Embed Size (px)
Citation preview

EL PROCESO FOTOSINTÉTICO
La conocida ecuación de la fotosíntesis.
luz y clorofila
CO2 + H20 C6H1206 + 6 02
Señala simplemente los compuestos iniciales y finales de una serie de reacciones. Desde
un principio, los bioquímicos y fisiólogos lo comprendieron así y trataron de dar
explicaciones del proceso de acuerdo con la lógica científica, suponiendo que el CO2 se
disuelve en el agua y forma H2C03, o bien basándose en la hipótesis de la formación de
aldehido fórmico (CH20), que se polimerizaría para formar glucosa (CH20)6; tales
fueron las hipótesis de Baeyer, Willstätter y otros.
Posteriormente, los estudios de Van Niel, corroborados por el uso de 018
, mostraron
que el O2 que se desprende en la fotosíntesis viene totalmente del agua, o sea que ésta
sufre una fotólisis y es su H2 el que se incorpora al CO2.
6C02 + 6H2O18
C6H12O6 + 6O218
Con éste y otros descubrimientos (Hill, etc.) se dio una nueva hipótesis sobre la
fotosíntesis, hipótesis que incluía la presencia activa de moléculas transportadoras de
energía, como el ATP y aceptoras - transportadoras de electrones e y H2, el
dinucleótido de adenina nicotinamida o NAD (antiguamente llamada nucleótido de
difosfopiridina o DPN) y el dinucleótido de adenina nicotinamida fosfatado o NADP
(antiguamente llamado nucleótido de trifosfopiridina o TPN), así como la riboflavina
y citocromos; de este modo, el H2 llevado en forma activa por una molécula de
NADPH2, gracias a la energía aportada por el ATP (energía que en principio venía de
la luz y que había sido transformada por la clorofila), podría ser integrado en una
molécula para producir glucosa.
En los últimos tiempos, el descubrimiento del efecto Emerson desencadenó una
serie de estudios, que culminaron en una nueva teoría de la fotosíntesis. El efecto
Emerson consiste en lo siguiente: si se ilumina un cloroplasto (o un fragmento activo
de c1oroplasto llamado cuantosoma) con luz monocromótica, se observa una caída en

el rendimiento cuántico con longitudes de onda mayores de 680 nm, pero la eficiencia
del cuantosoma es restaurada si se aplica simultáneamente una longitud de onda más
corta. Esto plantea la existencia de dos procesos fotoquímicos diferentes y la
existencia de dos clases de clorofila a, una con absorción máxima a 673 nm (Chla
673) y otra a 683 nm (Chla 683). Al parecer, existe todavía otra: P700.
La fotosíntesis incluye, pues, dos grupos de pigmentos (I y II); la energía lumínica
que mueve al Sistema I se capta por la Chla 683, P700 y carotenoides; la que mueve al
Sistema II se capta por la Chla 673, clorofila b y ficobilinas.
Una descripción muy resumida del proceso de la fotosíntesis, como se entiende
ahora, se encuentra en la figura 22. Como ahí se presenta, la luz incide sobre la hoja y
excita dos diferentes sistemas fotosintéticos: el sistema de pigmentos PI y el sistema de
pigmentos PII. El sistema PI es excitado por la luz de más de 685 nm, y como
resultado de la absorción de fotones, los electrones de la c1orofila a y carotenoides
salen de las órbitas alejadas del núcleo para ser capturados por la ferredoxina
(ferredoxina es un nombre genérico que designa una proteína con hierro, pero sin
núcleo porfirínico); la ferredoxina pasa estos electrones a una reductasa que los
transmite al NADP, el cual queda reducido; para guardar el balance interno de la
molécula, el NADP, al aceptar un e-, debe también aceptar un H2, con lo cual queda
como NADPH.

