Embed Size (px)
Citation preview

UNIVERSIDAD DE CHILE
FACULTAD DE CIENCIAS QUIMICAS Y FARMACEUTICAS
HOSPITAL CLINICO DE LA UNIVERSIDAD DE CHILE
Departamento de Endocrinología
Laboratorio de Endocrinología y Biología de la Reproducción
“ROL DEL FACTOR DE CRECIMIENTO NERVIOSO
(NGF) EN LA REGULACION DE LA PROLIFERACION Y
ANGIOGENESIS DEL OVARIO”
Memoria para optar al título de Bioquímico
PATRICIA EMITA LOZADA GONZALEZ
PROFESOR PATROCINANTE DIRECTOR DE MEMORIA
Dr. Hernán Lara Peñaloza Dra. Carmen Romero Osses
Facultad de Cs. Químicas y Farmacéuticas Hospital Clínico Universidad de Chile
Santiago-Chile 2006

ii
“Esta tesis va dedicada a mi madre por su perseverancia, esfuerzo, y sacrificio en la vida y
por inculcarme valores hermosos. Gracias vieja por estar en todos los momentos de mi vida, por
acompañarme en las buenas y en las malas, por estar ahí cuando las fuerzas ya no estaban, y
me decías estudia que queda poco, y por repetirme cuan importante era un cartón, porque con
eso me podría defender en la vida. Para ti esto, porque tú fuiste el artífice. Gracias viejita”.

iii
ABREVIATURAS
ADN: Ácido Desoxirribunoucleico ARN: Ácido Ribonucleico. ARNm: Ácido ribonucleico mensajero. BDNF: Factor Neurotrófico Derivado del Cerebro. FSH: Hormona Folículo Estimulante. HIF-1 alfa: Factor inducible por hipoxia1 alfa. LH: Hormona Luteinizante. NGF: Factor de crecimiento nervioso. NT-3: Neurotrofina 3. NT 4/5: Neurotrofina 4/5. PCR: Reacción en cadena de la polimerasa. RT: Transcripción Reversa. tkA: receptor tirosina Kinasa A. VEGF: Factor de crecimiento de endotelio vascular.

iv
INDICE PAGINA
RESUMEN.......................................................................................................... V
SUMMARY ....................................................................................................... VII
INTRODUCCION................................................................................................ 9
HIPOTESIS....................................................................................................... 15
I OBJETIVOS .............................................................................................. 16
I. 1 OBJETIVO GENERAL ............................................................................... 16
I. 2 OBJETIVOS ESPECÍFICOS ...................................................................... 16
II MATERIALES Y MÉTODOS..................................................................... 17
III RESULTADOS .......................................................................................... 24
IV DISCUSION............................................................................................... 32
V CONCLUSIONES...................................................................................... 37
VI REFERENCIAS ......................................................................................... 38

v
RESUMEN
“Rol del factor de crecimiento nervioso (NGF) en la regulación de la proliferación y angiogénesis del ovario”
El factor de crecimiento nervioso (NGF) es una neurotrofina, que inicialmente fue
descrita en el cerebro, y que en los últimos años ha adquirido importancia, debido a que actúa en
sitios no-neuronales, entre ellos el ovario, donde se ha encontrado que NGF participa en el
desarrollo ovárico, proliferación y diferenciación de células somáticas y en la esteroidogénesis.
Una de las etapas importantes que ocurren durante el desarrollo folicular y del cuerpo lúteo es la
angiogénesis, una de las moléculas importantes en este proceso es el factor de crecimiento de
endotelio vascular (VEGF).
NGF aumenta la expresión de VEGF en diversos tipos celulares. VEGF es regulado en
forma directa por el gen inducible por hipoxia (HIF-1) y en forma indirecta por c-Myc. Además
c-Myc, es un gen involucrado en la proliferación celular. En este trabajo se investigó el rol que
cumple NGF en la regulación de VEGF y la proliferación celular. Para este fin se utilizaron
células de granulosa humana, provenientes de pacientes del programa de fertilización asistida,
las cuales se cultivaron por 2, 4, 8 y 18 horas con NGF (50 ng/ml). Se midieron los niveles del
ARNm de VEGF, c-Myc e HIF-1 alfa por RT-PCR. En este trabajo el ARNm de las isoformas
de VEGF 165 y 121 aumentaron a las 4 y 8 horas respectivamente, los niveles del ARNm de
HIF-1 alfa se mantuvieron inalterados a todos los tiempos estudiados. Los niveles de ARNm de
c-Myc aumentaron a las 8 y 18 horas por efecto de NGF. Por Inmunocitoquímica se determinó
Ki67, un marcador de proliferación celular. La proteína Ki67 aumentó a las 8 y 18 horas por
efecto de NGF, concordando con la expresión génica de c-Myc.
Los resultados indican que NGF presenta la capacidad de aumentar, tanto la expresión
del ARNm de VEGF, como la proliferación en forma independiente, y que este aumento en la
proliferación sería en una etapa posterior al alza de VEGF y que probablemente el VEGF
expresado en las células de granulosa humana en cultivo, éste involucrado en la proliferación
directamente, esto podría suponer que una falla en la producción de NGF por parte del folículo

vi
podría llevar a éste a un crecimiento y maduración deficiente, debido a que no ocurriría una
angiogénesis y proliferación adecuada.

vii
SUMMARY
“Role of nerve growth factor (NGF) in the regulation of
proliferation and angiogenesis of ovary”
The nerve growth factor (NGF) is a neurothrophin, initially described in the brain, that
has acquired importance in the last few years, because it has several functions in non-neuronal
tissues. One of these tissues is the ovary, where NGF participates in the ovarian development,
proliferation and differentiation of somatic cells and steroidogenesis. Angiogenesis is an
essential stage during follicular and luteal development, and one of the most important
molecules in this process is the vascular endothelial growth factor (VEGF).
NGF increases VEGF expression in different cell types. VEGF is directly regulated by
hypoxia inducible factor-1(HIF-1) and indirectly by c-Myc. c-Myc is also a gene involved in cell
proliferation. This work investigated the role of NGF in regulating VEGF and cell proliferation.
To this aim, granulosa cells obtained from patients undergoing in vitro fertilization were cultured
for 2, 4, 8 and 18 hours with NGF (50 ng/ml). The levels of VEGF, c-Myc and HIF-1alpha
mRNA were evaluated by RT-PCR. mRNA for VEGF 165 and VEGF 121 isoforms increase at 4
and 8 hours respectively. HIF-1alpha mRNA levels did not change at any time. NGF (50ng/ml)
induces c-Myc mRNA increase at 8 and 18 hours. Ki67 protein, a cell proliferation marker, was
determined by immunocytochemistry. Ki67 protein increases at 8 and 18 hours with NGF
stimulation, and no change was observed at 4 hours, agreeing with the genic expression of c-
Myc.
The results indicate that NGF is able to increase VEGF mRNA expression and cell
proliferation by independent ways, and that the increase in the proliferation would be a later
stage to VEGF's rise. It is probable that the VEGF expressed in cultured human granulosa cells
acts directly on cell proliferation, suggesting that fails in follicular NGF production might induce

viii
a deficient follicular growth and maturation, due to impairment in angiogenesis and cell
proliferation.

9
INTRODUCCION
El ovario es un órgano muy importante para la reproducción humana, debido a que es el
encargado de la producción de las células germinales en la mujer. Presenta muchos
compartimentos, con múltiples funciones y propiedades biológicas. La función principal de esta
gónada es la diferenciación y liberación del ovocito maduro (ovulación). Además el ovario tiene
la capacidad de producir esteroides, que le otorgan las características sexuales secundarias a la
mujer y soportan el embarazo (McGee E y Hsueh A, 2000). En la etapa de diferenciación se
produce el crecimiento y desarrollo del folículo, desde el momento que deja la población de
reserva, proceso que puede llevar en promedio 85 días (Vega M, 1997) desde un folículo
primordial hasta llegar a un folículo preovulatorio o de Graaf.
El folículo es la unidad básica del ovario, y esta compuesto por un ovocito, que está
rodeado por las células de granulosa (CG) que rodean al ovocito, la membrana basal y las células
de la teca que rodean a las CG. Los folículos primordiales están compuestos por capas de células
epiteliales conocidas como CG, que rodean al ovocito inmaduro, además a las CG las circunda
una matriz extracelular llamada lámina basal, el primer cambio que se observa en las CG, es el
paso de células planas a cúbicas, dando origen a los folículos primarios, además de la iniciación
de la actividad mitótica, produciendo con esto un aumento en el número de capas de CG, dando
origen a los folículos secundarios. En el estado tardío de la diferenciación del folículo primario
se produce el inicio de la expresión de receptores de la Hormona Folículo Estimulante (FSH) en
las CG, lo cuál tiene una importancia clave debido a que estos receptores estimulan la
proliferación de las CG y, por lo tanto del crecimiento folicular. El folículo para llegar a ser un
folículo preovulatorio, ovulatorio o de Graaf, debe pasar por varias etapas antes de ser maduro y
poder liberar al ovocito, requiere la formación de la teca a través de una migración de células
mesenquemáticas hacia la lámina basal, formación del antro, y principalmente la proliferación de
CG, esto último produce un gran aumento del tamaño del folicular (Vega M, 1997).
Las neurotrofinas son factores de crecimiento, que inicialmente fueron encontradas en
cerebro, estos neuropéptidos son importantes para el desarrollo y mantención del sistema
nervioso central y periférico (Wiesmann C y col, 2001), pero numerosas evidencias han apoyado

