Embed Size (px)
Citation preview

8/3/2019 Articulo Laser 5
http://slidepdf.com/reader/full/articulo-laser-5 1/6
Probing the differential effects of infrared light sources IR1072 andIR880 on human lymphocytes: Evidence of selective
cytoprotection by IR1072
Andrea Bradford a, Amelia Barlow a,b, Paul L. Chazot a,*
a School of Biological and Biomedical Sciences, University of Durham, South Road, Durham, Tyne & Wear DH1 3LE, United Kingdomb School of Pharmacy, University of Sunderland, Wharncliffe Road, Sunderland, Tyne & Wear SR2 3SD, United Kingdom
Received 9 March 2005; received in revised form 24 May 2005; accepted 24 May 2005
Abstract
Light therapy, both laser and LED, have been shown to provide clinical benefit in many therapeutic arenas. The effects of IR1072
and IR880 were investigated, using a range of single and multiple irradiation protocols, for their effect on freshly prepared human
lymphocytes stimulated with phytohemagglutinin. Viable cell numbers remained significantly higher after irradiation with IR1072
and were significantly lower after IR880 irradiation compared to untreated controls, following a daily single irradiation over a 5-day
period. Cell numbers were significantly higher after pre-treatment with IR1072 and exposure to UVA, compared to cells treated with
UVA only. Cells irradiated twice on Day 3 post-harvest with various wavebands confirm on Day 5, an increase in % cell viability
after IR1072, and IR1072 alternating with IR1268 irradiation, and a decrease in % cell viability after IR880 irradiation alone. Fur-
ther, wavebands tested displayed no significant differences compared to the control. Cells were collected after exposure on Days 3
and 5 with IR1072 and IR880 treatments and protein levels were compared using quantitative immunoblotting probed with an anti-
iNOS antibody. Following IR1072, but not IR880, treatment there was a 4.9 ± 2.1-fold higher iNOS protein expression in treated
cells compared to the control on Day 5 post-treatment.
Ó 2005 Elsevier B.V. All rights reserved.
Keywords: 1072 nm; 880 nm; UVA; PHA Blasts; Cytoprotection; iNOS; Apoptosis
1. Introduction
Sunlight is the most important and universal source
of non-ionising radiation essential for life on Earth.
Flora and fauna have adapted through evolution to
those components of sunlight which reach the surfaceof the planet having been filtered by the atmosphere.
Comparing the known photobiological effects of light
with the transmission spectrum of water shows that all
of these are contained within the peak of this spectrum
suggesting that atmospheric or intracellular water may
have been influential in determining the course of these
evolutionary processes (Fig. 1)
Solar ultraviolet (UV) is short wave high energy radi-
ation known to be damaging to cells and responsible for
photoageing and carcinogenesis [1,2], whereas IR isknown to be a beneficial therapeutic agent, for example
in the treatment of musculo-skeletal disorders and heal-
ing of indolent wounds [3,4]. In the laboratory, various
photo biological effects of infrared light have been ex-
plored, albeit dictated by the random commercial avail-
ability of predominantly laser light sources [5–10]. These
well-documented experiments have demonstrated
unequivocally that selected wavelengths of infrared light
1011-1344/$ - see front matter Ó 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.jphotobiol.2005.05.005
* Corresponding author. Tel.: +44 191 334 1305; fax: +44 191 334
1201.
E-mail address: [email protected] (P.L. Chazot).
Journal of Photochemistry and Photobiology B: Biology 81 (2005) 9–14
www.elsevier.com/locate/jphotobiol
ARTICLE IN PRESS

8/3/2019 Articulo Laser 5
http://slidepdf.com/reader/full/articulo-laser-5 2/6
have non-thermal photo biological effect. In 1998,
Menezes et al. [2] showed that non-thermal quantities
of IR light (700–2000 nm) induced a strong cellular de-
fence against solar UV toxicity in normal human fibro-
blasts. In 2001, Dougal and Kelly [11] demonstrated
that one single 5-min application of 1072-nm narrow
waveband light was effective in the treatment of herpes
labialis. 1072-nm light was chosen as it represents a peak
in the transmission spectrum of the water molecule.
Cold sores (herpes labialis) are known to be activated
by UV [12], which is known to suppress the immune de-
fence system.
