Embed Size (px)
Citation preview

1
Análisis molecular de la diversidad bacterina y de genes que participan en la
nitrificación y desnitrificación en la rizosfera de plantas que crecen en suelos
contaminados con metales pesados
Registro SIP: 20080628
Responsable: Dra. Janet Jan Roblero
RESUMEN
La contaminación por metales pesados en el suelo es un problema ambiental y tiene un
impacto negativo en la salud humana y en la agricultura. Para recuperar estos sitios, se
sugiere una cobertura vegetal con especies de plantas endémicas para estabilizar los
contaminantes y reducir su toxicidad. La rizosfera, como una interfase importante entre el
suelo y la planta, juega un papel muy importante en la fitorremediación de suelos
contaminados con metales pesados, en los cuales, se sabe que la comunidad microbiana
afecta la movilidad y biodisponibilidad de los metales para la planta. Además, estos suelos
generalmente se caracterizan por tener bajos niveles de N, P y Ca.
En este trabajo se exploró la diversidad bacteriana y de genes relacionados con el ciclo
del nitrógeno, particularmente nitrificación y desnitrificación, en la rizosfera de plantas que
crecen en jales de minas de Zacatecas, empleando técnicas moleculares y de cultivo. Se
construyeron librerías de genes ribosomales a partir de DNA de la rizosfera de Viguiera
linearis y Bahia xylopoda. La diversidad bacteriana deducida del análisis DGGE reveló
diferencias entre los perfiles de las rizosferas de las plantas y del suelo no rizosférico. Se
detectaron genes moduladores del ciclo del nitrógeno: nifH, nifD, amoA, nirK, nirS y nosZ;
y 16S rRNA de miembros de Nitrobacter. Únicamente en la rizosfera de Chlorix virgata se
detectó el ciclo completo (fijación, nitrificación y desnitrificación). Los genes que se
encontraron en prácticamente todas las rizosferas fueron los de la oxidación del amonio y
la oxidación del nitrito. El análisis de las secuencias de los genes amoA obtenidos de la
rizosfera de V. linearis se agruparon con secuencias de amoA de Nitrosospira multiformis,
Nitrosospira tenuis y Nitrovibrio tenuis. Este estudio nos da un panorama general de la
comunidad bacteriana en suelos contaminados con metales pesados y de algunas de sus
funciones potenciales en este ambiente.

2
INTRODUCCION
Contaminación de suelos por metales pesados
Los metales pesados en su estado de oxidación cero se caracterizan por tener
propiedades como la: ductibilidad y conductividad, mientras que como cationes forman
sales y cristales con propiedades químicas y físicoquímicas diversas. Los metales son
componentes naturales en el suelo y algunos de ellos son requeridos por las plantas
como micronutrientes o sólo son utilizados por un grupo limitado de microorganismos. Sin
embargo, hay metales que no tienen una función biológica específica y cuando superan
cierta concentración en el ambiente son clasificados como elementos potencialmente
tóxicos (EPT) (Alloway, 1995). Los metales pesados contaminantes más comunes son el:
Cd, Cr, Cu, Hg, Pb y Ni.
La contaminación de los suelos con metales pesados se aceleró drásticamente desde el
inicio de la revolución industrial, como resultado de actividades humanas tales como la
minería, metalurgia, electrodeposición, producción de energía y combustibles, así como el
uso de fertilizantes, aplicación de pesticidas, riego con aguas negras, generación de
basura doméstica, etc. La contaminación por metales pesados se ha convertido en uno de
los más graves problemas ambientales hoy en día.
Contaminación de suelos con residuos de minas
La minería es una de las actividades primarias de un país, que se ocupa de la extracción
y los procesos de refinamiento de los metales (Adriano, 2001).
México tiene una importante actividad minera, en donde destaca ocupando el segundo
lugar mundial en la producción de plata y como uno de los mayores productores de
bismuto, fluorita, celestita, arsénico, cadmio y grafito. Asimismo es un importante
productor de oro, zinc, cobre, hierro y manganeso. En el periodo de Enero-Agosto de
2006 la producción minero-metalúrgica alcanzó un impacto económico de 37,976 millones
de pesos. Los estados productores principales son: Sonora, Zacatecas, Durango, San
Luis Potosí y Chihuahua (Secretaría de Economía, 2006).
Los procesos de extracción de metales tienen deficiencias en la recuperación de los
elementos principales y acompañantes, por ello es común que los residuos mineros
contengan pequeñas cantidades de otros elementos tóxicos como el As, Bi, Cd, Cu, Mo,
Ni, Se, Cr, Pb (Adriano, 2001). Los materiales con bajas concentraciones de metales se

3
eliminan en forma de lodos. Sin embargo, estos residuos retienen una cierta cantidad de
metales pesados potencialmente tóxicos. Estos lodos son depositados usualmente a cielo
abierto en pilas que en México se denominan presas de jales.
Desafortunadamente en México no existe una legislación o un organismo que regule el
confinamiento y tratamiento de los residuos de minas de forma adecuada. Sólo existe un
pequeño apartado en el Reglamento de la Ley General del Equilibrio Ecológico y la
Protección al Ambiente en Materia de Residuos Peligrosos que dice: “La disposición final
de desechos peligrosos generados por la minería se efectuará en presas de jales y según
lo dispuesto en las normas técnicas ecológicas correspondientes. Estas presas de jales
podrán ubicarse en el lugar en que se generen dichos residuos, excepto arriba de
poblaciones o de cuerpos receptores ubicados a una distancia menor de 25 kilómetros”.
Estos materiales constituyen un problema de salud pública por la lixiviación de los metales
que contaminan los mantos freáticos que alimentan las fuentes de agua potable, se
introducen en la cadena alimenticia a través de algunos productos agrícolas y se
acumulan en animales para el consumo humano. Por otra parte, los residuos se pueden
dispersar por el viento hasta alcanzar asentamientos humanos (Ongley et al., 2003). Este
problema es especialmente grave en algunas regiones del estado de Zacatecas.
Ciclo del nitrógeno
El nitrógeno es un elemento ubicuo que constituye aproximadamente el 78% de la
composición de gases de la atmósfera de la tierra. Paradójicamente es un recurso
limitante para muchos de los organismos del planeta (Behar et al., 2005). Sus estados de
oxidación oscilan entre -3, en el amoníaco (NH3) y +5, en el nitrato (NO3-). Es el elemento
de mayor demanda para los microorganismos y las plantas, y es el cuarto elemento más
común en las células, alcanzando aproximadamente el 12% del peso seco de una célula.
Este elemento es un componente de los aminoácidos, ácidos nucleicos, aminoazúcares y
sus polímeros respectivos.
La disponibilidad del nitrógeno combinado es un factor limitante para la producción
primaria en muchos ecosistemas. El nitrógeno inorgánico como el amonio, el nitrato y el
nitrito se encuentran como sales muy solubles en el agua; consecuentemente se
distribuye en la ecosfera diluidos en soluciones acuosas, formando pequeños reservorios
de reciclado activo.

4
Las plantas, los animales y la mayoría de los microorganismos necesitan formas de
nitrógeno combinado para incorporarlos a su biomasa celular, pero la capacidad de fijar
nitrógeno atmosférico se restringe a un número limitado de bacterias y arqueas que lo
fijan de manera libre o en simbiosis. Muchos hábitats dependen de las plantas para el
suministro de carbono orgánico que pueda ser usado como fuente de carbono y energía,
pero también dependen de la fijación bacteriana de nitrógeno atmosférico. Las plantas no
podrían continuar con su metabolismo fotosintético si no dispusieran de nitrógeno
combinado de origen procariótico.
