Embed Size (px)
Citation preview

1
1 INTRODUÇÃO
A grandeza da biodiversidade Amazônica é bem conhecida, e dentre os
microorganismos que contribuem para a nossa megadiversidade estão os fungos.
Com o avanço dos estudos na área biotecnológica os organismos fúngicos
tornaram-se de grande interesse e vêm contribuindo com produtos e processos de
importância industrial. Na maioria das vezes, eles são vistos pela população como
prejudiciais, uma imagem que é dada pelas poucas espécies dentro do reino que
causam as micoses do homem e animais ou as que são responsáveis por doenças
em plantas cultivadas. Muitas pessoas associam os fungos com os bolores ou mofos
que invadem paredes úmidas das residências, artigos de couro ou ainda cobrem os
alimentos, como frutas e grãos armazenados. Algumas espécies de fungos podem
ainda ser associadas à culinária, como é o caso dos cogumelos (AZEVEDO, 2003).
Um número significativo de espécies produz substâncias de grande interesse
industrial, entre as quais podemos destacar antibióticos como a penicilina ou a
cefalosporina, vitaminas como a riboflavina, esteróides, ácido cítrico, etanol, enzimas
tipo celulases, proteases, amilases e muitas outras de valor industrial. Os fungos têm
grande importância agrícola e ecológica, decompondo restos vegetais, degradando
substâncias tóxicas e na simbiose com as plantas. Devido a sua versatilidade, os

2
fungos vêm sendo estudados, principalmente quanto à sua aplicabilidade
biotecnológica (AZEVEDO, 2003: ESPOSITO & AZEVEDO, 2004).
Dentre os vários potenciais biotecnológicos dos fungos ressalta-se o poder de
degradação da matéria orgânica. Os fungos são capazes de degradar a madeira,
sendo os da classe Basidiomycetes os mais eficientes degradadores, possuindo a
mais efetiva capacidade de biodegradação de materiais lignocelulósicos na natureza
(HATAKKA, 1994; HIGUCHI, 1990).
A biodegradação dos materiais lignocelulósicos por fungos é atribuída à ação
de uma série de enzimas e compostos de baixa massa molar extracelulares. A
degradação ocorre necessariamente de forma extracelular, uma vez que os
componentes dos lignocelulósicos devem ser inicialmente despolimerizados até
compostos menores que são susceptíveis ao transporte pela parede celular e ao
metabolismo intracelular dos fungos envolvidos (FERRAZ, 2004).
Castro e Silva et al. (1993) trabalhando com cepas de fungo de podridão
branca proveniente da Amazônia concluíram que o “pool” enzimático do P.
sanguineus o indica como potencial na descoloração de efluente Kraft de indústria
papeleira, uma vez que a presença de lignase ou lacase, juntamente com Mn-
peroxidase na presença de -glucosidase, são importantes para a degradação de
compostos cloroligninos (ESPOSITO et al., 1991; DURÁN, 1992).
As enzimas são os produtos microbianos mais explorados na indústria
biotecnólogica, pois apresentam uma série de vantagens no processamento de

3
alimentos e em muitos outros ramos da indústria manufatureira. Muitos produtos
alimentícios consumidos diariamente, entre os quais o pão e o queijo, são feitos com
a colaboração desse tipo especial de proteína.Também são empregadas na
indústria de produtos de limpeza, no processamento da polpa e do papel, na
preparação têxtil, e em aplicações médicas, onde com freqüência, substituem
compostos ou processos químicos. A vantagem da utilização de enzimas em muitos
processos deve-se à menor produção de subprodutos residuais, o que propicia a
obtenção de produtos de melhor qualidade e com menor probabilidade de poluição
do meio ambiente.
Outras vantagens da aplicação de enzimas estão relacionadas com o custo
de produção relativamente baixo, susceptibilidade de produção em larga escala em
fermentadores industriais, espectro amplo de características físico-químicas de
diferentes enzimas (geralmente relacionadas ao habitat e fisiologia do
microorganismo produtor), susceptibilidade de manipulação genética e por
representarem um recurso natural renovável.
O mercado mundial de enzimas movimenta milhões de dólares anualmente.
Esse mercado é estimado em cerca de U$ 1,5 bilhões de dólares, com um aumento
anual de 12% no volume de enzimas produzidas nos últimos 10 anos (CASTRO E
SILVA, SILVA & CAVALCANTE, 2002). Apesar do seu papel nos ecossistemas e
suas aplicações na biotecnologia, o conhecimento sobre os fungos amazônicos
degradadores de madeira ainda permanece num nível incipiente, sendo que poucos
dados na área sobre enzimas com o potencial industrial são disponíveis. A Região

4
Amazônica, apesar de sua grande biodiversidade microbiana, até o presente
momento não apresenta nenhuma participação nesse mercado.
Estudos da biodiversidade, visando espécies com potencial biotecnológico,
provavelmente poderão contribuir com a produção de enzimas com preço de
mercado compatível e melhoria das condições de vida da população. No entanto,
faz-se necessário maior investimento em pesquisas científicas e tecnológicas, a fim
de que esse potencial seja melhor explorado, pela indústria, alimentícia,
farmacológica, entre outras.
Pela importância dada às enzimas no mercado mundial, destaca-se a
necessidade do desenvolvimento de estudos enzimáticos a partir da biodiversidade
Amazônica. Assim, o presente trabalho visa contribuir com conhecimentos sobre os
aspectos de crescimento e atividade enzimática dos fungos P. sanguineus e
Trametes sp, degradadores de madeira, visando seu potencial enzimático.

5
2. OBJETIVOS
2.1. Geral
Avaliar os aspectos relacionados ao crescimento e atividade ligninolíticas dos
fungos amazônicos deterioradores de madeira, Pycnoporus sanguineus (L.:F.)
Murr e Trametes sp.
2.2. Específicos
Avaliar o crescimento micelial dos fungos em função do pH, temperatura e
meio de cultura;
Verificar a influência do teor de nitrogênio no crescimento micelial dos fungos;
Determinar a atividade ligninolíticas (lacase, fenoxidase, peroxidase e lignina
peroxidase) dos fungos;
Estudar a influência de diferentes fontes de carbono na atividade enzimática
dos fungos.

6
3 REVISÃO DE LITERATURA
3.1 Aspectos gerais dos fungos
Os fungos são seres eucariontes, heterotróficos, aclorofilados, aeróbios a
microaerófilos, uni ou pluricelulares, com parede celular composta geralmente de
quitina e/ou celulose, além de outros carboidratos complexos, com glicogênio como
substância de reserva, nutrição por absorção, podendo viver como sapróbios,
parasitas ou simbiontes com outros organismos (LACAZ, 2002; PUTZKE, 2004).
Possuem o corpo formado por um emaranhado de filamentos, denominados hifas, e
o seu conjunto recebe o nome de micélio. As hifas variam no diâmetro, espessura da
parede, localização do pigmento, etc. (PUTZKE, 2004). Constituem um grupo muito
grande e heterogêneo encontrado virtualmente em qualquer nicho ecológico. O
número de espécies fúngicas espalhadas pelo mundo é estimada em cerca de
1milhão e 500 mil sendo, que destas apenas foram descritas cerca de 74 mil
espécies. Excluindo-se os insetos, os fungos constituem os mais numerosos seres
vivos existentes (HAWKSWORTH, 1991; ESPOSITO e AZEVEDO, 2004).
Os fungos são organismos muito diferentes, se comparados a animais e
vegetais, sendo atualmente distribuído em três reinos: Fungos = (Myceteae),

7
Stramenopila e Protista. A variação morfológica é muito grande, existindo espécies
macro e microscópicas o que contribui para existência de muitos sistemas de
classificação que ainda hoje sofrem adições e alterações (PUTZKE, 2004). Com os
avanços nos estudos morfológicos, fisiológicos e de microscopia eletrônica entre
outros, surgiram novas propostas, sempre aceitas por alguns e rejeitadas por outros
(Tabela 1).
Tabela 1. Sistemas comparativos de classificação de fungos.
Fonte: Modificado de PUTZKE (2004)
Em relação ao seu habitat natural, os fungos apresentam-se como
cosmopolitas, ou seja, encontram-se distribuídos em todas as regiões do planeta,
porém de maior incidência em ambientes de clima tropical, quente e úmido. Podem
ser encontrados no solo, águas, sobre animais e vegetais, em alimentos naturais e
Hawkworth et al. (1983)
Hawksworth et al (1995),
Alexopoulos et al (1996)
Reino Fungi Divisão Myxomycota Divisão Eumycota Subdivisões:
Mastigomycotina
Zygomycotina
Ascomycotina
Basidiomycotina
Deuteromycotina
Reino Chromista Filo Hyphochytridiomycota Filo Labyrinthulomycota Filo Oomicota Reino Protozoa Filo Acrasiomycota Filo Dictyosteliomycota Filo Myxomycota Filo Plasmodiophoromycota Reino Fungi Filo Chytridiomycota Filo Zygomycota Filo Ascomycota Filo Basidiomycota
Reino Stramenopila Filo Oomycota Filo Hyphochytridiomycota Filo Labyrinthulomycota Reino Protistas Filo Dictyosteliomycota Filo Acrasiomycota Filo Myxomycota Filo Plasmodiophoromycota Reino Fungi Filo Chytridiomycota Filo Zygomycota Filo Ascomycota Filo Basidiomycota

8
industrializados (TEIXEIRA et al., 2001; BONONI,1999). São encontrados
praticamente em qualquer local do ambiente que nos cerca inclusive no ar onde
estruturas reprodutivas, na forma de esporos, podem desenvolver novas estruturas
vegetativas e reprodutivas quando encontram substrato adequado (PUTZKE, 2004).
3.2 Importância ecológica dos fungos
Apesar da utilização milenar dos fungos nos mais diversos ramos ainda é
comum serem associados a efeitos prejudiciais. A diversidade fúngica é muito ampla
sendo que existem algumas espécies que podem causar doenças aos animais, aos
vegetais e também ao homem, bem como a deteriorização de vários tipos de
materiais causando prejuízos econômicos (Figura 1). No entanto, a maioria das
espécies é benéfica para diversas áreas. (Figura 2).
Figura 1. Principais malefícios causados pelos fungos. Fonte: PUTZKE (2002)

9
Figura 2. Principais benefícios proporcionados pelos fungos. Fonte: PUTZKE (2002).
Os fungos juntamente com as bactérias heterotróficas são os principais
decompositores de materiais lignocelulósicos da biosfera. Vários fungos estão
associados a degradar dejetos e detritos de esgotos até um grau em que possam
ser considerados adubo ou substância equivalente. Ecologicamente podem ser
considerados os lixeiros do mundo, por degradarem todo tipo de restos orgânicos,
independente da origem, transformando-os em elementos assimiláveis pelas plantas
(NEUFELD,1997; PUTZKE, 2002, 2004). Como saprófitas, decompõem resíduos
complexos de plantas e animais, transformando-os em formas químicas mais
simples, que retornam ao solo. Tais substâncias são, então, absorvidas pelas

10
gerações vegetais subseqüentes. Desse modo, a atividade fúngica é amplamente
responsável pela fertilidade do solo.
3.3 Importância biotecnológica dos fungos
O interesse do homem pelos fungos vem desde a antiguidade em função da
eterna busca por alimento. Logo se descobriu que eles eram uma nova fonte de
alimentação, porém vários envenenamentos acidentais ocorreram e estes fungos
venenosos receberam o nome de “fermentos venenoso da terra” (MORAIS et al.,
2003). Muitos fungos já eram empregados desde a mais longínqua antiguidade,
quando o homem descobriu a metodologia e técnica de preparo do pão, do queijo e
das bebidas. Na época, ignorando o que provocava a fermentação, pelo menos o
pão e as bebidas alcoólicas surgiram praticamente em todos os cantos do planeta
independentemente. Durante centenas de anos, várias tribos indígenas aprenderam
com a natureza que determinadas espécies de fungos eram comestíveis, e de
excelente valor nutritivo. Europeus e asiáticos ainda hoje mantêm cogumelos em
suas dietas, especialmente porque a ciência tem descoberto qualidades ignoradas
por tanto tempo e que existem na maioria dos demais alimentos tais como: baixam o
nível de colesterol no sangue, têm substâncias anticancerígenas, são fontes de
aminoácidos (essenciais e não essenciais), contêm minerais como o cálcio, potássio,
iodo e fósforo, contêm várias vitaminas, nutrem e não engordam (PUTZKE, 2004).
Atualmente várias espécies de fungos são usadas em diversos tipos de
industria (alimentícia, farmacêutica, etc). Os cogumelos das espécies Agaricus

11
campestris, Morchella hortenois, Clavaria fava, Pleurotis sp. Boletus sp. e as trufas
do gênero Tuber sp., são os mais freqüentemente utilizados como alimentos. As
espécies de Penicillium roqueforti, Penicillium camemberti e Penicillium gordonzola
podem ser utilizados no preparo de queijos. Dentre as leveduras de maior
importância industrial destacam-se as linhagens fermentativas de Saccharomyces,
empregadas na indústria de panificação, e bebidas alcoólicas: Cerveja
(Saccharomyces cerevisiae e Saccharomyces carlsbetgensis), rum, vinho
(Saccharomyces ellipsoideus), saquê e produção industrial do etanol a partir da cana
de açúcar (NEUFELD, 1997).
A produção de moléculas com atividade farmacológica é uma das mais
exploradas em Biotecnologia, por empresas de países industrializados. Os
antibióticos são os produtos microbianos tradicionalmente mais explorados nesta
área, contudo novos compostos têm sido investigados em escala de screenig
farmacológico industrial, como agentes antitumorais e anti-câncer, inibidores
enzimáticos e agentes cardiovasculares. A aplicação de microrganismos na
produção de fármacos teve um desenvolvimento acelerado a partir da descoberta
dos antibióticos, na década de 30. A produção de penicilina de Penicillium notatun
ou o Penicillium chrysogenum, revolucionou toda a terapêutica antimicrobiana,
possibilitando a descoberta de outras drogas desse tipo, algumas com
especificidade para fungos, bactérias, protozoários, etc. (LACAZ, 2002). A tabela 2
mostra exemplos de vários fungos com esse potencial.

12
Tabela 2 - Exemplos de alguns antibióticos e microrganismos a eles relacionados.
Microrganismos Antibioticos
Aspergillus clavatus Clavalina
Aspergillus flavus Acido aspergílico
Aspergillus flavus, A. giganteus e A. parasiticus Flavicina
Aspergillus fumigatus Fumigacina (Acido belvólico)
Cephalosporium salmosynne Cefalosporina N
Chaetomium cochioides Chaetomina
Chaetomium iodinium Iodinina
Chaetomium violaceum Violaceína
Gliocadium sp. Gliotoxina
Micromonospora sp. Micronomosporina
Penicillium citrinum Citrinina
Penicillium cyclopium e Penicillium puberulum Ácido penicílico
Penicillium notatum Penicilina
Penicillium spinulosum Espinulosina
Proactinomyces albus e P. violaceus Proactinilomicina, Actinomicetina
Streptomyces antibioticus Actinomicina
Streptomyces levendulae Estreptomicina
Streptomyces griséus Estreptomicina
Streptomyces aureofaciens Clortetraciclina
Streptomyces venezuelensis Cloranfenicol
Streptomyces rimosus Oxitetraciclina
Streptomyces noursei Fungicidina
Streptomyces bachijoensis Tricomicina ( Cabimicina)
Streptomyces nodosus Anfotericina A e B
Fonte: LACAZ, 2002

13
3.4 Fungos deterioradores de madeira
Os fungos são capazes de atacar e degradar a madeira, em seus
ecossistemas terrestres naturais e especialmente os da classe Basidiomycetes tem
demonstrado maior eficiência no processo de biodegradação de material
lignocelulósicos (HATAKKA, 1994; HIGUCHI 1990, Apud SOARES, 1998). Estes
fungos podem ser saprófitas, parasitas, simbióticos e micorrízicos, sendo usados na
indústria alimentícia, na produção de enzimas, na biopolpação, no
biobranqueamento de efluentes, na formação de ectomicorrizas, etc. (HAMLYN et
al.,1998).
Hoje, acredita-se que existam mais de 200.000 espécies de Basidiomicetos
ligninocelulósicos, a maioria dos quais estariam nas regiões tropicais e subtropicais
do mundo. Os basidiomicetos lignocelulósicos estão classificados em duas grandes
ordens dos basidiomycetes: Agaricales e Aphyllophorales. Estes apresentam
tamanhos, formas, cores, e consistências diversificadas, podem ser microscópicos
ou ter até 20cm de diâmetro, presos ao substrato através de estirpe, sésseis ou na
forma de manchas regulares ou irregulares na superfície da madeira. São versáteis
e, num mesmo tronco, uma espécie pode apresentar mais de uma forma ou cor.
Crescem inicialmente dentro da madeira colonizando-a e suas hifas formam uma
rede, nem sempre visível a olho nu, mais ou menos extenso, preenchendo o lume
das células, passando de uma célula a outra através da parede celular e em alguns
casos destruindo a estrutura da lamela média. Após certo período de tempo, que
pode durar uns poucos dias até muitos anos, variável de espécie para espécie e
com as condições ambientais, emergem os basidiocarpos (corpos de frutificação).

14
São extremamente importantes como decompositores e principais responsáveis pela
ciclagem do carbono nos ecossistemas. Degradam os componentes da madeira:
celulose, hemicelulose e lignina, a partir dos quais obtêm energia para seu
crescimento e reprodução (BONONI, 1997).
Conforme o tipo de degradação que causam os fungos degradadores de
madeira são classificados em: a) Fungos de podridão branca (“white-rot”), os quais
atacam indistintamente os três principais componentes da madeira (celulose,
hemicelulose e lignina); b) Fungos de podridão parda (“brow-rot”), os quais
degradam a porção polissacarídica da madeira, causando poucas modificações na
lignina, sem alteração do anel aromático; c) Fungos de podridão mole (“soft-rot”), os
quais degradam lignina mais lentamente do que os polissacarídeos, produzindo um
amolecimento característico da superfície da madeira (RODRIGUEZ, 1990).
3.5 Estrutura da parede celular da madeira
A madeira é um biopolímero tridimensional, composto principalmente de
celulose, hemicelulose e lignina. Estes polímeros constituem a parede celular da
madeira e são responsáveis pelo comportamento da maioria de suas propriedades
tecnológicas. A parede celular vegetal é, por sua vez, composta de uma parede
celular primária e uma parede celular secundária. A parede celular primária, mais
externa, é uma camada fina, formada durante o desenvolvimento da célula. Sua
espessura é estimada em cerca de 0,1μm, e contém cerca de 9% de celulose

15
incrustada em uma matriz plástica amorfa de hemicelulose, pectina e lignina e cerca
de 70% ou mais de água (PRESTON,1974 apud CASTRO E SILVA, 1996).
A parede secundária, por outro lado, tem suas microfibrilas celulósicas
depositadas de forma espiralada com um ângulo com cerca de 50 – 70º em relação
ao eixo longitudinal da célula. Quando são formadas novas camadas, esta
orientação modifica-se e as microfibrilas começam a ser depositadas
espiraladamente em um ângulo menor que varia de 5 – 30º. Antes do
desenvolvimento final da célula ocorre nova mudança na orientação das
microfibrilas, com as últimas camadas dispostas similarmente às primeiras poucas
camadas, e cerca de 60 – 90º em relação ao eixo longitudinal. Assim, a parede
secundária possui três camadas mais ou menos distintas: camadas S1, S2 e S3
(Figura 3), compostas de diferentes proporções de celulose, lignina e hemicelulose
(CASTRO E SILVA et al., 1992).
Figura 3. Esquema tridimensional de uma parede celular vegetal.
Fonte: Adaptado de Côté (1967).

16
A camada S1 é relativamente fina, com cerca de 0,1 – 0,3 μm de espessura.
Nesta camada o arranjo das microfibrilas forma um ângulo entre 50 – 70º em relação
ao eixo da célula considerada. A camada intermediária S2 forma a porção principal
da célula, com espessura variando de 1 a 5 μm. O ângulo microfibrilar varia entre 10
– 30º em relação ao eixo, diminuindo com o aumento do comprimento da fibra. A
camada S3, mais interna, tem 0,1 μm de espessura e o ângulo das microfibrilas
varia muito, conforme a espécie de madeira considerada, oscilando entre 50 – 90º
(LEPAGE et al., 1986).
3.6 Composição química da parede celular
A composição química dos tecidos que compõem a madeira apresentam uma
variação quando comparados estruturalmente. A zona cambial, por exemplo, tem
uma composição química diferente daquela encontrada na região do cerne e
alburno. Na parede celular os componentes químicos da madeira apresentam-se
distribuídos em proporções diferentes nas diferentes camadas que compõem a
parede celular vegetal. A celulose, por exemplo, está presente em pequena
quantidade na lamela média, aumentando em sua proporção ao peso seco da
parede celular através do centro da camada S2 e decrescendo a partir deste ponto
até a camada S3. A hemicelulose ocorre em grande quantidade, especialmente na
camada S2. A lignina é dominante na lamela média composta. Como essa camada é
muito fina, proporcionalmente, a maior parte de lignina é encontrada na parede
secundária (Figura 3) (PRESTON, 1974 apud CASTRO E SILVA, 1996).

17
3.6.1 Celulose
A celulose é um polímero homogêneo não ramificado, cuja cadeia é
constituída de centenas de unidades monossacarídicas, de apenas um tipo, D.
glicose, ligadas covalentemente por ligações glicosídicas do tipo β . (1→4) (Figura
4). Os dois resíduos terminais de glicose da cadeia polimérica de celulose diferem
entre si pela natureza redutora de uma das extremidades. (SOARES, 1998).
Figura 4. Fragmento de celulose.
Fonte: ÂNGELO (2004).
Na madeira as moléculas de celulose compõem as microfibrilas, as quais por
sua vez são organizadas em um arranjo paralelo, formando fibrilas que se
entrelaçam, originando então as fibras de celulose. (FENGEL e WEGENER, 1989).
Duas regiões morfologicamente diferentes podem ser bem caracterizadas, quando
se analisa a fibra de celulose por difração de raios-x: uma região cristalina,
altamente organizada e uma região amorfa, desordenada. As propriedades físicas
das fibras de celulose, tais como resistência à tração mecânica, alongamento e
capacidade de sovatação, dependem do grau de organização (grau de
cristalinidade) das mesmas (DALMEIDA, 1982).

18
3.6.2 Hemicelulose
Constituem um grupo de substâncias poliméricas compostas de unidades
monossacarídicas diferentes. São polissacarídeos heterogêneos, contendo nas suas
estruturas tanto hexoses, quanto pentoses. Possuem cadeia ramificada e de baixa
massa molecular. A ligação glicosídica mais freqüente nas hemiceluloses é a ligação
do tipo β (1→4), porém ligações do tipo β (1→3) e α (1→4), também podem ser
encontradas (CHUA et al., 1982, 1983).
3.6.3 Lignina
Lignina é uma substância cuja molécula é extremamente complexa,
polimérica, heterogênea, composta basicamente de três tipos diferentes de unidades
monoméricas derivadas de 4- hidroxiarilpropernil álcoois, sendo eles: álcool trans-
para-cumárico (a) (unidade hidroxifenil), álcool trans-coniferílico (b) (unidade
guaiacil) e álcool trans-sinapílico (c) (unidade siringil) (Figura 5) (DALMEIDA, 1982;
FENGEL e WEGENEER, 1989; HIGUCHI, 1990).Tais unidades são distribuídas ao
acaso e ligadas covalentemente por pelo menos dez tipos de ligações químicas
diferentes. Estas incluem ligações tipo éter, alquil-aril, alquil-alquil e aril-aril, sendo
que a mais freqüente é a ligação do tipo β - 0 - 4, representando em torno de 50%
das ligações. São freqüentes também ligações do tipo – C – C –, como as ligações
difenil e diarilpropano, particularmente a ligação β – 1 (Figura 6). Portanto, os
grupamentos funcionais que, definem as características estruturais e químicas da
lignina são os grupos hidroxílicos de álcoois primários e secundários, hidroxílicos de

19
fenóis, eterificados e livres, grupos carboxílicos de diferentes tipos, metoxilas ligadas
a anéis aromáticos e insaturações. O caráter aromático da estrutura da lignina está
em torno de 51% (FENGEL e WEGENEER, 1989; HIGUCHI, 1990).
Figura 5. a) Álcool trans-p-cumárico (grupo p-hidroxifenil), b) Álcool trans-
coniferílico (grupo guaiacil) e c) Álcool trans-sinapílico (grupo siringil)
FREUDENBERG (1965).
A lignina apresenta uma molécula tridimensional estéreo quimicamente,
bastante complexa, amorfa oticamente inativa. Possui diferentes características
estruturais em função do tipo de madeira e diferentes regiões de um mesmo tecido.
Em geral, ligninas encontradas em madeiras de coníferas apresentam um alto teor
de álcool coniferílico (unidades guaiacil); ligninas encontradas em madeiras de
folhosas, apresentam quantidades equivalentes de álcool coniferílico e álcool
sinápilico (unidade siringil). Com base nestas constatações experimentais foram
sugeridos os termos lignina guaiacil e lignina guaiacil-siringil, que se denominam
respectivamente os dois tipos principais de lignina, quanto a sua posição (FENGEL
& WEGENEER, 1989).