The small, short-lived reactive molecule of nitric
oxide (NO) has emerged as a potent inhibitor of apopto-
sis. Inhibition of apoptosis by NO has been shown in a
variety of cells including B-lymphocytes [13], spleno-
cytes [14] and endothelial cells [15]. Nitric oxide (NO)
is formed by an enzyme-catalysed reaction between
molecular oxygen and L-arginine. NO is an important
molecule mediating a wide range of physiological and
pathophysiological processes. The amount of NO pro-
duction [16] may determine its role, as may the type of insult [17]. Three forms of NOS have been described
which show approximately 50% identity in amino acid
sequence [18]. NO derived from the inducible isoform
of nitric oxide synthase (iNOS) is an inflammatory prod-
uct. iNOS differs from endothelial NOS (eNOS) and
neuronal NOS (nNOS), as both eNOS and nNOS re-
quire calcium for activity, whereas calmodulin binding
to iNOS is so tight that addition of Ca2+ is not necessary
[19]. iNOS expression can be both upregulated [20–22]
or downregulated [20] in a variety of cell types depend-
ing on stimulus.
This study was designed to investigate the effect, if
any, of a series of narrow wavebands of light on human
lymphocytes in an attempt to determine the possible
photo biological response of these cells to light within
the near infrared spectrum and to consider the effects
on iNOS expression after the treatment of infrared light
in vitro.
2. Materials and methods
2.1. Cell preparation
Heparinised human whole blood was obtained from
healthy volunteers (with local ethical approval), and
peripheral blood mononuclear cells (PBMC) were sep-
arated using Lymphoprep (Axis-Shield Poc AS, Oslo,
Norway) and centrifuged at 400 g for 5 min. The
PBMCs were isolated from the interfacial layer,washed twice in RPMI without L-glutamine (Gibcoä)
and resuspended in RPMIcm (RPMI + 10% v/v fetal
calf serum + 1% penicillin/streptomycin + 1% L-gluta-
mine). Cell density was adjusted accordingly to
1 · 106 cells/ml with RPMI. 100 ll PHA (ÔLectinÕ, Sig-
ma) was added to the cells to make PHA Blasts. Cells
were incubated in 35-mm culture dishes in RPMI med-
ia at 37 °C in 5% CO2.
2.2. Experimental set-up
A series of multiple exposure protocols all of which
have shown therapeutic benefit in cold sore trials (results
not shown) were adopted for this study to show the flex-
ibility of the treatments. The five protocols were set-up
as follows:
1. PHA Blasts were exposed to infrared light source,
IR1072, on Days 3, 4 and 5 post-harvest. Using 35-
mm culture dishes, all cells were exposed to a single
3-min treatment of infrared light. Following daily
treatments, individual replicate cell samples were ana-
lysed for % cell viability on Day 5.
2. PHA Blasts were exposed to IR1072 and IR880 on
Days 3 and 5 for 5 · 3-min treatments and analysedon Day 5. Cell viability and iNOS expression was
determined after each treatment on Day 5.
3. PHA Blasts were exposed daily from Day 1 onwards
to a single 3-min dose of IR1072 and IR880. After
daily irradiation, cells were analysed for % cell
viability.
4. PHA Blasts were exposed to IR1072 on Day 3 for
4 · 3-min treatment and on Day 4 for a single
3-min treatment. Cells were then left for 4 h before
exposure to UVA for 40 min and cell viability was
then determined.
Fig. 1. Transmission spectrum of pure water [9].
10 A. Bradford et al. / Journal of Photochemistry and Photobiology B: Biology 81 (2005) 9–14
ARTICLE IN PRESS

8/3/2019 Articulo Laser 5
http://slidepdf.com/reader/full/articulo-laser-5 3/6
5. Cells were incubated until Day 3 in tissue culture
tubes and exposed to various wavebands for
2 · 3 min on Day 3. Wavebands included IR660,
IR880, IR950, IR1267, IR1072, IR1072 nm alternat-
ing with IR1268, IR1072 and IR1267 nm, 1-ls puls-
ing of IR1072 nm and 7-ls pulsing of IR1072 nm.
Cells were analysed for % cell viability immediatelyafter irradiation.
Notably for all protocols used, the temperature of all
the dishes was maintained at room temperature
throughout the IR and control treatments.
