Embed Size (px)
Citation preview

ALGAS MARINAS COMO INHIBIDORES
POTENCIALES DE PROCESOS
INVOLUCRADOS EN EL DESARROLLO DE
ALZHEIMER
TESIS
QUE PARA OBTENER EL GRADO DE
MAESTRÍA EN CIENCIAS EN MANEJO DE RECURSOS MARINOS
PRESENTA
BERNARDO VEYRAND QUIRÓS
LA PAZ, B.C.S., JUNIO DE 2016
INSTITUTO POLITECNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS



IV
El hecho de que un misterio esté más allá del alcance de la comprensión
humana no significa que no debamos intentar comprenderlo, hasta donde ello
sea posible, dentro del dominio del intelecto.
Taimni

III
Dedicatoria
A mi familia y amigos. Así como aquellas personas que presentan trastornos
neurológicos. Por no perder las esperanzas de que se encuentre la cura para sus
problemas o deficiencias, y al fin su cuerpo y mente sólo les pertenecerá a ustedes
sin temor a ser controlados por una enfermedad.

IV
Agradecimientos
En ésta ocasión el recorrido no fue tan largo como en la licenciatura, sin
embargo si más complejo y más significativo. Cumpliendo una meta personal con el
tema de tesis elegido y el poder desarrollarla en base a una idea propia. Considero
éste arduo trabajo como un peldaño importante para mi formación como futuro
investigador, en el que se ve reflejado no sólo mi trabajo y compromiso si no el de
mucha gente que intervino para que todo esto fuera posible. Aunque considero ésta
tesis como una etapa más compleja y madura creo que fueron tiempos muy
divertidos que siempre llevaré conmigo. Este proyecto reúne el trabajo y esfuerzo de
muchos, conjuntando sueños y anhelos de varias personas que se relacionaron a lo
largo de estos años por lo que el crédito no puede ser sólo mío.
Me gustaría comenzar agradeciendo a mi familia, esas personas que sin ellos
esto no hubiera sido posible. Mis padres quienes me apoyan constantemente con
sus palabras de ánimo y se han sentido orgullosos de mis logros. A mi madre, por
darme siempre su apoyo incondicional y aclarar mi mente siempre que lo requiero.
Por alentar mis sueños y enseñarme a volar, pero a la vez como mantener mis pies
en la tierra y recordar siempre quien soy. A mi padre, por seguir siendo mi ejemplo a
seguir, y ser aún el superhéroe en el que creo. Por enseñarme a ser crítico, tener
principios y siempre seguir adelante. El me instruyó en que uno no debe dejar
inconcluso algo y debe proponerse a terminar sus metas siempre. “Siempre termina
lo que empezaste”. A su vez si no fuera por su sueño de estudiar biología marina yo
no me hubiera inmiscuido en éste mundo, así como las geniales amistades que hizo
que ahora representan mi familia. Gracias por seguirme apoyando en mis metas y
por las enseñanzas de valores y sentido de la responsabilidad.
A mi pequeña hermana Dominique, que aunque ya es toda una chica
universitaria yo la sigo viendo pequeña. Aunque no intervino directamente en mi
formación académica si lo hace en mi vida cotidiana. Gracias por el apoyo y por
siempre confiar en mí. Por los buenos momentos que vivimos siempre, la compañía y
todo el amor que recibo de tu parte. Porque con ella crecí y sigo creciendo como
persona. Sin ti la vida sería muy aburrida.
A mis abuelos quienes son parte fundamental en mi vida, mi abuela
Esperanza quien siempre me recibió con una gran sonrisa y me preguntaba acerca
de mi carrera, así como a mi Abuelo Rodolfo que aunque ya no está con nosotros
siempre me impulsó a seguir adelante y se interesó por mi formación académica; sé
que estaría orgulloso de éste logro. A mi abuela Clara quien me alienta siempre y me
ayuda en lo que pueda.
A mi familia adoptiva, mis tíos biologuescos, en especial a Alberto y Lucía,
quienes siempre me apoyan y se han interesado en mi carrera siempre, además de

V
enseñarme que la familia no se crea por lazos sanguíneos sino por el apoyo y cariño
que te brindan siempre. Gracias por guiarme en el sentido de la biología marina y su
amabilidad y hospitalidad de siempre.
Y hablando de familia adoptiva, quisiera agradecer a mis amigos hermanos,
Roberto y Pablo (las perras) que siempre están ahí para mí y nos apoyamos en todo
lo que podamos. Gracias por éstos años de amistad y por seguirme soportando, así
como por las horas de diversión y horas insanas de videojuegos y buenas pláticas
sin sentido. A Priscila por ser buena amiga y siempre saber escucharme y
aconsejarme, así como tenernos paciencia siempre con nuestras tonterías y
majaderías. A Edmundo que aunque esté a distancia seguimos en contacto y
siempre es grato y divertido hablar con él. A Luna (Mibi) por seguirnos soportando
aún después de haber salido de la carrera y seguir cotorreando con nosotros. A
Fernando que aunque por diferentes ocupaciones ya no nos frecuentamos tanto, sé
que ahí está para apoyar. A Ilse por ser muy buena persona conmigo y mi amiga, por
escucharme y apoyarme en mis decisiones. A Koko que aunque la vida nos separó
fuiste y serás una parte importante en mi vida, por enseñarme a ver la vida desde
otro aspecto, comprenderla y apreciar los pequeños detalles y momentos
importantes, por enseñarme a querer cosas que creí no me gustaban, así como
ayudarme a comprender que no soy un robot sin sentimientos.
A Diana, por aparecer en un momento muy crucial de la vida, y apoyarme a
seguir adelante. Por mostrarme que siempre hay tiempo para divertirse y encontrar
espacios mágicos en la cotidianidad. Por encontrar momentos divertidos y elegantes.
Por ser tan talentosa y apasionada en su carrera que eso me motiva a ser mejor en
mi área y crecer como persona. Por iluminar mis malos días con una sonrisa y
motivarme con tus pláticas, pero sobre todo a enseñarme que somos un equipo, te
quiero.
A mis compañeros de generación y profesores que me impartieron clases
éstos años de maestría. A los amigos que hice en estos años, o con los que conviví
entre pasillos o cubículos. Sobre todo un agradecimiento importante a los miembros
del Laboratorio de Química de Algas en CICIMAR por su apoyo diario en mantener la
cordura, ya que si no se podía ayudar con algo al menos siempre tenían una sonrisa
que sacar. Un agradecimiento especial a Araceli, quién me salvó y ayudó en
múltiples ocasiones. A Elena, por consejos y ayuda así como ideas creativas.
Gracias por siempre sacarme alguna carcajada y por ser una persona muy original
que inspira mucha confianza. A la M.C. Elizabeth Rodríguez y M.C. Dora Arvizu por
consejo en laboratorio. A Ana, Ilse, Miguel, Valerie, Dania y Josué. Gracias por su
compañía y buenos momentos en el laboratorio así como fuera en las fiestas o
eventos. A Fany por ser mi amiga y por la compañía, gracias por los consejos de
hermana mayor y por la ayuda a la hora de la preparación de los experimentos.
Agradecimiento especial a Roman Makarov de Microbiología por su ayuda en el

VI
manejo del lector de microplacas. Y a Paola Tenorio por la confianza y la ayuda en la
parte de antioxidantes en el CIBNOR.
Quisiera agradecer por supuesto a mi comité de tesis; a mi Sensei, el Dr. Iván
Murillo, por haberme recibido y adoptado, por continuar las enseñanzas luego de ya
estar desde algún tiempo en el laboratorio e instruirme en el arte de los productos
naturales, coordinar el desarrollo y logística y estar abierto a mis locas y cambiantes
ideas para el desarrollo de la tesis y no perder la fe en que resultaría un trabajo
interesante. Al Dr. Mauricio Muñoz, que aunque “legalmente” no es mi director de
tesis yo siempre lo sentí como uno, quien siempre con buena fe y humor me apoyaba
e instruía en todo lo que podía, por su ayuda en la parte experimental y por siempre
corregirme o enseñarme técnicas nuevas. Siempre estuvo al pendiente del trabajo y
me mandaba constantemente información, revisiones o aportaciones importantes
para el trabajo. Al Dr. Sergio Martínez, la Dra. Noemí Águila y la Dra. Christine Band
por las molestias y por las revisiones de éste escrito gracias. Así como al Dr. Rafael
Riosmena por su ayuda en la identificación de organismos, que aunque se adelantó
en el camino de la vida, espero que donde esté siga estudiando lo que le gusta.
Por último me gustaría agradecer a las instituciones que hicieron esto posible.
Al Centro Interdisciplinario de Ciencias Marinas Instituto Politécnico por recibirme y
apoyar mi tema de tesis. Al IPN por el apoyo y financiación de proyecto por parte de
la Secretaría de Investigación y Posgrado del Instituto Politécnico Nacional. Así como
el apoyo por parte de los protocolos de proyectos de investigación; “Formulación y
evaluación de eficacia de cebos no convencionales para peces, moluscos y
crustáceos: Etapa de mejora continua”. Enero 2014-Diciembre 2014. SIP: 20144322.
Y el protocolo de investigación “Evaluación de algas marinas como fuente potencial
de compuestos para el tratamiento de enfermedades degenerativas e infecciosas:
Alzheimer, cáncer, artritis y dengue”. Enero 2015-Diciembre 2015. SIP: 20150538.
Gracias a CONACyT por la beca otorgada en éstos dos años así como a IPN por las
becas BEIFI y Beca Posgrado.
A todos los que han creído en mí, así como a los que no, así como todas las
personas que han sido transeúntes en esta historia denominada vida muchas
gracias. Si he olvidado a alguien, prometo beberme mis extractos de algas marinas
para prevenir el Alzheimer.

VII
ÍNDICE
LISTA DE TABLAS ................................................................................................... IX
LISTA DE FIGURAS .................................................................................................. X
GLOSARIO ............................................................................................................... XI
ABREVIATURAS .................................................................................................... XIII
RESUMEN .............................................................................................................. XIV
ABSTRACT .............................................................................................................. XV
1. INTRODUCCIÓN ................................................................................................... 1
2. ANTECEDENTES ................................................................................................... 6
2.1 Historia de la enfermedad de Alzheimer ............................................................ 6
2.2 Inhibidores de origen marino ............................................................................. 8
2.3 Inhibidores de acetilcolinesterasa .................................................................... 10
2.4 Actividad antioxidante e inhibidora de AChE ................................................... 11
3. JUSTIFICACIÓN ................................................................................................... 13
4. OBJETIVOS .......................................................................................................... 15
4.1 Objetivo general .............................................................................................. 15
4.2 Objetivos particulares ...................................................................................... 15
5. HIPÓTESIS ........................................................................................................... 15
6. MATERIAL Y MÉTODOS ...................................................................................... 15
6.1 Obtención de extractos .................................................................................... 15
6.2 Fraccionamiento del extracto etanólico ............................................................ 16
6.3 Pruebas de actividad ....................................................................................... 17
6.4 Determinación de cinética enzimática .............................................................. 19
6.5 Cálculos y análisis estadísticos ....................................................................... 20
6.6 Identificación estructural por análisis espectroscópico ..................................... 20
7. RESULTADOS ...................................................................................................... 21
7.1 Actividad antioxidante ...................................................................................... 21
7.2 Inhibición de acetilcolinesterasa ...................................................................... 23
7.3 Relación entre antioxidantes y actividad anticolinesterasa .............................. 25
7.4 Análisis de fracciones obtenidas de Eisenia arborea ....................................... 25
7.5 Relación en el efecto inhibidor de fracciones ................................................... 28
7.6 Caracterización de fracciones por análisis espectroscópico de IR ................... 28
8. DISCUSIÓN .......................................................................................................... 31

VIII
9. CONCLUSIONES ................................................................................................. 41
10. RECOMENDACIONES ....................................................................................... 42
10. BIBLIOGRAFÍA ................................................................................................... 43

IX
LISTA DE TABLAS
Tabla 1. Actividad antioxidantes de extractos etanólicos de algas marinas
presentados como porcentaje de inhibición, concentración máxima y la concentración
efectiva media………………………………………………………………………………21
Tabla 2. Valores resultantes en el ensayo de FRAP empleando extractos
algales…...…………………………………………………………………………………...22
Tabla 3. Inhibición de la enzima AChE empleando extractos etanólicos algales. Se
muestra el porcentaje de inhibición, la concentración máxima inhibitoria y la
concentración efectiva media………………………………………………………….…..24
Tabla 4. Cinética enzimática para la constante de inhibición (Ki) de los extractos y
fracciones empleadas de algas marinas contra AChE………………………………….25
Tabla 5. Actividad antioxidante de fracciones del alga parda Eisenia arborea
representados por el porcentaje de inhibición y su EC50 (g/mL).………………….....26
Tabla 6. Inhibición de la enzima AChE empleando fracciones de E. arborea.
Representación en porcentaje de inhibición y su EC50 (g/mL). ……………………..28
Tabla 7. Cinética enzimática para la constante de inhibición (Ki) de los extractos y
fracciones empleadas de algas marinas contra AChE………………………………….28

X
LISTA DE FIGURAS
Figura 1. Metabolitos marinos inhibidores de acetilcolinesterasa………………………9
Figura 2. Porcentaje de inhibición y EC50 de extracto crudo de E. arborea………….22
Figura 3. Porcentaje de inhibición de AChE empleando extractos etanólicos de algas
marinas a diferentes concentraciones……………………………………………………24
Figura 4. Análisis de varianza del porcentaje de inhibición de AChE de los diferentes
extractos etanólicos………………………….……………………………………………..24
Figura 5. Porcentaje de inhibición y EC50 de fracción 2 de E. arborea……………….27
Figura 6. Porcentajes de inhibición de DPPH empleando fracciones de E. arborea a
diferentes concentraciones………………………………………………………………...27
Figura 7. Espectro infrarrojo de las diferentes fracciones de Eisenia arborea………30
Figura 8. Estructura química de los inhibidores tacrina y donepezil………………….37
Figura 9. Estructuras químicas de dieckol y florofucofuroeckol……………………….39

XI
GLOSARIO
Absorbancia: Medida de la fracción de luz que es retenida por una muestra.
Acetilcolina: Neurotransmisor encargado de mediar la actividad sináptica del
sistema nervioso.
Acetilcolinesterasa: Enzima de la familia de las colinesterasas cuya función
principal es hidrolizar el neurotransmisor acetilcolina.
Acetilo: Radical derivado del ácido acético, que contiene un grupo metilo unido a un
grupo carbonilo con un electrón desapareado disponible.
Actividad biológica: Capacidad de una sustancia de alterar una o más funciones
fisiológicas en un sistema u organismo vivo.
Algas: Grupo heterogéneo de organismos autótrofos y fotosintéticos con estructuras
simples y sin tejidos diferenciados.
Amiloidogénico: Referente a que participa en la formación de placas amiloides.
Antioxidante: Molécula capaz de retardar o prevenir la oxidación de otras
moléculas.
Cromatografía: Técnicas de separación de compuestos basados en el reparto
diferencial entre dos fases inmiscibles.
Enzima: Proteína capaz de catalizar una reacción química.
Espectroscopia: Técnica analítica experimental que se basa en detectar la
absorción de la radiación electromagnética y relacionarla con los niveles de energía
implicados en la transición cuántica.
Estereotáxica: Técnica de radiocirugía que administra en una sesión una alta dosis
de irradiación focalizada en un lesión intracraneana.
Extracto: Producto concentrado que se obtiene de una planta u otro organismo en
base a una extracción con un disolvente conservando las propiedades de donde
proviene.
Florotaninos: Polifenoles derivados del floroglucinol presentes en algas pardas.
Inhibidor: Sustancia química que evita que un proceso o reacción se lleve a cabo.
Neurotransmisor: Biomolécula que transmite información entre neuronas por medio
de impulsos nerviosos.

XII
Ovillos neurofibrilares: Formaciones insolubles de proteínas acumuladas en las
neuronas, las cuales producen disfunción neuronal.
Placas amiloides: Depósitos extracelulares de -amiloide en el cerebro que se
asocian con la degeneración del sistema nervioso y la enfermedad de Alzheimer.
Proteína tau: Proteína que se encuentra en abundancia en el sistema nervioso y
radica en los axones neuronales, cuya función se relaciona con la unión de
microtúbulos.

XIII
ABREVIATURAS
ACh Acetilcolina
AChE Acetilcolinesterasa
A Péptidos -amiloides
DPPH 2,2-difenil-1-picrilhidracilo
DTNB 5,5’-dithiobis-(2-ácido nitrobenzóico)
EC50 Concentración efectiva media.
FRAP Del idioma inglés, ferric reducing antioxidant power (poder antioxidante
reductor de hierro)
HCl Ácido clorhídrico
IR Infrarrojo
Ki Constante de inhibición
Km Constante de Michaelis
mg Miligramos
mL Mililitros
mM Mili molar
TPTZ Tripiriditriazina
U Unidades de enzima
g Microgramos

XIV
RESUMEN
La enfermedad neurodegenerativa de Alzheimer conduce al déficit de memoria y la
muerte. Actualmente no existe ningún medicamento probado para curar o prevenir
dicha enfermedad, siendo un inconveniente la falta de conocimiento sobre los
mecanismos neuronales que subyacen a la patogénesis del problema. Los estudios
actuales aceptan la hipótesis amiloide, la cual describe la acumulación de la proteína
-amiloide en el sistema nervioso central. Los inhibidores para el sitio dual de la
acetilcolinesterasa son prometedores para el tratamiento del alzheimer. Éstos alivian
los déficits cognitivos y agentes modificadores de la enfermedad mediante la
inhibición de la agregación de placas amiloides así como el control de los niveles de
acetilcolina por medio del sitio de unión dual de la enzima. A su vez los péptidos
amiloides se han identificado como fuente de estrés oxidativo, adquiriendo un estado
de radical libre produciendo efectos neurotóxicos. El presente estudio propuso la
búsqueda de productos naturales de origen marino con potencial anti-alzheimer y
actividad antioxidante. Se determinó la actividad antioxidante por medio de los
ensayos colorimétricos del radical libre 2,2-difenil-1-picrilhidracilo y el poder
antioxidante reductor de hierro. El potencial anti-alzheimer fue determinado utilizando
un ensayo de inhibición de la enzima acetilcolinesterasa. Para ello se emplearon
doce algas marinas resultando más activas para antioxidantes Acanthophora
spicifera, Laurencia sp., Ganonema farinosum, Dictyopteris undulata y Eisenia
arborea. Para la inhibición enzimática de acetilcolinesterasa el alga roja Laurencia
sp. presentó el mayor porcentaje de inhibición así como la menor concentración
efectiva media, con una constante de inhibición de 4.06. El trabajo mostró una
relación estrecha entre la inhibición de acetilcolinesterasa y el efecto antioxidante del
2,2-difenil-1-picrilhidracilo, sugiriendo que la actividad podría deberse a compuestos
derivados del floroglucinol.
Palabras clave: Alzheimer, acetilcolinesterasa, antioxidantes, algas marinas,
Florotaninos.

