Embed Size (px)
Citation preview

1 序論:皮膚機能と老化
千秋達雄 老人総合研究所細胞化学部門
はじめに 一見すると皮膚は体表を覆う薄っぺらで、無味乾燥とした存在でしかない。老化に伴う脳や循環器系の障害が生命維持を脅かすことがあっても、皮膚の老化が直接生死に関わることは有りそうにない。ところが、皮膚は水や空気のように有って当り前、無ければたちまち生命維持が困難に曝される存在である。皮膚の機能の最も基本的なものは、物理的バリアとして乾燥や有害物質の侵入から生体を守ることで、それ無しには生きて行くことはできない。物理的バリアを破って侵入してきた有害物質や微生物や活性酸素に対しては、免疫系や抗酸化防御系が備わっている。それゆえ、皮膚は非常に多様な細胞から構成された複雑な機能を営む器官で、その詳しい働きについては未だ分からないことが多い。皮膚は外見的にも明らかな老年性変化を示すが、それがどのように進むか、またそれをどうすれば遅らせるかを知るには、まだまだ基礎的 ・ 応用的研究が必要である。本章では、皮膚の機能と老化について概観し、以後の章では各々の著者らの専門的見地から見た皮膚の機能や老化の局面をご解説いただこう。
1.1 皮膚の構造と機能 皮膚は、表面から順に表皮、真皮、皮下組織からなる3層構造を持っている(図 1.1)。
図 1.1 皮膚の構造と機能の模式図表皮は後で述べるように、分化の様相の異なる何層もの細胞から成る構造を持っている。真皮は表皮の支持組織として存在し、膠原線維、弾性線維のネットワーク、微小血管、

知覚神経などが走っている。皮下組織は脂肪に富み、外力に対するクッションとして、またエネルギー貯蔵や、断熱材しての働きがある。このような層状構造に埋まり込む形で汗腺、毛嚢、脂腺、立毛筋等が存在しているこれらは皮膚の付属器官とばれ、立毛筋以外はいずれも発生学的に表皮と同様に上皮細胞が分化したものである。
a) 表皮の細胞構成 表皮は分化の段階の異なる表皮細胞 (ケラチノサイト) が層状に重なってできており、いずれも細胞骨格タンパク質としてケラチンを含んでいる (図 1.2)。
図 1.2 表皮の細胞構成の模式図
一番内側の基底層では一層の円柱状細胞として存在し、その上に数層からなる有棘層がある。基底層と有棘層下層の細胞のみが増殖能を持っている。表皮細胞は次第に表面に向かって多角形から偏平な形状に変わりながら分化し、更に分化が進んだ顆粒層ではケラトヒアリンと呼ぶ顆粒が出現する。表皮細胞は顆粒層の外側で細胞核消失、ケラチン線維の凝集、周辺帯 (ComiHedEnvelope) 形成を含む急激な形態変化 (角化) を起こして角層を形成する。角層は垢として剥離するが、内側の表皮細胞が増殖 ・ 分化することで補われている。成人では基底層の細胞が角化するまで約一か月、垢として剥離するまで吏に約2週間かかると言われる。基底層には少数ながらメラニン細胞が存在し、複数の突起を伸ばして表皮細胞にメラニン顆粒を分配している。また、有棘層には少数ながらやはり複数の突起を持つランゲルハンス細胞が存在する。ランゲルハンス細胞は皮膚の免疫機能に重要な役割を演じており、マクロファージと同様に外部から侵入した抗原を補食後、リンパ節に移動して不細胞を活性化する。
b) 表皮細胞の分化 皮膚の機能の最も基本的なものは、物理的バリアとして乾燥や有害物質の侵入から生

体を守ることである。この役割を担っているのが表皮の角層である。それゆえ、表皮細胞が最終的に角化まで分化の過程を辿ることが、生命維持のために不可欠である。表皮を含む上皮系組織には細胞骨格タンパク質としてケラチンが存在する [1]。ケラチンには 30 種類に及ぶ分子種があり、酸性のタイプⅠと塩基性のタイプ I I のペアが、組織特
図 1.3 表皮細胞の分化と機能発現