La interconversión de los numerosos estados de oxidación del nitrógeno es
predominantemente biológica. Sin embargo, nuestro conocimiento de cómo fluye este
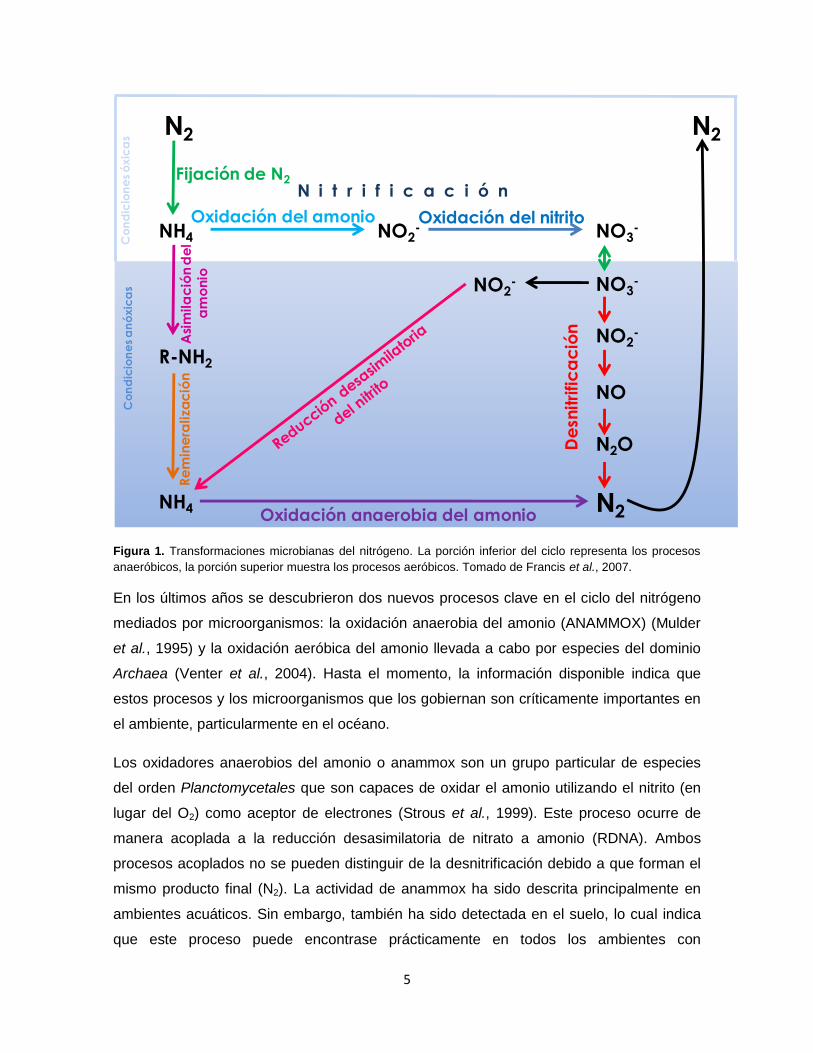
elemento en la Tierra ha cambiado drásticamente en los últimos años. La figura 1 muestra
un esquema general de los procesos que intervienen en el ciclo biogeoquímico del
nitrógeno. Las diversas transformaciones del nitrógeno permiten la circulación del
nitrógeno de la atmósfera a través de los ambientes terrestres y acuáticos. Este
movimiento del nitrógeno a través de la biosfera determina en gran medida la
productividad ecológica de los ambientes terrestres y marinos.
Como se observa en el ciclo del nitrógeno de la Figura 1, el reservorio más grande de
nitrógeno elemental (N2) es la propia atmósfera terrestre, de donde es fijado y reducido a
formas asimilables para otros organismos (NH3). Básicamente, el nitrógeno se mantiene
reducido dentro de los organismos, pero cuando los organismos mueren o lo excretan
puede ser nitrificado a nitrato, que a su vez se puede desnitrificar en condiciones anóxicas
y regresar a la atmósfera en su forma elemental.
A lo largo de este ciclo, los microorganismos participan en prácticamente todos los pasos
del ciclo. Particularmente, en la nitrificación participan dos grupos de bacterias, las
bacterias oxidadoras de amonio (AOB) y las bacterias oxidadoras de nitrito (NOB). La
nitrificación es un proceso aerobio estricto, quimiolitoautotrófico y está restringido a sólo
un pequeño grupo de bacterias dentro de Proteobacteria (Kowalchuck y Stephen, 2001).
Por otro lado, la desnitrificación es principalmente heterótrofa facultativa, ocurre bajo
tensiones muy bajas de oxígeno y está distribuida entre 50 géneros diferentes de
microorganismos que incluye miembros de Bacteria, Archaea y Eukarya (Zumft, 1997;
Risgaard-Petersen et al., 2006).
5
N2
Fijación de N2
NH4 NO2- NO3
-
NO
N2O
NO2-
N2
N2NH4
R-NH2
NO3-NO2
-
Oxidación del amonio Oxidación del nitrito
Oxidación anaerobia del amonio
De
snitri
fic
ac
ión
Asi
mila
ció
n d
el
am
on
io
Re
min
era
liza
ció
n
Co
nd
icio
ne
s ó
xic
as
Co
nd
icio
ne
s a
nó
xic
as
N i t r i f i c a c i ó n
Figura 1. Transformaciones microbianas del nitrógeno. La porción inferior del ciclo representa los procesos
anaeróbicos, la porción superior muestra los procesos aeróbicos. Tomado de Francis et al., 2007.
En los últimos años se descubrieron dos nuevos procesos clave en el ciclo del nitrógeno
mediados por microorganismos: la oxidación anaerobia del amonio (ANAMMOX) (Mulder
et al., 1995) y la oxidación aeróbica del amonio llevada a cabo por especies del dominio
Archaea (Venter et al., 2004). Hasta el momento, la información disponible indica que
estos procesos y los microorganismos que los gobiernan son críticamente importantes en
el ambiente, particularmente en el océano.
Los oxidadores anaerobios del amonio o anammox son un grupo particular de especies
del orden Planctomycetales que son capaces de oxidar el amonio utilizando el nitrito (en
lugar del O2) como aceptor de electrones (Strous et al., 1999). Este proceso ocurre de
manera acoplada a la reducción desasimilatoria de nitrato a amonio (RDNA). Ambos
procesos acoplados no se pueden distinguir de la desnitrificación debido a que forman el
mismo producto final (N2). La actividad de anammox ha sido descrita principalmente en
ambientes acuáticos. Sin embargo, también ha sido detectada en el suelo, lo cual indica
que este proceso puede encontrase prácticamente en todos los ambientes con

6
condiciones anóxicas que contengan nitrógeno inorgánico en diferentes estados de
oxidación (Francis et al., 2007).
Nitrificación
Winogradsky y su grupo en 1890 descubrieron el proceso de nitrificación asociado a un
grupo de bacterias quimiolitoautótrofas que oxidaban el amonio a nitrito (Nitrosomonas) y
otro grupo de bacterias que oxidaban el nitrito a nitrato (Nitrobacter). Incluso actualmente,
la mayoría de los investigadores coinciden en que las especies de Nitrosomonas y
Nitrobacter son las responsables casi enteramente del proceso de nitrificación en suelos.
Hasta ahora no se han encontrado bacterias autótrofas capaces de oxidar el amonio a
nitrato directamente.
Algunas bacterias heterótrofas y ciertos hongos también pueden oxidar formas reducidas
de nitrógeno a nitrito y nitrato, pero mientras que la oxidación de amonio es un proceso
del cual obtienen energía las bacterias autótrofas, la nitrificación en organismos
heterótrofos no contribuye significativamente en la obtención de energía (Tate, 1977). Sin
embargo los microorganismos autótrofos son, por mucho, los principales responsables de
la nitrificación en los ecosistemas naturales.
En los organismos autótrofos oxidadores de amonio, dos enzimas clave son la amonio
monooxigenasa (AMO) y la hidroxilamina óxidoreductasa (HAO). In vivo las dos enzimas
son codependientes una de la otra; AMO cataliza la oxigenación del amoniaco a
hidroxilamina:
NH3 + O2 + H+ + 2e- NH2OH + H2O
Los dos electrones requeridos en este proceso derivan de la oxidación de la hidroxilamina
a nitrito por la HAO:
NH2OH + H2O NO2- + 5H+ + 4e-
La enzima cataliza la oxidación de diferentes sustratos, debido a su función esencial en el
metabolismo energético de las AOB, la enzima probablemente es constitutiva.
Las tres subunidades de la Amo de las AOB autótrofas son codificadas por los genes
amoA, amoB y amoC del operón amo. El gen amoA codifica la subunidad que contiene el
sitio activo de la enzima, dentro de él, la región carboxilo terminal es un blanco adecuado

7
para sondas y oligonucleótidos que permiten distinguir entre AOB de las subclases beta- y
gamma-proteobacteria, así como entre los oxidadores de amonio y metano de la subclase
gamma-proteobacteria (Purkhold et al., 2000).
Desnitrificación
La desnitrificación es parte del aparato bioenergético de la célula bacteriana, dónde los
oxianiones de N como el nitrato, nitrito y los óxidos gaseosos de N como el NO y N2O son
utilizados en lugar del O2 como aceptores en el transporte de electrones de la
fosforilación.
Entre los procariotes la distribución taxonómica de la capacidad para desnitrificar no sigue
ningún patrón, ni está asociada a algún o algunos grupos filogenéticos en particular. La
reacción es realizada por una gran variedad de bacterias y arqueas (halófilos e
hipertermófilos). También se encontró un grupo de enzimas de desnitrificación en las
mitocondrias de algunos hongos y recientemente se descubrió que ocurre una
desnitrificación completa en una foraminífera béntica (Risgaard-Petersen et al., 2006). Es
notable la desnitrificación entre las enterobacterias las cuales reducen el nitrato a nitrito, el
cual posteriormente reducen a amonio (Einsle, 2004).