XV
ABSTRACT
Alzheimer’s disease is a progressive disorder that leads to memory deficits and
death. Currently there is no medication proven to cure or prevent this disease, being
the lack of knowledge a problem to understand the neuronal mechanisms underlying
the pathogenesis of the problem. Most studies have accepted the amyloid hypothesis,
in which the neurophatogenesis is believed to be caused by the accumulation of the
toxic amyloid- protein in the central nervous system. Dual binding site of
acetylcholinesterase inhibitors are promising for the treatment of Alzheimer’s disease.
These alleviate the cognitive deficits and modifying agents by inhibiting the -amiloid
peptide aggregation as well as control of acetylcholine levels through dual binding site
of the enzyme. Amyloid peptides also have been identified as a source of oxidative
stress, acquiring a state of free radical producing neurotoxic effects. This study
proposed the search of marine natural products with anti-alzheimer and antioxidant
potential activity. Antioxidant activity was determined using colorimetric assays: 2, 2-
diphenyl-1-picrylhydrazyl and the ferric reducing antioxidant power. Anti-Alzheimer’s
potential was determined using acetylcholinesterase enzyme inhibition assay. Twelve
seaweed were used, been more effective for antioxidant activity: Acanthophora
spicifera, Laurencia sp., Ganonema farinosum, Dictyopteris undulata and Eisenia
arborea. For enzyme inhibition of AChE, red alga Laurencia sp. showed the highest
inhibition and the lowest average effective concentration, with an inhibition constant of
4.06. This study demostrated a close relationship between inhibition of AChE and
antioxidant effect of 2, 2-diphenyl-1-picrylhydrazyl, suggesting that the activity could
be due to compounds derived from phloroglucinol.
Keywords: Alzheimer’s disease, acetylcholinesterase, anti-oxidants, marine algae,
phlorotannins.

1
1. INTRODUCCIÓN
El alzheimer es un desorden neurológico crónico progresivo e irreversible, que
depende tanto de factores genéticos como no genéticos (Gupta et al., 2011). Ésta
enfermedad se asocia con el envejecimiento, produciendo pérdida de la memoria,
deterioro intelectual así como trastornos de la personalidad y del comportamiento. Su
evolución es variable, con un diagnóstico de supervivencia entre 8-10 años
(Prasansukab y Tencomnao, 2013).
Esta enfermedad es la principal causa de demencia en adultos mayores, su
prevalencia aumenta con la edad siendo muy rara antes de los 40 años. Sin embargo
se presenta en el 3 % de mayores a 65 años, en el 20 % en los mayores de 75 años,
y en individuos con síndrome de Down que pasan los 50 años (Hardy y Higgins,
1992).
A medida que la enfermedad progresa, los pacientes con alzheimer pierden
gradualmente su capacidad de pensar y razonar con claridad, hacer juicios, resolver
problemas, comunicarse y cuidar de sí mismos. Los síntomas también incluyen
confusión, irritabilidad y agresión, cambios de humor, cambios de personalidad y la
conducta, problemas de atención y orientación espacial, problemas con el lenguaje, y
la pérdida de la memoria a largo plazo, afectando todo esto la vida diaria de una
persona. El alzheimer puede incluso conducir a la muerte de la persona afectada en
la fase final al causar desnutrición, la muerte cerebral y fallo de múltiples órganos
debido a la cantidad de las células nerviosas que han muerto (Brookmeyer et al.,
2007; Prasansukab y Tencomnao, 2013).
Durante mucho tiempo se creyó que el alzheimer y sus síntomas de demencia
eran una consecuencia inevitable debido al envejecimiento. Actualmente se han
hecho importantes adelantos en la investigación en cuanto a su etiología, diagnóstico
y tratamiento de la enfermedad, los cuales a su vez han servido para ampliar el
conocimiento de otras enfermedades neurodegenerativas relacionadas a la edad.
Inicialmente el alzheimer daña las neuronas de las regiones cerebrales implicadas en
el control de la memoria en el hipocampo (Muñoz-Pilar et al., 2005). Posteriormente

2
se extiende a la corteza cerebral, principalmente en las regiones responsables del
lenguaje y el razonamiento. Finalmente, se ven afectadas otras regiones cerebrales
atrofiándose, ocasionando que el paciente deje de responder al mundo exterior. En
cambio el envejecimiento normal no se asocia con la pérdida de un gran número de
células cerebrales (Pepys, 2006).
Después de varias décadas de estudio, el alzheimer es ahora considerado
como una enfermedad compleja que resulta de factores genéticos y fisiológicos, tales
como edad, sexo, antecedentes familiares y el síndrome de Down. Las causas de la
enfermedad aún se desconocen, así como los aspectos complejos de su bioquímica,
aunque sus características histopatológicas en el cerebro son bien conocidas (Merlini
y Belloti, 2003). Las características neuropatológicas de pacientes con EA son la
presencia de placas seniles y ovillos neurofibrilares en el cerebro que contienen la
proteína tau hiperfosforilada de forma anormal, y la pérdida de neuronas en el núcleo
basal de Meynert y el hipocampo. Las placas seniles son agregados de péptidos -
amiloides (A). Éstos péptidos son el principal constituyente de las placas seniles así
como agentes neurotóxicos importantes (Dinamarca et al., 2008; Dinamarca et al.,
2010).
Las proteínas asociadas a los depósitos amiloides son conocidas como
moléculas o proteínas chaperonas, en las cuales se incluyen la laminina,
apolipoproteína E y la acetilcolinesterasa (AChE). De hecho, se ha encontrado que la
AChE se emplea para interactuar con depósitos de A presentes en depósitos
difusos de pre-amiloides, interviniendo en la maduración de placas seniles y vasos
sanguíneos cerebrales. La mayor parte de la actividad AChE cortical presente en el
cerebro con alzheimer se asocia principalmente al núcleo amiloide de las placas
seniles en lugar de con el componente neurítico que se encuentra en la periferia
(Dinamarca et al., 2010).
La acetilcolinesterasa (AChE; EC. 3.1.1.7) es un miembro de la familia de las
enzimas / hidrolasas. Ésta enzima degrada el neurotransmisor excitador
acetilcolina (ACh) en la unión sináptica a una tasa catalítica extraordinariamente
rápida, con una segunda tasa constante casi tan rápida como una reacción por
difusión controlada. ACh es degradada a colina y acetato a través de un mecanismo

3
de hidrólisis, lo que resulta en la disminución de la transmisión en la sinapsis
nerviosa (Quinn, 1987; Sussman et al., 1991).
A su vez, la enzima AChE es una de las varias proteínas involucradas con los
depósitos de placas amiloides, asociándose predominantemente con la formación del
núcleo amiloide de las placas seniles maduras, depósitos de pre-amiloides y vasos
sanguíneos cerebrales correspondientes a cerebros con EA. La enzima es secretada
y se asocia con estructuras extracelulares, así como con la lámina basal sináptica en
la unión neuromuscular (Dinamarca et al., 2008).
Además de su actividad catalítica, la enzima AChE ejerce funciones
secundarias no colinérgicas relacionadas con su sitio de unión periférica sobre la
diferenciación, la adhesión celular, en la mediación de la transformación y deposición
de péptidos A. Se afirma que la AChE se une a través de su sitio periférico a la
forma no amiloide de la proteína -amiloide, actuando como una proteína chaperona
y a su vez como inducción de cambio conformacional a la forma amiloidogénica con
la formación de fibrillas de amiloide subsecuente (Rouleau, 2011).
Hay dos hipótesis para explicar la patología del alzheimer; la hipótesis
colinérgica sugiere que la disminución en la producción de ACh en la unión sináptica
contribuye a su aparición, mientras que la hipótesis amiloide sugiere que el desarrollo
de agregados tóxicos de péptidos -amiloides en el cerebro contribuye a su
progresión (Colombres et al., 2004; Fisher, 2011).
Una de las principales estrategias terapéuticas empleada contra el alzheimer
sintomático se basa en la hipótesis colinérgica empleando como blanco la AChE. Al
ser ésta una enzima de sustrato específico que degrada el neurotransmisor ACh en
la sinapsis nerviosa, se debe mantener un nivel óptimo de AChE en el cerebro para
un funcionamiento adecuado (Sussman et al., 1991). Sin embargo, el uso clínico de
inhibidores de la AChE a veces se limita debido a los efectos adversos en pacientes.
Por lo tanto, es necesario desarrollar nuevas terapias que sean más eficaces
(Seneviranthne, 2010).
Por otro lado, los péptidos -amiloides involucrados en la formación de las
placas seniles se han identificado como fuente de estrés oxidativo, adquiriendo un
estado de radical libre produciendo efectos neurotóxicos. Por lo que al inhibir el

4
efecto oxidante, mediante la captación de radicales libres, también se reducirán los
péptidos involucrados, lo que disminuirá la formación de las placas amiloides (Hardy
y Higgins, 1992). Evidencia reciente indica que el estrés oxidativo puede ocurrir
temprano en la progresión del alzheimer antes del desarrollo de placas seniles. La
interacción de mitocondrias anormales, transición de metales redox y estrés oxidativo
contribuyen como elementos para la generación de especies reactivas de oxígeno
(ERO). A su vez las ERO se producen como consecuencia de procesos metabólicos
aerobios. Por lo general éstos radicales libres son usualmente removidos o
inactivados por antioxidantes. La absoluta o relativa deficiencia de antioxidantes
puede llevar a un incremento de estrés oxidativo, el cual promueve el daño
neurológico (Yan et al., 1998; Burrit et al., 2002).
Las capacidades metabólicas y fisiológicas de los organismos marinos que les
permiten sobrevivir en un hábitat complejo les brindan un gran potencial para
producir metabolitos únicos que no se encuentran en ambientes terrestres; tanto las
algas como los invertebrados marinos sésiles son actualmente reconocidos como
una fuente potencial para el desarrollo de compuestos farmacéuticos (Moo y Freile,
2009). Actualmente los organismos más empleados son algas marinas, debido a su
efectividad en trabajos de inhibición enzimática para enfermedades neurológicas
(Seneviranthne, 2010). Especialmente son empleadas las algas pardas, debido a la
presencia de compuestos que actúan como buenos inhibidores de enzimas que
intervienen en enfermedades de origen neurológico (Barak et al., 1996; Buchholtz,
2006; Casey et al., 2007).
Las algas marinas desde tiempos antiguos se han considerado como alimento
tradicional en la dieta de pueblos asiáticos. Esto es debido a la composición que
presenta, siendo un alimento bajo en calorías con altas concentraciones de vitaminas
y minerales, así como con bajas concentraciones de ácidos grasos (Vidal et al.,
2006).
Las algas que se emplean como alimento se localizan en su mayoría en la
zona fótica, con una notable exposición a la radiación solar, lo que favorece a la
generación de radicales libres, lo que a su vez puede condicionar para la biosíntesis
de metabolitos secundarios con propiedades antioxidantes. Se sugiere que la

5
ausencia de daño oxidativo en los componentes estructurales de las algas y su
estabilidad frente a condiciones adversas se deben a la presencia de efectivos
antioxidantes (Yan et al., 1998; Burrit et al., 2002).
Diversos trabajos muestran una relación directa entre el consumo de algas
marinas y la prevención o tratamiento de patologías relacionadas con el estrés
oxidativo. Se ha demostrado que extractos de algas marinas tienen actividad
antioxidantes la cual se explica por diferentes mecanismos de acción en los que se
encuentran la capacidad captadora de radicales libres, la quelación de metales
activos desde el punto de vista redox, los mecanismos de donación y aceptación de
electrones, la capacidad de interrupción de la peroxidación lipídica y el incremento de
la actividad de enzimas antioxidantes (Vidal et al., 2006).
En el presente trabajo se dan a conocer los resultados de un estudio
destinado a evaluar la actividad antioxidante e inhibidora de la enzima AChE
empleando extractos etanólicos de algas marinas obtenidas en la Bahía de La Paz,
Baja California Sur, México, con el fin de inhibir procesos involucrados en el
desarrollo del alzheimer

6
2. ANTECEDENTES
2.1 Historia de la enfermedad de Alzheimer
En 1901 el doctor alemán Alois Alzheimer comenzó a trabajar con un paciente
con síntomas de déficit del lenguaje, paranoia, delirios y comportamiento agresivo.
Tras morir el paciente en 1906 se estudió el cerebro post-mortem encontrando
cambios arterioscleróticos, placas que denominaron seniles y fibrillas neurofibrilares
(Alzheimer, 1907). En 1910, Emil Kraepelin denominó “Enfermedad de Alzheimer”
para referirse a la causa de demencia senil. Sin embargo no fue hasta finales de la
década de los 50 que se incrementó el interés en las teorías e ideas generales
acerca del desarrollo en base a la biología molecular (Terry et al., 1964).
En 1963 se estudió el patrón histológico de las fibrillas y las placas; donde las
placas se interpretaron como núcleos fibrilares amiloideas, rodeados de axones y
dendritas que contenían los filamentos. Posteriormente en 1976, el Dr. Robert
Katzman revisó la frecuencia y mortalidad del alzheimer y destacó la necesidad
enfocar investigaciones en ésta área (Katzman, 1976).
Al observar que se presentaba una asociación de la degeneración de
neuronas y la pérdida de la memoria se marcó un hito, al haber un vínculo entre
síntomas clínicos y déficits colinérgicos específicos del cerebro, entablando el inicio
de la neuroquímica moderna. A partir de esto se buscaron nuevas respuestas para la
explicación del alzheimer. Esto dio origen al postulado de diferentes teorías, como la
“teoría colinérgica”, la cual establece la deficiencia de un neurotransmisor crítico, la
ACh, observando menor producción o una actividad amplificada en base a la AChE.
La disminución del nivel del neurotransmisor causa un deterioro de la
neurotransmisión colinérgica que lleva a la pérdida de las capacidades intelectuales
(Glenner y Wong, 1984; Arce et al., 2009).
Posteriormente los estudios histológicos en cerebros con alzheimer en
relación a la formación de placas, determinaron en 1984 que el bloque de
construcción del péptido amiloide se encontró conformado por la proteína -amiloide
la cuál forma las placas amiloides. El conjunto de ambas hipótesis postulan que la

7
AChE es responsable de varias acciones no catalíticas que incluyen la actividad pro-
agregación de A(Hatdy y Selkoe, 2002). Por lo que se estima que la enzima AChE
no sólo conduce a la descomposición aumentada del neurotransmisor ACh, si no que
a su vez participa en la formación de las placas amiloides. Promueve la agregación
de péptidos -amiloides mediante la formación de un complejo con las fibrillas de
crecimiento. Éstos complejos han demostrado ser más citotóxicos que las fibras
solas. Un dominio estructural de la AChE que promueve la formación de fibrillas de
péptido A ha sido identificado como el sitio periférico de la enzima (Singh, 2013).
Por ello, la inhibición de la AChE se presenta a modo de blanco específico como una
estrategia crítica para el tratamiento de EA (Inestrosa et al., 1996).
La AChE es un objetivo sensible para toxinas colinérgicas tanto naturales
como sintéticas. Entre los compuestos anti-AChE se han encontrado carbamatos e
inhibidores provenientes de glicoalcaloides derivados de plantas. Algunas microalgas
presentan toxinas altamente eficaces que bloquean el sitio activo de la AChE. A su
vez se ha encontrado que venenos como el de la serpiente mamba verde incluyen
péptidos neurotóxicos que bloquean la entrada del activo y los sitios periféricos de la
AChE (Matsunaga et al., 1989; Harel et al., 1995).
La inhibición de la AChE juega un papel crítico en la mejora de la
neurotransmisión colinérgica en el cerebro y la disminución de la formación de -
amiloide con el fin de proteger a las neuronas de la neurodegeneración. Se han
aislado anteriormente inhibidores de colinesterasas a partir de algas marinas
presentando evidencia de inhibición contra AChE y butiril-colinesterasa (BChE;
Kannan et al., 2014).
Inhibidores de la AChE, tales como; la rivastigmina, tacrina, donepezilo y
galantamina se han utilizado para el tratamiento moderado del alzheimer. Éstos
fármacos se consideran inhibidores reversibles de la AChE (Gupta et al., 2011).
Éstos compuestos alivian los déficits cognitivos actuando por inhibición de la
agregación del péptido -amiloide, a través de la unión tanto de los sitios aniónicos
catalíticos y periféricos. Éste sitio se denomina unión dual de la enzima AChE. El sitio
de unión dual de los inhibidores de AChE es prometedor para tratamientos de la
enfermedad de Alzheimer (Singh, 2013).

