Embed Size (px)
Citation preview

1
TUMOR SUPPRESSOR NF2 BLOCKS CELLULAR MIGRATION BY INHIBITING
ECTODOMAIN CLEAVAGE OF CD44
Monika Hartmann1*, Liseth M Parra1,2*, Anne Ruschel1, Sandra Schubert1, Yong Li1§, Helen
Morrison1, Andreas Herrlich2**and Peter Herrlich1**
1 Leibniz Institute for Age Research, Fritz Lipmann Institute, Jena, Germany
2 Harvard Institutes of Medicine, Renal Division, Brigham and Women’s Hospital, Harvard
Medical School, Boston, USA
§ current address: Anhui University School of Life Sciences, Hefei, China
corresponding author: Peter Herrlich, M.D., PhD. Professor of Molecular Biology, Leibniz Institute for Age Research, Beutenbergstr. 11, 07745 Jena, Germany Phone: 493641656257 e-mail: [email protected] * shared first authorship ** shared senior authorship conflict of interest statement: the authors do not disclose a conflict of interest.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

2
Abstract
Ectodomain cleavage (shedding) of transmembrane proteins by metalloproteases (MMPs)
generates numerous essential signaling molecules, but its regulation is not totally understood.
CD44, a cleaved transmembrane glycoprotein, exerts anti-proliferative or tumor-promoting
functions, but whether proteolysis is required for this is not certain. CD44-mediated contact
inhibition and cellular proliferation are regulated by counteracting CD44 C-terminal
interacting proteins, the tumor suppressor protein merlin (NF2) and ERM proteins (ezrin,
radixin, moesin). We show here that activation or overexpression of constitutively active
merlin, or downregulation of ERMs inhibited TPA-induced (as well as serum, HGF or PDGF)
CD44 cleavage by the metalloprotease ADAM10, while overexpressed ERM proteins
promoted cleavage. Merlin- and ERM-modulated Ras or Rac activity was not required for this
function. However, latrunculin (an actin-disrupting toxin) or an ezrin mutant unable to link to
actin inhibited CD44 cleavage, identifying a cytoskeletal C-terminal link as essential for
induced CD44 cleavage. Cellular migration, an important tumor property, depended on CD44
and its cleavage and was inhibited by merlin. These data reveal a novel function of merlin and
suggest that CD44 cleavage products play a tumor-promoting role. Neuregulin, an epidermal
growth factor ligand released by ADAM17 from its pro-form NRG1, is predominantly
involved in regulating cellular differentiation. In contrast to CD44, release of neuregulin from
its pro-form was not regulated by merlin or ERM proteins. Disruption of the actin
cytoskeleton however also inhibited NRG1 cleavage. This current study presents one of the
first examples of substrate-selective cleavage regulation.
Implications
Investigating transmembrane protein cleavage and their regulatory pathways has provided
new molecular insight into their important role in cancer formation and possible treatment.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

3
Introduction
The ubiquitously expressed surface glycoprotein CD44 is involved in a number of cellular
functions not all of which are understood in molecular terms. Its role in cell cycle control has
obtained most attention and it seems mechanistically best understood. Depending on
extracellular ligands, intercellular partner proteins and/or the inclusion of alternatively spliced
exon sequences, CD44 can act as a tumor suppressor and mediate contact inhibition, or can
act alternatively as a tumor promoter and metastasis inducer. Binding of high-molecular
weight hyaluronan causes cell cycle arrest and tumor suppression (1-3). To achieve cell cycle
arrest, the tumor suppressor protein merlin (neurofibromatosis type 2; Nf2) is recruited to the
cytoplasmic tail of CD44, a location from which it inhibits Ras and Rac dependent signaling
(1, 4). On the other hand CD44 can counteract the tumor suppressor p53. In order for p53 to
act as a tumor suppressor, CD44 expression needs to be downregulated (5). In addition, CD44
acts as co-receptor for receptor tyrosine kinases, the most prominent example being c-Met
which depends on the presence of a CD44 splice variant comprising exon v6 (6). This second
function of CD44 promotes tumor growth and metastasis formation (7-9).
The tumor suppressor protein merlin (Nf2), like CD44, is ubiquitously expressed in
mammals. Mice carrying one mutated nf2 allele are at risk of developing several types of
tumors (10). Merlin is kept inactive in proliferating cells, but is activated by
dephosphorylation at two serines upon cell-cell contact and/or hyaluronan binding ((2); and
unpublished data). Dephosphorylation is regulated by a signal transduction pathway emerging
from cell-cell and/or hyaluronan-cell contact and involves all ERM proteins in addition to
merlin. Interestingly, while merlin is activated by this process, the ERM proteins are
inactivated (11-13). Several types of analyses have led to proposals on how merlin could act
as a tumor suppressor, for example by inhibiting mitogenic signaling, activating the Hippo
pathway and/or promoting the establishment of adherens junctions (reviewed by (14)).
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

4
Several years ago it was discovered that CD44 – like numerous other transmembrane
proteins – is subject to ectodomain cleavage by metalloprotease activity (15, 16), now
identified as ADAM10 (A Disintegrin And Metalloprotease 10) (17-19). This is followed by
gamma secretase dependent release of the cytoplasmic tail, which promotes the expression of
proliferation-promoting genes in the nucleus. Given the tumor suppressive role of hyaluronan-
bound CD44, ectodomain cleavage would abolish this function. We therefore investigated
how ectodomain cleavage of CD44 might be regulated.
We report here, that it is the tumor suppressor protein merlin itself that prevents
CD44 cleavage, supporting the notion that proteolytic processing of CD44 promotes tumor
growth, and the hypothesis that naturally occurring Nf2 mutants that are prone to
malignancies may fail to inhibit CD44 ectodomain cleavage and thereby its tumor promoting
role. This cleavage regulation is specific to CD44, as we show that NRG1, the pro-form of the
epidermal growth factor ligand neuregulin, an ADAM17 substrate and major regulator of
cellular differentiation, is cleaved upon stimulation, but is not regulated by merlin or ERM
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

5
Materials and Methods
Reagents: DNA oligonucleotides (Metabion GmbH); TPA, DAPT, latrunculin B and
batimastat (BB94) (Calbiochem), PD98059 (Cell Signaling Technology), Angiotensin II
(ARIAD Pharmaceuticals, Inc), 4-OH-tamoxifen, EGF, FGF (Sigma), HGF and PDGF (R&D
Systems). CK-548 (Tocris).
Antibodies: Anti-FLAG (M2 and SIG1-25) (Sigma); phosphospecific antibodies against
Ezrin (Thr567)/Radixin (Thr564)/Moesin (Thr558) and p44/42 MAPK (Thr202/Tyr204) (Cell
Signaling Technology); ADAM10 (735-749) (Calbiochem or R&D Systems); Ezrin (3C12)
(Thermo Fisher Scientific), Ezrin (C-15), Moesin (C-15), Radixin (C-15), c-Myc (9E10), HA
(F-7), ,NF2 (C-19; C-18; B-12), ERK 1 (K-23), NRG antibody (C-20) and Actin (I-19) (Santa
Cruz Biotechnology Inc.); CD44 (IM7, Becton Dickinson) and α-tubulin (Abcam). Antibodies
directed against human CD44: for an N-terminal epitope H-CAM F4 (Santa Cruz), for a C-
terminal epitope ARP61023-P050 (Aviva Systems Biology). Rabbit polyclonal antibody
recognizing the N-terminus of APP was provided by Christoph Kaether (FLI, Jena,
Germany). All secondary antibodies (Dako).
Plasmids: The sequence encoding the standard isoform of rat CD44 was subcloned into the
NotI/XbaI sites of pFLAG-myc-CMV-21. CD44 mutants were generated by site-directed
mutagenesis. The primers are listed in Table 1. Plasmids encoding mouse pro-neuregulin-1
(NRG1) have been described (20). FLAG-tagged NRG1 was subcloned into EcoRI and XhoI
restriction sites of pFLAG-myc-CMV-21 (Sigma). Sequences encoding tagged CD44 were
subcloned from pFLAG-myc-CMV-21 vector into the EcoRI site of the pCDH-CMV-
MCS.Bsd viral vector. All constructs were verified by sequencing. For stable downregulation
of nf2 we used the viral vector pLV-H1-GIPZ (provided by Cui Yan and Helen Morrison,
Jena, Germany).
siRNA sequences: siRNA SMARTpools, cocktail of four siRNAs and control “Non-
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

6
TARGETplusTM Pool” were from Thermo Scientific Dharmacon. Other siRNA
oligonucleotides were from Applied Biosystems/Ambion. The sequences of the siRNA
oligonucleotides are listed in Table 2. For down-regulation of ERM proteins a mixture of
oligonucleotides targeting ezrin, radixin and moesin was used.
Definition of Growth Conditions: Low cell density (exponential growth) = density recorded
at 36 hrs after seeding of 1.25 x 104 cells per cm2 (NIH3T3) or 3.65 x 104 cells per cm2
(RPM-MC). High cell density (confluent growth condition) = density recorded at 36 hrs after
seeding of 3.5 × 104 cells per cm2 (NIH3T3 cells) or 10 × 104 cells per cm2 (RPM-MC cells).
Inhibition of Cleavage Conditions: Metalloprotease activity was blocked with 5 μM
batimastat (BB94; Calbiochem) 15 min prior to TPA stimulation. γ-secretase activity was
blocked by 5 μM DAPT (Sigma) or by 10 μM Compound E (Enzo).
Cell migration assays – Scratch wound assay: We isolated mouse embryonic fibroblasts
from mice with cd44flox/flox (CD44flox; GT(Rosa)26-Cre (B6/129) and immortalized these by
downregulation of p19ARF. CD44 gene deletion was achieved by treatment with tamoxifen.
Scratch-wound assays were performed in triplicates in 6-well plates at high cell density
(1.5x105 cells/well). 24 hours after seeding, cells were serum-starved for another 24 hours.
Scratches were introduced with a 200 µl pipette tip and cultures were re-supplied with serum-
containing medium. Where indicated, cleavage was inhibited by adding 5 µM batimastat.
Scratches were imaged at 10, 24 and 36 hours after scratching. Wound areas were quantified
using Photoshop and image J software.
Statistical Analysis: Intensity of immunoblot bands was quantified using Image J and Image
Lab® (Biorad, Hercules, CA) software. All values on histograms are reported as mean ±
standard deviation (SD). P-value < 0.05 (student’s T-test) was considered significant.
Cell lines, Transfections, TCA-DOC precipitations, Co-IP, Generation of cell lysates and
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

