Embed Size (px)
Citation preview

1
Världen är så stor så stor Själv är jag rätt liten
Alla som i världen bor
Allting som de andra tror Pappa mamma storebror
Världen är rätt liten
Mindre än man nånsin tror
Själv ska jag ta brorsans skor Och gå den sista biten Ända till där jätten bor
Ur Iris bok av Ann Jäderlund (Jäderlund, 2002)

2
Table of content List of articles……………………………………… 3 Abstract……………………………………………. 4 Glossary/abbreviations…………………………..… 5 Introduction……………………………………….. 6
The fly..................................................................... 8 A road map to Drosophila immunity.................. 10
Here is the enemy................................................ 10
Fundaments of an immune response................................ 12 Molecular patterns............................................... 12
Pattern recognition.............................................. 13 PGRPs..................................... 13 Other recognition receptors. 15 Blood cells and their functions................................. 16 Phagocytosis......................................................... 18 Wound closure............................................................ 20 The firearms…………………………………… 20
Antimicrobial peptides....................................... 20 The phenoloxidase cascade………………… 23
Communication.......................................................... 24 Signalling pathways.................................................. 25
Imd/Relish.......................................................... 25 JNK...................................................................... 28
Dpp/BMP........................................................... 29 Toll........................................................................ 30
Other biological processes………………………... 31 The making of a wing……………………… 31 Salt stress…………………………………... 34
Aim of this thesis………………………………… 37 Results and discussion……………………... 37 Acknowledgement………………………………... 43 References………………………………………... 44 Paper I-V

3
List of articles included in this thesis This Thesis is based on the following articles or manuscripts, which will be referred to in the text by their roman numerals. All published articles were re-printed with the permission of the respective publisher. I Werner, T., Borge-Renberg, K., Mellroth, P., Steiner, H., and
Hultmark, D. (2003) Functional diversity of the Drosophila PGRP-LC gene cluster in the response to lipopolysaccharide and peptidoglycan. J Biol Chem.
II Hedengren, M., Borge, K., and Hultmark, D. (2000)
Expression and evolution of the Drosophila attacin/diptericin gene family. Biochem Biophys Res Commun.
III Borge-Renberg, K., Williams, M, J., and Hultmark, D.
PGRP-LC activates blood cells through JNK, and not via Relish Manuscript
IV Borge-Renberg, K., Williams, M, J., Wiklund, M-L, and Hultmark, D.
PGRP-LC and Dredd interact with JNK signalling in Drosophila development and immunity Manuscript
V Keyser, P., Borge-Renberg, K., and Hultmark, D. (2007)
The Drosophila NFAT homolog is involved in salt stress tolerance. Insect Biochem Mol Biol.

4
Abstract In general the work behind this thesis has revolved around the interesting pattern recognition gene family PGRPs (peptidoglycan recognition proteins). In particular the transmembrane PGRP-LC and to investigate its multifaceted role in the immune response of the fruit fly. As a well characterized model organism living on, and surrounded by, a multitude of microorganisms, Drosophila melanogaster serves as a great tool to gain insights about innate immunity. The two pillars of Drosophila innate immunity are the humoral and the cellular defense. Together they are very potent and can vanquish many infections, but if one of these pillars is damaged, chances are that the defense will collapse and the organism will succumb to the infection.
The initial step in any immune response is to become aware of the pathogen. To accomplish this, innate immunity relies on recognizing common molecular building blocks necessary each group of microorganisms. One such building block is the bacterial cell wall component peptidoglycan. PGRPs are a widely spread gene family, and proteins of this family can bind peptidoglycan. We describe that there are 13 PGRP genes in Drosophila, one these codes for PGRP-LC. As it sits in the cell membrane in any of its three different splice forms, PGRP-LC can bind peptidoglycan, dimerize, and subsequently activate the imd/relish signalling pathway, and thereby trigger a vast production of antimicrobial peptides. These short peptides are the firearms of the humoral response. We identified three new inducible antimicrobial peptide genes, Diptericin B, Attacin C and Attacin D. Analyses of their sequences shed light on the evolution and relationship of these antimicrobial peptides
The antimicrobial peptides are potent weapons, but without a functional cellular response the animal is at loss. Animals lacking blood cells are gravely compromised. It is interesting to find that PGRP-LC is involved at this end of the immune response equation as well. We have found that PGRP-LC is able to activate blood cells and increase numbers of circulating cells, in a JNK (Jun N-terminal kinase) dependent manner. Intriguingly this activation is not dependent on Relish, the NF-kB transcription factor of the Imd/Relish pathway.
PGRP-LC activation funnels into both Imd/Relish and the JNK pathways. When PGRP-LC is lost, it appears that some basal, or background, JNK activation is lost. These effects are very mild, however the animal appears to become more sensitive to additional perturbations in this signalling pathway. This was the starting point when we started to re-evaluate Dredd, the caspase responsible for cleaving and activating Relish. Dredd also contributes to the JNK signalling pathway.

5
Glossary/ abbreviations Antimicrobial peptide protein able to kill or inhibit growth of microorganisms Apoptosis Self destruction of cells, often when something is not “wright” β−1,3-glucan Fungal cell wall component
CpG-DNA Phosphorylated DNA which is a common motif among bacteria
Crystal cell Type of blood cell Dpp Decapentaplegic, a TGF-β Dredd The caspase that is responsible for Relish activation
dsRNA double stranded RNA DAP-type peptidoglycan meso-diaminopimelic acid as crosslinker. Most G- some G+
Fat body Functional analog to the liver, production site for antimicrobial peptides G- Gram-negative bacteria G+ Gram-positive bacteria Haemocytes Blood cells
Haemocoel Body cavity Haemolymph The body fluid Imaginal disc Cluster from embryogenesis, to be used in the making of the adult fly Imd Signalling component of the Imd/Relish pathway JNK Jun N-terminal kinase Lamellocyte Type of blood cell LPS Lipopolysaccaride cell wall component of Gram- bacteria Lys-type peptidoglycan Lysin as cosslinker aminoacid Gram+ Lymph gland Haematopoetic tissue, site of blood cell proliferation NFAT Transcription factor belonging to the Rel family PAMP Pathogen associated molecular pattern Peptidoglycan Cell wall component of bacteria PGRP Peptidoglycan recognition protein Phagocytosis Uptake of microorganisms/particles for destruction Plasmatocyte Type of blood cell Phenoloxidase Catalyses the oxidation of phenolsmelanin. Toxic intermediates
PRR Pathogen recognition receptor, recognizes PAMPS Relish Transcription factor production of antimicrobial peptides
Toll Transmembrane receptor involved in embryogenesis and immunity

6
Introduction
Time has come for me to give my view on one corner of the scientific universe, the immune response of Drosophila melanogaster. It is a field within which one can learn much. Not just about the dynamics and statics of the immune system of a small insect, but of any organism. You come up close to the beauty and horror in the battle of life.
Higher vertebrates, have both innate and adaptive immune reactions. The fast innate, or immediate, response is based on a limited set of germline-encoded recognition receptors against molecular structures of microorganisms. The slower, precisely tailored, adaptive response utilizes the enormous variation of receptors, randomly designed by somatic recombination, and the clonal expansion of B- and T-cells. The powerful combination of adaptive and innate immune responses, is however a problem for the scientist. The two branches are greatly entangled, and it is therefore difficult to dissect out one part of the immune response from the other. For this purpose, Drosophila melanogaster, is a highly valuable tool. It can, and does, give us insights in how our own innate immune response works, and we are after all closest to our selves.
Analyses of Drosophila immunity can also help us understand how insects can be carriers, or vectors, of both viral and bacterial diseases, which plague mankind. The actual plague (through the fleas on rats), malaria (through mosquitoes), tick-borne encephalitis (TBE), borrelia, West Nile virus, and sleeping sickness are a few examples of such insect-transmitted diseases.
I have spoken about the beauty of insect immunity from a practical point of view, but it also shows its beauty in the different kinds of basic biological processes it allows us to encounter and study, such as movement and closure of epithelia, and movements and function of immune cells. Furthermore it kindly offers us to study many of these processes twice per animal thanks to metamorphosis. We may also feel like deciphering geniuses when we study the complexity of the signalling events that control tissue integrity, or an immune response.
The subject of my work are the signalling events, or the communications, that take place as the fly tries to regain homeostasis, whether it has been shifted by an infection or by a salty environment. There are many signalling events during an immune response; from the initial recognition of an intruder, and the intracellular components involved, to the actions that the different components of the immune

7
take in order to vanquish the enemy, and how the different elements cooperate. In particular my focus has been the signals downstream of the pathogen recognition receptor, PGRP-LC. I also pay some attention to the developmental aspects of the signalling cascades involved.
Well I say that beauty is in the eye of the beheld, and it is multifaceted.

8
The fly
How does it work? First we need a fruit fly. Fruit flies live in rather unhealthy surroundings where decomposing fruit grants a good feast, since Drosophila feed on yeasts and other microorganisms growing on these fruits.
The life cycle of a fruit fly. The larval stages are abbreviated with L1 to L3 for each of the three larval instar stages.

9
The adult female lays its fertilised eggs in this gooey mess, to ensure that her offspring are well provided for. The egg undergoes embryogenesis. The initial steps in the developmental processes are not guided by the egg or embryo and its genetic setup; instead they are dependent on the mother’s genetic setup through proteins and mRNA molecules she lays down in the egg (maternal contribution). If the female is a carrier for two mutant alleles in a maternally contributed gene, the embryo will also be mutant, and may die. An example of such a gene is Toll, which regulates the dorso-ventral polarity in the embryo (Anderson et al., 1985; Drier and Steward, 1997). In addition to this Toll is also an important immune gene.
In most cases the embryo is fortunate, and it can continue to develop and go through a process known as dorsal closure. During dorsal closure epithelial sheets spread and grow together in order to seal off the embryo from the rest of the world. (Reviewed in (Harden, 2002)). When dorsal closure fails, the embryo has a hole in the epithelia, and it dies. Movements of epithelia or other cells are common events in biology, and many parallels, molecular and functional, can drawn be between dorsal closure, wound healing, thorax closure and movement of blood cells.
A successful dorsal closure is essential for animal to become a small first instar larva, which hatches from the egg. The little first instar, eating-machine grows to become a second instar, and then a third instar larva, with successive molts in-between. As a late third instar larva, the animal temporarily stops feeding, and begins to pupariate. This is a bewildering event, where the entire animal practically disintegrates, and remakes itself from dormant imaginal cells and discs, into a whole new animal with a different body plan. It is now an adult which can mate and continue the life cycle at the beginning of this paragraph; a paragraph which has taken a minute or two for you to read, but in life it takes at least 9 days at 25oC for the fly to complete it.

10
A road map to Drosophi la immunity
Here is the enemy!
The fruit fly feed on the microorganisms, which grow on the decomposing fruits. This also means that they are at risk at all times. Friendly food can easily become furious foe, if the fly’s cuticle should break on the outside, or on the inside (gut or trachea). When that happens, any fungus or bacterium, will try to seize the opportunity to enter this Drosophila shaped feast hall, and the fly will die if it does not have a rapid and competent immune response. The enemy of a fruit fly does not only come in the microscopic size of microorganisms. Larger multicellular parasites are also a potential threat.
It is not possible to look at all kinds of bacterial, fungal, viral, nematode, wasp strains that there are on this planet. So instead we are using model bacteria, fungi, wasps, nematodes to infect our model organism the fruit fly. Hoping that this will tell us some, or maybe a lot, about how a generalised response to such an organism works.
Bacteria are typically divided into two groups, Gram-positive and Gram-negative bacteria. The difference between Gram-positive and Gram-negative is based on differences in bacterial cell wall construction (see figure), which also reflect how they stain by Gram-staining.

11
A cartoon illustrating the cell wall component peptidoglycan, and the cell walls of Gram-positive and Gram-negative bacteria.
Typically, cell walls of Gram-positive bacteria contain thick layers of
peptidoglycan and smaller amounts of teichoic acid; this cell wall faces the environment, and is lined with the inner membrane (Brock, 1994).

12
Most Gram-positive bacteria have Lys-type peptidoglycan, but not all (for instance bacteria from the genus Bacillus have DAP-type peptidoglycan) (Schleifer and Kandler, 1972). The cell walls of Gram-negative bacteria contain a thin layer of peptidoglycan, surrounded by an outer and an inner membrane (Brock, 1994). The outer membrane of Gram-negative bacteria also contains lipopolysaccaride, LPS. So not only can bacteria differ in the ratios between different cell wall components, but also how these components are exposed. Probably all Gram-negative have Lys-type peptidoglycan (Schleifer and Kandler, 1972).
In addition to this, bacteria make their peptidoglycan in different ways. The composition and structure of peptidoglycan is rather constant within the group of Gram-negative bacteria, but varies more in-between Gram-positive bacteria (Schleifer and Kandler, 1972), the composition of peptidoglycan can also vary depending on environmental factors (Vollmer et al., 2008). A more recent review about peptidoglycan structure was written by Vollmer et al., 2008 (Vollmer et al., 2008).
Fundaments of an immune response
Molecu lar pat terns
There are common features specific to the major groups of microorganisms. Microorganisms need to be made by, or contain, certain building blocks in order to be a functional bacteria, fungi, or virus. Other organisms do not need these specific features, and thus most do not have them. An example is the peptidoglycan of bacterial cell walls. The same basic principle can be applied to any organisms. These features can be utilized when trying to distinguish different organisms from each other, and this is precisely what occurs during the innate immune response of Drosophila and other organisms. This was the idea that Janeway had when he proposed that the innate immune response is based on pathogen associated molecular patterns, PAMPs, and pathogen recognition receptors, PRRs (Janeway, 1989), and that innate immunity was needed to prime the adaptive immunity. These pathogen recognition receptors are responsible for detecting the invader, and subsequently alerting and activating an efficient defense (if the host is fortunate). Examples of PAMPs are cell wall components of bacteria such as lipopolysaccaride, peptidoglycan, lipoteichoic acid, bacterial flagellin, β−1,3-glucan from fungal cell walls, or foreign genomes like bacterial CpG-DNA or double stranded RNA.

13
A cartoon illustrating the activation of the immune response. When no microorganisms are present (left), the immune system is inactive. When microorganisms have invaded the host (right) fragments of microorganisms can bind to receptors, and the immune response becomes activated.
There are however, to date, no known molecular features of other intruding multicellular organisms, like the parasitic wasp. Many of these PAMPs are recognized throughout many species, or hosts by pattern recognition receptors. Not surprisingly some of these pattern recognition receptors are similar throughout a wide range of organisms, either thanks to convergent evolution or due to a common origin.
Patte rn re cogni t ion
PGRPs
There is peptidoglycan in virtually all bacteria, but not in eukaryotes (Schleifer and Kandler, 1972), thus it fulfills the requirements for a PAMP. One important group of pattern recognition receptors is the peptidoglycan recognition protein, PGRP, family. (Reviewed in (Dziarski and Gupta, 2006b; Steiner, 2004). There are as many as thirteen different PGRP genes in the fruit fly genome (Werner et al., 2003; Werner et al.,

14
2000)(paper I), many of which have been shown to involved in the actual recognition event in the immune response. Where they activate the major signalling pathways; PGRP-LC is upstream of Imd/Relish (Choe et al., 2002; Gottar et al., 2002; Rämet et al., 2002b); and PGRP-SA and PGRP-SD are upstream of Toll (Leone et al., 2008; Michel et al., 2001). Yet others function as effector molecules like PGRP-SB1 (Mellroth and Steiner, 2006). Though there is an abundance of PGRP genes in Drosophila, the first PGRP was isolated from another insect, the silk moth Bombyx mori (Yoshida et al., 1996; Yoshida et al., 1986), as a protein capable of activating the phenoloxidase cascade (see below), and soon after in Tricoplusia ni as a peptidoglycan binding protein (Kang et al., 1998). PGRPs are found in a wide range of animals, but not in plants or nematodes, making them far from exclusive to insects. There are also PGRPs in humans, one is a secreted amidase, and the other three are bactericidal proteins (reviewed in (Dziarski and Gupta, 2006a)).
A crystal structure of PGRP-LCx (green), -LCa (blue) heterodimer with the peptidoglycan fragment TCT (red). From (Chang et al., 2006). Reprinted with permission from AAAS.
The peptidoglycan recognition domain is related to bacterial type 2
amidases, and some of them are functional enzymes capable of degrading peptidoglycan, like PGRP-SC1B, PGRP-SB1, PGRP-LB. As these enzymes hydrolyze the peptidoglycan, it removes some of the triggers for the immune response, thus attenuating the immune response (Bischoff et al., 2006; Mellroth et al., 2003; Zaidman-Remy et al., 2006) or it may result in antibacterial activity (Mellroth and Steiner, 2006)
One of these thirteen Drosophila PGRP proteins is PGRP-LC. As the transmembrane receptor for the Imd/Relish signalling pathway, with multiple spliced alternatives (x, a, and y) conveying the capability to

15
recognise different pathogen- associated epitopes, PGRP-LC is of great importance for the immune response of Drosophila (Paper I) (Choe et al., 2002; Gottar et al., 2002; Kaneko et al., 2004; Mellroth et al., 2005; Rämet et al., 2002b; Werner et al., 2003). It is also central to the work behind this thesis.
Depending on the ligand, both homo- and hetro-dimers can form (Chang et al., 2006; Mellroth et al., 2005; Werner et al., 2003). The PGRP-LCx splice variant can bind to both polymeric and monomeric peptidoglycan. As PGRP-LCx binds monomeric peptidoglycan it exposes and presents the peptidoglycan fragment to PGRP-LCa (which lacks the typical peptidoglycan-binding groove), and allow hetero-dimer formation (Chang et al., 2006; Mellroth et al., 2005). As a consequence of dimeristaion, PGRP-LC signals through Imd, and it results in the production and release of the firearms of the immune response, the antimicrobial peptides (Choe et al., 2005; Choe et al., 2002).
Another curious PGRP is PGRP-LF, a gene sitting as a neighbor to PGRP-LC on the chromosome. PGRP-LF is a transmembrane protein, which possesses not one but two PGRP domains. PGRP-LF can hamper PGRP-LC signalling (Maillet et al., 2008; Persson et al., 2007; Werner et al., 2003). (This protein is also discussed briefly below).
There are PGRPs upstream of Toll, the other major antimicrobial inducing signalling pathway, as well. Secreted PGRP-SA and PGRP-SD together with GNBP1 mediate peptidoglycan dependent activation of the Toll pathway (Bischoff et al., 2004; Gobert et al., 2003; Leone et al., 2008; Michel et al., 2001; Pili-Floury et al., 2004). PGRP-SA, is not only a pattern recognition molecule, but an enzyme, a carboxypeptidase (Chang et al., 2004).
Other pathogen recognition receptors
PGRPs are not the only pattern recognition molecules. As mentioned there are the β-glucan recognition proteins 1-3 (Gram negative binding protein, GNBP), and two genes, CG13422 and CG12780, which share homology to the N-terminal part of the GNBPs (Jiggins and Kim, 2006). GNBP1 recognize and hydrolyse peptidoglycan (Lys-type), and present is to PGRP-SA (Wang et al., 2006). Fungal components are recognized by GNBP3 (Gottar et al., 2006) to activate the serine protease cascade prior to Toll activation.