20
Figura 6. Estrutura da lignina em folhosa Fonte: NIMZ (1974).
Na madeira, como já citado anteriormente, a lignina pode ser encontrada nas
paredes primária e secundária das células, as quais detêm a maior fração da
quantidade total de lignina (70 - 90%) (Figura 7). Também é encontrada nos espaços
intercelulares, lamela média, que individualmente possui a maior concentração de
lignina, comparativamente às outras partes de um mesmo tecido. Nas paredes
celulares, a lignina aparece ligada a hemicelulose, formando uma espécie de
“cimento” no qual as fibras de celulose estão embebidas (Figura 8) (FENGEL e
WEGENER, 1989; ESAU, 1965). Na lamela média, forma uma espécie de retículo
com cadeias paralelas e intercruzadas, conferindo propriedades adesivas que
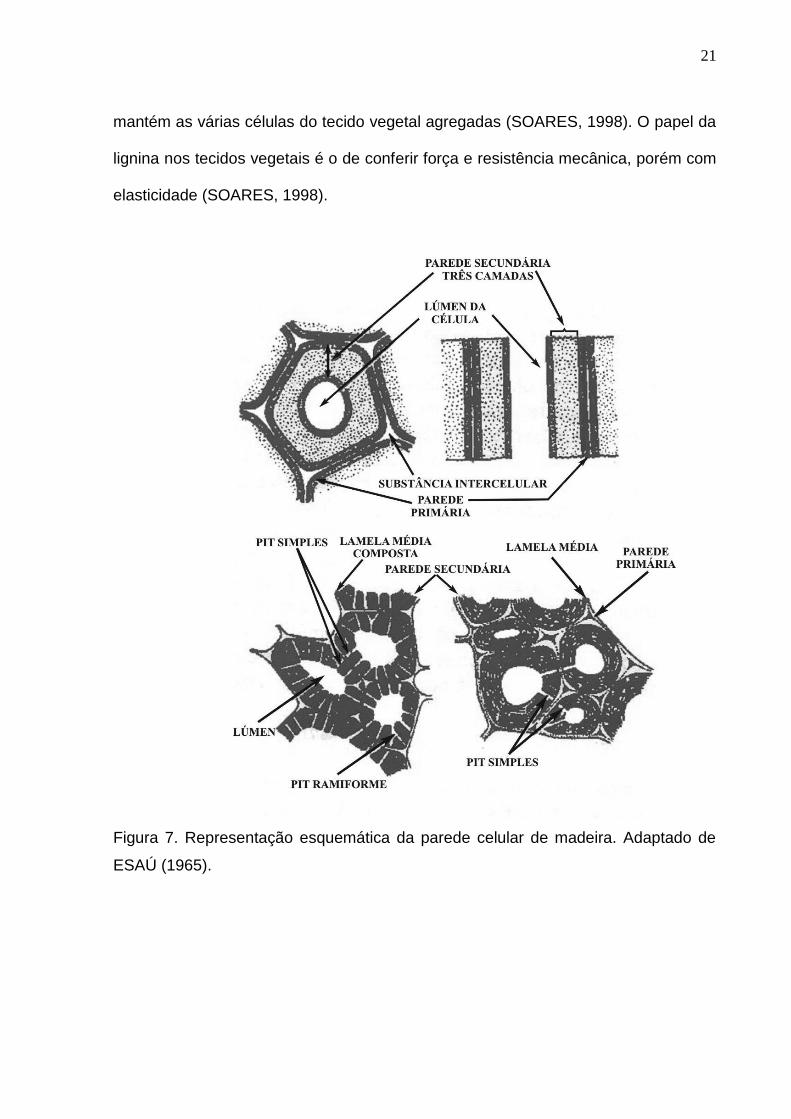
21
mantém as várias células do tecido vegetal agregadas (SOARES, 1998). O papel da
lignina nos tecidos vegetais é o de conferir força e resistência mecânica, porém com
elasticidade (SOARES, 1998).
Figura 7. Representação esquemática da parede celular de madeira. Adaptado de
ESAÚ (1965).

22
Figura 8. Sistemas de camadas na parede da célula de madeira: a) corte
ilustrativo; b) microscopia eletrônica de transmissão mostrando lamela média
(ML), a parede celular (S1, S2 e P e T) e o lume (W) das células de madeira.
Reproduzido com modificações de FENGEL e WEGENER (1989).
3.7 Enzimas Ligninolíticas
As pesquisas sobre a biodegradação da lignina têm se revestido de grande
interesse, em vista deste composto químico ser o mais abundante material renovável
próximo da celulose. Os fungos decompositores de madeira, mais especificamente
os de podridão branca, são organismos capazes de degradar a lignina para CO2 e
H2O, de modo que esses fungos usam este substrato como única fonte de carbono e
energia (KIRK & FARRELL, 1987). Portanto, estudos sobre a biodegradação da
lignina têm sido conduzidos principalmente usando-se fungos de podridão branca,
os quais produzem enzimas extracelulares modificadoras de lignina. Para
depolimerizar e mineralizar a lignina, esses fungos possuem um sistema oxidativo
não específico, que inclui várias oxidoredutases extracelular e metabólicos de baixo
peso molecular (SAPPARAT et al., 2002). Enzimas extracelulares envolvidas na
degradação da lignina e xenobióticos por fungos de podridão branca incluem vários

23
tipos de lacases, peroxidases e oxidases produtoras de H2O2 (CAMARERO et al.,
1999; VOLC et al., 1996; THURSTON,1994).
3.7.1 Fenoxidase (Lacase e Peroxidase)
3.7.1.1 Lacase (E.C 1.1.0.3.2)
A composição enzimática do sistema ligninolítico depende da espécie do
fungo, com a lacase sendo o componente comum (HATAKKA, 1994). Lacases são
principalmente glicoproteínas extracelulares ligadas ao cobre bivalente, o qual é
reduzido durante a oxidação de fenóis (Figura 9) e subseqüentemente oxidado
novamente ao estágio bivalente pelo oxigênio (FORSS et al., 1989). Lacase são
enzimas monoméricas, diméricas ou tetraméricas com pesos molecular entre 60 e
80 kDa e com teor de carboidrato entre 15 e 20% (THURSTON,1994).
Vários estudos têm demonstrado a participação da lacase em eventos
ligninolíticos significativos que incluem a oxidação de unidades não- fenólicas da
lignina, a geração de H2O2 requerida tanto para as atividades peroxidases como para
a formação de radical oxídrico (OH), e a produção de Mn3+ a partir de Mn2+ presente
nos lignocelulósicos.

24
Lacase (Cu2+)
Lacase red. (Cu1+)
O2
H2O PhOH
PhO•
Figura 9. Esquema de mecanismo de ação das lacases.
Fonte: FERRAZ (2004)
As lacases catalizam a polimerização oxidativa das unidades fenólicas da
lignina e uma variedade de compostos fenólicos e aminas aromáticas, os quais são
facilmente removidos por filtração. Sob certas condições clivam um grande número
de compostos fenólicos sintéticos e biológicos (ROY-ARCAND & ARCHIBALD,
1991). A reação normalmente envolve um mecanismo de remoção de um radical
mesomérico.
A seqüência de aminoácidos da lacase contém resíduos de uma cisteína e
dez histidinas os quais estão envolvidos na ligação dos átomos de cobre
(THURSTON,1994).
3.7.1.2 Peroxidase
Em um ciclo catalítico normal, a forma férrica da enzima é oxidada por
peróxido de hidrogênio para oxi-ferril, estado conhecido como Composto I. Este é
reduzido através de duplas transferências consecutivas de um elétron para o
Composto II (Figura 10). Fenóis ou aminas aromáticas são os típicos doadores de
elétrons para peroxidases, transformando-se em radicais livres, os quais
subseqüentemente derivam produtos estáveis (ESPÓSITO, 1992). As peroxidases
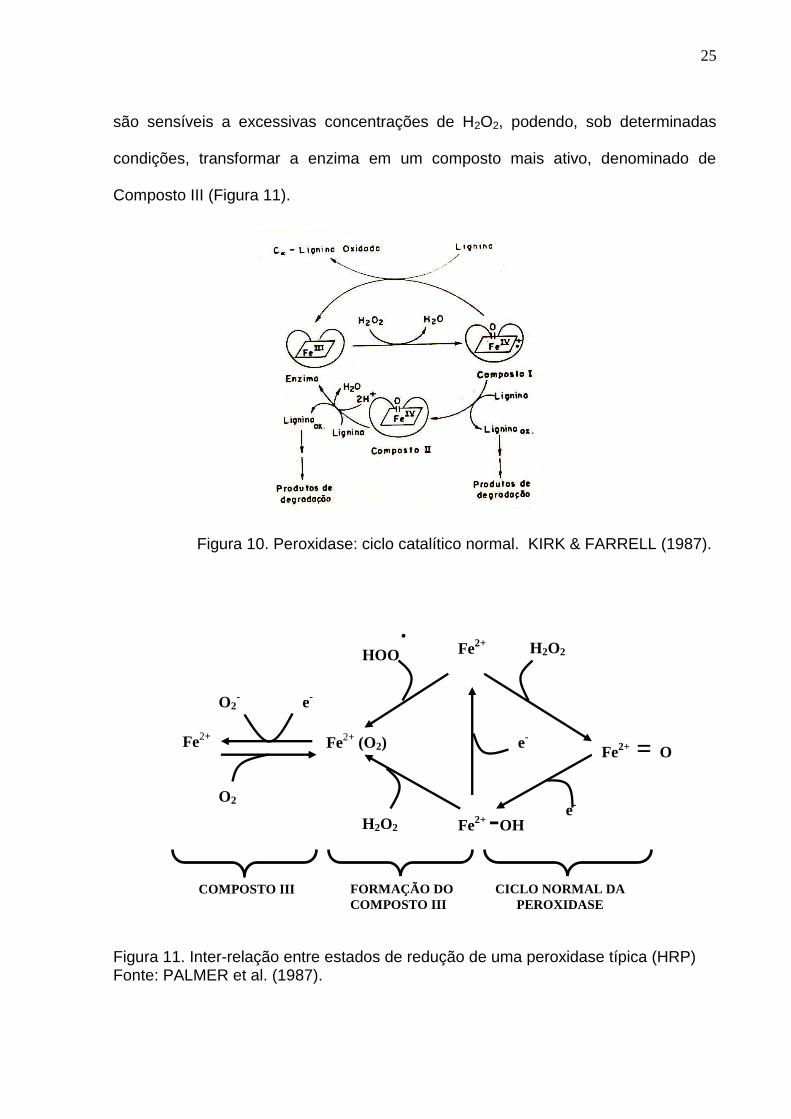
25
são sensíveis a excessivas concentrações de H2O2, podendo, sob determinadas
condições, transformar a enzima em um composto mais ativo, denominado de
Composto III (Figura 11).
Figura 10. Peroxidase: ciclo catalítico normal. KIRK & FARRELL (1987).
Figura 11. Inter-relação entre estados de redução de uma peroxidase típica (HRP) Fonte: PALMER et al. (1987).
COMPOSTO III
H2O2
H2O2
HOO·
e-
e-
O2
e- O2
-
Fe2+ -OH
Fe2+
Fe2+
(O2)
Fe2+
Fe2+
= O
FORMAÇÃO DO
COMPOSTO III
CICLO NORMAL DA
PEROXIDASE

26
Tanto a peroxidase como a lacase oxidam compostos fenólicos. Os produtos
resultantes da oxidação polimerizam espontaneamente, formando complexos
insolúveis, os quais podem ser removidos por precipitação, filtração ou centrifugação
(DAVIS & BURNS, 1978).
3.7.2 Lignina – Peroxidase ou Ligninase (LiP) ( E.C.1.11.1.14)
A lignina – peroxidase foi descoberta em cultura do fungo Phanerochaete
chrysosporium (GLENN, 1983; TIEN & KIRK,1983), e estaria relacionada a oxidação
de compostos monoméricos e diméricos derivados da lignina e despolimerização
parcial da mesma em presença de peróxido de hidrogênio.
As ligninases são proteínas N-monoméricas e O-glicosiladas com quatro
ligações dissulfêto, dois íons de cálcio e um de ferro protoporfirina IX como grupo
prostético.
As ligninases são uma classe de isoenzimas, de peso molecular variando de
38 a 43 kDa e ponto isoelétrico 3,5 (TIEN & KIRK, 1984; GOLD et al., 1984) que
requerem H2O2 para sua atividade. Na reação com H2O2 LiP forma um radical cátion
porfirina . LiP cataliza oxidações de um elétron de compostos fenólicos e não
fenólicos com potencial de redução de até 1,5 V ( KERSTEN et al., 1990) gerando
radicais fenox e radicais cátion aril. O principal produto de reação da LiP com H2O2 é
a LiP I intermediária (Figura 12). LiP I é reduzida via transferência de um elétron com

27
LiP II como intermediária. Neste ciclo catalítico o aromático reduzido, por exemplo,
álcool veratrílico, é oxidado para via radical cátion aril (HIGUCHI, 1989).
B
A
OCH3
HO
OH
OCH3O
OCH3
OCH3
OO
OCH3
OCH3
CH3
O
+OCH3
O
OCH3
OCH3
O
O
OCH3
HO
CHO
+
OHO
OH
OCH3
+O
OH
H
OCH3
OCH3
HO
OH
OCH3O
OCH3
OCH3
A1
A2 B1
Figura 12. Modelo de degradação da lignina por Li-P de Phanerochaete chrysosporium /H2O2. Fonte: KIRK e CULLEN (1998).
Portanto, as ligninases apresentam uma ampla especificidade para uma série
de compostos aromáticos, sendo capazes de oxidar fenóis a quinonas, sugerindo
um acoplamento oxidativo, permitindo a polimerização (CAMMAROTA, 1991).
3.7.3 Manganês–Peroxidase (Mn–P) (E.C.1.11.1.13)
Assim como para as ligninases as Mn–P também foram identificadas em
culturas de P. chrysosporium por Kuwahara et al. (1984), sendo dependente de
manganês e de H2O2 para sua atividade. São glicoproteínas com peso molecular
entre 43 – 49 kDa e ponto isoelétrico entre 4,2 e 4,9, contendo um mol de ferro

28
porfirinico IX / mol de enzima (GLENN & GOLD, 1985). Essas enzimas identificam-
se em torno de 57% com as LiP (SUNDARAMOORTHY et al., 1994). Em contraste
com as LiP, Mn-P contém um “loop” extra próximo ao sítio ativo e um único sítio é o
sítio de ligação do substrato proposto (MnII).
O principal produto da reação Mn(II) o qual é oxidado pela MnPI e MnPII para
Mn(III). Mn (III) é estabilizado formando complexos com ácidos orgânicos, por
exemplo, lactato ou oxalato e então oxida um substrato aromático gerando radicais
fenox ou radicais cátion aril (Figura 13). Tanto lignina peroxidase com manganês
peroxidase, são importantes na descoloração de efluente Kraft (MICHEL et al.,
1991).
Figura 13. Mn (II) é oxidada a Mn (III) por uma peroxidase dependente de Mn.
As enzimas ligninolíticas apresentam um grande potencial na
biotransformação de ligninas e seus derivados como também nos aspectos
ambientais (biorremediação). Essas enzimas têm um papel importante na
descontaminação de efluentes papeleiros e têxteis. Atualmente são utilizadas em
biossensores para detectar fenóis em ambiente contaminado (DURAN &
ESPOSITO, 2000).
2H+ + O2 + Mn
2+ Mn
3+ + H2O2

29
3.8 Aplicações ambientais dos fungos ligninolíticos.
Um dos grandes problemas que o mundo industrializado vem enfrentando nos
dias atuais é a contaminação de solos, lençóis freáticos e sedimentos por compostos
orgânicos tóxicos. Estes poluentes são uma potencial ameaça à saúde pública e ao
ambiente. Estima-se que o mercado de remediação de solo está em torno de US$
16 milhões/ano (VANDEVIVERE & VERSTRAETE, 2001). Os contaminantes mais
comuns são compostos organoclorados, hidrocarbonetos, policlorobifenóis e metais.
A biorremediação está se tornando muito pesquisada e testada principalmente no
caso de poluição por hidrocarbonetos. Os sistemas mais utilizados na
biorremediação são os microbiológicos, principalmente, bactérias, fungos
filamentosos e leveduras (SILVA & ESPOSITO, 2004).
Fungos ligninolíticos, onde se incluem os conhecidos fungos de podridão
branca, além de utilizar a citocromo P-450 monoxigenase e a epóxido hidrolase,
para degradar hidrocarbonetos aromáticos (BEZALEL et al, 1996), também utilizam
enzimas extracelulares ligninolíticas que conferem uma maior tolerância aos
poluentes em concentrações que normalmente seriam tóxicas para outros
microorganismos. Essas enzimas que estão envolvidas na degradação de ligninas,
incluem lacase, lignina peroxidase e manganês peroxidase; têm a capacidade de
mineralizar estes compostos (Figura 14), sendo esta uma grande vantagem para a
biorremediação (SILVA & ESPOSITO, 2004).

30
Figura 14. Reação das enzimas ligninolíticas na transformação e mineralização de hidrocarbonetos aromáticos. Fonte: CERNIGLIA (1997).
Outro aspecto do uso de fungos ligninolíticos na despoluição ambiental é o
tratamento biológico de efluentes. A industrialização da polpa e papel Kraft produz
uma grande quantidade de resíduos sólidos, líquidos e gasosos, extremamente
danosos ao ambiente. Neste contexto, a utilização de fungos ligninolíticos como um
método alternativo no tratamento de efluentes líquidos provenientes desse processo
parece ser de extrema relevância, tendo em vista que esses microrganismos
“trabalham praticamente de graça” e com extrema rapidez. Os processo de obtenção
de polpas celulósicas e de fabricação de papel são responsáveis pela maior carga
poluente, que atinge os ecossistemas na forma de efluentes ou resíduos sólidos,
altamente tóxicos e muitas vezes, mutagênicos, persistentes e/ou bioacumulativos
(ERIKSSON, 1990; MEHTA & GUPTA, 1991; ONYSKO, 1993 apud DURAN, 1997).
A utilização de fungos objetiva a descontaminação dos efluentes. A eficiência
desses processos esta diretamente relacionada à capacidade do fungo em degradar
a lignina (MEHTA & GUPTA, 1991; ONYSKO, 1993 apud DURAN & ESPOSITO,
1997).
Os efluentes podem causar toxicidade aguda ou crônica, mutagenicidade,
deficiência de oxigênio, eutroficação nos corpos de água receptores e, por
R
quinona
H2O
Enzimas
ligninolíticas
Clivagem do anel
aromático CO2 + H2O

31
conseqüência, modificações nas comunidades de plantas e animais (KAUSTSKY,
1992; LEONARDSSON, 1993 apud DURAN & ESPOSITO, 1997). A capacidade de
determinados fungos de descolorir efluente Kraft é amplamente documentada
(LANKINEN et al., 1991; CASTRO E SILVA et al., 1993: CAMMAROTA &
SANTANNA, 1992; FITZSIMONS ERIKSSON, 1990 dentre vários outros autores). O
fungo Phanerochaete chrysosporium foi testado no processo Mycor e proporcionou
uma redução marcado do cloro orgânico total (TOCl) em 70% e da cor em 80%
(CHANG et al., 1987). Entretanto, o alto consumo de oxigênio, o tempo de
resistência restrito e a perda de biomassa fúngica tornam este processo inviável
economicamente (CAMMAROTA & SANTANNA, 1992).
Vários estudos mostram a capacidade do fungo Phanerochaete
chrysosporium em degradar vários tipos de compostos aromáticos clorados e outros
organopoluentes (HAMMEL, 1989). O fungo Coleomycete, Stagonospora gigaspora
é capaz de mineralizar e solubilizar compostos derivados de lignina, removendo 90%
da cor de efluente de branqueamento CEH (BERGBAUER et al., 1992). O fungo
Trametes versicolor é capaz de reduzir consideravelmente organoclorados tóxicos
(PAICE et al.,1989; REID et al., 1990; ROY–ARCAND & ARCHIBALD, 1991).Em
outro estudo verificou-se que o fungo Lentinus edodes removeu 73% da cor de
efluentes fenólicos industriais e 30% de fenóis em 120 horas, sem fonte adicional de
carbono. (ESPÓSITO, 1992)
Considerando a grande diversidade de fungos no mundo, é possível antecipar
a existência de um grande número desses organismos com capacidade em degradar
poluentes e, portanto, com potencial para a micorremediação. Deve-se inclusive

32
levar em conta que grande parte dessa diversidade, se encontra em países tropicais
dos quais o Brasil é um dos maiores reservatórios (SILVA; MINTER, 1995). É
fundamental que pesquisas que descrevam e avaliem esse potencial sejam
efetivadas.

33
4 MATERIAIS E MÉTODOS
4.1 Coleta dos Fungos
Os fungos Pycnoporus sanguineus e Trametes sp com seus respectivos
substratos (porção de madeira usada como fonte de nutriente do fungo) foram
coletados, com auxílio de um facão, na BR 174 Km 41 (Amazonas) e no perímetro
urbano no Município de Manacapuru nos meses de fevereiro - maio de 2004 e
acondicionados em sacos de papel com as devidas informações sobre local, data de
coleta, coletor e tipo de substrato. Em seguida foram levados ao Laboratório de
Microbiologia da UTAM/EST-UEA, onde foram armazenados em câmara de
refrigeração científica em temperatura de 3.5ºC (INDREL RC 1500 D). No dia
seguinte após a coleta, os fungos foram inoculados em meio de cultura para em
seguida serem identificados e utilizados na pesquisa em questão.