2.3. Annexin V apoptosis kit
Cell viabilities were analysed using the Annexin V
Apoptosis Detection Kit (Autogen Bioclear, UK).
Apoptosis can be detected by the change in position
of phosphatidylserine (PS) in the cell membrane. In
non-apoptotic cells, most PS molecules are localised
at the inner layer of the plasma membrane, but soon
after inducing apoptosis, PD redistributes to the outer
layer of the membrane. Exposed PS can be easily de-
tected with Annexin V. Cells with bound Annexin V
showed green staining in the plasma membrane. Cells
that had lost membrane integrity showed red staining
(PI) throughout the cytoplasm and a halo of green
staining on the cell surface (plasma membrane) [23–
26]. Cells at 1 · 105 –1 · 106 per dish were rinsed and
resuspended in Assay Binding Buffer. Five microlitres
of Annexin V and 10 ll of propidium iodide (PI) were
added to the cells before incubating at room tempera-ture in the dark for 15–30 min. Cells were observed un-
der a dual filter set for FITC and rhodamine using
fluorescence microscopy, and counted blind by at least
two observers.
2.4. Western blotting analysis
Thawed cell pellet suspensions were homogenised on
ice with a Dounce homogeniser. The protein levels in
the cell suspension were determined using the Lowry
Assay [27] using bovine serum albumin as a standard.
Protein levels were adjusted to 10 lg protein was
loaded in each lane. Standard electrophoresis was per-
formed using a 6% polyacrylamide gel. Following elec-
trophoresis, the protein was transferred to
nitrocellulose (NC) membrane for 2.5 h at 50 V. The
NC membrane was blocked with 5% non-fat skimmed
milk in 1 · Tris buffered saline (TBS) containing 0.2%
Tween 20 (Sigma, UK) for 1 h at room temperature.
The NC membrane was incubated with primary anti-
body iNOS (dilution 1:2500) overnight at 4 °C. The
NC membrane was washed 4 · 10 min with wash buffer
(2.5% non-fat skimmed milk, 0.2% Tween 20 in TBS)
and incubated with anti-rabbit horseradish peroxi-
dase-linked secondary antibody (dilution 1:2000) for
1 h. The NC membrane was washed 4 · 10 min with
wash buffer. The protein bands from the NC were vis-
ualised using a substrate of 68 mM luminol, 1.25 mM
p-couramic acid, 30% hydrogen peroxide. The immu-
noblot was exposed to Hyperfilmä for 3 min in a film
cassette and were developed and fixed at room temper-ature. The protein bands were quantified using an
ImageQuantÒ densitometer in the linear range of the
film, to determine the relative iNOS expression. Optical
density values (standardised with b-actin as in [27])
were compared using a multiple ANOVA with a signif-
icance level of p < 0.05. Data were obtained from n = 3
individual replicate experiments.
2.5. Statistics
Apoptosis was measured using % cell viability, that
is,
% cell viability ¼ ½ðNo: of viable cellsÞ=ðNo: of total cellsÞ
à 100.
Data are given as the means ± standard deviation.
Comparisons between control and treated cells were
made by a multiple ANOVA and expressed as
mean ± SD, with a confidence interval of 95%. Statisti-
cal analysis was carried out using Prism 3.2.
2.6. Light sources
Both the 880- and 1072-nm light sources emitted mul-timode light of bandwidth less than 50 nm, continuous
mode of optical power 5 mW/cm2.
3. Results
Using a range of protocols, IR1072 treatment con-
sistently elicited a significant protective effect upon
PHA Blast survival. In contrast, IR880 was consis-
tently cytotoxic compared to control and IR1072 trea-
ted cells.
