623 0.2383 0.2234 0.2664 0.2925 0.1495 0.2463 0.0332 **** 0.8348 0.9220
R19 0.1647 0.2736 0.2782 0.2152 0.2683 0.2110 0.3403 0.1881 0.2200 0.1350 0.1805 **** 0.7549
R21 0.1763 0.1835 0.2085 0.2649 0.2811 0.2775 0.2989 0.2224 0.2834 0.1267 0.0812 0.2811 ****
A partir de la matriz de distancias genéticas de Nei (1978), el programa
PopGene permite obtener un dendrograma por poblaciones (figura 14). Este
dendrograma ofrece una disposición y agrupamiento de las poblaciones bastante
similar al dendrograma obtenido a partir de la matriz de similitud mediante el
método UPGMA. Sigue apareciendo como la población más alejada del resto la R09
(San Martín de la Virgen del Moncayo), al igual que se observaba en el dendrograma
del apartado 4.2. Esta población pertenece a la misma provincia biogeográfica que
la R12 (Soto del Real) (marcadas en el dendrograma con una flecha del mismo
color), con la cual, sin embargo, no se agrupa y de la que se encuentra bastante
alejada. Su posición en ambos dendrogramas podría deberse a que se tratara de un
error en la clasificación de la muestra o de una población resultante de un proceso
de hibridación, pero la falta de muestras de otras especies de Rosmarinus en este
análisis no permite confirmarlo, como ya se ha comentado previamente para el caso
de la población R19 (Tabernas).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 58
Fig. 14. Dendrograma basado en la distancia genética de Nei (1978): método UPGMA, índice de JACCARD [PopGene]. Las poblaciones que pertenecen a una misma provincia biogeográfica se han señalado con una elipse (en el caso de estar agrupadas juntas) o con flechas del mismo color para cada provincia.
Las otras dos poblaciones que aún perteneciendo a la misma provincia
biogeográfica no se han agrupado juntas son las poblaciones R01 (Turre) y R19. En
principio no sería de esperar este agrupamiento ya que según la clasificación de
partida se trata de dos poblaciones de, en principio, distintas especies, sin embargo
los resultados obtenidos en este análisis cuestionan este hecho.
En este estudio hay otras dos provincias biogeográficas que están
representadas por dos poblaciones distintas, las provincias V (poblaciones R07
[Villena] y R08 [Algimia de Almoacid]) y VII (poblaciones R04 [Valdeconcha] y R05
[Argamasilla de Alba]). En el caso de esta última provincia, las dos poblaciones que
la representan aparecen agrupadas juntas. En el caso de la otra provincia, VII, las
dos poblaciones que la representan aparecen en el mismo grupo, pero junto a otras
dos poblaciones correspondientes a otras dos provincias distintas. El resultado
observado en el dendrograma por muestras del apartado 4.2 confirma estos
resultados. En ese dendrograma las 4 muestras de las poblaciones R04 y R05 se

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 59
agrupaban juntas en un mismo cluster, mientras que las dos muestras de la
provincia R08 aparecían en grupos muy separados entre sí. En este dendrograma
por poblaciones este hecho se refleja en un menor grado de relación entre las
poblaciones de la provincia V que las de la VII.
También se aprecia que, según este análisis, las relaciones genéticas más
estrechas aparecen entre las poblaciones R12 (Soto del Real) y R15 (Torrejón el
Rubio), que también aparecían agrupadas juntas en el dendrograma por individuos.
Aunque ambas poblaciones pertenecen a provincias biogeográficas distintas (IX y X,
respectivamente), los lugares geográficos de recogida de muestra se encuentran a
una distancia de unos 300 km (ver figura 8), lo cual puede ser considerado como
una distancia relativamente pequeña si se tienen en cuenta las distancias medias de
las poblaciones incluidas en este trabajo.
4.4. Correlaciones entre distancias genéticas, geográficas y ambientales
4.4.1.‐ Correlación entre las distancias geográficas y genéticas
Las distancias entre los puntos de recogida de las muestras se utilizaron para
elaborar una matriz de distancias geográficas (Anejo III, tabla A4). Estas distancias
se estimaron de forma lineal sobre un mapa de la Península Ibérica. Hay que tener
en cuenta que entre dichos puntos se suceden diversos accidentes geográficos
(cordilleras, sierras, ríos, etc.) y ello afecta a las conclusiones que podamos sacar del
análisis de su correlación con la matriz de distancias genéticas.
La matriz de distancias genéticas obtenida en el apartado anterior y la matriz
de distancias geográficas elaborada se compararon mediante un test de Mantel
(1967), usando el programa NTSYS. El grado de correlación obtenido entre ambas
matrices fue de 0.2211 (P = 0,074), valor que resulta no ser significativo y además

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 60
bastante bajo, lo que pudiera estar justificado por los distintos accidentes
geográficos, que impiden el flujo genético entre poblaciones.
El hecho de que no haya una correlación entre la matriz de distancias
genéticas y geográficas, junto con el alto grado de diversidad de las muestras de
romero analizadas hasta el momento en este trabajo, ponen de manifiesto el alto
grado de relación entre las distintas poblaciones de romero de la Península, no
detectándose un proceso de aislamiento geográfico que reduzca el flujo génico.
Cuando esto ocurre, lo cual se ha detectado en diversos trabajos de análisis de
estructura poblacional de especies endémicas, las cuales suelen presentar una
reducida distribución geográfica, suele establecerse una alta correlación entre la
matriz de distancias genéticas y geográficas como un reflejo del aislamiento
geográfico de las poblaciones que conduce a una mayor diferenciación entre
poblaciones (Cardoso et al., 1998; Martín et al., 1999; Martín y Hernández‐Bermejo,
2000).
4.4.2.‐ Correlaciones entre las distancias ambientales y genéticas
Se quiso analizar el efecto que algunos factores ambientales pudieran tener
relación con la diversidad de las poblaciones estudiadas. Así se propusieron tres
variables a considerar: altitud, temperatura y precipitación anual (tabla 7).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 61
Tabla 7. Datos climáticos de los puntos de recogida de las muestras.
Población Término Provincia Datos ambientales
Alt. (m) Tm (ºC) Pm (mm)
R01‐00 Turre Almería 240 18,7 196 R04‐00 Valdeconcha Guadalajara 750 14,1 378 R05‐00 Argamasilla de Alba Ciudad Real 740 14,7 396 R06‐00 Alcaraz Albacete 1220 13,6 367 R07‐00 Villena Alicante 600 17,8 336 R08‐00 Algimia de Almoacid Castellón 700 17,0 442 R09‐00 San Martín de la Virgen del Moncayo Zaragoza 760 15,0 318 R10‐00 Loporzano Huesca 750 13,6 535 R11‐00 Leciñena Zaragoza 680 15,0 318 R12‐00 Soto del Real Madrid 1140 6,4 1326 R15‐01 Torrejón el Rubio Cáceres 460 16,1 523 R19‐01 Tabernas Almería 550 18,7 196 R21‐01 Conil de la Frontera / Chiclana Cádiz 20 17,1 603
Alt. (m) = Altitud Tm (ºC) = Temperatura media anual Pm (mm.) = Precipitación media anual (Ninyerola et al., 2005. http://opengis.uab.es/wms/iberia/index.htm)
A partir de los datos mostrados en la tabla 7, se calculó la matriz de distancia
ambiental conjunta (Anejo III, tabla A5), mediante el cálculo de las distancias
euclídeas, según la expresión:
( ) ( ) ( )2 2 212 1 2 1 2 1 210 m m m md Alt Alt x T T P P= − + − + −
donde los valores de la temperatura fueron ponderados (Tm x 10), con la finalidad
de manejar datos dentro de un mismo rango de valores. Igualmente, se calcularon
las matrices de distancias ambientales parciales para cada una de las variables
citadas (Anejo III, tablas A6, A7 y A8) a través de las expresiones:
( )212 1 2 1 2d Alt Alt Alt Alt= − = −
Altitud
( )212 1 2 1 2m m m md T T T T= − = −
Temperatura

