Embed Size (px)
Citation preview

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
73
1.11 สารพันธุกรรม (Genetic material) และองคประกอบทางเคมีของ DNA
1.11.1 ประวตัิและการคนพบสารพันธุกรรม
มนุษยไดนําความรูทางพันธุศาสตรมาใชนับพันปกอนที่จะเกิดวิชาพันธุศาสตรขึ้น เชน การคัดเลือกสายพันธุของสิ่งมีชีวิตเพ่ือนํามาใชทางการเกษตร มนุษยใชพันธุศาสตรในการปรับปรุงพันธุสิ่งตาง ๆ เพ่ือประโยชนของตนเองมานับพันป แตเพ่ิงจะสามารถอธิบายไดวาเหตุใดลูกจึงมักจะมีลักษณะเหมือนพอแม เหตุใดการปกชําจึงใหผลผลิตที่มีลักษณะคลายคลึงกับตนแมมากกวาการนําเมล็ดมาเพาะในชวงศตวรรษที่ 20 จากที่เมนเดลคนพบและอธิบายกฎเกณฑการถายทอดทางพันธุกรรม มาจนปจจุบันซึ่งสามารถปรับเปลี่ยนพันธุกรรมของสิ่งมีชีวิตโดยกระบวนการพันธุวิศวกรรม หน่ึงศตวรรษที่ผานมามีการคนพบสิ่งใหม ๆ วิธีการใหม ๆ เพ่ือพัฒนาความเขาใจทางพันธุศาสตรตลอดเวลาเชน
ป 1866 มีการตีพิมพผลงานของเมนเดล ป 1869 ฟรีดริช (Johann Friedrich Miescher) คนพบ nuclein (ซ่ึงปจจุบันพบวา คือ DNA) ในนวิเคลียส ป 1905 บาทีสัน ตั้งชื่อวทิยาศาสตรสาขานี้วาพันธุศาสตร ป 1908 กฎฮารดีไวเบิรกถูกสรางขึ้น ป 1909 Archibald Garrod พบความสัมพันธระหวาง DNA กับ Protein ป 1910 ยีนบนโครโมโซมเพศของแมลงหวี่ควบคุมลักษณะสีตา
ป 1912 Sir William Henry Bragg และลูกชาย Sir William Lawrence Bragg พบวา x – ray diffraction สามารถบอกโครงสรางอะตอมของผลึกได ป 1924 จากการศึกษาดวยกลอง microscope พบวาโครโมโซมมี DNA และโปรตีน ป 1927 พบวารังสีเอกซทําใหเกิดการกลายพันธุ ป 1928 กริฟฟท (Franklin Griffith) พบการสงสารพันธุกรรมขามตัวของแบคทีเรีย (transformation) และทราบวาสารพันธุกรรมเปน heat stable chemical ป 1931 แมคคลินทอค (Babara McClintock) พบการจัดเรียงตัวใหมของยีน ป 1941 บีเด้ิลและทาตั้ม ( George Beadle และ Edward Tatum) ตั้งกฎ หน่ึงยีนหนึ่งเอนไซม ป 1944 Oswald Avery, Maclyn McCarty และ Colin Macleod วเิคราะหวาสารที่ถูก transformation
ในการทดลองของกริฟฟทคือ DNA ป 1949 Erwin Chargaff พบวาจํานวน DNA และไนโตรจีนัสเบสเปลี่ยนแปลงตามสปชีส โดย A=T
และ G=C ในทุกสปชีส
เมนเดล

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
74
ป 1953 วัตสันและคริก ( James D. Watson และ Fransis H.C. Crick) เสนอโครงสราง DNA จาก X-ray diffraction และขอมูลจากชารกาฟฟ ซึ่งสงผลใหไดรับรางวัลโนเบลในป 1962 (รวมกับ Maurice Wilkins (ผูคนพบ DNA X-ray diffraction พรอมกับ Rosalind Franklin) และคลิกไดเสียชีวติในวนัที่ 28 กรกฏาคม 2547
ป 1958 เมเซลสันพบวาการจําลองตัวของ DNA เปนแบบกึ่งอนุรักษ ป 1961 Sidney Brenner และ F.H.C. Crick พบวา 3 นิวคลีโอไทดเปน 1 codon ที่จําเพาะตอ กรดอะมิโน ป 1964 ยานอฟกี้แสดงความสัมพันธระหวางยีนกับโพลีเปปไทด ป 1966 ไนเรนเบิรกวิเคราะหรหัสพันธกุรรมไดสมบูรณ ป 1970 สมิธ (Hamilton Smith) และนาธานแยกเอนไซมตัดจําเพาะไดอยางสมบูรณ ป 1972 Stanley Cohen และ Herbert Boyer สราง DNA สายผสมชนิดแรก ป 1977 คณะของแมกแซม และคณะของแซงเจอร พบการหาลําดับเบส ป 1978 Somatostatin เปน regulates human growth hormone โปรตีนตวัแรกที่ถูกใชใน recombinant technology ป 1982 พัลมิเตอรสราง “supermice” จากการฉีดยีนควบคุมการสราง growh hormone ใหหนูเล็ก ป 1985 มูลลิส พบวธิีเพ่ิมปริมาณ DNA นอกรางกายสิ่งมีชีวติที่เรยีกวา PCR ป 1988 เริ่มโครงการ Human genome project ป 2001 ตีพิมพผลงานโครงการ Human genome project (โครงการสําเร็จในเดือนมิถุนายน 2000)
การคนพบของเมนเดลบอกใหรูวาลักษณะทางพันธุกรรมถูกควบคุมโดยหนวยพันธุกรรม (ซ่ึงภายหลังเรียกวายีน) ที่สามารถถายทอดจากพอแมไปสูลูกได ในชวงเวลานั้นนักวิทยาศาสตรสวนมากเขาใจวาเปนสารพวกโปรตีนเพราะโปรตีนมีกรดอะมิโนมากถึง 20 ชนิด การเรียงตัวสลับไปมาของกรดอะมิโนนาจะเพียงพอตอการสรางโปรตีนหลากชนิดที่เซลลใช และนาจะมากพอที่จะควบคุมลักษณะของสิ่งมีชีวิตได เม่ือฟรีดริชไดคนพบนิวคลีอิน (สารที่อยูในนิวเคลียสและไมสามารถยอยไดดวย เปปซิน) รวมถึงการคนพบตอไปวานิวคลีอินมีคุณสมบัติเปนกรด (จึงเรียกวากรดนิวคลีอิกแทน) และ ฟลูเกนไดพัฒนาการยอมสีกรดนิวคลิอิคดวยสีฟุคซินทําใหทราบวากรดนิวคลิอิกนั้นเกาะอยูที่โครโมโซมจนกระทั่งกริฟฟทไดคนพบการสงถายโครโมโซม (transformation) ในแบคทีเรีย จึงเกิดแนวคิดวา “กรดนิวคลีอิกนาจะเปนสารพันธุกรรม”

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
75
การทดลองตาง ๆ เพื่อตรวจสอบวาชวีโมเลกุลใดที่เปนสารพนัธุกรรม
การทดลองของกริฟฟทโดยใช Streptococcus pneumoniae (Pneumococcus) ซ่ึงเปนแบคทีเรียที่ทําใหเกิดโรคปอดบวมในหนูมาทําการทดลอง แบคทีเรียกลุมน้ีมีหลายสายพันธุ และมีความแตกตางกันมากในการกอโรค อยางไรก็ตามอาจแบงแบคทีเรียกลุมน้ีออกเปน 2 พวกใหญ ๆ คือ
1. พวกที่ไมกอโรครุนแรง (avirulent) เปน S. pneumoniae พวกที่ไมมีแคบซูลหุม ( R ) 2. พวกที่กอโรคอยางรุนแรง (virulent) เปน S. pneumoniae พวกที่มแีคบซูลหุม ( S )
กริฟฟททําการทดลองโดยฉีดแบคทีเรยีแบบตาง ๆ เขาไปในหนูดังน้ี การทดลองที่ 1 ฉีดสายพันธุที่ไมมีแคบซูลหุม (ทําใหแบคทีเรยีน้ันมีลักษณะผวิขรขุระ) ใหกับหนู การทดลองที่ 2 ฉีดสายพันธุที่มีแคบซูลหุม (ทําใหแบคทีเรยีน้ันมีลักษณะผวิเรียบ) ใหกับหนู การทดลองที่ 3 นําสายพันธุที่มีแคบซูลหุมไปใหความรอน (ทําใหเซลลตาย) แลวฉีดใหกับหนู การทดลองที่ 4 นําสายพันธุที่มีแคบซูลหุมไปใหความรอนแลวใสสายพันธุที่ไมมีแคบซูลหุมลงไปกอนฉีด
ผลการทดลอง 1. ( R ) 2. ( S ) ตาย 3. (heated S) 4. (heated S + R) ตาย
ภาพที่ 1.56 การทดลองของกริฟฟทโดยใชแบคทีเรยี S. Pneumoniae ฉีดในหนู
นักเรียนคิดวากริฟฟทไดอะไรจากผลการทดลองครั้งน้ี …………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
76
นักเรียนคิดวาถาเจาะเลือดและนํามาตรวจสอบสายพันธุของแบคทีเรยีที่อยูในเลือดหนูแตละการทดลองนาจะมีผลเชนไร เหตุใดจึงเปนเชนนั้น การทดลองที่ 1 จะพบแบคทีเรียสายพนัธุ...…………………....…………………...................................
เพราะ ……………………………………………………………………………………………… การทดลองที่ 2 จะพบแบคทีเรียสายพนัธุ ……………….…….……..…………………………………….
เพราะ ……………………………………………………………………………………………… การทดลองที่ 3 จะพบแบคทีเรียสายพนัธุ …………………………………………………………………
เพราะ ……………………………………………………………………………………………… การทดลองที่ 4 จะพบแบคทีเรียสายพนัธุ ………………………………………………………………..
เพราะ ………………………………………………………………………………………………
การทดลองของกริฟฟทเปนการบอกใหทราบวามี “บางอยาง” ที่ควบคุมลักษณะทางพันธุกรรมที่สามารถถายทอดจากแบคทีเรียสายพันธุ S ไปยังสายพันธุ R (เพราะสายพันธุ R มีการเปลี่ยนแปลงใหมีคุณสมบัตเิชนเดียวกับสายพันธุ S) และสิ่งน้ันไมนาจะเปนโปรตนีเพราะความรอนนาจะทําใหโปรตีนเสียสภาพไปแลว การทดลองของกริฟฟทไดถูกปรับปรุงเพ่ือยืนยันวาสิง่ที่ถูกถายทอดจากแบคทีเรยีสายพันธุ S ไปยังสายพันธุ R คือ DNA โดยกลุมของเอเวอรี่ ไดทาํการทดลองใชเอนไซม DNase (ซ่ึงเปนเอนไซมที่ใชในการยอย/ทําให DNA เสียสภาพ) และโปรตเีอส (protease ซ่ึงเปนเอนไซมที่ใชในการยอยโปรตีน) ดังน้ี
1. สารสกัดหยาบจากแบคทีเรียสายพันธุ S ผสมกับ แบคทีเรยีสายพันธุ R 2. สารสกัดหยาบจากแบคทีเรียสายพันธุ S ผสมกับ แบคทีเรยีสายพันธุ R กับ DNase 3. สารสกัดหยาบจากแบคทีเรียสายพันธุ S ผสมกับ แบคทีเรยีสายพันธุ R กับ Protease
จากการทดลองนี้นักเรียนคิดวาสมมตฐิานของเอเวอรแีละคณะ เปนอยางไร ………………………………………………………………….………………………………………………………………………………………………………………………………………………………………ตัวแปรควบคุมคอื ……………ตัวแปรตนคือ …………………ตวัแปรตามคือ.............................. ผลการทดลอง
DNase Protease (1) (2) (3) แบคทีเรยีสายพันธุ …… แบคทีเรียสายพันธุ …… แบคทีเรยีสายพันธุ………..
แบคทีเรียสายพันธุ Rผสมกับสารสกัดจากแบคทีเรียสายพันธุ S