16
Blood c e l l s and the ir func t ions
We have a system where microorganisms have certain molecular features, which are recognized by the infected organism via its pattern recognition molecules. But many things remain to be done in order obtain inner peace. The blood cells of the insects need to become alerted about the presence of an invader. Then they have to try reach the enemy, attempt to devour it (phagocytose), or isolate it (encapsulate and melanise), and make sure that the fat body, which like all other organs is covered by a basal membrane, wakes up and starts to make the torpedoes, namely the antimicrobial peptides. A fly or larva with too few blood cells is not healthy and bacteria are living within it (Braun et al., 1998; Matova and Anderson, 2006). This underscores the importance of blood cells in the immune defence.
In the normal scenario, blood cells are present at all stages of development. In adults there is no known haematopoetic organ and there are many fewer blood cells. The cells found are phagocytic plasmatocytes (see below) (Lanot et al., 2001). All blood cells originate from two haematopoietic organs, the procephalic mesodermal anlage in embryos and the lymph gland in larva (Holz et al., 2003). At the third instar larval stage, the lymph gland can be divided into three distinct regions. The medulla holding progenitor cells, the cortex with mature cells, and the posterior signalling centre (Jung et al., 2005b). In the larva there are the three pools of blood cells, those that patrol the body cavity, the sessile ones that sit in a banded pattern below the cuticle (Lanot et al., 2001), and those in the lymph gland. A characteristic of an activated immune response is that the sessile cells are recruited into circulation (Lanot et al., 2001). A bacterial infection releases more blood cells that mere injury (Markus et al., 2005). Another sign of an activated cellular response is the appearance of lamellocytes (see below), which are barely present in a healthy larva, and the release of blood cells from the lymph glands. Rupture of/release from, the lymph gland is common phenomena after wasp infestation, but it also normally takes place during the prepupal stage (Lanot et al., 2001; Sorrentino et al., 2002).
Typically the circulating blood cells in Drosophila, are classified into three classes based on their function and morphology. These classes are plasmatocytes, lamellocytes and crystal cells. Reviewed in (Evans et al., 2003; Lavine and Strand, 2003; Meister, 2004; Ribeiro and Brehelin, 2006; Williams, 2007). The classification below is true for the approximately 5000 larval blood cells.

17
# Plasmatocytes: These cells are round, phagocytic cells which stain with P1 antibody (against the phagocytic receptor Nimrod) (Kurucz et al., 2007). The Plasmatocytes make up approximately 95% of all circulating cells, and they are 8-10µm in diameter.
# Lamellocytes: These cells are big flat cells (15-40 µm in diameter), which stain with the L1 antibody (Kurucz et al., 2003) and encapsulate particles too large to phagocytose. Lamellocytes are normally not detectable in healthy animals, but visible after the cellular response has become activated. These cells are only found in larvae. Lamellocytes are produced in response to parasitisation (Lanot et al., 2001; Sorrentino et al., 2002), and they may also be slightly induced by bacterial challenge, or wounding (Lanot et al., 2001; Markus et al., 2005).
# Crystal cells: Crystal cells are round cells with a diameter of 10-12 µm, carrying propenoloxidase crystals (Rizki and Rizki, 1959), which are released upon activation. Crystal cells can be visualised by heating the larva, since the phenoloxidase becomes activated by heat, and the cells turn black. This cell type corresponds to approximately 5% of the circulating haemocytes.
In addition to these cells there are also prohaemocytes. These cells
are smaller, 4-6 µm in diameter and are believed to give rise to additional plasmatocytes, crystal cells and possibly lamellocytes.
Exactly how the different classes are related to each other is a debated matter, but some facts are known. Expression of the GATA factor Serpent is necessary for the development of all blood cells (Lebestky et al., 2000). Depending on the ratio between Glial cell missing (Gcm1 and 2) and Lozenge, the Serpent expressing prohaemocytes can develop into plasmatocytes or crystal cells respectively. (Alfonso and Jones, 2002; Lebestky et al., 2000; Muratoglu et al., 2006). Furthermore, U-shaped inhibits crystal cell formation (Fossett et al., 2003), but is repressed by Serpent in combination with Lozenge (Fossett et al., 2003). Rupture and subsequent release of the phenoloxidase from crystal cells is dependent on the JNK pathway, small Rho GTPases and Eiger (Bidla et al., 2007).

18
The signal-induced molecular mechanisms behind lamellocyte differentiation are more of a puzzle. In response to wasp infestation, lamellocyte differentiation is dependent on collier, an orthologue of the mammalian early B-cell factor (Crozatier et al., 2004b). In addition, JAK/STAT, JNK activity and Toll signalling can induce lamellocyte formation (Huang et al., 2005; Lemaitre et al., 1995b; Luo et al., 1995; Qiu et al., 1998; Zettervall et al., 2004) (paper III).
In order to find an invader, the blood cells patrol the haemocoel, the body cavity, of the insect. These movements are probably both active, by cellular movements and changes in adhesion properties, and passive by floating around in the haemolymph, which is pumped around by the heart (the dorsal vessel). Some of the signalling pathways involved in cell movement regulate molecules needed for the cells to get a grip somewhere, ropes, tackles and pulleys, and components that allow the cell to let go. This calls for careful control and rearrangements of the cytoskeletal network. There are many similarities in how the haemocytes move, and the movements involved in both dorsal closure and wound closure. Therefore it is not surprising to find the small GTPases (Rac 1, 2 and Cdc42), and JNK signalling to be involved (Paladi and Tepass, 2004; Stramer et al., 2005; Williams et al., 2006).
Phagocytosis
During recent years, the similarities between mammalian and invertebrate (Drosophila) innate immunity have become more evident. One important mechanism of innate immunity is the clearance of microbes and apoptotic cells by phagocytic cells. In Drosophila, these tasks are mainly executed by the plasmatocytes, a class of blood cells. Empowered with blood cell derived phagocytic cells lines like the Drososphila S2 cells, which are easily manipulated by RNAi for knock down of genes (Hammond et al., 2000), a number of screens with slightly different focus on phagocytic interactions, have been executed (Kocks et al., 2003; Pearson et al., 2003; Philips et al., 2005; Rämet et al., 2002b; Stroschein-Stevenson et al., 2006). Using these cells, the scavenger receptor CI was cloned and identified as a third class of scavenger receptors involved in phagocytosis (Pearson et al., 1995; Rämet et al., 2001). The pathogen recognition receptor, PGRP-LC, has been implicated to be involved in phagocytosis (Rämet et al., 2002b). But the effects of knocking down PGRP-LC, in comparison to other phagocytic receptors, are relatively mild. Other components isolated as parts of the phagocytic machinery are β-COP, δ-COP, Actin, Rac,

19
Cdc42, and SCAR (Rämet et al., 2002b; Stuart et al., 2007). Of 600 proteins associated with the phagosome; 70% had a mammalian orthologue (Stuart et al., 2007). The molecules and mechanisms involved in Drosophila phagocytosis are reviewed by Stuart and Ezekowitz 2008 (Stuart and Ezekowitz, 2008).
Opsonis bind to foreign surfaces, and tag these objects for destruction or phagocytosis. In insects there is a group of thiolester-containing proteins (TEPs), which are related to C3 of the mammalian complement and α2-macroglobulin (Lagueux et al., 2000; Levashina et al., 2001). The secretion of these TEPs is induced upon infection. Once secreted, they bind to the surfaces of specific target microorganisms and promote phagocytosis. The different TEPs display different affinities for different microbes or parasites (Levashina et al., 2001; Stroschein-Stevenson et al., 2006). Though it is evident that TEP binding to microbial surfaces, targets the microbe for phagocytic uptake, it is not known with which blood cell receptors they co-operate.
In addition to opsonins, which tag particles for engulfment, there are cell-bound receptors. Croquemort and Peste are scavenger receptors of the same family as mammalian CD36. Both receptors are important for uptake of different microorganisms (Philips et al., 2005; Stuart et al., 2005), and in addition to this, Croquemort is required for phagocytosis of apoptotic cells (Franc et al., 1996).
The Nimrod C1, Eater, and Draper are cell surface receptors, containing multiple EGF-like repeats involved in phagocytosis of bacteria, and or apoptotic cells (Freeman et al., 2003; Kocks et al., 2005; Kurucz et al., 2007). In addition to these proteins, there are nine additional EGF-like repeat Nimrod family genes in Drosophila (Kurucz et al., 2007). It would not be surprising if some additional genes belonging to the Nimrod family are involved in phagocytosis.
Evidently, phagocytosis by blood cells is of great importance. They remove apoptotic bodies; re-sculpture the extracellular matrix casting moulds during metamorphoses, and kill bacteria. This is only the beginning. Phagocytosis by blood cells also appear to play a role in full activation of the fat body and the production of antimicrobial peptides (Brennan et al., 2007; Foley and O'Farrell, 2003); but haemocytes also make antimicrobial peptides themselves.
It was recently reported that phagocytosis could potentiate a peculiar and narrow kind of immunological memory in the fruit fly (Pham et al., 2007). If an animal was infected with a sub-lethal dose of Streptococcus pneumoniae, or Beauvaria bassiana, and allowed to clear the infection with the aid of phagocytic cells, the animal was specifically protected against future infections of the same species.

20
Wound c losure
Flies, like us are not always lucky. Sometimes they get hurt. Anyhow, when they are wounded, they have to do what we also do. First they need to make sure that the hole is temporarily closed so that they do not bleed to death. Secondly they have to start making new epithelia. Until a few years ago, very little was known about both of these events.
As in us, the problem of closing off the wound temporarily is solved by clotting proteins. To find out what is involved in clotting, the content of the clot and haemolymph have been subjected to proteomic approaches (Karlsson et al., 2004; Scherfer et al., 2004; Vierstraete et al., 2003). Two proteins from these screens, hemolectin and Fondue are indeed important for proper clot formation (Goto et al., 2003; Scherfer et al., 2006). Hemolectin displays many similarities to proteins involved the coagulation process in humans, such as the von Willebrand factor, coagulation factor V/VIII, and complement factors (Goto et al., 2001).
Now the wound has been temporarily closed, but it needs to be repaired permanently. The animal has to make new epidermis, and the epidermis has to spread, and close the gap. The animal needs the signals which control the migration of a cell, the ropes and pulleys. Not surprisingly this involves the same kind of pathways as in dorsal closure and in the activation of blood cells. JNK signalling is required for wound healing in embryos, larvae and adults (Bosch et al., 2005; Galko and Krasnow, 2004; Mattila et al., 2005; Rämet et al., 2002a). In addition to the JNK pathway, small GTPases like Cdc42, Rho and Rac 1 and 2 (Redd et al., 2004; Stramer et al., 2005), and the transcription factor Grainy head (Mace et al., 2005) have been shown to play an important roles for epithelial integrity.
Wound healing is of fundamental importance for any multicellular organism and the fundaments of wound healing are similar in many organisms. In mammals Rho, JNK signalling and Grainy head are also important (Ting et al., 2005), reviewed in (Harden, 2005), and (Martin and Parkhurst, 2004).
The f irearms
Antimicrobial peptides
Blood cells can phagocytose and encapsulate, but this one-on-one kind of warfare is not very efficient when there are many enemies. And if

21
there is one bacterium there will soon be many. Fortunately, there is more to the immune response than haemocytes. There are the antimicrobial peptides as well (reviewed in (Bulet et al., 2004; Cociancich et al., 1994; Otvos, 2002; Shai, 2002)). The first antimicrobial peptides, were described in the 1980s by work on the moth Cecropia, by Hans Boman and co-workers (Hultmark et al., 1980; Steiner et al., 1981). Since those pioneering days many more antimicrobial peptides have been isolated from insects to man. In the fruit fly 28 antimicrobial peptide genes (including eight lysozymes) from nine classes are expressed. Most antimicrobial peptides are short (often < 50 amino acids), charged (often cationic), and sometimes amphipathic peptides. These properties facilitate the interactions with negatively charged microbial cell walls and membranes. Some antimicrobial peptides, like cecropins (Steiner et al., 1988), exert their power directly by interacting with the membranes of microorganisms and thereby killing or inhibiting the growth of bacteria and fungi (as reviewed in (Shai, 2002). Others act on intracellular targets (as reviewed in (Otvos, 2002)). Due to the increase of antibiotic resistant bacteria of recent years, there has been an intensified interest in finding substitutes for the traditional antibiotics, but the killing mechanisms for a many of the antimicrobial peptides, are still poorly understood.
Depending on killing specificity, induction pattern and different evolutionary relationship to each other, the antimicrobial peptides can be divided into different classes. In Drosophila, the nine antimicrobial peptide classes are cecropins, drosomycins, attacins, diptericins, drosocin, metchnikovin, defencin, andropin, and lysoszymes. Lysozymes, also play a role as digestive enzymes. These classes of antimicrobial peptides can be grouped together based on relationships and mechanisms of actions. Below, I will give a brief discription of a selection of these groups, based on relevance to this thesis and Paper II.
Within the group of α-helical amphipatic peptides, we find the first isolated antimcirobial peptide, cecropin (Hultmark et al., 1980; Steiner et al., 1981). Active cecropins are approximately 30-40 amino acids long, α-helical, cationic, and amphipathic. They disrupt the membranes of microorganisms (Steiner et al., 1988), thereby killing the enemy. The cecropin killing spectrum is broad, killing both Gram-positive and Gram-negative bacteria at 0,1-10µM concentrations (Samakovlis et al., 1990). Even fungi are sensitive to cecropins (Ekengren and Hultmark, 1999).
Another group contain short (approximately 20 amino acids) proline-rich peptides, like drosocin and pyrrhocoricin (from Pyrrhocoris apterus). After first interacting with LPS, these peptides enter the cytoplasm and act on intracellular targets such as the bacterial 70-kDa heat shock protein, DnaK (Otvos et al., 2000);(Kragol et al., 2002;