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 62
( )212 1 2 1 2m m m md P P P P= − = −
Precipitación
Mediante la realización de un test de Mantel (1967) se calculó el grado de
correlación presente entre la matriz de distancias genéticas y las distintas matrices
ambientales mencionadas, total y parciales. El grado de correlación alcanzado fue
bajo para todas ellas (tabla 8).
Tabla 8. Correlaciones entre la matriz de distancias genética y las matrices ambientales (total y parciales).
Comparación entre matrices Correlación (r) Pvalue
d.ambiental total – d.genéticas ‐ 0,00039 0,9990 > 0,05 d.amb_altitud – d.genéticas 0,02182 0,8412 > 0,05 d.amb_temperatura – d.genéticas ‐ 0,18577 0,1096 > 0,05 d.amb_precipitación – d.genéticas 0,00321 0,9610 > 0,05
Estos resultados parecen indicar que no existe una relación clara entre los
parámetros ambientales considerados y las diferencias genéticas encontradas en las
poblaciones estudiadas en este trabajo. Hay que tener en cuenta que el grado de
diversidad genética reflejada en los análisis realizados hasta el momento en estas
muestras de romero es bastante alto y que se trata de una especie con una amplia
distribución geográfica y capaz de adaptarse a un amplio rango de condiciones
ambientales.
4.5. Diversidad genética
Para evaluar la diversidad genética de las muestras incluidas en este trabajo
se han realizado los siguientes análisis: estimación de la diversidad genética
mediante la expresión de Nei (1973) y el análisis de la varianza molecular (AMOVA).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 63
Estos estudios nos aportan información complementaria con el fin de
caracterizar la estructura genética de las poblaciones analizadas a nivel global, por
provincias biogeográficas y por población individual.
4.5.1. Diversidad genética según Nei
La diversidad genética según Nei (1973) viene dada por la expresión:
21 1 l u luh m P= − ∑ ∑ Nei (1973)
donde Plu es la frecuencia de la banda ‘u’ en el locus ‘l‘ y ‘m’ es el número de loci.
La diversidad genética (h), calculada con el programa PopGene según Nei
(1973), se estimó analizando las bandas generadas por los RAPDs, por un lado las
bandas totales (mono y polimórficas) y, por otro lado, las exclusivamente
polimórficas. Los resultados obtenidos se muestran en la tabla 9. En los cálculos
realizados con el programa PopGene se han llevado a cabo con la opción del
programa de equilibrio de Hardy‐Weinberg.
Diversidad genética para el total de poblaciones
En los resultados de la diversidad total encontramos un valor medio de ‘h’ de
0,2009 (d.e.: 0,1966), mientras que para la debida sólo a las bandas polimórficas ‘hp‘
toma un valor de 0,3105 (d.e.: 0,1590). Ambos valores son bajos, aunque era
esperado un valor mayor asociado a las bandas exclusivamente polimórfica.
Sería de esperar un valor más alto para una especie de distribución tan
amplia. Segarra‐Moragues et al. (2005) obtuvieron un valor de h = 0,38 para la
especie Borderea chouardii. No hay que olvidar que todos los valores de diversidad

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 64
genética calculados en este trabajo se han obtenido a partir de muestras que son
mezcla de varios individuos, lo cual reduce los valores de diversidad.
Diversidad genética por provincias biogeográficas
Para analizar los datos de la tabla 9, lo primero que hay que tener en cuenta
es que no es lo mismo considerar provincias con una o con dos poblaciones, y que
siempre hay sólo dos muestras de pool de individuos por población. Por tanto, estos
valores que se ofrecen son meramente indicativos y sirven para comparar
provincias con la misma representación.
Tabla 9. Diversidad genétcia según Nei (1973) para bandas totales y bandas polimórficas.
Provincias biogeográficas Poblaciones Bandas totales (h) Bandas polimórficas (hp)
Media d.e. Media d.e.
prov I [Pirenaica] R10 0,0975 0,1775 0,1506 0,2023
prov IV [Aragonesa] R11 0,0081 0,0580 0,0126 0,0721
prov V [Catalana – Valenciano ‐ Provenzal] R07 R08 0,1058 0,1875 0,1635 0,2126
prov VII [Castellano – Mestrazgo ‐ Manchega] R04 R05 0,0913 0,1661 0,1411 0,1893
prov VIII [Murciano ‐ Almeriense] R1 R19 0,1016 0,1788 0,1570 0,2024
prov IX [Carpetano – Ibérico ‐ Leonesa] R09 R12 0,1429 0,2125 0,2208 0,2298
prov X [Luso ‐ Extremadurense] R15 0,0244 0,0984 0,0377 0,1209
prov XI [Gaditano – Onubo ‐ Algarviense] R21 0,0081 0,0580 0,0126 0,0721
prov XII [Bética] R06 0,0569 0,1440 0,0879 0,1720
Como ya se explicó en el apartado de Material y Métodos, la Península Ibérica
queda dividida en 12 provincias biogeográficas, de las cuales son las provincias V
(Catalana – Valenciano – Provenzal), VII (Castellano – Mestrazgo – Manchega), VIII
(Murciano – Almeriense) y IX (Carpetano – Ibérico – Leonesa), las que cuentan con
al menos dos poblaciones por provincia biogeográfica en este trabajo. Sin embargo,
de éstas sólo las provincias VII y IX habrán de considerarse que cuenta con dos
poblaciones, porque de las otras dos provincias (V y VIII) hubo que eliminar la
poblaciones R07 y R19, respectivamente, debido a que ambas cuentan con una