10
la idea, de que estas afectan la sobrevida, diferenciación y proliferación de células no-
neuronales. Una de las neurotrofinas más prominentes, es el factor de crecimiento nervioso
(NGF). Entre los tejidos no neuronales en los cuales participa NGF, se encuentra el ovario,
donde también se ha observado la expresión de varios otros miembros de la familia de las
neurotrofinas, tales como BDNF, NT-3 y NT 4/5 y sus receptores durante el desarrollo ovárico
(Dissen y col, 1995). NGF tiene la capacidad de unirse a 2 receptores, uno de baja afinidad, p75
NGFR, el cual también puede unirse a otras neurotrofinas, y uno de alta afinidad, el receptor de
tirosina Kinasa A (trkA) (Tesarollo L, 1998). A través de estos receptores, NGF ejerce sus
funciones, y junto con otras neurotrofinas y sus receptores, está presente en el ovario humano
desde la etapa fetal (Anderson y col, 2002).
Ratas en edad pre-puberal, tratadas durante los primeros días de vida postnatal con
anticuerpos anti NGF, impiden el crecimiento de folículos antrales, retrasan la pubertad y alteran
la ciclicidad estral (Lara y col 1990). Se sugirió, que NGF era requerido para la ovulación y
desarrollo folicular temprano, debido a que la expresión del ARNm de NGF y trkA aumentan
transitoriamente antes de la ovulación en ovario de roedores (Dissen y col, 1996). Concordante a
esto, NGF también presenta la capacidad de aumentar la Cox-2, enzima involucrada en la
síntesis de la prostaglandina E-2, importante en la ovulación (Dissen GA y col, 2000). Además
en la ovulación, se interrumpen las uniones gap, produciendo primero un aumento en la
fosforilación de la conexina-43, y después una disminución de los niveles de la proteína
Conexina-43 (proteína constituyente de las uniones gap). NGF presenta la capacidad de inducir
una rápida fosforilación de la connexina-43 en células tecales (Mayerhofer A y col, 1996). En
ratones deficientes para el gen de NGF, se encontró la existencia de una reducción de folículos
primarios y secundarios, y de un aumento de ovocitos desnudos; encontrándose además que
NGF era importante en la proliferación de células de la teca y granulosa del ovario, visto por
ensayos de PCNA y BrdU (Dissen y col, 2001). Estos procesos ocurren en forma independiente
de las gonadotrofinas, donde niveles séricos de FSH y LH no se ven afectados en ratones
deficientes para el gen de NGF, todo esto sugiere que NGF estaría jugando un rol crucial en la
regulación del desarrollo de la función reproductiva (Dissen y col, 2001). También se ha
demostrado que las células de granulosa humana in vitro presentan la capacidad de responder a
NGF, regulando la esteroidogénesis (inhibiendo la luteinización in vitro) y aumentando la
expresión de receptores para FSH (Salas C y col, 2006). Lo interesante es que tanto el receptor

11
trkA, como su ligando NGF se expresan en las células de la granulosa de folículos pre-antrales y
antrales; por lo tanto NGF estaría involucrado en la esteroidogénesis ovárica, por una acción
autocrina (Salas C y col, 2006). Por otro lado, en este tipo celular, NGF también puede promover
la proliferación celular, visto por el método de Cell Titer (Salas C, 2002).
Se ha observado que NGF promueve la angiogénesis como parte de la cicatrización de
heridas (Graini G y col, 2004), así como también que participa en la capilarización gatillada por
isquemia (Emanueli C y col, 2002).
La angiogénesis es la formación de nuevos vasos sanguíneos a partir de los ya
existentes, es un proceso controlado que se lleva a cabo durante el ciclo menstrual femenino y en
la reparación de heridas. En un estado patológico, la angiogénesis ocurre en muchas
enfermedades, entre las cuales se pueden encontrar la retinopatía diabética, la artritis, progresión
tumoral entre otras. (Sullivan y col, 2003).
VEGF es un potente mitogéno del endotelio vascular, es una glico-proteína
homodimérica de 45 kda, (Geva E y Jaffe RB, 2000) que se une específicamente a receptores de
células endoteliales provocando proliferación. (Shibuya M, 2001). VEGF juega un rol crítico en
la función ovárica, debido a que induce la angiogénesis, que acompaña al desarrollo folicular y
del cuerpo lúteo (Redmer y col, 1996).
Nuestro grupo ha encontrado que NGF aumenta la expresión de VEGF en células de
granulosa humana en cultivo. Los niveles del ARNm de VEGF fueron evaluados por PCR
convencional, así como también por PCR de tiempo real; la proteína VEGF fue determinada en
los medios de cultivo por Elisa. En mamíferos existen 5 isoformas de VEGF, los cuales son
productos de splicing alternativo del gen de VEGF, estas isoformas son VEGF 121, 145, 165,
189 y 206. La isoforma 165 es la predominante, y junto con las isoformas 121 y 189 son las que
se encuentran en la mayoría de los tejidos y células (Geva E y Jaffe RB, 2000). En ovario normal
y cáncer de ovario las isoformas que se expresan mayoritariamente son las isoformas 121 y 165
(Fujimoto J y col, 1998); sin embargo, en cáncer ovárico epitelial se ha encontrado un aumento
significativo de las tres isoformas de VEGF (121, 165 y 189) por efecto de NGF (Campos X y
col, 2006).

12
La expresión de VEGF es regulada por el factor transcripcional inducible por hipoxia
(HIF-1), (Derý C y col, 2005; Semenza L, 2002) regulador esencial de la homeostasis del
oxígeno. HIF-1 está compuesto por 2 subunidades, HIF-1 alfa y HIF-1 beta. HIF-1 beta es la
subunidad que se expresa constitutivamente, en cambio el HIF-1 alfa es la subunidad inducible
(Semenza L, 2002). Como resultado de la deprivación de O2 (hipoxia), el crecimiento y la
viabilidad celular es reducida, es allí donde este factor es importante, porque se expresa en
condiciones de baja concentración de oxígeno, activando a más de 60 genes, implicados en
diferentes funciones celulares, tales como la angiogénesis, proliferación celular y sobrevida
celular. El aumento de la expresión de HIF-1 alfa, también puede producirse por efecto de
factores de crecimiento (Derý C y col, 2005).
c-Myc es un proto-oncogen, que media la transición de la fase G0-G1 y G1-S del ciclo
celular (Dang C, 1999). Mediante la técnica de hibridación in-situ, c-Myc se encontró en todas
las etapas del desarrollo folicular, menos en la etapa de folículo primordial, además se encontró
en la etapa de cuerpo lúteo. Estos datos indicarían, que la expresión de c-Myc podría jugar un rol
importante en los mecanismos moleculares de proliferación y apoptosis de los folículos
(Putowski y col, 1997); así como también en los procesos de angiogénesis durante la ciclicidad
del ovario, ya que coinciden con la expresión de VEGF en el mismo tejido (Fraser H, 2001) Se
ha encontrado que la sobreexpresión de c-Myc, promueve el crecimiento de vasos sanguíneos,
debido en parte a una inducción de VEGF. El mecanismo por el cual, c-Myc, controla la
expresión de factores angiogénicos, aún no está claro, sin embargo la regulación sobre VEGF
parece ser indirecta (Baudino T y col, 2002).
Ki 67 es una proteína, que esta estrictamente asociada a la proliferación celular, debido
a que esta presente en todas las etapas del ciclo celular (G1-S-G2-M) y ausente en G0 (Scholzen
T, 2000).
Con el objeto de esclarecer el mecanismo por el cual NGF actúa en el ovario, ya que se
ha demostrado que NGF, estaría participando en la esteroidogénesis, y en la regulación de la
expresión del receptor de FSH, y dado que NGF y su receptor trkA se expresan principalmente
en células de la granulosa de folículos pre-antrales como antrales (Salas C, y col 2006) al igual
que VEGF en ovario humano adulto, y debido a que NGF, en otros tejidos puede regular la
expresión de VEGF, en este estudio se evalúo el efecto de NGF sobre la regulación de VEGF y

13
la proliferación en células de granulosa humana, así como también se evalúo algunos factores de
transcripción, tales como HIF-1 alfa y c-Myc, involucrados en ambos procesos.

14
Resumiendo
• NGF es capaz de aumentar tanto los niveles de ARNm, como de la proteína de VEGF en
células de granulosa humana.
• NGF participa en la proliferación celular de las células somáticas del ovario.
• c-Myc es un gen que esta involucrado en la proliferación celular.
• Una molécula de señalización importante en la regulación de VEGF es HIF-1alfa.
De acuerdo a estos antecedentes, NGF estaría jugando un papel importante, en el
desarrollo folicular y por ende en la función de esta gónada,
Acorde a lo escrito anteriormente, se puede proponer la siguiente hipótesis:

15
HIPOTESIS
“El factor de crecimiento nervioso (NGF) regula la proliferación celular y
los niveles de ARNm de VEGF en células de granulosa humana en cultivo”.

16
I OBJETIVOS
I. 1 Objetivo General
Evaluar la acción de NGF en la expresión de VEGF y la proliferación de células de
granulosa humana a distintos tiempos de cultivo.
I. 2 Objetivos Específicos
I.2.1. Evaluar los niveles del ARNm de las isoformas de VEGF ( VEGF 121, VEGF 165
y VEGF 189) y los factores de transcripción HIF-1 alfa y c-Myc en células de granulosa
humana a las 2, 4, 8 y 18 horas de cultivo.
I.2.2. Determinación del efecto de NGF en la proliferación celular mediante la
cuantificación de Ki67 por Inmunocitoquímica, a distintos tiempos de cultivo.