8
Estudios in vitro indicaron que la enzima AChE tiene la capacidad de aumentar
la formación de fibrillas amiloides y agregaciones -amiloides. Se ha demostrado que
cuando ésta se infunde estereotáxicamente en la región CA1 en ratas, da lugar a la
formación de nuevas placas (Dinamarca et al., 2008).
Dinamarca et al. (2008) pusieron a prueba un análogo de hiperforina (IDN
5706) la cual libera cargas de AChE tanto de las fibrillas neuronales así como del
análogo AChE-A en ratones transgénicos. La hiperforina fue capaz de prevenir las
alteraciones de la memoria espacial causadas por neurotoxicidad inducida por A.
Esto reúne evidencia que indica el importante papel de la AChE en la neurotoxicidad
de las placas y la búsqueda de nuevos compuestos que disminuyen la AChE y A.
Ésta interacción es un agente terapéutico esencial para combatir el alzheimer.
Rouleau et al. (2011) sintetizaron inhibidores de AChE, probando in vitro la
inhibición de diferentes colinesterasas (AChE y BChE) y su relación en la prevención
de agregación de A. Obteniendo resultados que sugieren que los compuestos
intervienen simultáneamente en los sitios catalíticos y periféricos de AChE,
interviniendo en ambos procesos (Muñoz-Pilar et al., 2005).
2.2 Inhibidores de origen marino
Las algas marinas son reconocidas como fuentes ricas en compuestos
estructuralmente diversos biológicamente activas con gran potencial farmacéutico y
biomédico. Investigaciones han revelado que las algas marinas producen
compuestos con diversas actividades biológicas tales como anticoagulante, antiviral,
antialérgica, anticancerígena y antioxidantes. Además hay estudios que proporcionan
información sobre las propiedades neuroprotectoras de las algas marinas. Muchas
especies han sido utilizadas en dietas, así como remedios tradicionales en los países
orientales, y más recientemente en Europa y América. Por lo tanto, las algas marinas
tienen un gran potencial para ser utilizadas en la neuroprotección (Zarros, 2009;
Pangestuti y Kim, 2011).
Actualmente, no hay productos naturales marinos comerciales en uso clínico
con aplicación inhibidora de la AChE. Dado el éxito presente en compuestos

9
derivados de metabolitos secundarios de origen marino como compuestos
anticolinesterásicos, pueden resultar efectivos en las interacciones de la selectividad
y potencia en tratamientos con AChE para la EA (Newman y Cragg, 2007).
Los metabolitos marinos varían mucho en cuanto a estructura, masa y
composición química. En la actualidad sólo se conocen siete diferentes clases de
metabolitos marinos con actividad anticolinesterasa; onchidal, marinoquinolina,
dihidromonofarsesilacetona, pseudozoantoxantina, monooxofarnsesilacetona,
sargachromenol y ácido sargaquinóico (Fig. 1) (Stoddar et al., 2014).
Figura 1. Metabolitos marinos inhibidores de acetilcolinesterasa; 1. Onchidal, 2.
Marinoquinolina, 3. Dihidromonofarsesilacetona, 4. Pseudozoantoxantina, 5.
Monooxofarnsesilacetona, 6. sargachromenol y 7. Ácido sargaquinóico

10
Para la obtención de metabolitos secundarios de organismos marinos, las
opciones más empleadas son algas marinas, debido a su efectividad en trabajos de
inhibición enzimática para enfermedades neurológicas (Seneviranthne, 2010). En los
trabajos empleando algas marinas para la elaboración de extractos como inhibidores
enzimáticos para problemas de índole neurológico, se encuentran pocos de algas
rojas, de las cuales se han aislado compuestos como el inositol (Khotimchenko y
Vas’kovsky, 2004), siendo un buen inhibidor enzimático que puede intervenir en
enfermedades de origen neurológico (Barak et al., 1996; Casey et al., 2007).
Sin embargo las algas pardas han demostrado ser los organismos que
presentan un mejor efecto. Esto se puede observar en investigaciones como la de
Kang et al. (2013) donde demostraron que Ecklonia cava regula la expresión y
actividad de -y -secretasa, reduciendo los niveles de A estabilizando a las células
que presentaban neurotoxicidad.
2.3 Inhibidores de AChE
Estudios recientes muestran que especies de algas marinas presentaron
actividad inhibitoria específica contra AChE. De las especies que han resaltado
podemos encontrar: Caulerpa racemosa, Codium capitatum, Ulva fasciata, Halimeda
cuneata, Amphiora ephedraea, Amphiora bowerbankii, Dictyota hamifusa, Hypnea
valentiae, Padina gymnospora, Ulva reticulata, Gracilaria edulis, Ecklonia stolonifera
y Ishige okamurae (Pangestuti y Kim, 2011).
Autores como Myung et al. (2005) informaron que compuestos obtenidos de
algas marinas, como el dieckol y el florofucofluoroeckol poseen propiedades para
mejorar la memoria y la actividad inhibitoria contra AChE. Además Yoon et al. (2009)
tras trabajar con 27 algas marinas de Corea, encontraron que los extractos de
Ecklonia stolonifera mostraron actividad inhibitoria significativa, siendo dos esteroles
y ocho florotaninos obtenidos de ésta alga parda; de los cuales los compuestos
eckol, dieckol, y floroeckol demostraron actividad inhibitoria dependiendo de la dosis
selecionada contra AChE.

11
Kannan et al. (2013) evaluaron la actividad de extractos crudos y fracciones de
Ecklonia maxima como inhibidores de la enzima AChE empleando. Ésta
investigación sugiere que E. maxima puede ser útil como alimento funcional
empleándose como ingrediente en dietas para el manejo de desórdenes
neurológicos.
2.4 Actividad antioxidante e inhibidora de AChE
Las ERO son entidades químicas que incluyen radicales libres de oxígeno,
como los radicales anión superóxido (O2-), radicales hidroxilo (OH), el óxido nítrico
(NO), peroxinitrito y también especies no radicales, tales como H2O2 y oxígeno (O2).
En los organismos vivos, diferentes formas metabólicas pueden dar lugar a diversas
ERO endógenas de la respiración aeróbica normal (Gulcin et al., 2003).
El estrés oxidativo se define como el estado en el cual el nivel de ERO
sobrepasa las defensas antioxidantes del organismo, como resultado de un
desequilibrio entre los sistemas de formación y eliminación de especies reactivas de
oxígeno, donde son implicadas en diferentes enfermedades degenerativas como
cáncer, diabetes, enfermedad de Parkinson y Alzheimer (Finkel y Holbrook, 2000).
El cerebro es un órgano particularmente vulnerable al estrés oxidativo,
relacionado con la hipótesis de que este proceso está involucrado en los eventos
neurodegenerativos y la muerte neuronal (Behl, 1997). Algunos estudios sugieren
que los suplementos dietéticos con antioxidantes y captadores de radicales libres
han demostrado beneficios en frenar ligeramente el deterioro cognitivo (Morris et al.,
2002; Stutchbury y Munch, 2005).
El uso de antioxidantes ha sido reconocido como un método efectivo para
minimizar el efecto tóxico y patológico asociados con el estrés oxidativo ocasionado
por A Los antioxidantes son un grupo de moléculas reconocidas por su capacidad
para neutralizar los radicales libres, los cuales son compuestos que se emplean
como una alternativa para combatir las deficiencias asociadas al estrés oxidativo
(Ra-Ahn et al., 2012). La enzima AChE está asociada con la superficie de la
membrana extracelular, y juega un papel importante como una salvaguardia de las
células cerebrales. Recientemente se demostró que las placas seniles parecen

12
inducir procesos inflamatorios en el que se liberan especies de radicales de oxígeno
(Vina et al., 2004; Stutchbury y Munch, 2005).
El péptido A se ha identificado como una fuente de estrés oxidativo en el
cerebro debido a que adquiere un estado de radical libre contribuyendo al efecto de
toxicidad. Éste péptido a su vez induce a la citotoxicidad debido a que presenta una
acumulación intracelular de especies reactivas de oxígeno (ERO), el cual da lugar a
la peroxidación de los lípidos de membrana y a la muerte celular (Chang-Ho et al.,
2009; Ra-Ahn et al., 2012).
Existen investigaciones que establecen una relación entre la actividad
antioxidante y la actividad inhibidora de enzimas. En el trabajo de Mata et al. (2007)
evaluaron la actividad antirradicalaria, antioxidante e inhibidora de la AChE; así
como, la capacidad inhibitoria de extractos etanólicos y acuosos de cinco hierbas
aromáticas, las cuales se utilizan en preparaciones alimenticias tradicionales; hinojo
(Foeniculum vulgare), menta (Mentha spicata y Mentha vulgare), romero
(Rosmarinus officinalis) y tomillo (Thymus serpyllum), encontrando una actividad
simultánea entre la inhibición de la enzima AChE y la actividad antioxidante.
Ra-Ahn et al. (2012) realizaron ensayos con extractos etanólicos y fracciones
del alga parda Eisenia bicyclis mostrándose efectivos para reducir la muerte celular
ocasionada por la toxicidad inducida por A. A su vez éstas fracciones resultaron
activas para inhibir generaciones intracelulares de ERO y iones de calcio, los cuales
intervienen en el bloqueo de sinapsis.
Choi et al. (2014) aislaron inhibidores de AChE y BChE de recursos naturales
marinos a partir de 12 algas de costas coreanas, así como contra radicales libres.
Eisenia bicyclis fue la especie más efectiva para ambas actividades biológicas.

13
3. JUSTIFICACIÓN
La demencia afecta del 5 al 8 % de la población mayor a los 60 años,
prevaleciendo en un 20 % en los mayores de 75 años, siendo responsable de un 50
a un 75 % la enfermedad de Alzheimer de estos problemas del deterioro cognitivo
progresivo. La tendencia demográfica del siglo XXI demuestra un crecimiento
poblacional de edades superiores a los 60 años. El índice de las enfermedades
crónico degenerativas aumenta rápidamente conforme aumenta la proporción de
personas de mayor edad (Muñoz-Chacón, 2003).
Enfermedades con mayor incidencia en estos grupos de edad como la
demencia aún no cuentan con medidas preventivas, generando a su vez una
demanda de los servicios sociales que los países desarrollados están enfrentando
actualmente con gran dificultad, presentando mayor problema los países en vías de
desarrollo (Schilling et al, 2008; Martínez-Gil, 2009). El alzheimer constituye un
importante problema de salud pública por sus repercusiones en los pacientes, sus
familias, el sistema de salud y la sociedad en general (Hardy y Higgins, 1992; Karran
et al., 2011). Los pacientes con demencia requieren cuidados extensivos a un costo
elevado, siendo aproximadamente de 57,000 dólares por año (Karran et al., 2011).
En la actualidad, el alzheimer afecta a más de 26.6 millones de personas en
todo el mundo y su prevalencia está aumentando drásticamente cada año. Con el
aumento demográfico de la población, se espera que para el año 2050, el número de
pacientes con alzheimer se cuadruplique a más de 106 millones en todo el mundo,
estimando que 1 de cada 85 personas estarán viviendo con ésta enfermedad, por lo
que por es necesario el desarrollo de terapias efectivas (Hardy y Selkoe, 2002).
La búsqueda de una cura o prevención del alzheimer ha sido intensiva en los
últimos años, abarcando tanto aspectos genéticos, como fisiopatológicos,
ambientales y farmacéuticos; el tratamiento clínico de estas enfermedades se basa
en la mayoría de los casos en la mejora de la estimulación en los receptores
pertinentes, ya sea aumentando los niveles del neurotransmisor endógeno o por el
uso de sustancias que tienen una respuesta agonista similar (Houghton y Howes,
2005). Actualmente los principales avances en el tratamiento de la enfermedad de
Alzheimer han sido el uso de inhibidores de la acetilcolinesterasa, como la

14
galantamina, la huperzina A, la fisostigmina y sus derivados para aumentar los
niveles de acetilcolina en lugar de la utilización de compuestos colinérgicos. El uso
de inhibidores de enzimas que causan la prevención del alzheimer se encuentra en
vías de desarrollo (Choi, 2007; Birks, 2012).
Los efectos adversos más comunes de estos fármacos incluyen náuseas y
vómitos, ambos de los cuales están vinculados a la presencia de neuronas
colinérgicas en exceso. Efectos adversos menos comunes, secundarios incluyen
bradicardia, calambres musculares, disminución del apetito y pérdida de peso, así
como el aumento de la producción de ácido gástrico. Por lo tanto, se necesitan
nuevas clases e inhibidores potenciales más eficaces de la AChE para minimizar
estos efectos secundarios (Gupta et al., 2011).
Un método empleado actualmente es la disminución de depósitos de A, los
cuales han dado frutos, sin embargo aún no se tiene una fórmula segura. Una
alternativa para esto sería la inhibición de enzimas involucradas en los procesos de
producción de las placas amiloides y/o A, evitando así que se produzcan en lugar
de lidiar directamente con la eliminación de éste producto. El uso de productos
naturales de origen marino puede ser una opción útil para la aplicación contra
enfermedades como el alzheimer, donde se obtendrán metabolitos secundarios
capaces de contrarrestar los productos de esta enfermedad.
Las algas marinas al igual que otras plantas fotosintéticas tienen antioxidantes
capaces de protegerlos de los efectos dañinos, incluyendo daños en el ADN,
proteínas y lípidos por la radiación ultravioleta o la oxidación por las especies
reactivas de oxígeno, ya que están expuestos a la luz solar y altas concentraciones
de oxígeno en el mar. Antioxidantes sintéticos tales como el hidroxianisol butilado y
terc butilhidroquinona son conocidos por estar disponibles comercialmente, sin
embargo hay estudios que informan que resultan cancerígenos. Por lo tanto las
investigaciones centran su atención en el desarrollo de alternativas más seguras de
antioxidantes de fuentes naturales, incluyendo algas marinas (Ramjee et al., 2010;
Ali-Hassan et al., 2013).
A pesar de aún no lograr tener la cura para la enfermedad, una alternativa
para este tipo de enfermedades es ofrecer una progresión más lenta al padecimiento

15
y por lo tanto la posibilidad de mantener por más tiempo y con una mayor calidad las
capacidades funcionales de los pacientes.
4. OBJETIVOS
4.1 Objetivo general
Obtener un producto natural de origen algal con potencial inhibidor contra la enzima
acetilcolinesterasa, así como con actividad antioxidante para evitar la formación de
placas -amiloides.
4.2 Objetivos particulares
Analizar el efecto inhibitorio de diferentes extractos y fracciones algales contra
la enzima AChE.
Probar el efecto de extractos y fracciones como secuestrante de radicales
libres y efecto reductor de un sistema.
Aislar e identificar fracciones activas con actividad inhibidora.
Analizar la constante de inhibición enzimática de las reacciones.
5. HIPÓTESIS
Al metabolizar estructuras polifenólicas, las algas presentarán compuestos
que intervengan tanto en la inhibición de la enzima acetilcolinesterasa como en la
eliminación de radicales libres.
6. MATERIAL Y MÉTODOS
6.1 Obtención de extractos
Para la realización de los experimentos se trabajó con extractos de algas
marinas de la colección del Laboratorio de Química de Algas Marinas en las

16
instalaciones del Centro Interdisciplinario de Ciencias Marinas. Estas algas fueron
previamente recolectadas por medio de buceo SCUBA en diferentes localidades en
la Bahía de La Paz. El material recolectado se lavó y se le retiró el exceso de sal,
siendo posteriormente trasladado a las instalaciones del CICIMAR, donde fueron
secadas al sol, molidas y almacenadas hasta el momento de su utilización. Un
organismo de cada alga debidamente preservado fue enviado para su identificación
taxonómica con ayuda del Dr. Rafael Riosmena Rodríguez en el laboratorio de
Botánica Marina en la Universidad Autónoma de Baja California Sur.
Se emplearon los extractos etanólicos de 12 especies de algas, diez algas
rojas: Pterisiphonia bippinata (07-003-40), Acanthophora spicifera (07-004-40),
Gracilaria vermiculophylla (07-011-40), Chondracanthus canaliculatus (07-010-40),
Laurencia sp. (13-011-40), Liagora californica (06-008-40), Ganonema farinosum (04-
022-40), Gelidium robustum (06-033-40), Corallina vancouveriensis (06-025-40) y
Corallina sp. (06-024-40); y dos algas pardas: Dictyopteris undulata (06-028-40) y
Eisenia arborea (13-021-40). Estos extractos crudos fueron obtenidos realizando
extracciones con 100 mL de etanol destilado (EtOH) por cada 20 g de muestra,
filtrando la mezcla para extraer el residuo en dos ocasiones más empleando las
mismas condiciones. Los extractos resultantes se concentraron a sequedad en
rotavapor a 40°C (± 2°C) y se transfirieron a viales previamente tarados y
etiquetados.
6.2 Fraccionamiento del extracto etanólico
Una vez identificado el extracto 13-021-40 (Eisenia arborea) como el de mayor
potencial antioxidante se sometió a un fraccionamiento en columna empleando sílica
gel de fase normal y un sistema de elución con diferentes disolventes en gradiente
de polaridad; diclorometano (100 %), diclorometano-etanol (8:2, 1:1, 2:8), etanol (100
%), etanol-agua (1:1) y agua 100 %. A partir de este procedimiento, los eluatos
obtenidos fueron analizados por medio de cromatografía de capa fina. De acuerdo a
la variación en metabolitos secundarios encontrados se unieron los eluatos
semejantes formando cuatro fracciones (13-021-40 F1, 13-021-40 F2, 13-021-40 F3

17
y 13-021-40 F4), los cuales fueron concentrados a sequedad en rotavapor a 40°C (±
2°C).
6.3 Pruebas de actividad
Se realizaron ensayos colorimétricos de actividad secuestrante de radicales de 2,2-
difenil-1-picrilhidracilo (DPPH; Sharma y Bhat, 2009), el poder antioxidante reductor
de hierro (FRAP; Firuzi et al., 2005) y la actividad inhibidora de acetilcolinesterasa
(AChE; Rhee et al., 2001).
6.3.1 Actividad secuestrante de radicales
Para el ensayo colorimétrico se utilizó una solución metanólica de DPPH al
0.02 % (Sharma y Bhat, 2009). A partir de extractos y fracciones se preparó una
solución stock de 5 mg/mL, y se diluyeron secuencialmente para tener soluciones de
5.0, 2.5, 1.25, 0.625, 0.312, 0.156 y 0.078 mg/mL. Un mililitro de la solución de
prueba se mezcló con 4 mL de la solución de DPPH. La mezcla se dejó en oscuridad
durante media hora y posteriormente se determinó la absorbancia de la mezcla a una
longitud de onda de 517 nm utilizando un espectrofotómetro Milton Roy Company
SPECTRONIC 20D. Todos los ensayos se realizaron por triplicado. Se preparó un
blanco para obtener el factor de corrección por color y se utilizó ácido ascórbico
como control positivo. Se realizaron curvas de dosis-efecto obteniendo los
porcentajes de reducción del DPPH así como la EC50. Para ello se empleó la fórmula:
(𝐀𝐁 − 𝐀𝐌)
𝐀𝐁 𝐗 𝟏𝟎𝟎
Dónde:
AB: Absorbancia del blanco
AM: Absorbancia de la muestra corregida (AM=A-FC)
A: Absorbancia
FC: Factor de corrección de color

18
6.3.2 Poder de reducción antioxidante del hierro (FRAP)
Se emplearon los extractos etanólicos que mostraron mayor actividad en el
ensayo frente al DPPH, entre ellos los extractos de Acanthophora spicifera (07-004-
40), Laurencia sp. (13-011-40), Ganonema farinosum (04-022-40), Dictyopteris
undulata (06-028-40) y Eisenia arboea (13-021-40). A partir de cada extracto se
preparó una solución stock de 10 mg/mL utilizando etanol destilado como disolvente.
El ensayo se realizó en placas de microdilución de 96 celdas por adición de cada una
de las soluciones a probar y la solución de trabajo FRAP 2,4,6-tripiridil-s-triazina
(TPTZ) 10 mM en HCl 40 mM; FeCl3 20 mM y de buffer acetato (300 mM a un pH
3.6) preparada en fresco. La absorbancia fue leída a 595 nm después de 20
minutos, en un analizador semiautomático (Multiskan Spectrum, Thermo Scientific,
Finlandia). Las muestras y estándares se realizaron por cuadriplicado. Se utilizó una
curva de calibración de FeSO4 7H2O, y las actividades de los extractos de algas se
expresaron como mM de equivalentes de Fe (II) por gramo de muestra. Para la
determinación de los mili equivalentes se empleó la ecuación de la recta X= [(y-b)/m],
considerando que se partió de una muestra de 10 mg, siendo el FRAP expresado por
gramo de extracto.
6.3.3 Inhibición de la enzima acetilcolinesterasa
Se emplearon los extractos etanólicos que mostraron mayor actividad en el
ensayo frente al DPPH, entre ellos los extractos de A. spicifera (07-004-40),
Laurencia sp. (13-011-40), G. farinosum (04-022-40), D. undulata (06-028-40), E.
arborea (13-021-40), así como las fracciones obtenidas a partir del extracto etanólico
de E. arborea (13-021-40 F1, 13-021-40 F2, 13-021-40 F3 y 13-021-40 F4). La
actividad AChE se midió por el método de Ellman con modificaciones basadas en
Rhee et al. (2001). Para ello se prepararon soluciones las cuales se mencionan a
continuación: buffer A: 50 mM Tris.HCl, pH 8; buffer B: 50 mM Tris-HCl, pH 8, con
contenido de 0.1 % de suero de albúmina bovina (BSA); buffer C: 50 mM Tris.HCl,
pH 8, conteniendo NaCl al 0.1 M y MgCl2 6H2O al 0.02 M. Para el experimento se
empleó la enzima acetilcolinesterasa de anguila eléctrica (Tipo VI-s, en polvo
liofilizado, 292 U/mg sólido, 394 U/mg proteína) la cual se adquirió de Sigma Aldrich.