22
Kragol et al., 2001). The proline-rich antimicrobial peptides are mainly active on Gram-negative bacteria. This group of peptides is reviewed by Otvos, 2002 (Otvos, 2002).
Among the family of glycin-rich peptides, are the rather large attacins, and in Drosophila there are four attacins (approximately 200 amino acids). Attacins from other insects have been shown to bind to the cell wall component LPS, increase permeability of the outer membrane, inhibit synthesis of outer membrane proteins. Subsequently growing bacteria are incapable of proper divisions, resulting in long chains of dying bacteria (Carlsson et al., 1991; Carlsson et al., 1998; Engström et al., 1984; Hultmark et al., 1983). In Drosophila there are four attacins, A-D, two of which are described in paper II (Dushay et al., 2000; Hedengren et al., 2000; Åsling et al., 1995). At the N-terminus of attacins there is a short proline-rich stretch. Curiously a processed form of the Attacin C pro domain has itself been isolated in the body fluid, and the synthetic counterpart also displays activity against bacteria (Rabel et al., 2004). Glycin-rich antimicrobial peptides are generally active on gram-negative bacteria.
Diptericins are rather large (approximately 180-190 amino acids) chimeric proteins, with a N-terminal part similar to the proline-rich peptides, and an attacin like C-terminus. Like short proline-rich peptides and attacins, diptericins mainly kill Gram-negative bacteria. As we describe in paper II, there are two diptericins in the fruit fly, Diptericin, and Diptericin B (Hedengren et al., 2000; Lee et al., 2001; Wicker et al., 1990). As an antimicrobial peptide that is solely dependent on Imd /Relish signalling, diptericin is often used as a readout for signalling in the Imd/Relish pathway (Hedengren et al., 2000).
Antimicrobial peptides are found at 1-100 µM concentrations in the body fluid of insects, but they are produced by immune competent tissues elsewhere. The major quantities are synthesised by an organ named the fat body. The fat body is an adipose tissue, involved in energy storage, de-toxification and immune response, and it is coated by a basal membrane. In a third instar larva, the fat body occupies most of the body volume. Normally, when there is no sign of infection, there is no production of antimicrobial peptides from the fat body. This changes rapidly if the animal becomes infected. Within hours, many antimicrobial peptides are detected within the haemocoel (Rabel et al., 2004; Uttenweiler-Joseph et al., 1998), and the mRNA expression of for instance cecropin peak within 6 h post infection (Kylsten et al., 1990). The acute production of these peptides is essential for the survival of the fly, and mutants in the Imd/Relish and Toll signalling pathways die (Choe et al., 2002; Hedengren et al., 1999; Lemaitre et al., 1995a; Lemaitre et al., 1996; Leulier et al., 2000). Responsible for the acute

23
antimicrobial peptide gene transcription are the NF-κB transcription factors of these pathways; Relish in the Imd/Relish pathway, and Dif and Dorsal in the Toll pathway (De Gregorio et al., 2002b; Dushay et al., 1996; Ip et al., 1993; Lemaitre et al., 1995b). The impact of these transcription factors is evident; Relish mutants die if infected with one single bacterium (Hedengren et al., 1999). In addition to the imd/relish and Toll signalling pathways, other signalling cascades, like JNK, also modulate the humoral response (Delaney et al., 2006; Kallio et al., 2005; Park et al., 2004; Silverman et al., 2003; Tsuda et al., 2005).
Other than the systemic production of antimicrobial peptides from the fat body and haemocytes, the animal can also respond with locally induced antimicrobial peptide production in exposed epithelia like gut, reproductive tracts and salivary glands (Brey et al., 1993; Tingvall et al., 2001; Tzou et al., 2000; Önfelt Tingvall et al., 2001). The different expression and induction patterns call for other mechanisms, in addition to NF-κB transcription factors. This is reviewed by Uvell and Engström 2007 (Uvell and Engström, 2007).
The phenoloxidase cascade
One more visual way of getting rid of invaders is by activating the phenoloxidase cascade. The first PGRP was isolated as a phenoloxidase activator in the silk moth (Yoshida et al., 1996; Yoshida et al., 1986). During this toxic journey, where the road is the reward, phenoloxidase catalyzes the oxidation of phenols to quinones, which eventually undergo a non-enzymatic polymerisation, thus becoming eumelanin (brown-black). Reactive oxygen and nitrogen intermediates are formed in the process. The toxic properties of the intermediates formed are reviewed Nappi and Christensen 2005 (Nappi and Christensen, 2005), and the phenoloxidase cascade in immunity is reviewed in Cerenius and Söderhäll, 2004 (Cerenius and Söderhäll, 2004), and Christensen et al., 2005 (Christensen et al., 2005), Cerenius et al., 2008 (Cerenius et al., 2008). In short inactive pro-phenoloxidase (coded by the genes Black cells, Diphenol oxidase A2 and Diphenol oxidase A3) is stored in blood cells, and released into the haemolymph by JNK signalling dependent cell rupture and possible other mechanisms (Bidla et al., 2007). Upon wounding other stimuli, a serine protease cascade activates the pro-phenoloxidase-activating enzyme (Sp7 and MP1) (Castillejo-Lopez and Hacker, 2005; Leclerc et al., 2006; Tang et al., 2006). The phenoloxidase-activating enzyme is itself a serine protease, which cleaves and activates the pro-phenoloxidase to phenoloxidase.

24
Once activated the phenoloxidase cascade must be restricted to prevent excess melanisation. Hampering the phenoloxidase cascade is the job of a serpin (Serpin27A), serine protease inhibitor, which inhibit the pro-phenoloxidase-activating enzyme (De Gregorio et al., 2002a).
Though the phenoloxidase cascade is induced by wounding, microbial compounds, and parasitic attack, its function as a defence mechanism is disputed, and mutants are not severely compromised, compared to mutants in the Imd or Toll signalling pathways (Leclerc et al., 2006).
Communic ati on
I have mentioned the different organs involved in the immune response, the different kinds of blood cells, the fat body, the cuticle, and briefly the gut, and the different components involved such as the antimicrobial peptides, the phenoloxidase cascade, phagocytosis etc. But are they all individually combating the enemy, or do they work in concert, and how then is that achieved? One crucial problem for the insect is that the fat body, the major production site for antimicrobial peptides, is covered by a layer of extra-cellular matrix. So how can it detect the pathogen? Another spatial problem when I stared this work, was that PGRP-LC, the pattern recognition receptor upstream of the Imd/Relish signalling pathway responsible for most of the antimicrobial peptide production, had not been detected in the fat body (Paper III), the major site of antimicrobial peptide production .
An old idea supported by work in Manduca sexta was that blood cells could digest bacteria and supply soluble bacterial fragments like peptidoglycan or LPS to the fat body (Dunn et al., 1985; Taniai et al., 1997). More recent work has strengthened the link between blood cells and fat body (Agaisse et al., 2003; Bettencourt et al., 2004; Brennan et al., 2007; Dijkers and O'Farrell, 2007; Foley and O'Farrell, 2003).
It appears that proper bacterial phagocytosis by haemocytes, is necessary for full activation of the fat body (Brennan et al., 2007). Under specific conditions, like gut infection with Erwinia carotovora, blood cells are required for full fat body response (Basset et al., 2000; Foley and O'Farrell, 2003).
The nature of the signal activating the fat body in each case is not known, but some are suggested to be proteins (Bettencourt et al., 2004), and indeed some signals are proteins, like Unpaired 3 in JAK/STAT signalling (Agaisse et al., 2003), the cytokine/ ligand spätzle upstream of Toll, and recently an immune induced putative helical cytokine was

25
isolated (Malagoli et al., 2007). Yet other signals are not proteins. Nitric oxide produced in gut and blood cells provide the signal induced by gut infection of Erwinia carotovora (Foley and O'Farrell, 2003). The nitric oxide signalling in blood cells require calcineurin to transpond the signal to the fat body (Dijkers and O'Farrell, 2007). Or the signals may be microbial fragments, which are presented to the fat body, and are capable of inducing it.
Signalling pathways
There are many signalling pathways and together they control all the processes that take part in the immune response. Here I describe the components and regulation of the most important pathways concerning my work.
Imd/Reli sh
The imd/relish pathway is one of the major signalling highways in the immune response. Ablating this pathway renders the fly unable to produce many of the antimicrobial peptides, and the fly dies from infection by a single bacteria. Activation of the Imd/ Relish pathway is achieved through binding of the bacterial cell wall constituent, DAP-type peptidoglycan, to the transmembrane receptor PGRP-LC, peptidoglycan recognition protein LC. This allows the receptors to dimerise (Mellroth et al., 2005) and via the intracellular domain, the signal is transferred to Imd (immune deficient) (Choe et al., 2005). PGRP-LC activation may also be achieved by proteolyticly cleaving off the extracellular part of the molecule (Schmidt et al., 2007; Schmidt et al., 2008). The alerting signal is split up downstream of Imd, and carried further to BG4 and TAK1. Imd contains a death domain similar to that of mammalian RIP (receptor interacting protein which mediate both TNF or Fas induced NF-κB activation or apoptosis respectively) (Georgel et al., 2001). The protein BG4, (also known as Fadd, Fas associated death domain), contain both a death domain, with which it interacts with Imd, and a DID (death inducing domain), with which it interact with the caspase-8 homolog, Dredd. The BG4- Dredd, interaction, faciliate Dredd’s auto catalytic cleavage and activation (Hu and Yang, 2000). Both BG4 and Dredd mutants are more susceptible to bacteria in a similar manner as Relish (Elrod-Erickson et al., 2000; Leulier et al., 2000; Leulier et al., 2002; Naitza et al., 2002).

26
Provided that the NF-κb transcription factor Relish has become phosphorylated by the IκB-Kinase complex, Dredd can cleave it, and similar to the activation of mammalian p105 to p50, the DNA interacting Rel homology domain is released from the inhibitory ankyrin part (Stöven et al., 2000; Stöven et al., 2003). In contrast to the processing of p105 to p50, where the inhibitory part is trimmed off by a proteasome dependent degradation, the ankyrin domain of Relish remains in the cytoplasm (Stöven et al., 2000).
A number of events participate in the regulated activation of Relish. Caspar, (a Fas associated factor 1 homolouge) inhibits the Dredd dependent cleavage of Relish, and mutants display a constitutively active humoral immune response (Kim et al., 2006). Similarly another protein, Dnr1 (Defence repressor 1) also appears to regulate Relish processing, via RING-finger dependent interactions with Dredd (Foley and O'Farrell, 2004).
The ubiquitin-conjugating enzymes Bendless and UEVa1 appear to be important upstream of Tak1 for full IKK activation and the subsequent Relish activation as described by Zhou, et al. 2005. These authors also propose that Imd, BG4, and Dredd form a complex upstream of Tak1, that together with Bendless and UEVa1 is responsible for Tak1 activation.
However Relish activity is not merely controlled by Dredd. As mentioned, the signalling pathway split downstream of Imd. BG4 is recruited, and Tak1 (the MP3K, TGF-β activated kinase) is activated; through Tab2, and possibly the RING-finger protein Iap2, and the IκB-Kinase complex, becomes activated (Gesellchen et al., 2005; Huh et al., 2007; Kleino et al., 2005; Leulier et al., 2006; Valanne et al., 2007; Zhuang et al., 2006). The IκB-Kinase complex is a protein conglomerate of kenny (IKKγ) and ird5 (IKKα /β), capable of phosphorylating Relish. Both Tak1 and IκB-Kinase complex have been reported as needed for Relish signalling (Lu et al., 2001; Rutschmann et al., 2000; Silverman et al., 2000; Vidal et al., 2001; Wu and Anderson, 1998). However the direct involvement of Tak1, which also transpond signalling to the JNK pathway (Mihaly et al., 2001; Takatsu et al., 2000), has recently been questioned (Delaney et al., 2006). The activated REL-68 is free to move into the nucleus, bind to κB-sites in promoter regions of genes and promote their transcription. Many of Relish target genes are antimicrobial peptides, such as diptericin and cecropin.
The actions of an activated Relish also have other functions. Once activated, the Rel part can induce an accelerated proteasome dependent turnover of the protein, Tak1 (MAP3K) (Park et al., 2004), which is involved in both the Relish/ Imd pathway and the JNK pathway (Silverman et al., 2003; Vidal et al., 2001), thus dampening the effect of
27
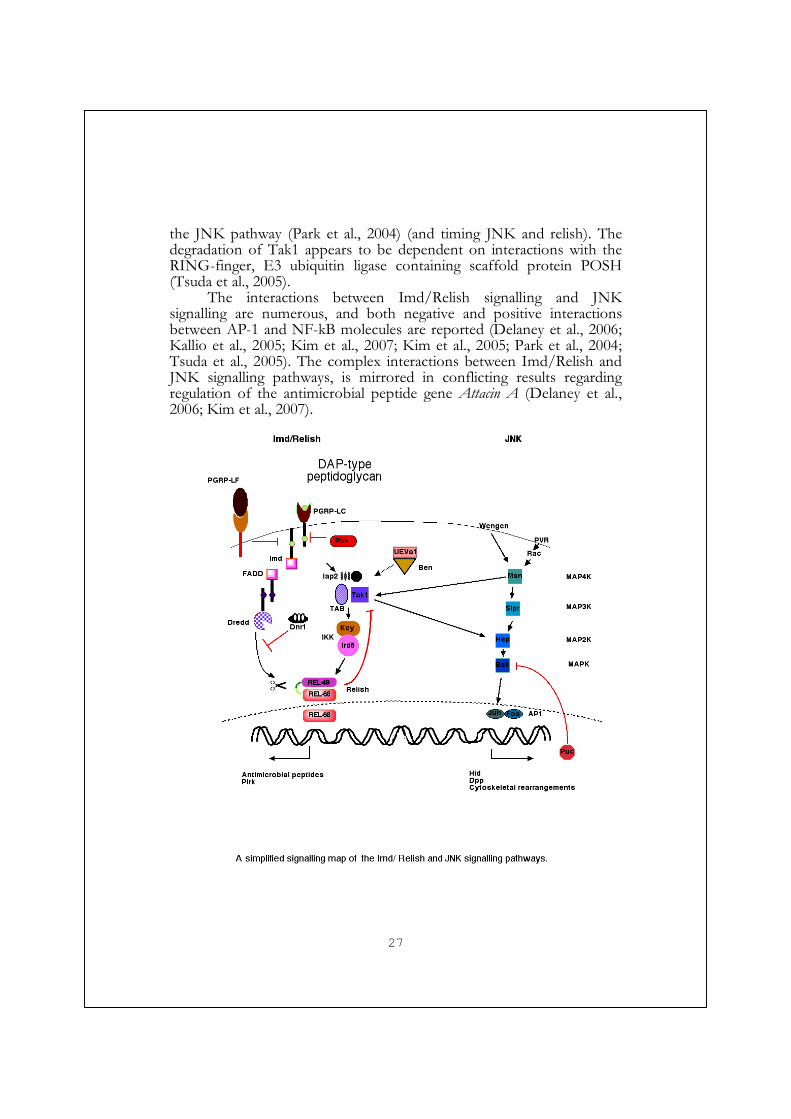
the JNK pathway (Park et al., 2004) (and timing JNK and relish). The degradation of Tak1 appears to be dependent on interactions with the RING-finger, E3 ubiquitin ligase containing scaffold protein POSH (Tsuda et al., 2005).
The interactions between Imd/Relish signalling and JNK signalling are numerous, and both negative and positive interactions between AP-1 and NF-kB molecules are reported (Delaney et al., 2006; Kallio et al., 2005; Kim et al., 2007; Kim et al., 2005; Park et al., 2004; Tsuda et al., 2005). The complex interactions between Imd/Relish and JNK signalling pathways, is mirrored in conflicting results regarding regulation of the antimicrobial peptide gene Attacin A (Delaney et al., 2006; Kim et al., 2007).

