Embed Size (px)
Citation preview

Universidade de Aveiro 2006
Departamento de Química
Ana Paula Mora Tavares
Produção de lacase para potencial aplicação como oxidante na indústria papeleira
tese apresentada à Universidade de Aveiro para cumprimento dos requisitos necessários à obtenção do grau de Doutor em Engenharia Química, realizada sob a orientação científica da Dra. Ana Maria Barreto Rebelo Xavier, Professora Auxiliar do Departamento de Química da Universidade de Aveiro e do Professor Dr. João Manuel da Costa Araúo Pereira Coutinho, Professor Associado do Departamento de Química da Universidade de Aveiro
Apoio financeiro da FCT e do FSE no âmbito do III Quadro Comunitário de Apoio

o júri
presidente Prof. Doutor Fernando Manuel Bico Marques professor catedrático do Departamento de Engenharia Cerâmica e Vidro da Universidade de Aveiro
Prof. Doutor José Joaquim C. Cruz Pinto professor catedrático do Departamento de Química da Universidade de Aveiro
Profª. Doutora Maria Ascensão Carvalho Fernandes de Miranda Reis professora associada do Departamento de Química da Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa
Prof. Doutor Dmitry Victorovich Evtuguin professor associado do Departamento de Química da Universidade de Aveiro
Prof. Doutor João Manuel da Costa Pereira Coutinho professor associado do Departamento de Química da Universidade de Aveiro
Profª. Doutora Maria Alice Zarur Coelho professora adjunta do Departamento Engenharia Bioquímica da Universidade Federal do Rio de Janeiro
Profª. Doutora Ana Maria Rebelo Barreto Xavier professora auxiliar do Departamento de Química da Universidade de Aveiro
- ii -

agradecimentos
À Professora Doutora Ana M.R.B. Xavier agradeço pela orientação de todo este trabalho, pelo seu apoio, pela sua confiança, pela convivência e pela amizade de todos estes anos. Ao Professor Doutor João A.P. Coutinho agradeço pela orientação deste trabalho, pelo seu apoio, pela sua confiança. À Professora Doutora Maria Alice Zarur Coelho agradeço pela orientação na realização da parte experimental de produção de lacase e de biopolímeros assim como a sua orientação na parte de modelação matemática dos resultados da optimização da produção da lacase e pela sua amizade. Ao Professor Doutor Dmitry Evtuguin agradeço pela sua orientação durante o trabalho realizado com o branqueamento da pasta de papel, pela realização das análises de GPC. Ao Doutor José Gamelas agradeço pelo seu apoio na parte experimental com a pasta de papel, pela realização das análises de voltametria cíclica, de RMN e de espectroscopia UV/Vis, além da sua amizade durante este trabalho. Ao Doutor Armindo Gaspar agradeço pelo seu apoio na parte experimental com a pasta de papel. À Professora Doutora Ana Barro-Timmons agradeço pela sua ajuda durante a realização do trabalho com os biopolímeros, pelas análises de ICP, termogravimétrica, de FTIR e análise elementar. Ao Professor Doutor José A. Lopes da Silva agradeço pela sua ajuda durante a realização do trabalho com os biopolímeros, pela análise de reologia e de açúcares neutros. Aos colegas e amigos de laboratório Margarida, Marta,Sofia, Daniel, Ana Sofia, Priscila, Alexandre, Juliana, Elisabete e Alzira pela amizade e pela ajuda durante a parte experimental desta tese. À PORTUCEL, Cacia, Portugal agradeço pela disponibilização da pasta de papel utilizada neste trabalho. À Universidade de Aveiro pelo apoio financeiro através da concessão da bolsa de Doutoramento concedida durante o primeiro ano de Doutoramento. À Fundação para Ciência e Tecnologia (FCT) pelo apoio financeiro através da concessão da bolsa de Doutoramento e pelo apoio financeiro dos deslocamentos em congressos Internacionais. Finalmente, desejo agradecer ao meu marido que sempre me apoiou durante a realização desta tese e aos meus pais e irmã que sempre me incentivaram na concretização deste trabalho.
- iii -

- iv -
palavras-chave
lacase, Trametes versicolor, modelação matemática, planeamento de experiências, biobranqueamento da pasta kraft, polioxometalatos, exopolissacarídeo.
resumo
A biocatálise enzimática apresenta um grande potencial para o branqueamento da pasta, como um tratamento alternativo ao convencional.Proporciona importantes benefícios ambientais além de introduzir propriedades únicas às fibras do papel. A chave para o sucesso na aplicação industrial está na identificação de novas enzimas e na pesquisa das condições específicas de trabalho.
Para este trabalho escolheu-se o fungo Trametes versicolor conhecido como fungo da podridão branca da madeira, que é capaz de produzir enzimas oxidativas, nomeadamente a enzima lacase. A primeira parte do trabalhoenvolveu a optimização da produção da lacase em matraz e em bioreactor e a modelação matemática dos resultados experimentais obtidos. Para a optimização da produção da lacase em matraz, diferentes indutores foram utilizados: cobre, 2.5-xilidina e uma mistura fenólica. Estudou-se também a situação de limitação de carbono. Para a fermentação em bioreactor fez-se umplaneamento factorial de experiências que consistiu de dois níveis (máximo e mínimo) e três factores: concentração de glucose, agitação e controlo do pHdo meio de cultura.
A segunda parte do trabalho envolveu a aplicação da lacase no branqueamento da pasta kraft para o fabrico de papel. Neste estudo, diferentes tipos de polioxometalatos (POMs) foram testados, juntamente com a lacase, como mediadores da deslenhificação da pasta kraft não branqueada. As condições do branqueamento estudadas foram a temperatura de reacçãoem um único estágio de branqueamento e o branqueamento em estágiosalternados: Este tipo de branqueamento permitiu uma deslenhificaçãosignificativa da pasta de papel com uma redução no índice kappa de cerca de 50% para o POM SiW11MnIII(H2O)O39
5-, após 5 estágios de branqueamento e,para o SiW11VVO40
5- após três estágios de branqueamento. Os espectros de UV-Vis e de RMN mostraram que estes dois POMs se mantiveram estáveisdurante o processo de branqueamento da pasta de papel.
Para além de produzir a lacase, o fungo T. versicolor também é conhecido como produtor de exopolissacarídeos (EPSs). A terceira parte deste trabalho constituiu na produção e optimização de um EPS. A optimização da produção do EPS foi feita através da selecção de cinco diferentes meios de cultura e doplaneamento factorial de experiências 22 com ponto central com o meio seleccionado. As variáveis estudadas no planeamento foram a concentraçãoinicial de glucose e o pH do meio de cultura. A concentração inicial de glucosefoi o principal factor para a optimização da produção do EPS e para ocrescimento celular.
Realizaram-se alguns estudos para conhecer a natureza do EPS queenvolveram análise elementar, análise dos iões por ICP, análise estrutural porFTIR, determinação do peso molecular por GPC, análise termogravimétrica,análise dos açúcares neutros e dos ácidos hexourónicos.

key words
laccase, Trametes versicolor, mathematical modelling, factorial design, kraft pulp biobleachig, polyoxometalates, exopolyssacharide.
abstract
Enzymatic biocatalyst has a significant potential for improving pulp biobleaching as an alternative treatment to conventional. It allows for the development of environmentally benign processes and may introduce unique properties into the paper fibre. The challenge for the success on the industrial application is the identification of new enzymes and the development of specific and optimized working conditions.
On this work the fungus Trametes versicolor, known as white rot fungus, which produces oxidative enzymes namely laccase was used. The first part ofthe work addresses the optimization of laccase production in Erlenmeyer andbioreactor and the mathematical modelling of the experimental results. To optimise the laccase production in Erlenmeyer different inducers were tested:copper, 2.5-xylidine and a phenolic mixture. The situation of carbon limitationwas also studied. For bioreactor fermentation a factorial design with two levels(maximum and minimum) and three factors: initial glucose concentration,agitation and medium pH was used.
The second part of the work involved the application of laccase on kraftpulp biobleaching. In this study different polyoxometalates (POMs) were tested, with laccase, as mediators on kraft pulp bleaching. The conditions of bleachingwere reaction temperature of the single bleaching stage and the multi-stage bleaching. The alternative treatment in a multi-stage process allowed a sustainable eucalypt kraft pulp delignification with more than 50% of kappa number reduction to POM SiW11MnIII(H2O)O39
5-, after five stages of bleaching, and to the POM o SiW11VVO40
5- after three stages of bleaching. The UV/VIS spectrum and RMN showed that these two POMs maintain stable during the biobleaching process of pulp paper.
In addition to the production of laccase the fungus T. versicolor is also known as exopolysaccharides (EPSs) producer. The third part of this workdeals with the optimization of the EPS production. This was optimized by a selection of five culture media and by a 22 factorial design with a central point of selected medium. The factors of factorial design were initial glucoseconcentration and initial media pH. The initial glucose concentration was found to be the most important factor in EPS production and also in cell growth.
Some studies were carried in order to characterize the EPS usingelementary analysis, ions analysis by ICP, structural analysis by FTIR, analysis of molecular weight by GPC, thermal gravimetric analysis, neutral sugars and hexouronic acids.
- v -

Índice Geral
Introdução.............................................................................................................. xx
Capítulo 1 - Revisão bibliográfica........................................................................ 1
1.1 A indústria de pasta de papel................................................................. 1
1.1.1 Composição da madeira.......................................................... 3
1.1.1.1 Celulose.................................................................... 4
1.1.1.2 Hemiceluloses ........................................................ 5
1.1.1.3 Lenhina .................................................................... 6
1.1.2 O processo de fabricação da pasta de papel............................ 11
1.1.3 Branqueamento da pasta kraft................................................. 12
1.1.3.1 O papel da lacase no branqueamento da pasta......... 14
1.1.3.2 Uso de mediadores .................................................. 15
1.1.3.2.1 Os polioxometalatos.................................. 18
1.2 A natureza dos fungos........................................................................... 22
1.2.1 Fungos da podridão branca da madeira................................... 24
1.2.2 O sistema lenhinolítico dos fungos da podridão branca da
madeira................................................................................. 24
1.2.3 O fungo Trametes versicolor.................................................. 26
1.3 Enzimas lenhinolíticas........................................................................... 27
1.3.1 Lenhina Peroxidase (LiP) ....................................................... 28
1.3.2 Manganês Peroxidades (MnP) ............................................... 29
1.3.3 Lacases .................................................................................. 29
1.3.3.1 Caracterização estrutural da lacase.......................... 29
1.3.3.2 Mecanismos de catálise da lacase........................... 31
1.3.3.3 Aplicações da lacase................................................. 32
1.3.3.4 Produção da lacase................................................... 33
1.3.4 Produção industrial de enzimas............................................... 36
1.4 Polissacarídeos produzidos por fungos.................................................. 37
1.4.1 Polissacarídeos....................................................................... 38
1.4.2 Produção de Exopolissacarídeos............................................. 42
1.4.2.1 Processo de recuperação de exopolissacarídeos
vi

de origem microbiana .............................................. 44
1.4.3 Compostos comercializados.................................................... 46
1.5 Planeamento de experiências e modelação matemática........................ 48
1.5.1 Planeamento factorial a dois níveis ....................................... 48
1.5.1.1 Cálculo dos principais efeitos e interacções............. 49
1.5.1.2 Metodologia de superfície de resposta.................... 50
1.5.2 Modelação matemática........................................................... 51
1.5.2.1 Cinética de crescimento microbiano........................ 52
1.5.2.2 Modelos para a formação do produto – A taxa
específica de produção............................................. 55
Capítulo 2 - Materiais e Métodos......................................................................... 57
2.1 Microorganismo e condições de manutenção........................................ 57
2.2 Preparação do inóculo para culturas líquidas........................................ 58
2.3 Fermentações com Trametes versicolor para a produção enzimática... 59
2.3.1 Meio de produção enzimática................................................. 59
2.3.2 Fermentações em matraz........................................................ 60
2.3.2.1 Estudo do efeito da concentração inicial de
glucose.................................................................. 60
2.3.2.2 Estudo do efeito da adição de cobre..................... 60
2.3.2.3 Fermentações com diferentes indutores................ 60
2.3.2.4 Fermentações com limitação por carbono ............61
2.3.2.5 Testes de estabilidade da lacase............................ 61
2.3.3 Fermentações em Bioreactor.................................................. 63
2.3.3.1 Planeamento factorial a dois níveis...................... 64
2.3.3.2 Modelação matemática da produção de
lacase em bioreactor............................................ 66
2.3.4 Métodos analíticos................................................................. 69
2.3.4.1 Determinação do consumo de substrato pelo
método DNS......................................................... 69
2.3.4.2 Determinação da actividade de lacase.................. 69
2.3.4.3 Determinação da actividade de manganês
vii

peroxidase (MnP) ................................................. 70
2.3.4.4 Determinação da actividade celulásica total......... 70
2.3.4.5 Determinação da proteína total pelo método
de Folin-Lowry..................................................... 71
2.3.4.6 Determinação da biomassa................................... 71
2.3.4.7 Determinação do pH............................................ 72
2.4 Aplicação da lacase na pasta kraft......................................................... 72
2.4.1 Preparação da pasta kraft........................................................ 72
2.4.2 Produção da lacase ................................................................. 72
2.4.3 Mediadores utilizados............................................................. 73
2.4.4 Branqueamento da pasta kraft em reactor............................... 73
2.4.5 Extracção alcalina da pasta..................................................... 75
2.4.6 Métodos analíticos.................................................................. 75
2.4.6.1 Determinação do índice kappa.................................... 75
2.4.6.2 Determinação da viscosidade intrínseca..................... 77
2.4.6.3 Determinação do estado de oxidação dos POMs por
espectroscopia no visível............................................ 77
2.4.6.4 Análise de espectroscopia de ressonância magnética
nuclear – RMN.......................................................... 78
2.4.6.5 Análise de voltametria cíclica................................... 79
2.5 Fermentações com Trametes versicolor para produção de
Exopolissacarídeo.................................................................................. 79
2.5.1 Meios de produção.................................................................. 79
2.5.2 Optimização do meio de cultura............................................. 80
2.5.3 Extracção do EPS do meio de cultura..................................... 82
2.5.4 Análise reológica do meio de cultura..................................... 82
2.5.5 Métodos analíticos para a caracterização do EPS................... 82
2.5.5.1 Espectroscopia de Infravermelho (FTIR)................. 82
2.5.5.2 Análise termogravimétrica....................................... 83
2.5.5.3 Análise dos açúcares neutros ................................... 84
viii

2.5.5.4 Determinação do peso molecular............................. 84
2.5.5.5 Determinação dos ácidos urónicos........................... 84
2.5.5.6 Análise elementar do EPS........................................ 85
Capítulo 3 - Resultados e discussão..................................................................... 86
3.1 Produção de lacase em matraz............................................................... 86
3.1.1 Efeito da concentração inicial de glucose............................... 86
3.1.2 Efeito da adição de cobre........................................................ 90
3.1.3 Produção de lacase com diferentes indutores......................... 92
3.1.4 Efeito da limitação por carbono com diferentes indutores......95
3.2 Produção de lacase em fermentador...................................................... 98
3.2.1 Optimização da produção da lacase por planeamento de
experiências.......................................................................... 98
3.2.2 Modelação matemática da produção de lacase em
bioreactor.............................................................................. 106
3.3 Aplicação da lacase para o biobranqueamento da pasta de papel.......... 112
3.3.1 Comparação dos mediadores da lacase para o
branqueamento da pasta.......................................................... 112
3.3.1.1 Ensaios de biobranqueamento da pasta.................... 112
3.3.1.2 Estudo de oxidação dos POMs pela lacase.............. 119
3.3.2 Branqueamento da pasta kraft com SiW11MnIII e lacase
SiW11VV e lacase num sistema de multi-estágios ............... 123
3.4 Produção de exopolissacarídeo (EPS) por Trametes versicolor............ 129
3.4.1 Selecção do meio de produção do EPS................................... 129
3.4.2 Optimização do meio de cultura YM para a produção de
EPS....................................................................................... 132
3.4.3 Análises do EPS...................................................................... 138
3.4.3.1 Análise dos açúcares neutros.................................... 138
3.4.3.2 Análise Termogravimétrica...................................... 139
3.4.3.3 Análise de cromatografia de permeação em gel
(GPC) ....................................................................... 140
3.4.3.4 Análise reológica do meio de cultura....................... 141
ix

3.4.3.5 Análise do EPS produzido no meio YM por
Espectrometria no Infravermelho com Transformada
de Fourier (FTIR) .................................................... 145
3.4.3.6 Análise do EPS por Espectrometria de Emissão
de Plasma Induzido (ICP) e por Análise elementar.. 145
Capítulo 4 – Conclusões........................................................................................ 147
Bibliografia............................................................................................................. 151
Anexos..................................................................................................................... 164
Lista de publicações............................................................................................... 168
x

LISTA DE ABREVIATURAS
ABTS Ácido 2,2’-azino-bis(3-etilbenzotriazolina-6-sulfónico)
ANOVA Análise de variância
BSA albumina de soro bovino
CBQ celobiose:quinona oxidoreductase
CDH celobiose dehidrogenase
CED cupri – etilenodiamina
DMCA N,N-dimetilacetamida
DNS ácido 3,5-dinitrosalicílico
E potencial redox efectivo
Epc potencial de pico catódico
Epa potencial de pico anódico
ENH eléctrodo normal de hidrogénio
EPR ressonância paramagnética electrónica
EPS exopolissacarídeo
FE fibrilas elementares
FTIR espectroscopia de infravermelho com transformada de
Fourier
GlOx glioxalacto oxidase
GPC cromatografia de permeação em gel
GPY meio de glucose, peptona e extracto de levedura
IPA isopolianiões
HBT 1-hidroxilbenzotriazol
HPA heteropolianiões
K constante de decaimento exponencial
Ks constante de saturação
K1 taxa de síntese da lacase
K2 taxa de decaimento da lacase
xi

L razão entre a taxa específica de crescimento no início da fase
de desaceleração e a taxa específica de crescimento na fase
exponencial (factor de sobrevivência)
LiP lenhina peroxidase
MCM meio completo para fungos
MDT meio definido para Trametes
MF mistura fenólica
MnP manganês peroxidase
MSR metodologia de superfícies de resposta
P produto
POM polioxometalato
RMN ressonância magnética nuclear
s segundo
S substrato
SLM sistema lacase-mediador
t tempo
TaK meio Tien and Kirk
TEMPO N-oxil-2,2,6,6-tetrametilpiperidina
UV/VIS espectrofotómetro de ultravioleta/visível
VAO álcool veratrílico oxidase
X biomassa
YM meio extracto de levedura e extracto de malte
α constante associada ao crescimento da biomassa
β constante não associada ao crescimento da biomassa
µ taxa específica de crescimento
ε coeficiente de extinção molar
ν velocidade de varrimento
λ comprimento de onda
xii

ÍNDICE DE TABELAS
Tabela 1.1 - Estrutura de alguns mediadores orgânicos da lacase........................... 16
Tabela 1.2 – Formas diferencial e integrada de várias equações de
crescimentoa ....................................................................................... 53
Tabela 2.1 – Composição do meio de cultura Tien & Kirk..................................... 58
Tabela 2.2 – Composição do meio de cultura MDT................................................ 59
Tabela 2.3 – Condições de cultura para a produção de lacase por Trametes
versicolor em Erlenmeyer................................................................... 62
Tabela 2.4 - Factores e níveis estudados no planeamento factorial 23.....................65
Tabela 2.5 – Matriz do planeamento de experiências 23......................................... 66
Tabela 2.6 - Mediadores utilizados no branqueamento da pasta kraft.................... 73
Tabela 2.7 – Cor e comprimento de onda do estado reduzido e do estado
oxidado dos POMs ............................................................................ 78
Tabela 2.8 – Composição dos meios de cultura usados na produção
do EPS ............................................................................................... 80
Tabela 2.9 - Factores e níveis estudados no planeamento factorial 22
com ponto central............................................................................... 81
Tabela 2.10 – Matriz do primeiro planeamento de experiências 22 com ponto
central............................................................................................... 81
Tabela 2.11 – Matriz do segundo planeamento de experiências 22 com ponto
central .............................................................................................82
Tabela 3.1- Produtividade de lacase e aumento da produtividade de lacase
para os indutores cobre, xilidina e mistura fenólica em
condições de limitação por carbono e sem limitação por
carbono.................................................................................................. 97
Tabela 3.2 – Planeamento de experiências 23 e repostas da produção da
lacase por Trametes versicolor........................................................... 99
Tabela 3.3 – Efeitos estimados e teste ANOVA para a produção de lacase
por Trametes versicolor através do planeamento de
experiências 23 ................................................................................... 100
Tabela 3.4 – Parâmetros cinéticos da fermentação de Trametes
xiii

versicolor em matraz.......................................................................... 107
Tabela 3.5 – Parâmetros para a produção de lacase em bioreactor por
Trametes versicolor........................................................................... 112
Tabela 3.6 – Branqueamento da pasta kraft catalisado por lacase (L) na
presença de diferentes mediadores a 45ºC......................................... 114
Tabela 3.7 – Branqueamento da pasta kraft catalisado por lacase (L) na
presença de diferentes mediadores a 60 ºC...................................... 116
Tabela 3.8 – Oxidação dos polioxometalatos pela lacase........................................ 121
Tabela 3.9 – Branqueamento da pasta kraft em estágios sequenciais
catalisado por SiW11MnIII e lacase ................................................. 124
Tabela 3.10 - Branqueamento da pasta kraft em estágios sequenciais
catalisado por SiW11VV e lacase...................................................... 127
Tabela 3.11 - Resultados do cultivo de Trametes versicolor nos meios de
cultura TaK, MCM e YM................................................................ 130
Tabela 3.12 – Planeamento de experiências 22 e repostas da produção de
EPS por Trametes versicolor........................................................... 135
Tabela 3.13 - Resultados para o segundo planeamento de experiências 22
e repostas da produção de EPS por Trametes versicolor................. 135
Tabela 3.14 - Efeitos individuais calculados e ANOVA para o
planeamento de experiências 22 ...................................................... 137
Tabela 3.15 - Açúcares neutros presentes no EPS produzido por
Trametes versicolor nos meios YM, MCM e TaK.......................... 139
Tabela 3.16 - Ácidos hexourónicos presentes no EPS produzido por
Trametes versicolor nos meios YM, MCM e TaK.......................... 139
Tabela 3.17 – Teor em iões e análise elementar do EPS produzido por
Trametes versicolor nos meios YM, MCM e TaK.......................... 146
xiv

ÍNDICE DE FIGURAS
Figura 1.1 – Representação da cadeia principal da estrutura de celulose
(Higuchi, 1993). .............................................................................. 4
Figura 1.2 – Diagrama estrutural da celulose mostrando as ligações β-(1,4) dos
resíduos de D-glucose e as ligações de hidrogénio inter e intra
molecular (a) e representação das microfibrilas da celulose (b)
(Adaptado de Voet & Voet, 1990)..................................................... 5
Figura 1.3 – Álcoois percursores da lenhina (Adaptado de Biermann, 1996)......... 7
Figura 1.4- Estrutura de uma lenhina resinosa (Sipilä et al., 2005)........................ 8
Figura 1.5 - Principais subestruturas na lenhina; β-O-4’ (a); β-O-4’ degradada
(b); α-O-4’ acíclica (c); β-5’ cíclica e α-O-4’ (d); α-2’/6’ e β-2’6’
(e); 5,5’ (f); 4-O-5’(g); β-1’ (h); β-β’ (i); éster α-HC-
hidroxicinamato de arilglicerilo (com ligação α-4) (j); éster γ-HC-
hidroxicinamato de arilglicerilo (com ligação γ-4) (k)
(Sjöström, 1981). .................................................................................. 10
Figura 1.6 - Representação esquemática da degradação da lenhina por
lacase num sistema com mediador (Adaptado de Bourbonnais
et al., 1998). ....................................................................................... 17
Figura 1.7– Representação bola-bastão e poliedro da unidade fundamental
de MO6 (Fernandez, 2003). ................................................................ 19
Figura 1.8 – Modelos poliédricos representam as três possíveis uniões entre
duas unidades octaédricas MO6. A) união de vértice, B)
união de aresta e C) união de face (Fernandez, 2003)........................ 19
Figura 1.9 - Representação poliédrica da estrutura do anião de Keggin. O
centro externo M12O40 encapsula a unidade interna,
representada pelo tetraedro XO4n- (Jeannin, 1998)............................ 20
Figura 1.10 - Ciclos redox do POM na reacção catalítica da oxidação
da lenhina (Balakshin et al., 2001).................................................... 21
Figura 1.11 – Ciclos redox no sistema catalítico de oxidação da
lenhina com POM-lacase-O2 (Balakshin et al., 2001)........................ 22
Figura 1.12 - Esquema da função desempenhada pelas enzimas degradadoras
xv

de materiais lenhinocelulósicos dos basidiomicetos (Adaptado de
Rajarathnam et al.,1992). ................................................................. 25
Figura 1.13 – O fungo Trametes versicolor na forma de “cauda de perú”.............. 26
Figura 1.14 - Diagrama de da estrutura da lacase de Trametes versicolor.
Os três domínios (T1, T2 e T3) estão mostrados a cores
diferentes. Os átomos de cobre são as esferas azuis e a porção
de hidrato de carbono são os modelos em bola-e-bastão. A
parte cinzenta é o N-terminal e a verde o C-
terminal. O sítio T1 é a esfera isolada. (Piontek et al., 2002)............ 30
Figura 1.15 – Esquema de reacção da lacase com um substrato
(Adaptado de Call & Muck, 1997). ................................................. 31
Figura 1.16 – Uma reacção típica da lacase com um difenol
(Adapado de Thurston, 1994). ........................................................ 32
Figura 1.17 – Representação de alguns monómeros de polissacarídeos................. 39
Figura 1.18- Representação dos monómeros de α e de β D- glucose..................... 39
Figura 1.19 – Diagrama molecular de um D-glucano de origem fúngica.
Representação das ligações glicosídicas (1→4) com
ramificações (1→6). ....................................................................... 39
Figura 1.20 - Conformação de tripla hélice da cadeia de β-(1→3)-D-glucanos..... 41
Figura 1.21 – Opções de recuperação e purificação de polissacaríeos de
Trametes versicolor (Adaptado de Cui & Chisti, 2003)..................... 45
Figura 1.22 – PSP comercializado pela JHS Natural Products............................. 47
Figura 1.23 - VPS comercializado pela JHS Natural Products............................... 47
Figura 1.24 – Vários perfis cinéticos de crescimento: (A) exponencial; (B)
logístico; (C) linear; (D) rápida-aceleração/lenta-
desaceleração (Adaptado de Mitchell et al., 2004).......................... 53
Figura 2.1 – Trametes versicolor após 7 dias de crescimento em placa de Petri... 57
Figura 2.2- Bioreactor usado na produção da lacase por Trametes versicolor....... 64
Figura 2.3 - Reactor e controlador de temperatura e de agitação usado no
branqueamento da pasta kraft de papel. ............................................. 74
Figura 2.4 - Condições do branqueamento da pasta kraft com os POMs SiW11MnIII
xvi

e SiW11VV através do sistema de multi-estágios............................. 75
Figura 2.5 - EPS obtido após precipitação por etanol, lavado e liofilizado......... 83
Figura 3.1 - Actividade de lacase produzida por Trametes versicolor em
diferentes concentrações iniciais de glucose. .................................... 87
Figura 3.2 - Consumo de glucose do meio de cultura durante a produção de
lacase por Trametes versicolor em diferentes
concentrações iniciais de glucose. .......................................................... 87
Figura 3.3 - Evolução da proteína total do meio de cultura durante a
produção de lacase por Trametes versicolor em diferentes
concentrações iniciais de glucose. ............................................... 88
Figura 3.4 – Evolução do pH do meio de cultura durante a produção de
lacase por Trametes versicolor em diferentes concentrações
iniciais de glucose............................................................................... 89
Figura 3.5 – Biomassa final de Trametes versicolor, máxima actividade de
lacase e pH do meio para diferentes concentrações
iniciais de glucose.............................................................................. 90
Figura 3.6 - Efeito da adição de cobre ao primeiro dia na produção da
lacase por Trametes versicolor............................................................. 92
Figura 3.7 – Efeito da adição de cobre ao terceiro dia na produção da
lacase por Trametes versicolor............................................................. 92
Figura 3.8 – (a) Produção de lacase por Trametes versicolor na presença de
cobre, xilidia (xil) e mistura fenólica (MF). (b)
amplificação da Figura 3.7 (a).............................................................. 94
Figura 3.9 - pH do meio de cultura (a) e consumo de glucose (b) durante a
produção de lacase por Trametes versicolor com
diferentes indutores. ............................................................................. 95
Figura 3.10 – Produção de lacase por Trametes versicolor na presença de
cobre e xilidia (xil) e de cobre, xilidia (xil) e mistura fenólica
(MF) com e sem limitação por carbono (LM)...................................... 96
Figura 3.11 - Diagrama de Pareto para o planeamento factorial 23......................... 101
Figura 3.12- Correlação entre os valores observados e os valores
previstos da produção da lacase....................................................... 101
xvii

Figura 3.13 – Superfícies de resposta para o planeamento de experiências 23........ 103
Figura 3.14 - Consumo de glucose e pH do meio durante a produção de
lacase por Trametes versicolor para uma concentração inicial
de glucose de 9 g/L e sem controlo de pH. .......................................... 104
Figura 3.15 - Dados experimentais e modelados para o cultivo de Trametes
versicolor em matraz com o meio com carbono.
(X) biomassa; (S) glucose. .................................................................. 109
Figura 3.16 - Dados experimentais e modelados para o cultivo de Trametes
versicolor em matraz com o meio com limitação
por carbono. (X) biomassa; (S) glucose.................................................. 109
Figura 3.17 - Dados experimentais e modelados para a produção de lacase
em bioreactor por Trametes versicolor em diferentes condições
de fermentação: F1, F2, F5 e F6 (meios com limitação por
carbono). Os dados experimentais estão representados por
símbolos e os modelados por linhas...................................................... 110
Figura 3.18 - Dados experimentais e modelados para a produção de lacase
em bioreactor por Trametes versicolor em diferentes condições
de fermentação: F3, F4, F7 e F8 (meios com carbono). Os dados
experimentais estão representados por símbolos e os modelados
por linhas............................................................................................... 110
Figura 3.19 – Dados experimentais e modelados para o consumo de glucose
e dados modelados para o crescimento da biomassa em
bioreactor por Trametes versicolor em diferentes condições de
fermentação: F3, F4, F7 e F8 (meios com glucose). Os dados
experimentais estão representados por símbolos e os
modelados por linhas. ........................................................................ 111
Figura 3.20 – Variação da viscosidade intrínseca e do índice kappa para o
sistema de branqueamento da pasta de papel a 45ºC com
diferentes mediadores. ....................................................................... 114
Figura 3.21 – Variação da viscosidade intrínseca e do índice kappa para o
sistema de branqueamento da pasta de papel a 60ºC com
diferentes mediadores. ....................................................................... 116
xviii

Figura 3.22 – Espectros de UV-Vis da solução de SiW11MnIII antes do
branqueamento (controlo), ao final do branqueamento a 45ºC
com lacase e a 60ºC com ou sem lacase: (a) controlo; (b)
L+SiW11MnIII 45ºC; (c) L+SiW11MnIII 60ºC; (d)
SiW11MnIII 60ºC. ............................................................................... 117
Figura 3.23 – Percentual de SiW11MnIII, SiW11VV e PW11MnIII no estado
oxidado ao final da reacção para as diferentes condições de
branqueamento. .................................................................................. 118
Figura 3.24 – Espectro UV-Vis para as soluções aquosas (3.0 mmol/L) de
SiW11MnII (A) e SiW11VIV (B) a pH=4.5 e com adição de
lacase (380 U/L) a temperatura ambiente e pressão
atmosférica (a). A: 4h (b); 8 h (c); 24 h (d); 50 h (e); B: 15
min. (b); 30 min. (c); 60 min. (d); 120 min. (e).................................. 119
Figura 3.25 – Figura 3.25 – Espectro de 51V RMN da solução original de
SiW11VV (a) da solução obtida após a reacção do SiW11VIV
(3.0mmol/l) com a lacase (b)..............................................................120
Figura 3.26 – Voltametria cíclica (v=10 mV/s) dos POMs SiW11VIV (a) e
SiW11MnII (b) ([POM]=1.0 mmol/l, pH=4.5,
25ºC)................................................................................................... 123
Figura 3.27 – Espectros de UV-VIS da solução de SiW11MnIII antes do
branqueamento (a); depois de um estágio de branqueamento
POM (b); depois de dois estágios de branqueamento POM-L
(c) seguido de uma semana de armazenamento do efluente (d);
depois de cinco estágios de
branqueamento POM-L-POM-L-POM (e)......................................... 126
Figura 3.28 – Espectros de UV-Vis da solução de SiW11VV antes do
branqueamento (a), depois de um estágio POM de
branqueamento (b), depois de dois estágios POM-L (45ºC) de
branqueamento (c), depois de dois estágios POM-L (60ºC) de
branqueamento (d), e depois de três estágios
POM-L(60ºC)-POM de branqueamento (e)....................................... 127
Figura 3.29 – Espectro de 51V RMN da solução obtida após dois estágios
xix

de branqueamento: SiW11V-L (60ºC)................................................ 128
Figura 3.30 – Crescimento da biomassa (a), consumo de açúcares redutores
(b), produção de EPS (c) durante a fermentação
de Trametes versicolor nos meios TaK, MCM e YM........................ 131
Figura 3.31 – Crescimento da biomassa (a), consumo de açúcares redutores
(b) e produção de EPS (c) durante o cultivo de Trametes
versicolor em diferentes concentrações
(g/L) de glucose (G) e valores de pH................................................. 133
Figura 3.32 – Superfície de resposta para o planeamento de experiências 22......... 134
Figura 3.33 – Superfície de resposta para a produção do EPS do segundo
planeamento de experiências 22 com ponto central......................... 138
Figura 3.34 – Análise termogravimétrica do EPS produzido nos meios
YM, MCM e TaK. ............................................................................. 140
Figura 3.35 – Análise de cromatografia de permeação em gel (GPC) do
EPS produzido nos meios YM, MCM e TaK..................................... 141
Figura 3.36 – Viscosidade aparente dos meios de culturas YM (a), MCM
(b) e TaK (c) durante a produção de EPS por Trametes
versicolor. ....................................................................................... 143
Figura 3.37 – Viscosidade aparente dos meios de cultura TaK, MCM e YM
durante a produção do EPS por Trametes versicolor......................... 144
Figura 3.38 - Concentração do EPS em relação à viscosidade aparente do
meio de cultura. .................................................................................144
Figura 3.39 - Espectro de absorção na região do Infravermelho do EPS
produzido por Trametes versicolor no meio YM............................. 145
xx

Introdução
INTRODUÇÃO
A indústria da pasta e do papel tem sofrido uma grande pressão no sentido do
desenvolvimento de novas tecnologias ambientalmente aceites e de poupança de energia.
Décadas de investigação na biodegradação de polímeros lenhinocelulósicos, no avanço das
ciências biológicas e nos estudos dos mecanismos das reacções enzimáticas, têm
conduzido ao sucesso da aplicação da biotecnologia no processamento das fibras de papel.
Hoje em dia, a biotecnologia pode oferecer à indústria da pasta e do papel grandes
vantagens que podem aumentar a eficiência dos processos já existentes, na manufactura da
pasta e do papel e aprimorar a qualidade do produto através de tecnologias ecológicas à
base de microorganismos e enzimas.
Devido à sua elevada especificidade, baixos custos de investimento e à sua origem
natural, as enzimas produzidas pelos fungos da podridão branca da madeira (entre eles o do
género Trametes) têm-se tornado uma potente ferramenta de pesquisa na caracterização de
fibras. Estas enzimas são biocatalisadores competitivos que podem reduzir ou substituir o
uso de compostos químicos agressivos, que tornam esta indústria poluente em muitas
operações do fabrico da pasta e do papel. Deste modo, estas enzimas constituem uma
alternativa ecológica potencial ao branqueamento químico da pasta de papel.
Aplicações de enzimas em grande escala, com um custo efectivo, podem ser
possíveis através do aumento da capacidade da produção das enzimas em grandes
quantidades. A aceitação destas tecnologias tem vindo a crescer com o conhecimento da
microbiologia, enzimologia, biologia molecular e com o aumento da experiência industrial.
Na indústria papeleira já há um número de aplicações biotecnológicas implementadas,
nomeadamente o biobranqueamento com enzimas hidrolíticas ou oxidativas (xilanase),
aumentos de controlo dos pitches com a lipase.
O cultivo de fungos do género Trametes também tem atraído um considerável
interesse comercial devido ao grande potencial na área terapêutica. Sabe-se que na
antiguidade muitos povos já utilizavam fungos com finalidade medicinal e as suas
aplicações são mantidas até hoje na medicina popular, principalmente nos países orientais.
Actualmente, a existência de substâncias terapeuticamente activas de fungos do género
Trametes tem sido comprovada, com efeitos como actividades antiinflamatória, antiviral,
antimicrobiana, antitumoral, entre outras. Dentre estas, a actividade antitumoral tem-se
xxi

Introdução
mostrado a mais interessante dado o número crecente de pesquisas envolvendo este tema e
a busca incessante do combate ao cancro. Uma das possibilidades de obtenção de
substâncias com potencialidades terapêuticas, é a partir da produção destes fungos em
cultura submersa. Neste tipo de processo, os fungos produzem um polissacarídeo
extracelular, ou exopolissacarídeo que devido às possibilidades de aplicação na indústria
farmacêutica, tem vindo a despertar grande interesse na sua produção.
Este trabalho teve como objectivo geral estudar o fungo da podridão branca da
madeira, Trametes (Coriolus) versicolor nos processos de degradação da lenhina, na
produção de enzimas lenhinolíticas e na produção de um exopolissacarídeo. Os objectivos
específicos foram: i) optimizar a produção da enzima lacase por Trametes versicolor em
matraz através da adição de diferentes indutores (Cu, xilidina e mistura fenólica) e do
emprego da limitação por carbono no meio de cultura; ii) Determinar os efeitos da
concentração inicial de glucose, pH inicial do meio de cultura e agitação na produção de
lacase po Trametes versicolor em bioreator, através do planejamento de experiências 23;
iii) Aplicação da lacase juntamente com diferentes mediadores no branqueamento da pasta
kraft de papel; iv) Optimizar a produção do exopolissacarído por Trametes versicolor
através da seleção de diferentes meios de cultura e posterior optimização do meio de
cultura seleccionando.
xxii

Revisão Bibliográfica
Capítulo 1 - Revisão bibliográfica
1.1 A indústria de pasta de papel
A indústria da pasta e do papel compreende uma grande e crescente porção da
economia mundial. A produção de pasta e de papel tem aumentado globalmente e irá
continuar a aumentar num futuro próximo, comprovado pelo aumento contínuo da
produção e do consumo de pasta e papel nas últimas décadas. Trata-se de uma indústria de
capital intensivo que acompanha de perto os desenvolvimentos tecnológicos, procurando
responder às exigências dos consumidores, nomeadamente através do cumprimento de
critérios ambientais.
A actividade principal desta indústria tem que ver com as várias etapas do processo
produtivo do papel iniciando-se na produção de madeira (a indústria papeleira portuguesa é
responsável pela gestão directa de cerca de 200.000 ha de floresta), a sua exploração e
transformação em pasta para papel, e a transformação de pasta em diferentes tipos de
papeis. Duas visões opostas se confrontam quanto ao futuro da indústria do papel. Os
defensores das novas tecnologias de informação e comunicação prevêem o declínio da
utilização do papel, enquanto a indústria do papel contrapõe com a experiência do passado,
afirmando que a procura de papel continuará a crescer a nível mundial, induzida em parte
pelas novas aplicações decorrentes do desenvolvimento da electrónica e das
telecomunicações (Gouveia, 2005).
Entre 1980 e 1997 a média anual de crescimento da produção de papel e cartão na
Europa foi de 3,3%, acompanhando o progresso económico. Estima-se que a procura de
papel, na Europa Ocidental, cresça a uma taxa estável de 2% ao ano até 2010. Em 1998, da
produção de papel na U.E., 52% refere-se a papel de uso gráfico, 38% a papel de
embalagem e os restantes 10% são papéis para usos sanitários e outras espécies (Gouveia,
2005).
Os maiores países produtores mundiais de pasta para papel localizam-se na
América do Norte (E.U.A. e Canadá), na Ásia (República Popular da China e Japão) e na
1

Revisão Bibliográfica
Europa. Os principais países produtores de pasta para papel de zonas económicas
emergentes são o Brasil (7ª posição mundial) e a Indonésia (9ª posição mundial). Portugal
ocupa a 16ª posição (Melo & Gouveia, 2005).
A população global utilizou aproximadamente 300 milhões de toneladas de papel
em 1996 (Akhtar et al. 1997), e as estimativas mostram que este número está a aumentar
gradualmente com o passar dos anos e que poderá chegar a 420 milhões de toneladas/ano
no ano de 2010, o que corresponde a uma taxa de crescimento de 2.4% ao ano (MBendi,
2005).
Em Portugal, a indústria papeleira é um sector estratégico para a economia
nacional, e tem apresentado um desempenho particularmente positivo ao longo dos últimos
20 anos. A produção de pasta ocupa a 6ª posição e a produção de papel a 12ª, em termos da
U.E., registando-se, entre 1992 e 1998, um crescimento médio anual de 1,2% e 2,9%,
respectivamente. Trata-se de uma indústria que exporta o grosso da sua produção para os
países da U.E., com predominância das pastas branqueadas de eucalipto ao sulfato, de
grande qualidade. No ano de 2000 a produção total de pasta para papel foi de 1 774 mil
toneladas (Gouveia, 2005) e em 2004 de 1 949 mil toneladas (CELPA, 2005).
A crescente consciência ambiental e a exigência de qualidade e segurança ao nível
dos produtos de origem florestal foram, numa fase inicial, os factores que levaram o sector
a actuar de uma forma ecologicamente responsável. Como em outras indústrias de grande
escala, a indústria papeleira exerce o seu próprio impacto no meio ambiente. Embora a
situação esteja a melhorar de ano para ano, as fábricas de pasta e de papel ainda libertam
compostos de enxofre no ar e descargas de efluentes que aumentam a eutrofização e que
têm efeitos tóxicos no biota em seu redor. Compostos organoclorados são formados
durante o branqueamento químico da pasta, e têm atraído muita atenção das indústrias e
dos investigadores.
Durante as últimas décadas, vários estudos têm sido desenvolvidos na área do
branqueamento do papel, de modo a desenvolver novas tecnologias sustentáveis para
minimizar a poluição ambiental, aumentar a qualidade da pasta e obter melhorias
económicas. A biotecnologia tem emergido com grande potencial tecnológico para a
modernização da indústria da pasta e do papel, com o uso de enzimas de origem
microbiológica. Devido aos grandes problemas ambientais, têm-se criado oportunidades no
2

Revisão Bibliográfica
ramo da biotecnologia para substituir o uso de cloro nas etapas do branqueamento da pasta,
reduzindo assim, uma parte dos compostos tóxicos, assim como a coloração escura dos
efluentes, que são gerados durante o processamento da pasta e do papel, minimizando o
impacto ambiental.
Actualmente, o objectivo geral da indústria da pasta e do papel é aumentar a
eficiência do processo, reduzindo os custos, desenvolvendo processos benignos ao meio
ambiente e aumentando a qualidade do produto. A biocatálise tem apresentado um grande
potencial para melhorar o processo tradicional de manufactura da pasta e do papel, além de
introduzir propriedades únicas à fibra do papel. As fibras podem ser modificadas por
enzimas ou microorganismos para melhorar os processos químicos ou mecânicos da
indústria papeleira. O grande desafio para o sucesso comercial é identificar enzimas,
microorganismos e as condições para desenvolver às suas aplicações. A actividade
catalítica e a estabilidade de potenciais enzimas podem ser melhoradas pelos novos
métodos que estão a ser desenvolvidos. Durante os últimos anos, o número de novas
aplicações de enzimas tem crescido e algumas têm alcançado aplicações comerciais ou
aproximações, onde se inclui o branqueamento com xilanase ou microorganismos,
deslenhificação directa com enzimas oxidativas, redução do consumo de energia com
celulases, redução de pitches com lipases entre outras (Viikari & Lantto, 2002).
O uso da enzima xilanase no branqueamento da pasta de papel foi o primeiro passo
essencial da aplicação da biotecnologia, demonstrando que as enzimas são um meio
tecnológico eficiente e que podem ser introduzidos nas plantas industriais já existentes,
sem grandes investimentos. Entretanto, verificou-se que o efeito da xilanase no
branqueamento é limitado e que se obtém somente uma redução de cerca de 25% dos
compostos químicos (Bourbonnais et al., 1997). Recentemente, outras enzimas como a
lipase e a glucanase também foram testadas em aplicações de grande escala (Messer et al.,
1997).
1.1.1 Composição da madeira
A madeira é um biocompósito natural constituído principalmente por fibras. É um
tecido biológico altamente organizado a nível celular e molecular. Sua composição
3

Revisão Bibliográfica
química apresenta cerca de 20-30% de lenhina, 65-80% de polissacarídeos (dos quais 40-
50% são celulose e 20-35% são hemiceluloses), de outras substâncias de baixo peso
molecular (1-3%) representadas essencialmente por extractáveis (compostos alifáticos:
alcanos, álcoois, ácidos gordos; terpenos e terpenóides: esteróis e compostos fenólicos:
ácidos fenólicos, flavonóides, taninos, lenhanos e estilbenos), de cinzas (menos que 1%) e
por compostos minerais presentes em pequenas quantidades (Sjöström, 1981). As
quantidades relativas dos componentes da madeira variam com o tipo de madeira e com o
tipo de parede celular. Geralmente classifica-se a madeira em dois tipos: as
gimnospérmicas ou madeira dura (hardwood), ex. folhosas e as angiospérmicas ou mole
(softwood), ex. resinosas.
1.1.1.1 Celulose
A celulose é um polissacarídeo, um homopolímero linear constituído por unidades
repetidas de celobiose β-D-glucopiranose, que são ligadas umas as outras por ligações
glicosídicas β-(1→4). As suas cadeias apresentam entre 4000 a 10000 unidades de D-
glucose anidra. A Figura 1.1 mostra a cadeia principal da celulose.
Figura 1.1 – Representação da cadeia principal da estrutura de celulose (Higuchi, 1993).
A celulose está estruturalmente ligada à hemicelulose e à lenhina, não sendo assim
um substrato facilmente acessível. Fisicamente, a celulose é um material sólido e branco
que existe no estado amorfo-cristalino. A forma cristalina é resistente ao ataque químico e
à degradação microbiana enquanto que a forma amorfa é primeiramente atacada e
degradada (Biermann, 1996).
As moléculas de celulose são lineares e ligam-se umas as outras por pontes de
hidrogénio intra e intermoleculares, formando fibrilas elementares (FE) (Figura 1.2).
4