17
II MATERIALES Y MÉTODOS
II.1 Obtención de muestras.
II.1.1 Células de granulosa.
Las células de granulosa humana fueron obtenidas, de aspirados de folículos
preovulatorios, de un total de 40 pacientes del programa de fertilización in vitro del Hospital
Clínico de la Universidad de Chile. Este trabajo contó con la aprobación del Comité de Ética de
la Institución y consentimiento informado de las pacientes.
II.1.2 Aislación y purificación de células de granulosa humana.
Los aspirados de los diferentes folículos de cada paciente, se centrifugaron a 1.200g por
7 minutos. Los precipitados contenían las células de granulosa humana, las células se purificaron
en una gradiente de Percoll al 50% (3 ml de Percoll y 3 ml de medio HAM F12 Dubelco (Sigma)
(bicarbonato de Sodio, 80 mg/L de gentamicina, 50 mg/L de penicilina G sódica (1000000 UI),
50 mg /L de estreptomicina, 5 mg/L de ketocononazol, pH 7,2-74), se centrifugaron por 45
minutos a 1.375g, con el fin de separar las células de granulosa de los glóbulos rojos.
Posteriormente las células, fueron llevadas a otro tubo que contenía, 1 ml de medio HAM F12
Dubelco, donde fueron resuspendidas (lavadas) y disgregadas, y sometidas a una centrifugación
de 1200g por 7 minutos. Luego los precipitados se resuspendieron en 3 ml de medio HAM F12
Dubelco libre de suero. Se realizó nuevamente una centrifugación a 1.200g por 7 minutos, el
precipitado correspondiente a las células de granulosa humana fueron resuspendidas en 1 ml de
medio HAM F12 Dubelco libre de suero. Las células fueron sometidas a un test de viabilidad
con Azul de Tripán (Sigma) (Tennant J. R, 1964). Para este propósito se realizó una solución
(1:1) de suspensión de células y Azul de Tripán (0.4%(p/v), la mezcla se incubó durante 5
minutos y se realizó el conteo de las células muertas y vivas en una cámara Neubauer con
microscopio óptico con aumento de 40X.

18
II.1.3 Cultivo de células de granulosa.
Las células de granulosa humana, fueron cultivadas en placas de 6 pocillos de 35 mm.
Los pocillos fueron recubiertos con gelatina al 1%, posteriormente se cultivaron 500.000 células
vivas por pocillo, en 3 ml de medio HAM F12 Dubelco libre de suero. Estas placas fueron
cultivadas por 18 horas a 37 ° C, en una estufa de cultivo, con un 100% de humedad, 95 % aire y
5 % de CO2. Después de 18 horas, se cambió el medio nuevamente por medio HAM F12, y se
agregó NGF en una concentración de 50 (ng/ml), obteniéndose células Control (sin NGF) y
estimuladas con (NGF), las células de granulosa fueron cultivadas por 2, 4, 8 y 18 horas. Al
término del cultivo, las células se resuspendieron en 1 ml de Trizol (Invitrogen), a continuación
fueron guardadas a –80° C para su posterior extracción de ARN total.
II.2. RT-PCR
II.2.1 Extracción de ARN.
El ARN total, fue extraído de 500.000 células de granulosa humana, para así obtener una
cantidad de ADN complementario (ADNc) óptimo para poder efectuar los análisis posteriores de
amplificación de este ADNc.
Al término del cultivo celular, a las células se les agregó un ml de Trizol (Invitrogen). Se
agregaron 0,2 ml de cloroformo y se mezcló por 10 segundos en vortex. Luego las muestras se
dejaron en reposo por 15 minutos a temperatura ambiente. Posteriormente, se centrifugaron a
12.000g por 15 minutos a 4° C. Se retiró la fase acuosa (fase superior) y se transfirió a un tubo
nuevo (400 ul aprox). Se agregó el mismo volumen de isopropanol, se agitó en vortex (hielo). Se
dejó precipitando el ARN toda la noche a –20° C. Al otro día, trabajando en hielo, se centrifugó
a 12.000g por 10 minutos a 4° C; se eliminó el sobrenadante por inmersión. Al precipitado que
contenía el ARN se le agregó 1 ml de etanol al 70%, esto con el fin de eliminar todo el
isopropanol restante y luego se centrifugó por cinco minutos a 12.000g a 4° C. El precipitado se
secó al aire por 5 minutos, el cual fue resuspendido en buffer TE (Tris-EDTA) 1x (15 ul). La
concentración y pureza del ARN se midió en un espectrofotómetro, a 260 nm y a 280 nm.

19
II.2.2. RT.
Para la realización del RT se usó 1 ug de ARN total. Se realizaron 2 mezclas. La primera
contenía la muestra, H20 DEPC, 0,5 ul de random hexamer Primers y 1 ul de dNTPs (10mM). La
primera mezcla se calentó por 5 minutos a 70° C. La segunda mezcla contenía 4 ul de buffer de
reacción 5X (250 mM Tris-HCl pH 8,3, 375 mM KCl y 15 mM MgCl2), 2 ul de DTT, 1 ul de
inhibidor de ribonucleasas y 1 ul de la enzima de transcriptasa reversa M-MLV (Invitrogen).
La mezcla 1 y 2 se juntaron y se incubaron por 1 hora a 37° C, deteniéndose la reacción por
congelación. Esta correspondió a la solución stock de ADNc.
Para la reacción de PCR semicuantitativo se tomaron 1 y 2 ul de muestra (Tabla 1)
dependiendo del gen a amplificar, por la reacción de la polimerasa en cadena (PCR), en 25 ul
totales de mezcla que contenían 0.15 ul de Taq Polimerasa (Biotools), 2.5 ul buffer 10X , 1 ul de
MgCl2, 0,5 ul de dNTP común para todos los PCR, y para cada PCR específico se agregaron
12,5 pmoles de partidores sentido y antisentido de VEGF, HIF-1 alfa y trkA, y 10 pmoles para
el gen c-Myc. Como gen constitutivo se usaron 12.5 pmoles de ß-actina. Para la ß-actina se
usaron 2 pares de partidores diferentes de 260 y 661 pb.
Los tubos con la mezcla de reacción se llevaron a un termociclador, para realizar la
reacción en cadena de la polimerasa. Los productos de PCR obtenidos fueron separados
en un gel de agarosa al 2 %, (TBE 1x) que contiene bromuro de etidio (1mg/ml). Las
bandas fueron semicuantificadas con un programa computacional USI (Un-Scan-it). Los
resultados de los niveles de ARNm se obtuvieron, calculando las unidades relativas,
correspondiente a la relación de los pixeles (gen especifico / ß-actina).
En la tabla 1 y 2 se detallan las secuencias específicas de los distintos partidores usados y los
programas para cada PCR.
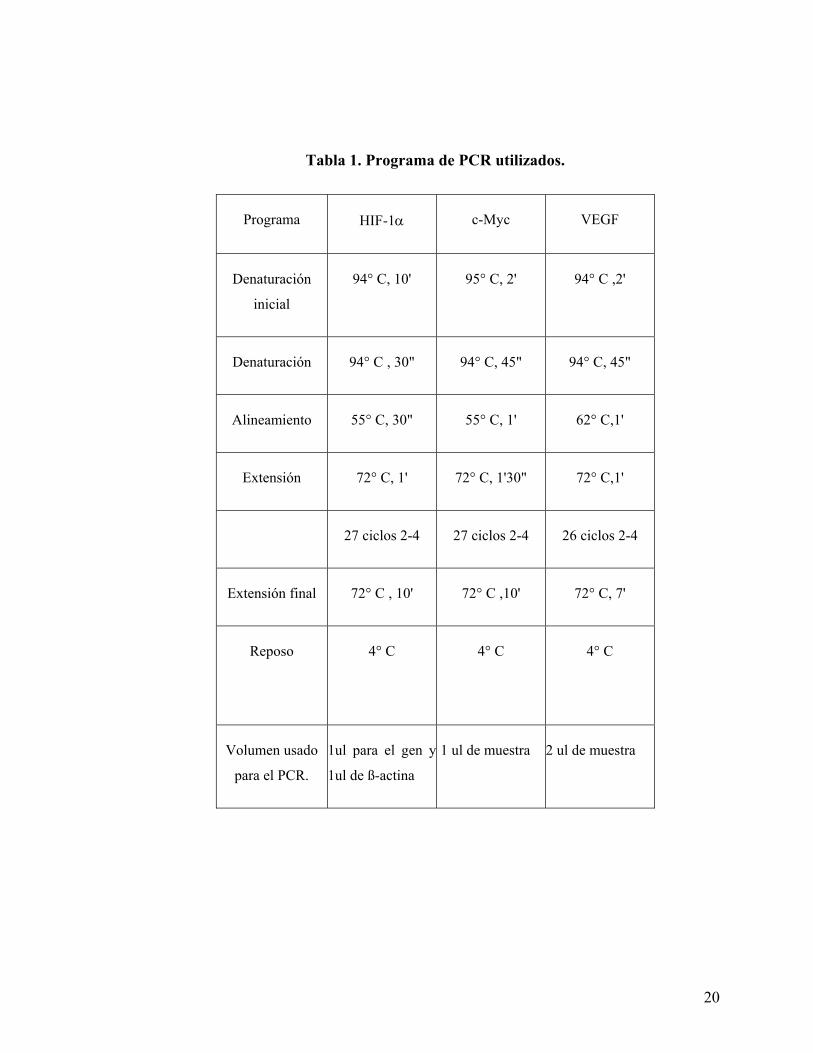
20
Tabla 1. Programa de PCR utilizados.
Programa HIF-1α c-Myc VEGF
Denaturación
inicial
94° C, 10' 95° C, 2' 94° C ,2'
Denaturación 94° C , 30" 94° C, 45" 94° C, 45"
Alineamiento 55° C, 30" 55° C, 1' 62° C,1'
Extensión 72° C, 1' 72° C, 1'30" 72° C,1'
27 ciclos 2-4 27 ciclos 2-4 26 ciclos 2-4
Extensión final 72° C , 10' 72° C ,10' 72° C, 7'
Reposo
4° C
4° C 4° C
Volumen usado
para el PCR.
1ul para el gen y
1ul de ß-actina
1 ul de muestra 2 ul de muestra
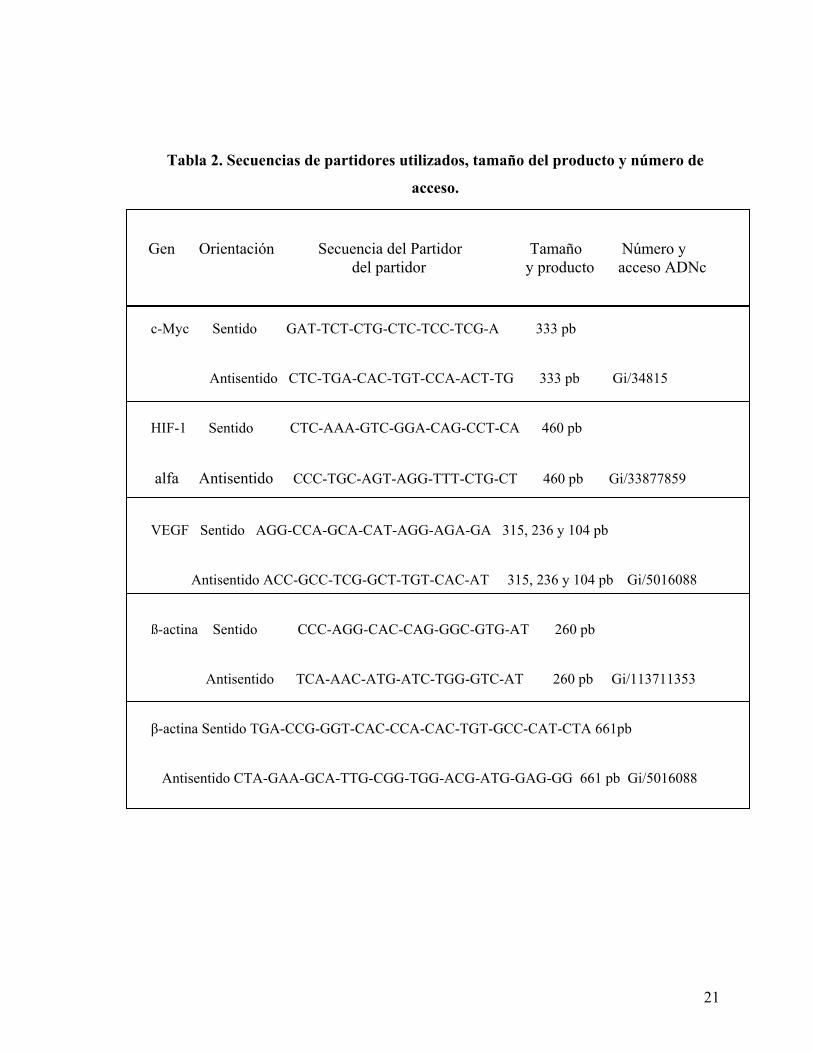
21
Tabla 2. Secuencias de partidores utilizados, tamaño del producto y número de
acceso.
c-Myc Sentido GAT-TCT-CTG-CTC-TCC-TCG-A 333 pb
Antisentido CTC-TGA-CAC-TGT-CCA-ACT-TG 333 pb Gi/34815
HIF-1 Sentido CTC-AAA-GTC-GGA-CAG-CCT-CA 460 pb
alfa Antisentido CCC-TGC-AGT-AGG-TTT-CTG-CT 460 pb Gi/33877859
VEGF Sentido AGG-CCA-GCA-CAT-AGG-AGA-GA 315, 236 y 104 pb
Antisentido ACC-GCC-TCG-GCT-TGT-CAC-AT 315, 236 y 104 pb Gi/5016088
ß-actina Sentido CCC-AGG-CAC-CAG-GGC-GTG-AT 260 pb
Antisentido TCA-AAC-ATG-ATC-TGG-GTC-AT 260 pb Gi/113711353
β-actina Sentido TGA-CCG-GGT-CAC-CCA-CAC-TGT-GCC-CAT-CTA 661pb
Antisentido CTA-GAA-GCA-TTG-CGG-TGG-ACG-ATG-GAG-GG 661 pb Gi/5016088
Gen Orientación Secuencia del Partidor Tamaño Número y del partidor y producto acceso ADNc