Figura 22.- Esquema del proceso fotosintético. La luz activa un sistema de pigmentos (P II),
efectuándose la fotólisis del agua. Los electrones así liberados pasan a través de aceptores, formando
un gradiente de disminución de energía, y con la energía liberada se sintetiza ATP. Otro sistema de
pigmentos (P I), al activarse posibilita la reducción de NADP, con ATP y NADPH. La planta puede
reducir el CO2 y fabricar alimento.
El proceso descrito deja dos incógnitas: ¿se restablece el sistema PI después de
perder e-?, ¿de dónde viene el H2 que va al NADP? Al incidir sobre el c1oroplasto la
luz excita también al sistema de pigmentos PII que, como sucede en PI, absorbe la
energía del fotón y emite electrones. La gran descarga de energía que significa esta
pérdida de electrones es empleada para romper la molécula de H20, desconociéndose
el mecanismo del fenómeno; los electrones son aceptados por un compuesto: la
plastoquinona. A partir de aquí se encuentra una cadena de aceptores intermediarios
de e-, que se van reduciendo y oxidando conforme aceptan y despiden e
- en un
gradiente descendente de energía, el cual termina con la aceptación de e- del sistema
de pigmentos PI, que restaura así su integridad intramolecular. En el paso de
electrones a través de aceptores con potenciales redox decrecientes, el exceso de
energía es empleado para efectuar la síntesis de ATP, molécula que lleva alto
contenido energético en su tercer P.

Como se ve, en este caso hay un viaje sin retorno de los electrones del sistema PII
al NADP, con formación de ATP, por lo que se llama una fotofosforilación acíclica.
En ciertas condiciones no se reduce al NADP y los e- (electrones) emitidos por la
ferredoxina van al citocromo b, por lo cual se habla de una fotofosforilación cíclica.
El proceso termina, pues, con la síntesis de dos moléculas con alta energía: el
NADPH2 y el ATP; para producirlas ha habido una fotó1isis de H20, y en resumen, la
energía lumínica ha quedado atrapada en la célula y se habla ahora de la generación
de energía metabólica.
La síntesis de moléculas con H2 en forma activa y con alta energía en forma de P,
capacita a la célula para sintetizar una molécula donde quede la energía
almacenada a su disposición; esta síntesis incluye la asimilación del CO2 del aire.
Las transformaciones de los compuestos químicos hasta llegar a glucosa y almidón,
incluyen el llamado ciclo de Calvin, que describe las transformaciones primarias. El
CO2 es aceptado por una molécula con 5 carbones (5C), llamada difosfato de
ribulosa y se forma una molécula de 6C que se rompe en dos moléculas de ácido
fosfoglicérico (3C cada una); estas transformaciones no necesitan luz y se llaman la
reacción oscura. Posteriormente, el ácido fosfoglicérico, por acción del NADPH y el
ATP formados en la reacción de la luz, pasan a formar al aldehido fosfoglicérico
(compuesto de 3C). Este puede tomar varios caminos, entre ellos formar difosfato
de fructosa (azucar de 6C, el cual se deriva posteriormente en glucosa, almidón y
sacarosa) o bien formar fosfato de ribosa, azúcar de 5C que se transformará en
difosfato de ribulosa para cerrar el ciclo (figura 23 A).
A partir de 1960 está en continua investigación un proceso fotosintético diferente
del tradicional. Descubierto en cultivos de piña, se comprobó que, una vez
realizados los procesos lumínicos, el primer carbohidrato formado en la fotosíntesis
no es el ácido fosfoglicérico, de tres carbonos, sino los ácidos málico y aspártico, que
contienen cuatro átomos de carbono. En general, son las plantas tropicales las que

presentan esta vía, llamada C4, por el número de carbonos del primer compuesto
formado en la fotosíntesis. Nótese como apartir de los ácidos málico y aspártico se
puede reintegrar al ciclo de Calvin al CO2, o bién pueden seguir dichos ácidos una
segunda ruta que es formar ácido pirúvico el cual posteriormente se transformará
en ácido fosfoenolpirúvico compuesto aceptor de CO2, reintegrandose de esta forma
al ciclo de las plantas C4.
de esta vía (C4 como primer producto de la fotosíntesis) en plantas C4 cultivadas son, además de
la piña, el sorgo, la caña de azúcar y el maíz, en las cuales se ha demostrado que la anatomía foliar
es también peculiar.
Como se aprecia en el diagrama de la figura 23b, el ácido fosfoenolpirúvico funciona
bombeando CO2 en altas concentraciones al ciclo de Calvin. La ventaja de las plantas C4 es, pues,
la presencia del ácido fosfoenolpirúvico que, al tener gran afinidad por el dióxido de carbono y
aceptarlo con gran eficiencia, causa un mayor rendimiento fotosintético de la planta.
Difosfato de ribulosa