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 65
única muestra por población y dificultan tanto los cálculos como las conclusiones
que se puedan sacar.
Como se puede apreciar, los valores medios de la diversidad varían dentro del
rango de 0,0081 (d.e.: 0,0580) a 0,1429 (d.e.: 0,2125) en el caso de bandas totales (h)
y de 0,0126 (d.e.: 0,0721) a 0,2208 (d.e.: 0,2298) para el caso de bandas polimórficas
(hp).
Por un lado, tanto en el primer caso como en el segundo, el resultado menor
corresponde a la provincia XI (Gaditano – Onubo – Algarviense) que está constituida
por una única población con dos muestras, la R21 (Conil de la Frontera / Chiclana);
esta población fue la que presentó en el dendrogama uno de los coeficientes de
similitud más altos entre muestras. En contraposición, tanto en el primer caso como
en el segundo, el resultado mayor corresponde a la provincia IX (Carpetano –
Ibérico – Leonesa) que está constituida por las poblaciones de San Martín de la
Virgen del Moncayo (R09) y Soto del Real (R12).
Diversidad genética para cada población
Como se aprecia en la tabla 10, los valores de las poblaciones que
corresponden a provincias en las que sólo se ha considerado una población siguen
siendo los mismos. A este nivel de estudio, no se pudo estimar ni la diversidad
genética total (h), ni por bandas polimórficas (hp) de las poblaciones de Villena
(R07) y Tabernas (R19), debido a que sólo se cuenta con una muestra para tales
poblaciones. Para el resto, los valores de la diversidad varían dentro del rango de
0,0081 (d.e.: 0,0580) a 0,0975 (d.e.: 0,1775) para el caso de bandas totales (h) y de
0,0126 (d.e.: 0,0721) a 0,1506 (d.e.: 0,2023) para el caso de bandas polimórficas (hp).
Son en general valores muy bajos a causa, como ya se ha explicado, del tipo de
muestras analizadas.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 66
Los valores más bajos son los alcanzados por las poblaciones de Leciñena
(R11) y de Conil de la Frontera / Chiclana (R21), mientras que el más alto por la
población de Loporzano (R10). Este resultado no ha de ser considerado como
representativo, debido a que el estudio por poblaciones está limitado por la
condición de las muestras manejadas en este trabajo, que son mezclas de varios
individuos. Para contar con resultados de mayor significación se habrán de realizar
estudios posteriores con muestras únicas de individuos y aumentando el tamaño de
muestra recogido por población.
Tabla 10. Diversidad para cada población según Nei (1973).
Población Bandas totales (h) Bandas polimórficas (hp)
Media d.e. Media d.e.
R01 0,0487 0,1348 0,0753 0,1622 R04 0,0569 0,1440 0,0879 0,1720 R05 0,0569 0,1440 0,0879 0,1720 R06 0,0569 0,1440 0,0879 0,1720 R07 0,0000 0,0000 0,0000 0,0000 R08 0,0812 0,1661 0,1255 0,1933 R09 0,0406 0,1244 0,0628 0,1508 R10 0,0975 0,1775 0,1506 0,2023 R11 0,0081 0,0580 0,0126 0,0721 R12 0,0325 0,1125 0,0502 0,1373 R15 0,0244 0,0984 0,0377 0,1209 R19 0,0000 0,0000 0,0000 0,0000 R21 0,0081 0,0580 0,0126 0,0721
En todos los casos ellos se ha partido de la suposición inicial de equilibrio de Hardy – Weinberg.
4.5.2. Análisis de la varianza molecular (AMOVA)
Los resultados aportados por el AMOVA (tabla 11) permiten hacer un reparto
de la variabilidad total detectada en los análisis de los marcadores moleculares
realizados en tres niveles distintos. Considerando la diversidad presente dentro de
las poblaciones consideradas, entre las poblaciones comprendidas dentro de una
misma zona o región establecida y entre las diferentes regiones.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 67
En este análisis se han considerado como regiones las provincias
biogeográficas especificadas en Material y Métodos (tabla 2). Hay que tener en
cuenta que la mayoría de estas provincias (‘regiones’) viene representada por una
única población y éstas a su vez por tan sólo dos muestras. Este hecho hace que,
aunque el análisis permite el reparto de la varianza en los tres niveles mencionados,
la discusión de los resultados en este trabajo se centre en la partición de la variación
entre y dentro de regiones ya que sería el caso que mejor se ajuste al tipo de datos
manejados.
Tabla 11. Resultado del AMOVA de poblaciones de romero a distintos niveles.
Componente variación % total de la varición
ANÁLISIS GLOBAL Variación entre grupos V(A): 1,03688679250 12.59 %
Variación entre poblaciones dentro de grupos V(B): 3,1261363620 37.96 %
Variación dentro de poblaciones V(C): 4,0727272720 49.45 %
ANÁLISIS DE POBLACIONES
Variación entre poblaciones V(A): 4,12531818180 50.32 % Variación dentro de poblaciones V(B): 4,07272727270 49.68 %
ANÁLISIS DE REGIONES
Variación entre regiones V(A): 3,23289550070 39.10 % Variación dentro de regiones V(B): 5,03461538460 60.90 %
El reparto de la variación en este nivel muestra una variación del 39%
atribuible a las diferencias entre regiones y del 61% a las encontradas dentro de
regiones.
En estudios similares de especies con polinización cruzada los valores de la
variación dentro de regiones suelen ser ligeramente superiores (p.e. Martín et al.,
1999; Martín y Hernández Bermejo, 2000), pero en este caso hay que tener en
cuenta que los valores son más bajos porque se están considerando regiones en las
que realmente sólo hay una población.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 68
Para poder abordar en profundidad este análisis sería necesario poder
estudiar un mayor número de muestras de cada población, o al menos tener
representadas todas las provincias biogeográficas con más de una población.
4.6. Conclusiones
Este trabajo ha permitido establecer la metodología para el uso de
marcadores moleculares tipo RAPD en el estudio de la diversidad genética presente
en las poblaciones de Rosmarinus officinalis distribuidas por una gran parte de la
Península Ibérica.
Los resultados obtenidos muestran la alta diversidad existente entre las
poblaciones analizadas. Asimismo los marcadores empleados han permitido
establecer la singularidad de las poblaciones de las que proceden las muestras de
semillas conservadas en el banco de Germoplasma de la UPM correspondientes a
las mismas poblaciones estudiadas, ya que ninguna de las muestras analizadas
mostraba el mismo patrón de marcadores, incluso entre muestras de la misma
población.
El tipo de muestras empleadas en este trabajo, mezcla de individuos de una
misma población, fue elegido porque permite abordar el análisis de un mayor
número de poblaciones al mismo tiempo sin que cada población quede reducida a
un único genotipo (correspondiente a sólo un individuo). Este tipo de muestras se
utilizan con frecuencia por ejemplo en el análisis de accesiones de bancos de
germoplasma (Martín et al., 1997). El empleo de este tipo de muestras ha permitido
en este trabajo poder establecer unos resultados preliminares acerca de las
diferencias interpoblacionales en romero a partir del análisis de unas pocas
muestras.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 69
El estudio de las poblaciones teniendo en cuenta la división en provincias
biogeográficas ha puesto de manifiesto ciertas diferencias entre las distintas
provincias aunque no de forma general en todos los casos. Los dendrogramas
obtenidos muestran por ejemplo un alto grado de relación entre las dos
poblaciones analizadas de la provincia VII, pero al mismo tiempo también revelan
pocas diferencias entre otras provincias, ya que aparecen muestras de poblaciones
de distintas provincias agrupadas juntas.
La falta de correlación observada entre la matriz de distancias genéticas y las
matrices de distancias geográficas y ambientales refuerza esta hipótesis de falta de
diferenciación entre las distintas provincias biogeográficas. Estos datos reflejarían
un alto grado de flujo génico entre las distintas poblaciones de romero de la
península que están además distribuidas de manera continua en una amplia zona,
dando como resultado un alto grado de diversidad. Si bien no parece haber una
diferencia significativa entre las distintas provincias, hay que tener en cuenta que en
este análisis ninguna muestra presentó una similitud del 100% con ninguna otra.
Para poder corroborar estos primeros resultados sería necesario ampliar el
número de poblaciones estudiadas por cada una de las provincias biogeográficas.
Una de las poblaciones incluidas en este estudio estaba inicialmente
determinada como Rosmarinus eriocalyx (población R19), y como tal ha sido
conservada la muestra correspondiente en el Banco de Germoplasma de la UPM.
Los análisis realizados en este trabajo no parecen confirmar esta determinación, ya
que la muestra de dicha población no aparece claramente separada del resto de
muestras de R. officinalis. La explicación pudiera estar, bien en un error en la
determinación del material vegetal en origen o en que se trate de un híbrido entre
ambas especies. Para poder confirmar la identificación de la población R19 y
comprobar que se trata realmente de un fenómeno de hibridación sería necesario
incluir en el análisis mediante marcadores moleculares una muestra de la que se
tuviera la certeza que corresponde a R. eriocalyx. Sin embargo, el hecho de que haya