異的に発現している ・ 基底層では塩基性の K5 と酸性の K14 のペアーが発現される (図1.3)。基底層より上の細胞では K5/K14 の発現が止まり、代わりに塩基性の K1 と酸性のK10 のペアが発現される。有棘層上層から顆粒層にかけては、角化に備えてプロフィラグリン、ロリクリン、インポルクリン等のマー力一タンパク質が発現する [2]。プロフイラグリンは多数のフィラグリンのユニットを含む高分子前駆体タンパク質として合成され、高度にリン酸化された形でケラトヒアリン顆粒に貯蔵される。脱リン酸化されるとプロテアーゼで分解されてフィラグリンが遊離される。フィラグリンはケラチン線維を凝集させる働きをするので、プロフィラグリンとして貯蔵しておけば、角化に必要になるまで時期尚早のケラチン線維の凝集を防いでおける。ロリクリン、インポルクリンは有棘層上層から顆粒層にかけて発現し、同じく有棘層上層から顆粒層で発現するトランスグルタミナーゼの主要な生体内基質タンパク質として、グルタミン残基とリジン残基の間でイソペプチド架橋されて角層細胞内に周辺帯を形成する ・ これら一連の遺伝子発現や酵素反応はカルシウムイオンで促進される [4]。穎粒層の細胞にはケラトヒアリン顆粒のほか、糖脂質の一種セラミドを含む層板顆粒も含まれており、角層細胞問に放出されて耐水性バリアを形成している [5,6]。角層細胞は、核を失っている点では既に死んだ細胞であるが、フィラグリンが内在するプロテアーゼの働きで分解されて、生じたペプチドやアミノ酸が天然の保湿成分 (NMF) として働き、角層のしなやかさを保つのに役立っている [6]。従って、角層バリアにおける水分保持には糖脂質とアミノ酸の両方が関わっている。従来より、角層のケラチンはプロテアーゼで限定的に分解され、且つシステイン残基間でジスルフィド架橋を形成していることが分かっていたが、最近になって角化に伴ってケラチンやフィラグリンがアルギニン残基で脱イミノ化されるこ

とも分かってきている [7]・
C) 真皮の構造と機能 真皮は組織学的に結合組織に属し、表皮細胞が緊密に詰まった表皮に比べると、細胞がまばらに存在している。主要なものは線維芽細胞で、その他はマクロファージ、肥満細胞等である。細胞外基質として膠原線維や弾性線維が走り、間隙をグリコザミノグリカンのゲルが埋めていいる (図 1.4)。
図 1.4 真皮の構造の模式図
これらの細胞外基質に埋り込む形で毛嚢、汗腺、脂腺等皮膚の付属器官と毛細血管と知覚神経のネットワークが存在する。膠原線維が皮膚に引っ張り強度を与えるのに対して、弾性繊維は力が加わって一時的に変形した皮膚が元の形に戻る弾性を与える。グリコザミノグリカンは真皮の乾燥重量の 0.2%しか存在しないが高い砲水力を持ち、組織中に水和した環境を保つほか、種々の活性物質の伝達にも働いていると思われる。このような細胞外基質成分の三次元構造は、線維芽細胞で合成された素材が細胞外で加工されてできたものである。真皮は、便宜的に上部の乳頭層と下部の網状層に分けて観察される。乳頭層は、表皮との境界の真皮乳頭の起伏に接した狭い部分で、比較的に細い膠原線維や弾性線維が散在している。細胞成分やプロテオグリカンの間質成分が多く、毛細血管系もよく発達している。この部分の膠原線維の直径は 0.3-3 μ m である。真皮の大半を占める網状層には細胞成分や血管系が乏しく、太い膠原線維や弾性線維が密集している。この部分の膠原線維の直径は 10-40 μ m である。膠原線維を構成するコラーゲンは細胞外基質の主要タンパク質で、乾燥重量の約 70%を占める。コラーゲン分子は、約 1000 個のアミノ酸からなるロープ状に絡み合った 3 本のα鎖が、1.5×300nm のスーパーヘリックス構造を作ったものである。コラーゲン分子は 1/4 長ずつずれて集合して膠原細線維 (径 10-300nm) を形成し、それが束ねられたものが膠原線維で、約70nm の周期で横紋構造が見られる。異なる遺伝子由来のα鎖が複数同定され、それら