La mayoría de los desnitrificantes son organismos aerobios heterótrofos que transfieren
equivalentes redox de la oxidación de una fuente de carbono a un óxido de nitrógeno bajo
condiciones anaerobias. Los desnitrificantes autótrofos utilizan compuestos inorgánicos
de azufre, hidrógeno, amonio o nitrato. La oxidación del Fe (II) en algunos
microorganismos también está acoplada a un proceso de desnitrificación completo
(Straub et al., 1996). La desnitrificación consiste en una serie de reacciones secuenciales
en donde primeramente el nitrato es reducido a nitrito, seguido por la reducción del nitrito
a óxido nítrico, el cual es transformado a óxido nitroso y finalmente es reducido a
nitrógeno molecular que es liberado a la atmósfera. Cada una de éstas etapas son
catalizadas por distintas enzimas (Knowles, 1982).
NO3- NO2
- NO N2O N2
Nitrato reductasa Nitrito reductasa Óxido nítrico reductasa Óxido nitroso reductasa
(Nar) (Nir) (Nor) (Nos)

8
La nitrito reductasa es la enzima clave de la desnitrificación ya que cataliza la reacción
que involucra un intermediario gaseoso para que el proceso de desnitrificación llegue a la
formación de nitrógeno molecular. Se conocen dos tipos de estas enzimas, una contiene
cobre (Nir-Cu) y la otra contiene un citocromo hemo (Nir-cd1). Las dos enzimas son
funcional y fisiológicamente similares, aunque excluyentes, es decir no se encuentran las
dos dentro de un mismo organismo (Glockner y Zumft, 1993).
La última etapa del proceso respiratorio desnitrificante consiste en la reducción del óxido
nitroso a nitrógeno molecular, la reacción es catalizada por la óxido nitroso reductasa
(Nos). Se sabe que ésta etapa puede ser considerada como un proceso respiratorio por sí
mismo, ya que muchas bacterias desnitrificantes son capaces de usar el óxido nitroso
como único aceptor de electrones durante la oxidación de la materia orgánica (Zumft,
1997).
METODOS Y MATERIALES
Secuenciación y análisis de las clonas de la librería de genes ribosomales
Se amplificó el inserto del gen 16S rRNA con los iniciadores M13-F (5’-
GTAAAACGACGGCCAG-3’) y M13-R (5’-CAGGAAACAGCTATGAC-3’) a partir del DNA
plasmídico de las clonas representativas. Los amplificados se purificaron con el paquete
de reactivos “DNA Purification Kit” (QIAGEN Gmbh). La secuenciación de los plásmidos
purificados se realizó de manera automatizada con un secuenciador ABI PRISM 310
Genetic Analyzer PE (Applied Biosystems). Se realizaron reacciones de secuenciación
por clona representativa a partir del extremo 5’ y 3’. El análisis de las secuencias
obtenidas se realizó de acuerdo a (Jan-Roblero et al. 2008).
Análisis de la región V3 del gen 16S rRNA
Para amplificar la región variable 3 del gen 16S rRNA se usaron los iniciadores Derecho 2
(GCG CCG CCG CGC GCG GGC GGG GCG GGG GCA CGG GGG GCC TAC GGG
AGG CAG CAG en negritas el broche de GC) y Reverso 3 (ATT ACC GCG GCT GCT
GG), bajo las condiciones de reacción descritas por el mismo autor (Muyzer et al., 1993).
Detección de genes relevantes del ciclo del nitrógeno
Enriquecimiento de bacterias moduladoras de los pasos del ciclo del nitrógeno.

9
Se sembraron 0.5 g suelo húmedo en medios para bacterias desnitrificadoras,
nitrificadoras (oxidadoras de amonio y oxidadoras de nitrito) y fijadoras de nitrógeno. Se
incubaron durante 5 días, tres semanas y 7 días respectivamente a 28ªC en oscuridad.
Detección por PCR de los genes relevantes del ciclo del nitrógeno
Se extrajo el DNA metagenómico a partir de los enriquecimientos mediante el protocolo
previamente descrito por Cullen y Hirsh (1988). Se amplificaron los genes que codifican
para la amonio monooxigenasa (amoA), nitrito reductasa dependiente de cobre (nirK) y
citocromo cd1 (nirS), óxido nítrico reductasa (nosZ) y los genes 16S rRNA del género
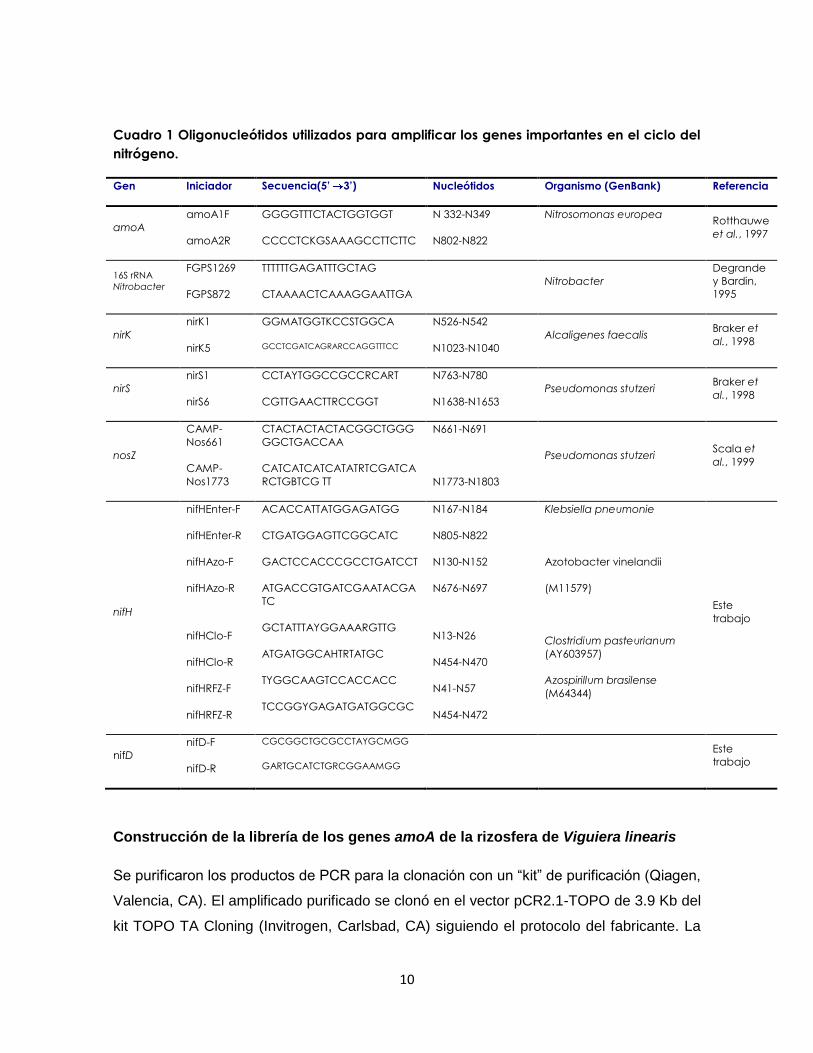
Nitrobacter. Los oligonucleótidos empleados se describen en el cuadro 1. El gen más
ampliamente utilizado para detectar la presencia de la capacidad potencial para fijar
nitrógeno es el gen nifH que codifica para la dinitrogenasa reductasa, una subunidad del
complejo enzimático responsable de esta actividad. Este gen está muy bien representado
en GenBank, por lo que es un blanco apropiado para estudiar la diversidad de este gen en
las bases de datos. También se emplea el gen nifD que codifica para una de las
subunidades de la proteína Fe-Mo. Debido a que el gen que codifica para la dinitrogenasa
(nifH) es muy poco conservado se diseñaron 4 juegos de iniciadores con el fin de evitar
degeneraciones y aumentar su especificidad. Se diseñaron a partir de un alineamiento
múltiple de secuencias completas de genes nifH disponibles en el banco de genes
(GenBank). Cada par de iniciadores amplifica uno de los cuatro grupos de genes nifH
reconocidos en el alineamiento (Cuadro 1).
10
Cuadro 1 Oligonucleótidos utilizados para amplificar los genes importantes en el ciclo del
nitrógeno.