El ciclo de Calvin en las plantas C4 se desarrolla en forma idéntica a la tradicional. Sin
embargo, en las plantas C3, la ribulosa difosfato también tiene afinidad por el oxígeno, el cual
bloquea la fotosíntesis y desencadena el proceso de fotorrespiración.
Fases de la fotosíntesis
La fotosíntesis es un proceso fundamental para los seres vivos, ya que permite la transformación de la energía luminosa procedente del Sol en energía química, que es almacenada para ser utilizada según las necesidades del organismo. A partir de elementos inorgánicos sencillos, el agua y el dióxido de carbono, se sintetiza la glucosa, que a su vez puede ser empleada para la producción de almidón, sacarosa y polisacáridos, asumiendo por tanto una función estructural; o bien puede servir de sustrato en las reacciones de respiración celular, con las que se obtiene energía (bajo la forma de moléculas de ATP); en ese caso
desempeña una función energética. En la fotosíntesis también se produce, mediante un proceso denominado fotofosforilación, cierta cantidad de ATP. No todos los seres vivos están en condiciones de realizar este proceso fundamental: esta capacidad sólo la poseen algunas bacterias, las cianobacterias, las algas y las plantas, que poseen las moléculas de clorofila necesarias para la absorción de la luz solar y las enzimas indispensables para realizar las reacciones fotosintéticas. La clorofila está ligada a las membranas del tilacoide, contenidas en el interior de unos orgánulos llamados cloroplastos (excepto en las bacterias, en las cuales el pigmento está asociado a invaginaciones de la membrana celular).

&& 2011
FACTORES QUE MODIFICAN LA FOTOSÍNTESIS
Los principales factores que modifican el proceso fotosintético son el CO2, la temperatura y
la luz.
El CO2 es la fuente del carbono para el alimento primario de la planta, a partir del cual se
sintetizan los demás compuestos. El contenido de CO2 en la atmósfera es de 0.03% y si bien casi
siempre el factor limitante es otro, hay casos en que la fotosíntesis se ve limitada por la cantidad
de CO2 a su disposición; esto sucede cuando hay una alta densidad de población del cultivo. En
invernadero, Molisch comprobó desde principios de este siglo que se obtienen tremendos
aumentos en rendimiento cuando se "abona" la atmósfera con CO2. Las plantas verdes presentan
respiración aerobia, pero soportan tensiones de O2 muy bajas y tensiones de CO2 muy altas: hasta
5 o 10%; sin embargo. Wilkins encontró que un aumento de más de 0.1 % de CO2 no influye en el
rendimiento, en tanto que Molisch probó con éxito concentraciones más altas. La absorción del
CO2 interfiere con la transpiración.
La temperatura es un factor importante en la fotosíntesis, pues si bien en la parte
fotoquímica no tiene efecto, si lo tiene y grande en la reacción oscura así como en la movilización
de los azúcares, y no debe olvidarse que la falta de remoción del producto, por movilización o uso,
trae consigo la detención de la reacción. En general, el mínimo para la fotosíntesis se encuentra a
0°C, el óptimo a los 40°C y el máximo a los 50°C. En realidad, al ir elevando la temperatura, el
óptimo se encuentra a los 40°C, pero si se mantiene esta temperatura largo tiempo, decrece la
producción de fotosintetizados, de modo que el óptimo real, que se sostiene indefinidamente, se
encuentra entre los 26 a 30°C.
La luz es fuente energética para la fotosíntesis; hoy se sabe que una planta puede estar
iluminada continuamente, y si bien la iluminación constante puede afectar los procesos
metabólicos, en algunas especies, Molisch comprobó que cuando no es éste el caso, la planta
puede iluminarse día y noche sin signos de agotamiento.