22
II.3. Inmunocitoquímica.
Se cultivaron 100.000 células de granulosa humana, provenientes de 11 pacientes, por 4,
8 y 18 horas en condiciones Control (sin NGF) y NGF (50 ng/ml), con un cultivo previo de 18
horas con solo medio HAM-F12 libre de suero en placas de 24 pocillos, cada muestra se realizó
en duplicado para un tiempo determinado. Posteriormente, las células se desprendieron por
pipeteo suave (mecánicamente), a continuación las células fueron esparcidas en portaobjetos,
donde posteriormente fueron fijadas con Metanol (Merck) por 15 minutos a 4° C, luego se lavó
el exceso de Metanol con PBS 1x (8 gr/L NaCl; 0,2 gr/L KCl; 1,44 gr/L Na2HPO4, 0,24 gr/L
KH2PO4; pH 7,4) por 5 minutos a 4° C, dejando finalmente las células en PBS-Azida 15 mM. Se
realizó la inmunocitoquímica para determinar la presencia de la proteína Ki67, para este fin las
células fueron lavadas 3 veces con PBS 1x, después se incubaron con perhidrol al 2% por 30
minutos a temperatura ambiente, con el fin de bloquear la actividad de peroxidasas endógenas,
luego las células se lavaron 3 veces con PBS 1x.
Para evitar la unión inespecífica del primer anticuerpo, las células fueron incubadas con
PBS-Leche durante 30 minutos a temperatura ambiente, sin lavar se colocó el primer anticuerpo
policlonal Ki67 Ab-3 (Lab Vision Corporation) en una dilución 1:100 en PBS-BSA 1%. Al otro
día se lavó 4 veces con PBS 1X por 2 minutos, posteriormente se aplicó el anticuerpo secundario
hecho en cabra biotinilado anti-conejo durante 10 minutos a temperatura ambiente, seguido de la
aplicación de Estreptavidina Peroxidasa por 10 minutos a temperatura ambiente, se lavó 4 veces
con PBS 1X. Finalmente se agregó a los portaobjetos el preparado (1 gota (40ul) de Dab
Cromógeno a 2 ml de sustrato Dab), como contratinción se usó Hematoxilina. Se contaron las
células positivas y las células negativas en un microscopio óptico con un aumento de 40X. Para
la expresión de los resultados se calcularon los porcentajes de células positivas Control y NGF
con respecto al total de las células (células positivas y negativas).

23
II.4.Análisis Estadístico
Tanto para la medición de los niveles de ARNm, como la inmunocitoquímica, se utilizó el
test de Student pareado, (Prueba de T) para la comparación de células Control con células
tratadas con NGF. Se consideró significativo valores de p<0,05.

24
III RESULTADOS
III.1. Determinación de los niveles de ARNm de VEGF por acción de
NGF 50 ng/ml.
En ovario humano normal se han detectado tres de las 5 isoformas de VEGF (VEGF
121, VEGF 165 y VEGF 189), por lo tanto, se determinaron los niveles del ARNm de VEGF de
las 3 isoformas, VEGF 189, VEGF 165 y VEGF 121, en 38 pacientes provenientes del programa
de fertilización asistida, en distintas condiciones de cultivo (2, 4, 8 y 18 horas), tanto para células
control y células estimuladas con NGF (50ng/ml). En algunos casos, solo se logró cuantificar la
expresión del ARNm de VEGF 165 y VEGF 121. Esto es, porque la isoforma de VEGF 189 en
algunas muestras se expresó muy poco. En la figura 1 se muestra un gel de agarosa
representativo de las muestras de PCR para las isoformas de VEGF.
L C N C N C N C N C(-)
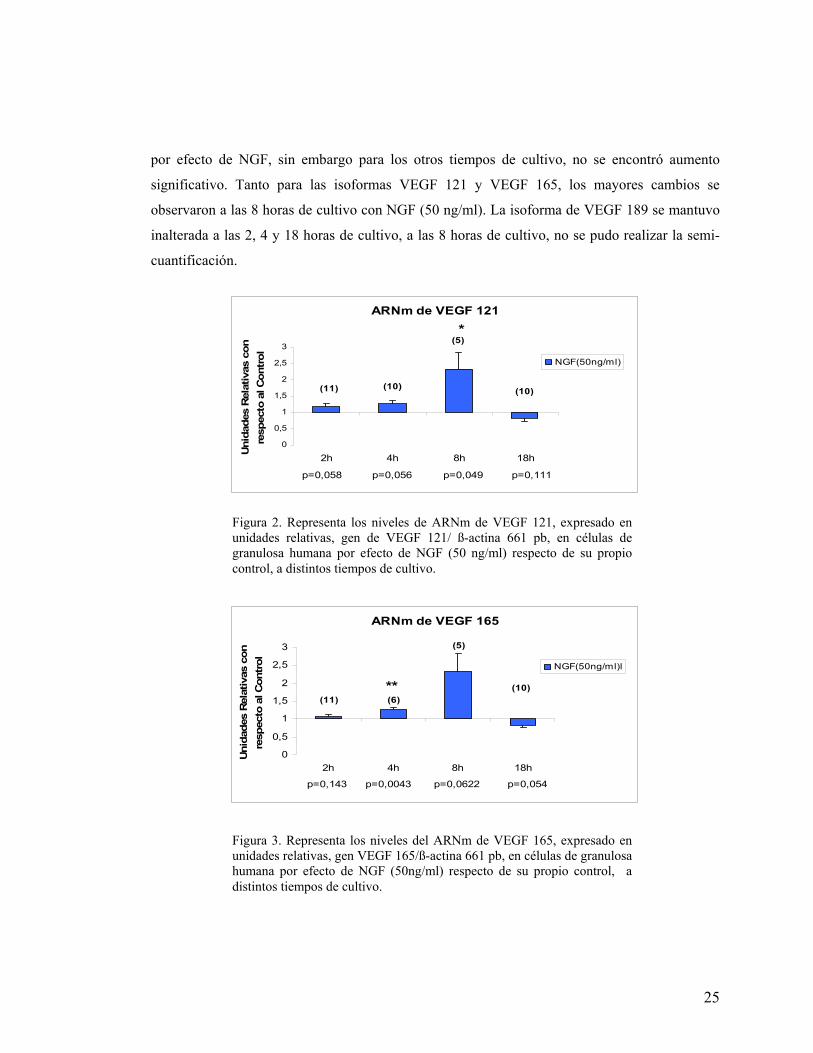
La figura 2, muestra un aumento significativo (*p<0,05) de la isoforma de VEGF 121,
por efecto de NGF a las 8 horas de cultivo, en tanto que a las 2 y 4 horas de estímulo, se observa
un incremento casi significativo (p=0,058 y p=0,052 respectivamente). Para la isoforma de
VEGF 165 (Figura 3), se observa a las 4 horas de cultivo, un aumento significativo (**p<0,01)
VEGF 189 308 pb VEGF 165 236 pb VEGF 121 104 pb
ß-actina 661 pb
Figura 1. RT-PCR Representativo de VEGF. Muestras Control (C) y Tratadas con NGF 50 ng/ml (N). La figura muestra los productos de PCR a 661, 308, 236, 104 pb, correspondientes a β-actina, VEGF 189, VEGF 165 y VEGF 121 L: corresponde al ladder y C (-) corresponde al control negativo.
25
por efecto de NGF, sin embargo para los otros tiempos de cultivo, no se encontró aumento
significativo. Tanto para las isoformas VEGF 121 y VEGF 165, los mayores cambios se
observaron a las 8 horas de cultivo con NGF (50 ng/ml). La isoforma de VEGF 189 se mantuvo
inalterada a las 2, 4 y 18 horas de cultivo, a las 8 horas de cultivo, no se pudo realizar la semi-
cuantificación.
ARNm de VEGF 121
(10)
(5)
(10)(11)
0
0,5
1
1,5
2
2,5
3
2h 4h 8h 18h
Uni
dade
s R
elat
ivas
con
re
spec
to a
l Con
trol NGF(50ng/ml)
p=0,058 p=0,056 p=0,049 p=0,111
*
ARNm de VEGF 165
(10)(11) (6)
(5)
0
0,5
1
1,5
2
2,5
3
2h 4h 8h 18h
Uni
dade
s R
elat
ivas
con
re
spec
to a
l Con
trol NGF(50ng/ml)l
p=0,143 p=0,0043
**
p=0,0622 p=0,054
Figura 2. Representa los niveles de ARNm de VEGF 121, expresado en unidades relativas, gen de VEGF 121/ ß-actina 661 pb, en células de granulosa humana por efecto de NGF (50 ng/ml) respecto de su propio control, a distintos tiempos de cultivo.
Figura 3. Representa los niveles del ARNm de VEGF 165, expresado en unidades relativas, gen VEGF 165/ß-actina 661 pb, en células de granulosa humana por efecto de NGF (50ng/ml) respecto de su propio control, a distintos tiempos de cultivo.