Revisão Bibliográfica
Várias FE de celulose são agregadas umas as outras formando as microfibrilas. Estas
microfibrilas são constituídas por regiões cristalinas rigorosamente ordenadas alternando
com regiões amorfas menos ordenadas. Microfibrilas formam as macrofibrilas e estas
formam fibras. As suas estruturas lineares e cristalinas são responsáveis pela rigidez
atribuída à madeira (Kirk & Fareel, 1987).
Os mais eficientes degradadores biológicos da celulose são os fungos filamentosos.
Tipicamente, as endoglucanases e as celobiohidrolases hidrolisam a celulose cristalina em
celobiose e a β-endoglucanase degrada a celobiose em glucose (Teeri, 1997).
(a)
Microfibrilas
(b)
Unidades de glucose
Ligações glicosídicas
Ligações de H
Figura 1.2 – Diagrama estrutural da celulose mostrando as ligações β-(1,4) dos resíduos de D-glucose e
as ligações de hidrogénio inter e intra molecular (a) e representação das microfibrilas da celulose (b) (Adaptado de Voet & Voet, 1990).
1.1.1.2 Hemiceluloses
As hemiceluloses são um grupo misto de polissacarídeos não celulósicos lineares
e/ou ramificado que compreende dois grandes grupos: pentosanos e hexosanos. Os
pentosanos são constituídos de unidades de pentoses (D-xilose, L-arabinose) e os
hexosanos são constituídos de unidades de hexoses (D-glucose, D-galactose, D-manose).
Também apresentam pequenas quantidades de L-ramnose, L-fucose e ainda desoxi-
5

Revisão Bibliográfica
hexoses e ácidos urónicos: ácido D-glucurónico, ácido 4-O-metil-D-glucurónico e ácido D-
galacturónico.
Os monossacarídeos ligam-se uns aos outros essencialmente por ligações
glicosídicas β-(1→4), mas também se podem encontrar ligações glicosídicas β-(1→3), β-
(1→6), α-(1→2), α-(1→3) e α-(1→6). As hemiceluloses e a lenhina são ligadas
covalentemente. Devido ao baixo grau de polimerização e à sua natureza amorfa, as
hemiceluloses são degradadas mais facilmente do que a celulose. Apesar disso, ainda é
necessário um sistema enzimático complexo para a sua degradação, devido à sua estrutura
variável e ramificada (Sjöström, 1981).
Tal como a celulose, grande parte da função das hemiceluloses é dar resistência à
parede celular, actuando como matriz de suporte para as microfibrilhas de celulose (Kirk &
Fareel, 1987). Fisicamente, são um material sólido e branco, raramente cristalino ou
fibroso. As hemiceluloses aumentam a resistência do papel e o rendimento da pasta. São
muito mais solúveis e susceptíveis a degradação química. O baixo peso molecular faz com
que sejam solúveis em soluções alcalinas diluídas a altas temperaturas, como acontece no
cozimento kraft (Biermann, 1996). A sua estrutura varia de acordo com o tipo de madeira.
Nas madeiras resinosas a hemicelulose consiste principalmente de glucomananas,
galactoglucomananas, arabinanas enquanto que nas madeiras folhosas consiste de
glucuroxilano e pequenas quantidades de glucomananas (2-5%).
1.1.1.3 Lenhina
A lenhina é o segundo maior componente celular da madeira. Ela estabelece as
ligações entre as fibras da madeira, conferindo firmeza e rigidez às fibras. Ela é encontrada
entre as células e a parede celular e é resistente ao ataque biológico, uma vez que não
apresenta ligações que sejam hidrolizáveis. É um polímero de estrutura amorfa, aromática,
altamente ramificada e insolúvel em água. Apresenta–se sob uma rede tridimensional com
ligações cruzadas (Sjöström, 1981).
É um heteropolímero complexo constituído por unidades de fenilpropano (C9) que
apresenta um elevado peso molecular (600-10000 kDa). Durante o tratamento químico, a
sua remoção permite que as fibras de celulose e hemicelulose sejam separadas facilmente
6

Revisão Bibliográfica
(Kirk & Cullen, 1998). Ela apresenta um carácter hidrofóbico, que na presença de pastas
inibe a absorção de água e o inchamento das fibras.
A lenhina é formada a partir dos álcoois p-cumarílico, coniferílico e sinapílico
(Kuad et al. 1997), os quais formam respectivamente as subunidades hidroxifenilo (H),
guaiacilo (G) e seringilo (S) que compõem a lenhina (Boudet et al., 1995). O álcool
coniferílico é percursor das unidades guaiacilo e o álcool sinapílico origina as unidades
seringilo e o álcool cumarílico dá origem às unidades p-hidroxifenilo (Kirk & Fareel,
1987). Estes percursores são formados a partir da glucose por uma variedade de reacções
enzimáticas que envolvem oxidações, reduções, aminações, descaboxilação entre outras. A
Figura 1.3 mostra os álcoois percursores da lenhina. A lenhina de madeiras moles é
constituída principalmente de unidade de G, enquanto que as duras de G e de S.
OH
CH
OCH3
CH
CH2OH
OH
CH
OCH3
CH
CH2OH
OH
CH
CH
CH2OH
H3CO
Álcool coniferílico Álcool sinapílicoÁlcool cumarílico
Figura 1.3 – Álcoois percursores da lenhina (Adaptado de Biermann, 1996).
Estudos recentes mostram que a polimerização desidrogenativa de monómeros de
lenhina é causada por uma classe de enzimas, as peroxidases ou o sistema peroxidase-
H2O2. Estas enzimas são capazes de oxidar um percursor, criando radicais livres
estabilizados por ressonância, que se polimerizam por acoplamento destes de radicais.
(Kilpeläinen et al., 1994). A combinação de radicais monoméricos com os grupos
fenólicos conduz à formação de um polímero linear, enquanto que as ramificações do
polímero ocorrem através da formação de estruturas benzil aril éter.
A Figura 1.4 mostra a estrutura da lenhina. A ligação dominante é do tipo β-O-4.
Mais de dois terços da unidade de fenilpropano na lenhina está ligada por ligações éter C-
7

Revisão Bibliográfica
O-C e as unidades restantes por ligações carbono-carbono C-C. Os principais tipos de
ligações são alquil-O-aril, alquil-O-alquil, aril-O-aril, alquil-alquil, aril-aril e alquil-aril.
Estas ligações explicam as fortes condições necessárias para a despolimerização da lenhina
e a incapacidade de originar a reversão para monómeros (Sjöström, 1981).
Figura 1.4- Estrutura de uma lenhina resinosa (Sipilä et al., 2005).
8

Revisão Bibliográfica
Os grupos funcionais da lenhina com maior influência sobre a reactividade da
lenhina consiste de grupos carbonilo, grupos hidroxilos fenólicos e grupos metoxilos
(Sjostron, 1981).
As lenhinas são diferentes consoante a espécie de madeira e variam de acordo com
o conteúdo de unidades de seringilo, guaiacilo e p-hidroxifenilo e podem ser classificadas
em várias classes de acordo com os seus elementos estruturais. A lenhina é covalentemente
associada à hemicelulose, na parede celular, por numerosos tipos de ligações. As mais
importantes são as ligações éter entre o carbono benzílico da lenhina e uma das partes do
hidrato de carbono e as ligações éster entre o carbono benzílico da lenhina e os resíduos de
ácidos (Hammel, 1997).
A lenhina é uma molécula altamente irregular que não tem nenhuma estrutura
precisa, mas contém uma série de variadas substruturas, conforme mostra a Figura 1.5.
Considerando que o polímero da lenhina é grande e altamente ramificado, deduz-se
que os mecanismos lenhinolíticos devam ser extracelulares. Estes mecanismos devem ser
oxidativos em lugar de hidrolíticos, uma vez que o polímero é interligado por ligações
estáveis de éter e de carbono-carbono. Como a lenhina consiste numa mistura
estereoirregular de unidades, os agentes lenhinolíticos certamente serão muito menos
específicos que os catalisadores biológicos típicos (Hammel, 1997). Somente uma classe
de microorganismo é capaz de degradar a lenhina, os basidiomicetes.
9

Revisão Bibliográfica
b c a
d
e f
g h i
j k
Figura 1.5 - Principais subestruturas na lenhina; β-O-4’ (a); β-O-4’ degradada (b); α-O-4’
acíclica (c); β-5’ cíclica e α-O-4’ (d); α-2’/6’ e β-2’6’ (e); 5,5’ (f); 4-O-5’(g); β-1’ (h); β-β’ (i); éster α-HC-hidroxicinamato de arilglicerilo (com ligação α-4) (j); éster γ-HC-hidroxicinamato de arilglicerilo (com ligação γ-4) (k) (Sjöström, 1981).
10

Revisão Bibliográfica
1.1.2 O processo de fabricação da pasta de papel
As pastas de madeira são o material de partida para a produção do papel. A
produção da pasta pode ser dividida em quatro partes principais: preparação de matérias-
primas, deslenhificação, branqueamento e sistema de tratamento de águas residuais.
Durante o processamento de produção da pasta, os seguintes processos podem ser
utilizados (Akhatar et al., 1997; Ander et al., 1998):
(i) mecânico → a madeira é destroçada, as cascas são trituradas e as fibras
reduzidas à uma pasta fibrosa denominada “pasta mecânica”. Neste processo não ocorre
uma separação completa das fibras dos demais constituintes, obtendo-se então uma pasta
com baixa qualidade, cuja aplicação é limitada.
(ii) termomecânico → a madeira, sob forma de aparas, sofre um aquecimento com
vapor (em torno de 140° C), seguindo para o processo de desfibramento em refinador a
disco. A pasta obtida desta forma resulta em celulose para a produção de papéis de melhor
qualidade, pois proporciona maior resistência mecânica.
(iii) semi-químico → neste processo acrescentam-se produtos químicos em baixas
porcentagens para facilitar ainda mais a desfibragem. O mais comum desses processos é
conhecido na Europa com a sigla NSSC (neutral sulphite semi chemical).
(iv) quimiotermicomecânico → a pasta derivada do processo mecânico sofre um
pré-tratamento com sulfito de sódio ou álcali antes da desfibragem.
(v) químico – kraft (ou ao sulfato) → é processo de fabricação da pasta de papel
mais utilizado. Pode ser utilizado em quase todas as espécies de madeira. A madeira é
tratada em vasos de pressão, denominados digestores, com hidróxido de sódio e sulfeto de
sódio. É um processo químico que visa dissolver a lenhina, preservando a resistência das
fibras, obtendo-se dessa maneira uma pasta forte (kraft significa forte em alemão), com
rendimento entre 50 - 60% (Kirk & Farrell, 1987).
(vi) químico – sulfito → é um processo em que os cavacos são cozidos em
digestores com um licor ácido, preparado com compostos de enxofre (SO2) e uma base
Ca(OH)2, NaOH, NH4OH etc. A pasta obtida desta maneira tem um rendimento entre 40 e
11

Revisão Bibliográfica
60 % e é de branqueamento muito fácil, apresentando uma coloração clara que permite o
seu uso mesmo sem ser branqueada. Este processo, que era muito utilizado para a
confecção de papéis para imprimir e escrever, está sendo substituído pelo processo sulfato
(principalmente após a introdução do dióxido de cloro no branqueamento), devido à
dificuldade de regeneração dos produtos químicos e as conseqüentes contaminações das
águas.
1.1.3 Branqueamento da pasta kraft
O branqueamento é um processo químico aplicado à pasta crua para aumentar a sua
brancura por remoção da lenhina residual da fibra. Neste processo deseja-se que um
mínimo possível ou nenhum dano ocorra aos polissacarídeos, particularmente a celulose,
que é a substância primária da parede celular da madeira e o constituinte chave das fibras
naturais do qual o papel é feito.
A determinação do conteúdo de lenhina na pasta é importante para a produção de
papel branco. O alto conteúdo de lenhina dificulta e encarece o branqueamento da pasta,
enquanto que o baixo conteúdo de lenhina pode favorecer o branqueamento (McDonald,
1970).
A determinação rápida do conteúdo de lenhina, durante o processamento da pasta,
nem sempre é fácil. A oxidação da madeira ou da pasta com soluções padrão de
permanganato de potássio constitui um método convencional para a determinação do
conteúdo de lenhina, e foi aprovado pela Technical Association of the Pulp and Paper
Industry (TAPPI). A lenhina da pasta reage com permanganato e a quantificação do
permanganato consumido é determinada por titulação com iodeto de potássio. Esse método
determina o Índice Kappa, que é o método industrial padrão de determinação do conteúdo
de lenhina. O Índice Kappa é definido como a quantidade de permanganato que é
consumido pela pasta. Uma vez que o permanganato é consumido pela lenhina, o Índice
Kappa é uma medida indirecta da percentagem de lenhina na pasta (Smook, 1989).
A maioria dos sistemas de deslenhificação são agressivos, particularmente quando
o nível de lenhina diminui, de forma que a deslenhificação a níveis desejáveis faz com que
ocorra alguma degradação nas propriedades da fibra.
12

Revisão Bibliográfica
Os agentes químicos oxidantes utilizados podem ser o hipoclorito de sódio
(NaClO), o oxigénio (O2), os peróxidos de hidrogénio (H2O2), o dióxido de cloro (ClO2), o
cloro (Cl2) e o ozono (O3).
Os estágios de branqueamento são designados por símbolos de acordo com o
agente químico utilizado: Z - ozono – usado na forma de gás; O – oxigénio; P – peróxido
de hidrogénio – em estado líquido em meio alcalino; D - dióxido de cloro – em solução
(ClO2); C – cloro; E - extração alcalina – usando NaOH; E/O - extração alcalina usando
oxigênio; E/P - extração alcalina usando peróxido; Q - Estágio ácido onde é adicionado um
agente quelante (EDTA ou DTPA);
Dependendo do agente de branqueamento e da sequência utilizada, o
branqueamento da pasta é denominado: STD - Standard (com uso de cloro molecular,
sequência (C+D)(EO)DED); ECF - Elementary chlorine free (sem uso do cloro molecular,
sequência D(EO)DED)¸ TCF - Totally chlorine free (sem uso de compostos clorados,
sequência OQP), entre outros (Dence & Reeve, 1996).
A sequência de branqueamento típico com ClO2 pode ser descrita com uma
sequência de operações do tipo DEDED em que são aplicados agentes químicos
sequencialmente, com extracções intermédias. Na primeira etapa (D) ocorre a oxidação e
fragmentação da lenhina, mas como a dissolução é apenas parcial, é necessário a etapa de
extracção alcalina posterior. Este procedimento é realizado duas vezes e termina com uma
terceira etapa com ClO2. No final do processo DEDED a pasta é lavada. A pasta branca
resultante é essencialmente constituída por celulose e hemiceluloses e contem quantidades
vestigiais de lenhina (0.1-0.3% no processo kraft) (Biermann, 1996; Reeve, 1989).
Uma das alternativas mais atractivas ao dióxido de cloro e ao cloro, no que respeita
ao impacto ambiental e aos custos, são o oxigénio, o peróxido de hidrogénio e o ozono.
Entretanto a limitação do uso destes compostos é geralmente a sua selectividade para a
lenhina (vs. celulose). A alta selectividade é importante para manter as propriedades de
resistência física da fibra. O oxigénio não é suficientemente selectivo para remover mais de
metade da lenhina residual da pasta kraft. O peróxido de hidrogénio não deslenhifica
efectivamente. O ozono é muito efectivo, no entanto limitações de transferência de massa
impostas pela necessidade de empregar ozono em concentrações diluídas de lenhina,
13

Revisão Bibliográfica
presentes dentro das fibras primárias da parede celulósica, dificultam a selectividade do
processo em escala industrial (Weinstock et al., 1997). No entanto, numerosas tecnologias
alternativas e de biobranqueamento estão a ser consideradas. Têm sido desenvolvidos
vários estudos visando obter um processo de tratamento da pasta que seja adaptável às
instalações já existentes e compatível com a legislação ambiental em vigor.
Estudos da biodegradação da lenhina têm grandes implicações não só no
entendimento do ciclo do carbono global, como também no desenvolvimento de técnicas
não prejudiciais ao meio ambiente para a remoção selectiva da lenhina da madeira na
indústria do papel. Diferentes tipos de microorganismos são capazes de degradar a
madeira, no entanto os fungos da podridão branca da madeira são os microorganismos
mais eficientes em degradar a lenhina, conforme discutido anteriormente. Estudos com os
fungos da podridão branca da madeira e suas enzimas têm revelado três tipos diferentes de
potenciais enzimas lenhinolíticas: lenhina peroxidase (LiP), manganês peroxidase (MnP) e
lacase. T. versicolor é um dos fungos mais extensivamente estudado na biodegradação da
lenhina. O uso de enzimas para a remoção da lenhina tem sido proposto como uma via para
a redução do consumo de dióxido de cloro na indústria do papel (Tolan & Thibault, 1997)
1.1.3.1 O papel da lacase no branqueamento da pasta
Muitas enzimas são conhecidas por actuar sobre a lenhina, especialmente as
oxidativas como é o caso da lacase. Ela pertence ao grupo da polifenol oxidase e esta
usualmente é considerada como sendo somente capaz de oxidar compostos fenólicos da
lenhina devido ao seu baixo potencial redox, quando comparada com a MnP ou LiP
(Kersten et al., 1990).
Nas últimas décadas, tem-se feito um forte esforço para se compreender os
mecanismos da biodegradação da lenhina pela lacase e para encontrar uma aplicação
enzimática industrial, principalmente no branqueamento da pasta kraft (Call & Mucke,
1997). Muitos estudos têm descrito a aplicação da lacase ao branqueamento de pasta kraft.
14

Revisão Bibliográfica
Katayama et al. (1989) produziram a primeira evidência de quebra do anel
aromático de um tipo de estrutura bifenílica da lenhina (DDVA; 2,2'-dihidroxi-3.3'-
dimetoxi - 5,5 '-dicarboxi bifenil) por T. versicolor.
1.1.3.2 Uso de mediadores
Juntamente com a lacase, certos compostos químicos, chamados de mediadores,
podem ser utilizados no branqueamento da pasta de papel, uma vez que o efeito catalítico
da lacase é insuficiente para muitos processos industriais. O tamanho molecular da lacase
não permite a sua penetração na parede celular do material lenhinocelulósico. O uso de
compostos mediadores torna possível a biodeslenhificação de pastas, uma vez que os
compostos não fenólicos da lenhina podem ser oxidados com o uso em conjunto da lacase
e de mediadores.
O sistema lacase-mediador (SLM) constitui uma alternativa promissora às
convencionais tecnologias de branqueamento da pasta de papel (Leonowicz et al. 2001).
Este sistema apresenta as seguintes vantagens (Smith & Ragauskas, 2005):
• Remove selectivamente a lenhina da pasta;
• Aumenta o rendimento da produção de pasta;
• Reduz o capital e despesas operacionais da deslenhificação;
• Minimiza a geração de ião cloreto durante o branqueamento;
• Reduz o uso de água fresca;
• Exibe verdadeiras propriedades catalíticas.
A Tabela 1.1 mostra o nome e a estrutura dos principais mediadores orgâncicos da
lacase já estudados.
Call (1994), mostrou que é possível obter mais de 55% de deslenhificação da pasta
kraft com lacase e mediador (1-hidroxilbenzotriazol (HBT)).
Na presença do mediador, HBT, um composto modelo de subestrutura de lenhina
15

Revisão Bibliográfica
β-O-4, não fenólico, foi oxidado pela lacase de T. versicolor originando quatro produtos.
Os resultados mostram que três tipos de reacções, quebra β-éter, quebra Cα-Cβ e oxidação
do Cα foram catalisadas pelo par lacase-HBT (Shingo, 1999).
O entendimento do mecanismo de oxidação da lenhina pelo SLM deverá facilitar a
comercialização desse sistema pela indústria do papel. Estes mediadores formam radicais
reactivos que podem penetrar nas fibras da pasta alcançando a lenhina melhor do que a
grande molécula de lacase. Deste modo a lenhina é degradada e sua repolimerização é
evitada. Estudos avaliam o papel de diferentes mediadores juntamente com a lacase,
principalmente em reacções de oxidação de compostos modelo da lenhina (Fabbrini et al.
2002; Bourbonnais et al., 1997; Castro, 2001). Os mediadores mais frequentemente
estudados são: HBT (Li et al. 1998; Castro et al., 2003; Majcherczyk et al., 1998¸
Bourbonnais et al., 1997), Ácido 2,2’-azino-bis(3-etilbenzotriazolina-6-sulfónico) (ABTS)
(Bourbonnais et al., 1997), N-oxil-2,2,6,6-tetrametilpiperidina (TEMPO), Ácido violúrico e
Polioxometalatos (Castro et al., 2003).
Tabela 1.1 - Estrutura de alguns mediadores orgânicos da lacase
Mediador Estrutura
1-hidroxilbenzotriazol (HBT)
Ácido 2,2’-azino-bis(3-etilbenzotriazolina-
6-sulfónico) (ABTS)
N-oxil-2,2,6,6-tetrametilpiperidina
(TEMPO)
Ácido violúrico
16

Revisão Bibliográfica
O mecanismo da oxidação da lenhina com SLM está apresentado na Figura 1.6,
onde os electrões que saem da lenhina são transferidos para o oxigénio, que apresenta o
papel de aceitador final de electrões. O mediador funciona como um transportador de
electrões difusível entre a lenhina e a enzima. Uma vez oxidado pela enzima, ele difunde-
se da porção enzimática e volta a oxidar a lenhina, que devido ao tamanho, não entra na
porção enzimática.
mediadorox mediadorred
lacase lacaseox
O2
H2O
lenhina lenhinaox
Figura 1.6 - Representação esquemática da degradação da lenhina por lacase num sistema com mediador (Adaptado de Bourbonnais et al., 1998).
Um estudo com uma série de lacases de origem fúngica demonstrou uma correlação
entre o potencial redox da enzima e actividade frente a um determinado substrato. A lacase
apresentou um potencial próximo a 0.7-0.8 V (vs. ENH). Este estudo mostrou que a
actividade da lacase face à lenhina em sistemas com mediador é dependente de uma
combinação de dois principais factores: o potencial redox da enzima e a estabilidade e
reactividade dos radicais gerados pela oxidação da lenhina com o mediador (Xu et al.,
1996).
A investigação por voltametria cíclica de alguns mediadores da lacase mostrou que
um importante papel na determinação do mecanismo da oxidação de um substrato pode
estar relacionado por um lado com a estabilidade da forma oxidada do mediador e por
outro com o seu potencial redox (Fabbrini et al., 2002).
Além da aplicação à pasta de papel, tem-se descrito o uso de mediadores e lacase
na degradação de hidratos de carbono aromático policíclicos, na síntese química e no
branqueamento de corantes têxteis (Bourbonnais et. al, 1998).
17

Revisão Bibliográfica
A deslenhificação catalítica das pastas kraft com POMs e lacase é uma tecnologia
não nociva ao meio ambiente e muito selectiva, que constitui uma alternativa promissora
ao convencional branqueamento (Call & Mucke, 1997).
1.1.3.2.1 Os polioxometalatos
As potencialidades económicas e ambientais dos polioxometalatos (POMs) tornam-
os muito promissores (Kozhevnikov, 1998). Outras capacidades específicas fazem dos
POMs uma nova tecnologia para o branqueamento e o processamento da pasta da madeira,
em particular (Evtuguin & Neto, 1997; Weinstock et al., 1997). Esta tecnologia promete
tanto vantagens económicas como ambientais sobre as actuais práticas na indústria
papeleira. O principal objectivo no uso de POMs é a sua aplicação em condições tais que
se consiga obter um alto nível de selectividade. Uma grande vantagem no uso dos POMs
como mediadores é que eles podem ser regenerados e reutilizados no processso de
deslenhificação.
Os POMs são caracterizados por apresentar um centro metálico, M, que é rodeado
por alguns átomos ou grupo de átomos. Nos POMs, os átomos ligantes são os átomos de
oxigénio. Em geral, eles apresentam unidades MOy, onde y indica o número de
coordenação de M. Além de M e O, outros elementos, que usualmente são designados
como X, podem fazer parte do estrutura do POM. Podem ser classificados como
isopolianiões (IPAs) ou heteropolianiões (HPAs).
Apresentam a fórmula geral:
[MmOY]m- (IPAs) (1.2)
[XxMmOY]n- (HPAs) (1.3)
onde x≤m, Mm = Mo, W, V, Nb, Ta e X variável = P, Si, Ge, B, As entre outros
(Gamelas et al., 2003). Xx representa o heteropoliátomo introduzido na estrutura e está
localizado no centro da molécula e Mm o adenoátomo. Os POMs apresentam uma estrutura
18

Revisão Bibliográfica
organizada em unidades discretas. Apesar das fórmulas 1.2 e 1.3 serem simples, a
composição de um grupo pode ser altamente complexa com vários elementos M fazendo
parte da estrutura. A unidade básica de construção desses POMs é o octaédro formado por
um metal rodeado de seis oxigénios (MO6). A Figura 1.7 mostra a unidade fundamental da
estrutura do POM.
Figura 1.7– Representação bola-bastão e poliedro da unidade fundamental de MO6 (Fernandez, 2003).
Essas unidades unem-se umas às outras. A Figura 1.8 mostra a união das unidades
de MO6.
Figura 1.8 – Modelos poliédricos representam as três possíveis uniões entre duas unidades octaédricas MO6. A) união de vértice, B) união de aresta e C) união de face (Fernandez, 2003).
Os POMs com estrutura de Keggin são os mais importantes e estudados
actualmente (Weinstock et al., 1997). A estrutura desses POMs é baseada em um tetraedro
19

Revisão Bibliográfica
central XO4, rodeado por doze octaedros MO6 arranjados em quatro grupos de três
octaedros cada (M3O13) (Pope, 1983). O heteroátomo encontra-se no centro formando a
estrutura tetraédrica XO4. Cada uma das unidades M3O10 compartilha os átomo de
oxigénio dos vértices com as outras três, formando assim o POM com estrutura de Keggin
e fórmula geral de:
[XM12O40]n- (XM12, forma abreviada) (1.4)
onde X é o heteroátomo (PV, AsV, GeIV, SiIV, BIII, FeIII; CoII, entre outros) e M é o adendo
átomo (MoIV; WVI). Os heteroátomos mais comuns nos aniões de Keggin são os
heteropolitungstatos [XW12O40]n- e os heteropolimolibdatos [XMo12O40]n-. O centro
externo de MmOY é livre de eléctrons metálicos se o arranjo é totalmente oxidado. A Figura
1.9 mostra a representação de um anião de Keggin.
XO4n-
M12O40
Figura 1.9 - Representação poliédrica da estrutura do anião de Keggin. O centro externo M12O40
encapsula a unidade interna, representada pelo tetraedro XO4n- (Jeannin, 1998).
Em solução, os potenciais de redução dos POMs com estrutura de Keggin são altos,
e por isso são reduzidos facilmente, mesmo em condições brandas, podendo depois ser
reoxidados pelo oxigénio, ar, ozono, H2O2, enzimas ou outro oxidante apropriado. Os
electrões são aceites pelos adenoiões do HPA (Sadakane & Steckhan, 1998). Entretanto,
apresentam algumas propriedades diferentes entre si como estabilidade, solubilidade,
acidez e potencial óxido-redução, que irão depender da espécie metálica (MoVI, WVI, etc.),
da presença ou ausência de algum outro metal incorporado, (VV, MnII, etc.) e do
heteroátomo central.
20

Revisão Bibliográfica
Uma característica dos POMs é a sua elevada solubilidade em água, que é
importante para a realização de experiências em catálise em fase líquida (Gamelas et al.,
2003; Hill, 1998 ). Outra característica bastante marcante é o potencial de óxido-redução,
que é mais influenciado pelo tipo de metal, do que pelo heteroátomo em si. Em relação aos
metais que compõe os POMs potenciais, o potencial redox é decrescente na seguinte
ordem: V >Mo>W. Os potenciais redox do POMs de Si são menores do que os de P. Uma
das maiores vantagens do uso dos POMs é que após serem reduzidos durante a reacção de
oxidação, eles podem ser reoxidados e reutilizados.
No branqueamento da pasta do papel, os POMs podem remover eléctrões dos
substratos orgânicos (reduzem-se) oxidando os fragmentos da lenhina dissolvida no licor
de branqueamento e ainda facilitando a transferência de eléctrões para o oxigénio. Na etapa
do branqueamento, aquece-se a altas temperaturas, uma mistura de água, pasta e POM
totalmente oxidado (POMoxi). Nestas condições de reacção, o POM é reduzido enquanto a
lenhina é oxidada. Após a reacção, o licor de branqueamento é separado da pasta e
reoxidado com o oxigénio conforme mostram as equações 1.5 e 1.6 (Katsoulis, 1998) e o
esquema apresentado na Figura 1.10:
Pasta + POMox → pasta branqueada + POMred +2H+ (1.5)
POMred + ½ O2 + 2H+ → POMox +CO2 + H2O (1.6)
O2
H2O
POMred POMox
lenhinaox lenhina
Figura 1.10 - Ciclos redox do POM na reacção catalítica da oxidação da lenhina (Balakshin et al. 2001).
Além dos mediadores orgânicos, os POMs também podem actuar como mediador
no processo de deslenhificação da pasta com lacase. A deslenhificação catalítica das pastas
kraft com POM e lacase (SLM) não é agressiva ao meio ambiente e é um processo
21

Revisão Bibliográfica
selectivo de deslenhificação. A Figura 1.11 mostra o mecanismo da oxidação da lenhina
pelo POM. A correspondente forma reduzida passa pela reoxidação directa pela lacase.
Do ponto de vista técnico, o SLM é eficiente e decompõe selectivamente a lenhina
residual sem significativa perda de viscosidade. Entretanto, uma das principais
desvantagens do SLM é a baixa estabilidade dos mediadores orgânicos (ABTS; HBT; etc.)
no sistema reaccional e a absorção de alguns mediadores coloridos na fibra.
Adicionalmente, considerando o alto custo dos mediadores, o SLM com mediadores
orgânicos não é viável ao contrário dos POMs que são muito efectivos como catalisadores
e podem ser reciclados no ciclo catalítico (Evtuguin et al., 1998).
Balakshin et al. (2001), demostraram pela primeira vez a possibilidade de usar o
sistema catalítico de POM com lacase para a deslenhificação da pasta kraft em condições
aeróbicas e a temperaturas moderadas (40-60 ºC). A lacase mostrou ser capaz de reoxidar
diferentes POMs, mesmo os que não são reoxidados pelo oxigénio a altas temperaturas.
O2
H2O
lacase lacaseox
POMox POMred
Lenhina Lenhinaox
Figura 1.11 – Ciclos redox no sistema catalítico de oxidação da lenhina com POM-lacase-O2 (Balakshin
et al., 2001).
1.2 A natureza dos fungos
Os fungos são organismos eucarióticos quimioorganotróficos. Apresentam dois
tipos morfológicos principais: os fungos filamentosos, assim chamados por formarem hifas
(ou filamentos revestidos de parede rígida) e as leveduras, fungos normalmente
unicelulares.
Os fungos constituem um grupo muito diversificado no que diz respeito à forma,
estrutura e capacidade metabólica. A maior parte desenvolve-se de matéria orgânica morta.
O seu crescimento é afectado por factores físicos e químicos como temperatura, humidade,
22

Revisão Bibliográfica
concentração de oxigénio, pH, micronutrientes, fontes de carbono e nitrogénio, entre
outros.
Nos fungos, os núcleos estão dispersos em um micélio (conjunto de hifas) contínuo
ou septado. Não possuem plastos ou pigmentos fotossintéticos e sua nutrição é obtida por
absorção. São saprófitas, parasitas facultativos ou biotróficos. São predominantemente
filamentosos, mas algumas espécies são leveduriformes.
Os fungos filamentosos são morfologicamente complexos e heterogéneos, e podem
exibir diferentes formas estruturais durante o seu ciclo de vida. A sua estrutura vegetativa
básica de crescimento é constituida por um filamento tubular, a hifa, que é originada a
partir de um único esporo reprodutivo (Papagianne, 2004). A colónia de um fungo
filamentoso é uma estrutura muito repetitiva. É composta de uma rede tridimensional
ramificada de hifas interconectadas e possui um indefinido número de pontos de
crescimento (Read, 1994). A área superficial relativa faz com que as hifas estejam bem
adaptadas para a absorção, segregação e excreção de substâncias. Quando crescem em
culturas submersas, os fungos filamentosos exibem diferentes formas morfológicas,
dispondo-se desde filamentos miceliares dispersos até densas massas miceliares agregadas,
chamadas de “pellets” (Papagianne, 2004). Os pellets são agregados de hifas apresentando
uma forma esférica ou elipsoidal, com uma estrutura interna variável. A forma particular
exibida é determinada não somente pelo material genético, mas também pelas condições
físicas e químicas do meio de cultura em que se encontram (Kossen, 2000; Znidarsic &
Pavko, 2001)
Reconhece-se a existência de três classes nesse grupo: basidiomicetes, ficomicetes
e ascomicetes. Os basidiomicetes (fungos superiores) são seres heterotróficos, necessitam
de meios eficientes de segregação de enzimas digestivas extracelulares. São providos de
talo pluricelular formado de hifas septadas. Seu micélio pode ter crescimento indefinido ou
formar, nos tipos mais evoluídos, corpos frutíferos de estrutura constante. A estrutura
reprodutora característica do grupo é o basídio, que é a extremidade de uma hifa dilatada,
uni ou pluricelular, e que emite ramificações, cada uma suportando em sua extremidade
um esporo, ou basidiósporo. Este grupo apresenta uma grande importância económica pois
compreende as espécies causadoras de uma série de doenças nas plantas cultivadas. Os
basidiomicetes superiores separam-se em dois grupos: os himenomicetes e os
gasteromicetes. Entre os primeiros estão os fungos destruidores da madeira.
23

Revisão Bibliográfica
1.2.1 Fungos da podridão branca da madeira
Os fungos da podridão branca da madeira e suas enzimas lenhinolíticas têm sido
muito estudados devido à sua grande eficiência em degradar a lenhina e as possíveis
aplicações na indústria papeleira (Akhtar et al., 1997), além de apresentar grande potencial
para a bioremediação de poluentes com estruturas similares a lenhina (Pointing, 2001).
Os substratos de crescimento para este tipo de fungo são a celulose e a
hemicelulose. A degradação da lenhina, que é altamente recalcitrante, ocorre no fim do
crescimento primário, através de um metabolismo secundário quando há deficiência em
nutrientes (Pointing, 2001). O ataque do fungo à lenhina é um processo oxidativo aeróbio,
onde são oxidados compostos fenólicos, metóxidos e alifáticos da lenhina através do
rompimento de anéis aromáticos e da criação de novos grupos carbonilo. Estas mudanças
na estrutura da lenhina resultam na sua despolimerização e na produção de dióxido de
carbono (Kirk & Farrell 1987).
Os fungos da podridão branca da madeira degradam a lenhina tanto selectivamente
como não selectivamente. Na degradação selectiva, tanto a lenhina como a hemicelulose
são significativamente mais degradas do que a celulose, enquanto que na degradação não-
selectiva, quantidades iguais de todos os componentes de lenhinocelulose são degradados.
Alguns fungos como Ganoderma applanatum, Heterobasidion annosum e Phellinus pini,
são capazes de realizar ambos os tipos de degradação (Blanchette, 1995).
1.2.2 O sistema lenhinolítico dos fungos da podridão branca da madeira
Vários tipos de enzimas lenhinolíticas, envolvidas na degradação da lenhina, são
produzidas pelos fungos da podridão branca da madeira. Além dessas, eles também podem
produzir celulases, xilanases e outras hemicelulases (Hatakka, 1994). Quase todos os
fungos da podridão branca da madeira produzem MnP e lacase, mas somente alguns
produzem LiP (Hatakka, 1994). LiP oxida as unidades não-fenólicas da lenhina (Kirk &
Farrell, 1987). MnP oxida Mn2+, que pode ser encontrado na madeira e no solo, a Mn3+. A
alta reactividade de Mn3+ faz oxidar os anéis fenólicos da lenhina em radicais livres
instáveis, que se decompõe espontaneamente (Hofrichter, 2002). A Lacase oxida os anéis
fenólicos em radicais fenoxílicos. Outras enzimas como a celobiose:quinona
oxidoreductase (CBQ), celobiose dehidrogenase (CDH), glioxalacto oxidase (GlOx),
24

Revisão Bibliográfica
glucose oxidase, álcool veratrílico oxidase (VAO) e algumas esterases também podem
apresentar algum papel no complexo processo da degradação da madeira, mas não actuam
sozinhas. Enquanto as enzimas lenhinolíticas oxidam compostos fenólicos e criam radicais
fenóxidos, os compostos não fenólicos são oxidados via catião radical (Call & Mucke,
1997; Garg & modi, 1999).
O sistema de degradação de materiais lenhinocelulósicos envolve uma série de
reacções. As espécies de basidiomicetos, incluindo as do género Trametes, são capazes de
realizar tal degradação, devido ao complexo enzimático lenhinocelulolítico específico. Um
esquema ilustrativo, onde se observa a função desempenhada pelas enzimas degradadoras
de materiais lenhinocelulósicos proposto por Rajarathnam et al. (1992) está apresentado na
Figura 1.12.
Celulose Hemicelulose Lenhina Proteínas
Forma alterada
Ácidos gordos
Glucose, xilose, etc.
Aminoácidos e peptídeos
Celubiose
Fenóis e compostos fenólicos
Indução de primórdios frutíferos
Fonte de C e energia
Glucose
Corpos frutíferos
Lipídeos
C1-celulase
Cx-celulase
β-Glucosidase
Hemicelulase Xilanase
Enzimas oxidativas Lacases Peroxidases lenhinases
Proteases Lipases Esterases
Figura 1.12 - Esquema da função desempenhada pelas enzimas degradadoras de materiais lenhinocelulósicos dos basidiomicetos (Adaptado de Rajarathnam et al., 1992).
25

Revisão Bibliográfica
1.2.3 O fungo Trametes versicolor
Trametes versicolor, por vezes classificado como Coriolus, Polyporus e Polysticus
é o mais comum habitante encontrado na madeira, causando a podridão branca
(Alexopoulus & Mims, 1979; Archibald et al., 1997). Este fungo pertence à classe dos
Basidiomycetes, ordem Polyporales, família Polyporaceae (Alexopoulus & Mims, 1979).
Existem cerca de 100 géneros nesta família.
É um cogumelo facilmente reconhecido pelo seu corpo frutífero (ou basidiocarpo)
que se apresenta em forma de “cauda de perú”, como mostra a Figura 1.13. As cores das
bandas podem variar dependendo da sua genética e do ambiente onde se encontram. A
maior parte das bandas são de cor castanha escura a clara, alternando- se com as bandas de
cores claras (as hifas).
Figura 1.13 – O fungo Trametes versicolor na forma de “cauda de perú”.
O seu ciclo de vida inicia-se com um esporo produzido pelo corpo frutífero
(cogumelo). Os esporos germinam e tornam-se hifas, que se agrupam formando o micélio
(fase vegetativa). Este por sua vez cresce originando o cogumelo (fase sexual)
(Alexopoulus & Mims, 1979).
T. versicolor é um dos mais eficientes fungos degradadores da madeira que
promove a deterioração simultânea da lenhina, celulose e hemicelulose (Tanaka et al.
26

Revisão Bibliográfica
1999). As enzimas oxidativas extracelulares produzidas por este fungo iniciam o ataque à
madeira. Esta espécie lenhinolítica é chamada de fungo da podridão branca porque confere
um aspecto esbranquiçado à madeira a medida que esta apodrece (Lewin & Goldstein,
1991). Estas características fazem com que este fungo e as suas enzimas lenhinolíticas
sejam estudados e/ou aplicados como alternativa ao branqueamento químico da pasta de
papel, na indústria têxtil para a remoção de cor (Swamy & Ramsay, 1999; Wong & Yu,
1999; Amaral et al. 2004), no tratamento de efluentes industriais (Modi et al. 1998) e na
bioremediação de uma série de poluentes orgânicos (Pointing, 2001).
Este fungo também é bastante usado na medicina tradicional do Este Asiático
devido às sua propriedades medicinais (Cui & Chisti, 2003). No Japão é chamado de
“Kawaratake” ou “cogumelo junto ao rio” e na China como “Yun Zhi” ou “ cogumelo
nuvem”. Na medicina tradicional este fungo é colhido, seco, moído e extraído com água
quente. Ele é usado no tratamento de infecções pulmonares, hepatite e em doentes com
cancro devido às suas propriedades imunomoduladoras (moduladoras do sistema
imunológico). Estas propriedades são atribuídas aos polissacarídeos do T. versicolor
(Sugiura et al. 1980; Ng, 1998). 1.3 Enzimas lenhinolíticas
Enzimas são proteínas globulares que actuam como catalisadores biológicos
conduzindo as reacções bioquímicas nas células dos organismos vivos. Estão grandemente
distribuídas na natureza. Quimicamente, as enzimas contêm uma ou mais cadeias de
centenas de aminoácidos numa complexa estrutura tri-dimensional, que é muito importante
para a sua acção. Como em toda a catálise, uma catálise enzimática é definida como uma
reacção de “parceria” que aumenta a velocidade de uma reacção química, sem que o
catalizador sofra mudanças em todo o processo. Esta definição implica que uma única
molécula de enzima é capaz de converter muitas moléculas de substrato durante o seu
tempo de vida. A taxa de conversão destas reacções é chamada de actividade de uma
enzima. As enzimas diferem dos catalisadores químicos em muitos aspectos, tais como: (i)
altas taxas de reacção; (ii) condições de reacção não agressivas e (iii) alta especificidade de
reacção (Call & Mucke, 1997). As enzimas são classificadas pela IUBMB (International
27

Revisão Bibliográfica
Union of Biochemistry and Molecular Biology) através do número EC, que as classifica de
acordo com a sua classe e com a reação que catalisa.
A interacção de uma enzima com o substrato pode ser pode ser ilustrada pelo
modelo “ chave -e- fechadura”. Somente um substrato (chave), que pode encaixar no
centro activo (fechadura), será convertido no centro catalítico, que é uma parte do centro
activo. A respeito desta imagem simplista, as reacções são certamente muito mais
complicadas e só podem ser descritas por modelos muito mais complexos. Ao contrário da
catálise química, as enzimas podem direccionar-se para reacções específicas através da
combinação de métodos altamente sofisticados com a engenharia genética e design
computacional de proteínas.
As enzimas são grandemente utilizadas como catalisadores na indústria, tal como
acontece na indústria da pasta do papel. Os fungos da podridão branca da madeira são os
principais produtores de enzimas lenhinolíticas. As principais enzimas lenhinolíticas são a
lacase, a manganês peroxidase e a lenhina peroxidase, mas outras enzimas como a CBQ, a
GlOx e algumas esterases também podem apresentarm um papel significativo no processo
de degradação da madeira.
1.3.1 Lenhina Peroxidase (LiP) (EC 1.11.1.14)
A LiP é uma glicoproteína hémica que catalisa uma variedade de compostos
fenólicos, não fenólicos, hidratos de carbono aromáticos e outros compostos que são
resistentes ao ataque microbiano. Requer peróxido de hidrogénio como cofactor e a
reacção ocorre através do mecanismo de oxidação de um electrão seguido por uma série de
reacções não enzimáticas (Garg & modi, 1999). Estas reacções incluem a degradação
oxidativa das ligações β-O-4, Cα-Cβ e outras ligações na lenhina. Os fungos segregam para
o meio de cultura várias isoenzimas que apresentam uma massa molecular entre 39 e 43
kDa e propriedades espectrais e catalíticas semelhantes (Kirk & Farell, 1987).
Alguns estudos mostram que, no T. versicolor, a LiP não é importante no
biobranqueamento da pasta de papel (Paice et al., 1993).
28

Revisão Bibliográfica
1.3.2 Manganês Peroxidase (MnP) (EC 1.11.1.13)
A MnP constitui o segundo grupo de glicoproteínas hémica que requer iões de
manganês livres para a sua actividade. A actuação desta enzima requer também a presença
de peróxido de hidrogénio. Inicialmente catalisa a oxidação do Mn2+ para o Mn3+, que
subsequentemente oxida vários compostos fenólicos (Garg & modi, 1999). Os fungos
também segregam várias isoenzimas da MnP para o meio de cultura, embora algumas
também possam estar ligadas à parede celular (Hatakka, 1994). A MnP purificada pode
desencadear a despolimerização de lenhinas sintéticas e também degradar clorolenhinas.
Verificou-se que inicialmente o sistema MnP pode igualar a extensão de branqueamento
obtida com o fungo, mas com o decorrer do tempo, os seus níveis diminuem.
1.3.3 Lacase (EC 1.10.3.2)
Lacase benzenodiol:oxigénio reductase, é uma polifenol oxidase, pertencente à
classe de enzimas oxidoreductase, também conhecida como oxidase de cobre azul. Pode
ser dividida principalmente em duas categorias segundo a sua origem: plantas e fungos. Foi
descoberta por Yoshida em 1883 nas plantas e caracterizada como um oxidase de cobre por
Bertrand em 1985 (Mayer & Staples, 2002). A lacase de fungos foi descoberta alguns anos
depois (Call & Mucke, 1997).
Os principais microorganismos produtores de lacase são os fungos da podridão
branca da madeira. T. versicolor é um dos mais estudados fungos da podridão branca da
madeira produtor de lacase (Garg & modi, 1999). Neste tipo de fungo, a lacase é produzida
dentro das células e excretada para o exterior dos filamentos das hifas.
O número de isoenzimas da lacase pode ser superior a 5, dependendo da espécie do
fungo e das condições ambientais de crescimento.
1.3.3.1 Caracterização estrutural da Lacase
A lacase pertence a família das enzimas de cobre azul, que pode ser definida pela
sua espectroscopia característica, pela sua sequência homologa e pela sua reactividade. Em
termos estruturais, esta enzima é uma glicoproteína que exibe uma heterogeneidade
adicional devido ao conteúdo variável de hidratos de carbono e de cobre (Yaropolov et al.,
29