Gen Iniciador Secuencia(5’ 3’) Nucleótidos Organismo (GenBank) Referencia
amoA
amoA1F
amoA2R
GGGGTTTCTACTGGTGGT
CCCCTCKGSAAAGCCTTCTTC
N 332-N349
N802-N822
Nitrosomonas europea
Rotthauwe
et al., 1997
16S rRNA
Nitrobacter
FGPS1269
FGPS872
TTTTTTGAGATTTGCTAG
CTAAAACTCAAAGGAATTGA
Nitrobacter
Degrande
y Bardin,
1995
nirK
nirK1
nirK5
GGMATGGTKCCSTGGCA
GCCTCGATCAGRARCCAGGTTTCC
N526-N542
N1023-N1040
Alcaligenes faecalis Braker et
al., 1998
nirS
nirS1
nirS6
CCTAYTGGCCGCCRCART
CGTTGAACTTRCCGGT
N763-N780
N1638-N1653
Pseudomonas stutzeri Braker et
al., 1998
nosZ
CAMP-
Nos661
CAMP-
Nos1773
CTACTACTACTACGGCTGGG
GGCTGACCAA
CATCATCATCATATRTCGATCA
RCTGBTCG TT
N661-N691
N1773-N1803
Pseudomonas stutzeri Scala et
al., 1999
nifH
nifHEnter-F
nifHEnter-R
nifHAzo-F
nifHAzo-R
nifHClo-F
nifHClo-R
nifHRFZ-F
nifHRFZ-R
ACACCATTATGGAGATGG
CTGATGGAGTTCGGCATC
GACTCCACCCGCCTGATCCT
ATGACCGTGATCGAATACGA
TC
GCTATTTAYGGAAARGTTG
ATGATGGCAHTRTATGC
TYGGCAAGTCCACCACC
TCCGGYGAGATGATGGCGC
N167-N184
N805-N822
N130-N152
N676-N697
N13-N26
N454-N470
N41-N57
N454-N472
Klebsiella pneumonie
Azotobacter vinelandii
(M11579)
Clostridium pasteurianum
(AY603957)
Azospirillum brasilense
(M64344)
Este
trabajo
nifD
nifD-F
nifD-R
CGCGGCTGCGCCTAYGCMGG
GARTGCATCTGRCGGAAMGG
Este
trabajo
Construcción de la librería de los genes amoA de la rizosfera de Viguiera linearis
Se purificaron los productos de PCR para la clonación con un “kit” de purificación (Qiagen,
Valencia, CA). El amplificado purificado se clonó en el vector pCR2.1-TOPO de 3.9 Kb del
kit TOPO TA Cloning (Invitrogen, Carlsbad, CA) siguiendo el protocolo del fabricante. La

11
transformación se realizó en células quimiocompetentes de la cepa DH10b de Escherichia
coli.
El crecimiento de las clonas se hizo en medio LB con ampicilina (50 μg/mL), X-gal (40 μg/
mL) e IPTG (0.5 mM). Las clonas con los plásmidos recombinantes se reconocieron por la
coloración blanca de la colonia y por su resistencia a ampicilina. Las clonas seleccionadas
se picaron con palillos de madera estériles y se sembraron en tubos con tapón de rosca
con 5 mL de medio LB adicionados con 5 µL de ampicilina (50 mg/mL) durante 24 h a
37°C. El DNA plasmídico se obtuvo por medio de la técnica de lisis alcalina (Sambrook et
al., 2002).
Se identificaron las clonas que contenían el inserto por liberación del mismo con la
endonucleasa de restricción EcoRI (Invitrogen, Carlsbad, CA) cuyas secuencias de corte
flanquean el sitio de inserción en el plásmido. Para priorizar los esfuerzos de
secuenciación se realizó un análisis RFLP con la enzima de restricción MboII. Se
secuenció una clona de cada perfil representativo. La secuenciación de los plásmidos
purificados se realizó de manera automatizada con un secuenciador ABI PRISM 310
Genetic Analyzer PE (Applied Biosystems). Se realizaron reacciones de secuenciación a
partir del extremo 5’.
Resultados
Análisis de la diversidad bacteriana de la rizosfera de Viguiera linearis y Bahia
xylopoda
Se obtuvo la secuencia parcial nucleotídica a partir de los extremos 5’ y 3’de
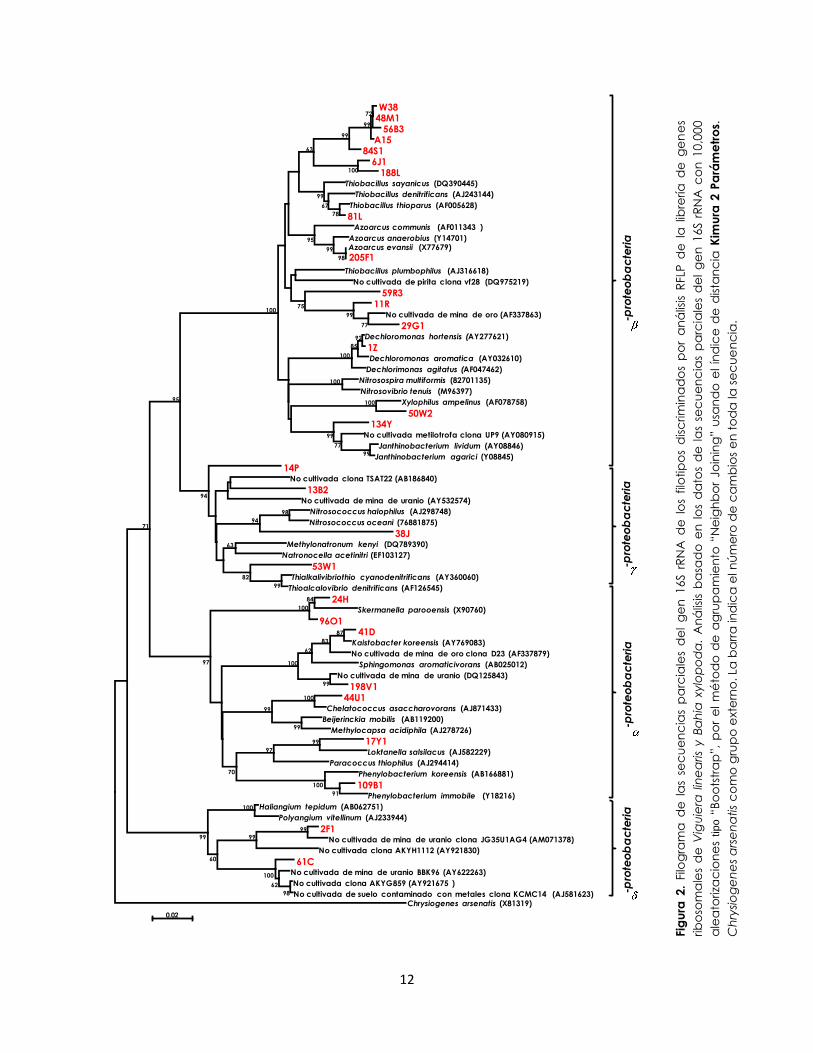
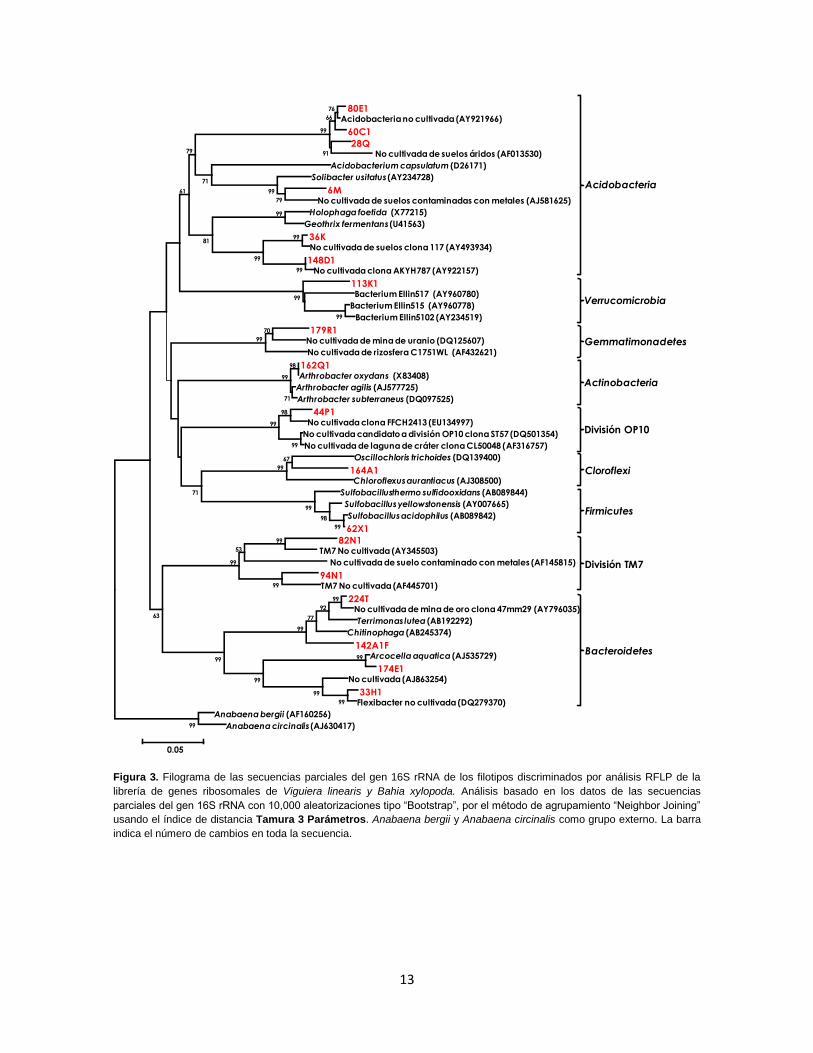
aproximadamente 1350 pb. Se identificaron 9 diferentes phyla en las librerías de genes
ribosomales de las dos rizosferas (Proteobacteria, Acidobacteria, Bacteroidetes,
Actinobacteria, Cloroflexi, Verrucomicrobia, Firmicutes, Gemmatimonadetes y la división
TM7), los cuales se exhiben en las Figuras 2 y 3.