7
analysis: See Supplement.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

8
Results
Increased cell density inhibits ectodomain cleavage of CD44
As has been reported previously, CD44 is subject to the metalloprotease ADAM10
dependent ectodomain cleavage and subsequent release of the cytoplasmic CD44 C-terminus
by gamma secretase (17-19). Aiming at understanding the regulation of CD44 ectodomain
cleavage, we introduced expression constructs encoding doubly-tagged (N- and C- terminal
tags) CD44 proteins into either NIH3T3, CD44-/- MEFs, RPM-MC or MDA-MB-231 cells,
and examined their proteolytic processing. The N-terminus of CD44 carried a FLAG tag, the
C-terminus a c-myc motif. RPM-MC human melanoma cells, as well as CD44-/- MEFs, do not
express endogenous CD44, which simplified detection of the transfected molecule, permitted
to introduce CD44 mutants, and allowed to analyze signaling pathways independently of
endogenous CD44. In NIH3T3 and MDA-MD-231 cells we also investigated the cleavage of
endogenous CD44. NRG1 was similarly tagged by N-terminal FLAG and C-terminal myc tag.
CD44 and NRG1 cleavage was induced by TPA, a phorbol ester that mimics diacylglycerol
and activates most protein kinase C (PKC) isoforms. Cleavage could also be induced by
serum factors, by HGF and PDGF, and, if cells carried the appropriate G protein-coupled
receptor, by angiotensin II. In most experiments gamma secretase activity was blocked using
the gamma secretase inhibitor, DAPT, in order to quantitate only the products of the first
processing event, ADAM dependent ectodomain cleavage. Omission of DAPT did not
significantly alter our principal results on regulation, but caused further processing of the C-
terminal ADAM dependent cleavage product (Supplemental Fig. S1). It is important to note
that in our analysis of CD44 and NRG1 cleavage we ensured that we focused exclusively on
processing of the substrates after their proper insertion into the plasma membrane. To
ascertain this, we carried out experiments very shortly after cell surface biotinylation,
showing that biotinylated ectodomains, solCD44E and neuregulin, are indeed released into
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

9
the supernatant (Supplemental Fig. S2), suggesting cleavage of substrates already present on
the cell surface. However, the effect of transport regulation on cleavage has not been
examined here.
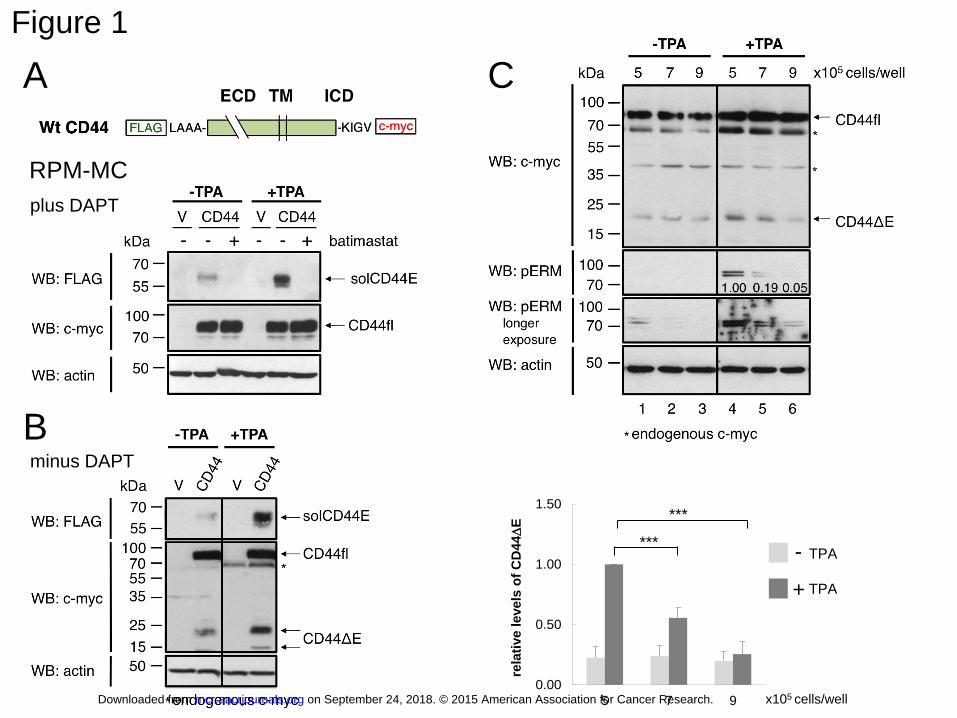
Figure 1A demonstrates the basic cleavage reaction: Transfected full-length CD44
molecule (CD44fl) is detected by antibodies directed against the C-terminal myc tag, and the
cleaved-off soluble ectodomain (solCD44E) is recognized by anti-FLAG antibodies. As
expected, vector-transfected control RPM-MC cells showed no staining (V). In the absence of
the cleavage stimulus (-TPA), there was relatively little spontaneous release of solCD44E, but
cleavage was strongly enhanced after treatment with TPA (Figure 1A). Both spontaneous and
induced cleavages were blocked by batimastat, an inhibitor of ADAM protease activity.
DAPT had no major effect on the result of ADAM dependent cleavage regulation (Figure 1B,
see also control experiments in Supplemental Fig. S1).
Because CD44 regulates contact inhibition of cells, we wondered whether its
cleavage regulation was dependent on cell density. While spontaneous cleavage was reduced
only slightly, TPA induced cleavage was markedly diminished by increasing cell density from
5- to 9 x 105 cells per well of a 6-well plate (for quantitation see the column diagram of three
independent experiments, Figure 1C). From previous reports, it has been known that high cell
density causes dephosphorylation of both merlin and its counterplayers, the ezrin-moesin-
radixin (ERM) proteins, by the same protein phosphatase-1 isoenzyme (2).
Dephosphorylation of ERM proteins deactivates them, whereas it activates merlin. In turn,
phosphorylation of both ERM proteins and merlin depends on protein kinase activity during
the exponential growth of cells (21-23). While little phospho-ERM could be detected in the
absence of a stimulus (left three lanes in Figure 1C), ERM proteins were strongly
phosphorylated upon TPA treatment of cells (compare lanes 1 and 4, Figure 1C). As
expected, phosphorylation of ERM proteins declined with increasing cell density, coinciding
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

10
with decreased CD44 cleavage (lanes 5 and 6, Figure 1C). Merlin dephosphorylation follows
exactly that of the ERM proteins (not shown, and Supplemental Fig. S3A).
The tumor suppressor protein merlin inhibits ectodomain cleavage of CD44
To investigate whether dephosphorylation of ERM proteins and reduced CD44
ectodomain cleavage with increasing cell density was not simply coincidental, we tested the
effect of overexpression of merlin or of ERM mutants (see below) on CD44 induced cleavage
in RPM-MC cells. To this end, we first examined the effect of a constitutively active merlin
mutant (NF2-S518A), which does not require dephosphorylation. These experiments were
done under low cell density conditions at which endogenous merlin is phosphorylated and
inactive, and activated ERM proteins drive proliferation. TPA induced solCD44E release in
the absence of transfected merlin (Figure 2A; compare lanes 1 and 4, WB: FLAG). However,
expression of the singly mutated active merlin (NF2-S518A) was sufficient to inhibit
solCD44E release in RPM-MC cells (Figure 2A; compare lanes 2 and 5, WB: FLAG; also
see column diagram of quantification of three independent experiments), while the phospho-
mimicking mutant S518D had no effect (see also similar data obtained in NIH3T3 cells,
Supplementary Fig. S4). Cleavage of the ADAM17 substrate neuregulin (NRG1) in the
same cells was not inhibited (Figure 2B). Quantitations have been compiled as column
diagram in Figure 2A/B. We can therefore conclude that the tumor suppressor merlin
specifically inhibits CD44 cleavage by ADAM10 and that merlin does not interfere with a
common signaling pathway addressing ADAM cleavage in general.
We had previously shown that contact inhibition requires the binding of
dephosphorylated active merlin to the C-terminus of CD44 via a membrane-proximal basic
amino acid sequence, known as the KR motif (Supplemental Fig. S3B; initially described as
an ezrin binding site; (1)). We therefore followed the idea that cleavage regulation of CD44
might also require merlin binding to the C-terminus. To investigate this, we compared
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

11
cleavage of wild type CD44 and the CD44 mutant with a complete deletion of the
intracellular domain, ICD (CD44ΔICD). First, we made sure, by confocal microscopy and
immunostaining, that the deletion mutant was properly inserted into the plasma membrane
(not shown). Second, we confirmed that it was still processed by ADAM10 (as is CD44 wt).
Release of solCD44E from CD44ΔICD was blocked by siRNA dependent downregulation of
ADAM10 (Figure 3A, compare lanes 1 and 2) or by addition of the ADAM inhibitor
batimastat (Figure 3A, lanes 1 vs 3).
We then compared the action of merlin mutants on cleavage of full-length CD44
(WtCD44; first six lanes, Figure 3B) and of CD44 with deletion of the ICD (CD44ΔICD;
lanes 7 to 12, Figure 3B; see also the upper column diagram and the loading scheme in
Supplemental Fig.S7) as well as on cleavage of the non-cleaved mutant CD44-KR-Mt (lower
column diagram, Figure 3B). Indeed, absence of the entire CD44 intracellular domain as well
as mutation of the KR motif prevented the inhibitory effect of merlin on cleavage. The
uppermost first panel in Fig. 3B, shows the level of endogenous (inactive) merlin and of the
transfected merlin mutants. The third panel shows detection of the released solCD44E, and
the fourth panel the full-length molecules (both detected with anti-FLAG antibodies). As
already shown in Figure 2, constitutively active NF2-S518A, but not NF2-S518D, inhibited
full-length CD44 cleavage (lanes 5 and 6, Figure 3B). Interestingly, total absence of the
cytoplasmic tail of CD44 caused significant spontaneous ectodomain cleavage (lane 7, Figure
3B), suggesting that the ICD suppressed spontaneous cleavage and was required for TPA
induced regulation. However, TPA was still able to increase cleavage to some extent
(compare lane 7 and 10, Figure 3B; also see discussion). Both spontaneous and induced
solCD44E release from the CD44ΔICD mutant was resistant to inhibition by constitutively
active merlin NF2-S518A (compare lanes 8 and 11, Figure 3B). Note that NF2-S518D had
little to no effect on this release (lanes 9 and 12, Figure 3B). A quantitation of three
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

