Embed Size (px)
Citation preview

aus dem Fachbereich Geowissenschaftender Universität Bremen
Nr.56
Harald Petermann
Magnetotaktische Bakterien und ihre Magnetosome
in Oberflächensedimenten des Südatlantiks
Berichte, Fachbereich Geowissenschaften, Universität Bremen, Nr. 56134 S., 93 Abb., 9 Tab., Bremen 1994
ISSN 0931-0800


Magnetotaktische Bakterien und ihre Magnetosome
in Oberflächensedimenten des Südatlantiks
Dissertation
zur Erlangung des Doktorgrades
am Fachbereich Geowissenschaften
der Universität Bremen
vorgelegt von
Harald Petermann
Bremen 1994


Inhaltsverzeichnis
1
1.1
2
2.1
2.1.1
2.1.2
2.2
2.2.1
2.2.1.1
2.2.1.2
2.2.2
2.2.3
2.2.4
2.2.5
2.2.6
3
4
4.1
4.2
4.3
4.3.1
4.3.2
4.4
5
5.1
5.2
5.3
5.3.1
Seite
Einleitung 1
Zielsetzung der Arbeit 2
Bakterien 3
Einführung 3
Morphologie 3
Stoffwechsel 3
Magnetotaktische Bakterien 4
Magnetosome 5
Bildung der Magnetosome 6
Magnetisierungsrichtung in Magnetosomen 6
Bewegung magnetotaktischer Bakterien im Magnetfeld 8
Magnetotaktische Organismen mit Eisen-Sulfid-Kristallen 10
Physiologie magnetotaktischer Bakterien: 11
Phylogenese magnetotaktischer Bakterien 12
Extrazellulär magnetitproduzierende Bakterien 12
Das Magnetodrom 13
Untersuchung der Schwimmbewegung magnetotaktischer Bakterien 19
Unterscheidung verschiedener Bewegungsformen 19
Schwimmgeschwindigkeiten 21
Bestimmung des magnetischen Moments magnetotaktischer Bakterien
aus der Schwimmbahn 22
Theoretischer Ansatz 22
Bestimmung des magnetischen Moments aus den untersuchten
Bakterienspuren 28
Bestimmung des magnetischen Moments in rotierenden Magnetfeldern 29
Tiefenverteilung magnetotaktischer Bakterien und geochemische
Zonierung in Wattsedimenten 34
Beschreibung des Arbeitsgebiets 34
Morphologie magnetotaktischer Bakterien in Wattsedimenten 35
Laboruntersuchung der Tiefenverteilung magnetotaktischer Bakterien in
Sedimentkernen aus dem Watt von Weddewarden 35
Geochemische Zonierung der oberflächennahen Sedimentschichten 38

5.3.2
5.4
5.5
5.6
6
6.1
6.2
6.3
6.4
6.5
6.6
6.7
6.8
6.9
6.10
7
7.1
7.2
7.3
7.4
7.5
7.6
8
8.1
8.1.1
8.1.2
"Reaktion der magnetotaktischen Bakterien auf Veränderung der
geochemischen Zonierung 39
Beprobung der Wattsedimente in situ 40
Bestimmung der geochemischen Zonierung und der Tiefenverteilung
magnetotaktischer Bakterien im gleichen Kern 42
Diskussion 44
Lebende magnetotaktische Bakterien in Sedimenten des Südatlantiks .46
Einführung 46
Behandlung der Proben 46
Bestimmung der Anzahl magnetotaktischer Bakterien im Sediment .47
Geographische Verteilung der Magnetbakterienkonzentration im
Südatlantik 49
Variation der Magnetbakterienkonzentration mit der Wassertiefe 51
Zusammenhang zwischen Magnetbakterienkonzentrationen und
Eintrag organischer Verbindungen ins Sediment 53
Zusammenhang zwischen Magnetbakterienkonzentration und Gehalt an
organischem Kohlenstoff und Stickstoff im Sediment.. 57
Zusammenhang zwischen Magnetbakterienkonzentration und dem
Nährstoffabbau im Sediment 59
Zusammenhänge zwischen der Magnetbakterienkonzentration
und anderen geochemischen Parametern 61
Tiefenverteilung magnetotaktischer Bakterien im Sediment 64
Extraktion und elektronenmikroskopische Untersuchung magne-
tischer Partikel aus Oberflächensedimenten des Südatlantiks 71
Extraktionsverfahren 71
Qualitative und quantitative Beurteilung der Extraktionsmethode 73
Elektronenmikroskopische Untersuchung 74
Elementanalyse im Elektronenmikroskop 79
Verteilung verschiedener Magnetosomformen in den Sedimenten des
Südatlantiks 81
Zusammenhang zwischen der Verteilung der Magnetosomformen
und dem Nährstoffangebot im Sediment 84
Gesteinsmagnetik 87
Messungen mit dem Alternating-Gradient-Magnetometer 87
Aufbau und Meßprinzip der AGM Apparatur 87
Probenpräparation 88

111
8.1.3 Messung 88
8.2 Das Auswerteprogramm HYSTEAR 91
8.3 Regionale Charakteristika magnetischer Komponentenspektren der
Oberflächensedimente im Südatlantik 93
8.4 Hystereseparameter der Oberflächensedimente des Südatlantiks 98
8.5 Messung der anhysteretischen und der isothermalen Remanenz 104
8.6 Sättigungsmagnetisierung der Oberflächensedimente
im östlichen Südatlantik 106
9 Zusammenfassung 108
10 Literaturverzeichnis 110
Anhang A
Aufnahme von Spuren magnetotaktischer Bakterien mittels Bildverarbeitung . 119
A.1
A.1.1
A.1.2
A.1.3
A.1.4
A.2
A.2.1
A.2.2
A.2.3
A.3
A.3.1
A.3.2
AA
Beschreibung der Bildverarbeitungskarte 119
Echtzeit-Bildverarbeitung 119
Bearbeitung von Bildsignalen in der Bildverarbeitungskarte 119
Abspeichern auf Festplatte 120
Ausdruck auf dem Laserdrucker 121
Technische Probleme bei der Bildverarbeitung 121
Genauigkeit der Ortsdarstellung 121
Verdrehung der Kamera 122
Konstanz der Helligkeit des Bildes 123
Bildverarbeitungs-Algorithmen zur Überlagerung mehrerer Bilder 124
Addition von Bildern 124
Überlagerung mehrerer Bilder mittels der XOR-Funktion 124
Aufzeichnen der Bakterienbahnen 125
Anhang B
Liste und Karten der Beprobungsstationen im Südatlantik 129


1
1 Einleitung
Für die zeitliche Einordnung der Schichtenabfolge von Sedimenten liefert die
Magnetostratigraphie zuverlässige Werte. Zur sicheren Bestimmung der Magnetfeldrichtungen ist aber die Kenntnis der Träger der remanenten Magnetisierungnotwendig. Während lange Zeit davon ausgegangen wurde, daß die Magnetomineralelithogen gebildet sind, erkannten KIRSCHVINK & LOWENSTAM (1979) erstmals dieBedeutung biogen gebildeter magnetischer Minerale und vermuteten, daß unteranderem magnetotaktische Bakterien einen maßgeblichen biogenen Beitrag zurremanenten Magnetisierung der Sedimente leisten können.
Magnetotaktische Bakterien wurden 1975 von BLAKEMORE in den Sedimenten dermarinen Salzsümpfe bei Woods Hole (Massachusetts) entdeckt. Sie fielen imLichtmikroskop durch ihre hohe Beweglichkeit und besonders dadurch auf, daß sie aufdem Objektträger immer in dieselbe Richtung schwammen. Weder ein chemischer Reiz,
noch die Einwirkung von Licht konnte dieses Phänomen erklären. Erst als dasMikroskop gedreht wurde und diese Bakterien daraufhin in eine neue Richtungschwammen, wurde deutlich, daß sie sich am magnetischen Feld orientieren. Wegenihrer am Magnetfeld ausgerichteten Bewegung wurden sie magnetotaktische Bakterien
genannt.
Sie bilden intrazellulär Magnetitkristalle - sogenannte Magnetosome - mit charak
teristischen Morphologien und einem engen Korngrößenspektrum von 40 bis< 200 nm.Aufgrund der Korngröße sind alle oder zumindest die überwiegende Mehrheit der
Magnetosome magnetische Einbereichsteilchen und besitzen eine hoheMagnetisierung.
Magnetotaktische Bakterien kommen in sehr unterschiedlichen Bereichen vor. Siesind in limnischen, brackischen sowie küstennahen marinen Sedimenten (BLAKEMORE,1982; MANN et al., 1990) und sogar in einem feuchten Moorboden (FASSBINDER et al.,1990) nachgewiesen worden. STOLZ et al. (1986) konnten lebende Magnetbakterienauch aus hemipelagischen Sedimenten des Santa Barbara Beckens vor Kalifornien inWassertiefen bis 600 m bergen. Ob diese Organismen auch in pelagischen Sedimentenleben und durch authigene Bildung magnetischer Kristalle zur Magnetisierung
beitragen, war aber nicht bekannt.
Fossile Magnetosome bleiben in nicht sulfidischen Sedimenten auch nach der Zersetzung der organischen Bakterienhülle erhalten. In pelagischen Sedimenten wurden
fossile Magnetosome erstmals von PETERSEN et al. (1986) an Kernen aus dem AngolaBecken anhand elektronenmikroskopischer Aufnahmen identifiziert. Bei weiterenUntersuchungen konnten fossile Magnetosome auch in Sedimenten anderer (hemi-)pelagischer Areale gefunden werden (McNEILL, 1990; STOLZ et al., 1990; VALI et al.,1989; YAMAZAKI et al., 1991). Einige Autoren schätzten, daß Magnetosome einenhohen Beitrag zur Gesamtmagnetisierung liefern.

2
1.1 Zielsetzung der Arbeit
Primäres Ziel dieser Arbeit war, zu klären, ob lebende magnetotaktische Bakterien in
pelagischen Sedimenten, speziell im Südatlantik, vorkommen. Dazu wurde eine
Apparatur aufgebaut, mit der sich magnetotaktische Bakterien anhand ihrer Bewegung
in definierten Magnetfeldern im Mikroskop eindeutig identifizieren lassen und mit dersich ihr magnetisches Moment bestimmen läßt. Vorbereitende Untersuchungen wurden
in Sedimenten des Weserästuars durchgeführt.
Während vier Fahrten mit dem Forschungsschiff METEOR sollte die Verteilung der
magnetotaktischen Bakterien in den Oberflächensedimenten des Südatlantiks bestimmt
werden.
Neben der Identifizierung der lebenden Magnetbakterien sollten elektronenmikro
skopische Untersuchungen an magnetischen Extrakten Aufschluß über die Verteilung
der Magnetosome in den südatlantischen Sedimenten geben. Dies ist möglich, da sichdie Magnetosome durch ihre charakteristische Morphologie von anderen magnetischen
Mineralen unterscheiden.
Anhand gesteinsmagnetischer Messungen sollten die Träger der Magnetisierung ins
gesamt charakterisiert werden. Durch den Vergleich verschiedener magnetischer Para
meter sollten die Konzentrationen sowie die mittlere Korngröße der magnetischen
Fraktion bestimmt und insbesondere der Anteil magnetischer Einbereichsteilchen, zu
denen die Magnetosome gehören, untersucht werden. Diese Ergebnisse sollten mit der
Verteilung von lebenden Magnetbakterien und Magnetosomen in Beziehung gesetzt
werden.
Diese Arbeit ist eingebunden in den Sonderforschungsbereich 261 'Der Südatlantik
im Spätquartär: Rekonstruktion von Stoffhaushalt und Stromsystemen' und soll einen
Beitrag liefern zum Teilprojekt 3 'Bildung und Stabilisierung physikalischer Signale im
Sediment'. Durch den Vergleich der Verteilungen der lebenden magnetotaktischen
Bakterien und ihrer Magnetosome mit geochemischen Stoff-Flüssen steht diese Arbeit inenger Verbindung zum Teilprojekt 2 'Umwandlungsprozesse und Stoff-Flüsse in
Sediment / Porenwassersystemen'.

3
2 Bakterien
2.1 Einführung
Am Beginn der Arbeit stehen einige allgemeine Bemerkungen über Bakterien, auf dieim weiteren Bezug genommen wird.
2.1.1 Morphologie
Unter dem Mikroskop kann man mehrere morphologische Grundformen von Bakterienunterscheiden. Zu den häufigsten gehören:
- Kokken: sie sind kugelförmig,
- Stäbchen: längliche, nicht gebogene Formen,
- Vibrios: längliche gebogene Bakterien,
- Spirillen: spiralförmig gewundene Bakterien.
Die Form der Bakterien ist nicht unveränderlich: Bei knappem Nährstoffangebotentwickeln sich die meisten Bakterien zu kokkoiden Kümmerformen zurück, bei sehralten Bakterienkulturen treten Riesenformen auf (SCHLEGEL, 1985; BAZYLINSKI, 1990).
Durch den Aufbau ihrer Zellwand lassen sich die Bakterien in zwei Gruppenunterteilen, die auch genetisch recht klar voneinander getrennt sind: Gram-positive undGram-negative Zellen, benannt nach einer Färbetechnik. Die Gram-positiven besitzenim Vergleich zu den Gram-negativen Bakterien eine stabilere Zellwand. Alle magnetotaktischen Bakterien gehören, soweit bekannt, zu den Gram-negativen Bakterien, wie
auch die Mehrzahl der marinen Bakterien.
Einige Bakterienarten können Dauerformen bilden, sogenannte Sporen. DieseSporen können sehr lange, auch bei ungünstigen Bedingungen, überdauern. Bei Gram
negativen Bakterien ist die Fähigkeit, Sporen auszubilden, nicht üblich (SCHLEGEL,1985).
Die meisten Bakterien sind unbeweglich, einige können sich gleitend fortbewegen,manche besitzen Flagellen, korkenzieherartig geformte Filamente. Ihre Fortbewegungerfolgt durch Drehung der Flagellen.
2.1.2 Stoffwechsel
Im Gegensatz zu den höheren Lebewesen, die beim Stoffwechsel organischeKohlenstoff-Verbindungen nur mit Sauerstoff auf- und abbauen können, gibt es beiBakterien eine große Vielfalt von Stoffwechseltypen.

4
Phototrophe Bakterien gewinnen ihre Energie aus Sonnenlicht, chemotrophe
Bakterien aus chemischen Reaktionen. Chemotrophe Bakterien werden weiter nach
ihrer Kohlenstoffquelle und nach ihrem Oxidations- / Reduktionsmittel unterschieden.
Autotrophe Bakterien beziehen den Kohlenstoff zum Aufbau der Zelle aus CO2.
Heterotrophe Bakterien oxidieren organische Kohlenstoffverbindungen, dazu können
sie verschiedene Oxidationsmittel benutzen, etwa 02' NOx, SOx, Mn (IV), Fe (111) oder
organische Verbindungen. Aerobe Bakterien verwenden 02' anaerobe Bakterien einesder anderen der angeführten Oxidationsmittel. Bakterien sind nicht auf einen
Stoffwechselweg beschränkt, meist können sie sich auf veränderte Umweltbedingungen
einstellen, indem sie zu einem anderen Stoffwechselweg übergehen.
Bezüglich des Verhaltens gegenüber Sauerstoff lassen sich vier Gruppen
unterscheiden:
- strikte Aerobier,
- mikroaerophile Bakterien,
- fakultativ anaerobe Bakterien,
- strikt anaerobe Bakterien.
Die beiden ersten Gruppen benötigen Sauerstoff, die zweite Gruppe toleriert nur
geringe 02-Konzentrationen, Gruppe drei kann sowohl in sauerstoffhaitiger, als auch in
sauerstofffreier Umgebung leben, die letzte Gruppe wird durch Sauerstoff oft irreversibel
geschädigt. Sauerstoff tritt in einigen sehr reaktionsfreudigen Verbindungen (03' H20 2)auf, die die Zelle schädigen. Nur Bakterien, die Enzyme besitzen, um diese
Verbindungen abzubauen, können bei Anwesenheit von Sauerstoff überleben.
2.2 Magnetotaktische Bakterien
Magnetotaktische Bakterien wurden 1975 von BLAKEMORE entdeckt. Bei der
mikroskopischen Untersuchung von Sedimenten mariner Salzsümpfe fiel eine Gruppe
schnell beweglicher Bakterien auf, die immer in dieselbe Ecke des Objektträgers
schwamm. Sie änderten ihre Bewegungsrichtung nur, wenn ein Handmagnet in ihre
Nähe gebracht wurde. Dies war nach Beobachtungen an Tauben (WALLCOTT et al.,
1974) der zweite Hinweis für den Einfluß von Magnetfeldern auf Organismen. DerVersuch einer Kultivierung der magnetotaktischen Bakterien stieß auf große
Schwierigkeiten, erst vier Jahre später konnte die erste Spezies, das Aquaspirillummagnetotacticum, Kurzbezeichnung MS-1, in Kultur gebracht werden (BLAKEMORE et al.,1979). Es dauerte weitere neun Jahre bis eine zweite Spezies in Reinkultur vorlag
(BAZYLINSKI et al., 1988). Die wesentlichen physiologischen Arbeiten über magnetotaktische Bakterien, die zu Beginn dieser Dissertation vorlagen, wurden an A. magnetotacticum durchgeführt.
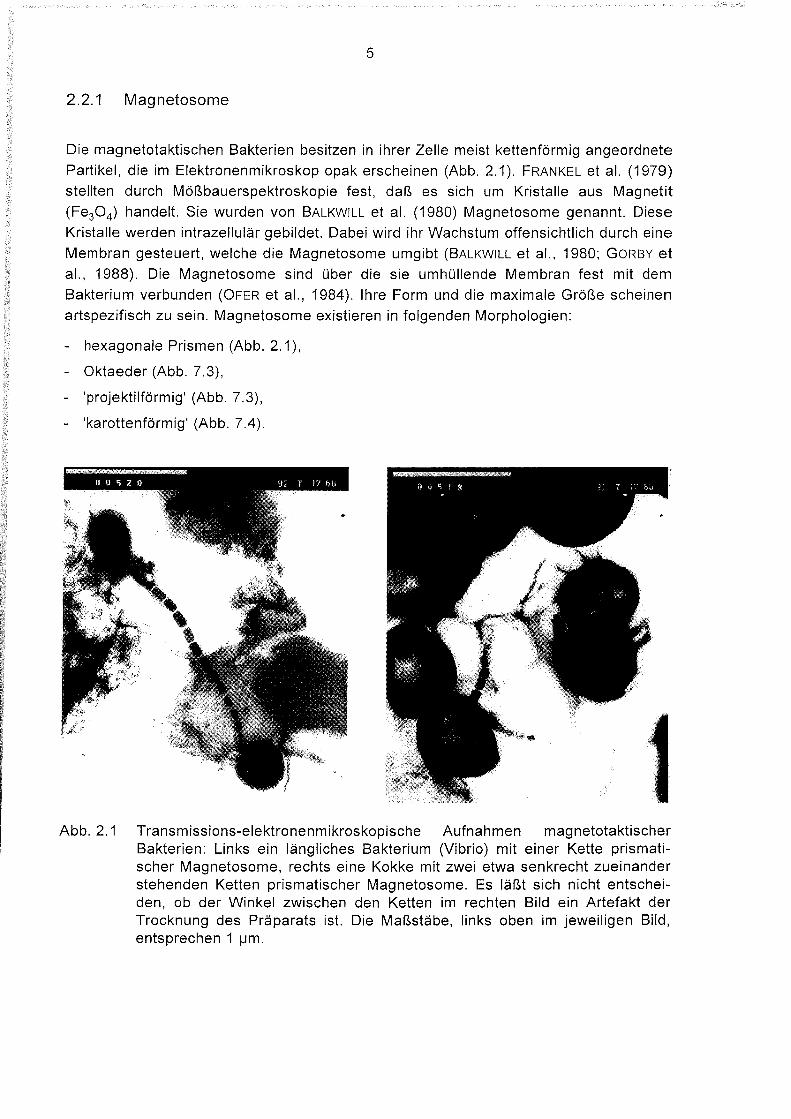
5
2.2.1 Magnetosome
Die magnetotaktischen Bakterien besitzen in ihrer Zelle meist kettenförmig angeordnete
Partikel, die im Elektronenmikroskop opak erscheinen (Abb. 2.1). FRANKEL et al. (1979)
stellten durch Mößbauerspektroskopie fest, daß es sich um Kristalle aus Magnetit
(Fe30 4) handelt. Sie wurden von BALKWILL et al. (1980) Magnetosome genannt. Diese
Kristalle werden intrazellulär gebildet. Dabei wird ihr Wachstum offensichtlich durch eine
Membran gesteuert, welche die Magnetosome umgibt (BALKWILL et al., 1980; GORBY et
al., 1988). Die Magnetosome sind über die sie umhüllende Membran fest mit dem
Bakterium verbunden (OFER et al., 1984). Ihre Form und die maximale Größe scheinen
artspezifisch zu sein. Magnetosome existieren in folgenden Morphologien:
- hexagonale Prismen (Abb. 2.1),
- Oktaeder (Abb. 7.3),
- 'projektilförmig' (Abb. 7.3),
- 'karottenförmig' (Abb. 7.4).
Abb. 2.1 Transmissions-elektronenmikroskopische Aufnahmen magnetotaktischerBakterien: Links ein längliches Bakterium (Vibrio) mit einer Kette prismatischer Magnetosome, rechts eine Kokke mit zwei etwa senkrecht zueinanderstehenden Ketten prismatischer Magnetosome. Es läßt sich nicht entscheiden, ob der Winkel zwischen den Ketten im rechten Bild ein Artefakt derTrocknung des Präparats ist. Die Maßstäbe, links oben im jeweiligen Bild,entsprechen 1 ~m.

6
2.2.1.1 Bildung der Magnetosome
Bei A. magnetotacticum findet die Bildung der Magnetosome in mehreren Schritten statt
(FRANKEL et 811., 1983). Dreiwertiges Eisen wird über einen Komplexbildner (Chelator,
z.B. Eisen (I11)-Quinat) in Lösung gebracht. In komplexierter Form besitzt Eisen (111) eine
erhöhte Löslichkeit in Wasser. In der Zelle wird das Eisen in zweiwertiger Form vom
Chelator abgegeben und in die Vesikel transportiert, die die Magnetosome umgeben.
Dort wird es zunächst als Eisenhydroxid abgelagert. Dabei handelt es sich vermutlich
um Ferrihydrit Fe20 3·nH20 (PAOLETTI & BLAKEMORE, 1988) oder Ferritin, ein komplexes
Protein mit etwa 6 nm großem Eisenhydroxidkern, das in biologischen Systemen sehr
verbreitet ist (ARTYMIUK et al., 1990; SMITH et al., 1990). Der Nachweis des Eisenhydro
xids gelang an Zellkulturen, die sich in der exponentiellen Wachstumsphase befanden,
also der Phase, in der sich alle Bakterien teilen und keine Bakterien absterben. Die
Bildung der Magnetosome erfolgt zeitversetzt nach der Zellteilung. Bakterien im frühen
exponentiellen Wachstumsstadium enthielten ausschließlich dieses nicht genauer
spezifizierbare Eisenhydroxid. Innerhalb der Magnetosom-Membran wird das
Eisenhydroxid anschließend in Magnetit umgewandelt. Die die Magnetosome
umgebende Membran besteht aus Proteinen und Lipiden, wobei zwei der Proteine nur in
dieser Membran und nicht in der Zellmembran nachgewiesen werden konnten (GORBY
et al., 1988). Diese Proteine steuern vermutlich die Synthese der Magnetosome. Der
Syntheseweg des Magnetits ist zur Zeit aber noch wenig erforscht.
2.2.1.2 Magnetisierungsrichtung in Magnetosomen
Die Magnetosome liegen recht genau im theoretisch berechneten Korngrößenbereich für
magnetische Einbereichsteilchen (BUTLER & BANERJEE, 1975; Abb. 2.3). Ihre interne
Magnetisierung zeigt demnach im wesentlichen in eine Richtung, und das Magnetosom
besitzt dadurch insgesamt maximale Magnetisierung (Abb. 2.2).
Abb.2.2Magnetisierung in einem quaderförmigen Magnetitkristall mit den Seitenlängen 180 nm·100 nm·100 nm. Die Pfeile geben die jeweilige mittlereMagnetisierungsrichtung des repräsentiertenVolumens an. Abbildung aus WILLIAMS & DUNLOP(1990).

7
Nur bei den prismatischen und oktaedrischen Formen gibt es Magnetosome mit
einem Achsenverhältnis von annähernd eins, die im Gebiet der Zweibereichsteilchenliegen. Ganze Ketten solcher Magnetosome dürften aber wiederum im Feld derEinbereichsteilchen liegen. Anschaulich formuliert kann man eine Kette mit n
Magnetosomen durch ein einzelnes Teilchen mit der n-fachen Länge, und einemAchsenverhältnis von 1/n ersetzen: Vier Würfel mit 2000 A Kantenlänge bilden eineKette mit 8000 A Länge und einem Achsenverhältnis von 0.25. Dieser Wert liegt im Feldfür Einbereichsteilchen.
Länge Ltnge[ pm ] r----r-----.-----,r-----r-----,[ A ]
superparamagnetischeTeilchen
1.00
0.80
0.80
0.40
0.2Q
0.100.08
0.08
0.04
EinbereichsTeilchen
ZweibereichsTeilchen
10000
8000
8000
4000
00
0.0 0.2 0.4 0.8 0.8 1.0
Verhältnis kleine I große Achse
Abb. 2.3 Domänenstatus quaderförmiger Magnetitkristalle mit quadratischer Grundfläche nach BUTLER & BANERJEE (1975). Auf der Abszisse ist dasAchsenverhältnis aufgetragen, auf der Ordinate die Länge. Die überwiegendeMehrzahl der Magnetosome (Kreuze) fällt in den Bereich derEinbereichsteilchen, einige mit einem Achsenverhältnis nahe eins sind nachdiesem Modell Zweibereichsteilchen. Meßwerte aus PETERSEN et al. (1989).
Magnetotaktische Bakterien können zwischen drei und mehreren hundertMagnetosomen enthalten (PETERMANN, 1989). Nimmt man an, daß die Magnetisierungin allen Magnetosomen homogen und parallel ist (Abb. 2.2), so liegt das magnetischeMoment eines Magnetbakteriums zwischen 10-16 und 10-13 Am2. Die meistenMagnetbakterien besitzen etwa 10 - 20 Magnetosome (MANN et al. , 1990a), dasmagnetische Moment sollte daher im Mittel bei einigen 10-15 Am 2 liegen.

8
2.2.2 Bewegung magnetotaktischer Bakterien im Magnetfeld
Als Magnetotaxie wird eine gerichtete Bewegung (Taxie) im Magnetfeld bezeichnet.Tatsächlich richten sich magnetotaktische Bakterien parallel zu einem anliegendenMagnetfeld aus. Die Bewegung längs der Feldlinien erfolgt durch aktive Fortbewegungmittels Flagellen, nicht durch passives Ausnutzen magnetischer Anziehungskräfte.
Flagellen bestehen aus einem etwa 20 nm dicken helical (korkenzieherförmig)gewundenen Proteinfilament, das über eine hakenförmige Struktur ('proximal hook',SCHLEGEL, 1985) in einer Scheibe verankert ist, die im Bakterienkörper steckt(ENGELHARDT et al., 1993). Diese Scheibe rotiert relativ zum Bakterienkörper. Durch dieRotation des Flagellums schraubt sich das Bakterium durch die Flüssigkeit und bewegtsich dabei vorwärts. Der Antrieb des Flagellenmotors erfolgt durch Diffusion vonProtonen ins Bakterium hinein. Um diese Diffusion zu ermöglichen, müssen dieBakterien die Protonenkonzentration in der Zelle niedriger halten als in der Umgebungund ständig Protonen gegen den Konzentrationsgradienten durch die Zellmembran nachaußen schleusen.
Wozu dient die Magnetotaxie? Magnetbakterien müssen sich nicht bewegen, um dieNährstoffzufuhr zu erhöhen (STEINBERGER, 1990). Da Bakterien sehr klein sind - dieMehrzahl ist etwa 0.5 bis 3 IJm groß - besitzen sie ein großes Verhältnis von Oberflächezu Volumen. Daher sorgen diffusive Prozesse für eine ausreichende Nährstoffzufuhr.Bewegen müssen sie sich aber, wenn sie sich in einem für sie nicht adäquatemLebensraum befinden. Sie versuchen dann ein günstigeres Milieu zu erreichen.Üblicherweise erfolgt dies in einer 'run and tumble'-Bewegung. Die Bakterien führenabwechselnd Taumelbewegungen um die eigene Achse und geradlinigeSchwimmbewegungen in die neue Richtung aus. Führt ihr Weg in ein chemischgünstigeres Milieu, so ist die Strecke, die sie zwischen zwei Taumelbewegungenschwimmen, länger, als wenn sie sich in chemisch ungünstigeres Milieu hineinbewegen.Dadurch nähern sie sich auf einem Zickzack-Weg dem Ort, der ausreichendeLebensbedingungen bietet.
Magnetotaktische Bakterien leben zum weit überwiegenden Teil in Sedimenten. Fallsdas Sediment aufgewirbelt wird und sie in die Wassersäule geraten, müssen sieversuchen, auf kürzestem Wege zurück ins Sediment zu gelangen. Aufgrund ihresmagnetischen Momentes werden sie im Erdmagnetfeld gedreht, anschließendschwimmen sie parallel zum Erdfeld ins Sediment zurück.

ChemotaktischesBakterium
9
MagnetotaktischesBakterium
(pb
"'c:> Erdmagnetfeld
~
Abb. 2.4 Vergleich zwischen den Schwimmbewegungen eines chemotaktischenBakteriums (links) und eines magnetotaktischen Bakteriums (rechts). Beideleben bevorzugt im Sediment. Während das chemotaktische Bakteriumabwechselnd Taumel- und Schwimm-Bewegungen ausführt und auf einemZickzack-Pfad ins Sediment gelangt, wird das magnetotaktische Bakteriumim Erdfeld gedreht und bewegt sich parallel zu den Feldlinien geradlinig insSediment hinein.
Das Erdmagnetfeld besitzt auf der nördlichen Hemisphäre der Erde eine nach untengerichtete, auf der südlichen Hemisphäre eine nach oben gerichtete Komponente (Abb.2.5). Der Winkel des Erdmagnetfeldes zur Erdoberfläche, die Inklination, nimmt mit
zunehmender geographischer Breite zu. An den magnetischen Polen sind dieMagnetfeldlinien senkrecht zur Erdoberfläche gerichtet, am Äquator horizontal. Dieüberwiegende Mehrzahl der Magnetbakterien der Nordhalbkugel schwimmt nach Nordenund damit nach unten ins Sediment, Magnetbakterien der Südhalbkugel schwimmenvorzugsweise nach Süden und damit ebenfalls nach unten (BLAKEMORE et al., 1980).Am Äquator finden sich ungefähr gleiche Anteile nordsuchender und südsuchenderMagnetbakterien (FRANKEL et al., 1981). Erst ab einer Inklination über sechs Grad setztsich eine der beiden Orientierungen durch (TORRES OE ARAUJO et al., 1990). Auch inhohen Breiten kommt immer ein kleiner Anteil magnetotaktischer Bakterien mit inverserPolarität vor. Das horizontale Erdmagnetfeld am Äquator bietet den Magnetbakteriennicht den Vorteil, auf direktem Wege ins Sediment zu gelangen. Wenn sie im Sedimentbleiben, haben sie aber auch keine Nachteile. Ein unmittelbarer Vorteil derMagnetotaxie am Äquator liegt nicht auf der Hand.

NÖRDLICHE HEMISPHÄRE
10
SÜDLICHE HEMISPHÄRE
NORDSUCHEND ~ SÜDSUCHEND SÜDSUCHEND NORDSUCHEND
Abb. 2.5 Orientierung magnetotaktischer Bakterien im Magnetfeld. Die Pfeile innerhalbder Bakterien geben jeweils die Richtung der Magnetisierung an, die dünnenPfeile außerhalb der Bakterien die Schwimmrichtung. Die MagnetfeldrichtungB ist durch die dicken Pfeile gekennzeichnet.
Zusätzlich zur Magnetotaxie zeigen die Magnetbakterien chemotaktisches Verhalten.Wenn das Milieu zu ungünstig wird, unterbrechen magnetotaktische Bakterien ihreSchwimmbewegung. SPORMANN & WOLFE (1984) berichteten dies erstmals, aber aucheigene Untersuchungen zeigen dies immer wieder. Besonders dann, wenn dieMagnetbakterien mit künstlichen Magnetfeldern weit aus dem Sediment geführt werden,tritt bei manchen Bakterien die chemotaktische Hemmung in Erscheinung. MATSUNAGAet al. (1991) geben an, daß ein Teil der von ihnen untersuchten Bakterien der magnetotaktischen Kultur AMB-1 unbeweglich war.
2.2.3 Magnetotaktische Organismen mit Eisen-Sulfid-Kristallen
Die in dieser Arbeit behandelten magnetotaktischen Bakterien bilden Magnetitkristalle.Daneben existieren magnetotaktische Organismen, die intrazellulär Eisensulfide
aufbauen.
DEMITRACK (1985) entdeckte bei der Beprobung der Sedimente des Teiches, an demBlakemore die ersten magnetotaktischen Bakterien gefunden hatte, Kristalle aus Greigit(Fe3S4), die eine Korngröße von etwa 100 nm und kuboidale Form besaßen. FARINA etal. (1990) fanden in stark sulfidischen Sedimenten einen vielzelligen magnetotaktischenOrganismus, der Partikel aus Eisensulfid enthielt. Durch Elektronenbeugung ließ sichdie chemische Zusammensetzung nicht eindeutig bestimmen. Es wurde vermutet, daßes sich um Pyrrhotit (Fe7SS) handelt. Bei einigen Partikeln ist ein Kern aus Eisensulfidvon einer Schicht aus Eisenoxid, vermutlich Magnetit, umgeben (FARINA et al., 1990).Gleichzeitig entdeckten MANN et al. (1990b) multizelluläre magnetotaktischeOrganismen, die Magnetosome aus Greigit mit Anwachsungen von Pyrit (FeS2) bilden.RODGERS et al. (1990) konnten elektronenmikroskopisch zeigen, daß es sich umprokaryontische (zellkernlose) Zellen handelt, die aber an einigen Stellen zusammengewachsen waren und sich als Aggregat synchron vorwärts bewegten.
11
Die Partikel aus Pyrrhotit oder Greigit zeigen selten solch perfekte Kristallflächen, wiesie bei Magnetosomen aus Magnetit angetroffen werden (siehe Kap. 7.3). Die Syntheseder Eisensulfide scheint nicht so exakt kontrolliert zu sein, wie die von Magnetit. DieKorngröße der Eisensulfide liegt bei 50 bis 100 nm. Da Magnetit und Greigit identische
Kristallstrukturen und ganz ähnliche magnetische Eigenschaften besitzen" istanzunehmen, daß es sich bei den Greigit-Teilchen ebenfalls um magnetischeEinbereichsteilchen handelt. Mittlerweile wurde auch über einzellige magnetotaktischeBakterien mit Magnetosomen aus Greigit berichtet (HEYWOOD et aL, 1990; HEYWOOD et
aL,1991).
Greigit ist thermodynamisch nicht stabil, er reagiert in reduzierendem Milieu zu Pyrit.Wie zuvor erwähnt, existieren sogar innerhalb der Zellen eines multizellulärenOrganismus Greigit und Pyrit nebeneinander (MANN et aL, 1990b). Die Bedeutung derbiogenen Eisensulfid-Mineralisationen für die Magnetisierung von Sedimenten läßt sich
bisher nicht abschätzen. Hier eröffnet sich der Sedimentmagnetik ein neues Feld.
2.2.4 Physiologie magnetotaktischer Bakterien:
Tabelle 2.1 Übersicht über die bis Anfang 1994 in Kultur vorliegenden magnetotaktischen Bakterien (N.D. bedeutet 'not determined').
Name Morphologie Vorkommen Stoffwechsel- Terminaler Referenztyp Elektronen-
akzeptor
MS-1 Spirillum Süßwasser heterotroph N03- (02) BLAKEMORE '79,(Fe (111)) BAZYLINSKI '83
GUERIN '92
MV-1 Vibrio Salzwasser heterotroph N20 BAZYLINSKI '88, '90
AMB-1 Spirillum Süßwasser heterotroph NO~- MATSUNAGA '91
MSR-1 Spirillum Süßwasser heterotroph N.D. SCHLEIFER '91
RS-1 Vibrio Süßwasser heterotroph soi- SAKAGUSHI '93
MC-1 Kokkus Brackwasser autotroph? N.D. MELDRUM '93aheterotroph
MV-2 Vibrio Brackwasser autotroph - MELDRUM '93b
MV-4 Vibrio Brackwasser heterotroph S2032- MELDRUM '93b
Der Stoffwechsel von Bakterien läßt sich nur an Reinkulturen untersuchen. Bis 1990ließen sich nur 2 Stämme magnetotakischer Bakterien in Kultur bringen. Erst in denletzten Jahren gelang es, einige weitere Stämme zu kultivieren, so daß sich ihre Zahlmomentan auf 8 beläuft. Neben zwei Stämmen, die unter aeroben Bedingungenwachsen (MATSUNAGA et aL, 1991; SCHLEIFER et aL, 1991), wurde ein magnetotaktisches Bakterium aus sulfidischem Milieu isoliert, das aber dennoch Magnetosomeaus Magnetit produziert (SAKAGUCHI et aL, 1993). Keine ausführliche Beschreibung gibtes von drei Isolaten, bei denen MELDRUM et aL (1993a, 1993b) Morphologie undWachstum der Magnetosome beschrieben. Diese Bakterien wurden in einem doppelten

12
Gradienten aus Sauerstoff und Schwefelwasserstoff (H2S) kultiviert. Dieses Verfahrenerscheint nach den Ergebnissen dieser Arbeit sehr sinnvoll und müßte noch weitereErfolge erbringen. Eine genauere Beschreibung des Stoffwechsels und derKulturbedingungen dieser Stämme liegt noch nicht vor.
2.2.5 Phylogenese magnetotaktischer Bakterien
Seit wenigen Jahren ist es möglich, anhand von vergleichenden Untersuchungen derErbsubstanz Bakterien stammesgeschichtlich einzuordnen und dabei den Grad derVerwandtschaft zu bestimmen. Untersuchungen an einem speziellen Teil der Erbsubstanz, der 16-S rRNA, lassen sich auch an magnetotaktischen Bakterien durchführen,die nicht in Kultur vorliegen (SCHLEIFER et al., 1991).
Durch Vergleich der Ähnlichkeit leicht mutierender Gensequenzen kann man dieuntersuchten Bakterienarten klassifizieren und im phylogenetischen Stammbaumeinordnen. Es stellte sich heraus, daß magnetotaktische Bakterien nicht einegeschlossene Gruppe bilden. Sie gehören zu verschiedenen Untergruppen derProteobakterien, zu denen auch viele unmagnetische Bakterienarten gezählt werden(SPRING et al., 1992).
Bakterien, die Eisensulfide produzieren, sind entwicklungsgeschichtlich weit entferntvon den magnetitbildenden Arten. Dies zeigt, daß sich die Magnetotaxie beiverschiedenen Bakteriengruppen unabhängig voneinander entwickelt hat (DELONG etal., 1993).
2.2.6 Extrazellulär magnetitproduzierende Bakterien
Neben den magnetotaktischen Bakterien gibt es noch eine weitere Gruppe magnetitproduzierender Bakterien. Sie baut durch Reduktion von Eisen organische Substanzoxidativ ab (LOVLEY et al., 1987). Dieser Stoffwechselweg ist sehr ineffizient. DerEnergieumsatz findet bei allen Lebewesen über das Molekül ATP (Adenosin-TriPhosphat) statt. Es besitzt drei Phosphat-Gruppen. Beim Abspalten einer Phosphatgruppe wird Energie frei. Die Energie, die bei der Oxidation von organischer Substanzmit Eisen (111) gewonnen wird, reicht jedoch nicht aus, um ein ATP-Molekül direkt zuregenerieren. Damit die Energie der Eisenreduktion ausreicht, muß über mehrereZwischenreaktionen ein entsprechendes elektrochemisches Potential aufgebaut werden.Eisenreduzierer verarbeiten daher große Mengen Fe (111). Um ein Gramm Zellsubstanzaufzubauen reduziert die Bakterienart Thiobacillus ferrooxidans 150 gEisen (FRITSCHE,1990). Deshalb bilden die magnetitproduzierenden Eisenreduzierer erhebliche Mengenan Magnetit. Auf die magnetischen Eigenschaften dieses Magnetits wird in Kapitel 8.3eingegangen.

13
3. Das Magnetodrom
Magnetotaktische Bakterien erscheinen im Lichtmikroskop meist strukturlos. Das Auf
lösungsvermögen des Mikroskops von etwa 0.5 IJm liegt in der Größenordnung derBakterien. Nur einzelne Exemplare sind größer als 5 IJm, die Mehrzahl der Kokken
besitzt einen Durchmesser von 1-2 IJm, Spirillen und Stäbchen erreichen meist 2-4 IJmLänge, sind aber unter 1 IJm breit.
Da sich die magnetotaktischen Bakterien in ihrer äußeren Form nicht von anderen
Bakterien unterscheiden, kann eine Identifizierung nur durch ihr Schwimmverhaltenparallel zu einem äußeren Magnetfeld erfolgen.
Abb. 3.1 Apparatur zur Beobachtung magnetotaktischer Bakterien ('Magnetodrom'),bestehend aus Mikroskop, drei orthogonalen Spulenpaaren, Stromversorgung, Videokamera, Videorecorder, Monitor und einem Computer zurAnsteuerung der Stromversorgung und zur Verarbeitung der Videobilder.
Zu diesem Zweck wurde eine Apparatur aufgebaut, mit der definierte Magnetfelderam Lichtmikroskop angelegt werden können. Dieses 'Magnetodrom' (Abb. 3.1) besteht
aus folgenden Komponenten:
- Lichtmikroskop,
- 3 orthogonale Spulenpaare,
- Stromquelle zur Erzeugung der Magnetfelder,
- Videokamera,

14
- Videorecorder mit Monitor und
- Computer mit Bildverarbeitungseinrichtung.
Lichtmikroskop
Es wurde ein inverses Lichtmikroskop Typ 'Zeiss Axiovert 10' verwendet. Neben derhervorragenden optischen Qualität war die mechanische Robustheit für die Wahl diesesGerätetyps ausschlaggebend. Die Konzeption des Geräts und das große Eigengewichtminimieren mechanische Schwingungen von Objekt und optischem System. Für dieUntersuchungen auf Schiffen ist dies, insbesonders bei der geringen Größe derUntersuchungsobjekte, ein wichtiger Gesichtspunkt.
Bei normalen Mikroskopen empfiehlt es sich, mit Deckglas zu arbeiten (Abb. 3.2), dasonst das Objektiv durch Kontakt mit der Probe beschmutzt werden kann. FürBeobachtungen mit Ölimmersion ist ein Deckglas unerläßlich. Im inversen Mikroskop istder Objektträger identisch mit dem Deckglas, man muß das Untersuchungsobjekt auchbei Verwendung von Ölimmersion nicht abdecken. Dadurch wird die Beobachtungwesentlich erleichtert.
I
~
M
o=====:J
M
:.--_I.c-_T'D
Abb. 3.2 Vergleich zwischen inversem (links) und normalem Mikroskop (rechts). Beiminversen Mikroskop übernimmt der Objektträger 0 gleichzeitig die Funktiondes Deckglases 0, das Mikroskopobjektiv M kann nicht in die Probe Peintauchen. Das Immersionsöl I für Beobachtungen mit dem 100x Objektivwird in der dargestellten Weise zugefügt.
Durch einen 'Long-distance-Kondensor' steht über dem Objektträger eine freie Höhevon 30 mm zur Verfügung. Die Dicke des Präparates spielt somit keine kritische Rolle,und es bleibt genügend Raum zur Arbeit am Objekt. Dieser Vorteil wird mit einemVerlust an Auflösungsvermögen erkauft. Das theoretische Auflösungsvermögen gehtvon 0.22 jJm auf 0.50 jJm zurück (GERLACH, 1985).
Zusätzlich besteht die Möglichkeit, hinter dem Objektiv eine Linse in den Strahlengang zu bringen und die Vergrößerung nochmals um den Faktor 2,5 zu erhöhen.Dadurch werden zwar Auflösungsvermögen und Tiefenschärfe herabgesetzt, aber der'Optovar-Vergrößerungsschieber' hat sich für die Bildverarbeitung sehr bewährt.
15
Um Magnetfeldinhomogenitäten durch das Mikroskop zu minimieren, mußte bei derZusammenstellung des 'Zeiss Axiovert 10' darauf geachtet werden, daß das Mikroskop
möglichst unmagnetisch ist. Aus diesem Grund wurde als Objekttisch statt des üblichenKreuztisches ein Gleittisch gewählt, bei dem eine Aluminiumplatte auf einem Ölfilm liegt.
Der Mikroskopkörper ist aus Aluminium gefertigt, alle Stahlschrauben wurden durchMessingschrauben ersetzt. Weitere Stahlteile wurden in der Institutswerkstatt ausMessing nachgebaut. In den Objektiven enthaltene Stahlfedern wurden vom Herstellerdurch Kupfer-Beryllium-Federn ersetzt. Nicht zu ersetzen waren ein Stahlring an derHalterung der Kondensorlinse und die Linsenhalterungen der Objektive, die zur genauenPositionierung der Linsen benötigt werden. Die dadurch am Beobachtungsort erzeugtenmagnetischen Störfelder konnten durch Abmagnetisieren der Objektive deutlichverringert werden.
Nach dem Abmagnetisieren erwerben die Objektive im Erdfeld innerhalb eines Tagesaber wieder eine Magnetisierung, aus der Störfelder resultieren (Zeile 2 in Tabelle 3.1).
Bei längerer Lagerung im Erdfeld ändert sich dieser Wert nur wenig. Durch dasangelegte Magnetfeld wird aufgrund der Suszeptibilität des Objektivs ein Störfelderzeugt. Dadurch wird das Magnetfeld am Probenort um etwa 2 % erhöht.
Die in Tabelle 3.1 angegebenen Störfelder verschiedener Komponenten des Mikroskops wurden mittels Förster-Sonden gemessen. Die Förster-Sonde integriert bei der
Messung über ein Volumen, in dem sich die Störfelder der Stahlbestandteile desObjektivs deutlich ändern. Daher liefert die Messung nur Näherungswerte.
Tabelle 3.1 Störfelder durch magnetische Bestandteile des Mikroskops amBeobachtung,sort. Die rechte Spalte gibt die maximalen Felddifferenzenan, die im Gesichtsfeld der Bildverarbeitungskarte auftreten. Bei denUntersuchungen werden Felder von etwa 100 IJT angelegt.
Remanente Störfeld [ IJT ] Gradient Störfeld Maximale FelddifferenzStörfelder [ IJT / cm ] [ IJT / 160 IJm ]
Objektiv vorEntmagnetisierung "'" 5 - 10 "",10 "'" 0.16
Objektiv nach ,
Entmagnetisierung "'" 0.5 - 1 "'" 1 "'" 0.016
restliche magnetischeTeile "'" 0.9 < 0.1 < 0.002
Für Routineuntersuchungen wurden Kondensor und Objektive nicht entmagnetisiert,sondern ihre Störfelder mittels der Abgleichpotentiometer kompensiert. Da es rechtaufwendig ist, die sehr kleinräumigen Feldinhomogenitäten exakt zu vermessen, sinddie Magnetbakterien selbst als Feldindikatoren benutzt worden. Das Feld wurde dabeiso lange abgeglichen, bis das Schwimmverhalten der Magnetbakterien anzeigte, daß esausreichend kompensiert war.

16
Spulenpaare
Zur Erzeugung homogener Magnetfelder wurden am Mikroskop drei zueinandersenkrecht stehende Spulenpaare angebracht. Wegen der kompakten Bauweise desAxiovert-Mikroskops konnte die klassische Helmholtz-Spulenkonfiguration (zweiparallele kreisförmige Spulen, Abstand = Spulenradius), die über einen größerenräumlichen Bereich homogene Felder erzeugt (CoLLINSaN, 1983), nicht eingesetztwerden. Der Beobachtungsraum unter dem Mikroskop ist jedoch so klein, daß dort auchmit den verwendeten rechteckigen Spulenpaaren ein hinreichend homogenesMagnetfeld erzeugt wird. Geringe Inhomogenitäten des Magnetfeldes sind, wie zuvorbeschrieben, vor allem durch die Aufmagnetisierungen eisenhaitiger Bestandteile desMikroskops bedingt.
Stromversorgung
Die Stromquelle wurde so konzipiert, daß jedes Spulenpaar unabhängig über eineneigenen Verstärker angesteuert wird. Um die Stromschwankungen gering zu halten, isteine Stromgegenkopplungs-Schaltung gewählt worden (nach ROHE, 1983 und TIETZE &
SCHENK, 1986). Der ausgegebene Strom wird dabei durch das Eingangssignal, dieWiderstandswerte des Spulenpaares und eines im Verstärker eingebautenFußwiderstandes bestimmt. Variationen der Stromstärke, die durch induzierteSpannungen entstehen, werden durch diese Schaltung gedämpft.
Bei konstantem Ansteuersignal schwankt die Stromstärke nach Erreichen der Arbeitstemperatur um weniger als 1 Prozent. Diese Schwankungen werden induktiv auf dieSpulen übertragen. Neben der Einstreuung durch die technischen Wechselströme imLabor trägt ein der Universität Bremen benachbarter Rundfunksendemast zumStörsignal bei. Zur Langzeitstabilisierung der Stromstärke wurde darauf geachtet, daßder Fußwiderstand besonders hohe Qualitätskriterien erfüllt.
Kompensation des Erdmagnetfeldes
Bevor man ein definiertes Magnetfeld zur Beobachtung der Magnetbakterien anlegt,muß das Erdmagnetfeld kompensiert werden. Im Labor wird das Erdmagnetfeld durchStahlarmierungen des Gebäudes verändert, auf Schiffen durch die Stahlwände, vorallem aber durch den Schiffskurs, der das Mikroskop immer wieder in eine neueRichtung zum Erdmagnetfeld bringt. Der Feldabgleich muß daher schnell und genauerfolgen. Das Magnetfeld läßt sich bei einer Maximalamplitude von 300 IJT in Schrittenvon 10-4 IJT einstellen.
Abgleichsignal und Ansteuersignal werden vor der Verstärkung addiert. Dazu wurdeeine Additionsschaltung benutzt (ROHE, 1983; TIETZE & SCHENK, 1986). Beide Strömefließen über ein und dasselbe Spulenpaar. Die Apparatur wird dadurch wesentlichhandlicher als bei Benutzung getrennter Spulen für Abgleich und Ansteuerung.

17
Ansteuersignal
Das Magnetfeld am Lichtmikroskop kann auf drei verschiedene Weisen erzeugt undgeregelt werden:
Analoger Funktionsgenerator:Eine im Verstärker eingebaute Platine mit einem analogen Funktionsgenerator (AnalogDevices) erzeugt auf zwei Kanälen jeweils eine um 90° phasenverschobene Sinusspannung. Legt man die Spannungen an zwei zueinander senkrecht stehendeSpulenpaare, so ergibt sich ein rotierendes Magnetfeld konstanter Stärke und ausreichender Homogenität, durch das magnetotaktische Bakterien gezwungen werden, aufKreisbahnen zu schwimmen.
Joystick:Ein handelsüblicher Joystick wurde so umgebaut, daß das Magnetfeld parallel zurhorizontalen Richtung anliegt, die durch den Joystick vorgegeben wird. Durch Drehendes Griffs kann auch die vertikale Feldkomponente angesprochen werden. Die
Schwimmrichtung der Magnetbakterien läßt sich somit in allen drei Raumrichtungensteuern. Der Joystick erwies sich als unerläßlich zum Erkennen von Magnetbakterien,die nicht kontinuierlich schwimmen (Kap. 6.3). Legt man mit dem Joystick ein
Magnetfeld in einer bestimmten Richtung an, so bewegen sich die Bakterien, falls sieschwimmen, in die jeweils vorgegebene Richtung. Ebenso wird die Präparation fürelektronenmikroskopische Aufnahmen sehr erleichtert.
Digitaler Funktionsgenerator:Um beliebige Schwimmbahnen definieren zu können, wurde der Personal-Computer miteinem programmierbaren Digital-Analog-Wandler ausgestattet. Das ausgewählte Modellstammt von der Firma Sorcus (Heidelberg), hat vier Kanäle und erlaubt die Ausgabe vonDaten in einem zeitlichen Abstand von 25 IJs; bei zwei Ausgabekanälen entspricht dies
maximal 20000 Spannungswerten pro Sekunde. Da die Digital-Analog-Karte eineneigenen Prozessor besitzt (Z-80), arbeitet sie unabhängig vom Rechner und muß nurgestartet werden.
Es können etwa 20000 Funktionswerte pro Periode ausgegeben werden,verteilt aufdie Anzahl der verwendeten Kanäle. Aus praktischen Gründen wurden für eine SinusPeriode nicht mehr als 500 Funktionswerte ausgegeben.
Videokamera
Um die Bewegung der Bakterien aufzuzeichnen, wurde eine schwarzweiß Videokameraan das Mikroskop angeschlossen. Beim verwendeten Modell 'Sony AVC-D7CE' handeltes sich um eine CCD-Kamera (charge coupled device) mit 756 x 581 Bildpunkten. DieKamera hat eine Auflösung von 380 Linien, die Verschlußzeit ist von 1/60 bis 1/10000

18
Sekunde einstellbar, wodurch die schnell beweglichen Magnetbakterien sehr scharfabgebildet werden können. Magnetfelder verzerren das Bild bei CCD-Kameras nicht.
Bildverarbeitungskarte
Eine Bildverarbeitungskarte 'MVP-AT' der Firma Matrox (München) ermöglicht es, imFalle einfacherer Algorithmen Bilder in Echtzeit zu bearbeiten. Auch diese Karte besitzteinen eigenen Prozessor. Er kann Bildbearbeitungsbefehle ausführen, die in einer'Imager-AT' genannten Softwarebibliothek mitgeliefert wurden. Die Befehle müssenallerdings vom Prozessor des Rechners einzeln aufgerufen werden, weshalb die Kartenicht im 'stand-alone-Betrieb' arbeiten kann. Zur Darstellung der (bearbeiteten) Bildersteht neben dem nachfolgend beschriebenen Fernsehmonitor ein weiterer Bildschirmzur Verfügung.
Die Bildverarbeitungskarte besitzt vier Bildspeicher für Bilder mit 512 x 512 Punkten.
Die Aufnahmen können in etwa 0.3 Sekunden auf die Festplatte des Rechnersübertragen werden. Die genaue Funktionsweise der Bildverarbeitungskarte ist imAnhang A dargestellt.
Fernsehmonitor
Das Videosignal wird direkt auf einem Fernsehschirm dargestellt. Insofern ist man beiden Untersuchungen nicht an das Mikroskop gebunden, die Beobachtungen können
auch über den Monitor erfolgen. Dies hat sich auf den Schiffsexpeditionen besondersbei stärkerem Seegang bewährt.
Auch für die Bildverarbeitung ist der Monitor unerläßlich. Die Aufnahmen werden überdie Tastatur des Rechners in Echtzeit bearbeitet, über den Monitor lassen sich dieeinzelnen Szenen auswählen und kontrollieren.
Videorecorder
Als preisgünstige Möglichkeit zur Speicherung großer Mengen von Bildern wurde einhandelsüblicher Videorecorder benutzt. Die aufgezeichneten Aufnahmen können späterzur Bildverarbeitung herangezogen werden.

19
4. Untersuchung der Schwimmbewegung magnetotaktischer Bakterien
Mit der Bildverarbeitungskarte (Kap. 3) lassen sich, durch Überlagerung aufeinander
folgender Bilder, Spuren der Schwimmbewegung magnetotaktischer Bakterien auf
zeichnen. Entsprechend der zeitlichen Auflösung der Videokamera von 50 Halbbildern
pro Sekunde werden je Sekunde 50 Positionen eines Bakteriums erfaßt. Da bei einem
Halbbild wird nur jede zweite Bildzeile belichtet wird, entsteht ein Abbild aus hellen unddunklen Streifen (Anhang A).
Die Auswertung der Bewegungsspuren erlaubt eine sichere Identifizierung der
Magnetbakterien und ermöglicht es, die Geschwindigkeit der Schwimmbewegung sowie
das magnetische Moment der Bakterien zu bestimmen.
4.1 Unterscheidung verschiedener Bewegungsformen
Abbildung 4.1 zeigt verschiedene Bewegungsspuren magnetotaktischer Bakterien in
homogenen, betragsmäßig konstanten Magnetfeldern. Bei den Szenen abis c wurde einmit konstanter Frequenz rotierendes Magnetfeld angelegt. Dabei bewegt sich in Szene a
eine große Anzahl magnetotaktischer Bakterien auf nahezu perfekt kreisförmigen
Bahnen. Diese Kreisbewegung wird von einer mehr oder weniger ausgeprägten
'Wackelbewegung' überlagert (b,c). Dieses seitliche Abweichen von der mittlerenBewegungsbahn ist ein periodischer Vorgang, wobei Amplitude und Periodendauer
(0.04 bis 0.28 s) der seitlichen Auslenkung für verschiedene Individuen ganz unter
schiedlich sind. Besonders deutlich wird dies in Abbildung 4.1 d. Hier wurde die
Feldrichtung abwechselnd nach oben und unten, dann nach links und rechts
vorgegeben
a) b)


20
c) d)
Abb. 4.1 Schwimmbahnen magnetotaktischer Bakterien. Jede Bahn ist mit 50 Bildernpro Sekunde aufgenommen. Der Maßstab ist für alle vier Bilder gleich undbeträgt 100 ~m. Bei den Szenen abis c waren rotierende Magnetfelderangelegt, bei Szene d zeigte die Feldrichtung abwechselnd nach oben undunten, dann nach links und rechts.
Ein Ausschnitt einer Kreisbahn, der bei stärkerer Vergrößerung aufgenommen wurde(Abb. 4.2), illustriert die teilweise extreme Auslenkung von der mittleren Bewegungs
richtung. In dieser Aufnahme wird deutlich, daß es sich dabei um eine helicale
Bewegung handelt (siehe Kapitel 4.3).
Abb. 4.2 Detailaufnahme der Bewegungsbahn einer Magnetokokke. Das Bakteriumdurchschwimmt hier nur einen kleinen Ausschnitt eines Kreises.