34
4.2 Isolamento dos fungos
Utilizando-se um estilete (bisturi), foi retirado de cada carpóforo cerca de 3mm
de uma de suas extremidades. A amostra do fungo foi submetida à assepsia em
álcool a 70%, hipoclorito a 3% e água destilada, por um período de 1–2 minutos em
cada solução. Em seguida cada amostra foi colocada sobre filtro de papel para
retirar o excesso de líquido presente. Posteriormente cada fragmento fúngico já
devidamente desinfetado e seco foi inoculado no centro de cada placa de Petri, com
o meio de cultura ágar-extrato de malte, procedimento este executado em ambiente
estéril (interior da Câmara de Fluxo Laminar). As placas de Petri foram identificadas,
lacradas com fita plástica (Parafilm “M”) e deixadas em temperatura ambiente 28º C
( 2ºC) por um período de sete dias. Decorrido esse período, foi realizada a
repicagem no mesmo tipo de meio para a obtenção de uma cultura pura, sendo que
a cultura de Trametes sp foi identificada com o código FSF11 e o P. Sanguineus
com o código PIC07. As duas culturas foram depositadas em câmara de refrigeração
científica (INDREL RC 1500 D) em temperatura de 3.5ºC.
4.3 Otimização das Condições de Crescimento do Fungo
4.3.1 Temperatura
No estudo do crescimento dos fungos Pycnoporus sanguineus e Trametes sp,
objeto desta pesquisa, foi utilizado o método da placa de Petri em triplicata, no qual

35
o avanço da fronteira micelial foi medido em função do tempo.
Para o estudo sobre a influência da temperatura no crescimento micelial, foi
retirado da periferia da cultura pura cerca de 5mm do micélio de Pycnoporus
sanguineus e Trametes sp os quais foram inoculados em placas de Petri (triplicata)
contendo meio Ágar –malte (2%). Estas culturas foram mantidas em incubadoras
com temperaturas pré-estabelecidas de 25º, 31º e 35º C e em temperatura ambiente
de 28ºC ( 2ºC). O crescimento foi mensurado através da progressão linear da
fronteira micelial, sendo as medidas tomadas em duas direções a cada 24 horas,
durante 12 dias. A mensuração do raio dessa colônia permite avaliar o crescimento
micelial, o qual ocorre pela extensão da ponta da hifa, enquanto que as partes mais
antigas das hifas são incapazes de crescimento. Embora a hifa mais antiga não seja
capaz de crescer, tem um importante papel de suportar o crescimento da ponta à
medida que novo protoplasma é formado através da hifa e transportado para a ponta
pela corrente citoplasmática ativa (MOORE-LANDECKER, 1982).
4.3.2 pH
Cada fungo foi inoculado em placa de Petri (triplicata) contendo o meio ágar-
extrato de malte. O pH do meio foi ajustado para a faixa de 4,0 – 7,0 utilizando-se
para cada caso NaOH 1,0 M ou H2SO4 1,0 M. Para evitar a degradação do ágar
abaixo do pH 5,0 o ajuste foi realizado após a autoclavagem. Procedida à
inoculação, as culturas foram mantidas em incubadora a 31º C, durante 12 dias.
Assim como realizado para a temperatura, o avanço da fronteira micelial foi
mensurado em duas direções a cada 24 horas, no período de 12 dias.

36
4.3.3 – Fonte de Nitrogênio
No estudo da fonte de nitrogênio mais adequada ao crescimento de
Pycnoporus sanguineus e Trametes sp utilizamos nitrato de sódio (NaNO3) e sulfato
de amônia ((NH4)2SO4)), as quais foram adicionadas ao meio ágar-malte nas
concentrações de 30mEq/L, 60mEq/L, 90mEq/L, 120mEq/L e 150mEq/L (fonte de
nitrogênio na forma de NO3- ou NH4
+, MMNO3- e MMNH4
+ respectivamente),
acrescidos de 2% de glicose como fonte de carbono. Após a inoculação de cerca de
5mm do micélio de cultura pura, as placas foram lacradas, identificadas e mantidas
em temperatura ambiente em torno de 28ºC durante 12 dias. O experimento foi
realizado em triplicata para cada concentração e também para a testemunha. O
avanço da fronteira micelial foi mensurado diariamente no período de 12 dias.
4.4 Taxa de Crescimento dos Fungos
A taxa de crescimento foi definida como a razão entre a média da progressão
da fronteira micelial e o número de dias de crescimento do fungo.
4.5 Análise Estatística
Para a análise estatística dos dados utilizou-se o software STATISTICA 6.0,
onde se obteve os parâmetros descritivos (média, desvio padrão, variância) dos
dados, teste de correlação entre as variáveis, e o teste ANOVA e de Tukey para
contrastes das médias.

37
4.6 Determinação das Atividades Enzimáticas
4.6.1 Meio de cultura e inóculo
Os fungos foram cultivados em meio líquido contendo:
20g de extrato de malte;
antibiótico;
1000mL de água destilada.
fonte de carbono: dextrose e serragem de madeira Hura creptans.
O meio de cultura foi previamente esterilizado em autoclave à 120º C por 15
minutos, deixando-se esfriar até uma temperatura adequada para manuseio. Em
seguida, em ambiente asséptico, colocou-se 150mL de meio de cultura em cada um
dos quatro recipientes (erlenmayer). Em dois erlenmayer adicionou-se 3g de
dextrose e nos outros dois 3g de serragem de Hura creptans, como fonte de
carbono. Posteriormente procedeu-se a inoculação dos fungos (PIC07 e FSF11)
utilizando-se cerca de 2 retângulos (1,5cm x 1,0cm) de micélio por frasco de 7 – 10
dias o qual foi retirado da região periférica da placa pois o crescimento é restrito a
extremidade da hifa . O experimento foi repetido sem adição de fonte de carbono o
qual foi usado como testemunha. Cada erlenmayer foi etiquetado (identificação) e
devidamente fechado com tampão fixado com fita crepe.
Os recipientes acrescidos de fonte de carbono e a testemunha foram
mantidos em cultura estacionária em temperatura ambiente (cerca de 26ºC 2)

38
sendo as mesmas cultivadas por um período de cinco dias, a pH inicial em torno de
5,6 - 6,0. O experimento foi repetido sob agitação a 180 rpm em temperatura
ambiente (cerca de 26ºC 2). Decorrido cinco dias de cultivo dos fungos, procedeu-
se à determinação das atividades enzimáticas em dias alternados, até o décimo
oitavo dia após a inoculação dos mesmos.
4.6.2 Fenoxidases (Lacase e Peroxidase)
A atividade enzimática para fenoloxidase e lacase foi determinada em caldo
de cultivo extrato de malte acrescido de 2% de glicose ou serragem da madeira de
Hura crepitans como fonte de carbono.
Para a atividade lacase foi utilizado a metodologia proposta por Ander &
Eriksson (1976). O método baseia-se na oxidação do substrato enzimático de
siringaldazina para sua forma de quinona, que apresenta absorção a 525 nm (ε =
65000 M-1 cm-1). Uma unidade de atividade de lacase ou peroxidase corresponde à
quantidade de enzimas necessárias para oxidar 1μmol de substrato por minuto.
Para a determinação da atividade lacase utilizou-se: 50μL de caldo de cultura
filtrado, 0,95 mL de tampão tartarato de sódio a pH = 4.2, 1 mL de siringaldazine
(solução estoque 5 mg/10 mL de etanol), 1 mL de H2O destilada.
Para atividade fenoxidase utilizou-se a metodologia da oxidação do substrato
enzimático siringaldazine que apresenta absorção a 460nm (ε = 29.400). Uma
unidade de atividade fenoxidase corresponde a quantidade de enzimas necessárias

39
para oxidar 1μmol de substrato por minuto. Para a determinação da atividade
fenoxidase utilizou-se: 0,95 mL de tampão tartarato de sódio a pH 4,2, 1 mL de H2O2
(2,0mM ), 50μL de caldo de cultura filtado, 1 mL de siringaldazine (solução estoque
5mg/10 mL de etanol). A determinação da Peroxidase foi feita pela diferença da
Fenoxidase menos a Lacase.
4.6.3 Lignina Peroxidase ( LiP ) ou Ligninase
A atividade enzimática para lignina peroxidase ou ligninase foi determinada
em caldo de cultivo extrato de malte acrescido de 2% de glicose ou serragem da
madeira de Hura crepitans como fonte de carbono e também em meio sem
acréscimo de fonte de carbono (controle).
Na avaliação da atividade Lignina Peroxidase foi utilizado o método proposto
por Tien-Kirk (1984): este método baseia-se na oxidação de álcool veratrílico a
aldeído veratrílico. Para a determinação da atividade lignina peroxidase utilizou-se:
550 μL de caldo de cultura filtrado, 200 μL de H2O2 2,0 mM e 250 μL de solução de
álcool veratrílico em tampão tartarato de sódio 0,4M (pH3,0). O aparecimento do
aldeído veratrílico foi determinado lendo-se a absorbância a 310 nm (ε = 9.300 M–1
cm-1 ). Uma unidade de Lignina Peroxidase corresponde a quantidade de enzima
que oxida 1μmol de álcool veratrílico por minuto.

40
5 RESULTADOS
5.1 Otimização das condições de crescimento dos fungos
5.1.1 Velocidade de crescimento em função do tempo
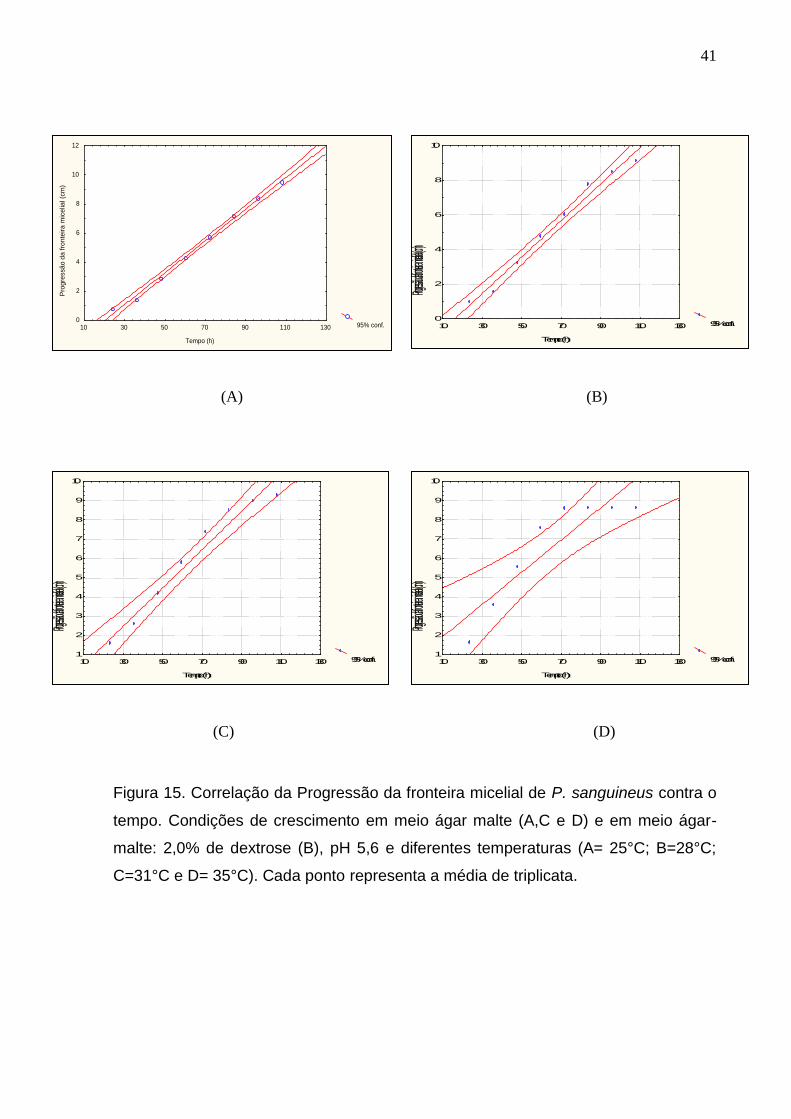
O avanço da fronteira micelial apresentou uma melhor resposta linear de
crescimento em função do tempo, entre 24 e 108 horas, e entre temperaturas de
25°C a 31°C em pH 5,2. Assim, a velocidade de crescimento fúngico foi obtida em
função da inclinação da reta no intervalo de tempo mencionado (Figura15).
5.1.2 Taxa de crescimento micelial
O crescimento é restrito à extremidade da hifa, a qual cresce mais ou menos
a uma taxa linear constante e, portanto, aumenta a margem da colônia a uma taxa
linear (MOORE-LANDECKER, 1982). No presente estudo a taxa de crescimento
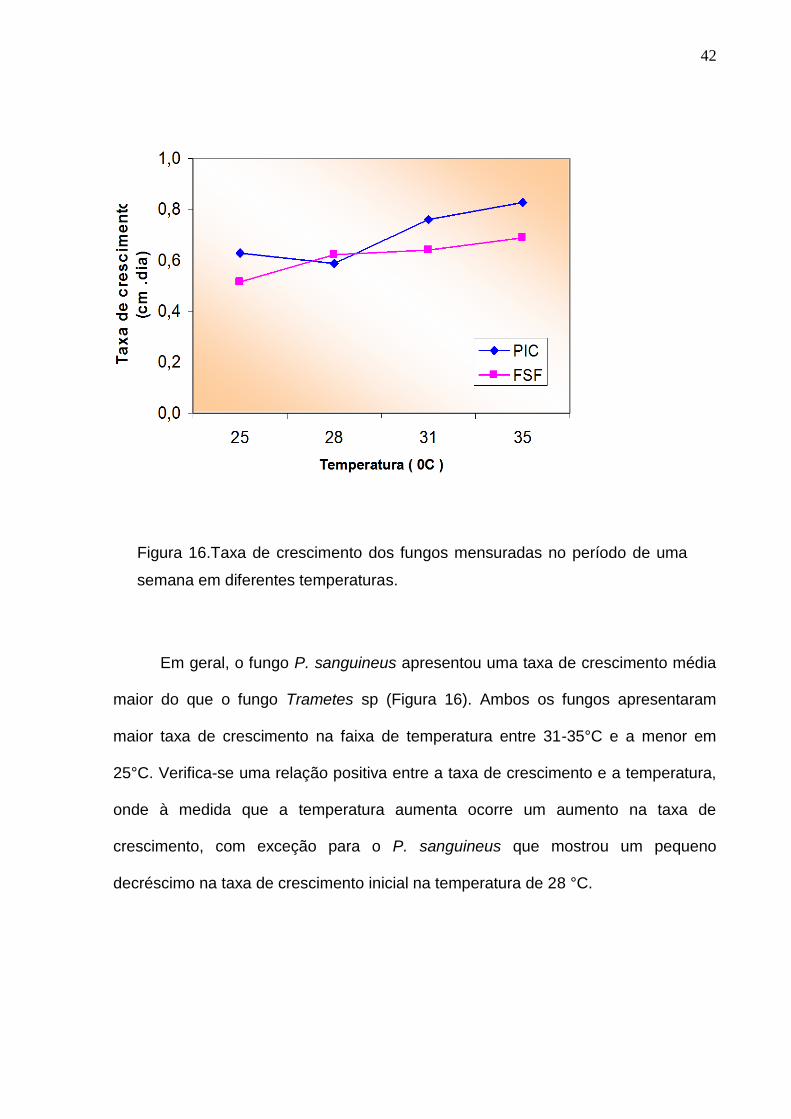
micelial apresenta-se diferentes para as temperaturas testadas (Figura 16).
41
(A) (B)
(C) (D)
Figura 15. Correlação da Progressão da fronteira micelial de P. sanguineus contra o
tempo. Condições de crescimento em meio ágar malte (A,C e D) e em meio ágar-
malte: 2,0% de dextrose (B), pH 5,6 e diferentes temperaturas (A= 25°C; B=28°C;
C=31°C e D= 35°C). Cada ponto representa a média de triplicata.
95% conf.
Tempo (h)
Pro
gre
ssã
o d
a f
ron
teira
mic
elia
l (c
m)
0
2
4
6
8
10
12
10 30 50 70 90 110 13095% conf.
Tempo (h)
Progressão
da fronteir
a micelial (c
m)
0
2
4
6
8
10
10 30 50 70 90 110 130
95% conf.
Tempo (h)
Progressão
da fronteir
a micelial (c
m)
1
2
3
4
5
6
7
8
9
10
10 30 50 70 90 110 130 95% conf.
Tempo (h)
Progressão
da fronteir
a micelial (c
m)
1
2
3
4
5
6
7
8
9
10
10 30 50 70 90 110 130
42
Figura 16.Taxa de crescimento dos fungos mensuradas no período de uma
semana em diferentes temperaturas.
Em geral, o fungo P. sanguineus apresentou uma taxa de crescimento média
maior do que o fungo Trametes sp (Figura 16). Ambos os fungos apresentaram
maior taxa de crescimento na faixa de temperatura entre 31-35°C e a menor em
25°C. Verifica-se uma relação positiva entre a taxa de crescimento e a temperatura,
onde à medida que a temperatura aumenta ocorre um aumento na taxa de
crescimento, com exceção para o P. sanguineus que mostrou um pequeno
decréscimo na taxa de crescimento inicial na temperatura de 28 °C.

43
5.1.3 Influência do pH no crescimento micelial dos fungos
Os fungos apresentaram um bom crescimento na faixa de pH considerada
(pH 4–7). No caso do Pycnoporus sanguineus o maior crescimento médio, em
termos absolutos, ocorreu no pH 5 e o menor no pH 4 (Tabela 03). O mesmo padrão
ocorreu para Trametes sp sendo que neste caso o valor médio absoluto para o
menor crescimento foi inferior àquele do P. sanguineus.
Tabela 03. Influência do pH no crescimento micelial dos fungos.
FUNGO pH
Cresc. micelial (cm)
Variância Desvio Padrão (Média)
Pycnoporus
sanguineus
4
5.01
5.023
2.24 5 6.90 5.747 2.39 6 6.88 6.104 2.47 7 6.64 6.026 2.45
Trametes sp
4
4.56
3.645
1.90 5 6.53 5.777 2.40 6 6.35 6.225 2.49 7 6.28 6.425 2.53
A Análise de variância para determinar se existe diferença significativa no
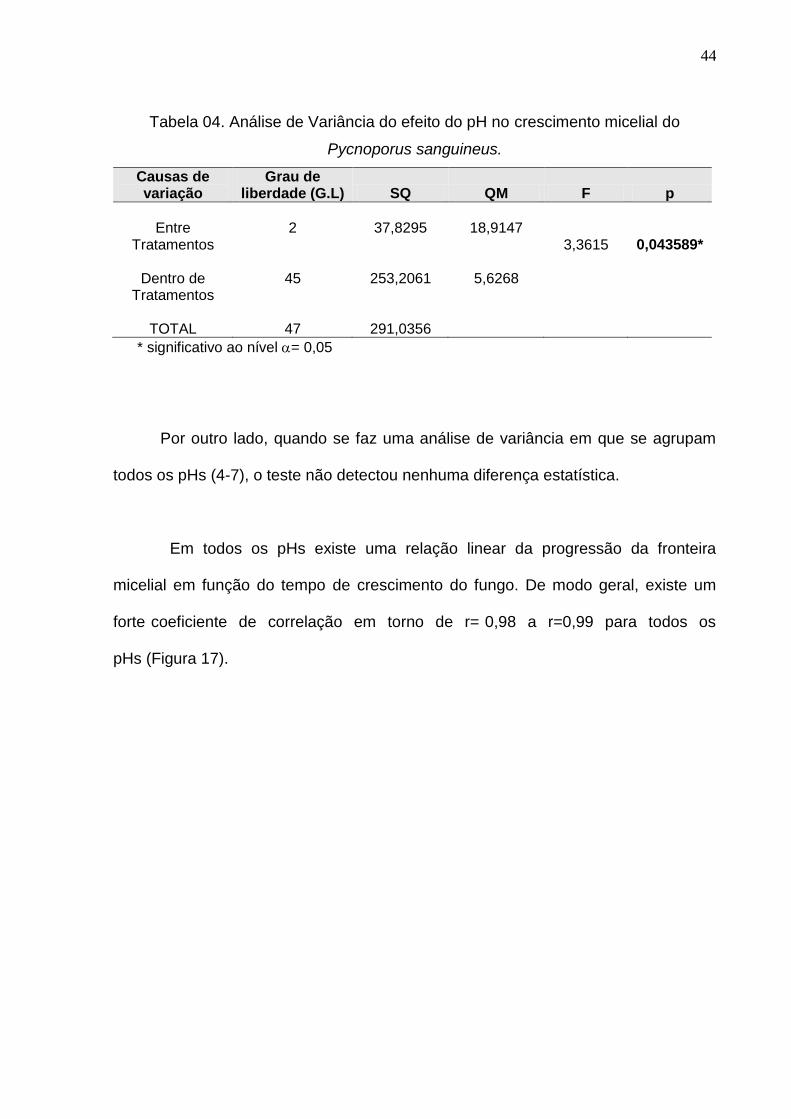
crescimento micelial entre os diferentes pHs (pH 4-6) evidenciou que ao nível de
95% de probabilidade existe diferença entre as médias obtidas (Tabela 04). Ou seja,
o crescimento micelial, medido pela progressão linear do avanço da fronteira
micelial, não é o mesmo. Teste de Tukey para comparação entre duas médias
mostrou diferença significativa no valor médio de crescimento para o pH4.
44
Tabela 04. Análise de Variância do efeito do pH no crescimento micelial do
Pycnoporus sanguineus.
Causas de variação
Grau de liberdade (G.L) SQ QM F p
Entre
Tratamentos
Dentro de Tratamentos
TOTAL
2
45
47
37,8295
253,2061
291,0356
18,9147
5,6268
3,3615
0,043589*
* significativo ao nível = 0,05
Por outro lado, quando se faz uma análise de variância em que se agrupam
todos os pHs (4-7), o teste não detectou nenhuma diferença estatística.
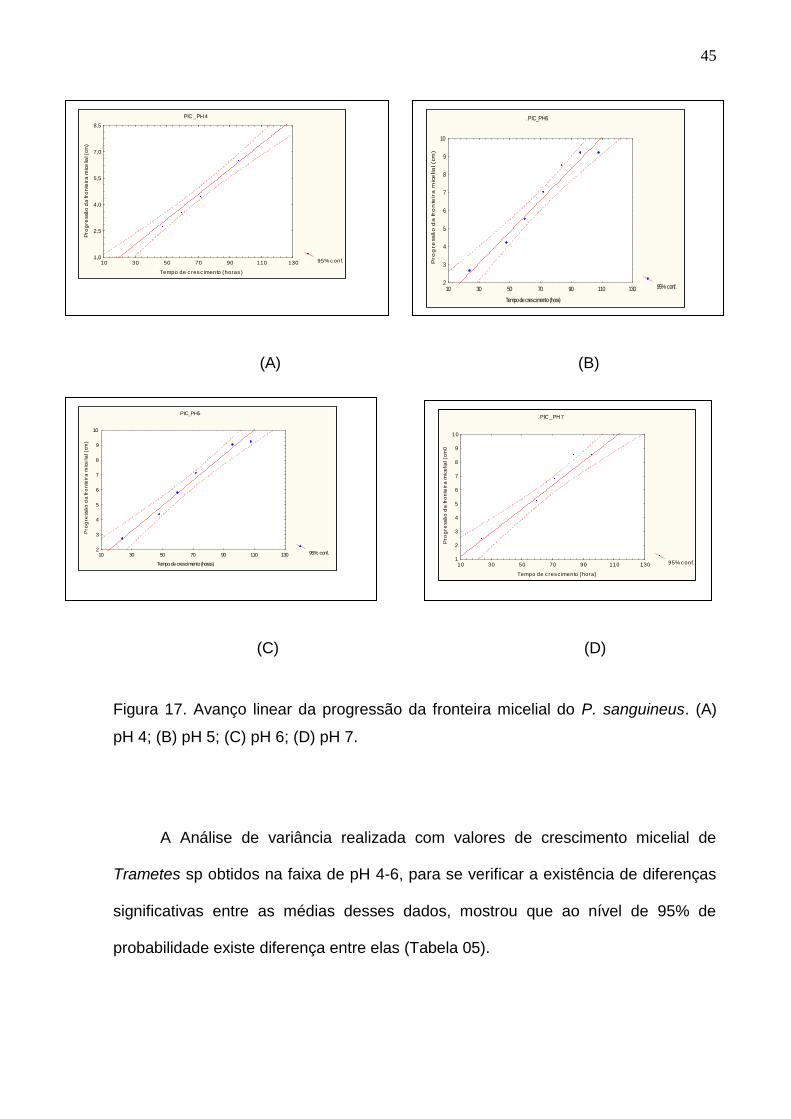
Em todos os pHs existe uma relação linear da progressão da fronteira
micelial em função do tempo de crescimento do fungo. De modo geral, existe um
forte coeficiente de correlação em torno de r= 0,98 a r=0,99 para todos os
pHs (Figura 17).
45
(A) (B)
(C) (D)
Figura 17. Avanço linear da progressão da fronteira micelial do P. sanguineus. (A)
pH 4; (B) pH 5; (C) pH 6; (D) pH 7.
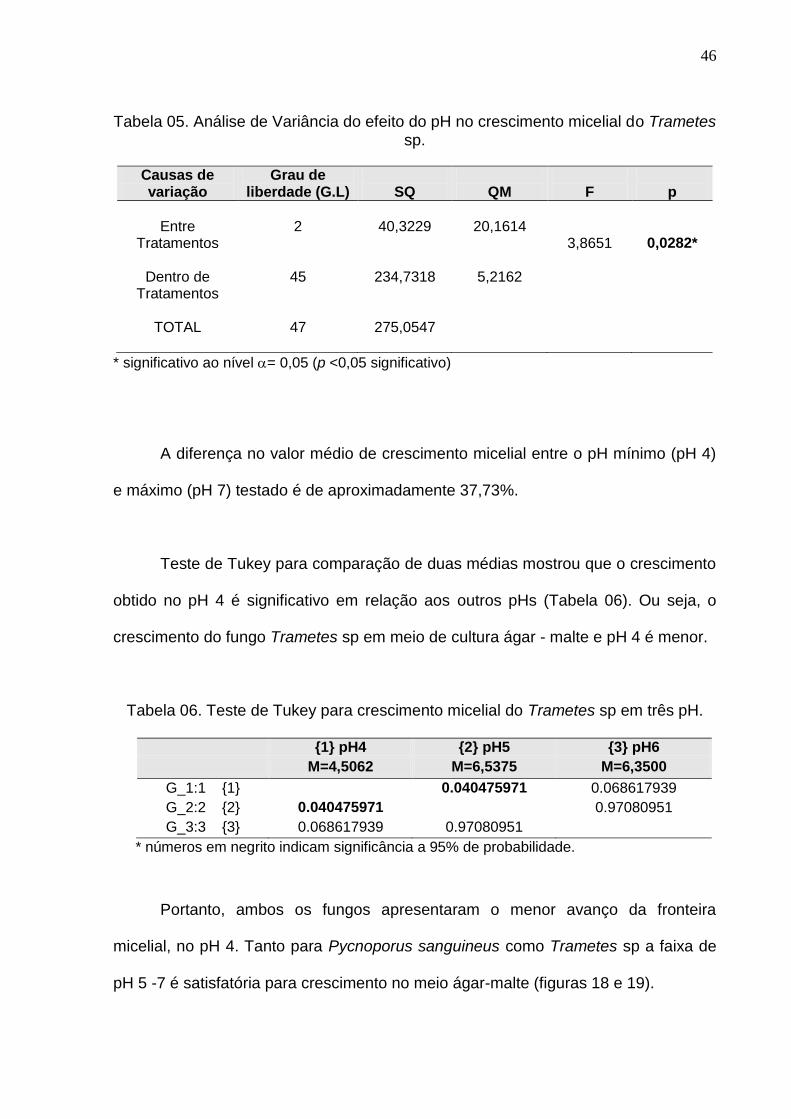
A Análise de variância realizada com valores de crescimento micelial de
Trametes sp obtidos na faixa de pH 4-6, para se verificar a existência de diferenças
significativas entre as médias desses dados, mostrou que ao nível de 95% de
probabilidade existe diferença entre elas (Tabela 05).
95% conf.
PIC_PH4
Tempo de c resc imento (horas )
Pro
gre
ssã
o d
a fr
on
teir
a m
ice
lial (
cm)
1,0
2,5
4,0
5,5
7,0
8,5
10 30 50 70 90 110 130
95% conf.
. PIC_PH6
Tempo de crescimento (hora)
Pro
gre
ssã
o d
a fro
nte
ira
mic
elia
l (cm
)
2
3
4
5
6
7
8
9
10
10 30 50 70 90 110 130
95% conf.
PIC_PH5
Tempo de crescimento (horas)
Pro
gre
ssã
o d
a fr
on
teir
a m
ice
lial (
cm)
2
3
4
5
6
7
8
9
10
10 30 50 70 90 110 130
95% c onf.
. PIC_PH7
Tempo de c res c imento (hora)
Pro
gre
ssã
o d
a fr
on
teir
a m
ice
lial (
cm0
1
2
3
4
5
6
7
8
9
10
10 30 50 70 90 110 130
46
Tabela 05. Análise de Variância do efeito do pH no crescimento micelial do Trametes sp.
Causas de variação
Grau de liberdade (G.L)
SQ
QM
F
p
Entre
Tratamentos
Dentro de Tratamentos
TOTAL
2
45
47
40,3229
234,7318
275,0547
20,1614
5,2162
3,8651
0,0282*
* significativo ao nível = 0,05 (p <0,05 significativo)
A diferença no valor médio de crescimento micelial entre o pH mínimo (pH 4)
e máximo (pH 7) testado é de aproximadamente 37,73%.
Teste de Tukey para comparação de duas médias mostrou que o crescimento
obtido no pH 4 é significativo em relação aos outros pHs (Tabela 06). Ou seja, o
crescimento do fungo Trametes sp em meio de cultura ágar - malte e pH 4 é menor.
Tabela 06. Teste de Tukey para crescimento micelial do Trametes sp em três pH.
{1} pH4 {2} pH5 {3} pH6
M=4,5062 M=6,5375 M=6,3500
G_1:1 {1} 0.040475971 0.068617939
G_2:2 {2} 0.040475971 0.97080951
G_3:3 {3} 0.068617939 0.97080951
* números em negrito indicam significância a 95% de probabilidade.
Portanto, ambos os fungos apresentaram o menor avanço da fronteira
micelial, no pH 4. Tanto para Pycnoporus sanguineus como Trametes sp a faixa de
pH 5 -7 é satisfatória para crescimento no meio ágar-malte (figuras 18 e 19).