12
independent experiments with CD44ΔICD is shown in the upper column diagram. Strong
cleavage coincided with the appearance of several solCD44E bands (Figure 3A and B). We
assume that this might be due to the presence of several ADAM10 cleavage sites on CD44.
The CD44KR-Mt mutant was barely inducible by TPA and not inhibited by merlin (see lower
column diagram in Figure 3B), as one would expect because the mutation destroys the merlin
binding site. For comparison, another ADAM10 substrate, the amyloid precursor protein,
APP, is shown in the second panel of Figure 3B. TPA induced cleavage of both, CD44 and of
APP (compare lanes 1 and 4, Figure 3B), however, cleavage of the ADAM17 substrate APP
was not affected by merlin (compare lanes 5, 6 and 11, 12, Figure 3B). Also cleavage of the
ADAM10 substrate c-Met was merlin-resistant (data not shown). Consistent with this neither
APP nor c-Met does, to our knowledge, carry a KR motif in its C-terminus.
Our results using the CD44 ICD deletion and the KR-Mt mutant suggest that merlin
interaction with the ICD of CD44 is necessary for its cleavage regulating function.
Importantly, CD44 was co-precipitated with merlin only if its ICD was intact (Figure 3C).
Mutation of the KR motif abolished merlin interaction (Supplemental Fig. S3B), identical to
the ICD deletion (Figure 3C). Similarly, the absence of merlin regulation on neuregulin
release (Figure 2B) and the resistance of APP, c-Met and NRG1 cleavage to regulation by
merlin further strengthens the idea of substrate specific regulation of ectodomain cleavage and
confirms our assertion that merlin exerts a direct specific effect on CD44 and its cleavage,
rather than interfering with a cleavage regulatory signaling pathway common to these three
substrates.
The action of ERM proteins and the mechanism of merlin dependent inhibition of CD44
ectodomain cleavage
CD44 C-terminally bound merlin mediates contact inhibition predominantly by
blocking Ras and Rac activity. This is counteracted by ERM proteins, which promote Ras and
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

13
Rac activation. A well-studied mechanism has shown that Ras is activated by ERM proteins
interacting with both Ras and the guanine-nucleotide exchange factor and activator of Ras,
son of sevenless (SOS) (24). Because active merlin inhibits CD44 ectodomain cleavage, we
were wondering whether its Ras inhibitory action was required for this effect and in turn,
whether cleavage required ERM protein-dependent Ras activation.
Upon downregulation of all three ERM proteins using siRNA, TPA induced cleavage
was indeed significantly reduced (detected by the reduced release of solCD44E [N-terminal
FLAG tag] and of the membrane-bound cleavage product CD44ΔE [C-terminal c-myc tag];
Figure 4A). Neuregulin release, however, was not affected by downregulation of ERMs
(Figure 4B). If the ERM-induced Ras activation were required for CD44 cleavage, a
constitutively active Ras pathway should bypass ERM protein requirement and cause
constitutive cleavage. We thus turned on the Ras pathway by transfecting an ERM-
independent dominant-active SOS (DA-SOS, tagged with HA to visualize its expression;
(25); called SOS-F in (26)) and compared its effect on cleavage in control cells (empty vector,
V), cells expressing CD44 full length (WtCD44) and cells carrying CD44 with a mutant KR
domain (CD44 KR-MT), the motif required for interaction of CD44 with merlin and ERM
proteins (see above) (1). In control cells lacking CD44 expression (empty vector control lanes
1 to 4 in Figure 4C; see also the experimental set-up table in Supplemental Fig.S7), DA-
SOS caused phosphorylation of a downstream target of Ras, Erk, to the same degree, as did
TPA stimulation (Figure 4C, compare lane 2 and 3). DA-SOS in the presence of TPA further
enhanced phospho-Erk (Figure 4C, lane 4), which might be explained, although does not
prove, by their different mechanisms of action: DA-SOS activates Ras, whereas TPA acts
downstream of Ras, adding to the activation of the pathway. DA-SOS did neither enhance
spontaneous (Figure 4C, lane 6) nor TPA induced (Figure 4C, lane 8) release of WtCD44
ectodomain (solCD44E). Cleavage of mutant CD44 KR-Mt barely responded to TPA
treatment (Figure 4C, compare lanes 9 and 11) and this block could not be overcome by DA-
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

14
SOS (lanes 10 and 12). The column diagram shows a quantitation of three independent
experiments. Finally, an inhibitor of MEK, a downstream effector of Ras, blocked Erk
phosphorylation, but did not affect CD44 cleavage (Figure 5A). We therefore conclude that
CD44 cleavage is independent of an active Ras-Erk signaling pathway.
Given that the SOS-Ras activating function of ERM proteins was not required for
CD44 ectodomain cleavage, we explored other putative options. To this end we exploited
transfections with ezrin mutants. Overexpression of ezrin mutants should compete out
endogenous ERM proteins. This is possible because in our cultured cells all three ERM
proteins are redundant with respect to Ras activation and this redundancy can be overcome by
overexpression of an active mutant of one of the ERMs. We therefore tested whether
overexpressed ezrin mutants could reveal the function of ERM proteins that was required for
CD44 cleavage. Figure 5B shows the effect of overexpressed ezrin wt or ezrin mutants. In the
absence of TPA stimulation we detected no CD44 cleavage irrespective of transfected ezrin
constructs (as detected by cleavage product CD44ΔE; lanes 1-5, Figure 5B). TPA induced
cleavage is shown in lanes 6-10. Neither wild type ezrin (Figure 5B, compare lanes 6 and 7)
nor a phospho-mimicking mutant T567D (Figure 5B, lane 8) affected CD44 cleavage,
suggesting that endogenous ERM proteins are so abundant that additional transfected ezrin
constructs made no difference. Somewhat surprisingly, however, the “inactive” ezrin T567A
mutant inhibited CD44 cleavage (Figure 5B, lane 9) suggesting that it competed with the
endogenous ERM proteins for a cleavage regulatory component.
A possible lead towards this putative component and the cleavage regulatory
function of ezrin was generated by the effect of the ezrin mutant R579A which cannot interact
with F-actin (27). This mutant inhibited CD44 cleavage (Figure 5B, lane 10), highlighting the
possible need for an actin link to achieve CD44 cleavage. Interestingly, in respect to its
inability to bind to actin, ezrin R579A mimics the cleavage inhibitory merlin whose C-
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