28
JNK
The Jun N-terminal kinase (JNK) pathway is one of the MAP kinase pathways, where the momentum is provided by protein kinases, which phosphorylate and activate other downstream kinases in a protein kinase cascade. The sequential chain of activating kinases, eventually lead to activation of a transcription factor. The core of the JNK pathway consist of the MAP2K (Hemipterous) (Glise et al., 1995; Sluss et al., 1996), which is upstream of the MAPK (Basket) (Riesgo-Escovar et al., 1996; Sluss et al., 1996), which in turn is responsible for the phosphorylation, and activation, of Jun in the AP-1 heterodimer transcription factor (Riesgo-Escovar et al., 1996; Sluss et al., 1996). Together the heterodimer of Jun, (Jra, Jun related antigen) and Fos (kayak), promote the expression of target genes. Examples of target genes are puckered, decapentapelgic (dpp), ance, chickadee (profilin), and hid. puckered encodes a phosphatase, which negatively regulates, or fine tune, the JNK pathway at the level of Basket, by removing the activating phosphate (Martin-Blanco et al., 1998). The influence of JNK on dpp expression is controlled by two mechanisms, by promoting the expression per se, but also by inhibiting the transcriptional repressor of anterior open, aop (Riesgo-Escovar and Hafen, 1997).
Many stimuli activate JNK signalling. As mentioned above, and as discussed in paper III, engagement of the cell surface receptor PGRP-LC activates the Imd pathway, and the components BG4, Tab2 and Iap2 ensure that the Tak1 (MAP3K) becomes activated. Tak1 can in turn phosphorylate Hemipterous, and thereby participate in JNK signalling (Mihaly et al., 2001; Takatsu et al., 2000).
After interactions between Eiger (the tumour necrosis factor) and Wengen (the TNF receptor), and through Misshapen (MAP4K), and Tak1 (MAP3K), JNK signalling is activated and induce apoptosis (Geuking et al., 2005; Igaki et al., 2002; Kanda et al., 2002; Mihaly et al., 2001; Moreno et al., 2002b).
PVR, a homologue to PGDF/VEGF receptor (platelet derived growth factor), is mainly involved in migratory and hematopoetic processes (Cho et al., 2002; Heino et al., 2001; Zettervall et al., 2004), and through the small GTPases Rac1/2 and possibly Cdc42, misshapen (MAP4K), and slipper (MAP3K), it can activate JNK (Ishimaru et al., 2004; Macias et al., 2004; Stronach and Perrimon, 2002).
JNK signalling participates in many processes; cytoskeletal rearrangements (F-actin polymerisation), apoptosis, or programmed cell death), and as mentioned it is also involved in regulating antimicrobial peptide production (Adachi-Yamada et al., 1999; Agnes et al., 1999;

29
Boutros et al., 2002; Jasper et al., 2001; Kallio et al., 2005; Kim et al., 2005; Takatsu et al., 2000).
Dpp/ BMP/ TGF-β
Decapentaplegic, or Dpp is a member of the bone morphogenic proteins, BMP, belonging to the TGF-β family. Decapentaplegic is in some contexts downstream of the JNK pathway, and in other contexts upstream. It is a multifaceted molecule with many functions and thus many components regulate its expression and impact. The name decapentaplegic, mean fifteen defects and reflects the many functions of the protein. The name refers to a series of mutant alleles affecting development and displaying defects in adult cuticular structures derived from one or more of the fifteen major imaginal discs (Spencer et al., 1982)
There are five different dpp transcripts, with different transcription start sites. In addition to Dpp there are other Drosophila BMPs; Glass bottom boat (Gbb), and screwdriver (Scw). Both are believed to potentiate Dpp signalling, but Gbb, unlike Scw, can also signal in absence of Dpp (Shimmi et al., 2005). (How Dpp signalling works, and how it implements its effects, is reviewed in (Affolter and Basler, 2007), and (O'Connor et al., 2006).)
BMPs are soluble, found as homo/heterodimers, and the transport efficiency as well as the capability to interact with their receptors, are modified by interactions with short gastrulation (Sog), tolloid (Tld), and twisted gastrulation (Tsg) (Eldar et al., 2002). BMPs signal by binding to class I, and class II cell surface receptor heterodimers, which are formed upon BMP binding. The class I receptors involved in BMP signalling are thickveins (tkv) and saxophone, and the class II receptors are punt, and wishful thinking, (Wit)
Interaction between Dpp and the receptors result in phosphorylation of Mad, (mothers against Dpp), and this allows Mad to interact with Medea. The Mad-Medea complex translocate into the nucleus, bind to enhancer regions and, together with Schnurri, counter-act the effect of the repressor Brinker, thus promoting Dpp-dependent transcription of genes (Torres-Vazquez et al., 2001; Torres-Vazquez et al., 2000). Examples of Dpp-regulated genes are bifid (Optomotor blind), vestigial, spalt major, and tinman.
The outcome of Dpp/BMP activation, depend on when, where and how. It is used as a patterning molecule, a molecule promoting cell survival and moderate proliferation, as well as processes involves in

30
cytoskeletal rearrangements like cell movement during dorsal and thorax closure (Martin-Blanco et al., 2000; Moreno et al., 2002a, Hou, 1997 #431; Zeitlinger et al., 1997).
Discontinous (spatially) or low levels of Dpp can induce apoptosis through the JNK pathway (Manjon et al., 2007), or exclusion and non-growth of these cells in the epithelia in question (Gibson and Perrimon, 2005; Shen and Dahmann, 2005). If however the apoptotic cells are kept alive, they produce Dpp and wingless, thus inducing increased growth of surrounding tissues (Ryoo et al., 2004).
Another group of proteins in the TGF-β family are the activins. In Drosophila this group consists of Activin−β and the activin-like protein Dawdle. Activins are TGF-β molecules and they signal through similar, and sometimes the same receptors as the BMPs. Baboon is a type I TGF-β /activin receptor, and Punt a class II receptor. Activation result in Sno-Medea-Smox interactions, which regulate gene expression and induces proliferation (Brummel et al., 1999; Takaesu et al., 2006).
Toll
The cell surface receptor Toll is an immunological icon. Not only is proper Toll signalling crucial during embryogenesis, which is how many of the components involved were isolated, (see introduction to the fly). Toll is also involved in the induction of some antimicrobial peptides, such as Drosomycin, it is necessary for an antifungal response (Lemaitre et al., 1996), and it controls proliferation of blood cell. Flies with a constitutive Toll signalling have increased numbers of blood cells or melanotic nodules (Gerttula et al., 1988; Huang et al., 2005; Qiu et al., 1998; Roth et al., 1991), and flies lacking the NF-κB transcription factors (Dif and Dorsal) in Toll signalling have very few (Matova and Anderson, 2006). The Toll signalling pathway has also been implicated in the defence against certain viruses (Zambon et al., 2005).
Two separate branches induced by microbial stimuli, activate Toll signalling. One branch is triggered by the bacterial cell wall component peptidoglycan, via the β-glucan recognition protein GNBP1, and the PGRPs, PGRP-SA, and PGRP-SD (Bischoff et al., 2004; Gobert et al., 2003; Leone et al., 2008; Michel et al., 2001; Pili-Floury et al., 2004). The other branch is also stimulated by microbial substances. Components of fungal cell wall, binds to the β-glucan recognition protein GNBP3 (Gottar et al., 2006). After stimulation of either branch, Toll signalling is activated through serine protease cascades (Gottar et al., 2006; Kambris et al., 2006), where eventually the Toll ligand spätzle is cleaved by spätzle

31
processing enzyme, SPE (Jang et al., 2006). The trimmed spätzle molecule can interact with Toll, and induce conformational changes, which promote signalling (Gangloff et al., 2008). (Reviewed (Anderson, 2000; Hultmark, 2003)). Upon activation the intracellular TIR domain (Toll/interleukin 1 receptor) containing part of Toll, interacts with the adaptor protein Myd88, which in turn interact with Tube and Pelle (a serine threonine kinase), through death domain interactions. This leads to the phosphorylation of the IκB by an unknown kinase, and the subsequent degradation of IκB by the proteasome. This in turn releases the inhibition of Dif/Dorsal and allows the dimer to enter the nucleus and activate transcription.
In addition to Toll there are eight more Toll like receptors, TLRs in the fruit fly (reviewed in (Leulier and Lemaitre, 2008)). Some of them are involved in the immune response. In humans there are ten toll-like receptors, TLRs, and in contrast to Drosphila Toll many of them interact directly with microbial structures in their functions in immunity.
Other biological processes
The making of a wing- ”Flyg fu la f luga f lyg!” 1
Already at the very beginning of this thesis, a fly was made, and now I want it to fly away, but in order for the fly to be able to fly, it has to have wings. How are they made, where do they come from?
The wing may look like a flat piece of gelatine leaf, but it is actually made from two apposed epidermal sheets covered by cuticular exoskeleton. This rather overly flappy material is supported by the veins, within which tracheae, nerves, haemolymph and blood cells may harbour.
Drosophila is a holometabolic animal. It means that seemingly the entire organism is disintegrated and remade during metamorphosis. You can’t make something out of nothing; this is true, also for holometabolic animals. Instead these animals are remade from dormant cells, put aside during embryogenesis. These sleeping beauties, imaginal cells, are often huddled together in highly organised imaginal discs, which are connected to the epithelia by a stalk. Eyes, legs, wings (approximately 50000 cells at late third instar), antennae, genitals and other structures, all have their
1 A swedish tounge twisting phrase ”Fly ugly fly, fly! -And the ugly fly flew.”

32
own imaginal discs. During metamorphosis, the cells in these discs will continue to proliferate, evert, spread and differentiate into the new body parts of the fly.
The wing disc, shaped like Sri Lanka, gives rise to wings and large parts of the thorax. Needless to say, it is complicated to make three-dimensional structures like wings and thorax from something flat and small. Looking at wing can give us insights into how signalling pathways interact, and it can help us to identify the molecular participants in signalling pathways. The wing can be smaller, larger, have chunks missing, have the wrong number of wing veins, or wrong spacing in between the veins, it all “means” something (Blair, 2007; Crozatier et al., 2004a; O'Connor et al., 2006). The longitudinal veins are formed more or less parallel to the anterior-posterior boundary, and the levels of hedgehog and Dpp provide critical information about the positioning of these veins.
A sketch, depicting a Drosophila wing. L1- L5 are the longitudinal veins, ACV is the anterior crossvein, PCV is the posterior crosswvein. And M is the marginal distal vein. A is anterior and P is posterior.

33
EGFR (epidermal growth factor receptor) is expressed early in the development of the longitudinal proveins, and it is believed to be necessary early for vein formation. Early proveins express EGFR targets, such as rhomboid and star, and simultaneously, loose the expression of the intervein marker DSRF (Drosophila serum response factor). Reduced or blocked EGFR signalling blocks vein formation in the adult. How EGFR signalling is localised to the provein region, and through which ligand (Spitz, Keren or Vein), is not clear. EGFR is vein reinforcing, by maintaining vein fate.
BMP (TGF-β) signalling is mediated by decapentaplegic or glass bottom boat. Dpp is expressed at an early time point as a stripe along the anterior posterior axis. Dpp levels influence the distance between L2– L3 and L4-L5 veins. Higher Dpp levels result in greater distances and lower levels decrease the distances. Dpp/BMP is also vein fate reinforcing. Loss of Dpp signalling in the longitudinal veins result in a short vein phenotype (de Celis, 1997; Ray and Wharton, 2001), and it is necessary for cross vein formation (Ralston and Blair, 2005). Ectopic Dpp expression can induce ectopic vein formation.
Hedgehog is a secreted growth factor and a functional homolog to the mammalian sonic hedgehog. It expressed in the posterior part of the wing disc as a consequence of engrailed expression. Hedgehog signalling is mediated through its receptors Patched and Smoothened. The L4 provein is formed by cells just posterior to the anterior-posterior boundary. The L3 vein is formed by cells receiving low hedgehog signals. High amplitude of the hedgehog signal promotes intervein specificity. These intervein cells signal to adjacent cells, which have received no or low hedgehog signal, to become provein cells. This means that cells receiving no hedgehog signal only can form veins if they are adjacent to cells, which received a high hedgehog signal. Collier is expressed in the L3-L4 intervein region in response to hedgehog signalling, and it inhibits vein formation (Vervoort et al., 1999). Collier probably works by repressing EGFR signalling and inducing DSRF in the intervein area. Increasing the hedgehog signal result in a greater distance between L3 and L4, whereas reducing the signal decreases this space.
Wingless is expressed along the dorsal ventral axis, or the future wing margin, the edge of the wing. This expression stems from Notch signalling at the boundary. Ectopic wingless signalling induces ectopic vein-like structures.
Notch signalling depends on its cell-bound ligands Delta and Serrate to organise the dorsal-ventral boundary of the wing. Delta is expressed in the ventral cells of the dorsal-ventral boundary, and Serrate is expressed in the dorsal cells. At the dorsal-ventral boundary, where

34
dorsal and ventral cells meet, the joint signalling of Serrate and Delta result in Notch dependent expression of Wingless.
Now we have successfully made a pair of wings (and thorax) with
all the veins in all the right places, with plenty of hairs or bristles covering it. Why all the hairs, you may wonder. They are sensory bristles providing the fly with data important for manoeuvring in the air. “-Och den fula flugan flög.”2
Salt stress
For a living cell to live well, the biochemical reactions that are a part of life need to work smoothly, and they do so at certain physical conditions (pH, ionic strength, and concentrations). At isotonic conditions the amount of osmolytes (solute particles like ions, or organic molecules) are optimal both within and outside the cell. If the osmolyte concentration outside the cell is to high (hypertonic), water will try to compensate for this difference and rush out of the cell, the cell will shrink and the concentration of particle/molecules within will increase. In the long run, hypertonicity can cause DNA damage, cell cycle arrest, or cell death through apoptosis (reviewed in (Ho, 2006)).
The osmolyte concentration surrounding, compared to within, an organism are often dissimilar (most often hypotonic), furthermore it may change. Thus the organism needs to be capable to maintain water homeostasis by responding to these changes. And they do so by an altered uptake/ production/ release of ions or non-ionic compounds, osmolytes. For this purpose, multicellular organisms have developed different organs, like our kidneys and the malphigian tubes of the insect, designed to secrete nitrogenous molecules, ions and toxic compounds (Chapman, 1998). By these means they can control the body fluid surrounding most cells, and provide a stable and favourable milieu. This is more difficult for single cell organisms, like the yeast Saccharomyces cerevisiae, which is totally exposed to the environment, and which has been extensively studied in its response to hypertonic stress.
2 A swedish tounge twisting phrase ”Fly ugly fly, fly! -And the ugly fly flew.”

35
In mammals, hypertonicity elicits phosphorylation and activation
of the transcription factor NFAT5. (Reviewed in (Aramburu et al., 2006; Ho, 2006; Jeon et al., 2006).) NFAT5 is a REL protein, and as such it belongs to the same family of proteins as Relish and p105 (signalling pathways-Imd/Relish). Like Relish, NFAT5 function as a dimer, and after activation it enters the nucleus and facilitates the transcription of target genes by binding to its specific DNA motif. The proteins coded by these genes help counter balance the efflux of water by increasing non-ionic osmolytes like sorbitol, myoinositol and taurine, thus luring water back into the cell. The importance of NFAT5 in response to hypertonicity in kidney function is evident. Mice with reduced NFAT function have renal problems, problems in concentrating urine, and the mutant animals die due to atrophy of kidney medulla. NFAT also play a role in response to hypertonicity in other regions than the kidney. NFAT

36
mRNA ubiquitously expressed, but the protein is enriched in thymus and activated lymphocytes. Animals with reduced NFAT function and hyperosmotic stress have lower lymphocyte numbers and T-lymphoid growth and survival; further more an impaired T-cell antibody response upon immunization. NFAT5 is also abundantly expressed in the brain, and its expression in neurons increases upon hypernatremia (high sodium levels) (Loyher et al., 2004).
Kidney development relies on series of reciprocal inductive signals, and the activin – follistatin system, play an important part in this, as well as in damage repair. This is reviewed in (Maeshima et al., 2008).
In Drosophila the malphigian tubules are the functional equivalents of the kidney. The primary urine is the result of the combined effort of both the principal malphigian cells and the stellate cells of the malphigian tubules. The recruitment of these cell types, the development of the malphigian tublues and the signalling events and molecules involved, share surprisingly many similarities with the development of the mammalian kidney (Jung et al., 2005a). But the malphigian tublues do not work alone in maintaining osmolyte homeostasis. The anal pads, is another (larval) structure involved in regulating homeostasis, by importing ions from the surrounding into the haemolymph.
The Drosophila genome encodes one NFAT, and in paper V (Keyser et al., 2007) we describe the characterisation of the Drosophila NFAT gene, and show that this Rel-protein, similar to mammalian NFAT5, is involved in the response to osmotic stress.