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
77
เอเวอรี่และคณะไดสรุปผลการทดลองวา DNA เปนสารที่ทําใหเกิดการถายทอดทางพันธุกรรม เพราะเมื่อแบคทีเรียสายพนัธุที่ไมมีแคปซูล (R) ไดรับ DNA ที่สกัดจากแบคทีเรียสายพันธุ S เขาไปแลวจะสามารถสรางแคบซูลไดเหมือนสายพันธุ S นอกจากการทดลองของกลุมกริฟฟทและกลุมเอเวอรี่แลวยังมีการทดลองของเฮอรชี่ (Herchey) ซ่ึงทําในไวรัส โดยการติดฉลากไวรัสดวยสารกัมมันตรังสีที่ฟอสฟอรัส (32P) ซ่ึงเปนองคประกอบของกรดนิวคลิอิก และที่ซัลเฟอร (35S) ซ่ึงเปนองคประกอบของโปรตีนหอหุม จากนั้นติดตามผลการถายทอดสารตาง ๆ ไปยังรุนลูก และพบรุนลูกที่มีฟอสฟอรัสที่ติดกัมมันตรังสี นอกจากนั้นก็มีการทดลองของคอลลาจและซิงเจอร (Collarge and Singer) ที่พบวา RNA ก็เปนสารพันธุกรรมที่ถายทอดลักษณะทางพันธุกรรมไปยังรุนลูกไดเชนกัน
ภาพประกอบการอธิบายการทดลองของเฮอรชีส ภาพที่ 1.58 ประกอบการอธิบายการทดลองของเฮอรชีส
ภาพที่ 1.57 การทดลองของเฮอรชี่ (Herchey) ซ่ึงทําในไวรสั โดยการติดฉลากไวรัส 1.11.2 องคประกอบทางเคมีของ DNA
จากการศึกษาวิจัยที่พบวา DNA เปนสารพันธุกรรม มีคําถามตอไปวา DNA สามารถเก็บขอมูลและถายทอดขอมูลจากรุนหนึ่งไปสูรุนหน่ึงไดอยางไร การศึกษาเบื้องตนพบวาบนสาย DNA ประกอบไปดวยยีน (gene) หลายยีน
ยีน หมายถึง หนวยนําลักษณะทางพันธุกรรม เปนหนวยที่ทําหนาที่ถายทอดลักษณะตาง ๆ ไปยังรุน (generation) ถัดไป
DNA มีหนวยยอยเปนกรดนิวคลีอิก (nucleic acid) มาเรียงตอกันเปนสายยาวซึ่งเรียกวาสายโพลีนิวคลีโอไทดหรือสาย DNA สายดีเอ็นเอ 2 เสนจับกันเปนเกลียวคูเวียนขวา (right - handed double helix structure)
ติดฉลากที่ S ซึ่งเปนองคประกอบของโปรตีน
ติดฉลากที่ P ซึ่งเปนองคประกอบของกรดนิวคลิอิก
ไมพบสารกัมมันตรังสี
พบสารกัมมันตรังสี

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
78
โครงสรางของดีเอ็นเอประกอบดวยโพลีนิวคลีโอไทด 2 เสน ซ่ึงเกิดจากการรวมตัวกันของหลายๆ นิวคลีโอไทด (nucleotides) นักวิทยาศาสตรชาวเยอรมันชื่อโคสเซล (A. Kossel) ไดศึกษาองคประกอบของกรดนิวคลีอิกของดีเอ็นเอ พบวามีไนโตรจีนัสเบส (nitrogenous base) เปนองคประกอบ 4 ชนิดคือ A T C และ G ตอมาเลวีน (A.T. Levene) พบวาไนโตรจีนัสเบสสรางพันธะกับนํ้าตาลดีออกซีไรโบส ซ่ึงเปนนํ้าตาลที่มีคารบอน 5 อะตอม และสรางพันธะกับหมูฟอสเฟต การศึกษาตอมาพบวาเบสใน RNA จะสรางพันธะกับนํ้าตาลไรโบสและฟอสเฟต ไนโตรจีนัสเบส
สามารถแบงไดเปน 2 กลุมใหญคือ 1. เบสไพริมิดีน (pyrimidine base) มีวงแหวน 1 วง ไดแก ไทมีน (thymine; T), ไซโทซีน (cytosine; C) ในดีเอ็นเอและยูราซิล (uracil; U) ในอารเอ็นเอ (RNA) ตําแหนงของเบสที่มีไนโตรเจนของเบสไพริมิดีน คือตําแหนงที่ 1 และ 3 จึงจัดเปนเบสชนิดที่มีไนโตรเจนเปนองคประกอบ (nitrogenous base) เบสไพริมิดีนมีสวนเบสตําแหนงที่ 1 เกาะกับนํ้าตาลที่ตําแหนงที่ 1 ดวยพันธะ β –N- glycosyl
2. เบสพิวรีน (purine base) มีวงแหวน 2 วง ไดแก อะดีนีน (adenine; A) และกวานีน (guanine; G) ตําแหนงของเบสที่มีไนโตรเจนของ adenine คือตําแหนงที่ 1, 3, 7 และ 9 เบสพิวรีน จัดเปนเบสชนิด ที่มีไนโตรเจนเปนองคประกอบ (nitrogenous base) สวนของเบสตําแหนงที่ 9 จะมาเกาะกับนํ้าตาลทีต่ําแหนงที่ 1 ดวย
พันธะ β –N- glycosyl
เบสและน้ําตาลรวมเรียกชื่อวา นิวคลีโอไซด (nucleoside) และถาเติมหมูฟอสเฟตเขาไปจะกลายเปน นิวคลีโอไทด (nucleotide) หรือกลาวไดวานิวคลีโอไทดเปน phosphatester ของ นิวคลิโอไซด
ตัวอยางเชน ถาเบสเปนไทมีนเกาะกับ นํ้ าตาล ดีออกซี ไรโบส จะ เรียกนิ วคลี โอไทด น้ี ว า ดีออกซี ไทมิ ดีนโมโนฟอสเฟต
(deoxythimidine monophosphate) หรือ ดีออกซีไทมิดีน 5’ ฟอสเฟต (deoxythymidine - 5’ - phosphate)
นํ้ าตาลไรโบส จะ เรี ยกนิวคลี โอไทด รวมกันว า ไทมิ ดีนโมโนฟอสเฟต (thimidine monophosphate หรือ thymidine - 5’ - phosphate) สวนยูราซิล (uracil) ซ่ึงพบในอารเอ็นเอ จะเปน ยูริดีน 5’โมโนฟอสเฟต (uridine- 5’ -phosphate)หรือ ยูริดีนโมโนฟอสเฟต (uridine monophosphate)
ภาพที่ 1.58 เบสพวิรีนและไพริมิดีน
โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
79
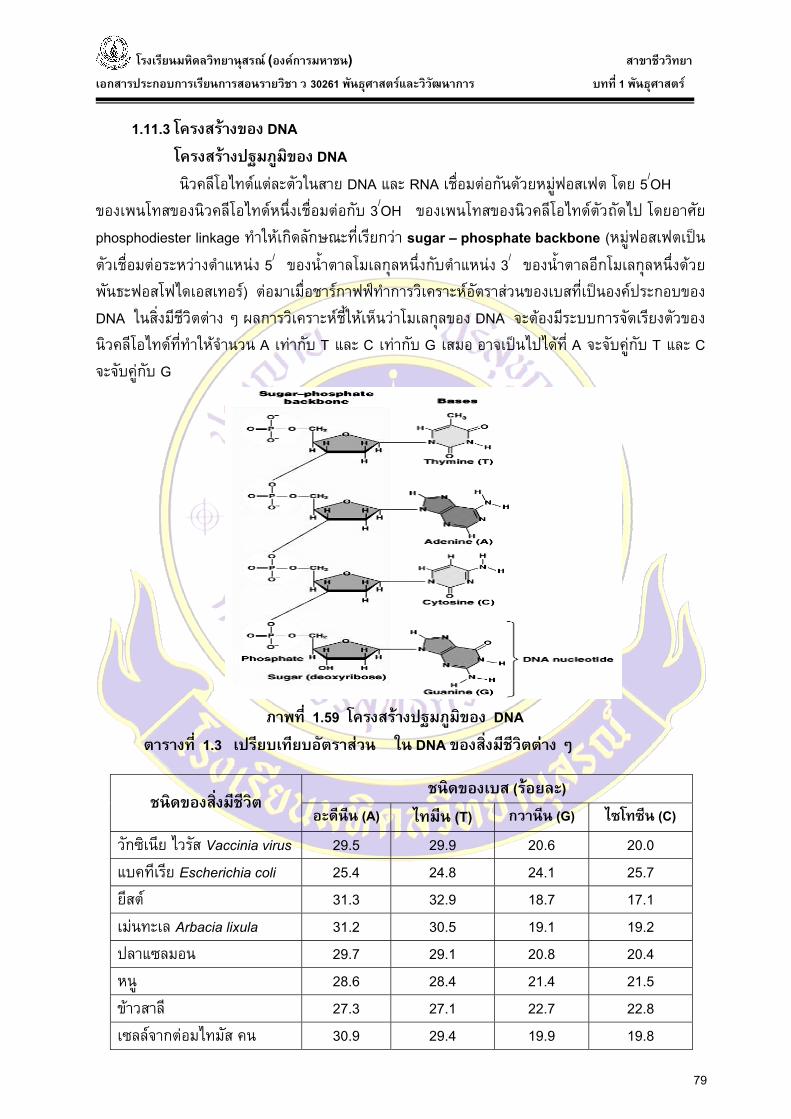
1.11.3 โครงสรางของ DNA โครงสรางปฐมภูมิของ DNA
นิวคลีโอไทดแตละตัวในสาย DNA และ RNA เชื่อมตอกันดวยหมูฟอสเฟต โดย 5/OH ของเพนโทสของนิวคลีโอไทดหน่ึงเชื่อมตอกับ 3/OH ของเพนโทสของนิวคลีโอไทดตัวถัดไป โดยอาศัย phosphodiester linkage ทําใหเกิดลักษณะที่เรียกวา sugar – phosphate backbone (หมูฟอสเฟตเปนตัวเชื่อมตอระหวางตําแหนง 5/ ของน้ําตาลโมเลกุลหน่ึงกับตําแหนง 3/ ของน้ําตาลอีกโมเลกุลหน่ึงดวยพันธะฟอสโฟไดเอสเทอร) ตอมาเมื่อชารกาฟฟทําการวิเคราะหอัตราสวนของเบสที่เปนองคประกอบของ DNA ในสิ่งมีชีวิตตาง ๆ ผลการวิเคราะหชี้ใหเห็นวาโมเลกุลของ DNA จะตองมีระบบการจัดเรียงตัวของ นิวคลีโอไทดที่ทําใหจํานวน A เทากับ T และ C เทากับ G เสมอ อาจเปนไปไดที่ A จะจับคูกับ T และ C จะจับคูกับ G
ภาพที่ 1.59 โครงสรางปฐมภูมิของ DNA
ตารางที่ 1.3 เปรียบเทียบอัตราสวน ใน DNA ของสิ่งมีชวีิตตาง ๆ
ชนิดของสิ่งมีชีวิต ชนิดของเบส (รอยละ)
อะดีนีน (A) ไทมีน (T) กวานีน (G) ไซโทซีน (C)
วักซิเนีย ไวรสั Vaccinia virus 29.5 29.9 20.6 20.0 แบคทีเรยี Escherichia coli 25.4 24.8 24.1 25.7 ยีสต 31.3 32.9 18.7 17.1 เมนทะเล Arbacia lixula 31.2 30.5 19.1 19.2 ปลาแซลมอน 29.7 29.1 20.8 20.4 หนู 28.6 28.4 21.4 21.5 ขาวสาล ี 27.3 27.1 22.7 22.8 เซลลจากตอมไทมัส คน 30.9 29.4 19.9 19.8