15
terminus also cannot interact with F-actin (28, 29). These observations propose that disrupting
F-actin would exert a similar inhibitory effect. We tested this assumption by adding an
increasing amount of latrunculin, an inhibitor known to block actin polymerization (30).
Short-term treatment (30 min) of the cells with 0.75 to 1.0 μg/ml of latrunculin indeed
inhibited CD44 cleavage (Figure 5C). This result made us wonder whether a link to the actin
cytoskeleton were specific for the ERM dependent substrate CD44. This was not the case:
NRG1 cleavage depended also on an intact actin cytoskeleton (Figure 5D).
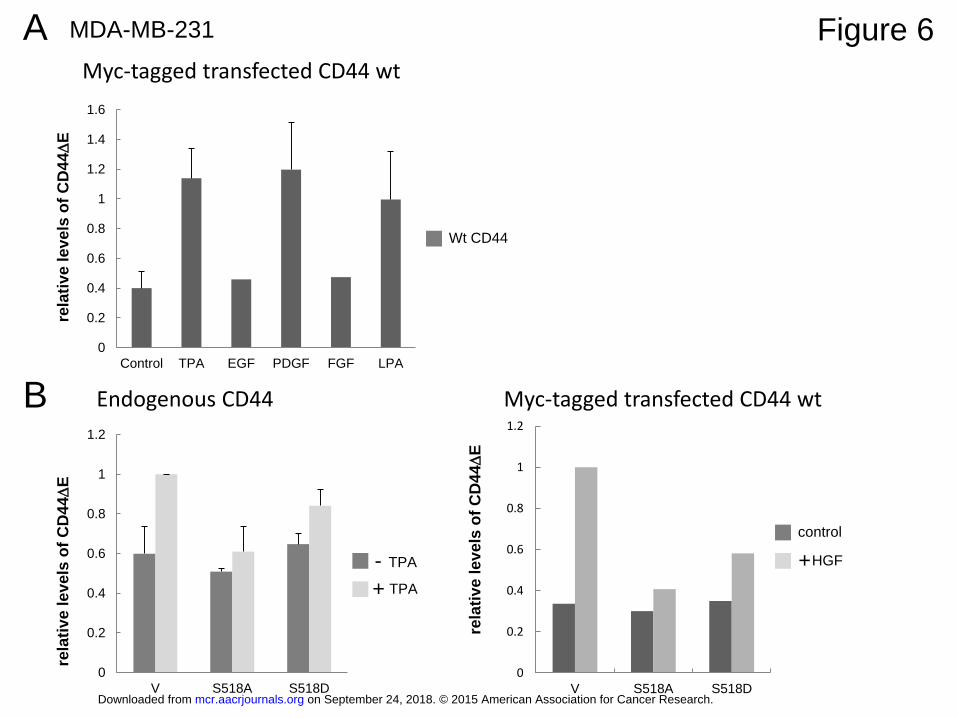
Physiologic stimuli induce ectodomain cleavage in different normal and cancer cells
TPA mimics a signaling process that is activated by numerous physiologic stimuli.
Therefore such stimuli should also result in ectodomain cleavage. This is indeed the case: in
HEK293T cells that that stably overexpress the angiotensin receptor (HEKNE in Figure 5D),
angiotensin II strongly induced neuregulin release. In the triple-negative breast cancer cell
line MDA-MB-231 we were able to induce either endogenous or transfected CD44 cleavage
by serum (see below in Figure 7B and C), HGF, PDGF, LPA and, moderately, by FGF and
EGF (Figure 6A and B). MDA-MB-231 cells were also responsive to TPA. TPA or HGF
induced cleavage of endogenous CD44 or overexpressed CD44 was inhibited by expression
of constitutively active merlin (S518A) (Figure 6B). CD44 cleavage was also induced in
MEF cells (see below: Figure 7A). We conclude that the pathway regulating CD44 cleavage
is addressed by many extracellular stimuli.
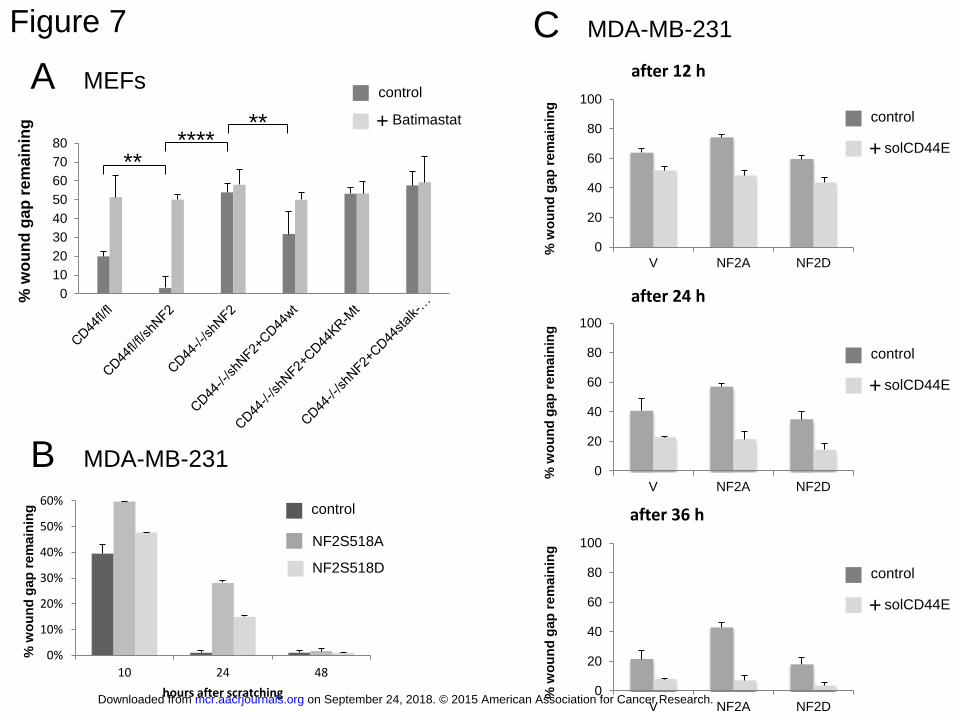
CD44 cleavage is required for cellular migration
According to our data, CD44 cleavage is a property of proliferating cells. This
property as well as its blockade by the tumor suppressor NF2 (contact inhibition) should also
be relevant for the control of cancer cells. We therefore explored whether CD44 cleavage was
needed for cancer-relevant cellular phenotypes related to their proliferative capabilities:
mobility and migration. To this end we subjected MEF and MDA-MB-231 cells grown in
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

16
confluent monolayers to scratch assays and measured their ability to close the scratch wound.
After plating and scratching cells were supplied with fetal calf serum that contains factors like
LPA that we have shown to induce ectodomain cleavage. Using MEFs from mice with floxed
CD44 alleles, we could compare cells expressing CD44 with CD44 null cells (after CRE
dependent excision) and we also could substitute the cells with CD44 mutants. Supplemental
Fig.6 shows photographic examples of the original scratch assay. The left picture of each
condition tested shows the scratch at time 0 and the original cell number as indicated in the
square. The right picture of each condition tested shows the same scratch wound after 24
hours and the percentage of the wound area still open. Quantitation of five experiments has
been compiled in Figure 7A. Treatment with the ADAM inhibitor Batimastat strongly
inhibited wound closure of migration competent cells. CD44 plus cells (CD44fl/fl) repaired the
scratch wound efficiently (remaining wound area 21 % after 24 hours, Supplemental Fig.S6
panel 1 and Figure 7A). Downregulation of merlin (NF2) by stably integrated shRNA
enhanced the migration (0% wound remaining, Supplemental Fig.S6 panel 2, less than 2 %
in the quantitation of Figure 7A). There was almost no wound closure by CD44 null cells
(CD44-/- after CRE induction, see supplemental Material and Methods, and Supplemental
Fig. S5). In these cells NF2 was also downregulated (Supplemental Fig.S6 panel 3 and
Figure 7A). Although CD44-/-:nf2+/+ was not obtained, the data suggest that NF2 did not
address a pathway other than CD44. However, re-introducing CD44 wt partly re-established
migration (remaining wound area 30 % after 24 hours in the absence of merlin,
Supplemental Fig.S6 panel 4 and Figure 7A). This partial rescue might be explained by the
fact that CD44wt cDNA overexpression does not generate certain splice variants of CD44 that
would be expressed under physiological conditions but are missing in CD44-/- cells. Most
importantly, stable transfection of the non-cleavable mutant CD44-KR-Mt as well as of a
CD44 mutant that lacks the stalk region including the ADAM cleavage site, could not rescue
the wound healing deficiency of CD44 null MEFs (Supplemental Fig. S6 panel 5 and 6 and
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

17
Figure 7A). These results clearly indicate that CD44 cleavage is required for a cancer
relevant phenotype, cellular migration, and that this function is negatively controlled by
merlin.
MDA-MB-231 cells closed the scratch wound faster than MEFs. At 24 hours after
scratching, wounds were totally closed (note that wound area normalized to the 0 time point;
Figure 7B). Expression of active merlin (NF2S518A) inhibited wound healing. Inactive
merlin (NF2S518D) had less effect (Figure 7B and C). Interestingly, spontaneous migration
was enhanced and the inhibition by merlin rescued by expression of solCD44E (Figure 7C)
suggesting a role of CD44 ectodomain cleavage in migration.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

18
Discussion
The activity state of both merlin and ERM proteins is controlled by signaling pathways that
address specific protein kinases (e.g. PAK) and phosphatases (e.g. PP1). A hyaluronan-CD44
dependent signaling pathway (or cell-cell contact) favors dephosphorylation of ERM proteins
and merlin, deactivating ERM proteins and activating merlin, thus establishing tumor
suppression capabilities. Growth factor stimulation, in turn, activates ERM proteins and
inactivates merlin, favoring growth and tumor development. In this context, our results on
CD44 cleavage inhibition by merlin permit the following conclusions:
• When activated by a cell density dependent signaling pathway, merlin prevents CD44
ectodomain cleavage, coinciding with and preserving contact inhibition of cells.
• Regulation by merlin represents an example of substrate-selective cleavage regulation.
NRG1 and APP, in our cells studied, are subject to substrate-specific regulation different
from CD44.
• CD44 cleavage appears to serve a tumor-promoting process by enhancing
proliferation/migration of cells, including cancer cells.
TPA induced CD44 shedding from the cell surface is regulated by ERM proteins and
merlin, in contrast to other ADAM substrates, such as NRG1, c-Met and APP. Interestingly,
in neural cells, purinergic P2X7 receptor induced APP cleavage required ERM proteins (31).
Rather than direct interaction of the APP ICD with ERM proteins, this event required
downstream signaling induced by ERM proteins. However, ERM activation was not always
associated with the induction of APP shedding. Nerve growth factor (NGF) and
benzoylbenzoyl ATP triggered ERM phosphorylation, but only the latter led to APP shedding
(31). Apparently, signaling pathways can diverge after ERM activation. Similar to our results
with CD44, TPA induced L-selectin shedding in lymphocytes was regulated by direct
interaction of ERM proteins with a proximal basic amino acid region in the cytoplasmic
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

19
domain of L-selectin (32). The important conclusion from these reports is that substrates are
specifically selected for cleavage.
Does this mean we can disregard other forms of regulation? Not at all. Regulation of
ADAM activity has been widely studied (33-36). As example, we have seen ADAM
activation by TPA in one of our experiments, where despite constitutive cleavage of the ICD-
less CD44, TPA was still able to increase cleavage to some extent (compare lane 7 and 10,
Figure 3B). However, CD44 is, in addition, specifically selected for cleavage on the substrate
level.
Importantly, regulation of ectodomain cleavage by a tumor suppressor protein has
not been observed previously. It suggests that CD44 cleavage serves a tumor-promoting
function. This notion is further strengthened by the documentation that CD44 cleavage
participates substantially in the regulation of cellular migration. Migration is inhibited by
merlin and can be rescued by soluble CD44 ectodomain (Figure 7). A role of CD44 in
cellular migration has been observed previously (15, 17, 37, 38, 43-46). For instance, CD44
promoted invasion of glial cell tumor cells by its ability to bind hyaluronan (37). CD44
mediated migration of pancreatic cancer cells in conjunction with MT1-MMP (38). It has
been suggested that cleavage is involved by the fact that inhibition of metalloproteases
reduced migration (15); conversely co-expression of metalloprotease with CD44 enhanced
migration (46). We have described elsewhere that CD44 homodimerization is a pre-condition
for ectodomain cleavage (Hartmann et al., 2015, submitted). Expectedly, ligation by anti-
CD44 antibodies induced metalloprotease dependent CD44 ectodomain release and migration
in the aggressive tumor cell line U251MG (17, 45). Ligation-induced cleavage and migration
was counteracted by expression of a dominant-negative Rac mutant (17, 45), suggesting that
Rac signaling preceded cleavage. Our results are compatible with these data. Merlin action on
cleavage did however need to interfere with Ras/Rac (below). Expression of soluble CD44
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