47
Figura 18. Crescimento micelial de P. sanguineus em meio ágar malte.
Figura 19. Crescimento micelial de Trametes sp em meio ágar malte.
pH 6
pH 4 pH 5
pH 7
pH 4 pH 5
pH 6 pH 7

48
5.1.4 Concentração de Nitrogênio
A tabela 07 mostra os valores médios da progressão da fronteira micelial para
Pycnoporus sanguineus e Trametes sp nas diferentes concentrações de nitrato de
sódio (NS) e sulfato de amônia (SA). No geral, verifica-se para P. sanguineus que a
média do crescimento micelial, definido como a progressão linear do micélio, foi
similar para todas as fontes de nitrogênio testadas, notadamente para o nitrato de
sódio.
Tabela 07. Parâmetros descritivos da progressão da fronteira micelial do Pycnoporus
sanguineus e Trametes sp em diferentes concentrações de nitrogênio.
FUNGO
Fonte
Conc. N
Cresc. micelial (cm)
Variância
Desvio
Nitrogênio (mEq./L) (média) Padrão
Pycnoporus sanguineus
Nitrato de Sódio
30
5.37
8.7183
2.9527
60 4.98 8.0713 2.8410
90 4.91 8.0969 2.8455
120 5.38 9.7654 3.1250
150 5.40 7.7620 2.7860
Sulfato de Amônia
30
6.38
6.2108
2.4921
60 6.22 6.6321 2.5753
90 6.13 7.7465 2.7833
120 5.86 7.5927 2.7555
150 5.10 7.3424 2.7097
Trametes sp
Nitrato de Sódio
30
6.28
7.9397
2.8177
60 5.36 7.5935 2.7556
90 4.75 5.7773 2.4036
120 5.16 9.2191 3.0363
150 4.54 8.0677 2.8404
Sulfato de Amônia
30
6.24
7.2399
2.6907
60 5.89 7.9115 2.8127
90 5.32 5.2343 2.2879
120 5.78 8.2993 2.8808
150 4.84 6.2521 2.5004
49
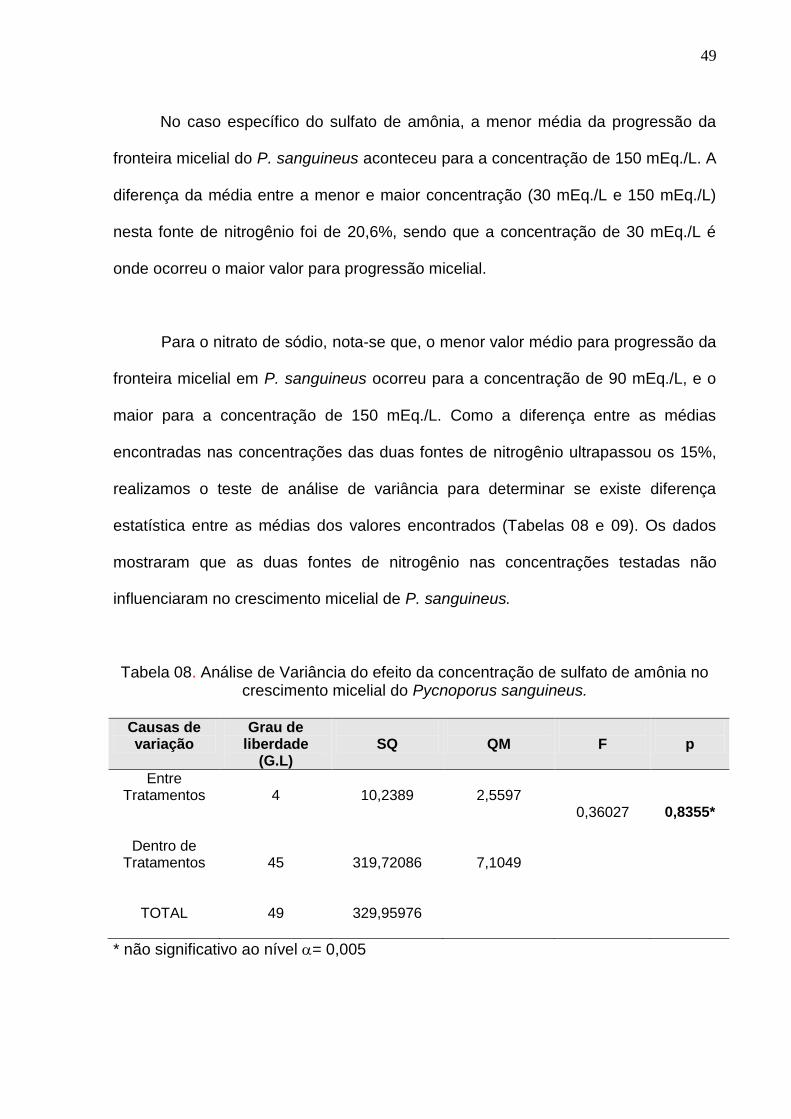
No caso específico do sulfato de amônia, a menor média da progressão da
fronteira micelial do P. sanguineus aconteceu para a concentração de 150 mEq./L. A
diferença da média entre a menor e maior concentração (30 mEq./L e 150 mEq./L)
nesta fonte de nitrogênio foi de 20,6%, sendo que a concentração de 30 mEq./L é
onde ocorreu o maior valor para progressão micelial.
Para o nitrato de sódio, nota-se que, o menor valor médio para progressão da
fronteira micelial em P. sanguineus ocorreu para a concentração de 90 mEq./L, e o
maior para a concentração de 150 mEq./L. Como a diferença entre as médias
encontradas nas concentrações das duas fontes de nitrogênio ultrapassou os 15%,
realizamos o teste de análise de variância para determinar se existe diferença
estatística entre as médias dos valores encontrados (Tabelas 08 e 09). Os dados
mostraram que as duas fontes de nitrogênio nas concentrações testadas não
influenciaram no crescimento micelial de P. sanguineus.
Tabela 08. Análise de Variância do efeito da concentração de sulfato de amônia no crescimento micelial do Pycnoporus sanguineus.
Causas de variação
Grau de liberdade
(G.L)
SQ
QM
F
p
Entre Tratamentos
Dentro de Tratamentos
TOTAL
4
45
49
10,2389
319,72086
329,95976
2,5597
7,1049
0,36027
0,8355*
* não significativo ao nível = 0,005
50
Tabela 09. Análise de Variância do efeito da concentração de nitrato de sódio no crescimento micelial do Pycnoporus sanguineus.
Causas de variação
Grau de liberdade
(G.L)
SQ
QM
F
p
Entre Tratamentos
Dentro de
Tratamentos
TOTAL
4
45
49
2,4755
399,5877
402,0632
0,61882
8,50186
0,07279
0,99004*
* não significativo ao nível = 0,005
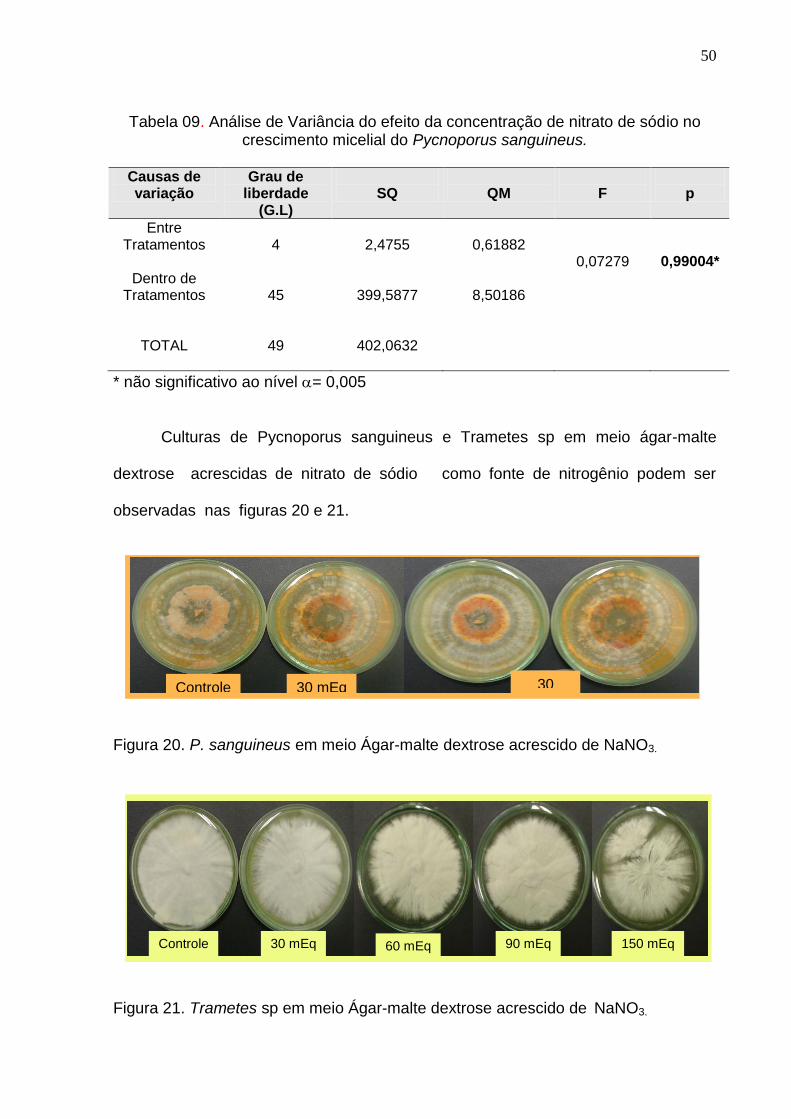
Culturas de Pycnoporus sanguineus e Trametes sp em meio ágar-malte
dextrose acrescidas de nitrato de sódio como fonte de nitrogênio podem ser
observadas nas figuras 20 e 21.
Figura 20. P. sanguineus em meio Ágar-malte dextrose acrescido de NaNO3.
Figura 21. Trametes sp em meio Ágar-malte dextrose acrescido de NaNO3.
30 mEq
Controle 30 mEq
150 mEq 90 mEq 60 mEq Controle 30 mEq

