Embed Size (px)
DESCRIPTION
Эволюция регуляторных систем у бактерий. М.С.Гельфанд Институт проблем передачи информации им. А.А.Харкевича РАН 40 лет журналу «Молекулярная биология» 25.4.2007. От эволюции генов и белков к эволюции геномов, метаболических и регуляторных систем. ~ 1000 бактериальных геномов - PowerPoint PPT Presentation
Citation preview

Эволюция регуляторных систем у бактерий
М.С.Гельфанд
Институт проблем передачи информации им. А.А.Харкевича РАН
40 лет журналу «Молекулярная биология» 25.4.2007

От эволюции генов и белков к эволюции геномов, метаболических и
регуляторных систем
~1000 бактериальных геномов
Проблема: как их анализировать?
Возможности:• сравнительная геномика
– Функциональная аннотация– Метаболическая реконструкция– Регуляторные системы
• Новый уровень изучения эволюции

Сравнительная геномика регуляторных систем: задачи
• Микроэволюция: динамика отдельных регуляторных сайтов
• Ко-эволюция факторов транскрипции и узнаваемых ими мотивов в ДНК– Изменение специфичности регуляторов
• Макроэволюция:– Расширение, сокращение, деление и слияние
регулонов– Перекоммутация регуляторных каскадов– Смена регуляторных систем

Методы: филогенетический футпринтинг
Регуляторные сайты выглядят как консервативные островки в выравниваниях 5’-областей
Пример: гены nrd у Yersinia+Erwinia,сайты связывания FNR, DnaA, NrdR
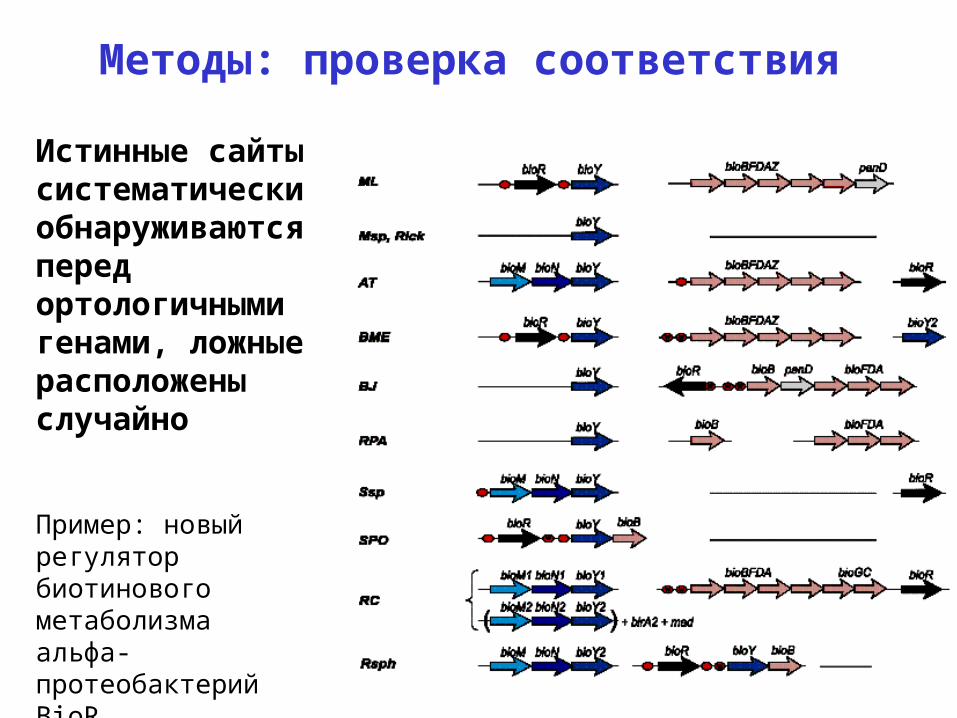
Методы: проверка соответствия
Истинные сайты систематически обнаруживаются перед ортологичными генами, ложные расположены случайно
Пример: новый регулятор биотинового метаболизма альфа-протеобактерий BioR

Рождение и исчезновение сайтов – очень динамический процесс
Сайты связывания NadR перед геном pncB

Расширение регулона: как FruR стал CRA
icdA
aceA
aceB
aceEF
pckA
ppsApykF
adhE
gpmApgk
tpiA
gapApfkAfbp
FructosefruKfruBA
eda
eddepd
Glucose
ptsHI-crr
Mannose
manXYZ
mtlDmtlAMannitol
гамма-протеобактерии
проекция экспериментально известных сайтов E.coli на родственные геномы: что окажется консервативно?