Revisão Bibliográfica
1994). Compreende entre 520-550 aminoácidos, apresenta um peso molecular que varia de
60-80 kDa e é constituída por 15-20% de hidratos de carbono (Thurston, 1994).
Usualmente as lacases de fungos contêm 4 átomos de cobre distribuídos em 3 sítios
de ligação diferentes, que são identificados por densidade electrónica. O Cobre é
coordenado por duas histidinas e uma cisteína, que se encontra em um arranjo triangular
plano. Cada ião de cobre parece desempenhar um papel importante no mecanismo
catalítico (Thurston, 1994; Solomon et al., 1996). O primeiro sítio, tipo 1 (T1), é
responsável pela cor azul da enzima devido à máxima absorvância a 605 nm e é
coordenado com a cisteína; o tipo 2 (T2) não exibe sinal no visível e funciona como
aceitador de um electrão e o tipo 3 (T3) incorpora dois centros de cobre fortemente
acoplados, sendo responsável pela banda a 330 nm e funciona como aceitador de dois
electrões (Solomon et al., 1996). Diferentemente das outras oxidases de cobre, o quarto
ligante é substituído pela fenilalanina. Ela está organizada em três domínios arranjados
sequencialmente. A Figura 1.14 mostra a estrutura tridimensional da lacase obtida pelo T.
versicolor.
Figura 1.14 - Diagrama de da estrutura da lacase de Trametes versicolor. Os três domínios (T1, T2 e T3) estão mostrados a cores diferentes. Os átomos de cobre são as esferas azuis e a porção de hidrato de carbono são os modelos em bola-e-bastão. A parte cinzenta é o N-terminal e a verde o C-terminal. O sítio T1 é a esfera isolada. (Piontek et al., 2002).
30

Revisão Bibliográfica
1.3.3.2 Mecanismos de catálise da Lacase
A lacase catalisa a oxidação de uma variedade de substratos como os polifenois,
diaminas e alguns compostos inorgânicos. As reacções de oxidação com a lacase envolvem
a transferência de electrões de uma molécula para outra. Por outro lado, a lacase sozinha
não é capaz de oxidar compostos não fenólicos.
As reacções de catálise são feitas através da oxidação de um electrão do substrato
reduzido, com concomitante redução de quatro electrões do oxigénio a água (Leitner et al.,
2002). Embora o exacto papel da lacase na degradação da lenhina esteja por esclarecer,
tem-se provado que, na presença de mediadores, compostos não fenólicos da lenhina
podem ser degradados. O aceitador de electrões da lacase é o cobre e não a proteína. No
centro trinuclear de cobre, os eléctrons são transferidos para a água.
O mecanismo de reacção ainda não está, como referido, completamente
esclarecido. A partir de estudos espectrocópicos e de Ressonância Paramagnética
Electrónica (EPR) mostrou-se que a enzima é primeiro completamente reduzida, e então o
oxigénio é reduzido a duas moléculas de água juntamente com a formação de radicais
intermediários e que existem diferentes locais de ligação ao cobre. Os cobres T1 e T2 estão
envolvidos na captura e transferência electrónica (oxidações do substrato, por transferência
de electrão, ocorrem no T1) e o T2 e o T3 participam na ligação do oxigénio (Thurston,
1994; Call & Muck, 1997). A Figura 1.15 propõe um esquema de reacção.
O2 + 4H+ + 4e 2 H⎯⎯ →⎯Lacase2O
Figura 1.15 – Esquema de reacção da lacase com um substrato (Adaptado de Call & Muck, 1997).
31

Revisão Bibliográfica
As reacções típicas (de todas as oxidases azuis) podem ser descritas de acordo com
a seguinte reacção (1.1):
2 DH2+O2 2D+2H⎯⎯ →⎯Lacase
2O
4DH+O2→4D+2H2O (1.1)
Este tipo de reacção pertence aos fundamentos da biologia/bioquímica no
metabolismo do oxigénio (Call & Muck, 1997).
Uma reacção típica da lacase está apresentada na Figura 1.16 onde um difenol sofre
uma oxidação, com a perda de um electrão, para formar um radical livre. Esta espécie pode
ser convertida em quinona numa segunda catálise enzimática.
OH
OH OH
O
O
e- e-O .
Figura 1.16 – Uma reacção típica da lacase com um difenol (Adapado de Thurston, 1994).
Independentemente da sua origem, a lacase pode ser fortemente inibida por muitos
aniões que são capazes de interagir com os sítios de cobre como a azida, o cianeto e o
fluoreto. Os agentes complexantes removem o cobre do centro activo inibindo a actividade.
1.3.3.3 Aplicações da lacase
As aplicações práticas da lacase têm conduzido a um incremento da pesquisa nas
fontes produtoras da enzima, nomeadamente fungos da podridão branca, e no uso de
mediadores, que facilitam a acção catalítica do sistema. Basicamente, todas as aplicações
da lacase podem estar relacionadas com uma propriedade da enzima, que é a habilidade
para produzir um radical livre a partir de um substrato adequado. É conhecido durante as
32

Revisão Bibliográfica
últimas décadas que a lacase catalisa a oxidação directa de fenóis e aminas tais como os
clorofenóis e corantes, conforme descrito nos itens acima. Actualmente tem-se estudado a
degradação de compostos xenobióticos como os pesticidas e efluentes domésticos e
industriais.
A remoção da lenhina é um processo que tem desencadeado uma grande quantidade
de investigações, especialmente devido à sua importância na indústria da pasta do papel.
Para se aumentar a produção de etanol combustível, lacase obtida pelo T. versicolor
foi expressa em Saccharomyces cerevisiae de modo a aumentar a sua resistência à
inibidores fenólicos nos hidrolisados lenhinocelulósicos (Larsson et al. 2001).
Em análise de fármacos, um novo método baseado na lacase foi desenvolvido para
distinguir morfina de codeína em amostras de medicamentos (Bauer et al., 1999).
Lacase imobilizada tem sido utilizada com sucesso na remoção de fenóis da uva
branca para a clarificação de vinhos (Servili et al. 2000).
Em bioremediação, as enzimas de fungos tem-se mostrado muito eficazes na
degradação de uma variedade de poluentes ambientais. A lacase tem aumentado a
eficiência de processos de bioremediação (Gelo et al., 1999; Pointing, 2001) para a
degradação de triclorofenol (Leontievsky, 2000), alcenos (Niku & Viikari, 2000), efluentes
industriais (Fukuda et al., 2001), descoloração de pigmentos (Yesilada et al., 2003;
Zamora et al., 2003, Amaral et al., 2004; Blanquez et al., 2004; Couto et al., 2004a) e
degradação de herbicidas (Mougin et al.,2001).
1.3.3.4 Produção da lacase
A produção da lacase por microorganismos é uma característica de muitos
basidiomicetos, particularmente os associados à decomposição da madeira, entre eles o T.
versicolor. São muitos os factores que afectam a sua produção. Nomeadamente:
• concentração de inóculo;
• composição do meio de cultura;
• concentração e proporções relativas de carbono e nitrogénio;
• pH, temperatura, taxa de arejamento e agitação do meio de cultura;
• presença de indutor;
• aunsência de inibidores.
33

Revisão Bibliográfica
Para que a aplicação da lacase seja realizada com sucesso, é necessário que a sua
produção seja feita em grandes quantidades. Por isso, além de se estudar os factores
ambientais da produção, muitos autores têm considerado a adição de indutores ao meio de
cultura. O indutor pode ser o substrato da própria enzima ou um composto estruturalmente
análogo. No caso da lacase os compostos aromáticos são os mais promissores. Deste modo,
pode-se aumentar a produção da lacase numa grande variedade de fungos lenhinolíticos
pela indução por compostos como 2.5-xilidina, álcool veratrílico, ácido felúrico, guaiacol,
etanol, cobre e produtos secundários da indústria do papel como a xilidina e
lenhinosulfonados, entre outros.
O composto 2.5-xilidina tem sido o mais estudado e mais comum indutor da
produção de lacase. Quando comparado com álcool veratrílico em fermentação semi-sólida
por T. versicolor, xilidina mostrou ser um melhor indutor (Couto et al., 2002). Um outro
estudo desenvolvido com T. versicolor em fermentador “air lift” comparou a adição de
xilidina, etanol e álcool veratrílico na produção da lacase e mostrou que todos os indutores
aumentaram a produção enzimática, entretanto a xilidina apresentou melhores resultados
de actividade enzimática (cerca de 1500U/L), com um aumento de 14 vezes em relação ao
controlo (nenhum indutor adicionado ao meio) (Rancaño et al., 2003). A indução por
etanol e álcool veratrílico apresentou um menor aumento na actividade da lacase, não
passando de 400 U/L. Bollag & Leonowicz (1984) também verificaram que a xilidina
estimulou a produção de lacase em Fomes annosus, Pholiota mutabilis, Pleurotus
ostreatus e T. versicolor. Com o fungo T. modesta, a adição de xilidina, entre muitos
indutores testados, foi a que conduziu a maiores actividades enzimáticas (Nyanhongo et
al., 2002). A adição de ABTS juntamente com xilidina na produção de lacase por Trametes
sp. mostrou ser um indutor mais eficiente, com um aumento na actividade de 3.59 vezes
em relação ao controlo, enquanto que a xilidina sozinha apresentou um aumento de 2.84
vezes (Jang et al., 2002). A indução da produção da lacase por Pycnoporus cinnabarinus
com lenhinosulfonatos e álcool veratrílico aumentou a actividade em cerca de 2 a 3 vezes
enquanto que o guaiacol não apresentou nenhuma indução. Obteve-se um aumento de 9
vezes na actividade da lacase quando se empregou xilidina como indutor e não se observou
nenhuma alteração na estrutura da lacase (Eggert et at. 1996).
A introdução de álcool veratrílico em diferentes meios de cultura aumentou a
produção da lacase desde 1.6 até 16 vezes em diferentes fungos utilizados. Similarmente a
34

Revisão Bibliográfica
adição de guaiacol aumentou a produção enzimática até 38 vezes. Entretanto, a maior
produção de lacase foi obtida com a adição de “preparados de lenhinha” (indulina AT,
polifom, reax e lenhinosulfonato) com um aumento na produção até 100 vezes com o
fungo Phlebia floridensis (Arora & Gill, 2000). A adição de pirogalol ao quarto dia da
fermentação com T. versicolor aumentou em 3 vezes a produção da lacase (Osiadacz et al.,
1999). A adição de álcool veratrílico aumentou 10 vezes a produção da lacase pelo fungo
do género Trametes (Mansur et al., 1997)
Como a lacase é uma enzima que contém cobre, estudos têm mostrado que a sua
adição ao meio de cultura, aumenta a produção da enzima. Giardina et al. 1999,
observaram que a adição de sulfato de cobre ao meio de cultura de P. ostreatus aumentou a
actividade da enzima e que este aumento foi proporcional à quantidade de cobre
adicionado, atingindo um valor máximo com 150 µM de sulfato de cobre. Outro estudo,
também com P. ostreatus, mostrou um aumento de 8 vezes na actividade da lacase com a
adição de cobre (1mM) ao meio de cultura (Baldrian & Gabriel, 2002). Com o fungo
Trametes, o cobre induziu produção da lacase nas seguintes espécies: T. versicolor (Collins
& Dobson, 1997), T. pubescens (Galhaup & Haltrich, 2001; Galhaup et al. 2002a), T.
modesta (Nyanhongo et al., 2002) e T. hirsuta (Couto et al., 2004b).
Como todos os fungos, os fungos filamentosos são heterotróficos. Isto significa que
eles necessitam de compostos orgânicos como fonte de carbono e de energia, embora
alguns casos indiquem que alguns fungos filamentosos possam fixar dióxido de carbono
(Papagianne, 2004). Deste modo, a composição do meio de cultura também é um factor
importante na produção da lacase. Jang et al. (2002) estudaram o efeito das seguintes
fontes de carbono: glucose, glicerol, xilose, dextrana e verificaram que a glucose foi a
melhor fonte de carbono para a produção de lacase por Trametes sp. Nyanhongo et al.
(2002) avaliaram diferentes concentrações de glucose na produção da lacase por T.
modesta e observaram que a concentração ideal foi de 0.87 g/L e que concentrações acima
de 1 g/L inibem a sua produção.
Outro importante factor na produção da enzima é o pH do meio de cultura. Ele
pode afectar a actividade e estabilidade da enzima (O’Callaghan et al. 2002). A maioria
dos fungos tolera o pH num intervalo alargado de 4-9, enquanto que a enzima é mais
sensível as variações do pH (Papagianne, 2004). A maior parte das enzimas extracelulares
são produzidas em maiores quantidades a pH próximo do pH ótimo da sua actividade.
35

Revisão Bibliográfica
Jonsson et al. (1997) mostrou que o meio de crescimento tamponado em pH 6.0 foi
necessário para se obter actividade de lacase em culturas líquidas durante o cultivo de
Pichia pastoris e que a manutenção do pH foi extremamente importante para a produção
de altas concentrações de lacase. Outro estudo mostrou que o pH inicial do meio cultura de
Trametes sp. afectou a produção da lacase. Dos diferentes valores de pH testados (de 3 a
9), o pH de 4.5 foi encontrado como o óptimo (Jonsson et al. 1997) enquanto que para T.
modesta o pH ideal foi de 6.9 (Nyanhongo et al. 2002).
Em fermentações submersas, a agitação é importante para se obter uma boa mistura
do meio evitando limitações na transferência de massa e de calor. Em processos aeróbicos,
a mistura é necessária para se garantir uma suficiente transferência de oxigénio.
1.3.4 Produção industrial de enzimas
A produção industrial de enzimas faz-se principalmente pelo processo de
fermentação submersa (em meio líquido), que envolve o crescimento de microorganismos
num meio rico e com elevadas concentrações de oxigénio. Actualmente este é o método
mais divulgado. Devido ao desenvolvimento de tecnologias de fermentação em grande
escala, a produção de enzimas microbianas representa uma parcela significativa nas
indústrias de biotecnologia. As fermentações em meio líquido podem ser divididas em
diversas fases nomeadamente: preparação de inóculo, formulação do meio de cultura e
condições de fermentação. Os processos mais utilizados para a produção são o processo de
fermentação em contínuo e o processo em “feed batch”. Parâmetros operacionais como
temperatura, pH, taxa de alimentação, consumo de oxigénio e formação de dioxido de
carbono são medidos e controlados para optimizar o processo de fermentação. O primeiro
passo para a remoção da enzima do meio é a remoção das células, que normalmente é feito
por centrifugação ou microfiltração. A maioria da enzimas industriais são extracelulares e
permanecem no meio fermentado após a remoção da biomassa. A enzima remanescente é
concentrada por evaporação, filtração por membrana ou cristalização, dependendo da sua
aplicação, e comercializada.
Por outro lado, as enzimas também podem ser produzidas por fermentações em
estado sólido. As fermentações de substratos sólidos ocorrem em materiais insolúveis em
água. O mais tradicional processo para a produção de enzimas é o Koji, que foi
desenvolvido a partir da arte oriental de fermentação de grãos de cereais e de soja, por
36

Revisão Bibliográfica
fungos. Apresenta como vantagens a simplicidade, a redução nos custos energéticos e no
volume do fermentador, facilidade em arejamento e a pequena quantidade de água usada
permite a obtenção de metabólitos de forma concentrada, tornando a recuperação dos
mesmos mais rápida e económica. Entretanto, apresenta os seguintes incovenientes:
lentidão, dificuldade em dissipar energia, em controlar a homogeneidade e parâmetros de
funcionamento e de operação contínua (Pereira, 1996). Estudos mais recentes (Schutyser et
al., 2003) mostram a utilização de modelos matemáticos para uma melhor tranferência de
calor e melhor distribuição de água por pulverização está a ser desenvolvido.
1.4 Polissacarídeos produzidos por fungos
Na medicina tradicional do Este Asiático têm-se usado diversos fungos com
propriedades medicinais. Modernas práticas clínicas, desenvolvidas nos países asiáticos,
aplicam preparados a partir de fungos. Os efeitos terapêuticos têm sido demonstrado para
muitos fungos tais como: Favolus alveolarius, Phellinus linteus, Agaricus campestris,
Pestalotiopsis sp., Lentinus edodes, Pleurotus ostreatus, Tricholoma sp., T. versicolor
entre outros (Cui & Chisti, 2003). Das terapêuticas com base nos fungos, os
polissacarídeos obtidos a partir do fungo T. versicolor já se encontram comercializados.
Ambos os polissacarídeos, intracelular e extracelular, de T. versicolor são fisiologicamente
activos (Cui & Chisti, 2003). Nas práticas medicinais da China e do Japão, o fungo T.
versicolor é colhido, seco, triturado e utilizado como chá. As propriedades de cura do
extracto de T. versicolor foram noticiadas por cientistas Japoneses e Chineses e deste modo
iniciou-se um extensivo controlo nas investigações clínicas do extracto de T. versicolor.
Os compostos primários activos produzidos por T. versicolor são polissacarídeos
ligados a proteínas, também chamados de glicoproteínas, os quais são constituídos por
glucanas β-(1→4) com quantidades menores de glucanas β-(1→3) e β-(1→6). Essas
glicoproteínas são de difícil extracção, de onde a maioria dos produtos comercialmente
disponíveis no mercado contém apenas cerca de 1-2% destes polissacarídeos com
excepção do Coriolus VPS (JHS Natural Products) que contém cerca de 34%.
37

Revisão Bibliográfica
1.4.1 Polissacarídeos
Os polissacarídeos são macromoléculas complexas de hidratos de carbono,
formadas por milhares de unidades monossacarídicas ligadas entre si por ligações
glicosídicas, unidas em longas cadeias lineares ou ramificadas (polímeros de açúcares).
Estas ligações são formadas pela eliminação de moléculas de água entre o grupo hidroxilo
hemiacetil de um resíduo e um grupo hidroxilo primário ou secundário do resíduo
adjacente (Sutherland, 1985). Os polissacarídeos possuem duas funções biológicas
principais: como constituíntes de reserva e como elementos estruturais. Eles diferem entre
si nas unidades de monossacarídeos que os constituem, no comprimento da cadeia e no
grau de ramificação. São classificados como homopolissacarídeos, quando constituído por
um só tipo de unidade monomérica, e de heteropolissacarídeo, quando são constituídos por
duas ou mais unidades monoméricas diferentes.
Os polissacarídeos são caracterizados em termos de composição (tipo e quantidade
dos monómeros), estrutura primária (disposição dos monómeros e tipo de ligações química
entre eles), média da massa molecular relativa (Mr) e tipo e disposição dos substituintes.
Estes parâmetros determinam as propriedades funcionais dos polissacarídeos tais como
solubilidade em água, viscosidade e comportamento Newtoniano (Dumitriu, 1998).
A unidade monossacarídica predominante na estrutura dos polissacarídeos é a D-
glucose, seguido por D-manose, D-frutose, D e L-galactose, D-xilose, D-arabinose, D-
glucosamina, D-galactosamina, D-ácido glucurónico, N-acetil ácido murámico e N-acetil
ácido neuraminico (Dumitriu, 1998).
Os monossacarídicos podem formar uma variedade de estruturas lineares ou
ramificadas, conterem um número pequeno ou grande de resíduos monossacarídicos e
serem homogéneos ou heterogéneos. A Figura 1.17 mostra alguns desses monómeros e a
Figura 1.18 mostra a unidade do monómero de α-D-glucose e β-D-gucose.
O grupo dos glucanos, polímeros formados por moléculas de glucose, é o mais
abundante dentro dos polissacarídeos, pois inclui a celulose, o amido, o glicogénio e
muitas outras macromoléculas produzidas pelos seres vivos. A unidade básica do β-
glucano é a uma estrutura repetida de glucose ligada por ligações beta (β) formando uma
cadeia linear. As ligações vão desde carbono 1 da molécula de glucose ao carbono 3 da
próxima (β-(1→3)), do carbono 1 ao carbono 4 (β-(1→4)) ou do carbono 1 ao carbono 6
(β-(1→6)). A maioria apresenta na cadeia principal as ligações β-(1→3), β-(1→4) ou
38

Revisão Bibliográfica
mistura β-(1→3), β-(1→4) com β-(1→6) nas cadeias laterais. A estrutura básica repetitiva
de um β-glucano com cadeias laterais β-(1→6) está mostrada na Figura 1.19.
Nos polissacarídeos de fungos, os β-glucanos mais importantes são os β-(1→3)-
glucanos com cadeias laterais β-(1→6). Além destes, muitos outros tipos de
polissacarídeos podem ser sintetizados por fungos, tais como outras estruturas de β-
glucanos, α−glucanos, mananos, galactanos, fosfogalactanos e vários heteropolímeros com
manose ou galactose na cadeira principal (Karnezis et al., 2000).
Figura 1.17 – Representação de alguns monómeros de polissacarídeos
Figura 1.18- Representação dos monómeros de α e de β D- glucose
Figura 1.19 – Diagrama molecular de um D-glucano de origem fúngica. Representação das ligações glicosídicas (1→4) com ramificações (1→6).
39

Revisão Bibliográfica
Os polissacarídeos podem apresentar ligações a vários outros componentes
químicos, como lipídeos, formando os lipopolissacarídeos, ou proteínas, formando as
glicoproteínas (associação covalente de glucanos a proteínas, sendo a cadeia principal da
molécula constituída de proteína) e proteoglicanos (cadeia principal constituída de
glucanos e ligações com proteínas). Os tipos de ligações entre os monómeros, a variação
de composição, tipo e quantidade de ramificações, influenciam na estrutura secundária e
terciária dos polissacarídeos determinando suas propriedades físicas e, consequentemente,
suas actividades biológicas (Daba & Ezeronye, 2003).
Devido à capacidade de certos polissacarídeos em formar géis, eles são utilizados
em diversos sectores industriais, como o alimentar, farmacêutico, cosmético, químico,
têxtil e petrolífero, na função de emulsificantes, texturizantes, estabilizantes, espessantes,
controladores da formação de cristais de gelo, agentes gelificantes e coagulantes,
lubrificantes, entre outros (Sutherland, 1990).
Nas últimas décadas, vários polissacarídeos têm apresentado efeitos biológicos
como propriedades anti-inflamatórias, imunoestimulante, antitrombótica, antidiabética e
como protector às infecções. O primeiro polissacarídeo apresentado como tendo actividade
biológica foi o que continha β-(1→3)-glucanos. O primeiro teste foi feito em 1969 onde a
utilização deste polissacarídeo em tumores transplantados fez com que o tumor parasse de
crescer (Paulsen, 2002).
Os polissacarídeos de origem fúngica podem ser extraídos do corpo frutífero, do
micélio e do caldo fermentado por fungos, podendo variar a composição química,
estrutural e a actividade antitumoral (Mizuno et al., 1995). A maioria dos polissacarídeos
produzidos por basidiomicetos e com actividade antitumoral são o β-(1→3)-D-glucanos,
também chamados de “pleuran” (Nosál’ová et al., 2001).
A acção antitumoral dos β-(1→3)-D-glucanos parece estar relacionada com a
conformação de tripla hélice da cadeia (a estrutura em hélice é mantida pelas ligações de
hidrogénio, Figura 1.20), com a razão entre o número de ramificações e com o
comprimento da cadeia. Por esta razão nem sempre os β-glucanos presentes em fungos
apresentam actividade antitumoral (Paulsen, 2002). Os β-glucanos apresentam diferentes
modos de acção quando comparados com as terapias convencionais, inibindo o
crescimento das células cancerígenas e reforçando o sistema imunitário. As diferenças na
actividade podem também estar relacionadas com a habilidade da molécula do
40

Revisão Bibliográfica
polissacarídeo em se solubilizar na água, tamanho da molécula e forma das ramificações.
No etanto, alguns β-glucanos insolúveis em água, solúveis em soluções alcalinas diluídas,
apresentam uma importante actividade antitumoral (Bohn & BeMillar, 1995).
Unidades de glucose
Ligações glicosídicas
Ligações de H
Figura 1.20 - Conformação de tripla hélice da cadeia de β-(1→3)-D-glucanos.
As proteínas ligadas aos glucanos apresentam um maior potencial imunológico do
que os correspondentes glucanos (Daba & Ezeronye, 2003)
De acordo com Gutiérrez et al. (1996), existe uma correlação entre os
polissacarídeos componentes da parede celular, ou seja, os extraídos do corpo frutífero,
com os polissacarídeos extracelulares produzidos por fungos. Verificou-se que os
polissacarídeos extraídos do corpo frutífero apresentaram os mesmos tipos de monómeros
e de ligações que os dos polissacarídeos extracelulares obtidos em cultura submersa,
havendo, no entanto, alguma variação na proporção dos tipos de ligações presentes nos
biopolímeros.
Vários efeitos fisiológicos têm sido descritos com o uso dos polissacarídeos de T.
versicolor. Alguns dos principais efeitos são: efeitos imunosupressivos da quimioterapia,
radioterapia e transfusão de sangue; antagonização da imunosupressão induzida por
tumores; inibição da proliferação de vários tipos de cancro; aumento do apetite e de
funções vitais; acalmia do sistema nervoso central entre outras. Em adição esses
polissacarídeos podem curar os dessarranjos intestinais e também beneficiar a terapia de
infecções microbianas que enfraquece a resposta imunológica.
41

Revisão Bibliográfica
1.4.2 Produção de Exopolissacarídeos
O termo exopolissacarídeo (EPS) é utilizado para designar os polissacarídeos que
se encontram exteriores à célula ou livres no meio circundante. Os monossacarídeos mais
comumente encontrados nos EPSs são D-glucose, D-galactose, D-manose, L-fucose e L-
ramnose (Rosado et al. 2003). Os EPSs microbianos oferecem várias vantagens como a
possibilidade de uma produção controlada, velocidade e rendimento elevados, maior
pureza e consistência na composição, conduzindo a produtos específicos. Eles podem ser
utilizados como emulsificantes, estabilizantes, agentes formadores de géis, adsorventes
selectivos e como aditivo no controlo reológico. Apesar do elevado custo de produção são
utilizados comercialmente na indústria alimentar, na medicina e na indústria do petróleo
(Dumitriu, 1998).
A biossíntese dos exopolissacarídeos produzido por fungos é ainda pouco
conhecida. Karnezis et al. (2000) em uma revisão sobre a síntese de β-glucanos,
verificaram que a maioria dos exopolissacarídeos isolados de fungos são β-glucanos e que
a biossíntese envolve uma série de reacções: iniciação, alongamento da cadeia e
ramificação. A etapa de alongamento é a mais estudada e é catalisada por uma glucano-
sintetase cujas propriedades são específicas para cada espécie de fungo.
Os polissacarídeos comercializados de T. versicolor são extractos de cogumelos ou
micélio cultivado em substrato sólido (Yadav & Tripathi, 1991) ou micélio produzido em
fermentações submersas (Ng, 1998; Kim et al., 2002a; Cui & Chisti, 2003). Os mais
importantes polissacarídeos que já estão aprovados clinicamente são o polissacaropeptídeo
Krestin (PSK) e o polissacaropeptídeo PSP. Ambos são obtidos a partir do micélio
cultivado em fermentações submersas. Em alguns casos, utilizou-se o micélio formado na
superfície de meio líquido estático, para extrair-se os compostos activos, no entanto este
tipo de cultura não é satisfatória, em termos de rendimento, para a sua produção comercial
(Cui & Chisti, 2003).
Normalmente utiliza-se a glucose ou a sacarose como fonte de carbono nas
fermentações submersas, entretanto estudos com outras fontes de carbono alternativas têm
sido realizados. Num estudo comparativo entre a sacarose e a maltose encontrou-se uma
diferença significativa na produção de um EPS durante o cultivo de Paecilomyces
japonica. A concentração máxima de EPS foi encontrada no meio com maltose, enquanto
42

Revisão Bibliográfica
que a máxima concentração de biomassa apresentou um comportamento inverso (Bae et al.
2001). Kim et al. (2002b) estudou o efeito de nove fontes de carbono e verificou que a
sacarose apresentou-se como sendo a melhor fonte de carbono na produção de EPS por P.
sinclairii.
As fontes de nitrogénio incluem a peptona, o extracto de levedura, a farinha de
amendoim e uréia. Os fosfatos são usados para tamponar o meio e o sulfato de magnésio é
normalmente o único nutriente inorgânico adicionado. As fermentações são normalmente
realizadas entre 24-28ºC sob condições aeróbicas. O pH na maioria das vezes não é
controlado durante o cultivo.
Em culturas submersas, as condições favorecem um tipo de morfologia, e as
condições hidrodinâmicas apresentam um papel importante. A cultura miceliar de T.
versicolor tende a ser altamente viscosa devido aos filamentos da biomassa em suspensão e
aos polímeros extracelulares dissolvidos no meio. Neste tipo de cultivo, muitos fungos
filamentosos podem crescer como micélios livres ou como pellets e a forma do
crescimento é determinada por diversos factores tais como: meio de crescimento,
concentração do inóculo e condições físicas (Sinha et al. 2001 a). A agitação do meio de
cultura apresenta uma variedade de efeitos nos fungos, que podem incluir a ruptura da
parede celular, mudanças na morfologia, variação na eficiência e taxa de crescimento e
variação na taxa de formação do produto desejado. Park et al. (2002) estudaram o efeito da
intensidade de agitação no crescimento miceliar e na produção de um exobiopolímero por
Cordyceps militaris e observaram que a maior agitação estudada (300 rpm) não favoreceu
a produção do exobiopolímero e sim a produção miceliar. Um aumento na produção foi
obtido quando uma agitação de 150 rpm foi empregue.
Durante a excreção do EPS, a mudança na reologia do cultivo é uma consequência
directa da formação do produto. Um aumento na viscosidade ocorre com o aumento da
concentração do EPS. Durante a produção do EPS, o meio de cultivo desenvolve
características não-Newtonianas e pode-se apresentar como um fluido pseudoplástico, onde
a viscosidade diminui com o aumento da tensão de corte. Estas características resultam em
mudanças no comportamento microbiano e formação do produto devido à falta de
homogeneidade em termos de mistura, oxigenação, e transferência de massa e de calor
durante a fermentação. Devido a estas condições, a qualidade dos polissacarídos poderá ser
43

Revisão Bibliográfica
heterogénea, ou seja, mudanças no tamanho da cadeia, nas ramificações e na reologia
(Dumitriu, 1998).
1.4.2.1 Processo de recuperação de exopolissacarídeos de origem microbiana
A recuperação de exopolissacarídeos de microorganismos é importante na
determinação dos custos de produção e das suas propriedades físico-químicas. Os
principais passos na obtenção de EPS produzido por fermentação são a extracção,
precipitação e purificação. O passo limitante no processo de recuperação do EPS é a
separação das células do meio fermentado devido à grande viscosidade do meio
fermentado e ao grau de associação do EPS às células (Ex. Cápsula) (Dumitriu, 1998).
A maioria dos polissacarídeos comerciais de T. versicolor usam somente o
polímero intracelular recolhido do cogumelo ou do micélio produzido em culturas
submersas. O cogumelo contém aproximadamente 59% dos polissacaropeptídeos em peso.
A biomassa miceliar contem cerca de 30% em peso seco. Entretanto, a composição deste
polissacarídeo é dependente do método de recuperação usado. A sua extracção com água
quente parece ser necessária para se remover os polímeros activos em quantidade
suficiente para uso comercial. Tipicamente, a biomassa é extraída com água quente e o
extracto obtido é concentrado por evaporação sob vácuo ou ultrafiltração. O concentrado é
então precipitado com sulfato de amónio ou álcool. Os precipitados são redissolvidos,
dializados e podem posteriormente ser purificado por métodos de cromatografia. A solução
do produto purificado é concentrada e seca num secador spray. A remoção de substâncias
de baixo peso molecular é importante uma vez que essas substâncias além de não
apresentarem actividade fisiológica, conferirem um odor desagradável ao produto final.
O processo típico de recuperação de glicoproteínas está mostrado na Figura 1.21. A
sua recuperação dá- se por agitação do micélio com água destilada a 98ºC durante cerca de
3h. O extracto é então concentrado por evaporação. A solução concentrada é saturada com
sulfato de amónio para a precipitação do polissacaropeptídeo. O precipitado é dissolvido
em água e dessalgado por diálise. Esta solução é concentrada a 5% do volume original.
Uma segunda precipitação com sulfato do amónio é realizada. O precipitado final é
dessalgado, dissolvido em água e purificado por cromatografia. Mais uma precipitação
44

Revisão Bibliográfica
com sulfato do amónio é feita e então o precipitado dessalgado é concentrado e seco num
secador spray (Cui & Chisti, 2003).
Trametes versicolor
(Micélio ou cogumelo)
Diálise Precipitação com sal (sulfato de amónio)
Pequenas moléculas
Biomassa Extracções múltiplas
(água, 80-98ºC)
Concentração (ultrafiltração, evaporação)
Precipitação com álcool (etanol)
Precipitado (polissacarídeo bruto)
Dissolução (água)
Extracções múltiplas (água, 80-98ºC)
Diálise
Cromatografia (opcional)
Concentração Secagem
(Spray-dry)
Pequenas moléculas (resíduos)
Figura 1.21 – Opções de recuperação e purificação de polissacarídeos de Trametes versicolor
(Adaptado de Cui & Chisti, 2003)
45

Revisão Bibliográfica
1.4.3 Compostos comercializados
Os compostos comerciais obtidos a partir dos polissacarídeos de T. versicolor são o
polissacaropeptídeo (glicoproteína), PSP e o polissacaropeptídeo Krestin, PSK. Ambos
polissacarídeos são obtidos a partir da extracção do micélio de T. Versicolor, são
respectivamente produtos Japonês e Chinês. Eles apresentam actividade fisiológica similar
mas estruturalmente são diferentes. PSK e PSP são produzidos a partir das linhagens CM-
101 e Cov-1 de T. versicolor, respectivamente. Estes compostos são produzidos em
fermentações descontínuas. O tempo de fermentação do PSK é de 10 dias, enquanto que a
produção do PSP é de 64h. PSK é recuperado através da extracção da biomassa com água
quente e sulfato de amónio enquanto que o PSP é recuperado por precipitação alcoólica a
partir do extracto obtido com água quente.
PSK foi comercializado pela Kureha Chemicals, Japão. Após extensivos testes
clínicos, o PSK foi aprovado para uso no Japão em 1977, e em 1985 foi considerado o
décimo nono da lista dos produtos comercializados com mais sucesso no mundo. Ele é
uma mistura de polissacarídeos ligados covalentemente a proteínas. A análise elementar do
PSK mostrou, aproximadamente, a seguinte composição: oxigénio 47.5%, carbono 40.5%,
hidrogénio 6.2% e nitrogénio 5.2%. O extracto pulverizado tipicamente contém 34–35% de
hidrato de carbono solúvel (91– 93% β-glucana), 28–35% proteína, ∼7% humidade, 6–7%
cinzas, e os resíduos são açúcares livres e amino ácidos. D-glucose é o principal
monossacarídeo presente além de fucose, galactose, manose e xilose. Apresenta ligações
glicosídicas α-1,4 e β-1,3 na porção do polissacarídeo (Cui & Chisti, 2003).
O PSP apareceu no mercado cerca de 10 anos após o PSK. PSP é usado para
estimular as funções de imunidade antes ou após aos tratamentos operatórios de cancro e
para auxiliar a imunidade durante o tratamento do cancro. D-glucose é o principal
monossacarídeo presente, no entanto também existem raminose e arabinose. A porção
polissacarídica é altamente ramificada. A Figura 1.22 mostra o produto PSP
comercializado.
46

Revisão Bibliográfica
Figura 1.22 – PSP comercializado pela JHS Natural Products.
O Coriolus VPS é um produto de T. versicolor também comercializado pela JHS
Natural Products e é utilizado na América do Norte. Quando comparado com outros
produtos, tem uma maior concentração de polissacarídeos activos. A Figura 1.23 mostra o
produto comercializado. Segundo a JHS Natural Products, o Coriolus VPS contém
compostos mais valiosos e mais difíceis de se obter, encontrados na parede e no micélio de
T. versicolor (JHS Natural Products, 2005).
Figura 1.23 - VPS comercializado pela JHS Natural Products.
47

Revisão Bibliográfica
1.5 – Planeamento de experiências e modelação matemática
1.5.1 Planeamento factorial a dois níveis
O planeamento de experiências é uma ferramenta importante para se determinar
quais os factores (variáveis) que são importantes num determinado processo, assim como
os seus limites inferior e superior e as suas interacções, obtendo-se informações relevantes
para a optimização deste processo.
No planeamento de experiências muitos factores podem ser variados
simultaneamente, de acordo com um plano experimental. Através dele, podem-se
determinar os factores que exercem maior influência no desempenho de um determinado
processo. Existem muitas vantagens do planeamento de experiências quando comparado
com outras aproximações estatísticas, como sejam:
Habilidade de detectar e estimar as interacções dos factores em estudo;
Possibilidade de reduzir o número de testes necessários para se obter informações
suficientes;
Redução no tempo e custo do processo;
Melhoria do processo.
Quando o objectivo principal é optimizar o sistema, ou seja maximizar ou minimizar
algum tipo de resposta, uma técnica conveniente a ser utilizada é a metodologia de
superfícies de resposta (MSR).
Os métodos estatísticos requerem que as observações sejam variáveis aleatórias
distribuídas independentemente, portanto as experiências devem ser realizados de modo
aleatório. Qualquer planeamento começa com uma série inicial de experiências com o
objectivo de definir os factores e os níveis importantes. Podem-se ter factores qualitativos
(tipo de equipamento, operador, catalisador, etc.) e quantitativos (temperatura,
concentração, pH do meio, etc.). Os resultados devem ser analisados e devem introduzir-se
modificações no planeamento experimental quando necessário. Antes de se iniciar as
experiências, os objectivos e os critérios devem estar bem estabelecidos, de modo a
permitir a escolha dos factores envolvidos, a faixa de variação dos factores, os níveis
escolhidos dos factores e a variável da resposta (Calado & Montgomery, 2003).
48

Revisão Bibliográfica
O planeamento factorial é uma técnica muito utilizada quando se tem dois ou mais
factores independentes. Ele permite uma combinação de todos os factores a todos os
níveis, obtendo-se uma análise de um factor, sujeito a todas combinações. Esta técnica é
importante para se medirem os efeitos, de um ou mais factores, na resposta de um
determinado processo. O usual é realizar um planeamento com dois níveis (um limite
máximo e um mínimo) no máximo de três níveis, uma vez que mais factores aumentariam
em demasiado o número de pontos experimentais, facto esse que se quer evitar quando se
deseja realizar um planeamento. O planeamento factorial é a única maneira de prever a
interacção entre os factores estudados. A primeira coisa a fazer num planeamento factorial
é determinar quais são os factores e as respostas de interesse para o sistema que se deseja
estudar. Dependendo do problema pode haver mais do que uma resposta de interesse.
Eventualmente essas repostas também podem ser qualitativas.
Um planeamento factorial requer a execução de experiências para todas as
possíveis combinações dos níveis dos factores. A representação de um planeamento
factorial em dois níveis é 2k, onde 2 significa o número de níveis e k o número de factores.
Por exemplo: um planeamento factorial com três factores é um 23 ou seja, 2×2×2 = 8
experiências. Um planeamento factorial com apenas dois factores é um 22 ou seja, 2×2 = 4
experiências.
Este tipo de planeamento é de grande utilidade na investigação preliminar, quando
se deseja saber se determinados factores têm ou não influência sobre a resposta. São
planeamentos simples de se executar e podem ser ampliados, quando se quer conhecer
melhor a relação funcional existente entre a resposta e os factores.
1.5.1.1- Cálculo dos principais efeitos e interacções
O efeito principal de um factor1 (F1), no caso de um planeamento factorial 22 (2
factores), é por definição a média dos efeitos (resposta) deste factor1 (nível superior e no
nível inferior) nos dois níveis do factor2 (F2). Sendo y1 a resposta observada no i-ésimo
ensaio, podemos escrever o efeito de F1 como:
F1 = (1/2) ((y2 - y1) + (y4 – y3)) (2.2)
onde
49

Revisão Bibliográfica
y1 = resposta de F2 (no nível superior) no nível inferior de F1 (+,-) y2 = resposta de F2 (no nível superior) no nível superior de F1 (+,+) y3 = resposta de F2 (no nível inferior) no nível inferior de F1 (-,-) y4 = resposta de F2 (no nível inferior) no nível superior de F1 (-,+) O resultado mostra que a resposta pode aumentar ou diminuir em X valores quando
F1 passa de seu nível inferior para seu nível superior.
Se não houvesse interacção, o efeito do F1 seria o mesmo para os dois níveis do F2.
Quando as respostas de F1 são diferentes nos dois níveis de F2, significa que ocorre
interacção entre os dois factores. O efeito da interacção é dado por F1 × F2 e é dado por:
F1 × F2 = (1/2) ((y4 – y3) - (y2 - y1)) (2.3)
As equações 2.2 e 2.3 mostram que tanto os efeitos principais quanto o efeito das
interacções são calculados utilizando-se todas as respostas observadas. Cada um dos
efeitos é a diferença de duas médias. Metade das observações pertence a uma das médias,
enquanto a metade restante aparece na outra média. Não há portanto informações ociosas
no planeamento de experiências. Essa é uma importante característica nos planeamentos
factoriais a dois níveis (Neto et al., 1995)
1.5.1.2 Metodologia de superfície de resposta
A metodologia de superfície de resposta é uma técnica de optimização baseada no
emprego de planeamento factorial. É utilizada quando as variáveis de resposta são
influenciadas por muitos factores independentes e o objectivo é optimizar essas respostas.
A primeira etapa no uso da metodologia de resposta é determinar a relação
matemática entre a variável de resposta e os factores independentes. A primeira etapa é
constituída pela modelação, normalmente feita ajustando-se modelos lineares ou
quadráticos a resultados obtidos a partir de planeamentos experimentais factoriais. O
deslocamento dá-se sempre ao longo do caminho de máxima inclinação de um
determinado modelo, trajectória na qual a reposta varia de forma mais pronunciada.
A relação mais simples é uma recta. Se esse polinómio de menor grau se ajustar
bem a resposta, então a função será dada por um modelo chamado de modelo de primeira
ordem ou linear:
50

Revisão Bibliográfica
Y = β0 + β1x1 + β2x2 + ... + βkxk (2.4)
onde β são as estimativas dos parâmetros do modelo e x representam os factores
codificados.
O procedimento do uso de superfícies de resposta é sequencial, isto é, quando se
está longe do ponto óptimo de resposta, o modelo de primeira ordem ajusta-se bem aos
dados. No entanto, quando se quer determinar um ponto óptimo deve-se procurar uma
estratégia operacional que leve a isso. Quando se encontra a possível região de óptimo, o
modelo de segunda ordem deve ser adoptado. Se houver uma curvatura no sistema então
um modelo de segunda ordem ou seja, quadrático deve ser utilizado, e a expressão geral
para dois factores será:
Y = β0 + β1x1 + β2x2 + β11x12 + β22x2
2 + β12x1x2 (2.5)
Na metodologia de superfícies de resposta o número de variáveis não é uma
restrição nem o número de respostas. Esta metodologia pode ser aplicada a qualquer
número de factores independentes e pode modelar simultaneamente várias respostas. Esta
característica é importante em muitas situações práticas, principalmente na indústria onde
vários critérios têm de ser satisfeitos ao mesmo tempo (Neto et al., 1995).
1.5.2 Modelação matemática
Modelos matemáticos são uma ferramenta útil para a optimização da produção e
aumento de escala de bioreactores, devido à quantidade de informações quantitativas sobre
o rendimento e produtividade da biomassa e produtos. Também desempenham um papel
importante na síntese e projecto de sistemas de controlo, além de poderem ser utilizados
em simulação do processo.
Os modelos não estruturados constituem uma classe de modelos onde não há
qualquer interesse de descrição da estrutura da população a modelar. No caso de uma
população microbiana, nada é dito acerca da composição ou qualidade da biomassa. O
crescimento é expresso unicamente como sendo o aumento da biomassa. Esse tipo de
modelos é usado na descrição de fenómenos de crescimento, em que num dado intervalo
51

Revisão Bibliográfica
de tempo, todas as propriedades extensivas aumentem com o mesmo factor (Takamatsu et
al., 1981).
Utiliza-se modelo estruturado quando a composição da população celular muda
significativamente e que esta mudança infuencia na cinética de crescimento. No caso da
biomassa, esta é compartimentada em pequenos números de componentes. Às vezes estes
componentes podem ser um componente sintético (ARN e percursores) e um componente
estrutural (ADN e proteínas) ou então um componente assimilatório e um componente
sintético (Bailey & Ollis, 1986).
Num modelo descritivo, utiliza-se um ajuste numérico de dados experimentais para
fins interpolativos. Deve ser usado somente dentro da região onde o modelo foi testado
experimentalmente. Um modelo preditivo tem como finalidade a extrapolação de dados.
Existem diferentes tipos de modelos matemáticos para microrganismos filamentosos
descritos na literatura. Normalmente, os modelos envolvem a taxa específica de
crescimento (µ) como uma função da concentração de substrato (S), produto (P) e
biomassa (X). Os modelos descrevem a conversão de substratos em produtos através das
concentrações conhecidas dos metabólitos. Em muitos processos, a formação do produto
está relacionada com o crescimento da biomassa através da equação de Luedeking–Piret ou
suas modificações.
Crescimento e formação de produtos de fungos filamentoso estão associados num
processo complexo e não completamente esclarecido. A estrutura multicelular do micélio,
a heterogeneidade morfológica e diferenças no comprimento da hifa ao longo da
fermentação fazem com que se torne difícil construir modelos matemáticos para
fermentações de fungos. Levando-se em conta estas variações no estado da biomassa,
características estruturais devem ser adicionadas nos modelos de crescimento e formação
de produto (Papagianni, 2004).
1.5.2.1 Cinética de crescimento microbiano
Vários modelos cinéticos têm sido descritos para o crescimento de fungos
filamentosos incluindo linear, exponencial, logístico e rápida-aceleração/lenta-
desaceleração no crescimento. As curvas típicas destes modelos estão mostradas na Figura
52