26
ARNm de VEGF 189
(10) (10) (7)
0
0,5
1
1,5
2
2,5
3
2h 4h 18h Uni
dade
s R
elat
ivas
con
re
spec
to a
l Con
trol
NGF (50ng/ml)
p=0.759 p=0,52 p=0,054
Figura 4. Representa los niveles del ARNm de VEGF 189, expresado en unidades relativas, gen de VEGF 189/ß-actina 661 pb, por efecto de NGF (50ng/ml), respecto de su propio control, a distintos tiempos de cultivo.

27
III.2. Determinación de los niveles de ARNm de HIF-1 alfa en células
de granulosa humana en situación control y por efecto de NGF
50ng/ml.
Debido a la existencia de varias evidencias, que demuestran que HIF-1 alfa participa en
la regulación de la expresión de VEGF, y que la expresión de HIF-1 alfa puede ser inducida por
factores de crecimiento (Derý C y col, 2005; Semenza L, 2002), se analizaron los niveles del
ARNm de HIF-1 alfa por efecto de NGF. Las células de granulosa humana, provenientes de 36
pacientes del programa de fertilización asistida, fueron cultivadas a distintos tiempos (2, 4, 8 y
18 horas), en ausencia y presencia de NGF (50 ng/ml).
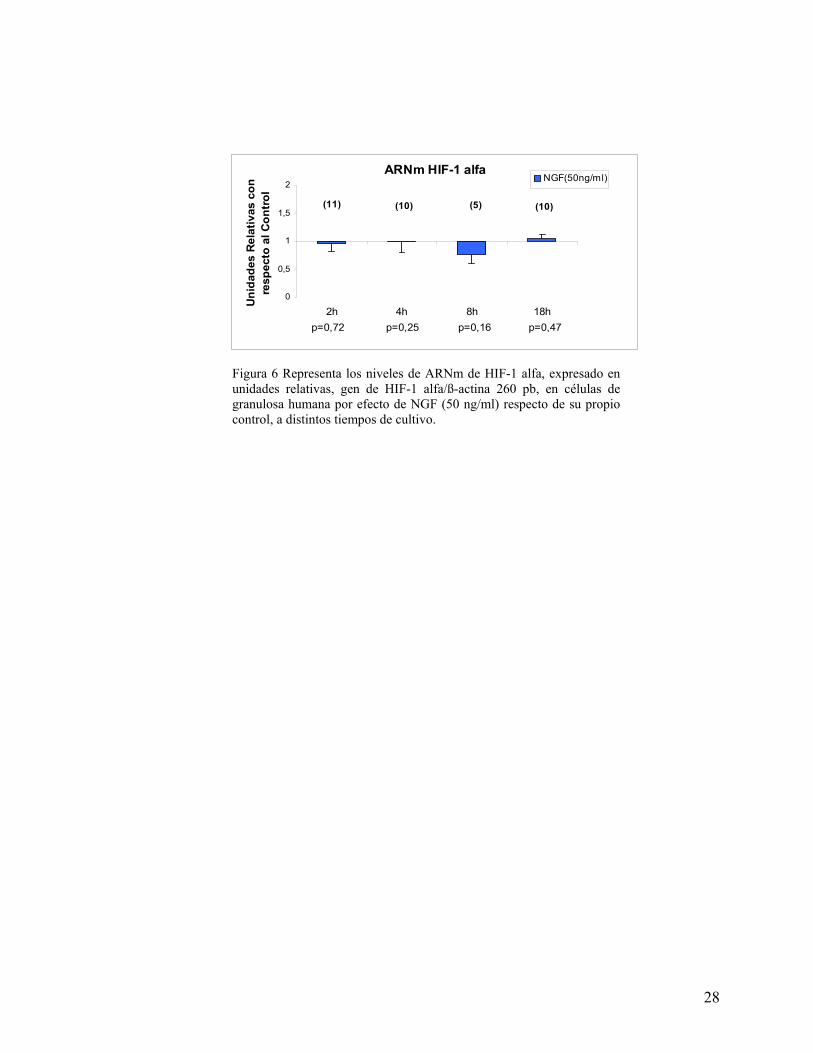
La figura 6 muestra que los niveles de ARNm de HIF-1alfa, no cambian por efecto de
NGF en las condiciones estudiadas (no hipóxicas).
Figura 5. RT-PCR representativo de HIF-1alfa y β-actina. L: ladder; C:control, N: tratadas con NGF; C(-): Control negativo
L C N C N C N C N C (-)
HIF-1 alfa 460 pb
ß-actina 260 pb
28
ARNm HIF-1 alfa
(10)(10)(11) (5)
0
0,5
1
1,5
2
2h 4h 8h 18h
Uni
dade
s R
elat
ivas
con
re
spec
to a
l Con
trol
NGF(50ng/ml)
p=0,72 p=0,25 p=0,16 p=0,47
Figura 6 Representa los niveles de ARNm de HIF-1 alfa, expresado en unidades relativas, gen de HIF-1 alfa/ß-actina 260 pb, en células de granulosa humana por efecto de NGF (50 ng/ml) respecto de su propio control, a distintos tiempos de cultivo.