19
La enzima liofilizada se disolvió en buffer A para preparar una solución stock de 1000
U/ml y posteriormente se diluyó en buffer B para tener la cantidad de 0.22 U/ml para
el ensayo en microplaca. Como sustrato se utilizó una solución de acetilcolina iodada
15 mM diluida en agua destilada. El reactivo de Ellman; 5,5’-dithiobis-(2-ácido
nitrobenzóico) fue adquirido de Sigma Aldrich. Para el ensayo se utilizó 3 mM
disuelto en el buffer A.
Para la medición de la actividad AChE se emplearon lecturas de placas de 96
celdas. Para realizar la reacción se mezclaron 25 L de 15 mM del sustrato
previamente diluida en buffer A, 125 L del reactivo de Ellman en buffer C (Tris-HCL
al 0.1 M con NaCl y 0.02 M MgCl2 6H2O al 3 mM), 50 L de buffer B (Tris-HCl
conteniendo 0.01 % de suero de albúmina bovina), 25 L de extracto de alga marina
empleado (5, 2.5, 1.25, 0.625, 0.312, 0.156 y 0.078 mg/mL). Una vez mezcladas se
agregaron 25 L de la enzima AChE a una concentración de 0.22 U/mL y se
realizaron las lecturas cada 13 segundos durante ocho ocasiones empleando una
longitud de onda de 405 nm. El porcentaje de inhibición se calculó en base la fórmula
empleada.
% 𝑖𝑛ℎ𝑖𝑏𝑖𝑐𝑖ó𝑛 =𝐴𝑏𝑠𝑜𝑟𝑏𝑎𝑛𝑐𝑖𝑎 𝑐𝑜𝑛𝑡𝑟𝑜𝑙−𝐴𝑏𝑠𝑜𝑟𝑏𝑎𝑛𝑐𝑖𝑎 𝑑𝑒 𝑚𝑢𝑒𝑠𝑡𝑟𝑎
𝐴𝑏𝑠𝑜𝑟𝑏𝑎𝑛𝑐𝑖𝑎 𝑐𝑜𝑛𝑡𝑟𝑜𝑙 X100
6.4 Determinación de cinética enzimática
Para la determinación de los parámetros cinéticos de Km (constante de
Michaelis) al sólo emplear un sustrato para el ensayo de actividad de AChE se
emplearon concentraciones de sustrato partiendo de supuestos conocidos de
manera bibliográfica, y se determinó la velocidad de la reacción empleando dichas
concentraciones. Para la obtención de estos parámetros se aplicó el método de
Lineweaver-Burk determinando el valor de Vmax y V0 por medio de regresión lineal
mediante Microsoft Excel 2010. El método de Lineweaver-Burk (dobles recíprocos)
interviene la ecuación de Michaelis Menten: 1/V0= Km/Vmáx. Ésta ecuación representa
la unión de la línea recta y=bx+a.

20
Para la determinación de la constante de inhibición (Ki) mediante regresión
lineal se aplicó la representación gráfica secundaria de la pendiente de cada recta
primaria en función de las concentraciones de cada extracto empleado y se utilizó la
ecuación para conversión de EC50 a Ki para determinación de actividad enzimática.
𝐾𝑖 =𝐸𝐶50
(𝑆/𝐾𝑚 + 1)
6.5 Cálculos y análisis estadísticos
Para las comparaciones cuantitativas de las diferentes actividades biológicas
se realizaron matrices de datos, cálculos y gráficas empleando el programa Microsoft
Excel 2013. Para los análisis estadísticos se empleó el Software STATISTICA 8, para
ver si existían diferencias significativas por cada prueba de actividad biológica. Para
el caso de los porcentajes de inhibición de DPPH y AChE se realizaron pruebas de
ANDEVA de dos vías para observar diferencias por tratamientos entre las diferentes
algas empleadas y las diferentes concentraciones. Para el caso de FRAP se realizó
una ANDEVA de una vía para ver diferencias entre grupos. Finalmente se realizó una
matriz de correlación de Pearson para observar la relación presente entre las
actividades antioxidantes DPPH y FRAP y la actividad inhibitoria de AChE.
6.6 Identificación estructural por análisis espectroscópico
Las fracciones de E. arborea 13-021-40 F1, 13-021-40 F2, 13-021-40 F3 y 13-
021-40 F4 fueron analizadas con el fin de observar los grupos funcionales presentes.
Para ello se empleó un análisis de espectroscopia de infrarrojo utilizando un
espectrómetro FTIR-ATR Modelo Spectrum TwoTM Perkin-Elmer y se leyó en un
espectro entre 4,000 y 500 cm-1 realizando una corrección por medio del Software
Spectrum de Perkin-Elmer. Los resultados obtenidos se analizaron y compararon con
bases de datos de espectros infrarrojos, así como con literatura especializada.
21
7. RESULTADOS
7.1 Actividad antioxidante
7.1.1 DPPH
Como se muestra en la Tabla 1, todos los extractos crudos sometidos al
ensayo presentaron actividad inhibitoria de DPPH. Sólo se consideraron los cinco
extractos más activos que alcanzaron un mayor porcentaje de inhibición, para
continuar con las siguientes pruebas, siendo las algas A. spicifera (90.28% ± 1.79),
Laurencia sp. (53.57% ± 3.87), G. farinosum (63.36% ± 1.20), D. undulata (94.69% ±
0.30) y E. arborea (94.69% ± 0.26). Las algas pardas mostraron un mayor efecto
inhibidor, observándose una diferencia significativa con respecto a la actividad de las
algas rojas estudiadas (ANOVA, F=14.326 P=0.000000).
Entre los extractos evaluados, el extracto etanólico del alga parda E. arborea
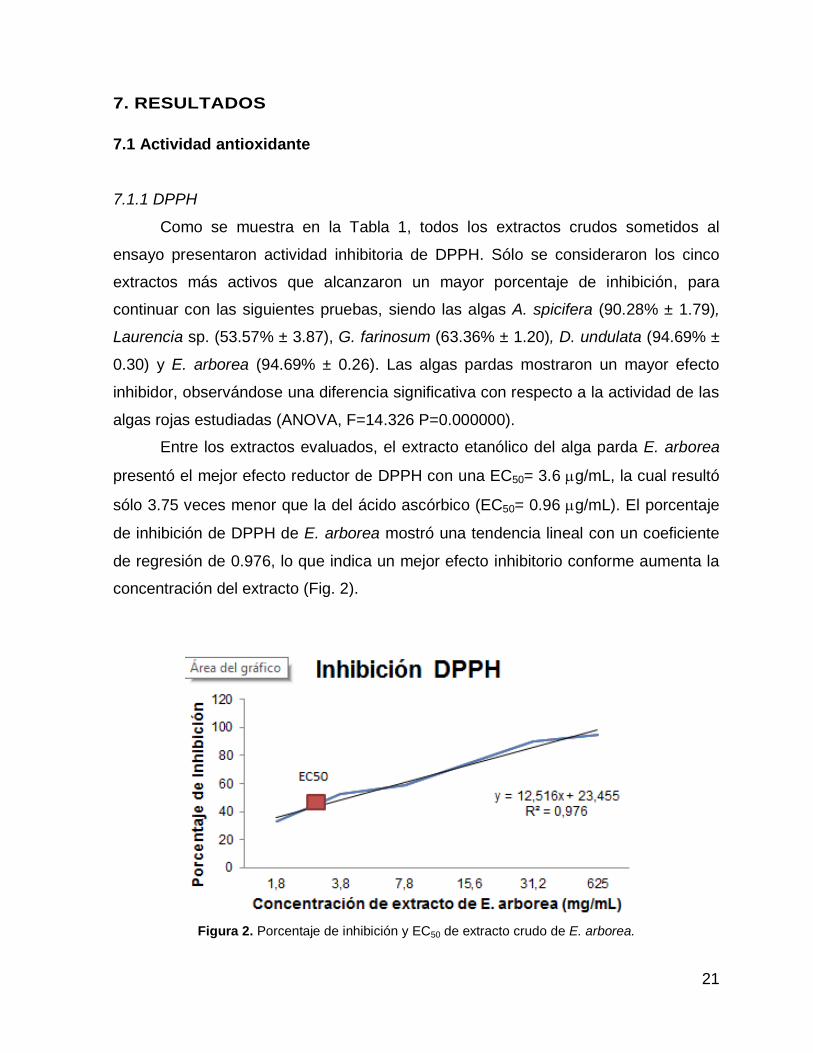
presentó el mejor efecto reductor de DPPH con una EC50= 3.6 g/mL, la cual resultó
sólo 3.75 veces menor que la del ácido ascórbico (EC50= 0.96 g/mL). El porcentaje
de inhibición de DPPH de E. arborea mostró una tendencia lineal con un coeficiente
de regresión de 0.976, lo que indica un mejor efecto inhibitorio conforme aumenta la
concentración del extracto (Fig. 2).
Figura 2. Porcentaje de inhibición y EC50 de extracto crudo de E. arborea.
22
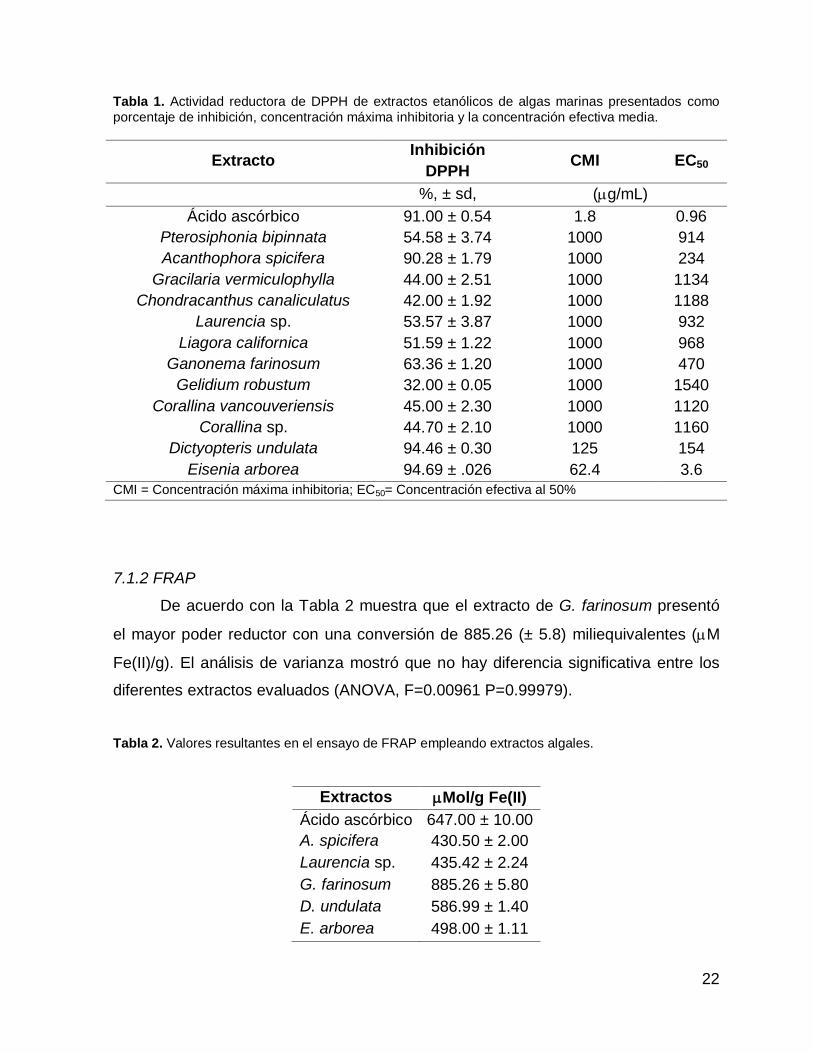
Tabla 1. Actividad reductora de DPPH de extractos etanólicos de algas marinas presentados como
porcentaje de inhibición, concentración máxima inhibitoria y la concentración efectiva media.
Extracto Inhibición
DPPH CMI EC50
%, ± sd, (g/mL)
Ácido ascórbico 91.00 ± 0.54 1.8 0.96
Pterosiphonia bipinnata 54.58 ± 3.74 1000 914
Acanthophora spicifera 90.28 ± 1.79 1000 234
Gracilaria vermiculophylla 44.00 ± 2.51 1000 1134
Chondracanthus canaliculatus 42.00 ± 1.92 1000 1188
Laurencia sp. 53.57 ± 3.87 1000 932
Liagora californica 51.59 ± 1.22 1000 968
Ganonema farinosum 63.36 ± 1.20 1000 470
Gelidium robustum 32.00 ± 0.05 1000 1540
Corallina vancouveriensis 45.00 ± 2.30 1000 1120
Corallina sp. 44.70 ± 2.10 1000 1160
Dictyopteris undulata 94.46 ± 0.30 125 154
Eisenia arborea 94.69 ± .026 62.4 3.6
CMI = Concentración máxima inhibitoria; EC50= Concentración efectiva al 50%
7.1.2 FRAP
De acuerdo con la Tabla 2 muestra que el extracto de G. farinosum presentó
el mayor poder reductor con una conversión de 885.26 (± 5.8) miliequivalentes (M
Fe(II)/g). El análisis de varianza mostró que no hay diferencia significativa entre los
diferentes extractos evaluados (ANOVA, F=0.00961 P=0.99979).
Tabla 2. Valores resultantes en el ensayo de FRAP empleando extractos algales.
Extractos Mol/g Fe(II)
Ácido ascórbico 647.00 ± 10.00
A. spicifera 430.50 ± 2.00
Laurencia sp. 435.42 ± 2.24
G. farinosum 885.26 ± 5.80
D. undulata 586.99 ± 1.40
E. arborea 498.00 ± 1.11
23
7.2 Inhibición de acetilcolinesterasa
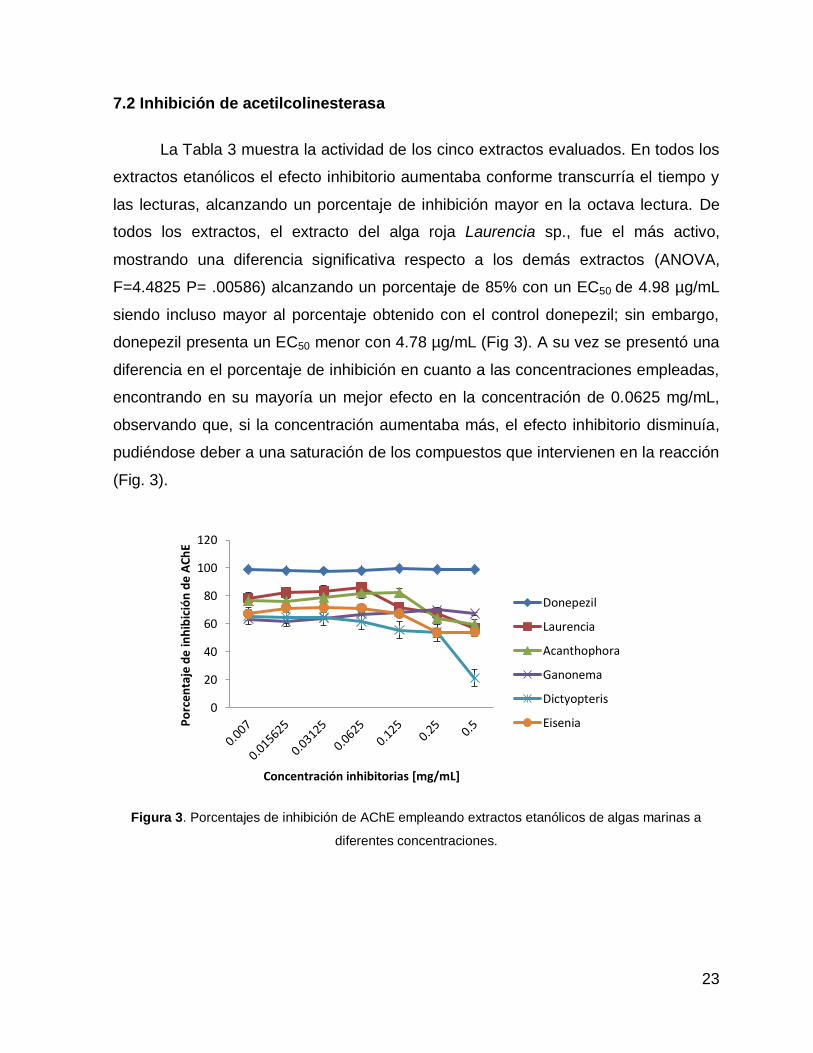
La Tabla 3 muestra la actividad de los cinco extractos evaluados. En todos los
extractos etanólicos el efecto inhibitorio aumentaba conforme transcurría el tiempo y
las lecturas, alcanzando un porcentaje de inhibición mayor en la octava lectura. De
todos los extractos, el extracto del alga roja Laurencia sp., fue el más activo,
mostrando una diferencia significativa respecto a los demás extractos (ANOVA,
F=4.4825 P= .00586) alcanzando un porcentaje de 85% con un EC50 de 4.98 µg/mL
siendo incluso mayor al porcentaje obtenido con el control donepezil; sin embargo,
donepezil presenta un EC50 menor con 4.78 µg/mL (Fig 3). A su vez se presentó una
diferencia en el porcentaje de inhibición en cuanto a las concentraciones empleadas,
encontrando en su mayoría un mejor efecto en la concentración de 0.0625 mg/mL,
observando que, si la concentración aumentaba más, el efecto inhibitorio disminuía,
pudiéndose deber a una saturación de los compuestos que intervienen en la reacción
(Fig. 3).
Figura 3. Porcentajes de inhibición de AChE empleando extractos etanólicos de algas marinas a
diferentes concentraciones.
0
20
40
60
80
100
120
Po
rce
nta
je d
e in
hib
ició
n d
e A
Ch
E
Concentración inhibitorias [mg/mL]
Donepezil
Laurencia
Acanthophora
Ganonema
Dictyopteris
Eisenia
24
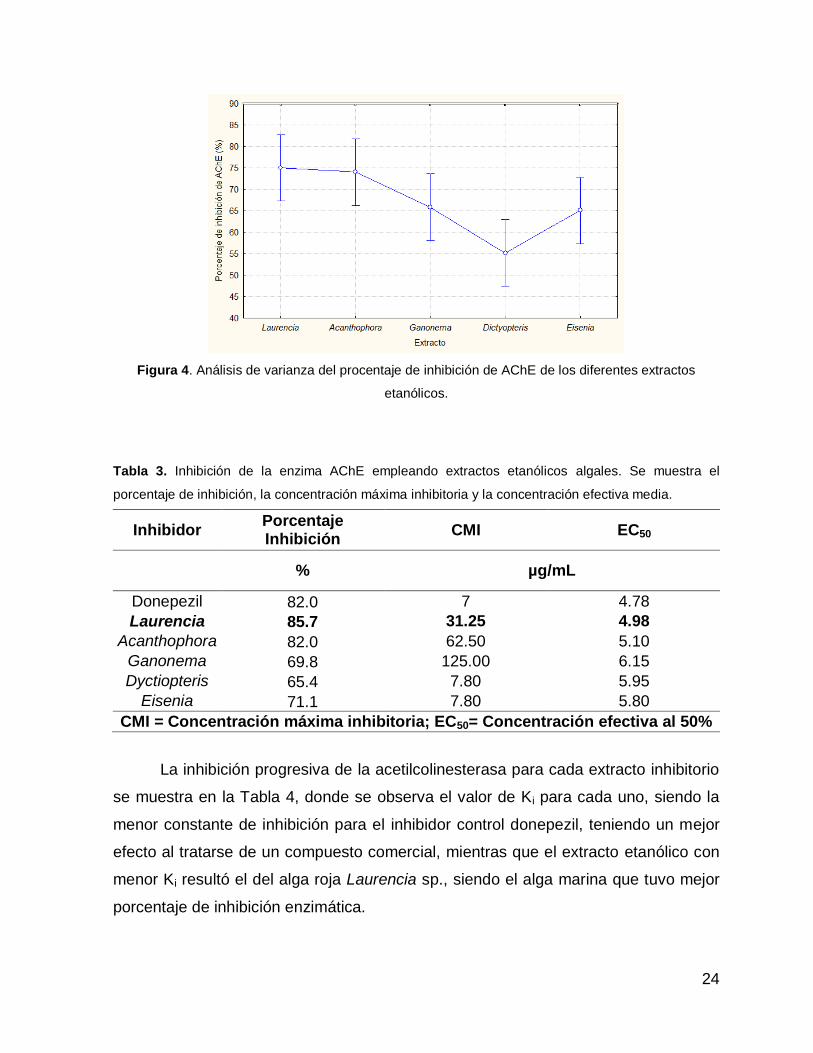
Figura 4. Análisis de varianza del procentaje de inhibición de AChE de los diferentes extractos
etanólicos.
Tabla 3. Inhibición de la enzima AChE empleando extractos etanólicos algales. Se muestra el
porcentaje de inhibición, la concentración máxima inhibitoria y la concentración efectiva media.
Inhibidor Porcentaje Inhibición
CMI EC50
% µg/mL
Donepezil 82.0 7 4.78
Laurencia 85.7 31.25 4.98
Acanthophora 82.0 62.50 5.10
Ganonema 69.8 125.00 6.15
Dyctiopteris 65.4 7.80 5.95
Eisenia 71.1 7.80 5.80
CMI = Concentración máxima inhibitoria; EC50= Concentración efectiva al 50%
La inhibición progresiva de la acetilcolinesterasa para cada extracto inhibitorio
se muestra en la Tabla 4, donde se observa el valor de Ki para cada uno, siendo la
menor constante de inhibición para el inhibidor control donepezil, teniendo un mejor
efecto al tratarse de un compuesto comercial, mientras que el extracto etanólico con
menor Ki resultó el del alga roja Laurencia sp., siendo el alga marina que tuvo mejor
porcentaje de inhibición enzimática.