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs IIVV.. RReessuullttaaddooss yy ddiissccuussiióónn
P á g i n a | 70
poblaciones, de las que hay certeza de que son realmente R.officinalis, y que
muestran muchas más diferencias, como es la población R09 de San Martín de la
Virgen del Moncayo, hacen pensar que la posición en el dendrograma de R19 se
deba más a un error de determinación que a un caso de hibridación.
Estos resultados ponen de manifiesto la importancia de la caracterización de
los recursos fitogenéticos y la necesidad de abordar el estudio de las muestras que
de una manera u otra van a ser sometidas a procesos de conservación (Callow et al.,
1997).
Desde la perspectiva de la conservación, la estimación exacta de la diversidad
genética dentro y entre poblaciones es de gran importancia para el correcto diseño
de estrategias de conservación y gestión de los recursos genéticos de una especie
(Martín y Hernández Bermejo, 2000). En este trabajo sólo se ha podido abordar el
análisis entre poblaciones, ya que como ha quedado reflejado en los resultados,
para poder establecer el análisis de diversidad intrapoblacional es necesario el
estudio de los individuos de la población de manera independiente.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 71
5. Bibliografía
LL II BBRROOSS YY AARRTT ÍÍ CCUULLOOSS
Aberturas Aguado, A. 1986. Las aromáticas en La Alcarria. Enciclopedia la de la
Provincia de Guadalajara. Caja Provincial de Guadalajara, Guadalajara.
Abramovic, H. y Abram, V. 2006. Effect of added rosemary extract on
oxidative stability of Camelina sativa oil. Acta Agriculturae Slovenica 87:
255 – 261.
Almela, L., Sánchez‐Muñoz, B., Ferández‐López, J.A., Roca, M.J. y Rabe, V.
2006. Liquid chromatographic‐mass spectrometric analysis of phenolics and
free radical scavenging activity of rosmery extract from different raw
material. Journal of Chromatography A 1120: 221‐229.
Atti‐Santos, A.C., Rossato, M., Fernandes Pauletti, G., Duarte Rota, L., Ciro
Rech, J., Regina Pansera, M., Angostini, F., Atti Serafini, L. y Moyna, P.
2005. Physico‐chemical evaluation of Rosmarinus officinalis L. essential oils.
Brazilian Archives of Biology and Technology. 48: 1035‐1039.
Bachmann, K. 2001. Evolution and the genetic analysis of populations: 1950‐
2000. Taxon 50: 7‐45.
Cardoso, MA, Provan, J, Powell, W, Ferreiras, PCG y de Oliveira, DE. 1998.
High genetic differentation among remmant populations of the
endangered Caesalpinia echinata Lam. (Leguminosae‐Caesalpinoideae).
Molecular Ecology., 7:601‐608.
Carvalho Jr. R. N., Moura, L.S., Rosa, P.T.V., Angela, M. y Meireles, A. 2005.
Supercritical fluid extraction from rosemary (Rosmarinus officinalis): Kinetic
data, extract´s global yield, composition, and antioxidant activity. Journal
of Supercritical Fluids 35: 197‐204.
Cronquist, A. 1981. An Integrated System of Classification of Flowering Plants.
Columbia Univ. Press, New York.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 72
De Bolòs, O. y Vigo, J. 1995. Flora dels Països Catalans. Vol. 3. Editorial Barcino,
Barcelona.
Del Bano, M.J., Lorente, J., Castillo, J., Benavente‐García, O., del Río J.A.,
Ortuno, A., Quirin, K.W. y Gerard, D. 2003. Phenolic diterpenes, flacones,
and rosmarinic acid distribution during the development of leaves, flowers,
ítems, and roots of Rosmarinus officinalis. Antioxidant activity. Journal of
Agricultural and Food Chemistry 51: 4247‐4253.
Dice, L. R. 1945. Measures of the amount of ecologic associaton between
species. Ecology 26: 297‐302.
Elamrani, A., Zrira, S., Benjilali, B., Berrada, M. 2000. A study of Moroccan
rosemary oils. Journal of Essential Oil Research 12: 487‐495.
Fernández‐Pola, J. 2001. Cultivo de Plantas Medicinales, Aromáticas y
Condimenticias. Ediciones Omega, Barcelona.
Ferreras, C. y Arozena, M.E. 1987. Guía Física de España. Vol.2. Los bosques.
Alianza Editorial, Madrid.
Font Quer, P. 1981. Plantas medicinales. El Dioscórides Renovado. Ed. Labor,
Barcelona.
Gawel, NJ, Jarret, R. 1991. A modified CTAB DNA extraction procedure for
Musa and Ipomea. Plant Molecular Biology Reports 9: 262‐266.
García Olmedo F, Sanz‐Magallón G y Marín Palma E. 2001. La agricultura
española ante los retos de la biotecnología. Colección Tablero. Instituto de
Estudios Económicos, Madrid.
González‐Andrés, F. 2001. La caracterización vegetal: objetivos y enfoques. En
González‐Andrés F. y Pita Villamil JM (eds.). Conservación y Caracterización
de Recursos Fitogenéticos. Escuela Universitaria de la Ingeniería Técnica
Agrícola INEA, Valladolid. pp: 189‐198.
Hayward, M.D. y Sackville Hamilton, N.R. 1997. Genetic diversity‐‐population
structure and conservation. En: Callow, J.A., Ford‐Lloyd, B.V. y Newbruy,
H.J. (eds.). Biotechnology and Plant Genetic Resources. Conservation and
Use. C.A.B. Internacional, Wallingford. pp: 49‐76.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 73
Herrera, C.M. 2004. Distribution ecology of pollen tubes: fine‐grained, labile
spatial mosaics in southern Spanish Lamiaceae. New Phytologist 161: 473‐
484.
Hidalgo, P.J, Ubera, JL, Tena, MT, Valcarcel, M. 1998. Determination of the
carnosic acid content in wild and cultivated Rosmarinus officinalis. Journal
of Agricultural and Food Chemistry 46: 2624‐2627.
Iriondo Alegría JM. 2001. Conservación de recursos fitogenéticos. En
González‐Andrés F. y Pita Villamil JM (eds.). Conservación y Caracterización
de Recursos Fitogenéticos. Escuela Universitaria de la Ingeniería Técnica
Agrícola INEA, Valladolid. pp: 15‐32.
Jaccard, P. 1908. Nouvelles recherches sur la distribution florale. Bulletin de la
Société vaudoise des Sciencies Naturalles 44: 223‐270.
Klaas, M. 1998. Applications and impact of molecular markers on evolutionary
and diversity studies in the genus Allium. Plant Breeding 117: 297‐308.
López González, G. 1982. Guía Verde INCAFO. Árboles y Arbustos. Vol. 4.
INCAFO, Madrid.
López González, G. 2001. Los Árboles y Arbustos de la Península Ibérica e Islas
Baleares (especies silvestres y las principales cultivadas). I. Ediciones Mundi‐
Prensa, Madrid.
Mantel, N. 1967. The detection of disease clustering and a generalized
regression aproach. Cancer Res. 27: 209‐220.
Martín, C., González‐Benito, M.E. y Iriondo, J.M. 1999. The use of genetic
markers in the identification and charaterization of three recently
discovered populations of a threatened plant species. Molecular Ecology 8:
S31‐S40.
Martín, C., Juliano, A., Newbury, H.J., LU, B‐R., Jackson, M.T. y Ford‐Lloyd,
B.V. 1997. The use of RAPD markers to facilitate the identification of Oryza
species within a germoplasm collection. Genetic Resources and Crop
Evolution, 44: 175‐183.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 74
Martín, J.P. y Hernández Bermejo, J.E. 2000. Genetic variation in the endemic
and endangered Rosmarinus tomentosus Huber‐Morath & Maire (Labiatae)
using RAPD markers. Heredity 85:434‐443.
Mendiola, M.A. 1989. Plantas aromáticas de la España Peninsular. Ediciones
Mundi‐Prensa, Madrid.
Morales, R. 2000. Diversidad en labiadas mediterráneas y macaronésicas.
Portugaliae Acta Biologica. 19: 31‐48.
Moreno, S., Scheyer, T., Romano, C.S. y Vojnov, A.A. 2006. Antioxidant and
antimicrobial activities of rosmery extracts linked to their polyphenol
composition. Free Radical Research 40: 223‐231.
Muñoz, F. 1987. Plantas medicinales y aromáticas. Estudio, cultivo y
procesado. Ediciones Mundi‐Prensa, Madrid.
Nei, M. 1973. Analysis of gene diversity in subdivided populations. Proc Natl
Acad Sci USA 70: 3321‐3323.
Nei, M. 1978. Estimation of average heterozygosity and genetic distances from
a small number of individuals. Genetics 89: 583‐590.
Nevo, E. 1998. Genetic diversity in wild cereals: regional and local studies and
their bearing on conservation ex situ and in situ. Genetic Resources and
Crop Evolution 45: 355‐370.
Nybon, H y Bartish, IV. 2000. Effects of life history traits and sampling
strategies on genetic diversity estimates obtained with RAPD markers in
plant. Perspectives in Plan Ecology. Evolution and Systematics, 3: 93‐144.
Ortíz‐Dorda, J., Martínez‐Mora, C., Correal, E., Simón, B. y Cenis, J.L. 2005.
Genetic structure of Atriplex halimus populations in the Mediterranean
basin. Annals of Botany 95: 827‐834.
Ramírez, P., R. García‐Risco, M., Santoyo, S., Señoráns, F.J., Ibáñez, E. y
Reglero, G. 2006. Isolation of functional ingredients from rosemary by
preparative‐supercritical fluid chromatography (Prep‐SFC). Journal of
Pharmaceutical and Biomedical Analysis 41: 1606‐1613.
Rivas‐Martínez et al. 1987. Memoria del mapa de series de vegetación de
España. ICONA. Madrid.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 75
Renau‐Morata, B., Nebauer, S.G., Sales, E., Allainguillaume, J., Caligari, P. y
Segura, J. 2005. Genetic diversity and structure of natural and manager
populations of Cedrus atlantica (Pinaceae) assessed using random amplified
polymorphic DNA. American Journal of Botany 92: 875‐884.
Rohlf, F.J. 2005. NTSYSpc. Numerical Taxonomy and Multivariate Analysis
System (versión 2.2). Getting Started Guide. State University of New York,
New York.
Rosúa, J.L. 1986. Contribución al estudio del género Rosmarinus L. en el
Mediterráneo occidental. Lagascalia 14: 179‐187.
Salido, S., Altarejos, J., Nogueras, M., Sánchez, A. y Luque, P. 2003. Chemical
composition and seasonal variations of rosemary oil from southern Spain.
Journal of Essential Oil Research 15: 10‐14.
Sambrook, J, Fritsch, E.F y Maniatis, T. 1989. Molecular Cloning: a Laboratory
Manual (2º ed.). Cold Spring Harbor Laboratory Press. Cold Spring Harbor,
New York.
Sanchis Duato, E.; Fos Causera, M. y Bordón Ferré, Y. 2003. Ecosistemas
Mediterráneos. Ed. Universidad Politécnica de Valencia, Valencia.
Scarascia‐Mugnozza GT y Perrino, P. 2002. The history of ex situ conservation
and use of plant genetic resources. IPGRI, Roma.
Segarra‐Moragues, J.G.; Palop‐Esteban, M.; González‐Candelas, F. y Catalán, P.
2005. On the verge of extinction: genetics of the critically endangered
Iberian plant species, Borderea chouardii (Dioscoreaceae) and implications
for conservation management. Molecular Ecology 14: 969‐982.
Torres Lamas M.E. 2001. Caracterización mediante marcadores moleculares
basados en ADN. En González‐Andrés F. y Pita Villamil JM (eds.).
Conservación y Caracterización de Recursos Fitogenéticos. Escuela
Universitaria de la Ingeniería Técnica Agrícola INEA, Valladolid. pp: 235‐252.
Tutin, T.G. et al. 1972. Flora Europaeae. Ed. Cambridge University Press.
Cambridge.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 76
Wang, C‐N., Möller, M. y Cronk, Q.C.B. 2004. Population genetic structure of
Titanotrichum oldhamii (Gesneriaceae), a subtropical bulbiliferous plant with
mixed sexual and asexual reproduction. Annals of Botany 93: 201‐209.
Westman, A.L. y Kresovich, S. 1997. Use of molecular marker techniques for
description of plant genetic variation. En Callow, J.A., Ford‐Lloyd, B.V. y
Newbruy, H.J. (eds.). Biotechnology and Plant Genetic Resources.
Conservation and Use. C.A.B. Internacional, Wallingford. pp: 9‐48.
Wilkie, S.E., Isaac, P.G. y Slater, R.J. 1993. Random amplified polymorphic DNA
(RAPD) markers for genetic analysis in Allium. Theorical and Applied
Genetics 86: 497‐504.
Zaouali, Y., Messaoud, C., Ben Salah, A., Boussaid, M. 2005. Oil composition
variability among populations in relationship with their ecological areas in
Tunisian Rosmarinus officinalis L. Flavour and Fragance Journal 20: 512‐520.
PPÁÁGG IINNAASS WWEEBB
Food and Agriculture Organization of the United Nations. Tratado
internacional sobre los recursos fitogenéticos para la alimentación y la
agricultura. Comisión de recursos genéticos para la alimentación y la
agricultura, 2001.
http://www.fao.org/
Ninyerola M., Pons X y Roure JM. 2005. Atlas Climático Digital de la Península
Ibérica. Metodología y Aplicaciones en Bioclimatología y Geobotánica.
Universidad Autónoma de Barcelona, Bellaterra
http://opengis.uab.es/wms/iberia/index.htm
Ministerio de Agricultura, Pesca y Alimentación. Premios Mejor Empresa
Alimentaria Española – Edición 2005. Dossier de prensa. Madrid, 16 de
noviembre de 2006.
http://www.mapya.es/