Общий предок Enterobacteriales
icdA
aceA
aceB
aceEF
pckA
ppsApykF
adhE
gpmApgk
tpiA
gapApfkAfbp
FructosefruKfruBA
eda
eddepd
Glucose
ptsHI-crr
Mannose
manXYZ
mtlDmtlAMannitol
гамма-протеобактерииEnterobacteriales

Общий предок Escherichia and Salmonella
icdA
aceA
aceB
aceEF
pckA
ppsApykF
adhE
gpmApgk
tpiA
gapApfkAfbp
FructosefruKfruBA
eda
eddepd
Glucose
ptsHI-crr
Mannose
manXYZ
mtlDmtlAMannitol
гамма-протеобактерииEnterobacterialesE. coli и Salmonella spp.

Исчезновение регуляторов – тоже динамический процесс
потеря RbsR у Y. pestis (ABC-транспортер тоже утерян)
Start codon of rbsD
RbsR binding site

Потеря регулятора и слияние регулонов
По-видимому, laci-X существовал у общего предка (Klebsiella – внешний геном)
Yersinia и Klebsiella: два регулона, GalR и Laci-X
Erwinia: один регулон, GalR

Изменение регулятора
Замена: вторжение регулятора из другого подсемейства (горизонтальный перенос),
с другим мотивом связывания
стрептококки, утилизация мальтозы/мальтодекстрина

Дупликация регулятора (и изменение мотива?) – альфа-проетобактерия, утилизация мальтозы/трегалозы

Регуляция гомеостаза железаЖелезо:• необходимый кофактор (лимитирующий во многих
экологических нишах)• опасно в больших концентрациях
Кишечная палочка (E. coli): FUR, регуляция транскрипции в ответ на железо:
• синтез сидерофоров• транспорт (сидерофоры, гем, Fe2+, Fe3+)• хранение (ферритины и т.п.)• железо-зависимые ферменты• синтез гема• синтез железо-серных кластеров
Похоже в сенной палочке (Bacillus subtilis)

Регуляция гомеостаза железа у альфа- протеобактерий
Эксперимент:
FUR/MUR: Bradyrhizobium, Rhizobium and Sinorhizobium
RirA (семейство Rrf2): Rhizobium and Sinorhizobium
Irr (семейство FUR): Bradyrhizobium, Rhizobium and Brucella
RirA IrrFeS heme
RirA
degraded
FurFe
Fur
Iron uptake systems
Siderophoreuptake
Fe / Feuptake Transcription
factors
2+ 3+
Iron storage ferritins
FeS synthesis
Heme synthesis
Iron-requiring enzymes
[iron cofactor]
IscR
Irr
[- Fe] [+Fe]
[+Fe][- Fe]
[+Fe][ Fe]-
FeS
FeS statusof cell