25
Tabla 4. Cinética enzimática para la constante de inhibición (Ki) de los extractos y fracciones
empleadas de algas marinas contra AChE.
Inhibidor Ki (M)
Donepezil 3,18
Laurencia sp. 4,06
A. spicifera 4,18
G. faninosum 5,41
D. undulata 5,18
E. arborea 4,92
7.3 Relación entre antioxidantes y actividad anticolinesterasa
El análisis de correlación de Pearson se realizó para investigar la relación
entre la actividad antioxidante, así como la actividad inhibidora de AChE. Los
resultados demostraron una fuerte correlación positiva entre los diferentes ensayos
biológicos. La actividad anti AChE presentó un coeficiente de regresión de 0.92
respecto a la actividad DPPH y un coeficiente de regresión de 0.60 respecto a FRAP.
Mientras que los ensayos de actividad antioxidante presentaron un mayor coeficiente
de regresión entre sí, explicando la relación entre DPPH y FRAP.
7.4 Análisis de fracciones obtenidas de Eisenia arborea
7.4.1 DPPH
Al mostrar el alga parda E. arborea el mejor efecto captador de radicales libres
entre los diferentes extractos se consideró para su fraccionamiento, obteniendo así
cuatro fracciones las cuales se denominaron 13-021-40 F1, 13-021-40 F2, 13-021-40
F3 y 13-021-40 F4. La fracción 2 (13-021-40 F2) presentó un mayor porcentaje de
inhibición de DPPH con 94.64 % ± 0.24 con un EC50 de 16 g/mL (Tabla 5). La
fracción 13-021-40 F2 también mostró una tendencia lineal con un R2 de 0.90 (Fig.
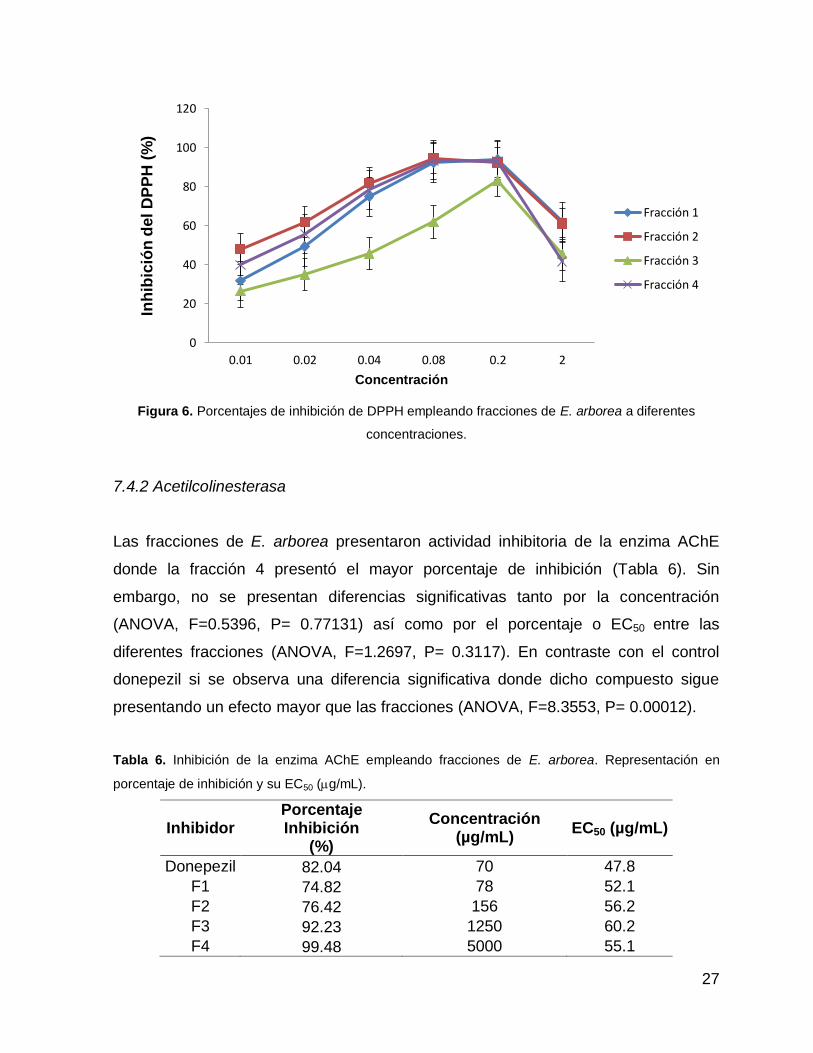
5). Sin embargo, los estadísticos indicaron que la fracción 3 se muestra diferencias
significativas con las demás fracciones, siendo la que presenta menor actividad
(ANOVA, F=18.094 P= .00003) (Fig. 6). A su vez se observaron diferencias
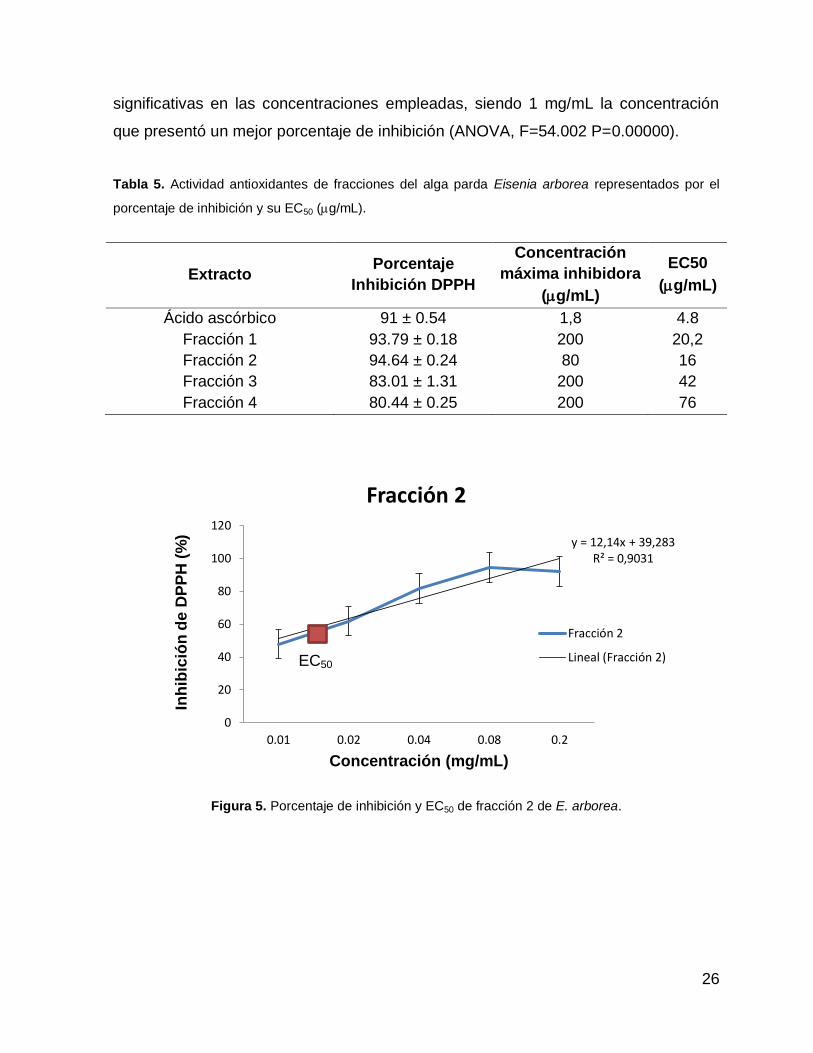
26
significativas en las concentraciones empleadas, siendo 1 mg/mL la concentración
que presentó un mejor porcentaje de inhibición (ANOVA, F=54.002 P=0.00000).
Tabla 5. Actividad antioxidantes de fracciones del alga parda Eisenia arborea representados por el
porcentaje de inhibición y su EC50 (g/mL).
Extracto Porcentaje
Inhibición DPPH
Concentración
máxima inhibidora
(g/mL)
EC50
(g/mL)
Ácido ascórbico 91 ± 0.54 1,8 4.8
Fracción 1 93.79 ± 0.18 200 20,2
Fracción 2 94.64 ± 0.24 80 16
Fracción 3 83.01 ± 1.31 200 42
Fracción 4 80.44 ± 0.25 200 76
Figura 5. Porcentaje de inhibición y EC50 de fracción 2 de E. arborea.
y = 12,14x + 39,283 R² = 0,9031
0
20
40
60
80
100
120
0.01 0.02 0.04 0.08 0.2
Inh
ibic
ión
de D
PP
H (
%)
Concentración (mg/mL)
Fracción 2
Fracción 2
Lineal (Fracción 2)EC50
27
Figura 6. Porcentajes de inhibición de DPPH empleando fracciones de E. arborea a diferentes
concentraciones.
7.4.2 Acetilcolinesterasa
Las fracciones de E. arborea presentaron actividad inhibitoria de la enzima AChE
donde la fracción 4 presentó el mayor porcentaje de inhibición (Tabla 6). Sin
embargo, no se presentan diferencias significativas tanto por la concentración
(ANOVA, F=0.5396, P= 0.77131) así como por el porcentaje o EC50 entre las
diferentes fracciones (ANOVA, F=1.2697, P= 0.3117). En contraste con el control
donepezil si se observa una diferencia significativa donde dicho compuesto sigue
presentando un efecto mayor que las fracciones (ANOVA, F=8.3553, P= 0.00012).
Tabla 6. Inhibición de la enzima AChE empleando fracciones de E. arborea. Representación en
porcentaje de inhibición y su EC50 (g/mL).
Inhibidor Porcentaje Inhibición
(%)
Concentración (µg/mL)
EC50 (µg/mL)
Donepezil 82.04 70 47.8
F1 74.82 78 52.1
F2 76.42 156 56.2
F3 92.23 1250 60.2
F4 99.48 5000 55.1
0
20
40
60
80
100
120
0.01 0.02 0.04 0.08 0.2 2
Inh
ibic
ión
del D
PP
H (
%)
Concentración
Fracción 1
Fracción 2
Fracción 3
Fracción 4

28
Respecto a los valores de la constante de inhibición, la fracción 4 de Eisenia
arborea presentó el menor valor de Ki coincidiendo en que ésta fracción presentó el
valor más alto del efecto inhibidor entre las fracciones obtenidas. El valor de Ki de
donepezil se presenta menor que el resto de las fracciones, sin embargo, éstos
valores no difieren mucho uno del otro siendo valores muy cercanos incluyendo el
inhibidor control (Tabla 7).
Tabla 7. Cinética enzimática para la constante de inhibición (Ki) de los extractos y fracciones
empleadas de algas marinas contra AChE.
Inhibidor Ki (M)
Donepezil 3,18
EaF1 4,33
EaF2 4,78
EaF3 5,24
EaF4 3,91
7.5 Relación en el efecto inhibidor de fracciones
El análisis de correlación de Pearson para las fracciones de E. arborea sugiere
una correspondencia entre el efecto antioxidante del DPPH y la actividad inhibidora
de la enzima acetilcolinesterasa, presentando un valor de R de 0.94. Esto indica una
relación positiva entre amas actividades, lo que sugiere que en conjunto tiene el
potencial para intervenir en procesos anti-alzheimer interviniendo en la formación de
las placas amiloides o en los procesos involucrados en la formación de éstas.
7.6 Caracterización de fracciones por análisis espectroscópico de IR
De las señales observadas en los espectros infrarrojos de las cuatro
fracciones 13-021-40 F1, 13-021-40 F2, 13-021-40 F3 y 13-021-40 F4 se pueden
apreciar en todas 13 bandas diagnósticas (Fig. 7), sin embargo, la fracción 4 muestra
una mayor intensidad en la mayoría de las bandas, pudiendo indicar una alta
concentración de compuestos. De las bandas diagnósticas se distinguen 4 bandas
de absorción principales: la banda centrada en 3380 cm-1 que corresponde a un

29
grupo hidroxilo, mientras que las bandas 1630 y 1480 a enlaces de grupo carboxilo.
En la zona denominada huella dactilar ubicada entre los 600 y 1400 cm-1 se puede
distinguir una banda de 1029 cm-1 indicando la presencia de un enlace C-O.
El análisis de espectro IR al mostrar bandas cercanas a 3400 cm-1 (OH), así como
1650-1450 cm-1 (fenil), 1200 y 1050 cm-1 (éter) y 820 (1,3,5-fenil sustituido) indica
resultados que sugieren que las fracciones de E. arborea presentan derivados de
floroglucinol. Sin embargo, entre las otras señales no identificadas pueden indicar la
presencia de alcanos, alcoholes, ésteres, éteres o ácidos carboxílicos. Los espectros
infrarrojos mostraron semejanza entre las cuatro fracciones, encontrándose
compuestos similares entre ellos, presentando las 13 bandas de absorción, así como
presentando señales propias de un compuesto orgánico indicadas por las bandas de
absorción principales.
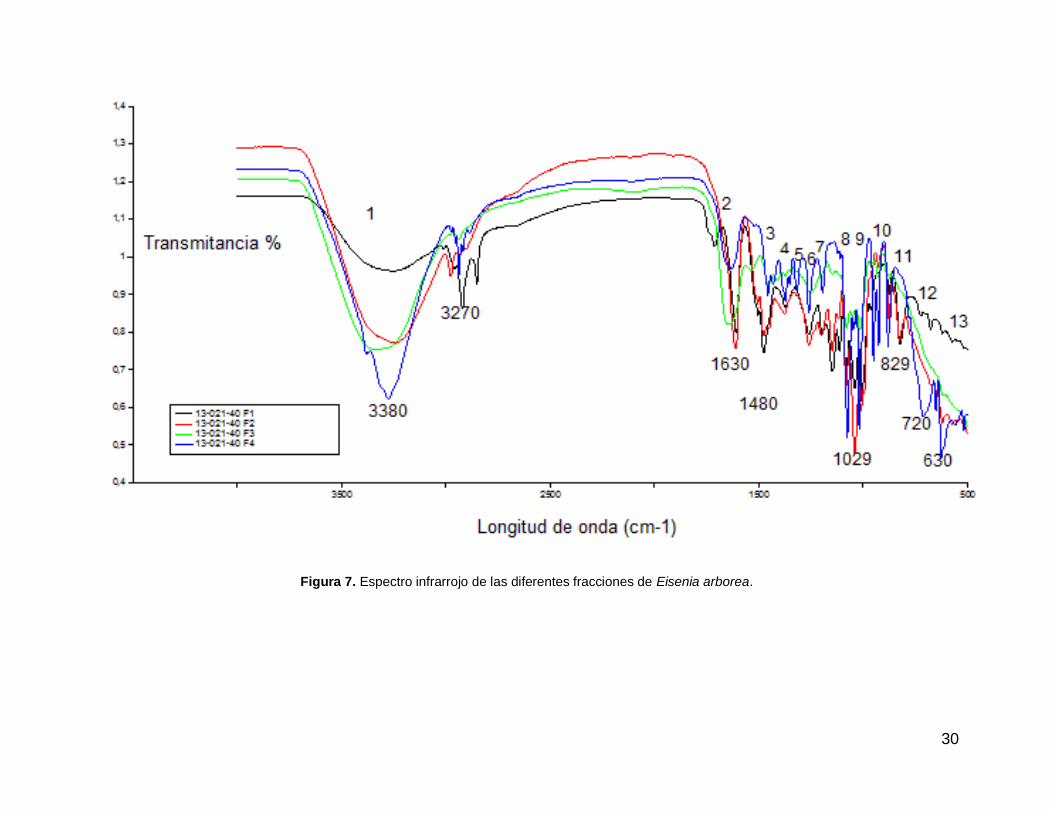
30
Figura 7. Espectro infrarrojo de las diferentes fracciones de Eisenia arborea.