21
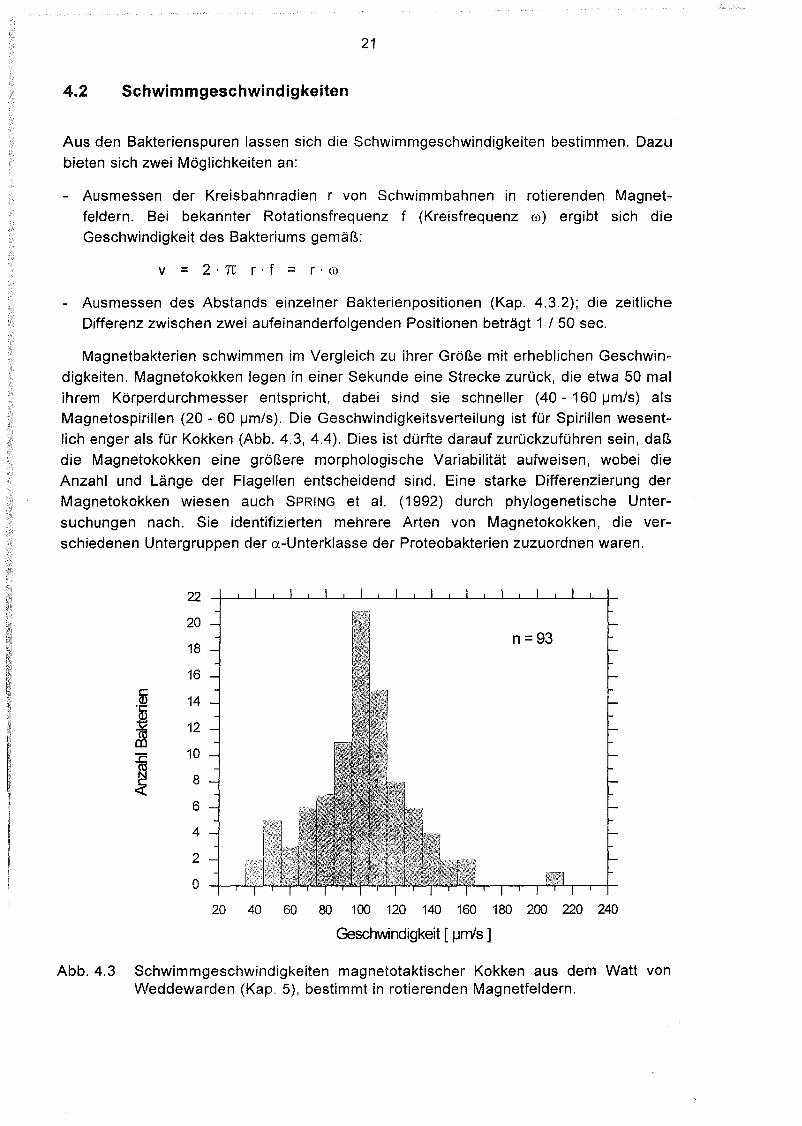
4.2 Schwimmgeschwindigkeiten
Aus den Bakterienspuren lassen sich die Schwimmgeschwindigkeiten bestimmen. Dazubieten sich zwei Möglichkeiten an:
- Ausmessen der Kreisbahnradien r von Schwimmbahnen in rotierenden Magnetfeldern. Bei bekannter Rotationsfrequenz f (Kreisfrequenz m) ergibt sich dieGeschwindigkeit des Bakteriums gemäß:
v = 2·1t r·f = r·m
- Ausmessen des Abstands einzelner Bakterienpositionen (Kap. 4.3.2); die zeitlicheDifferenz zwischen zwei aufeinanderfolgenden Positionen beträgt 1 / 50 sec.
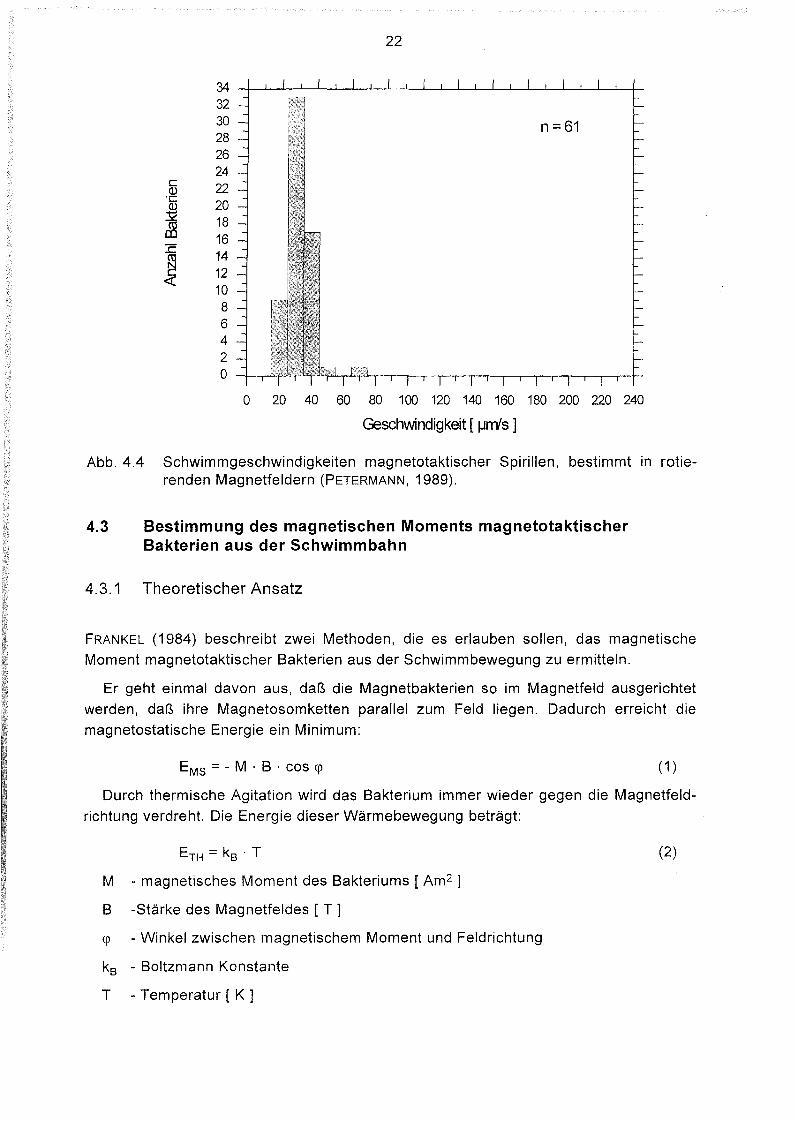
Magnetbakterien schwimmen im Vergleich zu ihrer Größe mit erheblichen Geschwindigkeiten. Magnetokokken legen in einer Sekunde eine Strecke zurück, die etwa 50 malihrem Körperdurchmesser entspricht, dabei sind sie schneller (40 - 160 IJm/s) alsMagnetospirillen (20 - 60 IJm/s). Die Geschwindigkeitsverteilung ist für Spirillen wesentlich enger als für Kokken (Abb. 4.3, 4.4). Dies ist dürfte darauf zurückzuführen sein, daßdie Magnetokokken eine größere morphologische Variabilität aufweisen, wobei dieAnzahl und Länge der Flagellen entscheidend sind. Eine starke Differenzierung derMagnetokokken wiesen auch SPRING et al. (1992) durch phylogenetische Untersuchungen nach. Sie identifizierten mehrere Arten von Magnetokokken, die verschiedenen Untergruppen der a-Unterklasse der Proteobakterien zuzuordnen waren.
22
20
18n=93
16c:Q) 14.e:
j 12
::E 10
~ 8
6
4
2
0
20 40 60 80 100 120 140 160 180 200 220 240
Geschwindigkeit [ lJm's ]
Abb.4.3 Schwimmgeschwindigkeiten magnetotaktischer Kokken aus dem Watt vonWeddewarden (Kap. 5), bestimmt in rotierenden Magnetfeldern.
22
34
3230 n =61282624
c 22Q)'e 20~~
1816
1:
~14121086420
0 20 40 60 80 100 120 140 160 180 200 220 240
Geschwindigkeit [ Ilm'S ]
Abb. 4.4 Schwimmgeschwindigkeiten magnetotaktischer Spirillen, bestimmt in rotierenden Magnetfeldern (PETERMANN, 1989).
4.3 Bestimmung des magnetischen Moments magnetotaktischerBakterien aus der Schwimmbahn
4.3.1 Theoretischer Ansatz
FRANKEL (1984) beschreibt zwei Methoden, die es erlauben sollen, das magnetische
Moment magnetotaktischer Bakterien aus der Schwimmbewegung zu ermitteln.
Er geht einmal davon aus, daß die Magnetbakterien so im Magnetfeld ausgerichtet
werden, daß ihre Magnetosomketten parallel zum Feld liegen. Dadurch erreicht die
magnetostatische Energie ein Minimum:
EMS =- M . B . cos cp (1 )
Durch thermische Agitation wird das Bakterium immer wieder gegen die Magnetfeld
richtung verdreht. Die Energie dieser Wärmebewegung beträgt:
ETH =kB . T
M - magnetisches Moment des Bakteriums [ Am 2 1
B -Stärke des Magnetfeldes [ T 1
cp - Winkel zwischen magnetischem Moment und Feldrichtung
kB - Boltzmann Konstante
T - Temperatur [ K ]
(2)

23
Der Ausrichtungsgrad der Magnetbakterien im Magnetfeld unter Einfluß der Wärme
bewegung läßt sich nach FRANKEL (1984) mit den gleichen Gesetzen beschreiben, die
für die Orientierung einzelner atomarer magnetischer Momente im Magnetfeld gelten.
Der Mittelwert des Cosinus des Winkels zwischen magnetischem Moment (Ausrichtung
der Magnetosomkette) und Magnetfeld folgt der Langevin Funktion L:
< cos cp > =coth ( ß) - 1 / ß=L ( ß) (3)
wobei ß als Quotient aus magnetostatischer und thermischer Energie definiert ist:
ß =( M . B ) / ( ks . T ) (4)
In stärkeren Magnetfeldern sollten die Bakterien demnach besser ausgerichtet
werden. Mit der Annahme, daß sie sich parallel zu ihrem magnetischen Moment
bewegen, sollten sie bei höheren Feldstärken eine größere Geradeausgeschwindigkeit
erreichen:
VII =va . < cos cp > (5)
Va ist die Eigengeschwindigkeit des Bakteriums, VII die effektive Geschwindigkeit in
Feldrichtung. Bei bekannter Magnetfeldstärke läßt sich das magnetische Moment dann
aus dem Mittelwert des Winkels zwischen Schwimm- und Feldrichtung sowie der daraus
resultierenden Schwimmgeschwindigkeit berechnen.
Als zweites Verfahren zur Bestimmung der magnetischen Momente hat Frankel
(1984) die U-Turn-Methode vorgeschlagen. Wird das Magnetfeld umgepolt, so müssen
die Magnetbakterien einen Bogen von 180 0 schwimmen, um der Feldrichtung folgen zu
können. Aus der Weite dieses Bogens soll sich das magnetische Moment des
Bakteriums bestimmen lassen.
Unmittelbar nach der Umpolung des Magnetfeldes beträgt der Winkel cp zwischen der
Feldrichtung und dem magnetischen Moment des Bakteriums etwa 1800• Das Bakterium
erfährt ein magnetisches Drehmoment DMAG :
DMAG =M . B . sin cp (6)
(7)
Die Drehung des Magnetbakteriums in die Feldrichtung, wird durch die viskose
Reibung der umgebenden Flüssigkeit abgebremst. Das Drehmoment der viskosen
Reibung ist dem magnetischen Drehmoment entgegengesetzt. Für ein kugelförmiges
Bakterium läßt sich das Drehmoment der viskosen Reibung DvlS beschreiben durch:
DVIS =8 . TC . r3 . 11 . dcpfdt
r - (effektiver) viskoser Radius des Bakteriums
11 - Viskosität der umgebenden Flüssigkeit
d<pfdt - Winkelgeschwindigkeit, mit der das Bakterium in Feldrichtung gedreht wird

24
Für ein kugelförmiges Bakterium gilt in erster Näherung (dq>/dt = konst.):
M . B . sin q> = 8 . 7t . r3 . 11 . dq>/dt (8)
Andererseits läßt sich die Geschwindigkeitskomponente der Schwimmbewegung
senkrecht zum Magnetfeld beschreiben durch:
v.L = va . sin q>
Aus den Gleichungen (8) und (9) folgt:
v.L =vo·"Co·dq>/dt
"Co faßt alle Konstanten zusammen:
"Co = (8 . 7t . r3 . 11) / ( M . B )
(9)
(10)
(11 )
Wenn das Magnetfeld umgepolt wird, ändert sich der Winkel um 180 0• Die Weite W
des nach Umpolung des Magnetfeldes geschwommenen Kreisbogens ergibt sich als
Integral über die Gechwindigkeit senkrecht zum Magnetfeld:
t1 7t
W = Iv, . dt = Ivo . ~o . dep =Vo . ~o . • (12)
oDaraus läßt sich das magnetische Moment berechnen:
M = (8 . 7t2 . r3 . 11 . va) / ( W . B ) (13)
Beide Ansätze wurden mit dem Magnetodrom überprüft. Die Magnetfelder lassen sich
nach Stärke und Richtung vorgeben (Kap. 3), die Schwimmbewegungen mit Hilfe der
Bildverarbeitung exakt ausmessen.
Abbildung 4.5 zeigt eine Bakterienspur, die im folgenden exemplarisch ausgewertet
wird. Aus den (zentralen) Pixelpositionen des Magnetbakteriums können nach
Umrechnen des Maßstabs Geschwindigkeiten, Beschleunigungen, Winkel und Winkel
änderungen des Bakteriums sowie der zeitliche Verlauf und die Ortsabhängigkeit dieser
Parameter bestimmt werden.
Die digitalisierten Pixel positionen der Bakterienspur (Abb. 4.5) sind in Abbildung 4.6
dargestellt. Aus den Ziffern in den Positionssymbolen läßt sich der Verlauf der
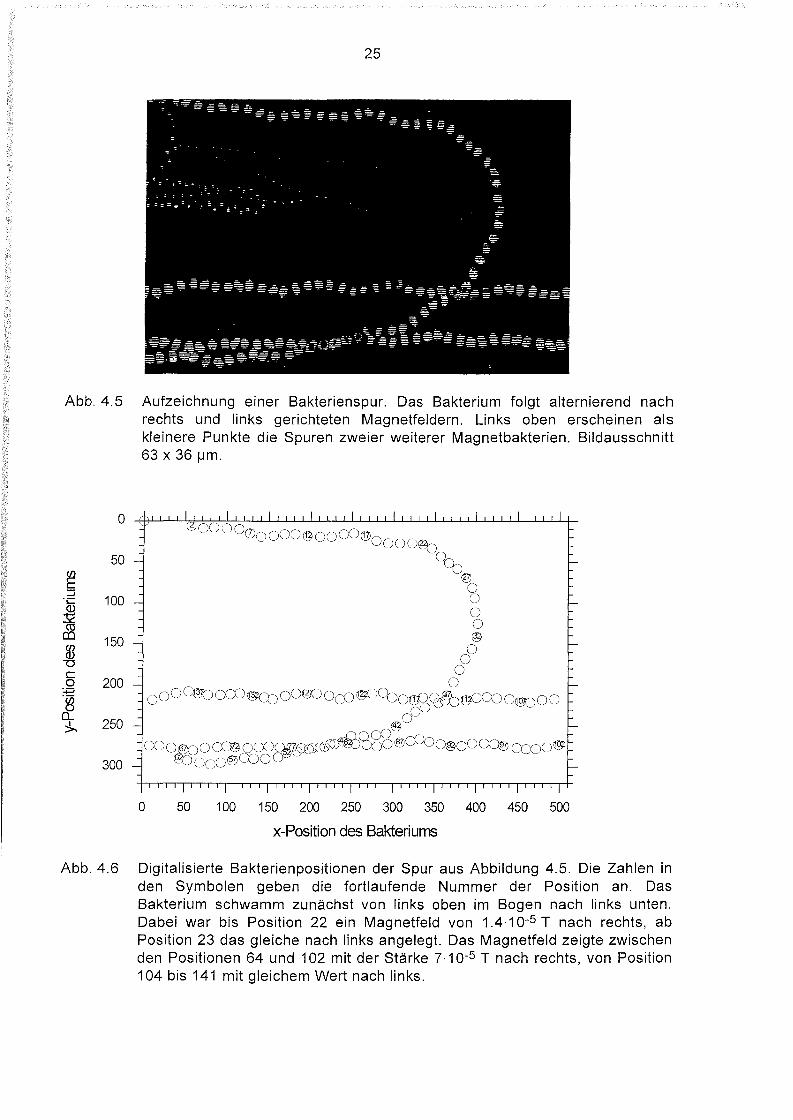
Bewegung ablesen. Von Position 2 bis 22 war ein Magnetfeld der Stärke 1.4.10-5 T nach
rechts, von Position 23 bis 62 mit gleicher Stärke nach links gerichtet. Von Position
64 bis 102 zeigte ein Feld der Stärke 7.10-5 T nach rechts, zwischen Position
104 und 141 ein Feld gleicher Intensität nach links. Die daraus berechneten momen
tanen Geschwindigkeiten sind in Abbildung 4.7 aufgetragen.
25
Abb. 4.5 Aufzeichnung einer Bakterienspur. Das Bakterium folgt alternierend nachrechts und links gerichteten Magnetfeldern. Links oben erscheinen alskleinere Punkte die Spuren zweier weiterer Magnetbakterien. Bildausschnitt63 x 36 IJm.
~::J
100'C
~co
a:J 150~
"'0C0 200:p.~
0....250>-
300
o 50 100 150 200 250 300 350 400 450 500
x-Position des Bakteriums
Abb. 4.6 Digitalisierte Bakterienpositionen der Spur aus Abbildung 4.5. Die Zahlen inden Symbolen geben die fortlaufende Nummer der Position an. DasBakterium schwamm zunächst von links oben im Bogen nach links unten.Dabei war bis Position 22 ein Magnetfeld von 1.4.10-5 T nach rechts, abPosition 23 das gleiche nach links angelegt. Das Magnetfeld zeigte zwischenden Positionen 64 und 102 mit der Stärke 7.10-5 T nach rechts, von Position104 bis 141 mit gleichem Wert nach links.
Zeitdauer [ s l0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4
120 --P-uJ..U.J..LJ.u..u.J.J..U..Lilu.w.w.W.1.U.llJeukUJ..U.J..J.u.uw..w.I..wu.l.l..U.J-
IJ)
-- 100~
o 10 20 30 40 ~ 60 70
Nurrmer der Bakterienposition
26
Zeitdauer [ s l1.2 1.4 1.6 1.8 2.0 2.2 2.4 2.6 2.8 3.0
120 -f=J..U.llJ.l.u..u.w.ulllWJ.uJ.uJ..U.J..LuL.u.l.lliJ.JJ.u..u.w.ul.JJ..U..IJL1.U.L.UJ..U.J..u.L..wI..1.U..l.J4-
100
60
40
20
60 70 00 ro 10J 110 120 130 140 150
Nurrmer der Bakterienposition
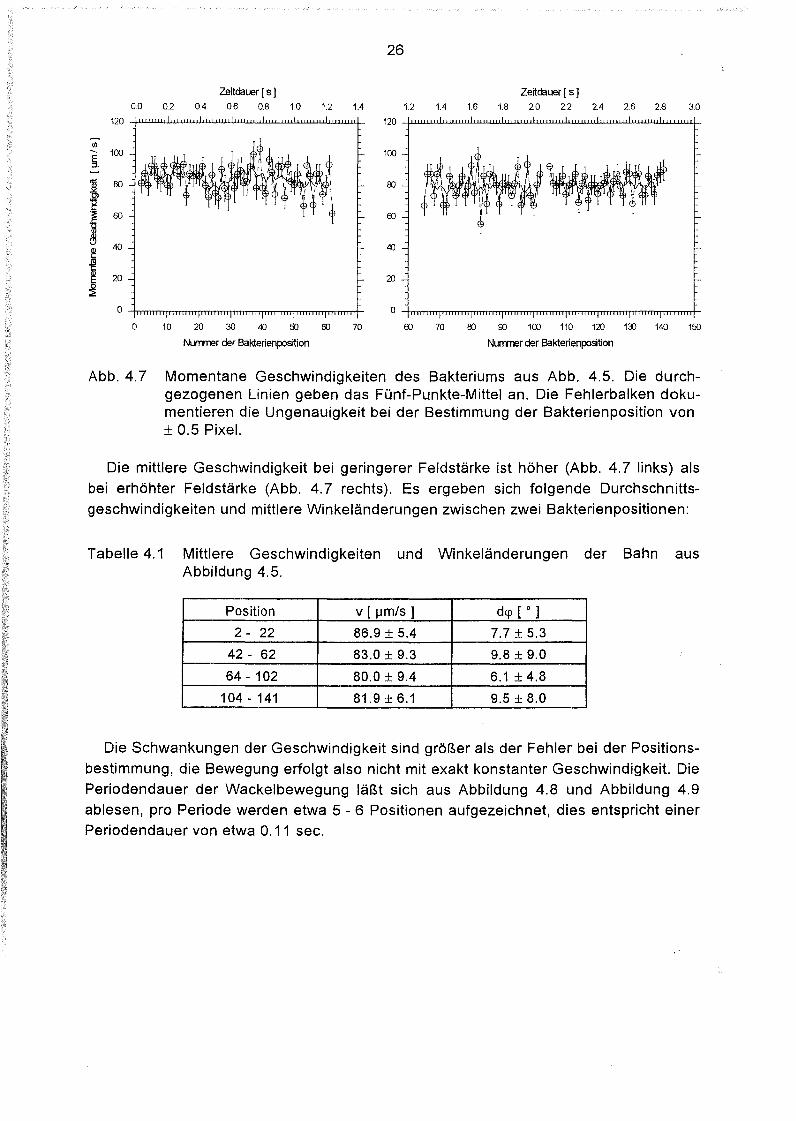
Abb. 4.7 Momentane Geschwindigkeiten des Bakteriums aus Abb. 4.5. Die durchgezogenen Linien geben das Fünf-Punkte-Mittel an. Die Fehlerbalken dokumentieren die Ungenauigkeit bei der Bestimmung der Bakterienposition von± 0.5 Pixel.
Die mittlere Geschwindigkeit bei geringerer Feldstärke ist höher (Abb. 4.7 links) alsbei erhöhter Feldstärke (Abb. 4.7 rechts). Es ergeben sich folgende Durchschnittsgeschwindigkeiten und mittlere Winkeländerungen zwischen zwei Bakterienpositionen:
Tabelle 4.1 Mittlere Geschwindigkeiten und Winkeländerungen der Bahn ausAbbildung 4.5.
Position v [ IJm/s ] dcp [ 0 ]
2 - 22 86.9 ± 5.4 7.7 ± 5.3
42 - 62 83.0 ± 9.3 9.8 ± 9.0
64 - 102 80.0 ± 9.4 6.1 ± 4.8
104 - 141 81.9 ± 6.1 9.5 ± 8.0
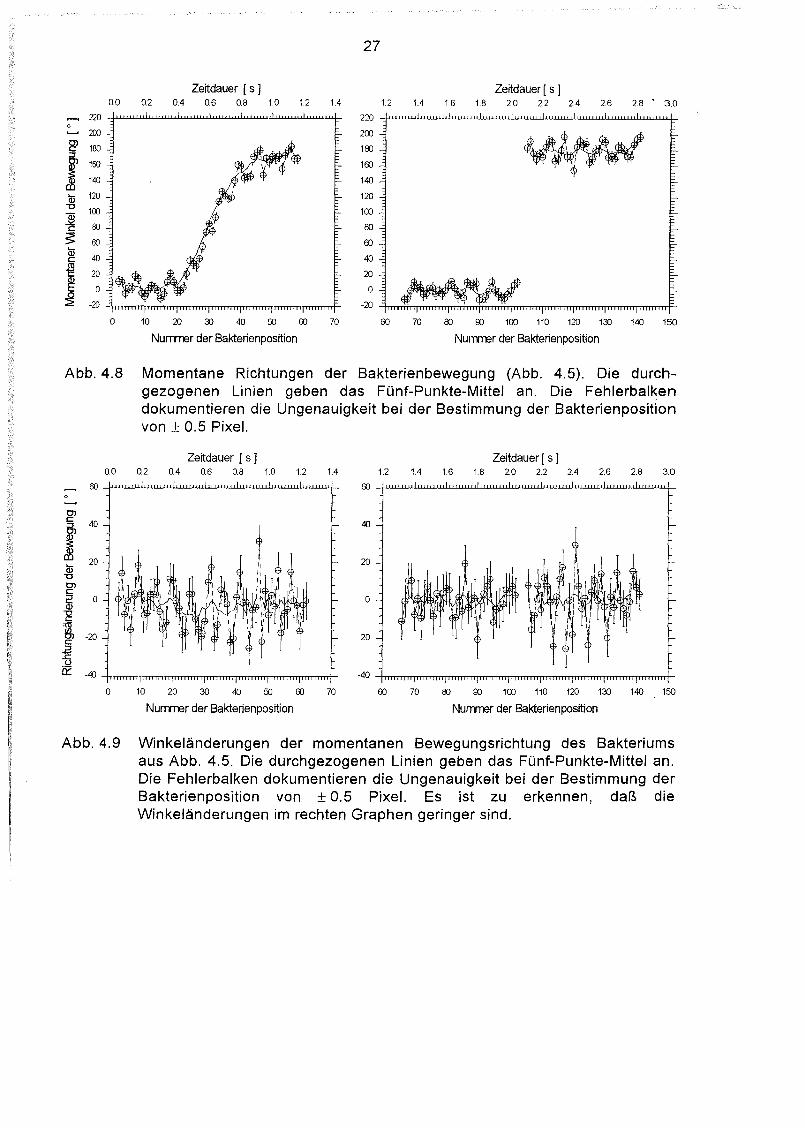
Die Schwankungen der Geschwindigkeit sind größer als der Fehler bei der Positionsbestimmung, die Bewegung erfolgt also nicht mit exakt konstanter Geschwindigkeit. DiePeriodendauer der Wackelbewegung läßt sich aus Abbildung 4.8 und Abbildung 4.9ablesen, pro Periode werden etwa 5 - 6 Positionen aufgezeichnet, dies entspricht einerPeriodendauer von etwa 0.11 sec.
27
Zeitdauer [s 10.0 0.2 0.4 0.6 0.8 10 1.2 1.4
Zeitdauer [ s112 1.4 16 18 2.0 2.2 2.4 2.6 28' 3.0
2~ -j-u-u=uL..cUllUu1n.JJ.LLU...L.u.LU.U...L.u.u.w.iliu.~J.uu"-L1.WJ.uu"-L1.WJ.uu.J-U.W4-
200
180
160
1~
1~
100
80
60
40
~
o
-~ imrnTn'TjTTTTTTTTl"jTTilriTmTfrrrrrITrfTTTT'"lTTTfTTTTTrrrrpTTTTTTrrfTmrrmrnmmnf-o m ~ ~ ~ ~ 00 m
NUll'Tll3r der Bakterienposition60 70 80 00 100 110 1~ 1~ 140 150
Nurrrrer der Bakterienposition
Abb.4.8 Momentane Richtungen der Bakterienbewegung (Abb. 4.5). Die durchgezogenen Linien geben das Fünf-Punkte-Mittel an. Die Fehlerbalkendokumentieren die Ungenauigkeit bei der Bestimmung der Bakterienpositionvon ± 0.5 Pixel.
Zeitdauer [s 10.0 0.2 0.4 0.6 0.8 1~ 12 1.4
Zeitdauer [ s 11.2 14 16 18 2.0 22 2.4 2.6 2.8 3.0
70 80 00 100 110 1~ 1~ 140 150
NUll'Tll3r der Bakterienposition
40
o
~
-20
10 20 30 40 ~ 00 70
NUll'Tll3r der Bakterienposmon
o
~ 00 -t'-'-UJ..l..Uu.1..u.w.u.J..u....ll.LU..lu.uw.wL.ull.LU.huJ..w.ulJJJ.lll.1.Uf-o
Abb. 4.9 Winkeländerungen der momentanen Bewegungsrichtung des Bakteriumsaus Abb. 4.5. Die durchgezogenen Linien geben das Fünf-Punkte-Mittel an.Die Fehlerbalken dokumentieren die Ungenauigkeit bei der Bestimmung derBakterienposition von ± 0.5 Pixel. Es ist zu erkennen, daß dieWinkeländerungen im rechten Graphen geringer sind.
28
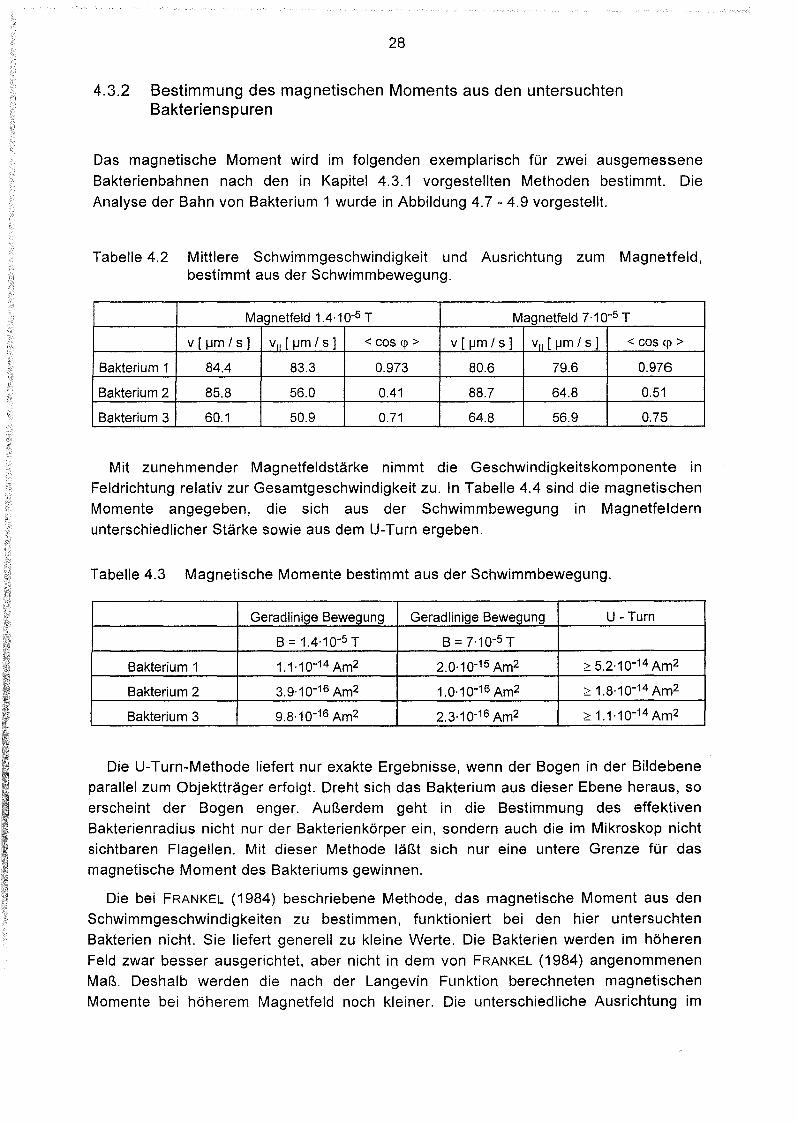
4.3.2 Bestimmung des magnetischen Moments aus den untersuchtenBakterienspuren
Das magnetische Moment wird im folgenden exemplarisch für zwei ausgemessene
Bakterienbahnen nach den in Kapitel 4.3.1 vorgestellten Methoden bestimmt. Die
Analyse der Bahn von Bakterium 1 wurde in Abbildung 4.7 - 4.9 vorgestellt.
Tabelle 4.2 Mittlere Schwimmgeschwindigkeit und Ausrichtung zum Magnetfeld,bestimmt aus der Schwimmbewegung.
Magnetfeld 1.4.10-5 T Magnetfeld 7.10-5 T
v [ IJm / s ] VII [ IJm / s] < Gas <p > V [ IJm / s ] VII [ IJm / s] < Gas <p >
Bakterium 1 84.4 83.3 0.973 80.6 79.6 0.976
Bakterium 2 85.8 56.0 0.41 88.7 64.8 0.51
Bakterium 3 60.1 50.9 0.71 64.8 56.9 0.75
Mit zunehmender Magnetfeldstärke nimmt die Geschwindigkeitskomponente in
Feldrichtung relativ zur Gesamtgeschwindigkeit zu. In Tabelle 4.4 sind die magnetischen
Momente angegeben, die sich aus der Schwimmbewegung in Magnetfeldern
unterschiedlicher Stärke sowie aus dem U-Turn ergeben.
Tabelle 4.3 Magnetische Momente bestimmt aus der Schwimmbewegung.
Geradlinige Bewegung Geradlinige Bewegung U - Turn
B=1.4·10-5 T B = 7.10-5 T
Bakterium 1 1.1.10-14 Am2 2.0,10-15 Am2 ;::: 5.2.10-14 Am2
Bakterium 2 3.9.10-16 Am2 1.0'10-16 Am2 ;::: 1.8.10-14 Am2
Bakterium 3 9.8.10-16 Am2 2.3,10-16 Am2 ;::: 1.1.10-14 Am2
Die U-Turn-Methode liefert nur exakte Ergebnisse, wenn der Bogen in der Bildebene
parallel zum Objektträger erfolgt. Dreht sich das Bakterium aus dieser Ebene heraus, soerscheint der Bogen enger. Außerdem geht in die Bestimmung des effektiven
Bakterienradius nicht nur der Bakterienkörper ein, sondern auch die im Mikroskop nichtsichtbaren Flagellen. Mit dieser Methode läßt sich nur eine untere Grenze für das
magnetische Moment des Bakteriums gewinnen.
Die bei FRANKEL (1984) beschriebene Methode, das magnetische Moment aus den
Schwimmgeschwindigkeiten zu bestimmen, funktioniert bei den hier untersuchten
Bakterien nicht. Sie liefert generell zu kleine Werte. Die Bakterien werden im höheren
Feld zwar besser ausgerichtet, aber nicht in dem von FRANKEL (1984) angenommenenMaß. Deshalb werden die nach der Langevin Funktion berechneten magnetischen
Momente bei höherem Magnetfeld noch kleiner. Die unterschiedliche Ausrichtung im

29
Magnetfeld wird offensichtlich nicht nur von der thermischen Agitation gestört. Um eintieferes Verständnis dafür zu finden, muß man reale Schwimmbahnen betrachten.
Die Schwimmbahnen, die man durch die Bildverarbeitung erhält, sind eine Projektionder dreidimensionalen Bahn auf eine Ebene. Die magnetotaktischen Bakterien bewegensich mittels Flagellen vorwärts (Kap. 2.2). Die schraubende Bewegung der helicalenFlagellen sorgt für den Vortrieb der Bakterien. (Bei den schraubenförmig gebautenSpirillen wird die Vorwärtsbewegung außerdem durch die Rotation des Körpersunterstützt. )
Da der Drehimpuls des Bakteriums und des vom Bakterium bewegten Wasserskonstant (gleich Null) sein muß, rotieren Flagellen und Körper mit unterschiedlichenWinkelgeschwindigkeiten in entgegengesetzter Richtung.
FLAGELLUM
cAbb. 4.10 Schematische Darstellung der Rotationen von Flagellum und Bakterienkörper
bei der Vorwärtsbewegung der Magnetbakterien.
Wenn ein Bakterium unsymmetrisch gebaut ist, das heißt, wenn die Längsachse desBakterienkörpers (bei kugelförmigen Bakterien definiert durch die Richtung desmagnetischen Moments) und die Achse der Flagellen nicht in einer Linie liegen, kommtes zu der in Abbildung 4.1 und Abbildung 4.2 dargestellten Wackelbewegung(STEINBERGER, 1990). Dies ist beispielsweise gegeben, wenn die Flagellen exzentrischam Körper angebracht sind. Der Bakterienkörper bewegt sich auf einer helicalen Bahn.In der Projektion erscheint dies als Wackelbewegung.
Die Orientierung des Bakteriums im Magnetfeld wird nicht nur von der thermischenAgitation gestört, sondern auch von der Eigenbewegung des Bakteriums. Daher mußder Ansatz von FRANKEL (1984) versagen.
4.4 Bestimmung des magnetischen Moments in rotierendenMagnetfeldern
In rotierenden Magnetfeldern konstanter Stärke und sukzessiv erhöhter Frequenzwerden die in Abbildung 4.12 gezeigten Schwimmbahnen beobachtet. Bei niedrigenFrequenzen (f < 1 Hz => co < 6 Hz) können alle Magnetbakterien dem rotierenden Feldfolgen und schwimmen auf Kreisbahnen; der Winkel zwischen Schwimm- und Feldrichtung ist klein und in etwa konstant. Mit zunehmender Frequenz vergrößert sichdieser Winkel, die Kreisbahnen werden enger. Ab einer - für jedes Bakterium unterschiedlichen - kritischen Frequenz können die Bakterien dem Feld nicht mehr folgen.

30
Der Winkel zwischen Schwimm- und Feldrichtung ist nicht mehr konstant, die Bakterienbewegen sich auf 'spaghettiförmigen' Bahnen (Abb. 4.12).
Abb. 4.11 Schematische Darstellung der Drehung eines Magnetbakteriums im rotierenden Magnetfeld. M bezeichnet das magnetische Moment, B die momentane Richtung des Magnetfeldes, cp den Winkel zwischen beiden Richtungen.Bei konstanter Stärke und niedriger Winkelgeschwindigkeit des Feldes ist derWinkel cp konstant.
Aus der Änderung des Bewegungsverhaltens bei der kritischen Frequenz läßt sich,ausgehend von Formel (8), das magnetische Moment der Bakterien herleiten. Solangedie Winkelgeschwindigkeit des Bakteriums dcp/dt gleich der Winkelgeschwindigkeit desrotierenden Feldes co ist, gilt:
M . B . sin cp = 8 . 1t . r3 . 11 . co (14)
Wird bei konstanter Stärke die Rotationsfrequenz des Feldes erhöht, so vergrößert
sich der Winkel cp zwischen Feld und magnetischem Moment. Für cp =90° erreicht dasmagnetische Drehmoment seinen maximalen Wert. Wird die Frequenz co des Feldesüber diesen kritischen Wert hinaus erhöht, dann wird die viskose Reibung so groß, daßdas Bakterium dem Feld nicht mehr folgen kann. Es ergeben sich 'spaghettiförmi,ge'Spuren. Für die kritische Frequenz ist sin cp == 1, Gleichung (14) vereinfacht sich zu:
M . B = 8 . 1t . r3 . 11 • co .'I krlt
und ergibt nach M aufgelöst:
M =8 . 1t . r3 . 11 . co ' / B'I krlt
(15)
(16)
31
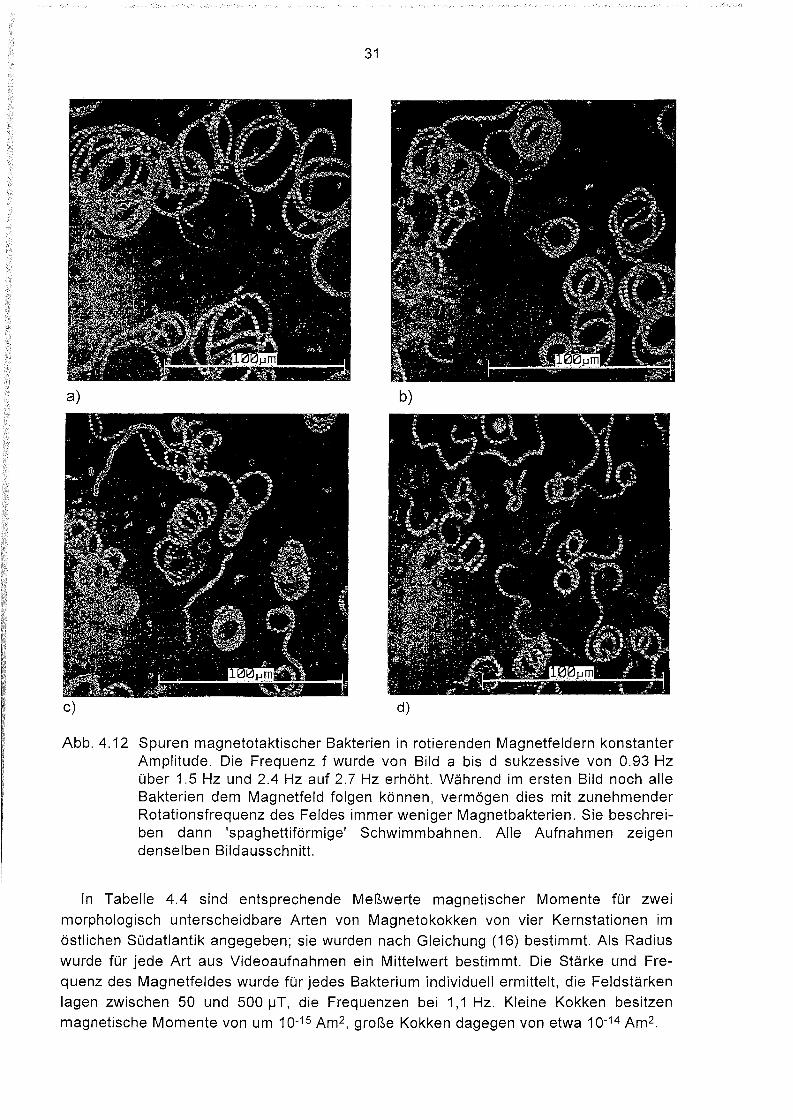
a) b)
c) d)
Abb. 4.12 Spuren magnetotaktischer Bakterien in rotierenden Magnetfeldern konstanterAmplitude. Die Frequenz f wurde von Bild abis d sukzessive von 0.93 Hzüber 1.5 Hz und 2.4 Hz auf 2.7 Hz erhöht. Während im ersten Bild noch alleBakterien dem Magnetfeld folgen können, vermögen dies mit zunehmenderRotationsfrequenz des Feldes immer weniger Magnetbakterien. Sie beschreiben dann 'spaghettiförmige' Schwimmbahnen. Alle Aufnahmen zeigendenselben Bildausschnitt.
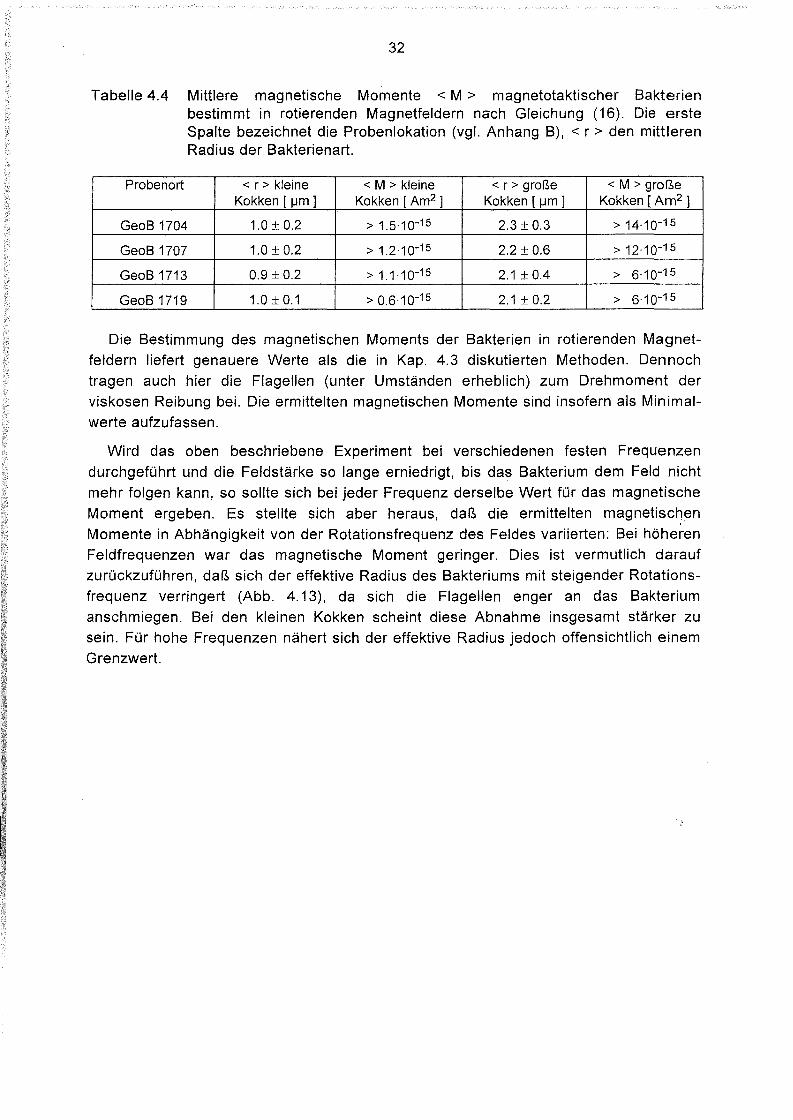
In Tabelle 4.4 sind entsprechende Meßwerte magnetischer Momente für zwei
morphologisch unterscheidbare Arten von Magnetokokken von vier Kernstationen im
östlichen Südatlantik angegeben; sie wurden nach Gleichung (16) bestimmt. Als Radius
wurde für jede Art aus Videoaufnahmen ein Mittelwert bestimmt. Die Stärke und Fre
quenz des Magnetfeldes wurde für jedes Bakterium individuell ermittelt, die Feldstärken
lagen zwischen 50 und 500 IJT, die Frequenzen bei 1,1 Hz. Kleine Kokken besitzen
magnetische Momente von um 10-15 Am2 , große Kokken dagegen von etwa 10-14 Am 2.
32
Tabelle 4.4 Mittlere magnetische Momente < M > magnetotaktischer Bakterienbestimmt in rotierenden Magnetfeldern nach Gleichung (16). Die ersteSpalte bezeichnet die Probenlokation (vgl. Anhang B), < r> den mittlerenRadius der Bakterienart.
Probenort < r> kleine < M > kleine < r> große < M > großeKokken [ IJm ] Kokken [ Am2 ] Kokken [ IJm] Kokken [ Am2 1
GeoB 1704 1.0 ± 0.2 > 1.5.10-15 2.3 ± 0.3 > 14'10-15
GeoB 1707 1.0 ± 0.2 > 1.2'10-15 2.2 ± 0.6 > 12.10-15
GeoB 1713 0.9 ± 0.2 > 1.1.10-15 2.1 ± 0.4 > 6.10-15
GeoB 1719 1.0±0.1 > 0.6,10-15 2.1 ± 0.2 > 6.10-15
Die Bestimmung des magnetischen Moments der Bakterien in rotierenden Magnetfeldern liefert genauere Werte als die in Kap. 4.3 diskutierten Methoden. Dennochtragen auch hier die Flagellen (unter Umständen erheblich) zum Drehmoment derviskosen Reibung bei. Die ermittelten magnetischen Momente sind insofern als Minimal
werte aufzufassen.
Wird das oben beschriebene Experiment bei verschiedenen festen Frequenzen
durchgeführt und die Feldstärke so lange erniedrigt, bis das Bakterium dem Feld nichtmehr folgen kann, so sollte sich bei jeder Frequenz derselbe Wert für das magnetischeMoment ergeben. Es stellte sich aber heraus, daß die ermittelten magnetisc~~n
Momente in Abhängigkeit von der Rotationsfrequenz des Feldes variierten: Bei höherenFeldfrequenzen war das magnetische Moment geringer. Dies ist vermutlich daraufzurückzuführen, daß sich der effektive Radius des Bakteriums mit steigender Rotationsfrequenz verringert (Abb. 4.13), da sich die Flagellen enger an das Bakterium
anschmiegen. Bei den kleinen Kokken scheint diese Abnahme insgesamt stärker zusein. Für hohe Frequenzen nähert sich der effektive Radius jedoch offensichtlich einemGrenzwert.
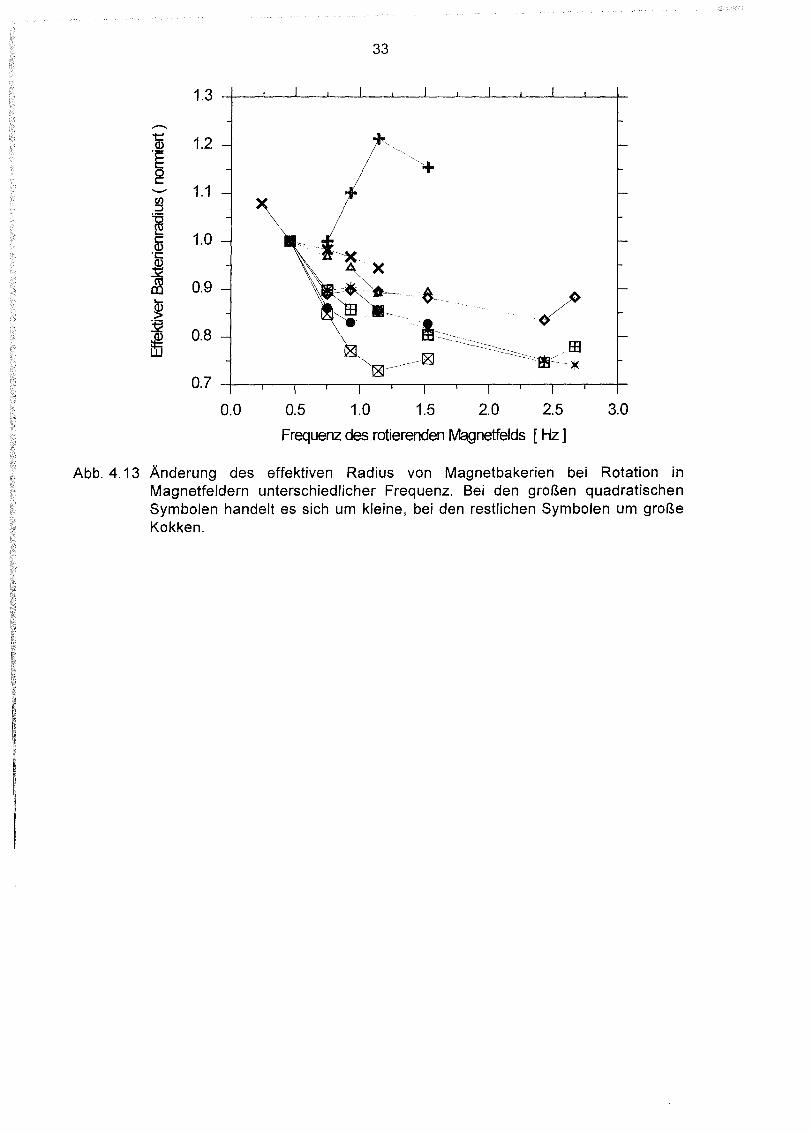
1.3
--1::: 1.2Q)
.~
~........ 1.1f/):::J
~1.0ID
'e:
~&i 0.9...g2~ 0.8~
0.7
0.0
33
0.5 1.0 1.5 2.0 2.5 3.0
Frequenz des rotierenden rv1agnetfelds [Hz]
Abb. 4.13 Änderung des effektiven Radius von Magnetbakerien bei Rotation inMagnetfeldern unterschiedlicher Frequenz. Bei den großen quadratischenSymbolen handelt es sich um kleine, bei den restlichen Symbolen um großeKokken.

34
5 Tiefenverteilung magnetotaktischer Bakterien und geochemischeZonierung in Wattsedimenten
Vorbereitende Arbeiten für die Expeditionen mit dem Forschungsschiff METEOR
fanden im Nordseewatt von Weddewarden bei Bremerhaven statt. Dabei wurde vorallem die Tiefenverteilung der Magnetbakterienpopulationen im Sediment untersucht.Das Arbeitsgebiet wurde gewählt, weil dort die Arbeitsgruppe Geochemie desFachbereichs Geowissenschaften der Universität Bremen bereits umfangreicheVorarbeiten durchgeführt hatte. So war das Gebiet geologisch kartiert (FISCHER, 1990;SAGEMANN, 1990), wobei die Schichtung der obersten zwei Meter spezifisch unterBerücksichtigung der Korngrößenverteilung beschrieben wurde. Auf einer etwa 16 m2
großen Fläche im Schlickwatt sind außerdem die jahreszeitlichen Variationen
geochemischer Parameter, insbesondere von Sauerstoff, Nitrat, Eisen, Sulfat,Schwefelwasserstoff und Schwermetallen, detailliert analysiert worden (SAGEMANN inVorb., SKOWRONEK in Vorb.). Die dabei gewonnenen Ergebnisse lassen sich mit denTiefenverteilungen magnetotaktischer Bakterien in Beziehung setzen.
Zudem wurden dort Magnetbakterien gesammelt, die
- für die Charakterisierung im Transmissionselektronenmikroskop (Kap. 2.2),
- für die Bewegungsanalyse im Uchtmikroskop (Kap. 4) und
- für Testzwecke bei der Beprobung im Südatlantik (Kap. 6.11)
Verwendung fanden.
5.1 Beschreibung des Arbeitsgebiets
Der Ort Weddewarden liegt auf der rechten Weserseite, an der nördlichen Stadtgrenzevon Bremerhaven, das untersuchte Wattgebiet bei 53,6°N und 8,5°E zwischen vierBuhnen (Steinwällen), die zur Regulierung des Flußlaufes angelegt wurden (Abb. 5.4links). Während der größte Teil des Untersuchungsgebiets aus Feinsandwatt besteht,befindet sich das Probengebiet von SAGEMANN und SKOWRONEK im feinkörnigerenSchlickwatt.
Der mittlere Tidenhub bei Weddewarden beträgt 3.4 m. Das untersuchte Areal stehtbei mittlerem Hochwasser etwa 0.5 - 2 m unter Wasser. Bei Niedrigwasser fällt dasSediment nicht vollständig trocken, in den Vertiefungen der Rippelmarken bleibt dasWasser zurück. Dort wurden die Proben genommen. Dabei ist darauf geachtet worden,daß nicht in der Nähe von Kotspuren des Wattwurms Arenico/a marina (REINECK, 1978)beprobt wurde. Verglichen mit anderen Arealen im Nordseewatt ist die Dichte dieserKotspuren hier gering.
Die Salinität des Gebiets variiert stark, nach LÜNEBURG et al. (1975) liegt es in dermesohalinen Zone (Salinität zwischen 5 und 18 %0). SAGEMANN und SKOWRONEKbestimmten in ihrem Probengebiet nahe der Fahrrinne der Weser je nachWasserführung Salzgehalte zwischen 5 und 22 %0. Die Temperatur schwankt ebenfalls

35
stark. Bei intensiver Sonneneinstrahlung kann sich das dunkle Watt an der Oberflächeauf über 30°C aufheizen. Die Erwärmung führt zu einer starken Zunahme derbenthischen Grabungsaktivität. Dadurch wird das Sediment wesentlich stärkerdurchwühlt als nach längeren Phasen kühlen Wetters. Die Beprobungen wurden nur
durchgeführt bei bedecktem Himmel, niedrigen Lufttemperaturen (von 2 bis 10°C) undnach einer längeren Periode schlechten Wetters. Die genaue Vorgehensweise bei derBeprobung wird in Kap. 5.3 bis 5.5 erläutert.
5.2 Morphologie magnetotaktischer Bakterien in Wattsedimenten
Es wurden vier verschiedene morphologische Grundformen magnetotaktischer
Bakterien identifiziert: Kokken, Spirillen, Vibrionen und Stäbchen (vgl. Kap. 2.1). Amhäufigsten finden sich kleine Kokken (Durchmesser bei 1 IJm), die auf Grund desbeschränkten Auflösungsvermögens im Lichtmikroskop keine innere Struktur erkennenlassen. Besonders auffällig waren große, schnell bewegliche Kokken, die mit dem vonMOENCH (1988) beschriebenen Bilophococcus magnetotacticus identisch sein könnten.Die in deutlich geringerer Zahl vertretenen kleinen Spirillen sind etwa 2 IJm lang und0.5 IJm breit. Weiter wurden einzelne Exemplare größerer Spirillen mit einem im
Lichtmikroskop auffälligen lichtbrechenden 'Querstreifen' sowie wenige kleinere undgrößere Vibrionen und Stäbchen beobachtet. Das früher untersuchte Magnetobacteriumbavaricum (PETERMANN et al., 1990) konnte in diesen Sedimenten nicht nachgewiesenwerden.
5.3 Laboruntersuchung der Tiefenverteilung magnetotaktischerBakterien in Sedimentkernen aus dem Watt von Weddewarden
Über die Bedingungen, unter denen magnetotaktische Bakterien leben, war zu Beginnder Dissertation wenig bekannt. Lediglich zwei Stämme dieser Bakterien konnten bisdahin kultiviert werden (BLAKEMORE et al., 1979; BAZYLINSKI et al., 1988), nur ein Stammwar ausführlicher beschrieben. Um Einzelheiten der Lebensbedingungen zu klären, istdie Tiefenverteilung der Magnetbakterien in der Sedimentsäule zunächst im Laboruntersucht worden.
Mit einer Kolbenpipette (Brand, Typ 'Transferpettor') wurde aus unterschiedlichenTiefen 50 IJI Sedimentsuspension entnommen (Abb. 5.1). Die Pipette wird von oben ins
Sediment geführt. Ein Stempel gewährleistet, daß nur Probenmaterial aus einerbestimmten Schicht in die Pipette eindringen kann. Die Sedimentsuspension wird aufeinen Objektträger gegeben und im Durchlichtmikroskop (siehe Kap. 3) untersucht. DieProbe erscheint zunächst opak. Durch das Porenwasser bildet sich aber um dasSediment ein durchsichtiger Tropfenrand. Nach Anlegen eines statischen, homogenenMagnetfeldes schwimmen die magnetotaktischen Bakterien in der vorgegebenenRichtung an eine Seite des Tropfenrandes und können dort beobachtet werden. Wennweniger als 100 Magnetbakterien am Rand erscheinen, lassen sich die schnell
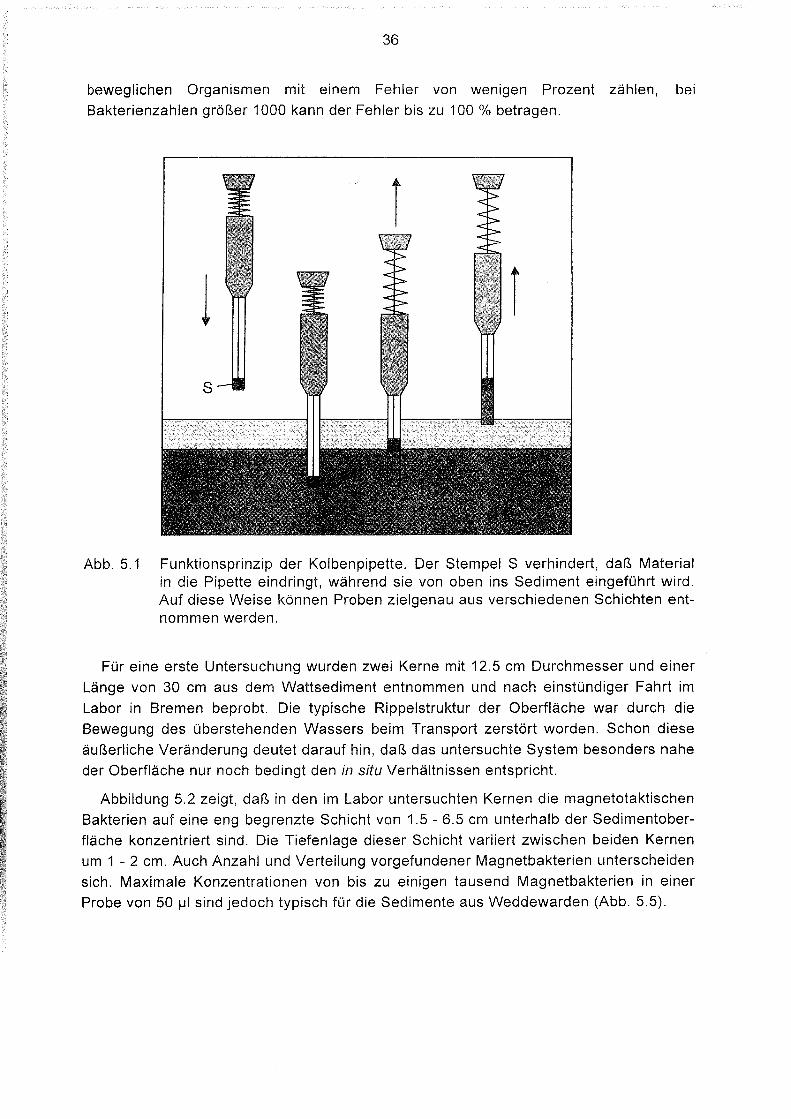
36
beweglichen Organismen mit einem Fehler von wenigen Prozent zählen, bei
Bakterienzahlen größer 1000 kann der Fehler bis zu 100 % betragen.
1s
Abb. 5.1 Funktionsprinzip der Kolbenpipette. Der Stempel S verhindert, daß Materialin die Pipette eindringt, während sie von oben ins Sediment eingeführt wird.Auf diese Weise können Proben zielgenau aus verschiedenen Schichten entnommen werden.
Für eine erste Untersuchung wurden zwei Kerne mit 12.5 cm Durchmesser und einer
Länge von 30 cm aus dem Wattsediment entnommen und nach einstündiger Fahrt im
Labor in Bremen beprobt. Die typische Rippelstruktur der Oberfläche war durch die
Bewegung des überstehenden Wassers beim Transport zerstört worden. Schon diese
äußerliche Veränderung deutet darauf hin, daß das untersuchte System besonders nahe
der Oberfläche nur noch bedingt den in situ Verhältnissen entspricht.
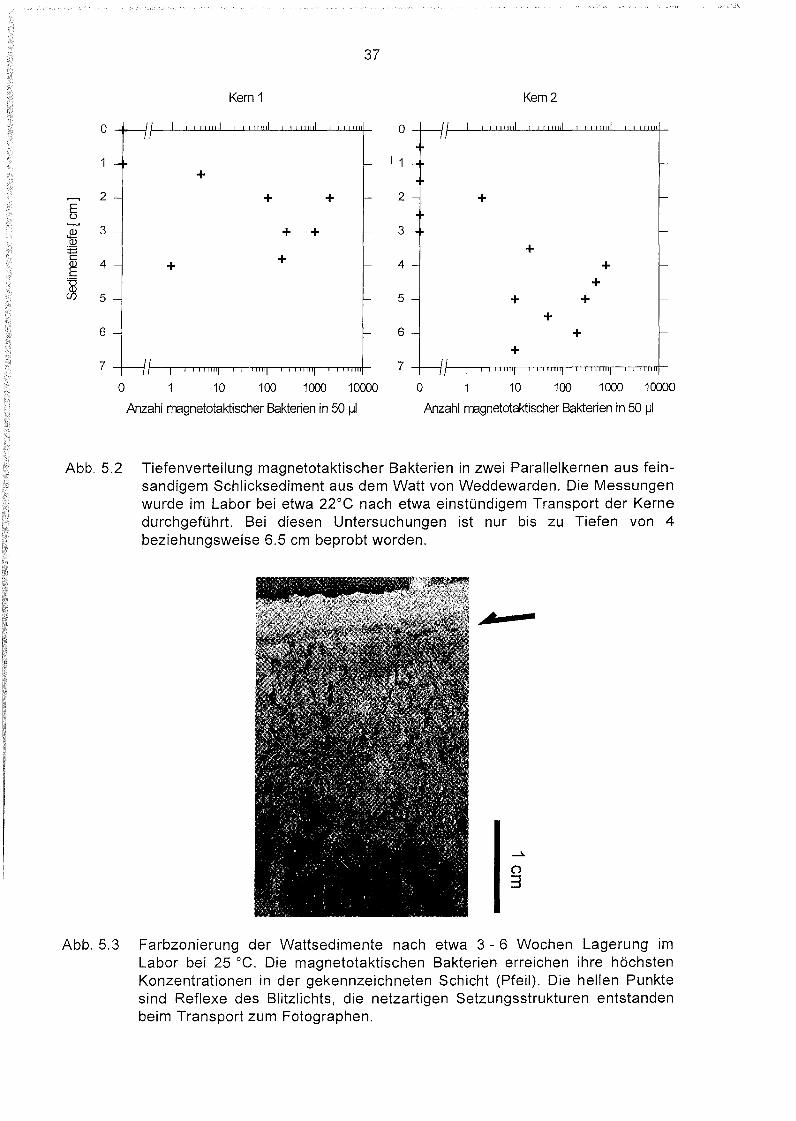
Abbildung 5.2 zeigt, daß in den im Labor untersuchten Kernen die magnetotaktischen
Bakterien auf eine eng begrenzte Schicht von 1.5 - 6.5 cm unterhalb der Sedimentoberfläche konzentriert sind. Die Tiefenlage dieser Schicht variiert zwischen beiden Kernen
um 1 - 2 cm. Auch Anzahl und Verteilung vorgefundener Magnetbakterien unterscheiden
sich. Maximale Konzentrationen von bis zu einigen tausend Magnetbakterien in einer
Probe von 50 IJI sind jedoch typisch für die Sedimente aus Weddewarden (Abb. 5.5).
37
++
+
++
+
Kem2
+
+
+
o 1 10 100 1000 10000
Anzahl magnetataktischer Bakterien in 50 IJI
Kern 1
0
I 1+
+ + 2
+ + 3
+ + 4
5
6
7
o 1 10 100 1000 10000
Anzahl magnetataktischer Bakterien in 50 IJI
2
o
3
4
5
7
6
Abb. 5.2 Tiefenverteilung magnetotaktischer Bakterien in zwei Parallelkernen aus feinsandigem Schlicksediment aus dem Watt von Weddewarden. Die Messungenwurde im Labor bei etwa 22°e nach etwa einstündigem Transport der Kernedurchgeführt. Bei diesen Untersuchungen ist nur bis zu Tiefen von 4beziehungsweise 6.5 cm beprobt worden.
Abb. 5.3 Farbzonierung der Wattsedimente nach etwa 3 - 6 Wochen Lagerung imLabor bei 25 oe. Die magnetotaktischen Bakterien erreichen ihre höchstenKonzentrationen in der gekennzeichneten Schicht (Pfeil). Die hellen Punktesind Reflexe des Blitzlichts, die netzartigen Setzungsstrukturen entstandenbeim Transport zum Fotographen.

38
Bei längerer Lagerung der Kerne im Labor stellte sich eine neue, engeregeochemische Zonierung im Sediment ein. Sie ist vermutlich eine Folge der erhöhtenTemperatur und der damit einhergehenden Erhöhung der biologischen Aktivität
(SAGEMANN, mündl. Mitteilung, 1994) und war deutlich erkennbar an einer Verengungder typischen Farbschichtung in den Wattsedimenten (Abb. 5.3). Die Magnetbakterien
befanden sich dann in einer wesentlich dünneren Schicht, die näher an der Oberflächelag. Die Tiefenverteilung der Bakterien wird offensichtlich durch die geochemischeZonierung im Sediment bestimmt.
5.3.1 Geochemische Zonierung der oberflächennahen Sedimentschichten
Die geochemische Tiefenzonierung oberflächennaher Sedimente spiegelt imwesentlichen Stoffwechselaktivitäten von Mikroorganismen wider. Ein Modell für diese
Zonierung wird bei FROELICH et al. (1979) beschrieben. Verschiedene heterotropheBakterien, das sind Bakterien, die organische Kohlenstoffverbindungen alsKohlenstoffquelle nutzen, konkurrieren um organische Verbindungen. Der oxidativeAbbau dieser Verbindungen liefert je nach Reaktionspartner (terminaler Elektronenakzeptor) unterschiedlich viel Energie. Vereinfacht läßt sich der Abbau organischenMaterials durch die nachfolgenden Reaktionsgleichungen beschreiben. Dabei ist dieÄnderung der Gibb'schen freien Enthalpie ~Go unter Standardbedingungen angegeben,die bei Abbau eines Mols der angegebenen chemischen Verbindung frei wird. Ihre
Zusammensetzung entspricht dem Redfield-Verhältnis (REDFIELD et al., 1963). VonSauerstoff (02) über Nitrat (N03-), Mangan (Mn02), Eisen (Fe20 3) und Sulfat (S042-)
nimmt die Energie ab, die aus den Reaktionen gewonnen werden kann.
(CH20)106 (NH3)16 (H3P04) + 138 02 ~ 106 CO2 + 16 HN03+ H3P04 + 122 H20
LlGo = 56370 kJ / Mol == 3190 kJ / Mol Glukose
(CH20)106 (NH3)16 (H3P04) + 94.4 HN03 ~ 106 CO2+ 55.2 N2+ H3P04 + 177.2 H20
LlGo = 53540 kJ / Mol == 3030 kJ / Mol Glukose
(CH20)106 (NH3)16 (H3P04) + 236 Mn02 ~ 106 CO2+ 236 Mn2++ 8 N2+ H3P04+ 366 H20
LlGo=51600 kJ / Mol == 2920 kJ / Mol Glukose
(CH20)106(NH3)16(H3P04)+212Fe203+848H+~ 424 Fe2++ 106C02+ 16 NH3+ H3P04+520H20
LlGo =24510 kJ / Mol == 1410 kJ / Mol Glukose
(CH20)106 (NH3)16 (H3P04) + 53 SOl- ~ 106 CO2+ 16 NH3+ 53 S2- + H3P04+ 106 H20
LlGo = 6710 kJ / Mol == 380 kJ / Mol Glukose
Bakterien, die aus der Zersetzung organischer Verbindungen mehr Energie gewinnenkönnen, sind in der Lage, Bakterienarten zu verdrängen, die einen Stoffwechselwegbenutzen, der weniger Energie liefert. Nitratreduzierende Bakterien können erst beigeringen Sauerstoffkonzentrationen dominierend werden, Sulfatreduzierer in nochgrößeren Tiefen. In groben Zügen läßt sich die geochemische Tiefenzonierung durch dieAbfolge der beschriebenen Abbauprozesse der verschiedenen physiologischenBakteriengruppen (die sich in ihrem Stoffwechsel unterscheiden) erklären. VieleBakterien sind allerdings fähig organische Verbindungen mit mehreren der oben

39
angeführten Elektronenakzeptoren abzubauen. Sie benutzen dann den in der jeweiligen
Sedimenttiefe vorhandenen optimalen Akzeptor.
5.3.2 Reaktion der magnetotaktischen Bakterien auf Veränderung dergeochemischen Zonierung
Während der Lagerung von Wattsediment im Labor bauen die Organismen weiterhin
organische Verbindungen ab und reduzieren dabei die oben beschriebenen terminalen
Elektronenakzeptoren 02, N03-, Mn02, Fe20 3 und SO/-. Bei den Redox-Reaktionen
wird Sauerstoff von den Elektronenakzeptoren abgespalten und auf die organischen
Verbindungen übertragen. Gleichzeitig dringt von oben Luftsauerstoff in das Sediment
ein und steht einerseits direkt zur Oxidation organischen Materials zur Verfügung,
andererseits werden die reduzierten Elektronenakzeptoren reoxidiert. Diese können
anschließend genutzt werden, um wiederum organische Substanz oxidativ abzubauen.
Im Labor ist die Sauerstoffzufuhr allerdings wesentlich geringer als im Watt, da kaum
turbulente Strömung auftritt. In den ersten Tagen nach Versuchsbeginn sind noch die
Bewegungsspuren kleiner Arthropoden (Corophium volutator) und Würmer erkennbar,
doch innerhalb weniger Tage geht deren bioturbate Aktivität zurück. Auch der hohe
Sauerstoffeintrag durch das Überströmen bei Flut entfällt im Labor. Die Sauerstoffzufuhr
erfolgt hier überwiegend diffusiv und erreicht nur Tiefen von einem bis wenigen
Millimetern. Dadurch wird die geochemische Zonierung insgesamt enger.
Die Magnetbakterien müssen auf diese neuen Umweltbedingungen durch Änderung
ihres Aufenthaltsortes oder ihres Stoffwechsels reagieren. Sie können in freiem Wasser
mit einer Geschwindigkeit von etwa 30 bis 200 IJm/s schwimmen (Abb. 4.3,4.4). In
ungestörten Sedimenten liegen diese Geschwindigkeiten bei niedrigeren Werten. Dabei
scheinen Magnetokokken nur schwer entgegen ihrer bevorzugten Orientierung zum
Magnetfeld schwimmen zu können. Spirillen können zwar leicht vorwärts und rückwärts
schwimmen, verfügen dafür über keinen Mechanismus, um Hindernisse effektiv zu
passieren. Wenn die Magnetbakterien im Sediment nicht in der Lage sind, der sich
verengenden geochemischen Zonierung entgegen ihrer bevorzugten Schwimmrichtung
zu folgen, so müssen sie zwangsläufig ihren Stoffwechsel umstellen und einen anderen
Elektronenakzeptor zur Atmung benutzen. Denkbar wäre eine Umstellung von N03- auf
Fe (111) (vgl. Abb. 5.6). Falls sie dazu nicht in der Lage sind oder falls dieser Abbauweg
zu wenig Energie liefert (BLAKEMORE, 1982; PAOLETTI & BLAKEMORE, 1986), sterben sie
ab, nachdem ihre Energiereserven aufgebraucht sind.
Dennoch steigt in vielen Proben nach etwa zwei Wochen die Zahl der
Magnetbakterien innerhalb der obersten 10 Millimeter des Sediments stark an.
Anreicherungskulturen mit ~1 05 magnetotaktischen Bakterien und mehr pro 50 IJI bilden
sich aus.