の組み合わせの違いをもとに I 型から XII 型までのタイプが見つかっている。皮膚のコラーゲンの約 75%を I 型、25%を I I I 型が占めており、残りは微量成分である。コラーゲンはフィブロネクチンと呼ぶ接着タンパク質を介して真皮の細胞との結合を保っている。弾性線維は、横紋のない線維状構造 (micro-f ibri l ) とその間隙を充填する無定形構造を持ち、それぞれフィブリリンとエラスチンとからなっている。エラスチンは真皮の総タンパク質の約 2%を占め、ランダムコイル状のペプチド鎖が鎖間架橋してゴムのように伸縮自在なネットワーク構造を作っている。グリコザミノグリカンは、直鎖状の酸性多糖類の総称で、真皮に多いのはヒアルロン酸とデルマタン硫酸である。ヒアルロン酸以外のグリコザミノグリカンはコアタンパク質に多数結合してプロテオグリカンと呼ぶ親水性巨大分子を形成している。
d) 複合組織としての皮膚 表皮と真皮は発生学的にも形態学的にも独立の存在に見えるが、機能的には密接に影響しあっている組織である。表皮と真皮を隔てる基底膜は、表皮細胞から分泌されたIV 型コラーゲンのネットワークの重層構造に、接着タンパク質やプロテオグリカンが結合してできている。表皮側では基底層の表皮細胞の半接着斑 (ヘミデスモソーム) と接触し、真皮側では VII 型コラーゲンからなる係留線維や弾性線維と接触している。表皮細胞が基底層から有頼層に移行することは、基底膜の直接的影響から離れて分化への道を辿ることであり、これに伴って K5/K14 ペアから K1/K10 ペアヘとケラチン遺伝子の発現交代が起こる。ところが、表皮細胞だけを培養しても直ちに重層化や K1/K1Oの発現が始まるとは限らない。真皮由来の要因以外に、培養器材や他の表皮細胞との接着状態、培地のカルシウムイオン濃度等が複雑な影響を与えるので、培養系で表皮細胞を正常な皮膚のように角化させることは難しい。また) 培養系に置いた線維芽細胞は分裂や遊走を示すが、真皮のような三次元構造を作らせることは容易でない。これらの問題点に対処するため、コラーゲンゲル中で繊維芽細胞を生やして作った代替真皮の上に表皮細胞を撒き、気相に曝しながら培養して重相角化させる皮膚の三次元培養法が開発されている [8]。創傷治癒の過程では、表皮細胞と線維芽細胞の外にもマクロファージ、好中球、血管内皮細胞、平滑筋細胞も巻き込んだサイトカインネットワークと細胞外基質の協調的相互作用で、創傷部位付近の細胞の増殖、分化、遊走が誘発される [9]。その際に表皮細胞と線維芽細胞は、上述のような角化と細胞外基質の形成という本来の働き以外に、白らもサイトカインを分泌してネットワークの構築に参画する [8]。表皮細胞には通常でも他の組織よりはるかに高レベルの IL-1 が含まれ [1O,11]、炎症反応を促進する [12]。線維芽細胞では表皮細胞増殖因子 (KGF) が多量に誘導されて表皮細胞の増殖が促進されたり [13]、表皮細胞の作る表皮増殖因子 (EGF) で繊維芽細胞の増殖が促進される等の相互作用が営なまれる。増殖が促進された表皮細胞は、通常には発現されない過増殖型ケラチンペア K6/K16 を発現する [14]。また、血小板由来増殖因子(PDGF)[15] やコラゲナーゼ [16]の産生も報告されている。
1.2 内因性老化と光老化

皮膚とその付属器官は、白髪、しわ、たるみ、しみ等外から容易に観察できる老年性変化を示すが、これは個体の加齢に依存した内因性の要因と外部から皮膚に加えられた傷害要因との両方の帰結として現われたものである [17-19]。同一人あるいは同一個体でも部位による程度の差が大で、屋外での作業を日課としている人の陽に曝された部分は、30 歳でも老年者に見られる深いしわやしみが見られるのに反して、老年者でも着衣で光線から保護された部分にはそのような所見が見られない。陽に曝された結果としてもたらされた皮膚の退行性変化を光老化 (Photoaging) と呼び、内因性老化(Intrinsic Aging) と区別している。光老化には内因性老化が病的に誇張された側面がなくもないが、これを一般的概念とするのは誤りである。光老化で悪役を演じるのは紫外線である。紫外線が皮膚におけるビタミン D の合成を促進することから、嘗て日光浴は健康を増進すると推奨されてきた。