Распре-деление
факторов по
геномам

Семейство FUR/MUR/Irr
MBNC03003593RB2654 19538
AGR C 620
RL mur
Nwi 0013RPA0450
BJ furROS217 18337
Jann 1799SPO2477
STM1w01000993MED193 22541
OB2597 02997SKA53 03101Rsph03000505ISM 15430
GOX0771ZM01411
Saro02001148Sala 1452
ELI1325OA2633 10204
PB2503 04877CC0057
Rrub02001143Amb1009Amb4460
SM murMBNC03003179
BQ fur2BMEI0375
Mesorhizobium sp. BNC1 (I)
Sinorhizobium meliloti
Bartonella quintana
Rhodopseudomonas palustris
Bradyrhizobium japonicum
Caulobacter crescentus
Zmomonas mobilisy
Rhodobacter sphaeroides
Silicibacter sp. TM1040Silicibacter pomeroyi
Agrobacterium tumefaciens
Rhizobium leguminosarum
Brucella melitensis
Mesorhizobium sp. BNC1 (II)
Rhodobacterales bacterium HTCC2654
Nitrobacter winogradskyiNham 0990 Nitrobacter hamburgensis X14
Jannaschia sp. CC51Roseovarius sp.217
Roseobacter sp. MED193Oceanicola batsensis
HTCC2597
Loktanella vestfoldensis SKA53
Roseovarius nubinhibens ISM
Gluconobacter oxydans
Erythrobacter litoralis
Novosphingobium aromaticivoransSphinopyxis alaskensis RB2256
Oceanicaulis alexandrii HTCC2633
Rhodospirillum rubrum
Parvularcula bermudensis HTCC2503
Magnetospirillum magneticum (I)
EE36 12413Sulfitobacter sp. EE-36
ECOLIPSEAE
NEIMAHELPY
BACSUHelicobacter pylori: sp|O25671
Bacillus subtilis: P54574sp|
Neisseria meningitidis: sp|P0A0S7Pseudomonas aeruginosa: sp|Q03456
Escherichia coli: P0A9A9sp|
Fur
Magnetospirillum magneticum (II)
RHE_CH00378Rhizobium etli
PU1002 04436Pelagibacter ubique HTCC1002
MU
R
Irr
FU
R
E.coli
B.subtilis
регуляция генов транспортеров Mn
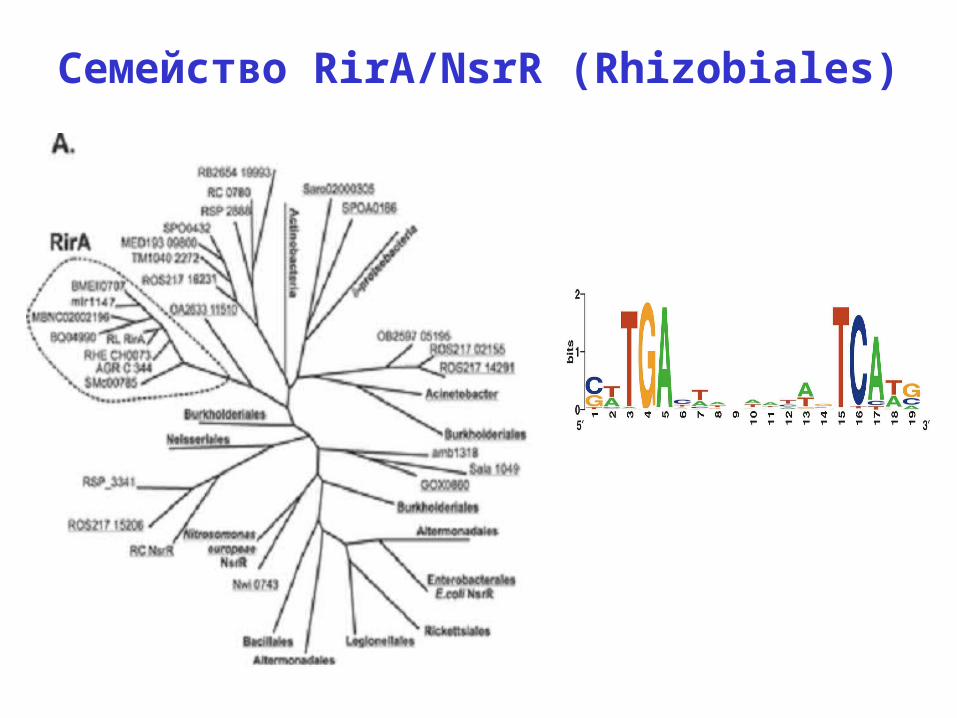
Семейство RirA/NsrR (Rhizobiales)