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
80
ในชวงที่นักฟสิกสไดนําเทคนิค X-ray diffraction มาใชในการหาโครงสรางของสารเคมีตาง ๆ น้ันไดมีนักฟสิกสชาวอังกฤษชือ่ วิลคินส (M.H.F. Wilkins) และแฟรงกิน (Rosallin Franklin) ไดลองหาโครงสรางของ DNA ดวยเทคนิคนี้ และสามารถแปรผลไดวา โครงสรางของ DNA มีลักษณะเปนเสนยาว มีลักษณะซ้ําๆ กันไมสมํ่าเสมอ ไมขึ้นกับองคประกอบและลําดับของเบส มีลักษณะเปนเกลียวคูและประกอบดวยสายโพลีนิวคลีโอไทดมากกวาหนึ่งสาย ที่มีเบสหันเขาขางในและสายที่เชื่อมระหวางหมูฟอสเฟตและน้ําตาลอยูดานนอก
ภาพที่ 1.60 เครื่องยิงรังสีเอกซที่ใชทํา x-ray diffraction รายงาน การคนพบ DNA X-ray diffraction
ก.
ข.

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
81
ความรูเสริม : การศึกษาโครงสรางของ DNA โดยเทคนิค X-ray diffraction ทําไดโดยการทําให DNA อยูในสภาพผลึกกึ่งเหลว แลวนํามาวางไวระหวางเครื่องยิงรังสีเอกซ และฉากหลังซ่ึงเปนแผนฟลม จากนั้นยิงรังสีผานผลึก DNA อนุภาคที่เขาไปชน DNA จะเกิดการเบี่ยงตัวออกและวิ่งไปตกกระทบบนแผนฟลม ซ่ึงจะสามารถอานผลไดตอไป การอานผลจะเปรียบเทียบกับโพลีเมอรที่มีลักษณะตาง ๆ เชน เปนเสนเกลียวแบบตาง ๆ
แบบจําลอง DNA x-ray
ผลท่ีไดจากการทํา x-ray diffraction ของโพลีเมอรรูปแบบตาง
แสดงการตกกระทบบนแผนฟลม
ผลท่ีไดจากการทํา DNA x-

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
82
จากการศึกษาของพอลิ่ง (Linus Pauling) พบวามีพันธะไฮโดรเจนเกิดขึ้นระหวางเบส ของสายโพลีนิวคลีโอไทดสองสาย ทําใหสายโพลีนิวคลีโอไทดสองสายนี้จับกันได โดย G ทําพันธะ 3 กับ C และ A ทําพันธะคูกับ T อยางไรก็ตามพันธะนี้ถูกทําลายไดดวยความรอนปานกลาง
โครงสรางทตุิยภูมิของ DNA จากขอมูลที่นาสนใจ 4 ประการอันไดแก
1. DNA เปนโมเลกุลที่มีสายยาวประกอบดวยนิวคลีโอไทด 4 ชนิดเชื่อมกันดวยหมูพันธะระหวางหมูฟอสเฟตกับนํ้าตาล
2. เบส A มีปริมาณเทากับ T และเบส G มีปริมาณเทากับ C 3. โครงสรางของโมเลกุล DNA เปนเกลียว ประกอบดวยสาย
โพลีนิวคลีโอไทดมากกวา 1 สาย มีเบสหันเขาดานในและสายเชื่อมระหวางหมูฟอสเฟตและน้ําตาลอยูขางนอก
4. พันธะระหวางคูเบสถูกทําลายไดดวยความรอนปานกลาง พันธะระหวางเบส A กับ T เปนพันธะคู และ C กับ G เปนพันธะสาม
ขอมูลทั้ง 4 ประการนี้ทําใหวัตสันและคริกสรางแบบจําลองโครงสรางของ DNA ขึ้น โดยใหสายโพลีนิวคลีโอไทด 2 สายเรียงตัวสลับทิศทางกัน โดยทําใหนิวคลีโอไทดที่ปลายขาง 3/ ของสายหนึ่งจับกับนิวคลีโอไทดที่ปลายขาง 5/ ของอีกสายหนึ่ง โดยพันธะระหวางคูเบส ทําใหเกิดเปนลักษณะเรียงตัวขนานกลับทิศทาง (antiparallel) มีนํ้าตาลและหมูฟอสเฟตอยูรอบนอกของโมเลกุล สายโพลีนิวคลีโอไทดที่เกิดขึ้นนี้มีลักษณะเปนเกลียวคูเวียนขวา (double helix) หรือหมุนตามเข็มนาฬิกา
โครงสราง DNA ที่เกิดขึ้นมีลักษณะคลายบันไดเวียน โดยมีนํ้าตาล ดีออกซีไรโบสกับหมูฟอสเฟตเปนราวบันได และบันไดแตละขั้นคือเบสหนึ่งคู ในการเวียนแตละรอบเกลียวจะมีจํานวนคูเบสเทากัน โดยเกลียวจะเวียนหมุนครบรอบยาว 3.4 นาโนเมตร (nm) ทุกๆ 10 คูเบส ทําใหเบสแตละคูอยูหางกัน 0.34 nm เสนผาศูนยกลางของเกลียวคูมีคาเทากับ 2 nm การพันเกลียวทําใหเกิดรอง 2 ขนาดเรียกวา รองใหญ (major groove) และ รองเล็ก (minor groove)
ภาพที่ 1.61 การเรียงตวัแบบ antiparallel ของ DNA
โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
83
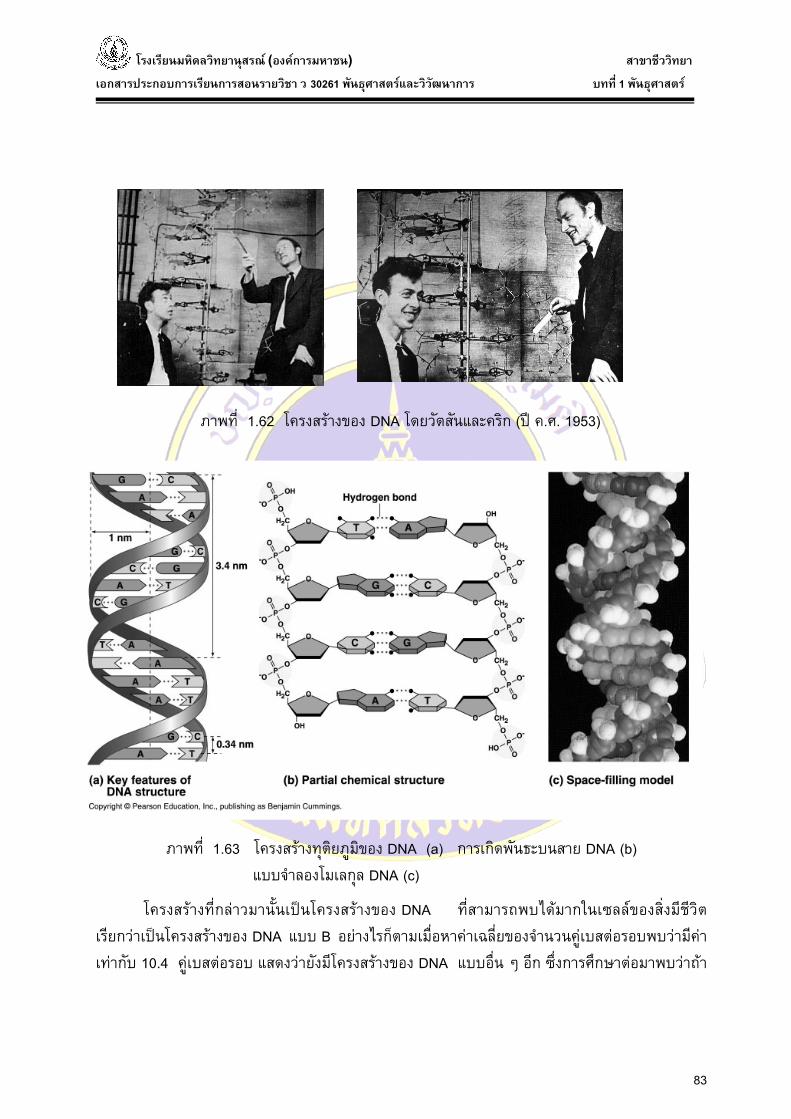
ภาพที่ 1.62 โครงสรางของ DNA โดยวตัสันและคริก (ป ค.ศ. 1953)
ภาพที่ 1.63 โครงสรางทุติยภูมิของ DNA (a) การเกิดพันธะบนสาย DNA (b)
แบบจําลองโมเลกุล DNA (c)
โครงสรางที่กลาวมานั้นเปนโครงสรางของ DNA ที่สามารถพบไดมากในเซลลของสิ่งมีชีวิต เรียกวาเปนโครงสรางของ DNA แบบ B อยางไรก็ตามเมื่อหาคาเฉลี่ยของจํานวนคูเบสตอรอบพบวามีคาเทากับ 10.4 คูเบสตอรอบ แสดงวายังมีโครงสรางของ DNA แบบอ่ืน ๆ อีก ซ่ึงการศึกษาตอมาพบวาถา