20
ectodomain, however, prevented cleavage (Hartmann et al., 2015, submitted) which seems to
contradict our finding that expression of soluble CD44 enhanced migration. We have no
straight forward explanation for this observation. Further analyses will be needed. The
influence on migration may depend on the substratum the cells are placed on, on the time
course of adhesion/de-adhesion and on adhesion molecules other than CD44.
The cleavage-promoting role of ERM proteins matches their overexpression in
tumors (39-41). However, we ruled out that ERM-induced cleavage regulation requires their
activation of the Ras and Rac pathway by demonstrating that an ERM-independent
constitutive activation of Ras did not influence CD44 cleavage. Ezrin mutants defective in
activating guanine nucleotide exchange factors, e.g. R579A; T567A, prevented induced
cleavage (Figure 5B; and not shown). Based on the dominant-negative effect of the ezrin
actin-link mutant R579A and our results using actin-disrupting latrunculin, we hypothesize
that a CD44 F-actin link plays a role in the induction of its proteolysis. We have shown
previously that short-term treatment with latrunculin causes only highly specific pathway
disruptions (42). What the F-actin link might contribute is currently speculative. Does it
support the assembly of CD44 and its protease ADAM10 in the plane of the plasma
membrane? Interestingly, neuregulin release was also sensitive to an actin poison. We do
however not know whether and how NRG1 is linked to F-actin.
Another intriguing observation has been reported: the induction of CD44 cleavage by
treating cells with hyaluronate oligosaccharides (16). Low molecular hyaluronan does not
cause activation of merlin, while high-molecular weight hyaluronan does (1, 3). We assume
that the oligosaccharides activate cleavage-inducing signaling pathways in a CD44-
independent manner (47). The oligosaccharides induce metalloprotease expression in the
absence of CD44 (47). Also the hyaluronate receptor TLR-4 (48) may cause the phenotype
observed.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

21
How does merlin interfere with CD44 cleavage? If ERM-induced Ras activity is not
needed for cleavage regulation, one could assume that merlin also does not act through its
Ras-inhibiting function. Our merlin mutant data indeed prove this to be correct. Ras inhibition
by merlin requires that the protein is dephosphorylated at position S518 ((2); and unpublished
data). Dephosphorylation of S518 does not suffice for tumor suppression (but is followed by a
second dephosphorylation at S272 upon cell-cell contact; unpublished data). However, the
merlin mutant NF2-S518A used in our studies, mimicking the single dephosphorylation,
sufficed to inhibit CD44 cleavage. Because S272 dephosphorylation does not occur in
growing cells (unpublished data), our data indicate that full tumor suppressor activity of
merlin, and thus Ras pathway blockade, was not required for inhibition of CD44 cleavage.
How then does merlin act? Merlin does not carry a C-terminal F-actin binding domain. Thus,
by replacing ERM proteins on the CD44 C-terminus it likely disrupts the link to the actin
cytoskeleton. We consider this a plausible mechanism.
In the end, it is likely that ERM phosphorylation is needed to promote cleavage.
Structural studies of moesin showed that phosphorylation releases an inhibitory interaction of
its C- and N-terminus (43) In this context it is puzzling that inactive ezrin T567A exerts a
dominant-negative effect on cleavage. Interestingly, a corresponding inactive moesin mutant,
T558A, inhibits the formation of microvilli-like structures (44), suggesting that ezrin T567A
could still compete with a cleavage-relevant interaction partner.
We leave a few questions open that we cannot answer at this point. Only a fraction of
CD44 is subjected to induced (or spontaneous) cleavage (see also Figure 1 in ref. 42). One
does not need to demand complete cleavage because the reaction is to generate highly active
components, particularly evident in the release of growth factors. Mechanistically, partial
cleavage indicates however, that in addition to the ICD modification another condition must
be met to induce proteolysis. Possibly this condition is fulfilled if the CD44 ICD is deleted.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

22
Then no regulation is required and cleavage becomes constitutive (Figure 3B). Another open
and highly interesting question concerns the role of CD44 cleavage in vivo, particularly in
cancer. One might expect that tumors not only shed CD44 ectodomain at an increased rate
and via this mechanism generate CD44 ICD which drives the expression of proliferation
inducing genes in the nucleus, but also that cleavage products of the CD44 ectodomain might
exert additional defined roles in cancer.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

23
Acknowledgements
We are grateful for the support we received from the Leibniz Institute for Age Research and
the Jungstiftung (fellowship to MH). The administrative staff of the institute was very helpful.
Thanks are also due to our technician and lab manager Birgit Pavelka and to Christoph
Kaether who adviced us on APP and gave us reagents. A.H. was supported by NIDDK
R00DK077731, M.H. by a fellowship of the Jung Foundation, P.H. by DFGHE551.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

24
References
1. Morrison H, Sherman LS, Legg J, Banine F, Isacke C, Haipek CA, et al. The NF2 tumor suppressor gene product, merlin, mediates contact inhibition of growth through interactions with CD44. Genes & Development. 2001;15:968-80. 2. Jin H, Sperka T, Herrlich P, Morrison H. Tumorigenic transformation by CPI-17 through inhibition of a merlin phosphatase. Nature. 2006;442:576-9. 3. Tian X, Azpurua J, Hine C, Vaidya A, Myakishev-Rempel M, Ablaeva J, et al. High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature. 2013;499:346-9. 4. Morrison H, Sperka T, Manent J, Giovannini M, Ponta H, Herrlich P. Merlin/neurofibromatosis type 2 suppresses growth by inhibiting the activation of Ras and Rac. Cancer Res. 2007;67:520-7. 5. Godar S, Ince TA, Bell GW, Feldser D, Donaher JL, Bergh J, et al. Growth-inhibitory and tumor- suppressive functions of p53 depend on its repression of CD44 expression. Cell. 2008;134:62-73. 6. Orian-Rousseau V, Chen L, Sleeman JP, Herrlich P, Ponta H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes & Development. 2002;16:3074-86. 7. Todaro M, Gaggianesi M, Catalano V, Benfante A, Iovino F, Biffoni M, et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell stem cell. 2014;14:342-56. 8. Günthert U, Hofmann M, Rudy W, Reber S, Zöller M, Haussmann I, et al. A new variant of glycoprotein CD44 confers metastatic potential to rat carcinoma cells. Cell. 1991;65:13-24. 9. Matzke A, Herrlich P, Ponta H, Orian-Rousseau V. A five-amino-acid peptide blocks Met- and Ron-dependent cell migration. Cancer research. 2005;65:6105-10. 10. McClatchey AI, Saotome I, Mercer K, Crowley D, Gusella JF, Bronson RT, et al. Mice heterozygous for a mutation at the Nf2 tumor suppressor locus develop a range of highly metastatic tumors. Genes & Development. 1998;12:1121-33. 11. Fehon RG, McClatchey AI, Bretscher A. Organizing the cell cortex: the role of ERM proteins. Nature Reviews Molecular Cell Biology. 2010;11:276-87. 12. Bretscher A, Edwards K, Fehon RG. ERM proteins and merlin: integrators at the cell cortex. Nature Reviews Molecular Cell Biology. 2002;3:586-99. 13. Ivetić A, Ridley AJ. Ezrin/radixin/moesin proteins and Rho GTPase signalling in leucocytes. Immunology. 2004;112:165-76. 14. Li W, Cooper J, Karajannis MA, Giancotti FG. Merlin: a tumour suppressor with functions at the cell cortex and in the nucleus. EMBO reports. 2012;13:204-15. 15. Okamoto I, Kawano Y, Tsuiki H, Sasaki J, Nakao M, Matsumoto M, et al. CD44 cleavage induced by a membrane-associated metalloprotease plays a critical role in tumor cell migration. Oncogene. 1999;18:1435-46. 16. Sugahara KN, Murai T, Nishinakamura H, Kawashima H, Saya H, Miyasaka M. Hyaluronan oligosaccharides induce CD44 cleavage and promote cell migration in CD44-expressing tumor cells. The Journal of biological chemistry. 2003;278:32259-65. 17. Murai T, Miyazaki Y, Nishinakamura H, Sugahara KN, Miyauchi T, Sako Y, et al. Engagement of CD44 promotes Rac activation and CD44 cleavage during tumor cell migration. The Journal of biological chemistry. 2004;279:4541-50. 18. Anderegg U, Eichenberg T, Parthaune T, Haiduk C, Saalbach A, Milkova L, et al. ADAM10 is the constitutive functional sheddase of CD44 in human melanoma cells. The Journal of investigative dermatology. 2009;129:1471-82.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