40
5.4 Beprobung der Wattsedimente in situ
Aufgrund der in Kapitel 5.3 dargelegten Veränderungen der chemischen Zonierung
durch Transport und Lagerung der Sedimente im Labor wurde zur Bestimmung
ungestörter Tiefenverteilungen von Magnetbakterien eine Beprobung in situ, also im
Watt durchgeführt.
Abb. 5.4 Beprobung der Wattsedimente in situ. Die Proben wurden mit der Pipettedirekt aus dem Sediment entnommen. Das Lichtmikroskop wurde überBatterie betrieben, die Spulen dienten dabei als Vorwiderstand (HeIligkeitsregelung) der Beleuchtung und gleichzeitig zur Erzeugung einesMagnetfeldes.
Die Beprobungsstelle im feinsandig-schlickigen Watt lag nur wenige Meter von der
Uferbefestigung entfernt (Abb. 5.4), so daß innerhalb kurzer Zeit eine große Anzahl an
Untersuchungen möglich war. Die Proben wurden mit der Pipette direkt aus dem Watt
entnommen, unmittelbar auf den Objektträger gegeben und im Mikroskop untersucht.
Bei der Beprobung betrug die Lufttemperatur etwa 5°C.
Für die Gesamtheit der Magnetbakterien zeigt die Tiefenverteilung (Abb. 5.5) eine
maximale Häufigkeit von bis zu mehreren tausend Exemplaren pro 50 1-11. In den
obersten drei Millimetern, der aeroben Schicht, wurden nur sehr wenige
Magnetbakterien beobachtet « 10 in 50 1-11). Die überwiegende Mehrzahl meidet
offensichtlich die aerobe Schicht. Die höchste Konzentration wird zwischen 0.5 und 2
3 cm erreicht. SAGEMANN und SKOWRONEK ermittelten in ihrer Versuchsfläche, 300 m
von der Uferbefestigung, im selben Monat Eindringtiefen von 0.5 cm für Sauerstoff und
ab einer Tiefe von 1.8 cm Fe (11) Konzentrationen> 10 1-1 mol/I im Porenwasser. Bei
Annahme einer ähnlichen geochemischen Zonierung in beiden Flächen lebt demnach
die Mehrzahl der Magnetbakterien in der Nitratreduktionszone oder der obersten Schicht
der Eisenreduktionszone.
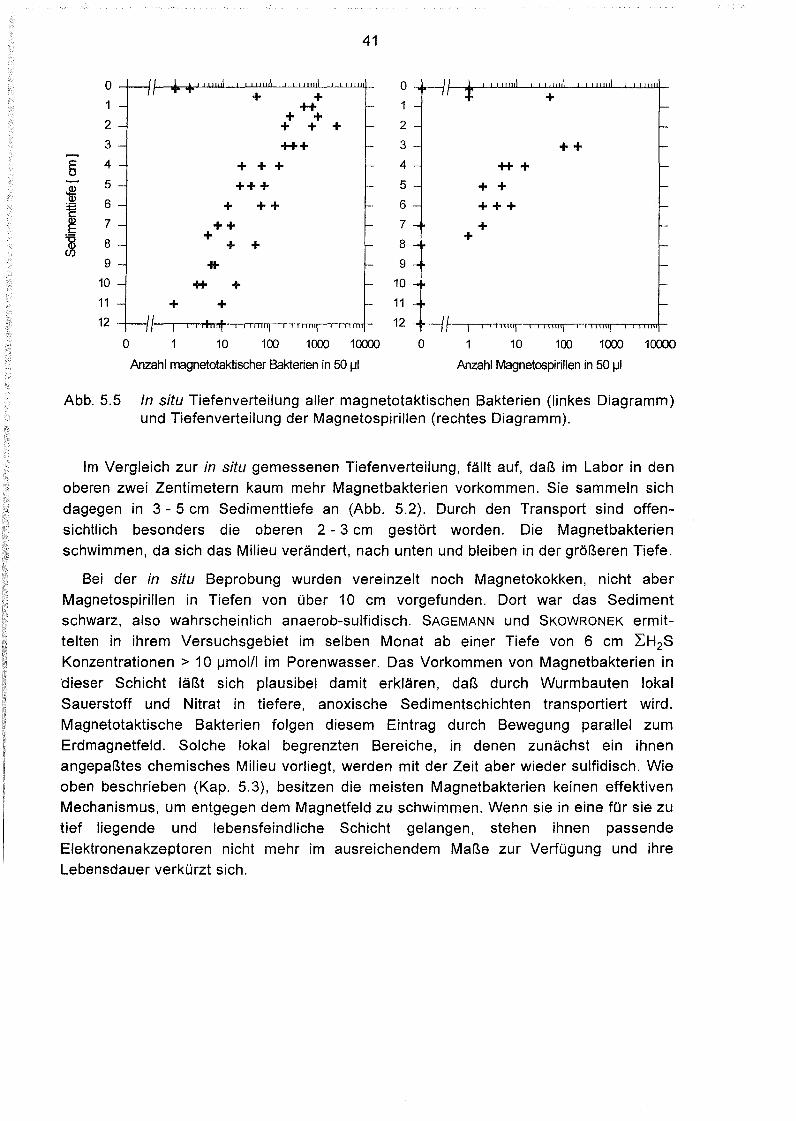
+++++
+ ++++
++
1 10 100 1000 10000
Anzahl Magnetospirillen in 50 IJI
41
0 0
1 + 1++2 + +
+ + + 2
3 +++ 3
6 4 + + + 4
'*5 +++ 5
~6 + ++ 6
~ 7 ++ 7
] 8 + 8+ +9 '* 9
10 -1+ + 10
11 + + 11
12 12
0 1 10 100 1000 10000 0
Anzahl magnetotaktischer Bakterien in 50 IJI
Abb. 5.5 In situ Tiefenverteilung aller magnetotaktischen Bakterien (linkes Diagramm)und Tiefenverteilung der Magnetospirillen (rechtes Diagramm).
Im Vergleich zur in situ gemessenen Tiefenverteilung, fällt auf, daß im Labor in denoberen zwei Zentimetern kaum mehr Magnetbakterien vorkommen. Sie sammeln sichdagegen in 3 - 5 cm Sedimenttiefe an (Abb. 5.2). Durch den Transport sind offensichtlich besonders die oberen 2 - 3 cm gestört worden. Die Magnetbakterienschwimmen, da sich das Milieu verändert, nach unten und bleiben in der größeren Tiefe.
Bei der in situ Beprobung wurden vereinzelt noch Magnetokokken, nicht aberMagnetospirillen in Tiefen von über 10 cm vorgefunden. Dort war das Sedimentschwarz, also wahrscheinlich anaerob-sulfidisch. SAGEMANN und SKOWRONEK ermittelten in ihrem Versuchsgebiet im selben Monat ab einer Tiefe von 6 cm LH2SKonzentrationen> 10 IJmol/1 im Porenwasser. Das Vorkommen von Magnetbakterien in'dieser Schicht läßt sich plausibel damit erklären, daß durch Wurmbauten lokalSauerstoff und Nitrat in tiefere, anoxische Sedimentschichten transportiert wird.Magnetotaktische Bakterien folgen diesem Eintrag durch Bewegung parallel zumErdmagnetfeld. Solche lokal begrenzten Bereiche, in denen zunächst ein ihnenangepaßtes chemisches Milieu vorliegt, werden mit der Zeit aber wieder sulfidisch. Wieoben beschrieben (Kap. 5.3), besitzen die meisten Magnetbakterien keinen effektivenMechanismus, um entgegen dem Magnetfeld zu schwimmen. Wenn sie in eine für sie- zutief liegende und lebensfeindliche Schicht gelangen, stehen ihnen passendeElektronenakzeptoren nicht mehr im ausreichendem Maße zur Verfügung und ihreLebensdauer verkürzt sich.

42
5.5 Bestimmung der geochemischen Zonierung und der Tiefenverteilungmagnetotaktischer Bakterien im gleichen Kern
Da Korngrößen sowie der Grad der Bioturbation und damit die geochemische Zonierung
in den Wattsedimenten lateral stark variieren, lassen sich Ergebnisse vonUntersuchungen an Sedimenten verschiedener Lokalitäten nur bedingt korrelieren. Umeinen direkten Vergleich der geochemischen Daten mit den Bakterienzahlen zu ermöglichen, wurde ihre Tiefenverteilung deshalb im selben Sedimentkern wie diegeochemischen Parameter bestimmt.
Messungen von Sauerstoffprofilen erfolgten in situ und im Labor, die Analysen von
Nitrat, Eisen, Sulfat und L:H 2S ausschließlich im Labor (SAGEMANN und SKOWRONEK,beide in Vorb.). Für letztere wurde das Sediment in Schichten von 0.25 - 2 cm Dickezerlegt, das Porenwasser der einzelnen Schichten ausgepreßt und anschließend
analysiert.
Die beobachtbaren Magnetbakterienzahlen gingen durch diese Segmentierungteilweise deutlich zurück. Vermutlich fiel das Sediment beim Zerschneiden teilweisetrocken, und die Magnetbakterien wurden so in ihrer Bewegungsfähigkeit beeinträchtigt,
daß sie sich anschließend im Lichtmikroskop nicht mehr identifizieren ließen. Um diesenEffekt zu minimieren, ist die Kolbenpipette vor Beprobung teilweise mit steril filtriertem
Wasser aus der Schicht knapp über der Sedimentoberfläche gefüllt worden. Fürsegmentiertes Material hat sich diese Methode bewährt, da das Sediment dadurchoptimal durchfeuchtet wird. Die ermittelten Magnetbakterienkonzentrationen sind jedochnur Mindestwerte. Obwohl die Proben sofort nach dem Segmentieren genommenwurden, kann nicht ausgeschlossen werden, daß ein Teil der Bakterien geschädigt
wurde.
Der Kern ist vor der Beprobung 4 Tage bei 4 oe im Labor gelagert worden. Dabei
wurde das überstehende Wasser täglich gewechselt und ständig umgerührt, um dennatürlichen Gegebenheiten möglichst nahe zu kommen. Die Mehrzahl der Magnet
bakterien in diesem Kern ist auf eine enge Schicht in einer Tiefe von 3 - 6cmkonzentriert. In diesem Horizont sind weder Sauerstoff noch Nitrat nachweisbar. InTiefen über 6 cm, in denen Sulfid vorhanden ist, geht die Magnetbakterienkonzentrationum Größenordnungen zurück. Einige wenige Magnetbakterien wurden jedoch noch inTiefen von 8 bis 11 cm in sulfidischem Milieu beobachtet. Auch um 1 cm Sedimenttiefesind Magnetbakterien vorhanden. Diese gehören offensichtlich einer anderenphysiologischen Gruppe an als die im Bereich erhöhter Konzentrationen gelöstenEisens. Entsprechend den unterschiedlichen geochemischen Milieus lassen sichvermutlich drei bis vier Gruppen von Magnetbakterien unterscheiden:
- mikroaerophile Nitratreduzierer,
- Nitratreduzierer, die im anaeroben Milieu leben,
- Bakterien in der Eisenreduktionszone und vermutlich
- sulfidtolerante Bakterien, die in der Sulfatreduktionszone überleben können.
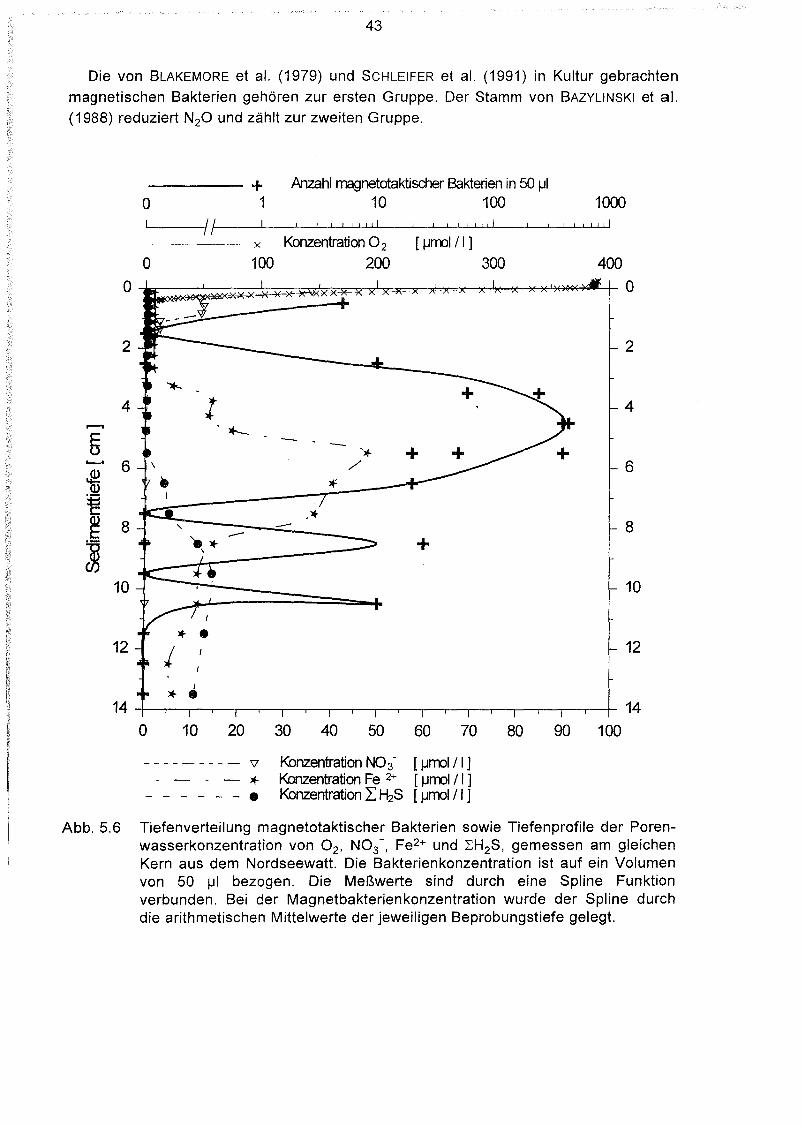
43
Die von BLAKEMORE et al. (1979) und SCHLEIFER et al. (1991) in Kultur gebrachtenmagnetischen Bakterien gehören zur ersten Gruppe. Der Stamm von BAZYLINSKI et al.(1988) reduziert N20 und zählt zur zweiten Gruppe.
o----- + Anzahl magnetotaktischer Bakterien in 50 IJI
1 10 100 1000
300[ IJrnol/ I]Konzentration°2
200100
L-_---j,/--,__..L--_"--...L.'.....J'L..L'LI!W'..LJ' 1'----1----J''--L'-L'-L'..L'.LI'[..LI_L..L----I-.....J'L..L'L''Ll'..LJ1I- 1/~~_--------- X
o 400O~-_L.---~~:-=~7v-=~=~~~~~**~~IL-j.-O
12
14100908070605040302010
* •
I* •
,o
12
2 2
'*-4 1 4
~ ' *-6 '*........ 6 /" 6~Q) *~ -*~ 8 - 8-~
+
10 10
--------- \l
- - - - *- - - - - - . Konzentration NO3- [ IJrnoI/l ]
Konzentration Fe 2+ [ IJrnoI/l ]Konzentration L lizS [lJrnoI/l]
Abb. 5.6 Tiefenverteilung magnetotaktischer Bakterien sowie Tiefenprofile der Porenwasserkonzentration von 02' N03-, Fe2+ und 2:H2S, gemessen am gleichenKern aus dem Nordseewatt. Die Bakterienkonzentration ist auf ein Volumenvon 50 IJI bezogen. Die Meßwerte sind durch eine Spline Funktionverbunden. Bei der Magnetbakterienkonzentration wurde der Spline durchdie arithmetischen Mittelwerte der jeweiligen Beprobungstiefe gelegt.

44
5.6 Diskussion
Magnetotaktische Bakterien kommen in den untersuchten Wattsedimenten des Weser
ästuars in großer Zahl vor. In den aeroben Schichten, 1 - 3 mm unterhalb der Sediment
oberfläche, konnten nur einzelne Magnetbakterien nachgewiesen werden. Die Mehrzahl
der Magnetbakterien lebt im anaeroben Milieu. In der Literatur wird jedoch meist davon
ausgegangen, daß der größte Teil der Magnetbakterien mikroaerophil ist. Nach den
geochemischen Analysen und Beprobungen auf Magnetbakterien am gleichen Kern
lassen sich im Watt drei bis vier Typen von Magnetbakterien unterscheiden.
Eine geringere Anzahl von Magnetbakterien lebt in der mikroaeroben Schicht, die
Mehrzahl im anaeroben Bereich und zwar in der Nitratreduktionszone oder im obersten
Bereich der Eisenreduktionszone, also der Schicht, in der zum ersten Male größere
Mengen gelösten Eisens im Porenwasser nachweisbar sind. In der Sulfatreduktionszone
kommen nur wenige Magnetbakterien vor. .
Das Ergebnis, daß so große Magnetbakterienkonzentrationen in der obersten Schicht
mit gelöstem Eisen im Porenwasser beobachtet werden (Abb. 5.6), läßt sich auf
unterschiedliche Weise deuten:
- Von dieser in den Wattsedimenten vorkommenden, zahlenmäßig starken physio
logischen Gruppe magnetotaktischer Bakterien ist bis 1993 noch keines in Kultur
gebracht worden. In der Literatur wurden bisher überwiegend Kulturen der
nitratreduzierenden Magnetbakterien behandelt. Arbeiten, die Vorkommen von
Magnetbakterien in natürlichen Sedimenten beschreiben, befaßten sich noch nicht
mit deren physiologischer Einteilung (SPARKS et al., 1986; MANN et al., 1990).
- Alternativ sind die in der Eisenreduktionsschicht beobachteten Magnetbakterien doch
primär Nitratreduzierer, können ihren Stoffwechsel aber umstellen und Eisen
reduzieren, um damit organischen Kohlenstoff zu oxidieren. SHORT & BLAKEMORE
(1986) beschreiben diesen Mechanismus für das mikroaerophile Magnetbakterium
Aquaspirillum magnetotacticum. Von den Autoren wird aber betont, der
Energiegewinn falle dabei so gering aus, daß das Bakterium kaum seinen
Stoffwechsel davon bestreiten könne. Nach GUERIN & BLAKEMORE (1992) ist für A.
magnetotacticum das Zellwachstum bei Abwesenheit anderer terminaler
Elektronenakzeptoren von der Eisenkonzentration abhängig. Aquaspirillum magnetotacticum vermag also offensichtlich den Stoffwechsel auf Eisenreduktion umzustellen.
Die plausiblere Erklärung dürfte sein, daß die Mehrzahl der magnetotaktischen Bak
terien neben der Nitratreduktion zur Eisenreduktion fähig ist.
Neue Veröffentlichungen dokumentieren, daß weitere Magnetbakterienstämme in
02 - H2S Gradienten kultiviert werden konnten (MELDRUM et al., 1993a, b). Es liegen
aber noch keine genauen physiologischen Daten vor. Der von MELDRUM et al. (1993a)
beschriebene Magnetococcus MC-1 und das Magnetovibrio MV-2 (MELDRUM et al.,
1993b) können in Gegenwart von H2S wachsen, während das magnetotaktische Vibrio

45
MV-4 (MELDRUM et al., 1993b) kein H2S toleriert. Diese neu in Kultur gebrachtenBakterien sollten zur großen Gruppe der Magnetbakterien gehören, die in der Eisenreduktionszone vorkommen. Nach den zitierten Autoren ist MC-1 chemoheterotroph,MV-2 dagegen chemoautolithotroph. MV-4 kann zwischen beiden Stoffwechselwegenwählen.
Chemoheterotrophie, also Energiegewinn aus biochemischer Umsetzung organischerVerbindungen, ist für Bakterien in Milieus mit steilen geochemischen Konzentrationsgradienten und damit einhergehenden hohen Stoffflüssen sehr effektiv. Ob Eisentatsächlich die Rolle des Elektronenakzeptors spielt, ist derzeit noch ungeklärt. In derEisenreduktionszone liegt Eisen schon in gelöster Form vor und sollte sich daher vonMagnetbakterien leicht in die Zelle aufnehmen lassen, so daß die Bildung derMagnetosome unter geringerem Energieaufwand erfolgen kann.
Überraschend und neu bei Magnetbakterien ist der chemoautolithotropheStoffwechselweg von MV-2 und MV-4. Chemoautolithotrophie bedeutet, daß zumEnergiegewinn anorganische Verbindungen oxidiert und CO2 als Kohlenstoffquel/egenutzt wird. Welche Bedeutung diese Art des Stoffwechsels für die magnetotaktischenBakterien besitzt, ist offen. Für chemoautolithotrophe Bakterien sind steile Konzentrationsgradienten lebensnotwendig, weil nur dann anorganische Verbindungen, die sichauf unterschiedlichen Redox-Potentialen befinden, räumlich so nahe beieinander liegen,daß sie durch Diffusion den Bakterien in ausreichendem Umfang zur Verfügung stehen.Die chemische Energie, die in der Potentialdifferenz steckt, wird von den Bakterien
genutzt.

46
6 Lebende magnetotaktische Bakterien in Sedimenten des Südatlantiks
6.1 Einführung
Schon bald nach der Entdeckung magnetotaktischer Bakterien (BLAKEMORE, 1975)
wurde erkannt, daß sie zur Sedimentmagnetisierung beitragen können (KIRSCHVINK &
LOWENSTAM, 1979). Den ersten Nachweis von fossilem biogenem Magnetit in der
Tiefsee erbrachten PETERSEN et al. (1986) an Sedimenten von vier DSDP-Bohrungen
aus dem Angola Becken. Der fossile biogene Magnetit wurde elektronenmikroskopischdurch Vergleich mit den Magnetosomen lebender Magnetbakterien identifiziert. STOLZ
et al. (1986) wiesen lebende Magnetbakterien in hemipelagischen Sedimenten desSanta Barbara Beckens vor Kalifornien aus Tiefen bis zu 600 m nach. Sowohl
PETERSEN et al. als auch STOLZ et al. vermuteten, daß biogener Magnetit wesentlichzur Sedimentmagnetisierung beiträgt.
Zu Beginn dieser Arbeit war offen, ob Magnetbakterien in Tiefseesedimenten vor
kommen oder ob der von PETERSEN et al. nachgewiesene biogene Magnetit aus
anderen Quellen eingetragen wurde. Auch die Größe des Beitrags biogenen Magnetits
zur Sedimentmagnetisierung war in Einzelheiten unbekannt.
6.2 Behandlung der Proben
Im Rahmen dieser Arbeit konnten während vier Fahrten mit dem Forschungsschiff
METEOR Proben aus den obersten Schichten des Sediments genommen und auf
lebende Magnetbakterien hin untersucht werden (Anhang B: Abb. 1 - 4).
Die Sedimente wurden mit folgenden drei Geräten beprobt:
- Multicorer,
- Großkastengreifer und
- Schwerelot.
MulticorerBei diesem Probenahmegerät bleibt die Sedimentoberfläche exzellent erhalten, das
Sediment gelangt samt dem in situ überstehenden Bodenwasser an Bord. Dadurch wird
das Milieu, in dem die Bakterien leben, nur minimal verändert. Bei 47 von 86 Stationenstanden Multicorer Rohre zur Verfügung, um Tiefenprofile der Magnetbakterienver
teilung zu bestimmen (Anhang B: Tabelle 1).
GroßkastengreiferAn 59 von 86 Stationen wurde der Großkastengreifer eingesetzt. Daraus konnten dreiverschiedene Proben gewonnen werden: eine Oberflächenprobe aus einer Fläche von
25 cm2 und einer Tiefe bis etwa 0.5 cm, eine Sammelprobe von etwa 300 cm3

47
Sediment aus den tieferen Schichten (0.5 bis ~ 40 cm), die mit Oberflächenwasserüberschichtet wurden, und Spritzenproben (Volumen 10 ml) in Tiefenabständen von5 cm.
SchwerelatMulticorer und Kastengreifer dringen maximal 0.5 m ins Sediment ein. Einzelne Probenaus Sedimenttiefen bis 10m wurden aus den Schwerelotkernen entnommen, an denendie Arbeitsgruppe Geochemie ihre Untersuchungen durchführte. Diese Kerne wurdensofort nach der Segmentierung in Stücke von einem Meter Länge bei 4 oe gekühlt.
Kühlung der ProbenIm Laufe der verschiedenen Fahrten stellte sich heraus, daß die Proben vor und auchwährend der mikroskopischen Untersuchung möglichst auf in situ Temperatur gehaltenwerden sollten. Auf der ersten Expedition M 16/2 war noch keine Kühlung vorgesehen,während der zweiten Reise M 20/2 wurden die Proben behelfsmäßig gekühlt. Auf denFahrten M 23/1 und M 23/2 sind die Proben dann bei niedrigen Temperaturen gelagert
und untersucht worden. Die Multicorer Rohre wurden in einem Standzylinder inEiswasser aufbewahrt. Die Temperatur im Sediment betrug dabei 2 bis maximal 10 oe.Der Objekttisch des Mikroskops wurde mit einer Wasser-Umlaufkühlung auf niedrigenTemperaturen gehalten. Dadurch blieb die Temperatur des untersuchten Sedimenttropfens unter etwa 10 oe. Anschließend an die Untersuchung wurden die Proben imKühlraum bei 4 oe gelagert und im Kühlcontainer nach Bremen transportiert.
6.3 Bestimmung der Anzahl magnetotaktischer Bakterien im Sediment
Der Nachweis der Magnetbakterien erfolgt im Lichtmikroskop durch ihre Bewegung indefinierten äußeren Magnetfeldern (Kap. 4.1).
Mit einer Kolbenpipette vom Typ 'Transferpettor' (Brand) wird 50 1-11 Sedimentsuspension auf einen Glasobjektträger gegeben. Die Suspension ist anfangs undurchsichtig,nach kurzer Zeit bildet aber das Porenwasser einen Rand um das Sediment, der mit
dem Durchlichtmikroskop (Kap. 3.) gut beobachtbar ist. Die Magnetbakterien bewegensich parallel zu einem angelegten Magnetfeld aus dem optisch opaken Sediment an denTropfenrand. Dort werden sie gezählt. Bei einer für Magnetbakterien eher geringenGeschwindigkeit von 30 I-Im/s (vgl. Abb. 4.3 & 4.4) dauert es etwa 3 Minuten, bis allebeweglichen Bakterien den Rand erreicht haben.
Je nach Korngröße und Zusammensetzung des Sediments lassen sich Magnetbakterien, die an den Rand des Tropfens gelangen, mehr oder weniger leicht erkennen.Die Beobachtung in grobkörnigem Sediment ist sehr einfach. In nannofossilreichenSedimenten ist die Identifikation schwieriger, da sich die kleinen Partikel auch sehr naheam Tropfenrand ablagern. Außerdem scheint es nur wenig freien Porenraum zu geben,der den Bakterien ein Durchkommen ermöglicht. Versuche einer Beobachtung

48
in der Tropfenmitte führten nicht zu besseren Ergebnissen. Roll- und Stampfbewegungen des Schiffes mobilisieren die Sedimentpartikel immer wieder, so daß sich(Magnet-) Bakterien dort nur schwer identifizieren lassen.
Damit sich die maximal mögliche Anzahl von Magnetbakterien an den Tropfenrandbewegt, wurde zunächst ein rotierendes Magnetfeld angelegt. Durch passive Rotation insolchen Magnetfeldern sollen Magnetbakterien von Sedimentpartikeln, an denen siehaften, gelöst werden. Anschließend wurde ein homogenes Gleichfeld von etwadoppelter lokaler Erdfeldstärke eingestellt. Die Vertikalkomponente dieses Feldes zeigte
meist nach unten, so daß sich die Magnetbakterien knapp oberhalb des Objektträgersbewegten und damit in der Fokusebene des Mikroskops lagen. Falls es sich um sehr
feinkörniges, nannofossilreiches Sediment handelte, wurde die Vertikal komponentezunächst mit geringer Amplitude nach oben eingestellt, so daß die Magnetbakterienungehindert im freien Wasser oberhalb der sedimentierten Partikel zum Tropfenrandgelangen konnten. Nach einiger Zeit wurde die Vertikalkomponente wieder nach unteneingestellt, um alle Magnetbakterien in die Fokusebene zu bringen.
Die Magnetfeldrichtung wurde anschließend mit dem Joystick (Kap. 3.) vorgegeben.
Manuell läßt sich der Zeitpunkt der Änderung der Feldrichtung am besten auf die
Reaktion der Bakterien abstimmen. So lassen sich auch sehr langsam beweglicheMagnetbakterien oder solche, die nur eine kurze Strecke zurücklegen und dann fürlängere Zeit unbeweglich bleiben, am sichersten identifizieren.
Die Beobachtungen wurden sowohl mit Phasenkontrast als auch im Hellfeld durchgeführt. Im Phasenkontrast erscheint das Objekt kontrastreicher und größer, aberschlechter aufgelöst als im Hellfeld. Zur Identifikation hat sich aber, vor allem innannofossilreichen Sedimenten, das Hellfeldbild besser bewährt.
Im Laufe der Untersuchungen wurde die Dauer, die zur Durchmusterung einesObjektträgers verwendet wurde, auf knapp eine Stunde verlängert, da sich während derFahrt M 23/1 zeigte, daß einige Magnetbakterien erst nach dieser langen Zeit amTropfenrand erscheinen. Die geringe Größe mancher Magnetbakterien und die langeDauer der Beobachtung sowie das Nachlassen der Konzentration des Beobachtersbedingen, daß Magnetbakterien übersehen werden können, die Zählung insofern alsoetwas zu niedrige Werte liefert.
Da die Untersuchungen ohne Deckglas stattfanden, Luftsauerstoff also ungehindert inden untersuchten Tropfen eindringen kann, können obligat anaerobe Bakterien nurschlecht beobachtet werden. Sie verlassen das Sediment nicht und können nach
Kontakt mit Luftsauerstoff irreparable Schäden davontragen. Aerobe und fakultativanaerobe Bakterien können dagegen mit der verwendeten Beobachtungstechnik gut
erfaßt werden.
Unbewegliche Bakterien, die sich nicht entlang der Magnetfeldlinien bewegen, lassensich mit der gewählten experimentellen Methodik nicht identifizieren. Während derExpedition M 20/2 sind Versuche durchgeführt worden, eventuell vorhandeneunbewegliche Magnetbakterien durch ein starkes magnetisches Gradientenfeld passivzu bewegen. Dazu wurde ein Objekttisch angefertigt, in dem zwei starkePermanentmagnete antiparallel angebracht sind. Die Magnetbakterien sollten mit etwa

49
1 IJm/s angezogen werden und daher leicht zu beobachten sein. Es gelang jedoch nicht,ein Magnetbakterium auf diese Weise zu nachzuweisen. Falls sie dennoch vorhandensind, haften sie wahrscheinlich an Sedimentpartikeln. Diese sind entweder zu schwer,um im Gradientenfeld bewegt zu werden, oder andere Sedimentpartikel blockieren ihreBewegung. Der Versuch die Bakterien anzufärben, wurde wieder verworfen, da sich dieFarbstoffe immer auch an die feinen Sedimentpartikel anlagern und sich daher dieBakterien nicht besser vom Sediment abheben.
6.4 Geographische Verteilung der Magnetbakterienkonzentration imSüdatlantik
Abbildung 6.1 zeigt eine vereinfachte geographische Verteilung der Magnetbakterien
konzentrationen in den oberflächennahen Sedimenten des Südatlantik. Die Darstellungfaßt die Ergebnisse von vier Expeditionen mit dem Forschungsschiff METEOR
zusammen. Eine Tabelle der beprobten Stationen mit geographischer Position,Wassertiefe, Beprobungsgerät und maximaler Magnetbakterienzahl findet sich inAnhang B, ebenso auch vier Karten mit den Stationsnummern. Die Stationsnumern sindvierstellig, die ersten beiden Ziffern bezeichnen die Fahrt, die letzten beiden Ziffern dieStation. Die Probennummern der Fahrt M 16/2 beginnen mit den Ziffern 15.. , für M 20/2mit 17.. , für M 23/1 mit 20.. und mit 21.. für M 23/2.
In Abbildung 6.1 und den folgenden Grafiken (Abb. 6.2 - 6.15) ist für jede beprobteStation jeweils die maximale Anzahl magnetotaktischer Bakterien angegeben, die ineinem durch die verwendete Pipette vorgegebenen Volumen von 50 IJI bestimmt wurde.Jeder Kern wird daher durch den Maximalwert der Magnetbakterienkonzentration imTiefenprofil repräsentiert. Dabei ist zu beachten, daß die Profile an den einzelnenStationen, je nach Anzahl der zu bearbeitenden Kerne, nicht in gleich dichten Tiefenabständen beprobt werden konnten.
Die höchsten Konzentrationen (bis zu 30 000/50 IJI) magnetotaktischer Bakterienwerden an den Kontinentalrändern insbesondere im Auftriebsgebiet vor Namibia undSüdafrika angetroffen. Am Kontinentalrand südöstlich vor Brasilien waren deutlichweniger «100/50 1-11), vor der Nordostküste Brasiliens auf der ersten Fahrt M 16/2 keine
Magnetbakterien nachzuweisen. An den Stationen im zentralen Atlantik wurden nureinzelne «10/50 1-11) aktiv schwimmende Magnetbakterien gefunden. Die einzigeLokation vor der Südostküste Brasiliens, bei der mehr als 100 Magnetbakterien in 50 1-11
Sedimentsuspension nachgewiesen werden konnten, ist die Station GeoB 2127 aufeinem Seamount der Vit6ria-Trindade-Kette. Dort, aber auch auf einem zweitenbeprobten Seamount (Station GeoB 1726 auf dem Walfisch Rücken) wurden imVergleich zu den umliegenden Stationen erhöhte Konzentrationen an Magnetbakterienfestgestellt (20/50 IJI). Es ist zu vermuten, daß bedingt durch lokalen Auftrieb anSeamounts erhöhte Nährstoffkonzentrationen vorhanden sind und die Magnetbakterienzahl dadurch zunimmt. Technische Schwierigkeiten bei der Kernnahme aufSeamounts bedingten, daß intakte Proben nur in den beiden genannten Fällengewonnen werden konnten.
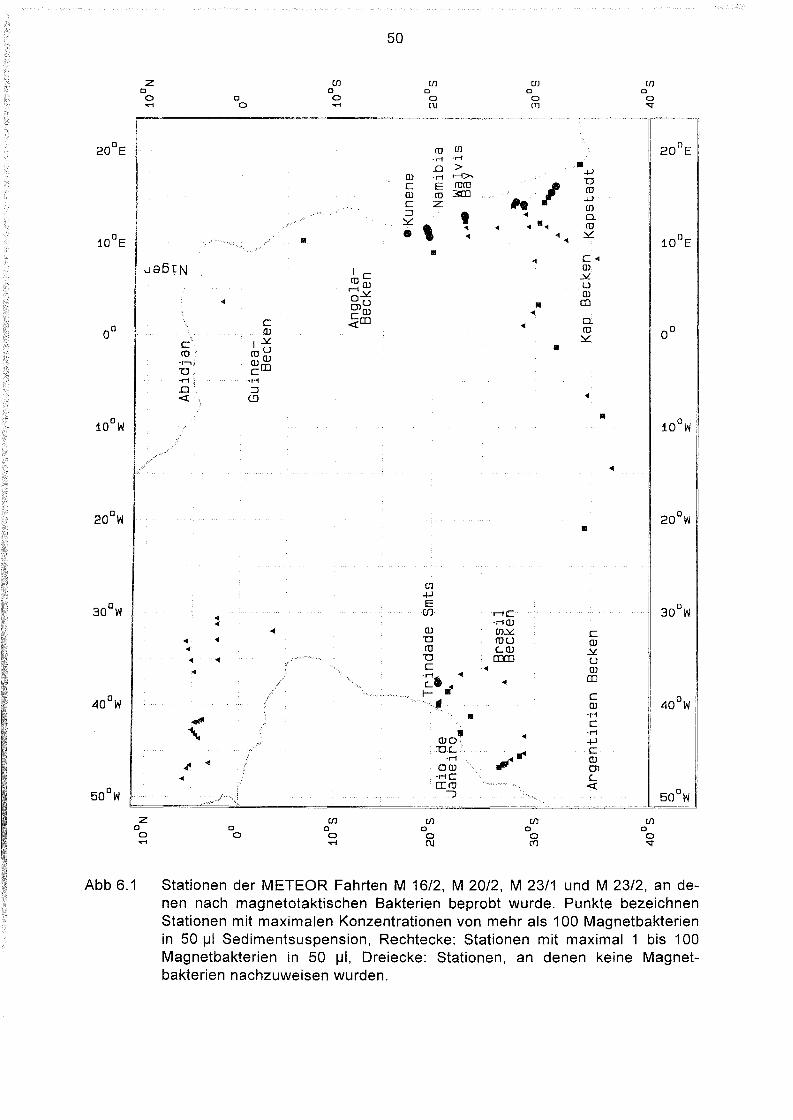
50
z UJ UJ UJ UJ0 0 0 0 0
0 0 0 0 0 0.... 0 .... ru (Tl "<t
200
E m U) 20 0 E·rl ·rl
.0 > •0) ·rl rl»-, .j..J
C E mm , u0) m 3lIl m
.j..JC Z -. • U)::J , ... 0..~ ....... m, ... ...
100
E • ... ...... ~100 E• ..
... c ...Ja5~N 'c
0)
.YmO) Url..:-.c 0)
... Du • mD10) ...
CC m 0..<I: ...
0° 0) m 0°,.Y ~
•CO m U0)0)
TI c m·rl ·rl.0 ::J<I: t!J ...
10°1'1•
100 W
...
20°1'1 200
W•
30°1'1 ......... ......:... ......
40°1'14'l'
'\
~...
...
zoo....
oo
UJoo....
Ul.j..J
ECf)
0)
TIcuUC
....
•0)0
·Ul:..···rl
00)·rlCrrcu
J
UJooru
11
r-iC'riO)(J)~
COU1:..0)
rD:Il
UJoo(Tl
C0)
.YU0)
mC0)·rl
C·rl.j..J
C0)
D1I:..<I:
UJoo
""Abb 6.1 Stationen der METEOR Fahrten M 16/2, M 20/2, M 23/1 und M 23/2, an de
nen nach magnetotaktischen Bakterien beprobt wurde. Punkte bezeichnenStationen mit maximalen Konzentrationen von mehr als 100 Magnetbakterienin 50 IJI Sedimentsuspension, Rechtecke: Stationen mit maximal 1 bis 100Magnetbakterien in 50 IJI, Dreiecke: Stationen, an denen keine Magnetbakterien nachzuweisen wurden.
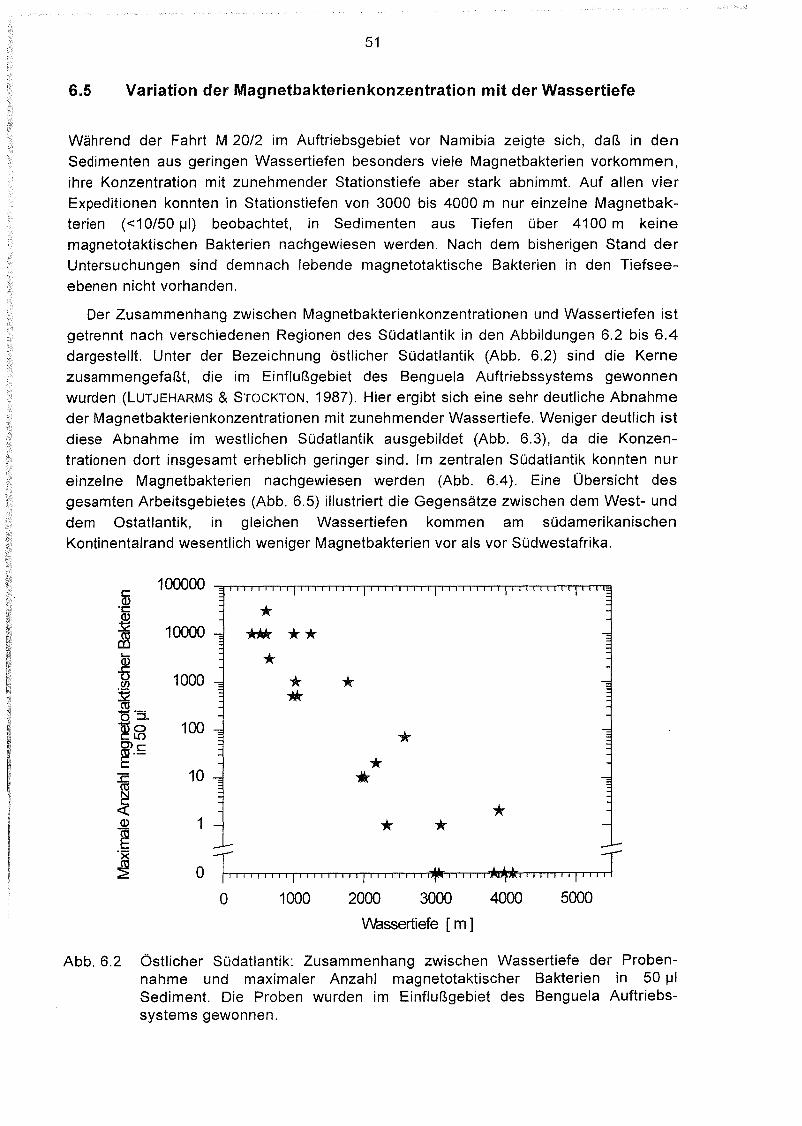
51
6.5 Variation der Magnetbakterienkonzentration mit der Wassertiefe
Während der Fahrt M 20/2 im Auftriebsgebiet vor Namibia zeigte sich, daß in den
Sedimenten aus geringen Wassertiefen besonders viele Magnetbakterien vorkommen,
ihre Konzentration mit zunehmender Stationstiefe aber stark abnimmt. Auf allen vier
Expeditionen konnten in Stationstiefen von 3000 bis 4000 m nur einzelne Magnetbak
terien «10/50 1-11) beobachtet, in Sedimenten aus Tiefen über 4100 m keinemagnetataktischen Bakterien nachgewiesen werden. Nach dem bisherigen Stand der
Untersuchungen sind demnach lebende magnetotaktische Bakterien in den Tiefsee
ebenen nicht vorhanden.
Der Zusammenhang zwischen Magnetbakterienkonzentrationen und Wassertiefen ist
getrennt nach verschiedenen Regionen des Südatlantik in den Abbildungen 6.2 bis 6.4
dargestellt. Unter der Bezeichnung östlicher Südatlantik (Abb. 6.2) sind die Kerne
zusammengefaßt, die im Einflußgebiet des Benguela Auftriebssystems gewonnen
wurden (LUTJEHARMS & STOCKTON, 1987). Hier ergibt sich eine sehr deutliche Abnahme
der Magnetbakterienkonzentrationen mit zunehmender Wassertiefe. Weniger deutlich ist
diese Abnahme im westlichen Südatlantik ausgebildet (Abb. 6.3), da die Konzen
trationen dort insgesamt erheblich geringer sind. Im zentralen Südatlantik konnten nur
einzelne Magnetbakterien nachgewiesen werden (Abb. 6.4). Eine Übersicht des
gesamten Arbeitsgebietes (Abb. 6.5) illustriert die Gegensätze zwischen dem West- und
dem Ostatlantik, in gleichen Wassertiefen kommen am südamerikanischen
Kontinentalrand wesentlich weniger Magnetbakterien vor als vor Südwestafrika.
100000CQ)'C
~ 10000~....
~ 1000~ctlö"5..~~ 100C>c~.-
::c 10
~Q) 1~
~ 0
**** ***
* *1k
**..
** *-r: I I I I I I f f I I I I I I I I I I I I I I I I f I I I~ I I I I I I Ja~", I I I I f f f I I I ,ro 1000 2000 3000 4000
VVassertiefe [m]
5000
Abb. 6.2 Östlicher Südatlantik: Zusammenhang zwischen Wassertiefe der Probennahme und maximaler Anzahl magnetotaktischer Bakterien in 50 1-1 1
Sediment. Die Proben wurden im Einflußgebiet des Benguela Auftriebssystems gewonnen.
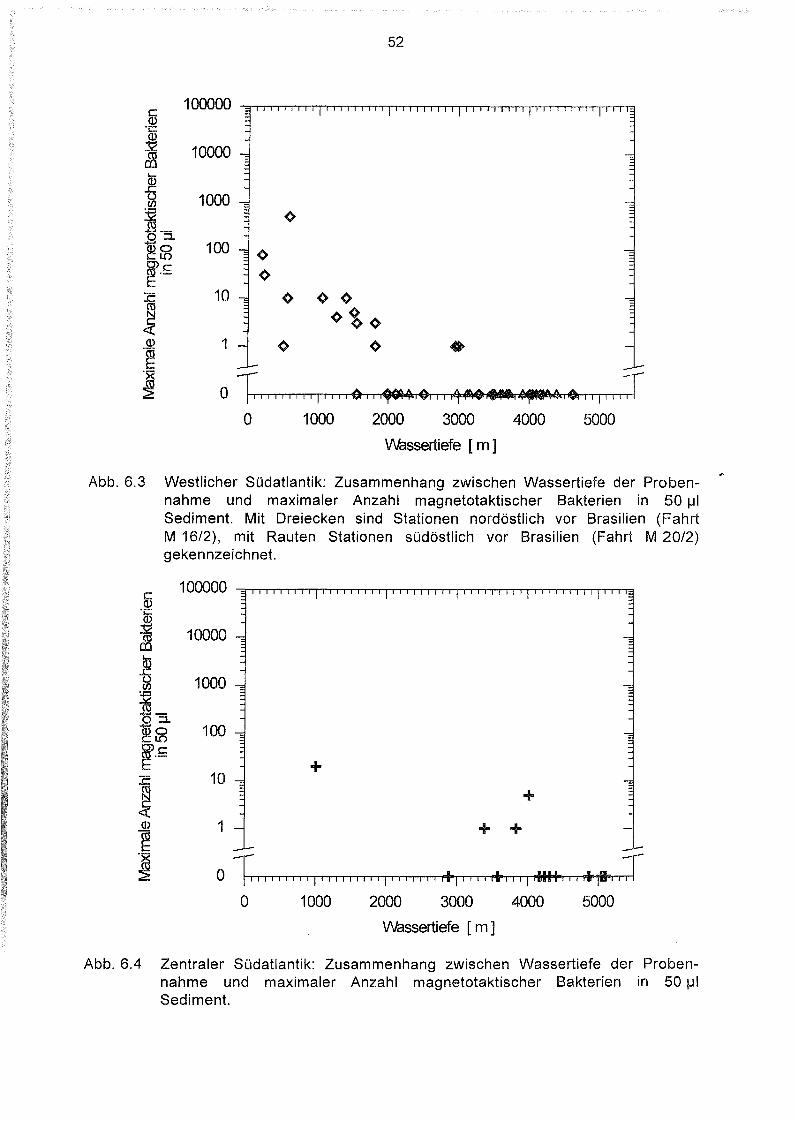
52
c: 100000a.>'C
~ 10000&i.....a.>'5 1000(J)
~o:::J.~f6 100O)c:~.-
:E 10
~a.> 1~
~ 0 -r:, ,,,,,,,1' , , , () , , ,~(',~, , , ,'t 901. :1",~ 1,~ , , I ' ,,ro 1000 2000 3000 4000
Wdssertiefe [m]
5000
Abb. 6.3 Westlicher Südatlantik: Zusammenhang zwischen Wassertiefe der Probennahme und maximaler Anzahl magnetotaktischer Bakterien in 50 ~I
Sediment. Mit Dreiecken sind Stationen nordöstlich vor Brasilien (FahrtM 16/2), mit Rauten Stationen südöstlich vor Brasilien (Fahrt M 20/2)gekennzeichnet.
+ +1
10
100
o "l:"""" I' , I , , I I , , I' , I , I , , r+I' , , , .+, , I 1,11111, , , +t!"' Ir
1000
10000
100000 -=-r-.-r-r-.-r-r-.-r-r-.-r-r-.-r-r-.-r-r-.-r-r-.-r-r-.-r-r-.-r-r..,.....,..,...,.....,..,...,.....,..,...,.....,..,...,.....,..,...,.....,..,...,.....,..,....-r=l
o 1000 2000 3000 4000
Wdssertiefe [m]
5000
Abb. 6.4 Zentraler Südatlantik: Zusammenhang zwischen Wassertiefe der Probennahme und maximaler Anzahl magnetotaktischer Bakterien in 50 ~I
Sediment.
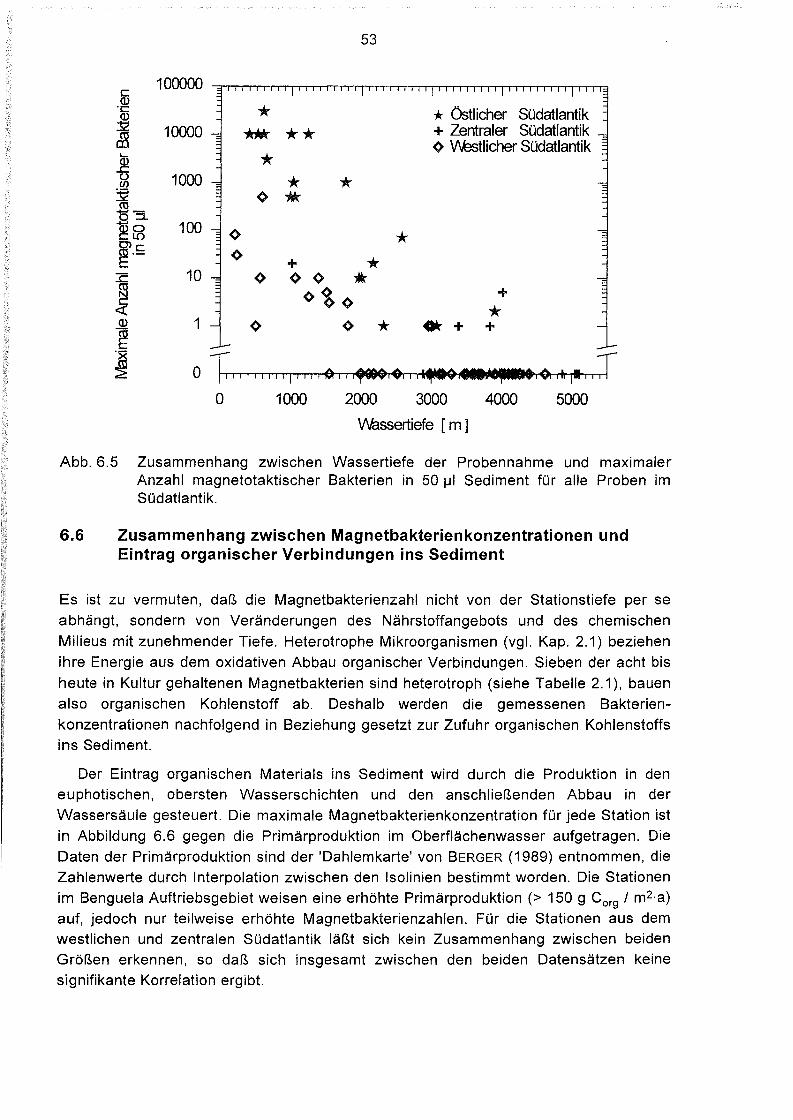
53
100000
* ÖStlicher Südatlantik10000 ** + Zentraler Südatlantik
o \J\kstlicher Südatlantik
1000 * *0 1*
100
*+ *10 0 o 0 ..o~o +
*1 0 0 * .+ +
55'C
!~~0"5..~ß[.~
~(])
~
~ 0 l:, I I " '" I' " I 0 I I 1~1~11,~I'.;~1I18.,Oi 1IlIIIIJ
o 1000 2000 3000 4000 5000
\Aßssertiefe [m]
Abb.6.5 Zusammenhang zwischen Wassertiefe der Probennahme und maximalerAnzahl magnetotaktischer Bakterien in 50 1-/1 Sediment für alle Proben imSüdatlantik.
6.6 Zusammenhang zwischen Magnetbakterienkonzentrationen undEintrag organischer Verbindungen ins Sediment
Es ist zu vermuten, daß die Magnetbakterienzahl nicht von der Stationstiefe per se
abhängt, sondern von Veränderungen des Nährstoffangebots und des chemischenMilieus mit zunehmender Tiefe. Heterotrophe Mikroorganismen (vgl. Kap. 2.1) beziehen
ihre Energie aus dem oxidativen Abbau organischer Verbindungen. Sieben der acht bisheute in Kultur gehaltenen Magnetbakterien sind heterotroph (siehe Tabelle 2.1), bauenalso organischen Kohlenstoff ab. Deshalb werden die gemessenen Bakterien
konzentrationen nachfolgend in Beziehung gesetzt zur Zufuhr organischen Kohlenstoffsins Sediment.
Der Eintrag organischen Materials ins Sediment wird durch die Produktion in deneuphotischen, obersten Wasserschichten und den anschließenden Abbau in derWassersäule gesteuert. Die maximale Magnetbakterienkonzentration für jede Station istin Abbildung 6.6 gegen die Primärproduktion im Oberflächenwasser aufgetragen. DieDaten der Primärproduktion sind der 'Dahlemkarte' von BERGER (1989) entnommen, dieZahlenwerte durch Interpolation zwischen den Isolinien bestimmt worden. Die Stationenim Benguela Auftriebsgebiet weisen eine erhöhte Primärproduktion (> 150 g Corg I m2 'a)auf, jedoch nur teilweise erhöhte Magnetbakterienzahlen. Für die Stationen aus demwestlichen und zentralen Südatlantik läßt sich kein Zusammenhang zwischen beidenGrößen erkennen, so daß sich insgesamt zwischen den beiden Datensätzen keinesignifikante Korrelation ergibt.
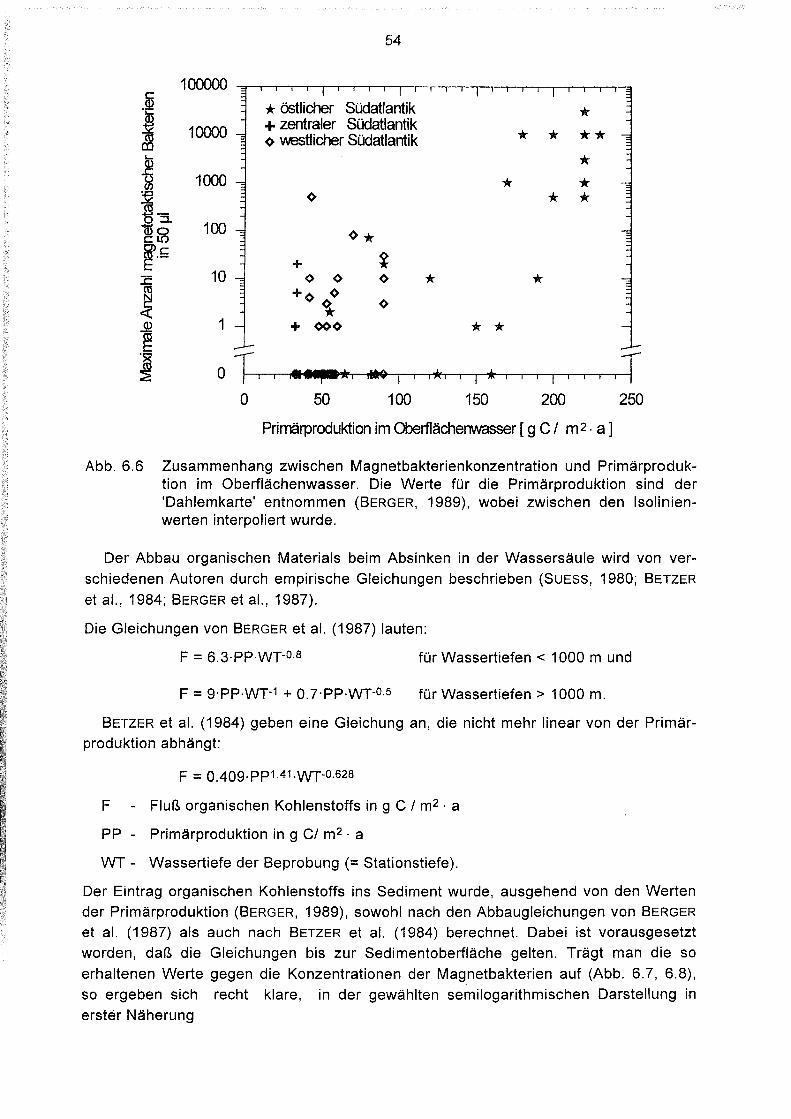
54
250
*******
*
200
*
*
*
* *
150
*
* östlieher Südatlantik+ zentraler Südatlantik~ \NeStlieher Südatlantik
o T"-....,i••I4I.N1I.~I.llr1I*fr,-rtilH.....,-.,-,-.,rlc*fr,-.,-,-'lrlIr*'--r--r-.,-.,-,-.,r--r--r-r-,To 50 100
1
10
100
1000
10000
Primärproduktion im OJerflächenwasser [ 9 C/ m2. a ]
Abb. 6.6 Zusammenhang zwischen Magnetbakterienkonzentration und Primärproduktion im Oberflächenwasser. Die Werte für die Primärproduktion sind der'Dahlemkarte' entnommen (BERGER, 1989), wobei zwischen den Isolinienwerten interpoliert wurde.
Der Abbau organischen Materials beim Absinken in der Wassersäule wird von ver
schiedenen Autoren durch empirische Gleichungen beschrieben (SUESS, 1980; BETZER
et al., 1984; BERGER et al., 1987).
Die Gleichungen von BERGER et al. (1987) lauten:
F = 6.3·PP·WT-o.8 für Wassertiefen < 1000 mund
F =9·PP·WT-1 + 0.7·PP·WT-o.5 für Wassertiefen > 1000 m.
BETZER et al. (1984) geben eine Gleichung an, die nicht mehr linear von der Primärproduktion abhängt:
F = OA09·Pp1.41.WT-o.628
F - Fluß organischen Kohlenstoffs in g C I m2 . a
pp - Primärproduktion in g CI m2 . a
WT - Wassertiefe der Beprobung (= Stationstiefe).
Der Eintrag organischen Kohlenstoffs ins Sediment wurde, ausgehend von den Wertender Primärproduktion (BERGER, 1989), sowohl nach den Abbaugleichungen von BERGER
et al. (1987) als auch nach BETZER et al. (1984) berechnet. Dabei ist vorausgesetztworden, daß die Gleichungen bis zur Sedimentoberfläche gelten. Trägt man die so
erhaltenen Werte gegen die Konzentrationen der Magnetbakterien auf (Abb. 6.7, 6.8),so ergeben sich recht klare, in der gewählten semilogarithmischen Darstellung in
erster Näherung
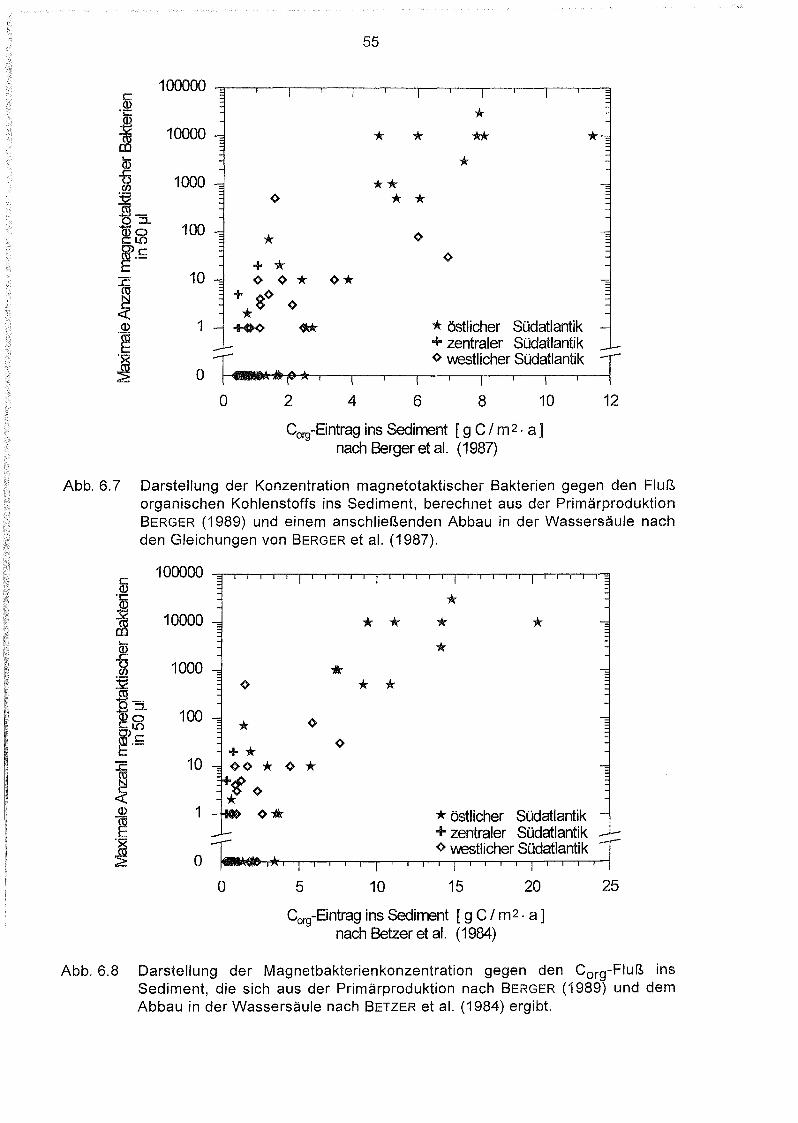
55
100000C(J)
*'e;
! 10000 * * **l...
*(J)
-5 1000 **cn~ * *t'tl0"5..
100wo* ~CL()
~,f;+ * ~
..c 10 ~ ~ * ~*
~+ $~
~
*(J) 1 ~ ~ * östlicher Südatlantik
~ + zentraler Südatlantik
~ o Tnlllll\~ ~ *~ westlicher Südatlantik =rI I
0 2 4 6 8 10 12
Corg-Eintrag ins Sediment [g C / m 2 . a ]nach Berger et al, (1987)
Abb. 6.7 Darstellung der Konzentration magnetotaktischer Bakterien gegen den Flußorganischen Kohlenstoffs ins Sediment, berechnet aus der PrimärproduktionBERGER (1989) und einem anschließenden Abbau in der Wassersäule nachden Gleichungen von BERGER et al. (1987).
100000C(J)
'e; *~ 10000 * * *&il...
*(J)
-5 1000 ...cn~ * *t'tl0-..... :::L.
100~o ~O)L()
~.f; ~
..c 10 ~ *~(J) 1 * östlicher Südatlantik~ + zentraler Südatlantik
~ o ~\.'*i : ~tli,ch,eriS~d~l~nt:k i =rI I I i
0 5 10 15 20 25
Corg-Eintrag ins Sediment [g C / m 2 , a ]nach Betzer et al. (1984)
Abb. 6.8 Darstellung der Magnetbakterienkonzentration gegen den Corg-Fluß insSediment, die sich aus der Primärproduktion nach BERGER (1989) und demAbbau in der Wassersäule nach BETZER et al. (1984) ergibt.