51
A progressão da fronteira micelial do Trametes sp em diferentes
concentrações de nitrogênio também mostrou similaridade nos valores absolutos
(Tabela 07). Por outro lado, o maior valor médio de crescimento micelial aconteceu
na concentração de 30 mEq./L do nitrato de sódio.
Diferentemente do que aconteceu com o P. sanguineus nota-se que a
diferença no valor médio da progressão da fronteira micelial de Trametes sp entre a
menor e maior concentração foi de 27,71% para o nitrato de sódio e 22,4% para o
sulfato de amônia.
Com o intuito também de verificar a existência ou não de diferenças
significativas na progressão da fronteira micelial entre as concentrações nas duas
fontes de nitrogênio, ou seja, testar a hipótese de que as médias das populações de
onde essas amostras provieram são iguais, foi aplicado o teste de análise de
variância.
Análise de variância mostrou que as médias entre as concentrações dentro de
cada fonte de nitrogênio, não são significativas (Tabelas 10 e 11). Portanto, para o
fungo Trametes sp as duas fontes de nitrogênio nas concentrações testadas não
exercem nenhuma influência no sentido de aumentar ou diminuir a produção
micelial.
52
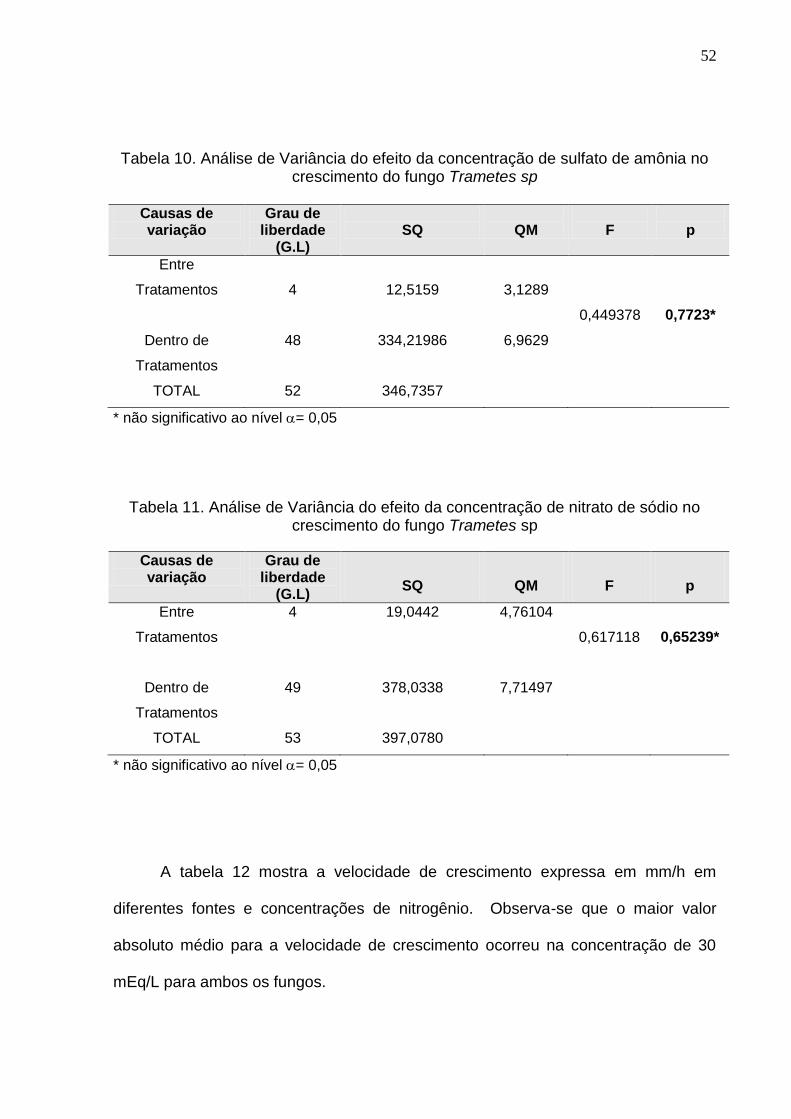
Tabela 10. Análise de Variância do efeito da concentração de sulfato de amônia no crescimento do fungo Trametes sp
Causas de variação
Grau de liberdade
(G.L)
SQ
QM
F
p
Entre
Tratamentos
Dentro de
Tratamentos
TOTAL
4
48
52
12,5159
334,21986
346,7357
3,1289
6,9629
0,449378
0,7723*
* não significativo ao nível = 0,05
Tabela 11. Análise de Variância do efeito da concentração de nitrato de sódio no
crescimento do fungo Trametes sp
Causas de variação
Grau de liberdade
(G.L)
SQ
QM
F
p
Entre
Tratamentos
Dentro de
Tratamentos
TOTAL
4
49
53
19,0442
378,0338
397,0780
4,76104
7,71497
0,617118
0,65239*
* não significativo ao nível = 0,05
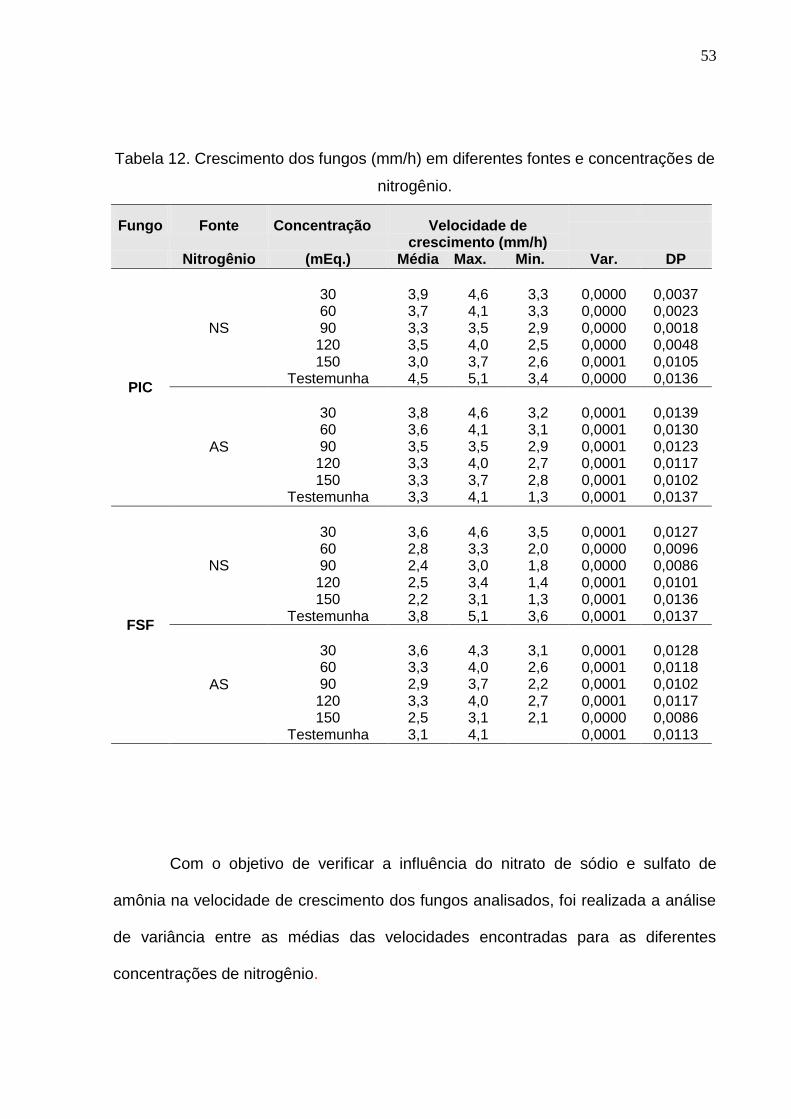
A tabela 12 mostra a velocidade de crescimento expressa em mm/h em
diferentes fontes e concentrações de nitrogênio. Observa-se que o maior valor
absoluto médio para a velocidade de crescimento ocorreu na concentração de 30
mEq/L para ambos os fungos.
53
Tabela 12. Crescimento dos fungos (mm/h) em diferentes fontes e concentrações de
nitrogênio.
Fungo
Fonte
Concentração
Velocidade de crescimento (mm/h)
Nitrogênio (mEq.) Média Max. Min. Var. DP
PIC
NS
30
3,9
4,6
3,3
0,0000
0,0037
60 3,7 4,1 3,3 0,0000 0,0023 90 3,3 3,5 2,9 0,0000 0,0018
120 3,5 4,0 2,5 0,0000 0,0048 150 3,0 3,7 2,6 0,0001 0,0105
Testemunha 4,5 5,1 3,4 0,0000 0,0136
AS
30
3,8
4,6
3,2
0,0001
0,0139
60 3,6 4,1 3,1 0,0001 0,0130 90 3,5 3,5 2,9 0,0001 0,0123
120 3,3 4,0 2,7 0,0001 0,0117 150 3,3 3,7 2,8 0,0001 0,0102
Testemunha 3,3 4,1 1,3 0,0001 0,0137
FSF
NS
30
3,6
4,6
3,5
0,0001
0,0127
60 2,8 3,3 2,0 0,0000 0,0096 90 2,4 3,0 1,8 0,0000 0,0086
120 2,5 3,4 1,4 0,0001 0,0101 150 2,2 3,1 1,3 0,0001 0,0136
Testemunha 3,8 5,1 3,6 0,0001 0,0137
AS
30
3,6
4,3
3,1
0,0001
0,0128
60 3,3 4,0 2,6 0,0001 0,0118 90 2,9 3,7 2,2 0,0001 0,0102
120 3,3 4,0 2,7 0,0001 0,0117 150 2,5 3,1 2,1 0,0000 0,0086
Testemunha 3,1 4,1 0,0001 0,0113
Com o objetivo de verificar a influência do nitrato de sódio e sulfato de
amônia na velocidade de crescimento dos fungos analisados, foi realizada a análise
de variância entre as médias das velocidades encontradas para as diferentes
concentrações de nitrogênio.
54
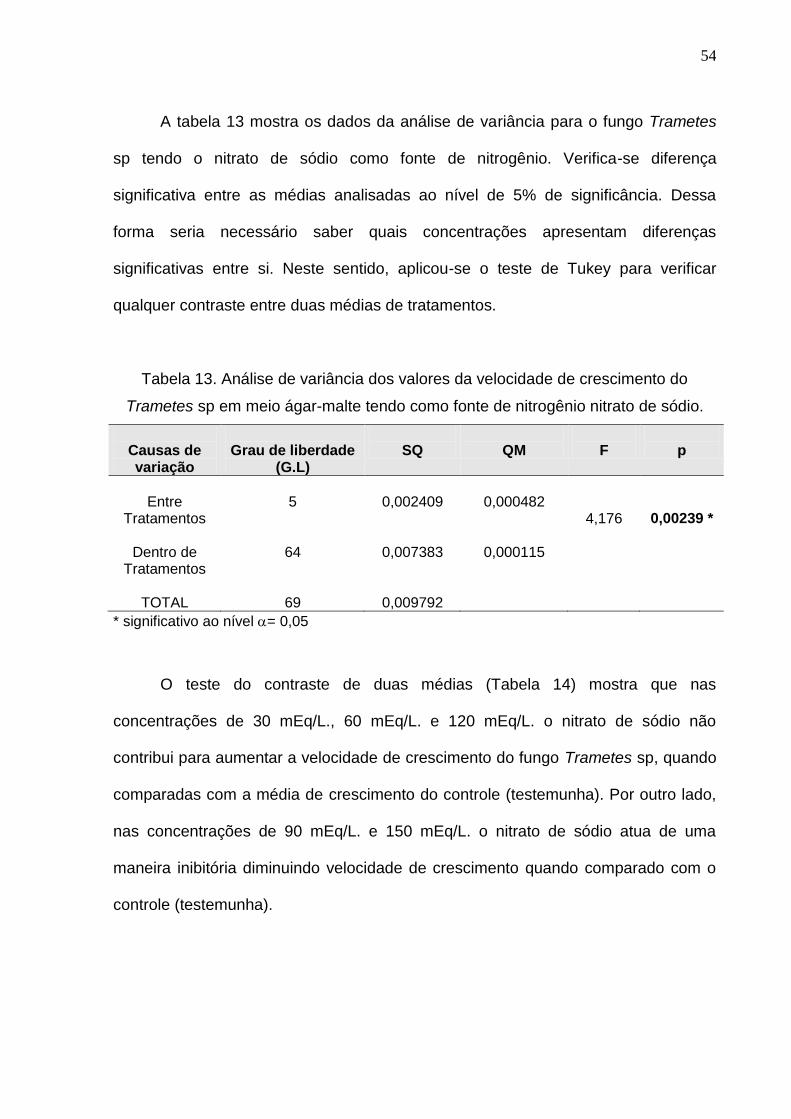
A tabela 13 mostra os dados da análise de variância para o fungo Trametes
sp tendo o nitrato de sódio como fonte de nitrogênio. Verifica-se diferença
significativa entre as médias analisadas ao nível de 5% de significância. Dessa
forma seria necessário saber quais concentrações apresentam diferenças
significativas entre si. Neste sentido, aplicou-se o teste de Tukey para verificar
qualquer contraste entre duas médias de tratamentos.
Tabela 13. Análise de variância dos valores da velocidade de crescimento do
Trametes sp em meio ágar-malte tendo como fonte de nitrogênio nitrato de sódio.
Causas de variação
Grau de liberdade
(G.L)
SQ
QM
F
p
Entre
Tratamentos
Dentro de Tratamentos
TOTAL
5
64
69
0,002409
0,007383
0,009792
0,000482
0,000115
4,176
0,00239 *
* significativo ao nível = 0,05
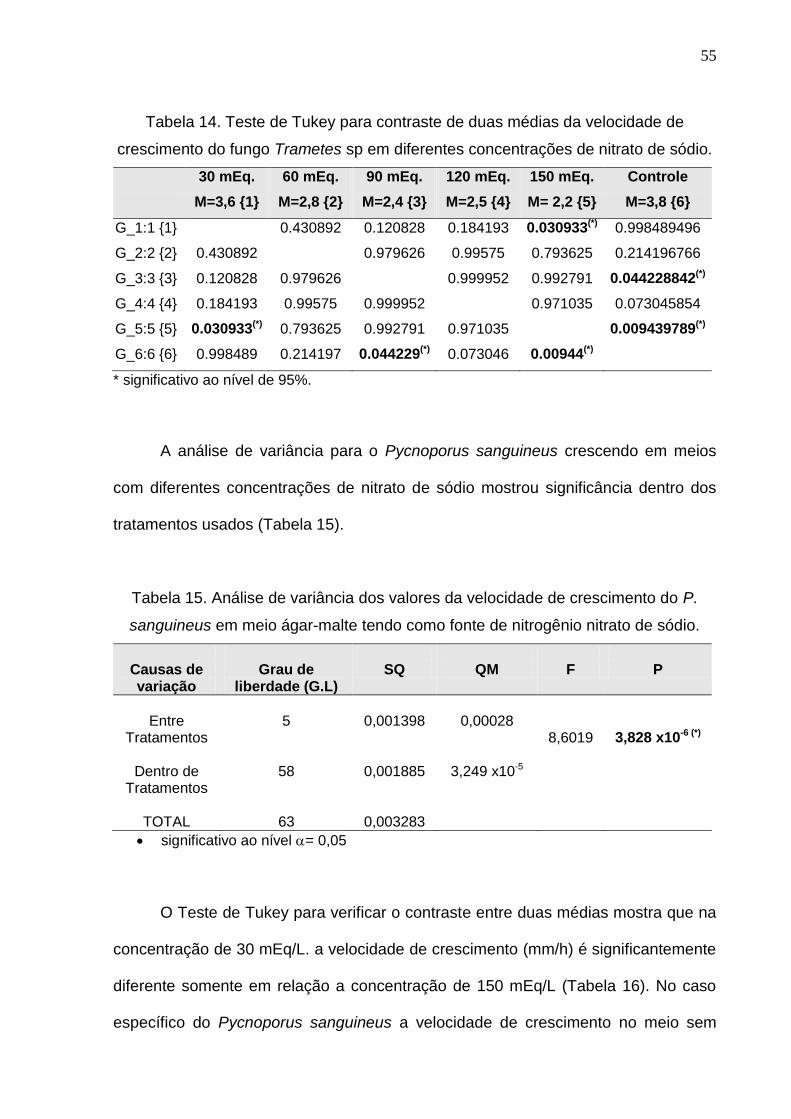
O teste do contraste de duas médias (Tabela 14) mostra que nas
concentrações de 30 mEq/L., 60 mEq/L. e 120 mEq/L. o nitrato de sódio não
contribui para aumentar a velocidade de crescimento do fungo Trametes sp, quando
comparadas com a média de crescimento do controle (testemunha). Por outro lado,
nas concentrações de 90 mEq/L. e 150 mEq/L. o nitrato de sódio atua de uma
maneira inibitória diminuindo velocidade de crescimento quando comparado com o
controle (testemunha).
55
Tabela 14. Teste de Tukey para contraste de duas médias da velocidade de
crescimento do fungo Trametes sp em diferentes concentrações de nitrato de sódio.
30 mEq. 60 mEq. 90 mEq. 120 mEq. 150 mEq. Controle
M=3,6 {1} M=2,8 {2} M=2,4 {3} M=2,5 {4} M= 2,2 {5} M=3,8 {6}
G_1:1 {1} 0.430892 0.120828 0.184193 0.030933(*) 0.998489496
G_2:2 {2} 0.430892 0.979626 0.99575 0.793625 0.214196766
G_3:3 {3} 0.120828 0.979626 0.999952 0.992791 0.044228842(*)
G_4:4 {4} 0.184193 0.99575 0.999952 0.971035 0.073045854
G_5:5 {5} 0.030933(*) 0.793625 0.992791 0.971035 0.009439789(*)
G_6:6 {6} 0.998489 0.214197 0.044229(*) 0.073046 0.00944(*)
* significativo ao nível de 95%.
A análise de variância para o Pycnoporus sanguineus crescendo em meios
com diferentes concentrações de nitrato de sódio mostrou significância dentro dos
tratamentos usados (Tabela 15).
Tabela 15. Análise de variância dos valores da velocidade de crescimento do P.
sanguineus em meio ágar-malte tendo como fonte de nitrogênio nitrato de sódio.
Causas de variação
Grau de
liberdade (G.L)
SQ
QM
F
P
Entre
Tratamentos
Dentro de Tratamentos
TOTAL
5
58
63
0,001398
0,001885
0,003283
0,00028
3,249 x10-5
8,6019
3,828 x10-6 (*)
significativo ao nível = 0,05
O Teste de Tukey para verificar o contraste entre duas médias mostra que na
concentração de 30 mEq/L. a velocidade de crescimento (mm/h) é significantemente
diferente somente em relação a concentração de 150 mEq/L (Tabela 16). No caso
específico do Pycnoporus sanguineus a velocidade de crescimento no meio sem
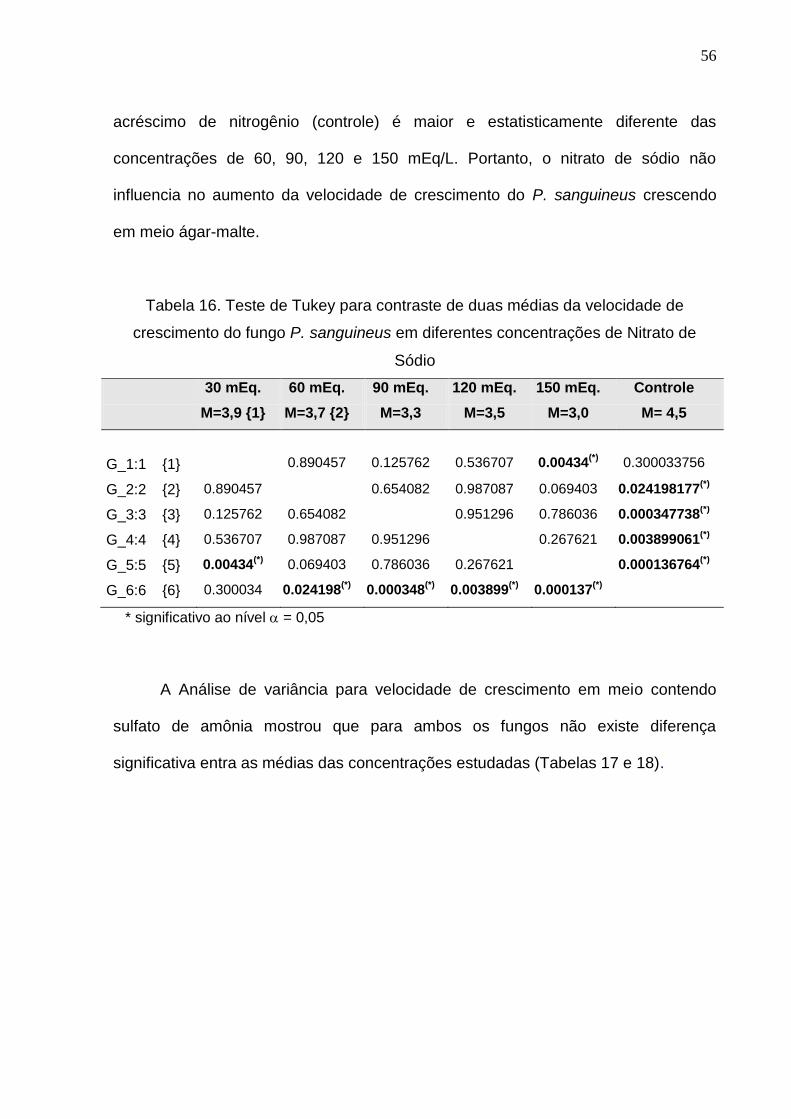
56
acréscimo de nitrogênio (controle) é maior e estatisticamente diferente das
concentrações de 60, 90, 120 e 150 mEq/L. Portanto, o nitrato de sódio não
influencia no aumento da velocidade de crescimento do P. sanguineus crescendo
em meio ágar-malte.
Tabela 16. Teste de Tukey para contraste de duas médias da velocidade de
crescimento do fungo P. sanguineus em diferentes concentrações de Nitrato de
Sódio
30 mEq. 60 mEq. 90 mEq. 120 mEq. 150 mEq. Controle
M=3,9 {1} M=3,7 {2} M=3,3 M=3,5 M=3,0 M= 4,5
G_1:1 {1}
0.890457
0.125762
0.536707
0.00434(*)
0.300033756
G_2:2 {2} 0.890457 0.654082 0.987087 0.069403 0.024198177(*)
G_3:3 {3} 0.125762 0.654082 0.951296 0.786036 0.000347738(*)
G_4:4 {4} 0.536707 0.987087 0.951296 0.267621 0.003899061(*)
G_5:5 {5} 0.00434(*)
0.069403 0.786036 0.267621 0.000136764(*)
G_6:6 {6} 0.300034 0.024198(*)
0.000348(*)
0.003899(*)
0.000137(*)
* significativo ao nível = 0,05
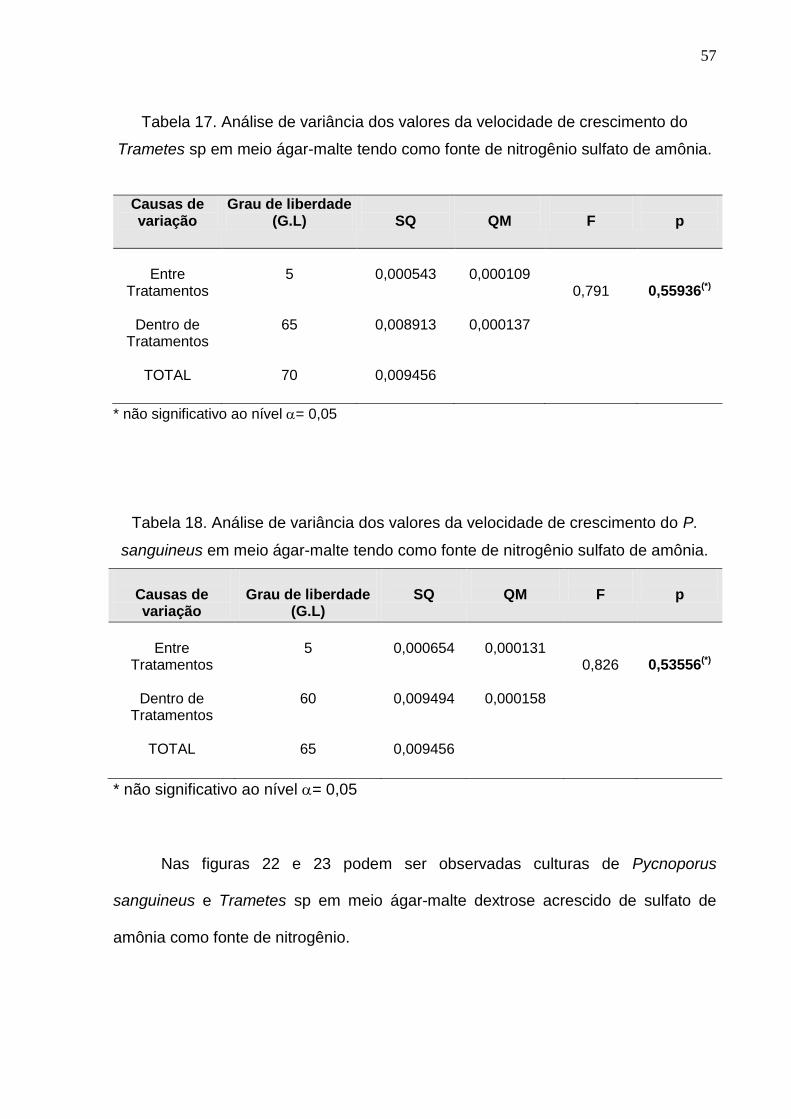
A Análise de variância para velocidade de crescimento em meio contendo
sulfato de amônia mostrou que para ambos os fungos não existe diferença
significativa entra as médias das concentrações estudadas (Tabelas 17 e 18).
57
Tabela 17. Análise de variância dos valores da velocidade de crescimento do
Trametes sp em meio ágar-malte tendo como fonte de nitrogênio sulfato de amônia.
Causas de variação
Grau de liberdade (G.L)
SQ
QM
F
p
Entre
Tratamentos
Dentro de Tratamentos
TOTAL
5
65
70
0,000543
0,008913
0,009456
0,000109
0,000137
0,791
0,55936(*)
* não significativo ao nível = 0,05
Tabela 18. Análise de variância dos valores da velocidade de crescimento do P.
sanguineus em meio ágar-malte tendo como fonte de nitrogênio sulfato de amônia.
Causas de variação
Grau de liberdade
(G.L)
SQ
QM
F
p
Entre
Tratamentos
Dentro de Tratamentos
TOTAL
5
60
65
0,000654
0,009494
0,009456
0,000131
0,000158
0,826
0,53556(*)
* não significativo ao nível = 0,05
Nas figuras 22 e 23 podem ser observadas culturas de Pycnoporus
sanguineus e Trametes sp em meio ágar-malte dextrose acrescido de sulfato de
amônia como fonte de nitrogênio.