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VV.. BBiibblliiooggrraaffííaa
P á g i n a | 77
PPRROOGGRRAAMMAASS II NNFFOORRMMÁÁTT II CCOOSS
NTSYS. Copyright © 1986‐2006. Numerical Taxonomy and Multivariate Analysis
System (version 2.2). Exeter Software. Applied Biostatistics Inc.
PopGene. Copyright © 1997. Freeware for Population Genetic Analysis (version
1.31). A joint project of Molecular Biology and Biotechnology Center
(University of Alberta) and Center for International Forestry Research.
WinAMOVA. Copyright © 1992‐1993. Analysis of Molecular Variance (version
1.55). Laurent Excoffier. Genetics and Biometry Laboratories. University of
Geneva.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 78
6. Anejos
Anejo I. Relación de las soluciones empleadas durante en los ensayos de
laboratorio.
Tampón CTAB
Composición Cantidad
2% hexadeciltrimetilamonio bromuro (CTAB, p/v) 10 g
100 mM tris‐HCl, pH 8,0 6,05 g en 500 ml (medir pH=8)
1,4 M NaCl 40,90 g
20 mM EDTA 3,72 g
Tampón de extracción.
Composición Cantidad
CTAB buffer (p/v) 20 mL bisulfito sódico (3,8 g/L) 0,076 g β‐mercaptoetanol (0,2% v/v) 40 μL PVP (1% p/v) 0,2 g
10 x TBE (para 1 litro).
Composición Cantidad Tris 109,0 g Ácido bórico 55,6 g 0,5 M Na2 EDTA, pH=8,0 40,0 mL
Na2EDTA a concentración 0,5 M y pH = 8,0 (para 1 litro).
Composición Cantidad
Na2 EDTA – 2H2O 18,6 g Disolvemos en 70 mL de H2O Ajustar pH=8,0 con 10M NaOH (no se disuelve hasta que no llega a este pH). Añadir agua hasta 100 mL*
* agua destilada y exenta (Bi‐hidro 10m) BiBraun Medical S.A.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 79
10 x Tampón de carga.
Composición Cantidad
Ficoll 400 20 g 0,2 M EDTA 37,5 mL Azul de bromofenol 100 mg Agua destilada 42, 4 mL
Marcador de pares de bases.
Composición Cantidad
Agua destilada esterilizada 800 μL 10 x tampón de carga 150 μL marcador 50 μL