しかし、紫外線には殺菌作用があることが知られているように、皮膚の細胞にも様々な傷害を与え、DNA に修復されない変異を起こした場合は、前癌状態を経て悪性腫瘍に移行するリスクが高くなる。オゾン層の破壊が進んでいる状況で、紫外線が 1%増えると偏平上皮癌が 2%増えるという推測もあり、世界的に皮膚癌が増えることが予想される。日常生活でも帽子や長袖服の着用や、サンスクリーンの使用で強い日光に当たるのを避ける工夫が望ましい [20]。現代では、ビタミン D も食品から必要量を摂取可能である。陽に当たると表皮のメラニン細胞でメラニン合成の第一段階に働くチロシナーゼが活性化され、表皮のメラニン分布が増加する [21]。これは浅黒い日焼けとして観察され、紫外線傷害の軽減に働く。これはタンパク質リン酸化酵素の活性化と遺伝子発現誘導を介した防衛反応であるが、日光浴の効用を支持するものではない。光老化に特徴的なのは日光性弾力線維症と呼ぶ弾性線維の変性である[22]。発症初期には真皮乳頭層でエラスチン様に染まる線維状構造の太さや絡み合いの増加として見られる。更に進むと、このような構造体が真皮網状層までも埋め尽くすように広範に見られる。弛みのできた皮膚に張りを持たせようと弾性線維が増え過ぎて、却って弛みを増してしまう悪循環を起こしたためと考えられる。これに伴ってコラーゲン [23-25]含量が減少すると言われている。最近、これに伴って弾性線維と結合して存在するプロテオグリカン分子種と、膠原線維と結合して存在するプロテオグリカン分子種の染色強度が増減することも報告された[26]。内因性老化で見られる変化としては、表皮と真皮の境界面の起伏の平坦化や表皮基底層の細胞の大きさや形態の不規則化が見られる [27](表紙の図を参照)。このため、基底層の接着面積が減少した老人の皮膚では、力が加わった時表皮と真皮が剥がれやすくなっている。表皮細胞の代謝回転や角層細胞の剥離速度も低下する [28]。メラニン細胞[29]やランゲルハンス細胞 [30]も減少する。真皮は厚さが減少し、単位平面積当たりのコラーゲン量も減少する [31]。真皮乳頭層の細い弾性線維は退化し、張りのない弛んだ外観を与える [26]。これに反して真皮網状層では弾性線維の数、太さ、分岐のいずれもが増加して乱雑な分布状態を示す [32]。真皮の細胞密度 [27]やプロテオグリカン含量 [33]は減少し、皮膚感覚 [34]や皮脂 [35] の産生も低下し、汗腺 [27]や血管系の退化 [27,36]に伴って皮膚温の調節能も低下する。器官全体と!して免疫能が低下し、創傷治癒遅延を来す。老人の皮膚は乾燥しているといわれる。湿度の低い冬に、皮膚の乾燥感から来る痒みを訴える老人も多い。この様な老
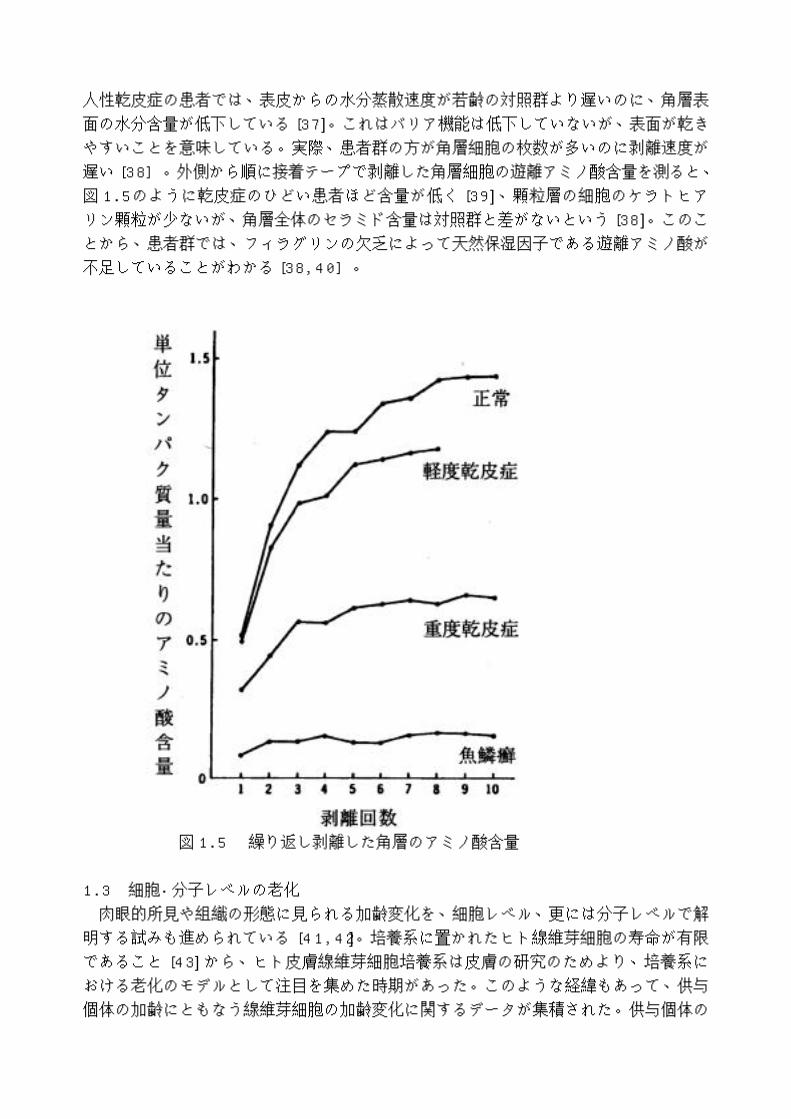
人性乾皮症の患者では、表皮からの水分蒸散速度が若齢の対照群より遅いのに、角層表面の水分含量が低下している [37]。これはバリア機能は低下していないが、表面が乾きやすいことを意味している。実際、患者群の方が角層細胞の枚数が多いのに剥離速度が遅い [38]。外側から順に接着テープで剥離した角層細胞の遊離アミノ酸含量を測ると、図 1.5 のように乾皮症のひどい患者ほど含量が低く [39]、顆粒層の細胞のケラトヒアリン顆粒が少ないが、角層全体のセラミド含量は対照群と差がないという [38]。このことから、患者群では、フィラグリンの欠乏によって天然保湿因子である遊離アミノ酸が不足していることがわかる [38,40]。
図 1.5 繰り返し剥離した角層のアミノ酸含量
1.3 細胞 ・ 分子レベルの老化 肉眼的所見や組織の形態に見られる加齢変化を、細胞レベル、更には分子レベルで解明する試みも進められている [41,42]。培養系に置かれたヒト線維芽細胞の寿命が有限であること [43]から、ヒト皮膚線維芽細胞培養系は皮膚の研究のためより、培養系における老化のモデルとして注目を集めた時期があった。このような経緯もあって、供与個体の加齢にともなう線維芽細胞の加齢変化に関するデータが集積された。供与個体の