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
84
ภาพที่ 1.64 โครงสรางของ DNA แบบตาง ๆ
DNA สูญเสียนํ้าบางสวนไปจะทําใหโครงสรางเกลียวคูเปลี่ยนแปลงไปมี 11-12 คูเบสตอรอบเกลียว (แทนที่จะเปน 10 คูเบส) มีเสนผาศูนยกลางเกลียว 2.3 nm โครงสรางลักษณะนี้ถูกเรียกวา โครงสราง
ของ DNA แบบ A นอกจากนี้ยังมีโครงสรางของ DNA แบบที่เกลียวเวียนซายดวย ซ่ึงคนพบโดย Alexander Rich ลักษณะเดนของ DNA แบบน้ีคือ มีเบสพิวรีนสลับกับไพริมิดีน ทําใหมีลักษณะคลายสลับฟนปลา (zigzag) 1 รอบเกลียวจะประกอบดวย 12 คูเบสมีเสนผาศูนยกลางเกลียว 1.8 nm เรียก DNA ลักษณะนี้วา Z-DNA
ดีเอ็นเอกับอารเอ็นเอลวนแตเปนกรดนวิคลีอิก ดังน้ันกรดนิวคลีอิกจะประกอบดวย เบส 5 ชนิด โดยที่
ไมไดแยกวาเปน ดีเอ็นเอหรืออารเอ็นเอ อยางไรก็ตามดีเอ็นเอ และ อารเอ็นเอก็มีความแตกตางกัน เชน 1. ดีเอ็นเอตางจากอารเอ็นเอ คือ นํ้าตาลในดีเอ็นเอเปนดีออกซีไรโบส และในอารเอ็นเอเปน ไรโบส 2. เบสในอารเอ็นเอ เปน ยูราซิล (U) แทนที่ไทมีน (T) ทีอยูบนดีเอ็นเอ 3. อารเอ็นเอมีโพลีนิวคลีโอไทด 1 เสน สวนดีเอ็นเอมีโพลีนิวคลีโอไทด 2 เสน พันเปนเกลียวคูเวียน
ขวา 4. ดีเอ็นเอพบในนิวเคลียสของไมโทคอนเดรียและเซลล สวนอารเอ็นเอพบในนิวเคลียสและ
ไซโทพลาซึม
คุณสมบัติของ DNA 1. การเสียสภาพ (denaturation) และการคืนสภาพ (renaturation) : พันธะระหวางเบสของดีเอ็นเอ
สามารถถูกทําลายไดดวยความรอนและสารเคมี การเสียสภาพเนื่องจากการใหความรอนน้ันจะทําใหพันธะไฮโดรเจนสลาย แตเม่ืออุณหภูมิลดลงเบสอาจกลับมาจับกันดวยพันธะไฮโดรเจนได อีกจากคุณสมบัตขิอน้ีสามารถประยุกตใชโดยทําใหเกิดเทคนิค hybridization เพ่ือตรวจสอบความคลายคลึงของดีเอ็นเอจากสองแหลง

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
85
2. การดูดกลืนแสง : สารพันธุกรรมสามารถดูดกลืนแสงอัลตราไวโอเลตไดที่ความยาวชวงแสง 260 นาโนเมตร
3. ปฏิกิริยาตอกรด-เบส : พันธะไฮโดรเจนของคูเบสจะเสถียร (คงตัว) ในชวง pH 4.0 ถึง 11.0 ถามากหรือนอยกวานี้ดีเอ็นเอจะเสียสภาพ
4. sedimention : ดีเอ็นเอที่มีองคประกอบและขนาดแตกตางกันจะมีความหนาแนนแตกตางกัน ทําใหการตกตะกอนแตกตางกนัดวย
ดีเอ็นเอประกอบดวยนวิคลโีอไทด 4 ชนิดคือ นิวคลโีอไทดที่มีเบสเปน ATCG ความแตกตางของโมเลกุล DNA อยูที่การจัดเรียงลําดับเบสเชน นิวคลีโอไทดสองโมเลกุลตอกันจะมีลําดับเบสที่แตกตางถึง 16 แบบคือ
A
A AA T
A TA C
A CA G
A GA T AT T TT T CT T GT C AC C TC C CC C GC G AG G TG G CG G GG
ตารางที่ 1.65 แสดงชนิดของการเรียงลําดับของเบสทั้ง 4 ชนิด
จํานวนชนิดของการเรียงลําดับของเบสหาไดจากสูตร 4n เม่ือ n คอื จํานวนนิวคลีโอไทดเรียงตอกันเชน
2 นิวคลีโอไทดเรียงตอกัน 42 = 16 แบบ 3 นิวคลีโอไทดเรียงตอกัน 43 = 64 แบบ
สายดีเอ็นเอคนมีความยาว 3 X 109 คูเบส จะมีความแตกตางของดีเอ็นเอไดมากมายถึง 4300,000,000 แบบ ซ่ึงเปนจํานวนที่มากกวาประชากรบนโลกนี้ดังน้ันโอกาสที่จะมีคนสองคนที่มีพันธุกรรมเหมือนกันนั้นนอยมาก

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
86
1.3 คุณสมบตัิของสารพนัธุกรรม
การพิสูจนวาดีเอ็นเอเปนสารพันธุกรรมตองมีสมบัติสําคัญ คือ 1. ตองสามารถเพิ่มจํานวนตัวเองไดโดยมีลักษณะเหมือนเดิมเพ่ือสามารถถายทอดลักษณะทาง
พันธุกรรมจากรุนพอแมไปยังรุนลูกได 2. สามารถควบคุมใหเซลลสังเคราะหสารตางๆเพื่อแสดงลักษณะทางพันธุกรรมใหปรากฎ 3. ตองสามารถเปลี่ยนแปลงไดบาง ซ่ึงการเปลี่ยนแปลงที่เกิดขึ้นอาจกอใหเกิดสิ่งมีชีวิตสปชีส
ใหมๆ ขึ้น ดีเอ็นเอสามารถเพิ่มจํานวนตัวเองไดโดยกระบวนการจําลองตัวหรือการลอกแบบ (DNA replication) และสามารถสรางโปรตีนได โดยดีเอ็นเอผานกระบวนการถอดรหัส (transcription) จะได พรีอารเอ็นเอ (preRNA) หรือเอ็มอารเอ็นเอที่ยังไมสามารถทํางานได จะตองผานกระบวนการตัดตออารเอ็นเอ (RNA processing) และการเติม (modification) ไดแก การตัดอินทรอน (intron คือ อารเอ็นเอที่ไมทํางาน) ออกเหลือแตเอกซอน (exon คืออารเอ็นเอที่ทํางานได) แลวตอกับอีกเอกซอนหน่ึงกลายเปน เอ็มอารเอ็นเอที่พรอมจะทํางาน (mature mRNA) ซ่ึงกระบวนการนี้เกิดขึ้นในนิวเคลียส เอ็มอารเอ็นเอที่ตัดตอแลวจะถูกสงออกไปยังไซโทพลาสซึมที่ไรโบโซม (ribosome) จากนั้นเกิดกระบวนการแปลรหัส (translation) ซ่ึงเปนกระบวนการสงผานสารพันธุกรรม กอใหเกิดการแสดงออกของยีนในรูปการสรางโปรตีน) โดยทีอารเอ็นเอ (tRNA) จะนํากรดอะมิโนมาตอกันเปนโปรตีน เรียกวา โพลีเปปไทด (polypeptide) สรางโปรตีนแลวเปลี่ยนรูปไปเปนเอนไซม (enzyme) หรือโปรตีนชนิดอ่ืน ๆ เพ่ือใชในกิจกรรมของเซลลตอไป ภาพที่ 1.66 กระบวนการ DNA expression