12
W3848M1
56B3A15
84S16J1
188LThiobacillus sayanicus (DQ390445)
Thiobacillus denitrificans (AJ243144)
Thiobacillus thioparus (AF005628)
81LAzoarcus communis (AF011343 )
Azoarcus anaerobius (Y14701)Azoarcus evansii (X77679)
205F1
Thiobacillus plumbophilus (AJ316618)
No cultivada de pirita clona vf28 (DQ975219)
59R311R
No cultivada de mina de oro (AF337863)
29G1Dechloromonas hortensis (AY277621)
1ZDechloromonas aromatica (AY032610)
Dechlorimonas agitatus (AF047462)
Nitrosospira multiformis (82701135)
Nitrosovibrio tenuis (M96397)
Xylophilus ampelinus (AF078758)
50W2134Y
No cultivada metilotrofa clona UP9 (AY080915)
Janthinobacterium lividum (AY08846)
Janthinobacterium agarici (Y08845)
14PNo cultivada clona TSAT22 (AB186840)
13B2No cultivada de mina de uranio (AY532574)
Nitrosococcus halophilus (AJ298748)
Nitrosococcus oceani (76881875)
38JMethylonatronum kenyi (DQ789390)
Natronocella acetinitri (EF103127)
53W1Thialkalivibriothio cyanodenitrificans (AY360060)
Thioalcalovibrio denitrificans (AF126545)
24HSkermanella parooensis (X90760)
96O141D
Kaistobacter koreensis (AY769083)
No cultivada de mina de oro clona D23 (AF337879)
Sphingomonas aromaticivorans (AB025012)
No cultivada de mina de uranio (DQ125843)
198V144U1
Chelatococcus asaccharovorans (AJ871433)
Beijerinckia mobilis (AB119200)
Methylocapsa acidiphila (AJ278726)
17Y1Loktanella salsilacus (AJ582229)
Paracoccus thiophilus (AJ294414)
Phenylobacterium koreensis (AB166881)
109B1Phenylobacterium immobile (Y18216)
Haliangium tepidum (AB062751)
Polyangium vitellinum (AJ233944)
2F1No cultivada de mina de uranio clona JG35U1AG4 (AM071378)
No cultivada clona AKYH1112 (AY921830)
61CNo cultivada de mina de uranio BBK96 (AY622263)
No cultivada clona AKYG859 (AY921675 )
No cultivada de suelo contaminado con metales clona KCMC14 (AJ581623)Chrysiogenes arsenatis (X81319)
100
92
85
100
100
100
77
99
72
99
99
77
99
99
78
67
99
98
99
95
75
63
100
99
87
83
62
100
100
84
100
100
91
100
98
62
100
99
99
60
99
99
99
99
99
98
97
70
97
94
82
63
94
95
71
0.02
-pro
teo
ba
cte
ria
-pro
teo
ba
cte
ria
-pro
teo
ba
cte
ria
-pro
teo
ba
cte
ria
Fig
ura
2.
Filo
gra
ma
de
la
s se
cu
en
cia
s p
arc
iale
s d
el
ge
n 1
6S r
RN
A d
e l
os
filo
tip
os
dis
crim
ina
do
s p
or
an
ális
is R
FLP
de
la
lib
rería
de
ge
ne
s
rib
oso
ma
les
de
Vig
uie
ra l
ine
aris
y B
ah
ia x
ylo
po
da
. A
ná
lisis
ba
sad
o e
n l
os
da
tos
de
la
s se
cu
en
cia
s p
arc
iale
s d
el
ge
n 1
6S r
RN
A c
on
10
,00
0
ale
ato
riza
cio
ne
s tip
o “
Bo
ots
tra
p”,
po
r e
l m
éto
do
de
ag
rup
am
ien
to “
Ne
igh
bo
r Jo
inin
g” u
san
do
el
índ
ice
de
dis
tan
cia
Kim
ura
2 P
ará
me
tro
s.
Ch
rysi
og
en
es
ars
en
atis
co
mo
gru
po
exte
rno
. La
ba
rra
ind
ica
el n
úm
ero
de
ca
mb
ios
en
to
da
la s
ec
ue
nc
ia.
13
80E1Acidobacteria no cultivada (AY921966)
60C1
28QNo cultivada de suelos áridos (AF013530)
Acidobacterium capsulatum (D26171)
Solibacter usitatus (AY234728)
6MNo cultivada de suelos contaminadas con metales (AJ581625)
Holophaga foetida (X77215)
Geothrix fermentans (U41563)
36KNo cultivada de suelos clona 117 (AY493934)
148D1No cultivada clona AKYH787 (AY922157)
113K1Bacterium Ellin517 (AY960780)
Bacterium Ellin515 (AY960778)
Bacterium Ellin5102 (AY234519)
179R1No cultivada de mina de uranio (DQ125607)
No cultivada de rizosfera C1751WL (AF432621)
162Q1Arthrobacter oxydans (X83408)
Arthrobacter agilis (AJ577725)
Arthrobacter subterraneus (DQ097525)
44P1No cultivada clona FFCH2413 (EU134997)
No cultivada candidatoa división OP10 clona ST57 (DQ501354)
No cultivada de laguna de cráter clona CL50048 (AF316757)
Oscillochloris trichoides (DQ139400)
164A1Chloroflexusaurantiacus (AJ308500)
Sulfobacillusthermo sulfidooxidans (AB089844)
Sulfobacillusyellowstonensis (AY007665)
Sulfobacillusacidophilus (AB089842)
62X1
82N1TM7 No cultivada (AY345503)
No cultivada de suelo contaminado con metales (AF145815)
94N1TM7 No cultivada (AF445701)
224TNo cultivada de mina de oro clona 47mm29 (AY796035)
Terrimonaslutea (AB192292)
Chitinophaga (AB245374)
142A1FArcocella aquatica (AJ535729)
174E1No cultivada (AJ863254)
33H1Flexibacter no cultivada (DQ279370)
Anabaena bergii (AF160256)
Anabaena circinalis (AJ630417)
99
99
99
99
92
99
77
99
99
98
99
76
66
91
99
99
99
99
99
99
99
99
98
71
99
70
99
99
67
99
99
98
99
53
99
99
79
99
99
81
71
71
63
79
61
0.05
Acidobacteria
Verrucomicrobia
Gemmatimonadetes
Actinobacteria
División OP10
Cloroflexi
Firmicutes
División TM7
Bacteroidetes
Figura 3. Filograma de las secuencias parciales del gen 16S rRNA de los filotipos discriminados por análisis RFLP de la
librería de genes ribosomales de Viguiera linearis y Bahia xylopoda. Análisis basado en los datos de las secuencias
parciales del gen 16S rRNA con 10,000 aleatorizaciones tipo “Bootstrap”, por el método de agrupamiento “Neighbor Joining”
usando el índice de distancia Tamura 3 Parámetros. Anabaena bergii y Anabaena circinalis como grupo externo. La barra
indica el número de cambios en toda la secuencia.
