Embed Size (px)
Citation preview

452 KARRIERE, KÖPFE & KONZEPTE
BIOspektrum | 04.12 | 18. Jahrgang
DOI: 10.1007/s12268-012-0208-6© Springer-Verlag 2012
ó Methan ist ein potentes Treibhausgas, dasaber auch zur regenerativen Energiegewin-nung genutzt wird. Ein Großteil der biologi-schen Methanemission ist auf Acetat-abbau-ende methanogene Archaeen zurückzufüh-ren. Entgegen der großen Diversität derMikroorganismenwelt wurde diese Stoff-wechseleigenschaft bislang nur in zwei Gat-tungen gefunden, Methanosarcina und Metha-nosaeta. In der Methanogenese mit Acetat alsSubstrat bilden sie neben Methan auch redu-ziertes Ferredoxin und Heterodisulfid. DasRecycling dieser beiden Produkte innerhalbder Zelle erfolgt über eine membrangebun-dene Atmungskette, die in Methanosarcina-Arten gut, in Methanosaeta-Arten kaum unter-sucht ist. Methanosarcina mazei nutzt für dieFerredoxinoxidation eine Hydrogenase (Ech).Der in dieser Reaktion produzierte Wasser-stoff diffundiert von der Innenseite der Mem-bran auf die Außenseite und wird dort voneiner zweiten Hydrogenase oxidiert, die soProtonen im extrazellulären Raum freisetzt.Die Elektronen werden über ein membran-lösliches Phenazinderivat auf die terminaleReduktase übertragen, die das Heterodisul-fid reduziert und ebenfalls Protonen in denextrazellulären Raum entlässt. Damit stelltebis vor Kurzem die Ech-Hydrogenase das letz-te Glied der Atmungskette dar, für das eineIonentranslokation und somit eine Beteili-gung an der Energiekonservierung zwar pos-tuliert, aber nicht biochemisch belegt war. Die
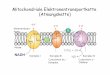
Entwicklung eines enzymatischen Systemszur Ferredoxinreduktion [1, 2] ermöglichte esschließlich, Ferredoxin-abhängige Ionen-translokation an invertierten Membranvesi-keln zu messen und so zu zeigen, dass auchdie Ech-Hydrogenase an der Etablierung einesIonengradienten beteiligt ist [3]. Die Ech-Hydrogenase bildet demnach eine neue Kop-plungsstelle im methanogenen Stoffwechsel(Abb. 1), die im Zuge der Ferredoxinoxida-tion Protonen in den extrazellulären Raumpumpt und so zur Energetisierung der Zyto-plasmamembran beiträgt.
Methanosaeta-Arten hingegen besitzen kei-ne Hydrogenasen und können daher nichtden beschriebenen Weg zur Ferredoxinoxi-dation nutzen. Membranfraktionen vonMethanosaeta thermophila zeigen allerdingshohe Aktivitäten der Atmungskette Acetat-abbauender Methanogener, der Ferredo-xin:Heterodisulfid-Oxidoreduktase [4]. DieFrage ist nun: Welches ist das Ferredoxin-oxi-dierende Enzym?
Analysiert man die bis jetzt sequenziertenMethanosaeta-Genome, so fällt auf, dass sichin allen Genomen Gene finden, die für eineF420H2-Dehydrogenase codieren. DiesesEnzym ist eigentlich an der methylotrophenMethanogenese beteiligt und sorgt dort fürdie Oxidation des Kofaktors F420H2. Metha-nosaeta-Arten sind allerdings nicht in derLage, auf methylierten Verbindungen zuwachsen, und erwartungsgemäß kann die
Membranfraktionvon Methanosaetathermophila auchkein F420H2 umset-zen. Dennoch werdendie Gene, die für die
F420H2-Dehydrogenase codieren, stark expri-miert (unveröffentlicht). Versucht man, überDatenbankvergleiche einen anderen Elektro-nendonor mit der F420H2-Dehydrogenase zuverknüpfen, so fällt auf, dass die F420H2-Dehy-drogenase große Ähnlichkeit zur NADH-Dehy-drogenase der aeroben Atmungskette auf-weist. Interessanterweise fehlen im Genomvon Methanosaeta-Arten Gene, die für dieModule der NADH- und F420H2-Oxidationcodieren. Stattdessen ist eines der hydrophi-len Proteine der F420H2-Dehydrogenase C-ter-minal mit einer Lysin-reichen Sequenz ver-längert. Diese ist bei physiologischem pH posi-tiv geladen und könnte somit gut mit demnegativ geladenen Elektronenüberträger Fer-redoxin inter agieren (Abb. 1). In Methano-saeta-Stämmen könnte also die ungewöhn -liche „F420H2/ NADH“-Dehydrogenase dasgesuchte Ferredoxin-oxidierende Enzym derAtmungskette sein. ó
Literatur[1] Welte C, Deppenmeier U (2012) Protononen-Translokationin Methanogenen. BIOspektrum 2:159–161[2] Welte C, Kallnik V, Grapp M et al. (2010) Function of Echhydrogenase in ferredoxin-dependent, membrane-bound elec-tron transport in Methanosarcina mazei. J Bacteriol 192:674–678[3] Welte C, Krätzer C, Deppenmeier U (2010) Involvement ofEch hydrogenase in energy conservation of Methanosarcinamazei. FEBS J 277:3396–3403[4] Welte C, Deppenmeier U (2011) Membrane-bound elec-tron transport in Methanosaeta thermophila. J Bacteriol193:2686–2670
Korrespondenzadresse:Dr. Cornelia WelteInstitut für Mikrobiologie und BiotechnologieRheinische Friedrich-Wilhelms-Universität BonnMeckenheimer Allee 168D-53115 BonnTel.: [email protected]
VAAM-Promotionspreis 2012
Acetat-abbauende methanogene Archaeen –Neues aus der Atmungskette
CORNELIA WELTE
INSTITUT FÜR MIKROBIOLOGIE UND BIOTECHNOLOGIE, UNIVERSITÄT BONN
Cornelia WelteJahrgang 1984. 2003–2008 Bio-logiestudium an der UniversitätBonn und der Lunds Universitet,Schweden. 2008–2011 Promo-tion am Institut für Mikrobiologieund Biotechnologie in Bonn beiProf. Dr. U. Deppenmeier, dortseit 2011 Postdoc.
¯ Abb. 1: Schema der Atmungsketten von Methanosarcina mazei (oben)und Methanosaeta thermophila (unten) beim Wachstum auf Acetat.Elektronen werden über reduziertes Ferredoxin (Ferredoxinred) in dieAtmungsketten eingeschleust. Ms. mazei oxidiert Ferredoxinred über dieProtonen-translozierende Ech-Hydrogenase. Mt. thermophila könnte hier-für die F420H2-Dehydrogenase (Fpo-Komplex) mit der Untereinheit Fpolund deren Lysin-reichen C-Terminus (KKK) als Elektroneneintrittsstellenutzen.
![Kapitel 6) Ctitratcyklus+Atmungskette [Kompatibilitätsmodus]biochemietrainingscamp.de/stoff/ci/der_malat_shuttel.pdf · Pyruvat 2. Malat 3. Aspartat ... GOT = ASAT = Glutamat–](https://img.pdfslide.tips/doc/110x75/5d6451b488c993e91b8bcc85/kapitel-6-ctitratcyklusatmungskette-kompatibilitaetsmodusbi-pyruvat-2-malat.jpg)