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
87
1.3.1 การจําลองโมเลกลุของ DNA (DNA replication) การสังเคราะหดีเอ็นเอหรือการจําลองตัวของดีเอ็นเอ คือการสังเคราะหดีเอ็นเอใหมใหเหมือน ดีเอ็นเอคูเดิมที่เปนแมแบบ (template)
ภาพที่ 1.67 การจําลองตัวของ DNA
การจําลองดีเอ็นเอ (replication) มี 3 แบบจําลอง (model) คือ 1. วิธีอนุรักษ (conservative model) คือ การที่ดีเอ็นเอสรางสายใหมทั้งโมเลกุลใหเหมือนสาย
เกา โดยที่อนุรักษสายเกาเอาไวทั้งโมเลกุล (โพลีนิวคลีโอไทด 2 สายเกาจับกนัเองและโพลนิีวคลีโอไทดสองสายใหมจะจับกันเอง)
2. วิธีก่ึงอนุรักษ (semiconservative model) คือ การที่ดีเอ็นเอสายเกาแยกเปน 2 สาย (strand) แลวสราง ดีเอ็นเอสายใหมอีก 1 เสน มารวมกันเปน 1 โมเลกุลดีเอ็นเอ เรียกเปนการอนุรักษโมเลกุลเดิม (conserve) ไวครึ่งหน่ึง
3. ดิสเพอรซีฟ โมเดล (dispersive model) คือ การที่โมเลกุลดีเอ็นเอสายเกาแตกหักออกทุกครึ่งรอบแลวสรางโมเลกุลดีเอ็นเอสายใหมเติมเขาไป
ภาพที่ 1.68 รูปแบบการจาํลองตัวของ DNA ที่คาดวานาจะเปนไปได

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
88
การทดลองเพื่อพิสูจนวาแบบจําลองแบบใดถูกตองน้ัน นักชีวเคมี 2 คน คือ เมเซลสัน (M. Meselson) และสทาฮล (F.W. Stahl) ศึกษาโดยการเพาะเลี้ยงแบคทีเรียในอาหารที่มีกัมมันตภาพรังสีของไนโตรเจน (15N) และเพาะเลี้ยงแบคทีเรียรุนตอมาในอาหารที่มีไนโตรเจนธรรมดา (14N) เม่ือทําการตรวจสอบ พบวาปริมาณสารกัมมันตภาพรังสีของไนโตรเจนในรุนลูกพบวาลดลงไปครึ่งหน่ึง และเหลือ 1 ใน 4 เทานั้นในแบคทีเรียรุนลูก ดังนั้นจึงสรุปการทดลองไดวาการจําลองตัวเองของดีเอ็นเอเปนแบบ เซมิคอนเซอรเวทีฟ
รูปแสดงการทดลองเพื่อพิสูจนวาดเีอ็นเอมีการจําลองตวัรูปแบบใด
ภาพที่ 1.69 การทดลองเพื่อพิสูจนวา DNA มีการจําลองตัวรูปแบบใด
ในแบคทีเรียซ่ึงมี 1 โครโมโซม (chromosome) จะเร่ิมตนจําลองตัวเองที่จุดเริ่มตนของ การเรพพลิเคชั่น (origin of replication หรือ Ori) เม่ือเกิดการเรพพลิเคชั่น จุดเริ่มตน (origin) จะแยกออกจากกันเห็นเปนรอยแยก เรียกวา เรพพลิเคชั่นฟอรค (replication fork) ซ่ึงเมื่อเสร็จสิ้นการจําลอง ดีเอ็นเอ 1 โมเลกุลจะแยกเปน 2 โมเลกุล สวนในยูคาริโอท (eukaryote) มีการเกิดการเรพพลิเคชั่นเปนแบบ ไบไดเร็คชั่นนอลเรพพลิเคชั่น (bidirectional replication) และมีจุดเริ่มตนสรางดีเอ็นเอหลายจุด สรางมาชนกันแลวตอกันเปนโมเลกุลดีเอ็นเอที่ยาวขึ้น
ภาพที่ 1.70 การจําลอง DNA ของยูคาริโอต

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
89
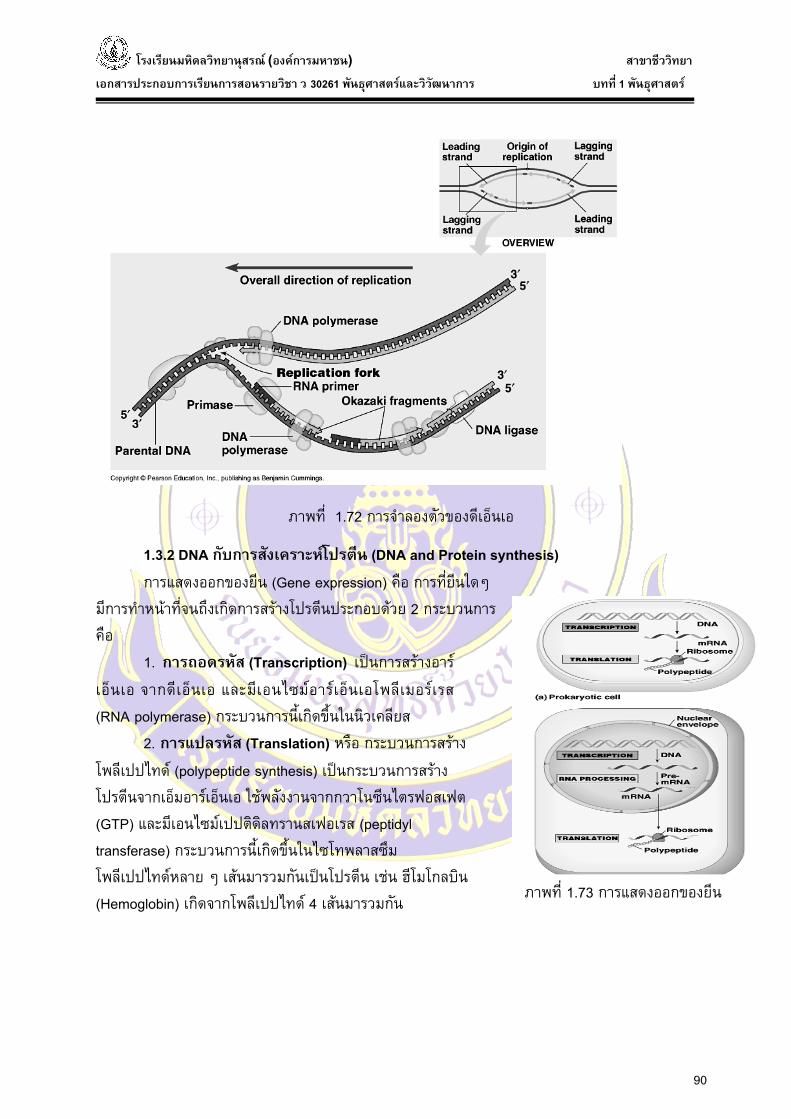
ดีเอ็นเอและเอ็มอารเอ็นเอถูกสรางจากปลาย 5’ ไปยัง 3’ เสนที่สามารถสรางติดตอกันไปเรื่อยๆ โดยไมหยุดเรียกวาเสนนํา (Leading strand) สวนดีเอ็นเอเสนที่สรางแบบไมตอเน่ือง ซ่ึงดีเอ็นเอเสนสัน้ๆ น้ี เรียกวา Okazaki fragment ซ่ึงจะถูกเชื่อมใหตอกันโดยเอนไซมดีเอ็นเอไลเกส DNA ligase ใหเกิดเปนสายโพลนิีวคลีโอไทดที่สมบูรณ เรยีกเสนดีเอ็นเอเสนน้ีวา เสนตาม (Lagging strand) โดยเอนไซมที่ใชในการสรางเสนตาม มีดังน้ี
1. เฮลิเคส (helicase หรือ helix-destabilizing protein) เปนเอนไซมที่สลายพันธะไฮโดรเจน ทําใหโมเลกุล ดีเอ็นเอมีการคลายเกลียวคูออกจากกันเปน สายเดี่ยว 2 สายโดยอาศัยพลังงานจากการสลาย ATP 2. โปรตีนเกาะสายเดี่ยว (single strand DNA binding protein : SSB หรือ DBP) จะจับกับดีเอ็นเอสายเดี่ยวที่แยกออกจากกัน เปนตัวปองกันไมใหดีเอ็นเอสายเดี่ยวกลับไปจับกันอีก จะไดไมเกิดเปนโมเลกุลดีเอ็นเอเกลียวคูเหมือนเดิม และปองกัน DNA สายเดี่ยวไมใหถูกยอยโดยเอนไซม nuclease 3. DNA gyrase หรือ topoisomerase ทําหนาที่ คลายปมเหนือจุดแยก (replication fork) โดยการตัด DNA สายใดสายหนึ่งออก เพ่ือใหคลายเกลียวไดแลวจึงตอกลับใหม 4. ไพรเมส (primase) ทําหนาที่สรางอารเอ็นเอ เริ่มตน (RNA primer) (primosome : ไพรเมอรและเอนไซมที่รวมกันอยูที่จุดแยก) 5. ดีเอ็นเอโพลเีมอรเรส I (DNA polymerase I) ทําหนาที่ในการตอสายโพลีนิวคลีโอไทดใหยาวขึน้และตรวจสอบลําดับเบสที่ผิดพลาดและกําจัดลําดับเบสที่ผิดพลาดออกไป รวมถึงการกําจัด RNA primer
6. ดีเอ็นเอโพลีเมอรเรส III (DNA polymerase III) เปนเอนไซมหลักในการจําลองตัวของ DNA 7. ดีเอ็นเอไลเกส (DNA ligase) ทําหนาที่เชื่อมดีเอ็นเอเสนสั้น ๆ เขาดวยกัน (โดยการสรางพันธะฟอสโฟไดเอสเทอรเชื่อมจุดที่ขาด)
ภาพที่ 1.71 แสดงการเกิดเสนนําและเสนตาม
โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
90
1.3.2 DNA กับการสังเคราะหโปรตีน (DNA and Protein synthesis) การแสดงออกของยีน (Gene expression) คือ การที่ยีนใดๆ
มีการทําหนาที่จนถึงเกิดการสรางโปรตนีประกอบดวย 2 กระบวนการ คือ
1. การถอดรหัส (Transcription) เปนการสรางอารเอ็นเอ จากดีเอ็นเอ และมีเอนไซมอารเอ็นเอโพลีเมอรเรส (RNA polymerase) กระบวนการนี้เกิดขึ้นในนิวเคลียส
2. การแปลรหัส (Translation) หรือ กระบวนการสรางโพลีเปปไทด (polypeptide synthesis) เปนกระบวนการสรางโปรตีนจากเอ็มอารเอ็นเอ ใชพลังงานจากกวาโนซีนไตรฟอสเฟต (GTP) และมีเอนไซมเปปตดิิลทรานสเฟอเรส (peptidyl transferase) กระบวนการนี้เกิดขึ้นในไซโทพลาสซึม โพลีเปปไทดหลาย ๆ เสนมารวมกันเปนโปรตีน เชน ฮีโมโกลบิน (Hemoglobin) เกิดจากโพลีเปปไทด 4 เสนมารวมกัน
ภาพที่ 1.72 การจําลองตัวของดีเอ็นเอ
ภาพที่ 1.73 การแสดงออกของยีน