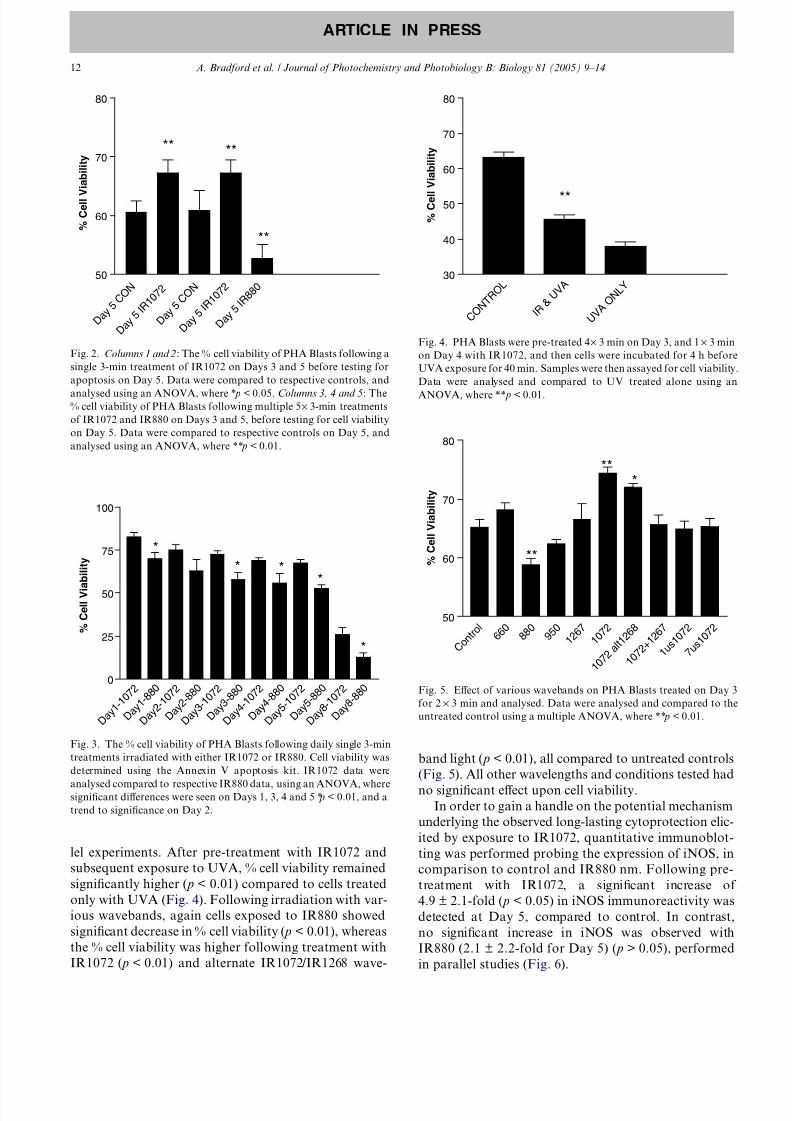
Following irradiation with IR1072, % cell viability
significantly increased on Day 5 ( p < 0.05) comparedto the control data following both a single and multiple
5 · 3-min treatment protocol on Days 3 and 5 (Fig. 2).
In the next protocol, cells irradiated with 5 · 3 min of
IR1072 and IR880, the % cell viability significantly de-
creased after treatment with IR880 both on Day 5
( p < 0.01) compared to cells treated with IR1072 (Fig.
2). The daily treatment protocol elicited a significant de-
crease in % cell viability for IR880 treated cells over an
8-day period [Day 1 ( p < 0.01), Day 3 ( p < 0.01), Day 4
( p < 0.05), Day 5 ( p < 0.05) and Day 8 ( p < 0.05)], com-
pared to those irradiated with IR1072 (Fig. 3), in paral-
A. Bradford et al. / Journal of Photochemistry and Photobiology B: Biology 81 (2005) 9–14 11
ARTICLE IN PRESS
8/3/2019 Articulo Laser 5
http://slidepdf.com/reader/full/articulo-laser-5 4/6
lel experiments. After pre-treatment with IR1072 and
subsequent exposure to UVA, % cell viability remained
significantly higher ( p < 0.01) compared to cells treated
only with UVA (Fig. 4). Following irradiation with var-
ious wavebands, again cells exposed to IR880 showed
significant decrease in % cell viability ( p < 0.01), whereas
the % cell viability was higher following treatment with
IR1072 ( p < 0.01) and alternate IR1072/IR1268 wave-
band light ( p < 0.01), all compared to untreated controls
(Fig. 5). All other wavelengths and conditions tested had
no significant effect upon cell viability.In order to gain a handle on the potential mechanism
underlying the observed long-lasting cytoprotection elic-
ited by exposure to IR1072, quantitative immunoblot-
ting was performed probing the expression of iNOS, in
comparison to control and IR880 nm. Following pre-
treatment with IR1072, a significant increase of
4.9 ± 2.1-fold ( p < 0.05) in iNOS immunoreactivity was
detected at Day 5, compared to control. In contrast,
no significant increase in iNOS was observed with
IR880 (2.1 ± 2.2-fold for Day 5) ( p > 0.05), performed
in parallel studies (Fig. 6).