![Kapitel 6) Ctitratcyklus+Atmungskette [Kompatibilitätsmodus]biochemie-trainings-camp.de/stoff/ci/Ctitratcyklus.pdf · das Enzym: Citratsynthase reguliert durch mitochondriales ATP,](https://img.pdfslide.tips/doc/110x75/5d4e6d6f88c99303708bac76/kapitel-6-ctitratcyklusatmungskette-kompatibilitaetsmodusbiochemie-trainings-campdestoffci.jpg)


















![Kapitel 6) Ctitratcyklus+Atmungskette [Kompatibilitätsmodus]biochemietrainingscamp.de/stoff/ci/Ctitratcyklus.pdf · TPP Liponsäure NAD FAD CoA ( Vergleiche die Pyruvat-Dehydrogenase)](https://img.pdfslide.tips/doc/110x75/5e1287ca50baaa02a71d88e6/kapitel-6-ctitratcyklusatmungskette-kompatibilittsmodusbi-tpp-liponsure.jpg)



![8 Durch Grubenwasseranstieg entstehendes · PDF fileRuhrgebiet (Quelle: Grundbau-Taschenbuch, Teil 3, 2010 [118]) Jahr nach Abbauende 1 2 3 4 5 Prozentualer Anteil der Senkung im Jahr](https://img.pdfslide.tips/doc/110x75/5a7fd4ac7f8b9a571e8c08ff/8-durch-grubenwasseranstieg-entstehendes-quelle-grundbau-taschenbuch-teil.jpg)