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
91
ภาพที่ 1.74 กระบวนการ transcription และ translation
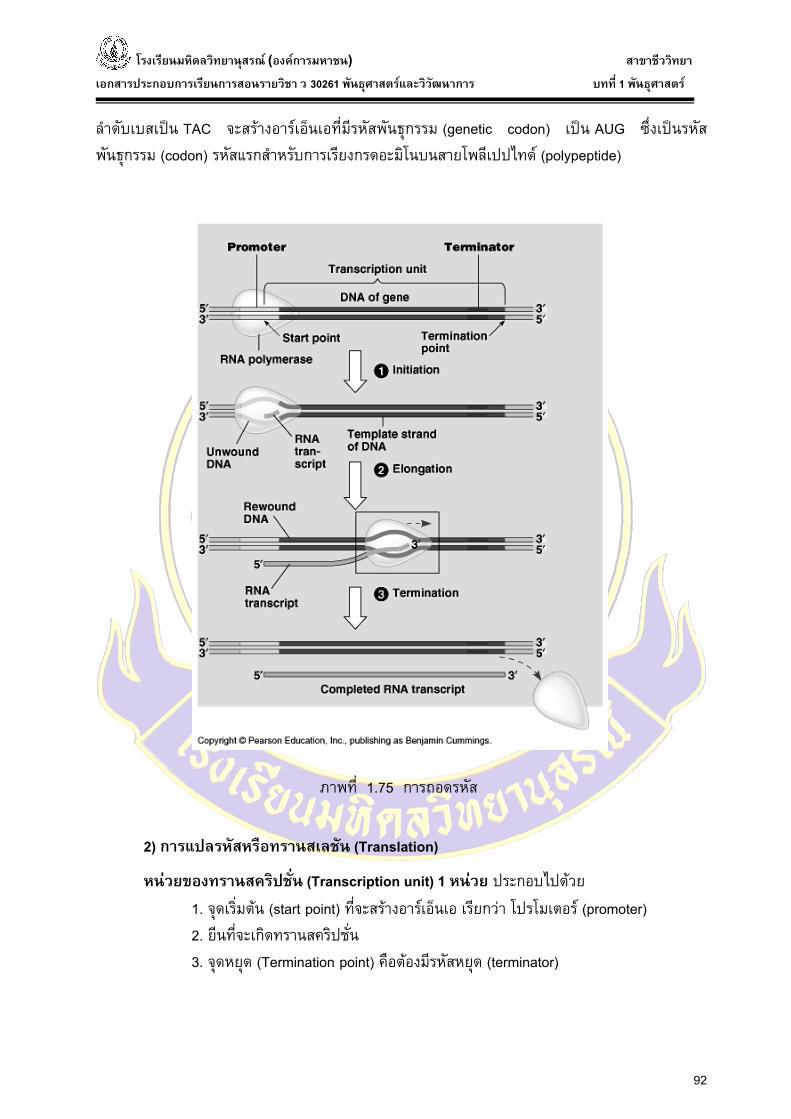
1) การถอดรหัส หรือการทรานสคริบชัน (Transcription)
ในการสรางอารเอ็นเอจะมีเอนไซมที่ทําหนาที่สรางอารเอ็นเอ คือ เอนไซมอารเอ็นเอโพลีเมอรเรส ซ่ึงเปนเอนไซมที่มีขนาดใหญ โดยสวนหัวของมันจะทําหนาที่ทําลายพันธะไฮโดรเจนระหวางเบสของ ดีเอ็นเอ ทําใหดีเอ็นเอแยกกันเปนสายเดี่ยว แลวมีการสรางโพลีนิวคลีโอไทด โดยนําเอา นิวคลีโอไทด ไตรฟอสเฟต (NTP = nucleotide triphosphate) 4 ชนิดคือ UTP ATP CTP และ GTP เขามาตอกันเปน สายอารเอ็นเอ ตําแหนงบนดีเอ็นเอที่เอนไซม อารเอ็นเอโพลีเมอรเรส เขาเกาะเรียกวา โปรโมเตอร (promoter) ในการสรางอารเอ็นเอแตละรอบนั้นดีเอ็นเอ 1 โมเลกุลจะสรางอารเอ็นเอ ได 1 โมเลกุลมี โพลีนิวคลีโอไทดเพียงเสนเดียว ดีเอ็นเอใหม 2 โมเลกุลบนดีเอ็นเอจะมีบริเวณที่จําเพาะเจาะจงสําหรับการสรางอารเอ็นเอ โดยบนสายโคดดิ้ง (Non-coding strand) ของดีเอ็นเอมีบริเวณที่เรียกวา พริบนาว บอกซ (Pribnow box) ซ่ึงเปนบริเวณที่อารเอ็นเอโพลีเมอรเรสเกาะบนดีเอ็นเออยางจําเพาะ ดีเอ็นเอเสนที่มี
โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
92
ลําดับเบสเปน TAC จะสรางอารเอ็นเอที่มีรหัสพันธุกรรม (genetic codon) เปน AUG ซ่ึงเปนรหัสพันธุกรรม (codon) รหัสแรกสําหรับการเรียงกรดอะมิโนบนสายโพลีเปปไทด (polypeptide)
2) การแปลรหัสหรือทรานสเลชัน (Translation)
หนวยของทรานสคริปชัน่ (Transcription unit) 1 หนวย ประกอบไปดวย 1. จุดเริ่มตน (start point) ที่จะสรางอารเอ็นเอ เรียกวา โปรโมเตอร (promoter) 2. ยีนที่จะเกดิทรานสคริปชั่น 3. จุดหยุด (Termination point) คือตองมีรหัสหยุด (terminator)
ภาพที่ 1.75 การถอดรหัส

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
93
เริ่มตนกระบวนการทรานสคริปชั่น (initiation of transcription) ตองมีอารเอ็นเอโพลีเมอรเรส เขาเกาะกอน แลวมีกระบวนการยืดยาว (elongation) หมายถึงการตอกันของนวิคลีโอไทดไตรฟอสเฟต (NTP = nucleotide triphosphate) 4 ชนิดคือ UTP ATP CTP และ GTP เขามาตอกันเปนอารเอ็นเอไปเรื่อย ๆ จนถึงรหัสหยุด (terminator) คือการสิ้นสุดกระบวนการถอดรหัส (termination) ดังน้ันในแตละครั้งโมเลกุลอารเอ็นเอที่ถูกสรางจะมีเสนเดียวจากการเริ่มตนของ 1 โมเลกุลของดีเอ็นเอ
ภาพที่ 1.76 กระบวนการถอดรหัส ผลผลติของอารเอ็นเอ มีดังน้ี
1. อาร อารเอ็นเอ หรือไรโบโซมอล อารเอ็นเอ (rRNA = ribosomal RNA) เม่ือรวมกับโปรตนีจะกลายเปนไรโบโซม (ribosome) ตําแหนงของอารอารเอ็นเออยูที่ไรโบโซม หนาที่เปนสวนประกอบของ ไรโบโซมซึ่งจะทําหนาที่สรางโปรตีน 2. ที อารเอ็นเอ หรือ ทรานสเฟอร อารเอ็นเอ (tRNA = transfer RNA) โครงสรางคลายใบโคลฟเวอร (clover leaf) มี 3 ลูป (loop) โดยปลาย 3’ เปนที่เกาะของกรดอะมิโน สวนปลายลูปหน่ึงเปนตําแหนงของ ทีอารเอ็นเอ (ที่นํากรดอะมิโน) เกาะกับ เอ็มอารเอ็นเอ เพ่ือนํากรดอะมิโนมาตอกันเปนโพลีเปปไทด
3. เอ็ม อารเอ็นเอ หรือเมสเซ็นเจอร อารเอ็นเอ (mRNA = messenger RNA) มี เบส 3 ตัวแรกคือ AUG อยูที่เอ็ม อารเอ็นเอ โดยที่เอ็มอารเอ็นเอถูกสรางในนิวเคลียส และถูกสงตอเพ่ือไปใชงานยังไซโทพลาสซึม เอ็มอารเอ็นเอทําหนาที่นําขอมูลจากดีเอ็นเอไปยังไรโบโซมเพื่อระบุชนิดของกรดอะมิโนในสายโพลีเปปไทด
ภาพที่ 1.77 tRNA
ภาพที่ 1.78 mRNA