年齢が高いほど培養系での寿命が短く [44]、増殖因子の刺激に対する反応性が低い[45]。細胞表面の受容体に対する EGF の結合は加齢に伴って低下し、老人 (67-82 歳)由来の線維芽細胞の受容体含量は若者 (24-32 歳) 由来の細胞の 1/4、新生児由来の細胞の 1/40 に減少し、EGF が受容体と結合した複合体の細胞内への取り込みも遅延する[46]。また、老齢個体由来の線維芽細胞はコラーゲン mRNA 含量 [47]やコラーゲン分泌 [48]が低下する。若齢由来の細胞が増殖刺激を受けるとコラゲナーゼ等の分解酵素系が活性化されて細胞外基質の部分的分解と修復を起こした後、元の不活性状態に戻るのに対して、老齢由来の細胞では分解酵素系の発現が持続的に高く、それを抑制する阻害因子の発現は低下している [49,50]。このような遺伝子レベルの加齢変化は、内因性老化に見られる真皮細胞外基質の退行性変化をもたらしていると思われる。表皮細胞についても、供与個体の加齢に依存したコロニー形成能や増殖刺激に対する反応性の低下が見られており [41]、遺伝子発現との関連 [51]も示唆されているが、概してまだデータが不足である。
1.4 レチノイン酸による皮膚傷害の予防と改善 先進諸国における平均寿命の延びと、オゾン層破壊による紫外線量の増加によって、長期にわたる紫外線被曝による皮膚癌発生率の増加が予想され、光線による傷害の予防と治療対策が重要課題になってきている。期待される治療薬としてビタミン A 類縁化合物のレチノイン酸の効果が注目されており、ここ十数年間にかなりのデータが集積されている [52-56]。レチノイン酸は細胞核内受容体と複合体を形成して DNA 上の認識配列に結合して多様な分化を誘導することが知られており、皮膚科領域ではトレチノインと呼ばれている。O.0O1-0.1%トレチノインを含むクリームまたはクリームのみを長期間塗付し続けて、臨床的及び組織化学的所見から効果が判定されている。6 ヵ月間の投与により、小しわ、しみ、あれ、たるみ等の臨床的所見に有意な改善が見られ(図 1.6)、
図 1.6 トレチノイン塗布による皮膚傷害の改善

組織化学的には角層の厚みの減少、顆粒細胞の総数増加、表皮全体の厚さの増加、表皮メラニン含量の減少等が共通して見られているが、臨床的所見を説明するような真皮の組織像の改善は認められていない [53]。しかし、1O-12 ヵ月間トレチノイン塗付を続けた場合は、抗 I 型コラーゲンによる免疫組織化学染色 [54] や電子顕微鏡的観察 [55]で、真皮コラーゲン産生促進が見られている。また、トレチノインの内因性老化に対する効果に関する情報は少ないが、68-79 歳の白人女性の片足の大腿部にトレチノインを9 ヵ月塗付した実験で、元老化した皮膚に見られたと同様な表皮の厚みの増加以外に、平坦化した表皮と真皮の境界の起伏の増加と、基底細胞や有様細胞の形態の規則化が観察されている [56]。
おわりに 皮膚の機能的構造、外観や組織像に見られる老化像と細胞 ・ 分子レベルの背景、レチノイン酸への期待等についてざっと述べた。はじめに述べたように、皮膚はあって当り前で有難味の判らない器官であるが、生体を外界から守るためにきわめて多様な役割を演じている。皮膚に常在する細胞の動態は近隣の細胞との接触ばかりでなく、接触のない細胞から分泌されるサイトカイン類の影響下にあり、正常なプロセスがどのうように進むかについても解明が待たれる問題が残されている。今後に予想される人口学的並びに地球環境的状況の変化による皮膚傷害の増加に対処するため、更なる基礎的、応用的研究の継続が必要である。