Revisão Bibliográfica
1.24. As equações que descrevem estes modelos estão apresentadas na Tabela 1.2 (Ikasari
& Michell, 2000).
Bio
mas
sa
fase lag Tempo
Figura 1.24 – Vários perfis cinéticos de crescimento: (A) exponencial; (B) logístico; (C) linear; (D)
rápida-aceleração/lenta-desaceleração (Adaptado de Mitchell et al., 2004)
Tabela 1.2 – Formas diferencial e integrada de várias equações de crescimentoa
Forma diferencial Forma integrada
Linear K
dtdX
= (2.6) X = Kt + X0 (2.10)
Exponencial X
dtdX µ=
(2.7) teXX µ0= (2.11)
Logística ⎟⎟⎠
⎞⎜⎜⎝
⎛−=
mXXX
dtdX 1µ
(2.8) ( )( ) t
m
m
eXXXX µ−−+
=1/1 0
(2.12)
Duas fases X
dtdX µ= , t< ta
(2.9a) teXX µ0= , t< ta (2.13a)
( )[ XLedtdX
attK −−= µ ] ,
t≥ ta
(2.9b) ( )( ⎥⎦⎤
⎢⎣⎡ −= −− tatK
A eKLXX 1exp µ ) ,
t≥ ta
(2.13b)
a X é a biomassa microbiana, t é o tempo, K é a taxa linear de crescimento, µ é a taxa específica de crescimento, X0 é a biomassa inicial, Xm é a máxima possível biomassa, ta é o tempo quando se inicia a fase de desaceleração do crescimento, L é a razão entre a taxa específica de crescimento no início da fase de desaceleração e a taxa específica de crescimento na fase exponencial e k é ia taxa específica do decaimento exponencial (Mitchell et al., 2004).
53

Revisão Bibliográfica
Quando a cinética de crescimento é directamente proporcional à concentração da
biomassa, traduz-se pela seguinte equação:
dtdX
X1
=µ (2.14)
Esta equação é válida para meios definidos, nos quais a taxa específica de
crescimento durante a fase exponencial apresenta um valor constante para um dado
microorganismo em determinadas condições de fermentação.
A tradução matemática da dependência de µ com diversos factores (substrato,
biomassa, produto, etc.) tem originado os mais diversos modelos. O proposta de Monod
(1942) para a taxa específica de crescimento é o mais conhecido e utilizado nas áreas de
biotecnologia e microbiologia. Não inclui o efeito das diferenças entre as células, nem as
alterações da composição celular. Ele representa a dependência de µ com a concentração
de um substrato limitante e tem a forma de uma equação hiperbólica:
)()(max
tSKtS
s +=
µµ (2.15)
onde µmax é a taxa específica de crescimento máxima, Ks é o parâmetro de afinidade do
microorganismo com o substrato (constante do substrato) e S a concentração do substrato.
Quanto maior a constante do substrato menor é a sua afinidade.
O trabalho de Ikasari & Mitchel (2000) desenvolveu o crescimento de um fungo
filamentoso em fermentação semi-sólida considerando duas fases de crescimento,
conforme mostrado nas equações 2.9a e 2.9b. Neste modelo cinético, a fase exponencial de
crescimento é seguida por uma fase de desaceleração no crescimento. A parte entre
parênteses da equação 2.10a representa a taxa específica de crescimento durante a fase de
desaceleração, que diminui devido a dois factores. Primeiro uma repentina desaceleração é
assumida no instante em que termina a fase exponencial para iniciar a fase de
desaceleração com o parâmetro L. Segundo, a posterior desaceleração é seguida por um
decaimento exponencial do crescimento, descrito pelo termo K.
54

Revisão Bibliográfica
1.5.2.2 Modelos para a formação do produto – A taxa específica de produção O crescimento de microorganismos é muitas vezes acompanhado pela formação de
produtos. Essa formação pode estar associada ou não ao crescimento. Os produtos podem
estar solúveis na cultura, podem emergir numa forma gasosa ou ser um componente celular
não excretado. Neste caso, há necessidade de ruptura e extracção nas células.
A formação do produto pode ocorrer de três formas diferentes:
a) associada ao crescimento: ocorre quando o produto é formado juntamente com o
crescimento celular (metabolito primário);
b) dissociada do crescimento: ocorre quando o produto só começa a formar-se no fim
da fase exponencial de crescimento (metabolito secundário);
c) cinética mista: ocorre quando o produto começa a se formar a meio da fase
exponencial de crescimento.
Segundo o modelo de Pirt (1975), o produto está associado ao metabolismo energético
como catabolito da fonte de substrato limitante. Este considera que a fonte de carbono
limitante é utilizada para a formação de produto, para a formação de biomassa e para a
manutenção celular. O modelo relaciona a taxa específica de consumo de substrato (qs)
com a taxa específica de formação de produto (qp), conforme a equação abaixo:
qp = YP/S qs = mYYY
SPSX
SP/
/
/ +µ (2.16)
O modelo também relaciona a formação de produto com o crescimento dizendo que
a formação de produto por unidade de tempo é função da taxa específica de formação deste
produto e da concentração da biomassa.
Xqdt
dPp= (2.17)
O modelo de Luedeking e Piret (2000) é um modelo muito útil pois relaciona todas
as situações reais. Ele é baseado na seguinte equação:
55

Revisão Bibliográfica
XdtdX
dtdP
βα += (2.18)
onde α é a constante associada ao crescimento e β é a constante não associada ao
crescimento e sim com a concentração de biomassa. Dividindo toda a equação pela
concentração de biomassa vem:
βαµ +=dt
dPX1 (2.19)
56

Materiais e Métodos
Capítulo 2 - Materiais e Métodos
2.1 Microorganismo e condições de manutenção
Este trabalho foi realizado com o fungo da podridão branca da madeira, Trametes
versicolor obtido pelo Instituto Nacional de Engenharia, Tecnologia e Inovação (INETI,
Portugal). Manteve-se a cultura estoque (mãe) em placas de Petri com o meio sólido Tien
and Kirk, TaK (Tien & Kirk, 1988). Armazenou-se a cultura a 4 ºC e mensalmente
transferiu-se para uma nova placa. Fez-se a transferência através de um pequeno corte
(com uma ansa previamente estéril) do meio sólido juntamente com o fungo crescido para
uma nova placa de crescimento. A composição do meio de cultura TaK está descrita na
Tabela 2.1. O fungo crescido em placa de Petri durante 7 dias é mostrado na Figura 2.1.
Utilizou-se este meio de manutenção nos ensaios de produção enzimática e de produção do
EPS.
Figura 2.1 – Trametes versicolor após 7 dias de crescimento em placa de Petri.
57

Materiais e Métodos
Tabela 2.1 – Composição do meio de cultura Tien & Kirk
Concentração (g/L) Nutriente
Meio líquido Meio sólido
Glucose 10 10
Extracto de malte 10 10
Peptona 2 2
Extracto de
levedura 2 2
Asparagina 1 1
KH2PO4 2 2
Tiamina 0.001 0.001
MgSO4 1 1
Agar - 20
2.2 Preparação do inóculo para culturas líquidas
Para a preparação do inóculo, fez-se uma cultura de T. versicolor crescida durante 7
dias em placas de Petri contendo o meio sólido Tak. Adicionaram-se 10,0 mL do meio de
cultura específico em cada placa de Petri. Com uma ansa de inoculação previamente
estéril, removeu-se o micélio crescido no meio sólido e suspendeu-se no meio líquido.
Transferiu-se a suspensão de micélios da placa para um matraz, a fim de obter-se uma
suspensão concentrada de micélios, o inóculo. Usaram-se determinações de peso seco,
descritas seguidamente, para determinar a concentração da suspensão. Filtrou-se 8,0 mL
desta suspensão de micélios em papel de filtro de fibra de vidro (GF/C, 0.45 µm), colocou-
se o papel de filtro numa lâmpada de infravermelho durante 2 h. Determinou-se a
concentração da suspensão e calculou-se o volume de inóculo necessário para se obter uma
concentração inicial de 70 mg/L de micélios. Para se garantir a homogeneidade da
suspensão, realizaram-se testes preliminares, fazendo-se várias réplicas da mesma
suspensão e os respectivos pesos secos e o erro obtido foi inferior a 15%. Usou-se este
método de inoculação em todas as experiências deste trabalho nos meios de cultura
específicos utilizados.
58

Materiais e Métodos
2.3 Fermentações com Trametes versicolor para a produção enzimática
2.3.1 Meio de produção enzimática
Para a produção da lacase em matraz e em bioreactor utilizou-se o meio definido
para Trametes (MDT) (Roy & Archibald, 1993) com algumas modificações que serão
definidas posteriormente, consoante o tipo de experiência a ser realizada. A composição do
meio MDT é apresentada na Tabela 2.2. Também se inclui na composição deste meio 1,0
mL/L da solução de elementos vestigiais apresentada na Tabela 2.2. Neste trabalho
adicionou-se Tween 80 (0.5% m/v) para estimular a excreção da enzima. Nas fermentações
sem controlo de pH, o pH do meio de cultura foi ajustado em 5.0 antes de ser autoclavado.
A glucose foi autoclavada separadamente para evitar reacção de Maillard (formação de
produtos castanhos, Crueger & Crueger, 1990). Todos os meios de cultura, soluções e
materiais foram esterilizados a 120ºC durante 20 minutos.
Tabela 2.2 – Composição do meio de cultura MDT
Composição do meio MDT Composição da solução de elementos
vestigiais do meio MDT
Nutriente Concentração
(mM) Nutriente Concentração
(µM)
Glucose 83 FeSO4.7H2O 20
Glutamina 5 CuSO4.5H2O 2
NaCl 5 ZnCl2 5
KH2PO4 5 CoCl2.6H2O 6
MgSO4.7H2O 1 NiCl2.6H2O 0.1
CaCl2 0.1 MnSO4.H2O 20
2,2-dimetilsuccinato 10 (NH4)6MO7O24.4H2O 0.5
59

Materiais e Métodos
2.3.2 Fermentações em matraz
Para optimizar a produção da lacase, fizeram-se modificações no meio de cultura
MDT. Variou-se a concentração inicial de glucose, adicionaram-se diferentes indutores e
empregou-se a estratégia da limitação por carbono. Estas modificações estão mais detalhas
nos tópicos abaixo e na Tabela 2.3. Para cada experiência, determinou-se a proteína total, o
valor de pH do meio de cultura, actividade da lacase, concentração de glucose e
concentração final de biomassa.
Realizaram-se as fermentações em matraz de 500 mL com 250 mL de meio de
cultura. Introduziram-se os frascos numa câmara de incubação a 28ºC com agitação orbital
de 180 rpm durante 12 dias.
2.3.2.1 Estudo do efeito da concentração inicial de glucose
Modificou-se a concentração inicial de glucose do meio MDT para as seguintes
concentrações: 0, 1.5, 2, 3, 5 e 9 g/L. 2.5-xilidina (30µM) foi adicionada como indutor da
lacase ao terceiro dia de fermentação.
2.3.2.2 Estudo do efeito da adição de cobre
Adicionou-se CuSO4.5H2O ao meio MDT obtendo-se concentrações de 2.5, 16.0,
75.0 e 150 µM no primeiro dia da experiência para testar o seu efeito como micronutriente
e no terceiro dia da experiência para testar o seu efeito como indutor da lacase. A
concentração de 75.0 µM como micronutriente foi seleccionada para as experiências
posteriores com o meio MDT.
2.3.2.3 Fermentações com diferentes indutores
Para se avaliar o efeito da adição de indutores na produção da lacase, 2.5-xilidina
(30 µM), uma mistura fenólica (450 mg/L) e etanol (20 e 40 g/L) foram testados no meio
MDT, para além do cobre em diferentes concentrações (já descrito em 2.3.2.2). A mistura
60

Materiais e Métodos
fenólica era composta por 150 ppm (1:1:1 mg/L) dos seguintes corantes: Procion Orange
MX-2R, Remazol Red 3B e Remazol Black GF. Três experiências foram realizadas com
estes indutores:
1. adição de xilidina ao terceiro dia;
2. adição de xilidia ao terceiro dia e de cobre (75.0 µM) ao primeiro dia;
3. adição de xilidia e mistura fenólica ao terceiro dia e de cobre (75.0 µM) ao
primeiro dia.
2.3.2.4 Fermentações com limitação por carbono
Nos estudos com limitação por carbono a glucose foi removida do meio MDT após
3 dias de cultivo. Ao terceiro dia, o meio de cultura foi filtrado esterilmente em papel de
fibra de vidro (GF/C, 0.45 µm) sob vácuo e a biomassa retida no filtro foi transferida para
um novo meio MDT sem glucose. As três experiências realizadas no item 2.3.2.3 foram
também realizadas com limitação por carbono.
2.3.2.5 Testes de estabilidade da lacase
O efeito do pH na estabilidade da lacase foi avaliado para pH 3.0 e pH 4.5. Uma
amostra do meio de cultura, ao final da fermentação, com uma actividade de lacase inicial
de 2500 U/L foi colectada. Esta amostra foi então incubada a 28 °C durante 3 dias em
tampão citrato/fosfato (0.05 mM/0.1 mM) com um pH de 3.0 e também com um pH de 4.5.
A actividade da lacase remanescente foi medida a cada dia nas condições padrões.
61

Materiais e Métodos
Tabela 2.3 – Condições de cultura para a produção de lacase por Trametes versicolor em matraz
Fonte de
carbono
Suplemento do meio Indutores
Glucose (g/L) Cobre (µM) Cobre
(µM)
Xilidina
(mM)
Mistura fenólica
(mg/L)
9.0 - - - -
- - - 30.0 -
1.5 - - 30.0 -
3.0 - - 30.0 -
5.0 - - 30.0 -
9.0 - - 30.0 -
9.0 2.5 - - -
9.0 16.0 - - -
9.0 75.0 - - -
9.0 150.0 - - -
9.0 - 2.5 - -
9.0 - 16.0 - -
9.0 - 75.0 - -
9.0 - 150.0 - -
9.0 - 75.0 30.0 -
- - 75.0 30.0 -
9.0 - 75.0 30.0 450.0
- - 75.0 30.0 450.0
62

Materiais e Métodos
2.3.3 Fermentações em Bioreactor
As fermentações foram efectuadas no bioreactor BIOLAB, B. BRAUN (capacidade
de 1L) operado em descontínuo. O bioreactor apresenta controlo de temperatura, a qual foi
mantida a 28ºC através de uma resistência de aquecimento, agitação mecânica e controlo
de pH. O arejamento fez-se através de uma alimentação contínua de ar comprimido filtrado
num filtro estéril com porosidade de 0.2 µm. Nas experiências com controlo de pH, a
fermentação foi monitorada continuamente com eléctrodo de pH, e o pH foi mantido em 5
por adição automática (através de duas bombas peristálticas) com uma solução de NaOH
1M ou com uma solução de H3PO4 1M.
Inoculou-se o fermentador com biomassa crescida durante três dias em matraz (o
crescimento da biomassa em matraz fez-se de acordo com o item 2.2). Adicionou-se 2.5-
xilidina (concentração final de 30.0 µM) juntamente com o inóculo. As experiências
decorreram durante 9 dias e o volume de trabalho foi de 1.0 L. Retiraram-se cerca de 5 mL
de amostra com seringas estéreis em cada dia e removeram-se os vestígios de células e
fragmentos celulares das amostras por filtração sob vácuo com papel de fibra de vidro ou
por centrifugação numa centrifuga Ependorff a 10000 rpm durante 10 minutos.
Determinaram-se a actividade da lacase, o pH do meio (nas experiências sem controlo de
pH), a concentração de glucose,a proteína total e no fim do ensaio a concentração da
biomassa final.
As experiências no bioreactor foram realizadas com base no planeamento
experimental que está descrito no item a seguir. Neste planeamento analisou-se o efeito da
concentração inicial de glucose, o efeito do controlo de pH do meio e o efeito da agitação
na actividade da lacase.
63

Materiais e Métodos
Figura 2.2- Bioreactor usado na produção da lacase por Trametes versicolor.
2.3.3.1 Planeamento factorial para a produção de lacase em bioreactor
Para optimizar a produção da lacase em bioreactor utilizou-se o planeamento
experimental (23) sem repetição, com dois níveis e três variáveis (Box et al., 1978)
perfazendo um total de 8 experiências. Neste planeamento experimental escolheram-se três
factores: controlo de pH do meio de cultura, concentração inicial de glucose e agitação. Os
factores estudados para avaliar a produção da lacase e seus respectivos níveis estão
apresentados na Tabela 2.4. Os sinais (-1) e (+1) representam os níveis inferior e superior
dos factores, respectivamente. A concentração inicial de glucose apresenta um limite
inferior de 0 g/L e um limite superior de 9 g/L (a concentração dos outros componentes do
meio MDT permanece constante), a agitação apresenta um limite inferior de 100 rpm e um
limite superior de 180 rpm. Para o pH este estudo foi qualitativo, considerando-se o limite
inferior o não controlo de pH (pH permanece em ± 3.0) e o limite superior o controlo de
pH em 5.0. A matriz do planeamento experimental encontra-se na Tabela 2.5. Os
resultados foram analisados estatisticamente de acordo com o planeamento pré-
estabelecido a fim de verificar-se qualitativamente o efeito dos factores e suas interacções
64

Materiais e Métodos
na produção da lacase. Os cálculos dos principais efeitos e das interacções entre os
factores, a análise estátistica e os gráficos foram realizados com o software statistica
version 5.5 (StatSoft, inc.), considerando-se os níveis de significância de 10%. A resposta
do planeamento foi a máxima actividade da lacase. Os resultados foram expressos em
gráficos de superfície de resposta, gráfico de pareto, gráfico de valor observado vs valor
previsto, assim como na tabela de análise de variância (ANOVA).
As experiências foram realizadas de forma aleatória. A melhor condição para a
produção da lacase, estimada pela MSR, foi realizada experimentalmente a fim de se
confirmar os resultados previstos.
A equação (2.1) descreve o modelo de regressão utilizado no planeamento factorial
com 3 variáveis e inclui os termos de interacção:
Ŷŷ = β0 + β1xG+β2xpH + β3xA + β12xG pH + β13xG A + β23xpH A (2.1)
onde:
Ŷŷ a resposta, i.e. a produção da lacase;
xG, xpH e xA as variáveis independentes.
e os coeficientes da regressão são:
β0 o termo de interceptação;
β1, β2 e β3o os coeficientes dos efeitos lineares;
β12, β13, β23 os coeficientes dos efeitos de interacção.
Tabela 2.4 - Factores e níveis estudados no planeamento factorial 23
Níveis Factores
(-1) (+1)
Concentração de glucose g/L (G) 0 9
Controlo de pH (pH) sem com
Agitação rpm (A) 100 180
65

Materiais e Métodos
Tabela 2.5 – Matriz do planeamento de experiências 23
Factores Originais Factores Codificados Experiência
A (rpm) pH G (g/L) A
(rpm)
pH G
(g/L) F1 180 sem 0 + - -
F2 100 com 0 - + -
F3 180 sem 9 + - +
F4 180 com 9 + + +
F5 100 sem 0 - - -
F6 180 com 0 + + -
F7 100 sem 9 - - +
F8 100 com 9 - + +
2.3.3.2 Modelação matemática da produção de lacase em bioreactor
Para fazer a modelação matemática em bioractor foi necessário realizar
experiências em matraz uma vez que não foi possível determinar a concentração de
biomassa ao longo da fermentação em bioreactor. Este fungo quando se encontra em meio
líquido apresenta-se como pellets fazendo com que a cultura não seja homogénea o que
não permite a determinação directa da concentração da biomassa. Fizeram-se as
fermentações em 12 matrazes, onde cada matraz correspondeu a 1 dia de fermentação, ou
seja, cada matraz representou a biomassa de 1 dia de crescimento do fungo. Cada dia, 1
matraz foi retirado do agitador e o meio de cultura foi filtrado em papel de fibra de vidro
sob vácuo e a biomassa retida no filtro foi medida por peso seco. De acordo com as
experiências do bioreactor foi necessário realizar 2 fermentações em matraz: a primeira
com glucose no meio e a segunda com supressão de glucose.
Os modelos da fermentação em bioreactor foram baseados nos parâmetros de
crescimento estimados a partir das fermentações em matraz. Para determinar os parâmetros
em matraz, o modelo de crescimento descrito por Mitchell (Mitchell et al., 2000) para
crescimento de fungos filamentos em fermentações semi-sólida foi utilizado. Os
66

Materiais e Métodos
parâmetros de crescimento da biomassa foram determinados de acordo com as equações
2.2 e 2.3. Este modelo considera duas fases de crescimento. A primeira, uma fase
exponencial, que é descrita pela equação de Monod (eq. 2.2a). A segunda, uma fase de
desaceleração no crescimento, que é representada pela equação de Mitchell (eq. 2.2b). O
consumo de substrato e produção da enzima são representados pelas equações 2.3 e 2.4,
respectivamente. A produção de lacase foi considerada como sendo parcialmente associada
ao crescimento, e utilizou-se o modelo de Luedeking e Piret (Luedeking & Piret, 2000)
para descrever.
( ) XSKS
dtdX
S⎟⎟⎠
⎞⎜⎜⎝
⎛+
= maxµ para t < ta (2.2a)
( )( XLeSKsS
dtdX tatk −−⎟
⎠⎞
⎜⎝⎛
+= maxµ ) para t ≥ ta (2.2b)
dtdX
YdtdS
SX /
1=− (2.3)
XdtdX
dtdP βα += (2.4)
onde
X: biomassa (g/L) S: substrato (g/L) t: tempo (dias) ta: tempo do fim da fase exponencial (dia 3), µmax taxa específica de crescimento máxima (d-1) Ks: constante de saturação (g/L) L: factor de sobrevivência K: constante de decaimento exponencial (d-1) α: constante associada ao crescimento (U/g) β: constante não associada ao crescimento (U g-1 d-1)
Os parâmetros calculados para a fermentação em matraz foram usados como
constantes nas equações descritas no modelo do bioreactor. Nestas equações acrescentou-
se o parâmetro A que representa a mudança ambiental do matraz para o bioreactor.
O modelo da produção de lacase, do crescimento de biomassa e de consumo de
substrato em bioreactor está representado pelas equações 2.5-2.8.
67

Materiais e Métodos
( ) XSKSA
dtdX
S⎟⎟⎠
⎞⎜⎜⎝
⎛+
= maxµ para t < ta (2.5)
( )( XLeSKs
SAdtdX tatk −−⎟
⎠⎞
⎜⎝⎛
+= maxµ ) para t ≥ ta (2.6)
dtdX
YdtdS
SX /
1=− (2.7)
PKeKdtdP X
21 −= (2.8)
onde,
K1: taxa de síntese da lacase (U por g cel por dia) K2:taxa de decaimento da lacase (dia-1)
As equações diferenciais foram resolvidas usando o programa MATLAB (The
Mathworks Inc.). Os parâmetros µmax, KS, YX/S, K, L, A, K1 e K2 foram estimados com o
programa MATLAB Optimization Toolbox através da minimização do quadrado dos
resíduos entre os dados experimentais e os modelados, usando a função objectiva mostrada
na equação (2.9). Os programas utilizados encontram-se no anexo 5. Utilizou-se o método
Simplex-Nelder & Mead no procedimento de optimização. Os erros experimentais foram
estimados em 10% através das replicatas realizadas no shaker. A significância estatística
dos parâmetros estimados foi determinada pelo test t-student com um intervalo de
confiança de 90%.
[ ] [ ]( ) [ ] [ ]( ) [ ] [ ](∑∑ −+−+−ne
i
np
jPPSSXXMinimize 2
modexp2
modexp2
modexp ) (2.9)
onde,
np: número de parâmetros
ne: número de experiências
subscrito exp: dados experimentais
subscrito mod: dados modelados
68

Materiais e Métodos
2.3.4 Métodos analíticos
2.3.4.1 Determinação do consumo de substrato pelo método DNS
Determinou-se a concentração de substrato (glucose) quantificando a redução do
ácido 3,5-dinitrosalicílico (DNS) por reacção com os açucares redutores, o que se detecta
por mudança de cor, ou seja, por variações da absorvância no visível (λ=540 nm) (Miller,
1959).
A 1.0 mL de amostra devidamente diluída adicionou-se 1.0 mL de reagente de
DNS. Agitou-se no vortex e aqueceu-se a mistura em banho-maria (água a ferver) durante
5.0 minutos. Após este tempo, colocou-se o tubo contendo a mistura em banho gelado.
Adicionaram-se 10.0 mL de água destilada, agitou-se o tubo e leu-se a absorvância a 540
nm.
Para a realização da curva de calibração (Anexo 1) procedeu-se do mesmo modo,
substituindo a amostra por soluções de glucose de concentração conhecida (0.10-1.00 g/L).
2.3.4.2 Determinação da actividade da Lacase
Mediu-se a actividade da lacase nos meios fermentados através da oxidação do
ABTS pela enzima tal como descrito por Ander e Messner, 1998. Adicionaram-se 100 µL
de extracto enzimático ao tampão citrato/fosfato (0,05M/0,1M, pH 4.5) com ABTS (0,4
mM), que estavam previamente termostatizados a 40ºC durante 15 min., perfazendo um
volume final de 2.0 mL.
A actividade da lacase é determinada através da taxa de formação do produto da
oxidação do ABTS pela enzima em espectrofotómetro (Jenway 6405 UV/VIS) a 420nm. O
radical catiónico do ABTS possui um coeficiente de extinção molar (ε) a 420 nm de 36 000
M-1cm-1. Uma unidade de lacase corresponde à quantidade de enzima existente que oxida
1µmol de substrato por minuto. Para a conversão de abs/s em U/L efectua-se o seguinte
cálculo através da equação 2.10:
69

Materiais e Métodos
ε10f60Abs∆
LU 6
dil. ×××= (2.10)
onde,
∆ Abs: valor medido no espectrofotómetro 60: conversão de segundo para minuto fdil: factor de diluição da amostra 106: conversão de µL para L ε: coeficiente de extinção molar (M-1cm-1)
2.3.4.3 Determinação da actividade da Manganês peroxidase (MnP)
Mediu-se a actividade da MnP por 2 métodos. No primeiro, a actividade foi
determinada seguindo a variação da densidade óptica resultante da formação do complexo
malonato-Mn3+ a 270 nm, com 50 mM de tampão malonato de sódio pH 5.2, 0.2 mM de
MnSO4 e 0.1 mM de H2O2, com um coeficiente de extição molar de 11.59 mM-1cm-1
(Addleman et al., 1993). No segundo, a actividade foi determinada medindo a oxidação do
vermelho de fenol por variação da densidade óptica a 431 nm. A mistura reaccional
continha 0.2mM de MnSO4, 0.1 mM de H2O2, tampão malonato de sódio pH 4.5, 50 mM e
0.067 mM de vermelho de fenol, com um coeficiente de extição molar de 22751M-1cm-1
(Roy & Archibald, 1993).
2.3.4.4 Determinação da actividade celulásica total
Este método quantifica os açúcares redutores libertados por hidrólise enzimática da
celulose (Mandels et al. 1976).
Num tubo de ensaio colocou-se 1.0 mL de tampão citrato 0.05M pH4.8 e uma tira
de papel equivalente nº1. Adicionou-se 0.5 mL da amostra de extracto enzimático.
Incubaram-se os tubos de ensaio durante 1h a 50 ºC. Em seguida, adicionou-se 1.0 mL do
reagente de DNS. Colocaram-se os tubos num banho de água a ferver por 5 min. e em
seguida adicionaram-se 10.0 mL de água destilada. Mediu-se a absorvância a 540 nm. Uma
70

Materiais e Métodos
unidade de actividade enzimática corresponde à quantidade de enzima que liberta 1 mmol
de açúcar redutor por minuto.
2.3.4.5 Determinação da proteína total pelo método de Folin-Lowry
Este método baseia-se na reacção do “biureto” das proteínas com o cobre, em
condições fortemente alcalinas, com a formação dum complexo entre o cobre e os
aminoácidos aromáticos das proteínas, o qual é reduzido a um heteropolímero azul, por
adição do ácido fosfomolibdicofosfotúngstico. Desta reacção resulta a coloração azul
intensa (Haris & Angal, 1989) que pode ser quantificada espectrofotometricamente a 750
nm. Este método é sensível a concentrações de proteína acima de 0.25 mg mL-1.
Reagentes stock: A - 1% m/v de sulfato de cobre (CuSO4.5H2O);
B - 2% m/v de tartarato de sódio potássio;
C - 0.2M de hidróxido de sódio;
D - 4% m/v de carbonato de sódio.
A 49.0mL do reagente C adicionam-se 49.0mL do reagente D, 1.0 mL do reagente
A e 1.0 mL do reagente B. Estes formam o reagente F, a solução cobre-alcalina,
extemporânea.
O reagente E é obtido por adição de 10.0mL do reagente Folin-Ciocalteau a
10.0mL de água. A 0.50mL de amostra, adicionaram-se 2.5 mL de reagente E deixando
reagir durante 10 minutos. De seguida, adicionaram-se 0.25mL de reagente F ficando 30
minutos a reagir. A proteína total foi determinada pela conversão da absorvância a 750 nm
contra um branco (0.5 mL de tampão da amostra em lugar da própria amostra) pela recta
de calibração respectiva (Anexo 2).
Para a realização da curva de calibração procedeu-se do mesmo modo, substituindo
a amostra por soluções de proteína (BSA) de concentração conhecida.
2.3.4.6 Determinação da biomassa
Calculou-se a concentração da biomassa por determinação do peso seco. No fim de
cada experiência filtrou-se o meio de cultura em filtros de microfibra de vidro (GF/C, 0.45
71

Materiais e Métodos
µm) sob vácuo. A biomassa retida no filtro foi seca em estufa a 104 ºC durante 72 h. Ao
fim desse período, os filtros foram arrefecidos num exsicador durante 2 a 3 horas e só
depois pesados O peso seco final da biomassa fez-se por subtracção do peso da membrana
seca ao peso da membrana com a biomassa. Esse método quantifica todas as células e é
indicado para meios com altas concentrações celulares e para fungos filamentosos.
2.3.4.7 Determinação do pH
Fez-se a determinação do pH do meio de cultura através de um medidor de pH
(Crison, micropH 2000), previamente calibrado com soluções tampão conhecidas (pH = 4
e pH = 7).
2.4 Aplicação da lacase na pasta kraft 2.4.1 Preparação da pasta kraft
As experiências de branqueamento da pasta de papel foram realizadas com pasta
kraft de Eucalyptus globulus fornecida pela indústria papeleira PORTUCEL, Cacia,
Portugal. A pasta apresentava um índice kappa de 12.4 e viscosidade intrínseca de 1180
cm3g-1.
Em todas as experiências, antes do branqueamento, a pasta kraft seca foi
impregnada em água destilada durante uma noite. Acertou-se o pH em 4.5 utilizando uma
solução de H2SO4 (0.8 M) e em seguida filtrou-se a pasta sob vácuo para posterior
utilização no branqueamento.
2.4.2 Produção da lacase
A lacase utilizada nas experiências de branqueamento da pasta foi produzida em matraz
ou em bioreactor nas condições optimizadas. Utilizou-se uma concentração de 6U de
lacase por g de pasta.
72

Materiais e Métodos
2.4.3 Mediadores utilizados
Os mediadores utilizados no branqueamento da pasta de papel encontram-se na
Tabela 2.6. Os mediadores orgânicos (Tabela 2.6) utilizados foram reagentes comerciais,
fornecidos pela Aldrich Chem. Comp. (Madrid). Os polioxometalatos foram sintetizados
em laboratório. Os métodos de síntese dos polioxometalatos estão descritos por Gamelas et
al. (2005).
Tabela 2.6 – Mediadores utilizados no branqueamento da pasta kraft.
Mediadores inorgânicos (POM) Mediadores orgânicos
[SiW11MnIII(H2O)O39]5− ou (SiW11MnIII)* ABTS
[PW11MnIII(H2O)O39]4− ou (PW11MnIII)* HBT
[SiW11VVO40]5- ou (SiW11VV)* TEMPO
[BCoIII W11(H2O)O35]6- ou (BW11CoIII)* Ácido violúrico
(PW9)2MnIIIO54 ou (PW9)2MnIII*
[SiCoIIIW11(H2O)O39]5- ou (SiW11CoIII)*
* Forma abreviada do POM
2.4.4 Branqueamento da pasta kraft em reactor
As experiências de branqueamento foram realizadas num reactor PARR modelo
4843 com 250 mL de volume total, equipado com um sistema automático de controlo de
temperatura, pressão e agitação mecânica. O reactor e o controlador podem ver-se na
Figura 2.3. A velocidade de agitação foi mantida constante. Em todas as experiências,
adicionou-se um tampão fosfato (Na2HPO4) de pH 4.5 com uma concentração final de
0.1M. A pasta kraft (7.2g à base de pasta seca), a lacase, os mediadores e a solução tampão
fosfato foram introduzidos no reactor à temperatura ambiente, com um volume final de
trabalho de 113 mL. O reactor foi pressurizado com oxigénio. O tempo requerido para
atingir a temperatura de trabalho não foi contabilizado no tempo de branqueamento. No
final da reacção, o reactor foi rapidamente arrefecido com água fria, a pasta kraft filtrada
73

Materiais e Métodos
sob vácuo e neste filtrado analisou-se a actividade da lacase. Finalmente a pasta foi lavada
com sucessivas adições de água destilada e foi novamente filtrada sob vácuo. Fez-se então
a extracção alcalina da pasta (descrição no item 2.4.5) e colocou-se ao ar livre para secar
durante 1 semana, para posterior análises.
Inicialmente fez-se a selecção dos mediadores. As experiências foram realizadas
em uma única etapa de branqueamento com temperatura de 45 ºC com lacase e os
seguintes mediadores: ABTS, HBT, ácido violúrico, TEMPO, SiW11MnIII, PW11MnIII e
SiW11VV e com a temperatura de 60ºC com lacase e os seguintes mediadores: SiW11MnIII,
PW11MnIII, SiW11VV, ácido violúrico, BW11CoIII, (PW9)2MnIII e Si W11CoIII.
As reacções de branqueamento decorreram durante 4h. A pressão de oxigénio foi
mantida em 0.3 MPa. Os testes em branco foram realizados com lacase sem a adição de
mediador.
Figura 2.3 – Reactor e controlador de temperatura e de agitação usado no branqueamento da pasta
kraft de papel.
As experiências com multi-estágios foram realizadas com os POMs SiW11MnIII,
SiW11VV e PW11MnIII. O primeiro estágio de branqueamento foi realizado com uma
temperatura de 110ºC e P0(O2) = 0.6 MPa durante 2h. Ao fim deste estágio, o reactor foi
arrefecido com água fria e despressurizado. Em seguida, iniciou-se o segundo estágio com
adição de lacase, temperatura de 45ºC e P0(O2) = 0.3 MPa durante 4h. Os estágios foram
alternados até perfazer um total de 5 estágios conforme mostra a Figura 2.4.
74

Materiais e Métodos
1º estágio 2º estágio 3º estágio 4º estágio 5º estágio
45ºC, 4h 0.3MPa lacase
110ºC 2h
0.6MPa
110ºC 2h
0.6MPa
45ºC, 4h 0.3MPa lacase
110ºC 2h
0.6MPa
Figura 2.4 – Condições do branqueamento da pasta kraft com os POMs SiW11MnIII e SiW11VV através do sistema de multi-estágios.
2.4.5 Extracção alcalina da pasta
A extracção alcalina da pasta foi realizada a 70ºC durante 1h, com uma solução de
NaOH (concentração final de 2 g/L). O volume de solução no reactor foi de 71 mL. No
final da reacção, arrefeceu-se o reactor, lavou-se a pasta com sucessivas adições de água
destilada (sob vácuo) até se obter a neutralidade no filtrado. Por fim, esfarelou-se a pasta e
colocou-se numa caixa, onde foi guardada e seca à temperatura ambiente durante 7 dias,
para posteriores análises de índice kappa e de viscosidade intrínseca.
2.4.6 Métodos analíticos
2.4.6.1 Determinação do índice kappa
Determinou-se o índice kappa da pasta através do método padrão TAPPI T 236cm-
99. Este método envolve a oxidação da pasta com o permanganato de potássio, no qual
permite determinar o grau de deslenhificação da pasta. O permanganato reage com a
lenhina da pasta e não com os polissacarídeos. O índice kappa é o volume (mL) de uma
solução de permanganato 0.1N consumido por uma grama de pasta. Os resultados são
corrigidos para 50% do consumo de permanganato adicionado.
75

Materiais e Métodos
Reagentes: Solução de permanganato de potássio (0.1 N) (KMnO4)
Solução de tiossulfato de sódio (0.2 N) (Na2S2O3)
Pesam-se 1.1 a 1.3 g da pasta seca. Adicionam-se 125 mL de água destilada à pasta
que é triturada. Adicionam-se sob agitação contínua 25 mL do KMnO e 25 mL do H SO .
Após 1
álculo do índice kappa:
Solução de iodeto de potássio (1.0 N) (KI)
Ácido sulfúrico (4.0 N) (H2SO4)
Indicador (solução de amido, 2 –3 %)
4 2 4
0 minutos sob agitação, adicionam-se 5 mL de KI, algumas gotas do amido
(indicador) e titulou-se com Na2S2O3. Determinou-se o volume gasto. O teste branco foi
feito sem a pasta.
C
( )( ) ( )( )twpK p −+××= −× 25013.0110 500093.0 (2.11)
( )( )1.0
Nabp ×−= (2.12)
Onde :
K = índice kappa
uantidade de KMnO4 efectivamente consumido no teste (mL)
(amostra) (g)
umido no teste do branco (mL)
p = q
w = peso da pasta
a = quantidade de Na2S2O3 consumido no teste (mL)
b = quantidade de Na2S2O3 cons
N = normalidade do Na2S2O3
t = temperatura (ºC).
76

Materiais e Métodos
2.4.6.2 Determinação da viscosidade intrínseca
Determinou-se a viscosidade intrínseca da pasta através do método padrão SCAN –
CM15:99. Este método é utilizado para determinar a degradação da celulose da pasta
durante o seu branqueamento ou cozimento.
Pesam-se cerca de 0.16 – 0.18 g de pasta, adicionam-se 25 mL de água destilada, 3
fios de cobre e agita-se o frasco durante 40 minutos a 600 rpm. Em seguida, adiciona-se 25
mL de solução aquosa de cupri – etilenodiamina 1M (CED). Completa-se o volume do
frasco com uma solução de CED e água destilada 1:1, tapa-se sem deixar bolhas de ar e
agita-se novamente por 30 minutos a 600 rpm. Por fim, determinou-se o tempo de
escoamento da solução de CED com a pasta no viscosímetro capilar a uma temperatura
controlada de 25º C.
Cálculo da viscosidade intrínseca
mvth ××
=η (2.13)
onde,
η: viscosidade intrínseca
h: constante do viscosímetro (s -1)
t: tempo de escoamento (s)
v: volume do frasco (mL)
m: massa de pasta (g)
2.4.6.3 Determinação do estado de oxidação dos POMs por espectroscopia no visível
Todas as medidas espectrofotométricas foram realizadas num espectrofotómetro
(Jasco V-560 UV/Vis) com célula de 10 mm a temperatura ambiente. As medidas de todas
as amostras foram determinadas imediatamente após a reacção de branqueamento.
Determinou-se o estado de oxidação dos POMs ao final das experiências com um
único estágio e temperaturas de 45 e 60ºC. Nas reacções a 110ºC não foi preciso
determinar o estado de oxidação uma vez que o POM encontra-se totalmente no estado
reduzido. A Tabela 2.7 mostra as cores dos POMs no estado reduzido e no estado oxidado.
77

Materiais e Métodos
Realizaram-se também estudos da re-oxidação dos POMs reduzidos SiW11MnII,
SiW11MnII e SiW11VIV por reacções com lacase sem pasta de papel. Num matrás, colocou-
se a solução aquosa do POM no estado reduzido e lacase, a temperatura ambiente e pressão
atmosférica sob agitação. Retiraram-se amostras a determinados intervalos de tempo e
mediu-se o estado de oxidação do POM por espectro de UV/Vis. Este mesmo
procedimento foi realizado no reactor de branqueamento da pasta a pressão de 0.3 MPa e
agitação mecânica a temperatura ambiente. Em ambos estudos, o POM apresentava uma
concentração de 3.0 mM, o tampão fosfato 0.1 M (pH=4.5) e a lacase numa concentração
de 380 U/L ou 1330 U/L.
Tabela 2.7 – Cor e comprimento de onda (λ) do estado reduzido e do estado oxidado dos POMs
Cor e λ (nm) POM
Estado reduzido Estado oxidado
SiW11Mn Amarela* Cor de rosa (495)
SiW11V Púrpura (490) amarela*
BW11Co Cor de rosa (550) Verde (680)
(PW9)2Mn Amarela* Castanho escuro (600)**
PW11Mn Amarela* Cor de rosa (490)
* Não apresenta máximo de absorvância bem definido; ** apresenta um ombro de absorvância
2.4.6.4 Análise de espectroscopia de ressonância magnética nuclear - RMN
As análises de RMN foram feitas num espectrómetro Brucker MSL 400 (105.2
MHz, 9.4 T, 298 K). Os desvios químicos são referenciados em relação ao VOCl3 líquido.
Fizeram-se as análises de RMN para os POMs SiW11MnIII e SiW11VV para verificar se
ocorreu alguma alteração molecular durante o branqueamento da pasta de papel.
78

Materiais e Métodos
2.4.6.5 Análise de voltametria cíclica
As medidas de voltametria cíclica foram feitas em um analisador electroquímico
BAS 100BW, utilizando uma célula convencional de 3 eléctrodos fornecida pela BAS Inc.
O eléctrodo de trabalho foi um eléctrodo de carbono vítreo (BAS) e os eléctrodos de
referência e auxiliar foram Ag/AgCl e Pt, respectivamente. A superfície do eléctrodo de
trabalho foi polida com alumina (0.3 µm) e lavada com água destilada antes de cada
experiência. As medidas foram feitas a temperatura ambiente, num intervalo de potencial
de +1300 mV a 0 mV. As soluções foram desgaseificadas com azoto puro durante 5
minutos antes de serem medidas e mantidas com azoto gasoso durante a voltametria. As
soluções dos POMs tinham uma concentração de 1.0×10-3 M e tampão fosfato (KH2PO4)
0.5 M (pH=4.5).
2.5 Fermentações para produção do Exopolissacarídeo
2.5.1 Meios de produção
Estudaram-se 5 meios de cultura diferentes para seleccionar o melhor meio de
cultura para a produção do exopolissacarídeo. A composição de cada meio de cultura é
apresentada na Tabela 2.8. As experiências foram realizadas em matraz es de 500 mL com
250 mL de meio de cultura, a 28ºC e 180 rpm durante 9 dias. O inóculo tinha uma
concentração inicial de 70 mg/L, conforme descrito no item 2.2. O consumo de açúcares
redutores, o crescimento da biomassa e a viscosidade do meio de cultura foram
monitorados. Retiraram-se cerca de 5 mL de amostra diariamente. Removeram-se as
células e fragmentos celulares das amostras por filtração com papel de fibra de vidro sob
vácuo. Determinou-se a viscosidade do meio e o consumo de açúcares reductores. No fim
de cada ensaio, determinaram-se a concentração da biomassa (peso seco) e a concentração
do EPS (peso seco, item 2.5.3) depois de devidamente separados por centrifugação (15000
rpm, 4ºC e 20 min.).
79

Materiais e Métodos
2.5.2 Optimização do meio de cultura
Após a selecção do meio de cultura mais favorável para a produção do EPS, fez-se
a sua optimização através de um primeiro planeamento de experiências 22 com um ponto
central perfazendo um total de 5 experiências. Fizeram-se as fermentações em shaker com
matraz de 500 mL contendo 250 mL do meio de cultura YM durante 8 dias. A temperatura
foi mantida a 28ºC e a agitação foi de 180 rpm. Neste planeamento escolheram-se as
variáveis concentração inicial de glucose e pH do meio de cultura. A glucose tinha um
nível inferior de 5 g/L, um nível superior de 15 g/L e o ponto central de 10 g/L. O pH tinha
um nível inferior de 4, um nível superior de 7 e o ponto central de 5.5, como mostra a
Tabela 2.9. A composição dos outros componentes do meio de cultura permaneceu
constantes
Tabela 2.8 - Composição dos meios de cultura usados na produção do EPS
Meios de cultura Componente (g/L)
MDT GPY YM MCM TaK
Glucose 9 40 10 20 10
Extracto de malte - - 3 - 10
Extracto de levedura - 3 3 2 2
Peptona - 10 5 2 2
KH2PO4 0.68 - - 0.46 2
K2HPO4 - - - 1 -
MgSO4.7H2O 0.25 - - - 1
Asparagina - - - 1
Tiamina 0.081 - - - 0.001
NaCl 0.28 - - - -
CaCl2 0.015 - - - -
Dimetilsuccinato 0.0013 - - - -
MDT, Meio definido para Trametes GPY, Meio de glucose, peptona e extracto de levedura (Khondkar et al., 2002) YM, Meio extracto de levedura e extracto de malte (Kim et al., 2002a) MCM, Meio completo para fungos (Kim et al., 2002a) TaK, meio Tien & Kirk
80