37
Aim of this thesis
The intention of my PhD project, has been to study the signalling
events, revolving around a recently dicovered gene family of pattern recognition receptors, the PGRPs, and in particular the transmembrane receptor PGRP-LC. I have also been involved in trying to understand the Rel protein NFAT.
Results and discussion
Paper I
In this paper we describe the lay-out of the PGRP domain rich, PGRP-LC locus. We found that PGRP-LC indeed, by alternative splicing, have the capability of utilising three of these five different PGRP domains. For these three variants, PGRP-LCx, PGRP-LCy, and PGRP-LCa, the intracellular and transmembrane domain is the same, but onto it different peptidoglycan recognition domains are fused. By specifically knocking down the expression of the different splice forms in a cell line, we addressed their function and their capability to induce the antimicrobial peptides expression, in response to various stimuli.
We found that the different splice forms indeed have different epitope specificity. PGRP-LCx was capable of inducing antimicrobial production, as a consequence of binding to peptidoglycan (DAP-type). PGRP-LCa displayed a different recognition pattern. We found that PGRP-LCa was capable of detecting LPS, this was highly surprising, and being aware of the possibility of contaminated compounds, we carefully investigated this hypothesis, however our conclusions were unchanged.
An interesting twist was the later discovery that PGRP-LCa recognises a peptidoglycan fragment found in the commercial LPS preparation (Kaneko et al., 2004). Later experiments revealed that PGRP-LCa binds to a peptidoglycan fragment also known as tracheal cytotoxin, (TCT) after the peptidoglycan fragment already has bound by PGRP-LCx. PGRP-LCx then present the specific PGRP-LCa epitope to PGRP-LCa. This reveals that the basic idea with different recognition epitopes still is true.
We found no effect of reducing PGRP-LCy levels. The function of the protein remains an enigma. Does the y-splice form have a function, or is merely a waste product of evolution. There are of course an enormous set of compounds from microbes one could try to see if

38
PGRP-LCy binds to them, but without any particular idea, it seems futile. The abundance of PGRP-LCy mRNA is much lower than that of its splice siblings PGRP-LCx and PGRP-LCa. One idea that I had was that it was more abundant in haemocytes than in other tissues, and some real time PCR data do indicate this, but the amounts of PGRP-LCy compared to the others splice variants are still miniscule. PGRP-LCx is clearly the most abundant of them all.
The remaining two PGRP domains make up a separate gene, PGRP-LF, making it the only PGRP with two PGRP domains. We also noted that reducing the amounts of PGRL-LF, resulted in an increased induction of antimicrobial peptides upon infection. This suggested that PGRP-LF could act as an inhibitor.
This dampening effect of PGRP-LF has been observed again (Maillet et al., 2008; Persson et al., 2007). Persson et al. 2007, also demonstrated that both PGRP domains are capable of interacting with peptidoglycan. Why would the fly need an inhibitor of the Imd/Relish pathway that work on the outside, and one that binds to peptidoglycan? It seems like a much smarter idea to control the signalling pathway from within, as is the case with for instance Pirk. One possibility is if PGRP-LF bind to, and hamper, PGRP-LC in the absence of peptidoglycan, and when the animal is infected with bacteria, peptidoglycan bind to both PGRP-LC and PGRP-LF, thus releasing PGRP-LC from its inhibitor.
If PGRP-LF is a true negative regulator, it belongs to an increasing group of identified proteins, which are there to keep the actions of the Imd/Relish signalling pathway in place, or locked up, when no pathogens are around. Examples of negative regulators of the Imd/relish pathway are Dnr1, Pirk, and Caspar (Foley and O'Farrell, 2004; Kim et al., 2006; Kleino et al., 2008).
Paper II
Paper II focuses on three new inducible antimicrobial peptides, Diptericin B, Attacin C and Attacin D, and how studying them, shed more light on how different antimicrobial peptides might be related to one another. Analyses of their sequences add strength to the idea that these glycine-rich antimicrobial peptides have evolved from a common ancestor and are probably also related to proline-rich peptides such as drosocin
This idea has gained extra support by the fact that a processed form of the N-terminal pro domain of Attacin C has been isolated in haemolymph, and that its synthetic counterpart displays activity against bacteria (Rabel et al., 2004).

39
Attacin D lacks a signal peptide and appears to be an intracellular peptide. It would be interesting to see if it has any effect on Gram- negative intracellular bacteria, like Francisella tularensis.
The lack of interest from the scientific community, on the actual mechanisms of antimicrobial peptides, is hard for me to understand. Both from a curiosity point of view, and from a public health point of view. The potential knowledge of the mechanisms behind different antimicrobial peptides function could be useful in the making of new replacements for the decreasing list of effective antibiotics. Instead the focus seem be on adding one more antimicrobial peptide to the increasing list. Maybe I should have faith in the second wave of interest, which normally comes about after a slightly stagnant period when it was believed that everything was known.
Result s and di scuss ion paper III
In paper III, we have analysed the role of PGRP-LC in controlling blood cell number and activation. When PGRP-LCx is reintroduced into a mutant background, blood cell numbers increase from a slightly repressed level. Overexpressing PGRP-LCx within blood cells, using blood cell Gal4 drivers, has substantial effects on cell number, ratio of lamellocytes, size of the lymph gland, and also affects the banded pattern of sessile cells. This blood cell activation is dependent on the JNK signalling cascade. When we suppress the JNK cascade, by over-expressing a negative regulator, or by knocking down JNK components, blood cell numbers are reduced, as is the lamellocyte fraction. However Relish does not seem to be necessary for the proliferative and activating effect of PGRP-LCx in haemocytes, since overexpressing PGRP-LCx in a Relish mutant background barely affects the haemocyte-, sessile band-, lymph gland- phenotypes achieved by PGRP-LCx overexpression.
This puts a pathogen recognition receptor upstream of the cellular events that do take place during an immune response. Events that do need JNK signalling. One blood cell function that PGRP-LC has been suggested to be involved with is the phagocytosis of bacteria. Based on my own experiments (done in context of paper I), and the mild effects seen in published work (Rämet et al., 2002b), I have regarded PGRP-LCs involvement in phagocytosis as a side effect of it being a peptidoglycan binding protein. Allowing the bacteria to stay close for a longer period of time, thus increasing the chances of engaging the phagocytic machinery. The results of paper III and IV, with the cytoskeletal phenotypes observed has made me slightly more inclined to believe that PGRP-LC may be involved in phagocytosis.

40
Result s and di scuss ion paper IV
In paper IV we dig deeper into the consequences of PGRP-LC signalling, and we relocate (in part) the caspase Dredd, to the JNK pathway. PGRP-LC mutants have mild wing defects, such that the posterior cross veins are not complete, and they also display slight abdominal fusion problems. An incomplete posterior cross vein is a common phenotype for animals with reduced Dpp levels. These mild effects were reduced when PGRP-LCx was reintroduced into the mutant background. If however the PGRP-LC1 mutation were combined with either imd or Dredd mutants, drastic effects on wing shape could be seen. Wings from these animals are missing large portions along the anterior and posterior borders. Imd; PGRP-LC1 double mutants also have additional problems, with very poor survival, long generation time, and crooked legs. Many die as pharate adults inside the pupal case, with portions of the cuticle missing. Another common defect is aberrant abdominal fusion, and some displayed thorax closure problems. These are problems associated with JNK and/or Dpp signalling defects. Other indicative evidence also suggests that PGRP-LC may contribute, in a small way, in dpp expression. These indications should also be tested by other means, like more thorough genetic interaction studies, and by measuring Dpp levels or activity.
Forced expression of PGRP-LCx in wings and thorax using the Pannier- or apterous- Gal4 drivers, resulted in similar phenotypes as seen with JNK and Dpp expression. The phenotypes observed included overactive thorax closure, thicker bristles, wrong bristle type and positioning, and apoptosis in wings.
Forced expression of PGRP-LCx in haemocytes increased haemocyte number and induced lamellocyte formation, as discussed in paper III, and it also increased cell size, induced JNK activity, as well as induced F-actin formation (as detected in plasmatocytes by immunohistochemistry). All of these overexpression phenotypes were reduced or lost, when the experiment was performed in a dredd mutant background. Evidently Dredd must also participate in the JNK signalling pathway, as well as cleaving and activating Relish. But where? Based on experiments in a cell line by Zhou et al., 2005 (Zhou et al., 2005) these authors suggested that Dredd contribute to JNK signalling together with Imd, BG4, UEVa1 and Bendless by activating Tak1 (Zhou et al., 2005). dredd mutants do not only block the effect of overexpression PGRP-LC, but also that of Imd, and Basket overexpression as well, and dredd mutants reduce the amount of active Basket. This suggests that Dredd is involved in JNK signalling at more than one level, both upstream and

41
downstream of basket, or maybe parallel to basket. Alternatively that Dredd in addition to its role in Relish activation and its new role in the JNK pathway also have additional strings on its bow, What if one of these strings is an F-actin string and Dredd is involved in regulating its polymerisation? The great increase of F-actin formation in response to PGRP-LC is blocked in Dredd mutants.
I find Dredd and its possible functions and interaction partners intriguing. Do all aspects to Dredds functions need an active caspase, or do some aspects merely require a scaffold function. Dredd has recently been linked with RING finger domain proteins like, Dnr1 and Iap2 (Foley and O'Farrell, 2004; Huh et al., 2007; Leulier et al., 2006). Also I am very interested in RING finger domain contain proteins, like Iap1 (thread), and their involvement in cytoskeletal rearrangements. Iap1 and 2 are previously known as apoptotic inhibitors, but have recently been linked to actin polymerisation and immunity respectively (Geisbrecht and Montell, 2004; Huh et al., 2007; Kleino et al., 2005; Leulier et al., 2006; Oshima et al., 2006kg; Valanne et al., 2007).
Result s and di scuss ion paper V
In this paper we characterised the Drosophila NFAT gene, by creating and exposing mutants to various treatments, forced expression of the gene, and by expression pattern analyses. The NFAT gene is a member of the REL-family, and it is a homologue to the mammalian NFATs. Contrary to our initial hopes there was no obvious immune related phenotype of NFAT mutants. However complete null mutants for NFAT die faster than wild type animals, if put on a high salt diet (hyper tonic), and NFAT mutant larval anal pads swell if the larva is submerged in hypotonic solution. Drosophila NFAT appear to be important to maintain salt homeostasis, as is the mammalian NFAT5, and is not obviously involved in the immune response.
NFAT has been noted as a hit in many overexpression screens based on different organs/cell types and involving many signalling pathways (Abdelilah-Seyfried et al., 2000; Buszczak et al., 2007; Kraut et al., 2001; Mukherjee et al., 2006; Pena-Rangel et al., 2002; Singh et al., 2005; Zhu et al., 2005). In my eyes, one signalling pathway emerges out from the fog of signalling components, and that is the involvement of TGF-β signalling components. NFAT overexpression display similar phenotypes as proteins associated with TGF-β/ Dpp signalling pathways, in all but one (of eight) screens. In the last case the phenotype observed was enhanced by NFAT (Mukherjee et al., 2006), but they also discuss

42
that this phenotype had in previous studies been suppressed by hypomorphs of the Dpp signalling.
Why has this attracted my attention? Baboon, is the activin class I receptor in Drosophila, and similar to NFAT baboon mutants display enlarged anal pads (Brummel et al., 1999). However they do so regardless of the tonicity of the environment. In addition to this, the activin system has a proliferative and photoreceptor axon-targeting role in the brain (Brummel et al., 1999; Takaesu et al., 2006; Zhu et al., 2008). In mammals the Activin-Follistatin system is important for proper development of the kidney, and both molecules are induced upon damage. The development of the kidney and malpighian tubules display many similarities (Jung et al., 2005a).
NFAT5, in addition to its importance in kidneys, also appear to be important in the brain, and the expression levels here are relatively high. This puts NFAT5 and baboon in different animals in similar tissues, at slightly similar conditions.
It would be interesting if there is a link between NFAT in Drosophila and activin signalling. As discussed in Paper V, in the attempts to unravel the function of NFAT, a microarray on NFAT overexpressing animals, as well as Nfat mutants, was carried out. These data were inconclusive, and none of the known homologues to NFAT5 target genes were hits. The micro-array might have been more informative if it had been performed under different parameters (after hypertonic stress or using different Gal4 drivers), but it is also noteworthy that activin B was not on the list of genes which the microarray was able to detect.

43
Acknowledgements -Aaahhmmm, thank you very much. So I haven’t prepared a speech, but thank you. -Who do I want to thank? Jag vill tacka min familj, mamma, pappa och Per, Karin, Ingemar, Björn, Lina och lilla Vera. Tack! -I want to thank the producer and director, the conservative rebel Dan, for excellent casting of the crew, and for support and guidance during the more difficult/challenging shoots. And for spreading a curious mentality. -I want to thank the rest of the crew, my co-actors, for an inspiring time and a great atmosphere during and in-between shoots. Anna-Karin, Calle, Ines, Ingrid, Jens-Ola, Jesper, Karin, Magda-Lena, Malin, Martin, Marika, Mazen, Mike, Monica, Pia, Shannon, Sofia, Svenja, Sören, Thomas. -Who else? -I want to thank the rest of the crew on location, both extras and permanent members, in our production team and in other productions teams. -I want to thank our coordinator of everything on location Maria. Tack Erik, du är den manlige huvudrolls innehavaren i mitt liv. Tack för allt ditt stöd. Jag älskar dig. Dina och Frank jag älskar er mina go-ungar. Ni är bäst.

44
References Abdelilah-Seyfried, S., Chan, Y. M., Zeng, C., Justice, N. J., Younger-Shepherd, S., Sharp, L. E., Barbel, S., Meadows, S. A., Jan, L. Y. and Jan, Y. N. (2000). A gain-of-function screen for genes that affect the development of the Drosophila adult external sensory organ. Genetics 155, 733-52. Adachi-Yamada, T., Gotoh, T., Sugimura, I., Tateno, M., Nishida, Y., Onuki, T. and Date, H. (1999). De novo synthesis of sphingolipids is required for cell survival by down-regulating c-Jun N-terminal kinase in Drosophila imaginal discs. Mol Cell Biol 19, 7276-86. Affolter, M. and Basler, K. (2007). The Decapentaplegic morphogen gradient: from pattern formation to growth regulation. Nat Rev Genet 8, 663-74. Agaisse, H., Petersen, U. M., Boutros, M., Mathey-Prevot, B. and Perrimon, N. (2003). Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury. Dev Cell 5, 441-50. Agnes, F., Suzanne, M. and Noselli, S. (1999). The Drosophila JNK pathway controls the morphogenesis of imaginal discs during metamorphosis. Development 126, 5453-62. Alfonso, T. B. and Jones, B. W. (2002). gcm2 promotes glial cell differentiation and is required with glial cells missing for macrophage development in Drosophila Dev Biol 248, 369-83. Anderson, K. V. (2000). Toll signaling pathways in the innate immune response. Curr Opin Immunol 12, 13-9. Anderson, K. V., Jurgens, G. and Nusslein-Volhard, C. (1985). Establishment of dorsal-ventral polarity in the Drosophila embryo: genetic studies on the role of the Toll gene product. Cell 42, 779-89. Aramburu, J., Drews-Elger, K., Estrada-Gelonch, A., Minguillon, J., Morancho, B., Santiago, V. and Lopez-Rodriguez, C. (2006). Regulation of the hypertonic stress response and other cellular functions by the Rel-like transcription factor NFAT5. Biochem Pharmacol 72, 1597-604. Basset, A., Khush, R. S., Braun, A., Gardan, L., Boccard, F., Hoffmann, J. A. and Lemaitre, B. (2000). The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an immune response. Proc Natl Acad Sci U S A 97, 3376-81. Bettencourt, R., Asha, H., Dearolf, C. and Ip, Y. T. (2004). Hemolymph-dependent and -independent responses in Drosophila immune tissue. J Cell Biochem 92, 849-63.

45
Bidla, G., Dushay, M. S. and Theopold, U. (2007). Crystal cell rupture after injury in Drosophila requires the JNK pathway, small GTPases and the TNF homolog Eiger. J Cell Sci 120, 1209-15. Bischoff, V., Vignal, C., Boneca, I. G., Michel, T., Hoffmann, J. A. and Royet, J. (2004). Function of the Drosophila pattern-recognition receptor PGRP-SD in the detection of Gram-positive bacteria. Nat Immunol 5, 1175-80. Bischoff, V., Vignal, C., Duvic, B., Boneca, I. G., Hoffmann, J. A. and Royet, J. (2006). Downregulation of the Drosophila immune response by peptidoglycan-recognition proteins SC1 and SC2. PLoS Pathog 2, e14. Blair, S. S. (2007). Wing vein patterning in Drosophila and the analysis of intercellular signaling. Annu Rev Cell Dev Biol 23, 293-319. Bosch, M., Serras, F., Martin-Blanco, E. and Baguna, J. (2005). JNK signaling pathway required for wound healing in regenerating Drosophila wing imaginal discs. Dev Biol 280, 73-86. Boutros, M., Agaisse, H. and Perrimon, N. (2002). Sequential activation of signaling pathways during innate immune responses in Drosophila Dev Cell 3, 711-22. Braun, A., Hoffmann, J. A. and Meister, M. (1998). Analysis of the Drosophila host defense in domino mutant larvae, which are devoid of hemocytes. Proc Natl Acad Sci U S A 95, 14337-42. Brennan, C. A., Delaney, J. R., Schneider, D. S. and Anderson, K. V. (2007). Psidin is required in Drosophila blood cells for both phagocytic degradation and immune activation of the fat body. Curr Biol 17, 67-72. Brey, P. T., Lee, W. J., Yamakawa, M., Koizumi, Y., Perrot, S., Francois, M. and Ashida, M. (1993). Role of the integument in insect immunity: epicuticular abrasion and induction of cecropin synthesis in cuticular epithelial cells. Proc Natl Acad Sci U S A 90, 6275-9. Brock, T., D., Madigan, M, T, Martinko, J, M., and Parker, J. (1994). Biology of microorganisms. New Jersey: Prentice Hall. Brummel, T., Abdollah, S., Haerry, T. E., Shimell, M. J., Merriam, J., Raftery, L., Wrana, J. L. and O'Connor, M. B. (1999). The Drosophila activin receptor baboon signals through dSmad2 and controls cell proliferation but not patterning during larval development. Genes Dev 13, 98-111. Bulet, P., Stocklin, R. and Menin, L. (2004). Anti-microbial peptides: from invertebrates to vertebrates. Immunol Rev 198, 169-84. Buszczak, M., Paterno, S., Lighthouse, D., Bachman, J., Planck, J., Owen, S., Skora, A. D., Nystul, T. G., Ohlstein, B., Allen, A. et al. (2007). The carnegie protein trap library: a versatile tool for Drosophila developmental studies. Genetics 175, 1505-31.