25
19. Stamenkovic I, Yu Q. Shedding light on proteolytic cleavage of CD44: the responsible sheddase and functional significance of shedding. The Journal of investigative dermatology. 2009;129:1321-4. 20. Herrlich A, Klinman E, Fu J, Sadegh C, Lodish H. Ectodomain cleavage of the EGF ligands HB-EGF, neuregulin1-beta, and TGF-alpha is specifically triggered by different stimuli and involves different PKC isoenzymes. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2008;22:4281-95. 21. Shaw RJ, Paez JG, Curto M, Yaktine A, Pruitt WM, Saotome I, et al. The Nf2 tumor suppressor, merlin, functions in Rac-dependent signaling. Developmental Cell. 2001;1:63-72. 22. Kissil JL, Johnson KC, Eckman MS, Jacks T. Merlin phosphorylation by p21-activated kinase 2 and effects of phosphorylation on merlin localization. The Journal of biological chemistry. 2002;277:10394-9. 23. Xiao G-H, Beeser A, Chernoff J, Testa JR. p21-activated kinase links Rac/Cdc42 signaling to merlin. The Journal of biological chemistry. 2002;277:883-6. 24. Geißler KJ, Jung MJ, Riecken LB, Sperka T, Cui Y, Schacke S, et al. Regulation of Son of sevenless by the membrane-actin linker protein ezrin. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:20587-92. 25. Aronheim A, Engelberg D, Li N, al-Alawi N, Schlessinger J, Karin M. Membrane targeting of the nucleotide exchange factor Sos is sufficient for activating the Ras signaling pathway. Cell. 1994;78:949-61. 26. Sibilia M, Fleischmann A, Behrens A, Stingl L, Carroll J, Watt FM, et al. The EGF receptor provides an essential survival signal for SOS-dependent skin tumor development. Cell. 2000;102:211-20. 27. Saleh HS, Merkel U, Geißler KJ, Sperka T, Sechi A, Breithaupt C, et al. Properties of an ezrin mutant defective in F-actin binding. Journal of Molecular Biology. 2009;385:1015-31. 28. Xu HM, Gutmann DH. Merlin differentially associates with the microtubule and actin cytoskeleton. Journal of Neuroscience Research. 1998;51:403-15. 29. Lallemand D, Saint-Amaux AL, Giovannini M. Tumor-suppression functions of merlin are independent of its role as an organizer of the actin cytoskeleton in Schwann cells. Journal of Cell Science. 2009;122:4141-9. 30. Morton WM, Ayscough KR, McLaughlin PJ. Latrunculin alters the actin-monomer subunit interface to prevent polymerization. Nature Cell Biology. 2000;2:376-8. 31. Darmellah A, Rayah A, Auger R, Cuif M-H, Prigent M, Arpin M, et al. Ezrin/radixin/moesin are required for the purinergic P2X7 receptor (P2X7R)-dependent processing of the amyloid precursor protein. The Journal of biological chemistry. 2012;287:34583-95. 32. Ivetić A, Florey O, Deka J, Haskard DO, Ager A, Ridley AJ. Mutagenesis of the ezrin-radixin-moesin binding domain of L-selectin tail affects shedding, microvillar positioning, and leukocyte tethering. The Journal of biological chemistry. 2004;279:33263-72. 33. Díaz-Rodríguez E, Montero JC, Esparís-Ogando A, Yuste L, Pandiella A. Extracellular signal-regulated kinase phosphorylates tumor necrosis factor alpha-converting enzyme at threonine 735: a potential role in regulated shedding. Mol Biol Cell. 2002;13:2031-44. 34. Xu P, Derynck R. Direct activation of TACE-mediated ectodomain shedding by p38 MAP kinase regulates EGF receptor-dependent cell proliferation. Molecular Cell. 2010;37:551-66. 35. Le Gall SM, Maretzky T, Issuree PDA, Niu X-D, Reiss K, Saftig P, et al. ADAM17 is regulated by a rapid and reversible mechanism that controls access to its catalytic site. Journal of Cell Science. 2010;123:3913-22.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

26
36. Adrain C, Zettl M, Christova Y, Taylor N, Freeman M. Tumor necrosis factor signaling requires iRhom2 to promote trafficking and activation of TACE. Science (New York, NY). 2012;335:225-8. 37. Jiang W, Zhang Y, Kane KT, Collins MA, Simeone DM, Pasca di Magliano M, et al. CD44 Regulates Pancreatic Cancer Invasion through MT1-MMP. Molecular cancer research : MCR. 2015. 38. Kim Y, Kumar S. CD44-mediated adhesion to hyaluronic acid contributes to mechanosensing and invasive motility. Molecular cancer research : MCR. 2014;12:1416-29. 39. Wang G, Mao W, Zheng S. MicroRNA-183 regulates Ezrin expression in lung cancer cells. FEBS Letters. 2008;582:3663-8. 40. Saito S, Yamamoto H, Mukaisho K-i, Sato S, Higo T, Hattori T, et al. Mechanisms underlying cancer progression caused by ezrin overexpression in tongue squamous cell carcinoma. PloS one. 2013;8:e54881. 41. Yu Y, Khan J, Khanna C, Helman L, Meltzer PS, Merlino G. Expression profiling identifies the cytoskeletal organizer ezrin and the developmental homeoprotein Six-1 as key metastatic regulators. Nature Medicine. 2004;10:175-81. 42. Sperka T, Geißler KJ, Merkel U, Scholl I, Rubio I, Herrlich P, et al. Activation of Ras requires the ERM-dependent link of actin to the plasma membrane. PloS one. 2011;6:e27511. 43. Pearson MA, Reczek D, Bretscher A, Karplus PA. Structure of the ERM protein moesin reveals the FERM domain fold masked by an extended actin binding tail domain. Cell. 2000;101:259-70. 44. Oshiro N, Fukata Y, Kaibuchi K. Phosphorylation of moesin by rho-associated kinase (Rho-kinase) plays a crucial role in the formation of microvilli-like structures. The Journal of biological chemistry. 1998;273:34663-6. 45. Schulz A, Geißler KJ, Kumar S, Leichsenring G, Morrison H, Baader SL. Merlin inhibits neurite outgrowth in the CNS. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2010;30:10177-86. 46. Kajita M, Itoh Y, Chiba T, Mori H, Okada A, Kinoh H, et al. Membrane-type 1 matrix metalloproteinase cleaves CD44 and promotes cell migration. The Journal of Cell Biology. 2001;153:893-904. 47. Fieber C, Baumann P, Vallon R, Termeer C, Simon JC, Hofmann M, et al. Hyaluronan-oligosaccharide-induced transcription of metalloproteases. Journal of Cell Science. 2004;117:359-67. 48. Taylor KR, Trowbridge JM, Rudisill JA, Termeer CC, Simon JC, Gallo RL. Hyaluronan fragments stimulate endothelial recognition of injury through TLR4. The Journal of biological chemistry. 2004;279:17079-84.
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

27
Figure legends
Figure 1: Cell density dependent regulation of CD44 ectodomain cleavage
A: TPA induced ADAM dependent ectodomain cleavage of CD44.
The CD44-negative cell line RPM-MC was transfected with a doubly tagged expression
construct of standard isoform of wild-type CD44 (CD44s). V = vector control. All samples
were treated with γ-secretase inhibitor DAPT (5 µM). 5 µM batimastat was added as
indicated. The cells were kept in logarithmic growth condition. Full-length (CD44fl) CD44
was detected by an antibody recognizing the C-terminal myc tag. The cleaved ectodomain
(solCD44E) was detected in culture supernatants after TCA-DOC (see Supplemental Material
and Methods) precipitation by an antibody against the N-terminal FLAG tag. Treatment of the
cells with 100 ng/ml phorbol ester (TPA) induced detectable cleavage within 15 min and led
to accumulation of solCD44E in 3-4 hours. Here TPA treatment was for 3 hours. Actin served
as loading control. The cleavage is inhibited by the metalloprotease inhibitor batimastat.
B: TPA induced ADAM dependent ectodomain cleavage of CD44 in the absence of γ-
secretase inhibition (no DAPT)
C: High cell density inhibits ectodomain cleavage of CD44.
RPM-MC cells transfected with tagged CD44 as in A, were seeded into 6-well plates at
different densities, as indicated. Cells were treated with 100 ng/ml TPA for 4 hours. The
amount of cell lysates loaded on the gel was normalized to actin levels. CD44 cleavage was
detected by the release of CD44∆E (membrane-bound cleavage product lacking the
ectodomain), detected by c-Myc immunoblot in cell lysates. TPA induced generation of the
membrane-bound cleavage product (CD44ΔE) was diminished with increasing cell density.
At the same time merlin (not shown) and the ERM proteins were dephosphorylated (middle
panels). The 62 kDa band (*) likely represents endogenous c-Myc; the 40 kDa band (*) is
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

28
unspecific. Representative blots are shown. The intensity of immunoblot bands was quantified
with Image J. Histograms show mean values of relative level of cleavage ± SD from three
independent experiments (triple asterisks, p= 0.000924 for 5x105 versus 7x105 and p=
0.000211 for 5x105 versus 9x105). Levels of phospho-ERM proteins relative to that at the
density of 5x105 are indicated within the immunoblot.
Figure 2: The tumor suppressor merlin (NF2) downregulates CD44 ectodomain cleavage
A: RPM-MC cells were co-transfected with tagged CD44 (as in Figure 1) and merlin mutant
expression constructs (or vector control, “V”). The cells were kept at low cell density so that
endogenous merlin is not activated. Therefore, the vector control lanes show TPA induced
cleavage (lanes 1 and 4) similarly to Figure 1A. The action of merlin can however be
determined by introducing a mutant that mimics dephosphorylation. The dephosphorylation-
mimicking merlin mutant NFS518A (constructs as described in ref 42) inhibited CD44
cleavage (shown for the released solCD44E and the residual membrane-bound fragment
CD44ΔE; compare lanes 2 and 5). The phospho-merlin mimicking inactive mutant NFS518D
did not affect the cleavage induction (lanes 3 and 6). Because of the similar migration of
merlin and CD44, these two proteins were detected on separate gels, and respectively two
subsequent loading controls are shown.
B: A corresponding experiment as in (A) shows the result for another ADAM substrate, the
neuregulin precursor NRG1. Merlin dependent inhibition was specific for CD44, as NRG1,
was not affected.. The histograms in A´/B show mean values of relative level of cleavage ±
SD from three independent experiments (triple asterisk, p=0.000337).
Figure 3: The cleavage repression by merlin is substrate-specific and requires the
intracellular domain (ICD) of CD44
A: A complete deletion mutant of the CD44 ICD was transfected into RPM-MC cells.
Immune staining showed proper insertion into the plasma membrane (not shown). The cells
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