Materiais e Métodos
Tabela 2.9 - Factores e níveis estudados no planeamento factorial 22 com ponto central
Níveis
Factores (-1) 0 (+1)
Glucose inicial g/L (G) 5 10 15
pH do meio (pH) 4 5.5 7
A matriz do primeiro planeamento experimental encontra-se na Tabela 2.10. Os
resultados foram analisados estatisticamente de acordo com o planeamento preestabelecido
a fim de verificar qualitativamente o efeito dos factores e suas interacções na produção da
lacase. Os cálculos dos principais efeitos e das interacções entre as variáveis e os gráficos
foram realizados pelo software Statistica version 5.5 (StatSoft, inc.), considerando-se os
níveis de significância de 10%. A resposta do planeamento foi o peso seco do EPS. Os
resultados foram expressos em gráficos de superfície de resposta, assim como pela tabela
de análise de variância (ANOVA).
A equação 2.14 descreve o modelo de regressão utilizado no planeamento factorial
com 2 factores e inclui os termos de interacção:
Ŷŷ = β0 + β1xG+β2xpH + β12xG pH (2.14)
onde Ŷŷ é a resposta, i.e. o peso seco do EPS; xG, xpH são as variáveis
independentes e os coeficientes de regressão são: β0 o termo de interceptação; β1, β2 os
coeficientes dos efeitos lineares e β12, o coeficientes do efeito de interacção.
Tabela 2.10 – Matriz do primeiro planeamento de experiências 22 com ponto central
Factores originais Factores Codificados Experiência
pH G (g/L) pH G (g/L)
E1 7 5 + -
E2 4 15 - +
E3 4 5 - -
E4 7 15 + +
E5 5.5 10 0 0
81

Materiais e Métodos
No Segundo planeamento de experiências os níveis dos factores passaram a ser:
glucose com um nível inferior de 5 g/L, um nível superior de 25 g/L e um ponto central de
15 g/L e o pH com um nível inferior de 4, um nível superior de 7 e um ponto central de
5.5. Nesta experiência um outro ponto central foi adicionado com uma concentração de
glucose de 15 g/L e pH de 4.0 totalizando 6 experiências, como mostra a matriz apresentada
na Tabela 2.11. A resposta do planeamento foi o peso seco do EPS.
A equação 2.15 descreve o modelo de regressão utilizado no planeamento factorial
com 2 factores e inclui somente os efeitos lineares e quadráticos:
Ŷŷ = β0 + β1xG+β2xpH + β3xG2 + β4x2
pH (2.15)
onde Ŷŷ é a resposta, i.e. o peso seco do EPS; xG, xpH são as variáveis independentes; e os
coeficientes de regressão são: β0 o termo de interceptação; β1, β2 os coeficientes dos efeitos
lineares, β3, β4 os coeficientes dos efeirtos de segunda ordem.
Tabela 2.11 – Matriz do segundo planeamento de experiências 22 com ponto central
Factores originais Factores Codificados Experiência
pH G (g/L) pH G (g/L)
A1 5 4 - -
A2 5 7 - +
A3 15 5.5 0 0
A4 15 4 0 -
A5 25 4 + -
A6 25 7 + +
2.5.3 Extracção do EPS do meio de cultura
Para a extracção do EPS do meio de cultura, removeram-se as células por
centrifugação a 15000 rpm, 4ºC durante 20 min. Ao sobrenadante adicionou-se etanol (4
82

Materiais e Métodos
vezes), agitou-se e deixou-se durante uma noite a 4ºC. O EPS precipitado foi centrifugado
a 10000 rpm, 4ºC durante 20 minutos e o sobrenadante foi descartado. O EPS precipitado
foi sequencialmente lavado com etanol a 50, 60, 70 80 e 90%, liofilizado e o peso seco
determinado. A Figura 2.5 mostra o EPS obtido.
Figura 2.5 - EPS obtido após precipitação por etanol, lavagem e liofilização.
2.5.4 Análise reológica do meio de cultura
A viscosidade aparente dos meios de cultura foi determinada por análises em
reómetro (Rheometer AR 1000, T.A. Instruments), em diferentes tensões de corte e com
controlo de temperatura.
2.5.5 Métodos analíticos para acaracterização do EPS
2.5.5.1 Espectroscopia de Infravermelho com transformada de Fourier (FTIR)
Fez-se a análise de transformada de infravermelho num aparelho MATTSON 7000
FTIR (como transformação de função de Fourier). O espectro foi obtido a partir de 2 mg de
biopolímero em 200 mg de KBr. 64 scans foram acumulados com uma resolução de 4.0
cm-1. A espectroscopia de infravermelho utilizada foi na região do infravermelho médio,
que se estende num intervalo de 4000-400 cm-1.
2.5.5.2 Análise Termogravimétrica
A análise termogravimétrica do EPS foi realizada em um termo-analisador, TGA-
50 (Shmadzu), sob atmosfera de N2 para se investigar o comportamento térmico do
biopolímero. A taxa de aquecimento foi de 10 ºC/min e aqueceu-se até 700 ºC.
83

Materiais e Métodos
2.5.5.3 Análise dos açúcares neutros
A quantidade relativa dos monossacarídeos foi determinada por cromatografia gás-
líquido depois da hidrólise com ácido sulfúrico e derivatização a acetato de alditol
(Blakeney et al., 1983). Analisou-se as amostras derivatizadas num cromatógrafo gasoso
(Carlo Erba 6000).
2.5.5.4 Determinação do peso molecular
O peso molecular médio dos EPSs produzidos nos meios YM, TaK e MCM foi
determinado através da cromatografia de permeação em gel (GPC – gel permeation
chromatography). As análises de GPC foram realizadas em duas colunas 10µm MIXED B
300×7.5 mm protegidas por uma pré coluna Plgel 10µm (Polymer laboratories Ltd. UK)
usando um sistema PL-GPC 110 (Polymer laboratories Ltd. UK) e detector de índice de
refração. Dissolveu-se a amostra do EPS em N,N-dimetilacetamida (DMCA) contendo
0.5% (p/v) LiCl. Manteve-se as colunas e o sistema de injecção em 70ºC. Bombeou-se o
eluente (0.5% p/v LiCl em DMAC) com caudal de 0.9 mL/min. Como GPC é um método
relativo, é necessário fazer uma curva de calibração. Deste modo, calibrou-se as colunas do
GPC com o material de referência pululana. A curva de calibração se encontra no anexo 3.
2.5.5.5 Determinação dos ácidos urónicos
Determinaram-se os ácidos urónicos do EPS produzido nos meios YM, TaK e
MCM usando o método m-fenilfenol (Blumenkrantz &Asboe_Hansen, 1973).
Pesaram-se 2-3 mg do EPS que se incubaram durante 3 horas com 200µL de H2SO4
a 72% e hidrolisadas durante 1 hora com H2SO4 1M. Diluiu-se 7 vezes 0.5 mL do
hidrolisado com água destilada. Adicionaram-se 3 mL de solução de borato de sódio 12.5
mM em H2SO4 concentrado a tubos de ensaio arrefecidos em gelo. A cada tubo
adicionaram-se 500 µL de amostra e em seguida aqueceu-se em banho fervente durante 10
minutos. Arrefeceram-se em gelo os tubos e adicionou-se 100 µL de solução de m-
fenilfenol a 0.5%. Agitaram-se os tubos e deixou-se em ausência de luz durante 30
minutos. Leu-se a absorvância das amostra a 520 nm. As amostras padrão foram feitas com
84

Materiais e Métodos
concentrações conhecidas do ácido D-galacturónico. Determinou-se a quantidade de ácido
urónico presente na amostra por interpolação gráfica da curva padrão (Anexo 4).
2.5.5.6 Análise elementar do EPS
A percentagem relativa de C, N e H foi determinada através de um analisador
elementar Leco CHNS-932. Uma amostra sólida de 2 mg do EPS produzido nos meios
YM, MCM e TaK foi encapsulada e colocada no forno para se iniciar a combustão. Os
ajustes para o branco e calibração são aplicados ao sinal integrado final e as respostas são
obtidas como percentagem de peso de carbono, hidrogénio e nitrogénio.
85

Resultados e Discussão
Capítulo 3 – Resultados e discussão 3.1 – Produção de lacase em matraz 3.1.1 Efeito da concentração inicial de glucose
Para verificar o efeito da fonte de carbono na produção da lacase por T. versicolor
em fermentação descontínua, empregaram-se diferentes concentrações iniciais de glucose
no meio MDT. Adicionou-se 2.5-xilidina como indutor da lacase ao terceiro dia de cada
experiência.
A Figura 3.1 mostra o efeito da concentração inicial de glucose na produção da
lacase. Os resultados mostram que uma produção significativa de lacase se inicia a partir
do quarto dia de cultivo, com actividades variando entre 100 e 500 U/L. A sua produção
foi fortemente dependente da concentração inicial de glucose do meio de cultura. Durante
a fase inicial de produção da enzima, que ainda coincide com a fase de consumo de
glucose (para concentrações acima de 2 g/L), a produção de lacase é muito baixa (dia 3).
As maiores actividades de lacase (entre 600 e 650 U/L) foram obtidas a partir do sétimo
dia de cultivo, quando a glucose não estava presente no meio de cultura, ou seja em
condições de limitação por carbono, Figura 3.2. As menores actividades de lacase foram
detectadas para as maiores concentrações iniciais de glucose empregues (5 e 9 g/L),
mostrando que altos níveis de glucose reprimem a produção da lacase. Um aumento de
quase 2 vezes na actividade da lacase foi observado quando a concentração inicial de
glucose passou de 9 g/L para nenhuma glucose no meio. Galhaup et al. (2002b), mostrou
que quando a glucose está presente acima de certas concentrações, a síntese da lacase é
reprimida em T. pubescens. A repressão da glucose em fungos e leveduras é grandemente
conhecida e acredita-se que a glucose pode inibir ou induzir os genes de produção
enzimática (Ronne, 1995).
Os dados de proteína total do meio de cultura durante a produção da lacase em
diferentes concentrações iniciais de glucose estão apresentados na Figura 3.3.
Contrariamente aos resultados obtidos para a actividade da lacase, maiores quantidades
de proteína total foram determinadas para as maiores concentrações de glucose e além
86

Resultados e Discussão
disso, o perfil da evolução da proteína total não coincide com o perfil da produção da
lacase para uma mesma concentração de glucose. Esses resultados indicam que outras
proteínas estão a ser sintetizadas durante o cultivo do fungo e não é possível, desta
maneira, fazer uma correlação entre a concentração de lacase e de proteína total no meio
de cultura.
0
100
200
300
400
500
600
700
0 2 4 6 8 10 12 14
Tempo (dias)
Laca
se (U
/L)
0 g/L 1,5 g/L 2,0 g/L 3,0 g/L 5 g/L 9 g/L
Laca
se(U
/L)
Tempo (dias)
Figura 3.1 - Actividade de lacase produzida por Trametes versicolor em diferentes concentrações
iniciais de glucose.
0
2
4
6
8
10
0 2 4 6 8 10 12 14Tempo (dias)
Glu
cose
(g/L
)iiiii
0 g/L 1,5 g/L 2 g/L 3 g/L 5 g/L 9 g/L
Glu
cose
(g/L
)
Tempo (dias)
Figura 3.2 - Consumo de glucose do meio de cultura durante a produção de lacase por Trametes versicolor em diferentes concentrações iniciais de glucose.
A evolução do pH do meio de cultura para as diferentes concentrações de glucose
está apresentada na Figura 3.4. O perfil da evolução do pH mostra que existem dois
87

Resultados e Discussão
comportamentos distintos para as diferentes concentrações de glucose testadas. Em todos
os casos, o pH diminui de aproximadamente 5 até 3, enquanto o fungo consome a glucose
(Figura 3.2) e estabiliza assim que a glucose termina. Depois desta estabilização, inicia-se
um novo período associado à uma subida de pH, excepto no ensaio com maior
concentração de glucose (9 g/L) onde esta somente se esgota no penúltimo dia de
fermentação. Estes resultados indicam claramente uma associação entre a queda do pH
do meio de cultura e o consumo de glucose por T. versicolor, indicando uma mudança no
metabolismo do fungo quando ocorre o consumo total da fonte de carbono. Galhaup et al.
(2002b), também mostrou uma queda de pH de 5.0 até 3.6 durante a fase de crescimento
de T. pubenscens e que o pH volta a subir rapidamente quando a glucose é totalmente
consumida do meio. De acordo com vários estudos da literatura, esta queda de pH do
meio ocorre devido à síntese de vários ácidos orgânicos como o malónico, o oxálico, o
fumárico, o succínico entre outros. Estes são produzidos através do metabolismo primário
dos basidiomicetos (Roy & Archibald, 1993; Milagres et al. 2002; Humar et al., 2001;
Urzúa et al., 1998; Hofrichter et al., 1999) e o posterior aumento do pH indica a mudança
para um metabolismo secundário alternativo.
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0 2 4 6 8 10 12 14Tempo (dias)
Prot
eína
tota
l (m
g/m
L)iii
iiiii
0 g/L 1,5 g/L 2 g/L 3 g/L 5 g/L 9 g/L
Prot
eína
tota
l (m
g/m
L)
Tempo (dias)
Figura 3.3 - Evolução da proteína total do meio de cultura durante a produção de lacase por
Trametes versicolor em diferentes concentrações iniciais de glucose.
A Figura 3.5 apresenta, para cada ensaio, a máxima actividade da lacase obtida, o
respectivo valor de pH e a biomassa final para as diferentes concentrações de glucose
88
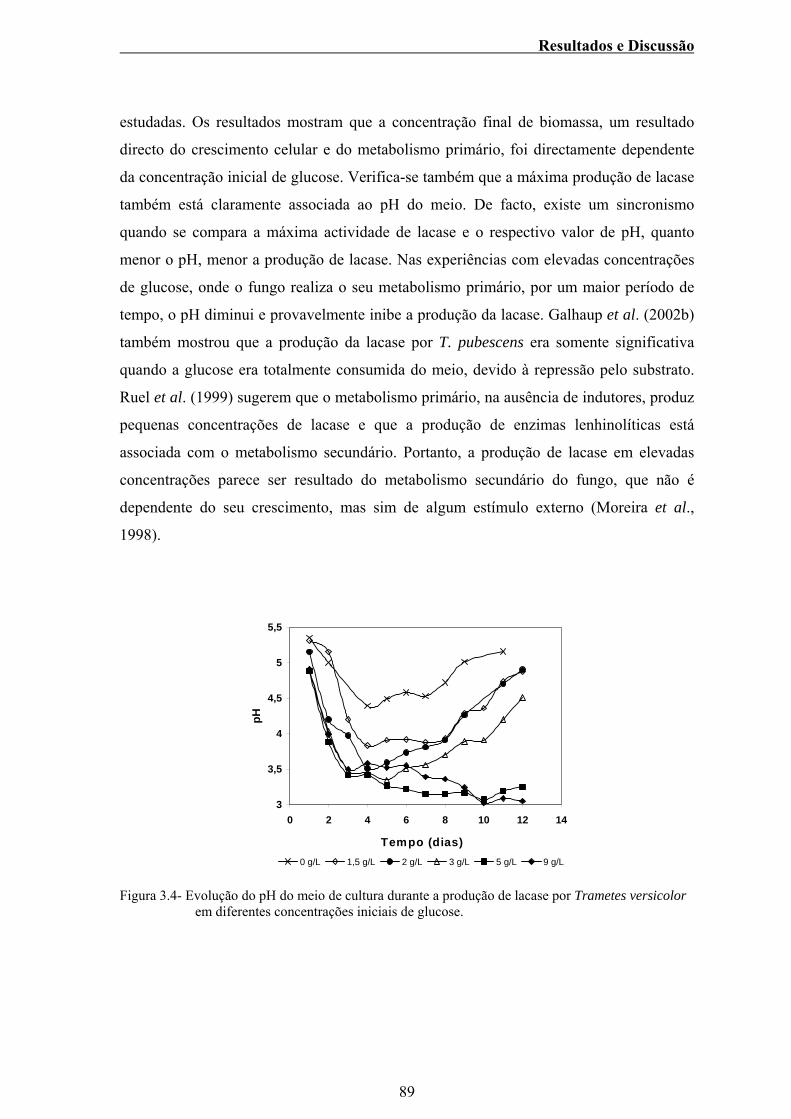
Resultados e Discussão
estudadas. Os resultados mostram que a concentração final de biomassa, um resultado
directo do crescimento celular e do metabolismo primário, foi directamente dependente
da concentração inicial de glucose. Verifica-se também que a máxima produção de lacase
também está claramente associada ao pH do meio. De facto, existe um sincronismo
quando se compara a máxima actividade de lacase e o respectivo valor de pH, quanto
menor o pH, menor a produção de lacase. Nas experiências com elevadas concentrações
de glucose, onde o fungo realiza o seu metabolismo primário, por um maior período de
tempo, o pH diminui e provavelmente inibe a produção da lacase. Galhaup et al. (2002b)
também mostrou que a produção da lacase por T. pubescens era somente significativa
quando a glucose era totalmente consumida do meio, devido à repressão pelo substrato.
Ruel et al. (1999) sugerem que o metabolismo primário, na ausência de indutores, produz
pequenas concentrações de lacase e que a produção de enzimas lenhinolíticas está
associada com o metabolismo secundário. Portanto, a produção de lacase em elevadas
concentrações parece ser resultado do metabolismo secundário do fungo, que não é
dependente do seu crescimento, mas sim de algum estímulo externo (Moreira et al.,
1998).
3
3,5
4
4,5
5
5,5
0 2 4 6 8 10 12 14
Tempo (dias)
pH
0 g/L 1,5 g/L 2 g/L 3 g/L 5 g/L 9 g/L
Tempo (dias)
Figura 3.4- Evolução do pH do meio de cultura durante a produção de lacase por Trametes versicolor em diferentes concentrações iniciais de glucose.
89

Resultados e Discussão
Figura 3.5 – Biomassa final de Trametes versicolor, máxima actividade de lacase e pH do meio
para diferentes concentrações iniciais de glucose.
0
200
400
600
800
1000
1200
1400
5, 8,
Glucose (g/L)
Bio
mas
sa (m
g L
acas
e (U
/L
0,0 1,5 2,0 3,0 0 0
)
Lacase Biomassa
3,0
3,2
3,5
3,7
3,9
4,1
4,4
4,6
pH
pH pH
Bio
mas
sa (m
g/L)
0,0 1,5 2,0 3,0 5,0 9,0
Glucose (g/L)
3.1.2 Efeito da adição de cobre
Como a lacase é uma proteína que contem cobre na sua estrutura, um aumento da
actividade da lacase em culturas suplementadas com cobre é esperado. A adição de cobre
em diferentes meios de cultura tem sido usada para aumentar a produção da lacase por
fungos (Collins & Dobson, 1997; Galhaup et al., 2002a; Giardina et al., 1999; Baldrian &
Gabriel, 2002).
Neste estudo, para se verificar o efeito da adição de cobre como micronutriente,
na produção da lacase por T. versicolor, diferentes concentrações iniciais (2.5, 16, 75 e
150 µM) de cobre foram empregues no meio de cultura MDT (Figura 3.6). Neste
conjunto de experiências, nenhum indutor foi adicionado para estimular a produção da
lacase. A concentração óptima de cobre encontrada foi de 75 µM. Esta conduziu a um
aumento de 3 vezes na produção da lacase quando comparado com o controlo, sem
adição de cobre. Para a maior concentração de cobre (150 µM), a actividade da lacase
diminuiu, provavelmente devido a um efeito tóxico do cobre no metabolismo deste
fungo.
Contrariamente aos resultados aqui obtidos, Collins & Dobson (1997) mostraram
que um aumento na concentração de cobre desde 0.4 até 400 µM promoveu um aumento
90

Resultados e Discussão
de 0.5 a 2.5 U/ml na actividade da lacase produzida por T. versicolor, respectivamente.
Um outro estudo com P. ostreatus também encontrou o mesmo resultado, onde o
aumento da concentração de cobre aumentou proporcionalmente a produção de lacase
(Giardina et al., 1999). Por outro lado, Couto et al. (2004b) estudaram o efeito da adição
de cobre na produção de lacase por T. versicolor e por T. hirsuta e verificaram que um
aumento muito pouco significativo ocorreu com T. versicolor e um aumento de 3 vezes
na actividade da lacase foi obtido com T. hirsuta. Isto pode significar que o efeito do
cobre sobre a produção de lacase é dependente do fungo e da estirpe do fungo utilizada.
O efeito da adição de cobre ao terceiro dia na produção, como indutor da lacase,
está mostrado na Figura 3.7. Para todas as concentrações de cobre estudadas, a produção
da lacase foi praticamente a mesma, apresentando uma actividade máxima de cerca de
170 U/L, que corresponde a um aumento de 105 U/L em relação ao controlo (65 U/L).
As actividades máximas obtidas pela adição de cobre como micronutriente
(adição ao meio de cultura) e como indutor (adição ao 3º dia) da lacase foram muito
semelhantes. Galhaup & Haltrich (2001) estudaram o efeito da adição de cobre em
diferentes dias de cultivo (0, 4 e 8 dias) na produção da lacase por T. pubescens e
observaram um ligeiro aumento na actividade (de 55 para 65 U/mL) quando o cobre foi
adicionado ao quarto dia de cultivo, em comparação ao primeiro dia, e uma drástica
diminuição na actividade foi observada pela adição do cobre ao oitavo dia.
Trupkin et al. (2003) também estudaram o efeito do cobre como indutor da lacase
e um aumento na produção da lacase foi mostrado com o aumento das concentrações de
cobre. O trabalho sugere que o cobre é tóxico para a maioria dos fungos, mas que ele
pode agir como indutor da produção de lacase por T. versicolor e que a produção da
lacase é um mecanismo de defesa em situações de stress oxidativo.
De acordo com os resultados obtidos da adição de cobre, seleccionou-se a adição
de cobre ao primeiro dia numa concentração de 75 µM para as posteriores experiências
de indução da produção da lacase por T. versicolor.
91

Resultados e Discussão
0
40
80
120
160
200
0 2 4 6 8 10 1
Laca
se (U
/L)ii
i
2
Tempo (dias)
2,5 uM 16 uM 75 uM 150 uM Controlo Figura 3.6 – Efeito da adição de cobre ao primeiro dia na produção da lacase por Trametes
versicolor.
0
40
80
120
160
200
0 2 4 6 8 10 12
Laca
se (U
/L)ii
i
Tempo (dias)Tempo (dias)
Tempo (dias)
Laca
se(U
/L)
Laca
se(U
/L)
2,5 uM 75 uM 150 uM Controlo Figura 3.7 - Efeito da adição de cobre ao terceiro dia na produção da lacase por Trametes
versicolor. 3.1.3 Produção de lacase com diferentes indutores
A indução e optimização da produção de lacase tem sido extensivamente estudada
com diferentes tipos de indutores e respectivas concentrações e tempo de adição (Muñoz
et al., 1997). A maior parte desses indutores são compostos aromáticos ou fenólicos,
92

Resultados e Discussão
muitas vezes relacionados com a lenhina ou seus derivados. Neste trabalho, avaliou-se
cobre, 2.5-xilidina (xil), uma mistura fenólica (MF) e etanol como indutores da lacase no
meio MDT por T. versicolor. Nestas condições estudadas não foram detectadas
actividades de MnP e de LiP.
A adição de etanol ao meio de cultura não favoreceu a produção da enzima para
as duas concentrações estudadas, 20 e 40 g/L. Embora o etanol não seja um composto
aromático, ele tem sido estudado como indutor da lacase. Em oposição aos nossos
resultados, Lee et al. (1999) mostraram que a adição de etanol (40 g/L) estimulou a
produção da lacase a níveis significativos, com um aumento de 2.6 U/mL quando
comparado com meio sem adição de etanol.
A Figura 3.8 (a) mostra o efeito dos diferentes indutores no meio MDT, contendo
9 g/L de glucose e a Figura 3.8 (b) mostra a amplificação da escala da Figura 3.8 (a) para
uma melhor visualização nas diferenças da produção da lacase com indução de cobre,
xilidina e cobre + xilidina. Os picos de actividade máxima de lacase foram obtidos em
diferentes dias, dependo do indutor ou da combinação entre eles, não havendo assim
nenhuma relação com o dia da actividade máxima e a adição de diferentes indutores.
A adição de xilidina produziu uma maior actividade de lacase quando comparada
com a adição de cobre (de 170 para 360 U/L), o que corresponde um aumento de 1.9
vezes. Muitos trabalhos mostram a indução de lacase por xilidina em fungos (Garzillo et
al., 1998; Koroljova-Skorobogat’ko et al., 1998; Couto et al., 2002). Entretanto, um
efeito cooperativo entre o cobre e a xilidina foi observado. A presença simultânea de
ambos os indutores levou a um maior aumento na produção de lacase (800 U/L) do que o
efeito individual de cada um deles (4.4 vezes em relação ao cobre e 2.3 em relação à
xilidina). O mesmo efeito de cooperação entre esses indutores foi observado na produção
de lacase por T. versicolor (Collins & Dobson, 1997) e por T. hirsuta e T. versicolor em
culturas sólida (Couto et al., 2004b)
93

Resultados e Discussão
0
1000
2000
3000
4000
5000
6000
0 2 4 6 8 10 12 14Tempo (dias)
Laca
se (
(U/L
)
Cu xil Cu+xil Cu+xil+MF Controlo
0
200
400
600
800
0 2 4 6 8 10 12 14
(b)
Laca
se(U
/L)
Tempo (dias)
(a)
Figura 3.8 – (a) Produção de lacase por Trametes versicolor na presença de cobre, xilidia (xil) e mistura fenólica (MF). (b) amplificação da Figura 3.8 (a).
Uma vez que a lacase actua como uma enzima oxidativa em compostos fenólicos,
uma complexa mistura fenólica (mistura de três corantes) foi empregada para favorecer o
aumento dos indutores previamente testados. Esta mistura fenólica foi utilizada por
Amaral et al. (2004) na descoloração de corantes têxteis e promoveu altas concentrações
de lacase por T. versicolor. Neste trabalho, obteve-se um aumento significativo na
actividade enzimática, de 850 U/L no meio com adição de cobre e xilidina, para 5500
U/L (6.5 vezes) na presença desta mistura fenólica (Figura 3.8).
Indutores com estruturas fenólicas parecem permitir a este tipo de fungo
reconhecer um sinal que conduz a uma intensiva resposta biológica que activa o
metabolismo secundário responsável pelo aumento da produção de enzimas
lenhinolíticas.
A Figura 3.9 mostra o valor de pH do meio de cultura e o consumo de glucose
durante a produção de lacase com os diferentes indutores. Com a adição de cobre, um
baixo consumo de glucose foi observado, com uma taxa de consumo de 0,48 g/d, quando
comparado com os demais indutores que apresentaram as seguintes taxas: 0.67 g/d para a
xilidina, 0.66 g/d para cobre, xilidina e mistura fenólica e de 0.93 g/d para cobre e
xilidina. Os resultados indicam que quando a xilidina está presente, o fungo consegue
assimilar mais rapidamente a glucose. Como nos resultados anteriores, o pH do meio
desce juntamente com o consumo da glucose até um valor aproximado e constante de 3.0,
94

Resultados e Discussão
excepto para o meio suplementado com cobre, onde o pH desceu e manteve-se constante
em 3.6, provavelmente devido à baixa taxa de consumo de glucose. No meio onde os três
indutores estavam presentes, o pH do meio aumentou muito rapidamente quando a
glucose foi totalmente consumida (dia 9).
2,9
3,4
3,9
4,4
4,9
5,4
5,9
0 2 4 6 8 10 12 14
Tempo (dias)
pH
Cu xil Cu + xil Cu + xil+MF
0
2
4
6
8
10
12
0 2 4 6 8 10 12 1
Tempo (dias)G
luco
se (g
/L)
4
Cu xil Cu+xil Cu+xil+MF
a b
Figura 3.9 – pH do meio de cultura (a) e consumo de glucose (b) durante a produção de lacase por Trametes versicolor com diferentes indutores.
3.1.4 Efeito da limitação por carbono com diferentes indutores
Como mostrado no item 3.1.1, a limitação por carbono (LC) estimulou a produção
da lacase por T. versicolor. Deste modo, fez-se a estratégia da limitação por carbono para
os melhores resultados da adição de indutores: cobre+xilidina (xil) e
cobre+xilidina+mistura fenólica (MF). Os resultados com e sem limitação por carbono
estão apresentados na Figura 3.10.
A actividade da enzima com a adição de cobre+xilidina aumentou cerca de 3
vezes ou 1850 U/L (de 850 para 2700 U/L) em condições de limitação por carbono. Um
efeito similar, embora menos significativo, também ocorreu quando a mistura fenólica foi
empregue (de 5520 para 6080 U/L), o que corresponde a um aumento de 560 U/L ou 1.1
vezes na actividade máxima da lacase. Além disso, observou-se ainda uma antecipação
de 2 dias na actividade máxima da lacase.
95

Resultados e Discussão
0
1
3
4
6
Laca
se (U
/L)ii
ii
500
000
500
000
7500
0 2 4 6 8 10 12 14Tempo (dias)
Cu+xil+LC Cu+xil+MF+LC Cu+xilCu+xil+MF Controlo
Laca
se(U
/L)
Figura 3.10 - Produção de lacase por Trametes versicolor na presença de cobre e xilidia (xil) e de cobre, xilidia (xil) e mistura fenólica (MF) com e sem limitação por carbono (LM).
A Tabela 3.1 mostra a produtividade e o aumento da produtividade da lacase para
os diferentes indutores com e sem limitação por carbono. O emprego da estratégia da
limitação por carbono e o emprego da mistura fenólica conduziram aos melhores
resultados da produtividade da lacase. Um aumento de 75.5 vezes em relação ao controlo
foi obtido nas condições de sem limitação por carbono e um aumento de 103.9 vezes para
combinação de todos os indutores nas condições de limitação por carbono.
Os resultados indicam que compostos fenólicos estimulam o fungo a desenvolver
mecanismos de protecção aos efeitos tóxicos, fazendo com que mais enzima seja
produzida de forma a tentar eliminar a toxicidade do meio onde se encontra e/ou pelo
facto do fungo reconhecer estruturas similares da madeira. É conhecido da literatura que
os compostos mais eficientes na indução de enzimas lenhinolíticas são os preparados de
lenhina e os compostos fenólicos tais como álcoois, aldeídos e ácidos (Muñoz et al.
1997). A indução desses compostos na produção da lacase vem da capacidade natural do
fungo T. versicolor em degradar a lenhina através de mecanismos específicos de ataque
às madeiras. A presença desse tipo de indutor faz com que o fungo os reconheça e
desenvolva um metabolismo de defesa ao stress oxidativo (Trupkin et al., 2003)
produzindo uma maior quantidade de enzima.
96

Resultados e Discussão
Tabela 3.1 - Produtividade de lacase e aumento da produtividade de lacase para os indutores cobre, xilidina e mistura fenólica em condições de limitação por carbono e sem limitação por carbono
Indutores Produtividade (U/ (L. dia)) Aumento da
produtividade
Sem limitação por carbono
Controlo 7.3 1.0
Cobre 31.8 4.4
xilidina 60.0 8.2
Cobre + xilidina 76.0 10.4
Cobre + xilidina + Mistura
fenólica
552.1 75.5
Com limitação por carbono
Cobre + xilidina 272.3 37.3
Cobre + xilidina + Mistura
fenólica
759.8 103.9
97

Resultados e Discussão
3.2 – Produção de lacase em fermentador
3.2.1 – Optimização da produção da lacase por planeamento de experiências
Após estudar-se as diferentes condições de produção da lacase por T. versicolor
em matraz, fez-se um planeamento de experiências com três factores: concentração
inicial de glucose, pH do meio de cultura e agitação, e dois níveis para aplicar em
fermentador. A finalidade é a optimização da produção enzimática, uma vez que a
produção enzimática é muito dependente das condições de cultivo. A variável de resposta
foi a máxima actividade de lacase obtida durante o processo de fermentação. O
planeamento totalizou 8 experiências com as diferentes combinações dos três factores em
estudo. Os ensaios foram efectuados em conformidade com o delineamento experimental
proposto no item 2.3.3.1. Para a glucose escolheu-se um limite inferior de 0 g/L e um
limite superior de 9 g/L, para a agitação um limite inferior de 100 rpm um limite superior
de 180 rpm e para o pH um limite inferior de 3.0 e um limite superior de 5.0, onde se
considerou o limite inferior o não controlo do pH e o limite superior o controlo do pH em
5.0 ± 0.1.
Os resultados das fermentações de T. versicolor pelo planeamento de experiências
estão resumido na Tabela 3.2. Os valores obtidos mostram que os meios com glucose (9
g/L) e sem controlo de pH (experiências F3 e F7) não favoreceram a formação da lacase,
resultando em actividades inferiores a 1000 U/L e com elevadas concentrações de
biomassa (≈ 1.5 g/L).
Obtiveram-se os melhores resultados quando glucose estava presente no meio de
cultura e o pH controlado (F4 e F7), o que correspondeu a actividades acima de 7000 U/L
para ambos valores de agitação. Quando se comparam experiências similares, com a
mesma concentração de glucose e a mesma agitação (F3 e F4; F7 e F8), onde somente se
varia o pH, obteve-se um aumento na actividade máxima da lacase de 16 vezes para uma
agitação de 100 rpm (de F3 para F4) e um aumento de 9 vezes para uma agitação de 180
rpm (de F7 para F8). Em todas as experiências a produtividade da lacase seguiu a mesma
tendência que a actividade máxima.
98

Resultados e Discussão
A biomassa final apresentou baixos valores para os meios sem glucose, de 0.3 a
0.6 g/L, o que era esperado uma vez que não havia fonte de carbono para o crescimento.
Para as experiências com glucose no meio, a biomassa final apresentou concentrações
próximas a 1.5 g/L.
Tabela 3.2 – Planeamento de experiências 23 e repostas da produção da lacase por Trametes
versicolor
Experiência Agitação
(rpm)
Controlo
de pH
Glucose
(g/L)
Biomassa
final
(g/L)
Máxima actividade de lacase
(U/L)
Produtividade de lacase (U/(L.d))
F1 180 3 0 0.37 1493 249
F2 100 5 0 0.60 2178 260
F3 180 3 9 1.51 444 37
F4 180 5 9 1.59 7113 889
F5 100 3 0 0.29 1410 117
F6 180 5 0 0.27 1781 184
F7 100 3 9 1.28 771 70
F8 100 5 9 1.54 7217 902
Os efeitos estimados de cada factor e suas interacções para o planeamento de
experiências estão descritos na Tabela 3.3. As respostas mostram que o controlo de pH,
concentração inicial de glucose e as suas interacções apresentam um grande efeito na
actividade máxima da lacase, enquanto que a agitação não apresenta nenhum efeito
significativo na produção da lacase. A significância dos efeitos estimados foi avaliada
através da análise de variância (ANOVA). Os testes ANOVA indicam que o modelo
linear descreve adequadamente o rendimento da síntese de lacase. O efeito significativo
de cada coeficiente foi determinado através do teste p (p < 0.1), considerando 90% de
intervalo de confiança. Baixos valores de p indicam um coeficiente muito significativo.
As variáveis que apresentaram um efeito significativo similar foram o pH e a interacção
Glucose × pH. O coeficiente de regressão (R2 = 0,99625) indica também que o modelo de
primeira ordem (linear) pode representar adequadamente os dados experimentais
referentes ao rendimento em lacase.
99
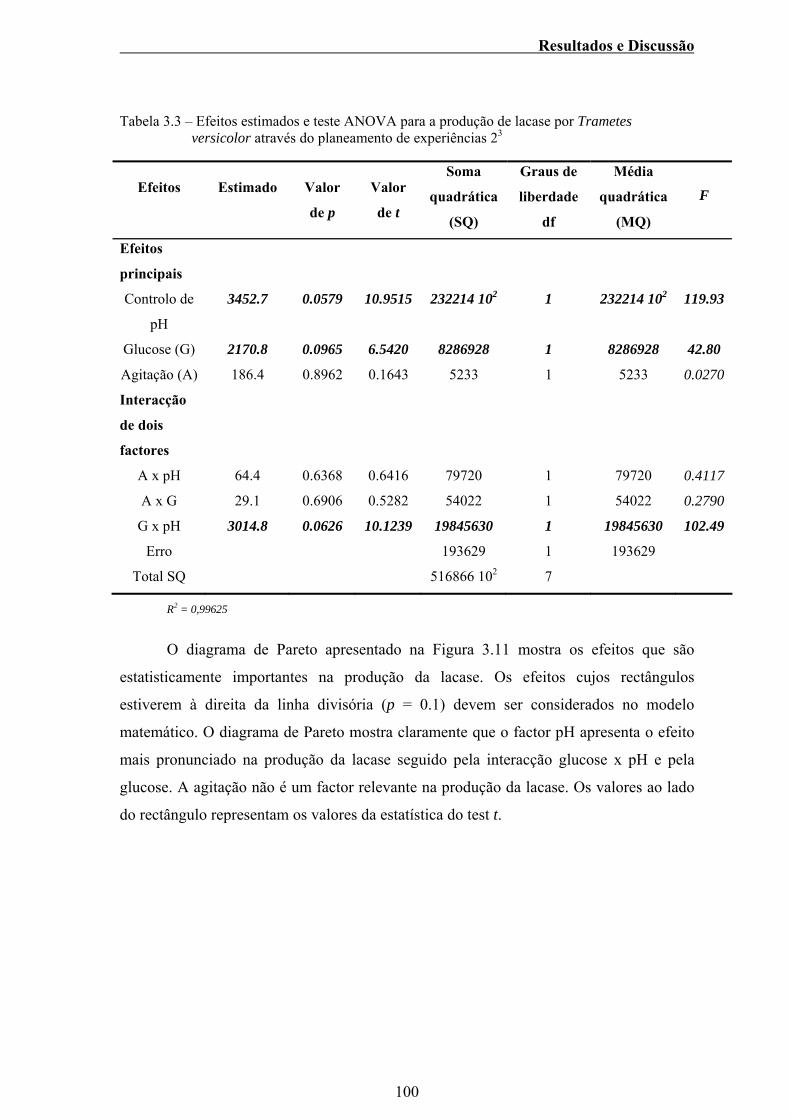
Resultados e Discussão
Tabela 3.3 – Efeitos estimados e teste ANOVA para a produção de lacase por Trametes versicolor através do planeamento de experiências 23
Efeitos Estimado Valor
de p
Valor
de t
Soma
quadrática
(SQ)
Graus de
liberdade
df
Média
quadrática
(MQ)
F
Efeitos
principais
Controlo de
pH
3452.7 0.0579 10.9515 232214 102 1 232214 102 119.93
Glucose (G) 2170.8 0.0965 6.5420 8286928 1 8286928 42.80
Agitação (A) 186.4 0.8962 0.1643 5233 1 5233 0.0270
Interacção
de dois
factores
A x pH 64.4 0.6368 0.6416 79720 1 79720 0.4117
A x G 29.1 0.6906 0.5282 54022 1 54022 0.2790
G x pH 3014.8 0.0626 10.1239 19845630 1 19845630 102.49
Erro 193629 1 193629
Total SQ 516866 102 7
R2 = 0,99625
O diagrama de Pareto apresentado na Figura 3.11 mostra os efeitos que são
estatisticamente importantes na produção da lacase. Os efeitos cujos rectângulos
estiverem à direita da linha divisória (p = 0.1) devem ser considerados no modelo
matemático. O diagrama de Pareto mostra claramente que o factor pH apresenta o efeito
mais pronunciado na produção da lacase seguido pela interacção glucose x pH e pela
glucose. A agitação não é um factor relevante na produção da lacase. Os valores ao lado
do rectângulo representam os valores da estatística do test t.
100

Resultados e Discussão
Pareto Chart of Standardized Effects; Variable: Laccase activityGráfico de Pareto; Variável: Actividade de lacase
2**(3-0) design; MS Residual=193628,6
p=,1
10,95115
Figura 3.11 – Diagrama de Pareto para o planeamento factorial 23.
Adicionalmente, a correlação entre os valores previstos no modelo para a
actividade máxima da lacase e os dados experimentais é apresentada na Figura 3.12. Os
resultados mostram que os dados estão muito próximos da linha, o que caracteriza um
bom ajuste, mostrando que o modelo linear descreve bem os dados experimentais.
Figura 3.12 - Correlação entre os valores observados e os valores previstos da produção da lacase.
Effect Estimate (Absolute Value)
,1643902
,5282018
,6416519
6,542022
10,1239
(1)pH
1by21 por 2
(2)Glucose
1by3
2by3
(3)Agitation
2 por 3
1 por 3
(3)agitação
14 -2 0 2 4 6 8 10 12
Efeito estimado (valor absoluto)
Observed vs. Predicted Values2**(3-0) design; MS Residual=193628,6
Pred
icte
d V
alue
s
Observed Values
-1000
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
-1000 0 1000 2000 3000 4000 5000 6000 7000 8000
Valo
res
prev
isto
s
Valores observados
Valores observados vs. valores previstos
101

Resultados e Discussão
Os resultados do planeamento de experiências apresentados acima podem também
ser visualizados através de uma superfície, chamada de superfície de resposta, que no
caso deste trabalho, é um planeamento factorial comum e por isso tem a forma de um
plano, visto que o modelo matemático é linear. As superfícies de resposta resultantes e a
respectiva relação matemática entre a variável de resposta (actividade da lacase; z) e as
variáveis independentes (factores) estão mostradas nas Figuras 3.12a-c. Os gráficos
mostram o efeito de duas variáveis enquanto que a terceira permanece constante. A
respectiva função que representa estas figuras é dada pela seguinte equação:
Actividade da lacase (U/L): 2931.55 - 220.69 pH - 1237.77 G - 11.40 A + 350.01
pH×G + 2.46 pH×A+0.46 G×A (3.1)
As Figuras 3.13a-c representam um gráfico tri-dimensional das seguintes
variáveis: concentração de glucose e pH (a uma agitação de 180 rpm), agitação e
concentração de glucose (a um pH de 3) e agitação e pH (a uma concentração de glucose
de 9 g/L). A cor castanha representa as elevadas actividades de lacase. O modelo de
primeira ordem (linear) ajusta bem a resposta e é mostrado nas equações apresentadas na
Figura 3.13. Na Figura 3.13a observa-se que quando se usa o nível inferior de ambos os
factores (glucose de 0 g/L e sem controlo de pH) a actividade da lacase cai atingindo o
mínimo valor observado no intervalo estudado (cerca de 400 U/L). As condições óptimas
encontradas para a produção da lacase foram uma concentração de glucose e um pH de
11 g/L e 5.2, respectivamente. Nessas condições, uma actividade de lacase de cerca de
10000 U/L é prevista pelo modelo. Após obter as condições optimizadas, realizou-se uma
nova experiência nas condições óptimas (glucose inicial de 11 g/L e pH 5.2). Nestas
condições, obteve-se a máxima actividade de lacase de 11403 U/L, que está de acordo
com o valor previsto pelo modelo, o que mostra a adequação da MSR utilizada.
As Figuras 3.13b e 3.13c mostram que a agitação não apresenta nenhum efeito
individual, nem interacção com os outros factores na produção da lacase, conforme já
discutido anteriormente.
102

Resultados e Discussão
478,437 1368,31 2258,184 3148,06 4037,93 4927,806 5817,68 6707,553 7597,427 8487,3 above
Superfície de resposta2**(3-0) design; MS Residual=193628,6
z=2931,55-220,6875*x-1237,7638*y+350,005*x*y+2,495*180*x+,45652*180*y-2051,55
464,267 624,259 784,252 944,244 1104,236 1264,23 1424,22 1584,213 1744,206 1904,2 above
Superfície de resposta2**(3-0) design; MS Residual=193628,6
z=2931,55-1237,7638*y-11,3975*x+350,0055*3,*y+2,4956*3,*x+,4565*x*y-662,0625
(rpm)
(g/L)
773,932 1440,156 2106,38 2772,604 3438,83 4105,05 4771,276 5437,5 6103,725 6769,95 above
Superfície de resposta2**(3-0) design; MS Residual=193628,6
z=2931,55-220,6875*y-11,3975*x+350,0055*8,*y+2,4956*x*y+,4565*8,*x-9902,11
(rpm)
Figura 3.13 - Superfícies de resposta para o planeamento de experiências 23.
103

Resultados e Discussão
Os resultados obtidos mostram claramente que é necessário o controlo do pH do
meio de cultura para a obtenção de elevadas actividades de lacase. Durante as
fermentações sem controlo de pH e com concentrações de glucose de 9 g/L, observou-se
que o pH diminui até um valor de 3.0, e que essa diminuição do pH ocorre
simultaneamente ao consumo de glucose (Figura 3.14), como aconteceu no cultivo em
matraz descrito anteriormente.
0
2
4
6
8
10
12
0 2 4 6 8 10 12 14
Tempo (dias)
Glu
cose
(g/L
)iiiii
3
3,4
3,8
4,2
4,6
5
pH
Glucose pH
Glu
cose
(g/L
)
Tempo (dias)
Figura 3.14 – Consumo de glucose e pH do meio durante a produção de lacase por Trametes versicolor para uma concentração inicial de glucose de 9 g/L e sem controlo de pH.
Conforme já discutido anteriormente há ácidos orgânicos que são produzidos
durante a fermentação. Os resultados mostraram que baixos valores de pH não são
favoráveis à produção de lacase por T. versicolor. Este facto pode ter duas explicações:
Pode o metabolismo da produção de lacase ser reprimido pelos baixos valores de pH; ou
mudanças conformacionais na estrutura tri-dimensional podem ser promovidas pelo
baixo pH, afectando o sítio activo da enzima e não permitindo assim a ocorrência de
reacções biocatalíticas de oxidação.
Com a finalidade de avaliar a perda de actividade da enzima a baixos pHs,
estudou-se a sua estabilidade para valores de pH de 3.0, 4.5 e 5.0 (controlo). A pH 3.0
observou-se uma perda de 50% na actividade da lacase durante o primeiro dia de
incubação, atingindo-se um valor de 80% de perda de actividade após três dias. Para o pH
de 4.5 observou-se uma perda de apenas 11% após os três dias de incubação. Para o
controlo, a pH 5.0, não foi observada qualquer redução na actividade da lacase durante os
três dias, o que mostra que pH de 5.0 é o ideal para a manutenção da actividade da lacase.
Quando se comparam os resultados para as fermentações com glucose, com e sem
104