58
Figura 22. Cultura do fungo P. sanguineus em meio Ágar-malte dextrose acrescido
de ((NH4)2SO4)).
Figura 23. Cultura do fungo Trametes sp em meio Ágar-malte dextrose acrescido
de ((NH4)2SO4)).
60 mEq
150 mEq Controle
120 mEq 90 mEq
30 mEq
60 mEq 150 mEq
controle
120 mEq
90 mEq
30 mEq
59
5.2 Atividade enzimática dos fungos
5.2.1 Atividade enzimática do P sanguineus em cultura estacionária
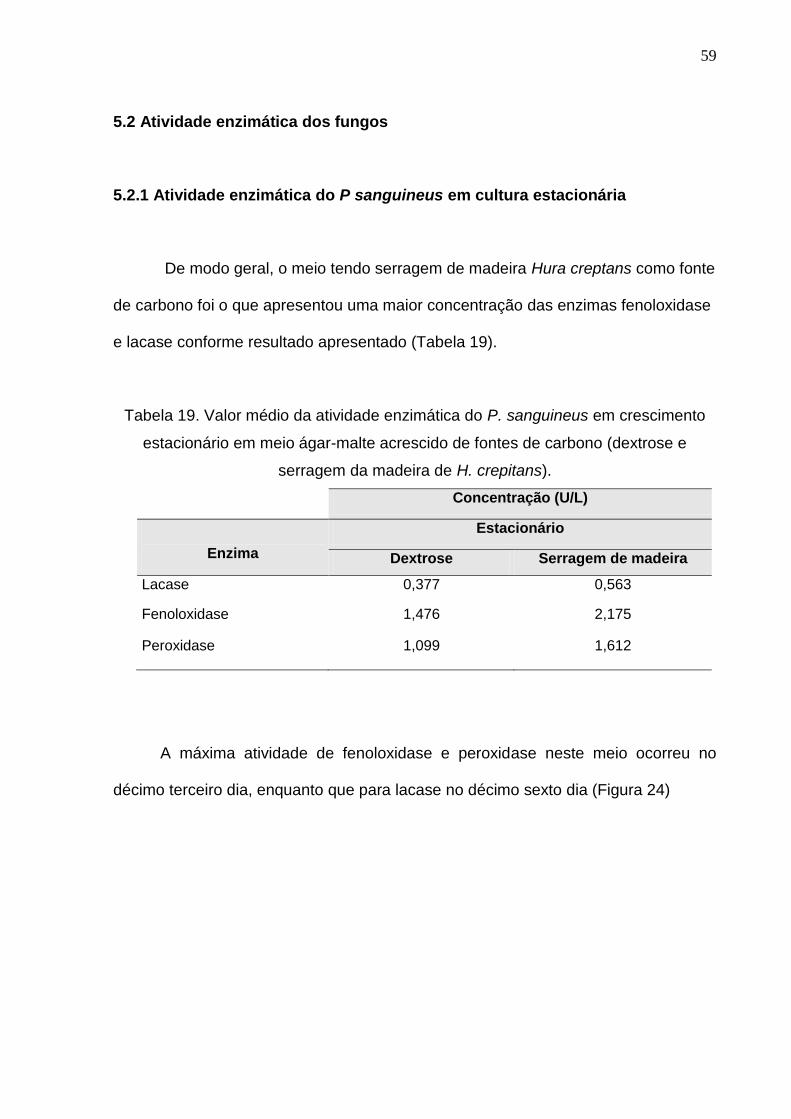
De modo geral, o meio tendo serragem de madeira Hura creptans como fonte
de carbono foi o que apresentou uma maior concentração das enzimas fenoloxidase
e lacase conforme resultado apresentado (Tabela 19).
Tabela 19. Valor médio da atividade enzimática do P. sanguineus em crescimento
estacionário em meio ágar-malte acrescido de fontes de carbono (dextrose e
serragem da madeira de H. crepitans).
Concentração (U/L)
Enzima
Estacionário
Dextrose Serragem de madeira
Lacase 0,377 0,563
Fenoloxidase 1,476 2,175
Peroxidase 1,099 1,612
A máxima atividade de fenoloxidase e peroxidase neste meio ocorreu no
décimo terceiro dia, enquanto que para lacase no décimo sexto dia (Figura 24)
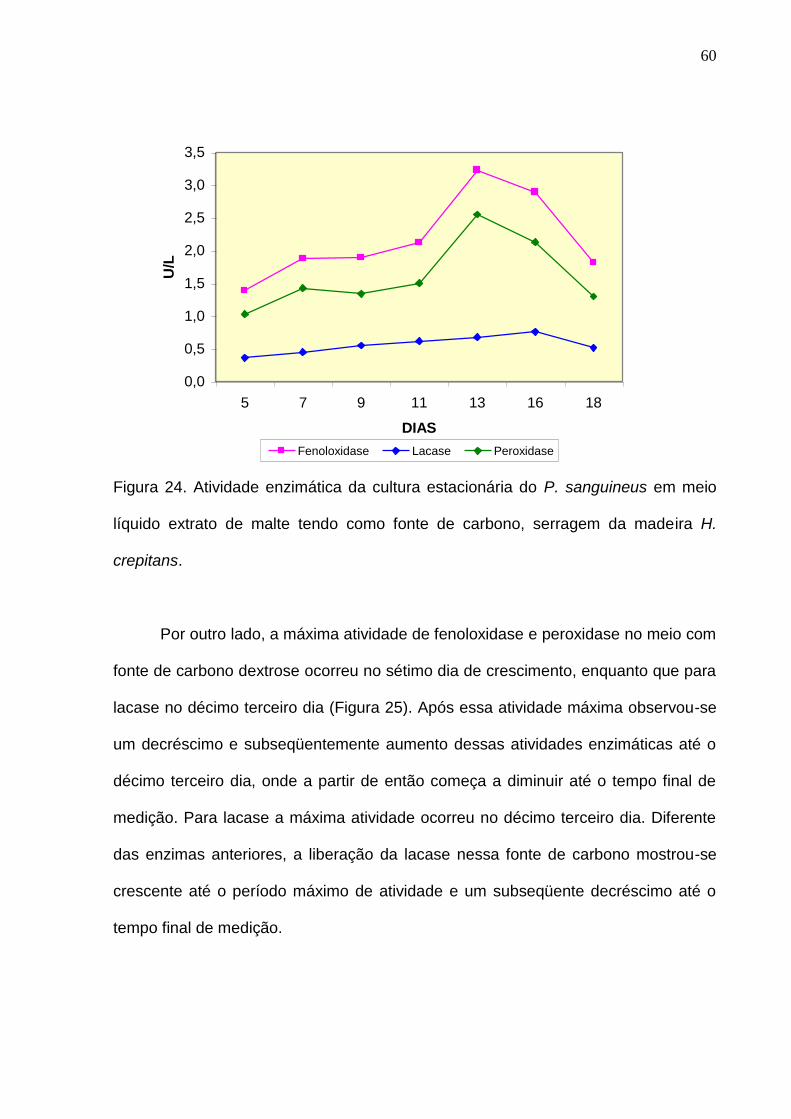
60
Figura 24. Atividade enzimática da cultura estacionária do P. sanguineus em meio
líquido extrato de malte tendo como fonte de carbono, serragem da madeira H.
crepitans.
Por outro lado, a máxima atividade de fenoloxidase e peroxidase no meio com
fonte de carbono dextrose ocorreu no sétimo dia de crescimento, enquanto que para
lacase no décimo terceiro dia (Figura 25). Após essa atividade máxima observou-se
um decréscimo e subseqüentemente aumento dessas atividades enzimáticas até o
décimo terceiro dia, onde a partir de então começa a diminuir até o tempo final de
medição. Para lacase a máxima atividade ocorreu no décimo terceiro dia. Diferente
das enzimas anteriores, a liberação da lacase nessa fonte de carbono mostrou-se
crescente até o período máximo de atividade e um subseqüente decréscimo até o
tempo final de medição.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
5 7 9 11 13 16 18
DIAS
U/L
Fenoloxidase Lacase Peroxidase
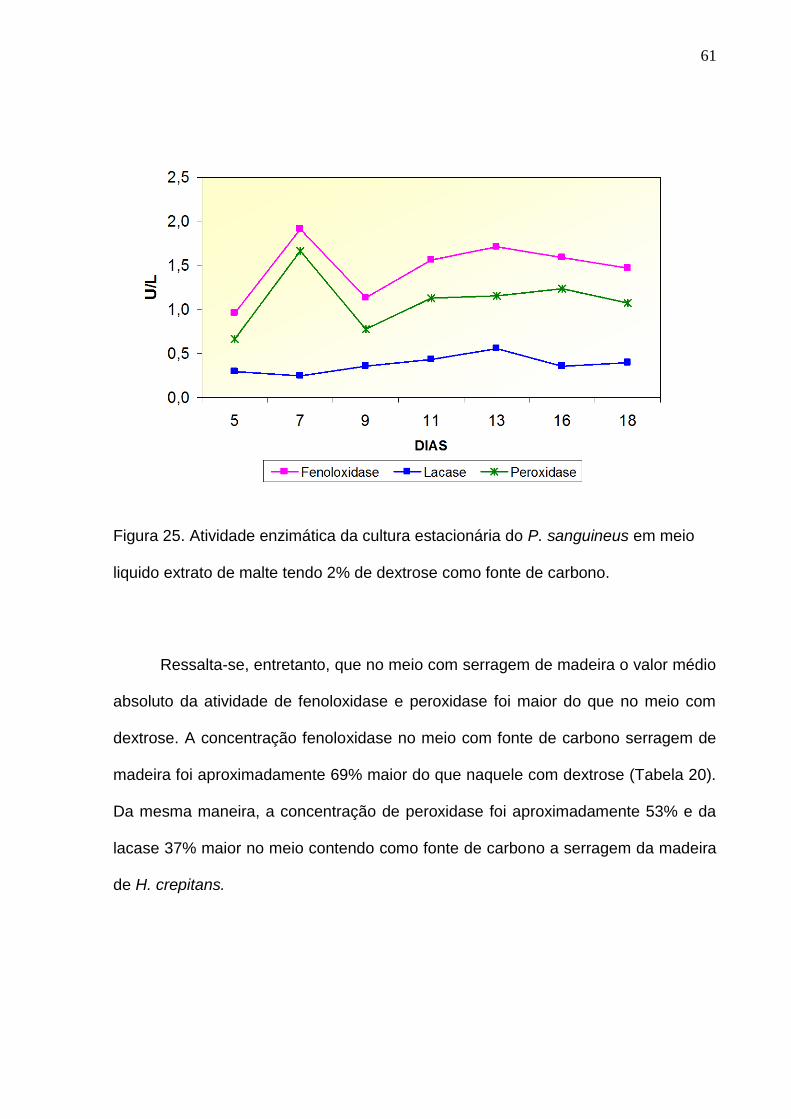
61
Figura 25. Atividade enzimática da cultura estacionária do P. sanguineus em meio
liquido extrato de malte tendo 2% de dextrose como fonte de carbono.
Ressalta-se, entretanto, que no meio com serragem de madeira o valor médio
absoluto da atividade de fenoloxidase e peroxidase foi maior do que no meio com
dextrose. A concentração fenoloxidase no meio com fonte de carbono serragem de
madeira foi aproximadamente 69% maior do que naquele com dextrose (Tabela 20).
Da mesma maneira, a concentração de peroxidase foi aproximadamente 53% e da
lacase 37% maior no meio contendo como fonte de carbono a serragem da madeira
de H. crepitans.
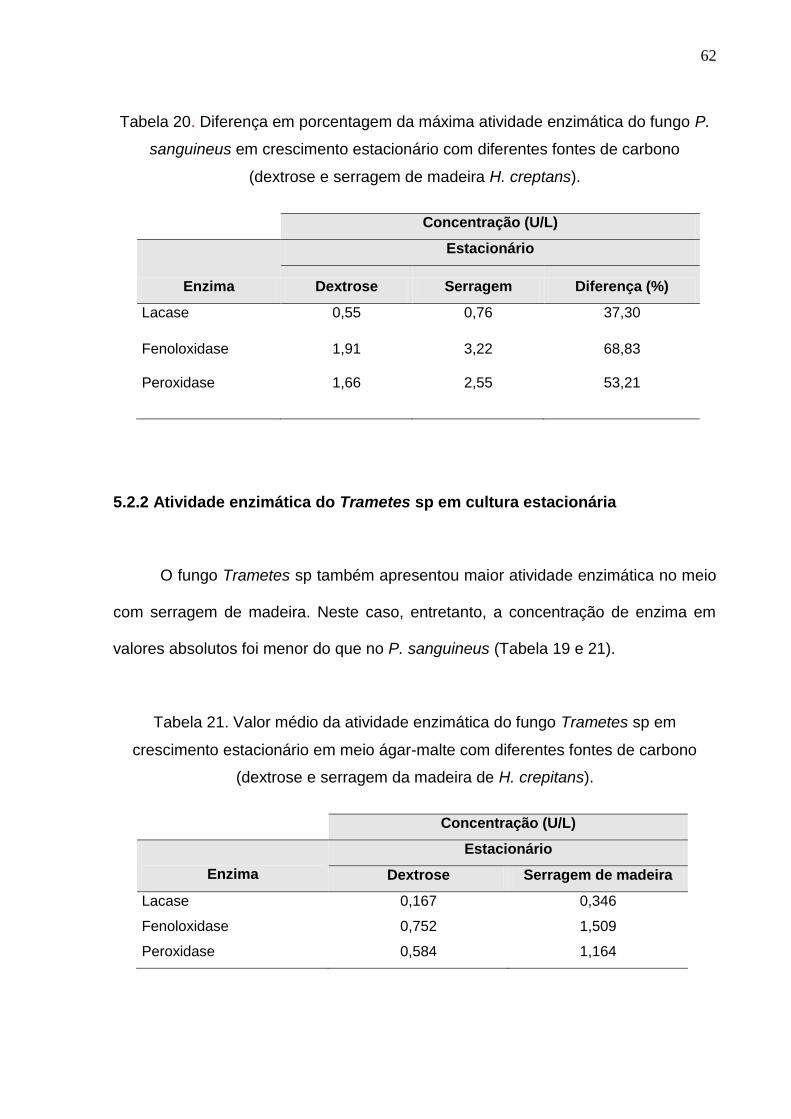
62
Tabela 20. Diferença em porcentagem da máxima atividade enzimática do fungo P.
sanguineus em crescimento estacionário com diferentes fontes de carbono
(dextrose e serragem de madeira H. creptans).
Concentração (U/L)
Enzima
Estacionário
Dextrose Serragem Diferença (%)
Lacase 0,55 0,76 37,30
Fenoloxidase 1,91 3,22 68,83
Peroxidase 1,66 2,55 53,21
5.2.2 Atividade enzimática do Trametes sp em cultura estacionária
O fungo Trametes sp também apresentou maior atividade enzimática no meio
com serragem de madeira. Neste caso, entretanto, a concentração de enzima em
valores absolutos foi menor do que no P. sanguineus (Tabela 19 e 21).
Tabela 21. Valor médio da atividade enzimática do fungo Trametes sp em
crescimento estacionário em meio ágar-malte com diferentes fontes de carbono
(dextrose e serragem da madeira de H. crepitans).
Concentração (U/L)
Enzima
Estacionário
Dextrose Serragem de madeira
Lacase 0,167 0,346
Fenoloxidase 0,752 1,509
Peroxidase 0,584 1,164

63
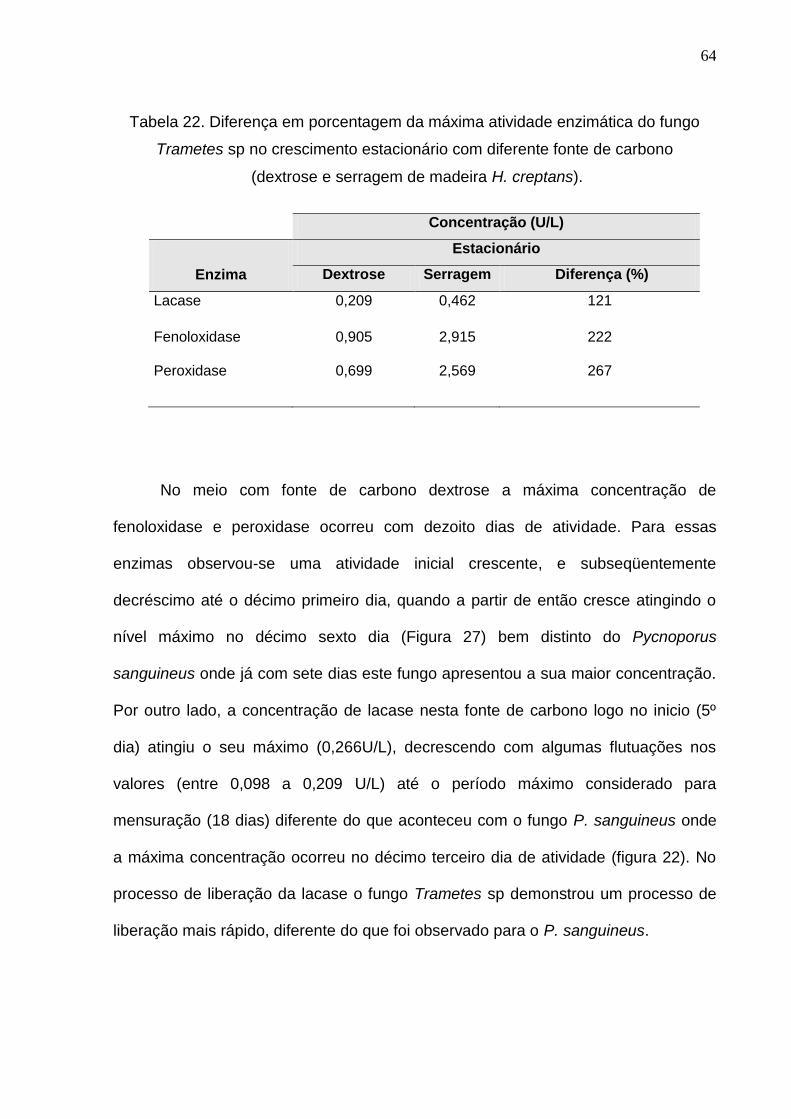
Para o crescimento em meio com fonte de carbono serragem de madeira a
maior concentração para fenoloxidase e peroxidase ocorreu no décimo oitavo dia de
crescimento (Figura 26) observando-se diferença em relação ao P. sanguineus que
com treze dias este fungo apresentou a sua maior concentração (Figura 24). A
máxima concentração de lacase em Trametes sp ocorreu no décimo sexto dia de
crescimento. Para este fungo a atividade enzimática máxima apresentou percentual
bem superior no meio acrescido de serragem chegando a apresentar duas vezes
mais concentração das enzimas fenoloxidase e peroxidase (Tabela 22). Verificou-se
que a lacase dobrou a concentração no meio acrescido de serragem. Este resultado
mostra que este fungo apresenta uma atividade bem mais agressiva no meio
acrescido de serragem da madeira de H. creptans do que o fungo P. sanguineus.
Figura 26. Atividade enzimática da cultura estacionária do Trametes sp em meio
líquido extrato de malte (fonte de carbono serragem de madeira H. crepitans).
64
Tabela 22. Diferença em porcentagem da máxima atividade enzimática do fungo
Trametes sp no crescimento estacionário com diferente fonte de carbono
(dextrose e serragem de madeira H. creptans).
Concentração (U/L)
Enzima
Estacionário
Dextrose Serragem Diferença (%)
Lacase 0,209 0,462 121
Fenoloxidase 0,905 2,915 222
Peroxidase 0,699 2,569 267
No meio com fonte de carbono dextrose a máxima concentração de
fenoloxidase e peroxidase ocorreu com dezoito dias de atividade. Para essas
enzimas observou-se uma atividade inicial crescente, e subseqüentemente
decréscimo até o décimo primeiro dia, quando a partir de então cresce atingindo o
nível máximo no décimo sexto dia (Figura 27) bem distinto do Pycnoporus
sanguineus onde já com sete dias este fungo apresentou a sua maior concentração.
Por outro lado, a concentração de lacase nesta fonte de carbono logo no inicio (5º
dia) atingiu o seu máximo (0,266U/L), decrescendo com algumas flutuações nos
valores (entre 0,098 a 0,209 U/L) até o período máximo considerado para
mensuração (18 dias) diferente do que aconteceu com o fungo P. sanguineus onde
a máxima concentração ocorreu no décimo terceiro dia de atividade (figura 22). No
processo de liberação da lacase o fungo Trametes sp demonstrou um processo de
liberação mais rápido, diferente do que foi observado para o P. sanguineus.
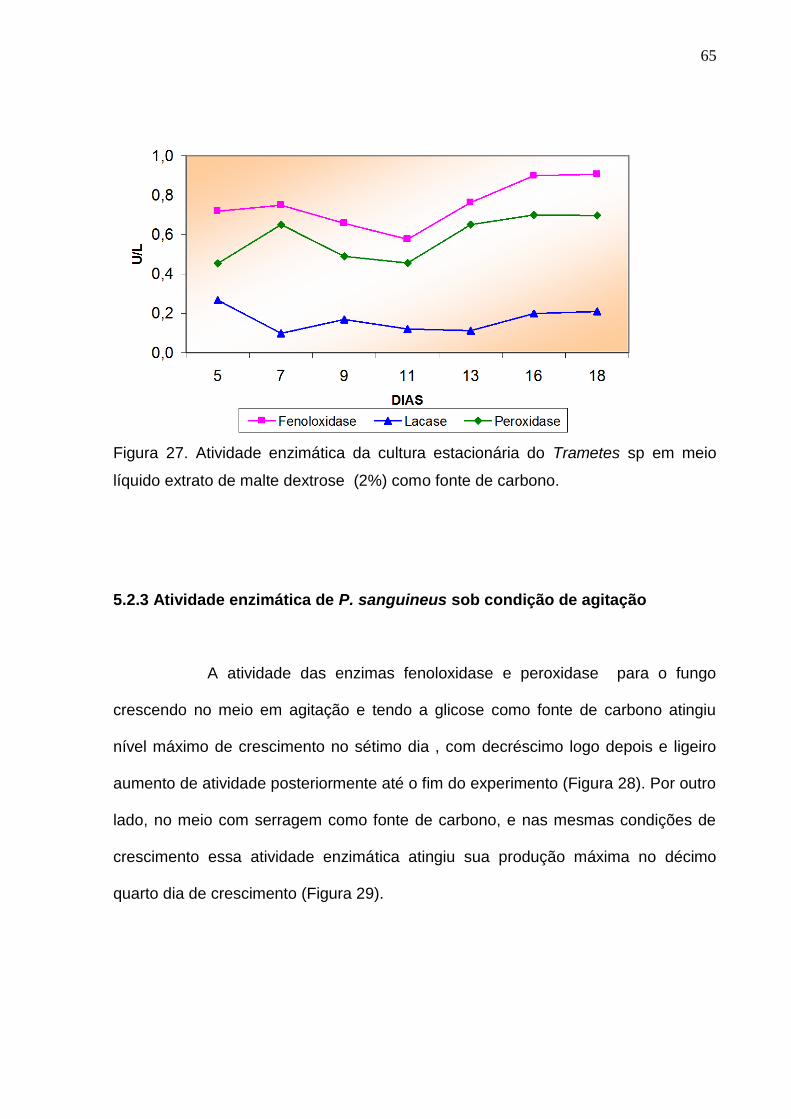
65
Figura 27. Atividade enzimática da cultura estacionária do Trametes sp em meio
líquido extrato de malte dextrose (2%) como fonte de carbono.
5.2.3 Atividade enzimática de P. sanguineus sob condição de agitação
A atividade das enzimas fenoloxidase e peroxidase para o fungo
crescendo no meio em agitação e tendo a glicose como fonte de carbono atingiu
nível máximo de crescimento no sétimo dia , com decréscimo logo depois e ligeiro
aumento de atividade posteriormente até o fim do experimento (Figura 28). Por outro
lado, no meio com serragem como fonte de carbono, e nas mesmas condições de
crescimento essa atividade enzimática atingiu sua produção máxima no décimo
quarto dia de crescimento (Figura 29).
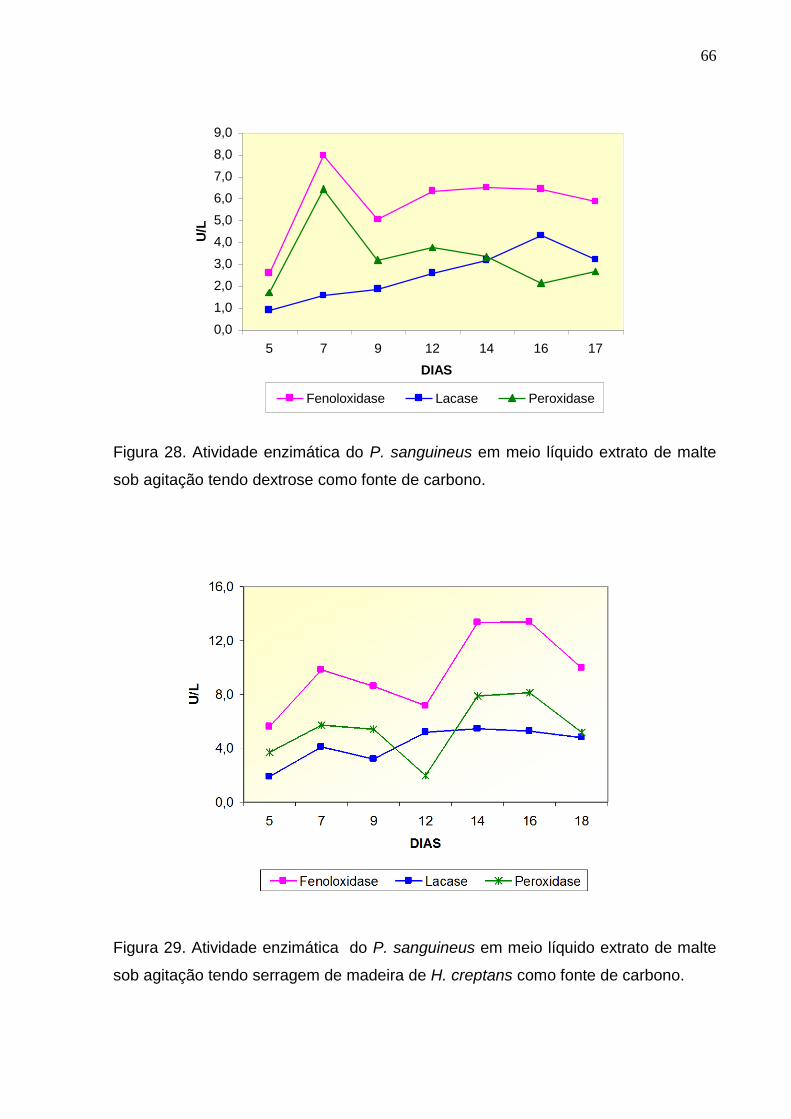
66
Figura 28. Atividade enzimática do P. sanguineus em meio líquido extrato de malte
sob agitação tendo dextrose como fonte de carbono.
Figura 29. Atividade enzimática do P. sanguineus em meio líquido extrato de malte
sob agitação tendo serragem de madeira de H. creptans como fonte de carbono.
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
9,0
5 7 9 12 14 16 17
DIAS
U/L
Fenoloxidase Lacase Peroxidase