56
lineare Zusammenhänge zwischen dem Eintrag organischen Kohlenstoffs und denMagnetbakterienkonzentrationen im Sediment. Die Werte für den COrg-Eintrag liegen fürdas Gebiet der Fahrt M 16/2, auf der keine lebenden Magnetbakterien identifiziertwerden konnten, in Abbildung 6.7 zwischen 0.5 und 1.2 g C/(m2 'a), in Abbildung 6.8zwischen 0.4 und 1.1 g C/(m2·a).
Bei Verwendung der Formel nach BETZER et al. (1984) ist die Streuung der Datenoffensichtlich deutlich geringer als für die Gleichungen nach BERGER et al. (1987). Auchin anderen Fällen hat sich die BETZER Gleichung besser bewährt. So ließen sich damit indrei Hochproduktionssystemen des östlichen Südatlantiks die Werte der Primärproduktion am besten aus den in Sinkstoffallen gemessenen Kohlenstoffflüssenrekonstruieren (ANDERSEN, in Vorb.).
Saisonalität des Nährstoffeintrages
Die Primärproduktionswerte der 'Dahlemkarte' (BERGER, 1989) sind über mehrere Jahregemittelt. Saisonale Daten sind seit 1987 nicht verfügbar. Der Satellit 'SeaWiff' mit dem'Coastal Zone Color Scanner', der die Biomassenverteilung anhand der Lichtabsorptionvon Chlorophyll messen soll, ist noch nicht gestartet. Der Einfluß der saisonalenVariation des Nährstoffeintrags ins Sediment kann daher hier nicht berücksichtigtwerden.
Die saisonale Variation ist aber beträchtlich. Während einer Hochproduktionsphase('Blüte') kann die Konzentration an Primärproduzenten so groß werden, daß sie sich zuAggregaten zusammenlagern. Diese Aggregate sinken wesentlich schneller ab alskleinere einzelne Partikel. Dadurch ist ihre Verweildauer in der Wassersäule geringerund somit auch die Zeitdauer, in der organische Substanzen gelöst und ins umgebendeWasser abgegeben werden. Die Menge der dabei remineralisierten Substanz ist nachSMITH et al. (1992) höher als bisher angenommen, da in den herabsinkendenAggregaten durch bakteriell induzierte Enzymtätigkeit ständig organischer Kohlenstoffabgebaut wird. Auf den Partikeln haftende Bakterien sondern Enzyme ab (Exoenzyme),die mehr organische Verbindungen zersetzen, als die Bakterien aufnehmen können.Dennoch liefern solche Aggregate den höchsten Nährstoffeintrag ins Sediment (LOCHTE& TURLEY, 1988). Nach TURLEY & LOCHTE (1990) dauert es etwa 6 Wochen, bis sie denBoden der Tiefsee erreichen. Dort nimmt die biologische Aktivität im Sediment zunächststark zu und erst mit fortschreitendem Abbau der Aggregate wieder ab.
Für das Gebiet der Expedition M 16/2 (Anhang B, Karte 1) liegen älterejahreszeitliche Daten der Chlorophyllabsorption aus den Jahren 1979 bis 1983 vor(CAMPBELL et al., 1992). Danach liegt das Maximum der Chlorophyllabsorption für diebeprobten Gebiete in den Monaten Mai bis Juli/August. Da die Fahrt M 16/2 Anfang Maistattfand, konnten Aggregate, die während des Chlorophyll Maximums gebildet wurden,zu diesem Zeitpunkt das Sediment noch nicht erreicht haben. Die Sedimente wurdenvermutlich beprobt, als leicht abbaubare organische Verbindungen im Sediment starkabgereichert waren. Dies liefert eine mögliche Erklärung dafür, daß während der FahrtM 16/2 keine aktiv beweglichen Magnetbakterien nachgewiesen werden konnten.

57
6.7 Zusammenhang zwischen Magnetbakterienkonzentration und Gehaltan organischem Kohlenstoff und Stickstoff im Sediment
Der Gesamtgehalt an organischem Kohlenstoff und Stickstoff im Sediment ist ein Maßfür das Angebot von Nährstoffen, das Bakterien im allgemeinen und Magnetbakterien imspeziellen zur Verfügung steht. Es stellt sich die Frage, ob der Zusammenhangzwischen den Magnetbakterienzahlen und den berechneten Werten des Eintrags anorganischem Kohlenstoff ins Sediment auch durch Messungen des Gehalts vonKohlenstoff und Stickstoff im Sediment untermauert werden kann.
Unter organischem Kohlenstoff faßt man solche Kohlenstoffverbindungen zusammen,die im Gegensatz zum karbonatischen Kohlenstoff bei einer Temperatur von 1200 oeverbrennen. Nicht alle organischen Kohlenstoffverbindungen sind jedoch für Mikroorganismen abbaubar. Huminstoffe und Kerogene lassen sich beispielsweise vonBakterien nur sehr schwer zersetzen. Eine diesbezügliche genauere Spezifizierung desorganischen Kohlenstoffs ist nicht verfügbar. Stickstoff hingegen liegt größtenteils in fürMikroorganismen abbaubaren Substanzen vor. Nur ein kleinerer Anteil kann inTonmineralen gebunden vorliegen (MÜLLER, 1977). Dieser ist den Bakterien nichtzugänglich.
In den Abbildungen 6.9 und 6.10 ist die Magnetbakterienkonzentration aufgetragengegen den Gehalt an organischem Kohlenstoff und Stickstoff im obersten Sedimentzentimeter (P.J. MÜLLER, unveröffentlichte Daten). Entsprechende Messungen für dieProben der Fahrten M 23/1 und M 23/2 waren vor Abschluß dieser Arbeit noch nichtverfügbar.
Die Zunahme der Magnetbakterienkonzentration mit steigendem Gehalt an organischem Kohlenstoff ist ähnlich deutlich ausgeprägt wie die Korrelation mit den Stickstoffgehalten. Mehr als 100 Magnetbakterien pro 50 1-11 fanden sich nur in Sedimenten miteinem Corg-Gehalt >10 mg/cm3 und NtocGehalten >1.5 mg/cm3 . Allein zwei Stationender Fahrt M 20/2, GeoB 1704 (Benguela Auftriebsgebiet, 400 m Wassertiefe) mit demhöchsten und die Station GeoB 1726 auf dem Walfisch Rücken mit dem niedrigstenCorg- und Ntot-Gehalt aller Stationen, fallen etwas aus dem linearen Trend dersemilogarithmischen Darstellungen. An den Stationen der Expedition M 16/2 sind dieKohlenstoff- (1.9 bis 3 mg/cm3) und Stickstoffgehalte (0.3 bis 0.5 mg/cm3) jeweils sehrniedrig.
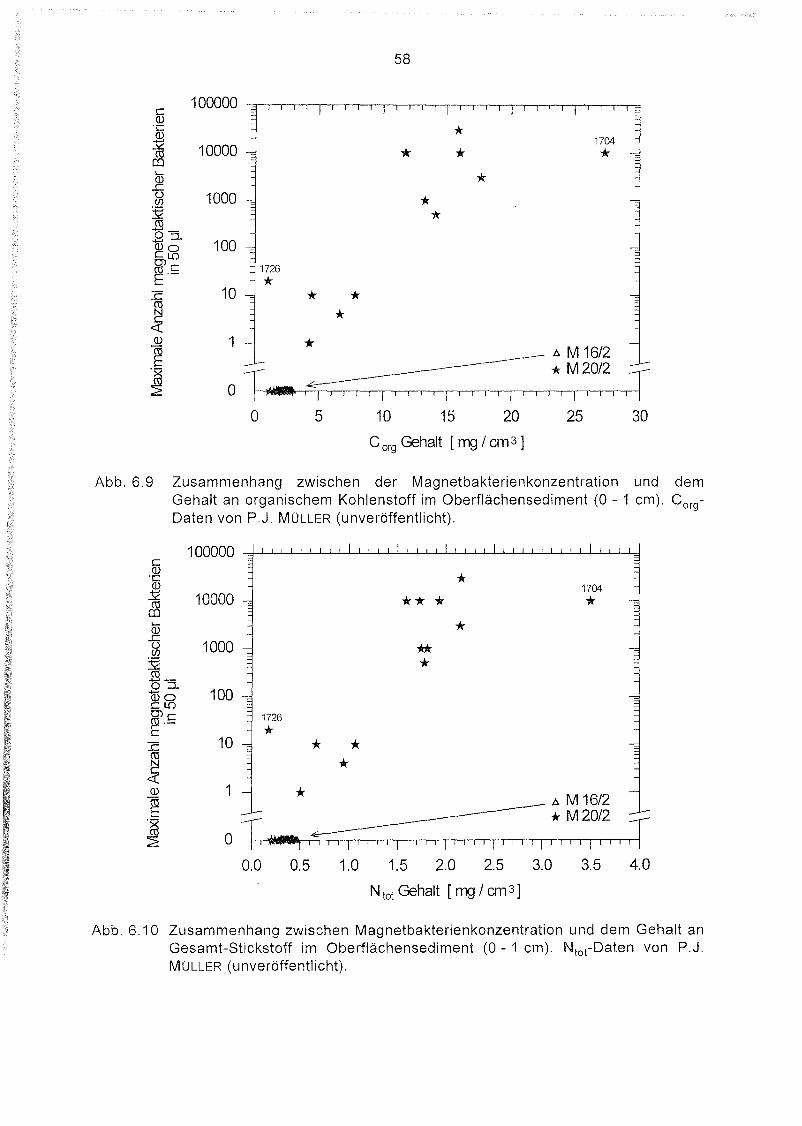
58
100000CQ)'C '*~ 1704
&l 10000 * '* *I-
*Q).L:0 1000 *(f)
~ *co0-...... :::::L
100WoCL!)Olc~.-
.L: 10 * *coN *~Q) 1 *~ h. M16/2
~ ol=~* M20/2 ,fI , I I I
0 5 10 15 20 25 30
Corg Gehalt [mg / cm 3 ]
Abb. 6.9 Zusammenhang zwischen der Magnetbakterienkonzentration und demGehalt an organischem Kohlenstoff im Oberflächensediment (0 - 1 cm). CorgDaten von P.J. MÜLLER (unveröffentlicht),
100000CQ)
*'CQ) 1704~ 10000 ** * *&lI-
*Q).L:
10000 **(f)
~ *coO:::::L
100wOCL!)Olc~.-
.L: 10 * *co
*~Q) 1 * h. M16/2~
~ -r 4""1 '
* M20/2 ,J0 I I , 1 I
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0
Ntot Gehalt [mg 1cm 3 ]
Abb. 6.10 Zusammenhang zwischen Magnetbakterienkonzentration und dem Gehalt anGesamt-Stickstoff im Oberflächensediment (0 - 1 cm). Ntot-Daten von P.J.MÜLLER (unveröffentlicht).
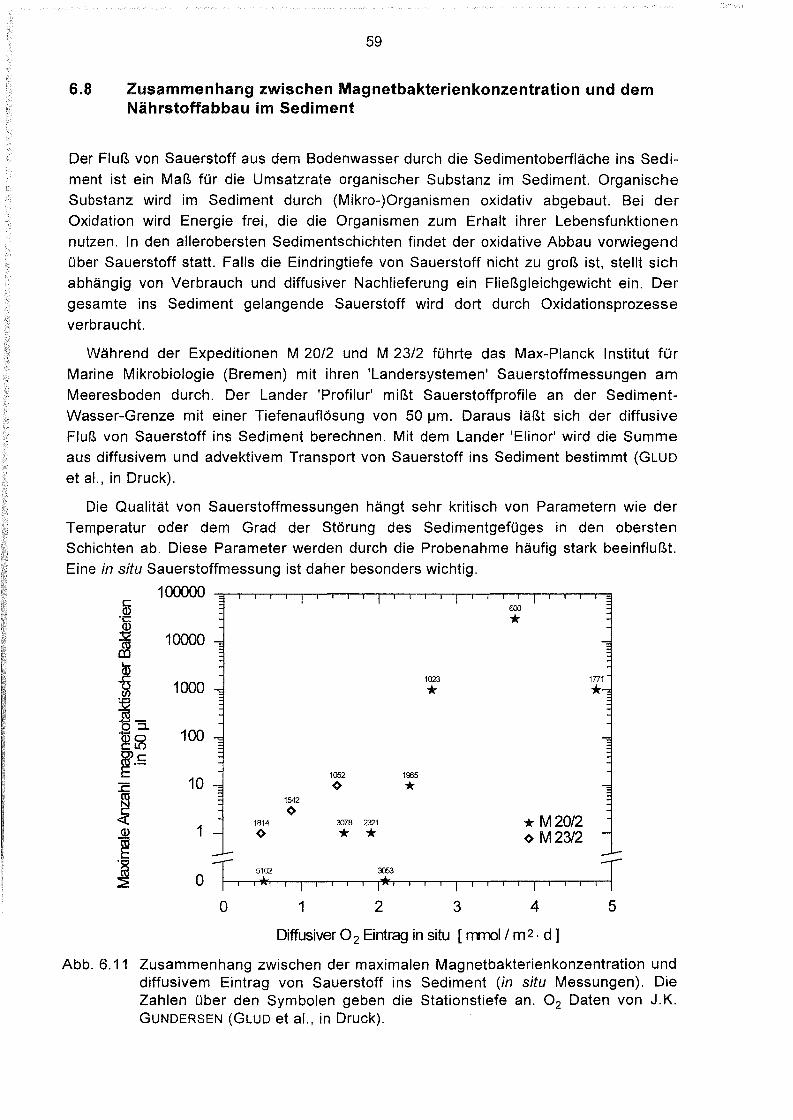
59
6.8 Zusammenhang zwischen Magnetbakterienkonzentration und demNährstoffabbau im Sediment
Der Fluß von Sauerstoff aus dem Bodenwasser durch die Sedimentoberfläche ins Sediment ist ein Maß für die Umsatzrate organischer Substanz im Sediment. OrganischeSubstanz wird im Sediment durch (Mikro-)Organismen oxidativ abgebaut. Bei derOxidation wird Energie frei, die die Organismen zum Erhalt ihrer Lebensfunktionennutzen. In den allerobersten Sedimentschichten findet der oxidative Abbau vorwiegendüber Sauerstoff statt. Falls die Eindringtiefe von Sauerstoff nicht zu groß ist, stellt sichabhängig von Verbrauch und diffusiver Nachlieferung ein Fließgleichgewicht ein. Dergesamte ins Sediment gelangende Sauerstoff wird dort durch Oxidationsprozesseverbraucht.
Während der Expeditionen M 20/2 und M 23/2 führte das Max-Planck Institut fürMarine Mikrobiologie (Bremen) mit ihren 'Landersystemen' Sauerstoffmessungen amMeeresboden durch. Der Lander 'Profilur' mißt Sauerstoffprofile an der SedimentWasser-Grenze mit einer Tiefenauflösung von 50 IJm. Daraus läßt sich der diffusiveFluß von Sauerstoff ins Sediment berechnen. Mit dem Lander 'Elinor' wird die Summeaus diffusivem und advektivem Transport von Sauerstoff ins Sediment bestimmt (GLUDet al., in Druck).
Die Qualität von Sauerstoffmessungen hängt sehr kritisch von Parametern wie derTemperatur oder dem Grad der Störung des Sedimentgefüges in den oberstenSchichten ab. Diese Parameter werden durch die Probenahme häufig stark beeinflußt.Eine in situ Sauerstoffmessung ist daher besonders wichtig.
100000 --:::r-....---.-.,-...,...-,.---,--r-....---.-.,-...,...-,.---,--,.-....---.-.,-...,...-,.---,--,.-....---.---.--::ICQ)'C
~ 10000&lL..
~ 1000~ctl0"5-
100~ß~.5
1: 10
~1542
~
11814
Q) ~
~
~ o1, 5102
,*, I
0 1
1023
*
1052 1965
~ *3078 2321 * M20/2* * ~ M2312
3053 ,=r1*' I I
2 3 4 5
Diffusiver O 2 Eintrag in situ [mnoll m 2 . d]
Abb. 6.11 Zusammenhang zwischen der maximalen Magnetbakterienkonzentration unddiffusivem Eintrag von Sauerstoff ins Sediment (in situ Messungen). DieZahlen über den Symbolen geben die Stationstiefe an. 02 Daten von J.K.GUNDERSEN (GLUD et al., in Druck).
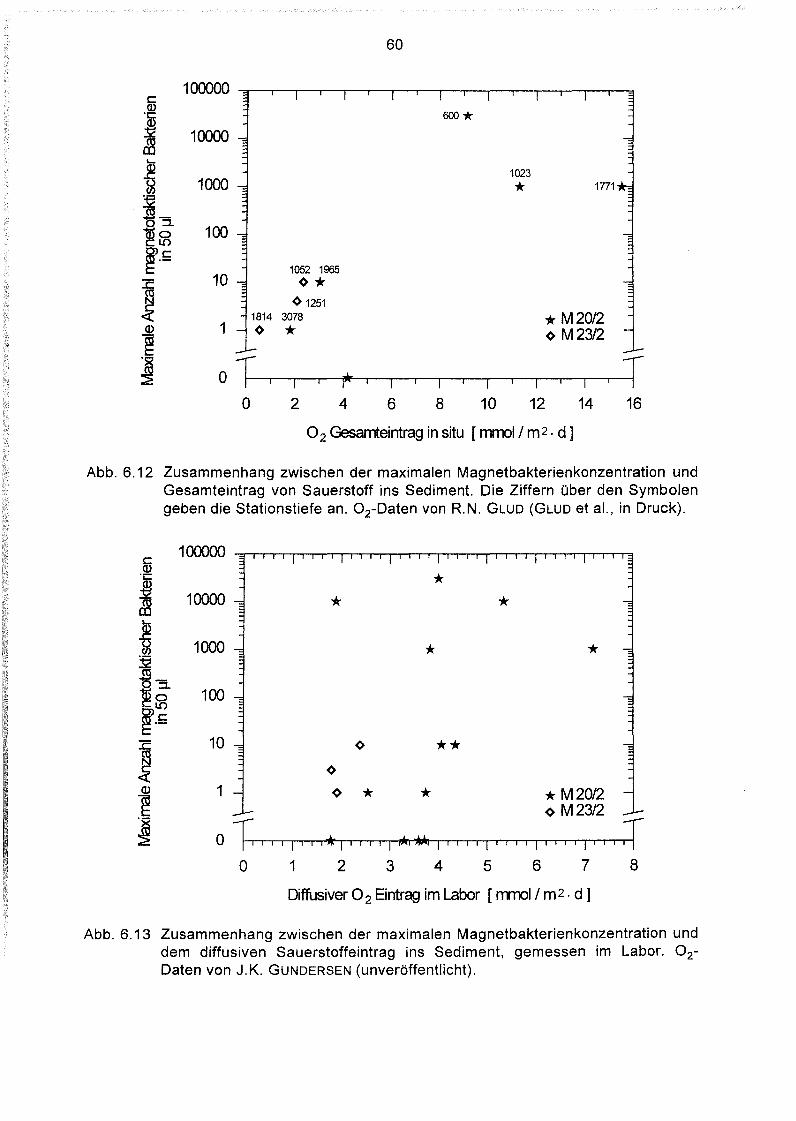
60
100000CQ}'C
~ 10000~
~ 1000~m0"5-
100~~~.f;
1052 1965::c 10 ~*
~ ~ 12511814 3078
Q} 1 ~ *~
~ 01= I 1*0 2 4 6
1023
*
* M20/2~ M2312
I I I I =r8 10 12 14 16
O2 Gesamteintrag in situ [mrrol / m2 . d ]
Abb. 6.12 Zusammenhang zwischen der maximalen Magnetbakterienkonzentration undGesamteintrag von Sauerstoff ins Sediment. Die Ziffern über den Symbolengeben die Stationstiefe an. 02-Daten von R.N. GLUD (GLUD et al., in Druck).
10000
1000
100
10
~
1 ~ * * * M20/2~ M2312
o -r: I , I I I I ,* I' , I , I ,*,. I i I i I I I I I I I I I i I I i I i Jo 1 2 3 4 5 6 7 8
Diffusiver O2 Eintrag im Labor [mrol / m2 . d ]
Abb. 6.13 Zusammenhang zwischen der maximalen Magnetbakterienkonzentration unddem diffusiven Sauerstoffeintrag ins Sediment, gemessen im Labor. 02Daten von J.K. GUNDERSEN (unveröffentlicht).

61
Die Abbildungen 6.11 und 6.12 verdeutlichen den Zusammenhang zwischenbeobachteter Magnetbakterienzahl und gemessenem Sauerstoffeintrag. Bei hohenUmsatzraten organischen Materials, also erhöhter bakterieller Aktivität, ist demnach
auch eine größere Anzahl magnetotaktischer Bakterien im Sediment nachweisbar. Anden Stationen der Fahrt M 23/2 im westlichen Südatlantik (Anhang B: Karte 4) liegen diediffusiven Sauerstoffflüsse zwischen 0.4 und 1.5 mmol/(m2 . d). Die Werte im östlichen
Südatlantik (Fahrt M 20/2) liegen bei ähnlicher Wassertiefe höher (2.2 bis4.8 mmol/(m2·d)). Die biologische Aktivität vor Brasilien bleibt also deutlich unter der imAuftriebsgebiet vor Namibia. Die Magnetbakterienkonzentration folgt generell diesemTrend. Allerdings sind beträchtliche Streuungen, möglicherweise aufgrund kleinräumigerHeterogenitäten im Sediment vorhanden.
Aus Abbildung 6.13 läßt sich ablesen, um wieviel schlechter die Anzahl magnetischerBakterien im Sediment mit den im Labor gemessenen Sauerstoffflüssen korreliert. Diesmuß vermutlich damit erklärt werden, daß die Magnetbakterien weniger sensibel aufVeränderungen reagieren, die aus der Probenentnahme vom Meeresboden und demTransport des Sediments resultieren als die Sauerstoffflüsse.
6.9 Zusammenhänge zwischen der Magnetbakterienkonzentration undanderen geochemischen Parametern
In diesem Kapitel werden die Konzentrationen der Magnetbakterien Parametern derPorenwasserprofile von Nitrat, Eisen und Mangan gegenübergestellt. Diese drei Stoffebauen zusammen mit Sauerstoff und Sulfat die geochemische Zonierung im Sedimentauf (Kap. 5.3.1).
Die Porenwasserprofile für Nitrat wurden an Bord gemessen. Das Kernmaterial wurde
unmittelbar, nachdem es an Deck war, bei 4°C gelagert. Die obersten Sedimentschichten wurden in Scheiben von 0.5 cm Dicke zerlegt. Das Porenwasser dieserSegmente wurde ausgepreßt und anschließend chemisch auf N03- analysiert. DieProben für die Mangan- und Eisenanalysen wurden an Bord genommen und später imLabor in Bremen gemessen.
Nitrat wird im Zuge des Abbaus organischer Stickstoffverbindungen mit Sauerstoffgebildet, die maximale Nitratkonzentration wird meist etwa in der Tiefe erreicht, in dersämtlicher Sauerstoff abgebaut ist. Von diesem Maximum aus diffundiert Nitrat in derSedimentsäule nach oben und unten, wo es wieder zersetzt wird. Mindestens drei der
acht in Kultur vorliegenden Magnetbakterienstämme können Nitrat oder NOx
Verbindungen zum Abbau organischer Substanz benutzen (siehe Tabelle 2.1).Abbildung 6.14 zeigt die Korrelation zwischen der Konzentration an Magnetbakterienund dem vom Maximum abwärts gerichteten Nitratfluß. Die Werte streuen stark. Diesdürfte unter anderem auf die relativ großen Probenabstände bei der geochemischenBeprobung zurückzuführen sein. Dennoch erkennt man tendenziell einen Anstieg derMagnetbakterienzahlen mit zunehmendem, nach unten gerichtetem Nitratfluß. Wie in
Kapitel 6.10 genauer ausgeführt wird, lebt eine große Zahl magnetotaktischer Bakterienin der Denitrifikationszone, also in der Schicht unterhalb des Nitratmaximums, in der
Nitrat abgebaut wird.
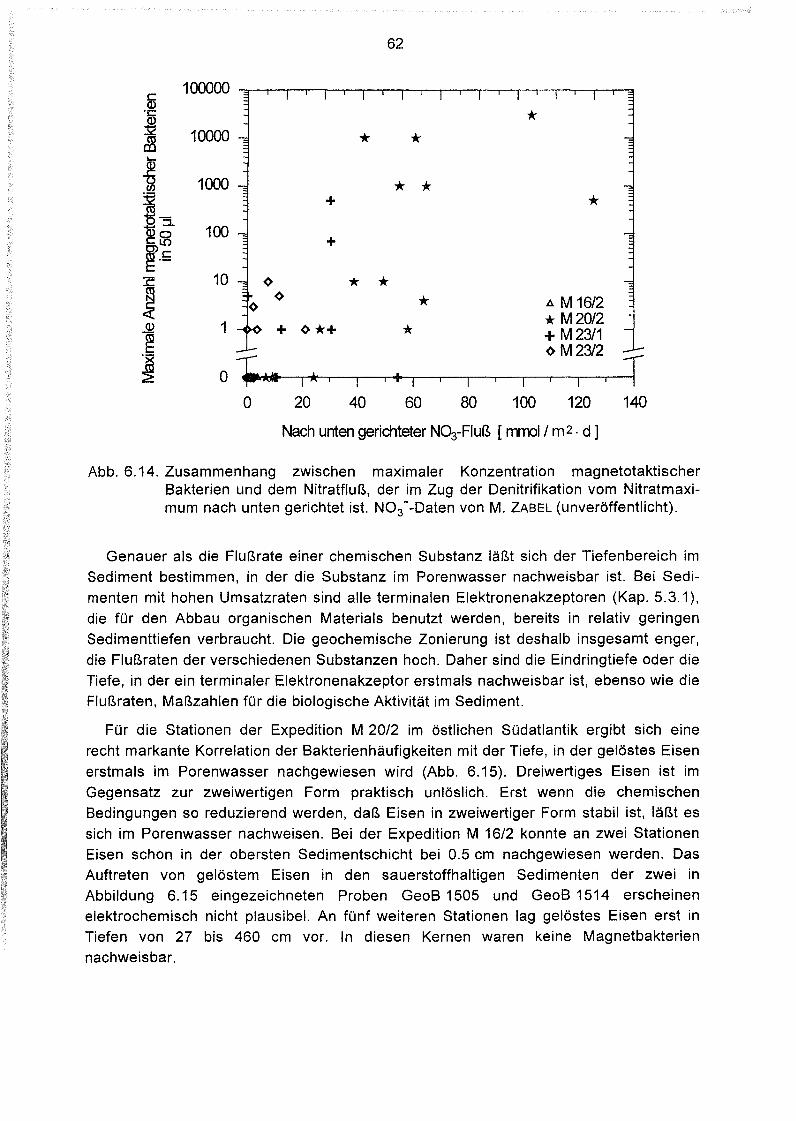
62
100000IDoe::
*~ooסס1 * *~
L-
~ 1000 * *~ + *15-~::J. 100g +~o~
:E 10 ~ * *~
~
* AM 16/2* M20/2(I) 1 + ~*+ * + M2311
~ ~ M2312
~ O~~ =r1* i + II 1 1 1
0 20 40 60 80 100 120 140
Nach unten gerichteter N03-Fluß [rrmol/ m 2 . d ]
Abb. 6.14. Zusammenhang zwischen maximaler Konzentration magnetotaktischerBakterien und dem Nitratfluß, der im Zug der Denitrifikation vom Nitratmaximum nach unten gerichtet ist. N03- -Daten von M. ZABEL (unveröffentlicht).
Genauer als die Flußrate einer chemischen Substanz läßt sich der Tiefenbereich imSediment bestimmen, in der die Substanz im Porenwasser nachweisbar ist. Bei Sedimenten mit hohen Umsatzraten sind alle terminalen Elektronenakzeptoren (Kap. 5.3.1),die für den Abbau organischen Materials benutzt werden, bereits in relativ geringen
Sedimenttiefen verbraucht. Die geochemische Zonierung ist deshalb insgesamt enger,die Flußraten der verschiedenen Substanzen hoch. Daher sind die Eindringtiefe oder dieTiefe, in der ein terminaler Elektronenakzeptor erstmals nachweisbar ist, ebenso wie dieFlußraten, Maßzahlen für die biologische Aktivität im Sediment.
Für die Stationen der Expedition M 20/2 im östlichen Südatlantik ergibt sich einerecht markante Korrelation der Bakterienhäufigkeiten mit der Tiefe, in der gelöstes Eisen
erstmals im Porenwasser nachgewiesen wird (Abb. 6.15). Dreiwertiges Eisen ist imGegensatz zur zweiwertigen Form praktisch unlöslich. Erst wenn die chemischenBedingungen so reduzierend werden, daß Eisen in zweiwertiger Form stabil ist, läßt essich im Porenwasser nachweisen. Bei der Expedition M 16/2 konnte an zwei StationenEisen schon in der obersten Sedimentschicht bei 0.5 cm nachgewiesen werden. DasAuftreten von gelöstem Eisen in den sauerstoffhaitigen Sedimenten der zwei inAbbildung 6.15 eingezeichneten Proben GeoB 1505 und GeoB 1514 erscheinenelektrochemisch nicht plausibel. An fünf weiteren Stationen lag gelöstes Eisen erst inTiefen von 27 bis 460 cm vor. In diesen Kernen waren keine Magnetbakterien
nachweisbar.
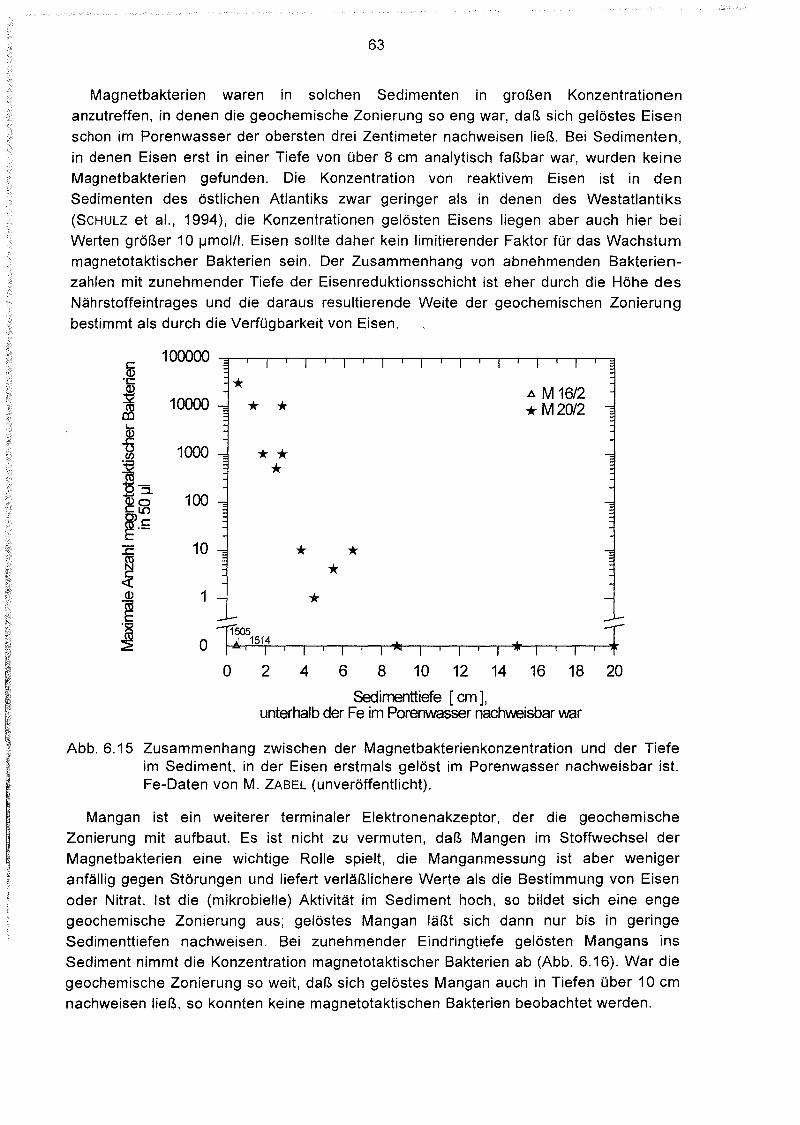
63
Magnetbakterien waren in solchen Sedimenten in großen Konzentrationenanzutreffen, in denen die geochemische Zonierung so eng war, daß sich gelöstes Eisenschon im Porenwasser der obersten drei Zentimeter nachweisen ließ. Bei Sedimenten,in denen Eisen erst in einer Tiefe von über 8 cm analytisch faßbar war, wurden keineMagnetbakterien gefunden. Die Konzentration von reaktivem Eisen ist in denSedimenten des östlichen Atlantiks zwar geringer als in denen des Westatlantiks(SCHULZ et al., 1994), die Konzentrationen gelösten Eisens liegen aber auch hier beiWerten größer 10 IJmol/l. Eisen sollte daher kein limitierender Faktor für das Wachstummagnetotaktischer Bakterien sein. Der Zusammenhang von abnehmenden Bakterienzahlen mit zunehmender Tiefe der Eisenreduktionsschicht ist eher durch die Höhe desNährstoffeintrages und die daraus resultierende Weite der geochemischen Zonierungbestimmt als durch die Verfügbarkeit von Eisen.
20
.6. M16/2* M20/2
I
4
*
* **
*
2I * I * I
6 8 10 12 14 16 18
Sedimenttiefe [cm],unterhalb der Fe im Porenwasser nachweisbar war
***
o~505( 1514
I I
c: ooסס10(])'C
~ooסס1
~s...
~ 1000
~0-~ß 100
~.5:c 10
~(]) 1~
~ 0
Abb. 6.15 Zusammenhang zwischen der Magnetbakterienkonzentration und der Tiefeim Sediment, in der Eisen erstmals gelöst im Porenwasser nachweisbar ist.Fe-Daten von M. ZABEL (unveröffentlicht).
Mangan ist ein weiterer terminaler Elektronenakzeptor, der die geochemischeZonierung mit aufbaut. Es ist nicht zu vermuten, daß Mangen im Stoffwechsel derMagnetbakterien eine wichtige Rolle spielt, die Manganmessung ist aber wenigeranfällig gegen Störungen und liefert verläßlichere Werte als die Bestimmung von Eisenoder Nitrat. Ist die (mikrobielle) Aktivität im Sediment hoch, so bildet sich eine engegeochemische Zonierung aus; gelöstes Mangan läßt sich dann nur bis in geringeSedimenttiefen nachweisen. Bei zunehmender Eindringtiefe gelösten Mangans insSediment nimmt die Konzentration magnetotaktischer Bakterien ab (Abb. 6.16). War diegeochemische Zonierung so weit, daß sich gelöstes Mangan auch in Tiefen über 10 cmnachweisen ließ, so konnten keine magnetotaktischen Bakterien beobachtet werden.
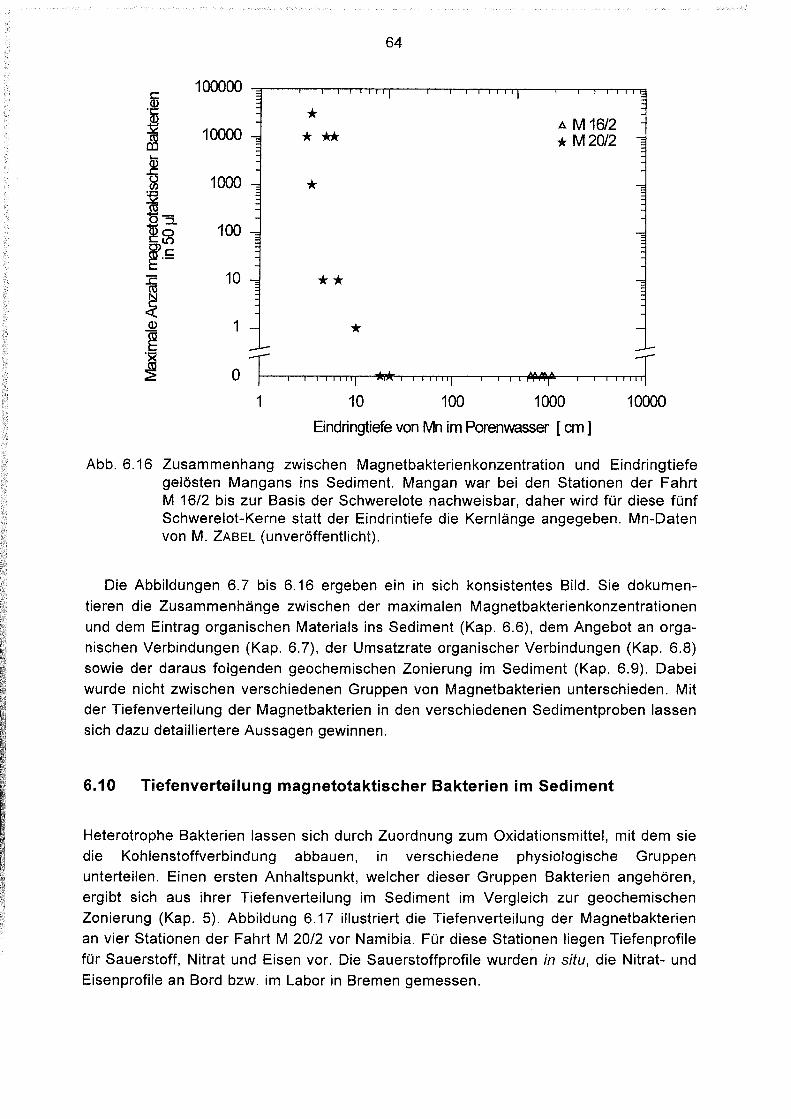
1IIIII1 ~iiiiilil
c: 100000Q)'C
! 10000...~ 1000~ctl0-_::::L
100~fiS[.~
::E 10
~Q) 1~
~ 0 -r1
64
****
*
**
*
10 100
Ä M 16/2* M20/2
j I I I ,'PAtt
1000Eindringtiefe von Mn im Porenwasser [cm]
Abb. 6.16 Zusammenhang zwischen Magnetbakterienkonzentration und Eindringtiefegelösten Mangans ins Sediment. Mangan war bei den Stationen der FahrtM 16/2 bis zur Basis der Schwerelote nachweisbar, daher wird für diese fünfSchwerelot-Kerne statt der Eindrintiefe die Kernlänge angegeben. Mn-Datenvon M. ZABEl (unveröffentlicht).
Die Abbildungen 6.7 bis 6.16 ergeben ein in sich konsistentes Bild. Sie dokumentieren die Zusammenhänge zwischen der maximalen Magnetbakterienkonzentrationenund dem Eintrag organischen Materials ins Sediment (Kap. 6.6), dem Angebot an organischen Verbindungen (Kap. 6.7), der Umsatzrate organischer Verbindungen (Kap. 6.8)sowie der daraus folgenden geochemischen Zonierung im Sediment (Kap. 6.9). Dabeiwurde nicht zwischen verschiedenen Gruppen von Magnetbakterien unterschieden. Mitder Tiefenverteilung der Magnetbakterien in den verschiedenen Sedimentproben lassensich dazu detailliertere Aussagen gewinnen.
6.10 Tiefenverteilung magnetotaktischer Bakterien im Sediment
Heterotrophe Bakterien lassen sich durch Zuordnung zum Oxidationsmittel, mit dem siedie Kohlenstoffverbindung abbauen, in verschiedene physiologische Gruppenunterteilen. Einen ersten Anhaltspunkt, welcher dieser Gruppen Bakterien angehören,ergibt sich aus ihrer Tiefenverteilung im Sediment im Vergleich zur geochemischenZonierung (Kap. 5). Abbildung 6.17 illustriert die Tiefenverteilung der Magnetbakterienan vier Stationen der Fahrt M 20/2 vor Namibia. Für diese Stationen liegen Tiefenprofilefür Sauerstoff, Nitrat und Eisen vor. Die Sauerstoffprofile wurden in situ, die Nitrat- undEisenprofile an Bord bzw. im Labor in Bremen gemessen.
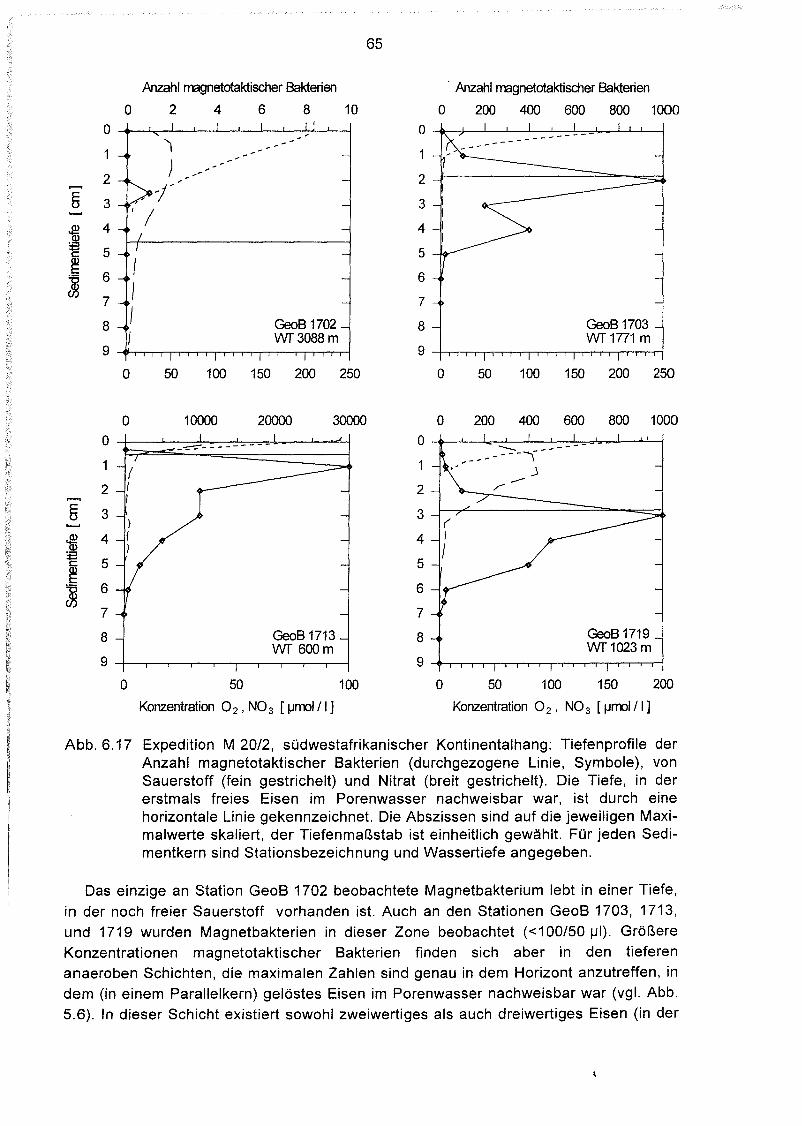
65
Anzahl magnetotaktischer Bakterien
o 2 4 6 8 10
Anzahl magnetotaktischer Bakterien
o 200 400 600 800 1000
GeoB 1703wr 1771 m
2
3
4
5
6
7
8
9 --f--,--,-,.-,--,....,..,..-,.-,-......,........,..~-.--r--r-r-..-.--r-r-r--i
Geo81702wr3088m
{
III
0--+----'...---''-...I.---'----'----'--'-..L..L--'-----1
1
2
3
4
5
6
7
8
9 .......,.,....,,.,.....,-,-,-,-,-,--,....,..,..-,.-,-......,....,-,-,...,...,...,...,.--l
o 50 100 150 200 250 o 50 100 150 200 250
o 10000 20000 30000 o 200 400 600 800 1000
GeoB 1719wr 1023 m
(
I
I
o -+---L---J..c--L.-..l.-----l.-..-L.~~.l.--....L.!....-i
1
2
3
4
5
6
7
8
9'-+-.......,......,.......-'--"--'--'-'---"--'-'----r-1r-r--..,.-,--.-r-I
GeoB 1713wr 600 m
(I
o -t---l--~_--'-:::-=-=".,..4-,~--'-..-..."JL.i-~- - - - - - - - --
1
2
3
4
5
6
7
8
9-t---,----,-...--.---r----r-.-...--r--l
o 50 100
Konzentration O2 , NO 3 [lJmoI/l]
o 50 100 150 200
Konzentration O2 , NO 3 [lJrnol / l ]
Abb. 6.17 Expedition M 20/2, südwestafrikanischer Kontinentalhang: Tiefenprofile derAnzahl magnetotaktischer Bakterien (durchgezogene Linie, Symbole), vonSauerstoff (fein gestrichelt) und Nitrat (breit gestrichelt). Die Tiefe, in dererstmals freies Eisen im Porenwasser nachweisbar war, ist durch einehorizontale Linie gekennzeichnet. Die Abszissen sind auf die jeweiligen Maximalwerte skaliert, der Tiefenmaßstab ist einheitlich gewählt. Für jeden Sedimentkern sind Stationsbezeichnung und Wassertiefe angegeben.
Das einzige an Station GeoB 1702 beobachtete Magnetbakterium lebt in einer Tiefe,in der noch freier Sauerstoff vorhanden ist. Auch an den Stationen GeoB 1703, 1713,und 1719 wurden Magnetbakterien in dieser Zone beobachtet «100/50 IJI). GrößereKonzentrationen magnetotaktischer Bakterien finden sich aber in den tieferenanaeroben Schichten, die maximalen Zahlen sind genau in dem Horizont anzutreffen, indem (in einem Parallelkern) gelöstes Eisen im Porenwasser nachweisbar war (vgl. Abb.5.6). In dieser Schicht existiert sowohl zweiwertiges als auch dreiwertiges Eisen (in der
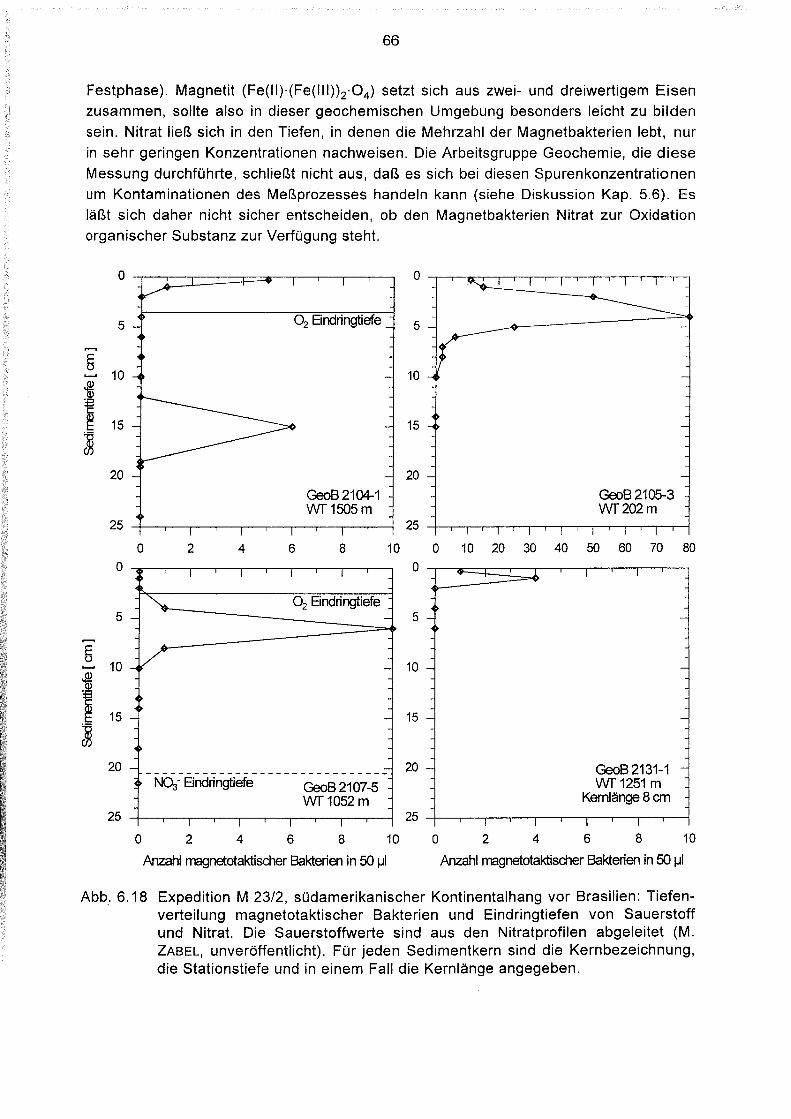
66
Festphase). Magnetit (Fe(II)·(Fe(lIl)h·04) setzt sich aus zwei- und dreiwertigem Eisenzusammen, sollte also in dieser geochemischen Umgebung besonders leicht zu bildensein. Nitrat ließ sich in den Tiefen, in denen die Mehrzahl der Magnetbakterien lebt, nurin sehr geringen Konzentrationen nachweisen. Die Arbeitsgruppe Geochemie, die diese
Messung durchführte, schließt nicht aus, daß es sich bei diesen Spurenkonzentrationenum Kontaminationen des Meßprozesses handeln kann (siehe Diskussion Kap. 5.6). Esläßt sich daher nicht sicher entscheiden, ob den Magnetbakterien Nitrat zur Oxidationorganischer Substanz zur Verfügung steht.
0
Ü:1 Eindringtiefe 5
10
15
20
GeoB2104-1 GeoB2105-3WT1505m WT202m
25
4 6 8 10 0 10 20 30 40 50 60 70 80
I I I0
I I
02 Eindringtiefe
=======~- 5-
-
- 10- -
- 15- -
20 - ~ 20 -
GeoB2107-5WT1052m
N03- EindringtiefeGeoB2131-1 WT1251 m
Kernlänge 8 cmI , I I 25 -!-------,---rl-.,--,.-,-.,---.--,-,------,,--,---1
o 2 4 6 8 10 0 2 4 6 8 10
Anzahl magnetotaktischer Bakterien in 50 ~I Anzahl magnetotaktischer Bakterien in 50 ~I
25
Abb. 6.18 Expedition M 23/2, südamerikanischer Kontinentalhang vor Brasilien: Tiefenverteilung magnetotaktischer Bakterien und Eindringtiefen von Sauerstoffund Nitrat. Die Sauerstoffwerte sind aus den Nitratprofilen abgeleitet (M.ZASEL, unveröffentlicht). Für jeden Sedimentkern sind die Kernbezeichnung,die Stationstiefe und in einem Fall die Kernlänge angegeben.

67
Vier Tiefenprofile magnetotaktischer Bakterien in Kernen der Fahrt M 23/2 vor derSüdostküste Brasiliens sind in Abbildung 6.18 dargestellt. Durch die Verbesserung derBeprobungstechnik ließen sich diese Sedimente bis in Tiefen über 20 cm untersuchen,ohne daß vorher die darüberliegenden Sedimentschichten abgetragen werden mußten.Im Gegensatz zu den Kernen vor Namibia (Fahrt M 20/2, Abb. 6.17) konnten vorBrasilien so an zwei Stationen (GeoB 2104 und GeoB 2125) auch in einer Sedimenttiefevon 15 cm noch einzelne Magnetbakterien «10/50 1-11) nachgewiesen werden. Für dieseKerne liegen bisher nur Nitratprofile vor (M. ZASEL, unveröffentlicht). Die Eindringtiefevon Sauerstoff wurde aus den Nitratprofilen abgeschätzt und in der Tiefe angesetzt, inder Nitrat die maximale Konzentration erreichte.
Außer bei GeoB 2107 wurde bei allen Kernen mit lebenden Magnetbakterien vorBrasilien in der obersten Probe eine geringe Anzahl von Magnetbakterien angetroffen.Diese Bakterien sind vermutlich aerob. An den Stationen GeoB 2104, GeoB 2107 undwahrscheinlich auch GeoB 2105 erreichen die Magnetbakterien ihre höchste Dichte imanaeroben Sediment. Es ist anzunehmen, daß die Schichten größter Bakteriendichte inder Nitratreduktionszone liegen. Messungen der Konzentrationen gelösten Eisens imPorenwasser stehen noch aus. Insofern ist eine Einordnung der Tiefenverteilung in diegeochemische Zonierung derzeit nur bedingt möglich.
Nicht dargestellt sind in Abbildung 6.18 eine Reihe von Kernen, in denen nur einzelneMagnetbakterien (1-4 Individuen) nachgewiesen werden konnten (siehe Anhang B:Tabelle 1). Sie kamen ausschließlich in den obersten zwei Sedimentzentimetern vor,lebten also wie die Bakterien der Stationen GeoB 1702 und GeoB 2131 in der aerobenSchicht.
Nach diesen Befunden muß zwischen aeroben und (fakultativ) anaeroben Magnetbakterien unterschieden werden. Aerobe Magnetbakterien fanden sich in fast allenKernen, in denen überhaupt Magnetbakterien nachzuweisen waren. Ihre Konzentrationwar (mit Ausnahme von Kern GeoB 1703) immer kleiner 25 pro 50 1-11. AnaerobeMagnetbakterien traten nur in den nährstoffreichen Sedimenten auf. Dort erreichten sievielfach aber beträchtliche Konzentrationen (bis 30000/501-11).
Nach der Tiefenverteilung magnetotaktischer Bakterien sind zwei Gruppen vonKernen zu unterscheiden (Abbildung 6.19). In den Kernen der Gruppe A waren Magnetbakterien nur in den aeroben Schichten nachweisbar, in den Kernen der Gruppe Bwurden zusätzlich (fakultativ) anaerobe Magnetbakterien beobachtet, die dort auch diehöchsten Konzentrationen stellten. Große Zahlen (fakultativ) anaerober Magnetbakterien wurden ausschließlich in nährstoffreichen Sedimenten angetroffen. Der CorgGehalt betrug dann über 10 mg/cm3 , der Ntot-Gehalt über 1.5 mg/cm3.
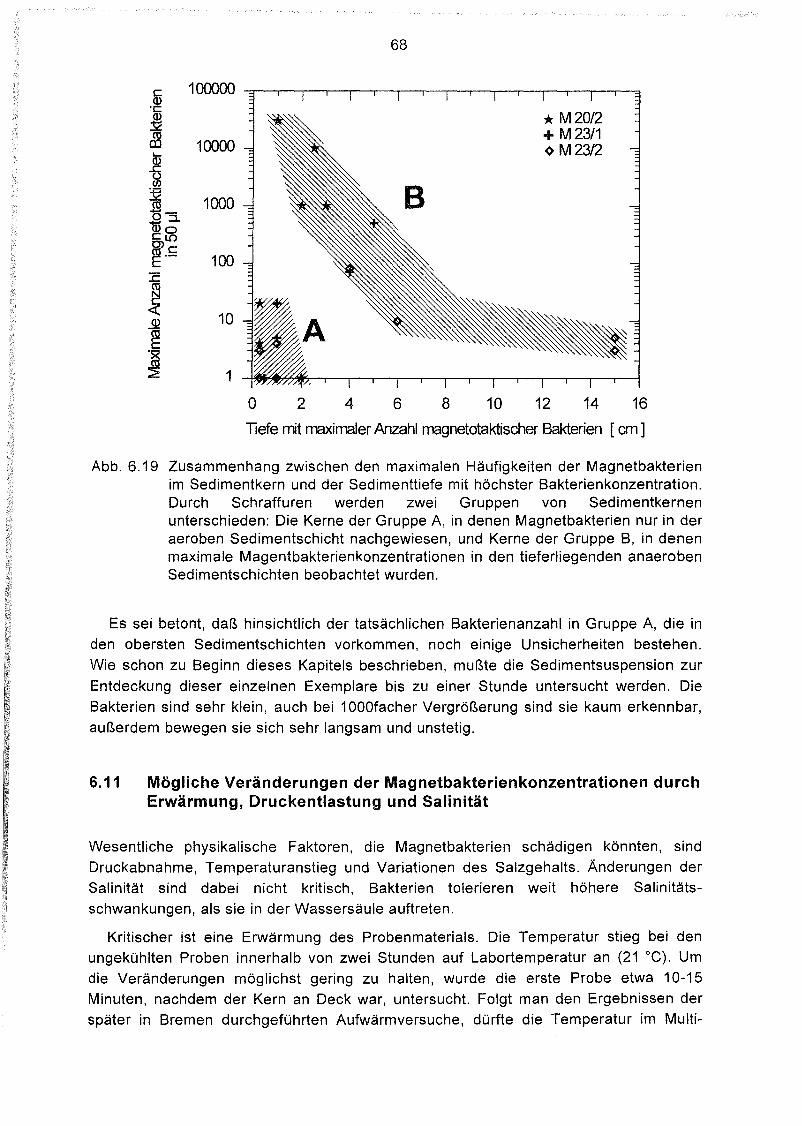
68
100000
10000
1000
100
10
1
o 2 4 6 8 10 12 14 16
liefe mit maximaler Anzahl magnetotaktischer Bakterien [cm]
Abb. 6.19 Zusammenhang zwischen den maximalen Häufigkeiten der Magnetbakterienim Sedimentkern und der Sedimenttiefe mit höchster Bakterienkonzentration.Durch Schraffuren werden zwei Gruppen von Sedimentkernenunterschieden: Die Kerne der Gruppe A, in denen Magnetbakterien nur in deraeroben Sedimentschicht nachgewiesen, und Kerne der Gruppe B, in denenmaximale Magentbakterienkonzentrationen in den tieferliegenden anaerobenSedimentschichten beobachtet wurden.
Es sei betont, daß hinsichtlich der tatsächlichen Bakterienanzahl in Gruppe A, die inden obersten Sedimentschichten vorkommen, noch einige Unsicherheiten bestehen.Wie schon zu Beginn dieses Kapitels beschrieben, mußte die Sedimentsuspension zur
Entdeckung dieser einzelnen Exemplare bis zu einer Stunde untersucht werden. DieBakterien sind sehr klein, auch bei 1000facher Vergrößerung sind sie kaum erkennbar,
außerdem bewegen sie sich sehr langsam und unstetig.
6.11 Mögliche Veränderungen der Magnetbakterienkonzentrationen durchErwärmung, Druckentlastung und Salinität
Wesentliche physikalische Faktoren, die Magnetbakterien schädigen könnten, sindDruckabnahme, Temperaturanstieg und Variationen des Salzgehalts. Änderungen derSalinität sind dabei nicht kritisch, Bakterien tolerieren weit höhere Salinitätsschwankungen, als sie in der Wassersäule auftreten.
Kritischer ist eine Erwärmung des Probenmaterials. Die Temperatur stieg bei denungekühlten Proben innerhalb von zwei Stunden auf Labortemperatur an (21 Oe). Um
die Veränderungen möglichst gering zu halten, wurde die erste Probe etwa 10-15Minuten, nachdem der Kern an Deck war, untersucht. Folgt man den Ergebnissen derspäter in Bremen durchgeführten Aufwärmversuche, dürfte die Temperatur im Multi-

69
corerrohr zum Zeitpunkt der ersten Probennahme kaum über 15 oe gelegen haben. Bei
den Expeditionen M 23/1 und M 23/2 wurden die Multicorerrohre im Labor gekühlt. Eine
Erwärmung bis auf 12-14 oe, maximal 16 oe konnte nicht vermieden werden. Diese
Erwärmung erfolgt hauptsächlich in der Zeit, in der der Kern durch die warmen
oberflächennahen Wasserschichten gezogen wird.
Bakterien der untersuchten Tiefseesedimente sind in ihrem Lebensraum nie Tempe
raturen über 4 oe ausgesetzt. Es ist anzunehmen, daß einige sich an die niedrigen
Temperaturen adaptiert haben, somit zu den psychrophilen Bakterien gehören.
Psychrophile Bakterien wachsen im allgemeinen bei Temperaturen bis 20 oe(SCHLEGEL, 1985). Wie eine Literaturrecherche zum Stichwort 'Psychrophilie' in allen
wesentlichen biologischen Zeitschriften von 1991 bis 1993 ergab, ist davon ausgehen,
daß praktisch alle Bakterien eine Erwärmung auf 15 oe überleben. Es wurde zwar
mehrfach von Bakterienkulturen mit Temperaturoptima bei 7 oe berichtet, dennoch
vermehrten sich auch diese Arten sich noch bei 13 oe (z.B.: ARAKI, 1992). Die meisten
psychrophilen Bakterien wachsen aber tatsächlich auch bei 20 oe (z.B.: Luo, 1992,
SHETTY et al., 1992, VISHNIAC & KURTZMAN, 1992, YANO et al., 1992). Die Tempera
turen, bei denen sie überleben, sollten höher liegen.
Die Lagerung und Untersuchung der Kerne bei Raumtemperatur erscheint daher
zunächst problemlos. Die Tatsache, daß die Magnetbakterien aus dem Südatlantik nur
in gekühltem Zustand überleben, ist in diesem Zusammenhang erwähnenswert, sie
scheinen 'stark' psychrophil zu sein und lassen sind bei Temperaturen um 20 oe nach
wenigen Tagen nicht mehr nachzuweisen.
Ein gewisser Einfluß der Temperaturerhöhung auf die Magnetbakterienzahlen ist
auch bei sofortiger Beobachtung nicht auszuschließen. Die Magnetbakterienzahlen in
den Proben der Fahrt M 23/2 sind bei gleichen 02-Flüssen leicht erhöht gegenüber den
Werten von Fahrt M 20/2 (Abb. 6.13). Nur bei der erstgenannten Fahrt wurden Probe
und Objektträger während der Beobachtung gekühlt.
Über die Druckempfindlichkeit magnetotaktischer Bakterien gibt es bisher keine
Untersuchungen. Allgemein lassen sich Bakterien nach ihrer Druckempfindlichkeit in
folgende Gruppen unterteilen (JANNASCH & TAYLOR, 1984; DEMING, 1986; YAYANOS,
1986):
- barosensitive Bakterien:optimales Wachstum bei Athmospärendruck (1 atm), kein Wachstum bei Drückenüber 200 atm möglich,
- barotolerante Bakterien:optimales Wachstum bei Athmospärendruck (1 atm), Wachstum bei zu Drücken von400 bis 600 atm und darüber möglich,
- barophile (druckliebende) Bakterien:optimales Wachstum bei Druck über 1 atm,
- obligat barophile Bakterien:kein Wachstum bei Athmosphärendruck, Wachstum nur unter höheren Drücken.

70
Verschiedene Autoren schätzen den Anteil barophiler Bakterien in der Tiefsee unterschiedlich ein (JANNASCH & TAYLOR, 1984; COLWELL, 1986; TURLEY & LOCHTE 1990).Einige Dutzend Kulturen barophiler Bakterien (YAYANOS, 1986), aber nur sehr wenigeKulturen obligat barophiler Bakerien (z.B.: YAYANOS & DIETZ, 1983) liegen in Kultur vor.Dabei ist zu beachten, daß keine Untersuchung gefunden wurde, bei denen dieBakterien permanent unter in situ-Druck gehalten wurden, was zu einer Verfälschungder Versuchsergebnisse führen kann.
Ein möglicher Grund für das Absterben barophiler Bakterien unter Normaldruck wirdvon DELONG und YAYANOS (1985, 1986) angegeben. Die Zellmembran wird bei Temperaturerniedrigung und in geringerem Maße auch bei Druckerhöhung weniger fließfähig.Barophile Bakterien ersetzen daher die membranbildenden gesättigten Phospholipidedurch mehrfach ungesättigte Phospholipide. Diese ungesättigten Fette sind fließfähiger.Unter Normaldruck und Normaltemperatur ist die Zellmembran dieser Bakterienoffensichtlich nicht mehr stabil genug.
In der Arbeit von YAYANOS (1986) wird festgestellt, daß barophile Bakterien oft auchpsychrophil sind und daß eine Dekompression bei gleichzeitiger Temperaturerhöhungdie Bakterien viel stärker schädigt. Deshalb führt er den Terminus 'obligat barophil beieiner Temperatur von ... ' ein. In dieser Arbeit wird gezeigt, daß Bakterien mit zunehmender Tiefe empfindlicher gegen Druckentlastung werden. Die erste Kultur, die sich auchbei niedrigen Temperaturen nicht unter Athmosphärendruck vermehrte, stammte auseiner Tiefe von 6350 m, jedoch mußten dabei alle diese Bakterien für etwa 15-30Minuten auf einen Druck von 1 atm dekomprimiert werden.
Bei YAYANOS & DIETZ (1982) wird angegeben, mit welchen Raten ein barophilerBakterienstamm aus 5700 m Tiefe, in Abhängigkeit von Druckerniedrigung undTemperaturerhöhung die Fähigkeit verliert, Kolonien zu bilden. Berechnet man in einemAnalogieschluß, daß eventuell existierende barophile Magnetbakterien mit denselbenRaten auch ihre Fähigkeit verlieren, aktiv zu schwimmen, so müßten bei den gekühltenProben (Kap. 6.1) mehr als 80 % der Bakterien nachweisbar sein. Die Magnetbakterienmüssen nach diesen Berechnungen bei Druckentlastung ihre Schwimmfähigkeit früherverlieren, als der angeführte barophile Bakterienstamm die Fähigkeit verliert Kolonien zubilden. Im anderen Falle sind die vorgefundenen Konzentrationen (:s; 10 Individuen proml) magnetotaktischer Bakterien in den Tiefseesedimenten real.
Der in Abbildung 6.13 dargestellte Zusammenhang zwischen der Magnetbakterienkonzentration und dem in situ gemessenen diffusiven Sauerstoffeintrag zeigt, daß dieMagnetbakterienzahlen für unterschiedliche Gebiete und gleiche Wassertiefen unterschiedlich sind. Dies gilt auch im Tiefenbereich bis 2000 m, in denen nach TARDENT(1979) und YAYANOS (1986) Barophilie bei Bakterien keine Rolle spielt. Innerhalb dereinzelnen untersuchten Gebiete nehmen die Bakterienkonzentrationen mit zunehmenderTiefe auch bis 2000 m Tiefe deutlich ab (Abb. 6.2 - 6.4).