Resultados e Discussão
controlo de pH (F3 e F4; F7 e F8), verifica-se pela actividade obtida que houve uma
redução correspondente de 90-93% na actividade da lacase para as fermentações sem
controlo de pH. Estes resultados parecem indicar que a hipótese considerada em relação à
pouca estabilidade da lacase a valores de pH baixos seja mais favorável, embora não
excluam por completo que haja alguma inibição metabólica de produção da lacase pelo
fungo nestas condições.
Estudos recentes, mostram que a estabilidade da lacase é muito dependente do pH
e da temperatura (Palonen et al., 2003; Galhaup et al. 2002a, Souza et al., 2002).
Nyanhongo et al. (2002) mostra que a actividade da lacase produzida por Trametes
modesta foi significativamente inibida a pH 3 e acima de 5, mostrando um óptimo a pH
4. Jonsson et al. (1997) observaram que pH abaixo de 4 é prejudicial para a produção da
lacase e sugere que essa perda é devido à susceptibilidade para proteases ácidas nessas
condições.
Galhaup et al. (2002b) relatam que para Trametes pubescens, quando a
concentração de glucose está presente no meio de cultura acima de um valor crítico (0.25
g/L), a síntese da lacase é reprimida e que a produção somente aumenta quando a glucose
é esgotada no meio. Neste estudo em bioreactor, quando o pH do meio não era controlado
e com glucose inicial de 9 g/L, a máxima produção da lacase foi obtida quando se
esgotou a glucose. No entanto, quando se controlou o pH, mesmo com 9 g/L de glucose
no meio cultura, obteve-se um grande aumento na actividade da lacase no meio.
Contrariamente ao mostrado por Galhaup et al. (2002b) para T. pubescens, a produção de
lacase não é inibida pela presença de glucose em altas concentrações, mas sim pelo baixo
valor do pH do meio de cultura, que é resultante do consumo de glucose. É importante
salientar que o estudo de Galhaup et al. (2002b) não foram efectuados em biorreactor e
portanto só estudaram situações sem controlo de pH.
Os resultados mostrados em bioreactor explicam as altas produções de lacase em
matraz em condições de limitação por carbono, quando se estuda o efeito de diferentes
concentrações iniciais de glucose. Pode-se afirmar que quando não havia glucose no
meio, em condições de limitação por carbono, se obtiveram as maiores produções de
lacase devido ao mais elevado valor de pH, e as menores actividades observadas com as
105

Resultados e Discussão
maiores concentrações de glucose não são devidas aos mecanismos de repressão por
glucose, mas sim ao baixo valor de pH do meio nestas condições.
3.2.2 – Modelação matemática da produção de lacase em bioreactor
Os resultados até aqui apresentados mostram a importância da adição de
indutores, assim como da concentração inicial de glucose no meio de cultura e do
controlo de pH. Para estudar o comportamento do fungo em bioreactor, nomeadamente o
crescimento celular, o consumo de glucose e a produção de lacase, foi-se avaliar o efeito
da limitação e ausência de limitação por carbono em matraz. A determinação dos
parâmetros cinéticos da fermentação em matraz foi necessária, uma vez que a biomassa
não pode ser medida directamente ao longo do tempo no bioreactor. De facto o fungo
cresce em agregados (pellets) quando se encontra num meio líquido, pelo que as culturas
não são homogéneas. Para se estimar os parâmetros da biomassa em bioreactor, assumiu-
se o crescimento em condições similares em matraz. Realizaram-se fermentações em 12
matrazes nas mesmas condições onde cada matraz correspondeu a um dia da
fermentação. Deste modo, os parâmetros cinéticos de crescimento foram obtidos e
posteriormente utilizados para a modelação matemática do bioreactor.
Para o desenvolvimento do modelo de crescimento celular, consumo de substrato
e produção enzimática em matraz, foi necessário considerar a existência de duas fases de
crescimento miceliar: uma fase rápida de crescimento exponencial seguida por uma fase
lenta de desaceleração no crescimento. Na fase exponencial de crescimento, ocorre o
metabolismo primário que está associado ao aumento da biomassa. Na fase de
desaceleração do crescimento, um diferente perfil de enzimas é produzido devido às
condições necessárias para o desenvolvimento de diferentes vias metabólicas. Este
modelo é descrito pelas equações 2.2a e 2.2b apresentadas anteriormente. Foi proposto
por Ikasari & Mitchell (2004), para fermentação em estado sólido, e é aqui adaptado para
fermentação em meio líquido, uma vez que o crescimento celular apresenta um perfil
muito semelhante ao estudado em estado sólido. As equações do modelo para as duas
fases foram desenvolvidas separadamente sendo o modelo ajustado aos dados
experimentais.
106

Resultados e Discussão
Os parâmetros cinéticos para a produção da lacase por T. versicolor com e sem
limitação por carbono, em matraz, estão apresentados na Tabela 3.4.
Tabela 3.4 – Parâmetros cinéticos da fermentação de Trametes versicolor em matraz
Parâmetros cinéticos Meio sem limitação por
carbono
Meio com limitação por
carbono
µmax (d-1) 0.87 0.82
Ks (g/L) 4.21 1.30
YX/S (g/g) 0.17 0.38
L 0.37 0.41
k (d-1) 0.0921 0.253
α (U/ g biomassa) 0.059 0.147
β (U/ g biomassa. d) 62.9 499
Os resultados da Tabela 3.4 mostram que a taxa específica de crescimento
máxima, µmax, é muito similar para ambos os meios de cultura: 0.87 dia-1 para o meio com
carbono e 0.82 dia-1 para o meio com limitação por carbono. Por outro lado, a constante
de saturação do substrato (Ks) é 3 vezes maior para o meio com carbono, uma vez que
esta constante está relacionada com a biomassa máxima obtida (1.7 g/L para o meio com
carbono e 0.8 g/L para o meio com limitação por carbono). O valor do parâmetro k,
constante de decaimento exponencial, obtida no meio com limitação por carbono, é 2.7
vezes maior do que no meio com carbono, o que indica que ocorre uma grande
desaceleração no crescimento celular quando o carbono está presente. Uma vez que
durante a fase exponencial de crescimento, o meio de cultura é o mesmo, o parâmetro L
(razão entre a taxa específica de crescimento no início da fase de desaceleração e a taxa
específica de crescimento durante a fase exponencial prévia), é semelhante para ambos os
meios.
Para a produção da lacase, o modelo de Luedeking-Piret (equação 2.4, capítulo 2)
foi adoptado, uma vez que a formação da lacase não é produto do metabolismo primário,
107

Resultados e Discussão
mas sim do metabolismo secundário. Os resultados apresentados na Tabela 3.4 mostram
que o valor da constante associada ao crescimento, α, é muito baixo (α = 0.059 ou α =
0.147 U/g de biomassa) quando comparado com o valor de β, constante não associada ao
crescimento (β = 62,9 ou β = 499 U g-1 de biomassa d-1), o que mostra que em ambos os
meios a produção da lacase somente se inicia na fase de desaceleração lenta no
crescimento, que se inicia após o terceiro dia. Este modelo de produção de lacase está de
acordo com a definição das duas fases distintas para o crescimento da biomassa
previamente adoptado.
Os resultados do modelo, juntamente com os dados experimentais para os meios
com e sem limitação por carbono para cultivo em matraz, estão apresentados nas Figuras
3.15 e 3.16. Os modelos utilizados correlacionam com grande aproximação o crescimento
da biomassa, o consumo de substrato e a síntese da lacase em cultivo submerso não
homogéneo.
Os dados das Figuras 3.15 e 3.16 mostram que a produção da lacase em matraz
somente tem inicio a partir do 5º dia de cultivo, quando o crescimento da biomassa se
encontra na fase de desaceleração, tanto para o meio com carbono como para o meio com
limitação por carbono, conforme já mostrado através dos parâmetros α e β.
Os resultados da modelação matemática em bioreactor para as oito experiências
realizadas com planeamento factorial (Tabela 2.5), onde se avalia a concentração da
biomassa, consumo de substrato e produção de lacase, são mostrados nas Figuras 3.17-
3.19. Assim como na modelação dos resultados em matraz, também aqui se obteve uma
boa descrição dos dados experimentais. O perfil da produção de lacase no meio com
limitação por carbono (F1, F2, F5 e F6) e sem limitação por carbono (F3, F4, F7 e F8) é
mostrado nas Figuras 3.17 e 3.18, respectivamente. A Figura 3.19 mostra a modelação
matemática do consumo de glucose e do crescimento da biomassa. O modelo utilizado
parece permitir uma previsão adequada do perfil de crescimento da biomassa, uma vez
que a curva do modelo descreve correctamente o único valor acessível
experimentalmente que é o da biomassa final.
Nas melhores condições de fermentação, ou seja, para o meio com glucose e
controlo de pH (F4 e F8), a produção da lacase atinge uma actividade máxima ao 5º dia,
apesar do meio de cultura apresentar ainda glucose e crescimento de biomassa. Para
108

Resultados e Discussão
meios sem controlo de pH (F3 e F7) o perfil da actividade da lacase apresenta um
aumento contínuo sem chegar a ser atingido um pico de actividade máxima.
0
0,4
0,8
1,2
1,6
2
0 2 4 6 8 10 12 14
Tempo (dias)
Bio
mas
sa (g
/L)
0
2
4
6
8
10
12
Glu
cose
(g/L
)
Experimental X Calculado X Experimental S Calculado S
0100200300400500600700800
0 2 4 6 8 10 12 14
Tempo (dias)
Laca
se (U
/L)
Experimental Calculado
Figura 3.15- Dados experimentais e modelados para o cultivo de Trametes versicolor em matraz com o meio com carbono. (X) biomassa; (S) glucose.
0,00
0,20
0,40
0,60
0,80
1,00
0 2 4 6 8 10
Tempo (dias)
Bio
mas
sa (
g/L)
0
2
4
6
8
10
12
14
Glu
cose
(g/
L)
Experimental X Calculado X Experimental S Calculado S
0
500
1.000
1.500
2.000
2.500
3.000
0 2 4 6 8 1
Tempo (dias)
Laca
se (U
/L)
Experimental Calculado
0
Figura 3.16 - Dados experimentais e modelados para o cultivo de Trametes versicolor em matraz com o meio com limitação por carbono. (X) biomassa; (S) glucose.
De acordo com os resultados modelados e a concentração da biomassa final, para
as experiências com glucose, o crescimento da biomassa não foi influenciado pelas
condições estudadas, em todos os casos, a concentração final de biomassa obtida foi de
1.5 g/L aproximadamente indicando que o aumento na actividade da lacase não foi
devido ao aumento da biomassa, mas sim às diferenças nas condições de cultivo. O perfil
de crescimento da biomassa foi o mesmo para todas as condições estudadas, com a fase
estacionária a ter inicio no final da fermentação.
109

Resultados e Discussão
0
500
1000
1500
2000
2500
0 2 4 6 8 10
Tempo (dias)
Laca
se (U
/L)
0
500
1000
1500
2000
2500
0 2 4 6 8 10
Tempo (dias)
Laca
se (U
/L)
F1 F2
Figura 3.17 - Dados experimentais e modelados para a produção de lacase em bioreactor por
Trametes versicolor em diferentes condições de fermentação: F1, F2, F5 e F6 (meios com limitação por carbono). Os dados experimentais estão representados por símbolos e os modelados por linhas.
0
500
1000
1500
2000
2500
0 2 4 6 8 10
Tempo (dias)
Laca
se (U
/L)
0
500
1000
1500
2000
2500
0 2 4 6 8 10
Tempo (dias)
Laca
se (U
/L)
F5 F6
0
200
400
600
800
0 2 4 6 8 10Tempo (dias)
Laca
se (U
/L)m
0
2000
4000
6000
8000
0 2 4 6 8 10
Tempo (dias)
Laca
se (U
/L)
0
200
400
600
800
0 2 4 6 8 10
Tempo (dias)
Laca
se (U
/L
F3 F4
0
2000
4000
6000
8000
0 2 4 6 8 10Tempo (dias)
Laca
se (U
/L)
F8F7)
Figura 3.18 - Dados experimentais e modelados para a produção de lacase em bioreactor por
Trametes versicolor em diferentes condições de fermentação: F3, F4, F7 e F8 (meios com carbono). Os dados experimentais estão representados por símbolos e os modelados por linhas.
110

Resultados e Discussão
0
2
4
6
8
10
0 2 4 6 8 10Tempo (dias)
Glu
cose
(g/L
)
0,0
0,4
0,8
1,2
1,6
2,0
Bio
mas
sa (g
/L)
Sexp Scalc Xcalc Xfinal
0
2
4
6
8
10
0 2 4 6 8 10
Tempo (dias)
Glu
cose
(g/L
)
0,0
0,4
0,8
1,2
1,6
2,0
Bio
mas
sa (g
/L)
Sexp Scalc Xcalc Xfinal
0
2
4
6
8
10
0 2 4 6 8 10Tempo (dias)
Glu
cose
(g/L
)
0,0
0,4
0,8
1,2
1,6
2,0
Bio
mas
sa (
g/L)
i
Sexp Scalc Xcalc Xfinal
0
2
4
6
8
10
12
0 2 4 6 8 10
Tempo (dias)G
luco
se (g
/L)
0,0
0,4
0,8
1,2
1,6
2,0
Bio
mas
sa (g
/L)
Sexp Scalc Xcalc Xfinal
F7 F8
F3 F4
Figura 3.19 - Dados experimentais e modelados para o consumo de glucose e dados modelados para o crescimento da biomassa em bioreactor por Trametes versicolor em diferentes condições de fermentação: F3, F4, F7 e F8 (meios com glucose). Os dados experimentais estão representados por símbolos e os modelados por linhas.
Os parâmetros cinéticos para a síntese da lacase (K1), degradação da lacase (K2) e
o parâmetro A estão apresentados na Tabela 3.5. Para as fermentações com limitação por
carbono, o parâmetro A foi tomado como 1, uma vez que a mudança do matraz para o
ambiente do bioreactor pode afectar somente o crescimento do fungo e não a síntese
enzimática e sob estas condições nenhum crescimento da biomassa foi observado. Dos
resultados apresentados na Tabela 3.5, é possível afirmar que a síntese de lacase por T.
versicolor é essencialmente determinada pela relação entre a presença de carbono no
meio de cultura e o controlo de pH, uma vez que nestas experiências foram obtidos
valores elevados de K1 (F4 e F8). Também se pode verificar que a influência da agitação
na fermentação não determina a produção enzimática no intervalo estudado. Os valores
de K2 apresentam um perfil similar aos obtido por K1, i.e. altos valores de K1 indicam
altos valores de K2, como esperado. Este resultado indica a existência de altos níveis de
inativação enzimática.
111

Resultados e Discussão
Tabela 3.5 – Parâmetros para a produção de lacase em bioreactor por Trametes versicolor
Experiência A K1 (U/ (g biomassa.d) K2 (d-1)
F1 1 1443 0.263
F2 1 1293 0.335
F3 0.51 14.3 0.026
F4 0.54 1932 0.667
F5 1 771.3 0.039
F6 1 564.4 0.084
F7 0.85 78.9 0.377
F8 0.64 2088 1.55
3.3 – Aplicação da lacase para o biobranqueamento da pasta de papel 3.3.1 – Comparação dos mediadores da lacase para o branqueamento da pasta
3.3.1.1 – Ensaios de biobranqueamento da pasta Embora a lacase seja uma enzima oxidativa, quando directamente aplicada nas
pastas não proporciona uma oxidação eficaz. A sua aplicação no branqueamento das
pastas tem sido muito estudada, mas ainda não se conseguiram resultados satisfatórios.
De facto a lacase sozinha apresenta um baixo potencial de oxidação-redução, podendo
apenas oxidar os fragmentos fenólicos da lenhina (Fabbrini et al., 2002). No entanto,
ultimamente a aplicação da lacase tem sido estudada em sistemas com mediadores
desempenhando o papel de reoxidante dos mediadores.
Neste estudo, fez-se uma comparação da aplicação da lacase nas pastas com
diferentes mediadores orgânicos, já estudados por diversos autores (Bourbonnais &
Paice, 1996; Bourbonnais et al., 1997; Bourbonnais et al., 1998; Call & Mucke, 1997;
Fabbrini et al., 2002, Ander et al., 1998; Li et al., 1999) e com mediadores inorgânicos,
nomeadamente polioxometalatos (Balakshin et al., 1998; Balakshin et al., 2001; Gamelas
et al. 2005). As principais vantagens dos POMs em relação aos mediadores orgânicos são
112

Resultados e Discussão
os altos potenciais redox, a capacidade de reoxidação, alta selectividade (capacidade de
deslenhificação com um mínimo de degradação dos polissacarídeos).
Todos os testes foram realizados a 45ºC e pH 4.5 que é aceitável do ponto de vista
da estabilidade/actividade da lacase. A redução do índice kappa traduz a oxidação dos
compostos insaturáveis nas pastas e a perda de viscosidade intrínseca está directamente
associada à degradação dos polissacarídeos da pasta. Esta perda de viscosidade não é
desejável para o fabrico de papel, uma vez que altera as propriedades mecânicas da pasta
branqueada. É o balanço deste binómio que dita a eficiência do processo de
branqueamento.
Na Tabela 3.6 e na Figura 3.20 encontram-se resultados dos branqueamentos da
pasta de papel com lacase e sem mediador; com lacase (L) e mediadores orgânicos
TEMPO, ABTS, HBT e ácido violúrico; e com lacase e mediadores inorgânicos POMs
(PW11MnIII, SiW11MnIII e SiW11VV). Conforme o previsto, quando a lacase se encontra
sem mediador no sistema de branqueamento de pasta de papel, os resultados da redução
no índice kappa são baixos, com apenas 11% de redução, quando comparados com o
sistema lacase-mediador (SLM), onde a redução no índice kappa variou de 13.0 a 26.2%,
dependendo do mediador utilizado. Houve sempre uma deslenhificação semelhante ao
trabalhar com este sistema SLM, sendo o melhor resultado obtido pelo ácido violúrico,
que apresenta uma maior redução no índice kappa e simultaneamente uma perda de
viscosidade de 8.4%, o que corresponde a uma boa selectividade de oxidação. Dos
mediadores testados, o TEMPO apresentou uma baixa viscosidade da pasta de 575 cm3/g
(perda de viscosidade de 51.4%), indicando uma extensa degradação dos polissacarídeos,
não sendo portanto, um mediador eficaz.
Todos os POMs estudados nestas condições apresentaram uma reduzida
deslenhificação, o que não permite distinguir qual o POM que apresenta a melhor
capacidade de branqueamento da pasta nestas condições. É importante notar que o pH
utilizado encontra-se dentro do intervalo de estabilidade (3-7) destes POMs (Tourné et
al., 1970).
113

Resultados e Discussão
Tabela 3.6 – Branqueamento da pasta kraft catalisado por lacase (L) na presença de diferentes mediadores a 45ºC
Sistema de branqueamento
Índice Kappa
Viscosidade Intrínseca
(cm3/g)
Redução do índice kappa
(%)
Perda de viscosidade
intrínseca (%)
POM oxidado
(%)
Pasta kraft (controlo) 12.3 1185 - - -
Lacase 10.9 1170 11.0 1.4 -
TEMPO+L 10.5 575 14.2 51.4 -
ABTS+L 10.2 1130 16.2 4.6 -
HBT+L 10.1 1100 17.5 7.2 -
Ácido violúrico +L 9.0 1085 26.2 8.4 -
PW11MnIII+L 10.2 1160 17.0 2.0 4.0
SiW11MnIII+L 10.3 1155 15.8 2.6 47
SiW11VV+L 10.6 1145 13.0 3.0 100
02468
101214
Past
a
Laca
se
L+TE
MPO
L+Si
W11
Mn
L+AB
TS
L+H
BT
L+Ac
.Vi
olúr
ico
Visc
osid
ade
intr
ínse
ca (c
m3/
g)
0200400600800100012001400
Índice kappa Viscosidade intrínseca
Índi
ce k
appa
L +
SiW
11M
nIII
L +
SiW
11VV
Índi
ce k
appa
L+S
iW11
MnI
II
Figura 3.20 – Variação da viscosidade intrínseca e do índice kappa para o sistema de branqueamento da pasta de papel a 45 ºC com diferentes mediadores.
Com a finalidade de se aumentar o grau de deslenhificação no SLM, uma vez que
os branqueamentos com POMs são referidos a elevadas temperaturas (Gaspar et al.
2003), acima de 100ºC, fez-se uma nova experiência com 60ºC no sistema reacional,
dado que a lacase não permite operar a temperaturas mais elevadas. Testaram-se ainda
114

Resultados e Discussão
outros POMs tais como: BW11CoIII, (PW9)2MnIII e SiW11CoIII. Conforme os resultados
apresentados na Tabela 3.7 e na Figura 3.21 o aumento da temperatura do sistema de 45
ºC para 60 ºC não favoreceu a deslenhificação da pasta ocorrendo um aumento no índice
kappa quando se utilizaram os POMs PW11MnIII+L e SiW11MnIII+L. Somente para o
POM SiW11VV+L foi observado um ligeiro aumento na redução do índice kappa, de 13%
para 15%. Os demais mediadores estudados também não apresentaram reduções
significativas no índice kappa. A menor redução do índice kappa com o aumento da
temperatura para os sistemas PW11MnIII+L e SiW11MnIII+L é justificado pelo estado de
oxidação destes POMs no fim da reacção. De facto a 60 ºC a concentração de POM no
estado oxidado é menor. Este facto pode ser explicado pela menor eficiência da lacase na
reoxidação dos POMs a 60 ºC possivelmente devido à sua inactivação ou sua parcial
desnaturação a esta temperatura mais elevada. O ligeiro aumento na redução do índice
kappa conseguido com o POM SiW11VV está relacionado com a sua grande facilidade em
ser reoxidado pela lacase, apresentando uma percentagem de reoxidação de 79% no fim
do branqueamento.
O processo de oxidação da lenhina ocorre pela acção da forma oxidada do POM,
sendo a forma reduzida resultante, reoxidada directamente pela lacase. As condições
termodinâmicas das reacções redox podem ser apresentadas da seguinte forma:
Eo lenhina < Eo POM < Eo lacase (~0.8 V) ( Balakshin et al., 2001) (3.2)
Para se compreender melhor este processo de oxidação – redução dos POMs pela
lacase fizeram-se espectros UV-Vis no final dos branqueamentos e determinou-se a
percentagem de POM no estado oxidado. Além disso, fez-se um estudo em separado, sem
a pasta de papel, de reoxidação directa dos POMs pela lacase.
115

Resultados e Discussão
Tabela 3.7 – Branqueamento da pasta kraft catalisado por lacase na presença de diferentes mediadores a 60 ºC
Sistema de branqueamento
Índice kappa
Viscosidade Intrínseca
(cm3/g)
Redução do índice kappa (%)
Perda de viscosidade
intrínseca (%)
POM oxidado
(%)
Pasta kraft
(controlo) 12,3 1190 - -
-
Lacase 11,1 1150 7,5 2,8 -
B W11CoIII+L 11,3 1110 9,0 6,7 -
(PW9)2III +L 10,9 1140 11,9 3,9 -
SiCoW11 +L 10,2 1060 16,5 10,6 -
Ac. violúrico+L 8,9 1110 27,1 6,5 -
SiW11MnIII 10,4 1080 15,1 9,0 12
PW11MnIII+L 10,7 1140 12,7 4,3 4
SiW11MnIII+L 10,5 1100 13,8 7,3 33
SiW11VV+L 10,4 1130 15,50 3.0 79
7,5
8,5
9,5
10,5
11,5
12,5
Pas
ta
L+B
CoW
11
Laca
se
L +(
PW
)2M
nIII
L+P
W11
Mn
L+S
iW11
MnI
II
SiW
11M
nIII
L+S
iCoW
11
L+A
c. v
iorú
lico
L +
SiW
11V
v
Índi
ce K
appa
Índice kappa Viscosidade Íntrinseca
950
1000
1050
1100
1150
1200
Visc
osid
ade
intr
ínse
ca (c
m3/
g)
L+PW
11M
nIII
SiW
11M
nIII
L+S
iW11
MnI
I
L+S
iW11
CoI
II
L+S
iW11
VV
L+BW
11C
oIII
L+(P
W) 2
MnI
II
Figura 3.21 – Variação da viscosidade intrínseca e do índice kappa para o sistema de branqueamento da pasta de papel a 60 ºC com diferentes mediadores.
116

Resultados e Discussão
Na Figura 3.22 pode ver-se o espectro de absorção do SiW11MnIII 100% no estado
oxidado (controlo) e os resultados obtidos com este POM no final das diferentes
condições de branqueamentos (testes a 45 ºC e a 60 ºC). A banda característica deste
POM tem um máximo de absorção a 490nm permitindo calcular a percentagem deste
POM que permanece no estado oxidado no final de cada ensaio. Os resultados mostram
que o aumento deste POM no estado oxidado seguiu a seguinte ordem: SiW11MnIII a 60
ºC (12 % no estado oxidado) < L+ SiW11MnIII a 60 ºC (33 % no estado oxidado) < L+
SiW11MnIII a 45 ºC (47 % no estado oxidado). Para o branqueamento sem lacase (60ºC)
uma pequena quantidade de POM no estado oxidado foi encontrado. Quando se
adicionou lacase ao sistema de branqueamento com o POM, a percentagem de POM no
estado oxidado aumentou para 33% a 60 ºC e para 47% a 45 ºC. Estes resultados
mostram que a oxidação da pasta é mais eficiente quando para além do POM se usa a
lacase. Por outro lado, os resultados mostram ainda que a oxidação do POM pela lacase é
mais eficiente a 45 ºC do que a 60 ºC, o que justifica os resultados apresentados
anteriormente.
0
1
2
3
100 200 300 400 500 600 700 800 900
�
Abs
.
a
bcd
Abso
rvân
cia
Comprimento de onda (nm)
Figura 3.22 – Espectros de UV-Vis da solução de SiW11MnIII antes do branqueamento (controlo)
e ao final do branqueamento em diferentes condições de reacção: (a) controlo; (b)
L+SiW11MnIII a 45ºC; (c) L+SiW11MnIII a 60ºC; (d) SiW11MnIII a 60ºC.
A Figura 3.23 compara o estado de oxidação no final da reacção para os POMs
PW11MnIII, SiW11MnIII e SiW11VV. Estes resultados também mostram que para qualquer
117

Resultados e Discussão
POM, o aumento da temperatura faz com que uma menor concentração de POM seja
oxidada, devido à perda de actividade da lacase com a subida da temperatura. O SiW11VV
é o POM que se reoxida de um modo mais eficaz atingindo 100% de reoxidação a 45ºC,
seguido do SiW11MnIII que atinge 47% a 45ºC e finalmente o PW11MnIII que apenas
chega a 4% de reoxidação a 45ºC.
Com a finalidade de verificar se a lacase apresentava perda de actividade com o
aumento da temperatura, a sua actividade foi determinada ao fim de cada branqueamento.
Para as experiências com SiW11MnIII, a lacase apresentou uma perda na actividade de
21% (de 380.5 U/L inicial para 300 U/L no final) para o branqueamento a 45ºC e de 58%
(de 380.5 U/L inicial para 158 U/L no fim) para o branqueamento a 60ºC. Estes
resultados estão de acordo com a percentagem de SiW11MnIII no estado oxidado a 45ºC e
a 60ºC, confirmando a perda da actividade da lacase com o aumento da temperatura. Nos
sistemas com lacase e com os POMs PW11MnIII e SiW11VV não foi possível determinar a
actividade da lacase no final da reacção, uma vez que estes POMs reagem com o
substrato que avalia a actividade enzimática.
0
20
40
60
80
100
POM
no
esta
do o
xida
do
(%)ii
iiiiii
ii
POM
(con
trol
o)
L+Si
W11
VV45
ºC
L+Si
W11
VV60
ºC
L+Si
W11
MnI
II45
ºC
L+Si
W11
MnI
II 60
ºC
SiW
11M
nIII
60ºC
L+PW
11M
nIII
45ºC
L+PW
11M
nIII
60ºC
POM
no
esta
do o
xida
do (%
)
Figura 3.23 –Percentual de SiW11MnIII, SiW11VV e PW11MnIII no estado oxidado ao final da reacção para as diferentes condições de branqueamento.
118

Resultados e Discussão
119
3.3.1.2 Estudo de oxidação dos POMs pela lacase
No estudo comparativo de oxidação do SiW11VIV, SiW11MnII e PW11MnII pela
lacase avaliou-se a eficiência do sistema catalítico na ausência da pasta de papel. As
condições das reacções foram seleccionadas levando-se em consideração a estabilidade
da lacase e dos POMs (temperatura ambiente, pH=4.5, concentração de POM 3.0
mmol/L, actividade inicial de lacase de 380 U/L). O estado de oxidação dos POMs
também foi monitorado através dos espectros de UV-Vis e os resultados são apresentados
na Figura 3.24. A forma reduzida do SiW11MnII apresenta uma coloração amarela, que se
foi alterando gradualmente para cor de rosa (forma oxidada) com o decorrer da reacção.
A banda de transição tem um máximo de absorção ao comprimento de onda de 490-495
nm (Figura 3.24A). Do mesmo modo, a cor rosa escuro característica do SiW11VIV na
forma reduzida alterou-se para amarela, a cor da forma oxidada, apresentando uma banda
característica no espectro visível com um máximo de absorção em 490 nm (Figura
3.24B). Um comportamento semelhante foi observado para o PW11MnII, embora com
uma baixíssima taxa de reoxidação (espectro não apresentado). A oxidação do SiW11VIV
para SiW11VV pela lacase também foi monitorada por espectroscopia 51V RMN, que
confirma que a estrutura do SiW11VV (Figura 3.25) manteve-se inalterada.
0.0
0.5
1.0
300 400 500 600 700
Wavelength (nm)
Abso
rban
ce
a
b
c
d
e
0
0.5
1
1.5
2
2.5
300 400 500 600 700
A B
Wavelength (nm)
Abs
orba
nce
a
b
c
de
Figura 3.24 – Espectro UV-Vis para as soluções aquosas (3.0 mmol/L) de SiW11MnII (A) e SiW11VIV (B) a pH=4.5 e com adição de lacase (380 U/L) a temperatura ambiente e pressão atmosférica (a). A: 4h (b); 8 h (c); 24 h (d); 50 h (e); B: 15 min. (b); 30 min. (c); 60 min. (d); 120 min. (e).
Abs
orvâ
ncia
Abso
rvân
cia
Comprimento de onda (nm) Comprimento de onda (nm)
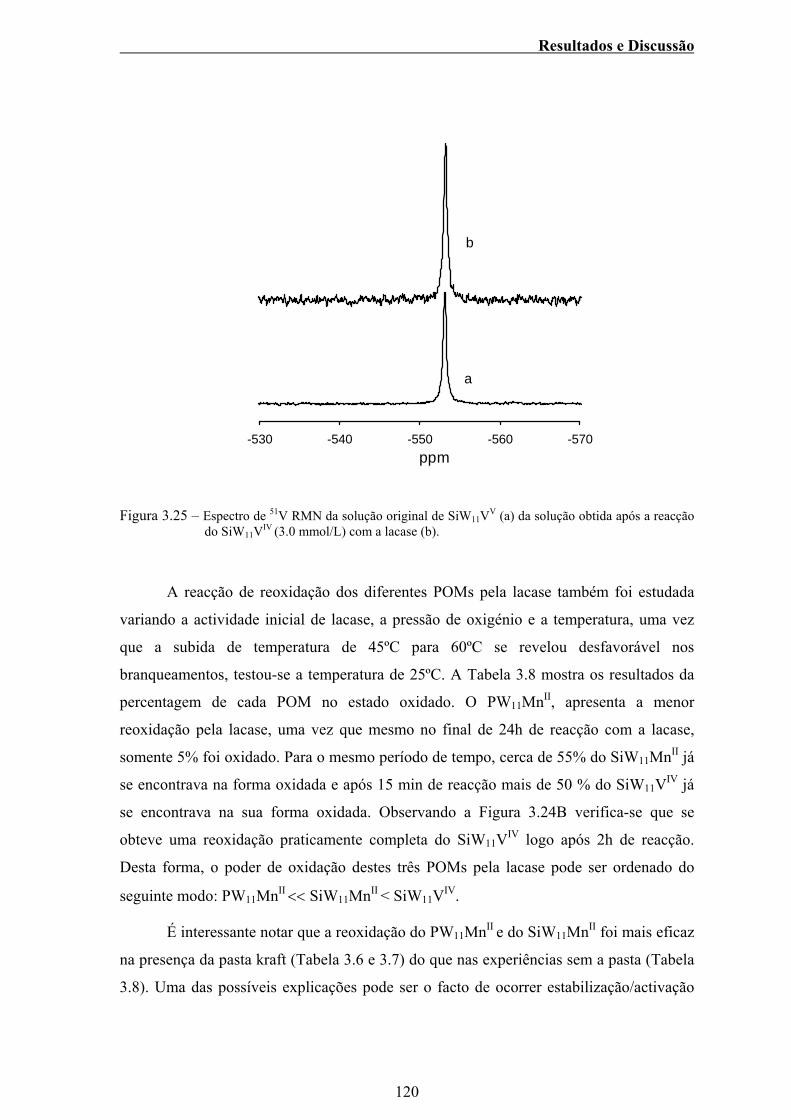
Resultados e Discussão
-570-560-550-540-530ppm
a
b
Figura 3.25 – Espectro de 51V RMN da solução original de SiW11VV (a) da solução obtida após a reacção do SiW11VIV (3.0 mmol/L) com a lacase (b).
A reacção de reoxidação dos diferentes POMs pela lacase também foi estudada
variando a actividade inicial de lacase, a pressão de oxigénio e a temperatura, uma vez
que a subida de temperatura de 45ºC para 60ºC se revelou desfavorável nos
branqueamentos, testou-se a temperatura de 25ºC. A Tabela 3.8 mostra os resultados da
percentagem de cada POM no estado oxidado. O PW11MnII, apresenta a menor
reoxidação pela lacase, uma vez que mesmo no final de 24h de reacção com a lacase,
somente 5% foi oxidado. Para o mesmo período de tempo, cerca de 55% do SiW11MnII já
se encontrava na forma oxidada e após 15 min de reacção mais de 50 % do SiW11VIV já
se encontrava na sua forma oxidada. Observando a Figura 3.24B verifica-se que se
obteve uma reoxidação praticamente completa do SiW11VIV logo após 2h de reacção.
Desta forma, o poder de oxidação destes três POMs pela lacase pode ser ordenado do
seguinte modo: PW11MnII << SiW11MnII < SiW11VIV.
É interessante notar que a reoxidação do PW11MnII e do SiW11MnII foi mais eficaz
na presença da pasta kraft (Tabela 3.6 e 3.7) do que nas experiências sem a pasta (Tabela
3.8). Uma das possíveis explicações pode ser o facto de ocorrer estabilização/activação
120

Resultados e Discussão
da lacase com os compostos fenólicos removidos da degradação parcial da lenhina. Mai
et al. (2000), mostrou que a presença de compostos fenólicos ou derivados da lenhina
aumentam a estabilidade e actividade da lacase, e sugere que esta estabilização pode ser
devido à ligação dos compostos ao centro activo da enzima ou a outra parte da cadeia da
proteína. Deste modo, certamente a reoxidação do PW11Mn e do SiW11Mn na presença da
pasta de papel, consegue ser mais eficaz devido à lacase se encontrar mais estável.
Os resultados mostram claramente que o efeito do aumento da concentração de
lacase de 380 U/L para 1330 U/L foi o mais significativo. O aumento da pressão de
oxigénio de 0.02 MPa para 0.3 MPa também melhorou a oxidação do POM pela lacase
como se vê no resultado obtido com o SiW11MnII. A temperatura apresentou um efeito
menor na reacção de oxidação deste mesmo POM. A temperatura máxima de reacção foi
limitada em 45ºC nestas experiências, uma vez que ocorreu uma diminuição na actividade
da lacase em temperaturas superiores a 50ºC. O POM que apresentou melhores resultados
quando oxidado pela lacase foi o SiW11VIV chegando até 82% após 15 min. de reacção. O
SiW11MnII também apresentou resultados significativos, uma vez que se oxidou em cerca
de 50% após 4 horas nas melhores condições de actividade inicial de lacase, pressão de
O2 e de temperatura. O PW11MnII apresentou um resultado insignificante uma vez que só
chegou a 5% de oxidação após 24 horas.
Tabela 3.8 – Oxidação dos polioxometalatos pela lacase em diferentes condições de reacção
POM Lacase (U/L) Pressão de O2 (MPa) Temperatura (ºC) POM oxidado (%)
SiW11MnII 380 ~0.02 25 16 (55)a
SiW11MnII 1330 ~0.02 25 35 (84)a
SiW11MnII 1330 0.3 25 48 (88)a
SiW11MnII 1330 0.3 45 52 (95)a
SiW11MnII 380 0.3 45 36 (69)a
SiW11VIV 380 ~0.02 25 52 (72)b
SiW11VIV 1330 ~0.02 25 82 (96)b
PW11MnII 380 ~0.02 25 <1 (5)a
a Medidas após 4 h ou após 24 h (dados entre parênteses). b Medidas após 15 min ou após 30 min (dados entre parênteses).
121

Resultados e Discussão
Considerando-se os potenciais redox destes POMs, a difícil oxidação do
PW11MnII pela lacase pode smer justificada pelo seu evado potencial (E=+0.91 V vs.
ENH) (Zhang et al., 1995). Uma vez que o potencial redox da lacase é muito próximo
deste, cerca de +0.8-0.9 V (vs. ENH) (Solomon et al., 1996; Mayer & Staples, 1996), este
POM é dificilmente reoxidado por esta enzima. Para os outros dois POMs, as diferenças
significativas no comportamento de oxidação do SiW11MnII e do SiW11VIV não podem
ser formalmente explicadas pelos potenciais redox descritos na literatura, uma vez que
são muito próximos. Para o SiW11V o potencial redox efectivo (E=(Epc+Epa)/2) do par
redox VV/IV foi apresentado como +0.64 V (vs. ENH, pH=5) (Altenau et al., 1975) e, em
outro estudo, como +0.72 V (vs. ENH, pH=4.5) (Harmalker & Pope, 1986).
Recentemente, Weinstock et al. (1997) mostrou o valor de potencial rdox efectivo de E =
+0.69 V para pH=5. Para o SiW11Mn o potencial redox do par MnIII/II foi medido como
+0.65 V (vs. ENH) (Tourné et al., 1970) e, mais recentemente, como +0.70-0.73 V (vs.
ENH) (Zhang et al., 1995; Sadakane & Steckhan, 1999; Sadakane & Steckhan, 1996).
Desta forma, uma comparação dos dados sobre os potenciais redox destes dois POMs não
é clara para se avaliarem as diferenças nos resultados. Além disso, sabe-se que os
potenciais redox dos POMs podem ser muito sensíveis a factores tais como o pH, os
contra iões e outros (Grigoriev et al., 2000).
Para se tentar esclarecer as diferenças nos potenciais redox desses dois POMs
(SiW11MnII e SiW11VIV) fez-se uma análise de voltametria cíclica em condições similares
às utilizadas nas experiências de branqueamento. Pelos resultados apresentados na Figura
3.26, pode-se observar que o par VV/IV no SiW11VIV é eletroquimicamente reversível
(Figura 3.26a) apresentando um E=+0.67 V (Epa=+0.70 V, Epc=+0.64 V, Epa-Epc=60 mV).
O correspondente par MnIII/II no SiW11Mn é quase - reversível (Figura 3.24b) e apresenta
um E=+0.76 V com Epa=+0.88 V, Epc=+0.64 V, e Epa-Epc=240 mV (vs. ENH). Deste
modo, a fácil oxidação do SiW11VIV pela lacase está relacionada com a sua alta
reversibilidade e com o baixo potencial de oxidação (Epa=+0.70V) do vanádio, quando
comparado com o do manganês, que apresenta um potencial de oxidação de Epa=+0.88V,
o que torna a sua reoxidação pela lacase mais difícil do que para o vanádio.
122

Resultados e Discussão
020040060080010001200
Potential (mV) vs. Ag/AgCl
1 µAa
b
Potencial (mV) vs. Ag/AgCl
Figura 3.26 – Voltametria cíclica (v=10 mV/s) dos POMs SiW11VIV (a) e SiW11MnII (b) ([POM]=1.0 mmol/L, pH=4.5, 25ºC).
3.3.2 – Branqueamento da pasta kraft com SiW11MnIII e lacase SiW11VV e lacase num sistema de multi-estágios
Como mostrado nos resultados anteriores, os POMs SiW11MnIII e SiW11VV são
facilmente reoxidados pela lacase a baixas temperaturas. Entretanto, sabe-se que estes
POMs actuam de forma mais eficiente quando as reacções de branqueamento com pasta
ocorrem a temperaturas superiores a 100ºC. Para se tentar solucionar esta
incompatibilidade de temperaturas óptimas, para o POM e para a lacase, realizou-se um
estudo catalítico de deslenhificação da pasta de papel em estágios separados e alternados.
Realizou-se o primeiro estágio de branqueamento com a temperatura óptima para o POM
(estágio POM a 110ºC) na ausência de lacase, seguido de um segundo estágio de
reoxidação do POM com a adição de lacase (estágio L) em difentes temperaturas (30, 45
e 60 ºC). Neste processo utilizaram-se os POMs SiW11MnIII, SiW11VV e PW11MnIII.
Os resultados obtidos com os estágios sequenciais de branqueamento com o
SiW11MnIII estão mostrados na Tabela 3.9. O tratamento prosseguiu até um total de 5
estágios POM e L alternados. Com um único estágio de branqueamento, o estágio POM,
pouca redução no índice kappa foi observada, resultando numa deslenhificação de apenas
23%, além do POM estar predominantemente no estado reduzido no final da reacção
(somente 4% encontrava-se no estado oxidado).
123

Resultados e Discussão
Quando a lacase foi adicionada no segundo estágio (L), um aumento de 5% no
grau de deslenhificação foi obtido e o POM passou a estar 68% no estado oxidado no
final do segundo estágio, o que foi importante para se obter uma maior deslenhificação
no estágio posterior. Adicionalmente, realizou-se um espectro de UV-vis do efluente
deste branqueamento depois de armazenado por uns dias (Figura 3.27). Nesta altura
quase 100% do POM se encontrava no estado oxidado. De qualquer modo, os dois
estágios de branqueamento mostraram melhores resultados, com um aumento de
aproximadamente duas vezes no grau de deslenhificação, do que um único estágio de
POM+L 45ºC (já apresentados na Tabela 3.8) por outro lado, demostrou uma menor
selectividade devido a maior perda na viscosidade.
Tabela 3.9 – Branqueamento da pasta kraft em estágios sequenciais catalisado por SiW11MnIII
(POM) e lacase
Sistema de
branqueamento
Índice
Kappa
Viscosidade
Intrínseca
(cm3/g)
Redução no
índice kappa
(%)
Perda de
viscosidade
Intrínseca
(%)
SiW11MnIII
(%, oxid.)
Pasta kraft 12.3 1185 - - -
Sem catalisador 10.6 1140 14 4 -
POM 9.5 1130 23 5 4*
POM-L (45ºC) 8.9 1080 28 9 68* (98)**
POM-L-(45ºC)-
POM
6.9 1075 44 9 7*
POM-L-(45ºC)-
POM-L(45ºC)-POM
6.0 1020 51 14 11*
POM-L (30ºC)-
POM
7.1 1065 42 10 -
POM-L (60ºC)-
POM
7.5 1065 39 10 -
PW11Mn-L 9.0 1085 27 8 4*
* Dados para o último estágio da sequência.** Solução armazenada durante uma semana.
Ao fim do terceiro estágio de branqueamento (POM-L-POM), uma grande
deslenhificação foi obtida, com 44% de redução no índice kappa e ao mesmo tempo
124

Resultados e Discussão
houve baixa perda de viscosidade, apenas 9%. A adição de mais um estágio POM na
sequência de branqueamento não demonstrou uma perda de viscosidade tão significativa
como quando se adicionou um estágio L. Este facto sugere que os estágios L são
responsáveis por uma maior perda de viscosidade. De facto esta enzima, para além de
oxidar o POM, oxida também a celulose pelo que promove a perda de viscosidade da
pasta de papel (Thurston, 1994; Accunzo & Galli, 2003).
Após 5 estágios consecutivos de branqueamento (POM-L-POM-L-POM),
obtveram-se os melhores resultados, com um índice kappa de 6.0, que corresponde mais
de 50% de deslenhificação da pasta, com apenas 14% de perda de viscosidade da pasta
(Tabela 3.9).
Para avaliar se a temperatura do estágio L apresenta algum efeito neste processo
de branqueamento em multi-estágios, testou-se a temperatura do estágio L também a
30ºC. Os resultados mostraram que a variação da temperatura no estágio L de 45ºC para
30 ºC e 60ºC não apresentaram nenhum benefício para os três estágios de branqueamento
da pasta (Tabela 3.9). Isto pode ser explicado pela menor reoxidação do SiW11MnII a
30ºC do que a 45ºC e pela parcial desnaturação da lacase a 60ºC.
Os espectros UV-Vis, apresentados na Figura 3.27, foram determinados para um
estágio, para dois estágios e para cinco estágios de branqueamento. Observa-se que nos
branqueamentos que não terminam num estágio L, o POM encontra-se praticamente todo
no estado reduzido (Figura 3.27 b, e).
O PW11MnIII também foi testado nas experiências de multi-estágios. Com dois
estágios de branqueamento, o grau de deslenhificação obtido foi similar ao SiW11MnIII
nas mesmas condições experimentais (Tabela 3.9). No entanto, a reoxidação do
PW11MnIII após o estágio com a lacase foi muito baixa, apenas 4% no estado oxidado, o
que não justificou o uso deste catalisador com 3 e 5 estágios de branqueamento.
É importante notar que a diminuição do índice kappa da pasta não branqueada é
determinada não somente pela oxidação da lenhina, mas também devido à hidrólise dos
resíduos de ácido hexourónico (HexA) ligados aos polissacarídeos da pasta. A pasta kraft
de eucalipto contém cerca de 30-40 mmol de HexA/kg de pasta seca, o que corresponde a
3-4 unidades de índice kappa (Furtado et al., 2001). Então, a 110ºC e pH 4.5 uma parcial
125