29
were treated with TPA for 3 hours. Cleavage (lane 1) was absent upon downregulation of
ADAM10 (lane 2; see reduced expression of the ADAM10 precursor A10P) or treatment with
batimastat (lane 3). C = control siRNA.
B: Cleavage of CD44ΔICD is resistant to merlin inhibition.
CD44 wild type or CD44 lacking the ICD, both tagged with N-terminal FLAG, were co-
transfected with plasmids encoding merlin mutants (or vector) as in Figure 2. solCD44E is
detected by anti-FLAG. In the same cells the cleavage of endogenous Alzheimer Precursor
Protein, APP, was determined by immunoblot using an ectodomain specific antibody
recognizing released solAPPE. CD44 cleavage was quantitated from three independent
experiments, shown in the column diagram (triple asterisk, p=0.000870). The full-length
molecules of WtCD44 and CD44ΔICD show the size difference only in 6% gel. The ICD
deletion mutant generated significant amounts of solCD44E in the presence or absence of
TPA. This indicates that the ICD represses cleavage and is needed for regulated processing.
Mutation of the KR motif of the ICD prevents induced cleavage and was, like the ICD
deletion, resistant to merlin (NF2, see column diagrams).
C: We had shown in the past that activated merlin exerts its function upon binding to a basic
amino acid stretch in the membrane proximal part of the cytoplasmic domain of CD44.
Indeed, merlin co-precipitates CD44 only in the presence of the cytoplasmic tail (experiment
shown was done in NIH3T3 cells). The KR mutant does not interact with merlin
(Supplemental Fig. S3B).
Figure 4: ERM proteins promote CD44 cleavage
A: Downregulation of all three ERM proteins reduce CD44 cleavage.
RPM-MC cells were transfected as in Figure 1A. Cells were grown at low density. The
expression of ERM proteins was downregulated in about 98 % of the cells by a mixture of
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

30
small interfering RNAs (siRNAs) targeting all three members of the ERM protein family,
ezrin, radixin and moesin (non-targeting siRNA “C” was used as control). Downregulation of
ERM proteins inhibited ectodomain cleavage of CD44.
B: NRG1 cleavage is resistant to downregulation of ERM proteins.
Set-up of the experiment as in A except that double-tagged NRG1 was transfected as cleavage
substrate.
C: Constitutive activation of the Ras pathway does not stimulate CD44 cleavage.
Overexpression of a HA-tagged dominant active SOS mutant (DA-SOS, 24) that is
permanently membrane-associated, activates Ras – as detected by phosphorylated Erk -
independently of the presence of ERM proteins and TPA treatment. Histogram shows mean
values of relative level of solCD44E ± SD from three independent experiments (Wt: V versus
DA-SOS, p= 0.53955; KR-Mt: V versus DA-SOS, p=0.932614).
Figure 5: Mechanism of ERM protein dependent CD44 cleavage
A: Inhibition of the Ras pathway does not affect CD44 cleavage.
NIH3T3 cells were seeded in 6-well plates at low cell density. The cells were transfected as in
Figure 1A. Cells were pre-treated with increasing concentrations of MEK1 inhibitor
(PD98059) for 30 minutes and afterwards stimulated with 100 ng/ml TPA for 4 hours. The
effect of PD98059 was confirmed by its inhibition of Erk phosphorylation. Inhibition of the
MEK-ERK pathway had no effect on CD44 cleavage.
B: Effect of overexpressed ezrin mutants on CD44 cleavage.
RPM-MC cells were co-transfected with plasmids encoding N-terminal FLAG and C-terminal
HA-tagged CD44, and plasmids encoding myc-tagged ezrin mutants (described in ref. 42):
T567A (inactive), T567D (active), R579A (not interacting with F-actin) and treated as in
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