71
7 Extrak~ion und elektronenmikroskopische Untersuchllrl{.Jmagnetischer Partikel aus Oberflächensedimenten des Südatlantiks
Elektronenmikroskopische Untersuchungen erlauben es, fossile Magnetosome anhand
ihrer Morphologie eindeutig zu identifizieren, Mit dieser Methode gelang PETERSEN etal. (1986) im Angola Becken der erste Nachweis von Magnetosomen in Tiefseesedimenten. Bei weiteren Untersuchungen (u.a. VALI et al., 1989; YAMAZAKI et al.,1991) wurden Magnetofossilien mit Hilfe des Elektronenmikroskops auch in anderen
pelagischen Sedimenten gefunden.
In dieser Arbeit sind Oberflächensedimente der Expeditionen M 16/2 und M 20/2sowie in geringerem Umfang auch Proben aus größerer Sedimenttiefe magnetischextrahiert und die Extrakte im Transmissions-Elektronenmikroskop (TEM) untersuchtworden. Dabei wurde in erster Linie überprüft, ob Magnetosome auch in denSedimenten vorkommen, in denen keine lebenden Magnetbakterien nachweisbarwaren. Im Gegensatz zu üblichen gesteinsmagnetischen Untersuchungen lassen sichmit dieser Methode fossile Magnetosome eindeutig von anderen magnetischenPartikeln unterscheiden. Allerdings ist die Präparation relativ aufwendig. Nur äußerstgeringe Probenmengen, die nicht notwendigerweise repräsentativ sind, könnenuntersucht werden.
7.1 Extraktionsverfahren
Das Extraktionsverfahren wird im folgenden detailliert beschrieben, da der Erfolg derExtraktion wesentlich von ihrer sorgfältigen Durchführung abhängt.
Drei Kubikzentimeter der zu untersuchenden Probe werden mit 200 ml Aqua bidestin ein Becherglas gegeben, zerstoßen und mit einem Hochleistungs-Ultraschallgerätdispergiert. Vorversuche ergaben, daß bei einer abgestrahlten Schalleistung von 50Watt die Magnetosomketten von den anderen Partikeln gelöst werden, ihreKettenstruktur aber noch erhalten bleibt. Die Zugabe einer Spatelspitze des
Dispergierungsmittels Na4(P04h·10H20 verhindert ein Ausflocken insbesondere derTonfraktion .
Die Suspension wird in einen Scheidetrichter gefüllt, mittels einer Schlauchpumpeumgewälzt und dabei an einem Magneten (Abb. 7.1) vorbeigeführt. Um ein gravitativesAbsetzen der magnetischen Partikel zu vermeiden, wird die Suspension im Scheidetrichter mit Aqua bidest auf einen Liter verdünnt und durch Luftzufuhr ständig inBewegung gehalten. Die Umwälzung der Suspension erfolgt möglichst langsam, damitdie magnetischen Teilchen lange im Einflußbereich des Magneten bleiben. Dadurcherhöht sich die Menge an extrahiertem Material. Der verwendete Magnet aus KobaltSamarium-Legierung erzeugt ein Magnetfeld bis zu 300 mT. Um den lokalenFeldgradienten zu erhöhen, ist auf dem Magneten ein Finger aus weichmagnetischemEisen aufgesetzt, der in einer Teflonhülse steckt. Nach einer Extraktionsdauer von
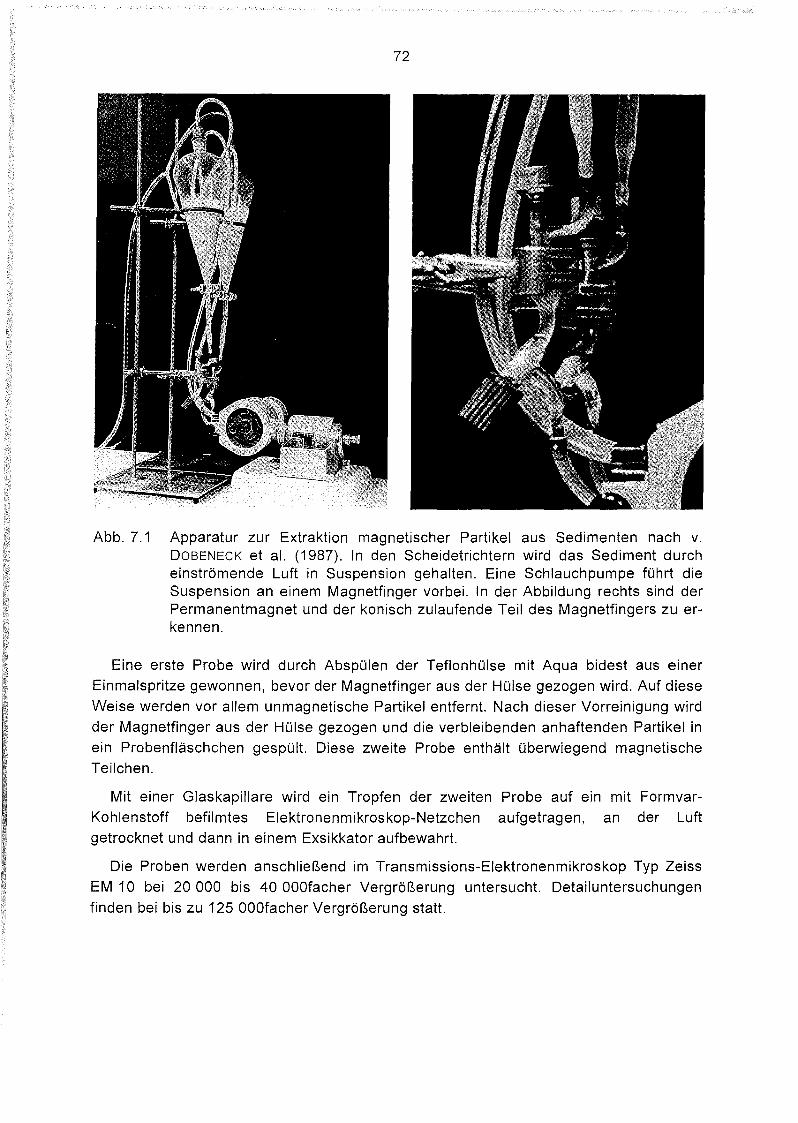
72
Abb. 7.1 Apparatur zur Extraktion magnetischer Partikel aus Sedimenten nach v.DOBENECK et al. (1987). In den Scheidetrichtern wird das Sediment durcheinströmende Luft in Suspension gehalten. Eine Schlauchpumpe führt dieSuspension an einem Magnetfinger vorbei. In der Abbildung rechts sind derPermanentmagnet und der konisch zulaufende Teil des Magnetfingers zu erkennen.
Eine erste Probe wird durch Abspülen der Teflonhülse mit Aqua bidest aus einerEinmalspritze gewonnen, bevor der Magnetfinger aus der Hülse gezogen wird. Auf dieseWeise werden vor allem unmagnetische Partikel entfernt. Nach dieser Vorreinigung wirdder Magnetfinger aus der Hülse gezogen und die verbleibenden anhaftenden Partikel inein Probenfläschchen gespült. Diese zweite Probe enthält überwiegend magnetischeTeilchen.
Mit einer Glaskapillare wird ein Tropfen der zweiten Probe auf ein mit FormvarKohlenstoff befilmtes Elektronenmikroskop-Netzchen aufgetragen, an der Luftgetrocknet und dann in einem Exsikkator aufbewahrt.
Die Proben werden anschließend im Transmissions-Elektronenmikroskop Typ ZeissEM 10 bei 20 000 bis 40 OOOfacher Vergrößerung untersucht. Detailuntersuchungenfinden bei bis zu 125 OOOfacher Vergrößerung statt.

73
7.2 Qualitative und quantitative Beurteilung der Extraktionsmethode
Das Extraktionsverfahren soll den Gehalt an Magnetosomen in den Sedimentenmöglichst quantitativ erfassen. Dazu muß bei der Extraktion verschiedener Sedimentproben mit derselben Apparatur zunächst vor allem eine Kontamination vermiedenwerden. Da die Magnetosome extrem klein sind (50 - 130 nm), wurde nach jederExtraktion sehr großer Wert gelegt auf die penible Reinigung aller benutzten Komponenten. Im Laufe der Arbeit wurden zweimal Kontrolluntersuchungen durchgeführt, beidenen jeweils Aqua bidest ohne Sediment umgewälzt wurde. Nach drei Tagen warkeinerlei Trübung des Wassers durch Schmutzpartikel festzustellen, und bei der elektronenmikroskopischen Untersuchung wurden keine Magnetosome nachgewiesen.Dennoch ist zwar nicht auszuschließen, daß Magnetosome zwischen verschiedenenExtraktionen verschleppt wurden, ihre Konzentration bleibt aber in jedem Fall sehrgering.
Wesentlichen Einfluß auf das Ergebnis der Extraktion haben die einzelnenArbeitsschritte, sie sollen daher im folgenden näher beschrieben werden:
Bei einer optimalen Schalleistung (Abstrahlleistung) des Ultraschallgeräts von 50Watt bleiben vorhandene Magnetosomketten weitgehend erhalten. Intakte Kettenlassen sich bei der Durchsicht des TEM-Netzchens wesentlich leichter erkennen alsCluster oder einzelne Magnetosome. Bei niedrigerer Ultraschallenergie, lösen sich nichtalle Magnetosome von den Sedimentpartikeln, an denen sie haften.
Generell bleiben Ketten prismatischer Magnetosome (Abb. 7.4) besser erhalten als
Ketten oktaedrischer Magnetosome (Abb. 7.3). Projektil- und karottenförmigeMagnetosome liegen außerhalb der Bakterienzelle nicht in Ketten vor. Energetisch istes günstiger, wenn die Magnetosome nicht in Ketten vorliegen, sondern sich antiparallel ausrichten. Das Aufbrechen der Ketten erfordert jedoch eine Aktivierungsenergie. Diese ist bei den prismatischen Magnetosomen am höchsten.
Die Kraft, die ein Magnetfeld auf magnetische Mineralkörner ausübt, ist abhängigvom Magnetfeldgradienten und dem magnetischen Moment des Partikels. Dasmagnetische Moment ist bei kleinen Teilchen proportional zum Volumen (0::: r3). Dieviskose Reibung des Wassers bremst die suspendierten Teilchen ab, während sie vom
Magnetfinger angezogen werden. Bei laminarer Strömung ist die Reibungskraft auf einkugelförmiges Teilchen nach Stokes proportional zum Teilchenradius. In derExtraktionsapparatur stellt sich bei langsamen Umwälzgeschwindigkeiten laminareStrömung ein. Die Reynoldszahl für die gegebene Geometrie liegt unterhalb deskritischen Wertes, bei dem die laminare in eine turbulente Strömung umschlägt. Da diemagnetische Kraft mit r3, die entgegenstehende viskose Kraft dagegen mit r1 ansteigt,werden größere Teilchen bevorzugt extrahiert. Untersuchungen an extrahiertemMagnetit bezogen sich daher bisher meist auf größere Kornklassen (u.a. CANFIELD &
BERNER, 1987). Ist die Umwälzgeschwindigkeit gering, so bleiben die magnetischenPartikel relativ lange im Bereich der Anziehung durch den Magneten und auch kleinereTeilchen werden effektiv extrahiert. Die Umwälzgeschwindigkeit darf jedoch einenbestimmten Wert nicht unterschreiten, da sich sonst in den Ecken der Apparatur zuviel

74
Sediment absetzt. Im Verlauf der Extraktion wurden solche Bodensätze immer wiederaufgewirbelt.
Wenn magnetische Minerale sich durch gegenseitige Anziehung zu Aggregatenzusammenlagern, lassen sie sich leichter aus der Suspension extrahieren. Einzelneund an nichtmagnetische Partikel gelagerte Magnetosome werden dagegen wenigervollständig erfaßt. Die Ausbeute dieser Teilchen kann durch Extraktion über mehr alsdrei Tage ein wenig erhöht werden. Aus Zeitgründen wurde die Dauer aber auf drei bisvier Tage beschränkt. Daher bestimmt der Grad der Dispergierung der Magnetominerale mit das Ergebnis der Extraktion. Die zugegebene Menge des Dispergierungsmittels wirkt je nach Zusammensetzung des Sediments (tonig bis karbonatisch)unterschiedlich.
Das Abspülen der Hülse erfolgt mit Aqua bidest aus einer Einmalspritze. Da sich dieWirkung des Wasserstrahis nur schlecht dosieren und reproduzieren läßt, geht einunterschiedlich großer Anteil an Magnetosomen bei der Vorreinigung verloren. Dieser
Anteil ist abhängig von der Art des Sediments, speziell von der Menge unmagnetischerMatrix, die an der Hülse haftet.
Es kann nur ein einziger Tropfen der Extraktion auf die 3 mm großen TEM-Netzchenaufgetragen werden. Er wurde mit einer dünnen Kapillare aus dem Probenfläschchenentnommen. Die Verteilung der magnetischen Partikel im Fläschchen ist aber, wieschon optisch an schwarzen Konglomeraten magnetischer Teilchen erkennbar, nichthomogen. Zwar wurde versucht, möglichst gleiche Probenmengen zu erhalten - derTropfen wurde immer zum selben Zeitpunkt nach Befüllen des Fläschchens in gleicherHöhe über dem Gefäßboden entnommen - doch wird durch die ungleichmäßigeVerteilung der Partikel eine quantitative Bestimmung der Konzentration biogenen
Magnetits insgesamt wohl am stärksten beeinträchtigt.
7.3 Elektronenmikroskopische Untersuchung
Eine chemische Behandlung der Proben zur Kontrastierung mußte nicht vorgenommenwerden, da die Magnetosome durch die hohe Kernladungszahl des Eisens bereits sehrkontrastreich sind.
Abbildung 7.2 zeigt eine TEM-Aufnahme von Magnetosomen unterschiedlicherMorphologie und dasselbe Bild nach Bearbeitung mit einem Gradientenfilter, der dieKanten betont. Helligkeitsunterschiede der Magnetosome haben zwei Ursachen: DieDicke, die bestimmt, wieviele der Elektronen absorbiert werden, und die Ausrichtungdes Kristalls, die maßgeblich dafür ist, wieviele Elektronen durch Bragg-Streuungverloren gehen.
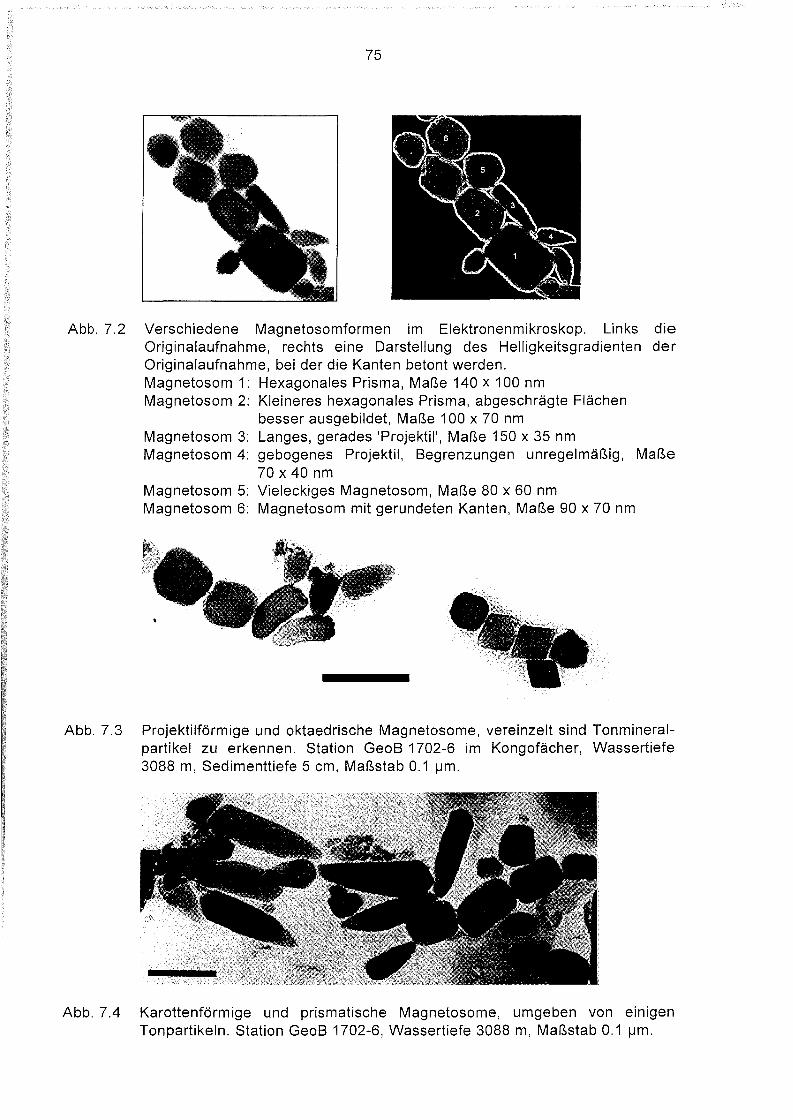
75
Abb. 7.2 Verschiedene Magnetosomformen im Elektronenmikroskop. Links dieOriginalaufnahme, rechts eine Darstellung des Helligkeitsgradienten derOriginalaufnahme, bei der die Kanten betont werden.Magnetosom 1: Hexagonales Prisma, Maße 140 x 100 nmMagnetosom 2: Kleineres hexagonales Prisma, abgeschrägte Flächen
besser ausgebildet, Maße 100 x 70 nmMagnetosom 3: Langes, gerades 'Projektil', Maße 150 x 35 nmMagnetosom 4: gebogenes Projektil, Begrenzungen unregelmäßig, Maße
70 x 40 nmMagnetosom 5: Vieleckiges Magnetosom, Maße 80 x 60 nmMagnetosom 6: Magnetosom mit gerundeten Kanten, Maße 90 x 70 nm
Abb. 7.3 Projektilförmige und oktaedrische Magnetosome, vereinzelt sind Tonmineralpartikel zu erkennen. Station GeoB 1702-6 im Kongofächer, Wassertiefe3088 m, Sedimenttiefe 5 cm, Maßstab 0.1 I-lm.
Abb. 7.4 Karottenförmige und prismatische Magnetosome, umgeben von einigenTonpartikeln. Station GeoB 1702-6, Wassertiefe 3088 m, Maßstab 0.1 I-lm.

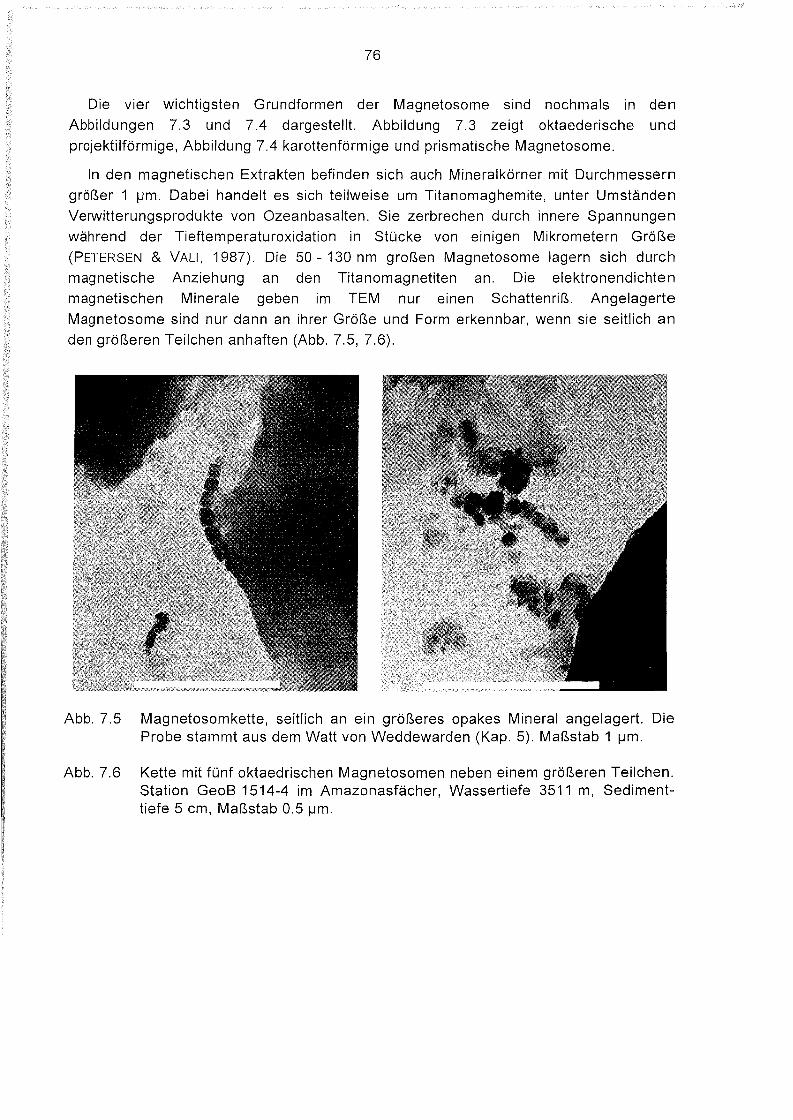
76
Die vier wichtigsten Grundformen der Magnetosome sind nochmals in den
Abbildungen 7.3 und 7.4 dargestellt. Abbildung 7.3 zeigt oktaederische und
projektilförmige, Abbildung 7.4 karottenförmige und prismatische Magnetosome.
In den magnetischen Extrakten befinden sich auch Mineralkörner mit Durchmessern
größer 1 ~m. Dabei handelt es sich teilweise um Titanomaghemite, unter Umständen
Verwitterungsprodukte von Ozeanbasalten. Sie zerbrechen durch innere Spannungen
während der Tieftemperaturoxidation in Stücke von einigen Mikrometern Größe
(PETERSEN & VALI, 1987). Die 50 - 130 nm großen Magnetosome lagern sich durch
magnetische Anziehung an den Titanomagnetiten an. Die elektronendichten
magnetischen Minerale geben im TEM nur einen Schattenriß. Angelagerte
Magnetosome sind nur dann an ihrer Größe und Form erkennbar, wenn sie seitlich an
den größeren Teilchen anhaften (Abb. 7.5, 7.6).
Abb. 7.5 Magnetosomkette, seitlich an ein größeres opakes Mineral angelagert. DieProbe stammt aus dem Watt von Weddewarden (Kap. 5). Maßstab 1 ~m.
Abb. 7.6 Kette mit fünf oktaedrischen Magnetosomen neben einem größeren Teilchen.Station GeoB 1514-4 im Amazonasfächer, Wassertiefe 3511 m, Sedimenttiefe 5 cm, Maßstab 0.5 ~m.

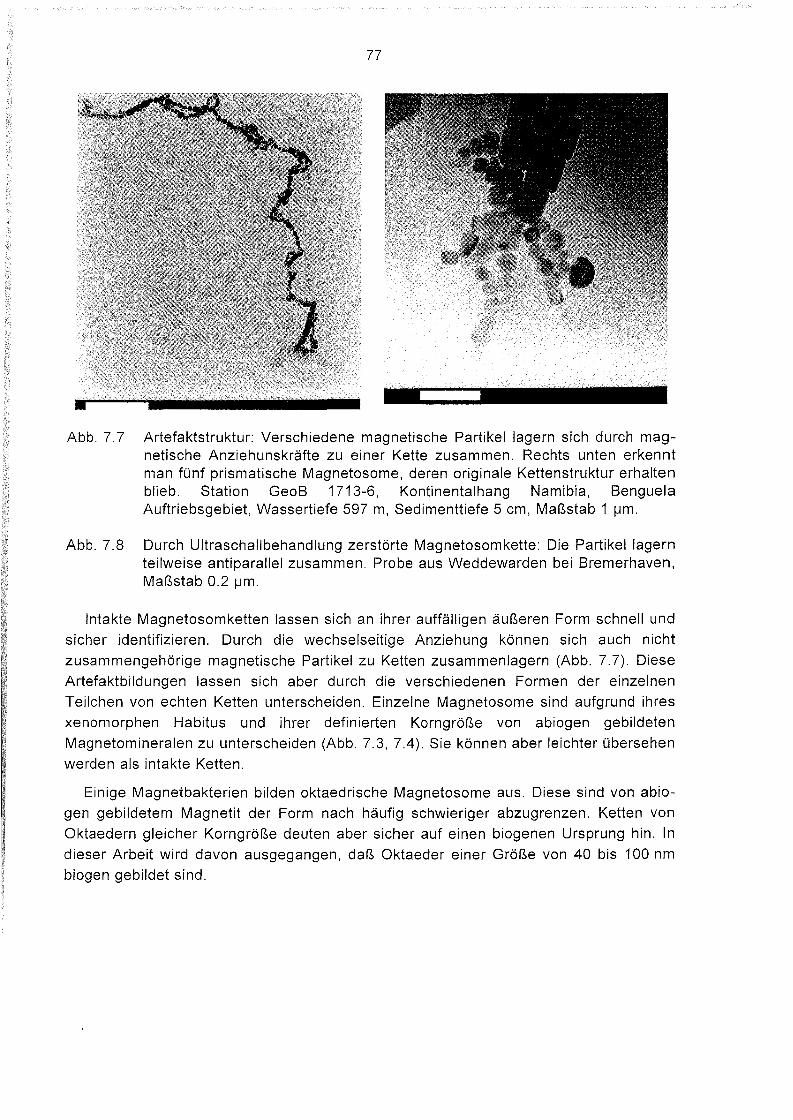
77
Abb. 7.7 Artefaktstruktur: Verschiedene magnetische Partikel lagern sich durch magnetische Anziehunskräfte zu einer Kette zusammen. Rechts unten erkenntman fünf prismatische Magnetosome, deren originale Kettenstruktur erhaltenblieb. Station GeoB 1713-6, Kontinentalhang Namibia, BenguelaAuftriebsgebiet, Wassertiefe 597 m, Sedimenttiefe 5 cm, Maßstab 1 ~m.
Abb. 7.8 Durch Ultraschallbehandlung zerstörte Magnetosomkette: Die Partikel lagernteilweise antiparallel zusammen. Probe aus Weddewarden bei Bremerhaven,Maßstab 0.2 IJm.
Intakte Magnetosomketten lassen sich an ihrer auffälligen äußeren Form schnell und
sicher identifizieren. Durch die wechselseitige Anziehung können sich auch nicht
zusammengehörige magnetische Partikel zu Ketten zusammenlagern (Abb. 7.7). Diese
Artefaktbildungen lassen sich aber durch die verschiedenen Formen der einzelnenTeilchen von echten Ketten unterscheiden. Einzelne Magnetosome sind aufgrund ihres
xenomorphen Habitus und ihrer definierten Korngröße von abiogen gebildeten
Magnetomineralen zu unterscheiden (Abb. 7.3, 7.4). Sie können aber leichter übersehenwerden als intakte Ketten.
Einige Magnetbakterien bilden oktaedrische Magnetosome aus. Diese sind von abiogen gebildetem Magnetit der Form nach häufig schwieriger abzugrenzen. Ketten von
Oktaedern gleicher Korngröße deuten aber sicher auf einen biogenen Ursprung hin. In
dieser Arbeit wird davon ausgegangen, daß Oktaeder einer Größe von 40 bis 100 nmbiogen gebildet sind.

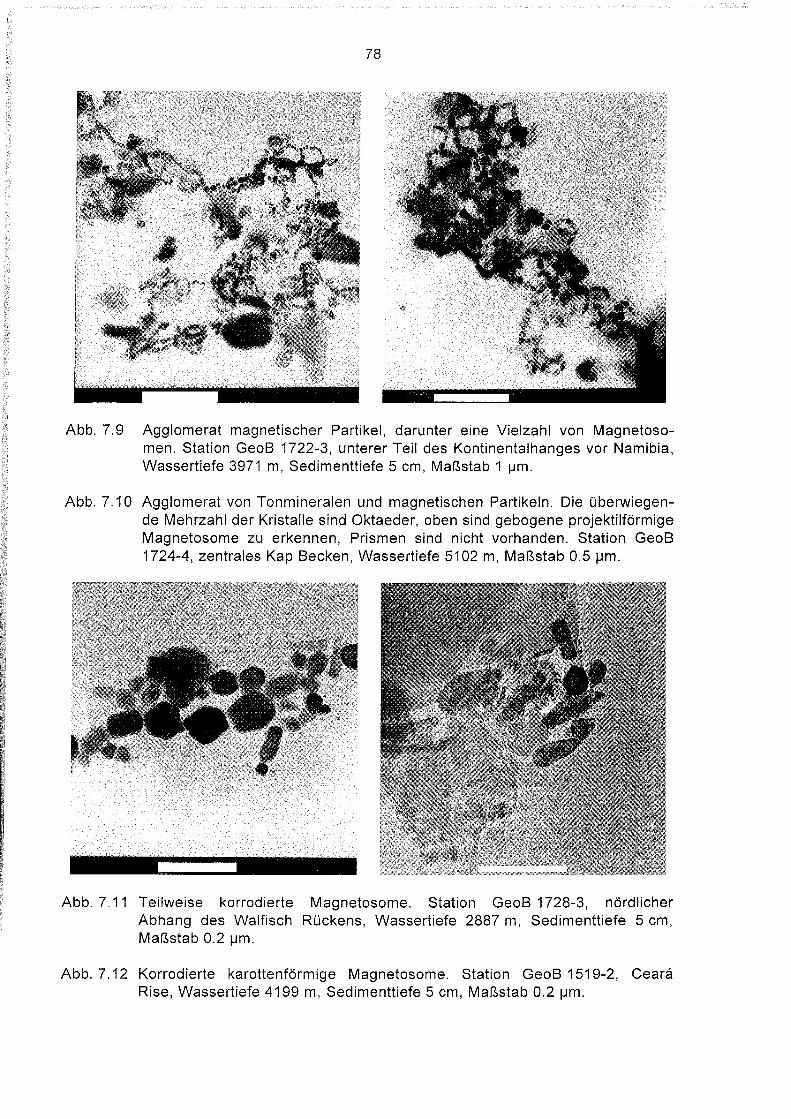
78
Abb. 7.9 Agglomerat magnetischer Partikel, darunter eine Vielzahl von Magnetosomen. Station GeoB 1722-3, unterer Teil des Kontinentalhanges vor Namibia,Wassertiefe 3971 m, Sedimenttiefe 5 cm, Maßstab 1 I-lm.
Abb. 7.10 Agglomerat von Tonmineralen und magnetischen Partikeln. Die überwiegende Mehrzahl der Kristalle sind Oktaeder, oben sind gebogene projektilförmigeMagnetosome zu erkennen, Prismen sind nicht vorhanden. Station GeoB1724-4, zentrales Kap Becken, Wassertiefe 5102 m, Maßstab 0.5 I-lm.
Abb. 7.11 Teilweise korrodierte Magnetosome. Station GeoB 1728-3, nördlicherAbhang des Walfisch Rückens, Wassertiefe 2887 m, Sedimenttiefe 5 cm,Maßstab 0.2 I-lm.
Abb. 7.12 Korrodierte karottenförmige Magnetosome. Station GeoB 1519-2, CearaRise, Wassertiefe 4199 m, Sedimenttiefe 5 cm, Maßstab 0.2 I-lm.


79
Die äußere Form der Magnetosome und damit ihr Erkennungsmerkmal kann durchKorrosion verändert sein. VALI & KIRSCHVINK (1989) berichten über dieses Phänomen
für miozäne Proben aus dem Angola Becken (Sedimenttiefe 25 - 50 m). Abbildung 7.11zeigt leicht korrodierte prismatische Magnetosome, Abbildung 7.12 stärker korrodierte
karottenförmige Magnetosome. In den Sedimenten sind auch abgerundete magnetischePartikel nachweisbar, die sich nicht eindeutig als Magnetosome klassifizieren lassen.Zur chemischen Stabilität von Magnetosomen siehe Kapitel 7.6.
Die vergleichende quantitative Untersuchung von Proben verschiedener Stationenim TEM wird durch die geringe Größe der Magnetosome und die Qualität derverwendeten TEM-Netzchen erschwert. Das Netzchen besitzt eine untersuchbare
Fläche von etwa 2 x 2 mm. Dies entspricht bei 20 OOOfacher Vergrößerung einem Feldvon 40 x 40 m; ein Magnetosom besitzt in diesem Maßstab eine Größe von 1 - 2 mm.Da sich die magnetischen Teilchen gegenseitig anziehen, sind sie in der Regel sehrinhomogen verteilt.
Um qualitativ gleichwertige Netzchen zu verwenden, wurde auf fertig befilmteNetzchen zurückgegriffen. Die Folie besteht aus Formvar, das mit Kohlenstoffbeschichtet ist, um statische Aufladungen im TEM zu vermeiden. Leider war die
Qualität dieser Netzchen nicht immer zufriedenstellend. Bei der Untersuchung platztedie Folie oftmals auf, so daß diese Stellen nicht mehr untersucht werden konnten. DieNetzchen waren auch gelegentlich mit einer für Elektronen undurchlässigen Schichtverschmutzt, deren Herkunft nicht geklärt werden konnte. Eigene Bemühungen,Netzchen herzustellen, waren nicht erfolgreicher. So konnten nicht alle Probengleichmäßig untersucht werden.
Zusammenfassend ist festzustellen, daß Magnetosome mit der beschriebenenMethode sicher identifiziert werden können, es aber erhebliche Schwierigkeitenbereitet, die Konzentration bakteriellen Magnetits in den untersuchten Sedimenten
genauer zu bestimmen.
Insgesamt sind die Magnetosomkonzentrationen im östlichen Südatlantik (FahrtM 20/2) höher als an den Stationen im äquatorialen Westatlantik (M 16/2). Auffällig ist,daß hohe Magnetosomzahlen auch in großen Wassertiefen angetroffen wurden (Abb.7.9, 7.10), an denen keine lebenden Magnetbakterien nachweisbar waren.
7.4 Elementanalyse im Elektronenmikroskop
Um zur Identifikation der extrahierten Partikel nicht nur auf die Form des Schattenrisses·im TEM-Bild angewiesen zu sein, wurden am Elektronenmikroskop Zeiss EM 420(Fachbereich Physik der Universität Bremen) EDAX-Analysen der untersuchtenMinerale vorgenommen. Dabei wird die untersuchte Probe durch den Elektronenstrahldes Elektronenmikroskops teilweise ionisiert. Elektronen werden vorzugsweise aus derinnersten, der K-Schale herausgeschlagen. Den freiwerdenden Platz besetzenElektronen höherer Schalen. Die Energie, die beim Übergang aus höheren in die
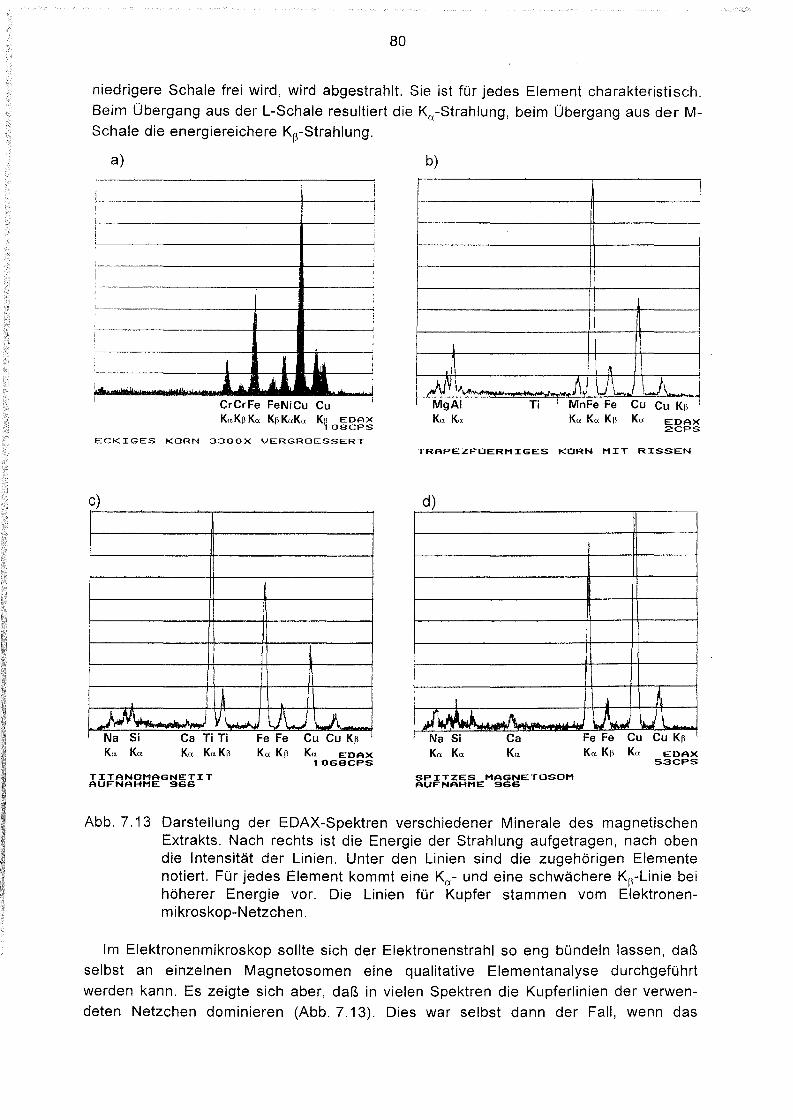
80
niedrigere Schale frei wird, wird abgestrahlt. Sie ist für jedes Element charakteristisch.
Beim Übergang aus der L-Schale resultiert die Ku-Strahlung, beim Übergang aus der M
Schale die energiereichere Kß-Strahlung.
a) b)
TRAPEZFOERMIGES KORN MIT RISSEN
MnFe Fe Cu Cu KrJKu Ku KrJ K" EDAX
2CPS
--.-_ .._--
I
II I
,n i A
I ~'Vl\~, PIJ \j\_1 \J,I JCrCrFe FeNiCu Cu
KuKpKu K~KuK" Ku EDAX10acPS
ECKIGES KORN 3300X VERGROESSERT
I I
! IIi j
!
i:, ,,
i
i ,,i
i i I,
." .I " J i
c) d)
Fe Fe ß
Ku Kß K" EDAXS3CPS
Na S,Ku Ku
I I I .I .~J ...~ ... J,. 1\ J~ U l_
I Cu Cu KFe Fe Cu Cu KJlKu Kß Ku EDAX
1068CPS
Ca TI TIKu K"Kp
II
I
I ~-J\w'V~ .,J V~ JlJ .A..
TITANOMAGNETITAUFNAHME 966
SPITZES MAGNETOSOMAUFNAHME 966
Abb. 7.13 Darstellung der EDAX-Spektren verschiedener Minerale des magnetischenExtrakts. Nach rechts ist die Energie der Strahlung aufgetragen, nach obendie Intensität der Linien. Unter den Linien sind die zugehörigen Elementenotiert. Für jedes Element kommt eine Ku- und eine schwächere Kß-Linie beihöherer Energie vor. Die Linien für Kupfer stammen vom Elektronenmikroskop-Netzchen.
Im Elektronenmikroskop sollte sich der Elektronenstrahl so eng bündeln lassen, daß
selbst an einzelnen Magnetosomen eine qualitative Elementanalyse durchgeführt
werden kann. Es zeigte sich aber, daß in vielen Spektren die Kupferlinien der verwen
deten Netzchen dominieren (Abb. 7.13). Dies war selbst dann der Fall, wenn das

81
analysierte Mineral in der Mitte einer Masche des Netzchens lag und somit einenAbstand von etwa 50 IJm zum Kupfersteg hatte. Die Abstrahlcharakteristik scheint nichtso gut zu sein, wie zunächst angenommen. Daher erhält man immer eine Information,die über die Umgebung des Teilchens integriert. So läßt sich auch der in den Spektren
enthaltene 'Untergrund' von Na-, Mg-, AI-, Si- und Ca-Linien erklären, der durch
Tonminerale entsteht.
Die beiden ersten Spektren in Abbildung 7.13 zeigen, daß die magnetische Mineralfraktionen im Sediment im Detail sehr heterogen sind. Das untersuchte Mineral inAbbildung 7.13a ist eine Eisen-Chrom-Nickelverbindung, das Mineral in Abbildung7.13b enthält neben Eisen geringe Anteile von Mangan. Über die Herkunft dieserbeiden Minerale kann nur spekuliert werden. Sie sind in keinem Fall repräsentativ. Dasdritte Spektrum (Abb. 7.13c) stammt von einem Titanomagnetit. Dieses Mineral isttypisch für ozeanische Basalte. Das vierte Spektrum (Abb. 7.13d) stammt von einemMagnetosom. Es enthält keine nachweisbaren Spuren von Titan.
Die EDAX-Analyse des benutzten Elektonenmikroskops kann nur Elemente nachweisen, die schwerer sind als Sauerstoff. Titanbeimischungen und Eisensulfide lassensich damit sicher identifizieren. Dagegen läßt sich Magnetit aber nicht von anderen
Eisenoxiden oder -hydroxiden unterscheiden. Diese Verbindungen tragen aber einewesentlich geringere Magnetisierung als Magnetit und können sehr schlecht extrahiertwerden. Daher ist anzunehmen, daß das Magnetosom in Abbildung 7.13d aus Magnetitbesteht. Über den Oxidationszustand lassen sich keine Aussagen treffen; Magnetit
enthält Fe(") und Fe("I) und kann zu Maghemit (y-Fe20 3) oxidiert werden.SCHMIDBAUER und PETERSEN (pers. Mitteilung, 1992) haben Mößbauer Untersuchungenan Magnetosomen durchgeführt. Dabei wandelte sich bei Lagerung an Luft der
gesamte Magnetit innerhalb von 14 Tagen in Maghemit um.
7.5 Verteilung verschiedener Magnetosomformen in den Sedimentendes Südatlantiks
Aus den zuvor ausführlich dargelegten Gründen kann mit dem Elektronenmikroskop dieExistenz von Magnetosomen im Sediment nachgewiesen, ihre Konzentration aber nurschwer quantifiziert werden. Im Gegensatz dazu läßt sich die relative Häufigkeitverschiedener Magnetosomformen recht einfach bestimmen. Dazu wurden dieentwickelten TEM-Negative in einem Mikrofiche Lesegerät durchgesehen. Diese fürGesteinsschliffe übliche Methode erlaubt es, einen großen Bildausschnitt mit ausreichender Vergrößerung zu untersuchen. Die Magnetosome wurden nach folgendenvier Formen unterschieden:
- Oktaeder (Abb. 7.3),
- Projektile (Abb. 7.3) und Karotten (Abb. 7.4),
- Prismen (Abb. 7.4),
- abgerundete Formen (Abb. 7.3).
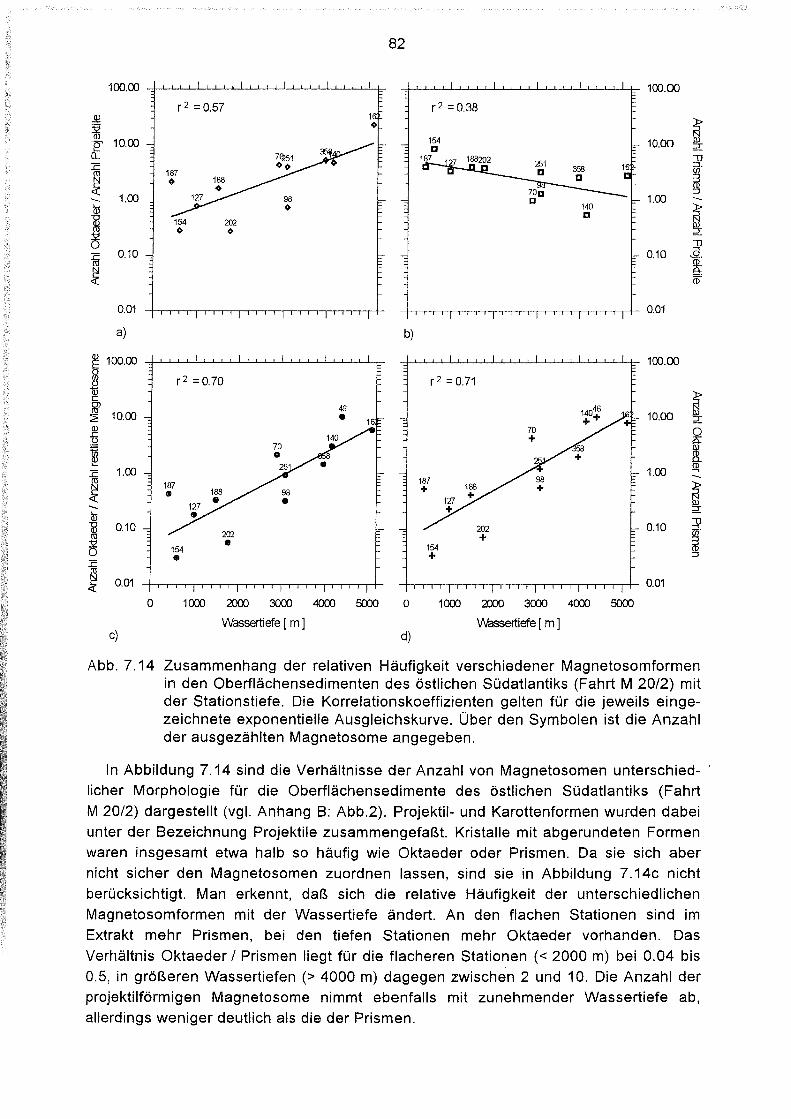
82
0.10
10.00
1.00
-=t-'-L..J-L..J-L..J-L..J-L..J-L..J-L..J-L..J-LJ.-LJ.-'--'--'--'-+ 100.00
187~
0.01
r 2 =0.57
0.10
10.00
1.00
1000 2000 3000 4000 5000
wassertiefe [ m1
b)
-=t-'-L..J-L..J-L..J-L..J-L..J-L..J-L..J-L..J-L..J-L..J-LJ.-L..J-l::- 100.00
-+-r-,,-,,-,,-,,-,,-,,-,,-,,-,,-,,-,,-,,-t- 0.01
202
•
r 2 = 0.70
154
•
187
•
o
1.00
0.10
0.01
1000 2000 3000 4000 5000 0
wassertiefe [ m1~ ~
a)
~ 100.00 -::j-'---L..-L...J....L...L-...L..L...L..L.l-LL...JL.L...l--L..-L...J....L...L-...L-'-'-.I-J=--
~IDc
j' 10.00aJ
~
~
Abb. 7.14 Zusammenhang der relativen Häufigkeit verschiedener Magnetosomformenin den Oberflächensedimenten des östlichen Südatlantiks (Fahrt M 20/2) mitder Stationstiefe. Die Korrelationskoeffizienten gelten für die jeweils eingezeichnete exponentielle Ausgleichskurve. Über den Symbolen ist die Anzahlder ausgezählten Magnetosome angegeben.
In Abbildung 7.14 sind die Verhältnisse der Anzahl von Magnetosomen unterschiedlicher Morphologie für die Oberflächensedimente des östlichen Südatlantiks (FahrtM 20/2) dargestellt (vgl. Anhang B: Abb.2). Projektil- und Karottenformen wurden dabeiunter der Bezeichnung Projektile zusammengefaßt. Kristalle mit abgerundeten Formenwaren insgesamt etwa halb so häufig wie Oktaeder oder Prismen. Da sie sich abernicht sicher den Magnetosomen zuordnen lassen, sind sie in Abbildung 7.14c nichtberücksichtigt. Man erkennt, daß sich die relative Häufigkeit der unterschiedlichenMagnetosomformen mit der Wassertiefe ändert. An den flachen Stationen sind imExtrakt mehr Prismen, bei den tiefen Stationen mehr Oktaeder vorhanden. DasVerhältnis Oktaeder / Prismen liegt für die flacheren Stationen « 2000 m) bei 0.04 bis0.5, in größeren Wassertiefen (> 4000 m) dagegen zwischen 2 und 10. Die Anzahl derprojektilförmigen Magnetosome nimmt ebenfalls mit zunehmender Wassertiefe ab,allerdings weniger deutlich als die der Prismen.
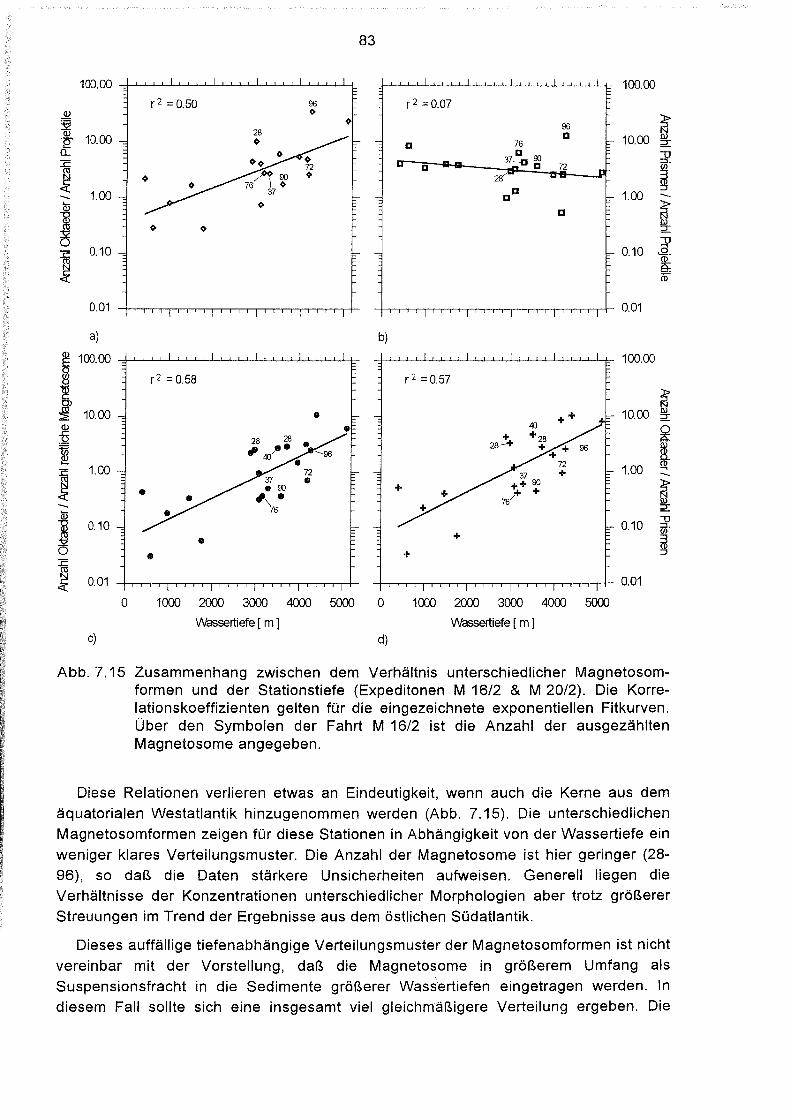
83
0.10
~~ 10.00 ::T
\J:::So
~t::- 1.00 --
~2:
~c- 0.10 .Q.
i§;CD
96EI
E:I
131!1
76E:I
37-e 9J
2~ EI 72
r 2 :::: 0.07
EI
-;:j--'LL..JLL..J1--'--'--'--'--"--'--'--'--'--"--,--,--,--,--,I--'--'--'--'--"--t:- 100.00
-t-r,....,,....,I,....,,....,'I,....,,....,",....,,....,",....,,....,'I+ 0.01
1.00
0.01 --+-rr-rr-rr-rr-rr-rr-rr-rr-r-rr-rr-rr-rrl-
100.00 -;:j--'~~~~~~~'--'-'--'-'--'-'--'-'--'-Y=--
~
~(j)
'e' 10.000..
~
~10.00 2:
I~--
0.10
1.00
+28-+
+
1000 2000 3000 4000 5000
Wassertiefe [ m]
r 2 :::: 0.57
+
+
b)
-t-r,....,,....,,....,,....,,....,,....,,....,,....,,....,,....,,....,,....,rl- 0.01
r 2 :::: 0.58
•
o
0.10
1.00
1000 2000 3000 4000 5000 0
\,f\f,;jssertiefe [ m]
~ ~
a)
~ 100.00 -=I-'---'--'---'---'--'--'-J.....J.....J-l......I.-'-J-...l.-L--'--'-J.....J.....J-l......I.-'-J--I=-
i~ 10.00
-E~
~
Abb. 7.15 Zusammenhang zwischen dem Verhältnis unterschiedlicher Magnetosomformen und der Stationstiefe (Expeditonen M 16/2 & M 20/2). Die Korrelationskoeffizienten gelten für die eingezeichnete exponentiellen Fitkurven.Über den Symbolen der Fahrt M 16/2 ist die Anzahl der ausgezähltenMagnetosome angegeben.
Diese Relationen verlieren etwas an Eindeutigkeit, wenn auch die Kerne aus demäquatorialen Westatlantik hinzugenommen werden (Abb. 7.15). Die unterschiedlichenMagnetosomformen zeigen für diese Stationen in Abhängigkeit von der Wassertiefe einweniger klares Verteilungsmuster. Die Anzahl der Magnetosome ist hier geringer (2896), so daß die Daten stärkere Unsicherheiten aufweisen. Generell liegen dieVerhältnisse der Konzentrationen unterschiedlicher Morphologien aber trotz größererStreuungen im Trend der Ergebnisse aus dem östlichen Südatlantik.
Dieses auffällige tiefenabhängige Verteilungsmuster der Magnetosomformen ist nichtvereinbar mit der Vorstellung, daß die Magnetosome in größerem Umfang alsSuspensionsfracht in die Sedimente größerer Wassertiefen eingetragen werden. Indiesem Fall sollte sich eine insgesamt viel gleichmäßigere Verteilung ergeben. Die

84
Absinkgeschwindigkeit eines einzelnen Magnetosoms durch die Wassersäule läßt sich
näherungsweise nach dem Stokeschen Gesetz berechnen. Beschreibt man ein
Magnetosom als Kugel mit einem Radius von 80 nm, so wird der Absinkprozess durch
folgende Kräfte bestimmt:
FGEWICHT = (PMAGNETIT - PWASSER) . VMAGNETOSOM . 9
FREIBUNG =6 . 1t . 11 . rMAGNETOSOM . VS1NK
F - Kraft
P - Dichte
V - Volumen
9 - Erdbeschleunigung
11 - Viskosität von Wasser
r - Radius
v - Geschwindigkeit
Daraus ergibt sich für die Sinkgeschwindigkeit der Magnetosome in der Wassersäule:
VS1NK =((PMAGNETIT - PWASSER) . VMAGNETOSOM . g) I (6·1t·11· rMAGNETosoM)
VS1NK =((5.2 - 1)'103 kg/m3) . (80.10-9 m)3 . 10 m/s I (6·1t·10-3 kg/m s . 40.10-9 m)
VS1NK =2.85.10-8 m/s =2.46.10-3 m/d =0.90 m/a
Ein Magnetosom benötigt nach diesen Annahmen 1.1 Jahre um einen Meter abzu
sinken. Die Vertikalgeschwindigkeiten der Meeresströmungen sind vielfach wesentlich
höher. WARREN & SPEER (1991) berechnen für das Angola Becken aufwärts gerichtete
Wasserbewegungen mit Geschwindigkeiten von 3.10-7 m/s bei 4000 m Wassertiefe und
von 8.10-7 m/s bei 2400 m Wassertiefe. Einzelne Magnetosome, aber auch ganze
Magnetosomketten könnten, da ihre Absinkgeschwindigkeit unter diesen Werten liegt,
fast beliebig weit verfrachtet werden. Die gefundene gute Sortierung der verschiedenen
Magnetosomformen läßt insofern den Schluß zu, daß die Magnetosome auch in
Tiefseesedimenten ganz überwiegend authigen gebildet werden.
7.6 Zusammenhang zwischen der Verteilung der Magnetosomformenund dem Nährstoffangebot im Sediment
Trägt man die Verhältniszahlen der verschiedenen Magnetosomformen gegen den Corg
Gehalt im Sediment (Abb. 7.16) oder den diffusiven Fluß von Sauerstoff ins Sediment
auf (Abb. 7.17), so läßt sich der Zusammenhang zwischen den Magnetosomformen und
dem Nährstoffgehalt oder dem Abbau von Nährstoffen im Sediment betrachten.
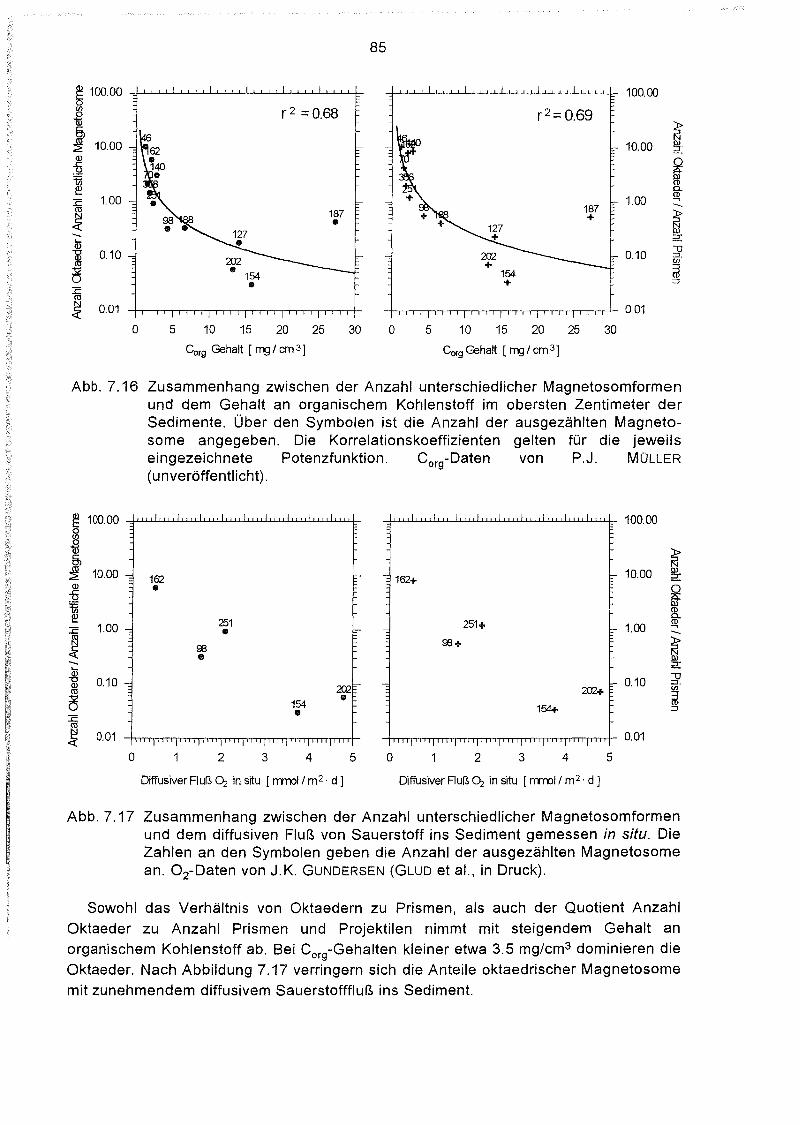
85
10.00
0.10
1.00
5 10 15 20 25 30
Corg Gehalt [rrg / cm 3 ]
o5 10 15 20 25 30
Corg Gehalt [ rrg / crn 3 ]
o0.01
Abb. 7.16 Zusammenhang zwischen der Anzahl unterschiedlicher Magnetosomformenund dem Gehalt an organischem Kohlenstoff im obersten Zentimeter derSedimente. Über den Symbolen ist die Anzahl der ausgezählten Magnetosome angegeben. Die Korrelationskoeffizienten gelten für die jeweilseingezeichnete Potenzfunktion. Corg-Daten von P.J. MÜLLER(unveröffentlicht) .
0.10
10.00
1.00
54
154+
32
251+
00+
o
--:J-Ll-U.U-U.U-U.Uu..L.UL.J..J...LL.J..J...Lu..w..u..w..u..w..u..w..u..w..u..w..i:- 100.00
-rn",..,rn-nrrrT"rnTrTlTrrrrTTrrTTrrTTTTTTTT"TTnTTTT-r- 0.01
543
154•
2o0.01 -rn,.,.,..,""-'-ITTrTT"ITTnTTnTTnT"TTn,..,....,,..,....,,,,..,,,,..,rl-
Diffusiver Ruß 0:2 in situ [rrrrol / m2 . d] Diffusiver Fluß 0:2 in situ [rrrrol / m2 . d]
Abb. 7.17 Zusammenhang zwischen der Anzahl unterschiedlicher Magnetosomformenund dem diffusiven Fluß von Sauerstoff ins Sediment gemessen in situ. DieZahlen an den Symbolen geben die Anzahl der ausgezählten Magnetosomean. 02-Daten von J.K. GUNDERSEN (GLUD et al., in Druck).
Sowohl das Verhältnis von Oktaedern zu Prismen, als auch der Quotient AnzahlOktaeder zu Anzahl Prismen und Projektilen nimmt mit steigendem Gehalt anorganischem Kohlenstoff ab. Bei Corg-Gehalten kleiner etwa 3.5 mg/cm3 dominieren dieOktaeder. Nach Abbildung 7.17 verringern sich die Anteile oktaedrischer Magnetosomemit zunehmendem diffusivem Sauerstofffluß ins Sediment.

86
Magnetbakterien können nach der geochemischen Zone, in der sie leben, in verschiedene physiologische Gruppen eingeteilt werden (Kapitel 5.5). In den marinen Sedimenten muß man zwischen unterschiedlichen Gruppen von Magnetbakterien differenzieren (Kapitel 6.10). Während im Auftriebsgebiet vor Namibia anaerobe Magnetbakterien aus geochemisch tieferen Schichten die größte Konzentration aufweisen,waren Magnetbakterien in den pelagischen Ablagerungen des zentralen Atlantiks - wennüberhaupt - vor allen im obersten Zentimeter des Sediments nachweisbar. DieseBakterien sind vermutlich aerob.
Die bisher in Kultur gebrachten aeroben Magnetbakterien besitzen oktaedrischeMagnetosome (BLAKEMORE et al., 1979; MATSUNAGA et al., 1991; SCHLEIFER et al.,1991), die (fakultativ) anaeroben Magnetbakterien dagegen Prismen oder projektilförmige Magnetosome (BAZYLINSKI et al., 1988; SAKAGUCHI et al., 1993). Die beiMELDRUM et al. (1993a und 1993b) beschriebenen Magnetbakterien sind physiologischkaum untersucht. Es wird beschrieben, daß die Bakterien mikroaerophile Bänder bilden,gleichzeitig wird aber vermutet, daß der Stamm MV-2 zur gleichen Art gehört, wie deranaerobe Stamm MV-1 (BAZYLINSKI et al., 1988). Auch die Toleranz gegenüber H2S läßtvermuten, daß diese Stämme (fakultativ) anaerob sind. Die Bakterien dieser Stämmebesitzen prismatische Magnetosome.
TEM-Aufnahmen von magnetotaktischen Bakterien aus der anaeroben Schicht imWatt von Weddewarden zeigen, daß keines der untersuchten Magnetbakterien oktaedrische Magnetosome bildet.
BLAKEMORE et al. (1985) vermuteten, daß Magnetosome als Paläoredoxanzeiger verwendet werden könnten. Dabei gingen sie davon aus, daß alle Magnetbakterienmikroaerophil sind und postulierten, daß die Magnetosome als Indikatoren für mikroaerobes Milieu dienen könnten. Mit der Kultivierung des ersten anaeroben Magnetbakterienstammes (BAZYLINSKI et al., 1988) wurde diese Vorstellung widerlegt. Es gibtvermutlich sehr viele verschiedene Arten von Magnetbakterien. Die bisher in Kulturgebrachten Stämme liefern daher nur einen Einblick in die Vielfalt der Stoffwechselwegeder Magnetbakterien.
Nach den Ergebnissen dieser Arbeit scheint ein Zusammenhang zwischen Stoffwechsel und Magnetosomform zu existieren. Sollte sich weiter bestätigen, daß Mehrzahlder aeroben Magnetbakterien Oktaeder bilden, die anaeroben Magnetbakteriendagegen Prismen, Projektile oder Karotten, so könnten die Magnetosome als Paläoredoxanzeiger verwendet werden, und es ließen sich mit der hier formulierten Unterscheidung aerobe und anaerobe Milieus in Oberflächensedimenten rekonstruieren.
Da das Vorkommen anaerober Magnetbakterien auf nährstoffreiche Sedimentebeschränkt ist, könnten prismatische Magnetosomformen auch als Indikatoren für diePaläoproduktivität dienen.