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
94
ไรโบโซม (Ribosome) ในยูคาริโอท (eukaryote) ประกอบดวย 2 หนวยยอย (subunit) คือ ขนาดเล็ก 40S และขนาดใหญ 60S เชือ่มรวมกันมีขนาดของไรโบโซม เปน 80S สวนใน โปรคาริโอท (prokaryote) น้ันก็ประกอบดวย 2 หนวยยอย เชนเดียวกันคือขนาดเล็ก 30S และขนาดใหญ 50S เชือ่รวมกันมีขนาดของไรโบโซมเปน 7 0 S ( S = S v e d b e r g u n i t ) ที่ไรโบโซมมีตําแหนงตาง ๆ ดังน้ี
1. เปปติดิลไซต หรือพีไซต (peptidyl site = P site) เปนที่เกาะของทีอารเอ็นเอที่นํากรดอะมิโนหลายๆ ตวั เราเรียกทีอารเอ็นเอ ชนิดนี้วา เปปติดิล - ทีอารเอ็นเอ (peptidyl - tRNA)
2. อะมิโนเอซิลไซต หรือเอไซต (Aminoacyl site = A site) เปนที่เกาะของทีอารเอ็นเอที่นํากรดอะมิโน 1 ตัว เรียกทีอารเอ็นเอชนิดนี้วาอะมิโนเอซิล - ทีอารเอ็นเอ (aminoacyl - tRNA)
3. รองเอ็ม อารเอ็นเอ (mRNA groove) เปนรองสําหรับให เอ็มอารเอ็นเอพาดผานและเกาะ เพราะภายในไรโบโซม มีตาํแหนงสําหรับ เอ็มอารเอ็นเอ (mRNA - binding site) ทําหนาที่เปนชองสําหรับใหเอ็มอารเอ็นเอสอดผาน
4. เอ็กซิทไซต หรืออีไซต (Exit site = E site) เปนที่ออกของทีอารเอ็นเอภายหลังหมดหนาที่นํากรดอะมิโนมาตอในสายโพลีเปปไทดเรียบรอยแลว
ในการสรางโปรตีนจะมีเอนไซมที่ใชในการสรางโปรตีนคือ เอนไซม เปปติดิลทรานสเฟอเรส (peptidyl transferase) โดยที่ เอ็มอารเอ็นเอ จะมีรหัสสรางกรดอะมิโนจํานวนมาก และมารวมกันกลายเปนสายโพลีเปปไทดในที่สุด
ระบบการเผาผลาญที่บกพรองมาแตกําเนิด (Inborn Error of Metabolism) เปนการคนพบของการรอด (A.E.Garrod, 1909) พบวาความบกพรองทางพันธุกรรมที่เปนมาตั้งแตกําเนิด มีสาเหตุจากยีนเกิดการกลายพันธุ (mutation single gene) มีผลทําใหเอนไซมผิดปกติ เกิดเปนความบกพรอง (defective enzyme) เชน เม่ือเอนไซมที่สรางกรดอะมิโนฟนิลอะลานีน( phenylalanine) ผิดปกติ จะทําใหเกิดเปนโรคฟนิลคีโตนูเรีย (Phenylketonuria) ซ่ึงในกระบวนการสรางมีเอนไซม 2 ชนิด โดยเอนไซมชนิดหนึ่งจะสรางสาร อัลแคปตอน (Alkapton) เม่ือเกิดความผิดปกติของเอนไซมชนิดนี้ ก็จะทําให สารอัลแคปตอน ไมถูกเปลี่ยนไปเปนผลผลิตทําให สารอัลแคปตอนถูกสะสมเพิ่มขึ้นเกิดเปนโรค อัลแคปโตนูเรีย (Alkaptonuria) คือ ปสสาวะเปนสีดํา
ภาพที่ 1.79 แบบจําลองของไรโบโซม

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
95
มีเอนไซมอีกชนิดหนึ่งจะสรางสารเมลานินสรางเม็ดสีที่ผิวหนัง (melanin) เม่ือเกิดความผิดปกติจะทําใหเกิดโรคอัลบินิซึม (Albinism) (โรคผิวเผือก) โดยสรุป จากความบกพรองทางพันธุกรรมทําให เกิดระบบการเผาผลาญที่บกพรองมาแตกําเนิด และมีผลใหให Garrod เปนผูคนพบความสัมพันธระหวางยีนและเอนไซม
บีดเดิ้ล และทาทั้ม (G. W. Beadle & E.L.Tatum, 1940) เขาทําใหสายพันธุนิวโรสปอรรา (Neurospora) เกิดกลายพันธุ โดยการ ใชรังสีเอ็กซ (X - ray) หลังจากยีนกลายพันธุไป เอนไซมที่เฉพาะเจาะจงถูกสรางผิดปกติ ทําใหไมสามารถสรางผลผลิตในขั้นตอนของมันได น่ันคือเกิดการเปลี่ยนแปลงทางพันธุกรรม ในระบบการสรางกรดอะมิโนอารจินีน (Biosynthesis pathway ของ arginine)
จาก 1 ยีนสามารถสราง 1 เอนไซมที่จําเพาะเจาะจง ตอมาไดมีการคนพบวาเอนไซมเปนโปรตนี จึงเปลี่ยนจาก 1 ยีนสราง 1 โปรตีน และตอมาในยคุที่ศึกษาถึงระดับโมเลกลุพบวาโปรตีนประกอบดวย โพลีเปปไทด จีงเปลี่ยนเปน 1 ยีนสราง 1 โพลีเปปไทด
ยีนมีหนวยยอยเปนซิสตรอน (cistron) โดย 1 ซิสตรอน สามารถสราง 1 โพลีเปปไทด ดังน้ัน ถา 1 ยีนเกิดกลายพันธุไป (one gene mutation) ทําใหเกิดความผิดปกตขิองเอนไซมขึ้นได
ดีเอ็นเอจะมีความยาวเทาใดก็ได และมีหลาย ๆ ยีนอยูบนดีเอ็นเอแตละยีนจะมีชิน้สวนของดีเอ็นเอ ซ่ึงประกอบดวย เบส, นํ้าตาล และกลุมฟอสเฟตโดยทีนํ้่าตาล และกลุมฟอสเฟตตอกันเปนแกนสันหลังของดีเอ็นเอ (sugar phosphate backbone)
รหัสพันธุกรรม ดีเอ็นเอเปนแมพิมพในการสังเคราะหเอ็มอารเอ็นเอ ดังนั้นขอมูลทางพันธุกรรมในดีเอ็นเอจะ
ถายทอดใหกับเอ็มอารเอ็นเอ การเรียงลําดับของนิวคลีโอไทดชนิดตางๆ ของเอ็มอารเอ็นเอจึงเปนตัวกําหนดการเรียงลําดับของกรดอะมิโนเพ่ือสังเคราะหโปรตีน
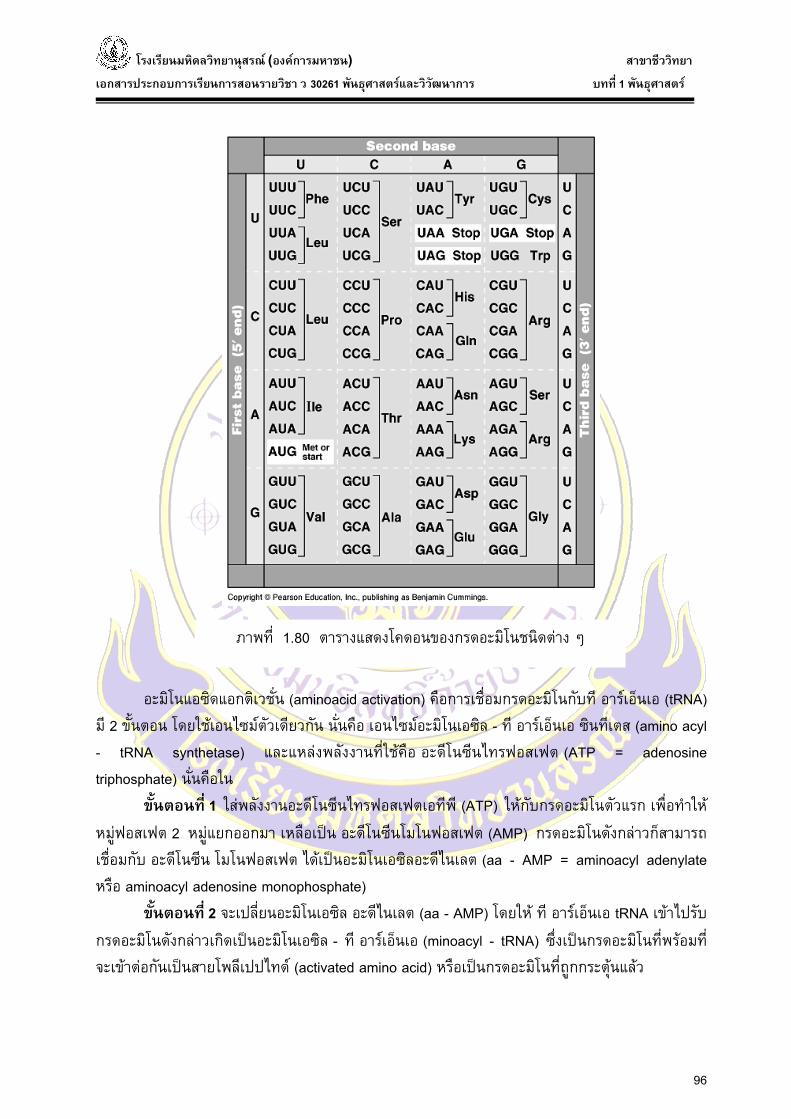
กรดอะมิโนที่ใชในการสังเคราะหโปรตีนมีประมาณ 20 ชนิด ดังน้ันการกําหนดรหัสพันธุกรรม (genetic code) ในสายเอ็มอารเอ็นเอนั้นจะประกอบดวยนิวคลีโอไทด 3 ตัว ตอ 1 โคดอน (codon) หรือ 1 รหัสพันธุกรรม โดย 1 โคดอนจะจําเพาะตอกรดอะมิโน 1 ตัว นิวคลีโอไทดบนดีเอ็นเอและอารเอ็นเอมีอยางละ 4 ตัว ดังน้ันรหัสพันธุกรรมที่เปนรหัสบนเอ็มอารเอ็นเอ มีทั้งหมดไดเปน 43=64 ตัวแตกรดอะมิโนทั้งหมดมี 20 ตัว ดังน้ันจะมีโคดอนที่ซํ้ากันและเปลี่ยนไป เชน เปลี่ยนไปเปนรหัสหยุด (stop codon) ที่มี 3 ตัว คือ UAA UAG และ UGA ในการซ้ําจะมี 2 แบบ คือ
1. แบบทีเ่บสตําแหนงที่ 3 (third base) เปนเบสอะไรก็ไดที่ไมมีการแยกกลุม ก็จะใหกรดอะมิโนชนิดเดียวกัน
2. แบบที่เบสตําแหนงที่ 3 มีการแยกกลุมของเบส น่ันคือ เบสในกลุมไพริมิดีน (pyrimidine) คือ U และ C และกลุม พิวรีน (purine) คือ A และ G ก็จะใหกรดอะมิโนคนละชนิดกัน จึงตองมีการแยกกลุมของเบส
โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
96
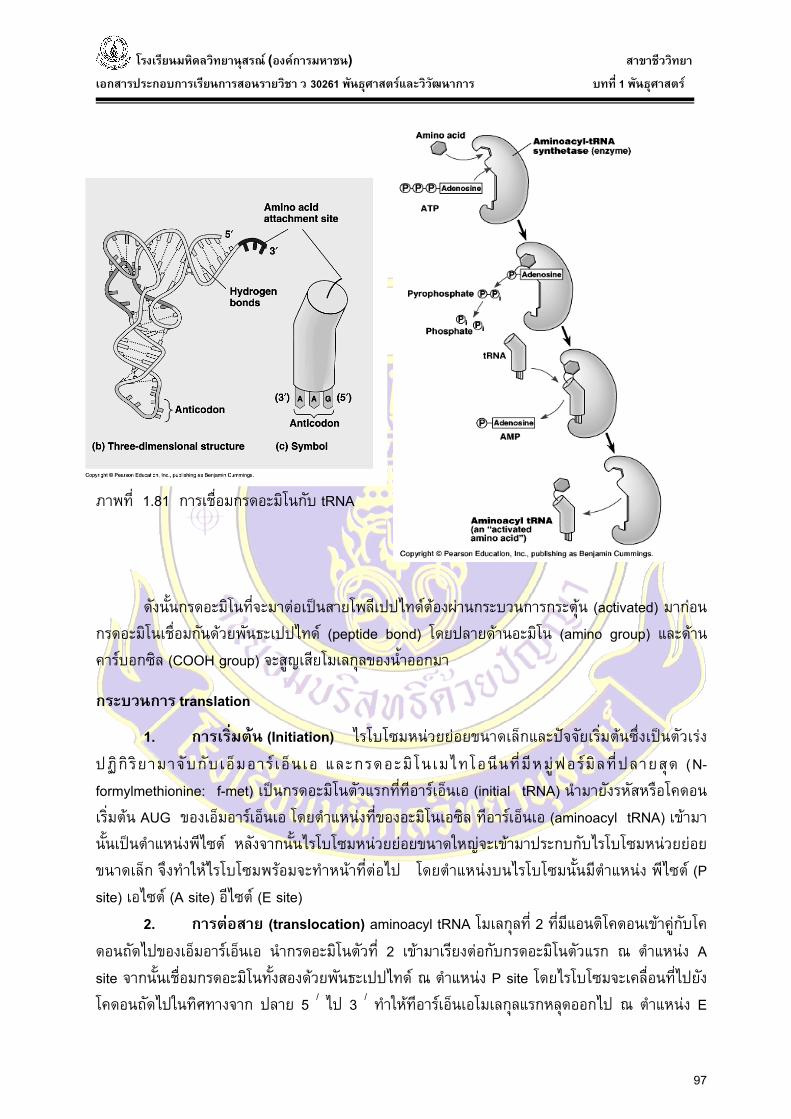
อะมิโนแอซิดแอกติเวชั่น (aminoacid activation) คือการเชื่อมกรดอะมิโนกับที อารเอ็นเอ (tRNA)
มี 2 ขั้นตอน โดยใชเอนไซมตัวเดียวกัน น่ันคือ เอนไซมอะมิโนเอซิล - ที อารเอ็นเอ ซินทีเตส (amino acyl - tRNA synthetase) และแหลงพลังงานที่ใชคือ อะดีโนซีนไทรฟอสเฟต (ATP = adenosine triphosphate) น่ันคือใน
ข้ันตอนที่ 1 ใสพลังงานอะดีโนซีนไทรฟอสเฟตเอทีพี (ATP) ใหกับกรดอะมิโนตัวแรก เพ่ือทําใหหมูฟอสเฟต 2 หมูแยกออกมา เหลือเปน อะดีโนซีนโมโนฟอสเฟต (AMP) กรดอะมิโนดังกลาวก็สามารถเชื่อมกับ อะดีโนซีน โมโนฟอสเฟต ไดเปนอะมิโนเอซิลอะดีไนเลต (aa - AMP = aminoacyl adenylate หรือ aminoacyl adenosine monophosphate)
ข้ันตอนที่ 2 จะเปลี่ยนอะมิโนเอซิล อะดีไนเลต (aa - AMP) โดยให ที อารเอ็นเอ tRNA เขาไปรับกรดอะมิโนดังกลาวเกิดเปนอะมิโนเอซิล - ที อารเอ็นเอ (minoacyl - tRNA) ซ่ึงเปนกรดอะมิโนที่พรอมที่จะเขาตอกันเปนสายโพลีเปปไทด (activated amino acid) หรือเปนกรดอะมิโนที่ถูกกระตุนแลว
ภาพที่ 1.80 ตารางแสดงโคดอนของกรดอะมิโนชนิดตาง ๆ
โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
97
ภาพที่ 1.81 การเชื่อมกรดอะมิโนกับ tRNA
ดังน้ันกรดอะมิโนที่จะมาตอเปนสายโพลเีปปไทดตองผานกระบวนการกระตุน (activated) มากอน
กรดอะมิโนเชื่อมกันดวยพันธะเปปไทด (peptide bond) โดยปลายดานอะมิโน (amino group) และดานคารบอกซิล (COOH group) จะสูญเสียโมเลกุลของน้ําออกมา
กระบวนการ translation
1. การเร่ิมตน (Initiation) ไรโบโซมหนวยยอยขนาดเล็กและปจจัยเริ่มตนซึ่งเปนตัวเรงปฏิกิ ริยามาจับกับเ อ็มอาร เ อ็นเอ และกรดอะมิ โนเมไทโอนีนที่ มีห มูฟอร มิลที่ ปลายสุด (N-formylmethionine: f-met) เปนกรดอะมิโนตัวแรกที่ทีอารเอ็นเอ (initial tRNA) นํามายังรหัสหรือโคดอนเริ่มตน AUG ของเอ็มอารเอ็นเอ โดยตําแหนงที่ของอะมิโนเอซิล ทีอารเอ็นเอ (aminoacyl tRNA) เขามาน้ันเปนตําแหนงพีไซต หลังจากนั้นไรโบโซมหนวยยอยขนาดใหญจะเขามาประกบกับไรโบโซมหนวยยอยขนาดเล็ก จึงทําใหไรโบโซมพรอมจะทําหนาที่ตอไป โดยตําแหนงบนไรโบโซมนั้นมีตําแหนง พีไซต (P site) เอไซต (A site) อีไซต (E site)
2. การตอสาย (translocation) aminoacyl tRNA โมเลกลุที่ 2 ที่มีแอนติโคดอนเขาคูกับโคดอนถัดไปของเอ็มอารเอ็นเอ นํากรดอะมิโนตัวที ่ 2 เขามาเรียงตอกับกรดอะมิโนตัวแรก ณ ตําแหนง A site จากนั้นเชื่อมกรดอะมิโนทั้งสองดวยพันธะเปปไทด ณ ตําแหนง P site โดยไรโบโซมจะเคลื่อนที่ไปยังโคดอนถัดไปในทิศทางจาก ปลาย 5 / ไป 3 / ทําใหทีอารเอ็นเอโมเลกุลแรกหลดุออกไป ณ ตําแหนง E