La luz es esencial para la planta desde dos puntos de vista: a) para que la planta sintetice
clorofila, b) porque es la energía primaria que la clorofila transformara en energía química. Desde
el primer punto de vista, la planta exige una cantidad mínima de luz para que la protoclorofilida
pase a clorofilida y luego a clorofila, de modo que la falta de luz determina clorosis; conforme se
aumenta la intensidad lumínica, se incrementa la síntesis de clorofila, hasta un nivel considerado
adecuado, un aumento en intensidad lumínica por encima del nivel adecuado propicia la rápida
destrucción de la clorofila que su síntesis, hasta llegar a clorosis. Toda planta tiene un umbral, a
partir del cual el aumento en intensidad de luz es nocivo; experimentos ya antiguos de Shantz han
mostrado que la papa, la lechuga y el rábano se desarrollaron mejor cuando recibieron la mitad de
la luz óptima que requiere el maíz para su desarrollo. La intensidad lumínica óptima para un buen
crecimiento y tamaño foliar máximo fluctúa de 2500 a 3000 bujias-pie.
De los factores de la fotosíntesis, la propia cantidad de clorofila no es limitante por lo
general y se ha visto (en algodón, melón, pepino y otros cultivos) que a una planta se le pueden
quitar hasta 25% de sus hojas sin afectar el rendimiento. Sin embargo, los factores edáficos a veces
pueden determinar deficiencias serias de minerales esenciales para la fotosíntesis, como el Fe, Mn,
y Mg, bajo estas condiciones sí se limita seriamente la cantidad de clorofila. En clima desértico es
habitual que se tenga una deficiencia de Fe y exceso de luz, lo que determina una limitada
proporción de clorofila en hojas de cultivos como el peral, provocando clorosis en estas.