87
8 Gesteinsmagnetik
In den Kapiteln 6 und 7 sind Versuche beschrieben worden, die Verteilung lebenderMagnetbakterien sowie fossilen bakteriellen Magnetits in den Sedimenten desSüdatlantiks auf der Basis von licht- und elektronenmikroskopischen Untersuchungenzu ermitteln. Um diese Ergebnisse abzusichern, wurden gesteinsmagnetische Untersuchungen mit dem neuerworbenen 'Alternating-Gradient-Magnetometer' (AGM) anOberflächenproben der Expeditionen M 16/2 und M 20/2 durchgeführt. Die aus denHysteresemessungen gewonnenen gesteinsmagnetischen Parameter wurden mitStandardverfahren (DAY et al., 1977) interpretiert. Mit einem von V. DOBENECK (1993)eingeführten ModelIierverfahren ist darüber hinaus versucht worden, die Magnetomineralogie der Sedimente und die Anteile verschiedener magnetischer Fraktionen zucharakterisieren. Des weiteren wurden Messungen der anhysteretischen und derisothermalen remanenten Magnetisierung durchgeführt.
8.1 Messungen mit dem Alternating-Gradient-Magnetometer
8.1.1 Aufbau und Meßprinzip der AGM Apparatur
Mit dem Alternating-Gradient-Magnetometer (Princeton Measurement Corporation)lassen sich Hystereseschleifen sowie Backfield Kurven für Sedimentproben von einem
Gewicht bis zu etwa 10 - 20 mg messen. Dabei wirken auf die Probe zwei unabhängigeMagnetfelder, ein homogenes Feld, das sie aufmagnetisiert, und ein Gradientenfeld,das eine Kraft auf sie ausübt (Abb. 8.1). Die Kraft, mit der die Probe vomGradientenfeld bewegt wird, ist proportional zur Probenmagnetisierung. Die Auslenkungwird über die Glasstäbe des Probenhalters auf einen Piezokristall übertragen und dortgemessen.
s
H P
Abb. 8.1 Funktionsskizze des Alternating-Gradient-Magnetometers (AGM). F bezeichnet die Feldspule, die die Probe P aufmagnetisiert, G die Gradientenspule,die eine magnetische Kraft (Auslenkung) auf die Probe ausübt. I?erProbenhalter H ist an zwei Glasstäben S aufgehängt. Sie übertragenwährend der Messung eine periodische Kraft auf einen Piezokristall, die proportional zur Auslenkung der Probe ist.

88
Um die Meßgenauigkeit zu optimieren, wird als Gradientenfeld ein Wechselfeldbenutzt, dessen Frequenz der Eigenfrequenz des Systems Probenhalter - Probe entspricht. Bei der Anregung mit dieser Frequenz schwingt die Probe mit größtmöglicher
Auslenkung und erzeugt im Piezokristall ein maximales Meßsignal. Je nach Masseschwingen die Proben während der Messung mit einer Frequenz von 300 bis 500 Hzbei einer Auslenkung von wenigen Mikrometern.
8.1.2 Probenpräparation
Um Kontaminationen und Verschleppungen zwischen verschiedenen Proben zu vermeiden, wurde eine Präparationstechnik konzipiert, die nur Einwegmaterialien benutzt.Das Probengefäß besteht aus einem drei Millimeter hohen Stück Trinkhalm, das aufeine kreisrunde Kunststoffolie geklebt wird. Auf der Unterseite der Folie ist die
Probennummer vermerkt. Jedes Probengefäß ist erst unmittelbar vor Benutzunghergestellt worden, um eine magnetische Verschmutzung durch Eisenstaub aus derLuft zu vermeiden.
Das Sediment wurde in leicht feuchtem Zustand mit einem Mikrospatel in dasProbengefäß gefüllt. Um ein möglichst repräsentatives Meßergebnis zu erhalten,wurden kleinere Mengen aus verschiedenen Partien der Sedimentprobe entnommen.Aufgrund der Spulenkonfiguration des AGM dürfen die Probenmaße 3x3x2 mm3 nicht
übersteigen. Dadurch und durch die Stabilität des Probenhalters ergibt sich dieBeschränkung auf 10 - 20 mg Sediment. Die Probe wird im Exsikkator getrocknet undanschließend mit einem Plastikklebstoff verklebt. Dadurch soll die Probe fixiert undstabilisiert werden, um Bewegungen der magnetischen Partikel bei der Messung zuverhindern.
8.1.3 Messung
Das Probengefäß wird zur Messung mittels Silikonfett auf den Probenhalter (Abb. 8.1)geklebt. Da die Probe beim Meßvorgang vibriert, muß sorgfältig auf eine stabileBefestigung geachtet werden.
Für alle Proben wurden zwei Hystereseschleifen ermittelt, die erste bei einerMaximalfeldstärke von 0.3 T (Tesla), die zweite bei einer Maximalfeldstärke von 1 T.Während mit der ersten Messung bevorzugt niederkoerzitive Minerale, vor allemMagnetit, erfaßt werden, lassen sich bei der Messung bis 1 Tauch höherkoerzitiveMinerale, insbesondere Hämatit (a-Fe20 3), nachweisen. Um eine möglichst guteAuflösung der Kurven zu erreichen, wurden die Hystereseschleife bis 0.3 T in Schrittenvon 2 mT, die der Messung bis 1 T in Schritten von 5 mT gemessen. Eine Hystereseschleife mit 600 Meßwerten wird in etwa drei Minuten durchlaufen. Die Gerätedrift istentsprechend gering.
An allen Proben wurde außerdem eine Backfield Kurve ermittelt. Dazu wird dieProbe zunächst in einem Feld von 0.3 Taufmagnetisiert und anschließend mit höheren
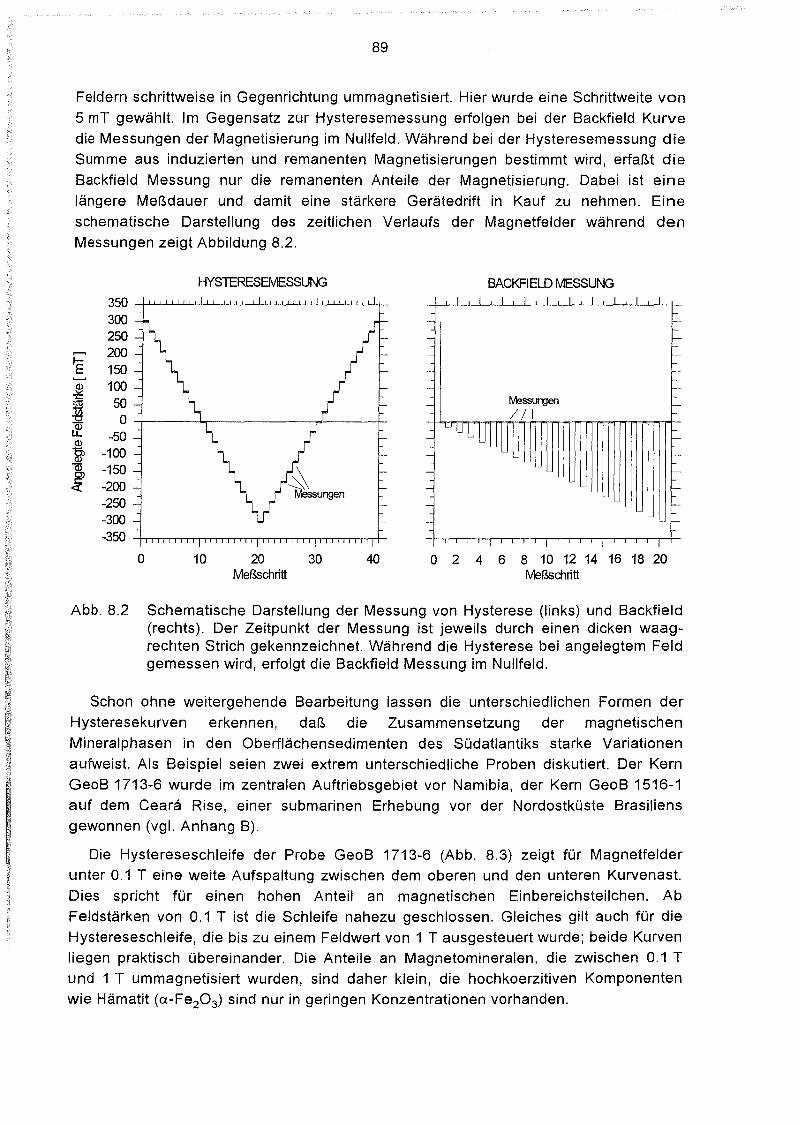
89
Feldern schrittweise in Gegenrichtung ummagnetisiert. Hier wurde eine Schrittweite von
5 mT gewählt. Im Gegensatz zur Hysteresemessung erfolgen bei der Backfield Kurvedie Messungen der Magnetisierung im Nullfeld. Während bei der Hysteresemessung dieSumme aus induzierten und remanenten Magnetisierungen bestimmt wird, erfaßt die
Backfield Messung nur die remanenten Anteile der Magnetisierung. Dabei ist einelängere Meßdauer und damit eine stärkere Gerätedrift in Kauf zu nehmen. Eineschematische Darstellung des zeitlichen Verlaufs der Magnetfelder während denMessungen zeigt Abbildung 8.2.
HYSTERESEMESSUNG BACKFIELD MESSUNG
rv1essurgen/ / I
o 2 4 6 8 10 12 14 16 18 20rv1eßschritt
4020 30Meßschritt
10o
350 -t-'-.L.L.L.L.L.LLLLLLLLLLLLLLLLJ-LLl-L.L..1..LLJ..LLLLLL.L.Lt-
300250
.......... 200
'E 150~
~ 100
~ ~~ -t-------'-..-------~---+-
-50
~ -100~ -150
<i:: -200-250-300
-350 -h-rrrrrrrrrrrrrrnrrn-rrT--rTT--rTT"""'TTTTTTn-I--
Abb. 8.2 Schematische Darstellung der Messung von Hysterese (links) und Backfield(rechts). Der Zeitpunkt der Messung ist jeweils durch einen dicken waagrechten Strich gekennzeichnet. Während die Hysterese bei angelegtem Feldgemessen wird, erfolgt die Backfield Messung im Nullfeld.
Schon ohne weitergehende Bearbeitung lassen die unterschiedlichen Formen derHysteresekurven erkennen, daß die Zusammensetzung der magnetischenMineralphasen in den Oberflächensedimenten des Südatlantiks starke Variationenaufweist. Als Beispiel seien zwei extrem unterschiedliche Proben diskutiert. Der KernGeoB 1713-6 wurde im zentralen Auftriebsgebiet vor Namibia, der Kern GeoB 1516-1auf dem Ceara Rise, einer submarinen Erhebung vor der Nordostküste Brasiliensgewonnen (vgl. Anhang B).
Die Hystereseschleife der Probe GeoB 1713-6 (Abb. 8.3) zeigt für Magnetfelderunter 0.1 T eine weite Aufspaltung zwischen dem oberen und den unteren Kurvenast.Dies spricht für einen hohen Anteil an magnetischen Einbereichsteilchen. AbFeldstärken von 0.1 T ist die Schleife nahezu geschlossen. Gleiches gilt auch für dieHystereseschleife, die bis zu einem Feldwert von 1 T ausgesteuert wurde; beide Kurvenliegen praktisch übereinander. Die Anteile an Magnetomineralen, die zwischen 0.1 Tund 1 T ummagnetisiert wurden, sind daher klein, die hochkoerzitiven Komponentenwie Hämatit (a-Fe20 3) sind nur in geringen Konzentrationen vorhanden.
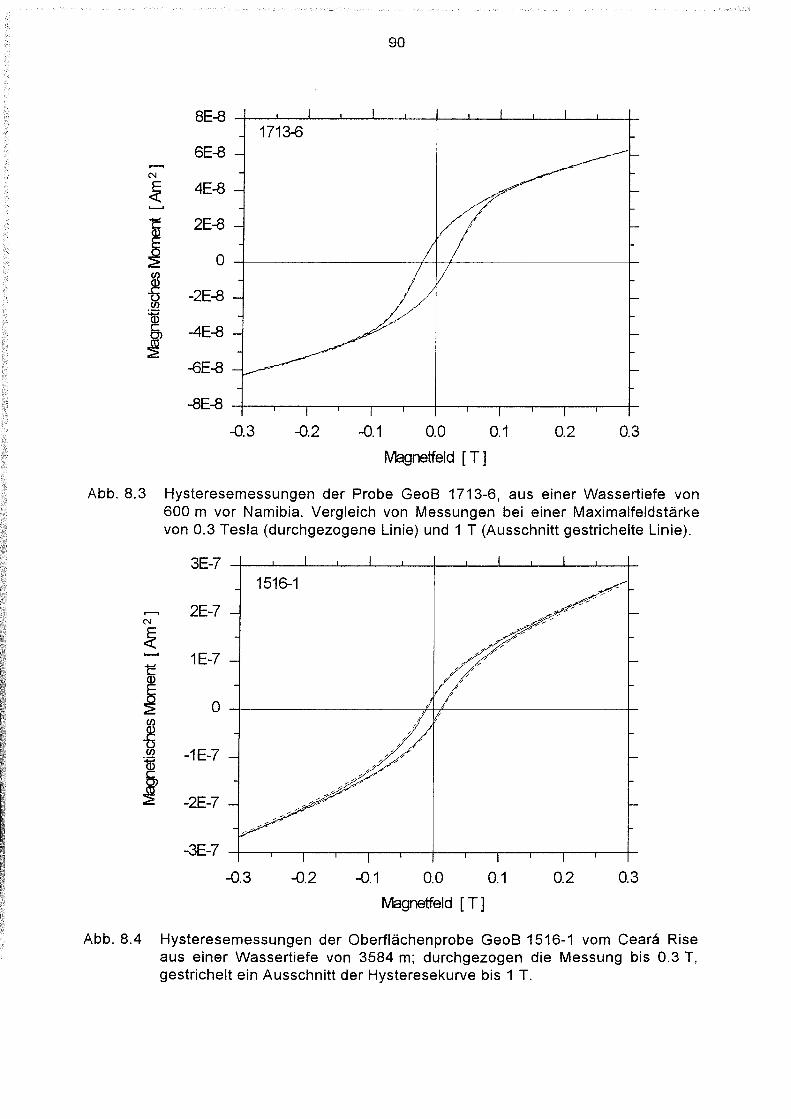
90
8E-81713-6
6E-8N
~ 4E-8
C 2E-8Q)
~ 0
~ -2E-8:mc -4E-80>
~-6E-8
-8E-8
-0.3 -0.2 -0.1 0.0 0.1 0.2 0.3
Magnetfeld [T]
Abb. 8.3 Hysteresemessungen der Probe GeoB 1713-6, aus einer Wassertiefe von600 m vor Namibia. Vergleich von Messungen bei einer Maximalfeldstärkevon 0.3 Tesla (durchgezogene Linie) und 1 T (Ausschnitt gestrichelte Linie).
3E-71516-1
2E-7N
~C
1E-7Q)
~ 0
~Cf) -1E-7:mc~ -2E-7
-3E-7
-0.3 -0.2 -0.1 0.0 0.1 0.2 0.3
Magnetfeld [T]
Abb. 8.4 Hysteresemessungen der Oberflächenprobe GeoB 1516-1 vom Ceara Riseaus einer Wassertiefe von 3584 m; durchgezogen die Messung bis 0.3 T,gestrichelt ein Ausschnitt der Hysteresekurve bis 1 T.
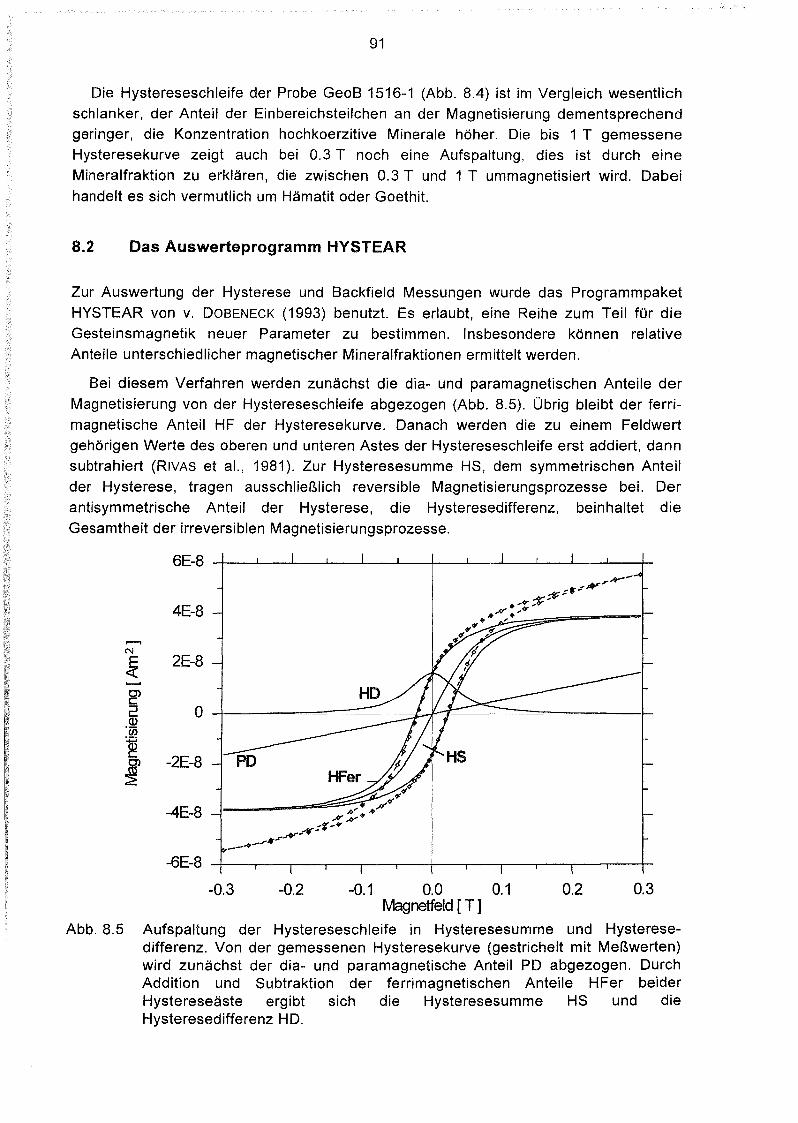
91
Die Hystereseschleife der Probe GeoB 1516-1 (Abb. 8.4) ist im Vergleich wesentlichschlanker, der Anteil der Einbereichsteilchen an der Magnetisierung dementsprechendgeringer, die Konzentration hochkoerzitive Minerale höher. Die bis 1 T gemesseneHysteresekurve zeigt auch bei 0.3 T noch eine Aufspaltung, dies ist durch eine
Mineralfraktion zu erklären, die zwischen 0.3 T und 1 T ummagnetisiert wird. Dabeihandelt es sich vermutlich um Hämatit oder Goethit.
8.2 Das Auswerteprogramm HYSTEAR
Zur Auswertung der Hysterese und Backfield Messungen wurde das ProgrammpaketHYSTEAR von v. DOBENECK (1993) benutzt. Es erlaubt, eine Reihe zum Teil für dieGesteinsmagnetik neuer Parameter zu bestimmen. Insbesondere können relativeAnteile unterschiedlicher magnetischer Mineralfraktionen ermittelt werden.
Bei diesem Verfahren werden zunächst die dia- und paramagnetischen Anteile derMagnetisierung von der Hystereseschleife abgezogen (Abb. 8.5). Übrig bleibt der ferrimagnetisehe Anteil HF der Hysteresekurve. Danach werden die zu einem Feldwertgehörigen Werte des oberen und unteren Astes der Hystereseschleife erst addiert, dannsubtrahiert (RIVAS et al., 1981). Zur Hysteresesumme HS, dem symmetrischen Anteil
der Hysterese, tragen ausschließlich reversible Magnetisierungsprozesse bei. Derantisymmetrische Anteil der Hysterese, die Hysteresedifferenz, beinhaltet dieGesamtheit der irreversiblen Magnetisierungsprozesse.
6E-8 -1--.l...---L_-l.-_.l...---L_-I-_.l-----L_....l.-_.L..----L_-1-
4E-8
2E-8
o
-2E-8
-4E-8
0.30.2-0.2
-6E-8
-0.3 -0.1 0.0 0.1Magnetfeld [ T]
Abb. 8.5 Aufspaltung der Hystereseschleife in Hysteresesumme und Hysteresedifferenz. Von der gemessenen Hysteresekurve (gestrichelt mit Meßwerten)wird zunächst der dia- und paramagnetische Anteil PD abgezogen. DurchAddition und Subtraktion der ferrimagnetischen Anteile HFer beiderHystereseäste ergibt sich die Hysteresesumme HS und dieHysteresedifferenz HO.

92
Nach v. DOBENECK (1993) kann die Hysteresesumme homogener Mineralfraktionenin guter Näherung durch eine einzige Tangens-Hyperbolicus-Funktion, die Hysteresedifferenz durch eine einzige (modifizierte) Secans-Hyperbolicus-Funktion approximiertwerden.
Ist die Konzentration der magnetischen Minerale in Sedimenten so gering, daß sichihre wechselseitige Beeinflussung vernachlässigen läßt, so kann die Hystereseschleifeeiner Probe aufgefaßt werden als Summe der Hystereseschleifen aller enthalteneneinzelnen Minerale. Ziel der ModelIierung mit dem Programm HYSTEAR ist es, dieHysteresekurve additiv aus Komponenten verschiedener Koerzitivfeldstärken zusam
menzusetzen. Als Ergebnis erhält man Komponentenspektren für die Hysteresesummeund die Hysteresedifferenz, wobei die jeweiligen Komponenten charakterisiert werdendurch das
. Halbwertsfeld der Hysteresesumme:
Feldstärke, bei der die Hysteresesumme die Hälfte ihres Maximalwertes erreicht, unddas
- Halbwertsfeld der Hysteresedifferenz:
Feldstärke, bei der die Hysteresedifferenz auf die Hälfte des Wertes zurückgeht, derim Nullfeld gemessen wurde. Dieser Wert liefert nach v. DOBENECK (1993) dieselbeInformation wie die Remanenzkoerzitivfeldstärke.
Diese Parameter werden bestimmt, nachdem para- und diamagnetische Anteile,ermittelt nach dem Gesetz von WEISS & FORRER (1929), von den Hystereswertenabgezogen wurden. Dabei wird die Restkrümmung der Kurve bei hohen Feldwerten biszu unendlich großen Feldwerten extrapoliert. Die Halbwertsfelder werden deshalb imweiteren als wahre Halbwertsfelder (true median fields) bezeichnet.
Die verwendeten Funktionen, der Tangens hyperbolicus und der Secans hyper
bolicus, liefern kein orthonormierbares Funktionensystem. Eine eindeutige Zerlegungder Hystereseschleife in verschiedene Komponenten (Elementarkurven) ist dahergrundsätzlich nicht möglich, die Kurve muß gefittet werden. Als Fit-Algorithmus dientdas Remez Verfahren. Zunächst wird eine Elementarkurve so ausgewählt, daß sie diegemessenen Daten möglichst gut approximiert. Der Anteil dieser Elementarkurve wirdvon der vorhandenen Kuve abgezogen. Die nächste Elementarkurve wird an dasverbleibende Residuum angepaßt. Die einzelnen Elementarkurven einer solchenAnfangsverteilung werden anschließend so lange ausgetauscht, bis die Verbesserungder Kurvenanpassung einen bestimmten Wert unterschreitet.
Bei allen Fitprozeduren werden die Modellkurven so lange iterativ an die Meßwerteangenähert, bis die Differenz zwischen Meßwerten und Fitkurve ein (lokales) Minimumerreicht. Die mit dem Programm gefundene Approximation beschreibt aber nicht injedem Fall die richtige Verteilung magnetischer Mineralkomponenten im Sediment.Nach v. DOBENECK (1993) haben Berechnungen mit zufälligen Anfangsbedingungen(Monte-Carlo-Verfahren) zu unterschiedlichen Ergebnissen geführt, die sich aber in derGüte der Approximation kaum unterschieden. Das verwendete Remez Verfahren liefert

93
vor allem stabile und in der Regel einfache Lösungen. Bei der Interpretation derSpektren muß daher beachtet werden, daß die Lösungen niemals eindeutig sind.
Dadurch erhält man Komponentenspektren. Jede Komponente wird charakterisiertdurch ihr Halbwertsfeld, das ist die Magnetfeldstärke, bei der Hysteresesummen unddie Hysteresedifferenzen den halben Maximalwert erreichen. Im Programm HYSTEARwird versucht, durch Variation der Anteile dieser Komponenten die gemesseneHystereseschleife zu approximieren und dadurch die Mineralzusammensetzung zurekonstruieren.
Neben der chemischen Zusammensetzung beeinflussen aber auch der Gitteraufbau,die Korngröße, die Kornform und die Stärke innerer Spannungen die magnetischenEigenschaften eines Minerals.
Beispielsweise ist Magnetit von magnetotaktischen Bakterien weitgehend
spannungsfrei, da die Synthese der Magnetosome sehr kontrolliert erfolgt. SeineKoerzitivfeldstärke ist dadurch geringer als die von Iithogenem Magnetit derselbenKorngröße und Kornform. Titanomagnetit, der durch Verwitterung von Ozeanbasalten
ins Sediment eingetragen wird, sowie terrigener (Titano-) Magnetit haben höhereKoerzitivfeldstärken, (Titano-) Magnetit aus einem explosiven Vulkanismus, ewa vonInselbögen, besitzt aufgrund der sehr hohen inneren Spannungen die höchstenKoerzitivfeldstärken.
Ziel einer Analyse von Hysteresekomponenten ist es also, sowohl verschiedeneMagnetominerale zu identifizieren, als auch Variationen ihrer kritischen Parameter,insbesondere der Korngrößen zu unterscheiden.
8.3 Regionale Charakteristika magnetischer Komponentenspektren derOberflächensedimente im Südatlantik
Alle gemessenen Hysteresekurven wurden mit dem zuvor beschriebenen ProgrammHYSTEAR von v. DOBENECK (1993) modelliert. Dabei sind jeweils Spektren für 30 Komponenten berechnet worden, um verschiedene magnetische Mineralphasen voneinander zu trennen. Typischerweise sind zwei bis drei, nur in Extremfällen mehr als fünfPhasen zu erwarten.
Geographisch benachbarte Proben ergaben ähnliche Spektren, Proben aus anderen
Meeresgebieten liefern davon verschiedene Spektren. Für die Sedimente der FahrtenM 16/2 und M 20/2 ließen sich sechs Gebiete anhand der Zusammensetzung ihrerMagnetominerale unterscheiden. Exemplarische Spektren von Proben dieser sechsGebiete sind in den Abbildungen 8.6 und 8.7 dargestellt. Dabei wurde darauf geachtet,daß jeweils eine wenig verrauschte Meßkurve ausgewählt wurde, die eineentsprechend gute Approximation zuließ.
Die Probe GeoB 1508 vom westlichen Abhang des Mittelatlantischen Rückens vorder Nordostküste Brasiliens aus einer Wassertiefe von 3685 m weist eine sehr breiteVerteilung von Spektralklassen auf. Für die Hysteresesumme lassen sich drei bis vierGruppen von Spektralklassen zusammenfassen. Bedeutende Anteile (~ 38 %) liegen inden Spektralklassen 3 - 10, der weichmagnetischen Fraktion mit Halbwertsfeldern

94
zwischen 6 und 30 mT. Etwa 35 % liegen in den Spektralklassen 12 - 17 mit Halbwertsfeldern von 40 bis 60 mT, der Rest verteilt sich auf die höheren Spektralklassen 18 bis28 (Halbwertsfelder 70 - 400 mT). Die weitere Aufspaltung dieser hochkoerzitivenKlasse ist etwas problematisch, da sie in den anderen Kernen aus diesem Gebiet einuneinheitliches Bild zeigte. Für die Hysteresedifferenz lassen sich drei Gruppen vonSpektralklassen klar unterscheiden, eine weichmagnetische Komponente in denKlassen 5 - 9 (38 %), eine Komponente mit Halbwertsfeldern zwischen 30 und 60 mT(35 %) und ein hochkoerzitiver Anteil mit Halbwertsfeldern größer 70 mT. Auffällig ist,daß bei der Hysteresedifferenz hier, wie auch in den anderen Spektren keine
bedeutenden weichmagnetischen Anteile nachweisbar waren. Das Maximum dermittelkoerzitiven Phase liegt bei Halbwertsfeldern um 40 mT und damit niedriger als beiden Proben der anderen Gebiete.
Der Kern GeoB 1513 aus dem Amazonasfächer (Wassertiefe 3621 m) weistebenfalls drei Komponenten auf. Niederkoerzitive Anteile (Halbwertsfelder unter 30 mT)liegen bei etwas über 20 %. Der mittelkoerzitive Anteil trägt zu 55 % zum magnetischenSignal bei. Er hat sein Maximum bei 55 bis 60 mT, ein verglichen mit den anderenProben hoher Wert.
Der Kern GeoB 1519 vom Hang des Ceara Rise (Wassertiefe 4199 m) zeigt ein
Spektrum, das sich in etwa zwischen den beiden zuvor beschriebenen einordnen läßt.Der niederkoerzitive Anteil (25 - 30%) ist höher als in Probe GeoB 1513, der mittelkoerzitive Anteil ist wie dort dominierend (50%), die Halbwertsfelder betragen hier um
50 mT.
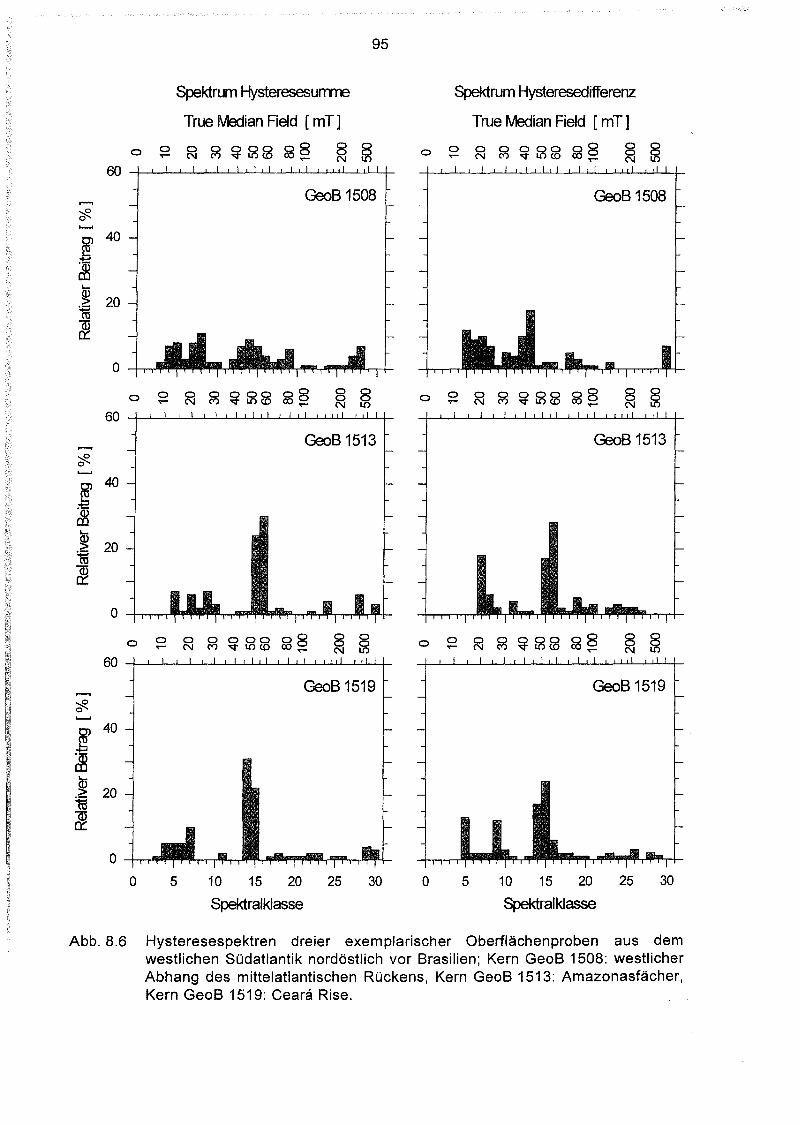
95
Spektrum Hysteresesunme Spektrum Hysteresedifferenz
True Median Field [mT] True Median Field [mT]
0 0 0 ~Sfgg2§ 8 § 0 0 0 ~ Sf gg g8 8 §..- N N ..... N ..- N
60
GeoB 1508 GeoB 1508'cf2.
~40
.....~...~ 20iä&
0
0 0 ~ Sf gg g8 8 8 0 0 ~Sfgg2@ 8 00 0 0..- N ..... N ll) ..... N N ll)
60
GeoB 1513 GeoB 1513~0
E 40
~...~ 20iä~
0
0 0 0 000 00 0 8 0 0 ~Sfgg2§ 8 §0 co O 0 0..- N C") "<t ll) co ..... N ll) ..- N N
60
GeoB 1519 GeoB 1519~0
~40
.....~
~ 20iä~
0
0 5 10 15 20 25 30 0 5 10 15 20 25 30
Spektralklasse Spektralklasse
Abb.8.6 Hysteresespektren dreier exem plarischer Oberflächenproben aus demwestlichen Südatlantik nordöstlich vor Brasilien; Kern GeoB 1508: westlicherAbhang des mittelatlantischen Rückens, Kern GeoB 1513: Amazonasfächer,Kern GeoB 1519: Ceara Rise.
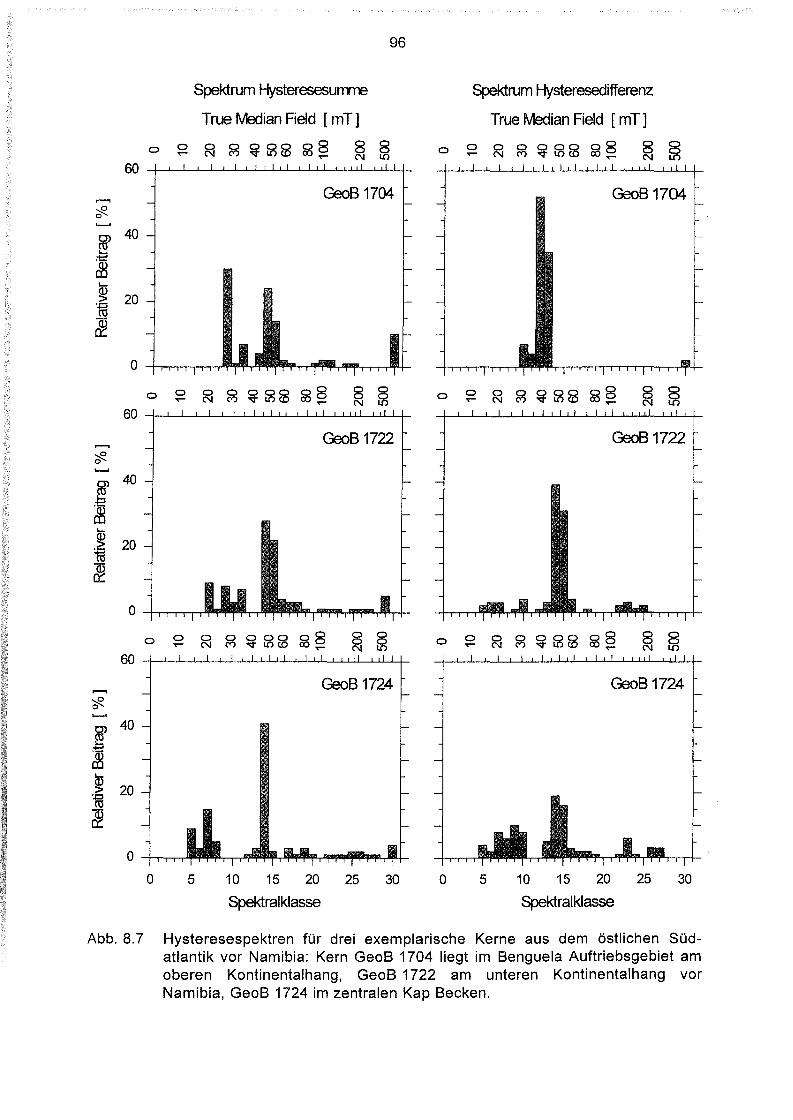
96
Spektrum Hysteresesurrme
True Median Field [mT]
o ~ ~ ~ ~ f5g g § ~ §
GeoB 1704
Spektrum Hysteresedifferenz
True Median Field [mT]
o ~ ~ ~ ~ f5g g § ~ §
GeoB 1704
~o
...~ 20
~
~o
...~ 20
&
o 5
GeoB 1722
8 8N L()
GeoB 1724
10 15 20 25 30
Spektralklasse
o 5
GeoB 1722
8 8N LO
GeoB 1724
10 15 20 25 30
Spektralklasse
Abb. 8.7 Hysteresespektren für drei exemplarische Kerne aus dem östlichen Südatlantik vor Namibia: Kern GeoB 1704 liegt im Benguela Auftriebsgebiet amoberen Kontinentalhang, GeoB 1722 am unteren Kontinentalhang vorNamibia, GeoB 1724 im zentralen Kap Becken.
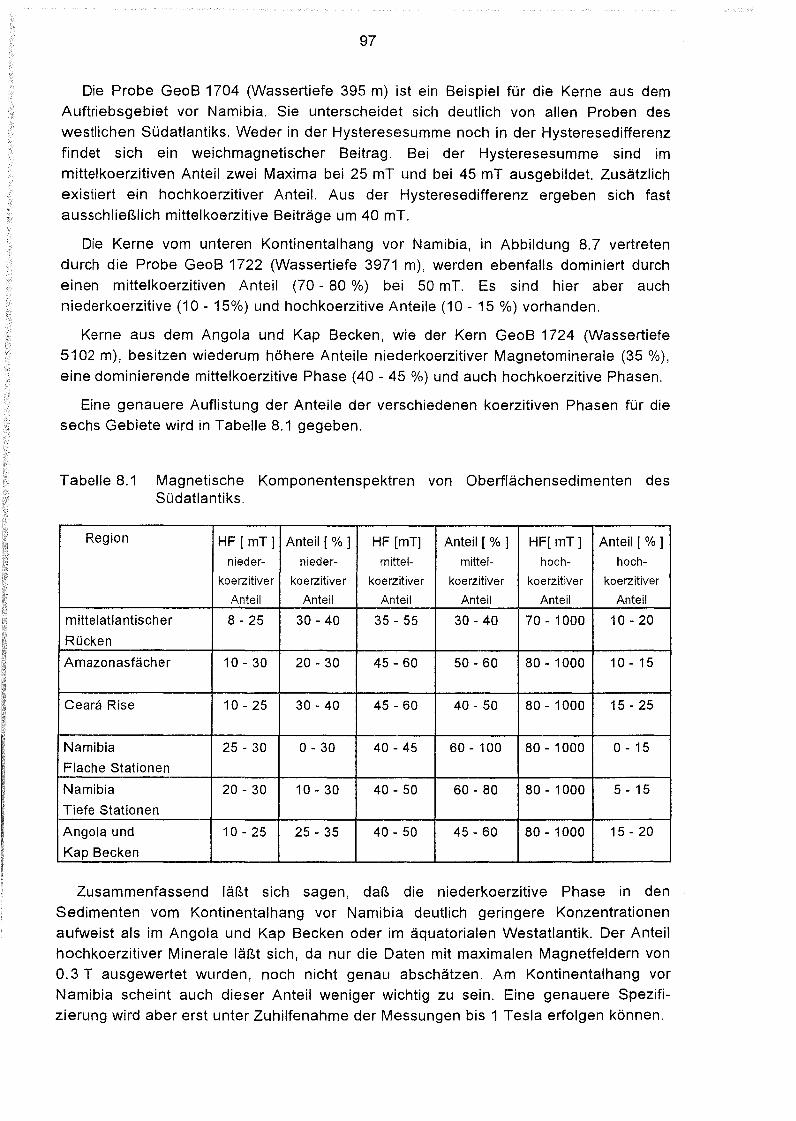
97
Die Probe GeoB 1704 (Wassertiefe 395 m) ist ein Beispiel für die Kerne aus demAuftriebsgebiet vor Namibia. Sie unterscheidet sich deutlich von allen Proben deswestlichen Südatlantiks. Weder in der Hysteresesumme noch in der Hysteresedifferenzfindet sich ein weichmagnetischer Beitrag. Bei der Hysteresesumme sind immittelkoerzitiven Anteil zwei Maxima bei 25 mT und bei 45 mT ausgebildet. Zusätzlich
existiert ein hochkoerzitiver Anteil. Aus der Hysteresedifferenz ergeben sich fastausschließlich mittelkoerzitive Beiträge um 40 mT.
Die Kerne vom unteren Kontinentalhang vor Namibia, in Abbildung 8.7 vertretendurch die Probe GeoB 1722 (Wassertiefe 3971 m), werden ebenfalls dominiert durcheinen mittelkoerzitiven Anteil (70 - 80 %) bei 50 mT. Es sind hier aber auchniederkoerzitive (10 - 15%) und hochkoerzitive Anteile (10 - 15 %) vorhanden.
Kerne aus dem Angola und Kap Becken, wie der Kern GeoB 1724 (Wassertiefe5102 m), besitzen wiederum höhere Anteile niederkoerzitiver Magnetominerale (35 %),
eine dominierende mittelkoerzitive Phase (40 - 45 %) und auch hochkoerzitive Phasen.
Eine genauere Auflistung der Anteile der verschiedenen koerzitiven Phasen für diesechs Gebiete wird in Tabelle 8.1 gegeben.
Tabelle 8.1 Magnetische Komponentenspektren von Oberflächensedimenten desSüdatlantiks.
Region HF[mT] Anteil [% ] HF [mT] Anteil [% ] HF[ mT] Anteil [%]
nieder- nieder- mittel- mittel- hoch- hoch-
koerzitiver koerzitiver koerzitiver koerzitiver koerzitiver koerzitiver
Anteil Anteil Anteil Anteil Anteil Anteil
mittelatlantischer 8 - 25 30 - 40 35 - 55 30 - 40 70 - 1000 10 - 20
Rücken
Amazonasfächer 10 - 30 20 - 30 45 - 60 50 - 60 80 - 1000 10- 15
Ceara Rise 10 - 25 30 - 40 45 - 60 40 - 50 80 - 1000 15 - 25
Namibia 25 - 30 0-30 40 - 45 60 - 100 80 - 1000 0-15
Flache Stationen
Namibia 20 - 30 10- 30 40 - 50 60 - 80 80 - 1000 5 - 15
Tiefe Stationen
Angola und 10- 25 25 - 35 40 - 50 45 - 60 80 - 1000 15 - 20
Kap Becken
Zusammenfassend läßt sich sagen, daß die niederkoerzitive Phase in denSedimenten vom Kontinentalhang vor Namibia deutlich geringere Konzentrationenaufweist als im Angola und Kap Becken oder im äquatorialen Westatlantik. Der Anteilhochkoerzitiver Minerale läßt sich, da nur die Daten mit maximalen Magnetfeldern von0.3 T ausgewertet wurden, noch nicht genau abschätzen. Am Kontinentalhang vorNamibia scheint auch dieser Anteil weniger wichtig zu sein. Eine genauere Spezifizierung wird aber erst unter Zuhilfenahme der Messungen bis 1 Tesla erfolgen können.

98
In den Sedimenten, die im küstennahen Auftrieb vor Namibia liegen und in denen diehöchsten Konzentrationen an Magnetbakterien nachgewiesen werden konnten, istpraktisch nur die mittelkoerzitive Fraktion nachweisbar. Die Bedeutung dieser Phasenimmt mit zunehmendem Abstand vom Küstenauftrieb ab.
Die niederkoerzitive Phase, mit Halbwertsfeldern von 10 - 30 mT, wird vermutlichdurch (Titano-) Magnetit und (Titano-) Maghemit in Form von magnetischen Mehrbereichsteilchen oder Pseudo-Einbereichsteilchen gebildet. Auch superparamagnetische Partikel, wie der extrazellulär gebildete Magnetit der in Kapitel 2.2.6 beschriebenen Bakterien, können durch Zusammenlagern eine niederkoerzitive remanenteMagnetisierung tragen.
Die mittelkoerzitive Fraktion, mit Halbwertsfeldern von 40 - 60 mT, wird durch Einbereichsteilchen dominiert. Als Mineralphasen kommen auch hier primär nur Magnetit,Maghemit, Titanomagnetit oder Titanomaghemit in Betracht. Eine Unterscheidung nachTitangehalt und Oxidationsgrad kann nicht getroffen werden. Erst künftige thermomagnetische Messungen werden darüber Aufschluß geben können.
Die hochkoerzitive Fraktion, mit Halbwertsfeldern von 70 - 1000 mT, ist hauptsächlich Hämatit zuzuordnen, der Halbwertsfelder bis zu einigen hundert Millitesla besitzt.Nicht identifizieren lassen sich die Eisenhydroxide (u.a. Goethit) mit Halbwertsfeldern
von einigen Tesla. Sie können mit den verwendeten Feldstärken nicht von den paramagnetischen Mineralen getrennt werden. Eine Identifizierung dieser Phasen istebenfalls thermomagnetisch möglich.
v. DOBENECK (1993) ordnete dem biogenen Magnetit magnetotaktischer BakterienHalbwertsfelder von 25 bis 30 mT zu, wodurch sie sich von den anderen Einbereichsteilchen deutlich unterscheiden ließen. Für die flachen Stationen im Auftriebsgebiet vorNamibia ergaben sich solche Werte zwar bei der Hysteresesumme, nicht aber bei derHysteresedifferenz (Abb. 8.7, Tabelle 8.1). Dort wurden aber die höchsten Magnetbakterienkonzentrationen nachgewiesen und auch die elektronenmikroskopischen Aufnahmen zeigten hohe Konzentrationen an biogenem Magnetit. KIRSCHVINK (pers.Mitteilung, 1994) schlägt vor, diese Ergebnisse wie folgt zu interpretieren: In den hieruntersuchten Oberflächensedimenten sind die Ketten der Magnetbakterien noch überwiegend erhalten, während in den von V. DOBENECK (1993) analysierten Sedimentenaus größerer Tiefe die Ketten häufiger zerstört sind. Liegen die Magnetosome nichtmehr in Ketten, sondern in Clustern vor, dann gehen die Halbwertsfelder auf 25 bis30 mT zurück.
8.4 Hystereseparameter der Oberflächensedimente des Südatlantiks
Die Charakterisierung von Sedimenten anhand gesteinsmagnetischer Messungen kannnur in sehr einfachen Fällen eindeutig sein. Eine Vielzahl unterschiedlicher Parameter,wie die chemische Zusammensetzung, die Korngröße, die Kornform, innere Spannungen, die magnetische Vorgeschichte und Wechselwirkungen zwischen den Partikeln,beeinflussen die magnetischen Eigenschaften. Grundsätzlich muß deshalb eine
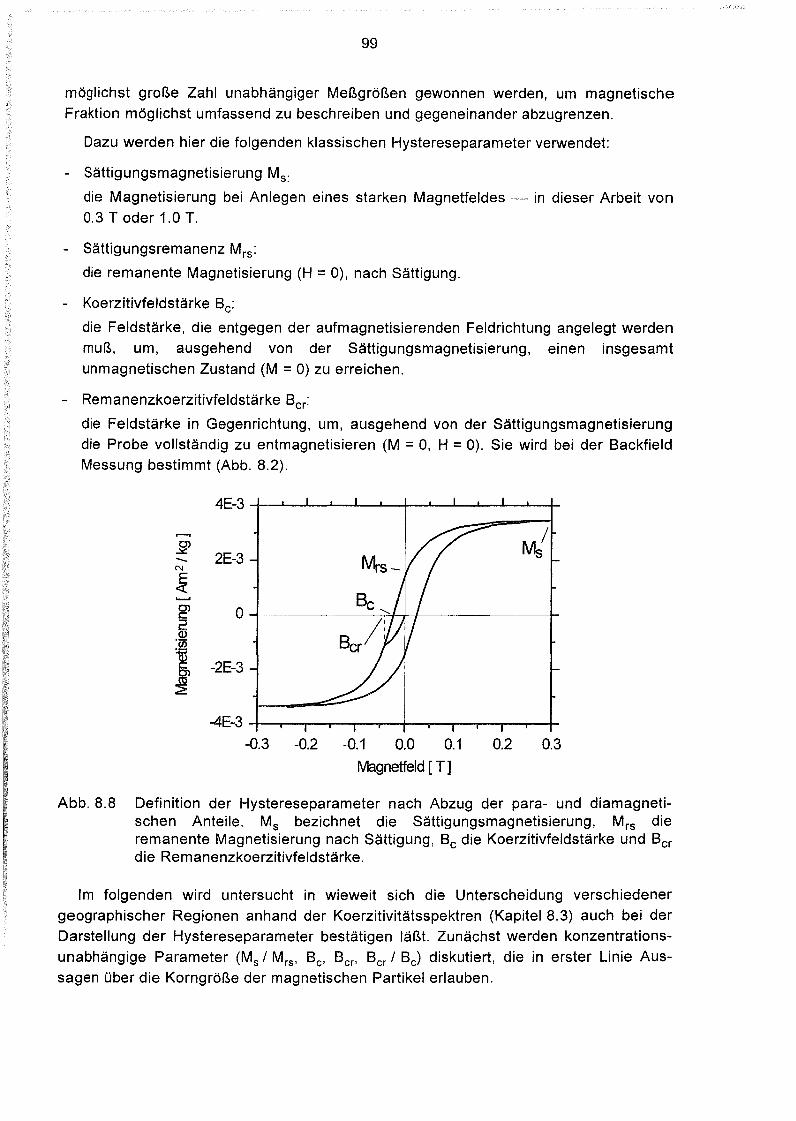
99
in dieser Arbeit von
möglichst große Zahl unabhängiger Meßgrößen gewonnen werden, um magnetischeFraktion möglichst umfassend zu beschreiben und gegeneinander abzugrenzen.
Dazu werden hier die folgenden klassischen Hystereseparameter verwendet:
- Sättigungsmagnetisierung Ms:
die Magnetisierung bei Anlegen eines starken Magnetfeldes0.3 T oder 1.0 T.
- Sättigungsremanenz Mrs :
die remanente Magnetisierung (H =0), nach Sättigung.
- Koerzitivfeldstärke Be:
die Feldstärke, die entgegen der aufmagnetisierenden Feldrichtung angelegt werdenmuß, um, ausgehend von der Sättigungsmagnetisierung, einen insgesamtunmagnetischen Zustand (M =0) zu erreichen.
- Remanenzkoerzitivfeldstärke Ber:
die Feldstärke in Gegenrichtung, um, ausgehend von der Sättigungsmagnetisierungdie Probe vollständig zu entmagnetisieren (M = 0, H = 0). Sie wird bei der BackfieldMessung bestimmt (Abb. 8.2).
0.30.2-0.1 0.0 0.1
I\Aagnetfeld [ T ]
-4E-3 -t--.-r--,..---.,..--r--r----r-.----r---r--.-t-
-0.3 -0.2
4E-3
~2E-3 Mf-- Mrs-N
~Cl 0c:2.~
~-2E-3c:
I
Abb.8.8 Definition der Hystereseparameter nach Abzug der para- und diamagnetischen Anteile. Ms bezichnet die Sättigungsmagnetisierung, Mrs dieremanente Magnetisierung nach Sättigung, Be die Koerzitivfeldstärke und Berdie Remanenzkoerzitivfeldstärke.
Im folgenden wird untersucht in wieweit sich die Unterscheidung verschiedenergeographischer Regionen anhand der Koerzitivitätsspektren (Kapitel 8.3) auch bei derDarstellung der Hystereseparameter bestätigen läßt. Zunächst werden konzentrationsunabhängige Parameter (Ms / Mrs ' Bc ' Bcp Bcr / Bc) diskutiert, die in erster Linie Aussagen über die Korngröße der magnetischen Partikel erlauben.
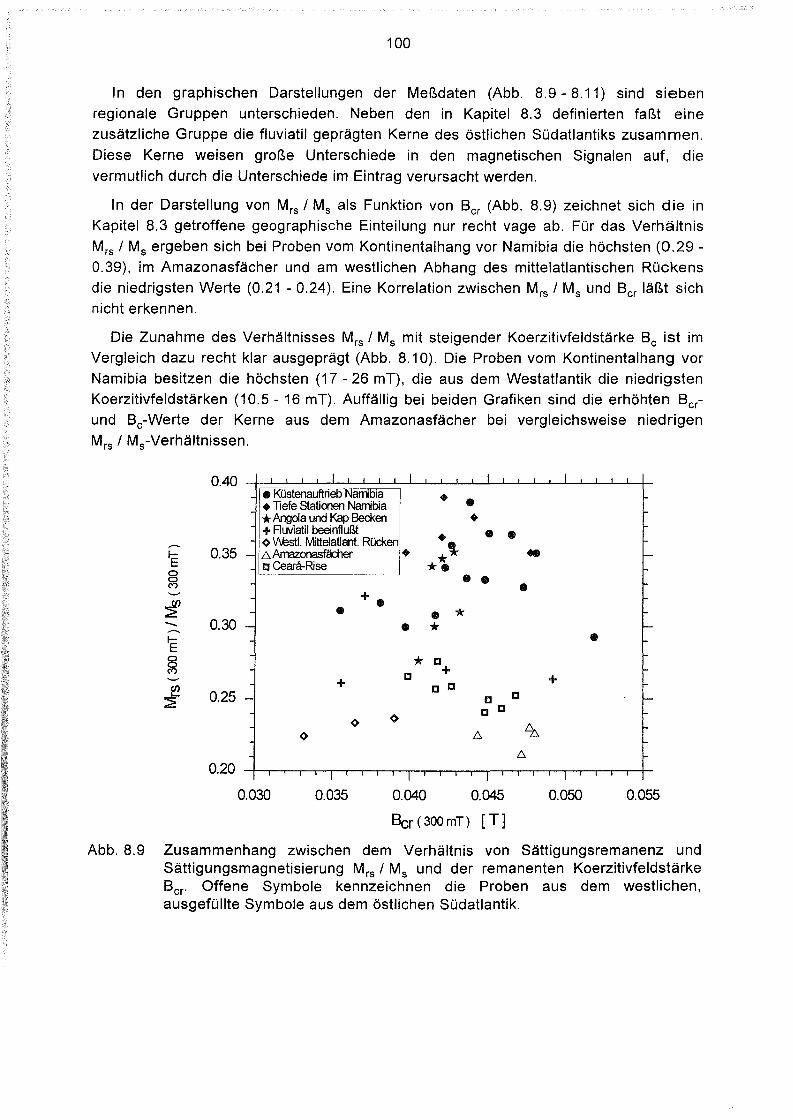
100
In den graphischen Darstellungen der Meßdaten (Abb. 8.9 - 8.11) sind siebenregionale Gruppen unterschieden. Neben den in Kapitel 8.3 definierten faßt einezusätzliche Gruppe die fluviatil geprägten Kerne des östlichen Südatlantiks zusammen.Diese Kerne weisen große Unterschiede in den magnetischen Signalen auf, dievermutlich durch die Unterschiede im Eintrag verursacht werden.
In der Darstellung von Mrs I Ms als Funktion von Ber (Abb. 8.9) zeichnet sich die inKapitel 8.3 getroffene geographische Einteilung nur recht vage ab. Für das VerhältnisMrs I Ms ergeben sich bei Proben vom Kontinentalhang vor Namibia die höchsten (0.29 0.39), im Amazonasfächer und am westlichen Abhang des mittelatlantischen Rückensdie niedrigsten Werte (0.21 - 0.24). Eine Korrelation zwischen Mrs I Ms und Ber läßt sichnicht erkennen.
Die Zunahme des Verhältnisses Mrs I Ms mit steigender Koerzitivfeldstärke Be ist imVergleich dazu recht klar ausgeprägt (Abb. 8.10). Die Proben vom Kontinentalhang vorNamibia besitzen die höchsten (17 - 26 mT), die aus dem Westatlantik die niedrigstenKoerzitivfeldstärken (10.5 - 16 mT). Auffällig bei beiden Grafiken sind die erhöhten Berund Be-Werte der Kerne aus dem Amazonasfächer bei vergleichsweise niedrigenMrs I Ms-Verhältnissen.
0.40 I I I I• Küstenauftrieb Nanibia •• liefe Stationen Nanibia •*Angola und Kap Becken •+Ruviatil beeinflußt • • •~ Vl.estl. Mitte/atlant. Rücken
*\'E 0.35 - ,6,.Amazonasfächer • .- I-aCeara-Rise *.8 • •(V) •
~+ •• • *- 0.30 - • * I-
'E •8 * a(V)
a ++ +
f!? 0.25 -a a
a:2 a I-
~a a
,6,. ~
,6,.0.20
I I I I
0.030 0.035 0.040 0.045 0.050 0.055
Ber (300 mT) [TlAbb. 8.9 Zusammenhang zwischen dem Verhältnis von Sättigungsremanenz und
Sättigungsmagnetisierung Mrs I Ms und der remanenten KoerzitivfeldstärkeBer. Offene Symbole kennzeichnen die Proben aus dem westlichen,ausgefüllte Symbole aus dem östlichen Südatlantik.
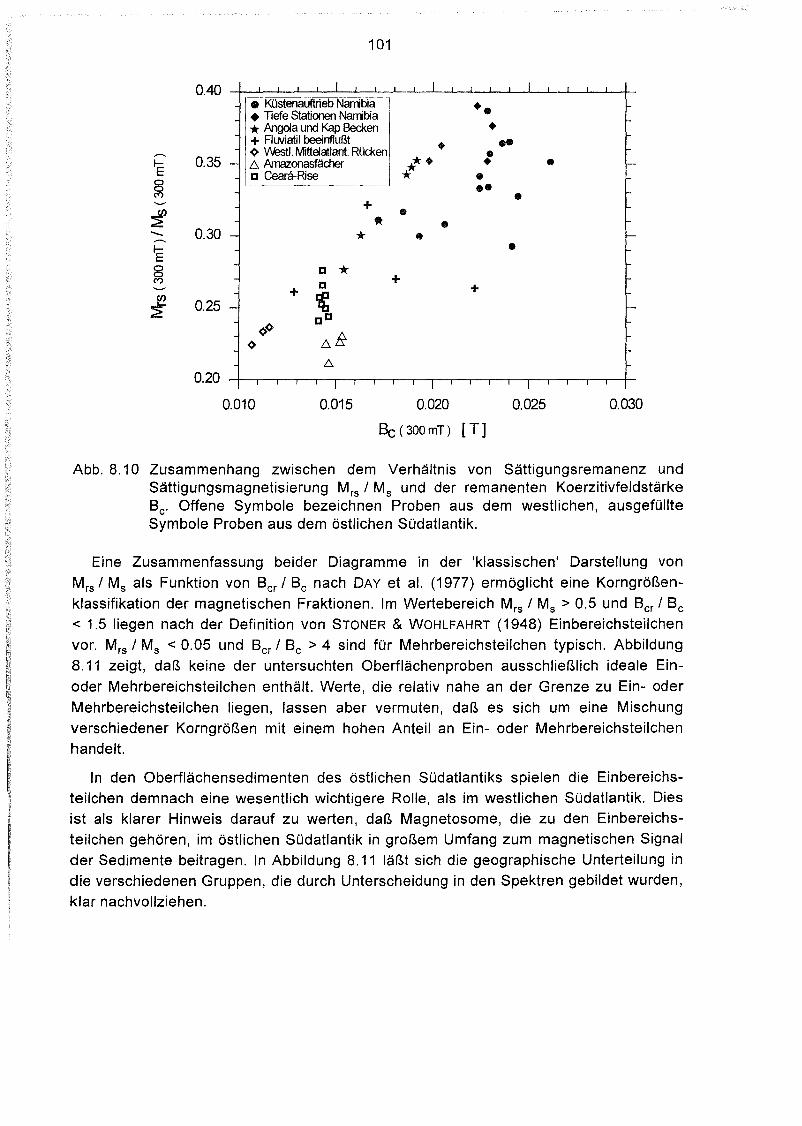
101
0.40 I I I
eKüstenauftrieb Nanlbla + •• Trefe Stationen Narribia* Angola und Kap Becken ++ FhNiatil beeinflußt + ..
0.35 -~ IJIkstI. Mttelatlant. Rücken
**+ •'E b. Amazonasfächer + •e Ceara-Rise *: •8 --_.._--,--------- ••C") •+ •~ • •- 0.30 - * •
'E •g e *e +
+ +~ 0.25 - ~ I-:2: ee
~~b.~~
b.0.20
I I I
0.010 0.015 0.020 0.025 0.030
Be (300mT) [Tl
Abb. 8.10 Zusammenhang zwischen dem Verhältnis von Sättigungsremanenz undSättigungsmagnetisierung Mrs / Ms und der remanenten KoerzitivfeldstärkeBc ' Offene Symbole bezeichnen Proben aus dem westlichen, ausgefüllteSymbole Proben aus dem östlichen Südatlantik.
Eine Zusammenfassung beider Diagramme in der 'klassischen' Darstellung vonM rs / Ms als Funktion von Bcr / Bc nach DAY et al. (1977) ermöglicht eine Korngrößenklassifikation der magnetischen Fraktionen. Im Wertebereich Mrs / Ms > 0.5 und Bcr / Be
< 1.5 liegen nach der Definition von STONER & WOHLFAHRT (1948) Einbereichsteilchenvor. Mrs / Ms < 0.05 und Bcr / Bc > 4 sind für Mehrbereichsteilchen typisch. Abbildung8.11 zeigt, daß keine der untersuchten Oberflächenproben ausschließlich ideale Einoder Mehrbereichsteilchen enthält. Werte, die relativ nahe an der Grenze zu Ein- oderMehrbereichsteilchen liegen, lassen aber vermuten, daß es sich um eine Mischungverschiedener Korngrößen mit einem hohen Anteil an Ein- oder Mehrbereichsteilchenhandelt.
In den Oberflächensedimenten des östlichen Südatlantiks spielen die Einbereichsteilchen demnach eine wesentlich wichtigere Rolle, als im westlichen Südatlantik. Diesist als klarer Hinweis darauf zu werten, daß Magnetosome, die zu den Einbereichsteilchen gehören, im östlichen Südatlantik in großem Umfang zum magnetischen Signalder Sedimente beitragen. In Abbildung 8.11 läßt sich die geographische Unterteilung indie verschiedenen Gruppen, die durch Unterscheidung in den Spektren gebildet wurden,klar nachvollziehen.
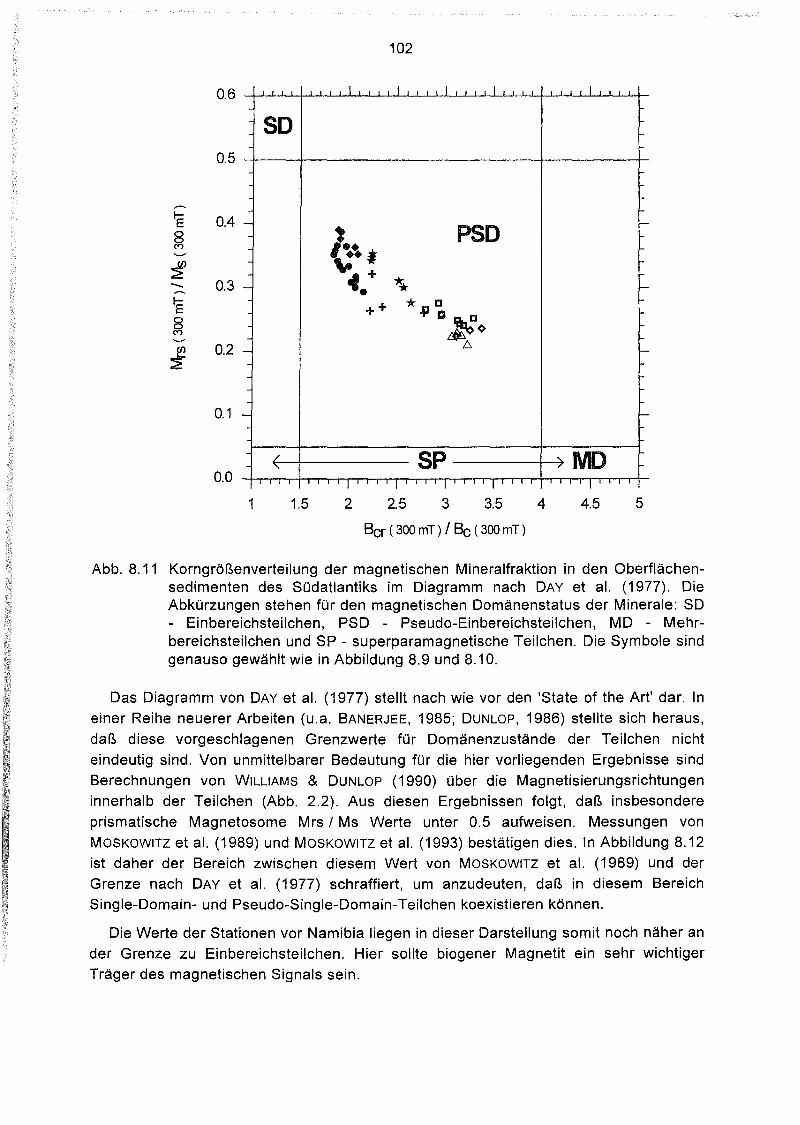
102
0.6
0.5
'E 0.48C"')
~- 0.3
'E8C"')
-I 0.2
0.1
0.0
I I I I I
SO
-) PSO I-,....\e++
- -e+**
I-•++ * [J
.p EI ~LJi;.~
t::..I--
- I-
/ SP .... MD.....
I I I I I
1 1.5 2 2.5 3 3.5 4
Ber (300 mT) I Be (300 mT)
4.5 5
Abb. 8.11 Korngrößenverteilung der magnetischen Mineralfraktion in den Oberflächensedimenten des Südatlantiks im Diagramm nach DAY et al. (1977). DieAbkürzungen stehen für den magnetischen Domänenstatus der Minerale: SO- Einbereichsteilchen, PSD - Pseudo-Einbereichsteilchen, MD - Mehrbereichsteilchen und SP - superparamagnetische Teilchen. Die Symbole sindgenauso gewählt wie in Abbildung 8.9 und 8.10.
Das Diagramm von DAY et al. (1977) stellt nach wie vor den 'State of the Art' dar. Ineiner Reihe neuerer Arbeiten (u.a. BANERJEE, 1985; DUNLOP, 1986) stellte sich heraus,daß diese vorgeschlagenen Grenzwerte für Domänenzustände der Teilchen nichteindeutig sind. Von unmittelbarer Bedeutung für die hier vorliegenden Ergebnisse sindBerechnungen von WILLIAMS & DUNLOP (1990) über die Magnetisierungsrichtungeninnerhalb der Teilchen (Abb. 2.2). Aus diesen Ergebnissen folgt, daß insbesondereprismatische Magnetosome Mrs I Ms Werte unter 0.5 aufweisen. Messungen vonMOSKOWITZ et al. (1989) und MOSKOWITZ et al. (1993) bestätigen dies. In Abbildung 8.12ist daher der Bereich zwischen diesem Wert von MOSKOWITZ et al. (1989) und derGrenze nach DAY et al. (1977) schraffiert, um anzudeuten, daß in diesem BereichSingle-Domain- und Pseudo-Single-Domain-Teilchen koexistieren können.
Die Werte der Stationen vor Namibia liegen in dieser Darstellung somit noch näher ander Grenze zu Einbereichsteilchen. Hier sollte biogener Magnetit ein sehr wichtigerTräger des magnetischen Signals sein.
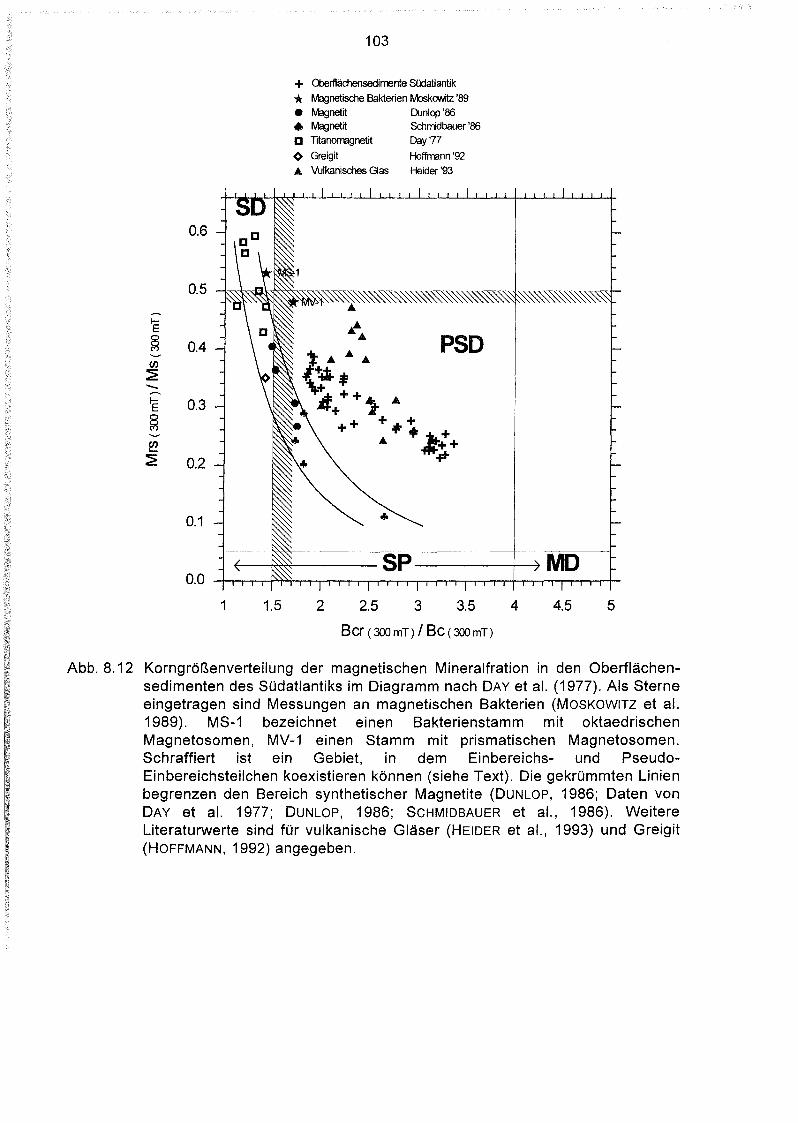
103
+ Cl>erflächensedirrente SOdatlantik* Wlagnetische Bakterien Moskowitz '89
• Wlagnetit DJnlop '86• Wlagnelit Sdmidbauer '86D Trtanomagnetit Day '77
~ Greigit Hoffmann '92... Vulkanisches Glas Heider '93
----~MD0.0 -h--r-r--.---f'
0.5~~~~~~~~~ ~~...
'E ......~
... PSD0.4 j .........Cf)
~
\~*-'E 0.3 4+++~ ...~ • ++ + +~ 4' +.l+~ ... ++~ 0.2 ++
0.6
0.1
1 1.5 2 2.5 3 3.5 4 4.5 5
Bcr (300mT) / Be (300mT)
Abb. 8.12 Korngrößenverteilung der magnetischen Mineralfration in den Oberflächensedimenten des Südatlantiks im Diagramm nach DAY et al. (1977). Als Sterneeingetragen sind Messungen an magnetischen Bakterien (MOSKOWITZ et al.1989). MS-1 bezeichnet einen Bakterienstamm mit oktaedrischenMagnetosomen, MV-1 einen Stamm mit prismatischen Magnetosomen.Schraffiert ist ein Gebiet, in dem Einbereichs- und PseudoEinbereichsteilchen koexistieren können (siehe Text). Die gekrümmten Linienbegrenzen den Bereich synthetischer Magnetite (DUNLOP, 1986; Daten vonDAY et al. 1977; DUNLOP, 1986; SCHMIDBAUER et al., 1986). WeitereLiteraturwerte sind für vulkanische Gläser (HElDER et al., 1993) und Greigit(HOFFMANN, 1992) angegeben.