46
Carlsson, A., Engström, P., Palva, E. T. and Bennich, H. (1991). Attacin, an antibacterial protein from Hyalophora cecropia, inhibits synthesis of outer membrane proteins in Escherichia coli by interfering with omp gene transcription. Infect Immun 59, 3040-5. Carlsson, A., Nystrom, T., de Cock, H. and Bennich, H. (1998). Attacin--an insect immune protein--binds LPS and triggers the specific inhibition of bacterial outer-membrane protein synthesis. Microbiology 144 ( Pt 8), 2179-88. Castillejo-Lopez, C. and Hacker, U. (2005). The serine protease Sp7 is expressed in blood cells and regulates the melanization reaction in Drosophila Biochem Biophys Res Commun 338, 1075-82. Cerenius, L., Lee, B. L. and Söderhäll, K. (2008). The proPO-system: pros and cons for its role in invertebrate immunity. Trends Immunol. Cerenius, L. and Söderhäll, K. (2004). The prophenoloxidase-activating system in invertebrates. Immunol Rev 198, 116-26. Chang, C. I., Chelliah, Y., Borek, D., Mengin-Lecreulx, D. and Deisenhofer, J. (2006). Structure of tracheal cytotoxin in complex with a heterodimeric pattern-recognition receptor. Science 311, 1761-4. Chang, C. I., Pili-Floury, S., Herve, M., Parquet, C., Chelliah, Y., Lemaitre, B., Mengin-Lecreulx, D. and Deisenhofer, J. (2004). A Drosophila pattern recognition receptor contains a peptidoglycan docking groove and unusual L,D-carboxypeptidase activity. PLoS Biol 2, E277. Chapman, R. F. (1998). Excretion and salt and water regulation. In The insects- Structure and function, pp. 478-508. Cambridge: Cambridge University Press. Cho, N. K., Keyes, L., Johnson, E., Heller, J., Ryner, L., Karim, F. and Krasnow, M. A. (2002). Developmental control of blood cell migration by the Drosophila VEGF pathway. Cell 108, 865-76. Choe, K. M., Lee, H. and Anderson, K. V. (2005). Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc Natl Acad Sci U S A 102, 1122-6. Choe, K. M., Werner, T., Stöven, S., Hultmark, D. and Anderson, K. V. (2002). Requirement for a peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in Drosophila. Science 296, 359-62. Christensen, B. M., Li, J., Chen, C. C. and Nappi, A. J. (2005). Melanization immune responses in mosquito vectors. Trends Parasitol 21, 192-9. Cociancich, S., Bulet, P., Hetru, C. and Hoffmann, J. A. (1994). The inducible antibacterial peptides of insects. Parasitol Today 10, 132-9.

47
Crozatier, M., Glise, B. and Vincent, A. (2004a). Patterns in evolution: veins of the Drosophila wing. Trends Genet 20, 498-505. Crozatier, M., Ubeda, J. M., Vincent, A. and Meister, M. (2004b). Cellular immune response to parasitization in Drosophila requires the EBF orthologue collier. PLoS Biol 2, E196. de Celis, J. F. (1997). Expression and function of decapentaplegic and thick veins during the differentiation of the veins in the Drosophila wing. Development 124, 1007-18. De Gregorio, E., Han, S. J., Lee, W. J., Baek, M. J., Osaki, T., Kawabata, S., Lee, B. L., Iwanaga, S., Lemaitre, B. and Brey, P. T. (2002a). An immune-responsive Serpin regulates the melanization cascade in Drosophila. Dev Cell 3, 581-92. De Gregorio, E., Spellman, P. T., Tzou, P., Rubin, G. M. and Lemaitre, B. (2002b). The Toll and Imd pathways are the major regulators of the immune response in Drosophila. Embo J 21, 2568-79. Delaney, J. R., Stöven, S., Uvell, H., Anderson, K. V., Engström, Y. and Mlodzik, M. (2006). Cooperative control of Drosophila immune responses by the JNK and NF-kappaB signaling pathways. Embo J 25, 3068-77. Dijkers, P. F. and O'Farrell, P. H. (2007). Drosophila calcineurin promotes induction of innate immune responses. Curr Biol 17, 2087-93. Drier, E. A. and Steward, R. (1997). The dorsoventral signal transduction pathway and the Rel-like transcription factors in Drosophila. Semin Cancer Biol 8, 83-92. Dunn, P. E., Dai, W., Kanost, M. R. and Geng, C. X. (1985). Soluble peptidoglycan fragments stimulate antibacterial protein synthesis by fat body from larvae of Manduca sexta. Dev Comp Immunol 9, 559-68. Dushay, M. S., Roethele, J. B., Chaverri, J. M., Dulek, D. E., Syed, S. K., Kitami, T. and Eldon, E. D. (2000). Two attacin antibacterial genes of Drosophila melanogaster. Gene 246, 49-57. Dushay, M. S., Åsling, B. and Hultmark, D. (1996). Origins of immunity: Relish, a compound Rel-like gene in the antibacterial defense of Drosophila. Proc Natl Acad Sci U S A 93, 10343-7. Dziarski, R. and Gupta, D. (2006a). Mammalian PGRPs: novel antibacterial proteins. Cell Microbiol 8, 1059-69. Dziarski, R. and Gupta, D. (2006b). The peptidoglycan recognition proteins (PGRPs). Genome Biol 7, 232. Ekengren, S. and Hultmark, D. (1999). Drosophila cecropin as an antifungal agent. Insect Biochem Mol Biol 29, 965-72. Eldar, A., Dorfman, R., Weiss, D., Ashe, H., Shilo, B. Z. and Barkai, N. (2002). Robustness of the BMP morphogen gradient in Drosophila embryonic patterning. Nature 419, 304-8.

48
Elrod-Erickson, M., Mishra, S. and Schneider, D. (2000). Interactions between the cellular and humoral immune responses in Drosophila. Curr Biol 10, 781-4. Engström, P., Carlsson, A., Engström, A., Tao, Z. J. and Bennich, H. (1984). The antibacterial effect of attacins from the silk moth Hyalophora cecropia is directed against the outer membrane of Escherichia coli. Embo J 3, 3347-51. Evans, C. J., Hartenstein, V. and Banerjee, U. (2003). Thicker than blood: conserved mechanisms in Drosophila and vertebrate hematopoiesis. Dev Cell 5, 673-90. Foley, E. and O'Farrell, P. H. (2003). Nitric oxide contributes to induction of innate immune responses to gram-negative bacteria in Drosophila. Genes Dev 17, 115-25. Foley, E. and O'Farrell, P. H. (2004). Functional dissection of an innate immune response by a genome-wide RNAi screen. PLoS Biol 2, E203. Fossett, N., Hyman, K., Gajewski, K., Orkin, S. H. and Schulz, R. A. (2003). Combinatorial interactions of serpent, lozenge, and U-shaped regulate crystal cell lineage commitment during Drosophila hematopoiesis. Proc Natl Acad Sci U S A 100, 11451-6. Franc, N. C., Dimarcq, J. L., Lagueux, M., Hoffmann, J. and Ezekowitz, R. A. (1996). Croquemort, a novel Drosophila hemocyte/macrophage receptor that recognizes apoptotic cells. Immunity 4, 431-43. Freeman, M. R., Delrow, J., Kim, J., Johnson, E. and Doe, C. Q. (2003). Unwrapping glial biology: Gcm target genes regulating glial development, diversification, and function. Neuron 38, 567-80. Galko, M. J. and Krasnow, M. A. (2004). Cellular and genetic analysis of wound healing in Drosophila larvae. PLoS Biol 2, E239. Gangloff, M., Murali, A., Xiong, J., Arnot, C. J., Weber, A. N., Sandercock, A. M., Robinson, C. V., Sarisky, R., Holzenburg, A., Kao, C. et al. (2008). Structural insight into the mechanism of activation of the toll receptor by the dimeric ligand spatzle. J Biol Chem 283, 14629-35. Geisbrecht, E. R. and Montell, D. J. (2004). A role for Drosophila IAP1-mediated caspase inhibition in Rac-dependent cell migration. Cell 118, 111-25. Georgel, P., Naitza, S., Kappler, C., Ferrandon, D., Zachary, D., Swimmer, C., Kopczynski, C., Duyk, G., Reichhart, J. M. and Hoffmann, J. A. (2001). Drosophila immune deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis. Dev Cell 1, 503-14. Gerttula, S., Jin, Y. S. and Anderson, K. V. (1988). Zygotic expression and activity of the Drosophila Toll gene, a gene required maternally for embryonic dorsal-ventral pattern formation. Genetics 119, 123-33.

49
Gesellchen, V., Kuttenkeuler, D., Steckel, M., Pelte, N. and Boutros, M. (2005). An RNA interference screen identifies Inhibitor of Apoptosis Protein 2 as a regulator of innate immune signalling in Drosophila. EMBO Rep 6, 979-84. Geuking, P., Narasimamurthy, R. and Basler, K. (2005). A genetic screen targeting the tumor necrosis factor/Eiger signaling pathway: identification of Drosophila TAB2 as a functionally conserved component. Genetics 171, 1683-94. Gibson, M. C. and Perrimon, N. (2005). Extrusion and death of DPP/BMP-compromised epithelial cells in the developing Drosophila wing. Science 307, 1785-9. Glise, B., Bourbon, H. and Noselli, S. (1995). hemipterous encodes a novel Drosophila MAP kinase kinase, required for epithelial cell sheet movement. Cell 83, 451-61. Gobert, V., Gottar, M., Matskevich, A. A., Rutschmann, S., Royet, J., Belvin, M., Hoffmann, J. A. and Ferrandon, D. (2003). Dual activation of the Drosophila toll pathway by two pattern recognition receptors. Science 302, 2126-30. Goto, A., Kadowaki, T. and Kitagawa, Y. (2003). Drosophila hemolectin gene is expressed in embryonic and larval hemocytes and its knock down causes bleeding defects. Dev Biol 264, 582-91. Goto, A., Kumagai, T., Kumagai, C., Hirose, J., Narita, H., Mori, H., Kadowaki, T., Beck, K. and Kitagawa, Y. (2001). A Drosophila haemocyte-specific protein, hemolectin, similar to human von Willebrand factor. Biochem J 359, 99-108. Gottar, M., Gobert, V., Matskevich, A. A., Reichhart, J. M., Wang, C., Butt, T. M., Belvin, M., Hoffmann, J. A. and Ferrandon, D. (2006). Dual detection of fungal infections in Drosophila via recognition of glucans and sensing of virulence factors. Cell 127, 1425-37. Gottar, M., Gobert, V., Michel, T., Belvin, M., Duyk, G., Hoffmann, J. A., Ferrandon, D. and Royet, J. (2002). The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 416, 640-4. Hammond, S. M., Bernstein, E., Beach, D. and Hannon, G. J. (2000). An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 404, 293-6. Harden, N. (2002). Signaling pathways directing the movement and fusion of epithelial sheets: lessons from dorsal closure in Drosophila. Differentiation 70, 181-203. Harden, N. (2005). Cell biology. Of grainy heads and broken skins. Science 308, 364-5. Hedengren, M., Borge, K. and Hultmark, D. (2000). Expression and evolution of the Drosophila attacin/diptericin gene family. Biochem Biophys Res Commun 279, 574-81.

50
Hedengren, M., Åsling, B., Dushay, M. S., Ando, I., Ekengren, S., Wihlborg, M. and Hultmark, D. (1999). Relish, a central factor in the control of humoral but not cellular immunity in Drosophila. Mol Cell 4, 827-37. Heino, T. I., Karpanen, T., Wahlstrom, G., Pulkkinen, M., Eriksson, U., Alitalo, K. and Roos, C. (2001). The Drosophila VEGF receptor homolog is expressed in hemocytes. Mech Dev 109, 69-77. Ho, S. N. (2006). Intracellular water homeostasis and the mammalian cellular osmotic stress response. J Cell Physiol 206, 9-15. Holz, A., Bossinger, B., Strasser, T., Janning, W. and Klapper, R. (2003). The two origins of hemocytes in Drosophila. Development 130, 4955-62. Hu, S. and Yang, X. (2000). dFADD, a novel death domain-containing adapter protein for the Drosophila caspase DREDD. J Biol Chem 275, 30761-4. Huang, L., Ohsako, S. and Tanda, S. (2005). The lesswright mutation activates Rel-related proteins, leading to overproduction of larval hemocytes in Drosophila melanogaster. Dev Biol 280, 407-20. Huh, J. R., Foe, I., Muro, I., Chen, C. H., Seol, J. H., Yoo, S. J., Guo, M., Park, J. M. and Hay, B. A. (2007). The Drosophila inhibitor of apoptosis (IAP) DIAP2 is dispensable for cell survival, required for the innate immune response to gram-negative bacterial infection, and can be negatively regulated by the reaper/hid/grim family of IAP-binding apoptosis inducers. J Biol Chem 282, 2056-68. Hultmark, D. (2003). Drosophila immunity: paths and patterns. Curr Opin Immunol 15, 12-9. Hultmark, D., Engström, A., Andersson, K., Steiner, H., Bennich, H. and Boman, H. G. (1983). Insect immunity. Attacins, a family of antibacterial proteins from Hyalophora cecropia. Embo J 2, 571-6. Hultmark, D., Steiner, H., Rasmuson, T. and Boman, H. G. (1980). Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur J Biochem 106, 7-16. Igaki, T., Kanda, H., Yamamoto-Goto, Y., Kanuka, H., Kuranaga, E., Aigaki, T. and Miura, M. (2002). Eiger, a TNF superfamily ligand that triggers the Drosophila JNK pathway. Embo J 21, 3009-18. Ip, Y. T., Reach, M., Engström, Y., Kadalayil, L., Cai, H., Gonzalez-Crespo, S., Tatei, K. and Levine, M. (1993). Dif, a dorsal-related gene that mediates an immune response in Drosophila. Cell 75, 753-63. Ishimaru, S., Ueda, R., Hinohara, Y., Ohtani, M. and Hanafusa, H. (2004). PVR plays a critical role via JNK activation in thorax closure during Drosophila metamorphosis. Embo J 23, 3984-94.

51
Janeway, C. A., Jr. (1989). Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol 54 Pt 1, 1-13. Jang, I. H., Chosa, N., Kim, S. H., Nam, H. J., Lemaitre, B., Ochiai, M., Kambris, Z., Brun, S., Hashimoto, C., Ashida, M. et al. (2006). A Spatzle-processing enzyme required for toll signaling activation in Drosophila innate immunity. Dev Cell 10, 45-55. Jasper, H., Benes, V., Schwager, C., Sauer, S., Clauder-Münster, S., Ansorge, W. and Bohmann, D. (2001). The genomic response of the Drosophila embryo to JNK signaling. Dev. Cell 1, 579-86. Jeon, U. S., Kim, J. A., Sheen, M. R. and Kwon, H. M. (2006). How tonicity regulates genes: story of TonEBP transcriptional activator. Acta Physiol (Oxf) 187, 241-7. Jiggins, F. M. and Kim, K. W. (2006). Contrasting evolutionary patterns in Drosophila immune receptors. J Mol Evol 63, 769-80. Jung, A. C., Denholm, B., Skaer, H. and Affolter, M. (2005a). Renal tubule development in Drosophila: a closer look at the cellular level. J Am Soc Nephrol 16, 322-8. Jung, S. H., Evans, C. J., Uemura, C. and Banerjee, U. (2005b). The Drosophila lymph gland as a developmental model of hematopoiesis. Development 132, 2521-33. Jäderlund, A. (2002). Iris bok. Stockholm Alfabeta Bokförlag AB Stockholm Kallio, J., Leinonen, A., Ulvila, J., Valanne, S., Ezekowitz, R. A. and Rämet, M. (2005). Functional analysis of immune response genes in Drosophila identifies JNK pathway as a regulator of antimicrobial peptide gene expression in S2 cells. Microbes Infect 7, 811-9. Kambris, Z., Brun, S., Jang, I. H., Nam, H. J., Romeo, Y., Takahashi, K., Lee, W. J., Ueda, R. and Lemaitre, B. (2006). Drosophila immunity: a large-scale in vivo RNAi screen identifies five serine proteases required for Toll activation. Curr Biol 16, 808-13. Kanda, H., Igaki, T., Kanuka, H., Yagi, T. and Miura, M. (2002). Wengen, a member of the Drosophila tumor necrosis factor receptor superfamily, is required for Eiger signaling. J Biol Chem 277, 28372-5. Kaneko, T., Goldman, W. E., Mellroth, P., Steiner, H., Fukase, K., Kusumoto, S., Harley, W., Fox, A., Golenbock, D. and Silverman, N. (2004). Monomeric and polymeric gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway. Immunity 20, 637-49. Kang, D., Liu, G., Lundstrom, A., Gelius, E. and Steiner, H. (1998). A peptidoglycan recognition protein in innate immunity conserved from insects to humans. Proc Natl Acad Sci U S A 95, 10078-82.