31
Figure 1A. Overexpressed mutant ezrins compete with endogenous ERM proteins for
cleavage-relevant interactions.
C: Block of actin polymerization inhibits CD44 cleavage
RPM-MC cells were transfected with a plasmid encoding wild-type CD44 and treated with
DAPT as in Figure 1A. To block actin polymerization (F-actin) latrunculin B was added at the
concentrations indicated. 30 min after addition of latrunculin B cells were stimulated with 100
ng/ml TPA for 30 minutes.
D: Neuregulin release is inhibited by interference with F-actin
HEKNE wt cells (HEK293T-AT1R +pB-Flag-NRG1-EGFP) were pretreated with a vehicle
control (DMSO) or with the Arp2/3 inhibitor CK-548 in increasing concentrations (0-15 μM)
alone or prior to angiotensin II (1 nM) stimulation. NRG1 cleavage was detected by
immunoblot using NRG1 antibodies. Angiotensin II was able to induce NRG1 cleavage in the
absence (lane 2), but not in the presence of CK-548 (lanes 6-8).
Figure 6: Induction of CD44 cleavage by physiologic stimuli in the breast cancer cell MDA-
MB-231
A: Growth factor dependent cleavage induction. MDA-MB-231 cells were transfected with
the double tagged CD44 construct as in Figure 1A.
B: Merlin inhibits TPA or HGF induced CD44 cleavage in MDA-MB-231 cells. The left
panel shows the detection of cleavage of endogenous CD44 expression using human CD44
specific antibodies. In the right panel cells had been transfected with tagged CD44.
A, B: Cells were treated for 1 hour with 50 ng/ml of growth factors, or 100 nM TPA.
Figure 7: Merlin inhibits cellular migration through block of CD44 cleavage
A: Quantitation of 3 series of scratch assays using immortalized mouse embryonic fibroblasts
from mice with floxed cd44 alleles (CD44fl/fl; GT(Rosa)26-CRE (B6/129) and derivatives of
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
32
these cells. Examples of the original photographs are shown in Supplemental Fig.6. CD44fl/fl
cells express the endogenous CD44. CD44fl/fl/shNF2 = the same cells with stable
downregulation of merlin. After CRE induced disruption of the cd44 alleles (CD44-/-) the
cells were infected with viral constructs encoding CD44 wt or the non-cleavable mutants:
CD44-KR-Mt and CD44stalk-del. Supplemental Fig.5 shows the levels of merlin and CD44
in the analyzed MEFs lines.
B and C: Scratch assays using MDA-MB-231 cells. The cells were infected by lentiviral
merlin constructs where indicated. In (C) rescue of cleavage dependent migration by
expression of the soluble CD44 ectodomain. To resolve the time course better, the
temperature was reduced to 30o C.
Tables
Table 1: Primers used for mutagenesis
Site-directed mutagenesis:
Forward primer 5’-3’ Reverse Primer 5’-3’
CD44 KR-Mt cagtagggcagcgtgtgggcaggcggcggcgctggtgatc gatcaccagcgccgccgcctgcccacacgctgccctactg
CD44ΔICD gcattgctgtcaacTGAaggagaaggtg caccttctcctTCAgttgacagcaatgc
CD44Δstalk gtgatggcaccTGGCTTATCATC GATGATAAGCCAGGTGCCATCAC
Table 2: siRNA oligonucleotides
Target GenBank Accession No: Sense strand sequence, 5’-...-3’
human ADAM10 NM_001110 GCUAAUGGCUGGAUUUAUU
GGACAAACUUAACAACAAU
CCCAAAGUCUCUCACAUUA
GCAAGGGAAGGAAUAUGUA
luciferase (control) CGUACGCGGAAUACUUCGATT
human ezrin NM_003379 AACCCCAAAGAUUGGCUUUCC
human radixin NM_002903 AAGCAGUUGGAAAGGGCACAA
human moesin NM_002444 AGAUCGAGGAACAGACUAAGA
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
A
Figure 1
B
C
0.00
0.50
1.00
1.50
5 7 9
rela
tive
le
ve
ls o
f C
D4
4D
E
-TPA
+TPA
***
***
x105 cells/well
TPA
TPA
-
+
RPM-MC
plus DAPT
minus DAPT
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
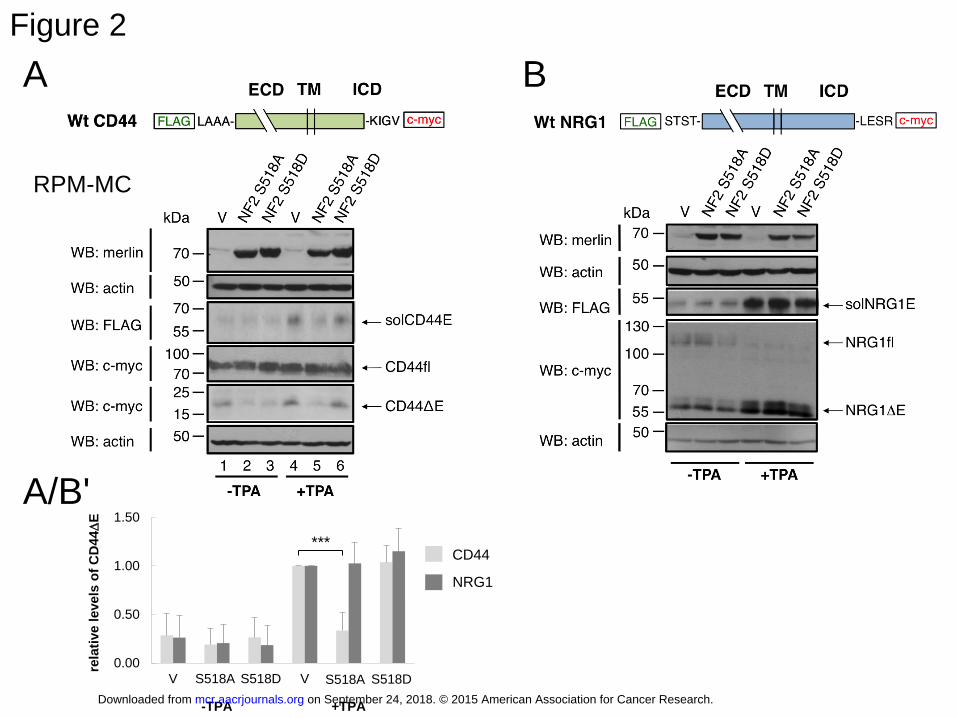
Figure 2
A B
0.00
0.50
1.00
1.50
V S518A S518D V NF2 A NF2 D
rela
tive
le
ve
ls o
f C
D4
4D
E
CD44
NRG1
***
-TPA +TPA
CD44
NRG1
S518A S518D
A/B'
RPM-MC
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
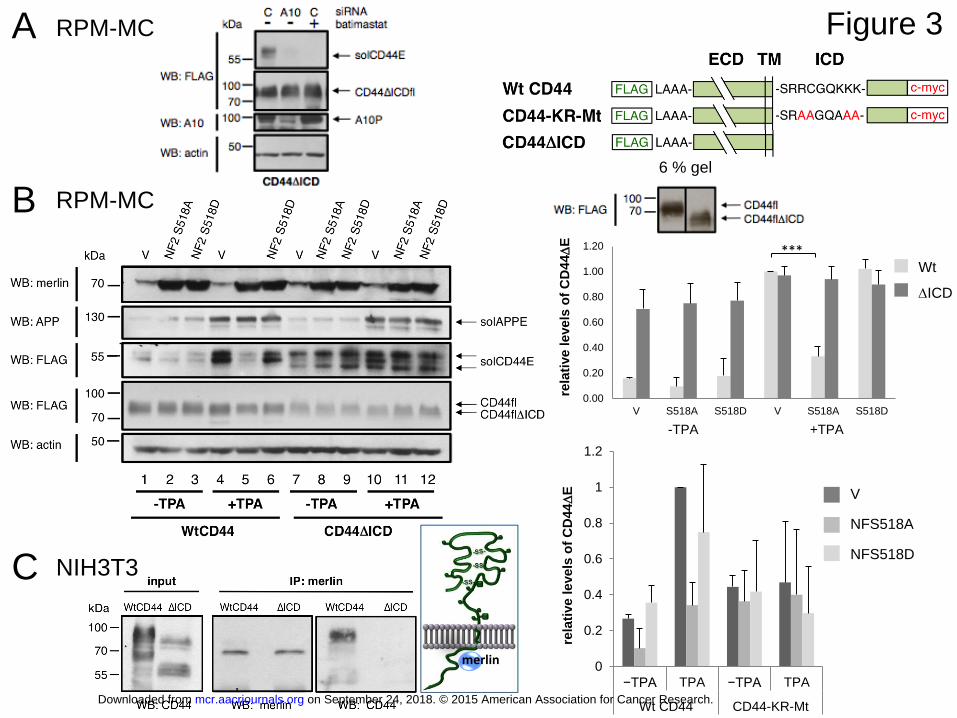
Figure 3
6 % gel
NFS518A
NFS518D
V
B RPM-MC
Wt
DICD
0.00
0.20
0.40
0.60
0.80
1.00
1.20
V S518A S518D V S518A S518D
rela
tive
le
ve
ls o
f C
D4
4D
E
***
-TPA +TPA
0
0.2
0.4
0.6
0.8
1
1.2
−TPA TPA −TPA TPA
Wt CD44 CD44-KR-Mt
rela
tive
le
ve
ls o
f C
D4
4D
E
A RPM-MC
C
merlin
NIH3T3
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
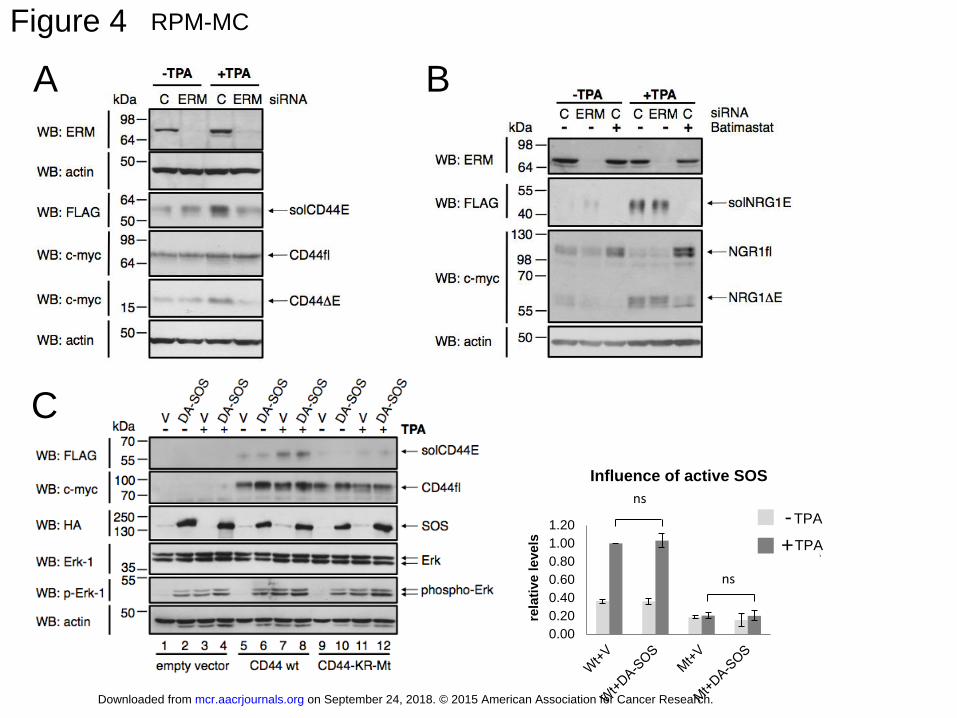
Figure 4
A
RPM-MC
0.00
0.20
0.40
0.60
0.80
1.00
1.20re
lati
ve
le
ve
ls
Influence of active SOS
-TPA
+TPA
ns
ns
TPA
TPA
-
+
C
B
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
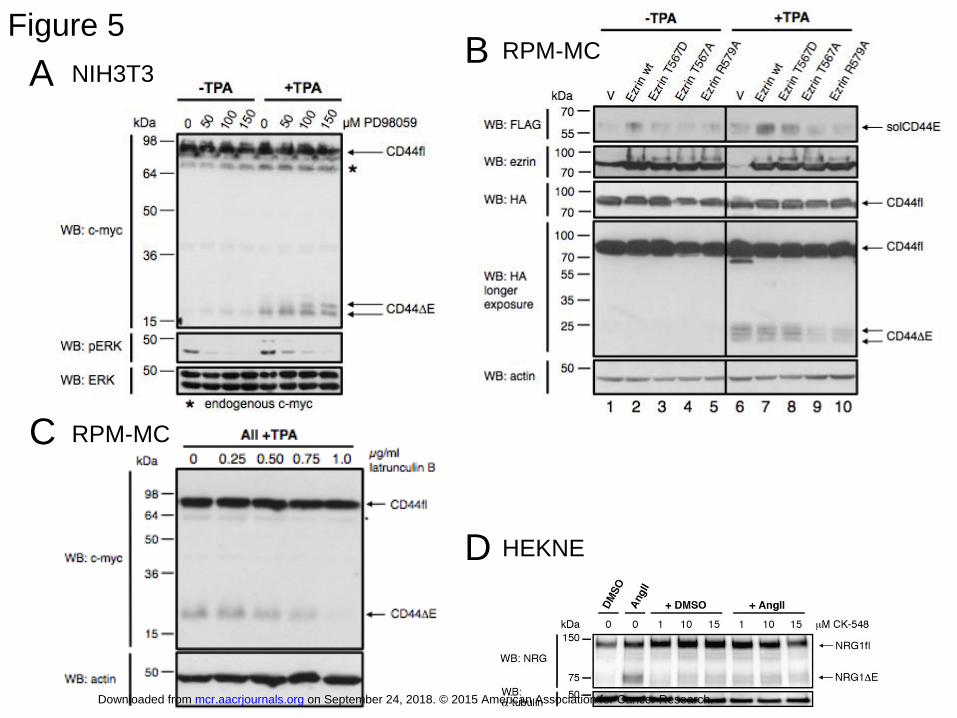
Figure 5
HEKNE
F
D
A
C
B NIH3T3
RPM-MC
RPM-MC
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
0
0.2
0.4
0.6
0.8
1
1.2
V S518A S518D
rela
tive levels
of
CD
44D
E
Figure 6 A
B
Myc-tagged transfected CD44 wt
Endogenous CD44
MDA-MB-231
control
HGF + TPA
TPA
-
+
Myc-tagged transfected CD44 wt
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Control TPA EGF PDGF FGF LPA
rela
tive levels
of
CD
44D
E
Wt CD44
0
0.2
0.4
0.6
0.8
1
1.2
V S518A S518D
rela
tive leve
ls o
f C
D44D
E
Wt CD44
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T
MDA-MB-231 B
0
10
20
30
40
50
60
70
80
% w
ou
nd
gap
rem
ain
ing
-BATIMASTAT
+BATIMASTAT**
**** **
Figure 7
A MEFs
0
20
40
60
80
100
V NF2A NF2D
% w
ou
nd
ga
p r
em
ain
ing
after 24 h
- solCD44
+ solCD44
0
20
40
60
80
100
V NF2A NF2D
% w
ou
nd
ga
p r
em
ain
ing
after 36 h
- solCD44
+ solCD44
control
Batimastat +
0
20
40
60
80
100
V NF2A NF2D
% w
ou
nd
ga
p r
em
ain
ing
after 12 h
- solCD44
+ solCD44
solCD44E
control
+
solCD44E
control
+
solCD44E
control
+
MDA-MB-231 C
0%
10%
20%
30%
40%
50%
60%
10 24 48
% w
ou
nd
ga
p r
em
ain
ing
hours after scratching
NF2S518A
control
NF2S518D
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T

Published OnlineFirst February 4, 2015.Mol Cancer Res Monika Hartmann, Liseth M. Parra, Anne Ruschel, et al. BY INHIBITING ECTODOMAIN CLEAVAGE OF CD44TUMOR SUPPRESSOR NF2 BLOCKS CELLULAR MIGRATION
Updated version
10.1158/1541-7786.MCR-15-0020-Tdoi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2015/02/05/1541-7786.MCR-15-0020-T.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/early/2015/02/04/1541-7786.MCR-15-0020-TTo request permission to re-use all or part of this article, use this link
on September 24, 2018. © 2015 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on February 4, 2015; DOI: 10.1158/1541-7786.MCR-15-0020-T





