METABOLISMO DE LOS HIDRATOS DE CARBONO.
En la fotosíntesis, la planta fabrica ácido fosfoglicérico directamente al asimilar el CO2, y
luego, por acción de NADPH y ATP sintetizados con energía lumínica, reduce al ácido
fosfoglicérico, pasándolo a aldehído. A partir de éste, la planta fabrica fosfatos de hexosa: glucosa-
fosfato y fructosa-fosfato.
Estos son los responsables de las transformaciones de los hidratos de carbono, ya que la
energía necesaria para ello es dada por el radical con P, que en realidad procede del ATP
sintetizado en la fotosíntesis.
El azúcar central de la planta es glucosa-l-fosfato, del cual pueden formarse los principales
hidratos de carbono, la glucosa se forma probablemente en la fotosíntesis en forma directa. La
manera más general de su formación es por acción de una fosfatasa sobre la glucosa-l-fosfato. El
catabolismo de la glucosa es la respiración, proceso muy importante. La sacarosa, azúcar de
transporte, es anabolizada a partir de la unión de glucosa-l-fosfato con la sacarosa. Su catabolismo
puede ser por unión con el ácido fosfórico en presencia de una enzima, o bién por acción de la
sacarosa, con la intervención de la enzima hidrolítica que rompe la molécula en dos.
El almidón se constituye a partir de la glucosa-1-fosfato y su catabolismo es también por
dos caminos: uno, catabolizado por una fosforilasa, y el otro por la enzima hidrolítica amilasa.
Los monosacáridos más importantes en la planta, se pueden dividir en: a) Pentosas la más
rerpresentativa es la Ribosa (carbohidrato de 5C) y en b) Hexosas (carbohidratos de 6C) la más
importante la glucosa, también están la fructosa y la galactosa.
Respecto a los Disacáridos el más representativo es la sacarosa, el cual se forma por la unión de
glucosa + fructosa, además existe la maltosa conformada por la unión de la glucosa + glucosa.
Un trisacárido importante lo es la Rafinosa. En tanto que un polisacárido sobresaliente lo es la
hexosana denominado almidón, celulosa e inulina en tanto que una pentosana importante lo es el xilano.
Respecto a hidratos de carbono compuestos lo más representativos son: pectinas, gomas, mucílagos,
glucósidos, taninos.
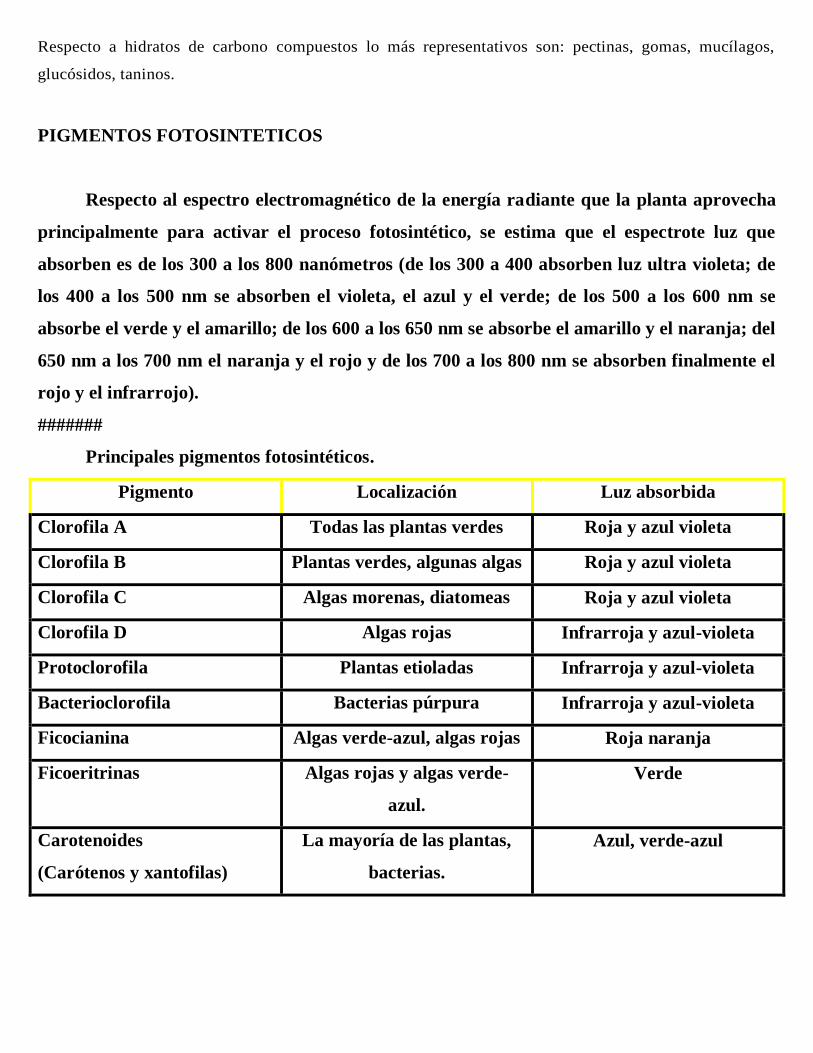
PIGMENTOS FOTOSINTETICOS
Respecto al espectro electromagnético de la energía radiante que la planta aprovecha
principalmente para activar el proceso fotosintético, se estima que el espectrote luz que
absorben es de los 300 a los 800 nanómetros (de los 300 a 400 absorben luz ultra violeta; de
los 400 a los 500 nm se absorben el violeta, el azul y el verde; de los 500 a los 600 nm se
absorbe el verde y el amarillo; de los 600 a los 650 nm se absorbe el amarillo y el naranja; del
650 nm a los 700 nm el naranja y el rojo y de los 700 a los 800 nm se absorben finalmente el
rojo y el infrarrojo).
#######
Principales pigmentos fotosintéticos.
Pigmento Localización Luz absorbida
Clorofila A Todas las plantas verdes Roja y azul violeta
Clorofila B Plantas verdes, algunas algas Roja y azul violeta
Clorofila C Algas morenas, diatomeas Roja y azul violeta
Clorofila D Algas rojas Infrarroja y azul-violeta
Protoclorofila Plantas etioladas Infrarroja y azul-violeta
Bacterioclorofila Bacterias púrpura Infrarroja y azul-violeta
Ficocianina Algas verde-azul, algas rojas Roja naranja
Ficoeritrinas Algas rojas y algas verde-
azul.
Verde
Carotenoides
(Carótenos y xantofilas)
La mayoría de las plantas,
bacterias.
Azul, verde-azul