52
Karlsson, C., Korayem, A. M., Scherfer, C., Loseva, O., Dushay, M. S. and Theopold, U. (2004). Proteomic analysis of the Drosophila larval hemolymph clot. J Biol Chem 279, 52033-41. Keyser, P., Borge-Renberg, K. and Hultmark, D. (2007). The Drosophila NFAT homolog is involved in salt stress tolerance. Insect Biochem Mol Biol 37, 356-62. Kim, L. K., Choi, U. Y., Cho, H. S., Lee, J. S., Lee, W. B., Kim, J., Jeong, K., Shim, J., Kim-Ha, J. and Kim, Y. J. (2007). Down-regulation of NF-kappaB target genes by the AP-1 and STAT complex during the innate immune response in Drosophila. PLoS Biol 5, e238. Kim, M., Lee, J. H., Lee, S. Y., Kim, E. and Chung, J. (2006). Caspar, a suppressor of antibacterial immunity in Drosophila. Proc Natl Acad Sci U S A 103, 16358-63. Kim, T., Yoon, J., Cho, H., Lee, W. B., Kim, J., Song, Y. H., Kim, S. N., Yoon, J. H., Kim-Ha, J. and Kim, Y. J. (2005). Downregulation of lipopolysaccharide response in Drosophila by negative crosstalk between the AP1 and NF-kappaB signaling modules. Nat Immunol 6, 211-8. Kleino, A., Myllymaki, H., Kallio, J., Vanha-Aho, L. M., Oksanen, K., Ulvila, J., Hultmark, D., Valanne, S. and Rämet, M. (2008). Pirk is a negative regulator of the Drosophila imd pathway. J Immunol 180, 5413-22. Kleino, A., Valanne, S., Ulvila, J., Kallio, J., Myllymaki, H., Enwald, H., Stöven, S., Poidevin, M., Ueda, R., Hultmark, D. et al. (2005). Inhibitor of apoptosis 2 and TAK1-binding protein are components of the Drosophila Imd pathway. Embo J 24, 3423-34. Kocks, C., Cho, J. H., Nehme, N., Ulvila, J., Pearson, A. M., Meister, M., Strom, C., Conto, S. L., Hetru, C., Stuart, L. M. et al. (2005). Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell 123, 335-46. Kocks, C., Maehr, R., Overkleeft, H. S., Wang, E. W., Iyer, L. K., Lennon-Dumenil, A. M., Ploegh, H. L. and Kessler, B. M. (2003). Functional proteomics of the active cysteine protease content in Drosophila S2 cells. Mol Cell Proteomics 2, 1188-97. Kragol, G., Hoffmann, R., Chattergoon, M. A., Lovas, S., Cudic, M., Bulet, P., Condie, B. A., Rosengren, K. J., Montaner, L. J. and Otvos, L., Jr. (2002). Identification of crucial residues for the antibacterial activity of the proline-rich peptide, pyrrhocoricin. Eur J Biochem 269, 4226-37. Kragol, G., Lovas, S., Varadi, G., Condie, B. A., Hoffmann, R. and Otvos, L., Jr. (2001). The antibacterial peptide pyrrhocoricin inhibits the ATPase actions of DnaK and prevents chaperone-assisted protein folding. Biochemistry 40, 3016-26.

53
Kraut, R., Menon, K. and Zinn, K. (2001). A gain-of-function screen for genes controlling motor axon guidance and synaptogenesis in Drosophila. Curr Biol 11, 417-30. Kurucz, E., Markus, R., Zsamboki, J., Folkl-Medzihradszky, K., Darula, Z., Vilmos, P., Udvardy, A., Krausz, I., Lukacsovich, T., Gateff, E. et al. (2007). Nimrod, a putative phagocytosis receptor with EGF repeats in Drosophila plasmatocytes. Curr Biol 17, 649-54. Kurucz, E., Zettervall, C. J., Sinka, R., Vilmos, P., Pivarcsi, A., Ekengren, S., Hegedus, Z., Ando, I. and Hultmark, D. (2003). Hemese, a hemocyte-specific transmembrane protein, affects the cellular immune response in Drosophila. Proc Natl Acad Sci U S A 100, 2622-7. Kylsten, P., Samakovlis, C. and Hultmark, D. (1990). The cecropin locus in Drosophila; a compact gene cluster involved in the response to infection. Embo J 9, 217-24. Lagueux, M., Perrodou, E., Levashina, E. A., Capovilla, M. and Hoffmann, J. A. (2000). Constitutive expression of a complement-like protein in toll and JAK gain-of-function mutants of Drosophila. Proc Natl Acad Sci U S A 97, 11427-32. Lanot, R., Zachary, D., Holder, F. and Meister, M. (2001). Postembryonic hematopoiesis in Drosophila. Dev Biol 230, 243-57. Lavine, M. D. and Strand, M. R. (2003). Haemocytes from Pseudoplusia includens express multiple alpha and beta integrin subunits. Insect Mol Biol 12, 441-52. Lebestky, T., Chang, T., Hartenstein, V. and Banerjee, U. (2000). Specification of Drosophila hematopoietic lineage by conserved transcription factors. Science 288, 146-9. Leclerc, V., Pelte, N., El Chamy, L., Martinelli, C., Ligoxygakis, P., Hoffmann, J. A. and Reichhart, J. M. (2006). Prophenoloxidase activation is not required for survival to microbial infections in Drosophila. EMBO Rep 7, 231-5. Lee, J. H., Cho, K. S., Lee, J., Yoo, J., Lee, J. and Chung, J. (2001). Diptericin-like protein: an immune response gene regulated by the anti-bacterial gene induction pathway in Drosophila. Gene 271, 233-8. Lemaitre, B., Kromer-Metzger, E., Michaut, L., Nicolas, E., Meister, M., Georgel, P., Reichhart, J. M. and Hoffmann, J. A. (1995a). A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense. Proc Natl Acad Sci U S A 92, 9465-9. Lemaitre, B., Meister, M., Govind, S., Georgel, P., Steward, R., Reichhart, J. M. and Hoffmann, J. A. (1995b). Functional analysis and regulation of nuclear import of dorsal during the immune response in Drosophila. Embo J 14, 536-45. Lemaitre, B., Nicolas, E., Michaut, L., Reichhart, J. M. and Hoffmann, J. A. (1996). The dorsoventral regulatory gene cassette

54
spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86, 973-83. Leone, P., Bischoff, V., Kellenberger, C., Hetru, C., Royet, J. and Roussel, A. (2008). Crystal structure of Drosophila PGRP-SD suggests binding to DAP-type but not lysine-type peptidoglycan. Mol Immunol 45, 2521-30. Leulier, F. and Lemaitre, B. (2008). Toll-like receptors--taking an evolutionary approach. Nat Rev Genet 9, 165-78. Leulier, F., Lhocine, N., Lemaitre, B. and Meier, P. (2006). The Drosophila inhibitor of apoptosis protein DIAP2 functions in innate immunity and is essential to resist gram-negative bacterial infection. Mol Cell Biol 26, 7821-31. Leulier, F., Rodriguez, A., Khush, R. S., Abrams, J. M. and Lemaitre, B. (2000). The Drosophila caspase Dredd is required to resist gram-negative bacterial infection. EMBO Rep 1, 353-8. Leulier, F., Vidal, S., Saigo, K., Ueda, R. and Lemaitre, B. (2002). Inducible expression of double-stranded RNA reveals a role for dFADD in the regulation of the antibacterial response in Drosophila adults. Curr Biol 12, 996-1000. Levashina, E. A., Moita, L. F., Blandin, S., Vriend, G., Lagueux, M. and Kafatos, F. C. (2001). Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae. Cell 104, 709-18. Loyher, M. L., Mutin, M., Woo, S. K., Kwon, H. M. and Tappaz, M. L. (2004). Transcription factor tonicity-responsive enhancer-binding protein (TonEBP) which transactivates osmoprotective genes is expressed and upregulated following acute systemic hypertonicity in neurons in brain. Neuroscience 124, 89-104. Lu, Y., Wu, L. P. and Anderson, K. V. (2001). The antibacterial arm of the Drosophila innate immune response requires an IkappaB kinase. Genes Dev 15, 104-10. Luo, H., Hanratty, W. P. and Dearolf, C. R. (1995). An amino acid substitution in the Drosophila hopTum-l Jak kinase causes leukemia-like hematopoietic defects. Embo J 14, 1412-20. Mace, K. A., Pearson, J. C. and McGinnis, W. (2005). An epidermal barrier wound repair pathway in Drosophila is mediated by grainy head. Science 308, 381-5. Macias, A., Romero, N. M., Martin, F., Suarez, L., Rosa, A. L. and Morata, G. (2004). PVF1/PVR signaling and apoptosis promotes the rotation and dorsal closure of the Drosophila male terminalia. Int J Dev Biol 48, 1087-94. Maeshima, A., Miya, M., Mishima, K., Yamashita, S., Kojima, I. and Nojima, Y. (2008). Activin A: autocrine regulator of kidney development and repair. Endocr J 55, 1-9.

55
Maillet, F., Bischoff, V., Vignal, C., Hoffmann, J. and Royet, J. (2008). The Drosophila peptidoglycan recognition protein PGRP-LF blocks PGRP-LC and IMD/JNK pathway activation. Cell Host Microbe 3, 293-303. Malagoli, D., Conklin, D., Sacchi, S., Mandrioli, M. and Ottaviani, E. (2007). A putative helical cytokine functioning in innate immune signalling in Drosophila melanogaster. Biochim Biophys Acta 1770, 974-8. Manjon, C., Sanchez-Herrero, E. and Suzanne, M. (2007). Sharp boundaries of Dpp signalling trigger local cell death required for Drosophila leg morphogenesis. Nat Cell Biol 9, 57-63. Markus, R., Kurucz, E., Rus, F. and Ando, I. (2005). Sterile wounding is a minimal and sufficient trigger for a cellular immune response in Drosophila melanogaster. Immunol Lett 101, 108-11. Martin, P. and Parkhurst, S. M. (2004). Parallels between tissue repair and embryo morphogenesis. Development 131, 3021-34. Martin-Blanco, E., Gampel, A., Ring, J., Virdee, K., Kirov, N., Tolkovsky, A. M. and Martinez-Arias, A. (1998). puckered encodes a phosphatase that mediates a feedback loop regulating JNK activity during dorsal closure in Drosophila. Genes Dev 12, 557-70. Martin-Blanco, E., Pastor-Pareja, J. C. and Garcia-Bellido, A. (2000). JNK and decapentaplegic signaling control adhesiveness and cytoskeleton dynamics during thorax closure in Drosophila. Proc Natl Acad Sci U S A 97, 7888-93. Matova, N. and Anderson, K. V. (2006). Rel/NF-kappaB double mutants reveal that cellular immunity is central to Drosophila host defense. Proc Natl Acad Sci U S A 103, 16424-9. Mattila, J., Omelyanchuk, L., Kyttala, S., Turunen, H. and Nokkala, S. (2005). Role of Jun N-terminal Kinase (JNK) signaling in the wound healing and regeneration of a Drosophila melanogaster wing imaginal disc. Int J Dev Biol 49, 391-9. Medzhitov, R. and Janeway, C. A., Jr. (1997). Innate immunity: the virtues of a nonclonal system of recognition. Cell 91, 295-8. Meister, M. (2004). Blood cells of Drosophila: cell lineages and role in host defence. Curr Opin Immunol 16, 10-5. Mellroth, P., Karlsson, J., Hakansson, J., Schultz, N., Goldman, W. E. and Steiner, H. (2005). Ligand-induced dimerization of Drosophila peptidoglycan recognition proteins in vitro. Proc Natl Acad Sci U S A 102, 6455-60. Mellroth, P., Karlsson, J. and Steiner, H. (2003). A scavenger function for a Drosophila peptidoglycan recognition protein. J Biol Chem 278, 7059-64. Mellroth, P. and Steiner, H. (2006). PGRP-SB1: an N-acetylmuramoyl L-alanine amidase with antibacterial activity. Biochem Biophys Res Commun 350, 994-9.

56
Michel, T., Reichhart, J. M., Hoffmann, J. A. and Royet, J. (2001). Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature 414, 756-9. Mihaly, J., Kockel, L., Gaengel, K., Weber, U., Bohmann, D. and Mlodzik, M. (2001). The role of the Drosophila TAK homologue dTAK during development. Mech Dev 102, 67-79. Moreno, E., Basler, K. and Morata, G. (2002a). Cells compete for decapentaplegic survival factor to prevent apoptosis in Drosophila wing development. Nature 416, 755-9. Moreno, E., Yan, M. and Basler, K. (2002b). Evolution of TNF signaling mechanisms: JNK-dependent apoptosis triggered by Eiger, the Drosophila homolog of the TNF superfamily. Curr Biol 12, 1263-8. Mukherjee, T., Schafer, U. and Zeidler, M. P. (2006). Identification of Drosophila genes modulating janus kinase/signal transducer and activator of transcription signal transduction. Genetics 172, 1683-97. Muratoglu, S., Garratt, B., Hyman, K., Gajewski, K., Schulz, R. A. and Fossett, N. (2006). Regulation of Drosophila friend of GATA gene, u-shaped, during hematopoiesis: a direct role for serpent and lozenge. Dev Biol 296, 561-79. Naitza, S., Rosse, C., Kappler, C., Georgel, P., Belvin, M., Gubb, D., Camonis, J., Hoffmann, J. A. and Reichhart, J. M. (2002). The Drosophila immune defense against gram-negative infection requires the death protein dFADD. Immunity 17, 575-81. Nappi, A. J. and Christensen, B. M. (2005). Melanogenesis and associated cytotoxic reactions: applications to insect innate immunity. Insect Biochem Mol Biol 35, 443-59. O'Connor, M. B., Umulis, D., Othmer, H. G. and Blair, S. S. (2006). Shaping BMP morphogen gradients in the Drosophila embryo and pupal wing. Development 133, 183-93. Oshima, K., Takeda, M., Kuranaga, E., Ueda, R., Aigaki, T., Miura, M. and Hayashi, S. (2006). IKK epsilon regulates F actin assembly and interacts with Drosophila IAP1 in cellular morphogenesis. Curr Biol 16, 1531-7. Otvos, L., Jr. (2002). The short proline-rich antibacterial peptide family. Cell Mol Life Sci 59, 1138-50. Otvos, L., Jr., O, I., Rogers, M. E., Consolvo, P. J., Condie, B. A., Lovas, S., Bulet, P. and Blaszczyk-Thurin, M. (2000). Interaction between heat shock proteins and antimicrobial peptides. Biochemistry 39, 14150-9. Paladi, M. and Tepass, U. (2004). Function of Rho GTPases in embryonic blood cell migration in Drosophila. J Cell Sci 117, 6313-26. Park, J. M., Brady, H., Ruocco, M. G., Sun, H., Williams, D., Lee, S. J., Kato, T., Jr., Richards, N., Chan, K., Mercurio, F. et al. (2004). Targeting of TAK1 by the NF-kappa B protein

57
Relish regulates the JNK-mediated immune response in Drosophila. Genes Dev 18, 584-94. Pearson, A., Lux, A. and Krieger, M. (1995). Expression cloning of dSR-CI, a class C macrophage-specific scavenger receptor from Drosophila melanogaster. Proc Natl Acad Sci U S A 92, 4056-60. Pearson, A. M., Baksa, K., Rämet, M., Protas, M., McKee, M., Brown, D. and Ezekowitz, R. A. (2003). Identification of cytoskeletal regulatory proteins required for efficient phagocytosis in Drosophila. Microbes Infect 5, 815-24. Pena-Rangel, M. T., Rodriguez, I. and Riesgo-Escovar, J. R. (2002). A misexpression study examining dorsal thorax formation in Drosophila melanogaster. Genetics 160, 1035-50. Persson, C., Oldenvi, S. and Steiner, H. (2007). Peptidoglycan recognition protein LF: a negative regulator of Drosophila immunity. Insect Biochem Mol Biol 37, 1309-16. Pham, L. N., Dionne, M. S., Shirasu-Hiza, M. and Schneider, D. S. (2007). A specific primed immune response in Drosophila is dependent on phagocytes. PLoS Pathog 3, e26. Philips, J. A., Rubin, E. J. and Perrimon, N. (2005). Drosophila RNAi screen reveals CD36 family member required for mycobacterial infection. Science 309, 1251-3. Pili-Floury, S., Leulier, F., Takahashi, K., Saigo, K., Samain, E., Ueda, R. and Lemaitre, B. (2004). In vivo RNA interference analysis reveals an unexpected role for GNBP1 in the defense against Gram-positive bacterial infection in Drosophila adults. J Biol Chem 279, 12848-53. Qiu, P., Pan, P. C. and Govind, S. (1998). A role for the Drosophila Toll/Cactus pathway in larval hematopoiesis. Development 125, 1909-20. Rabel, D., Charlet, M., Ehret-Sabatier, L., Cavicchioli, L., Cudic, M., Otvos, L., Jr. and Bulet, P. (2004). Primary structure and in vitro antibacterial properties of the Drosophila melanogaster attacin C Pro-domain. J Biol Chem 279, 14853-9. Ralston, A. and Blair, S. S. (2005). Long-range Dpp signaling is regulated to restrict BMP signaling to a crossvein competent zone. Dev Biol 280, 187-200. Rämet, M., Lanot, R., Zachary, D. and Manfruelli, P. (2002a). JNK signaling pathway is required for efficient wound healing in Drosophila. Dev Biol 241, 145-56. Rämet, M., Manfruelli, P., Pearson, A., Mathey-Prevot, B. and Ezekowitz, R. A. (2002b). Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature 416, 644-8. Rämet, M., Pearson, A., Manfruelli, P., Li, X., Koziel, H., Gobel, V., Chung, E., Krieger, M. and Ezekowitz, R. A. (2001).