D a y 5
C O N
D a y 5 I R
1 0 7 2
D a y 5
C O N
D a y 5 I R
1 0 7 2
D a y 5 I R
8 8 0
50
60
70
80
**
****
** % C
e l l V i a b i l i t y
Fig. 2. Columns 1 and 2: The % cell viability of PHA Blasts following a
single 3-min treatment of IR1072 on Days 3 and 5 before testing for
apoptosis on Day 5. Data were compared to respective controls, and
analysed using an ANOVA, where * p < 0.05. Columns 3, 4 and 5: The% cell viability of PHA Blasts following multiple 5 · 3-min treatments
of IR1072 and IR880 on Days 3 and 5, before testing for cell viability
on Day 5. Data were compared to respective controls on Day 5, and
analysed using an ANOVA, where ** p < 0.01.
D a y 1
- 1 0 7
2
D a y 1
- 8 8 0
D a y 2
- 1 0 7
2
D a y 2
- 8 8 0
D a y 3
- 1 0 7
2
D a y 3
- 8 8 0
D a y 4
- 1 0 7
2
D a y 4
- 8 8 0
D a y 5
- 1 0 7
2
D a y 5
- 8 8 0
D a y 8
- 1 0 7
2
D a y 8
- 8 8 0
0
25
50
75
100
*
* **
*
% C
e l l V i a b i l i t y
Fig. 3. The % cell viability of PHA Blasts following daily single 3-min
treatments irradiated with either IR1072 or IR880. Cell viability was
determined using the Annexin V apoptosis kit. IR1072 data were
analysed compared to respective IR880 data, using an ANOVA, wheresignificant differences were seen on Days 1, 3, 4 and 5 * p < 0.01, and a
trend to significance on Day 2.
C O N T
R O L
I R & U V A
U V A O N L
Y30
40
50
60
70
80
**
% C
e l
l V i a b i l i t y
Fig. 4. PHA Blasts were pre-treated 4· 3 min on Day 3, and 1 · 3 min
on Day 4 with IR1072, and then cells were incubated for 4 h before
UVA exposure for 40 min. Samples were then assayed for cell viability.
Data were analysed and compared to UV treated alone using an
ANOVA, where ** p < 0.01.
C o n t r o l
6 6 0
8 8 0
9 5 0
1 2 6 7
1 0 7 2
1 0 7 2
a l t 1
2 6 8
1 0 7 2
+ 1 2 6 7
1 u s 1
0 7 2
7 u s 1
0 7 2
50
60
70
80
**
**
*
% C
e l l V i a b i l i t y
Fig. 5. Effect of various wavebands on PHA Blasts treated on Day 3
for 2 · 3 min and analysed. Data were analysed and compared to the
untreated control using a multiple ANOVA, where ** p < 0.01.
12 A. Bradford et al. / Journal of Photochemistry and Photobiology B: Biology 81 (2005) 9–14
ARTICLE IN PRESS

8/3/2019 Articulo Laser 5
http://slidepdf.com/reader/full/articulo-laser-5 5/6
4. Discussion
This study has identified an ex vivo method by which
immune cell viability may be improved in the presence
of adversity. In this instance, the adverse events were
the stress of being cultured outside the human body
and, secondly, being exposed to an insult, namely
UVA light. Many authors have suggested the concept
that a particular range of wavelengths has therapeutic
benefit [6–8,10,28–30]. Biostimulation is the commonest
means by which therapeutic efficacy is sought. Whilst
the wavelengths in the 855–905 nm range may stimulate
fibroblast proliferation [9], importantly light in thisrange also appears to be lymphotoxic as shown by our
studies. The cytotoxic and protective effects upon the
cells are rapid as the analysis was carried out within
2 h of exposure to the IR light and both effects were long
lasting, being observed at least 2 days post-treatments.