Семейство IscR

Регуляция генов в функциональных
подсистемах
Rhizobiales
Bradyrhizobiaceae
Rhodobacteriales
The Zoo (likely ancestral state)
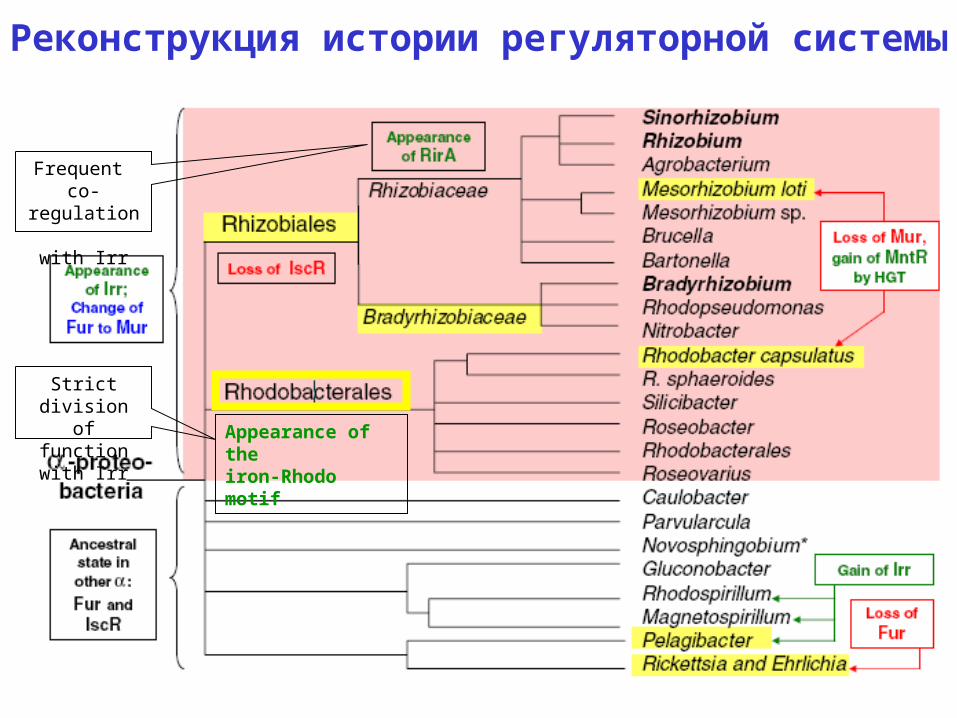
Реконструкция истории регуляторной системы
Appearance of theiron-Rhodo motif
Frequent co-regulation
with Irr
Strict division of function
with Irr

Все мотивы и одна очень привлекательная
гипотеза:
• Кросс-узнавание мотивов FUR и IscR в общем предке.
• Когда FUR стал MUR, а IscR пропал у Rhizobiales, возникший RirA (из семейства Rrf2 с совсем другим общим консенсусным мотивом) «завладел» этими сайтами.
• Iron-Rhodo мотив узнаетс IscR: поддается прямой проверке
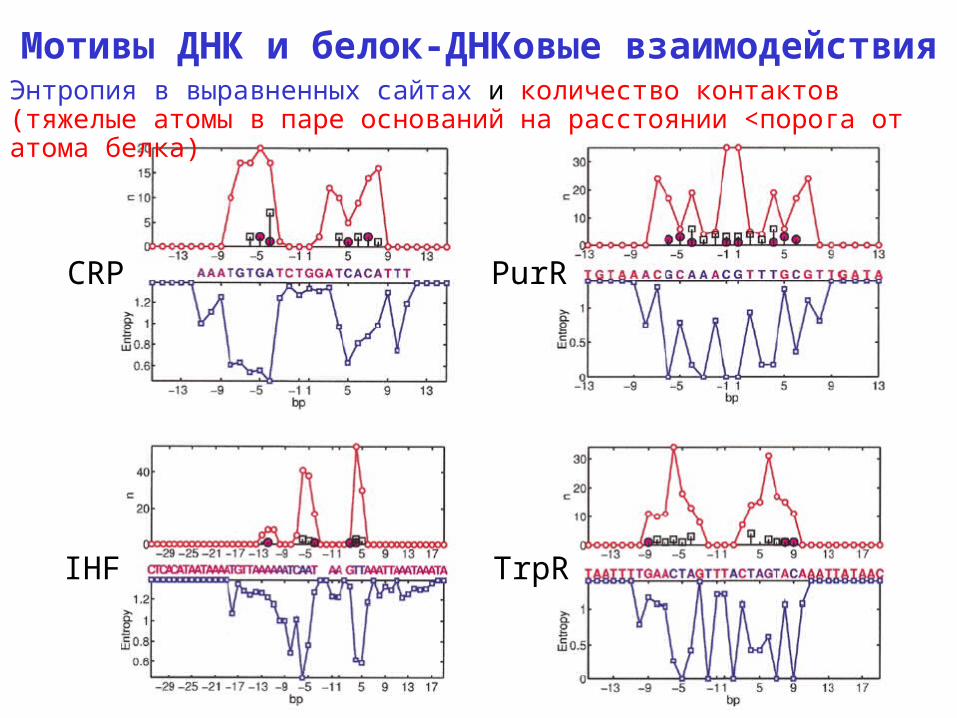
Мотивы ДНК и белок-ДНКовые взаимодействия
CRP PurR
IHF TrpR
Энтропия в выравненных сайтах и количество контактов (тяжелые атомы в паре оснований на расстоянии <порога от атома белка)