29
III. 3. Estudio de la acción de NGF (50ng/ml), sobre los niveles de ARNm
de c-Myc en células de granulosa humana.
Se evaluaron los niveles de ARNm de c-Myc a las 2, 4, 8 y 18 horas de cultivo, con el
fin de determinar la participación de este gen en la ruta de señalización de NGF, ya que hay
antecedentes que indican que c-Myc puede ser un gen involucrado en la regulación de la
expresión de VEGF (Baudino T y col, 2002) y en la proliferación celular (Dang C, 1999). Por lo
tanto, era importante evaluar, si NGF regularía su expresión en células de granulosa humana.
Para realizar estos experimentos se utilizaron células de granulosa provenientes de 39 pacientes.
La figura 7 corresponde a un RT-PCR representativo del gen c-Myc, se puede observar la
presencia de 2 bandas, una de aproximadamente 333 pb, que correspondería a c-Myc y otra
banda que correspondería a la β-actina 661 pb.
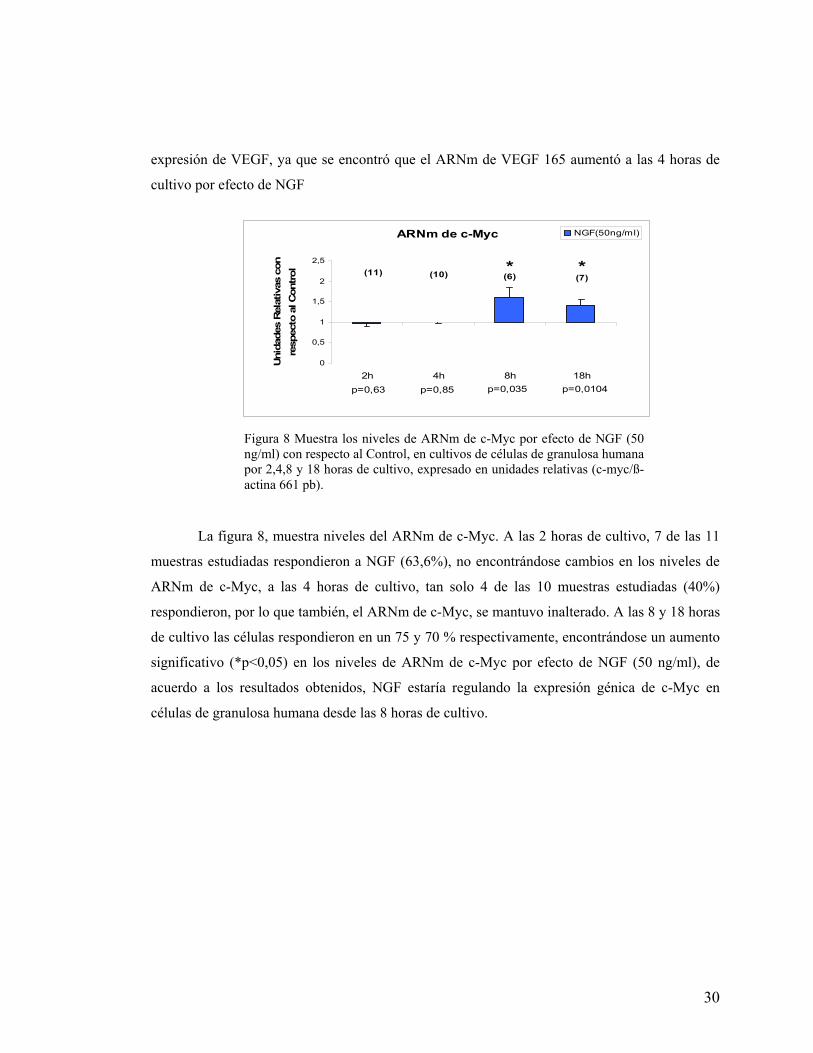
Se encontró, que los niveles del ARNm de c-Myc aumentaron a las 8 y 18 horas de
cultivo, por efecto de NGF (50ng/ml), y que a las 2 y 4 horas de cultivo, no presentó cambios
por efecto de NGF. Al ser c-Myc un gen que actúa en la transición G1-S del ciclo celular, estos
datos indicarían que NGF estaría regulando la expresión de c-Myc, y que este gen podría estar
involucrado, mas bien en la regulación de la proliferación celular, que en la regulación de la
Figura 7, RT-PCR Representativo del gen c-Myc (333 pb), ß-actina (661 pb). Muestras Control (C) y Tratadas con NGF (N). L: corresponde al ladder y C (-) corresponde al control negativo
661 pb
Ladder C N C N C N C N C(-)
333 pb
30
expresión de VEGF, ya que se encontró que el ARNm de VEGF 165 aumentó a las 4 horas de
cultivo por efecto de NGF
ARNm de c-Myc
(7)(6)(10)(11)
0
0,5
1
1,5
2
2,5
2h 4h 8h 18h
Uni
dade
s R
elat
ivas
con
re
spec
to a
l Con
trol
NGF(50ng/ml)
* *
p=0,63 p=0,85 p=0,035 p=0,0104
La figura 8, muestra niveles del ARNm de c-Myc. A las 2 horas de cultivo, 7 de las 11
muestras estudiadas respondieron a NGF (63,6%), no encontrándose cambios en los niveles de
ARNm de c-Myc, a las 4 horas de cultivo, tan solo 4 de las 10 muestras estudiadas (40%)
respondieron, por lo que también, el ARNm de c-Myc, se mantuvo inalterado. A las 8 y 18 horas
de cultivo las células respondieron en un 75 y 70 % respectivamente, encontrándose un aumento
significativo (*p<0,05) en los niveles de ARNm de c-Myc por efecto de NGF (50 ng/ml), de
acuerdo a los resultados obtenidos, NGF estaría regulando la expresión génica de c-Myc en
células de granulosa humana desde las 8 horas de cultivo.
Figura 8 Muestra los niveles de ARNm de c-Myc por efecto de NGF (50 ng/ml) con respecto al Control, en cultivos de células de granulosa humana por 2,4,8 y 18 horas de cultivo, expresado en unidades relativas (c-myc/ß-actina 661 pb).
31
III.5 Inmunocitoquímica de Ki67
Para confirmar los resultados de c-Myc, y así comprobar que NGF actúa promoviendo la
proliferación celular, se estudió un marcador de proliferación celular Ki67. Esta proteína esta
presente en todos las etapas del ciclo celular (G1-S-G2-M) y ausente en la etapa G0. Para este
propósito, fueron estimuladas 100.000 células Control y con NGF (50ng/ml) por 4, 8 y 18 horas.
Estas células fueron sometidas a una Inmunocitoquímica para Ki67. Para tal efecto se utilizaron
muestras de 11 pacientes.
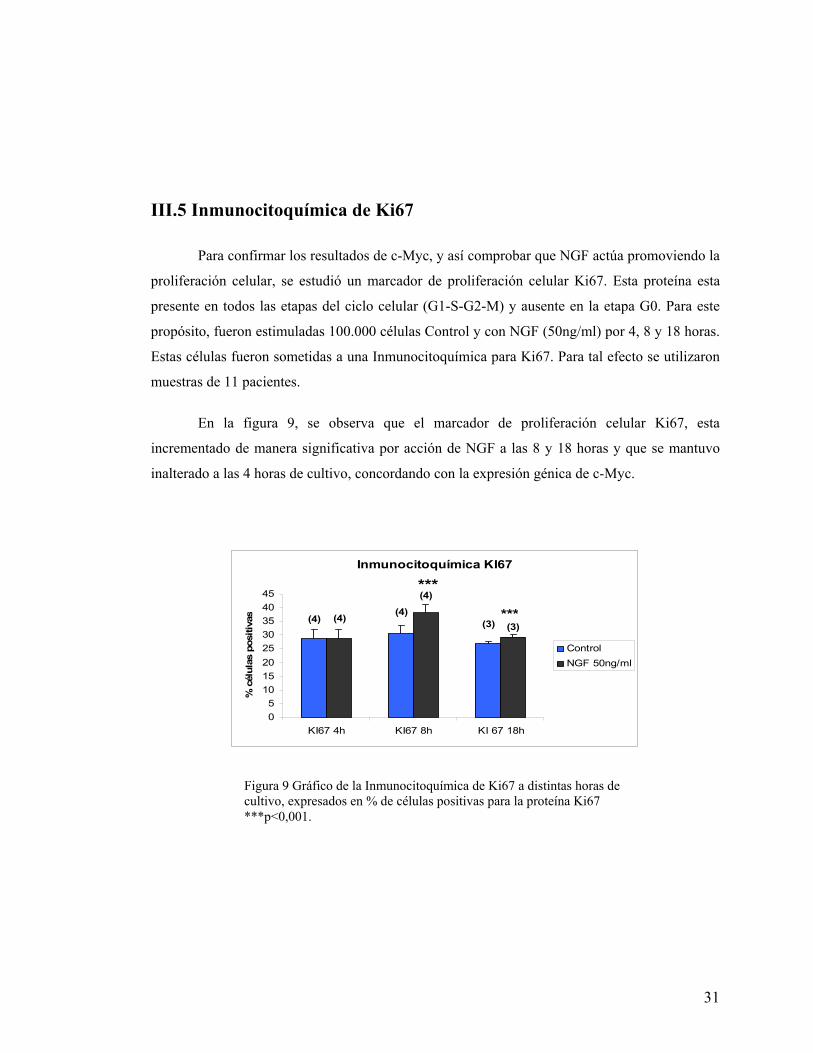
En la figura 9, se observa que el marcador de proliferación celular Ki67, esta
incrementado de manera significativa por acción de NGF a las 8 y 18 horas y que se mantuvo
inalterado a las 4 horas de cultivo, concordando con la expresión génica de c-Myc.
Inmunocitoquímica KI67
(3)(4)
(4)(3)
(4)
(4)
05
1015202530354045
KI67 4h KI67 8h KI 67 18h
% c
élul
as p
ositi
vas
ControlNGF 50ng/ml
***
***
Figura 9 Gráfico de la Inmunocitoquímica de Ki67 a distintas horas de cultivo, expresados en % de células positivas para la proteína Ki67 ***p<0,001.

32
IV DISCUSION
El factor de crecimiento nervioso (NGF) esta involucrado en la angiogénesis (Emanuelli C y
col, 2002; Calza y col, 2001), proceso importante en el desarrollo folicular y del cuerpo lúteo
(Redmer y col, 1996).
Una de las moléculas involucradas en la angiogénesis es VEGF (Redmer y col, 1996;
Ferrara N, 2001). Debido a que NGF induce la expresión de VEGF (Middeke M y col, 2002), y
estimula la proliferación celular, se evalúo el efecto de NGF en la regulación de estos procesos,
en células de granulosa humana en cultivo a diferentes tiempos, así como también se evalúo la
expresión génica de HIF-1 alfa y c-Myc, moléculas que intervienen en dichos procesos.
• NGF y su relación con VEGF y c-myc.
En este trabajo la expresión del ARNm de VEGF de las isoformas de VEGF 165 y 121
aumentaron a las 4 como a las 8 horas respectivamente, por efecto de NGF 50 ng/ml,
corroborando así con resultados preliminares, en los cuales en el modelo de granulosa humana
en cultivo, NGF aumenta tanto el ARNm como la proteína de VEGF 121 y 165 a las 8 horas de
estímulo con NGF 50 (ng/ml). Por otra parte, se observó un incremento muy pequeño del ARNm
de VEGF a las 2 horas de estímulo con NGF, no siendo significativo, lo que indicaría que NGF
estimula la transcripción de VEGF en forma temprana.
Estos resultados confirmarían el efecto de NGF en la regulación de la expresión de
VEGF y, por lo tanto se podría decir que NGF, estaría involucrado indirectamente en el
fenómeno de la angiogénesis. En un trabajo anterior se encontró que NGF (50ng/ml), inducía el
aumento del ARNm de VEGF a las 18 horas de cultivo (Muñoz Y, 2002). En el presente estudio
no se observó un aumento del ARNm de VEGF a las 18 horas de estimulo con NGF, esto podría
explicarse por diferencias en el grado de luteinización de las células de granulosa obtenidas para
el estudio.