14
Nitrificación
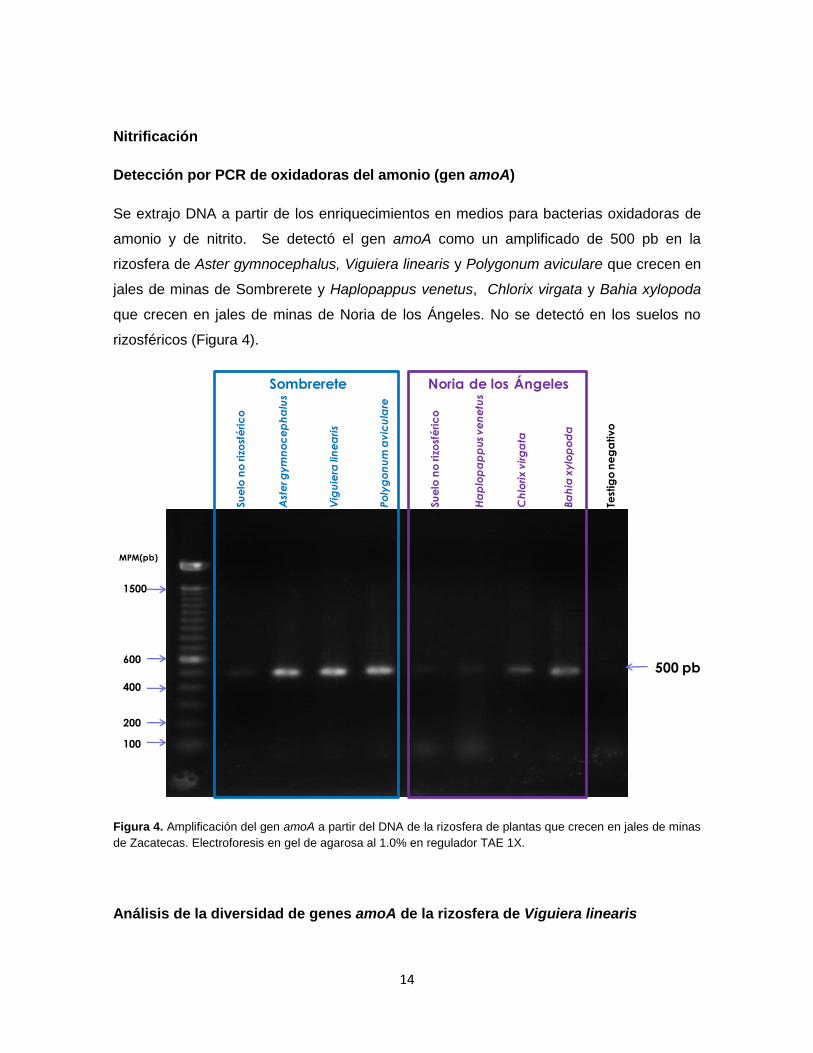
Detección por PCR de oxidadoras del amonio (gen amoA)
Se extrajo DNA a partir de los enriquecimientos en medios para bacterias oxidadoras de
amonio y de nitrito. Se detectó el gen amoA como un amplificado de 500 pb en la
rizosfera de Aster gymnocephalus, Viguiera linearis y Polygonum aviculare que crecen en
jales de minas de Sombrerete y Haplopappus venetus, Chlorix virgata y Bahia xylopoda
que crecen en jales de minas de Noria de los Ángeles. No se detectó en los suelos no
rizosféricos (Figura 4).
1500
600
400
200
100
MPM(pb)
500 pb
Sombrerete Noria de los Ángeles
Su
elo
no
riz
osf
éri
co
Ast
erg
ym
no
ce
ph
alu
s
Vig
uie
ra li
ne
ari
s
Po
lyg
on
um
av
icu
lare
Su
elo
no
riz
osf
éri
co
Ha
plo
pa
pp
us
ve
ne
tus
Ch
lori
xv
irg
ata
Ba
hia
xy
lop
od
a
Test
igo
ne
ga
tiv
o
Figura 4. Amplificación del gen amoA a partir del DNA de la rizosfera de plantas que crecen en jales de minas
de Zacatecas. Electroforesis en gel de agarosa al 1.0% en regulador TAE 1X.
Análisis de la diversidad de genes amoA de la rizosfera de Viguiera linearis

15
En la Figura 5 es una imagen de la restricción con la enzima de restricción EcoRI, la
banda de 4000 pb corresponde al vector y la banda de 500 pb es el inserto liberado.
10000
4000
1000750
500
250
MPM(pb)
3900 pb
500 pb
Figura 5. Restricción con EcoRI para liberar el inserto del vector 2.1 pCR TOPO.
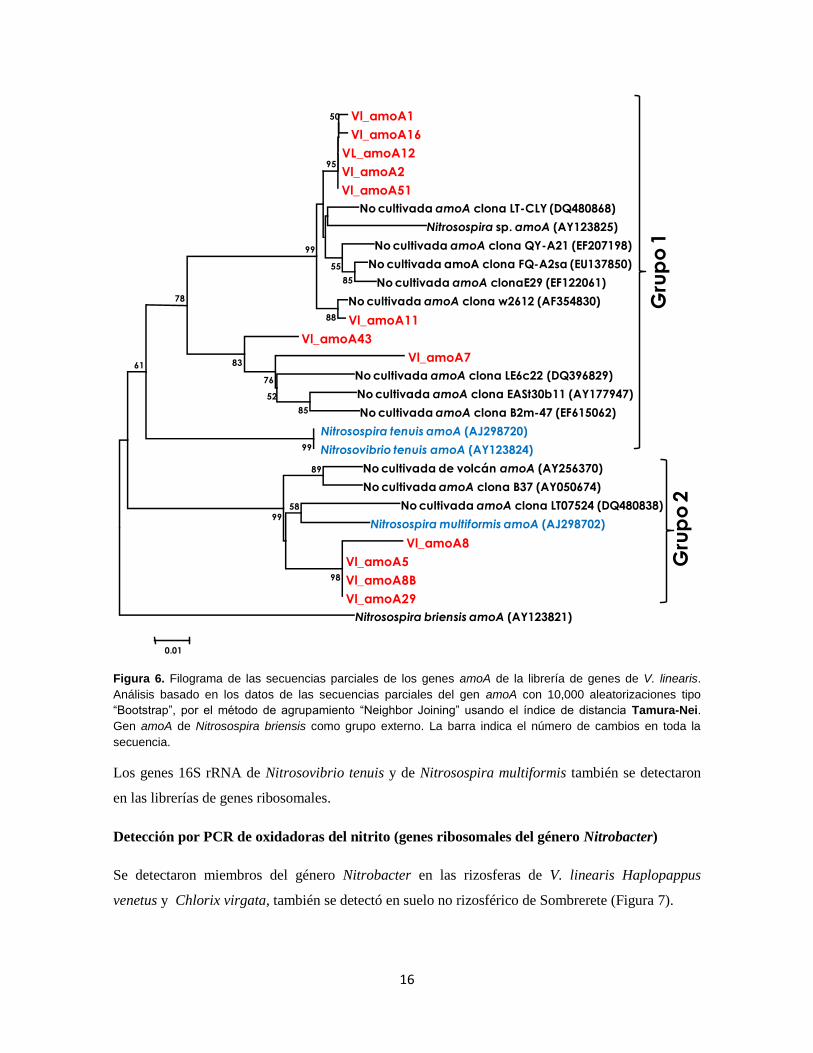
Aproximadamente 500 nucleótidos derivados de la secuencia de los genes amoA de la
librería de V. linearis se utilizaron para hacer un análisis de las relaciones filogenéticas
con secuencias del banco de genes (Figura 6). El árbol filogenético nos permitió
reconocer 2 grupos principales. El grupo 1 comprende genes amoA de los
microorganismos Nitrosospira tenuis y Nitrovibrio tenuis, el grupo 2 incluye genes amoA
de Nitrosospira multiformis. La secuencia de nuestras clonas se ubicaron en ambos
grupos.
Los genes 16S rRNA de Nitrosovibrio tenuis y de Nitrosospira multiformis también se detectaron
en las librerías de genes ribosomales.