This study clearly demonstrates that light in the 1050–
1100 nm range improves cell viability following both
single and multiple treatment protocols. Maintaining
lymphocyte viability in the presence of adverse factors
is of significance as bacterial endo- and exo-toxins are
leucotoxic factors, the effect of which, may be reduced
by the irradiation of the inflammatory cells by
1072 ± 25-nm light. It has long been postulated thatIR light has a protective effect against UVA, however,
the exact range of wavelengths has been unknown.
These present results suggest 1072 ± 25-nm light is pro-
tective against some of the damaging effects of UVA.
This concurs with the clinical utility of this wavelength
in treating cold sores (e.g. [11] and further unpublished
clinical observations). Although significant, the protec-
tion is incomplete and therefore requires further optimi-
sation. There are likely to be missing elements, including
other cell types and mediators in this ex vivo model
which are naturally present in vivo. Photo-modulation
of the immune response is a potential therapeutic tool
yet to be fully evaluated. The protective effect of 1072-
nm light against UV damage is an important finding
which potentially could reduce the skin damage induced
by PUVA in the treatment of psoriasis. Additional ther-
apeutic benefits would be applicable to any pathology
which responds to more resilient lymphocytes.Nitric oxide has been shown to be a potent inhibitor
of apoptosis in a variety of cell types [31]. NO diffuses
very rapidly both through water and cell membranes,
and iNOS is produced more rapidly and efficiently than
eNOS and nNOS. iNOS can function without the eleva-
tion of intracellular calcium levels and its activity is rap-
idly inducible in immune cells, for example, primarily
activated monocytes and macrophages, following expo-
sure to cytokines and microbial products [32]. Biochem-
ically, these present results show that iNOS has been
upregulated in a wavelength-dependent fashion, in com-
parison to untreated controls. NO is believed to act as
an inhibitor of apoptosis by two distinct mechanisms:
first through a cGMP-dependent mechanism where
NO acts either at the level of caspase-3-like protease
activation or upstream of this event to prevent the acti-
vation of the protease; second, NO also inhibits the
activity of the caspase-3-like protease by S -nitrosylation
of the enzyme. Suppression of caspase-3-like activity
then rescues the cell from programmed cell death [1].
We report the first evidence that IR1072 and IR880
elicit opposing effects upon lymphocyte viability ex vivo,
the former being protective and the latter wavelength
cytotoxic. Furthermore, we provide the first demonstra-
tion that IR1072 protects against UV-mediated lympho-toxicity. Preliminary biochemical evidence shows a
wavelength-dependent induction of iNOS, which may
offer a candidate protective mechanism underlying
IR1072-induced long-term preconditioning in immune
cells, and warrants further investigation.
Acknowledgements
The authors thank the volunteers for donating blood
and especially Dr. Anne Cunningham for her technical
expertise. Funding for this study was from Virulite
Ltd. (UK).
References
[1] L. Marrot, J.P. Belaidi, F. Lejeune, J.R. Meunier, D. Asselineau,
F. Bernerd, Photostability of sunscreen products influences the
efficiency of protection with regard to UV-induced genotoxic or
photoageing-related endpoints, Brit. J. Dermatol. 151 (2004)
1234–1244.
[2] S. Menezes, B. Coulomb, C. Lebreton, L. Dubertret, Non-
coherent near infrared radiation protects normal human dermal
fibroblasts from solar ultraviolet toxicity, J. Invest. Dermatol. 111
(1998) 629–633.
Fig. 6. Effect of IR treatment upon iNOS protein expression levels
PHA Blasts were exposed daily to 1 · 3-min infrared source, IR1072 or
IR880 and assayed on Days 3 and 5, for iNOS protein expression using
quantitative immunoblotting with a selective anti-iNOS antibody
(Autogen Bioclear, UK). Immunoblots were re-probed and standar-
dised with a b-actin antibody (Sigma, UK). Lane 1, control cells (Day
5); Lane 2, IR1072-treated cells (Day 5); Lane 3, IR880-treated cells
(day 5). Data were compared by compared by a multiple ANOVA with
a level of significance set at p < 0.01.