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 80
Anejo II. Imágenes de algunos geles con los productos de amplificación de RAPDs
obtenidos durante la fase experimental.
Fig. A1. Primer OPF_13 (03.05.06).
Fig. A2. Primer OPO_07 (11.05.06).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 81
Fig. A3. Primer OPO_10 (10.05.04).
Fig. A4. Primer OPO_15 (11.05.06).
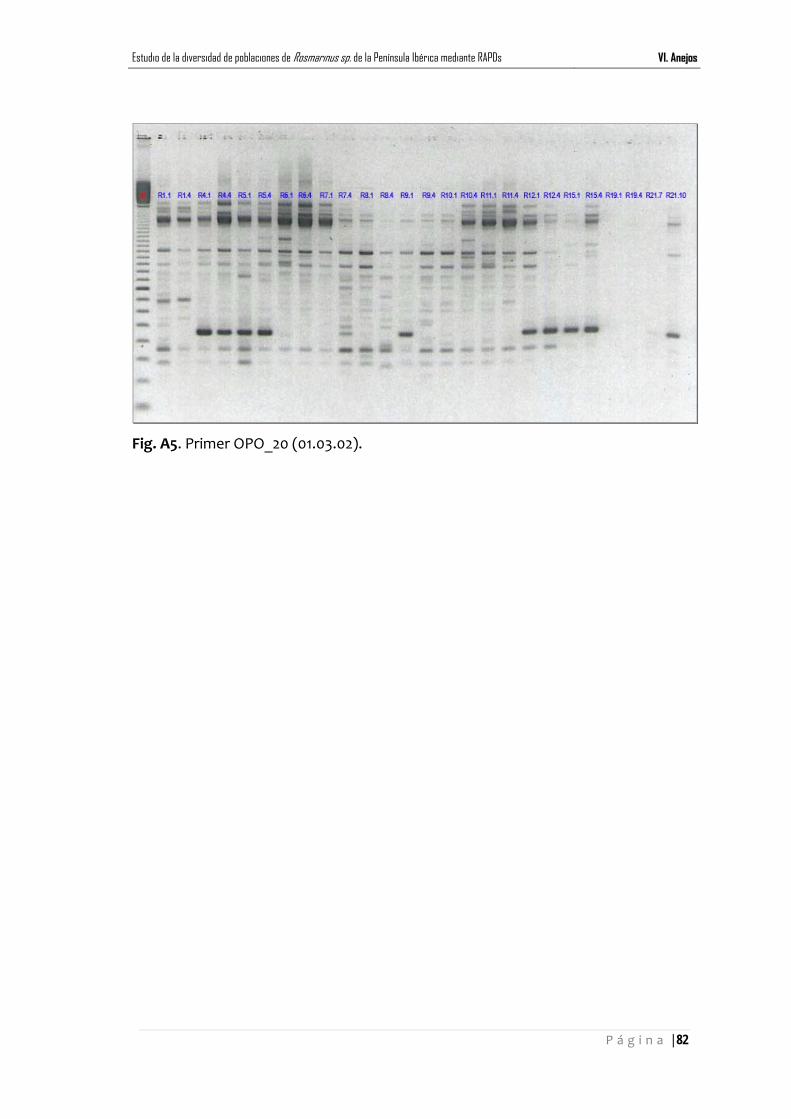
Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 82
Fig. A5. Primer OPO_20 (01.03.02).

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 83
Anejo III. Tablas de los productos de amplificación y de sus análisis.
Tabla A1. Matriz de datos – RAPDs (NTSYSpc 2.2). Muestras eliminadas R7.4 y R19.1.
51 marcadores Q. T. R1.1 R1.4 R4.1 R4.4 R5.1 R5.4 R6.1 R6.4 R7.1 R8.1 R8.4 R9.1 R9.4 R10.1 R10.4 R11.1 R11.4 R12.1 R12.4 R15.1 R15.4 R19.4 R21.7 R21.10
OPF 13 07 1.1 1.4 4.1 4.4 5.1 5.4 6.1 6.4 7.1 8.1 8.4 9.1 9.4 10.1 10.4 11.1 11.4 12.1 12.4 15.1 15.4 19.4 21.7 21.10 OPF 13_1800 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPF 13_1700 P 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 0 0 OPF 13_1500 P 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPF 13_1300 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPF 13_1200 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPF 13_950 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPF 13_650 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
OPO 07 14 1.1 1.4 4.1 4.4 5.1 5.4 6.1 6.4 7.1 8.1 8.4 9.1 9.4 10.1 10.4 11.1 11.4 12.1 12.4 15.1 15.4 19.4 21.7 21.10 OPO 07_1300 P 0 0 1 1 1 1 1 1 0 0 1 1 1 0 1 1 1 0 0 0 0 0 1 1 OPO 07_1200 P 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 0 1 1 0 0 OPO 07_1100 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 07_1000 P 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 07_950 P 1 0 1 0 1 1 1 1 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 0 OPO 07_900 P 0 0 0 1 0 1 1 1 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 OPO 07_850 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 07_800 P 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 OPO 07_700 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 07_675 P 0 0 1 0 1 0 1 1 0 0 0 1 0 0 0 0 0 1 1 1 1 0 1 1 OPO 07_550 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 07_500 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 07_325 P 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
continúa …

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 84
Tabla A1. Matriz de datos – RAPDs (NTSYSpc 2.2). Muestras eliminadas R7.4 y R19.1.
…continuación
OPO 10 09 1.1 1.4 4.1 4.4 5.1 5.4 6.1 6.4 7.1 8.1 8.4 9.1 9.4 10.1 10.4 11.1 11.4 12.1 12.4 15.1 15.4 19.4 21.7 21.10 OPO 10_2300 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 10_1400 P 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 OPO 10_1200 P 1 0 0 0 1 1 0 1 1 0 0 1 1 0 0 1 1 0 0 0 0 0 0 0 OPO 10_950 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 10_900 P 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 OPO 10_850 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 10_775 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 10_650 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 10_600 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
OPO 15 07 1.1 1.4 4.1 4.4 5.1 5.4 6.1 6.4 7.1 8.1 8.4 9.1 9.4 10.1 10.4 11.1 11.4 12.1 12.4 15.1 15.4 19.4 21.7 21.10 OPO 15_2000 P 1 1 1 1 1 1 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 15_1800 P 1 1 1 1 1 1 0 0 1 0 0 0 0 1 1 1 1 1 1 1 1 0 1 1 OPO 15_1400 P 0 0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 OPO 15_1350 P 1 1 1 1 1 1 1 1 1 0 1 0 0 1 1 1 1 1 1 1 1 1 0 0 OPO 15_1000 M 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 OPO 15_950 P 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 OPO 15_800 M º 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
continúa …