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
98
site จากนั้นทีอารืเอ็นเอโมเลกุลที่ 3 ที่มีแอนติโคดอนเขาคูกับโคดอนลําดับถัดไป นํากรดอะมิโนตัวที่ 3 เขาจับกับเอ็มอารเอ็นเอตรงโคดอนที่วาง แลวสรางพันธะเพปไทดระหวางกรดอะมิโนตัวที่ 2 กับตวัที่ 3
3. กระบวนการสิ้นสุดการสังเคราะห (termination) เม่ือไรโบโซมเคลื่อนมาทีละรหัสจนเจอรหัสหยุด (stop codon) คือ UAA UAG UGA รหัสใดรหัสหนึ่งจะไมมีอะมิโนเอซิลทีอารเอ็นเอเขามาเกาะ โดยมีแฟคเตอรปลดปลอย (release factor) เขามาแทนจึงทําให สายโพลีเปปไทด หลุดออกมาและหยุดการแปลรหัส ไรโบโซมหนวยยอยขนาดเล็กและขนาดใหญแยกออกจากกันและเอ็มอารเอ็นเอหลุดจากไรโบโซม
ภาพที่ 1.82 กระบวนการ translocation
โพลีโซม (polysome) การสังเคราะหโปรตีนสามารถทําไดพรอม ๆ กันหลาย ๆ สายโดยอาศัย mRNA เพียงสายเดียว
บริเวณที่มีเหตุการณเชนน้ีถูกเรียกวาโพลีโซม (polysome)
ภาพที่ 1.83 โพลีโซม

โรงเรียนมหิดลวิทยานุสรณ (องคการมหาชน) สาขาชีววิทยา เอกสารประกอบการเรียนการสอนรายวิชา ว 30261 พันธุศาสตรและวิวัฒนาการ บทที่ 1 พันธุศาสตร
99
ความแตกตางการแสดงออกของยีนในสิ่งมีชีวิต ในยูคาริโอทการควบคุมการแสดงออกของยีนในเซลล เร่ิมในนิวเคลียสยีนในดีเอ็นเอ ผาน
กระบวนการ ทรานสคริปชัน และ กระบวนการตัดตออารเอ็นเอ (RNA processing) การตัดอินทรอน (intron) ออกแลวตอเอกซอน (exon) เขาดวยกัน จากนั้นก็เติมแคบ (cap) ที่ปลาย 3’ โดยที่ ปลาย 5’ tail เปนโพลี A (poly A) คือ A หลายตัวเติมลงไป ไดเปน อารเอ็นเอที่สามารถทํางานไดแลวสงไปยัง ไซโทพลาสซึม ผานกระบวนการ ทรานสเลชัน เกิดเปนสายโพลีเปปไทด
ในโปรคารีโอตการสังเคราะหโปรตีนจะเกิดขึ้นพรอม ๆ กับการถอดรหัส mRNA เน่ืองจาก โปรคารีโอตไมมีเยื่อหุมนิวเคลียสอยางพวกยูคารีโอต และ mRNA ของมันก็ไมมีอินทรอน
ภาพที่ 1.84 การสรางโปรตีนในโปรคารโีอต





