A. Bradford et al. / Journal of Photochemistry and Photobiology B: Biology 81 (2005) 9–14 13
ARTICLE IN PRESS

8/3/2019 Articulo Laser 5
http://slidepdf.com/reader/full/articulo-laser-5 6/6
[3] C. Webb, M. Dyson, W.H. Lewis, Stimulatory effect of 660-nm
low level laser energy on hypertrophic scar-derived fibroblasts:
possible mechanisms for increase in cell counts, Laser. Surg. Med.
22 (5) (1998) 294–301.
[4] Y. Ceylan, S. Hizmetli, Y. Silig, The effects of infrared laser and
medical treatments on pain and serotonin degradation products in
patients with myofascial pain syndrome: a controlled trial,
Rheumatol. Int. 24 (2003) 260–263.
[5] D.J. Castro, R.P. Abergel, C. Meeker, R.M. Dwyer, M.A.
Lesavoy, J. Uitto, Effects of the Nd:YAG laser on DNA synthesis
and collagen production in human skin fibroblast cultures, Ann.
Plast. Surg. 11 (1983) 214–222.
[6] R.P. Abergel, E.J. Zaragoza, R.M. Dwyer, J. Uitto, Diiferential
effects of Nd:YAG laser on collagen and elastin production by
chick embryo aortae in vitro. Relevance to laser angioplasty for
removal of atherosclerotic plaques, Biochem. Biophys. Res.
Commun. 131 (1985) 462–468.
[7] T.M. Mendez, A.L. Pinheiro, M.T. Pacheco, P.M. Nascimento,
L.M. Ramalho, Dose and wavelength of laser light have influence
on the repair of cutaneous wounds, J. Clin. Laser Med. Surg. 22
(2004) 19–25.
[8] M. Kreisler, A.B. Christoffers, B. Willershausen, B. dÕHoedt,
Effect of low-level GaA1As laser irradiation on the proliferation
rate of human periodontal ligament fibroblasts: an in vitro study,
Clin. Periodontol. 30 (2003) 353–358.
[9] A.N. Pereira, P. Eduardo Cde, E. Matson, M.M. Marques, Effect
of low-power laser irradiation on cell growth and procollagen
synthesis of cultured fibroblasts, Laser. Surg. Med. 31 (2002) 263–
267.
[10] M. Kreisler, A.B. Christoffers, H. Al-Haj, B. Willershausen, B.
dÕHoedt, Low level 809-nm diode laser-induced in vitro stimula-
tion of the proliferation of human gingival fibroblasts, Laser.
Surg. Med. 30 (2002) 365–369.
[11] G. Dougal, P. Kelly, A pilot study of treatment of herpes labialis
with 1072-nm narrow waveband light, Clin. Exp. Dermatol. 26
(2001) 149–154.
[12] R. Korner, F. Bahmer, R. Wigand, The effect of infrared laser
rays on herpes simplex virus and the functions of humanimmunocompenent cells, Hautarzt 40 (1989) 350–354.
[13] J.B. Mannick, K. Asano, K. Izumi, E. Kieff, J.S. Stamler, Nitric
oxide produced by human B lymphocytes inhibits apoptosis and
Epstein-Barr virus reactivation, Cell 79 (1994) 1137–1146.
[14] A.M. Genaro, S. Hortelano, A. Alvarez, C. Martinez, L. Bosca,
Splenic B lymphocyte programmed cell death is prevented by
nitric oxide release through mechanisms involving sustained Bcl-2
levels, J. Clin. Invest. 95 (1995) 1884–1890.
[15] S. Dimmeler, J. Haendeler, M. Nehls, A.M. Zeiher, Suppression
of apoptosis by nitric oxide via inhibition of interleukin-1beta-
converting enzyme (ICE)-like and cysteine protease protein
(CPP)-32-like proteases, J. Exp. Med. 185 (1997) 601–608.
[16] M. Ding, J.L. Wong, N.E. Rogers, L.J. Ignarro, R.R. Voskuhi,
Gender differences of inducible nitric oxide production in SJL/J
mice with experimental autoimmune encephalomyelitis, J. Immu-nol. 77 (1997) 99–106.
[17] I.L. Campbell, Exacerbation of lymphocytic choriomeningitis in
mice treated with the inducible nitric oxide synthase inhibitor
aminoguanidine, J. Immunol. 71 (1996) 31–36.
[18] C.J. Lowenstein, S.H. Snyder, Nitric oxide, a novel biologic
messenger, Cell 70 (1992) 705–707.