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 85
Tabla A1. Matriz de datos – RAPDs (NTSYSpc 2.2). Muestras eliminadas R7.4 y R19.1.
…continuación
OPO 20 15 1.1 1.4 4.1 4.4 5.1 5.4 6.1 6.4 7.1 8.1 8.4 9.1 9.4 10.1 10.4 11.1 11.4 12.1 12.4 15.1 15.4 19.4 21.7 21.10 OPO 20_2000 P 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1
OPO 20_1650 P 0 1 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0
OPO 20_1600 P 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0
OPO 20_1450 P 1 1 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 0 0 1 1 0 1 1
OPO 20_1300 P 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0
OPO 20_1250 P 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 1 0 1 0 1 1 1 1
OPO 20_1200 P 1 1 1 1 1 1 1 1 1 1 0 1 1 0 1 0 0 1 1 1 1 1 1 1
OPO 20_1150 P 0 1 0 0 0 0 1 0 0 0 1 0 0 1 0 1 1 1 0 1 0 1 0 0
OPO 20_1125 P 1 0 0 0 0 1 0 1 1 1 0 1 1 1 1 0 0 0 0 0 0 0 0 0
OPO 20_900 P 0 0 1 1 1 1 0 1 0 1 1 0 0 0 1 1 1 0 0 0 0 0 0 0
OPO 20_700 P 1 1 0 0 0 0 0 0 0 1 0 0 1 0 1 1 1 1 1 1 1 1 1 1
OPO 20_600 P 0 0 0 0 0 0 0 1 1 1 1 1 0 1 1 1 1 1 0 0 0 0 0 0
OPO 20_550 P 0 0 0 0 0 0 1 1 1 1 1 0 1 0 1 0 0 0 0 0 0 1 0 0
OPO 20_500 P 1 0 0 0 0 0 1 1 1 1 1 0 0 0 1 0 0 0 0 0 0 0 1 1
OPO 20_450 P 0 0 1 1 1 1 1 0 1 1 0 1 1 1 0 0 0 1 1 1 1 0 1 1
* Q = cantidad de bandas generadas por marcador T = tipo de banda M = monomófica P = polimórfica.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 86
Tabla A2. Matriz de similitud de JACCARD (NTSYSpc 2.2).
R1.1 R1.4 R4.1 R4.4 R5.1 R5.4 R6.1 R6.4 R7.1 R8.1 R8.4 R9.1 R9.4 R10.1 R10.4 R11.1 R11.4 R12.1 R12.4 R15.1 R15.4 R19.4 R21.7 R21.10
R1.1 1,000 R1.4 0,838 1,000 R4.1 0,675 0,711 1,000 R4.4 0,744 0,784 0,806 1,000 R5.1 0,756 0,707 0,816 0,842 1,000 R5.4 0,825 0,732 0,750 0,868 0,829 1,000 R6.1 0,698 0,690 0,667 0,732 0,705 0,727 1,000 R6.4 0,721 0,600 0,651 0,714 0,727 0,750 0,833 1,000 R7.1 0,842 0,744 0,675 0,789 0,756 0,780 0,698 0,762 1,000 R8.1 0,769 0,718 0,650 0,763 0,690 0,714 0,674 0,738 0,865 1,000 R8.4 0,643 0,718 0,650 0,718 0,651 0,636 0,756 0,780 0,725 0,744 1,000 R9.1 0,725 0,634 0,692 0,718 0,775 0,756 0,674 0,738 0,769 0,744 0,619 1,000 R9.4 0,744 0,650 0,625 0,692 0,707 0,732 0,651 0,674 0,744 0,763 0,595 0,681 1,000 R10.1 0,763 0,806 0,684 0,806 0,725 0,750 0,667 0,651 0,861 0,784 0,737 0,737 0,667 1,000 R10.4 0,829 0,738 0,674 0,738 0,750 0,773 0,696 0,756 0,786 0,805 0,721 0,682 0,698 0,714 1,000 R11.1 0,738 0,732 0,667 0,775 0,829 0,767 0,652 0,711 0,698 0,674 0,714 0,674 0,651 0,750 0,773 1,000 R11.4 0,714 0,750 0,643 0,795 0,805 0,744 0,630 0,689 0,714 0,690 0,732 0,651 0,628 0,769 0,750 0,974 1,000 R12.1 0,750 0,838 0,718 0,838 0,756 0,738 0,738 0,682 0,795 0,769 0,683 0,725 0,659 0,861 0,705 0,780 0,800 1,000 R12.4 0,750 0,789 0,718 0,838 0,800 0,738 0,738 0,682 0,750 0,725 0,605 0,683 0,659 0,763 0,705 0,738 0,756 0,892 1,000 R15.1 0,769 0,861 0,737 0,811 0,732 0,756 0,756 0,622 0,725 0,700 0,619 0,659 0,634 0,784 0,682 0,714 0,732 0,917 0,865 1,000 R15.4 0,750 0,789 0,718 0,789 0,756 0,738 0,738 0,644 0,707 0,683 0,568 0,643 0,619 0,718 0,705 0,698 0,714 0,842 0,944 0,917 1,000 R19.4 0,700 0,784 0,585 0,737 0,667 0,651 0,732 0,674 0,700 0,718 0,675 0,595 0,650 0,711 0,698 0,732 0,750 0,789 0,838 0,763 0,789 1,000 R21.7 0,750 0,744 0,718 0,790 0,756 0,738 0,738 0,644 0,707 0,725 0,605 0,683 0,659 0,675 0,744 0,698 0,714 0,795 0,842 0,865 0,892 0,700 1,000 R21.10 0,725 0,718 0,692 0,763 0,732 0,714 0,714 0,622 0,683 0,700 0,581 0,659 0,634 0,650 0,721 0,674 0,690 0,769 0,816 0,838 0,865 0,675 0,971 1,000

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 87
Tabla A3. Matriz de similitud de DICE (NTSYSpc 2.2).
R1.1 R1.4 R4.1 R4.4 R5.1 R5.4 R6.1 R6.4 R7.1 R8.1 R8.4 R9.1 R9.4 R10.1 R10.4 R11.1 R11.4 R12.1 R12.4 R15.1 R15.4 R19.4 R21.7 R21.10
R1.1 1,000 R1.4 0,912 1,000 R4.1 0,806 0,831 1,000 R4.4 0,853 0,879 0,892 1,000 R5.1 0,861 0,829 0,899 0,914 1,000 R5.4 0,904 0,845 0,857 0,930 0,907 1,000 R6.1 0,822 0,817 0,800 0,845 0,827 0,842 1,000 R6.4 0,838 0,750 0,789 0,833 0,842 0,857 0,909 1,000 R7.1 0,914 0,853 0,806 0,882 0,861 0,877 0,822 0,865 1,000 R8.1 0,870 0,836 0,788 0,866 0,817 0,833 0,806 0,849 0,928 1,000 R8.4 0,783 0,836 0,788 0,836 0,789 0,778 0,861 0,877 0,841 0,853 1,000 R9.1 0,841 0,776 0,818 0,836 0,873 0,861 0,806 0,849 0,870 0,853 0,765 1,000 R9.4 0,853 0,788 0,769 0,818 0,829 0,845 0,789 0,806 0,853 0,866 0,746 0,925 1,000 R10.1 0,866 0,892 0,813 0,892 0,841 0,857 0,800 0,789 0,925 0,879 0,848 0,848 0,800 1,000 R10.4 0,907 0,849 0,806 0,849 0,857 0,872 0,821 0,861 0,880 0,892 0,838 0,811 0,822 0,833 1,000 R11.1 0,849 0,845 0,800 0,873 0,907 0,868 0,789 0,831 0,822 0,806 0,833 0,806 0,789 0,857 0,872 1,000 R11.4 0,833 0,857 0,783 0,886 0,892 0,853 0,773 0,816 0,833 0,817 0,845 0,789 0,771 0,870 0,857 0,987 1,000 R12.1 0,857 0,912 0,836 0,912 0,861 0,849 0,849 0,811 0,886 0,870 0,812 0,841 0,794 0,925 0,827 0,877 0,889 1,000 R12.4 0,857 0,882 0,836 0,912 0,889 0,849 0,849 0,811 0,857 0,841 0,754 0,812 0,794 0,866 0,827 0,849 0,861 0,943 1,000 R15.1 0,870 0,925 0,848 0,896 0,845 0,861 0,861 0,767 0,841 0,824 0,765 0,794 0,776 0,879 0,811 0,833 0,845 0,957 0,928 1,000 R15.4 0,857 0,882 0,836 0,882 0,861 0,849 0,849 0,784 0,829 0,812 0,725 0,783 0,765 0,836 0,827 0,822 0,833 0,914 0,971 0,957 1,000 R19.4 0,824 0,879 0,738 0,848 0,800 0,789 0,845 0,806 0,824 0,836 0,806 0,746 0,788 0,831 0,822 0,845 0,857 0,882 0,912 0,866 0,882 1,000 R21.7 0,857 0,853 0,836 0,822 0,861 0,849 0,849 0,784 0,829 0,841 0,754 0,812 0,794 0,806 0,853 0,822 0,833 0,886 0,914 0,928 0,943 0,824 1,000 R21.10 0,841 0,836 0,818 0,866 0,845 0,833 0,833 0,767 0,812 0,824 0,735 0,794 0,776 0,788 0,838 0,806 0,817 0,870 0,899 0,912 0,928 0,806 0,986 1,000