Detección por PCR de oxidadoras del nitrito (genes ribosomales del género Nitrobacter)
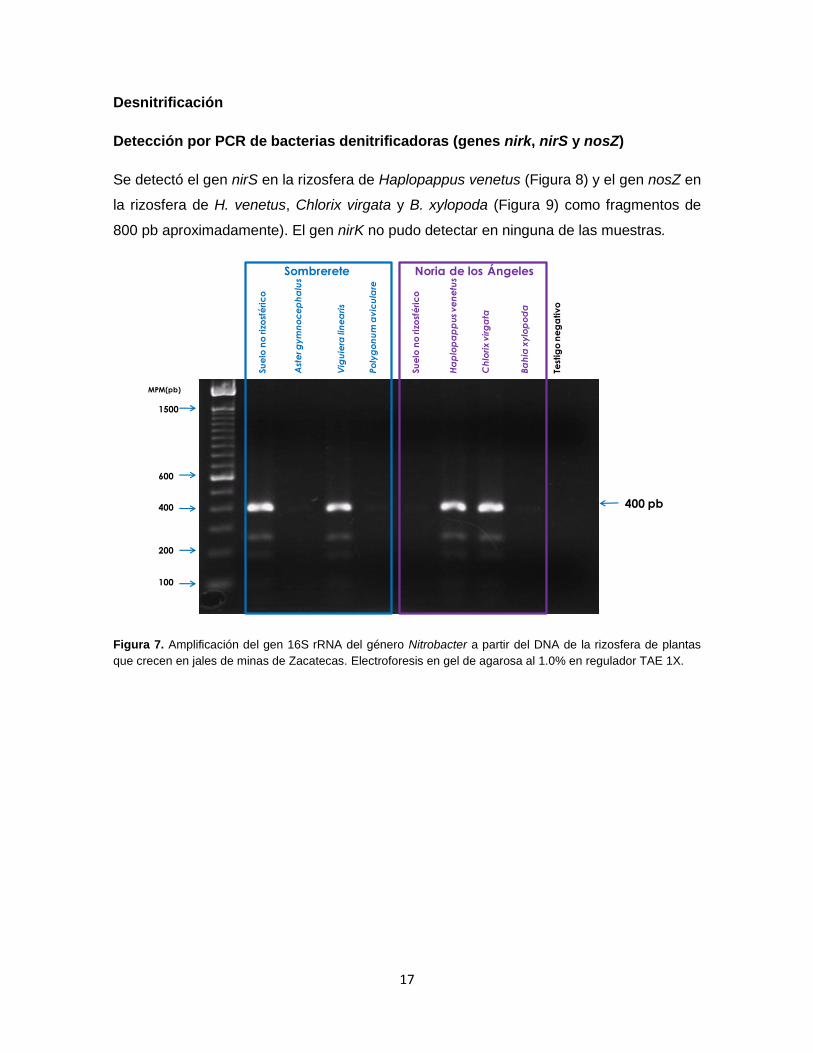
Se detectaron miembros del género Nitrobacter en las rizosferas de V. linearis Haplopappus
venetus y Chlorix virgata, también se detectó en suelo no rizosférico de Sombrerete (Figura 7).
16
Vl_amoA1
Vl_amoA16
VL_amoA12
Vl_amoA2
Vl_amoA51
No cultivada amoA clona LT-CLY (DQ480868)
Nitrosospira sp. amoA (AY123825)
No cultivada amoA clona QY-A21 (EF207198)
No cultivada amoA clona FQ-A2sa (EU137850)
No cultivada amoA clonaE29 (EF122061)
No cultivada amoA clona w2612 (AF354830)
Vl_amoA11
Vl_amoA43
Vl_amoA7
No cultivada amoA clona LE6c22 (DQ396829)
No cultivada amoA clona EASt30b11 (AY177947)
No cultivada amoA clona B2m-47 (EF615062)
Nitrosospira tenuis amoA (AJ298720)
Nitrosovibrio tenuis amoA (AY123824)
No cultivada de volcán amoA (AY256370)
No cultivada amoA clona B37 (AY050674)
No cultivada amoA clona LT07524 (DQ480838)
Nitrosospira multiformis amoA (AJ298702)
Vl_amoA8
Vl_amoA5
Vl_amoA8B
Vl_amoA29
Nitrosospira briensis amoA (AY123821)
98
89
5899
50
95
88
85
55
99
99
85
52
76
83
78
61
0.01
Gru
po
1G
rup
o 2
Figura 6. Filograma de las secuencias parciales de los genes amoA de la librería de genes de V. linearis.
Análisis basado en los datos de las secuencias parciales del gen amoA con 10,000 aleatorizaciones tipo
“Bootstrap”, por el método de agrupamiento “Neighbor Joining” usando el índice de distancia Tamura-Nei.
Gen amoA de Nitrosospira briensis como grupo externo. La barra indica el número de cambios en toda la
secuencia.
Los genes 16S rRNA de Nitrosovibrio tenuis y de Nitrosospira multiformis también se detectaron
en las librerías de genes ribosomales.
Detección por PCR de oxidadoras del nitrito (genes ribosomales del género Nitrobacter)
Se detectaron miembros del género Nitrobacter en las rizosferas de V. linearis Haplopappus
venetus y Chlorix virgata, también se detectó en suelo no rizosférico de Sombrerete (Figura 7).
17
Desnitrificación
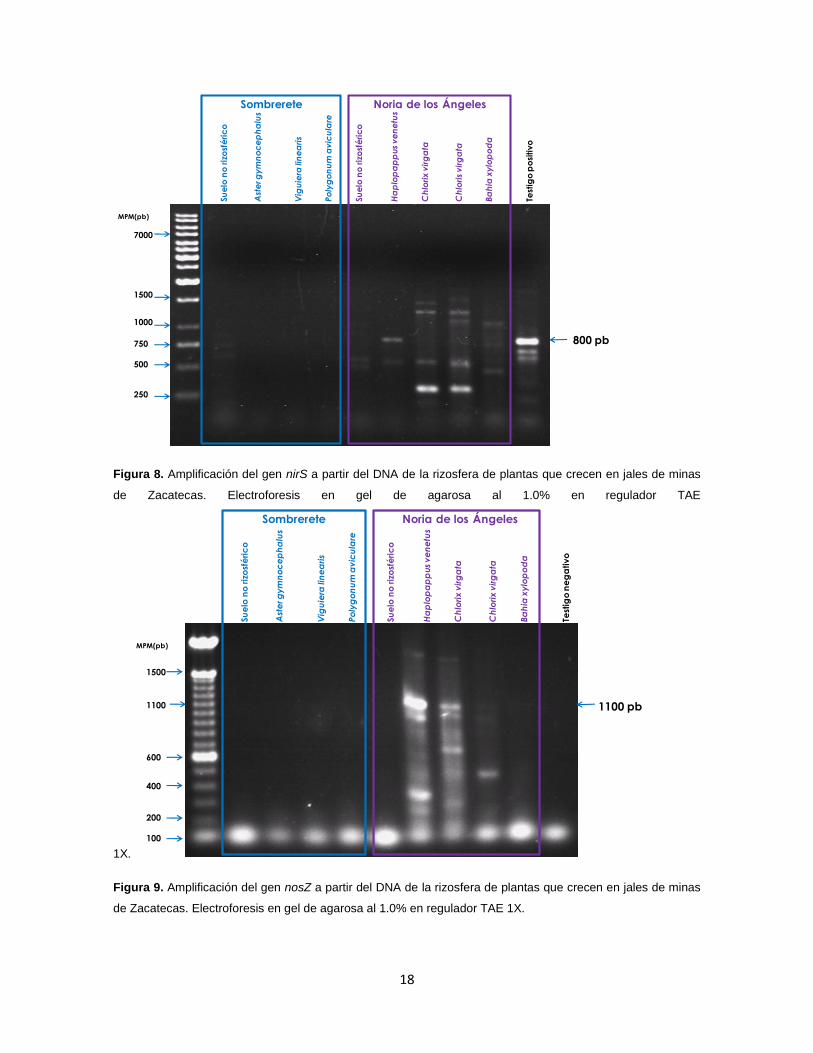
Detección por PCR de bacterias denitrificadoras (genes nirk, nirS y nosZ)
Se detectó el gen nirS en la rizosfera de Haplopappus venetus (Figura 8) y el gen nosZ en
la rizosfera de H. venetus, Chlorix virgata y B. xylopoda (Figura 9) como fragmentos de
800 pb aproximadamente). El gen nirK no pudo detectar en ninguna de las muestras.
Sombrerete Noria de los Ángeles
1500
600
400
200
100
MPM(pb)
400 pb
Su
elo
no
riz
osf
éri
co
Ast
erg
ym
no
ce
ph
alu
s
Vig
uie
ra li
ne
ari
s
Po
lyg
on
um
av
icu
lare
Su
elo
no
riz
osf
éri
co
Ha
plo
pa
pp
us
ve
ne
tus
Ch
lori
xv
irg
ata
Ba
hia
xy
lop
od
a
Test
igo
ne
ga
tiv
o
Figura 7. Amplificación del gen 16S rRNA del género Nitrobacter a partir del DNA de la rizosfera de plantas
que crecen en jales de minas de Zacatecas. Electroforesis en gel de agarosa al 1.0% en regulador TAE 1X.
18
800 pb
7000
1500
1000
750
500
250
MPM(pb)
Sombrerete Noria de los Ángeles
Su
elo
no
riz
osf
éri
co
Ast
erg
ym
no
ce
ph
alu
s
Vig
uie
ra li
ne
ari
s
Po
lyg
on
um
avic
ula
re
Su
elo
no
riz
osf
éri
co
Ha
plo
pa
pp
us
ve
ne
tus
Ch
lori
xv
irg
ata
Ch
lori
sv
irg
ata
Ba
hia
xy
lop
od
a
Test
igo
po
sitivo
Figura 8. Amplificación del gen nirS a partir del DNA de la rizosfera de plantas que crecen en jales de minas
de Zacatecas. Electroforesis en gel de agarosa al 1.0% en regulador TAE
1X.