[19] AC. Gorren, B. Mayer, The versatile and complex enzymology of
nitric oxide synthase, Biokhimiya 63 (1997) 734–743.
[20] C. Nathan, Perspectives series: nitric oxide and nitric oxide
synthases, J. Clin. Invest. 100 (1997) 2417–2423.
[21] E.H. Sinz, P.M. Kochanek, C.E. Dixon, R.S.B. Clark, J.A.
Carcillo, J.K. Schiding, M. Chen, S.R. Wisniewski, J.M. Carlos,
D. Williams, S.T. Dekosky, S.C. Watkins, D.W. Marion, T.R.
Billiar, Inducible nitric oxide synthase is an endogenous neuro-
protectant after traumatic brain injury in rats and mice, J. Clin.
Invest. 104 (1999) 647–656.
[22] Y. Wang, Y. Guo, S.X. Zhang, W.-J. Wang, W. Bao, R. Bolli,
Ischemic preconditioning upregulates inducible nitric oxide syn-
thase in cardiac monocytes, J. Mol. Cell. Cardiol. 34 (2002) 5–15.
[23] S.J. Martin, C.P. Reutelingsperger, A.J. McGahon, J.A. Rader,
R.C. van Schie, D.M. LaFace, D.R. Green, Early redistribution
of plasma membrane phosphatidylserine is a general feature of
apoptosis regardless of the initiating stimulus: inhibition by
overexpression of Bcl-2 and Abl, J. Exp. Med. 182 (1995) 1545–
1556.
[24] V.A. Fadok, D.R. Voelker, P.A. Campbell, J.J. Cohen, G.L.
Bratton, P.M. Henson, Exposure of phosphatidylserine on the
surface of apoptotic lymphocytes triggers specific recognition and
removal by macrophages, J. Immunol. 148 (1992) 2207–2216.
[25] V.A. Fadok, J.S. Savill, D.L. Haslett, G.L. Bratton, P.A.
Doherty, P.A. Campbell, P.M. Henson, Different populations of
macrophages use either the vitronectin receptor or the phospha-
tidylserine receptor to recognize and remove apoptotic cells, J.
Immunol. 149 (1992) 4029–4035.
[26] V.A. Fadok, D.J. Laszlo, P.W. Noble, L. Weinstein, D.W.H.
Riches, P.M. Henson, Particle digestibility is required for induc-
tion of the phosphatidylserine recognition mechanism used by
murine macrophages to phagocytose apoptotic cells, J. Immunol.
151 (1993) 4274–4285.
[27] P.L. Chazot, O.V. Godukhin, A. McDonald, T.P. Obrenovitch,
Spreading depression-induced preconditioning in the mouse
cortex: differential changes in the protein expression of ionotropicnicotinic acetylcholine and glutamate receptors, J. Neurochem. 83
(2002) 1235–1238.
[28] D.J. Castro, R.P. Abergel, K.J. Johnston, G.E. Adomian, R.M.
Dwyer, J. Uitto, M.A. Lesavoy, Wound healing: biological
effects of Nd:YAG laser on collagen metabolism in pig skin in
comparison to thermal burn, Ann. Plast Surg. 11 (1983) 131–
140.
[29] L. Almeida-Lopes, J. Rigau, R.A. Zangaro, J. Guidugli-Neto,
M.M. Jaeger, Comparison of the low level laser therapy effects
on cultured human gingival fibroblasts proliferation using
different irradiance and same fluence, Laser. Surg. Med. 29
(2001) 179–184.
[30] G. Baxter, Therapeutic Lasers: Theory and Practice, Churchill
Livingstone, Edinburgh, 1994.
[31] Y-M. Kim, R.V. Talanian, T.R. Billiar, Nitric oxide inhibitsapoptosis by preventing increases in caspase-3-like activity via two
distinct mechanisms, J. Bio. Chem. 272 (1997) 31138–31148.
[32] Q-W. Xie, C. Nathan, The high-output nitric oxide pathway: role
and regulation, J. Leukocyte Biol. 56 (1994) 576–582.
14 A. Bradford et al. / Journal of Photochemistry and Photobiology B: Biology 81 (2005) 9–14
ARTICLE IN PRESS





