31
8. DISCUSIÓN
Las pruebas de actividad antioxidante empleando el modelo de ensayo de
DPPH mostraron que el alga parda Eisenia arborea tuvo el mejor efecto secuestrante
de radicales libres, con un porcentaje de reducción del 94.69 % y una EC50 de 3.6
g/mL (Tabla 1), una alta actividad en contraste con trabajos reportados de algas
pardas del mismo orden, tal es el caso de Laminaria sp. (EC50 de 860 g/mL) y
Undaria sp. (EC50 de 420 g/mL) (Ismail y Tan, 2002).
El efecto antioxidante de algas pardas de la familia Lessoniaceae es
reconocido y respaldado por varias investigaciones, sobresaliendo los géneros
Ecklonia y Eisenia. Como el trabajo de Senevirathne et al. (2006) donde obtuvieron
un efecto inhibitorio contra DPPH, presentando el alga parda Ecklonia cava un valor
EC50 de 20 g/mL. De la misma familia se ha empleado el alga Eisenia bicyclis, de la
cual extractos y fracciones resultaron muy activos con una EC50 de 2.48 g/mL, que
también fueron efectivos para reducir la muerte celular ocasionada por toxicidad
inducida por A; esto mismo resultó activo para inhibir tanto generaciones
intracelulares de radicales libres de EROS, así como la formación de iones de calcio
(Ra-Ahn et al., 2012; Choi et al., 2014).
A pesar de no haber presentado Eisenia arborea el mejor efecto inhibidor de
AChE en el presente trabajo (EC50 de 5.80 g/mL) otros autores afirman que algas
pardas de la misma familia Lessoniaceae presentan actividad que interpretan como
un importante potencial; Kannan et al. (2013) evaluaron la actividad del alga Eclonia
maxima como inhibidores de AChE obteniendo valores de EC50 de 62.61 a 150.8
g/mL. Los autores sugieren que con éstos resultados el alga puede ser útil como
alimento funcional o como ingrediente para dietas en el manejo de desórdenes
neurológicos. El alga Eisenia bicyclis también se ha empleado en otros trabajos
obteniendo un porcentaje de inhibición del AChE del 68% utilizando una
concentración de 25 g/mL (Choi et al., 2014).
Yoon et al. (2008) realizaron análisis de 27 algas para probar su actividad
contra acetilcolinesterasa, encontrando a Ecklonia stolinifera como la más activa,

32
donde obtuvieron diferentes compuestos derivados de ésta, siendo los más activos
eckstolonol (42.66 g/mL), eckol (20.56 g/mL), florofucofuroeckol-A (4.89 g/mL),
dieckol (17.11 g/mL), 2- floroeckol (38.13 g/mL) y 7-floroeckol (21.11 g/mL).
La actividad antioxidante presente en algas pardas puede deberse a la
presencia de diferentes compuestos como fucovantinas, aminoácidos, carotenoides,
polisacáridos, terpenos, polifenoles, florotaninos y flavonoides, los cuales han sido
reportados anteriormente (Ye et al., 2008; Cornish y Garbary, 2010). Algunos de ellos
han sido aislados de E. arborea como los florotaninos 6.6’-bieckol y el 6,8’-bieckol,
que son derivados del floroglucinol pudiendo estar presenten en el extracto etanólico
(Sugiura et al., 2008).
A pesar de no tener un efecto antioxidante tan eficaz como las algas pardas,
las algas rojas también han sido otro grupo estudiado por su actividad y composición
estructural. Trabajos como el de Vidal et al., (2006) establecen que las algas marinas
rojas son efectivas en la captación de radicales libres si se emplean altas
concentraciones de extracto, habiendo una relación directa entre la concentración y
la actividad biológica, tal es el caso del alga Bryothamnion triquetrum la cual presenta
un valor de EC50 de 4,660 g/mL. Esto coincide con nuestra investigación en donde
las algas rojas que mostraron un mejor efecto antioxidante presentan igual un EC50
alto, como el caso de Laurencia sp. teniendo el mismo EC50 de 932 g/mL,
Ganonema farinosum con EC50 de 470 g/mL y Acanthophora spicifera con EC50 de
234 g/mL siendo ésta última una mejor actividad inhibitoria al presentar un EC50
reducido en comparación. Las algas rojas B. triquetrum, Laurencia sp. y A. spicifera
podrían presentar efecto similar al ser pertenecientes a la misma familia
Rhodomelaceae y tener propiedades similares, sin embargo G. farinosum difiere al
ser de la familia Liagoraceae, coincidiendo con el resto sólo a nivel de clase,
perteneciendo todas a Florideophyceae.
Otros trabajos como el de Heo et al. (2006) establecen un bajo porcentaje de
inhibición antioxidante, donde tras realizar un estudio empleando 26 especies de
algas rojas, sólo sobresale el alga Polysiphonia japonica con un porcentaje del
94.92% de eliminación de DPPH, sin embargo, éste trabajo difiere del presente
estudio al emplear un extracto orgánico de origen metanólico en lugar de ser

33
etanólico. A su vez el estudio no muestra el EC50 del extracto empleado, por lo que
no se determina que tan efectiva es la concentración empleada o que cantidad se
requiere para tener el efecto obtenido.
Estudios demostraron que los efectos antioxidantes en algas rojas pueden
deberse a compuestos como el ácido p-cumárico, cinámico y ferúlico (Fallarero et al.,
2006). A su vez la actividad también puede atribuírsele a otros metabolitos con
función antioxidante como los carotenos y el ácido ascórbico (Yan et al., 1998). A su
vez el efecto de la actividad de los compuestos polifenólicos presentes en los
extractos podría ser favorecida por la presencia del ácido ascórbico formando una
reacción antioxidante potenciada, tal y como se ha descrito por otros autores en
modelos experimentales in vitro (Vieira et al., 1998).
Los compuestos fenólicos son componentes muy importantes por su
capacidad de captación, debido a sus grupos hidroxilo. Investigadores han informado
que los compuestos fenólicos están asociados con la actividad de eliminación de
radicales libres de DPPH. Un gran número de estudios señalan que los polifenoles
de algas marinas se asocian con la actividad antioxidante y juegan un papel
importante en la estabilización de la peroxidación de lípidos (Yen et al., 1993; Heo et
al., 2006). Los fenoles son particularmente antioxidantes eficaces para los ácidos
grasos poliinsaturados; de hecho, transfieren fácilmente un átomo de hidrógeno a los
radicales piróxilo lipídicos formando un grupo ariloxi, el cual es incapaz de actuar
como un portador de cadena, formando pareja con otro radical eliminando rápido el
proceso de radical (Ruberto et al., 2001).
En el presente estudio, el alga E. arborea mostró el mayor efecto antioxidante,
lo cual puede atribuirse al hecho de que es un alga marina con gran exposición a
factores dañinos, suponiendo que la función de las moléculas antioxidantes en algas
es brindarles protección contra la radiación UV o daño fotodinámico, así como altas
concentraciones de oxígeno que conducen a la formación de radicales libres y otros
agentes oxidativos (Amsler, 2008).
Investigaciones han demostrado que las algas manifiestan una relación entre
la actividad neuroprotectora y la actividad antioxidante. No obstante, aún se trabaja
en la total comprensión de los mecanismos moleculares que sustentan estas

34
propiedades neuroprotectoras (Fallarero et al., 2003). La citotoxicidad mediante un
tratamiento A se ha encontrado asociada con el estrés oxidativo. Varios marcadores
de exceso de estrés oxidativo, tales como el aumento de ERO, la acumulación de
productos oxidados como carbonilos de proteínas de la oxidación de proteínas y
aldehídos e isoprostanos de la peroxidación lipídica, confirma el papel directo de A
en el daño oxidativo asociado con alzheimer (Miranda et al., 2000; Shih-Huang et al.,
2008). Aunque el uso de antioxidantes ha sido reconocido como un método eficaz en
la reducción de los efectos patológicos y tóxicos asociados con el estrés oxidativo
(Ono et al., 2006).
Al evaluar la capacidad antioxidante de una muestra, es recomendable
combinar al menos dos métodos, al estar éstos basados en distintos principios. En
éste caso se utilizó FRAP al medir la capacidad de reducción de Fe, y el método de
DPPH que determina la capacidad de captación de radicales libres. Los resultados
de capacidad antioxidante se presentan de diferente manera al estar basados en
métodos diferentes, así, FRAP se expresa en mol/g mientras que el DPPH está
basado en la determinación de parámetros cinéticos, considerando los gramos
necesarios para capturar el radical, siendo así cuanto más antioxidante sea una
muestra, su valor de EC50 se verá reducido (Pérez-Jiménez y Saura-Calixto, 2007).
Los resultados para la prueba de FRAP en nuestra investigación no mostraron
diferencias significativas entre algas rojas y pardas (ANOVA, F=0.00961 P= .99979),
sin embargo, otros trabajos si presentan a las algas pardas como las que tienen
mejores efectos de conversión del sistema, como las algas Turbinaria conoides y
Turbinarua ornata las cuales se muestran como muy buenos antioxidantes debido a
sus polisacáridos sulfatados, destacando el fucoidan, laminaran y alginato. A su vez
se presentan los carotenoides como estructuras abundantes con efectos
antioxidantes, tales como la fucoxantina que se encuentra de manera abundante y
presenta muy notable actividad biológica. Éstos compuestos presentan buenos
efectos antioxidantes, sin embargo, se desarrollan de manera diferente a los
florotaninos, por lo que el sistema FRAP se muestra como complemento para la
prueba de DPPH para observar en conjunto la actividad antioxidante como posible

35
tratamiento para el alzheimer (Chandini et al., 2008; Chattopadhay et al., 2010;
Kelman et al., 2012).
La inhibición de enzimas colinesterasas es una de las aproximaciones más
comunes para el tratamiento del alzheimer. Se emplea como control de
neurotransmisores en el cerebro. Sin embargo, en la última década se ha empleado
también como blanco para observar una disminución en la neurodegeneración la cual
parte de la formación de placas -amiloides (Hodges, 2006; Mukherjee et al., 2007).
En nuestra investigación obtuvimos que el alga roja Laurencia sp. mostró el
mejor efecto inhibidor de la enzima AChE con un porcentaje del 85.70% así como un
EC50 de 4.987 g/mL, siendo un valor casi cercano al control empleado donepezil
con un EC50 de 4.780 g/mL (Tabla 3). Syad et al. (2012) reportaron que el alga roja
Gelidiella acerosa alcanzó un porcentaje de inhibición de AChE del 54.18 %
empleando 487.80 g/mL de extracto bencénico, obteniendo un EC50 de 432.61
g/mL. En su investigación ningún alga roja presentó un EC50 tan alto, sin embargo,
coincide con nuestro trabajo en que para presentar un porcentaje mayor al 50% se
emplea una concentración mayor, siendo las más parecidas Laurencia sp. con una
concentración de 31.25 g/mL para un 85.70% de inhibición y Acanthophora spicifera
alcanzando un porcentaje del 82% con una concentración de 625 g/mL. Esto de la
misma manera se puede corroborar con el trabajo de Senevirathne et al. (2010)
donde establecieron que al utlizar extractos de algas rojas como Porphyra tenera se
deben emplear extractos de 1 mg/mL si se quiere obtener porcentajes de inhibición
mayores al 60 %. En otros trabajos presentan incluso menores efectos de inhibición
de algas rojas, como el caso de Ghannadi et al. (2013) donde establecieron que para
Gracilaria corticata y Gracilaria salicornia se tiene un EC50 de 9.5 y 8.7 mg/mL
respectivamente, siendo una concentración muy elevada para tomar en cuenta el
rendimiento del alga empleada.
Se ha informado previamente que tanto los alcaloides y flavonoides son
buenos inhibidores de la AChE, compuestos que pueden ser encontrados en algas
rojas. Una estrategia de tratamiento para mejorar las funciones colinérgicas para
aumentar la cantidad de acetilcolina disponible en las sinapsis entre las neuronas
colinérgicas. (Andrade et al., 2005; Jung y Park 2007). Por otra parte, Hodges (2006)

36
demostró que la inhibición de AChE juega un papel clave no sólo en la mejora de la
neurotransmisión colinérgica en el cerebro sino también en la reducción de la
agregación -amiloide, factor clave en la enfermedad de Alzheimer.
El donepezil así como otros inhibidores empleados actualmente han
presentado otro tipo de actividades funcionales para secretasas (Zimmermann et al.,
2004). Un informe reciente también demostró que los inhibidores selectivos de BChE
reducen el procesamiento de la proteína precursora amiloide y el nivel de A in vivo e
in vitro (Greig et al., 2001). Esta información sugiere que los efectos pueden
derivarse de la interacción de estos fármacos con la cascada amiloide, influyendo en
la expresión y el procesamiento metabólico de la proteína precursora amiloide (APP)
así como ralentizar las principales consecuencias patológicas de agregación
(Mukherjee et al., 2007).
En diferentes trabajos se han empleado plantas terrestres como objeto de
estudio sobresaliendo entre ellas el ginseng (Angelica sinensis), las cuales han
demostrado la relación y efecto contra la inhibición de la enzima acetilcolinesterasa y
el potencial antioxidante (Chang Ho et al., 2009). Ésta relación aún no es muy
estudiada en algas marinas, por lo que es importante probar extractos y sus
componentes activos los cuales podrían surgir como antioxidantes naturales,
fármacos anticolinesterásicos alternativos, o los cuales podrían servir como puntos
de partida en la síntesis de inhibidores más efectivos de la AChE (Shih-Huang et al.,
2008; Nour et al., 2014).
La inhibición de la actividad anticolinesterasa ha demostrado en algunos
estudios una relación fuertemente positiva con efecto antioxidante, tomando como
modelo el ensayo de DPPH, como el caso de Ali-Hassan et al. (2013) donde en su
investigación empleando el fruto Canarium odontophyllum, muestran una correlación
con un R de 0.913. Así mismo el trabajo de Amoo et al. (2012) indicaron una relación
positiva entre la actividad antioxidante y las propiedades inhibitorias de AChE de
algunas plantas medicinales africanas; así como Jo et al. (2012) donde reportaron
una correlación positiva entre las actividades previamente mencionadas, empleando
la semilla y el pericarpio de la planta de té Camellia sinensis. En nuestra
investigación se manejó una matriz de correlación entre los efectos antioxidantes

37
tomando en cuenta tanto FRAP como DPPH, así como el efecto inhibidor de AChE,
siendo el valor de R no tan alto para la relación entre FRAP y AChE de 0.60,
mientras que para la inhibición de AChE y DPPH muestra un valor de R= 0.92 siendo
similar a lo obtenido en los demás trabajos.
A su vez autores mencionan la relación entre el efecto de inhibición de AChE y
el efecto antioxidante empleando inhibidores sintéticos comerciales, como es el caso
de Priyanka et al. (2013) quienes observaron el efecto de donepezil sobre la
inhibición enzimática y actividad antioxidante en células de ratón que presentaban
daño por el envejecimiento. Saxena et al. (2008) mostraron que compuestos
comerciales como el donepezil y la tacrina, además de presentar efecto inhibitorio de
AChE también presentan efecto con supresión del estrés oxidativo. Diferentes
autores realizaron análisis de cerebros de ratones en estado de estrés oxidativo y
déficit de memoria, donde observaron la mejoría para ambos casos empleando
donepezil como inhibidor (Saxena et al., 2008; Cornelli, 2010).
Figura 8. Estructura química de los inhibidores tacrina y donepezil.
La actividad biológica que presentan el extracto etanólico y las fracciones de
E. arborea difieren; esto se debe a que las sustancias activas se van separando y
concentrando en ciertas fracciones, siendo por eso que las fracciones tienden a
inhibir la actividad enzimática con mayor efecto que el extracto crudo en sí. Este
mismo problema se puede ver reflejado en otro tipo de trabajos, como lo son
actividad antibacteriana o anti-incrustante donde puede presentarse un gran
problema al no emplearse una metodología muy sensible o si se utilizan bajas