104
8.5 Messung der anhysteretischen und der isothermalen Remanenz
Ein sehr sensibler Indikator für die Identifizierung magnetischer Einbereichsteilchen istdas Verhältnis der anhysteretischeri remanenten Magnetisierung (ARM) zur isothermalen remanenten Magnetisierung (IRM) (KING et al., 1982). Beide Parameter wurden anProben mit einem Volumen von etwa 10 cm3 gemessen. Einschränkungen, wie sieaufgrund der geringen Probengröße für die AGM-Messungen eine Rolle spielen, sindalso ohne Bedeutung. Die ARM wird den Proben im Erdmagnetfeld (0.04 mT) durch einLaborfeld aufgeprägt. Hier wurde eine Wechselfeldstärke von maximal 100 mT angelegt. Die IRM der Proben resultiert aus einem Gleichfeld von 100 mT. MagnetischeEinbereichsteilchen besitzen ein hohes, Mehrbereichsteilchen ein niedriges ARM / IRM
Verhältnis.
In der graphischen Darstellung des Verhältnisses ARM / IRM in Abhängigkeit vomVerhältnis Ber / Be lassen sich die Oberflächensedimente des östlichen und westlichenSüdatlantiks klar unterscheiden (Abb. 8.13). Das ARM / IRM-Verhältnis liegt im Westatlantik bei Werten zwischen 0.045 und 0.07, im Ostatlantik dagegen zwischen 0.065und 0.105. Für Proben aus dem östlichen Südatlantik wurden Ber / Be-Verhältnisse <2.2ermittelt, für solche aus dem westlichen Teil Ber / Be-Werte zwischen 2.8 und 3.4. DieErgebnisse der fluviatil beeinflußten Kerne aus dem östlichen Südatlantik liegen
zwischen diesen beiden Wertebereichen.
Die Oberflächensedimente des östlichen und westlichen Südatlantiks lassen sichebenso eindeutig durch das Verhältnis der Suszeptibilität der ARM zur SchwachfeldSuszeptibilität unterscheiden (Abb. 8.14). Die Suszeptibilität der ARM ist definiert alsQuotient der ARM und des Gleichfeldes, in dem die ARM erworben wurde.Ausgleichsgeraden ergeben ein durchschnittliches Verhältnis XARM / X von etwa 41, fürdie Proben aus dem Ostatlantik im Vergleich zu etwa 7 im Westatlantik. Werte um 50sind typisch für Einbereichsteilchen (R. THOMPSON, mdl. Mitt., 1994)
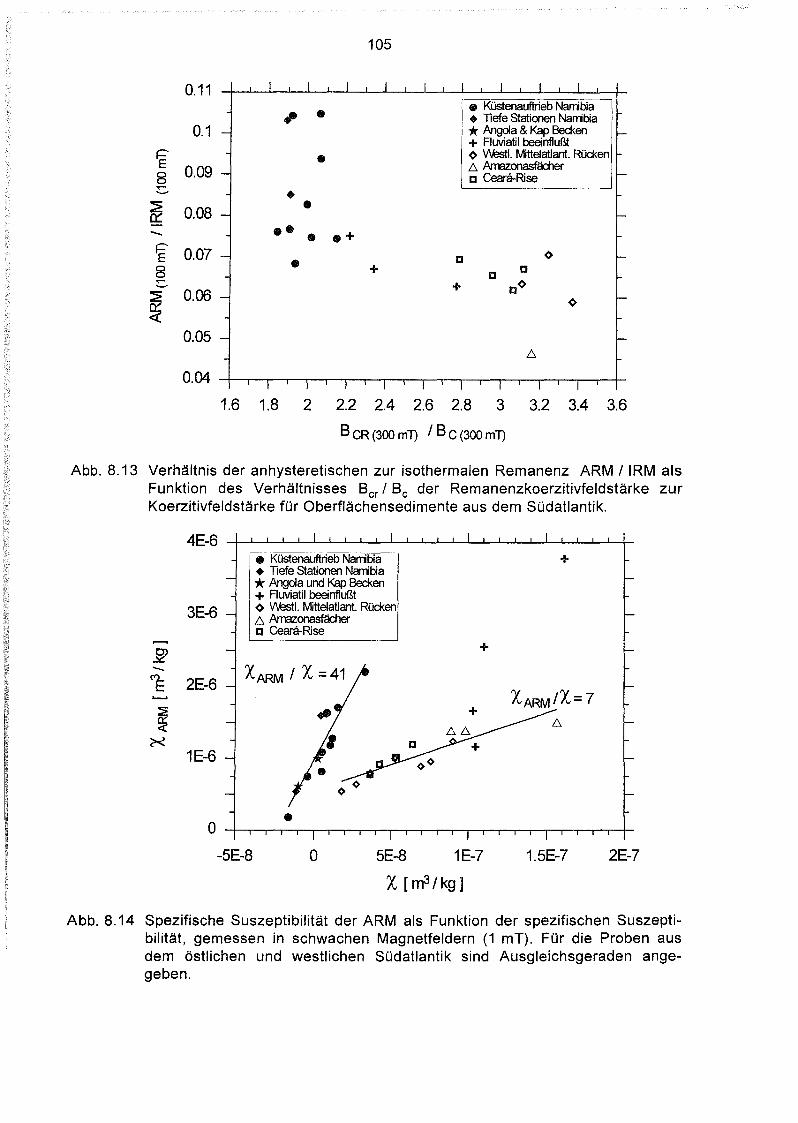
0.11
0.1 -
~0.09 -8
G
~ 0.08 --~ 0.07 -8G
~0.06 -
0.05 - -
3.61.6
0.04 -;---r--r--I-r-'I--r--r,-'---"1,""""'.-r--1 r--'-,,..--r-,"-,,--r-',---"1----1t-
1.8 2 2.2 2.4 2.6 2.8 3 3.2 3.4
BCR (300 m1) / BC (300 m1)
Abb. 8.13 Verhältnis der anhysteretischen zur isothermalen Remanenz ARM fiRM alsFunktion des Verhältnisses Ber f Be der Remanenzkoerzitivfeldstärke zurKoerzitivfeldstärke für Oberflächensedimente aus dem Südatlantik.
3E-6
~-1: 2E-6.......
~~
1E-6
• Küstenauftrieb Nanibia• liefe Stationen Nanibia* Angola und Kap Becken+ Ruviatil beeinflußt~ IAestI. Mittelatlant. Rücken!:":. AmazonasfächerIJ Geara-Rise
XARM / X=41
+
+
•2E-71.5E-75E-8 1E-7
X [m3/kg]
oO-t--,-......-r--r--,--.,--,---.--...--.----r--r--r--.--.,--,----r--r--.---,-......-r--r--r--I-
-5E-8
Abb. 8.14 Spezifische Suszeptibilität der ARM als Funktion der spezifischen Suszeptibilität, gemessen in schwachen Magnetfeldern (1 mT). Für die Proben ausdem östlichen und westlichen Südatlantik sind Ausgleichsgeraden angegeben.
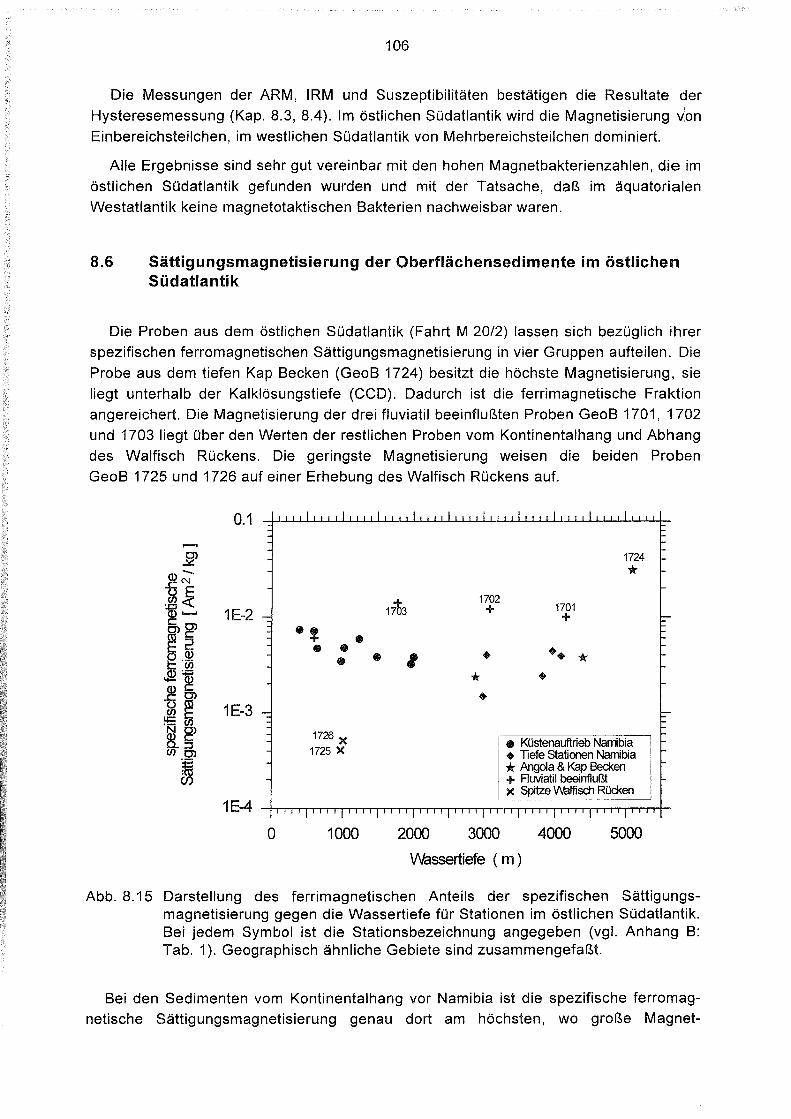
106
Die Messungen der ARM, IRM und Suszeptibilitäten bestätigen die Resultate derHysteresemessung (Kap. 8.3, 8.4). Im östlichen Südatlantik wird die Magnetisierung vonEinbereichsteilchen, im westlichen Südatlantik von Mehrbereichsteilchen dominiert.
Alle Ergebnisse sind sehr gut vereinbar mit den hohen Magnetbakterienzahlen, die im
östlichen Südatlantik gefunden wurden und mit der Tatsache, daß im äquatorialenWestatlantik keine magnetotaktischen Bakterien nachweisbar waren.
8.6 Sättigungsmagnetisierung der Oberflächensedimente im östlichenSüdatlantik
Die Proben aus dem östlichen Südatlantik (Fahrt M 20/2) lassen sich bezüglich ihrerspezifischen ferromagnetischen Sättigungsmagnetisierung in vier Gruppen aufteilen. DieProbe aus dem tiefen Kap Becken (GeoB 1724) besitzt die höchste Magnetisierung, sieliegt unterhalb der Kalklösungstiefe (CCD). Dadurch ist die ferrimagnetische Fraktionangereichert. Die Magnetisierung der drei fluviatil beeinflußten Proben GeoB 1701, 1702und 1703 liegt über den Werten der restlichen Proben vom Kontinentalhang und Abhangdes Walfisch Rückens. Die geringste Magnetisierung weisen die beiden ProbenGeoB 1725 und 1726 auf einer Erhebung des Walfisch Rückens auf.
i-. Küstenauftrieb NanibiaI • Tiefe Stationen Namibia* Angola &Kap Becken+ Ruviatil beeinflußtX Spitze V\alfisch Rücken
0.1
~0.>-"§N._~
1?t3~---. 1E-2
[~ .~ •• •g.~ • • I~~~01 1E-3q::~._ en~g> 1726 X
enS, 1725 X
~
1702+
*•
1701+
1724
*
o 1000 2000 3000 4000
Wassertiefe (m)
5000
Abb. 8.15 Darstellung des ferrimagnetischen Anteils der spezifischen Sättigungsmagnetisierung gegen die Wassertiefe für Stationen im östlichen Südatlantik.Bei jedem Symbol ist die Stationsbezeichnung angegeben (vgl. Anhang B:Tab. 1). Geographisch ähnliche Gebiete sind zusammengefaßt.
Bei den Sedimenten vom Kontinentalhang vor Namibia ist die spezifische ferromagnetische Sättigungsmagnetisierung genau dort am höchsten, wo große Magnet-
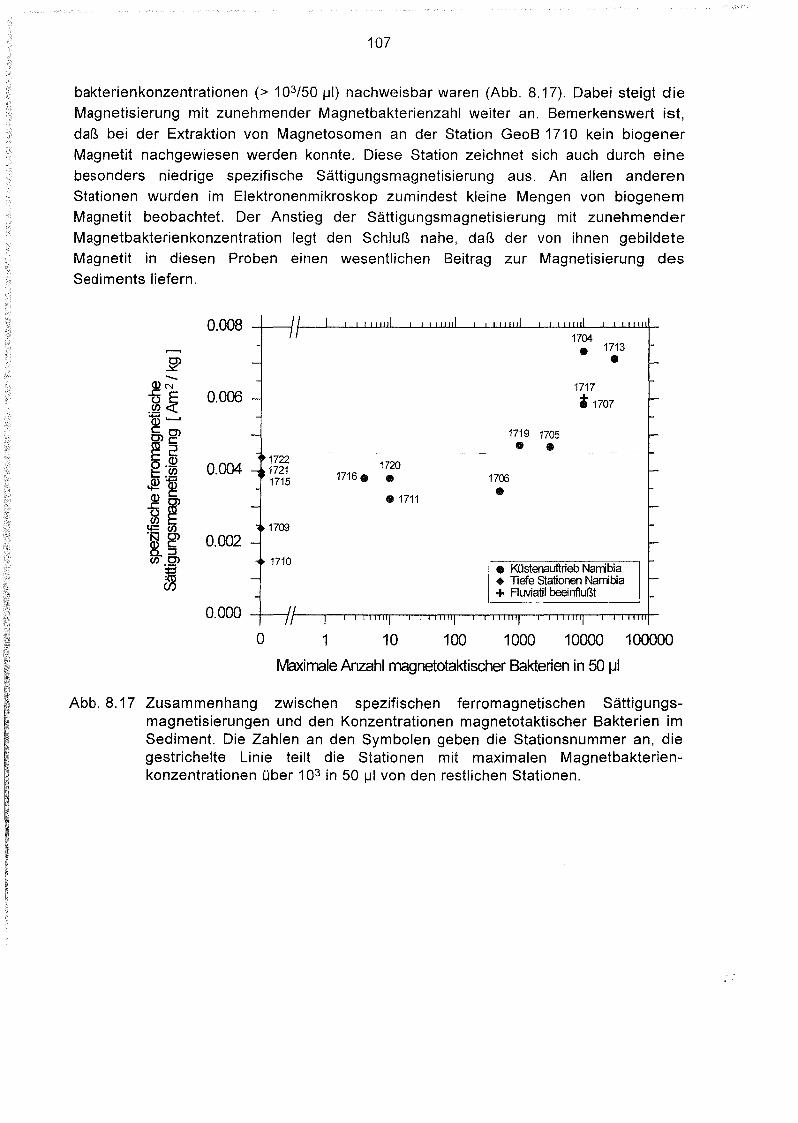
107
bakterienkonzentrationen (> 103/50 1-11) nachweisbar waren (Abb. 8.17). Dabei steigt dieMagnetisierung mit zunehmender Magnetbakterienzahl weiter an. Bemerkenswert ist,daß bei der Extraktion von Magnetosomen an der Station GeoB 1710 kein biogenerMagnetit nachgewiesen werden konnte. Diese Station zeichnet sich auch durch eine
besonders niedrige spezifische Sättigungsmagnetisierung aus. An allen anderenStationen wurden im Elektronenmikroskop zumindest kleine Mengen von biogenemMagnetit beobachtet. Der Anstieg der Sättigungsmagnetisierung mit zunehmenderMagnetbakterienkonzentration legt den Schluß nahe, daß der von ihnen gebildeteMagnetit in diesen Proben einen wesentlichen Beitrag zur Magnetisierung desSediments liefern.
1717
i 1707
1713
•
10000 100000
1719 1705
• •
1000
1706
•
I • Küstenauftrieb Nanibia
f
• Tiefe Stationen Nambia+ RlNiatil beeinflußt
10010
17201716. •
• 1711
1
1710
1709
172217211715
o
0.006
0.004
0.002
0.000
0.008
Maximale Anzahl magnetotaktischer Bakterien in 50 IJI
Abb. 8.17 Zusammenhang zwischen spezifischen ferromagnetischen Sättigungsmagnetisierungen und den Konzentrationen magnetotaktischer Bakterien imSediment. Die Zahlen an den Symbolen geben die Stationsnummer an, diegestrichelte Linie teilt die Stationen mit maximalen Magnetbakterienkonzentrationen über 103 in 50 1-11 von den restlichen Stationen.

108
9. Zusammenfassung
Das Vorkommen lebender magnetotaktischer Bakterien und ihrer fossilen magnetischen
Partikel wurde in Sedimenten des Nordseewatts und des Südatlantiks untersucht.
Zur Identifikation lebender Bakterien wurde eine Apparatur entwickelt, mit der an
einem Lichtmikroskop homogene Magnetfelder (bis 300 I-IT) beliebiger Richtung vorge
geben werden können. Die magnetotaktischen Bakterien bewegen sich parallel zu
diesen Magnetfeldern und können anhand ihres Schwimmverhaltens von anderen
Bakterien unterschieden werden. Aus den Bewegungsspuren läßt sich das magnetische
Moment einzelner Magnetbakterien bestimmen. Für zwei morphologisch deutlich
unterscheidbare kokkoide Formen wurde ein mittleres magnetisches Moment von
10-14 Am 2 bzw. 10-15 Am 2 ermittelt.
Vorarbeiten für die Untersuchungen im Südatlantik wurden im Nordseewatt durchge
führt. Magnetotaktische Bakterien kommen dort in den obersten Zentimetern der Watt
sedimente mit maximalen Konzentrationen von einigen 1000 Exemplaren in 50 1-11
Sedimentsuspension vor. Durch Vergleich der Tiefenverteilung der Magnetbakterien mit
Porenwasserprofilen von O2, N03-, Mn2+, Fe2+ und SO/- zeigte sich, daß die höchsten
Magnetbakterienkonzentrationen in der Nitratreduktionszone oder der obersten Eisen
reduktionszone vorkommen. In der Sulfatreduktionszone sind hingegen nur noch
vereinzelt Magnetbakterien nachweisbar. Die große Konzentration der Magnetbakterien
in der Eisenreduktionsschicht, in einer Tiefe, in der Nitrat nur noch in Spuren und
vermutlich nur lokal vorkommt, konnte hier erstmals nachgewiesen werden. Eisen
reduktion ist bei Magnetbakterien bisher nur einmal indirekt, als untergeordneter Stoff
wechselprozess beobachtet worden (GUERIN & BLAKEMORE, 1992).
Bei den Untersuchungen im Südatlantik konnten lebende Magnetbakterien erstmalig
in pelagischen Sedimenten nachgewiesen werden. Während vier Fahrten mit dem
Forschungsschiff METEOR wurde die Verteilung dieser Bakterien in den Oberflächen
sedimenten bestimmt. Ihre Anzahl nahm mit zunehmender Wassertiefe stark ab. In
Sedimenten aus Wassertiefen über 3000 m konnten nur einzelne «10/50 1-11), in Tiefen
über 4100 m gar keine Magnetbakterien gefunden werden. Es zeigten sich aber auch
deutliche regionale Unterschiede. Während sich im östlichen Südatlantik, speziell im
Küstenauftriebsgebiet vor Namibia, hohe Konzentrationen magnetotaktischer Bakterien
nachweisen ließen (bis zu 30 000/50 1-11), war ihre Zahl im westlichen Südatlantik
deutlich geringer. Es bestehen damit auch klare Zusammenhänge zwischen den
Magnetbakterienkonzentrationen und den Gehalten an organischem Material. Mit
steigendem Corg - und Gesamtstickstoffgehalt nimmt die Magnetbakterienzahl zu. Die
gleiche Entwicklung ist auch bei Zunahme des diffusiven Flusses von Sauerstoff in das
Sediment zu beobachten.
Lebende Magnetbakterien kamen nur in den obersten 15 cm der Sedimentsäule vor,
wobei die überwiegende Mehrzahl in den obersten 5 cm lebte. Bezüglich der Tiefen
verteilung der Magnetbakterien in der Sedimentsäule lassen sich zwei Typen von
Sedimenten unterscheiden. In den aeroben Sedimenten der nährstoffarmen Gebiete
konnten, wenn überhaupt, nur geringe Anzahlen magnetotaktischer Bakterien
beobachtet werden und zwar nur in den obersten zwei Zentimetern. In nährstoffreichen

109
Arealen konnten die höchsten Magnetbakterienkonzentrationen dagegen in den tieferenanaeroben Schichten nachgewiesen werden. Die Magnetbakterienzahl erreichte indiesen Sedimenten teilweise sehr hohe Werte (bis zu 30000/50 1J1).
Fossile Magnetosome konnten im Gegensatz zu den lebenden magnetotaktischen
Bakterien in fast allen Oberflächensedimenten des Südatlantiks nachgewiesen werden.Die Identifikation erfolgte nach magnetischer Extraktion der Sedimente imTransmissions-Elektronenmikroskop anhand ihrer charakteristischen Morphologie, dereng begrenzten Korngröße und, falls diese erhalten war, anhand der Kettenstruktur.Generell läßt sich festhalten, daß in den Proben aus dem östlichen Südatlantik mehr
Magnetosome vorkommen, als im äquatorialen Westatlantik, eine genauere Bestimmung der Konzentration ist aber kaum möglich.
Die relativen Anteile verschiedener Magnetosomformen lassen sich recht zuverlässig
bestimmen. Obwohl die sehr kleinen Magnetosome praktisch beliebig weit verfrachtet,werden können, wurden in den nährstoffreichen Sedimenten im zentralenAuftriebsgebiet vor Namibia überwiegend prismatische, in nährstoffarmen Sedimentendes offenen Ozeans überwiegend oktaedrische Magnetosome gefunden.
Von den acht bisher in Kultur vorliegenden Magnetbakterienstämmen sind drei aerobbzw. mikroaerophil; diese Bakterien besitzen oktaedrische Magnetosome. Die anderenfünf Stämme sind (fakultativ) anaerob und besitzen prismatische bzw. projektilförmigeMagnetosome. Da in den nährstoffreichen Sedimenten die maximalen Magnetbakterien
zahlen in den anaeroben Sedimentschichten, in den nährstoffarmen Sedimenten
dagegen in den aeroben Schichten anzutreffen waren, scheint ein Zusammenhangzwischen Magnetosomform und Stoffwechsel des Bakteriums zu bestehen. Falls sichdieser Zusammenhang auch in weiteren Untersuchungen bestätigt, könnte die Magnetosomform als Indikator für Paläo-Redoxbedingungen und Paläoproduktivitäten benutzt
werden.
Die Auswertung der gesteinsmagnetischen Untersuchungen (Hysterese- undBackfield Messung, Bestimmung der anhysteretischen und der isothermalenremanenten Magnetisierung sowie der Suszeptibilität) ergaben, daß sich anhand der
gesteinsmagnetischen Parameter sechs Regionen unterscheiden lassen: westlicherAbhang des mittelatlantischen Rückens, Amazonasfächer, Ceara Rise, Angola Beckenbzw. Kap Becken, unterer Kontinentalhang vor Namibia, oberer Kontinentalhang vorNamibia. Die magnetischen Minerale in den Sedimenten des östlichen Südatlantiksbesitzen eine kleinere durchschnittliche Korngröße als die der Sedimente desäquatorialen Westatlantiks. Der höhere Anteil der feinkörnigeren magnetischen Fraktionin den Sedimenten des östlichen Südatlantiks legt den Schluß nahe, daß dieMagnetosome dort einen größeren Anteil an der Magnetisierung haben als imäquatorialen Westatlantik. Die gesteinsmagnetischen Untersuchungen können abernicht sicher zwischen Magnetosomen und anderen Einbereichsteilchen unterscheiden.In den Sedimenten vom Kontinentalhang vor Namibia war die Konzentration der magnetischen Einbereichsteilchen am höchsten. In diesem Bereich wiesen genau die Kernedie höchste Sättigungsmagnetisierung auf, in denen die größten Magnetbakterienzahlengefunden wurden. Im oberen Teil des Kontinentalhanges vor Namibia scheinen dieMagnetosome tatsächlich die wichtigsten Träger der Magnetisierung zu sein.

110
10 Literaturverzeichnis
ANDERSEN, N. (1994), Biogeochemische Charakterisierung von Sinkstoffen und
Sedimenten aus ostatlantischen Produktionssystemen mit Hilfe von
Biomarkern. Berichte aus dem Fachbereich Geowissenschaften, Universität
Bremen, (in Vorb.).
ARAKI, T. (1992), Changes in rates of synthesis of individual proteins in a psychrophilic
bacterium after a shift in temperature. Canadian Journal of Microbiology, 37,
840-847.
ARTYMIUK, P.J., BAUMINGER, E.R., HARRISON, P.M., LAWSON, D.M., NOWIK, 1., TREFFRY,
A. & YEWDALL, S.J. (1990), Ferritin: A model system for iron biomineralisation.
In: Frankei, R.B. & Blakemore, R.P. (Eds.), Iron Biominerals. Plenum Press,
New York, 269-294.
BALKWILL, D.L., MARATEA, D. & BLAKEMORE, R.P. (1980), Ultrastructure of a
magnetotactic spirillum. Journal of Bacteriology, 141, 1399-1408.
BAZYLINSKI, D.A. & BLAKEMORE, R.P. (1983), Denitrification and assimilatory nitrate
reduction in Aquaspirillum magnetotacticum. Applied and Environmental
Microbiology, 46, 1118-1124.
BAZYLINSKI, D.A. (1990), Anaerobic production of single-domain magnetite by the
marine, magnetotactic bacterium, strain MV-1. In: Frankei, R.B. & Blakemore,
R.P. (Eds.), Iron Biominerals. Plenum Press, New York, 69-77.
BAZYLINSKI, D.A., FRANKEL, R.B. & JANNASCH, H. (1988), Anaerobic magnetite
production by a marine, magnetotactic bacterium. Nature, 334, 518-519.
BERGER, W.H. (1989), Global maps of ocean productivity. In: Berger, W.H., Smetacek,
V.S. & Wefer, G. (Eds.), Productivity of the ocean: present and past. Dahlem
Workshop Reports, J. Wiley and Sons, Chicester, 429-455.
BERGER, W.H., FISCHER, K., LAI, C., & Wu, G. (1987), Ocean productivity and organic
carbon flux. I. Overview and maps of primary production and export production,
University of California, San Diego, SIO Reference 87-30.
BETZER, P.R., SHOWERS, W.J., LAWS, E.A., WINN, C.D., DITuLLlO, G.R. & KROOPNICK,
P.M. (1984), Primary productivity and particle fluxes on a transect of the
Equator at 153°Win the Pacific Ocean. Deep-Sea Research, 31,1-11.
BLAKEMORE, R.P. (1975), Magnetotactic bacteria. Science, 190,377-379.
BLAKEMORE, R.P. (1982), Magnetotactic bacteria. Annual Review of Microbiology, 36,
217-238.
BLAKEMORE, R.P., FRANKEL, R.B. & KALMIJN, A.J. (1980), South-seeking magnetotactic
bacteria in the southern hemisphere. Nature, 286, 384-385.

111
BLAKEMORE, RP., MARATEA, D. & WOLFE, RS. (1979), Isolation and pure culture of a
freshwater magnetic spirillum in chemically defined medium. Journal of
Bacteriology, 140, 720-729.
BLAKEMORE, RP., SHORT, K.A, BAZYLINSKI, D.A, ROSENBLATT, C. & FRANKEL, RB.
(1985), Microaerobic conditions are required for magnetite formation within
Aquaspirillum magnetotacticum. Geomicrobiology Journal, 4, 53-71.
BLEIL, U., SPIESS, V. & WEFER, G. (1994), Geo Bremen SOUTH ATLANTIC 1993, Cruise
No. 23, 4 February - 12 April 1993, METEOR-Berichte, Universität Hamburg,
94-1, 261.
BUTLER, & BANERJEE, S.K. (1975), Theoretical single-domain grain size range in
magnetite and titanomagnetite. Journal of Geophysical Research, 80, 4049
4058.
CAMPBELL, J.W. & AARUP, T. (1992), New production in the North Atlantic derived from
seasonal patterns of surface chlorophyll. Deep-Sea Research, 39, 1669-1694.
CANFIELD, D.E. & BERNER, R.A (1987), Dissolution and pyritization of magnetite in
anoxie marine sediments. Geochimica et Cosmochimica Acta, 51, 645-659.
CHANG, S.-B.R, STOLZ, J.F., KIRSCHVINK, J.L. & AWRAMIK, S.M. (1989), Biogenie
magnetite in stromatolites. 11. Occurrence in ancient sedimentary environments.
Precambrian Research, 43, 305-315.
CHERKIS, N.Z., FLEMING, H.S. & BROZENA, J.M. (1989), Bathymetryof the South Atlantic
Ocean 3° S to 40° S, Williams & Heintz Map Corporation, Capitol Heights.
COLLINSON, D.W. (1983), Methods in rock magnetism and palaeomagnetism, techniques
and instrumentation. Chapman and Hall, London - New York, 503.
DAY, R, FULLER, M. & SCHMIDT, V.A (1977), Hysteresis properties of titanomagnetites:
grain-size and compositional dependence. Physics of the Earth and Planetary
Interiors, 13, 260-267.
DELONG E.F. & YAYANOS AA (1985), Adaption of the membrane lipids of a deep-sea
bacterium to changes in hydrostatic pressure. Science, 228, 1101-1102.
DELoNG E. F. & YAYANOS AA (1986), Biochem ical function and ecological significance
of novel bacteria lipids in deep-sea procaryotes. Applied and Environmental
Microbiology, 51, 730-737.
DELoNG, E.F., Frankei, RB. & Bazylinski, D.A (1993), Multiple evolutionary origins of
magnetotaxis in bacteria. Science, 259, 803-806.
DEMING J.W. (1986), Ecological strategies of barophilic bacteria in the deep ocean.
Microbiological Sciences, 3, 205-211.
DEMITRACK, A (1985), A search for bacterial magnetite in sediments of Eel Marsh,
Woods Hole. In: Kirschvink, J.L., Jones, D.D. & MacFadden, B.J., (Eds),
Magnetite Biomineralization and Magnetoreceptionin Organisms. Plenum Press,
New York, 625-645.

112
DUNLOP, D.J. (1986), Hysteresis properties of magnetite and their dependence on
particle size: A test of pseudo-single-domain remanence models. Journal of
Geophysical Research, 91, 9569-9584.
ENGELHARDT, H., SCHUSTER, S.C. & BÄUERLEIN, E. (1993), An archimedian spiral: The
basal disk of the Wolinella flagellar motor. Science, 362, 1046-1048.
ERICSON, D.B. & WOLLIN, G. (1968), Pleistocene c1imats and chronology in deep-sea
sediments. Science, 162,1227-1234.
FARINA, M., ESQUIVEL, D.M.S. & L1NS DE BARROS, H.G.P. (1990), Magnetotactic iron
sulfur crystals from a magnetotactic microorganism. Nature, 343, 256-258.
FASSBINDER, J.W.E., STANJEK, H. & VALI, H. (1990), Occurence of magnetic bacteria in
soil. Nature, 343,161-163.
FISCHER, J. & MOSER, H. (1956), Die Nachbildung von Magnetisierungskurven durch
einfache algebraische oder transzendente Funktionen. Archiv für Elektrotechnik,
42, 286-299.
FISCHER, J. (1990), Geologische Kartierung des südlichen Wurster Watts - Kartierung
Weddewarden. Unveröffentlichte Diplom-Arbeit, Fachbereich
Geowissenschaften der Universität Bremen.
FRANKEL, RB. (1984), Magnetic guidance of organims. Annual Review of Biophysics
and Bioengeneering, 13, 85-103.
FRANKEL, RB., BLAKEMORE, RP. & WOLFE, RS. (1979), Magnetite in freshwater
magnetotactic bacteria. Science, 203, 1355-1356.
FRANKEL, RB., BLAKEMORE, RP., TORRES DE ARAUJO, F.F., ESQUIVEL, D.M.S. & DANON,
J. (1981), Magnetotactic bacteria at the geomagnetic equator. Science, 212,
1296-1270.
FRANKEL, RB., PAPAEFTHYMIOU, G.C., BLAKEMORE, RB. & O'BRIEN, W. (1983), Fe304
precipitation in magnetotactic bacteria. Biochimica et Biophysica Acta, 763, 147
159.
FRITSCHE, W. (1990), Mikrobiologie. UTB Taschenbuch, Fischer, Jena, 497.
FROELlCH, P.N., KLiNKHAMMER, G.P., BENDER, M.L., LUEDTKE, N.A., HEATH, G.R,
CULLEN, 0., HARTMAN, B. & MAYNARD, V. (1979), Early oxidation of organic
matter in pelagic sediments of the Eastern Equatorial Atlantic: Suboxic
Diagenesis. Geochimica et Cosmochimica Acta, 43, 1075-1090.
GERLACH, D. (1985), Das Lichtmikroskop. Thieme, Stuttgart - New York, 571.
GLUD, RN., GUNDERSEN, J.K., J0RGENSEN, B.B., REVSBECH, N.P. & Schulz, H.D. (1994),
Diffusive and total oxygen uptake of deep-sea sediments in the south east
Atlantic Ocean: in situ and laboratory measurements. Deep-Sea Research, (in
press).
GORBY, Y.A., BEVERIDGE, T.J. & BLAKEMORE, RP. (1988), Characterization of the
Bacterial magnetosome membrane. Journal of Bacteriology, 170, 834-841.

113
GUERIN, W.F. & BLAKEMORE, RP. (1992), Redox cycling of iron supports growth and
magnetite synthesis by Aquaspirillum magnetotacticum. Applied and
Environmental Microbiology, 58, 1102-1109.
GUNDERSEN J.K. & GLUD RN. (1994), In situ oxygen Dynamics and pH-profiles. In:
BLEIL, U., SPIESS, V. & WEFER, G. (Eds.), Geo Bremen SOUTH ATLANTIC
1993, Cruise No. 23,4 February - 12 April 1993. METEOR-Berichte, Universität
Hamburg, 94-1,159-165.
HALGEDAHL, S.L. & FULLER, M. (1980), Magnetic domain observations of nucleation
processes in fine particles of intermediate titanomagnetite. Nature, 288, 70-72.
HElDER, F., KÖRNER, U. & BITSCHENE, P. (1993), Volcanic ash particles as carriers of
remanent magnetization in deep-sea sediments from the Kerguelen Plateau.
Earth and Planetary Science Letters, 118, 121-134.
HEYWOOD, B.R, BAZYLINSKI, D.A, GARRAT-REED, A, MANN., S. & FRANKEL, RB. (1990),
Controlled biosynthesis of greigite (Fe3S4) in magnetotactic bacteria.
Naturwissenschaften, 77, 536-538.
HEYWOOD, B.R, MANN, S. & FRANKEL RB. (1991), Structure, morphology and growth of
biogenie greigite (Fe3S4)' Symposium Proceedings of the Materials Research
Society, 218,93-108.
HOFFMANN, V. (1992), Greigite (Fe3S4): Magnetic properties and first domain
observations. Physics of the Earth and Planetary Interiors, 70, 288-301.
JANNASCH H.W. & TAYLOR C.D. (1984), Deep-sea microbiology. Annual Reviews in
Microbiology, 38, 487-514.
KING, J., BANERJEE, S.K., MARVIN, J. & ÖZDEMIR, Ö. (1982), A comparison of different
magnetic methods for determining the relative grain size of magnetite in natural
materials: some results from lake sediments. Earth and Planetary Science
Letters, 59, 404-419.
KIRSCHVINK, J.L. & LOWENSTAM, H.A (1979), Mineralization and magnetization of chiton
teeth: Palaeomagnetic, sedimentological and biologie implications of organic
magnetite. Earth and Planetary Science Letters, 44, 193-204.
KREBS, W. (1991), Grauwertbilder auf dem Laserdrucker, Großes Arbeitsvohaben,
Bildverarbeitung. Institut für digitale und theoretische Bildverarbeitung,
Universität Bremen, 75.
LOCHTE, K. & TURLEY, C.M. (1988), Bacteria and cyanobacteria associated with
phytodetritus in the deep sea. Nature, 333, 67-69.
LOVLEY, D.R, STOLZ, J.F., NORD, G.L., Jr. & PHILLlPS, E.J.P. (1987), Anaerobic
production of magnetite by a dissimilatory iron-reducing microorganism. Nature,
330, 252-254.

114
LÜNEBURG, H., SCHAUMANN, K. & WELLERSHAUS, S. (1975), Physiographie des Weser
Ästuars (Deutsche Bucht). Veröff. Inst. Meeresforsch. Bremerhaven, 15, 195
226.
Luo, J. H. (1992), Extracting and characterizing luciferase fram Photobacterium
phosphoreum. Chinese Journal of Microbiology and Immunology (Bejing), 12,
25-28.
LUTJEHARMS, J.RE. & STOCKTON, P.L. (1987), Kinematics of the upwelling front off
Southern Africa, In: Payne, AI.L., Gullard, J.A & Brink, K.H., The Benguela and
comparable ecosystems. South African Journal of Marine Sciences, 5, 35-49.
MANN,S., SPARKS, N.H.C. & BOARD, RG. (1990a), Magnetotactic bacteria: microbiology,
biomineralisation, palaeomagnetism and biotechnology. Advances in Microbial
Physiology, 31, 125-181.
MANN,S., SPARKS, N.H.C., FRANKEL, RB., BAZYLINSKI, D.A, JANNASCH, H.W. (1990b),
Biomineralisation of ferrimagnetic greigite (Fe3S4) an iron pyrite (FeS2) in a
magnetotactic bacterium. Nature, 343, 258-261.
MATSUDA, T, ENDO, J., OSAKABE, N., TONOMURA, A & ARII, T (1983), Morphology and
structure of biogenic magnetite particles. Nature, 302, 411-412.
MATSUNAGA, T., SAKAGUCHI, T & TADOKORO, F. (1991), Magnetite formation by a
magnetic bacterium capable of growing aerobically. Applied and Environmental
Biotechnology, 35, 651-655.
McNEILL, D.F. (1990), Biogenic magnetite from surface holocene carbonate sediments,
Great Bahama Bank. Journal of Geophysical Research, 95, 4363-4371.
MELDRUM, F.C., MANN, S., HEYWOOD, BR., FRANKEL, RB. & BAZYLINSKI, D.A (1993a),
Electron micrascopy study of magnetosomes in a cultured coccoid
magnetotactic bacterium. Proceedings of the Royal Society of London, Series B,
251, 231-236.
MELDRUM, F.C., MANN, S., HEYWOOD, B.R, FRANKEL, RB. & BAZYLINSKI, D.A (1993b),
Electron microscopy study of magnetosomes in two cultured vibrioid
magnetotactic bacteria. Proceedings of the Royal Society of London, Series B,
251, 237-242.
MOENCH, T.T. (1988), Bilophococcus magnetotacticus gen. nov. sp. nov., a motile,
magnetic coccus. Antonie van Leeuwenhoek, 54, 483-496.
MOON, T & MERRILL, RT (1985), Nucleation theory and domain states in multidomain
magnetic material. Physics of the Earth and Planetary Interiors, 37, 214-222.
MOSKOWITZ, B.M., FRANKEL RB., BAZYLINSKI, D.A, JANNASCH H.W. & LOVLEY D.R
(1989), A comparison of magnetite particles produced anaerobically by
magnetotactic and dissimilatory iron-reducing bacteria. Geophysical Research
Letters, 16, 665-668.

115
MOSKOWITZ, B.M., FRANKEL, RB. & BAZYLINSKI, D.A. (1993), Rock magnetic criteria for
the detection of biogenic magnetite. Earth and Planetary Science Letters, 120,
283-300.
MÜLLER, P.J. (1977), C/N ratios in Pacific deep-sea sediments: Effect of inorganic
ammonium and organic nitrogen compounds sorbed by clays. Geochimica et
Cosmochimica Acta, 41, 765-776.
O'REILLY, W. (1984), Rock and mineral magnetism. Chapman and Hall, New York, 223,
73.
OFER, S., NOWIK, 1., BAUMINGER, ER., PAPAEFTHYMIOU, G.C., FRANKEL, RB. &
BLAKEMORE, RB. (1984), Magnetosome dynamics in magnetotactic bacteria.
Biophysical Journal, 46, 57-64.
PAOLETTI, L.C. & BLAKEMORE, RB. (1986), Hydroxamate production by Aquaspirillum
magnetotacticum. Journal of Bacteriology, 167, 73-76.
PAOLETTI, L.C. & BLAKEMORE, RB. (1988), Iron reduction by Aquaspiril/um
magnetotacticum. Current Microbiology, 17, 339-342.
PETERMANN, H. (1989), Bestimmung des magnetischen Moments magnetotaktischer
Bakterien. Unveröffentlichte Diplomarbeit, Institut für Allgemeine und
Angewandte Geophysik, Universität München, 75.
PETERMANN, H., WEISS, D.G., BACHMANN, L. & PETERSEN, N. (1990), Motile behaviour
and determination of the magnetic moment of magnetic bacteria in rotating
magnetic fields. In: Alt, W. & Hoffmann, G. (Eds.), Biological Motion. Lecture
Notes in Biomathematics, 89, Springer, Berlin, 387-395.
PETERSEN, N. & VALI , H. (1987), Observation of shrinkage cracks in ocean floor
titanomagnetites. Physics of the Earth and Planetary Interiors, 46, 197-205.
PETERSEN, N., V. DOBENECK, T. & VALI, H. (1986), Fossil bacterial magnetite in deep-sea
sediments from the South Atlantic Ocean. Nature, 320, 611-615.
PETERSEN, N., WEISS, D.G. & VALI , H. (1989), Magnetic bacteria in lake sediments. In:
F.J. Lowes et al. (Eds.), Geomagnetism and Paleomagnetism. Kluwer Academic
Publishers, 231-241.
PYE, K. (1987), Aeolian dust and dust deposits, Academic Press Inc. London, 334, 78.
REDFIELD, A.C., KETCHUM, B.H. & RICHARDS, F.A. (1963), The influence of organic
matter on the composition of the sea water,.ln: Hili, M.N. (Ed.), The Sea.
Interscience, 2, New York, 26-77.
REINECK, H.E. (1978), Das Watt, Ablagerungsraum und Lebensraum, Kramer Frankfurt/
Main, 185.
RIVAS, J., ZAMARRO, J.M., MARTIN, E. & PEREIRA, C. (1981), Simple approximation for
magnetization curves and hysteresis loops. IEEE Transactions on Magnetics,
17,1498-1501.

116
RODGERS, F.G., BLAKEMORE, RP., BLAKEMORE, N.A, FRANKEL, RB., BAZYLINSKI, D.A.,
MARATEA, D. & RODGERS, C. (1990), Intercellular structure in a many-celled
magnetotactic prokaryote. Archives of Microbiology, 154, 18-22.
ROHE, K.H. (1983), Elektronik für Physiker. B.G. Teubner Studienbücher, Stuttgart, 248.
SAGEMANN, J. (1990), Geologische Kartierung des südlichen Wurster Watts - Kartierung
Weddewarden. Unveröffentlichte Diplom-Arbeit, Fachbereich
Geowissenschaften der Universität Bremen.
SAGEMANN, J., (in Vorb.), Saisonale Variationen von Porenwasserprofilen,
Nährstoffflüssen und Reaktionen in intertidalen Sedimenten des Weser-Ästuars.
Berichte aus dem Fachbereich Geowissenschaften der Universität Bremen.
SAKAGUCHI, T, BURGESS, J.G. & MATSUNAGA, T (1993), Magnetite formation by a
sulfate-reducing bacterium. Nature, 365,47-49.
SCHLEGEL, H.G. (1985), Allgemeine Mikrobiologie. Thieme, Stuttgart-New York, 571.
SCHLEIFER, K.H., SCHÜLER, D., SPRING S., WEIZEGGER, M., AMANN, R, LUDWIG, W. &
KÖHLER, M. (1991), The genus magnetospirillum gen. nov. description of
Magnetospirillum gryphiswaldense sp. nov. and transfer of Aquaspirillum
magnetotacticum to Magnetospirillum magnetotacticum combo nov.. Systematic
and Applied Microbiology, 14, 379 - 385.
SCHMIDBAUER, E. & SCHEMBERA, N. (1986), Magnetic hysteresis properties and
anhysteretic remanent magnetization of spherical Fe304 particles in the grain
size range 60-160 nm. Physics of the Earth and Planetary Interiors, 46, 77-83.
SCHULZ, H.D., DAHMKE, A, SCHINZEL, U., WALLMANN, K. & ZABEL, M. (1994), Early
diagenetic processes, fluxes and reaction rates in sediments of the South
Atlantic, Geochimica et Cosmochimica Acta, 58, 2041-2060.
SHETTY, TS., SETTY, T.M.R & RAVISHANKAR, C.N. (1992), Temperature growth
response of spoilage bacteria isolated from indian oil-sardine (Sardinella
longiceps) stored in chilIed sea-water. Fishery Technology, 29, 158-161.
SHORT, K.A & BLAKEMORE, RP. (1986), Iron respiration-driven proton translocation in
aerobic bacteria. Journal of Bacteriology, 167, 729-731.
SKOWRONEK, F. (1994), Frühdiagenetische Stoff-Flüsse gelöster Schwermetalle an der
Oberfläche von Sedimenten des Weser Ästuars, Berichte aus dem Fachbereich
Geowissenschaften der Universität Bremen. 106 (in Druck).
SMITH, D.C., SIMON, M., ALLREDGE, AL. & AZAM, F. (1992), Intense hydrolytic enzyme
activity on marine aggregates and implications for rapid particle dissolution.
Nature, 359, 139-142.
SMITH, J.M.A, ANDREWS, S.C., GUEST, J.R & HARRISON, P.M. (1990), Bacterioferritin: A
microbial iron-storage protein? In: Frankei, RB. & Blakemore, RP. (Eds.), Iron
Biominerals. Plenum Press, New York, 325-337.

117
SPARKS, N.H.C., COURTAUX, L., MANN, S. & BOARD, RG. (1986), Magnetotactic bacteria
are widely distributed in sediments in the U.K.. FEMS Microbiology Letters, 37,
305-308.
SPORMANN, AM. & WOLFE, RS. (1984), Chemotactic, magnetotactic and tactile
behaviour in a magnetic spirillum. FEMS Microbiology Letters, 22, 171-177.
SPRING, S., AMANN, R., LUDWIG, W., SCHLEIFER, K.H. & PETERSEN, N. (1992),
Phylogenetic diversity and identification of nonculturable magnetotactic bacteria.
Systematic and Applied Microbiology, 15, 116-122.
STACEY, F.D. & BANERJEE, S.K. (1974), The physical principles of rock magnetism.
Developments in Solid Earth Geophysics, 5, Elsevier Scientific Publishing
Company, Amsterdam, 195.
STEINBERGER, B. (1990), Über die Bewegung von magnetischen Bakterien in
Magnetfeldern. Diplomarbeit am Institut für Allgemeine und Angewandte
Geophysik der Universität München, 102.
STOLZ, J.F., , D.R & HAGGERTY, S.E. (1990), Biogenic Magnetite and the Magnetization
of Sediments. Journal of Geophysical Research, 95, 4355-4361.
STOLZ, J.F., CHANG, S.-B. R & KIRSCHVINK, J.L. (1986), Magnetotactic bacteria and
single-domain magnetite in hemipelagic sediments. Nature, 321, 849-851.
STONER, E.C. & WOHLFAHRT, E.P. (1948), A mechanism of magnetic hysteresis in
heterogeneous alloys. Philosophical Transactions of the Royal Society of
London, Series A, 240, 599-642.
SUESS, E. (1980), Particulate organic carbon flux in the oceans: Surface productivity and
oxygen utilization. Nature, 288, 260-263.
TARDENT, P. (1979), Meeresbiologie, Georg Thieme Verlag, Stuttgart, 32, 381.
TIETZE, U. & SCHENK, C. (1986), Halbleiter-Schaltungstechnik. Springer, Berlin, 860.
TORRES OE ARAUJO, F.F., GERMANO, F.A, GONCALVES, L.L., PIRES, M.A & FRANKEL
R.B. (1990), Magnetic polarity fractions in magnetotactic bacterial populations
near the Geomagnetic Equator. Biophysical Journal, 58, 549-555.
TURLEY, C.M. & LOCHTE, K. (1990), Microbial response to the input of fresh detritus to
the deep-sea bed. Palaeogeography, Palaeoclimatology, Palaeoecology, 89, 3
23.
V. DOBENECK (1993), Neue Ansätze zur Messung und Interpretation der magnetischen
Hysterese von Tiefseesedimenten, Verlag Marie L. Leidorf, Buch am Erlbach,
190.
v. DOBENECK, T., PETERSEN, N. & VALI , H. (1987), Bakterielle Magnetofossilien.
Geowissenschaften in unserer Zeit, 5, 27 - 35.
VALI, H. & KIRSCHVINK, J.L. (1989), Magnetofossil dissolution in a palaeomagnetically
unstable deep-sea sediment. Nature, 339, 203-206.

118
VALI , H., V. DOBENECK, T, AMARANTIDIS, G" FÖRSTER, 0., MORTEANI, G., BACHMANN, L.
& PETERSEN, N. (1989), Biogenic and lithogenic magnetic minerals in Atlantic
and Pacific deep-sea sediments and their paleomagnetic significance.
Geologische Rundschau, 78, 753-764.
VISHNIAC, H.S. & KURTZMAN, C.P. (1992), Cryptococcus antarcticus, new species and
Cryptococcus albidosimilis, new species: basidioblastomycetes from antarctic
soils. International Journal of Systematic Bacteriology, 42,547-553.
WALLCOTT, C. & GREEN, R.P. (1974), Orientation of homing pigeons altered by a change
in the direction of an applied magnetic field. Science, 184, p. 180.
WARREN, B.A & SPEER, K.G. (1991), Deep circulation in the Eastern South Atlantic
Ocean. Deep-Sea Research, 38, Suppl. 1, 281-322.
WEFER, G. & FISCHER, G. (1993), Seasonal patterns of vertical flux in equatorial and
coastal upwelling areas of the eastern Atlantic. Deep-Sea Research, 40, 1613
1645.
WEISS, P. & FORRER, R. (1929), Loi d'approche a la saturation. Annales de Physique,
12, 274-287.
WILLIAMS, W. & DUNLOP, D.J. (1990), Some effects of grain shape and varying external
magnetic fields on the magnetic structure of small grains of magnetite. Physics
of the Earth and Planetary Interiors, 65, 1-14.
YAMAZAKI, T, KATSURA, I. & MARUMO, K. (1991), Origin of stable remanent
magnetization of siliceous sediments in the Central Pacific. Earth and Planetary
Science Letters, 105, 81-93.
Yano, Y., Murayama, F., Kataho, N., Tachibana, M. & Nakamura, T (1992), Quality
control on intermediate temperature conditioning of beef by measuring
cadaverine and hypoxanthine. Animal Science and Technology, 63, 72-81.
YAYANOS, AA & Dietz, AS. (1982), Thermal inactivation of a deep-sea barophilic
bacterium, Isolate CNPT-3. Applied and Environmental Microbiology, 43, 1481
1489.
YAYANOS, AA & Dietz, AS. (1983), Death of a hadal deep-sea bacterium after
decompression. Science, 220, 497-498.
YAYANOS, AA (1986), Evolutional and ecological implication of the properties of deep
sea barophilic bacteria. Proceedings of the National Academy of Sciences of the
USA, 83, 9542-9546.

119
Anhang A
Aufnahme von Spuren magnetotaktischer Bakterien mittels Bildverarbeitung
A.1 Beschreibung der Bildverarbeitungskarte
Mit der verwendeten MVP-AT Karte, die seit 1988 auf dem Markt ist, kann in Verbindungmit einem gewöhnlichen AT-Rechner Echtzeit-Bildverarbeitung durchgeführt werden.Das Bildsignal wird von der Videokamera direkt in die Bildverarbeitungskarte gegeben.Von dort kann es - auch unverarbeitet - auf dem zugehörigen Monitor dargestelltwerden.
Zur Bearbeitung stehen vier Bildspeicher mit 512 x 512 Bildpunkten (Pixel) zurVerfügung. Weiterhin ist ein RAM-Speicher zur Abspeicherung von Bildausschnitten undvon Software-Filtern zur Bearbeitung der Bilder vorhanden.
A.1.1 Echtzeit-Bildverarbeitung
Die Datenmenge, die bei Echtzeit-Bildverarbeitung anfällt, ist sehr hoch. Die Videokamera liefert pro Sekunde 25 Bilder mit jeweils 262 kByte. Eine Festplatte mit 500MByte würde damit in 76.3 Sekunden vollgeschrieben. Eine sinnvolle Bearbeitung kanndaher nur erfolgen, wenn die Bilderflut schon in der Bildverarbeitungskarte reduziertwird. Nur fertig prozessierte Bilder wurden deshalb auf der Festplatte des Rechnersabgespeichert.
Die MVP-AT Karte ist langsamer als heute erhältliche Bildverarbeitungskarten. Umwirklich in Echtzeit zu arbeiten, darf die Ausführung eines Bildverarbeitungsbefehls nichtlänger als 40 Millisekunden benötigen. Dies ist hier nur für einfachere Befehle der'lmager-AT'-Software der Fall. Es ist nicht möglich, in 40 ms mehrere Befehle auf einBild anzuwenden. Dennoch gelang es, Spuren der Bakterienbahnen aufzuzeichnen.
A.1.2 Bearbeitung von Bildsignalen in der Bildverarbeitungskarte
Das Bildsignal wird als Analogsignal über ein Koaxialkabel in die Bildverarbeitungskartegegeben. Vor der Digitalisierung des Bildes können die Helligkeit (offset) und derKontrast (gain) des Bildes optimiert werden. Durch Einstellung des Offsetsignals läßtsich beispielsweise eine langsame Helligkeitsänderung der Lichtquelle kompensieren.
Bei der Digitalisierung wird aus dem rechteckigen Kamerabild mit Seitenverhältnis 4:3ein Teilbild herausgeschnitten. Zunächst läuft das Bild durch ein numerisches Filter, die'Input-Look-up-Table', durch die den Graustufen des Bildes andere Werte zugewiesenwerden können (Abb. A.1). Auf diese Weise läßt sich beispielsweise der Kontrast desBildes erhöhen, indem ein bestimmter Grauwertbereich gespreizt wird.