67
5.2.4 Atividade enzimática do Trametes sp sob condição de agitação
Para a atividade de fenoloxidase e peroxidase em meio tendo glicose como
fonte de carbono, o nível máximo de atividade ocorreu no décimo segundo dia de
crescimento. O padrão de produção dessas enzimas nesta fonte de carbono,
mostrou-se crescente até atingir o seu nível máximo e com posterior decréscimo,
com flutuações, até o final do período de medição (Figura 30). A produção de lacase
atingiu seu nível máximo ao nono dia de atividade do fungo Trametes sp, e posterior
decréscimo até o final do período considerado para crescimento (Figura 30). Por
outro lado, a produção dessa enzima no meio com serragem como fonte de carbono
atingiu seu nível máximo com dezesseis dias de crescimento (Figura 31).
A produção de fenoloxidase e peroxidase em meio com serragem como fonte
de carbono atingiu seu nível máximo, assim como a lacase, no décimo sexto dia de
crescimento do fungo.
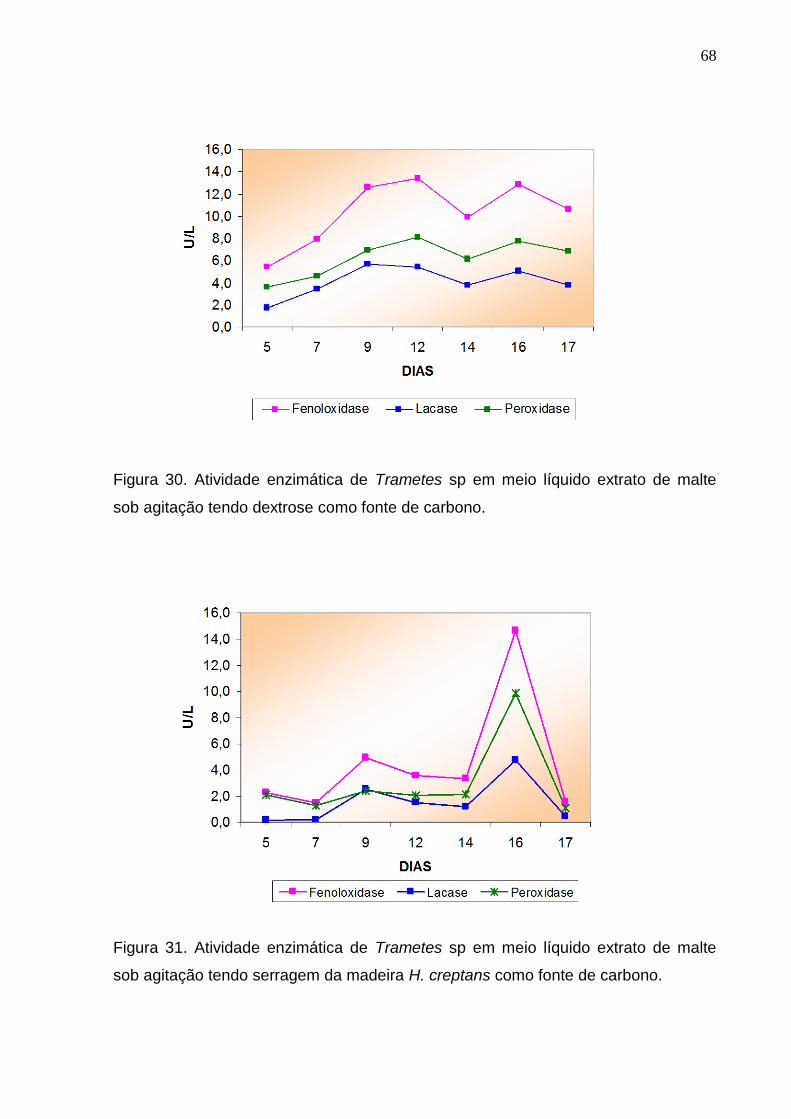
68
Figura 30. Atividade enzimática de Trametes sp em meio líquido extrato de malte
sob agitação tendo dextrose como fonte de carbono.
Figura 31. Atividade enzimática de Trametes sp em meio líquido extrato de malte
sob agitação tendo serragem da madeira H. creptans como fonte de carbono.
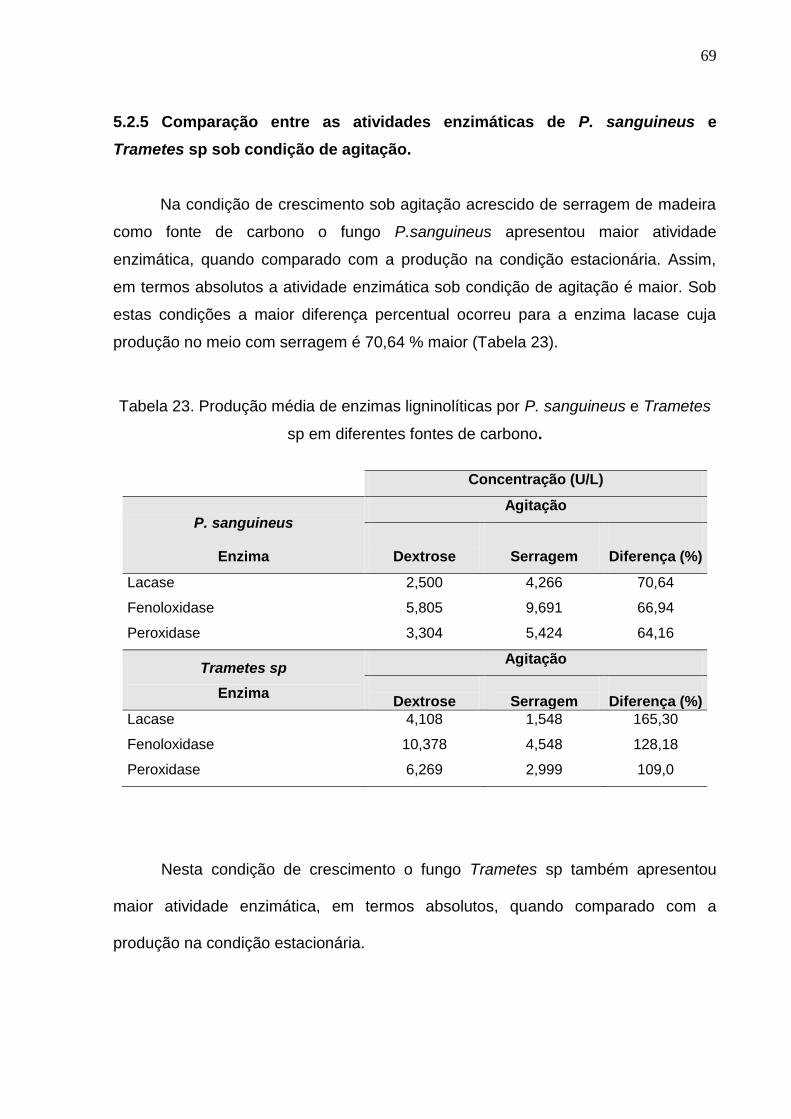
69
5.2.5 Comparação entre as atividades enzimáticas de P. sanguineus e
Trametes sp sob condição de agitação.
Na condição de crescimento sob agitação acrescido de serragem de madeira
como fonte de carbono o fungo P.sanguineus apresentou maior atividade
enzimática, quando comparado com a produção na condição estacionária. Assim,
em termos absolutos a atividade enzimática sob condição de agitação é maior. Sob
estas condições a maior diferença percentual ocorreu para a enzima lacase cuja
produção no meio com serragem é 70,64 % maior (Tabela 23).
Tabela 23. Produção média de enzimas ligninolíticas por P. sanguineus e Trametes
sp em diferentes fontes de carbono.
Concentração (U/L)
P. sanguineus
Enzima
Agitação
Dextrose
Serragem
Diferença (%)
Lacase 2,500 4,266 70,64
Fenoloxidase 5,805 9,691 66,94
Peroxidase 3,304 5,424 64,16
Trametes sp
Enzima
Agitação
Dextrose
Serragem
Diferença (%)
Lacase 4,108 1,548 165,30
Fenoloxidase 10,378 4,548 128,18
Peroxidase 6,269 2,999 109,0
Nesta condição de crescimento o fungo Trametes sp também apresentou
maior atividade enzimática, em termos absolutos, quando comparado com a
produção na condição estacionária.

70
A atividade enzimática também foi maior no meio com glicose como fonte de
carbono e condição de agitação (Tabela 23). A maior diferença percentual ocorreu
para a enzima lacase cuja produção no meio com glicose é 165% maior.
5.3 Atividade Ligninase dos fungos Pycnoporus sanguineus e Trametes sp
De modo geral ambos os fungos apresentaram alta atividade de ligninase,
sendo que P. sanguineus, apresenta máxima atividade dessa enzima (391,95 U/L)
no décimo primeiro dia de crescimento em meio com serragem da madeira de Hura
crepitans como fonte de carbono e em condição estacionária. Enquanto que nessas
mesmas condições, mas sob agitação a maior atividade ocorreu no final do período
de crescimento (décimo sexto dia) do experimento. Em termos de valor médio de
produção enzimática o P. sanguineus no meio com serragem como fonte de carbono
e em condição estacionária, apresentou maior atividade de ligninase (Figura 32). Por
outro lado, no crescimento sob agitação este fungo apresentou maior atividade desta
enzima no meio acrescido de dextrose como fonte de carbono. Ressalta-se,
entretanto, que apesar da maior atividade em meio contendo serragem de madeira
como fonte de carbono e em condição estacionária, o fungo Pycnoporus sanguineus
parece não ser exigente no quesito fonte de carbono uma vez que a produção dessa
enzima mostrou-se alta mesmo no meio usado como controle, ou seja sem
acréscimo de nenhuma fonte de carbono (Figura 32).
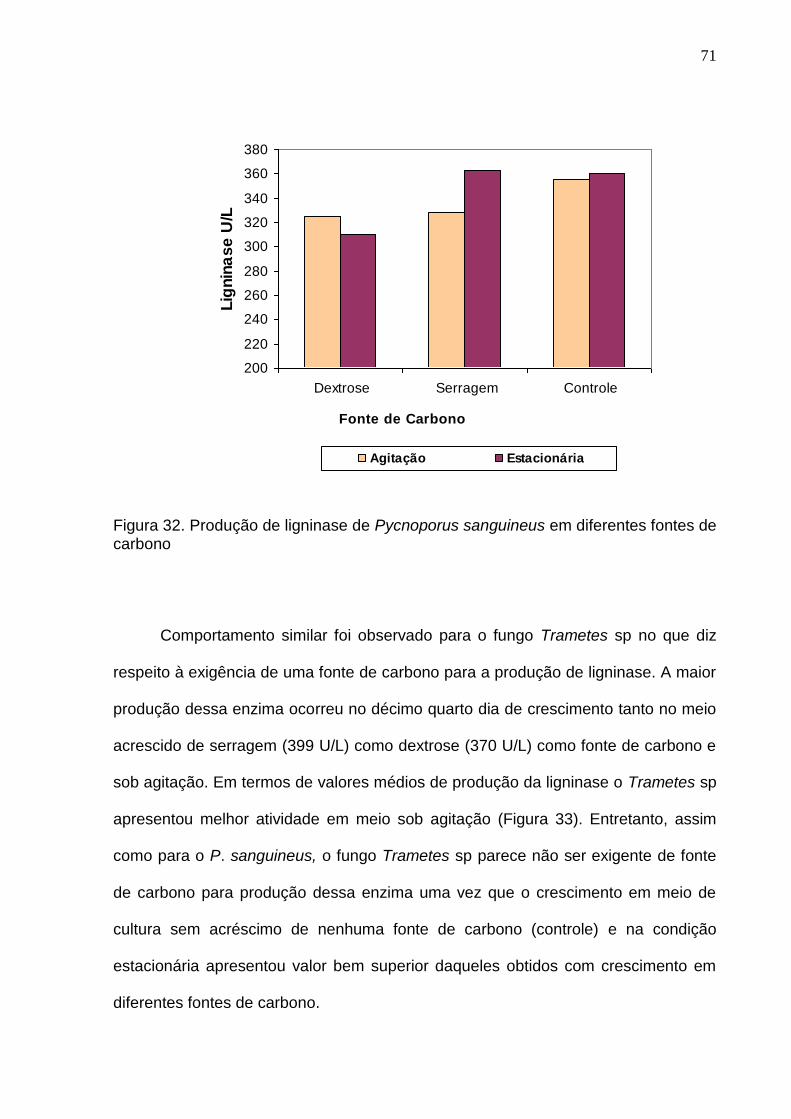
71
Figura 32. Produção de ligninase de Pycnoporus sanguineus em diferentes fontes de carbono
Comportamento similar foi observado para o fungo Trametes sp no que diz
respeito à exigência de uma fonte de carbono para a produção de ligninase. A maior
produção dessa enzima ocorreu no décimo quarto dia de crescimento tanto no meio
acrescido de serragem (399 U/L) como dextrose (370 U/L) como fonte de carbono e
sob agitação. Em termos de valores médios de produção da ligninase o Trametes sp
apresentou melhor atividade em meio sob agitação (Figura 33). Entretanto, assim
como para o P. sanguineus, o fungo Trametes sp parece não ser exigente de fonte
de carbono para produção dessa enzima uma vez que o crescimento em meio de
cultura sem acréscimo de nenhuma fonte de carbono (controle) e na condição
estacionária apresentou valor bem superior daqueles obtidos com crescimento em
diferentes fontes de carbono.
200
220
240
260
280
300
320
340
360
380
Dextrose Serragem Controle
Fonte de Carbono
Lig
nin
as
e U
/L
Agitação Estacionária
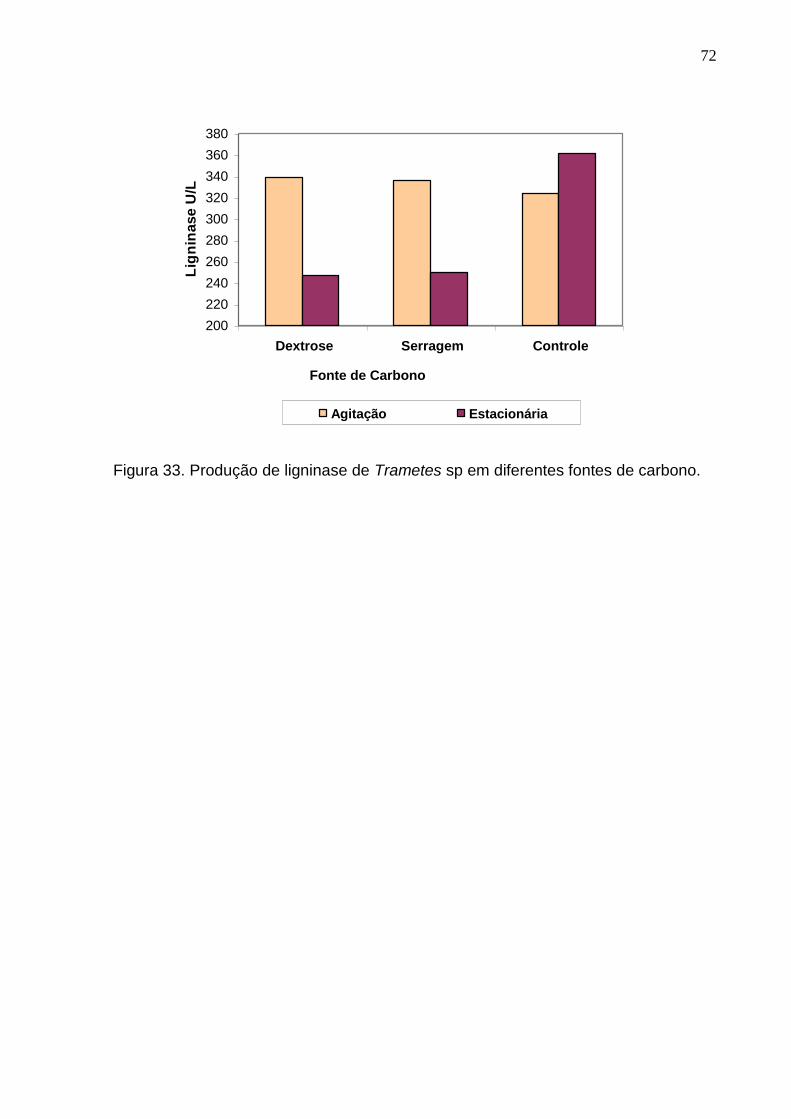
72
Figura 33. Produção de ligninase de Trametes sp em diferentes fontes de carbono.
200
220
240
260
280
300
320
340
360
380
Dextrose Serragem Controle
Fonte de Carbono
Lig
nin
ase U
/L
Agitação Estacionária

73
6. DISCUSSÃO
O método de mensuração linear para avaliar o crescimento dos fungos pode
ser considerado viável, simples, econômico, prático além de que permite observar as
mudanças na colônia do fungo no meio cultivo por um certo período de tempo.
6.1 Temperatura
A mensuração do raio da colônia permite avaliar o crescimento micelial, o
qual ocorre pela extensão da ponta da hifa, enquanto que as partes mais antigas
das hifas são incapazes de crescimento. Embora a hifa mais antiga não seja capaz
de crescer, tem um importante papel de suportar o crescimento da ponta à medida
que novo protoplasma é formado através da hifa e transportado para a ponta pela
corrente citoplasmática ativa (MOORE-LANDECKER, 1982). É este transporte ativo
dos componentes sintetizados que faz possível a rápida taxa de crescimento da
ponta da hifa, o qual para Pycnoporus sanguineus em cultura é em torno de 0,7 a
0,8 mm e Trametes sp entre 0,5 a 0,6 mm crescendo em meio Ágar-malte a
temperatura de 31ºC.

74
Ferraz (1991) observou que a velocidade de crescimento apresenta certa
variabilidade numa mesma condição de cultivo daí a necessidade de se conduzir um
experimento em condições padronizadas.
Os fungos amazônicos mostram um padrão de crescimento crescente à
medida que aumenta a temperatura o que confirma a tendência já amplamente
documentada na literatura cientifica da extrema importância desse parâmetro na
determinação da quantidade e taxa de crescimento de um organismo. O aumento da
temperatura em geral aumenta a atividade enzimática (MOORE-LANDECKER,
1982), uma vez que o movimento das moléculas da enzima e do substrato
aumentam, provocando colisões entre enzima e substrato resultando na formação
de mais produto.
6.2 pH
Outro requisito físico que afeta o crescimento do fungo é o pH, pois pode
influenciar a disponibilidade de certos íons metálicos. Estes podem formar
complexos que se tornam insolúveis em certas faixas de pH. No caso dos fungos
Pycnoporus sanguineus e Trametes sp a relação positiva de crescimento micelial
com o aumento do pH, de uma certa forma mostra que o crescimento no pH 4
(baixo) pode estar diretamente relacionado ao resultado direto da grande
disponibilidade de ferro, enquanto no pH alto (5 -7) pode ser devido o aumento da
atividade enzimática com o pH ótimo alto.

75
Na faixa de pH 5 -7 ocorreu o melhor crescimento de ambos os fungos, sendo
que na faixa de 5-6 verificou-se o máximo crescimento. Este resultado mostra que o
pH apresenta influência no crescimento fúngico, portanto torna-se necessário o
controle do pH. Castro e Silva (1996) também observou similar efeito do pH para os
fungos Antrodia albida e Tyromyces sp, os quais preferiram pH pouco ácido com
uma faixa de pH 5-8 e o ótimo em pH 6.
6.3 Nitrogênio
Os resultados do estudo da fonte de nitrogênio mais adequada ao
crescimento de Pycnoporus sanguineus e Trametes sp indicaram que ambos os
fungos utilizam indiscriminadamente NO3- ou NH4
+ como fonte de nitrogênio. A
utilização de NO3- ou NH4
+ como fonte de nitrogênio proporciona a vantagem de se
empregar qualquer dos dois íons (FERRAZ, 1991), enquanto que vários fungos
necessitam de uma forma específica de nitrogênio, uréia ou aminoácidos (MOORE-
LANDECKER, 1982). Ressalta-se, entretanto, que a velocidade de crescimento
máxima dos fungos Pycnoporus sanguineus e Trametes sp ocorreu na concentração
de 30 mEq /L de nitrogênio.
O nitrogênio presente na madeira e seu papel na deterioração da mesma tem
sido estudado há muito tempo sendo comprovado que fungos xilófagos degradam a
madeira a despeito do seu baixo teor de nitrogênio, sugerindo-se que autólise e re-
uso do nitrogênio é um mecanismo pelo qual o fungo pode conservar a pouca fonte

76
de nitrogênio na madeira (ERIKSSON et al., 1980). Estudos têm mostrado que a
degradação é aumentada se o nitrogênio é adicionado, mas uma alta concentração
parece ser inibitória. Keyser et al. (1978) apud Eriksson et al. (1980), por outro lado,
mostraram que a degradação da lignina parece acontecer somente nas condições
de falta de nitrogênio. Os dados obtidos para o nitrogênio, neste estudo indicam ou
sugerem que os fungos Pycnoporus sanguineus e Trametes sp, apresentam bom
crescimento micelial em todas as concentrações de nitrogênio testadas. Ressalta-se,
entretanto, que esse resultado diz respeito ao avanço da fronteira micelial, e não à
degradação da celulose ou da lignina encontrada na madeira. Pelo menos para os
fungos testados, o nitrogênio não é um fator limitante para a produção micelial.
Recomendam-se estudos complementares da mistura de várias fontes de nitrogênio
para se determinar se as mesmas proporcionam um melhor crescimento do que
(NH4)2SO4 ou NaNO3 sozinhos. Eriksson et al. (1980), mostraram que a mistura de
uréia e NH4H2PO4 proporcionou melhor crescimento para fungos testados, uma vez
que com uréia o nível de pH é mantido mais constante e próximo do pH ótimo para
crescimento dos fungos. Esses autores mostram que com NH4H2PO4 sozinho ocorre
uma queda no nível de pH devido à amônia. Entretanto para os fungos Pycnoporus
sanguineus e Trametes sp testados neste estudo, a amônia não é fator limitante
para produção micelial, uma vez que eles mostraram bom crescimento com
(NH4)2SO4 em todas as concentrações testadas.
6.4 Atividade enzimática
Nem todos os fungos causadores de podridão branca apresentam atividade
de lacase e nem lignina-peroxidase. Cruz (2002), não detectou essas duas enzimas

77
no cultivo de Ceriporiopsis subvernispora mesmo utilizando vários substratos com
diferentes potenciais de oxidação. No presente estudo, tanto P. sanguineus como
Trametes sp, fungos causadores de podridão branca apresentaram maior atividade
enzimática em cultura estacionária, e no meio com serragem como fonte de carbono
produzindo, ligninase-LiP, lacase, fenoloxidase e peroxidase. A produção de LiP por
esses fungos classifica-os como potencial para descoloração de efluente Kraft, pois,
lacase aparentemente pode atacar ou despolimelizar porções fenólicas da lignina,
enquanto que LiP parece ser capaz de oxidar estruturas não fenólica da lignina
(PEREZ & JEFFRIES, 1990). Ligninase também catalisa várias, aparentemente
diferentes, reações não estereoseletivas o que indica o envolvimento de radicais
livres, provavelmente radicais cátion (KIRK & FARRELL, 1987).
Portanto, o “pool” enzimático da cultura do P. sanguineus indica-o como
potencial para descoloração de efluente Kraft uma vez que a presença de ligninase
ou lacase juntamente com Mn-peroxidase na presença de -gluconidase são
importantes na degradação de clorolignina (ESPÓSITO et al., 1991; DURAN, 1992;
CASTRO E SILVA et al., 1993).
A maior atividade enzimática dos fungos no meio com serragem indica que o
crescimento de P. sanguineus e Trametes sp, utilizando substratos insolúveis como
celulose, necessita de um contato direto entre micélio e o substrato em questão.
A maior atividade da ligninase (LiP), em meio de cultura com essa única fonte
de carbono, nos leva a sugerir que alguns compostos derivados do substrato
ligninocelulósico funcionam como indutores para produção de peroxidases

78
ligninolíticas dos fungos Pycnoporus sanguineus e Trametes sp. Os níveis de
atividade de MnP e LiP podem estar relacionados às concentrações de compostos
aromáticos solúveis em água que são liberados no meio (KAPICH et al., 2004).
A atividade máxima das enzimas fenoloxidase e peroxidase em meio
contendo glicose já nos primeiros dias de crescimento, mostra que o fungo
Pycnoporus sanguineus usa imediatamente esse componente na produção dessas
enzimas, decrescendo a produção à medida que o meio vai exaurindo desse
elemento. Por outro lado, a produção no meio com serragem como fonte de carbono,
o fungo precisa buscar esse elemento no material lignocelulósico, cuja
acessibilidade não é tão fácil uma vez que o fungo precisa encontrá-lo na parede
celular dos elementos xilemáticos.
A produção de lacase também pode ser estimulada pela adição de Cu (II) em
concentração milimolar ao meio de cultura com glicose (GALHAUP et al., 2002). No
meio com glicose como principal fonte de carbono, a produção de lacase por P.
sanguineus e Trametes sp somente começou quando a glicose foi consumida do
meio de cultura, uma característica também mencionada por Galhaup et al. (2002),
para o fungo Trametes pubescens.
Um possível efeito regulatório envolvendo intermediários do processo de
biodegradação não pode ser descartado, muito embora não tenha sido objeto do
presente estudo. Entretanto sabe-se que metabólitos relacionados em vários termos
estrutural à lignina podem estimular ou inibir a atividade ligninolítica (MESTER et al.,
1995; HOILANEN et al., 1996). Neste contexto, a presença de 3,4-dimetoxibenzil

79
álcool (álcool veratrílico) pode ser significativa. Embora no presente estudo não
tenhamos caracterizado a produção do álcool veratrílico, sabe-se que esta
substância pode ser “sintetizada por vários fungos, a partir de fenilalanina, e seu
aparecimento nas culturas está associado intimamente com a atividade ligninolítica”
(SOARES, 1998). Este metabólico atuaria como mediador da ação de lignase, onde
essa lignina catalisaria a formação do respectivo cátion radical de álcool veratrílico,
que por sua vez poderia atuar como um agente transferidor de elétrons para outras
estruturas distantes ou não acessíveis do sítio ativo da enzima (AKAMATSU et
al.,1990; HARVEY et al., 1986; KIRK & FARELL., 1987; POP et al., 1990).
No caso dos fungos Pycnoporus sanguineus e Trametes sp, no meio com
serragem como fonte de carbono eles foram mais lentos para atingir níveis mais
elevados da atividade enzimática, mantendo esses níveis, principalmente o P.
sanguineus, por um período ligeiramente maior (dias).
No caso específico da lacase, estudos demonstraram que a sua atividade
pode ser mais ampla, atuando na degradação de estruturas fenólicas e não fenólicas
e que sua capacidade de polimerização pode ser reorientada para a
despolimerização através de substâncias que atuariam como mediadores da ação
da lacase (BOURBONNAIS et al., 1995; BOURBONNAIS & PAICE, 1990). É
possível que tais metabólitos mediadores estejam presentes na serragem de
madeira de Hura creptans, devido ao maior nível de lacase observado neste
substrato.