33
NGF y la proliferación en células de granulosa humana.
La proliferación de las células de granulosa humana, es un fenómeno importante en la
diferenciación y maduración del folículo; en este estudio se midió la proliferación celular,
determinando los niveles de ARNm de c-Myc por RT-PCR, gen involucrado en procesos de
proliferación y evaluando la proteína Ki67, marcador de proliferación, por inmunocitoquímica;
encontrándose que tanto c-Myc como Ki67, aumentaron en células de granulosa humana a las 8
y 18 horas de cultivo por efecto de NGF 50 (ng/ml). La expresión de la proteína Ki67, está
estrictamente asociada a la proliferación celular, debido a que está presente en todas las etapas
activas del ciclo celular (G1, S, G2 y M) y ausente en G0 (Scholzen T, 2000). c-Myc es un
proto-oncogen, que esta implicado en las transiciones de G0/G1 y G1/S del ciclo celular. Al
eliminar ambos alelos de c-Myc por recombinación homologa, se provoca una marcada
prolongación del tiempo de duplicación en las células (Dang Chi V ,1999). Estos resultados
indicarían, que NGF estaría involucrado, en la proliferación de las células de granulosa humana,
y que esta proliferación podría ser mediada a través de c-Myc, interviniendo directamente en las
transiciones de las fases G0/G1 y G1/S. Este aumento de la proliferación, podría estar mediada
por el incremento directo o indirecto de c-Myc, para dilucidar si NGF induciría en forma directa
la expresión de c-Myc, se deberían realizar experimentos que inhiban directamente a NGF,
usando un anticuerpo contra NGF o usando un inhibidor del receptor de alta afinidad de trkA
(K252a).
En otro trabajo, al estimular células de granulosa humana con el factor de crecimiento
epidermal (EGF) y el factor de crecimiento de fibroblastos (FGF), dos factores de crecimiento,
disminuyeron los tiempos de duplicación de las células de 48 h a 16-20 horas respectivamente, y
por medición de incorporación de Timidina-[H], se midió que tanto EGF, como bFGF son
potentes mitógenos, debido a que estimulan fuertemente la iniciación de la síntesis de ADN
(Gospodarowicz D y col, 1977), lo que podría indicar que, NGF podría estar cumpliendo el
mismo rol que estos factores de crecimiento en la granulosa, disminuyendo el tiempo de
duplicación y aumentando la proliferación celular. NGF sería un regulador del crecimiento,
diferenciación y sobrevida de las células por su influencia sobre proteínas del ciclo celular
(Hughes AL y col, 2000).

34
VEGF e HIF-1 alfa.
Una molécula de señalización importante en la regulación de VEGF es HIF-1alfa. Los
resultados de HIF-1 alfa concuerdan con la literatura, debido a que los niveles de ARNm de
HIF-1 alfa, se mantienen inalterados al colocar, un estímulo del tipo factor de crecimiento, tal
como el factor IGF-1 (factor de crecimiento parecido a la insulina tipo I). En células HCT116
(células humanas de carcinoma de colon), por efecto de IGF-1, el ARNm de HIF-1 alfa no
cambia, pero si la expresión de su proteína, por el contrario los niveles del ARNm de VEGF
aumentan (Fukuda R y col, 2002), en base a estos antecedentes se podría suponer que NGF en
células de granulosa humana estaría provocando el mismo efecto sobre HIF-1 alfa, cambiando la
expresión de la proteína, no así la del ARNm. Es por esto, que se evalúo la proteína de HIF-1
alfa por la técnica de Inmunocitoquímica y Western Blot. Debido a problemas con el anticuerpo,
solo se consiguió ver una marca significativa en una sola muestra, donde se observó una mayor
tinción en las células de granulosa estimuladas con NGF a las 2 horas de cultivo, comparadas
con el Control (Datos no mostrados), es por esta razón que no se puede sacar una conclusión,
solo se puede decir que NGF podría estar estimulando este factor en una etapa inicial, y que este
factor a su vez actuaría sobre el promotor de VEGF activando su transcripción. Además de esto,
para observar el cambio en la expresión de HIF-1 alfa se deberían realizar experimentos en
condiciones de hipoxia, debido a que en condiciones no hipóxicas la proteína de HIF-1 alfa es
rápidamente degradada (Dery M y col, 2005). Debido a que en esta tesis se trabajó con cultivos
de dos y más horas, es muy posible no encontrar cambios en los niveles del ARNm de HIF-1
alfa, por lo tanto, en el futuro deberían realizarse experimentos a tiempos más cortos en
condiciones no-hipóxicas.

35
NGF, VEGF, c-Myc y proliferación celular
Según los resultados, se podría proponer que NGF sería una molécula que actuaría
aumentando tanto la expresión de VEGF, así como la proliferación de las células de granulosa en
cultivo en forma independiente, ya que el aumento de VEGF sería en una etapa anterior al de la
proliferación, esto es debido a que el incremento de los niveles de ARNm de la isoforma de
VEGF 165, se observó desde las 4 horas de cultivo, en cambio la proliferación aumentó desde
las 8 horas, visto tanto por el gen c-Myc como por el marcador de proliferación celular Ki67.
En un estudio, en el cual ratones fueron sometidos a una hipofisectomía y
posteriormente expuestos a PMSG, un análogo a LH (inductor de ovulación), y a un anticuerpo
contra uno de los receptores para VEGF (VEGFR2 ), al medir la proliferación en células de
granulosa por el método de cuantificación de Ki67, la proliferación disminuyó en aquellos
ratones expuestos al Ab anti-VEGF-R2 mas PMSG con respecto a los ratones expuestos
solamente a PMSG, esto indicaría que la activación de VEGF-R2 es requerida para la
proliferación de las células de granulosa (Zimmermann R y col, 2003), por lo tanto se podría
hipotetizar, que es necesario que haya una expresión de VEGF, para que se produzca
proliferación. VEGF es un potente mitógeno del endotelio vascular, que se une específicamente
a los receptores de las células endoteliales, induciendo proliferación (Shibuya M, 2001). En
consecuencia VEGF podría regular en forma autocrina, debido a la existencia de receptores
VEGFR-1 y VEGFR-2 en células de granulosa de granulosa luteinizadas (Toshiaki E y col,
2001). Todo este proceso podría en parte estar gobernado bajo la acción de NGF.
La relación existente entre c-Myc y VEGF no es clara, al parecer c-Myc regularía a
VEGF en forma indirecta (Baudino T y col, 2002), los resultados encontrados en este trabajo,
indicarían que: a).-NGF aumenta c-Myc y que este evento esta relacionado con el aumento de la
proliferación celular y b).- la expresión génica de VEGF precede a la de c-Myc, pero no se
puede descartar que a tiempos mayores de 8 horas, VEGF pudiera aumentar por el aumento de c-
Myc. Para evaluar de mejor forma, si NGF activa a VEGF y posteriormente a c-Myc y de esta
forma provocar proliferación celular, se podría usar un anticuerpo para VEGF, que actuaría
como un inhibidor de la acción de VEGF, con esto evaluar c-Myc y la proliferación celular
mediante el método de cuantificación de Ki67.

36
En conclusión NGF es una molécula que estaría involucrada, tanto en el aumento de los
niveles de ARNm de VEGF como en la proliferación de las células de granulosa, esto último
sería en una etapa posterior al aumento de VEGF.
Es probable, que el VEGF expresado en las células de granulosa, esté involucrado en la
proliferación de las células directamente, esta regulación autocrina por parte de VEGF sobre las
células, permitiría al folículo crecer y diferenciarse hasta lograr su maduración, ya que en un
modelo “in vivo”, se demostró que al administrar un anticuerpo truncado para VEGF, que actúa
como un inhibidor de la acción de VEGF, encontraron una disminución en la proliferación
celular del compartimiento de células de granulosa de los folículos pre-antrales y antrales (Wulff
C y col, 2001)
VEGF participaría en la proliferación y en la angiogénesis del folículo. Esta nueva red de
vasos sanguíneos, proveería un incremento del suplemento de gonadotropinas, oxígeno,
precursores de esteroides, además de otras sustancias para el crecimiento del folículo (Wullf C y
col, 2001). En base a esto se podría suponer que una deficiencia de parte del folículo de producir
NGF, podría llevar al folículo a una cascada de señales inhibitorias para su crecimiento y
provocar una alteración en el desarrollo folicular y en la fertilidad debido a que no ocurre una
eficiente angiogénesis, y el folículo no sería capaz de desarrollarse y crecer.

37
V CONCLUSIONES
• NGF aumenta la expresión génica de la isoforma de VEGF 121 a las 8 horas de
cultivo.
• NGF aumenta la expresión génica de la isoforma de VEGF 165 a las 4 horas de
cultivo.
• NGF no aumenta la expresión génica de la isoforma de VEGF 189 a ningún tiempo
estudiado.
• NGF aumenta dos marcadores de proliferación celular Ki67 y c-Myc en células de
granulosa humana a las 8 y 18 horas de cultivo.
• NGF no cambia la expresión génica de HIF-1 alfa en las condiciones estudiadas.
• NGF en forma independiente estaría implicado, tanto en el aumento de VEGF como
en la proliferación de las células de granulosa humana.
Estos resultados podrían apoyar la importancia de NGF como un regulador de la función
ovárica, no solo por su efecto en la esteroidogénesis, inducción de los receptores de FSH, sino
que también, NGF estaría participando en la proliferación celular de la granulosa ovárica, así
como también en la angiogénesis, proceso importante durante el desarrollo folicular y del cuerpo
lúteo.