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 88
Tabla A4. Matriz de distancias geográficas (Km.).
R01 R04 R05 R06 R07 R08 R09 R10 R11 R12 R15 R19 R21
R01 0,00
R04 521,60 0,00
R05 326,00 215,16 0,00
R06 228,20 299,92 104,32 0,00
R07 260,80 365,12 254,28 169,52 0,00
R08 469,44 299,92 332,52 312,96 208,64 0,00
R09 723,72 247,76 449,88 508,56 508,56 339,04 0,00
R10 808,48 404,24 580,28 612,88 560,72 358,60 182,56 0,00
R11 743,28 345,56 515,08 547,68 495,52 293,40 143,44 65,200 0,00
R12 606,36 123,88 280,36 378,16 475,96 423,80 286,88 462,92 410,76 0,00
R15 632,44 378,16 371,64 469,44 625,92 658,52 586,80 762,84 710,68 299,92 0,00
R19 45,640 521,60 312,96 221,68 286,88 489,00 730,24 821,52 756,32 593,32 599,840 0,00
R21 534,64 743,28 567,24 638,96 717,20 886,72 991,04 1134,48 1069,28 730,24 521,60 489,00 0,00
Tabla A5. Matriz de distancias ambientales conjunta.
R01 R04 R05 R06 R07 R08 R09 R10 R11 R12 R15 R19 R21
R01 00,00
R04 543,45 00,00
R05 540,00 21,45 00,00
R06 996,11 470,16 481,00 00,00
R07 386,37 160,10 155,44 622,19 00,00
R08 521,92 86,24 65,15 526,48 145,95 00,00
R09 535,40 61,49 80,58 462,81 163,43 139,20 00,00
R10 636,47 202,19 192,92 513,91 298,54 188,71 256,50 00,00
R11 458,10 92,63 98,45 542,40 86,65 127,18 80,00 265,70 00,00
R12 1449,84 1027,98 1015,77 965,02 1133,44 993,12 1080,68 883,36 1111,33 00,00
R15 394,97 324,85 307,77 776,25 234,22 253,46 363,52 325,53 300,91 1056,70 00,00
R19 310,00 274,30 278,75 693,36 148,93 288,63 245,67 430,10 182,08 1280,68 340,15 00,00
R21 462,93 764,48 749,55 1223,49 638,54 698,80 793,26 749,87 719,21 1337,38 447,33 668,43 00,00

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 89
Tabla A6. Matriz de distancias ambientales – Altitud.
R01 R04 R05 R06 R07 R08 R09 R10 R11 R12 R15 R19 R21
R01 00,00 R04 543,45 00,00 R05 540,00 21,45 00,00 R06 996,11 470,16 481,00 00,00 R07 386,37 160,10 155,44 622,19 00,00 R08 521,92 86,24 65,15 526,48 145,95 00,00 R09 535,40 61,49 80,58 462,81 163,43 139,20 00,00 R10 636,47 202,19 192,92 513,91 298,54 188,71 256,50 00,00 R11 458,10 92,63 98,45 542,40 86,65 127,18 80,00 265,70 00,00 R12 1449,84 1027,98 1015,77 965,02 1133,44 993,12 1080,68 883,36 1111,33 00,00 R15 394,97 324,85 307,77 776,25 234,22 253,46 363,52 325,53 300,91 1056,70 00,00 R19 310,00 274,30 278,75 693,36 148,93 288,63 245,67 430,10 182,08 1280,68 340,15 00,00 R21 462,93 764,48 749,55 1223,49 638,54 698,80 793,26 749,87 719,21 1337,38 447,33 668,43 00,00
Tabla A7. Matriz de distancias ambientales – Temperatura.
R01 R04 R05 R06 R07 R08 R09 R10 R11 R12 R15 R19 R21
R01 00,00 R04 543,45 00,00 R05 540,00 21,45 00,00 R06 996,11 470,16 481,00 00,00 R07 386,37 160,10 155,44 622,19 00,00 R08 521,92 86,24 65,15 526,48 145,95 00,00 R09 535,40 61,49 80,58 462,81 163,43 139,20 00,00 R10 636,47 202,19 192,92 513,91 298,54 188,71 256,50 00,00 R11 458,10 92,63 98,45 542,40 86,65 127,18 80,00 265,70 00,00 R12 1449,84 1027,98 1015,77 965,02 1133,44 993,12 1080,68 883,36 1111,33 00,00 R15 394,97 324,85 307,77 776,25 234,22 253,46 363,52 325,53 300,91 1056,70 00,00 R19 310,00 274,30 278,75 693,36 148,93 288,63 245,67 430,10 182,08 1280,68 340,15 00,00 R21 462,93 764,48 749,55 1223,49 638,54 698,80 793,26 749,87 719,21 1337,38 447,33 668,43 00,00
Tabla A8. Matriz de distancias ambientales ‐ Precipitación.
R01 R04 R05 R06 R07 R08 R09 R10 R11 R12 R15 R19 R21
R01 00,00 R04 543,45 00,00 R05 540,00 21,45 00,00 R06 996,11 470,16 481,00 00,00 R07 386,37 160,10 155,44 622,19 00,00 R08 521,92 86,24 65,15 526,48 145,95 00,00 R09 535,40 61,49 80,58 462,81 163,43 139,20 00,00 R10 636,47 202,19 192,92 513,91 298,54 188,71 256,50 00,00 R11 458,10 92,63 98,45 542,40 86,65 127,18 80,00 265,70 00,00 R12 1449,84 1027,98 1015,77 965,02 1133,44 993,12 1080,68 883,36 1111,33 00,00 R15 394,97 324,85 307,77 776,25 234,22 253,46 363,52 325,53 300,91 1056,70 00,00 R19 310,00 274,30 278,75 693,36 148,93 288,63 245,67 430,10 182,08 1280,68 340,15 00,00R21 462,93 764,48 749,55 1223,49 638,54 698,80 793,26 749,87 719,21 1337,38 447,33 668,43 00,00

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 90
Anejo IV. Fórmulas utilizadas.
Fórmulas de los coeficientes de similitud de JACCARD y DICE.
ijaS
a b c=
+ + Jaccard (1908)
Donde:
a: nº de bandas comunes
b: nº de bandas sólo presentes en “i”
c: nº de bandas sólo presentes en “j”
22ij
aSa b c
=+ +
Dice (1945)
Donde:
a: nº de bandas comunes
b: nº de bandas sólo presentes en “i”
c: nº de bandas sólo presentes en “j”
En ambos coeficientes (Jaccard y Dice) lo que se viene a expresar es qué
porcentaje tienen en común las muestras. Así, en el numerador figura el número de
bandas comunes entre dos muestras comparadas, mientras que en el denominador
se recoge el número de bandas diferentes entre las dos muestras, el número de
bandas presentes en una muestra y no en la segunda, y viceversa. La diferencia
entre uno y otro, es que en el coeficiente de Dice se da un mayor peso a las bandas
comunes, por lo que los valores obtenidos a través de dicho coeficiente son un poco
mayores a los obtenidos con Jaccard.

Estudio de la diversidad de poblaciones de Rosmarinus sp. de la Península Ibérica mediante RAPDs VVII.. AAnneejjooss
P á g i n a | 91
Fórmula del cálculo de distancias genéticas según Nei (1978).
lnA A A A
X YXYD G G G⎛ ⎞
= − ⎜ ⎟⎝ ⎠ Nei (1978)
donde GX y GY son las medias de (2 nx Jx – 1)/(2 nx – 1) y (2 ny Jy – 1)/(2 ny – 1) de los r
loci estudiados, respectivamente, y GXY = JXY.





