1100 pb
1500
1100
600
400
200
100
MPM(pb)
Sombrerete Noria de los Ángeles
Su
elo
no
riz
osf
éri
co
Ast
erg
ym
no
ce
ph
alu
s
Vig
uie
ra li
ne
ari
s
Po
lyg
on
um
av
icu
lare
Su
elo
no
riz
osf
éri
co
Ha
plo
pa
pp
us
ve
ne
tus
Ch
lori
xv
irg
ata
Ch
lori
xv
irg
ata
Ba
hia
xy
lop
od
a
Test
igo
ne
ga
tiv
o
Figura 9. Amplificación del gen nosZ a partir del DNA de la rizosfera de plantas que crecen en jales de minas
de Zacatecas. Electroforesis en gel de agarosa al 1.0% en regulador TAE 1X.

19
En general, se detectaron más genes en las rizosferas que en el suelo no rizosférico. En
las rizosferas de las plantas de Sombrerete sólo se detectaron los genes relacionados al
paso de la nitrificación. En las rizosferas de las plantas de Noria de los Ángeles se
detectaron todos los pasos del ciclo. Sólo en la rizosfera de Chlorix virgata, que crece en
jales de minas de Noria de los Ángeles, se detectó el ciclo del nitrógeno completo (fijación
de nitrógeno, nitrificación y desnitrificación). En el suelo no rizosférico de Noria de los
Ángeles no se detectó ninguno de los genes buscados. Los pasos del ciclo del nitrógeno
que se encontraron en la mayoría de los suelos fueron la oxidación del amonio y la
oxidación del nitrito.
Diversidad de la región V3 de los genes 16S rRNA de los enriquecimientos
A diferencia de la DGGE a partir del DNA directo, donde no se observó una extensa
diversidad, las DGGEs de los enriquecimientos mostraron amplia diversidad en todos los
suelos (Figuras 10 y 11). También se observó una clara diferencia entre los perfiles
desplegados por los diferentes enriquecimientos. Los medios donde se observó la mayor
diversidad fue el de oxidadores de amonio y oxidadores de nitrito, esta observación
coincide con los resultados obtenidos en la detección por PCR. El medio con la diversidad
más pobre fue el medio para desnitrificadoras. El efecto rizosférico es muy claro en todos
los enriquecimientos. En el medio para bacterias fijadoras del nitrógeno, las rizosferas de
V. linearis y B. xylopoda fueron las que presentaron los perfiles con un mayor número de
bandas. La rizosfera de Chlorix virgata, aunque fue la única en amplificar el gen nifH de
Clostridia, en la DGGE sólo se observaron 5 bandas. Hay dos bandas compartidas (las
dos bandas más intensas en Haplopapus venetus) en todos los suelos, incluyendo los
suelos no rizosféricos.

20
Figura 10. Diversidad bacteriana de la rizosfera de plantas que crecen en jales de minas de Zacatecas
deducida por PCR-DGGE de la región variable 3 del gen 16S rRNA enriquecimientos en medio para fijadoras
de nitrógeno y oxidadoras de amonio. * Suelo donde se detectó el gen responsable de la función respectiva.
La flecha indica el gradiente desnaturalizante.
Figura 11. Diversidad bacteriana de la rizosfera de plantas que crecen en jales de minas de Zacatecas
deducida por PCR-DGGE de la región variable 3 del gen 16S rRNA de los enriquecimientos en medio para
oxidadoras de nitrito y desnitrificadoras. * Suelo donde se detectó el gen responsable de la función respectiva.
La flecha indica el gradiente desnaturalizante.
* * * * * *

21
IMPACTO
El beneficio más sobresaliente de este proyecto fue en la formación de recursos
humanos, tuvimos dos tesis a nivel posgrado; una de las cuales próxima a titularse, y la
otra aún en proceso. También incorporamos a dos alumnas de licenciatura, bajo esta
línea de investigación. Además con los resultados generados en el presente proyecto
publicaremos un artículo de investigación científica, el cual estamos escribiendo.
REFERENCIAS
Adriano CD (2001) Trace elements in terrestrial environments. Segunda edición. Springer. New
York pp 867.
Alloway BJ (1995) Soil processes and the behaviour of metals. En: Alloway BJ, Heavy metals in
soils (Ed) Blackie A y P London, UK. pp11-37.
Behar A, Yuval B, Jurkevitch E (2005) Enterobacteria-mediated nitrogen fixation in natural
populations of the fruit fly Ceratitis capitata. Mol Ecol 14:2637-2643.
Braker G, Fesefeldt A, Witzel KP (1998) Development of PCR primer systems for amplification of
nitrite reductase genes (nirk and nirS) to detect denitrifying bacteria in environmental samples. Appl
Environ Microbiol 64:3769-3775.
Cullen DW, Hirsch PR (1988) Simple and rapid method for direct extraction of microbial DNA from
soil to PCR. Soil Biol Biochem 30:983-993.
Degrange V, Bardin R (1995) Detection and counting of Nitrobacter populations in soil by PCR.
Appl Environ Microbiol 61:2093-2098.
Einsle OKP (2004) Structural basis of denitrification. Biol Chem 385:875-883.
Francis CA, Beman JM, Kuypers MMM (2007) New processes and players in the nitrogen cycle: the
microbial ecology of anaerobic and archaeal ammonia oxidation. ISME J 1:19-27.
Glockner AJA, Zumft W (1993) Copper-containing nitrite reductase from Pseudomonas
aureofaciens is functional in a mutationally cytochrome cd1-free background (NirS) os
Pseudomonas stutzeri. Arch Microbiol 160:18-26
Jan-Roblero J, Posadas A, Zavala Díaz de la Serna J, García R, Hernández-Rodríguez C (2008).
Phylogenetic characterization of bacterial consortia obtained of corroding gas pipelines in Mexico.
World J. Microbiol. Biotechnol. 24:1775-1784.
Knowles R (1982) Denitrification. Microbiol Rev 46:43-70.
Kowalchuk GA, Stephen JR (2001). Ammonia-oxidizing bacteria: a model for molecular microbial
ecology. Annu Rev Microbiol 55:485-529.
Mulder A, Graaf AA, Robertson LA, Kuenen JG (1995) Anaerobic ammonium oxidation discovered
in a denitrifying fluized bed reactor. FEMS Microbiol Ecol 16:177-184.

22
Muyzer G, De Waall EC, Vitterlinden AG (1993) Profiling of complete microbial populations by
denaturing gradient gel electrophoresis analysis of poliymerase chain reaction-amplified genes
coding for 16S rDNA. Appl Environ Microbiol 59:695-700.
Ongley LK, Armienta A, Mango H (2003) Concentrations of heavy metals in soil, Zimapan, México.
J Phy IV France 107:983-986.
Purkhold U, Pommerening-Röser A, Juretschko S, Schi,id MC, Koops HP, Wagner M (2000)
Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and
amoA sequence analysis: implications for molecular diversity surveys. Appl Environ Microbiol
66:5368-5382.
Risgaard-Petersen N, Langezaal AM, Ingvardsen S, Schmid MC, Jetten MSM, Op den Camp HJM
(2006) Evidence for complete denitrification in a benthic foraminifer. Nature 443:93-96.
Rotthauwe JWK, Liesack W (1997) The ammonia monooxygenase structural gene amoA as a
functional marker: molecular fine-scale analysis of natural ammonia-oxidazing populations. Appl
Environ Microbiol 63:4704-4712.
Sambrook J, de Souza FA, Van Elsas JD (2002) Molecular method to assess the diversity of
Burkholderia species in environmental samples. Appl Environ Microbiol 68:595-603.
Scala DJ, Kerkhof LJ (1999) Diversity of nitrous oxide reductase (nosZ) genes in continental shelf
sediments. Appl Environ Microbiol 65:1681-1687.
Secretaría de Economía (2006) Disponible en: http://www.economia.gob.mx.
Straub KLM, Benz B, Widdel F (1996) Anaerobic, nitrate dependent microbial oxidation of ferrous
iron. Appl Environ Microbiol 62:1458-1460.
Strous M, Kuenen JG, Jetten MSM (1999) Key physiology of anaerobic ammonium oxidation. Appl
Environ Microbiol 65:3248-3250.
Tate RL (1977) Nitrification in histosols: a potencial role for the heterotrophic nitrifier. Appl Environ
Microbiol 33:911-914.
Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA (2004) Environmental
genome shotgun sequencing of the Sargasso Sea. Science 304:66-74
Zumft W (1997) Cell biology and molecular basis of denitrification. Microbiol Mol Biol Rev 61:533-
616.





