38
concentraciones del principio activo, en donde puede pasar inadvertida la actividad
del extracto (Gómez-Alcalá, 2012). También puede ocurrir que una fracción no sea
tan activa mientras que el extracto crudo sí por cuestiones de concentración, como el
caso de E. arborea con respecto a la prueba DPPH; esto puede ser por que se
pierde la actividad debido que al reaccionar los grupos funcionales originan
subproductos inactivos, o porque la actividad se debe a la sinergia entre los
componentes de la mezcla, perdiendo la efectividad al separarse (Bhakuni y Rawat,
2005).
Para los parámetros de la cinética de inhibición se obtuvo el mejor resultado
para el alga roja Laurencia sp. teniendo un valor de Ki de 4.06, sin embargo, resulta
mayor que el valor del control empleado, teniendo el donepezil un valor de Ki de
3.18. El valor dado de Ki para el donepezil difiere con el trabajo de Geerts et al.
(2005), los cuales establecen valores de Ki para inhibición de AChE en el cerebro,
variando dependiendo del organismo objetivo empleado; 2.3 g/g en ratas, 0.65 g/g
para ratones y 1.3 g/g en conejos.
Hay investigaciones donde los autores emplean insecticidas clorpirifós (CPO)
para la inhibición de acetilcolinesterasa obteniendo valores de Ki de 2.2 a 9.3, donde
la AChE de la anguila Torpedo californica presentó un valor de Ki de 8.0, siendo la
misma enzima que se empleó para la realización de este ensayo; esto coincidió en
cuanto a los valores obtenidos en nuestra investigación, siendo el valor de Ki más
alto de 5.41 perteneciente a Ganonema farinosum, lo que contrastó al inhibidor CPO
empleado (Forsberg y Puu, 1984; Amitai et al., 1998). Es difícil contrastar los
resultados obtenidos de las constantes de inhibición con otros trabajos con aplicación
anti AChE donde se emplean algas marinas, ya que casi no se manejan parámetros
cinéticos, empleando solo sus porcentajes de inhibición, así como su EC50.
Los extractos crudos se encuentran catalogados dentro de los productos
naturales ya que contienen una mezcla de diversos compuestos, por lo que su
composición, así como rendimiento puede variar, dependiendo de factores como lo
son el organismo empleado, así como el disolvente utilizado (Sarker et al., 2006).
El presente trabajo muestra que los espectros infrarrojos observados en las
fracciones del alga parda Eisenia arborea sugieren la presencia de florotaninos

39
derivados del floroglucinol, coincidiendo con autores como Nashamura et al. (1996),
quienes establecieron que análisis de IR en algas del género Eisenia han mostrado
derivados de florofucofuroeckol, en conjunto con otros métodos espectroscópicos
muestran evidencia de presentar polímeros como eckol, florofucofuroeckol A, dieckol
y 8,8’-bieckol (Fig. 9). Esto ha sido reportado con anterioridad en las especies
Eisenia bicyclis y Eisenia kurome (Fukuyama et al., 1985; Ragan, 1985).
Figura 9. Estructuras químicas de dieckol (A) y florofucofuroeckol (B).
Derivados florataninos obtenidos del género Ecklonia han presentado buenos
efectos para actividad antioxidante siendo de los más activos el florofucofuroeckol-B
y el floroglucinol, presentando resultados significativos similares al ácido ascórbico
(Shibata et al., 2008; Yotsu-Yamashita, et al., 2013). A su vez eckol obtenido de
Eisenia cava ha demostrado mecanismos de protección que se emplean contra
especies reactivas de oxígeno presentes en ratones (Park et al., 2008). Los
floratatinos extraídos de los géneros de algas pardas como Eisenia y Ecklonia han
demostrado tener efectos contra la enzima acetilcolinesterasa, donde el dieckol y el
florofucofuroeckol mostraron una regulación en los niveles óptimos de la enzima en
el cerebro de ratones, así como mejoramiento en habilidades de la memoria (Myung
et al., 2005). Éstos efectos podrían explicarse de manera biológica gracias a trabajos

40
como los de Geiselman y McConnel (1981) y Taniguchi et al. (1991) donde afirman
que los florataninos de diversos pesos moleculares desempeñan un papel importante
en las defensas químicas contra el herbivorismo en especies marinas.
Estos puntos sobre los florataninos refuerzan nuestra hipótesis planteada en el
trabajo donde proponemos que las estructuras polifenólicas de las algas presentarían
compuestos que intervinieran tanto en la inhibición de la enzima acetilcolinesterasa
así como en la eliminación de radicales libres. Esto se comprobó gracias a lo
obtenido por medio del análisis de IR, obteniendo compuestos del floroglucinol, con
un efecto para ambas pruebas inhibitorias, siendo corroborado por el análisis de
correlación de las fracciones de Eisenia arborea, el cual presentó una relación con
una R de 0.94, por lo que ésta alga parda se presenta como una buena estrategia
para combatir, así como prevenir la enfermedad de Alzheimer.
Las estructuras presentadas por los derivados del florofucofuroeckol (Fig 9.)
muestran similitudes estructurales con los compuestos marinos obtenidos
anteriormente que presentan actividad anticolinesterasa (Fig 1.). A su vez las
estructuras de éstos derivados florotaninos presentan semejanza con los inhibidores
comerciales tales como la tacrina y el donepezil (Fig. 8), siendo éstos últimos
también efectivos como antioxidantes. Por lo que la estructura de los florotaninos
presentes en las algas pardas podría emplearse para tratamientos duales donde se
empleen como modelos moleculares de interacción para ambos procesos,
involucrados en la enfermedad del Alzheimer (Stoddard et al., 2014).

41
9. CONCLUSIONES
1) Los doce extractos etanólicos de las especies estudiadas presentaron
actividad antioxidante por el método de DPPH, resaltando las algas
Acanthophora spicifera, Laurencia sp., Ganonema farinosum, Dictyopteris
undulata y E. arborea donde se encontró que el alga parda E. arborea tiene la
mayor actividad antioxidante.
2) Para la inhibición enzimática de AChE el alga roja Laurencia sp. presentó el
mejor porcentaje de inhibición así como el valor más bajo de EC50.
Presentando una constante de inhibición de Ki de 4.06.
3) Se mostró una relación estrecha entre la inhibición de la actividad
anticolinesterasa y el efecto antioxidante de DPPH.
4) Los extractos etanólicos empleados presentan tanto efecto antioxidante como
inhibidor enzimático, por lo que se presentan como una alternativa para la
obtención de fármacos contra la formación de placas amiloides involucradas
en la enfermedad de Alzheimer.
5) La investigación sugiere que los compuestos observados en las fracciones del
alga parda E. arborea son derivados del floroglucinol. Siendo un posible efecto
inhibitorio tanto para oxidantes así como enzimas.
6) La investigación propone que la hipótesis es acertada siendo los florotaninos
los que intervienen en ambos procesos inhibidores por lo que pudieran ser
una alternativa para el control de la enfermedad de Alzheimer.

42
10. RECOMENDACIONES
1. Emplear diferentes enzimas involucradas en la formación de las placas
amiloides, como lo son el grupo de las secretasas, así como glutaminil ciclasa
con el fin de inhibir los diferentes procesos de la formación de las placas
amiloides y desarrollar una estrategia “multi-target”.
2. Fraccionar, aislar y purificar extractos para obtener compuestos activos y su
efecto en los ensayos de inhibición enzimática.
3. Realizar docking molecular para observar la interacción de los sitios activos en
las diferentes enzimas empleadas.
4. Llevar a cabo un experimento en organismos vivos para analizar la fisiología
del cerebro y comprobar la prevención o disminución de placas amiloides.

43
10. BIBLIOGRAFÍA
Ali-Hassan S. H., J. R. Fry y M. F. Abu-Bakar. 2013. Antioxidative phytochemicals
and anti-cholinesterase activity of native kembayau (Canarium odontophyllum)
fruit of Sabah, Malaysian Borneo. J. Nutr. Food Sci. 4: 249.
Alzheimer, A. 1907. Over an eigenartig disease of the cerebal cortex. Allg. A.
Phychiat. 63; 146-148.
Amitai, G., D. Moorad, R. Adani y B. P. Doctor. 1998. Inhibition of
acetylcholinesterase and buryrylcholinesterase by chlorphyrifos-oxon.
Biochem. Pharmacol. 56: 293-299.
Amoo, S. O., A. O. Aremu, M. Moyo y V. S. Staden. 2012. Antioxidant and
acetylcholinesterase-inhibitory properties of long-term stored medicinal plants.
J. Evid Based Complementary Altern Med. 12: 87-96.
Amsler, C. D. 2008. Algal Chemical Ecology. Springer, U. S. A., 311 pp.
Andrade, M. T., J. A. Lima, A. C. Pinto, C. M. Rezende, M. P. Carvalho y R. A.
Epifanio. 2005. Indole alkaloids from Tabernaemontana australis (Muell. Arg.I
Miers that inhibit acetylcholinesterase enzyme. Bioorg. Med. Chem. 13: 4092-
4095.
Arce, P. M., R. I. Franco, G. C. Muñoz, C. Pérez, B López, M. Villaroya, G. M. López
y G. A. García. 2009. Neuroprotective and cholinergic properties of
multifunctional glutamic acid derivates for the treatments of Alzheimer’s
disease. J. Med. Chem. 22: 7249-7257.
Barak, Y., J. Levine, A. Glasman, A. Elizur y R. H. Belmark. 1996. Inositol treatment
of Alzheimer’s disease: a double blind, cross-over placebo controlled trial.
Prog. Neuropsychopharmacol Biol Psychiatry. 20 (4): 729-735.
Behl, C. 1997. Amyloid -protein toxicity and oxidative stress in Alzheimer’s disease.
Cell Tissue Research. 290: 471-480.
Benzie I. F. y J. J. Strain. 1996. The ferric reducing ability of plasma (FRAP) as a
measure of “Antioxidant Power”: The FRAP Assay. Analytical Biochemistry.
239: 70-76.

44
Bhakuni, D. S. y D. S. Rawat. 2005. Bioactive Marine Natural Products. Springer.
New Delhi, India. 400 pp.
Brookmeyer, R., E. Johnson, K. Ziegler-Graham y H. M. Arrighi. 2007. Forecasting
the global burden of Alzheimer’s disease. Alzheimer’s and Dementia. 3 (3):
186-191.
Buchholz, M., U. Heiser, S. Shilling, A. J. Niestroj, K. Zunkel y H. U. Demuth. 2006.
The first potent inhibitors for human glutaminyl cyclase: synthesis and
structure-activity relationship. J. Med. Chem. 49 (2): 664-677.
Burrit, D., J. Larkindale y C. L. Hurd. 2002. Antioxidant metabolism in the intertidal red
seaweed Stictosiphonia arbuscular following desiccation. Planta 215 (5): 829-
838.
Casey, K. F., R. J. Beninger y M. Leyton. 2007. The 30th Annual Meeting of the
Canadian College of Neurophychopharmacology. J. Psychiatry Neurosci. 32
(6): 439-440.
Chandini, S. K., P. Ganesan y N. Bhaskar. 2008. In vitro antioxidant activities of three
selected brown seaweeds of India. Food Chem. 107: 707–713.
Chang Ho, C., A. Kumaran y L. S. Hwang. 2009. Bio-assay guided isolation and
identification of anti-Alzheimer active compounds from the root of Angelica
sinensis. Food Chem. 114: 246-252.
Chattopadhyay, N., T. Ghosh, S. Sinha, K. Chattopadhyay, P. Karmakar y B. Ray.
2010. Polysaccharides from Turbinaria conoides: Structural features and
antioxidant capacity. Food Chem.118: 823–829.
Choi, B. W., G., Ryu, S. H. Park, E. S. Kim, J. Shin, S. Roh, H. C. Shin y B. H. Lee.
2007. Anticholinesterasa activity of plastoquinones from Sargassum
sagamianum: lead compounds for Alzheimer’s disease therapy. Phytother. 21:
423-426.
Choi, J. S., S. Haulader, S. Karki, H. J. Jung, H. R. Kim y H. A. Jung. 2014. Acetyl
and butyryl-cholinesterase inhibitory activities of the edible brown alga Eisenia
bicyclis. Arch Pharm Res. 38 (8): 1477-1487.

45
Colombres, M., J. P. Sagal y N. C. Inestrosa. 2004. An overview of the current and
novel drug for Alzheimer’s disease with particular reference to anti-
cholinesterase compounds. Curr. Pharm. Des. 10: 3121-3130.
Cornelli, U. 2010. Treatment of Alzheimer’s disease with a cholinesterase inhibitor
combined with antioxidants. Neurodegener Dis. 7: 193-202.
Cornish, M. L. y D. J. Garbary. 2010. Antioxidants from macroalgae: potential
applications in human health and nutrition. Algae 25 (4): 155–71.
Dinamarca, M. C., J. P. Sagal, R. A. Quintanilla, J. A. Godoy, M. S. Arrázola y N. C.
Inestrosa. 2010. Amyloid--Acetilcholinesterase complexes potentiate
neurodegenerative changes induces by the A peptide. Implications for the
pathogenesis Alzheimer’s disease. Mol Neurodegener. 5 (4): 1-15.
Dinamarca, M. C., M. Arrázola, E. Toledo, W. F. Cerpa, J. Hancke y N. C. Inestrosa.
2008. Release of acetylcholinesterase (AChE) from b-amyloid plaques
assemblies improves the spatial memory impairments in APP-transgenic mice.
Chem Biol Interact. 175: 142-149.
Ellman, G. L., K. D. Courtney, V. Andres y R. M. Featherstone. 1961. A new and
rapid colorimetric determination of acetylcholinesterase activity. Biochem.
Pharmacol. 7 (2): 88-95.
Fallarero, A., A. Vidal y P. Vuorela. 2002. Protective effect of the aqueous extract of
Bryothamnion triquetrum against aglycemia and chemical hypoxia-induced
damage in GT1-7 immortalized mice hypothalamic cell line. Eur. J. Pharm. Sci.
17 (1): 42.
Fallarero, A., J. J. Loikkanen, P. T. Männistö, O. Castañeda y A. Vidal. 2003. Effects
of aqueous extracts of Halimeda incrassata (Ellis) Lamouroux and
Bryothamnion triquetrum (S. G. Gmelim) Howe on hydrogen peroxide and
methyl mercury-induced oxidative stress in GT1-7 mouse hypothalamic
immortalized cells. Phytomedicine. 10 (1): 39-47.
Finkel, T. y N. J. Holbrook. 2000. Oxidants, oxidative stress and the biology of ageing.
Nature. 408: 239-247.

46
Firuzi, O; A. Lacanna, R. Petrucci, G. Marrosu y L. Saso. 2005. Evaluation of the
antioxidant activity of flavonoids by “Ferric reducing antioxidant power” assay
and cyclic voltammetry. Biochim. Biophys. Acta. 1721: 174-184.
Fisher, A. 2011. Cholinergic modulation of amyloid precursor protein processing with
emphasis on M1 muscarinic receptor: Perspectives and challenges in
treatment of Alzheimer’s disease. J. Neurochem. 1: 22-33.
Forsberg, A. y G. Puu. 1984. Kinetics for the inhibition of acetylcholinesterase from
the electric eel by some organophosphates and carbamates. Eur. J. Biochem.
140: 153-156.
Fukuyama, Y., I. Miaura, Z. Kinzyo, H. Mori, Y. Nakayama, M. Takahashi y M. Ochi.
1985. Eckols, novel phlorotannins with a dibenzo-p-diozins skeleton
possessing inhibitory effects on macroglobulin from the brown alga Ecklonia
kurome OKAMURA. Chem. Lett., 739-742.
Geerts, H., P. O. Guillaumat, C. Grantham, W. Bode, K. Anciaux y S. Sachak. 2005.
Brain levels and acetylcholinesterase inhibition with galantamine and donepezil
in rats, mice and rabbits. Brain Res.1033 (2): 186-193.
Geiselman, J. A. y O. J. McConnel. 1981. Polyphenols in brown algae Focus
vesiculosus and Ascophyllum nodosum: Chemical defenses against the marine
herbivorus snail Littorina littorea. J. Chem. Ecol. 7: 1115-1133.
Ghannadi, A., A. Plubrukarn, K. Zandi, K. Sartavi y A. Yegdaneh. 2013. Screening for
antimalarial and acetylcholinesterase inhibitory activities of some Iranian
seaweeds. Res. Pharm. Sci. 8 (2): 113-118.
Glenner, G. G. y C. W. Wong. 1984. Alzheimer’s disease: initial report of the
purification and characterization of a novel cerebrovascular amyloid protein.
Biochem. Biophys. Res. Commun. 120: 885-890.
Gómez-Alcalá, L. 2012. Efecto in vitro del extracto etanólico de Macrocystis pyrifera
sobre el desarrollo de microepífitos. Tesis de Licenciatura. Universidad
Autónoma de Baja California Sur. La Paz, B.C.S. 49 pp.
Greig, N. H., T. Utsuki y Q. S. Yu. A new therapeutic target in Alzheimer’s disease
treatment: attention to butytylocholinesterase. Curr. Med. Res. Opin. 17 (3):
159-165.

47
Gulcin, I., M. Oktay, E. Kirecci y O. I. Kufrevioglu. 2003. Screening of antioxidant and
antimicrobial activities of anise (Pimpinella anisum) seed extracts. Food Chem.
83: 371-382.
Gupta, S., A. Fallarero, P. Järvinen, D. Karlsson, M. S. Johnson, P. M. Vuorela y C.
G. Mohan. 2011. Discovery of dual binding site acetylcholinesterase inhibitors
identified by pharmacophore modeling and sequential virtual screening
techniques. Bioorg. Med. Chem. Let. 21: 1105-1112.
Hardy, J. A. y G. A. Higgins. 1992. Alzheimer’s disease: the amyloid cascade
hypothesis. Science. 256 (5054): 184-185.
Hardy, J. y D. J. Selkoe. 2002. The amyloid hypothesis of Alzheimers’s disease:
progress and problems on the road to therapeutics. Science. 297: 353-356.
Harel, M., G. J. Kleywegt, R. B. Rabelli, I. Silman y J. L. Sussman. 1995. Crystal
structure of an acetylcholinesterase-fascitium complex: interaction of a three-
fingered toxin from snake venom with its target. Structure. 3: 1355-1366.
Heo, S. J., S. H. Cha, K. W. Lee y Y. J. Jeon. 2006. Anioxidants Activities of Red
Algae from Jeju Island. Algae. 21 (1): 149-156.
Hodges, J. R. 2006. Alzheimer centennial legacy: origins, landmarks and the current
status of knowledge concerning cognitive aspects. Brain Jour. Neurol. 129
(11): 2811-2822.
Houghton, P. J. y M. J. Howes. 2005. Natural products and derivatives affecting
neurotransmission neurotransmission relevant to Alzhiemer’s and Parkinson’s
disease. Neurosignals. 14 (1-2): 6-22.
Inestrosa, N. C., A. Álvarez, C. A. Pérez, R. D. Moreno, M. Vicente y C. Linker. 1996.
Acetyl cholinesterase accelerates assembly of amyloid-beta-peptides into
Alzheimer’s fibrils: possible role of the peripheral site of the enzyme. Neutron.
16: 881-891.
Ingkaninan, K., C. M. De Best y R. Van Der Hejiden. 2000. High-performance liquid
chromatography with on-line coupled UV, mass spectrometric and biochemical
detection for identification of acetylcholinesterasa inhibitors from natural
products. J. Chromatogr. 872 (1-2): 61-73.