38
VI REFERENCIAS
• Anderson R, Robinson L, Brooks J, Spears N. 2002. “Neurotrophins and their receptors
are expressed in the human fetal ovary”. The J. Of Clin. Endocr and Metab 87(2):890-
897.
• Baudino T, Mckay C, Pendeville-Samain H, Nilsson J, Maclean K, White E, Davis A,
Ihle J, Cleveland J. 2002. “c-Myc is essential for vasculogenesis and angiogenesis
during development and tumor progression”. Genes and Development 16: 2530-2543.
• Calza L, Giardino L, Giuliani A, Aloe L, Levi-Montalcini R. 2001 “Nerve growth
factor control of neuronal expression of angiogenetic and vasoactive factors”. PNAS
98(7):4160-4165.
• Campos X, Muñoz Y, Selman A, Yazigi R, Moyano L, Weinstein-Oppenheimer C, Lara
HE, Romero C. 2006. “Nerve growth factor and its high-affinity receptor trkA
participate in the control of vascular growth factor expression in epithelial ovarian
cancer”. Gynecol Oncol Aug 24 Epub ahead of print.
• Cantarella G, Lempereur L, Presta M Ribatti R, Lombardo G, Lazarovici P, Zappala
G, Pafumi C, Bernardini R. (2002). “Nerve growth factor– endothelial cell interaction
leads to angiogenesis in vitro and in vivo” .FASEB J. 10.1096/fj.01–1000fje.
• Dang C. 1999. “c-Myc Target genes involved in cell growth, apoptosis, and
metabolism”. Molecular and Cellular Biology 19(1):1-11.
• Déry C, Michaud D, Richard D. 2005 “Hypoxia-inducible factor 1: regulation by
hypoxic and non-hypoxic activators”. Int J Biochem Cell Biol 37:535-540.
• Dissen G, Hill D, Costa M, Les Dees W, Lara HE, Ojeda S. 1996. “A role for trkA nerve
growth factor in mammalian ovulation”. Endocrinology 137(1):198-209.

39
• Dissen GA, Hirshfield A, Malamed S, Ojeda SR. 1995. “Expression of neurotrophins
and their receptors in the mammalian ovary is developmentally regulated changes at the
time of folliculogenesis”. Endocrinology 136(10):4681-4692.
• Dissen GA, Parrot JA, Skinner MK, Hill DF, Ojeda SR. 2000. “Direct effects of nerve
growth factor on thecal cells from antral ovarian follicles”. Endocrinology 141(12):
4736-4750.
• Dissen G, Romero C, Hirshfield A, Ojeda S. 2001. “Nerve growth factor is required for
early follicular development in the mammalian ovary”. Endocrinology 142(5): 2078-
2086.
• Dissen G, Romero C, Paredes A, Ojeda S. 2002. “Neurotrophic control of ovarian
development”. Microscopy Research and Technique 59:509-515.
• Emanueli C, Salis M, Pinna A, Graiani G, Manni L, Madeddu P. 2002. “Nerve growth
factor promotes angiogenesis and arteriogenesis in ischemic hindlimbs”. Circulation
106: 106:2257-2262.
• Ferrara N 2001.”Role of vacular endothelial growth factor in regulation of physiological
angiogenesis”. Am J Physiol Cell Physiol 280:C1358- C1366.
• Fraser HM, Wulff C 2001. “Angiogenesis in the primate ovary”. Reprod Fertil Dev 13,
557–566.
• Fujimoto J, Sajaguchi H, Hirose R, Ichigo S, Tamaya T. 1998. “Biologic implications of
the expression of vascular endothelial growth factor subtypes in ovarian carcinoma”.
Cancer 83(12): 2528-2533.
• Fukuda R, Hirota K, Fan F, Jung Y, Ellis L, Semenza G. 2002. “Insulin-like growth
factor 1 induces hypoxia-inducible factor 1-mediated vascular endothelial growth factor
expression, which is dependent on MAP Kinase and phosphadylinositol 3-kinase
signaling in colon cancer cells”. J. Biol. Chem. 277 (41) 38205-38211.

40
• Geva E and Jaffe RB. 2000. “Role of vascular endothelial growth factor in ovarian
physiology and pathology”. Fertil. Steril. 74(3):429-438.
• Gospodarowicz D, C. Ill, Birdwell C. 1977. “Effects of fibroblast and epidermal growth
factors on ovarian cell proliferation in vitro. I. Characterization of the response of
granulosa cells to FGF and EGF”. Endocrinology 100:1108-1120.
• Graiani G, Emanueli C, Desortes S, Pinna A, Figueroa C, Manni L, Madeddu P. 2004.
“Nerve growth factor promotes reparative angiogenesis and inhibits endothelial
apoptosis in cutaneous wounds of Type 1 diabetic mice”. Diabetología 47:1047-1054.
• Hughes AL, Gollapudi L, Sladek TL, Neet KE. 2000. “Mediation of nerve growth
factor-driven cell cycle arrest in PC12 cells by p53. Simultaneous differentiation and
proliferation subsequent to p53 functional inactivation”. J Biol Chem 275:37829-37837.
• Lara HE, Mcdonald J, Ojeda SR. 1990. “Involvement of nerve growth factor in female
sexual development”. Endocrinology 126(1):364-375.
• Mayerhofer A, Dissen GA, Parrot JA, Hill DF, Mayerhofer D, Garfield RE, Costa ME,
Skinner MK, Ojeda SR. 1996. “Involvement of nerve growth factor in the ovulatory
cascade: trkA receptor activation inhibits gap junctional communication between thecal
cells”. Endocrinology 137(12):5662-5670.
• McGee E and Hsueh J.W. A. 2000. “Initial and cyclic recruitment of ovarian follicles”.
Endocrine Reviews 21(2):200-214.
• Middeke M, Hoffmann S, Hassan I, Wulffnderlich A, Hofbauer LC, Zielke A.2002. “In
vitro and in vivo angiogenesis in PC12 pheochromocytoma cells is mediated by vascular
endothelial growth factor”. Exp Clin Endocrinol Diabetes 110:386-392.
• Muñoz J. 2003. “Relación entre el factor de crecimiento nervioso (NGF) y el factor de
crecimiento del edotelio vascular en cáncer ovárico epitelial”. Tesis para optar al Título
de Bioquímico. Anales de la Facultad de Ciencias Químicas y Farmacéuticas,
Universidad de Chile.

41
• Nakayama K, Kanzaki A, Hata K, Katabuchi H, Okamura H, Miyazaki K, Fukumoto M,
Takebayashi Y. 2002. “Hypoxia-inducible factor 1 alpha (HIF-1 α) gene expression in
human ovarian carcinoma”. Cancer Letters 176:215-223.
• Putowski LT, Skrzypczak M, Zielewicz J, Kaminski K, Jakowicki JA. 1997. “The
relevance of c-Myc to the physiology of the human ovary”. Gynecol. Endocrinol.
11(1):5-10.
• Redmer D, Reynolds L. 1996. “Angiogenesis in the ovary”. Rev Reprod. 1(3):182-192.
• Salas C. 2002. “Efecto del factor de crecimiento nervioso (NGF) en la proliferación y
secreción de esteroides en células de granulosa humana”. Tesis para optar al Título de
Químico Farmacéutico. Anales de la Facultad de Ciencias Químicas y Farmacéuticas,
Universidad de Chile.
• Salas C, Julio-Pieper M, Valladares M, Pommer R, Vega M, Mastronardi C, Kerr B,
Ojeda SR, Lara HE, Romero C. 2006. “Nerve growth factor-dependent activation of
trkA receptors in the human ovary results in synthesis of FSH Receptors and estrogen
secretion”. J Clin Endocrinol Metab. 91(6):2396-403.
• Shibuya M. 2001. “Structure and function of VEGF/VEGF receptor system
involved in angiogenesis”. Cell Struct and Funct 26:25-35.
• Semenza, L. 2002. “Signal transduction to hypoxia-inducible factor 1”. Biochemical
Pharmacology. 64:993-998.
• Sholzen T, Gerdes J. 2000. “The Ki67 Protein: From the known and the unknown”. J. Of
Cell Physiol 182:311-322.
• Sullivan DC and Bicknell R. 2003 “New molecular pathways in angiogenesis”
BritishJournal of Cancer 89:228-231.
• Tennant JR. 1964. “Evaluation of the trypan blue technique for determination of cell
viability”. Transplantation 12:685-694.

42
• Tesarollo L.1998. “Pleiotropic functions of neurotrophins in development”. Cytokine
and Growth Factor Reviews 9(2):125-138.
• Toshiaki E, Yoshimitsu K, Nishikawa A, Manase K, Shibuya M, Kudo R. 2001. “Cyclic
changes in expression of mRNA of vascular endothelial growth factor, its receptors Flt-1
and KDR/Flk-1, and Ets-1 in human corpora lutea”. Fertil. Steril 76(4):762-768.
• Vega M. 1997. “Bases biológicas de la función ovárica”.Editorial Mediterráneo. Chile
9-57.
• Wiesmann C and A. M de Vos. 2001. “Nerve growth factor: structure and function”.
Cell Mol Life Sci 58:748-759.
• Wulff C, Wiegand S, Saunders P, Scobie G, Fraser H. 2001. “Angiogenesis during
foliicular development in the primate and its inhibition by treatment with truncated ftl-
1fc (vascular endothelial growth factor trapA40 )”. Endocrinology 142(7): 3244-3254.
• Zimmermann R, Hartman T, Kavic S, Pauli S, Bohlen P, Sauer M, Kitajewski J. 2003.
“Vascular endothelial growth factor receptor 2-mediated angiogenesis is essential for
gonadotropin-dependent follicle development”. J. Clin. Invest. 112: 659-669.