80
Estudos têm mostrado que a produção de lacase está na dependência tanto
da fonte de carbono quanto na fonte de nitrogênio (GALHAUP at al, 2002;
GIANFREDO et al., 1999). No presente trabalho a maior produção da lacase foi
obtida para o Pycnoporus sanguineus, quando utilizada serragem como única fonte
de carbono e nitrogênio. Para o fungo Trametes sp, entretanto, a atividade é maior
em condições limitantes de nitrogênio, resultado este similar ao encontrado por
Eggert et al. (1996) para outros fungos.
Ressalta-se que as enzimas ligninase, manganês-peroxidase e lacase
envolvidas na degradação da lignina são altamente não especifica com respeito à
faixa do seu substrato. Esses dados são importantes quanto ao aspecto
biotecnológico, considerando-se que são capazes de transformar in vitro ou a
mineralizar ampla faixa de poluentes orgânicos recalcitrantes com estrutura similar a
da lignina (POINTING, 2001; REDDY, 1995). Dessa forma, as informações sobre os
fungos de podridão branca Pycnoporus sanguineus e Trametes sp obtidas neste
estudo com relação à atividade enzimática, sugerem que estes fungos apresentam
potencial para a biorremediação de áreas degradadas e aplicação na industria
papeleira para descoloração de efluente Kraft, entre outros.

81
CONCLUSÃO
Tendo em vista os resultados obtidos, conclui-se que:
Os fungos P. sanguineus e Trametes sp testados a diferentes
temperaturas, apresentaram um ótimo térmico na faixa de 30 –35°C,
mas com produção micelial a partir de 25°C;
Quanto à exigência de nitrogênio para crescimento micelial os fungos
P. sanguineus e Trametes sp mostraram-se tolerantes nas
concentrações usadas (30mEq/L, 60mEq/L, 90mEq/L, 120mEq/L e
150mEq/L);
A faixa de pH ótima para ambos os fungos é de 5 a 7;
Tanto para P. sanguineus quanto para Trametes sp a atividade
enzimática foi maior sob agitação;
A atividade enzimática de Pycnoporus sanguineus foi maior, no que
diz respeito às enzimas ligninolíticas, em meio de cultura

82
suplementado com serragem de madeira como fonte de carbono e
para Trametes sp com glicose como fonte de carbono, ambos sob
agitação;
Sugestões:
Mais estudos são necessários notadamente para verificar outros tipos
de substratos para crescimento e misturas destes, principalmente
testando madeiras com maior densidade, a fim de maximizar a
produção de enzimas ligninolíticas.
Considerando-se a maior atividade em crescimento sob agitação
urge-se necessário testar nova faixa de rotação por minuto a fim de
verificar a influência ou não desse parâmetro na atividade enzimática.
Tanto o fungo Pycnoporus sanguineus quanto o Trametes sp
possuem potencial para aplicação industrial notadamente na indústria
papeleira, considerando a produção de enzimas ligninolíticas
apresentadas;
Sugere-se, portanto que os fungos estudados têm potencial biotecnológico,
podendo-se testar a eficiência destes como degradadores de substâncias tóxicas,
sendo, entretanto, necessários estudos posteriores com a finalidade de explorar o
potencial destas fontes alternativas de enzimas.

83
REFERÊNCIAS BIBLIOGRÄFICAS
ANDER, P. & ERIKSSON, K.E. The Importance of Phenol-oxidase Activity in Lignin degradation by White-rot Fungus Sporotrichum pulverulentum. Arch. Microbiol. 109:1-8. 1976. AKAMATSU, Y.; Ma, D.B.; HIGUCHI, T.; SHIMADA, M. A Novel Enzymetic Decarboxylation of Oxalic Acid by the Lignin peroxidase System of White-rot Fungus Phanerochaete chrysosporium. FEBS Letters 269(1): 261-262. 1990.
AZEVEDO, J. L. Fungos: Genética e Melhoramento de Fungos na Biotecnologia. Revista Biotecnologia Ciência & Desenvolvimento. 2003. Disponível em: <http://www.biotecnologia.com.br/revista/bio01/1hp> Acesso em 05/02/2005. BERGBAUER, M; EGGERT, C; KALNOWSKI, G. Biotratment of pulp mill bleachery effluents with the Coelomycetous fungus Stagonospora gigaspora. Biotechnol.Lett. 14:317-325. 1992. BEZALEL, L.; HADAR, Y.; FU, P.P.; FREEMAN, J.P.; CERNIGLIA, C.E. Initial oxidation products in the metabolism of pyrene, anthracene, fluorine, and dibenzothiophene by the white rot fungus Pleurotus ostreatus. Appl. Environ. Microbiol., 62:2554-2559. 1996. BONONI, V.L.R. Zigomicetos, Bazidiomicetos e Deuteromicetos. Noções básicas de taxonomia e aplicações Biotecnologicas. Instituto de Botânica, secretaria do Estado de Meio Ambiente. São Paulo-SP. 1999. BONONI, V.L.R. Biodegradação de Organoclorados no solo por Basidiomicetos Lignocelulosíticos. In: Microbiologia Ambiental. Jaguariúna: EMBRAPA- CNPMA (243-268). 1997. BOURBONNAIS, R.; PAICE,M.G.; REID, I.D.; LANTHIER, R.; YAGHUCHI, M. Lignin oxidation by Laccase Isozymes from Trametes versicolor and Role of the Mediator 2,2-azinobis-(3-ethylbenzethiazoline-6-sulphonate) in Kraft Lignin Depolymerization. Appl. Environ. Microb. 61: 1876-1880. 1995.

84
BOURBONNAIS, R.; PAICE, M.G. Oxidation of non-Phenolic Substrates : An Expanded Role for Laccase in Lignin Biodegradation. FEBS Letters 267(1): 99-102. 1990. CAMARERO, S.; SARKAR, S.; RUIZ-DUEÑAS, F. J.; MARTINEZ, M. J.; MARTINEZ, A. T. Description of a versatile peroxidase involved in natural degradation of lignin that has both Mn-peroxidase and lignin-peroxidase substrate binding sites. J. Biol. Chem. 274 : 10324-10330. 1999. CAMMAROTA, M.C; SANT`ANNA, G.L. Decolorization of Kraft bleach plant E1 stage effluent in a fungal bioreactor. ENVIRON. TECHNOL. 13:65-71. 1992. CAMMAROTA, M.C. Avaliação de um Bio-Reator Fúngico para Remoção de Cor do Efluente do Branqueamento de Polpa Kraft. Dissertação de Mestrado. Escola de Engenharia Química, UFRJ. R.J. 1991. CASTRO E SILVA, A.; SILVA, M. B. C.; CAVALCANTI, M. A. FUNGOS: O inexplorado potencial enzimático da biodiversidade amazônica. 2002 Disponível em: <http;// www.geocities.com.br/ biodiversidade 2002/fungos>. Acesso em 2/02/2005. CASTRO E SILVA, A. Micromorfologia da Degradação de Madeira da espécie Amazônica Hura creptans L. por fungos ligninolíticos pertencentes a classe Hymenomycetes (Tese de Doutorado). Manaus: INPA/FUA. 1996. CASTRO E SILVA, A; ESPOSITO, E; FERAZ, A,; DURAN, N. Decay of Parkia opossitifolia in Amazônia by Pycnoporus sanguineus and potential use for effluent decolorization. Holzforschung 47: 361-368.1993. CASTRO E SILVA, A; SILVA, M.N; SILVA, M.B; VASCONCELLOS, F.J. 1992. Noções Básicas de Anatomia da Madeira. SEDUC/AM. INSTITUTO DE TECNOLOGIA DA AMAZÔNIA. MANAUS. 1992. CERNIGLIA, C. E. Fungal metabolism of polycyclic aromatic hidrocarbons: past, present and future applications in biorremediation. J. of Ind. Microbiol., 19: 324-333. 1997. CHANG, H.M; JOYCE, T.W; KIRK, T.K. 4th Intein symp.wood Pulping chem. Paris, 159. 1987. CHUA, M. G. S.; CHOI, S. e KIRK, T. K. Mycelium Binding and Depolymerizationof Sinthetic C- Labeled Ligning During Decomposition by Phanerochaete chrysosporium Holzforschung 37 (2): 55-61. 1983. CHUA, M. G. S.; CHEN, .C. L. e CHANG, H. M. C- NMR Spectroscpic Study of Spruce Lignin Degraded by Phanerochaete chrysosporium. Holzforschung 36, 165-172. 1982. CÔTÉ, W. A., Jr. Wood Ultrastructure – An Atlas of Electron Micrographs. Seattle:Univ. of Washington Press. 1967.

85
CRUZ, P. B. S. Estudos das enzimas produzidas por Ceriporiopsis subvermispora (Pilat) Gilbn & Ryv. Durante a biodegradação de Pinus taeda L. Dissertação de Mestrado. Faculdade de Engenharia Química de Lorena, SP. 2002. D'ALMEIDA, G. L. O em “Composição Química dos Materiais em Celulose e Papel –Tecnologia de Fabricação da Pasta Celulósica” vol. I Ed. IPT- São Paulo, SP pp42-54. 1982. DAVIS, S; BURNS, R.G. Decolorization of phenolic effluents by soluble and Immobilized phenol oxidases. Appl. Microbiol. Biotechnol. 32:721-729. 1978. DURAN, N. ESPOSITO, E. Potential applications of oxidative enzymes and phenoloxidases-like conpounds in wasterwater and soil tratment: A Review. Appl.Catalysis B. Environ., 28,83-99. 2000 DURAN, N.; RODRIGUEZ, J.; GÓMEZ, E., CAMPOS, V. & BAEZA, J. – Bimass photochemistry – XI: Photochemical and pre-treatment of cellulose and its fungal degradation. Biotechnol. Bioeng., 31: 215-219.1988. DURAN, N., ESPOSITO, E. Biodegradação de Lignina e tratamento de Efluentes por Fungos Ligninolíticos. In: Microbiologia Ambiental. Jaguariúna: EMBRAPA - CNPMA. 1997. DURÁN, N. Reduction of chemical oxygem demand plant effluent by a combination of photochemical and method. Proc. Second Symp. Chem. Lignin Other Wood Components, Campinas, SP. Brazil 3: 323-333.1992.
ERIKSSON, K.E. L. Biotechnology in the pulp and paper industry. Wood Science and Technology, v.24, p. 79-101,1990. ERIKSSON, K.; ERUNEWALD, A.; VALLANDER, L. Studies of growth conditions in wood for tree white-rot Fungi and their cellulas less mutants. Biotech. Bioeng. 22 : 363-376. 1980.
EGGERT, C.; TEMP.U.; DEAN, J.F.D.;ERISSON, K-E.L. The ligninolytic system of the white-rot fungus Pycnoporus cinnabarinus : purification and characterization of the laccase. Appl. Environ. Microb. 62: 1151-1158. 1996.
EGGERT, C.;TEMP,U.; ERIKSSON, K.E. Laccase-producing white-rot gungus lacking lignin peroxidase, and manganese peroxidase. In: JEFFRIES, T.; VTKARI, L. (ed). Enzyme for pulps and paper processing. Washington: american Chemical Society, 1996. p. 130-150. ( ACS Symposium Series, 655). 1996. ESAU, K. em “Plant Anatomy” 2th ed. John Wiley International - New York pp. 35, 49. 1965. ESPOSITO, E. e AZEVEDO, J.L. Fungos: uma introdução à biologia, bioquímica e biotecnologia. Caxias do Sul: EDUCS, 2004.

86
ESPOSITO, E. Processos Envolvidos no Tratamento Biológico de Efluentes: Utilização de um Biorreator de Leito Recheado. Dissertação de Mestrado. Universidade Estadual de Campinas-SP. 1992. ESPOSITO, E.; CANHOS, V. P.; DURÁN, N. Screening of lignin degrading fungi for removal of color from Kraff mill wasterwater with no additional extra carbon-source. Biotechnol. Lett 13: 571-576.1991 FENGEL, D. e WEGENER, G. in Wood: Chermistry, Ultrastructure, Reactions. Wlater de Gruyter, Berlin pp 66-75. 1989. FERRAZ. A. L. Fungos Decompositores de Materiais Lignocelulósicos. in: Fungos: uma introdução a biologia, bioquímica e biotecnologia /Orgs. Elisa Espósito e João Lúcio de Azevedo. Caxias do Sul: Educs. 2004. FERRAZ, A.L. Contribuição ao Estudo do Ascomiceto Chrysonilia sitophila: Biodegradação de madeira e seus componentes. Tese de Doutorado. UNICAMPI. SP. 131pp. 1991. FITZSIMONS, R; EK, M; ERIKSSOM, K.E. Anaerobic degradation of chlorinated organic compounds of different molecular massas in bleach plant efluents. Environ. Sci. Technol, 24:1744-1751. 1990. FORSS, K.; JOKINEN, K.; SAVOLANEN, M; WILLIANSON, H. Utilization of enzymes for effluent treatment in the pulp and paper industry. Paperi Ja Puu 10:1108-1110. 1989. FREUDENBERG, K. Liginin: Its constituition and formation fron p-hidroxycinmamyl alcohols. Science 148: 595. 1965. GALHAUP, C.;WAGNER, H.; HINTERSTOISSER, B.; HALTRICH, D. Increased production of laccase by the wood-degrading basidiomycetes Trametes pubescens. Enzymme and Microbial Technology 30 (4) : 529-536. 2002. GIANFREDO, L.; Xu, F.; BOLLAG, J.M. Laccases: a useful group of oxidoreductase enzymes. Bioremediation Journal 3: 1-25. 1999.
GLENN, J.K; GOLD, M.H. Purification and characterization of extracellular Mn (II)- Dependent Peroxidase from the liginin-degrading basidiomycete Phanerochaete chrysosporium. Arsh. Biochem. Biophis. 242:329-334. 1985. GLENN, J.K. An extracellular H2O2 – Requiring enzyme preparation involved in liginin biodegradation by the white-rot basidiomycete Phanerochaete chrysosporium. Biochen. Biophys. Res. Commun.114:1077-1083. 1983. GOLD, M.H; KUWAHARA, M; OHANA, A.; GLENN, J.K. Purification and characterization of an extracellular H2O2-requiring diaril propane oxygenase from the white-rot basidiomycete Phanerochaete chrysosporium Arsh. Biochem. Biophis, 234:353-358. 1984.

87
HAMLYN, P.F. Fungal biotechnology. British Mycol. Soc. Newletter 3: 45-56. 1998. HAMMEL, K.E. Organopullutant degradation by ligninolytic fungi. Enzyme microbi. Technol., 11:776-782. 1989. HATAKKA, A. Lignin-Modifying Enzymes from Selected White-Rot Fungi: Production and Role in Lignin Degradation. FEMS Microbiological Reviews 13, 125-135. 1994 HAWSKSWORTH, D. L. The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycological Research 95:641-655.1991. HIGUCHI, T. Lignin Biochemistry: Biosynthensis and Biodegradation Wood. Science Technology 24: 23-63. 1990. HIGUCHI, T. Mechanisms of liginin degradation by liginin peroxidase and laccase of white-rot fungi Biogenesis and Biodegradation of Plant cell polymers. ACS Simposium Series 399:482-502. 1989 HOILANEN, A M.; LUNDELL,T.; VARES, T.; HATAKKA, A . Managanese and Malonate are Individual Regulatiors for the Production of Lignin and Managanese Peroxidase Isozymes and in the Degradation of Lignin by Phlebia radiata. Appl. Microb. Biotech. 45:792-799. 1996. KAPICH, A. N.; PRIOR, A.; BOTHA, A.; GALKIN, S.; LUNDELL, T.; HATAKKA, A. Effect of lignocellulose-containing substrates on production of ligninolytic peroxidases in submerged culture of Phanerochaete chrysosporium. Enzyme and Microbial Techonology 34 (2) : 187-195. 2004. KAUTSKY, H. The impact of pulp-mill efluent on phytobenthic communites in the Baltic sea. Ambio, v.21, p. 308-313. 1992 KERSTEN, P.J.; KALYANARAMAN, B.; HAMMEL, K.E et al. Comparison of liginin peroxidase, horseradish peroxidase and laccase in the oxidation of methoxy-benzenes. Boichen. J. 268:475-480. 1990. KEYSER, P.; KIRK, T. K.; ZEIKUS, J.G.J. Bacteriol. 135 : 790-810.1978. KIRK,T.K; CULLEN, D. Enzimology and molecular genetcs of wood degradation by white-rot fungi. In; JOHN WILEV & SONS. Enviromentally Friendly Technologies for the Pulp and paper Industry. New York, p. 273-308. 1998. KIRK,T.K; FARRELL, R.L. Enzimática “combustion” :The microbial degradation of lignin.Ann. Rev. Microbiol. 41:465-505. 1987. KUWAHARA, M; GLENN, J.K; MORGAN, M.A et al. Separation and characterization of two extracellular H2O2 dependent oxidase from liginolytic cultures of Phanerochaet chrysosporium. FEBS Lett.169 (2) 247-250. 1984. LACAZ, C.L. et al.;. Tratado de Micologia Médica. 9ª ed. São Paulo: ARVIER. 2002

88
LANKINEN, V.P; INKEROINEN,M.M.; PELLINEN, J; HATAKKA, A.I. The on set of liginin modifyng enzimes. Decrease of AOX and color removal by white-rot fungi grown on bleach plant eflluent. Wood. Sci. Tech. 24:189-195. 1991 LEONARDSSON, K. Long-term ecological effect of bleaching pulp mill efluents on bentthic macrofauna in the Gulf of Bothnia. Ambio, v.22,p. 359-362. 1993. LEPAGE, E. S. et al.; 1986. Manual de Preservação de Madeiras. Vol. I. São Paulo: IPT, 1986. MEHTA, V.; GUPTA, J. K. Biotechnology in the pulp and paper industry. Research Industry, 36:161-16. 1991. MESTER, T.; JONG, E.; FILED, J.A . Manganese Regulation of Veratryul Alcohol in White-rot Fungi and its Indirect Efefcts on Lignin Peroxidase. Appl. Environ. Microb. 61(5): 1881-1887. 1995. MICHEL, FC; DASS, B; GRULKE, E.A et al. Role of manganese peroxidases and lignin peroxidases of Phanerochaetes chrysosporium in the decolorization of craft bleach plant effluent. Appl. Env. Microbiol. 57(8) : 2368-2375. 1991 MOORE-LANDECKER, E. Fundamentals of the FUNGI. New Jersey: Prentice-Hall, INC. Englewood Cliffs.578pp. 1982 MORAIS, A. M.L. et al.; FUNGOS: Ferramentas na saúde pública As utilidades de um antigo vilão. Revista Biotecnologia Ciência & Desenvolvimento. 2003. Disponível em < http://www.biotecnologia.com.br/revista/bio07/7_b.asp> Acesso em 05/02/2005. NEUFELD, P. Micologia. Curso de Pós-Graduação em Biologia Parasitária do Instituto Oswaldo Cruz. Rio de Janeiro - RJ. 1997. NIMZ, H. Beech lignin; Proposal of a constitutional scheme. Angew. Chem. 13(5):313-329.1974. ONYSKO, K. A. Biological bleaching of chemical pulps; a review. Biotechnology Advance, v.11, p.179-198, 1993. PAICE, M.G; JURASSEk, L; HO,C.; BOURBONNAIS, R; ARCHIBALD, F.S. Direct Biological Bleaching of Hardwood kraft pulp with fungus Coriolus versicolor. Tappi J. 72:217-221.1989. PALMER, J.M.; HARVEY, P. J.; SCHOEMAKER, H. E. The role of peroxidases, radical cation and oxigen in the degradatrion of liginin. Phil. Trans. R. Soc. London. A31: 495-502. 1987. PERZ, J. & JEFFRIES, T. W. Mineralization OF 14C- Ring-Labeled Synthetic Lignin Correlates with the Procuction of Lignin peroxidase not as Manganel Peroxidase and laccase. Appl. Environ. Microbiol., 56 : 1806-1812. 1990.

89
POINTING, S.B. Feasibility of biomediation by white-rot fungi. Appl. Microbiol Biotchenol 57 : 20-33. 2001. POP,J.L; KALYANARAMAN,B.; KIRK, T.K. Lignin Peroxidase oxidation of Mn2+ in the Presence of Varatryl Alcohol, Malonic or Oxalic Acid, and Oxygen. Biochemistry 29: 10475-10480. 1990. PRESTON, R. D. The physical biology of plant cell walls. London: Chapman & Hall Ltd., 1974.
PUTZKE, J.; PUTZKE, M.T.L. Os Reinos Dos Fungos. Vol. 2. Santa Cruz do Sul: EDUNISC, 2002. PUTZKE, J.; PUTZKE, M.T.L. Os Reinos Dos Fungos. Vol. 1. 2ed . Santa Cruz do Sul: EDUNISC, 2004.
REDDY, C. A. The potential for white –rot fungi in the treatment of pollutants. Cun. Opin. Biotechnol 6 : 320-28. 1995. REID, I.D; PEACE, M.G; HO,C.; JURASEK, L. Biological bleaching of softwood kraft pulp with the fungus Trametes (Coriolus) versicolor. Tappi J. 73:150-164. 1990. ROY-ARCAND, L; ARCHBALD, F.S. Direct Dechlorination of chlorophenolic compounds by laccase from Trametes (Coriolus) versicolor. Enzyme Microb. Techinol. 13:194-201. 1991 RODRIGUEZ, J. Estudo do sistema ligninolitico do ascomiceto Chrysonilia sitophila. Tese de Doutorado. Instituto de Química, UNICAMP. SP, 1990. SAPARRAT, M. C. N.; GUILLEN, F.; ARAMBARRI, A. M.; MARTINEZ, A. T.; MARTINEZ, A. T.. Induction, Isolation, and characterization of two laccases from the white rot basidiomycete Coriolopsis rigida. Appl. Env. Microb. 68 (4) : 1534-1540. 2002 SILVA, M.; MINTER, D.W. Fungi from Brazil recorded by Batista and co-workers. Mycol. Papers, 169:1-585. 1995. SILVA, M.; ESPOSITO, E. O papel dos fungos na recuperação ambiental. In Fungos: uma introdução à Biologia, Bioquímica e Biotecnologia. Orgs. Elisa Espósito e João Lúcio de Azevedo. Caxias do Sul: EDUCS, 2004. SOARES, C. H. L. Estudos Mecanisticos da Degradação de Efluentes de Indústrias de Papel e Celulose por Fungos Basidiomicetos Degradadores de Madeira. Tese de Doutorado. Universidade Estadual de Campinas – SP,1998. SUNDARAMOORTHY, M; KISHI,K; GOLD, M.H et al. Preliminary crystallografic analysis of manganese peroxidase from Phanerochaete chrysosporium. Mol. Biol. 238:845-848. 1994.

90
SZKLARZ, G. D.; ANTBUS, R. K.; SINSABAUGH, R. L.; LINKINS, A. Purification of Phenol Oxidases and peroxidase by Wood-rotting Fungi. Mycol. 81: 234-241. 1989. THURSTON, F. The structure and function of fungal laccases. Microbiology. 140 19-26. 1994. TEIXEIRA, M. F. S.; PORTO, A. L. F.; ROCHA, W. C. & FERNANDES, O. C. C. Produção de Compostos Bioativos de Interesse Industrial. Universidade do Amazonas. Fundação UNISOL. Manaus – AM, 2001 TIEN,M; KIRK , T. K. Lignin-degrading enzyme from the hymenomycete Phanerochaete chrysosporium burds. Science. 221:661-663. 1983. TIEN,M; KIRK , T. K. Lignin-degrading enzyme from Phanerochaete chrysosporium: Purification characterization and catalytic. Properties of a unique H2O2 - requering oxigenase. Proc. Nat. Acad. Sci. (USA): 81:2280-2289. 1984. VANDEVIVERE, P.; VERSTRAEDE, W. Environmental applications. In: Basic Biotechnology Edited by Colin Ratledge and Bjorn Kristianson. CAMBRIDGE Univ. Press, 531-557 pp. UK. 2001. VOLC, J.; KUBATOVA, E.; DANIEL, G.; PRIKRYLOVA, V. Only C –2 specific glucose oxidase activity is expressend in ligninolytic cultures of the white rot fungus Phanerochaete chrysosporium. 1996





