48
Ismail, A. y S. Tan. 2002. Antioxidant Activity of Selected Commercial Seaweeds.
Malays. J. Nutr., 8 (2): 167–77.
Jo, Y. H., H. G. Yulk, J. H. Kim, R. Kim y S. C. Lee. 2012. Antioxidant, tyrosinase
inhibitory and acetylcholinesterase inhibitory activities of green tea (Camellia
sinensis L.) seed and its pericarp. Food Sci. Biotechnol. 12: 761-768.
Jung M. y M. Park. 2007. Acetylcholinesterase inhibition by flavonoids from Agrimonia
pilosa. Molecules. 12: 2130-2139.
Kang, I. J., B. G. Jang, S In, B Choi, M Kim y MJ Kim. 2013. Phlorotannin-rich
Ecklonia cava reduces the production of beta-amyloid by modulating alpha-
and gamma-secretase expression and activity. Neurotoxicology. 34: 16-24.
Kannan, R. R., Aderogba, M.A., Ndhlala, A.R., Stirk WA y Van Staden J. 2013.
Acetylcholinesterase inhibitory activity of phlorotannins isolated from the brown
alga, Ecklonia maxima (Osbeck) Papenfuss. Food Res Int. 54:1250–4.
Kannan, R. R., M. G. Kulkarni, W. A. Stirky y J. Van Stade. 2014. Advances in algal
drug research with emphasis on enzyme inhibitors. Biotechnol. Adv. 32: 1364-
1381.
Karran, E., M. Mercken y B. De Strooper. 2011. The amyloid cascade hypothesis for
Alzheimer’s disease: an appraisal for the development of therapeutics. Nature.
10: 698-712.
Katzman, R. 1976. The prevalence and malignancy of Alzheimer’s disease. Arch.
Neurol. 33: 299-308.
Kelman, D., E. Kromkowski Posner, K. J. McDermind, N. K. Tabandera, P. R. Wright
y A. D. Wright. 2012. Antioxidant activity of hawaiian marine algae. Mar. Drugs.
10: 403-416.
Khotimchenko, S. V. y V. E. Vas’kovsky. 2004. An inositol-containing sphingolipid
from the Red Alga Gracilaria verrucosa. Russ. J. Bioorganic Chem. 30 (2):
168-171.
Martínez Gil, M. 2009. El Alzheimer. Editorial CSIC. España. 126 pp.
Mata, A. T., C. Proenßa, A. R. Ferreira, M. L. M. Serralheiro, J. M. F. Nogueira y M.
E. M. Araújo. 2007. Antioxidant and antiacetylcholinesterase activities of five
plants used as Portuguese spices. Food Chem. 103: 778-786.

49
Matsunaga, M., R. E. Moore, W. P. Niemczura y W. W. Carmichael. 1989. Anatoxin-a
(s), a potent anticholinesterase from Anabaena flos-aquae. J. AM. Chem. Soc.
111: 8021-8023.
Mayer, A. M., A. D. Rodríguez, R. G. Berlinck y M. T. Hamann. 2009. Marine
pharmacology in 2005-6: Marine compounds with anthelmintic, antibacterial,
anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal,
antituberculosis, and antiviral activities; affecting the cardiovascular, immune
and nervous systems, and other miscellaneous mechanisms of action.
Biochimica et Biophysica. 1790: 283-308.
Merlini, G. y V. Belloti. 2003. Molecular mechanism of amyloidosis. N Engl J Med.
349 (6): 583-596.
Miranda, S., C. Opazo, L. F. Larrondo, F. J. Muñoz, F. Ruiz, F. Leighton y N. C.
Inestrosa. 2000. The role of oxidative stress in the toxicity induced by amyloid
-peptide in Alzheimer’s disease. Prog. Neurobiol. 62: 633-648.
Moo, R.D. y Y. Freile. 2009. Actividad citotóxica y antiproliferativa in vitro de
macroalgas marinas de Yucatán, México. Cienc Mar 35 (4): 345-358.
Morris, M. C., D. A. Evans, J. L. Bienias, C. C. Tangey, D. A. Bennet y N. Aggarwal.
2002. Dietary intake of antioxidant nutrients and the risk of incident Alzheimer
disease in a biracial community study. JAMA. 287: 3230-3237.
Mukherjee, P., V. Kumar, M. Mal y P. J. Houghton. 2007. Acetylcholinesterase
inhibitors from plants. Phytomedicine. 14 (4): 289-300.
Muñoz-Chacón, Y. 2003. Demencia, el reto del presente siglo. Acta Med Costarric.
45 (2): 42-42.
Muñoz-Ochoa. 2006. Extracción y caracterización de polisacáridos con actividad
anticoagulante a partir de algas colectadas en Baja California Sur, México.
2006. Tesis de Maestría. CICIMAR-IPN, México. 64 pp.
Muñoz-Pilar, P., L. Rubio, E. García-Palomero, I. Dorronsoro, M. Monte-Millán, R.
Valenzuela, P. Usán, C. Austria, M. Bartolini, V. Andrisano, A. Bidon-Chanal,
M. Orozco, F. J. Luque, M. Medina y A. Martínez. 2005. Design, Synthesis,
and Biological evaluation of dual binding site acetylcholinesterase Inhibitors:

50
New Disease-Modifyng Agents for Alzheimer’s Disease. J. Med. Chem. 48
(23): 7223-7233.
Myung, C., H. Shin, H. Bao, S. Yeo, B. Lee y J. Kang. 2005. Improvement of memory
by dieckol and phlorofucofuroeckol in ethanol-treated mice: Possible
involvement of the inhibition of acetylcholinesterase. Arch. Pharm. Res. 28:
691-698.
Myung, C. S., H. C. Shin, H. Ying Bao, S. J. Yeo, B. Ho Lee y J. Seong Kang. 2005.
Improvement of Memory by dieckol and phlorofucofuroeckol in ethanol-treated
mice: Possible involment of the inhibition of acetylcholinesterase. Arch- Pharm
Res. 28 (6): 691-698.
Nakamura, T., K. Nagayama, K. Uchida y R. Tanaka. 1996. Antioxidant activity of
fhlorotannins isolated from the brown algae Eisenia bicyclis. Fisheries Sci. 62
(6): 923-926.
Newman, D. J. y G. M. Cragg. 2007. Natural products as sources of new drugs over
the last 25 years. J. Nar Prod. 70: 461-477.
Nour, A.H., M. Khan, A. Z. Sulaiman, T. Batool, A. H. Nour, M. M. Khan, y F. Kormin.
2014. In vitro anti-acetyl cholinesterase and antioxidant activity of selected
Malaysan plants. Asian J. Pharm Clin. Res. 7(3): 93-97.
Ono, K. y M. Yamada. 2006. Antioxidant compounds have potent anti-fibrillogenic and
fibril-destabilizing effects for alpha-synuclein fibrils in vitro. J. Neurochem. 97:
105-115.
Pangestuti, R. y S. K. Kim. 2011. Neuroprotective effects of marine algae. Mar.
Drugs. 9: 803-818.
Park, E., G. Ahn, N. Ho Lee, J. Mi Kim, J. Seok Yun, J.W. Hyun, Y. J. Jeon, M. B.
Wie, Y. Jae Lee, J. Woo Park y Y. Jee. 2008. Radioprotective properties of
eckol against ionizing radiation in mice. FEBS J. 582: 925-930.
Pepys, M. B. 2006. Amyloidosis. Annu Rev Med. 57: 223-241.
Pereira, L., A. Sousa, H. Coelho, A. M. Amado y P. J. A. Ribeiro-Claro. 2003. Use of
FTIR, FT-Raman and 13C-NMR spectroscopy for identification of some
seaweed phycocolloids. Biomol. Eng., 20: 223-228.

51
Pérez-Jiménez, J. y F. Saura-Calixto. 2007. Metodología para la evaluación de
capacidad antioxidante en frutas y hortalizas. V Congreso Iberoamericano de
tecnología postcosecha y agroexportaciones. 1150-1160.
Prasansukab, A. y T. Tencomnao. 2013. Amyloidosis in Alzheimer’s disease: The
toxicity of amyloid beta (A), mechanism of its accumulation and implications
of medicinal plants for therapy. J Evid Based Complementary Altern Med.
2013: 1-10.
Priyanka, H., R. V. Singh, M. Mishra, S. ThyagaRajan. 2013. Diverse age-relaed
effects of Bacopa monnieri and donepezil in vitro on cytokine production,
antioxidant enzyme activities, and intracellular targets in splenocytes of F344
male rats. Int. Immunopharmacol. 15: 260-274.
Quinn, D. N. 1987. Acetylcholinesterase-Enzyme structure, reaction dynamics, and
virtual transition-states. Chem. Rev. 87: 955-979.
Ra Ahn, B., H. E. Moon, H. R. Kim, H. Ah Jung y J. S. Choi. 2012. Neuroprotective
effect of edible brown alga Eisenia bicyclis on amyloid beta peptide-induced
toxicity in PC12 cells. Arc. Pharmacal Res. 35 (11): 1989-1998.
Ragan, M. A. 1985. The high molecular weight polypholoroglucinols of the marine
brown alga Fucus vesiculosus L.: degradative analysis. Can. K. Chem. 63:
295-303.
Ramjee, P., N. Y. Yoon y S. K. Kim. 2010. Anti-photoaging and photoprotective
compounds derived from marine organisms. Mar. Drugs. 8 (4): 1189-1202.
Rhee, I. K., M. V. de Meent, K. Ingkaninan y R. Verpoorte. 2001. Screening for
acetylcholinesterase inhibitors from Amaryllidaceae using silica gel thin-layer
chromatography in combination with bioactivity staining. J. Chromatogr. 915:
217-223.
Rouleau, J., B. I. Iorga y C. Guillou. 2011. New potent human acetylcholinesterase
inhibitors in the tetracyclic triterpene series with inhibitory potency on amyloid
aggregation. Eur. J. Med. Chem. 46: 2193-2205.
Ruberto, G., M. T. Baratta, D. M. Biondi y V. Amico. 2001. Antioxidant activity of
extracts of the marine algal genus Cystoseira in a micellar model system. J.
Appl. Physiol. 13: 403-407.

52
Sarker, S. D., Z. Latif y A. I. Gray. 2006. Methods in Biotechnology: Natural Product
Isolation. Satyajit D., Ed. Human Press Inc: NJ, USA. 528 pp.
Saxena, G., S. P. Singh, A. Agrawal y C. Nath. 2008. Effect of donepezil and tacrine
on oxidative stress in intracerebral streptozotocin-induced model of dementia
in mice. Eur. J. Pharmacol. 581: 283-289.
Schilling, S., U. Zeitschel, T. Hoffmann, U. Heiser, M. Francke, A. Kehlen, M. Holzer,
B. Hutter-Paier, M. Prokesch, M. Windisch, W. Jagla, D. Schlenzing, C.
Lindner, T. Rudolph, G. Reuter, H. Cynis, D. Montag, H. Demuth y S. Rossner.
2008. Glutaminyl cyclase inhibition attenuates pyroglutamate A and
Alzheimer’s disease-like pathology. Nat. Med. 14: 1106-1111.
Senevirathne, M., C. B. Ahn y J. Y. Je. 2010. Enzymatic extracts from edible red
algae, Porphyra tenera, and their antioxidant, anti-acetylcholinesterase, and
anti-inflamatory activities. Food Sci. Biotechnol. 19 (6): 1551-1557.
Seneviranthne, M., S. K. Kim, N. Siriwardhana, J. H. Ha, K. W. Lee y Y. J. Jeon.
2006. Antioxidant potential of Ecklonia cava on reactive oxygen species
scavenging, metal chelating, reducing power and lipid peroxidation inhibition.
Food Sci. Tech. Int. 12: 27–38.
Shanthi, N, T. Eluvakkal, y K. Arunkumar. 2014. Characterization of Galactose Rich
Fucoidan with Anticoagulation Potential Isolated from Turbinaria decurrens
Bory de Saint-Vincent occurring along the Coast of Gulf of Mannar (Pamban),
India. J. Pharmacog. Phytochem., 3(2): 132–37.
Sharma, O. P. y T. K. Bhat. 2009. DPPH antioxidant assay revisited. Food Chem. 113
(4): 1202-1205.
Shibata, T., K. Ishimaru, S. Kawaguchi, H. Yoshikawa y Y. Hama. 2008. Antioxidant
activities of phlorotannins isolated from Japanese Laminariaceae. J. Appl.
Phycol. 20: 705-711.
Shih-Hao, H., L. Chun-Mao y C. Been-Huang. 2008. Protective effects of Angelica
sinensis extract on amyloid -peptide-induced neurotoxicity. Phytomedicine.
15: 710-721.
Silva, T. M. A., L. G. Alves, K. C. S. De Queiroz, M. G. L. Santos, C. T. Marqués, S.
F. Chavante, H. A. O. Rocha y E. L. Leite. 2005. Partial characterization and

53
anticoagulant activity of a heterofucan from the brown seaweed Padina
gymnospora. Braz. J. Med. Biol. Res. 38: 523- 533.
Singh, M., M. Kaur, H. Kukreja, R. Chung, O. Silakari y D. Singh. 2013.
Acetylcholinesterase inhibitors as Alzheimer therapy: From nerve toxins to
neuroprotection. Eur. J. Med. Chem. 70: 165-188.
Stoddard, S. V., M. T. Hamann y R. M. Wadkins. 2014. Insights and ideas garnered
form marine metabolites for development of dual-function acetylcholinesterase
and amyloid- aggregation Inhibitors. Mar. Drugs. 12: 2114-2131.
Stutchbury, G. y G. Munch. 2005. Alzheimer´s associated inflammation, potential
drug targets and future therapies. J. Neural. Transm. 112: 429-453.
Sugiura, Y., K. Matsuda, T. Okamoto, M. Kakinawa y H. Amano. 2008. Anti-allergic
effects of the brown alga Eisenia arborea on Brown Norway rats. Fish. Sci., 74:
180-186.
Sussman, J. L., M. Harel, F. Frolow, C. Oefner, A. Goldman, L. Toker y I. Silman.
1991. Atomic structure of acetylcholinesterase from Torpedo californica: A
prototypic acetylcholine-binding protein. Science. 253: 872-879.
Syad, A. N., K. P. Shunmugiah y P. D. Kasi. 2012. Assessment of Anticholinesterase
activity of Gelidiella acerosa: implications for its therapeutic potential against
Alzheimer’s Disease. J. Evid. Based Complementary Altern. Med. 2012: 1-8.
Taniguchi, K., J. Kurata y M. Suzuki. 1991. Feeding deterrent effect of phlorotannins
from the alfa Eckloia stolonifera against the abalone Haliotis discus hannai.
Nippon Suisan Gakkaishi. 57: 2065-2071.
Terry, R. D., N. K. Gonatas y M. Weiss. 1964. Ultrastructural studies in Alzheimer’s
pre-senile dementia. Am. J. Pathol. 44: 269-297.
Vidal, A., A. Fallarero, E. R. Silva de Andrade-Wartha, A. M. de Oliviera e Silva, A. de
Lima, R. P. Torres, P. Vuorela y J. Mancini-Filho. 2006. Composición química
y actividad antioxidante del alga marina roja Bryothamnion triquetrum (S. G.
Gmelin) Howe. Braz. J. Pharm. Sci. 42 (4): 589-600.
Vieira, O., I. Escargueil-Blanc, O. Meilhac, J. P. Basile, J. Laranjinha, L. Almeida, R.
Salvayre y A. Negre-Salvarye. 1998. Eddext of dietary phenolic compounds on

54
apoptosis of human cultured endotelial cells introduced by oxidized LDL. Br. J.
Pharmacol. 123: 565-573.
Vina, J., A. Lloret, R. Orti y D. Alonso. 2004. Molecular bases of the treatments of
Alzheimer’s disease with antioxidants: prevention of oxidative stress. JMAM.
25: 117-223.
Yan, X., T. Nagata y X. Fan. 1998. Antioxidative activities in some common
seaweeds. Plants Foods Hum. Nutr. 52 (3): 253-262.
Yan, X., Y. Chuda, M. Suzuki y T. Nagata. 1999. Fucoxanthin as the major
antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci. Biotechnol.
Biochem. 63 (3): 605-607.
Ye, H., K. Wang, C. Zhou, J. Liu y X. Zeng. 2008. Purification, antitumor and
antioxidant activities in vitro of polysaccharides from the brown seaweed
Sargassum pallidum. Food Chem. 111: 428–432.
Yen, G. C. 1993. Relationship between antioxidant activity and maturity of peanut
hulls. J. Agric. Food Chem. 41: 67-70.
Yoon, N. Y., H. Y. Chung, H. R. Kim y J. E. Choi. 2008. Acetyl- and
butyrylcholinesterase inhibitory activities of sterols and phlorotannins from
Eclonia stolonifera. Fish. Sci. 74 (1): 200.
Yoon, N. Y., S. H. Lee, L. Young y S. K. Kim. 2009. Phlorotannins from Ishige
okamurae and their acetyl- and butyrylcholinesterase inhibitory effects. J.
Funct. Foods. 1: 331-335.
Yotsu-Yamashita, M., S. Kondo, S. Segawa, Y. C. Lin. H. Toyohara, H. Ito, K. Konoki,
Y. Cho y T. Uchida. 2013. Isolation and structural determination of two novel
phlorotannins from the Brown alga Ecklonia kurome Okamura, and their radical
scavenging Activities. Mar. Drugs. 11: 165-183.
Zarros, A. 2009. In which cases is neuroprotection useful? Adv. Altern. Think.
Neurosci. 1: 3-5.
Zimmermann, M., F. Gardoni y E. Marcello. 2004. Acetylcholinesterase inhibitors
increase ADAM10 activity by promoting its trafficking in neuroblastom cell
lines. J. Neurochem. 90 (6): 1489-1499.





