引用文献l . Fuchs, E. 1993. Epidermal differentiation and keratin gene expression. J. Cell Sci. 106 Suppl. 17: 197-208. 2. Steven, A. C., M. E. Bisher, D. R. Roop, and P. M. Steinert. 1990. Biosynthetic pathways of filaggrin and loricrin--two major protelns expressed by terminally differentiated epidermal keratinocytes. J. Struct. Biol. l 04: 1 50- 1 62.3. Steinert, P. M. and L. N. Marekov. 1995. The proteins elafin, filaggrin, keratin intermediate filaments, Ioricrin, and small proline-rich proteins I and 2 are isodipeptide cross-linked components of the human epidermal cornified cell envelope. J. Biol. Chem. 270: 17702- 1771 1 .4. Yuspa, S. H., H. Hennings, R. W. Tucker, S. Jaken, A. E. Kilkenny, and D. R. Roop. 1988. Signal transduction for proiiferation and differentiation in keratinocytes. [Review]. Ann. NY Acad. Sci. 548: 191-196. 5. Wertz, P. W., M. C. Miethke, S. A. Long, J. S. Straus, and D. T. Downing. 1985. Composition of ceramides from human stratum comeum and comedones. J. Invest. Drmatol. 84:410-41 2. 6. Rawlings, A. V., I. R. Scott, C. R. Harding, and P. A. Bowser. 1994. Stratum comeum moisturization at the molecular level. [Review]. J. Invest. Dermatol. 103:731-741. 7. Senshu, T., S. Kan, H. Ogawa, M. Manabe, and H. Asaga. 1996. Preferential deimination of keratin K I and filaggrin during the terminal differentiation of human epidermis. Biochem. Biophys. Res. Commun. 225:712-7 1 9. 8. Bell, E., S. Sher, B. Hull, C. Merri l l , S. Rosen, A. Chamson, D. Asselineau, L. Dubertret, B. Coulomb, C. Lapiere, B. Nusgens, and Y. Neveux. 1983. The reconstitution of living skin. J. Invest. Dermatol. 8 1 :2s- 10s. 9. Moulin, V. 1995. Growth factors in skin wound healing. Eur. J. Cel] Biol. 68: 1-7. 10. Hauser, C., J. H. Saurat, A. Schmitt, F. Jaunin, and J. M. Dayer. 1986. Interleukin I is present in normal human epidermis. J. Immunol. 1 36:33 1 7-3323.