Resultados e Discussão
hidrólise do HexA contribui para a diminuição do índice kappa. Este facto foi confirmado
pela experiência em branco sem a adição de nenhum catalisador (Tabela 3.9).
0
0,5
1
1,5
300 400 500 600 700
Wavelength (nm)
Abs
orba
nce
c
d
b
a
e
Comprimento de onda (nm)
Abso
rvân
cia
Figura 3.27 – Espectros de UV-VIS da solução de SiW11MnIII antes do branqueamento (a); depois
de um estágio de branqueamento POM (b); depois de dois estágios de branqueamento POM-L (c) seguido de uma semana de armazenamento do efluente (d); depois de cinco estágios de branqueamento POM-L-POM-L-POM (e).
O mesmo tratamento de branqueamento da pasta em multi-estágios foi aplicado
para o POM SiW11VV. Os resultados mostrados na Tabela 3.10 indicam que este POM foi
mais eficiente na deslenhificação da pasta do que o SiW11MnIII, entretanto apresentou
uma menor selectividade. Os resultados mais próximos entre estes dois POMs foram
obtidos com dois estágios de branqueamento (POM-L), Tabelas 3.9 e 3.10. Com apenas
três estágios de branqueamento, o SiW11VV apresentou o mesmo grau de deslenhificação,
cerca de 50%, do que o SiW11MnIII com cinco estágios de branqueamento. O aumento da
temperatura também não favoreceu o branqueamento.
126

Resultados e Discussão
Tabela 3.10 - Branqueamento da pasta kraft em estágios sequenciais catalisado por SiW11VV e lacase
Sistema de
branqueamento
Índice
Kappa
Viscosidade
Intrínseca
(cm3/g)
Redução no
índice kappa
(%)
Perda de
Viscosidade
Intrínseca (%)
SiW11VV
oxidado
(%)
Pasta Kraft 12.3 1185 --- --- ---
POM 9.0 1055 26 11 6
POM-L (45ºC) 8.2 1035 33 13 98
POM-L (60ºC) 7.7 1010 37 15 80
POM-L (45ºC)-POM 6.9 1005 44 15 15
POM-L (60ºC)-POM 6.2 935 50 21 12
Similarmente ao SiW11Mn, o SiW11V foi encontrado predominantemente na
forma reduzida após os estágios POM e POM-L-POM, como mostrado na Figura 3.28,
que apresenta os espectros de UV/Vis no final das reações. Após o estágio POM-L cerca
de 80-100 % do SiW11V ficou no estado oxidado que corresponde a uma taxa de
reoxidação mais rápida do que para o SiW11Mn nas mesmas condições reacionais. O
espectro de RMN confirma a estabilidade do SiW11V após as reacções em multi-estágios
(Figura 3.28).
Figura 3.28 – Espectros de UV-Vis da solução de SiW11VV antes do branqueamento (a), depois
de um estágio POM de branqueamento (b), depois de dois estágios POM-L (45ºC) de branqueamento (c), depois de dois estágios POM-L (60ºC) de branqueamento (d), e depois de três estágios POM-L (60ºC)-POM de branqueamento (e).
0
0.5
1
1.5
2
2.5
300 400 500 600 700 800
Wavelength (nm)
Abs
orba
nce
a
b
c
d
e
Comprimento de onda (nm)
Abso
rvân
cia
127

Resultados e Discussão
A menor selectividade do SiW11VV no branqueamento da pasta quando
comparada com o SiW11MnIII pode ser explicada pela parcial dissociação, com a
libertação dos iões VO2+ do SiW11VV em condições acidificadas que favorecem a
degradação dos polissacarídeos da pasta de papel. Emboras as espécies de VO2+ não
terem sido detectadas pela análise de RMN (Figura 3.29) ao fim do branqueamento, a sua
formação durante o branqueamento pode ter ocorrido. Esses iões causam a
despolimerização oxidativa da pasta de papel. Os iões livres VO2+ apresentam um
potencial de redução mais alto, Eº= 0.9 V, do que o POM correspondente, além de exibir
um forte actividade catalítica em reacções com compostos orgânicos. Deste modo, VO2+
pode ser considerado como uma das espécies catalíticas mais activas, além de apresentar
um efeito pronunciado na oxidação da celulose durante a catálise do branqueamento da
pasta com o SiW11VV (Shatalov et al., 2000).
-600-580-560-540-520-500ppm
Figura 3.29 – Espectro de 51V RMN da solução obtida após dois estágios de branqueamento:
SiW11V-L (60ºC).
128

Resultados e Discussão
3.4 – Produção de exopolissacarídeo (EPS) por Trametes versicolor
3.4.1 Selecção do meio de produção do exopolissacarídeo
Com a finalidade de seleccionar o melhor meio de cultura para a produção de EPS
por T. versicolor estudaram-se em cinco diferentes meios já descritos: meio Tien & Kirk
(TaK) (utilizado na manutenção do fungo); meio definido para Trametes (MDT) (meio
utilizado na produção da lacase); meio extracto de malte e levedura (YM); meio completo
para fungos (MCM) e meio glucose extracto de levedura e peptona (GPY).
Dos cinco meios testados no cultivo, nenhuma produção de EPS por T. versicolor
foi observada para os meios MDT e GPY. O meio MDT, por ser um meio definido e
pobre, provavelmente não possui as fontes de nutrientes necessárias para sintetizar o EPS
pelo fungo. Por outro lado, o meio GPY apresenta uma alta concentração de glucose (40
g/L) que provavelmente inibe ou reprime o mecanismo de produção do EPS pelo fungo.
Os outros três meios usados, TaK, YM e MCM mostraram-se promissores para a
produção do EPS.
A Figura 3.30 mostra o crescimento da biomassa (a), o consumo de açúcares
redutores (b) e a produção do EPS (c) para os meios MCM, TaK e YM. Os resultados
mostram que a concentração dos açúcares redutores diminui durante a fermentação com o
correspondente aumento da biomassa. A curva da biomassa (Figura 30a) apresenta uma
fase de crescimento linear da biomassa entre o segundo e o quarto dia de cultivo e uma
fase de desaceleração no crescimento entre o quarto e o sétimo dia de cultivo. Este perfil
de crescimento da biomassa, com duas fases, foi o mesmo observado durante a produção
de lacase pelo T. versicolor neste estudo.
A produção do EPS somente se iniciou ao quarto dia de cultivo (Figura 3.30c), o
que mostra que a formação deste EPS não é associada à fase exponencial de crescimento
da biomassa (Figura 3.30a). A produção do EPS desencadeia-se quando termina o
crescimento celular mais activo e as células iniciam a fase de desaceleração do
crescimento. Este comportamento é típico dos produtos que não são produzidos pelo
metabolismo primário, cuja produção é associada com o crescimento exponencial da
biomassa, mas sim dos produtos do metabolismo secundário. Usualmente, os produtos
129

Resultados e Discussão
secundários dos fungos filamentosos estão relacionados com a produção dos metabólitos
secundários no início da fase de desaceleração no crescimento (Waites et al. 2001).
Comparando-se a produção do EPS nos três meios de cultura (Figura 3.30c),
observa-se que a produção termina após o sétimo dia de cultivo para os três meios
estudados. A produção máxima é obtida numa concentração de aproximadamente 700
mg/L (Tabela 3.11) no último dia de cultivo para o meio YM, enquanto que para os
meios TaK e MCM a concentração máxima de EPS obtida foi de 590 e 460 mg/L,
respectivamente. É de salientar que para ambos os meios a produção do EPS terminou
antes do consumo total de substrato.
A produção do EPS obtida com o meio YM (700 mg/L; Tabela 3.11) foi superior
do que a mostrada por Kim et al. (2002a) para o cultivo de C. versicolor, cuja máxima
produção do EPS no meio YM foi de 504 mg/L.
Tabela 3.11 - Resultados do cultivo de Trametes versicolor nos meios de cultura TaK, MCM e
YM.
Meio de cultura Parâmetros de cultivo
TaK MCM YM
Concentração máxima da biomassa (g/L) 4.4 3.3 3.8
Concentração máxima do EPS (mg/L) 590 460 700
Máxima viscosidade aparente (mPa.s) 26.0 16.0 47.1
Máxima produtividade do EPS (g/L d-1) 0.073 0.068 0.094
Rendimento (g de biomassa/g de açúcares
redutores consumidos)
0.24 0.17 0.23
Máximo EPS/biomassa (g/g) 0.16 0.14 0.24
130

Resultados e Discussão
(a)
(c)
(b)
0,0
1,0
2,0
3,0
4,0
0 2 4 6 8 1
Bio
mas
sa (g
/L
0
)
0,0
5,0
10,0
15,0
20,0
25,0
0 2 4 6 8 1
Açú
care
s re
duto
res
(g/L
0
)
0
0,2
0,4
0,6
0,8
1
0 2 4 6 8 1
Tempo (dias)
EPS
(g/L
)
0
TaK MCM YM Figura 3.30 – Crescimento da biomassa (a), consumo de açúcares redutores (b), produção de EPS
(c) durante a fermentação de Trametes versicolor nos meios TaK, MCM e YM.
A Tabela 3.11 mostra os resultados do cultivo de T. versicolor para a produção do
EPS nos meios de cultivo TaK, MCM e YM. Embora a alta concentração e o rendimento
da biomassa (g de biomassa/g de açúcares redutores consumidos) sejam obtidos com o
meio TaK, a máxima concentração e a máxima produtividade de EPS são conseguidas
usando-se o meio YM, conforme discutido anteriormente. Calculando-se a razão máxima
131

Resultados e Discussão
de EPS/biomassa (g/g), claramente se mostra que o meio YM apresenta os melhores
resultados com 0.24 g/g que é 1.5 e 1.7 vezes superior quando comparado com os meios
TaK e MCM, respectivamente. Por outro lado, TaK é o melhor meio para a produção de
biomassa, com concentrações finais de 4.4 g/L, enquanto que os meios YM e MCM
obtiveram concentrações finais de biomassa de apenas 3.8 e 3.3 g/L, respectivamente.
Estudos com o fungo do género Tremella mostraram que o YM foi o melhor meio
para a produção de polissacarídeo extracelular em termos de rendimento do polímero,
tanto para cultivos líquidos como para cultivos sólidos, quando comparado com o meio
GPY e com o meio potato dextrose agar (PDA) (Khondkar et al. 2002).
3.4.2 Optimização do meio de cultura YM para a produção de EPS
Os resultados discutidos anteriormente indicaram que YM foi o melhor meio de
cultura para a produção de EPS por T. versicolor e por isso foi seleccionado para a
optimização da produção do EPS. Os resultados apresentados para a produção da lacase
mostraram que, a concentração de glucose como fonte de carbono e o pH do meio de
cultura como um factor físico, são as duas principais condições de cultivo que podem
afectar tanto o crescimento celular como a formação do produto. Para optimizar o pH e a
concentração de glucose para a produção do EPS, o cultivo de T. versicolor foi estudado
no meio YM, partindo-se um planeamento de experiências 22 com um ponto central. Para
a glucose escolheu-se um nível inferior de 5 g/L, um nível superior de 15 g/L e um ponto
central de 10 g/L. Para o pH testou-se um nível inferior de 4, um nível superior de 7 e um
ponto central de 5.5.
A Figura 3.31 mostra os resultados da produção do EPS por T. versicolor em
diferentes valores de pH e concentrações iniciais de glucose no meio de cultura,
conforme o planeamento de experiências. Os meios com a maior concentração inicial de
glucose apresentaram um maior crescimento celular e maior produção de biopolímero. Os
meios com valor de pH de 4 apresentaram um consumo de açúcares redutores mais lento
pelo fungo, entretanto este facto resultou numa maior quantidade de biomassa formada.
Para os meios com concentração inicial de glucose 15 g/L, nota-se claramente que o valor
de pH de 4 favoreceu a produção da biomassa enquanto que o pH de 7 favoreceu a
132

Resultados e Discussão
produção do EPS. Para os meios com concentração inicial de glucose de 5 g/L já não é
possível notar-se este facto, uma vez que a produção do EPS pelo fungo é muito
aproximada durante o cultivo.
0,0
1,0
2,0
3,0
4,0
5,0
6,0
0 2 4 6 8 10
Bio
mas
sa (g
/
0
4
8
12
16
20
0 2 4 6 8 1
Açú
care
s re
duto
res
(g/L
)(a)
0
0
200
400
600
800
0 2 4 6 8 1
Tempo (dias)
EPS
(mg/
L)
G5 pH4 G5 pH7 G10 pH 5.5 G15 pH4 G15 pH7
Bio
mas
sa (g
/L)
Açú
care
s re
duct
ores
(g/L
)EP
S (m
g/L)
(b)
(c)
0
Figura 3.31 – Crescimento da biomassa (a), consumo de açúcares redutores (b) e produção de EPS (c) durante o cultivo de Trametes versicolor em diferentes concentrações (g/L) de glucose (G) e valores de pH.
133

Resultados e Discussão
A optimização do EPS foi realizada através do planeamento de experiências 22
com um ponto central conforme se encontra no gráfico de superfície de resposta na
Figura 3.32 e na Tabela 3.12. A resposta do planeamento foi a máxima produção de EPS
produzida no decorrer do cultivo. De acordo com a Figura 3.32 o pH não exerce uma
grande influencia na máxima produção do EPS, ao contrário do aumento da concentração
inicial de glucose que apresentou um grande aumento na máxima produção do EPS.
366,961 398,288 429,615 460,942 492,27 523,597 554,924 586,251 617,579 648,906 above
Superfície de Resposta; Variável: EPS2**(2-0) design; MS Residual=4898,45
DV: EPS
EPS=334,52+5,97*G-15,1*pH+3,03*G*pH
(mg/L)
Figura 3.32 – Superfície de resposta para o planeamento de experiências 22.
De acordo com os resultados deste planeamento de experiências não foi possível
obter o ponto de máxima curvatura para a máxima produção de EPS com os níveis de pH
e de glucose seleccionados. Deste modo, um segundo planeamento de experiências foi
realizado, onde os níveis dos factores passaram a ser: glucose com um nível inferior de 5
g/L, um nível superior de 25 g/L e um ponto central de 15 g/L e o pH com um nível
inferior de 4, um nível superior de 7 e um ponto central de 5.5.
134
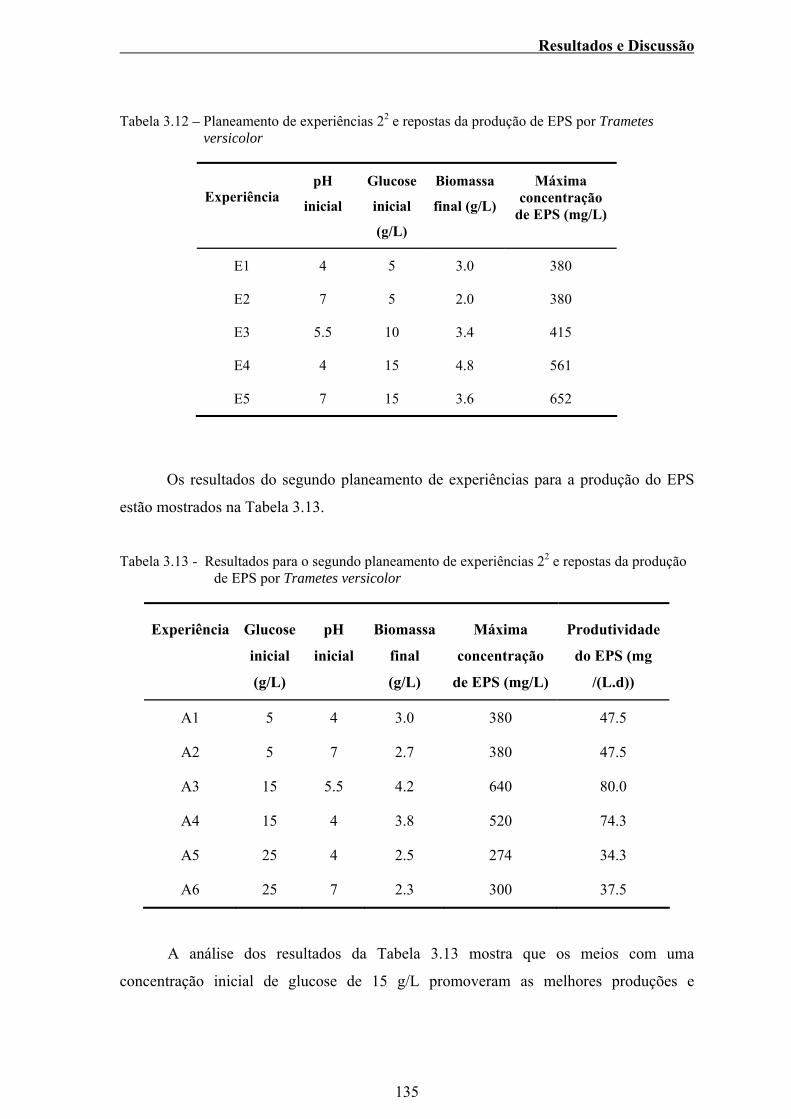
Resultados e Discussão
Tabela 3.12 – Planeamento de experiências 22 e repostas da produção de EPS por Trametes versicolor
Experiência pH
inicial
Glucose
inicial
(g/L)
Biomassa
final (g/L)
Máxima concentração
de EPS (mg/L)
E1 4 5 3.0 380
E2 7 5 2.0 380
E3 5.5 10 3.4 415
E4 4 15 4.8 561
E5 7 15 3.6 652
Os resultados do segundo planeamento de experiências para a produção do EPS
estão mostrados na Tabela 3.13.
Tabela 3.13 - Resultados para o segundo planeamento de experiências 22 e repostas da produção de EPS por Trametes versicolor
Experiência Glucose
inicial
(g/L)
pH
inicial
Biomassa
final
(g/L)
Máxima
concentração
de EPS (mg/L)
Produtividade
do EPS (mg
/(L.d))
A1 5 4 3.0 380 47.5
A2 5 7 2.7 380 47.5
A3 15 5.5 4.2 640 80.0
A4 15 4 3.8 520 74.3
A5 25 4 2.5 274 34.3
A6 25 7 2.3 300 37.5
A análise dos resultados da Tabela 3.13 mostra que os meios com uma
concentração inicial de glucose de 15 g/L promoveram as melhores produções e
135

Resultados e Discussão
produtividades do EPS, com 640 mg/L e 80 mg /(L.d) respectivamente, assim como o
melhor crescimento celular. Esses resultados indicam que esta concentração de glucose
foi a mais favorável para a produção do EPS provavelmente devido à uma concentração
celular elevada nestas condições. Para a maior concentração inicial de glucose, 25 g/L, a
produção do EPS não foi estimulada, resultando em baixas concentrações, menores que
300 mg/L e também em baixos crescimentos celulares. Elevadas concentrações de
glucose (acima de 20 g/L) provavelmente afectam o metabolismo da síntese do EPS por
T. versicolor assim como o próprio crescimento celular. Fang & Zhong (2002) sugerem
que na presença de altas concentrações de glucose (acima de 50 g/L) o crescimento do
fungo Ganoderma lucidum e a produção de um polissacarídeo intracelular foram inibidos
devido à pressão osmótica desfavorável do meio de cultura, entretanto a alta
concentração de glucose favoreceu a produção de polissacarídeo extracelular. Num outro
estudo, a produção do EPS pelo fungo Paecilomyces japonica também mostrou que a
concentração da fonte de carbono é um factor importante para a produção de um
biopolímero e para o crescimento do fungo, além de influenciar a morfologia do fungo e
a reologia do meio, que determina as características de transporte em altas concentrações
(acima de 40 g/L) (Sinha et al. 2001b).
Nesse segundo planeamento de experiências, o modelo de superfície de resposta
de segunda ordem é adoptado e mais um ponto no planeamento (ponto A4, Tabela 3.13)
foi adicionado. Este ponto correspondeu a mais um ponto central para não afectar as
estimativas usuais dos efeitos. O modelo usado contém somente os termos lineares (L) e
quadráticos (Q) dos efeitos principais. O modelo que contém os termos lineares e
quadráticos dos efeitos principais e a interacção dos dois factores também foi aplicado e
mostrou-se inadequado para representar os dados experimentais obtidos.
A significância de cada coeficiente foi determinada através do valor de p (p <
0.1), considerando-se 90% de intervalo de confiança, onde os baixos valores de p indicam
um valor significativo do efeito de cada factor (Tabela 3.14).
136

Resultados e Discussão
Tabela 3.14- Efeitos individuais calculados e ANOVA para o planeamento de experiências 22
Efeitos Estimado Valor de t Valor de p SS df MS F
Glucose
(L)
-93.0 -7.1538 0.0884 8649.00 1 8649.00 51.1775
Glucose
(Q)
-386.0 -12.1218 0.0524 24832.67 1 24832.67 146.9389
pH (L) 13.0 1.0000 0.500 169.00 1 169.00 1.0000
pH (Q) -227.0 -5.8205 0.1083 5725.44 1 5725.44 33.8784
Error 169.00 1 169.00
Total SS 97203.33 5
R2=0.99826
O gráfico de superfície de resposta (Figura 3.33) mostra o efeito da variação da
concentração inicial de glucose e do pH na produção do EPS. Tanto a variação da glucose
como a do pH afectaram a concentração máxima do EPS obtido. Neste caso foi possível
observar a curvatura do plano, ou seja, verificou-se a existência dos temos quadráticos no
modelo de regressão. As curvas de nível da Figura 3.33 servem para definir melhor a
região de máximo do rendimento. O máximo valor previsto para a produção do EPS foi
de 642.9 mg/L a uma concentração de glucose de 13.8 g/L e de pH de 5.5. A equação 3.3
representa o modelo da superfície de resposta para a produção do EPS (Figura 3.32):
EPS (mg/L) = -1274.28+53.25×G-1.93×G2+559.22×pH-50.44×pH2 (3.3)
137

Resultados e Discussão
-9,248 55,949 121,146 186,343 251,539 316,736 381,933 447,13 512,326 577,523 above
Superfície de Resposta; Variável: EPS2 factores, 6 experiências; MS Residual=169,
DV: EPS
Figura 3.33 – Superfície de resposta para a produção do EPS do segundo planeamento de experiências 22 com ponto central.
3.4.3 Análises do EPS
3.4.3.1 Análise dos açúcares neutros
A análise dos açúcares neutros presentes no EPS produzido nos três diferentes
meios YM, MCM e TaK foi realizada usando-se a metodologia de Blakeney et al. (1983)
e os resultados estão resumidos na Tabela 3.15. O açúcar predominante no EPS
produzido nos três meios de cultura foi a glucose, com quantidades relativas de 98%,
95% e 88% para os meios YM, TaK e MCM, respectivamente. Pequenas quantidades de
manose e xilose também foram detectadas no EPS produzido nestes meios em diferentes
quantidades. O EPS produzido no meio MCM contem 3% de outros açúcares, que
incluem a ramnose, a arabinose, a galactose e a fucose. Estes resultados indicam que o
meio de cultura influencia a composição dos açúcares neutros do EPS produzido por T.
versicolor. Entretanto, sem levar em consideração o meio de cultura, o açúcar
predominante é a glucose que está de acordo com os dados previamente publicados por
Miayazaki et al. (1974) para este fungo, onde o açúcar predominante foi a glucose com
138

Resultados e Discussão
um conteúdo estimado de 97.2%. Também Sugira et al. (1980) referiram que a glucose é
o principal açúcar do EPS extraído do micélio deste fungo.
Tabela 3.15- Açúcares neutros presentes no EPS produzido por Trametes versicolor nos meios
YM, MCM e TaK
Açúcares neutros (%, p/p) Meios
Glucose Manose Xilose Outros(*)
MCM 88% 7.4% 1.6% 3%
YM 98% 1.2% 0.8% traços (§)
TaK 95% 3.4% 1.6% traços (§)
(*) Ramnose, Arabinose, Galactose, Fucose (§) ≤ 0.1%
Os ácidos hexurónicos presentes no EPS produzido pelo T. versicolor foram
quantificados pelo total de ácido galacturónico (Anexo A4). Os resultados estão
apresentados na Tabela 3.16. Nota-se que o teor em ácidos hexurónicos do EPS
produzido no meio TaK é inferior aos dos outros EPSs produzidos nos meios YM e
MCM.
Tabela 3.16 - Ácidos hexourónicos presentes no EPS produzido por Trametes versicolor nos meios YM, MCM e TaK
Meio Ácidos Hexurónicos (mg/mg EPS)
MCM 59.6
YM 53.4
TaK 31.8
3.4.3.2 Análise Termogravimétrica
A análise termogravimétrica do EPS produzido nos meios YM, MCM e TaK pode
ser observada na Figura 3.34. O EPS foi termicamente estável até uma temperatura de
139

Resultados e Discussão
280°C para os meios TaK e YM e de 250ºC para o meio MCM. Acima destas
temperaturas a decomposição do EPS ocorreu em dois estágios: o primeiro ocorreu entre
aproximadamente 270ºC e 350ºC observando-se uma rápida perda de peso, que
representa uma perda de massa do EPS de 50%, 60% e 57% para o EPS produzido nos
meios YM, TaK e MCM, respectivamente; o segundo estágio de decomposição iniciou-se
a partir de 350°C até 560°C, onde uma decomposição mais lenta e gradual do EPS foi
registada, possivelmente devido à decomposição dos agregados de polissacarídeos. A
partir de 700°C nenhuma perda de peso foi observada, indicando um resíduo inorgânico
de 7.5 % para o EPS produzido nos meios YM e TaK e de 1.5% para o EPS produzido no
meio MCM.
0
20
40
60
80
100
120
0 200 400 600 800 1000
Temperatura (ºC)
Perd
a de
mas
sa (%
)
a
cb
Figura 3.34 – Análise termogravimétrica do EPS produzido nos meios YM (a), TaK (b) e MCM (c).
3.4.3.3 Análise de cromatografia de permeação em gel (GPC)
A distribuição do peso molecular médio do EPS obtido nos meios YM, MCM e
TaK está apresentada no cromatrograma da Figura 3.35. Os perfis da distribuição do peso
molecular foram muito semelhantes para o EPS produzido nos meios TaK e MCM,
enquanto que o EPS do meio YM apresentou um perfil de distribuição mais distinto, com
uma banda mais alargada o que pode representar a presença de agregados.
Os pesos moleculares médios obtidos foram: 67 KDa para o meio YM, 21 KDa
para o meio TaK e 25 KDa para o meio MCM. Comparando estes valores dos pesos
moleculares médios, com os polissacarídeos de T. versicolor PSK e PSP que apresentam
140
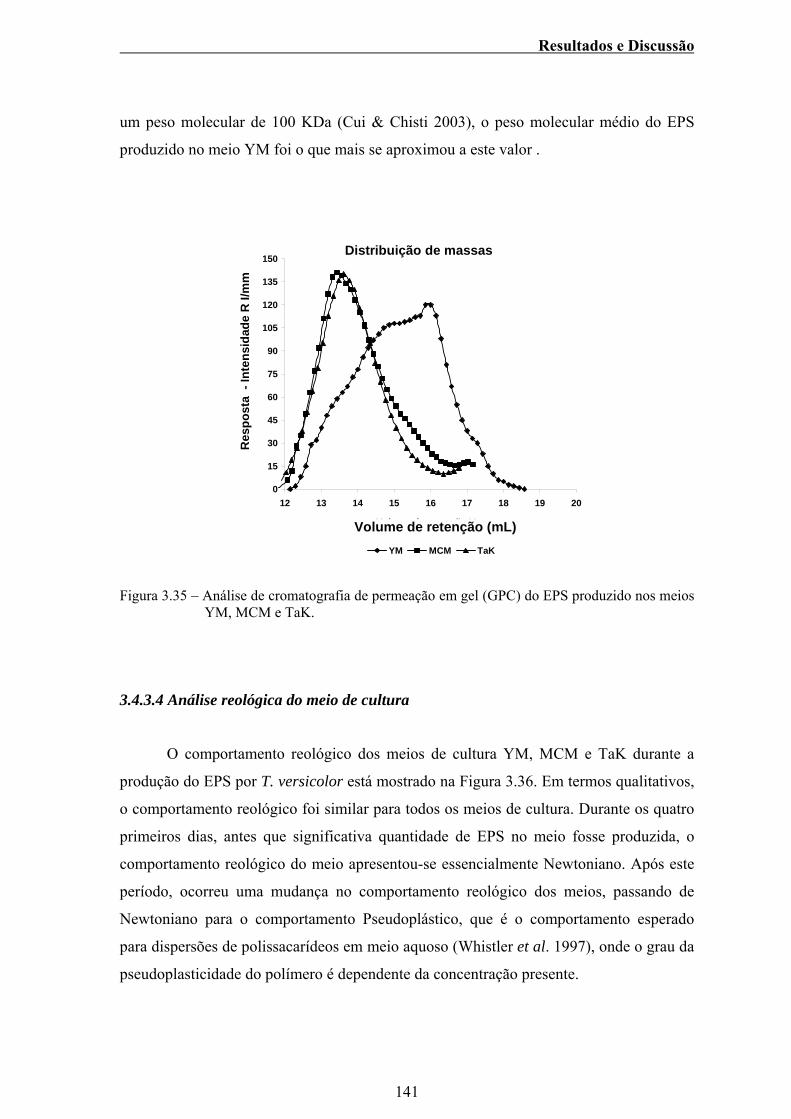
Resultados e Discussão
um peso molecular de 100 KDa (Cui & Chisti 2003), o peso molecular médio do EPS
produzido no meio YM foi o que mais se aproximou a este valor .
Distribuição de massas
0
15
30
45
60
75
90
105
120
135
150
12 13 14 15 16 17 18 19 20
Volume de retenção Vr
Res
post
a - I
nten
sida
de R
I/m
miii
iiiii
YM MCM TaK
Distribuição de massasR
espo
sta
-In
tens
idad
e R
I/m
m
Volume de retenção (mL)
Figura 3.35 – Análise de cromatografia de permeação em gel (GPC) do EPS produzido nos meios YM, MCM e TaK.
3.4.3.4 Análise reológica do meio de cultura
O comportamento reológico dos meios de cultura YM, MCM e TaK durante a
produção do EPS por T. versicolor está mostrado na Figura 3.36. Em termos qualitativos,
o comportamento reológico foi similar para todos os meios de cultura. Durante os quatro
primeiros dias, antes que significativa quantidade de EPS no meio fosse produzida, o
comportamento reológico do meio apresentou-se essencialmente Newtoniano. Após este
período, ocorreu uma mudança no comportamento reológico dos meios, passando de
Newtoniano para o comportamento Pseudoplástico, que é o comportamento esperado
para dispersões de polissacarídeos em meio aquoso (Whistler et al. 1997), onde o grau da
pseudoplasticidade do polímero é dependente da concentração presente.
141

Resultados e Discussão
A Figura 3.37 mostra a viscosidade aparente dos três meios de cultura durante
toda fermentação para uma taxa de cisalhamento de 10s-1. Os resultados mostram que a
viscosidade aparente do cultivo aumenta rapidamente a partir do quarto dia da
fermentação. Para o meio YM a viscosidade aparente aumentou continuamente até o
oitavo dia de cultivo, em concordância com os resultados obtidos de peso seco do EPS ao
longo da fermentação (3.31c).
Para os meios MCM e TaK a viscosidade aparente aumentou até ao sexto e sétimo
dia respectivamente (Figura 3.37), permanecendo a viscosidade constante até ao fim da
fermentação para o meio MCM enquanto que para o meio TaK houve uma ligeira
diminuição na viscosidade.
142
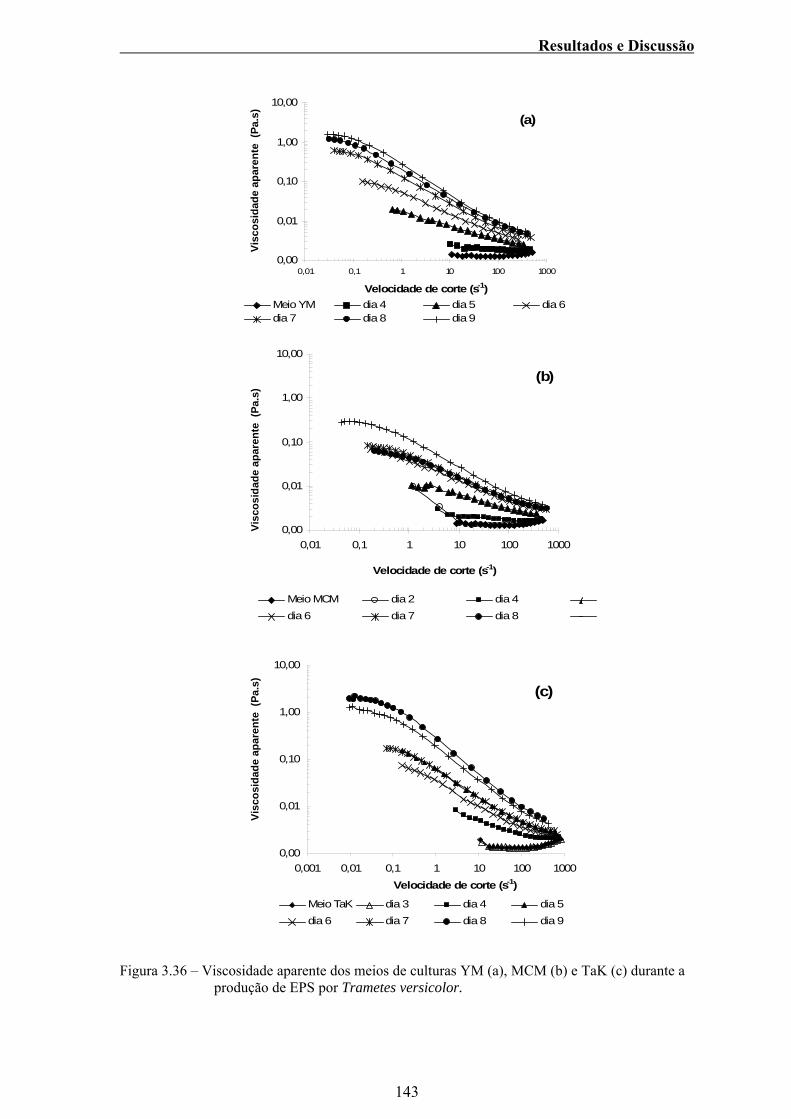
Resultados e Discussão
(a)
0,00
0,01
0,10
1,00
10,00
0,01 0,1 1 10 100 1000
Velocidade de corte (s-1)
Vis
cosi
dade
apa
rent
e (P
a.s)
Meio YM dia 4 dia 5 dia 6dia 7 dia 8 dia 9
Visc
osid
ade
apar
ente
(Pa
.s)
(b)
0,00
0,01
0,10
1,00
10,00
0,01 0,1 1 10 100 1000
Velocidade de corte (s-1)
Visc
osid
ade
apar
ente
(Pa.
s)i
Meio MCM dia 2 dia 4dia 6 dia 7 dia 8
Visc
osid
ade
apar
ente
(Pa
.s)
(c)
0,00
0,01
0,10
1,00
10,00
0,001 0,01 0,1 1 10 100 1000Velocidade de corte (s-1)
Visc
osid
ade
apar
ente
(P
Meio TaK dia 3 dia 4 dia 5dia 6 dia 7 dia 8 dia 9
Visc
osid
ade
apar
ente
(Pa
.s)
Figura 3.36 – Viscosidade aparente dos meios de culturas YM (a), MCM (b) e TaK (c) durante a produção de EPS por Trametes versicolor.
143

Resultados e Discussão
0,000
0,010
0,020
0,030
0,040
0 2 4 6 8Tempo (dias)
Visc
osid
ade
apar
ente
(P
a.s)
iiiiii
iiii
Visc
osid
ade
apar
ente
(Pa.
s)
10
TaK MCM YM
Figura 3.37 – Viscosidade aparente dos meios de cultura TaK, MCM e YM durante a produção do EPS por Trametes versicolor.
Comparando-se as Figuras 3.31c e 3.37 observa-se uma boa relação entre a
concentração do EPS e a viscosidade do meio de cultura. Uma vez que a quantificação do
EPS por peso seco é uma metodologia trabalhosa e que demora bastante tempo, a
avaliação da produção do EPS durante o tempo de fermentação pode ser determinada
através da análise reológica do meio de cultura, sendo esta metodologia com muito mais
precisão e rapidez. Deste modo, procurou-se uma correlação entre o peso seco do EPS e a
viscosidade aparente do meio de cultura YM e obteve-se uma correlação logarítmica
mostrada na Figura 3.38 e pela equação 3.4 (R2= 0.9903):
Y = 235.64 ln (X) + 1495.5 (3.4)
onde X é a concentração do EPS (mg/L) e Y é a viscosidade aparente (Pa.s).
y = 235,64Ln(x) + 1495,5R2 = 0,9903
0
200
400
600
800
1000
0 0,01 0,02 0,03 0,04 0,05Viscosidade aparente (Pa.s)
EPS
( mg/
L)iii
i
EPS
(mg/
mL)
Figura 3.38 – Concentração do EPS em relação à viscosidade aparente do meio de cultura.
144
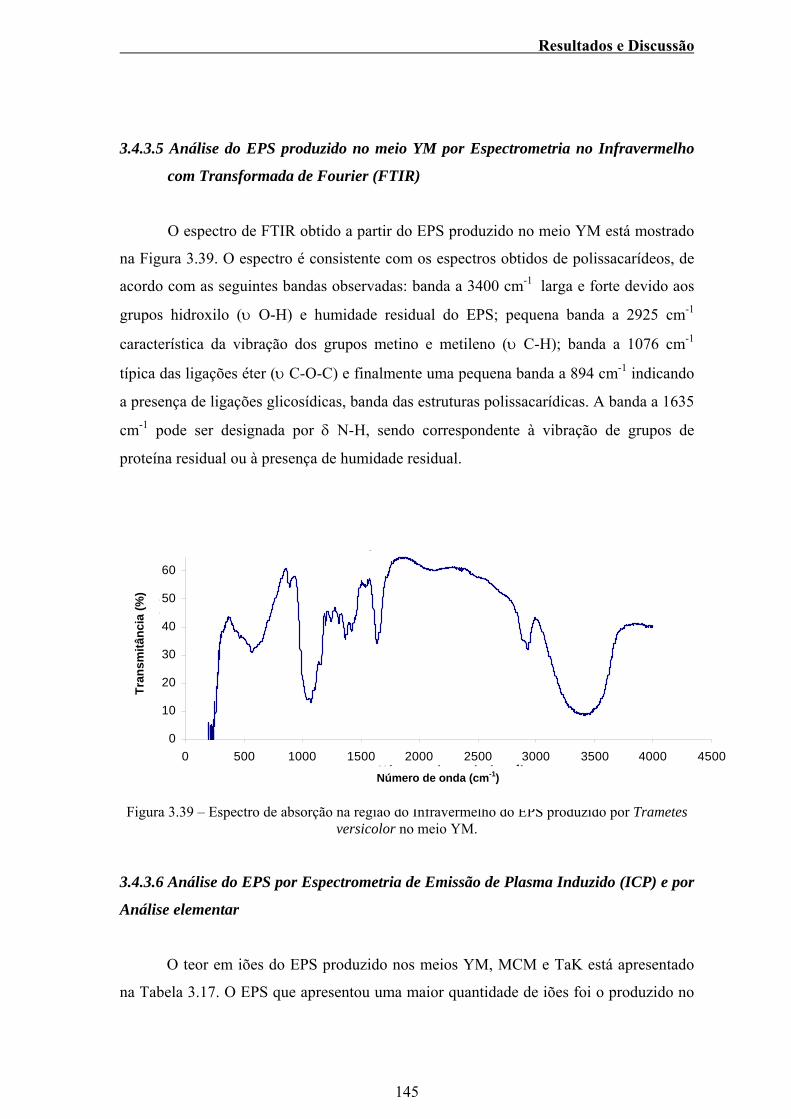
Resultados e Discussão
3.4.3.5 Análise do EPS produzido no meio YM por Espectrometria no Infravermelho
com Transformada de Fourier (FTIR)
O espectro de FTIR obtido a partir do EPS produzido no meio YM está mostrado
na Figura 3.39. O espectro é consistente com os espectros obtidos de polissacarídeos, de
acordo com as seguintes bandas observadas: banda a 3400 cm-1 larga e forte devido aos
grupos hidroxilo (υ O-H) e humidade residual do EPS; pequena banda a 2925 cm-1
característica da vibração dos grupos metino e metileno (υ C-H); banda a 1076 cm-1
típica das ligações éter (υ C-O-C) e finalmente uma pequena banda a 894 cm-1 indicando
a presença de ligações glicosídicas, banda das estruturas polissacarídicas. A banda a 1635
cm-1 pode ser designada por δ N-H, sendo correspondente à vibração de grupos de
proteína residual ou à presença de humidade residual.
Figura 3.39 – Espectro de absorção na região do Infravermelho do EPS produzido por Trametes versicolor no meio YM.
Espectro de IV da F6O
0
10
20
30
40
50
60
0 500 1000 1500 2000 2500 3000 3500 4000 4500-1)
Tran
smitâ
ncia
(%)
Tran
smitâ
ncia
(%)
Núm ero de onda (cmNúmero de onda (cm-1)
3.4.3.6 Análise do EPS por Espectrometria de Emissão de Plasma Induzido (ICP) e por
Análise elementar
O teor em iões do EPS produzido nos meios YM, MCM e TaK está apresentado
na Tabela 3.17. O EPS que apresentou uma maior quantidade de iões foi o produzido no
145

Resultados e Discussão
meio MCM, seguido pelo EPS produzido no meio TaK e no meio YM. Estes resultados
não puderam ser associados com a análise termogravimétrica uma vez que no meio MCM
o EPS apresentou uma menor quantidade de resíduos (1.5%) quando comparado com o
EPS produzido nos meios YM e TaK com 7.5% de massa residual (Figura 3.34).
Os dados da análise elementar do EPS mostram que este apresenta praticamente a
mesma composição em C, N e H para os três meios de produção. O EPS é composto por
uma maior percentagem em C, aproximadamente 35%, uma concentração em H entre 5 e
7% e um baixo teor em N com cerca de 1%.
Tabela 3.17 – Teor em iões e análise elementar do EPS produzido por Trametes versicolor nos meios YM, MCM e TaK
Iões (mg/g) (ICP) Análise elementar (%) Meios
P Fe Mg Ca Na K C H N
YM 2,08 0,15 0,15 1,52 0,69 2,29 34,45 5,65 0,9
MCM 8,10 0,27 8,32 2,57 0,91 10,4 35,98 7,05 0,97
TaK 8,66 0,08 5,24 6,75 0,09 1,01 35,91 6,4 1,1
146

Conclusões
Capítulo 4 - Conclusões Através deste trabalho, pode-se concluir que o fungo da podridão branca da
madeira, Trametes versicolor, foi capaz de produzir a enzima lacase em concentrações
significativas em matraz e em bioreactor:
I. A concentração inicial de glucose foi extremamente importante para a optimização da
produção da lacase em matraz, as menores actividades de lacase foram observadas
para as maiores concentrações iniciais de glucose (5 e 9 g/L), mostrando que altos
níveis de glucose reprimem a produção da lacase, enquanto que a limitação por
carbono favoreceu a produção enzimática;
II. O perfil da evolução da concentração de proteína total do meio de cultura não
coincide com o perfil da produção da lacase, o que indica que o fungo produz outras
proteínas certamente necessárias ao seu desenvolvimento;
III. A queda do pH durante a produção da lacase está directamente relacionada com o
consumo de glucose por T. versicolor devido à produção de ácidos orgânicos pelo
fungo;
IV. A adição do cobre ao meio de cultura foi mais favorável como micronutriente do que
como indutor da produção da lacase;
V. Dos diferentes indutores testados na produção da lacase, o etanol não favoreceu a
produção da enzima enquanto que o cobre, 2.5-xilidina e a mistura fenólica
apresentaram um aumento na actividade da lacase, e uma maior actividade foi obtida
quando foi empregue em simultâneo a mistura fenólica com 2.5 xilidina e cobre em
condições de limitação por carbono.
A optimização da produção da lacase em bioreactor por planeamento de experiências
apresentou as maiores actividades enzimáticas:
I. Controlo de pH, concentração inicial de glucose e as suas interacções apresentam um
grande efeito na actividade máxima da lacase;
147

Conclusões
II. A máxima actividade de lacase foi obtida quando o pH do meio de cultura estava
controlado e a concentração inicial de glucose era de 9 g/L;
III. A agitação não apresenta nenhum efeito significativo na produção da lacase;
IV. Com a modelação matemática da produção da lacase em bioreactor foi possível
determinar os parâmetros cinéticos da fermentação e de estimar-se o crescimento da
biomassa ao longo do tempo;
V. A lacase é produzida durante a fase de desaceleração do crescimento da biomassa, ou
seja, durante o metabolismo secundário do fungo.
Para além da optimização da produção, aplicou-se a lacase produzida pelo T. versicolor
no branqueamento da pasta kraft de papel onde diferentes mediadores da enzima e
diferentes condições experimentais foram avaliados:
I. Dos mediadores orgânicos (TEMPO, ABTS, HBT e ácido violúrico) e inorgânicos
POMs (PW11MnIII, SiW11MnIII, BW11CoIII, (PW9)2MnIII, SiW11CoIII e SiW11VV )
testados num único estágio de branqueamento, os POMs SiW11MnIII e SiW11VV
apresentaram os melhores resultados de branqueamento com uma maior
selectividade;
II. Os espectros de UV-Vis e de voltametria cíclica da soluções de SiW11MnIII e de
SiW11VV mostram que estes POMs são facilmente reoxidados pela lacase,
principalmente para o SiW11VV;
III. Os POMs actuam mais facilmente no branqueamento da pasta a temperaturas
elevadas (110ºC) enquanto que a lacase actua preferencialmente a baixas
temperaturas (45ºC);
IV. Os branqueamentos em estágios sequenciais, com a oxidação da lenhina pelo POM a
110ºC e a reoxidação do POM pela lacase num estágio separado a 45ºC,
apresentaram uma redução no índice kappa da pasta de cerca de 50% para o POM
SiW11MnIII com 5 estágios de branqueamento e para o SiW11VV com apenas 3
estágios de branqueamento, com uma pequena perda da viscosidade intrínseca
reduzida para ambos os POMs utilizados;
148