58
Drosophila scavenger receptor CI is a pattern recognition receptor for bacteria. Immunity 15, 1027-38. Ray, R. P. and Wharton, K. A. (2001). Context-dependent relationships between the BMPs gbb and dpp during development of the Drosophila wing imaginal disk. Development 128, 3913-25. Redd, M. J., Cooper, L., Wood, W., Stramer, B. and Martin, P. (2004). Wound healing and inflammation: embryos reveal the way to perfect repair. Philos Trans R Soc Lond B Biol Sci 359, 777-84. Ribeiro, C. and Brehelin, M. (2006). Insect haemocytes: what type of cell is that? J Insect Physiol 52, 417-29. Riesgo-Escovar, J. R. and Hafen, E. (1997). Drosophila Jun kinase regulates expression of decapentaplegic via the ETS-domain protein Aop and the AP-1 transcription factor DJun during dorsal closure. Genes Dev 11, 1717-27. Riesgo-Escovar, J. R., Jenni, M., Fritz, A. and Hafen, E. (1996). The Drosophila Jun-N-terminal kinase is required for cell morphogenesis but not for DJun-dependent cell fate specification in the eye. Genes Dev 10, 2759-68. Rizki, M. T. and Rizki, R. M. (1959). Functional significance of the crystal cells in the larva of Drosophila melanogaster. J Biophys Biochem Cytol 5, 235-40. Roth, S., Hiromi, Y., Godt, D. and Nusslein-Volhard, C. (1991). cactus, a maternal gene required for proper formation of the dorsoventral morphogen gradient in Drosophila embryos. Development 112, 371-88. Rutschmann, S., Jung, A. C., Zhou, R., Silverman, N., Hoffmann, J. A. and Ferrandon, D. (2000). Role of Drosophila IKK gamma in a toll-independent antibacterial immune response. Nat Immunol 1, 342-7. Ryoo, H. D., Gorenc, T. and Steller, H. (2004). Apoptotic cells can induce compensatory cell proliferation through the JNK and the Wingless signaling pathways. Dev Cell 7, 491-501. Samakovlis, C., Kimbrell, D. A., Kylsten, P., Engström, A. and Hultmark, D. (1990). The immune response in Drosophila: pattern of cecropin expression and biological activity. Embo J 9, 2969-76. Scherfer, C., Karlsson, C., Loseva, O., Bidla, G., Goto, A., Havemann, J., Dushay, M. S. and Theopold, U. (2004). Isolation and characterization of hemolymph clotting factors in Drosophila melanogaster by a pullout method. Curr Biol 14, 625-9. Scherfer, C., Qazi, M. R., Takahashi, K., Ueda, R., Dushay, M. S., Theopold, U. and Lemaitre, B. (2006). The Toll immune-regulated Drosophila protein Fondue is involved in hemolymph clotting and puparium formation. Dev Biol 295, 156-63.

59
Schleifer, K. H. and Kandler, O. (1972). Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol Rev 36, 407-77. Schmidt, R. L., Trejo, T. R., Plummer, T. B., Platt, J. L. and Tang, A. H. (2007). Infection-induced proteolysis of PGRP-LC controls the IMD activation and melanization cascades in Drosophila. Faseb J. Schmidt, R. L., Trejo, T. R., Plummer, T. B., Platt, J. L. and Tang, A. H. (2008). Infection-induced proteolysis of PGRP-LC controls the IMD activation and melanization cascades in Drosophila. Faseb J 22, 918-29. Shai, Y. (2002). Mode of action of membrane active antimicrobial peptides. Biopolymers 66, 236-48. Shen, J. and Dahmann, C. (2005). Extrusion of cells with inappropriate Dpp signaling from Drosophila wing disc epithelia. Science 307, 1789-90. Shimmi, O., Ralston, A., Blair, S. S. and O'Connor, M. B. (2005). The crossveinless gene encodes a new member of the Twisted gastrulation family of BMP-binding proteins which, with Short gastrulation, promotes BMP signaling in the crossveins of the Drosophila wing. Dev Biol 282, 70-83. Silverman, N., Zhou, R., Erlich, R. L., Hunter, M., Bernstein, E., Schneider, D. and Maniatis, T. (2003). Immune activation of NF-kappaB and JNK requires Drosophila TAK1. J Biol Chem 278, 48928-34. Silverman, N., Zhou, R., Stöven, S., Pandey, N., Hultmark, D. and Maniatis, T. (2000). A Drosophila IkappaB kinase complex required for Relish cleavage and antibacterial immunity. Genes Dev 14, 2461-71. Singh, A., Chan, J., Chern, J. J. and Choi, K. W. (2005). Genetic interaction of Lobe with its modifiers in dorsoventral patterning and growth of the Drosophila eye. Genetics 171, 169-83. Sluss, H. K., Han, Z., Barrett, T., Goberdhan, D. C., Wilson, C., Davis, R. J. and Ip, Y. T. (1996). A JNK signal transduction pathway that mediates morphogenesis and an immune response in Drosophila. Genes Dev 10, 2745-58. Sorrentino, R. P., Carton, Y. and Govind, S. (2002). Cellular immune response to parasite infection in the Drosophila lymph gland is developmentally regulated. Dev Biol 243, 65-80. Spencer, F. A., Hoffmann, F. M. and Gelbart, W. M. (1982). Decapentaplegic: a gene complex affecting morphogenesis in Drosophila melanogaster. Cell 28, 451-61. Steiner, H. (2004). Peptidoglycan recognition proteins: on and off switches for innate immunity. Immunol Rev 198, 83-96.

60
Steiner, H., Andreu, D. and Merrifield, R. B. (1988). Binding and action of cecropin and cecropin analogues: antibacterial peptides from insects. Biochim Biophys Acta 939, 260-6. Steiner, H., Hultmark, D., Engström, A., Bennich, H. and Boman, H. G. (1981). Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 292, 246-8. Stramer, B., Wood, W., Galko, M. J., Redd, M. J., Jacinto, A., Parkhurst, S. M. and Martin, P. (2005). Live imaging of wound inflammation in Drosophila embryos reveals key roles for small GTPases during in vivo cell migration. J Cell Biol 168, 567-73. Stronach, B. and Perrimon, N. (2002). Activation of the JNK pathway during dorsal closure in Drosophila requires the mixed lineage kinase, slipper. Genes Dev 16, 377-87. Stroschein-Stevenson, S. L., Foley, E., O'Farrell, P. H. and Johnson, A. D. (2006). Identification of Drosophila gene products required for phagocytosis of Candida albicans. PLoS Biol 4, e4. Stuart, L. M., Boulais, J., Charriere, G. M., Hennessy, E. J., Brunet, S., Jutras, I., Goyette, G., Rondeau, C., Letarte, S., Huang, H. et al. (2007). A systems biology analysis of the Drosophila phagosome. Nature 445, 95-101. Stuart, L. M., Deng, J., Silver, J. M., Takahashi, K., Tseng, A. A., Hennessy, E. J., Ezekowitz, R. A. and Moore, K. J. (2005). Response to Staphylococcus aureus requires CD36-mediated phagocytosis triggered by the COOH-terminal cytoplasmic domain. J Cell Biol 170, 477-85. Stuart, L. M. and Ezekowitz, R. A. (2008). Phagocytosis and comparative innate immunity: learning on the fly. Nat Rev Immunol 8, 131-41. Stöven, S., Ando, I., Kadalayil, L., Engström, Y. and Hultmark, D. (2000). Activation of the Drosophila NF-kappaB factor Relish by rapid endoproteolytic cleavage. EMBO Rep 1, 347-52. Stöven, S., Silverman, N., Junell, A., Hedengren-Olcott, M., Erturk, D., Engström, Y., Maniatis, T. and Hultmark, D. (2003). Caspase-mediated processing of the Drosophila NF-kappaB factor Relish. Proc Natl Acad Sci U S A 100, 5991-6. Takaesu, N., Hyman-Walsh, C., Ye, L., Wisotzkey, R., Stinchfield, M., O'Connor, M., Wotton, D. and Newfeld, S. J. (2006). dSno facilitates Baboon signaling in the Drosophila brain by switching the affinity of Medea away from Mad and toward dSmad2. Genetics. Takatsu, Y., Nakamura, M., Stapleton, M., Danos, M. C., Matsumoto, K., O'Connor, M. B., Shibuya, H. and Ueno, N. (2000). TAK1 participates in c-Jun N-terminal kinase signaling during Drosophila development. Mol Cell Biol 20, 3015-26.

61
Tang, H., Kambris, Z., Lemaitre, B. and Hashimoto, C. (2006). Two proteases defining a melanization cascade in the immune system of Drosophila. J Biol Chem 281, 28097-104. Taniai, K., Wago, H. and Yamakawa, M. (1997). In vitro phagocytosis of Escherichia coli and release of lipopolysaccharide by adhering hemocytes of the silkworm, Bombyx mori. Biochem Biophys Res Commun 231, 623-7. Ting, S. B., Caddy, J., Hislop, N., Wilanowski, T., Auden, A., Zhao, L. L., Ellis, S., Kaur, P., Uchida, Y., Holleran, W. M. et al. (2005). A homolog of Drosophila grainy head is essential for epidermal integrity in mice. Science 308, 411-3. Tingvall, T. O., Roos, E. and Engström, Y. (2001). The GATA factor Serpent is required for the onset of the humoral immune response in Drosophila embryos. Proc Natl Acad Sci U S A 98, 3884-8. Torres-Vazquez, J., Park, S., Warrior, R. and Arora, K. (2001). The transcription factor Schnurri plays a dual role in mediating Dpp signaling during embryogenesis. Development 128, 1657-70. Torres-Vazquez, J., Warrior, R. and Arora, K. (2000). schnurri is required for dpp-dependent patterning of the Drosophila wing. Dev Biol 227, 388-402. Tsuda, M., Langmann, C., Harden, N. and Aigaki, T. (2005). The RING-finger scaffold protein Plenty of SH3s targets TAK1 to control immunity signalling in Drosophila. EMBO Rep 6, 1082-7. Tzou, P., Ohresser, S., Ferrandon, D., Capovilla, M., Reichhart, J. M., Lemaitre, B., Hoffmann, J. A. and Imler, J. L. (2000). Tissue-specific inducible expression of antimicrobial peptide genes in Drosophila surface epithelia. Immunity 13, 737-48. Uttenweiler-Joseph, S., Moniatte, M., Lagueux, M., Van Dorsselaer, A., Hoffmann, J. A. and Bulet, P. (1998). Differential display of peptides induced during the immune response of Drosophila: a matrix-assisted laser desorption ionization time-of-flight mass spectrometry study. Proc Natl Acad Sci U S A 95, 11342-7. Uvell, H. and Engström, Y. (2007). A multilayered defense against infection: combinatorial control of insect immune genes. Trends Genet 23, 342-9. Valanne, S., Kleino, A., Myllymaki, H., Vuoristo, J. and Rämet, M. (2007). Iap2 is required for a sustained response in the Drosophila Imd pathway. Dev Comp Immunol 31, 991-1001. Wang, L., Weber, A. N., Atilano, M. L., Filipe, S. R., Gay, N. J. and Ligoxygakis, P. (2006). Sensing of Gram-positive bacteria in Drosophila: GNBP1 is needed to process and present peptidoglycan to PGRP-SA. Embo J 25, 5005-14. Werner, T., Borge-Renberg, K., Mellroth, P., Steiner, H. and Hultmark, D. (2003). Functional diversity of the Drosophila PGRP-

62
LC gene cluster in the response to lipopolysaccharide and peptidoglycan. J Biol Chem 278, 26319-22. Werner, T., Liu, G., Kang, D., Ekengren, S., Steiner, H. and Hultmark, D. (2000). A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc Natl Acad Sci U S A 97, 13772-7. Vervoort, M., Crozatier, M., Valle, D. and Vincent, A. (1999). The COE transcription factor Collier is a mediator of short-range Hedgehog-induced patterning of the Drosophila wing. Curr Biol 9, 632-9. Wicker, C., Reichhart, J. M., Hoffmann, D., Hultmark, D., Samakovlis, C. and Hoffmann, J. A. (1990). Insect immunity. Characterization of a Drosophila cDNA encoding a novel member of the diptericin family of immune peptides. J Biol Chem 265, 22493-8. Vidal, S., Khush, R. S., Leulier, F., Tzou, P., Nakamura, M. and Lemaitre, B. (2001). Mutations in the Drosophila dTAK1 gene reveal a conserved function for MAPKKKs in the control of rel/NF-kappaB-dependent innate immune responses. Genes Dev 15, 1900-12. Vierstraete, E., Cerstiaens, A., Baggerman, G., Van den Bergh, G., De Loof, A. and Schoofs, L. (2003). Proteomics in Drosophila melanogaster: first 2D database of larval hemolymph proteins. Biochem Biophys Res Commun 304, 831-8. Williams, M. J. (2007). Drosophila hemopoiesis and cellular immunity. J Immunol 178, 4711-6. Williams, M. J., Wiklund, M. L., Wikman, S. and Hultmark, D. (2006). Rac1 signalling in the Drosophila larval cellular immune response. J Cell Sci 119, 2015-24. Vollmer, W., Blanot, D. and de Pedro, M. A. (2008). Peptidoglycan structure and architecture. FEMS Microbiol Rev 32, 149-67. Wu, L. P. and Anderson, K. V. (1998). Regulated nuclear import of Rel proteins in the Drosophila immune response. Nature 392, 93-7. Yoshida, H., Kinoshita, K. and Ashida, M. (1996). Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J Biol Chem 271, 13854-60. Yoshida, H., Ochiai, M. and Ashida, M. (1986). Beta-1,3-glucan receptor and peptidoglycan receptor are present as separate entities within insect prophenoloxidase activating system. Biochem Biophys Res Commun 141, 1177-84. Zaidman-Remy, A., Herve, M., Poidevin, M., Pili-Floury, S., Kim, M. S., Blanot, D., Oh, B. H., Ueda, R., Mengin-Lecreulx, D. and Lemaitre, B. (2006). The Drosophila amidase PGRP-LB modulates the immune response to bacterial infection. Immunity 24, 463-73. Zambon, R. A., Nandakumar, M., Vakharia, V. N. and Wu, L. P. (2005). The Toll pathway is important for an antiviral response in Drosophila. Proc Natl Acad Sci U S A 102, 7257-62.

63
Zeitlinger, J., Kockel, L., Peverali, F. A., Jackson, D. B., Mlodzik, M. and Bohmann, D. (1997). Defective dorsal closure and loss of epidermal decapentaplegic expression in Drosophila fos mutants. Embo J 16, 7393-401. Zettervall, C. J., Anderl, I., Williams, M. J., Palmer, R., Kurucz, E., Ando, I. and Hultmark, D. (2004). A directed screen for genes involved in Drosophila blood cell activation. Proc Natl Acad Sci U S A 101, 14192-7. Zhou, R., Silverman, N., Hong, M., Liao, D. S., Chung, Y., Chen, Z. J. and Maniatis, T. (2005). The role of ubiquitination in Drosophila innate immunity. J Biol Chem 280, 34048-55. Zhu, C. C., Boone, J. Q., Jensen, P. A., Hanna, S., Podemski, L., Locke, J., Doe, C. Q. and O'Connor, M. B. (2008). Drosophila Activin- and the Activin-like product Dawdle function redundantly to regulate proliferation in the larval brain. Development 135, 513-21. Zhu, M. Y., Wilson, R. and Leptin, M. (2005). A screen for genes that influence fibroblast growth factor signal transduction in Drosophila. Genetics 170, 767-77. Zhuang, Z. H., Sun, L., Kong, L., Hu, J. H., Yu, M. C., Reinach, P., Zang, J. W. and Ge, B. X. (2006). Drosophila TAB2 is required for the immune activation of JNK and NF-kappaB. Cell Signal 18, 964-70. Åsling, B., Dushay, M. S. and Hultmark, D. (1995). Identification of early genes in the Drosophila immune response by PCR-based differential display: the Attacin A gene and the evolution of attacin-like proteins. Insect Biochem Mol Biol 25, 511-8. Önfelt Tingvall, T., Roos, E. and Engström, Y. (2001). The imd gene is required for local Cecropin expression in Drosophila barrier epithelia. EMBO Rep 2, 239-43.





