l I . Kupper, T. S., D. W. Ballard, A. O. Chua, J. S. McGuire, P. M. Flood, M. C. Horowitz, R. Langdon, L. Lightfoot, and U. Gubler. 1986. Human keratinocytes contain mRNA indistinguishable from monocyte interleukin I alpha and beta mRNA. Keratinocyte epidermal cell-derived thymocyte-activating factor is identical to interleukin I . J. Exp. Med. 164:209_5-2100. 12. Kupper, T. S. 1990. Immune and inflammatory processes in cutaneous tissues. Mechanisms and speculations [Review]. J. Clin. Invest. 86: 1783- 1789. 13. Wemer, S., K. G. Peters, M. T. Longaker, F. Fuller-Pace, M. J. Banda, and L. T. Williams. 1992. Large induction of keratinocyte growth factor expression in the dermis during wound healing. Proc. Natl. Acad. Sci. USA 89:6896-6900. 14. Jiang, C. K., T. Magnaldo, M. Ohtsuki, I. M. Freedberg, F. Bernerd, and M. Blumenberg. 1993. Epidermal growth factor and transforrning growth factor alpha specifically induce the activation- and hyperproliferation-associated keratins 6 and 16. Proc. Natl. Acad. Sci. USA 90:6786-6790. 15. Ansel, J. C., J. P. Tiesman, J. E. Olerud, J. G. Krueger, J. F. Krane, D. C. Tara, G. D. Shipley, D. Gilbertson, M. L. Usui, and C. E. Hart. 1993. Human keratinocytes are a major source of cutaneous platelet-derived growth factor. J. Clin. Invest. 92:67 1-678. 16. Saarialho Kere, U. K., S. O. Kovacs, A. P. Pentland, J. E. Olerud, H. G. Welgus, and W. C. Parks. 1993. Cell -matrix interactions modulate interstitial collagenase expression by human keratinocytes actively involved in wound healing. J. Clin. Invest. 92:2858-2866. 17. Kligman, A. M. and A. K. Balin. 1989. Aging of human skin. in Aging and Skin (Balin, A. K. and A. M. Kligman, eds.) pp. 1-42, Raven Press, New York. 18. Bhawan, J., C. H. Oh, R. Lew, K. S. Nehal, R. R. Labadie, A. Tsay, and B. A. Gilchrest. 1992. Histopathologic differences in the photoaging process in facial versus arm skin. American. J. Dermatopathol. 14:224-230. 19. Kurban, R. S. and A. K. Kurban. 1993. Common skin disorders of aging: diagnosis and treatment.[Review]. Geriatrics. 48:30-1 , 35-6, 39.
20. Green, H. A. and L. Drake. 1993. Aging, sun damage, and sunscreens. [Review]. Clinics Plast. Surg. 20: 1-8. 21. Gilchrest, B. A., H. Y. Park, M. S. Eller, and M. Yaar. 1996. Mechanisms of ultraviolet light-induced pigmentation. Photochem. Photobiol. 63: 1-10. 22. Kligman,L.H. 1989. Skin changes in photoaging: characteristics, prevention, and repair. in Aging and Skin (Balin, A. K. and A. M. Kligman, eds.) pp.33 1-346, Raven Press, New York. 23. Chen, V. L., R. Fleischmajer, E. Schwartz, M. Palia, and R. Timpl. 1986. Immunochemistry of elastotic material in sun-damaged skin. J. Invest. Dermatol. 87: 334=337. 24. Scwartz, E., F. A. Cruickshank, C. C. Christensen, J. S. Perlish, and M. Lebwohl. 1993. Collagen alterations in chronically sun-damaged human skin. Photochem. Photobiol. 58:84 1 -844. 25. Oikarinen, A. 1994. Aging of the skin connective tissue: how to measure the biochemical and mechanical properties of aging dermis. [Review]. Photodermatol. Photoimmunol. Photomed. lO:47-52. 26. Bernstein, E. F. B, L. W. Fisher, K. Li, R. G. LeBaron, E. M. Tan, and J. Uitto. 1995. Differential expression of the versican and decorin genes in photoaged and sun-protected skin. Comparison by immunohistochemical and northern analyses. Lab. Invest. 72:662-669. 27. Montagna, and K. Carlisle. 1979. Structural changes in aging human skin. J. Invest. Derrnatol. 73: 47-53. 28. Marks, R. 1989. Epidermal aging. in Aging and Skin (Balin, A. K. and A. M. Kligman, eds.) pp.277-283, Raven Press, New York. 29. Gilchrest, B. A., F. B. Blog, and G. Szabo. 1979. Effects of aging and chronic sun exposure on melanocytes in human skin. J. Invest. Dermatol. 73: 141-143. 30. Gilchrest, B. A., G. Murphy, and N. A. Soter. 1982. Effect of chronological aging and ultravilolet radiation on Langerhans' cells in human epidermis. J. Invest. Dermatol. 79, 85-88. 31. Shuster, S., M. M. Black, and E. McVitie. 1975. The influence of age and sex on skin thickness, skin collagen and density. Br. J. Dermatol. 93:639-643.