Детерминанты специфичности семейства LacI
Обучение: 459 белков, средняя длина: 338 аа, 85 групп специфичности
10 остатков в контакте с эффектором
6 остатков – межсубъединичные контакты
7 остатков в контакте с оператором
7 в области контакта с эффектором (5Ǻ<dmin<10Ǻ)
5 в зоне межсубъединичного контакта (5Ǻ<dmin<10Ǻ)
6 в зоне контакта с оператором (5Ǻ<dmin<10Ǻ)
– 44 SDPs
LacI from E.coli

Семейство факторов транскрипции CRP/FNR
FNR
HcpR
CooA
Gam ma
Desulfovibrio
Desulfovibrio
TGTCGGCnnGCCGACA
TTGTgAnnnnnnTcACAA
TTGTGAnnnnnnTCACAA
TTGATnnnnATCAA

Корреляция между нуклеотидами в мотиве и остатками в белке
• CooA в Desulfovibrio spp.• CRP в гамма-протобактериях• HcpR в Desulfovibrio spp. • FNR в гамма-протобактериях
DD COOA ALTTEQLSLHMGATRQTVSTLLNNLVRDV COOA ELTMEQLAGLVGTTRQTASTLLNDMIREC CRP KITRQEIGQIVGCSRETVGRILKMLEDYP CRP KXTRQEIGQIVGCSRETVGRILKMLEDVC CRP KITRQEIGQIVGCSRETVGRILKMLEEDD HCPR DVSKSLLAGVLGTARETLSRALAKLVEDV HCPR DVTKGLLAGLLGTARETLSRCLSRMVEEC FNR TMTRGDIGNYLGLTVETISRLLGRFQKYP FNR TMTRGDIGNYLGLTVETISRLLGRFQKVC FNR TMTRGDIGNYLGLTVETISRLLGRFQK
TGTCGGCnnGCCGACA
TTGTgAnnnnnnTcACAA
TTGTGAnnnnnnTCACAA
TTGATnnnnATCAA
Контактирующие: REnnnRTG: 1й аргининGA: глютамат и 2й аргинин

Эта кор-реляция сохра-
няется и в других членах
семейст-ва

Открытые проблемы
• Моделирование эволюции регуляторных систем– каталог элементарных событий– механизмы
• как рождаются сайты?• дупликации и горизонтальные переносы факторов и регулируемых оперонов
– оценки вероятностей– общие свойства:
• распределение размера семейств факторов транскрипции в геномах• геном-специфичные массовые дупликации («взрывы») и их причины• стабильные ядра и лабильные периферии регулонов: связь с метабооической
картой
• Коэволюция факторов транскрипции и мотивов ДНК: – Как эволюционируют мотивы? Что является движущей силой – мутации
фактора?– код ДНК-белкового узнавания – свой для кадого семейства? Предсказние
мотива по белку.
Все это на фоне трудоемкого сбора (литература, массовые эксперименты) и порождения (сравнительная геномика) данных, их неполноты и неточности

Кто это делал:
• А.А.Миронов (алгоритмы и программы)• А.Б.Рахманинова (структуры белков)• Дмитрий Родионов (BioR, NrdR, железо)• Ольга Лайкова (LacI, сахара)• Дмитрий Равчеев (FruR)• Ольга Калинина (специфичность/LacI)
• Leonid Mirny, MIT (ДНК-белковые контакты, специфичность)
• Andy Johnston, University of East Anglia (железо)
• Howard Hughes Medical Institute • РФФИ• РАН, программа “Молекулярная и клеточная биология”• INTAS





