Conclusões
V. O SiW11MnIII apresentou-se como sendo o catalisador mais selectivo enquanto que o
SiW11VV apresentou-se como sendo o mais efectivo no branqueamento da pasta
durante o branqueamento em estágios sequenciais.
Para a selecção, optimização e análise do exopolissacarídeo produzido por T.
versicolor foram estudados:
I. Os meios de cultura apresentam um papel importante na produção do
exopolissacarídeo. Dos cinco diferentes meios de cultura empregados (MCM, TaK,
YM, GPY e MDT), os meios MDT e GPY não favoreceram a produção do EPS
enquanto que os meios MCM, TaK e YM produziram EPS. Destes três, o meio YM
foi o que apresentou a maior produção de EPS;
II. As análises de reologia dos meios MCM, TaK e YM mostram que qualitativamente
estes três meios são similares, apresentando um comportamento pseudoplástico a
partir do quarto dia de fermentação. A análise de reologia do meio de cultura surge
como sendo uma metodologia rápida e eficaz para o acompanhamento da produção
do EPS ao longo da fermentação;
III. A metodologia do planeamento de experiências permitiu optimizar a produção do
EPS no meio de cultura YM. Para a maior e menor concentração inicial de glucose (5
e 25 g/L) a produção do EPS não foi favorecida. Com a concentração de 15g/L de
glucose é que se optimizou a produção de EPS;
IV. O açúcar predominante no EPS produzido nos meios de cultura MCM, TaK e YM é a
glucose embora também existam pequenas quantidades de manose e xilose. O EPS
produzido no meio MCM contem 3% de outros açúcares, que incluem ramnose,
arabinose, galactose e fucose;
V. O EPS foi termicamente estável até uma temperatura de 250-280°C para os meios
MCM e YM. Acima destas temperaturas a decomposição do EPS ocorreu em dois
estágios: no primeiro uma rápida perda de peso é observada, enquanto que no
segundo estágio se observou uma lenta decomposição. A partir de 700°C nenhuma
perda de peso é obtida;
VI. Os pesos moleculares médios do EPS nos diferentes meios são 67 KDa para o meio
YM, 21 KDa para o meio TaK e 25 KDa para o meio MCM;
149

Conclusões
VII. O EPS no meio YM apresenta grupos hidroxilas (υ O-H), grupos metino e metileno
(υ C-H), ligações de éter (υ C-O-C), ligações glicosídicas e ligações δ N-H, que
correspondem à vibração de grupos de proteína residual ou a presença de humidade
residual.
150

Bibliografia
Bibliografia
1. Accunzo, F., Galli, C. First evidence of catalytic mediation by phenolic compounds in the laccase-induced oxidation of lignin models. European Journal of Biochemistry 270: 3634-3640 (2003).
2. Addleman, K., Archibald, F.S. Kraft pulp bleaching and delignification by dikaryons and monokaryons of Trametes versicolor. Applied and Environmental Microbiology 59:266-273 (1993).
3. Akhtar, M., Blanchette, R. A., Kirt T. K. Fungal Delignification and Biomechanical Pulping of Wood. Advances in Biochemical Engineering/Biotechnology, (T. Scheper, ed.), Springer-Verlag, Berlin, Heidelberg (1997).
4. Alexopoulos, C.L., Mims, C.W. Introductory Micology, 3ºEd. Cap.21:429-533 (1979).
5. Altenau, J., Pope, M.T., Prados, R.A., So, H. Models for heteropoly blues. Degrees of valence trapping in vanadium(IV)- and molybdenum(V)-substituted Keggin anions Inorganic Chemistry 14: 417- 421 (1975).
6. Amaral, PFF, Fernandes DLA, Tavares APM, Xavier ABMR, Cammarota MC, Coutinho JAP and Coelho MAZ, Decolorization of Dyes from textile wastewater by Trametes versicolor. Environmental Technology 25:1313-1320 (2004).
7. Ander, P., Grabner, K., Kandioller, G., Srebotnik, E., Poppius-levlin, K., Semar, S., Anke, H., Niku-Paavola, M., Viikari, L., Messner, K. The in vitro 14C-labelled kraft pulp assay as used for evaluation of laccases and mediators intended for pulp bleaching, 7th International Conference of Biotechnology in Pulp and Paper Industry, Vancouver, A111-115 (1998).
8. Ander, P., Messner K, Oxidation of 1-hydroxybenzotriazole by Laccase and Lignin Peroxidase. Biotechnology Techniques 12(3):191-195 (1998).
9. Archibald, F.S., Bourbonnais, R., Jurasek, L, Paice, M.G, Reid, I.D. Kraft pulp bleaching and delignification by Trametes versicolor. Journal of Biotechnology 53: 215-236 (1997).
10. Arora D. S.; Gill, P. K. Laccase production by some white rot fungi under different nutritional conditions. Bioresource Technology, 73:283-285 (2000).
11. Bae, J.T., Park, J.P., Song, C.H., Yu, C.B., Park, M.K., Yun J.W. Effect of carbon source on the mycelial growth and exo-biopolymer production by submerged culture of Paecilomyces japonica. Journal of Bioscience and Bioengineering 91 (5): 522-524 (2001).
12. Bailey, J.E., Ollis, D.F. Biochemical Engineering Fundamentals. 2ª ed, MacGraw-Hill, NY, (1986).
13. Balakshin, M., Chen, C., Gratzl, J.S., Kirkman, A.G., Jakob, H. Kinetics of pulp delignification with laccase-mediator system. 5th European Workshop on Lignocellulosics and pulp. 585-588 (1998).
151

Bibliografia
14. Balakshin, M.Y., Evtuguin, D.V., Neto, C. P., Cavaco-Paulo, A. Polyoxometalates as mediators in the laccase catalyzed delignification. Journal of Molecular Catalysis B: Enzymatic 16: 131–140. (2001).
15. Baldrian P, Gabriel, J. Copper and cadmium increase laccase activity in Pleurotus ostreatus. FEMS Microbiology Letters 206: 69-74 (2002).
16. Bauer, C. G.; Kuehn, A.; Gajovic, N.; Skorobogatko, O.; Holt, P.-J.; Bruce, N. C.; Makower, A.; Lowe, C. R.; Schelier, F. New enzyme sensors for morphine and codeine based on morphine dehydrogenase and laccase. Fresenius Journal of Analytical Chemistry, 364:179-183. 1999
17. Biermann, J. Handbook of pulping and papermaking, 2nd ed. Academic Press (1996).
18. Blakeney, A.B., Harris P.J., Henry R.J., Stone B.A. A simple and rapid preparation of alditol acetates for monosaccharide analysis. Carbohydrate Research 113: 291-299 (1983).
19. Blanchette, R.A. Degradation of lignocellulose complex in wood. Canadian Journal of Botany 73: 999-1010 (1995).
20. Blánquez, P., Casas, N., Font, X., Gabarrell, X., Sarrà, M., Caminal, G., Vicent, T. Mechanism of textile metal dye biotransformation by Trametes versicolor. Water Research 38 (8): 2166-2172 (2004).
21. Blumenkrantz N, Asboe-Hansen G. New method for quantitative determination of uronic acids. Analytical Biochemistry, 54(2):484-489(1973).
22. Bohn, J.A., BeMiller, J.N. (1-3)- β-D-glucans as biological response modifiers: a review of structure-functional activity relationships. Carbohydrate Polymers 28: 3-14 (1995).
23. Bollag, J. M., Leonowicz, A. Comparative studies of extracellular fungal laccases. Applied Environmental Microbiology 48: 849-854 (1984).
24. Boudet, A.M., Lapierre, C., Grima-Pettenati, J. Biochemistry and molecular biology of lignification. New Phytologist 129: 203-236 (1995).
25. Bourbonnais, R., Paice, M. G., Reid, I. D., Lanthier, P., Yaguchi, M. Lignin Oxidation by Laccase Isozymes from Trametes versicolor and Role of the Mediator 2,2`-Azinobis(3-Ethylbenzthiazoline-6-Sulfonate) in Kraft Lignin Depolymerization. Applied and Environmental Microbiology 61(5): 1876–1880 (1995).
26. Bourbonnais, R., Paice, M.G., Enzymatic delignification of kraft pulp using laccase and a mediator. Tappi Journal 79(6): 199-204 (1996).
27. Bourbonnais, R., Paice, M.G., Freiermuth, B. Bodie,E., Borneman S. Reactivities of Various Mediators and Laccases with Kraft Pulp and Lignin Model Compounds. Applied and Environmental Microbiology 63(12): 4627–4632 (1997).
28. Bourbonnais, R., Leech, D., Paice, M.G., Electrochemical analysis of the interactions of laccase mediators with lignin model compounds. Biochimica et Biophysica Acta 1379: 381–390 (1998).
152

Bibliografia
29. Box, G.E.P., Hunter, W.G., Hunter, J.S., (1978) Statistics for experimenters. An introduction to design, data analysis and model building. New York, USA.
30. Calado, V., Montgomery, D. Planejamento de experimentos usando o Statistica. E-papers, Rio de Janeiro, ISBN 85-87922-83-1 (2003).
31. Call, H.P., PCT. World Patent Application: WO 94/29510 (1994).
32. Call H.P., Mucke I. History, overview and applications of mediated lignolytic systems, especially laccase-mediator-systems (Lignozym®-process). Journal of Biotechnology 53:163–202 (1997).
33. Castro, A., Estudo da biodegradação da lenhina pelo fungo Trametes versicolor, Tese de Doutoramento, Universidade de Aveiro, Departamento de Química, (2001).
34. Castro, A., Evtuguin, D. V., Xavier, A. M. B. Degradation of biphenil lignin model compounds by laccase of Trametes versicolor in the presence of 1-hydroxybenzotriazole and heteropolyanion [SiW11VO40]5. Journal of Molecular Catalysis B: Enzymatic 22:13-20 (2003).
35. CELPA, Associação da Indústria Papeleira, Boletim Estatístico 2004. Disponível em http://www.celpa.pt/images/pdf/boletim2004.pdf. APER
36. Collins, P.J., Dobson A.D.W. Regulation of laccase Transcription in Trametes versicolor. Applied Environmental and Microbiology 63(9):3444-3450 (1997).
37. Couto, S. R., Gundín, M., Lorenzo, M., Sanromán, M. A. Screening of supports and inducers for laccase production by Trametes versicolor in semi-solid-state conditions. Process Biochemistry 38: 249-/255 (2002).
38. Couto, S. R, Sanromán, M. A., Hofer, D., Gundín, M. Stainless steel sponge: a novel carrier for the immobilization of the white-rot fungus Trametes hirsuta for decolourisation of textile dyes. Bioresource Technology, 95(1): 67-72 (2004a).
39. Couto, S.R, Rosales, E., Gundín, M., Sanromán, M. Exploitation of a waste from the brewing industry for laccase production by two Trametes species. Journal of Food Engineering 64: 423–428 (2004b).
40. Crueger, W., Crueger A. (1990) Biotechnology, a text book of industrial microbiology. 2nd Ed. p.60, Sunderland, Sinauer Associates ISBN 0-87893-131-7.
41. Cui, J., Chisti, Y. Polysccharopeptides of Coriolus versicolor: physiological activity, uses and production. Biotechnology Advances 21: 109-122 (2003).
42. Daba, A.S., Ezeronye, O.U. Anti-cancer effect of polysaccharides isolated from higher basidiomycetes mushrooms African Journal of Biotechnology 2 (12) 672-678 (2003). (Disponível online em http://www.academicjournals.org/AJB).
43. Dence, C. W. and Reeve, D. W. (Eds.), Pulp Bleaching – Principles and Practice, 1996, Tappi Press, Atlanta.
153

Bibliografia
44. Domaille, P. J. The 1- and 2-dimensional tungsten-183 and vanadium-51 NMR characterization of isopolymetalates and heteropolymetalates. Journal of the American Chemical Society 106 (25): 7677-7687 (1984).
45. Dumitriu, S. Polysaccharides: structural diversity and functional versatility. New York: Marcel Dekker ISBN: 0-8247-0127-5 cap.8 (1998).
46. Eggert, C., Temp U., Eriksson K.E.L. The Ligninolytic System of the White Rot Fungus Pycnoporus cinnabarinus: Purification and Characterization of the Laccase. Applied and Environmental Microbiology 62(4): 1151–1158 (1996).
47. Evtuguin, D.V., Pascoal Neto, C. New polyoxometalate promoted method of oxygen delignification. Holzforschung 51(4): 338-342 (1997).
48. Evtuguin, D.V., Pascoal Neto, C., Pedrosa de Jesus, J.D. Bleaching of kraft pulp by oxygen in the presence of polyoxometalates. Journal of Pulp Paper Science 24(4): 133-140 (1998).
49. Fabbrini, M., Galli, C., Gentili, P. Comparing the Catalytic Efficiency of Some Mediators of Laccase Journal of Molecular Catalysis B: Enzymatic 16: 231-240 (2002).
50. Fang, Q.H., Zhong, J.J. Submerged fermentation of higher fungus Ganoderma lucidum for production of valuable bioactive metabolites—ganoderic acid and polysaccharide. Biochemical Engineering Journal 10: 61-65 (2002).
51. Fernandez, J.L. Theoretical Study of the Basicity and the Redox Properties of Heteropolyanions. Tese de Doutoramento, Departament de Química Física i Inorgànica de la Universitat Rovira i Virgili, Tarragona, (2003).
52. Fukuda, T., Uchida, H., Takashima, Y., Uwajima, T., Kawabata, T., Suzuki, M. Degradation of isphenol A by puried laccase from Trametes villosa. Biochemical and Biophysical Research Communication 284: 704– 706 (2001).
53. Furtado, F.P., Evtuguin, D.V., Gomes, T.M. Pulp & Paper Canada 102 (2001).
54. Galhaup, C., Haltrich, D. Enhanced formation of laccase activity by the white-rot fungus Trametes pubescens in the presence of copper. Applied and Microbiology Biotechnology 56: 225–232 (2001).
55. Galhaup, C., Goller, S. Peterbauer, C. K. Strauss, J. Haltrich, D.Characterization of the major laccase isoenzyme from Trametes pubescens and regulation of its synthesis by metal ions, Microbiology 148: 2159-2169 (2002a).
56. Galhaup, C., Wagner, H., Hinterstoisser, B. Haltrich, D. Increased production of laccase by the wood-degrading basidiomycete Trametes pubescens. Enzyme Microbiology and Technology 30: 529-536 (2002b).
57. Gamelas, J., Cavaleiro, A., Santos, I., Balula, M. Os polioxometalatos. Do anião de keggin às nanocápsulas. Boletim da Sociedade Portuguesa de Química, 90: 45-51 (2003).
58. Gamelas, J., Gaspar, A., Evtuguin, D.V., Pascoal Neto, C. Transition metal substituted polyoxotungstates for the oxygen delignification of kraft pulp. Applied Catalysis A: General 295 (2): 97-230 (2005).
154

Bibliografia
59. Garg, S.K., Modi, D.R. Decolorization of pulp-paper mill effluents by white-rot fungi. Critical Reviews in Biotechnology 19 (2): 85-112 (1999).
60. Garzillo, A.M.V., Colao, M.C., Caruso, C., Caporale, C., Celleti, D., Buonocore, V. Laccase from the white-rot fungus Trametes trogii. Applied Microbiology and Biotechnology 49: 545–551 (1998).
61. Gaspar, A., Evtuguin, D.V., Pascoal Neto, C. Oxygen bleaching of kraft pulp catalysed by Mn(III)-substituted polyoxometalates. Applied Catalysis A: General 239: 157–168 (2003).
62. Gelo, P.M., Kim, H., Butlin, N., Palmore, G. Electrochemical Studies of a Truncated Laccase Produced in Pichia pastoris. Applied and Environmental Microbiology, 65(12): 5515-5521 (1999).
63. Giardina, P., Palmieri, G., Scaloni, A., Fontanella, B., Faraco, V., Cennamo, G., Sannia, G. Protein and gene structure of a blue laccase from Pleurotus ostreatus. Biochemical Journal 341:655-663 (1999).
64. Gouveia, M. em http://www.gee.min-economia.pt/resources/docs/publicacao/Imprensa/ 20000601.pdf (2005).
65. Grigoriev, V., Hill, C. L., Weinstock, I. Role of cation size in the energy of electron-transfer to 1:1 polyoxometalate ion pairs {(M+)(Xn+VW11O40)}(8-n)- (M = Li, Na, K). Journal of American Chemical Society 122: 3544-3545 (2000).
66. Gutiérrez, A., Prieto, A., Martínez, A.T. Structural characterization of extracellular polysaccharides produced by fungi from the genus Pleurotus. Carbohydrate Research 281: 143-154 (1996).
67. Hammel, K.E. 2 Fungal Degradation of Lignin, Cab International Driven by Nature: Plant Litter Quality and Decomposition, pp. 33-45 (1997).
68. Harmalker, S. P., Pope, M. T. Heteropolyanions in nonpolar solvents as quasi-metalloporphyrins: slow proton transfer and kinetic stabilization of hydroxovanadium(IV). Journal of Inorganic Biochemistry 28: 85-95 (1986).
69. Harris, E.L.V., Angal, S. (1989) Protein purification methods: a practical approach, Oxford University Press.
70. Hatakka, A. Lignin-modifying enzymes from selected white-rot fungi: production and role in lignin degradation. FEMS Microbiology Reviews 13:125-135 (1994).
71. Hergert, H.L., Infrared spectra, in lignins: occurrance, formation, and reactions, (K.V. Sarkanen and C.H.Ludwigg, Eds) Chap. 7, Wiley intercience, New York. (1971).
72. Higuchi, T. Biodegradation mechanisms of lignin by white rot Bbasidiomycetes. Journal of Biotechnology 30:1-8 (1993).
73. Hill, C.L. Introduction: Polyoxometalatess multicomponent molecular vehicles to probe fundamental issues and practical problems. Chemical Reviews 98:1-2 (1998).
74. Hofrichter, M. Review: lignin conversion by manganese peroxidase (MnP). Enzyme Microbiology and Technology 30: 454-466 (2002).
155

Bibliografia
75. Humar., M. Petrič, M., Pohleven, F. Changes of the pH value of impregnated wood during exposure to wood-rotting fungi. Holz als Roh- und Werkstoff 59:288 – 293 (2001).
76. Ikasari, L., Michell, D. Two-Phase Model of the Kinetics of Growth of Rhizopus oligosporus in Membrane Culture. Biotechnology and Bioengineering 68(6): 619-627 (2000).
77. Jang, M., Ryu, W., Cho, M. Laccase production from repeated batch cultures using free mycelia of Trametes sp. Enzyme and Microbial Technology 30:741–746 (2002).
78. Jeannin, Y. P. The Nomenclature of Polyoxometalates: How To Connect a Name and a Structure. Chemical Reviews 98:51-76 (1998).
79. JHS Natural Products, disponível em http://www.jhsnp.com/store/vpscoriolusversicolor. html (consultado em fev. 2005).
80. Jönsson, L.J., Saloheimo M., Penttilä M., Laccase from the white-rot fungus Trametes versicolor: cDNA cloning of lcc1 and expression in Pichia pastori. Current Genetics 32: 425–430 (1997).
81. Karnezis, T., McIntosh, M., Wardak, A. Z., Stanisich, V.A., Stone, B.A. The Biosynthesis of b-Glycans. Trends in Glycoscience and Glycotechnology 12(66): 211–227 (2000).
82. Katayama, Y., Nishida, T., Morohoshi N., Kuroda, K. The metabolism of biphenyl structures in lignin by the wood-rotting fungus Coriolus versicolor. FEMS Microbiology Letters 61: 307-314 (1989).
83. Katsoulis, D.E. A Survey of Applications of Polyoxometalates. Chemical Reviews 98:359-387 (1998).
84. Kersten, P.J., Kalyanaraman, B., Hammel, K.E., Reinhammar, B., Kirk T.K. Comparison of lignin peroxidase, horseradish peroxidase and laccase in the oxidation of methoxybenzenes. Biochemical Journal 268:475–480 (1990).
85. Khondkar, P. Aidoo, K.E. Tester, R.F. Sugar profile of extracellular polysaccharides from different Tremella species. International Journal of Food Microbiology 79: 121-129 (2002).
86. Kilpelainen, I., Sipila, J., Brunow, G., Lundquist, K., Ede, R. Application of Two-Dimensional NMR Spectroscopy to Wood Lignin Structure Determination and Identification of Some Minor Structural Units of Hard- and Softwood Lignins. Journal of Agricultural and Food Chemistry 42: 2790-2794 (1994).
87. Kim, S.W., Hwang, H.J., Park, J.P, Cho, Y.J., Song, C.H., Yun, J.W. Myelial growth and exo-biopolymer production by submerged culture of various edible mushrooms under different media. Letters in Applied Microbiology 34: 56-61 (2002a).
88. Kim S.W., Hwang H.J., Xu, C.P., Na, Y.S., Song, S.K., Yun, J.W. Influence of nutritional conditions on the mycelial growth and exopolysaccharide production in Paecilomyces sinclairii. Letters in Applied Microbiology 34(6):389-93 (2002b).
156

Bibliografia
89. Kirk, T, Cullen, D. Enzymology and molecular genetics of wood degradation by white rot fungi In Young, R. A., Atkhar, M., (eds.) Environmentally Friendly Technologies for Pulp and Paper Industry, John Wiley & Sons, Inc (1998).
90. Kirk, T., Farrell, R. Enzimatic "combustion": the microbial degradation of lignin. Annual Review of Microbiology 41:465-505 (1987).
91. Kossen, N.W.F. The morphology of filamentous fungi. Advances in Biochemical Engineering and Biotechnology 70:1– 33 (2000).
92. Kozhevnikov, I.V. Catalysis by heteropoly acids and multicomponent polyoxometalates in liquid-phase reactions. Chemical Reviews 98:171-198 (1998).
93. Kuhad, R.C., Singh, A. Eriksson, K. Microorganisms and enzymes involved in the degradation of plant fiber cell walls in biotechnology in pulp and paper industry, Scheper, T., Springer.
94. Larsson, S., Cassland, P., Jönsson L.J. Development of a Saccharomyces cerevisiae Strain
with Enhanced Resistance to Phenolic Fermentation Inhibitors in Lignocellulose Hydrolysates by Heterologous Expression of Laccase. Applied and Environmental Microbiology. 67: 1163-1170 (2001).
95. Lee, I.Y., Jung, K.H., Lee, C.H., Park, Y.H. Enhanced production of laccase in Trametes versicolor by the addition of ethanol. Biotechnology Letters 21: 965-969 (1999).
96. Leitner, C., Hess J, Galhaup C, Ludwig R, Nidetzky B, Kulbe KD, Haltrich D. Purification and characterization of a laccase from the white-rot fungus Trametes multicolor. Applied Biochemistry and Biotechnology (98-100):497-507 (2002).
97. Leonowicz, A., Cho, N.S., Luterek, J., Wilkolazka, A., Wojtas-Wasilewska, M., Matuszewska, A., Hofrichter, M., Wesenberg, D., Rogalski, J. Fungal laccase: properties and activity on lignin. Journal of Basic Microbiology 41(3-4):185-227 (2001).
98. Leontievsky, A.A., Myasoedova, N.M., Baskunov, B.P., Evans, C.S., Golovleva, L.A. Transformation of 2,4,6-trichlorophenol by the white rot fungi Panus tigrinus and Coriolus versicolor. Biodegradation 11:331–340 (2000).
99. Leparulo-Loftus, M. Pope, M. T. Vanadium-51 NMR spectroscopy of tungstovanadate polyanions. Chemical shift and line-width patterns for the identification of stereoisomers. Inorganic Chemistry 26: 2112-2120 (1987).
100. Lewin, M., Goldstein I. Wood structure and composition, Marcel Dekker Inc. (1991).
101. Li, K., Helmã, R.F., Eriksson, K.E. Mechanistic studies of the oxidation of a non-phenolic lignin model compound by the laccase/1-hydroxybenzotriazole redox system. Biotechnology Applied Biochemistry 27: 239-243 (1998).
102. Li, K., Xu, F., Eriksson, K.E. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Applied and Environmental Microbiology 65:2654–2660 (1999).
157

Bibliografia
103. Luedeking, R. Piret, E.L. A kinetic study of the lactic acid fermentation. Batch process at controlled pH. Biotechnology and Bioengineering 67(6):393-401 (2000).
104. Majcherczyk, A., Johannes, C., Hüttermann, A. Oxidation of polycyclic aromatic hydrocarbons (PAH) by laccase of Trametes versicolor. Enzyme Microbiology and Biotechnology 22:335-341 (1998).
105. Mandels M, Andreotti, R., Roche, C. Measurement of saccharifying cellulase. Biotechnology Bioengineering Symposium 6:21-33 (1976).
106. Mansur, M., Suarez, T., Fernandez-Larrea, J. B., Brizuela, M. A. & Gonzalez, A. E. Identification of a laccase gene family in the new lignin-degrading basidiomycete CECT 20197. Applied and Environmental Microbiology 63:2637–2646 (1997).
107. Martinez-Inigo, M.J., Kurek, B. Oxidative Degradation of Alkali Wheat Straw Lignin by Fungal Lignin Peroxidase, Manganese Peroxidase and Laccase: A Comparative Study. Holzforschung 51:543-548 (1997).
108. Mayer, A.M., Staples, R.C. Laccase: new functions for an old enzyme. Phytochemistry 60:551-565 (2002).
109. MBendi, disponível em http://www.mbendi.co.za/indy/pulp/p0005.htm (consultado em fev. 2005).
110. McDonald, R.G. (Ed.) Pulp and paper manufacture. Vol. II, Chap.4, McGraw-Hill, New, York. (1970).
111. Melo, M., Gouveia, M. Pasta e Papel em Portugal. disponível em http://www.gee.min-economia.pt/resources/docs/publicacao/DT/dt37.pdf. (2005)
112. Messer, E., Srebotnik, E., Fiechter, A. Editorial. Journal of Biotechnology 53: 91–92 (1997).
113. Miyazaki, T., Yadomae, T., Sgiura, M., Ito, H., Fujii, K., Naruse, S. & Kunihisa, M. Chemical Structure of Antitumor Polysaccharide, Coriolan, produced by Coriolus versicolor. Chemical & Pharmaceutical Bulletin 22, 1739-1742 (1974).
114. Milagres, A.M.F., Arantes V., Medeiros C.L., Machuca A. Production of metal chelating compounds by white and brown-rot fungi and their comparative abilities for pulp bleaching. Enzyme and Microbial Technology 30:562–565 (2002).
115. Miller, G.L. Use of dinitrosalicilic acid reagent for determination of reducing sugar. Analytical Chemistry 31:426-431 (1959).
116. Mitchell, D.A., Meien, O.F., Krieger, N., Dalsenter, F.D.H. A review of recent developments in modeling of microbial growth kinetics and intraparticle phenomena in solid-state fermentation. Biochemical Engineering Journal 17:15-26 (2004).
117. Mizuno, T., Wang, G., Zhang, J., Kawagishi, H., Nishitoba, T., Li, J. Reishi, Ganoderma lucidum and Ganoderma tsugae: Bioactive substances and medicinal effects. Food Reviews International 11: 151-166 (1995).
158

Bibliografia
118. Modi, D.R., Chandra, H., Garg, S.K., Decolourization of bagasse-based paper mill effluent by the white-rot fungus Trametes versicolor. Bioresource Technology 66:79-81 (1998).
119. Moreira, M.T., Palma, C., Feijoo, G., Lema. J.M. Strategies for the continuous production of lignolytic enzymes in fixed and fluidised bed bioreactors. Journal of Biotechnology, 66:27-39 (1998).
120. Mougin, C., Boyer, F.D., Caminade, E., Rama, R. Cleavage of the diketonitrile derivative of the herbicide isoxa.utole by extracellular fungal oxidases. Journal of Agricultural and Food Chemistry 48: 4529–4534 (2000).
121. Mougin, C., Jolivalt C., Briozzo P.,·Madzak, C. Fungal laccases: from structure-activity studies Environmental Chemistry Letter 1:145–148 (2003).
122. Muñoz, C., Guillen, F, Martinez, A.T., Martinez, M.J. Laccase isoenzymes of Pleurotus eryngii: characterization, catalytic properties, and participation in activation of molecular oxygen and Mn2+ oxidation Applied and Environmental Microbiology 63: 2166-2174 (1997).
123. Neto, B.B., Scarmino, I., Bruns, R. Planejamento e optimização de experimentos, Editora da Unicamp, Campinas, SP, ISBN 85-268-0336-0 (1995).
124. Ng, T.B. A review of research on the protein-bound polysaccharide (polysaccharopeptide, PSP) from the mushroom Coriolus versicolor (Basidiomycetes: Polyporaceae). General Pharmacological 30 (1), 1-4 (1998).
125. Niku, P.M.L., Viikari, L. Enzymatic oxidation of alkenes. Journal of Molecular Catalysis B Enzymatic 10:435–444 (2000).
126. Nosal’ova, V., Bobek, P., Cerna, S., Galbavy, S., Stvrtina, S. Effects of pleuran (beta-glucan isolated from Pleurotus ostreatus) on experimental colitis in rats. Physiological Research 50(6):575-581 (2001).
127. Nyanhongo, G.S., Gomes, J., Gubitz G., Zvauya R., Read, J.S., Steiner, W. Production of laccase by a newly isolated strain of Trametes modesta. Bioresource Technology 84:259–263 (2002).
128. O’Callaghan, J., O’Brien M.M., McClean K., Dobson, A.D.W. Optimisation of the expression of a Trametes versicolor laccase gene in Pichia pastoris. Journal of Industrial Microbiology & Biotechnology 29:55–59 (2002).
129. Osiadacz, J., Al-Adhami, J.H., Bajraszewska, D., Fischer, P., Peczyñska-Czoch, W. On the use of Trametes versicolor laccase for the conversion of 4-methyl-3-hydroxyanthranilic acid to actinocin chromophore. Jounal of Biotechnology 72:141-149 (1999).
130. Paice, M.G., Reid, I.D., Bourbonnais, R., Archibald, F. S., Jurasek, L. Manganese peroxidase, produced by Trametes versicolor during pulpbleaching, demethylates and deligninfies kraft pulp. Applied and Environmental Microbiology 59 (1): 260-265 (1993).
131. Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnology Advances 22:189–259 (2004).
159

Bibliografia
132. Paulsen, B.S. Biologically active polysaccharides as possible lead compounds. Phytochemistry Reviews 1:379–387 (2002).
133. Park, J.P., Kim, Y.M., Kim, S.W., Hwang, H.J., Cho, Y.J., Lee, Y.S., Song, C.H. and Yun, J.W. Effect of agitation intensity on the exo-biopolymer production and mycelial morphology in Cordyceps militaris. Letters in Applied Microbiology 34: 433–438 (2002).
134. Pereira, N. P. Biotecnologia e Agricultura. Universidade Aberta. Lisboa. p. 101. (1996) ISBN: 972-674-182-3.
135. Piontek, K., Antorini, M., Choinowski, T. Crystal structure of a laccase from the fungus Trametes versicolor at 1.90-Å resolution containing a full complement of coppers. The journal of biological chemistry 277:40: 37663–37669 (2002).
136. Pointing, S.B. Feasibility of bioremediation by white-rot fungi. Applied Microbiology Biotechnology 57: 20-33 (2001).
137. Pope, M. T. Heteropoly and Isopoly Oxometalates, Springer-Verlag, Berlin. (1983).
138. Rancaño, G., Lorenzo, M., Molares, N., Rodriguez, S., Sanromán., M. Production of laccase by Trametes versicolor in an airlift fermentor. Process Biochemistry 39: 467-473 (2003).
139. Rajarathnam, S., Shashireka, M.N., Bano, Z. Biopotentialities of the Basidiomacromycetes. Advances in Applied Microbiology 37: 233-361 (1992).
140. Read, N.D. Cellular nature and multicellular morphogenesis of higher fungi In Ingran, D., Hudson, A. (eds.) Shape and form in plants and fungi, The linnean Society of London, Academic Press, Cap.15:251-269 (1994).
141. Reid, I., Paice, M., Ho, C., Jurasek, L. Biological bleaching of softwood kraft pulp with the fungus Trametes (Coriolus) versicolor. Tappi Journal 149-153 (1990).~
142. Ronne, H. Glucose repression in fungi. Trend in Genetics 1:12-17 (1995).
143. Rosado, F.R., Germano, S., Carbonero, E.R., Da Costa, S.M., Iacomini, M., Kemmelmeier, C. Biomass and exopysaccharide production in submerged cultures of Pleurotus ostreatoroseus Sing. and Pleurotus ostreatus “florida” (Jack.:Fr.) Kummer. Journal of Basic Microbiology 43 (3): 230-237 (2003).
144. Roy, B. Archibald, F. Effects of kraft Pulp and Lignin on Trametes versicolor Carbon Metabolism. Applied and Environmental Microbiology 59(6):1855-1863 (1993).
145. Ruel, K., Zhang, H., Niku-Paavola, M., Soloheimo, M., Moukha, S., Joseleau, J.P. In situ hybridization and immunocytochemistry in electron microscopy to study the expression of lignolytic enzymes in fungi growing in wood, 10th International Symposium on Wood and Pulping Chemistry, Yokohama, Japão, 1:534-538 (1999).
146. Sadakane, M., Steckhan, E. Investigation of the manganese-substituted α-Keggin-heteropolyanion K6SiW11O39Mn(H2O) by cyclic voltammetry and its application as oxidation catalyst. Journal of Molecular Catalysis A: Chemical 114: 221-228 (1996).
160

Bibliografia
147. Sadakane, M., Steckhan, E. Electrochemical properties of polyoxometalates as electrocatalysts Chemical Reviews 98:219-237 (1998).
148. Sadakane, M., Steckhan, E., Co-solvent Effects on the Redox Potential of Manganese-substituted -a-Keggin type Silicon Polyoxotungstate K6SiW11O39Mn(H2O): First Electrochemical Generation of the Manganese (V) Redox System in Aqueous Environment. Acta Chemical Scandinavica 53: 837 (1999).
149. Schutyser, M.A.I., Weber, F.J., Briels, W.J., Rinzema, A., Boom, R.M. Heat and water transfer in a rotating drum containing solid substrate particles. Biotechnology and Bioengineering 82 (5): 552-563 (2003).
150. Servili, M., DeStefano, G., Piacquadio, P., Sciancalepore, V. A novel method for removal phenols from grape must. American Journal of Enology and Viticulture. 51:357-361 (2000).
151. Shatalov, A.A., Evtuguin, D., Pascoal Neto, C. Cellulose degradation in the reaction system O2/heteropolyanions of series [PMo(12−n)VnO40](3+n)−. Carbohydrate Polymers 43:23-32 (2000).
152. Shingo, K., Masanori, A., Noriko, O., KyoKo, O., Takashi, I., Hideo, O. Degradation of a non-phenolic B-O-4 substructure and of polymeric lignin model compounds by laccase of Coriolus versicolor in the presence of 1-hydroxybenzotriazole. FEMES Microbiology Letters 170: 51-57 (1999).
153. Shreve, R.N., Brink, J.A. Indústrias de processos químicos. 4ª Ed. Editora Guanabara Koogan, 498-499. (1977)
154. Sigoillot, J., Petit, M., Ruel, K., Moukha, S., Comtat, J., Laugero, C., Joseleau, J:, Choudens, C., Asther, M. Enzymatic treatment with manganese peroxidadse from Phanerochatae chrysosporium for enhancing whear straw pulp characteristics Holzforschung, 51:549-556 (1997).
155. Sinha, J., Bae, J.T., Park, J.P., Kim, K.H., Song, C.H., Yun, J.W. Changes in morphology of Paecilomyces japonica and their effect on broth rheology during production of exo-biopolymers. Applied Microbiology and Biotechnology 56: 88-92 (2001a).
156. Sinha, J., Bae, J.T., Park, J.P., Song, C.H., Yun, J.W. Effect of substrate concentration on broth rheology and fungal morphology during exo-biopolymer production by Paecilomyces japonica in a bioreactor. Enzyme Microbiology and Technology 29: 392-399 (2001b).
157. Sipilä, J., Karhunen, P., Brunow G. em http://www.helsinki.fi/~orgkm_ww/Lignin.html, (2005).
158. Sjöström, E., Wood chemistry: Fundamentals and application, Academic Press Inc., Cap. 1, 3 e 4, (1981).
159. Smith, M., Ragauskas, A.J. em http://www.eere.energy.gov/industry/forest/pdfs/ biobleaching.pdf (2005).
161

Bibliografia
160. Smook, G.A. Thechical association of the pulp and paper industry (TAPPI), test method (T236) (30). In: Handbook for Pulping & Paper Technology (M.J. Kocurek, Tech.. Ed,.), 7th printing. TAPPI, Technology Park, Atlanta, Georgia. (1989).
161. Solomon, E. I., Sundaram, U. M. and Machonkin, T. E. Multicopper oxidases and oxygenases. Chemical Review 96: 2563-2605 (1996).
162. Souza, C. G. M., Zilly, A., Peralta, R. M. Production of laccase as the sole phenoloxidase by a Brazilian strain of Pleurotus pulmonarius in solid state fermentation. Journal of Basic Microbiology 42: 83–90 (2002).
163. Sugiura, M., Ohno, H., Kunihisa, M., Hirata, F., Ito, H. Studies on Antitumor Polysaccharides, Especially D-II, from Mycelium of C. versicolor. Japanese Journal of Pharmacology 30: 503-513 (1980).
164. Sutherland, I.W. Biosynthesis and composition of gram-negative bacterial extracellular and wall polysaccharides. Annual Review in Microbiology 39: 243-270 (1985).
165. Swamy, J., Ramsay, A. The evaluation of white rot fungi in the decolouration of textile dyes. Enzyme Microbiology and Technology 24:130-137 (1999).
166. Tanaka, H., Itakura, S., Enoki, A. Hydroxyl radical generation by an extracellular low-molecular-weight substance and phenol oxidase activity during wood degradation by the white-rot basidiomycete Trametes versicolor. Journal of Biotechnology 75(1):57-70 (1999).
167. Takamatsu, T., Shioya, S, Okuda, K. A comparison of unstructured growth models of micro-organisms. Journal of Fermentation Technology. 59 (2): 131-136 (1981).
168. Teeri, T. Crystalline cellulose degradation: new insight into the function of cellobiohydrolases. Trends in Biotechnology 15: 160-167 (1997).
169. Tien, M., Kirk, T.K. Lignin Peroxidase of Phanerochaete chrysosporium. Methods in Enzymology 161:238-247 (1988).
170. Thurston, C.F. The structure and function of fungal laccases. Microbiology 140:19-26 (1994).
171. Tolan, J.S., Thibault, L., Decreasing ECF bleaching costs with enzymes in a mill with oxygen delignification. Pulp Paper Can. 98 (12): 147-150 (1997).
172. Tourné, C.M., Tourné, G.F., Malik, S.A., Weakley, T.J.R. J. Triheteropolyanions containing copper(II), manganese (II), or manganese (III). Inorganic Nuclear Chemistry 32: 3875-3881 (1970).
173. Trupkin, S., Levin, L., Forchiassin, F., Viale, A. Optimization of a culture medium for ligninolytic enzyme production and synthetic dye decolorization using response surface methodology. Journal of Industrial Microbiology and Biotechnology 30:682–690 (2003).
174. Urzúa, U., Kersten, P.J., Vicuña, R. manganese peroxidase-dependent oxidation of glyoxylic and oxalic acids synthesized by Ceriporiopsis subvermispora produces
162

Bibliografia
extracellular hydrogen peroxide Applied and Environmental Microbiology, 64(1): 68–73 (1998).
175. Viikari, L., Lantto R. Biotechnology in the pulp and paper industry, Progress in Biotechnology, Elsevier Science B.V. vol. 21 p.v (2002)
176. Voet, D., Voet, J. Biochemistry, John Wiley and Sons, New York, pp.254-257 (1990).
177. Xu, F., Shin, W., Brown, SH., Wahleithner, J.A., Sundaram, U.M., Solomon, E.I. A study of a series of recombinant fungal laccases and bilirubin oxidase that exhibit significant differences in redox potential, substrate specificity, and stability Biochimica et Biophysica Acta 1292: 303-311 (1996).
178. Zamora, P., Pereira, C., Tiburtius, E.,. Moraes, S., Rosa, M., Minussi R., Durán, N. Decolorization of reactive dyes by immobilized laccase. Applied Catalysis B: Environmental 42(2):131-144 (2003).
179. Zing, R., Fletcher, I.J., Freierrmuth, B., Huber, K. Method for enhancing the activity of an enzyme, compounds showing enzyme activity enhancement, and detergents containing such compounds PCT. World patent application, WO 97/06224. February 1997.
180. Zhang, X., Pope, M.T., Chance, M., Jameson, G. High-valent manganese in polyoxotungstates. 1. Manganese (IV) Keggin derivatives. Polyhedron 14: 1381-1392 (1995).
181. Znidarsic, P., Pavko, A. Morphology of Filamentous Fungi. Food Technology and Biotechnology. 39 (3): 237–252 (2001).
182. Yadav, J.S., Tripathi, J.P. Optimization of cultivation and nutrition conditions and substrate pretreatment for solid substrate fermentation of wheat straw by Coriolus versicolor. Folia Microbiology 36:249– 301 (1991).
183. Yesilada, O., Asma, D., Cing, S. Decolorization of textile dyes by fungal pellets. Process Biochemistry 38: 933-/938 (2003).
184. Yaropolov, A.I., Skorobogatko, O.V., Vartanov, S.S., Varfolomeyev, S.D. Laccase Properties, catalytic mechanism and applicability. Applied Biochemistry and Biotechnology 49(3): 257-280 (1994).
185. Waites, M.J., Morgan, N.L., Rockey, J.S. & Hi Gton A.G. Industrial Microbiology, An Introduction. pp. 68. Blackwell Publishing, Oxford; ISBN 0-632-05307-0 (2001).
186. Weinstock, I. A.; Atalla, R. H.; Reiner, R. S.; Moen, M. A.; Hammel, K. E.; Houtman, C. J.; Hill, C. L.; Harrup, M. K. A new environmentally benign technology for transforming wood pulp into paper. Engineering polyoxometalates as catalysts for multiple processes. Journal of Molecular Catalysis A: Chemical 116: 59-84 (1997).
187. Whistler, R.L., BeMiller, J.N. Carbohydrate Chemistry for Food Scientists. pp.96. Eagan Press. St. Paul ISBN 0-913250-92-9 (1997).
188. Wong, Y., Yu, J. Laccase-catalyzed decolorization of synthetic dyes. Water Research 33(16): 3512-3520 (1999).
163

Anexos
ANEXOS
A1. Curva de calibração para o método de DNS de determinação de açúcares
redutores
Abs = 0,4341 Gluc - 0,0115R2 = 0,9995
0
0,1
0,2
0,3
0,4
0,5
0 0,2 0,4 0,6 0,8 1 1,2
Glucose (g/L)
Abs
i
Abs
orvâ
ncia
164

Anexos
A2. Curva de calibração para o método de Lowry de determinação de proteína total
Ptn = 0,7835 Abs - 0,177R2 = 0,9933
0
0,2
0,4
0,6
0,8
1
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6
Absorvância
Prot
eína
tota
l (m
g/m
L)iii
iiiiii
Prot
eína
tota
l (m
g/m
L)
165

Anexos
A3. Curva de calibração do GPC para a determinação do peso molecular médio do
EPS
Curva de calibração com pululana
Log Mp= -0,5864Vr + 13,614R2 = 0,9994
2
2,5
3
3,5
4
4,5
5
5,5
13 14 15 16 17 18 19 20
Vr, ml
RI r
esp
Res
post
a -
Inte
nsid
ade
R I/
mm
166

Anexos
A4. Curva de calibração para a determinação dos ácidos hexourónicos
Abs = 0,0091 Ác. Galac + 0,0054R2 = 0,9967
0,00
0,10
0,20
0,30
0,40
0,50
0 10 20 30 40 50
Ácido D-galacturónico (µg)
Abs
orvâ
ncia
iiii
Abs
orvâ
ncia
167

LISTA DE PUBLICAÇÕES
A.P.M. Tavares, M.A.Z. Coelho, J.A.P. Coutinho, A.M.R.B. Xavier. Optimization and modelling of laccase production by Trametes versicolor in bioreactor using statistical experimental design, Applied Biochemistry and Biotechnology, in press (2006). A.P.M. Tavares, M.S.M. Agapito, M.A.Z. Coelho, J. A. Lopes da Silva, A. Barros-Timmons, J.A.P. Coutinho, A.M.R.B. Xavier. Selection and optimization of culture medium for exopolysaccharide production by Coriolus (Trametes) versicolor, World Journal of Microbiology and Biotechnology, 21 (8-9) 1499-1507 (2005). A.P.M. Tavares, M.A.Z. Coelho, J.A.P. Coutinho, A.M.R.B. Xavier. Laccase improvement in submerged cultivation: induced production and kinetic modelling. Journal of Chemical Technology and Biotechnology, 80 (6) 669-676 (2005). J.A.F. Gamelas, A.P.M. Tavares, D.V. Evtuguin, A.M.B. Xavier. Oxygen bleaching of kraft pulp with polyoxometalates and laccase applying multi-stage process: comparative study. Journal of Molecular Catalysis B: Enzymatic, 33: 57-64 (2005). P.F.F. Amaral, D.L.A. Fernandes, A.P.M. Tavares, A.M.R.B. Xavier, M.C. Cammarota, J.A.P. Coutinho, M.A.Z. Coelho. Dye decolorization performed by Trametes versicolor from textile wastewater. Environmental Technology, 25: 1313-1320 (2004).
A.P.M. Tavares, J.A.F. Gamelas, A. Gaspar, D.V. Evtuguin, A.M.R.B. Xavier. A novel approach for the oxidative catalysis employing polyoxometalate-laccase system: application to the oxygen bleaching of kraft pulp. Catalysis Communications, 5 485-489 (2004).
168





