32. Tsuji, T. and T. Hamada. 1981 . Age related changes in human derTnal elastic fibers. British J. DerTnatol. 105:57 -63. 33. Longas, M. O., C. S. Russell, and X-Y. He. 1987. Evidence for structural changes in derrnatan sulfate and hyaluronic acid with aging. Carbohydr. Res. 159: 127-136. 34. Procacci, P. G. Bozza, G. Buzzelli, and M. D. Cortz. 1970. The cutaneous pricking pain threshold in old age. Gerontol. Clin. 12:213-218. 35. Jacobsen, E., J. K. Billings, R. A. Frantz, C. K. Kinney, M. E. Stewart, and D. T. Downing. 1985. Age related changes in sebaceous wax ester secretion rates in men and women. J. Invest. Dermatol. 85:483-485. 36. Kligman, A. M. 1979. Perspective and problems in cutaneous gerontology. J. Invest. Dermatol. 73:39-46. 37. Potts, R. O., E. M. Buras, and D. A. Chrisman. 1984. Changes with age in the moisture content of human skin. J. Invest. Dermatol. 82:97-100. 38. Hara, M., K. Kikuchi, M. Watanabe, M. Denda, J. Koyama, J. Nomura, I. Horii, and H. Tagami. 1993. Senile xerosis: functional, morphorogical, and biochemical studies. J. Geriatric Dermatol. I : 1 1 1 - 1 20. 39. Horii, I., Y. Nakayama, M. Obata, and H. Tagami. 1989. Stratum corneum hydration and amino acid content in xerotic skin. British. J. Dermatol. 121 :587-592. 40. Jacobson, T. M., K. U. Yuksel, J. C. Geesin, J. S. Gordon, A. T. Lane, and R. W. Gracy. 1990. Effects of aging and xerosis on the amino acid composition of human skin. J. Invest. Dermatol. 95:296-300. 41. Gilchrest, B. A. and M. Yaar. 1992. Ageing and photoageing of the skin: observations at the cellular and molecular level. [Review]. British J. Dermatol. 127 Suppl 41 :25-30. 42. West, M. D. 1994. The cellular and molecular biology of skin aging. [Review]. Arch. Dermatol. 130:87-95. 43. Hayflick, L., and P. S. Moorhead. 1961. The serial cultivation of human diploid cell strains. Exp. Cell Res. 37 :6 1 4-636. 44. Schneider, E. L., and Y. Mitsui. 1976. The relationship between in vitro cellular aging and in vivo human age. Proc. Natl. Acad. Sci. USA 73:3584-3588. 45. Plisko, A., and B. A. Gilchrest. 1983. Growth factor responsiveness of cultured human fibroblasts declines with age. J. Gerontol. 38:5 1 3-5 1 8. 46. Reenstra, W. R., M. Yaar, and B. A. Gilchrest. 1993. Effect of donor age on epidermal growth factor processing in man. Exp. Cell Res. 209: 1 18-122. 47. Furth, J. J. 1991 . The steady-state levels of type I collagen mRNA are reduced in senescent fibroblasts. J. Gerontol. 46:B 1 22-B 1 24. 48. Martin, M., R. el Nabout, C. Lafuma, F. Crechet, and J. Remy. 1990. Fibronectin and collagen gene expression during in vitro ageing of pig skin fibroblasts. Exp. Cell Res. 191 :8-1 3. 49. West, M. D., O. M. Pereira-Smith, and J. R. Smith. 1989. Replicative senescence of human skin fibroblasts correlates with a loss of regulation and overexpression of collagenase activity. Exp. Cell Res. 1 84: 1 38- 147. 50. Burke, E. M., W. E. Horton, J. D. Pearson, M. T. Crow, and G. R. Martin. 1994. Altered transcriptional regulation of human interstitial collagenase in cultured skin fibroblasts from older donors. Exp. Gerontol. 29:37 -53.
51. Gilchrest, B. A., M. Garmyn, and M. Yaar. 1994. Aging and photoaging affect gene expression in cultured human keratinocytes. Arch. Dermatol. 1 30:82-86. 52. Kligman, L. H., C. H. Duo, and A. M. Kligman. 1984. Topical retinoic acid enhances the repair of ultravilolet damaged connective tissue. Connect. Tissue Res. 12: 139-150. 53. Gilchrest, B. A. 1992. Retinoids and photodamage. [Review]. British J. Dermatol. 127 Suppl 41 : 14-20. 54. Griffiths, C. E., A. N. Russman, G. Majmudar, R. S. Singer, T. A. Hamilton, and J. J. Voorhees. 1993. Restoration of collagen formation in photodamaged human skin by tretinoin (retinoic acid). New

England J. Med. 329:530-535. 55. Yamamoto, O., J. Bhawan, G. Solares, A. W. Tsay, and B. A. Gilchrest. 1995. Ultrastructural effects of topical tretinoin on dermo-epidermal junction and papillary dermis in photodamaged skin. A controlled study. Exp. Dermatol. 4: 1 46- 1 54. 56. Kligman, A. M., D. Dogadkina, and R. M. Lavker. 1993. Effects of topical tretinoin on non-sun-exposed protected skin of the elderly. J. Am. Acad. Dermatol. 29:25-33.





