Grauwert
255
oOriginalbild
120
Grauwert
255
obearbeitetes Bild
Abb. A.1 Schematische Darstellung der Kontrastverstärkung mittels der 'Look-upTable'. Die Spreizung der Grauwerte muß nicht symmetrisch erfolgen.
Die Bearbeitung erfolgt auf einem der vier Bildspeicher. Er besteht aus 512 x 512Bildpunkten mit 8 Bit Helligkeitsinformation (256 Graustufen). Dabei muß beachtetwerden, daß ein Pixel nicht quadratisch ist, sondern ein Verhältnis Breite zu Höhe von0.88 besitzt.
Auf das Bild im Bildspeicher können die Bildverarbeitungsroutinen der 'Imager-AT'Software angewendet werden. Es sind drei Fälle zu unterscheiden:
- In der Testphase läßt sich die Wirkung der Bildverarbeitungsoperationen mit Hilfeeines vom Programm zur Verfügung gestellten Interpreters prüfen. Dieser Prozess istsehr langsam und kann nur auf stehende Einzelbilder angewandt werden.
- Die Bildverarbeitungsroutinen können auch in FORTRAN-Programme eingebundenwerden. Dadurch wird die Bildverarbeitung schneller. Da die CPU des Computersjeden Befehl einzeln an die Karte übergibt, genügt die Geschwindigkeit jedoch nicht,um in Echtzeit zu arbeiten.
- Bestimmte Befehle der 'Imager-AT'-Software erlauben es, durch einen einmaligenAufruf von der CPU des Rechners mehrere Bilder nacheinander zu bearbeiten. Nurmit diesen Befehlen konnte eine Echtzeit-Bildverarbeitung durchgeführt werden.
Das bearbeitete Bild erscheint auf dem zugehörigen Monitor. Vor der Ausgabe kannes durch eine 'Output-Look-up-Table' nochmals verändert werden.
A.1.3 Abspeichern auf Festplatte
Ein einfacher Befehl des Softwarepakets erlaubt die Abspeicherung ganzer Bilder aufFestplatte. Das Bild wird unformatiert zeilenweise abgespeichert und kann deshalb nichteinfach in Grafikprogramme eingelesen werden.
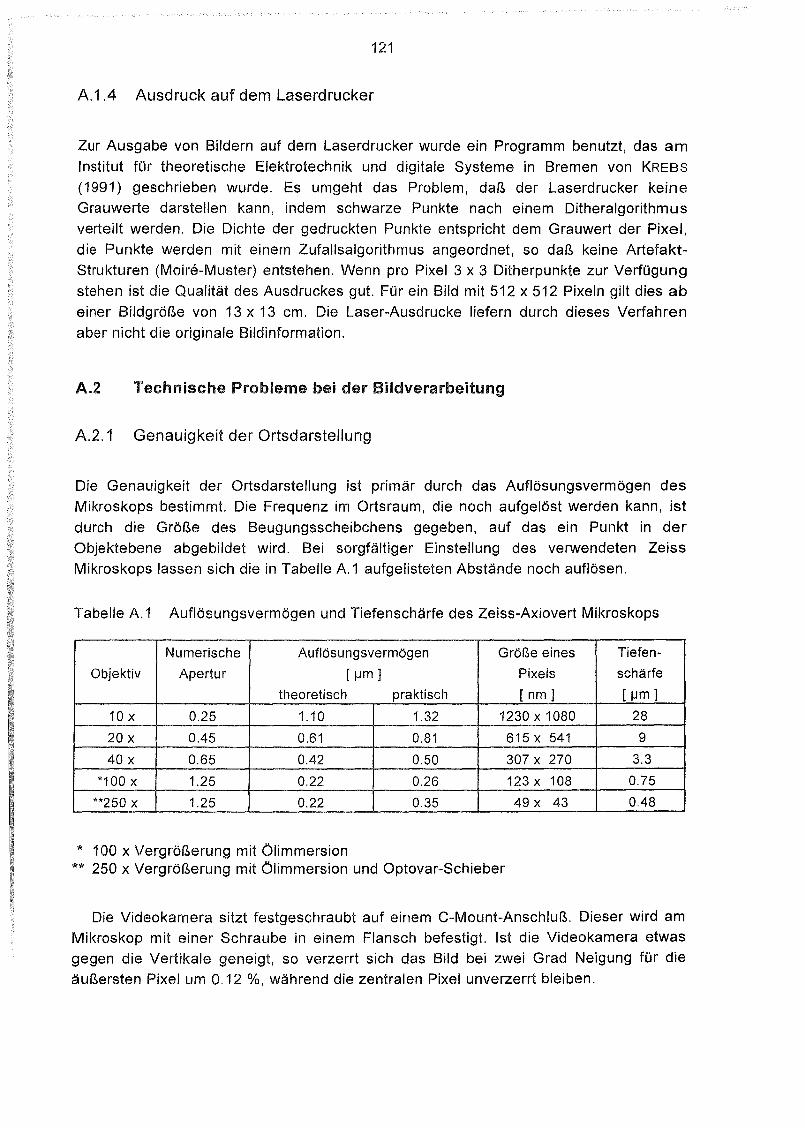
121
A.1.4 Ausdruck auf dem Laserdrucker
Zur Ausgabe von Bildern auf dem Laserdrucker wurde ein Programm benutzt, das amInstitut für theoretische Elektrotechnik und digitale Systeme in Bremen von KREBS
(1991) geschrieben wurde. Es umgeht das Problem, daß der Laserdrucker keineGrauwerte darstellen kann, indem schwarze Punkte nach einem Ditheralgorithmusverteilt werden. Die Dichte der gedruckten Punkte entspricht dem Grauwert der Pixel,
die Punkte werden mit einem Zufallsalgorithmus angeordnet, so daß keine ArtefaktStrukturen (Moire-Muster) entstehen. Wenn pro Pixel 3 x 3 Ditherpunkte zur Verfügung
stehen ist die Qualität des Ausdruckes gut. Für ein Bild mit 512 x 512 Pixeln gilt dies abeiner Bildgröße von 13 x 13 cm. Die Laser-Ausdrucke liefern durch dieses Verfahrenaber nicht die originale Bildinformation.
A.2 Technische Probleme bei der Bildverarbeitung
A.2.1 Genauigkeit der Ortsdarstellung
Die Genauigkeit der Ortsdarstellung ist primär durch das Auflösungsvermögen desMikroskops bestimmt. Die Frequenz im Ortsraum, die noch aufgelöst werden kann, ist
durch die Größe des Beugungsscheibchens gegeben, auf das ein Punkt in derObjektebene abgebildet wird. Bei sorgfältiger Einstellung des verwendeten ZeissMikroskops lassen sich die in Tabelle Ai aufgelisteten Abstände noch auflösen.
Tabelle Ai Auflösungsvermögen und Tiefenschärfe des Zeiss-Axiovert Mikroskops
Numerische Auflösungsvermögen Größe eines Tiefen-
Objektiv Apertur [ IJm ] Pixels schärfe
theoretisch praktisch [nm] [ IJm ]
10 x 0.25 1.10 1.32 1230 x 1080 28
20 x 0.45 0.61 0.81 615 x 541 9
40 x 0.65 0.42 0.50 307 x 270 3.3
*100 x 1.25 0.22 0.26 123x108 0.75
**250 x 1.25 0.22 0.35 49 x 43 0.48
* 100 x Vergrößerung mit Ölimmersion** 250 x Vergrößerung mit Ölimmersion und Optovar-Schieber
Die Videokamera sitzt festgeschraubt auf einem C-Mount-Anschluß. Dieser wird amMikroskop mit einer Schraube in einem Flansch befestigt. Ist die Videokamera etwasgegen die Vertikale geneigt, so verzerrt sich das Bild bei zwei Grad Neigung für dieäußersten Pixel um 0.12 %, während die zentralen Pixel unverzerrt bleiben.

122
Wenn ein Objekt nicht genau in der Brennebene liegt, erscheint es verschwommen,seine Größe wird verfälscht. Ein Punkt außerhalb der Brennebene wird als Scheibchenabgebildet. Die in Tabelle A.1 angegeben Tiefenschärfenbereiche sind so berechnet,daß das abgebildete Scheibchen die Größe eines Bildpunktes der Videokamera besitzt.
Befindet sich das Objekt mehr als 50 % des angegebenen Betrages aus der
Brennebene, dann erscheint es unscharf, die Objektgröße wird verfälscht (Abb. A.2).
o
1I I
I I
I!
! I
I I
I'--..:ta. i 1':-
G
d
Abb. A.2 Strahlengang einer Linse. Ein Punkt im Abstand d außerhalb derObjektebene 0 erscheint in der Zwischenbildebene Z auf die Breite Gverschmiert. Umgekehrt wird in einen Punkt im Gesichtsfeld des Betrachtersdie Information aus diesem Bereich abgebildet. Die Größe derVerschmierung kann aus der Distanz zum Brennpunkt und dem Winkel unterdem der äußerste Strahl einfällt (der numerischen Apertur) bestimmt werden.
TabelleA.2 Größe, auf die ein Punkt durch die verschiedenen Objektive inAbhängigkeit vom Abstand zur Fokusebene verschmiert wird.
Objektiv Numerische Winkel des äußersten Größe des Beugungsscheibchens /Apertur Strahls Abstand von der Fokusebene
10 x 0.25 14.5° 0.26
20 x 0.45 26.r 0.50
40 x 0.65 40.5° 0.86
100 x 1.25 55.4° 1.45
A.2.2 Verdrehung der Kamera
Die Magnetbakterien schwimmen parallel zum Feld, das ihnen durch die Feldspulen vorgegeben wird. Um den Winkel zwischen der Bewegungsrichtung der Bakterien und derFeldrichtung exakt zu bestimmen, ist eine möglichst genaue Ausrichtung derVideokamera wichtig. Deshalb mußte die Parallelität von Kamera und Feldspulen häufigüberprüft werden. Da die Orientierung der Videokamera an einer festen Kante desObjekttisches nicht möglich war (der unmagnetische Gleittisch besitzt keine festeKante), wurde dazu die mittlere Schwimmrichtung der Magnetbakterien selbst verwendet. Folgendes Verfahren hat sich bewährt:
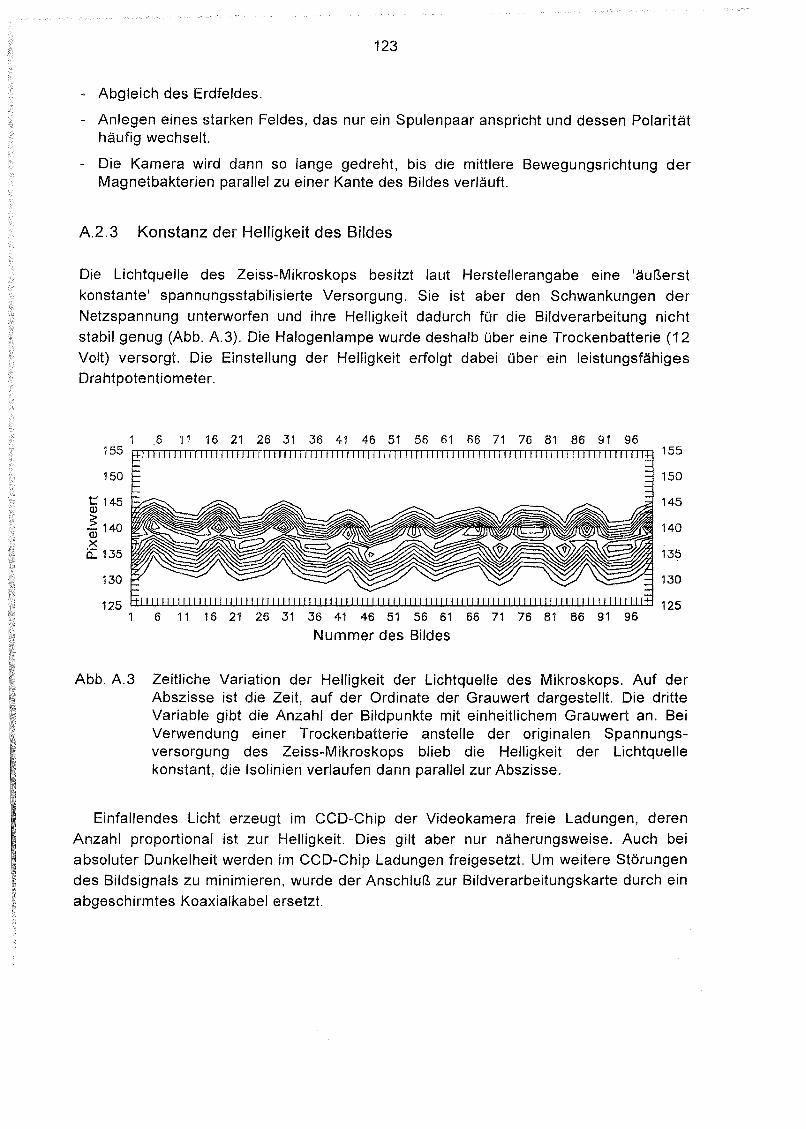
123
- Abgleich des Erdfeldes.
- Anlegen eines starken Feldes, das nur ein Spulenpaar anspricht und dessen Polaritäthäufig wechselt.
- Die Kamera wird dann so lange gedreht, bis die mittlere Bewegungsrichtung derMagnetbakterien parallel zu einer Kante des Bildes verläuft.
A.2.3 Konstanz der Helligkeit des Bildes
Die Lichtquelle des Zeiss-Mikroskops besitzt laut Herstellerangabe eine 'äußerst
konstante' spannungsstabilisierte Versorgung. Sie ist aber den Schwankungen derNetzspannung unterworfen und ihre Helligkeit dadurch für die Bildverarbeitung nichtstabil genug (Abb. A.3). Die Halogenlampe wurde deshalb über eine Trockenbatterie (12Volt) versorgt. Die Einstellung der Helligkeit erfolgt dabei über ein leistungsfähigesDrahtpotentiometer.
1 .6 11 16 21 26 31 36 41 46 51 56 61 66 71 76 81 86 91 96155 155
150
t: 145Q)
~ 140Q)
x0:: 135
130
150
145
140
135
130
125 1251 6 11 16 21 26 31 36 41 46 51 56 61 66 71 76 81 86 91 96
Nummer des Bildes
Abb. A.3 Zeitliche Variation der Helligkeit der Lichtquelle des Mikroskops. Auf derAbszisse ist die Zeit, auf der Ordinate der Grauwert dargestellt. Die dritteVariable gibt die Anzahl der Bildpunkte mit einheitlichem Grauwert an. BeiVerwendung einer Trockenbatterie anstelle der originalen Spannungsversorgung des Zeiss-Mikroskops blieb die Helligkeit der Lichtquellekonstant, die Isolinien verlaufen dann parallel zur Abszisse.
Einfallendes Licht erzeugt im CCO-Chip der Videokamera freie Ladungen, derenAnzahl proportional ist zur Helligkeit. Dies gilt aber nur näherungsweise. Auch beiabsoluter Dunkelheit werden im CCD-Chip Ladungen freigesetzt. Um weitere Störungendes Bildsignals zu minimieren, wurde der Anschluß zur Bildverarbeitungskarte durch einabgeschirmtes Koaxialkabel ersetzt.
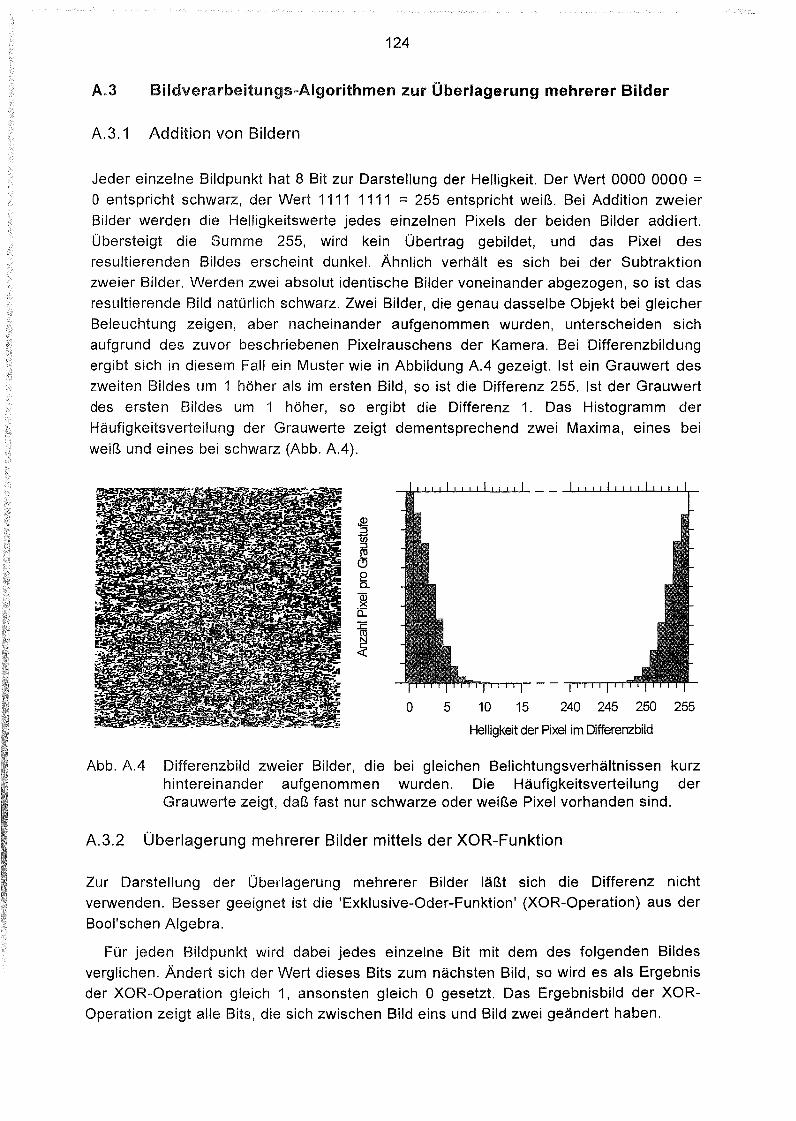
124
A.3 Bildverarbeitungs-Algorithmen zur Überlagerung mehrerer Bilder
A.3.1 Addition von Bildern
Jeder einzelne Bildpunkt hat 8 Bit zur Darstellung der Helligkeit. Der Wert 0000 0000 =o entspricht schwarz, der Wert 1111 1111 =255 entspricht weiß. Bei Addition zweier
Bilder werden die Helligkeitswerte jedes einzelnen Pixels der beiden Bilder addiert.Übersteigt die Summe 255, wird kein Übertrag gebildet, und das Pixel des
resultierenden Bildes erscheint dunkel. Ähnlich verhält es sich bei der Subtraktion
zweier Bilder. Werden zwei absolut identische Bilder voneinander abgezogen, so ist das
resultierende Bild natürlich schwarz. Zwei Bilder, die genau dasselbe Objekt bei gleicher
Beleuchtung zeigen, aber nacheinander aufgenommen wurden, unterscheiden sich
aufgrund des zuvor beschriebenen Pixelrauschens der Kamera. Bei Differenzbildung
ergibt sich in diesem Fall ein Muster wie in Abbildung A.4 gezeigt. Ist ein Grauwert des
zweiten Bildes um 1 höher als im ersten Bild, so ist die Differenz 255. Ist der Grauwert
des ersten Bildes um 1 höher, so ergibt die Differenz 1. Das Histogramm der
Häufigkeitsverteilung der Grauwerte zeigt dementsprechend zwei Maxima, eines bei
weiß und eines bei schwarz (Abb. A.4).
Helligkeit der Pixel im Differenzbild
240 245 250 25510 155o
Abb. AA Differenzbild zweier Bilder, die bei gleichen Belichtungsverhältnissen kurzhintereinander aufgenommen wurden. Die Häufigkeitsverteilung derGrauwerte zeigt, daß fast nur schwarze oder weiße Pixel vorhanden sind.
A.3.2 Überlagerung mehrerer Bilder mittels der XOR-Funktion
Zur Darstellung der Überlagerung mehrerer Bilder läßt sich die Differenz nichtverwenden. Besser geeignet ist die 'Exklusive-Oder-Funktion' (XOR-Operation) aus der
Bool'schen Algebra.
Für jeden Bildpunkt wird dabei jedes einzelne Bit mit dem des folgenden Bildesverglichen. Ändert sich der Wert dieses Bits zum nächsten Bild, so wird es als Ergebnis
der XOR-Operation gleich 1, ansonsten gleich 0 gesetzt. Das Ergebnisbild der XOROperation zeigt alle Bits, die sich zwischen Bild eins und Bild zwei geändert haben.
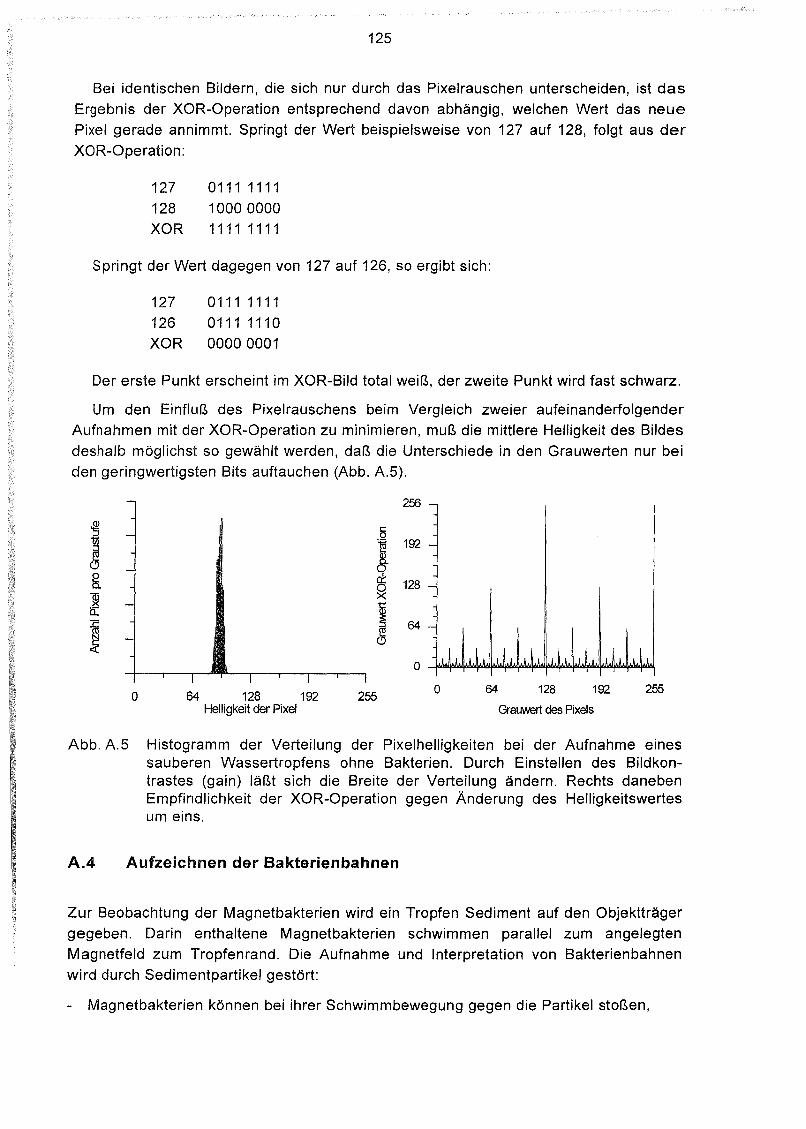
125
Bei identischen Bildern, die sich nur durch das Pixelrauschen unterscheiden, ist dasErgebnis der XOR-Operation entsprechend davon abhängig, welchen Wert das neuePixel gerade annimmt. Springt der Wert beispielsweise von 127 auf 128, folgt aus derXOR-Operation:
127 01111111128 1000 0000XOR 1111 1111
Springt der Wert dagegen von 127 auf 126, so ergibt sich:
127 01111111126 0111 1110XOR 00000001
Der erste Punkt erscheint im XOR-Bild total weiß, der zweite Punkt wird fast schwarz.
Um den Einfluß des Pixelrauschens beim Vergleich zweier aufeinanderfolgenderAufnahmen mit der XOR-Operation zu minimieren, muß die mittlere Helligkeit des Bildesdeshalb möglichst so gewählt werden, daß die Unterschiede in den Grauwerten nur beiden geringwertigsten Bits auftauchen (Abb. A.5).
255255128 192
Grauwert des Pixels
64o
256
.~~ 192
8~ 128x
I 64
64 128 192Helligkeit der Pixel
o
Abb. A.5 Histogramm der Verteilung der Pixelhelligkeiten bei der Aufnahme einessauberen Wassertropfens ohne Bakterien. Durch Einstellen des Bildkontrastes (gain) läßt sich die Breite der Verteilung ändern. Rechts danebenEmpfindlichkeit der XOR-Operation gegen Änderung des Helligkeitswertesum eins.
A.4 Aufzeichnen der Bakterienbahnen
Zur Beobachtung der Magnetbakterien wird ein Tropfen Sediment auf den Objektträgergegeben. Darin enthaltene Magnetbakterien schwimmen parallel zum angelegtenMagnetfeld zum Tropfenrand. Die Aufnahme und Interpretation von Bakterienbahnenwird durch Sedimentpartikel gestört:
- Magnetbakterien können bei ihrer Schwimmbewegung gegen die Partikel stoßen,

126
- die Partikel erscheinen auf dem Bild und erschweren die Interpretation.
Wegen der abnehmenden Dicke des Tropfens am Rand wird die Bewegung derMagnetbakterien dort zusätzlich behindert. Um zu gewährleisten, daß die Magnetbakterien in einem sedimentpartikelfreien Wassertropfen ausreichender Dickeschwimmen, hat sich folgende präparative Technik bewährt (Abb. A 7):
Der Sedimenttropfen wird auf den Objektträger gegeben und ein mit Wassergetränkter Streifen Filterpapier wird aufrecht neben den Sedimenttropfen gestellt. Aufdie vom Sedimenttropfen abgewandte Seite des Papierstreifens wird dann einWassertropfen gegeben und durch vorsichtige Zugabe von Wasser auf der demSedimenttropfen zugewandten Seite werden Sedimenttropfen und Papierstreifen
verbunden. Als Filterpapier haben sich handelsübliche ungebleichte Kaffeefilter bewährt,die Magnetbakterien können ihre großen Poren ungehindert passieren. Das zugegebene
Wasser stammt aus dem Aquarium, in dem die Bakterien leben. Es wurde derWasserschicht entnommen, die unmittelbar über dem Sediment liegt.
Magnetfeld )
o
Magnetfeld )
o
Abb. A 7 Um einen sauberen Hintergrund bei der Bildverarbeitung zu erhalten, werdendie Magnetbakterien aus der Sedimentsuspension S durch den Papierstreifen P in sauberes Wasser W 'geführt' (0 - Objektträger).
Das Wasser wird beim Entnehmen durch einen Membranfilter (porengröße 0.22 IJm)gesaugt, und dadurch vollständig gesäubert. Benutzt man Wasser von der Oberfläche,so wird die Beweglichkeit der Magnetbakterien deutlich verringert. Das Wasserunmittelbar über dem Sediment hat offensichtlich ein ähnliches chemisches Milieu wiedas Porenwasser. Nach diesen vorbereitenden Maßnahmen gelingen saubereAufnahmen der Bahnen der Bakterien.
Wie oben beschrieben, werden aufeinanderfolgende Bilder mit der XOR-Verknüpfungüberlagert. Nur dort, wo sich ein Magnetbakterium bewegt hat, findet eine Änderung derPixelhelligkeiten statt, im XOR-Bild entsteht ein heller Fleck. Der Rest des Bildes sollteim XOR-Bild schwarz erscheinen, wenn die Helligkeit nicht schwankt. Da die Beobachtung ohne Deckglas stattfindet, wird das einfallende Licht je nach momentaner Neigungder Oberfläche reflektiert. Dies und das zuvor diskutierte Pixel rauschen führen dazu,daß der Hintergrund des XOR-Bildes nicht völlig schwarz bleibt. Wird die Lichthelligkeitso eingestellt, daß die Helligkeitsschwankungen sich auf die geringerwertigen Bitsbeschränken, erscheint der Hintergrund ausreichend dunkel. Zusätzlich können, wiespäter in diesem Kapitel erläutert, diese Bits maskiert werden, was zu einer weiterenVerbesserung des Bildes führt (Abb. AB).
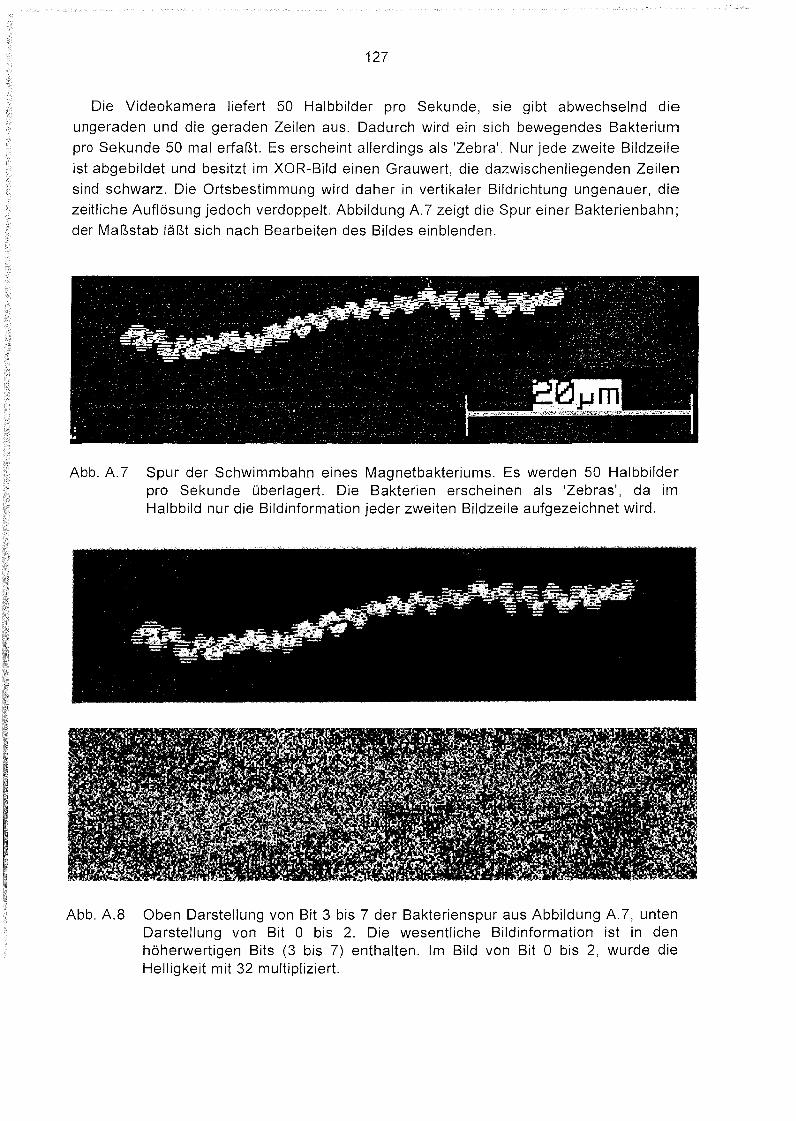
127
Die Videokamera liefert 50 Halbbilder pro Sekunde, sie gibt abwechselnd dieungeraden und die geraden Zeilen aus. Dadurch wird ein sich bewegendes Bakterium
pro Sekunde 50 mal erfaßt. Es erscheint allerdings als 'Zebra'. Nur jede zweite Bildzeile
ist abgebildet und besitzt im XOR-Bild einen Grauwert, die dazwischenliegenden Zeilen
sind schwarz. Die Ortsbestimmung wird daher in vertikaler Bildrichtung ungenauer, diezeitliche Auflösung jedoch verdoppelt. Abbildung A 7 zeigt die Spur einer Bakterienbahn;
der Maßstab läßt sich nach Bearbeiten des Bildes einblenden.
Abb. A 7 Spur der Schwimmbahn eines Magnetbakteriums. Es werden 50 Halbbilderpro Sekunde überlagert. Die Bakterien erscheinen als 'Zebras', da imHalbbild nur die Bildinformation jeder zweiten Bildzeile aufgezeichnet wird.
Abb. A8 Oben Darstellung von Bit 3 bis 7 der Bakterienspur aus Abbildung A7, untenDarstellung von Bit 0 bis 2. Die wesentliche Bildinformation ist in denhöherwertigen Bits (3 bis 7) enthalten. Im Bild von Bit 0 bis 2, wurde dieHelligkeit mit 32 multipliziert.


128
Aus den geschilderten Gründen ist die wesentliche Bildinformation in denhöherwertigen Bits der Helligkeitswerte enthalten. In Abbildung A8 ist das Bild der Bits
3 - 7 dem der Bits 0 - 2 gegenübergestellt. Es wird deutlich, daß die geringerwertigen
Bits kaum Information beinhalten. Deshalb empfiehlt es sich, auf diese Bits ganz zuverzichten. Dazu wurden sie maskiert, ihr Wert gleich Null gesetzt. Dies führte zu
rauschärmeren, saubereren Bildern in der XOR-Darstellung.
Durch Nachbearbeiten mit einem einfachen Filter (Abb. Ag), kann ein realistischeres
Bild erzeugt werden. Die Helligkeitsinformation der dunklen Zeilen, kann aber nicht
rekonstruiert werden. Ein solchermaßen gefiltertes Bild täuscht die zusätzliche
Information allerdings nur vor, daher wurde von dieser Option hier nicht weiter
Gebrauch gemacht.
Ursprungsbild
00 00 00 00 00
00 00 00 00 00
00 60 85 55 00
00 00 00 00 00
10 87 90 80 05
00 00 00 00 00
00 60 84 51 00
00 00 00 00 00
00 00 00 00 00
Operator
00 00 00
00 01 00
00 01 00
Ergebnisbild
00 00 00 00 00
00 60 85 55 00
00 60 85 55 00
10 87 90 80 05
10 87 90 80 05
00 60 84 51 00
00 60 84 51 00
00 00 00 00 00
00 00 00 00 00
Abb. Ag Wirkungsweise des Filteroperators, der aus einem Halbbild ein Pseudo-Vollbilderzeugt. Die Grauwerte der beschriebenen Zeile werden auf die darüberliegende Bildzeile kopiert, das Objekt wird um eine Zeile vergrößert.
Abb. A1 0 Bakterienspur aus Abb. A.7 nach Anwendung des Filteroperators nach Abb.Ag.

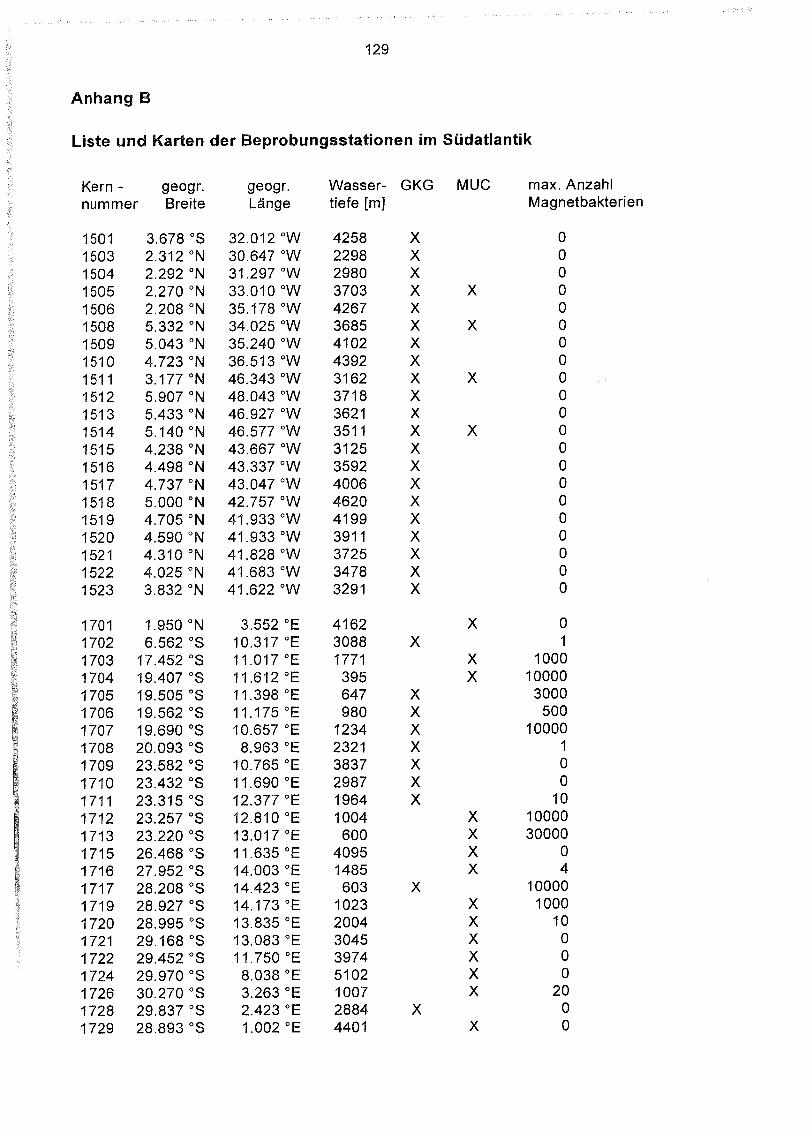
129
Anhang B
Liste und Karten der Beprobungsstationen im Südatlantik
Kern - geogr. geogr. Wasser- GKG MUC max. Anzahlnummer Breite Länge tiefe [m] Magnetbakterien
1501 3.678 oS 32.012°W 4258 X 01503 2.312 oN 30.647°W 2298 X 01504 2.292 oN 31.297°W 2980 X 01505 2.270 oN 33.010 0 W 3703 X X 01506 2.208 oN 35.178°W 4267 X 01508 5.332 ON 34.025°W 3685 X X 01509 5.043 oN 35.240 0 W 4102 X 01510 4.723 oN 36.513°W 4392 X 01511 3.177 oN 46.343°W 3162 X X 01512 5.907 oN 48.043°W 3718 X 01513 5.433 oN 46.927°W 3621 X 01514 5.140 oN 46.577°W 3511 X X 01515 4.238 oN 43.667°W 3125 X 01516 4.498 oN 43.337°W 3592 X 01517 4.737 oN 43.047°W 4006 X 01518 5.000 oN 42.757°W 4620 X 01519 4.705 oN 41.933°W 4199 X 01520 4.590 oN 41.933°W 3911 X 01521 4.310 oN 41.828°W 3725 X 01522 4.025 oN 41.683°W 3478 X 01523 3.832 oN 41.622°W 3291 X 0
1701 1.950 oN 3.552 oE 4162 X 01702 6.562 oS 10.317 oE 3088 X 11703 17.452 oS 11.017 oE 1771 X 10001704 19.407 oS 11.612 oE 395 X 100001705 19.505 oS 11.398 oE 647 X 30001706 19.562 oS 11.175 oE 980 X 5001707 19.690 oS 10.657 oE 1234 X 100001708 20.093 oS 8.963 oE 2321 X 11709 23.582 oS 10.765 oE 3837 X 01710 23.432 oS 11.690 oE 2987 X 01711 23.315 oS 12.377 oE 1964 X 101712 23.257 oS 12.810 oE 1004 X 100001713 23.220 oS 13.017 OE 600 X 300001715 26.468 oS 11.635 oE 4095 X 01716 27.952 oS 14.003 oE 1485 X 41717 28.208 oS 14.423 oE 603 X 100001719 28.927 oS 14.173 oE 1023 X 10001720 28.995 oS 13.835 oE 2004 X 101721 29.168 oS 13.083 oE 3045 X 01722 29.452 oS 11.750 oE 3974 X 01724 29.970 oS 8.038 OE 5102 X 01726 30.270 oS 3.263 oE 1007 X 201728 29.837 oS 2.423 oE 2884 X 01729 28.893 oS 1.002 oE 4401 X 0
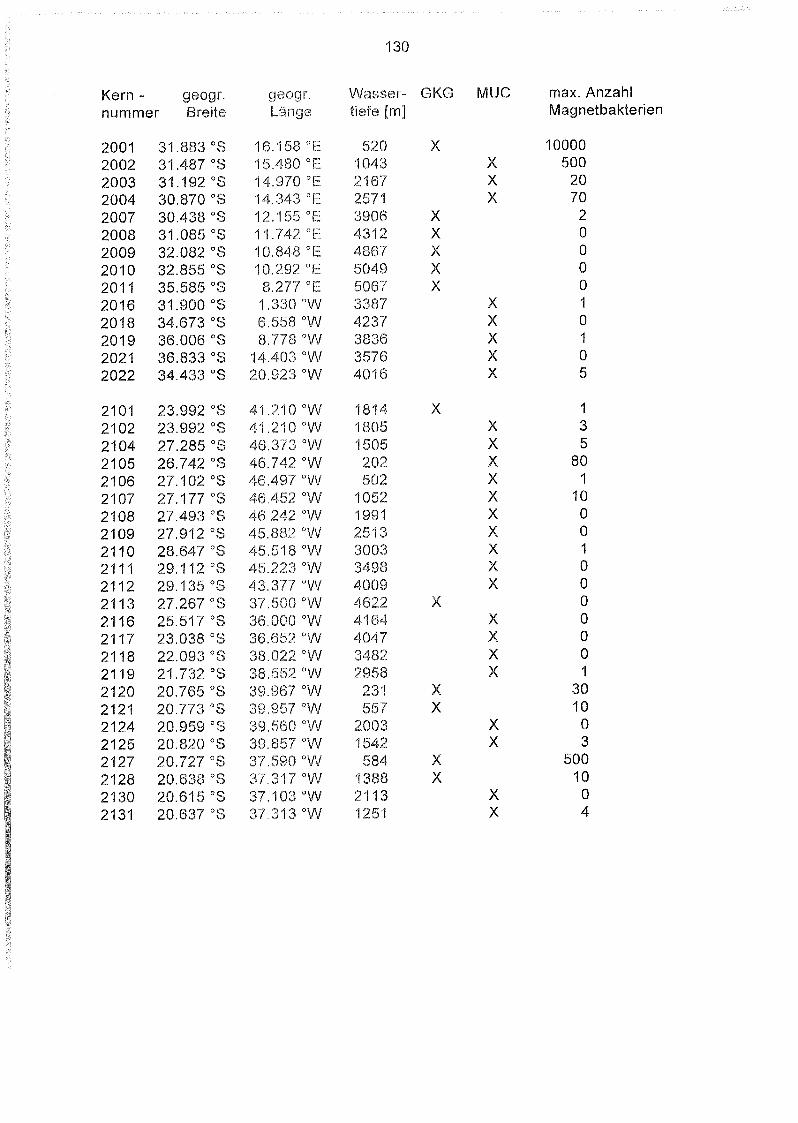
130
Kern - geogr. geogr. Wasser- GKG MUC max. Anzahlnummer Breite Länge tiefe [m] Magnetbakterien
2001 31.883 oS 16.158°E 520 X 100002002 31.487 oS 15.480 oE 1043 X 5002003 31.192 oS 14.970 2167 X 202004 30.870 oS 14.343 oE 2571 X 702007 30.438 oS 12.155 oE 3906 X 22008 31.085 oS 11.742 oE 4312 X 02009 32.082 oS 10.848 oE 4867 X 02010 32.855 oS 10.292 5049 X 02011 35.585 oS 8.277 oE 5067 X 02016 31.900 oS 1.330 oW 3387 X 12018 34.673 oS 6.558°W 4237 X 02019 36.006 oS 8.778°W 3836 X 12021 36.833 oS 14.403°W 3576 X 02022 34.433 oS 20.923°W 4016 X 5
2101 23.992 oS 41.210 0 W 1814 X 12102 23.992 oS 41.210 0 W 1805 X 32104 27.285 oS 46.3'73 °W 1505 X 52105 26.742 oS 46.742°W 202 X 802106 27.102 oS 46.497°W 502 X 12107 27.177 oS 46.452°W 1052 X 102108 27.493 oS 46.242°W 1991 X 02109 27.912 oS 45.882°W 2513 X 02110 28.647 oS 45.518°W 3003 X 12111 29.112 oS 45. °W 3498 X 02112 29.135 oS 43.377°W 4009 X 02113 27.267 oS 37.500 0 W 4622 X 02116 25.517 oS 36.000 0 W 4164 X 02117 23.038 oS 36.652°W 4047 X 02118 22.093 oS 38.022°W 3482 X 02119 21.732 oS 38.552°W 2958 X 12120 20.765 oS 39.96'7 °W 231 X 302121 20.773 oS 39.957°W 557 X 102124 20.959 oS 39.560 0 W 2003 X 02125 20.820 oS 39.857°W 1542 X 32127 20.727 oS 37.590 0 W 584 X 5002128 20.638 oS 37.3'1'7°W 1388 X 102130 20.615 oS 37.103°W 2113 X 02131 20.63'7 oS 37.313°W 1251 X 4
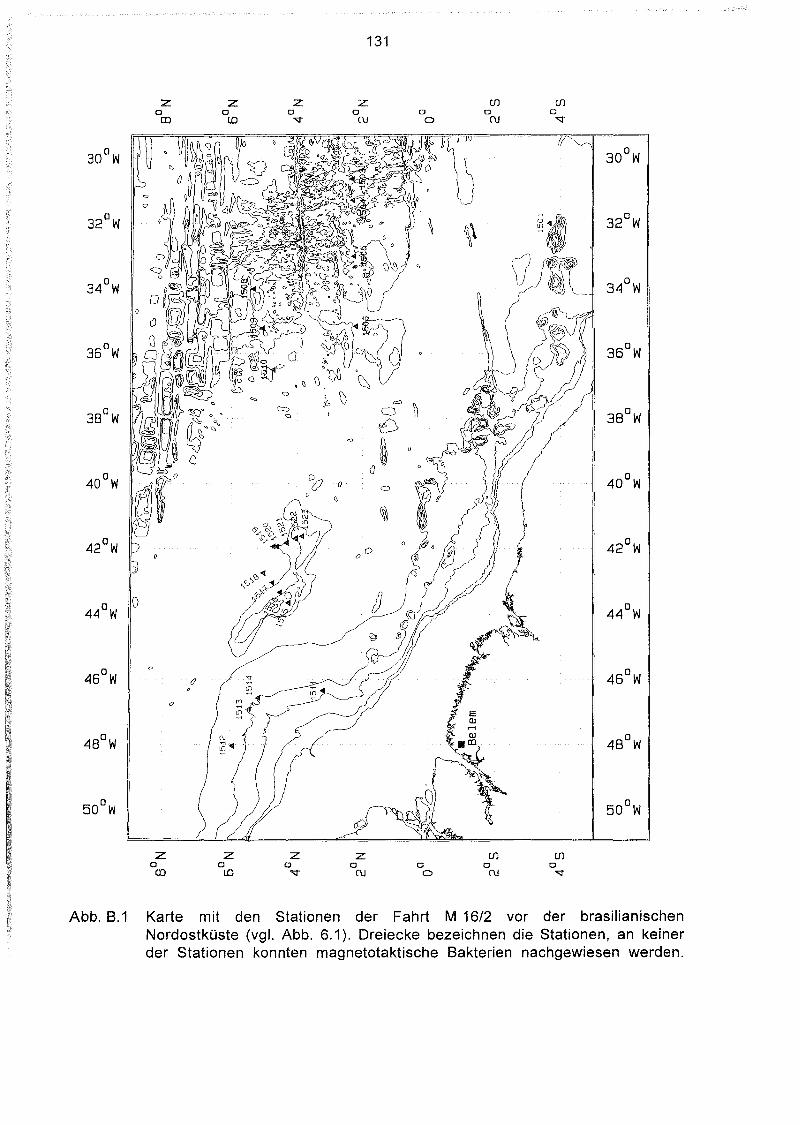
131
zoro
zolO
zo
""z
o(\J
oo
CDo
(\J
CDo
""
CDo
""CD
o(\J
oo
zo
(\J
zo
""z
olO
(J
o
zoro
32°W~~
32°W
V ~
34°W 34°W
400
W DO 0 0 40 0 W
0~
\Th
42 0 W0
0 42 0 W
44°W0
44°W
Abb. B.1 Karte mit den Stationen der Fahrt M 16/2 vor der brasilianischenNordostküste (vgl. Abb. 6.1). Dreiecke bezeichnen die Stationen, an keinerder Stationen konnten magnetotaktische Bakterien nachgewiesen werden.
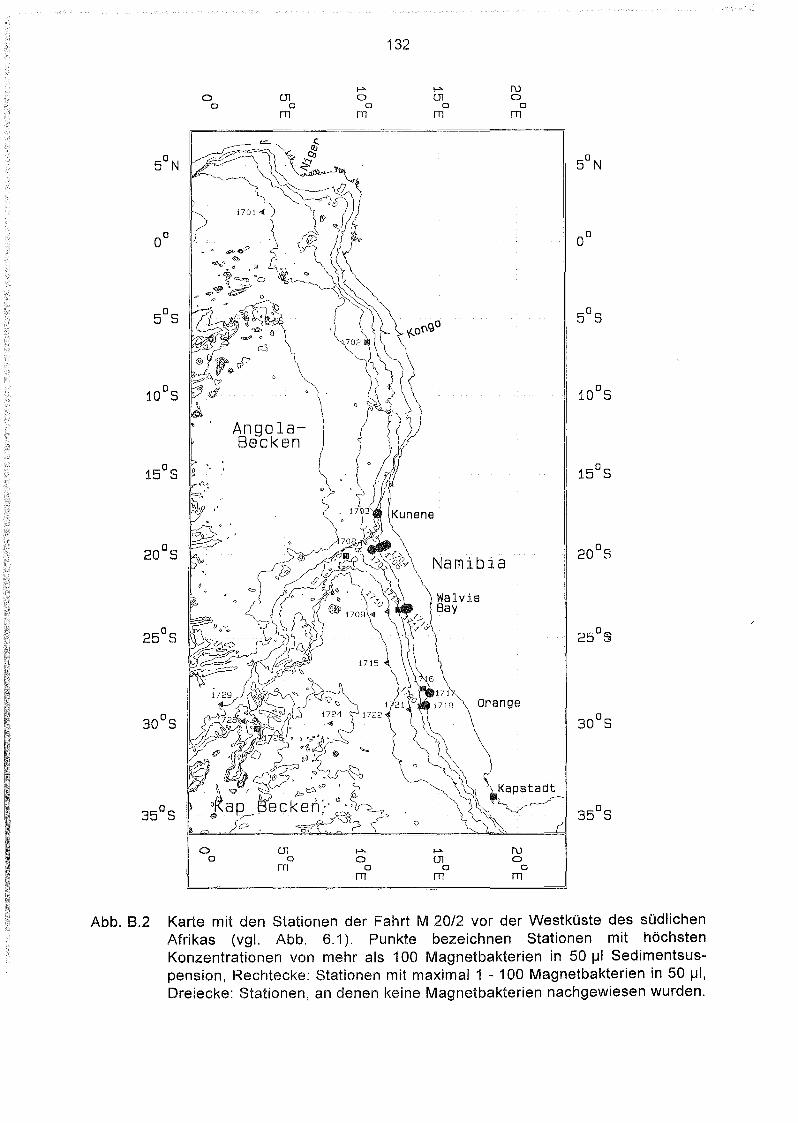
oo
oo
c.no
m
c.no
m
I-'"o
om
I-'"o
om
132
I-'"c.n
om
Namibia
...,.c.n
om
f\)o
om
f\)o
om
Abb. 8.2 Karte mit den Stationen der Fahrt M 20/2 vor der Westküste des südlichenAfrikas (vgl. Abb. 6.1). Punkte bezeichnen Stationen mit höchstenKonzentrationen von mehr als 100 Magnetbakterien in 50 \-11 Sedimentsuspension, Rechtecke: Stationen mit maximal 1 - 100 Magnetbakterien in 50 \-11,Dreiecke: Stationen, an denen keine Magnetbakterien nachgewiesen wurden.
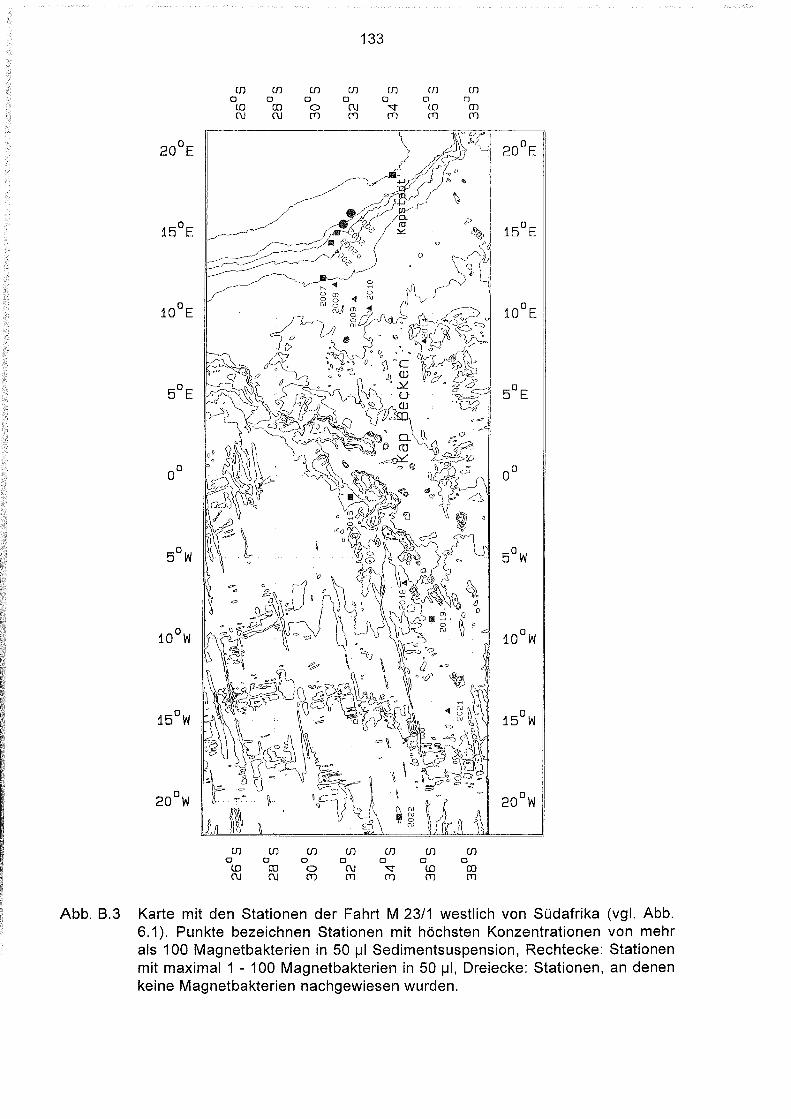
133
CD CD CD CD CD CD CD0 0 0 0 0 0 0lD m 0 (\J "<;J" lD m(\J (\J (Tl (Tl (Tl (Tl (Tl
200
E 200
E
/.'
150
Ep
150 E~
0
CD CD CD CD (J) CD CD0000000
lD m 0 (\J "<;J" lD m(\J (\J (Tl (Tl (Tl (Tl (Tl
Abb. 8.3 Karte mit den Stationen der Fahrt M 23/1 westlich von Südafrika (vgl. Abb.6.1). Punkte bezeichnen Stationen mit höchsten Konzentrationen von mehrals 100 Magnetbakterien in 50 1-11 Sedimentsuspension, Rechtecke: Stationenmit maximal 1 - 100 Magnetbakterien in 50 1-11, Dreiecke: Stationen, an denenkeine Magnetbakterien nachgewiesen wurden.
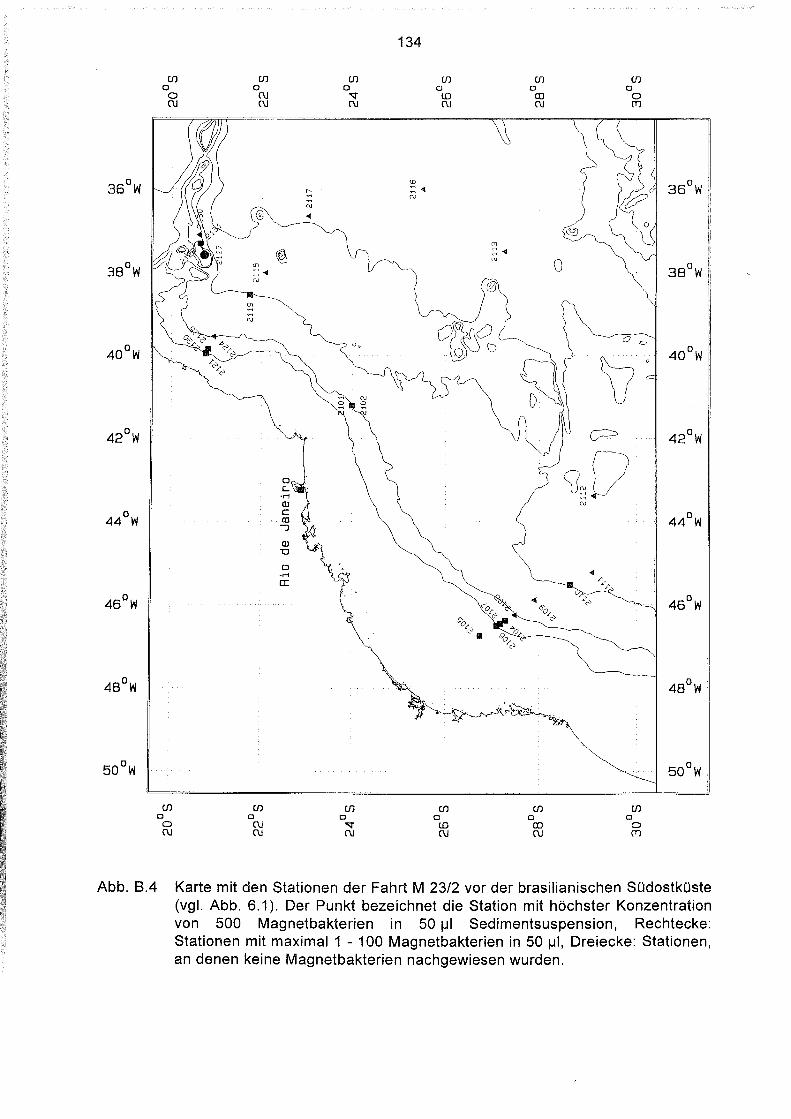
134
UJooC\J
UJo
C\JC\J
UJo"fC\J
UJo
lDC\J
UJo
coC\J
UJoo(T)
o~c) 0=
40 0W0. Q Q 400W
V0
42°W ~ 420W
')J"Dru
440W 440W
460
W ... 46°W6'0
"'0"'0
"'0.
4S0W 4S0W
UJooC\J
UJo
C\JC\J
UJo"fC\J
UJo
lDC\J
UJo
COC\J
UJoo(T)
Abb. BA Karte mit den Stationen der Fahrt M 23/2 vor der brasilianischen Südostküste(vgl. Abb. 6.1). Der Punkt bezeichnet die Station mit höchster Konzentrationvon 500 Magnetbakterien in 50 IJI Sedimentsuspension, Rechtecke:Stationen mit maximal 1 - 100 Magnetbakterien in 50 IJI, Dreiecke: Stationen,an denen keine Magnetbakterien nachgewiesen wurden.

Danksagung
Die vorliegende Arbeit wurde in der Arbeitsgruppe Geophysik des Fachbereichs 5 derUniversität Bremen angefertigt. Ich möchte allen, die durch ihre Zusammenarbeit,
Diskussionsbereitschaft und auch durch ihre moralische Unterstützung zum Gelingender Arbeit beigetragen haben, sehr herzlich danken.
Mein besonderer Dank gilt Herrn Prof. Dr. Ulrich Bleil für die Unterstützung undBetreuung während der vergangenen Jahre sowie für die sehr sorgfältige Durchsichtdes Manuskripts.
Herrn Prof. Dr. Bo Barker J0rgensen danke herzlich ich für seine Diskussionsbeiträge
und die Bereitschaft, diese Arbeit zu beurteilen.
Wesentliche Teile dieser Arbeit konnten nur durch die gute Zusammenarbeit mitRonnie Glud und Jens Gundersen vom Max-Planck-Institut für Marine Mikrobiologie,Bremen, Andreas Dahmke, Jens Sagemann, Frank Skowronek und Matthias Zabel vonder Arbeitsgruppe Geochemie sowie Herrn Dr. Peter Müller von der ArbeitsgruppeMeeresgeologie entstehen. Für die vielen Diskussionen und vor allem für dieÜberlassung ihrer Daten möchte ich ihnen allen ganz herzlich danken.
Besonders hervorheben möchte ich meine Freunde Nils Andersen, Tilo v. Dobeneck,Gerd Fischer, Mark Schwenke und Margarete Metzger. Für die Unterstützung und Hilfe,die ich durch sie erfahren habe, möchte ich mich ganz herzlich bedanken.
Ein besonderer Dank gilt auch meinen Eltern, die meine Ausbildung ermöglicht habenund meiner Freundin, die so viele Urlaube mit mir ausfallen lassen mußte.
Die Arbeit wurde finanziell vom Bundesministerium für Forschung und Technologie
(BMFT) und der Deutschen Forschungsgemeinschaft (DFG) im Rahmen desSonderforschungsbereiches 261 'Der Südatlantik im Spätquartär: Rekonstruktion von
Stoffhaushalt und Stromsystemen' unterstützt.


In dieser Reihe bereits erschienen:
Nr.l
Nr.2
Nr. 3
Nr.4
Nr.5
Nr.6
Nr.7
Nr.8
Nr.9
Wefer, G., E. Suess und Fahrtteilnehmer
Bericht über die POLARSTERN-Fahrt ANT IV/2, Rio de Janeiro - Punta Arenas,
6.11. - 1.12.1985.
60 Seiten, Bremen, 1986.
Hoffmann, G.
Holozänstratigraphie und KüstenlinienverIagerung an der andalusischen Mittelmeerkiiste.
173 Seiten, Bremen, 1988.
Wefer, G., U. BleH, P.J. Müller, H.D. Schulz, W.H. Berger, U. Brathauer, L. Bruck,
A. Dahmke, K. Dehning, M.L. Duarte-Morais, F. Fürsich, S. Hinrichs, K. Klockgeter,
A. Kölling, C. Kothe, J.F. Makaya, H. Oberhänsli, W. Oschmann, J. Posny, F. Rostek,
H. Schmidt, R. Schneider, M. Segl, M. Sobiesiak, T. Soltwedel, V. Spieß
Bericht über die METEOR-Fahrt M 6/6, Libreville - Las Palmas, 18.2. - 23.3.1988.
97 Seiten, Bremen, 1988.
Wefer, G., G.F. Lutze, T.J. Müller, O. Pfannkuehe, W. Schenke, G. Siedler, W. Zenk
Kurzbericht über die METEOR-Expedition Nr. 6, Hamburg - Hamburg,
28.10.1987 - 19.5.1988.
29 Seiten, Bremen, 1988.
Fischer, G.
Stabile Kohlenstoff-Isotope in partikulärer organischer Substanz aus dem Südpolarmeer
(Atlantischer Sektor).
161 Seiten, Bremen, 1989.
Berger, W.H. und G. Wefer
Partikelfluß und Kohlenstoffkreislauf im Ozean.
Bericht und Kurzfassungen über den Workshop vom 3.-4. Juli 1989 in Bremen.
57 Seiten, Bremen, 1989.
Wefer, G., U. Bleil, H.D. Schulz, W.H. Berger, T. Bickert, L. Bruck, u. Claussen,
A. Dahmke, K. Dehning, Y.H. Djigo, S. Hinrichs, C. Kothe, M. Krämer, A. Lücke,
S. Matthias, G. Meinecke, H. Oberhänsli, J. Pätzold, U. Pflaumann, U. Probst,
A. Reimann, F. Rostek, H. Schmidt, R. Schneider, T. Soltwedel, V. Spieß
Bericht über die METEOR - Fahrt M 9/4, Dakar - Santa Cruz, 19.2. - 16.3.1989.
103 Seiten, Bremen, 1989.
Kölling, M.
ModelIierung geochemischer Prozesse im Sickerwasser und Grundwasser.
135 Seiten, Bremen, 1990.
Heinze, P.-M.
Das Auftriebsgeschehen vor Peru im Spätquartär.
204 Seiten, Bremen, 1990.

Nr.l0
Nr.11
Nr.12
Nr.13
Nr.14
Nr.15
Nr.16
Nr.17
Nr. 18
Willems, H., G. Wefer, M. Rinski, B. Donner, H.-1. Bellmann, L. Eißmann, A. Müller,
B.W. Flemming, H.-C. Höfle, 1. Merkt, H. Streif, G. Hertweck, H. Kuntze, 1. Schwaar,
W. Schäfer, M.-G. Schulz, F. Grube, B. Menke
Beiträge zur Geologie und Paläontologie Norddeutschlands: Exkursionsführer.
202 Seiten, Bremen, 1990.
Wefer, G., N. Andersen, U. Blei!, M. Breitzke, K. Dehning, G. Fischer, C. Kothe,
G. Meinecke, P.J. Müller, F. Rostek, 1. Sagemann, M. Scholz, M. Segl, W. Thiessen
Bericht über die METEOR-Fahrt M 12/1, Kapstadt - Funchal, 13.3.1990 - 14.4.1990.
66 Seiten, Bremen, 1990.
Dahmke, A., H.D. Schulz, A. Kölling, F. Kracht, A. Lücke
Schwermetallspuren und geochemische Gleichgewichte zwischen Porenlösung und
Sediment im Wesermündungsgebiet.
BMFT-Projekt MFU 0562, Abschlußbericht.
121 Seiten, Bremen, 1991.
Rostek, F.
Physikalische Strukturen von Tiefseesedimenten des Südatlantiks und ihre Erfassung in
Echolotregistrierungen.
209 Seiten, Bremen, 1991.
Baumann, M.
Die Ablagerung von Tschemobyl-Radiocäsium in der Norwegischen See und in der Nordsee.
133 Seiten, Bremen, 1991.
Kölling, A.Frühdiagenetische Prozesse und Stoff-Flüsse in marinen und ästuarinen Sedimenten.
140 Seiten, Bremen, 1991.
SFB 261 (Hrsg.)
1. Kolloquium des Sonderforschungsbereichs 261 der Universität Bremen (14.Juni 1991):
Der Südatlantik im Spätquartär: Rekonstruktion von Stoftbaushalt und Stromsystemen.
Kurzfassungen der Vorträge und Poster.
66 Seiten, Bremen, 1991.
Pätzold, 1., T. Bickert, L. Brück, C. Gaedicke, K. Heidland, G. Meinecke, S. Mulitza
Bericht und erste Ergebnisse über die METEOR-Fahrt M 15/2, Rio de Janeiro - Vitoria,
18.1. -7.2.1991.
46 Seiten, Bremen, 1993.
Wefer, G., N. Andersen, W. Balzer, U. Bleil, L. Brück, D. Burda, A. Dahmke, B. Donner,
T. Felis, G. Fischer, H. Gerlach, L. Gerullis, M. Hauf, R. Henning, S. Kemle, C. Kothe,
R. Melyooni, F. Pototzki, H. Rode, 1. Sagemann, M. Schlüter, M. Scholz, V. Spieß,
U. Treppke
Bericht und erste Ergebnisse über die METEOR-Fahrt M 16/1, Pointe Noire - Recife,
27.3. - 25.4.1991.
120 Seiten, Bremen, 1991.

Nr.19
Nr. 20
Nr.21
Nr.22
Nr.23
Nr.24
Nr.25
Nr.26
Schulz, H.D., N. Andersen, M. Breitzke, D. Burda, K. Dehning, V. Diekamp, T. Felis,
H. Gerlach, R. Gumprecht, S. Hinrichs, H. Petermann, F. Pototzki, U. Probst, H. Rode,
l Sagemann, U. Schinzel, H. Schmidt, R. Schneider, M. Segl, B. Showers, M. Tegeler,
W. Thiessen, U. Treppke
Bericht und erste Ergebnisse über die METEOR-Fahrt M 16/2, Recife - Belem,
28.4. - 20.5.1991.
149 Seiten, Bremen, 1991.
Bemer,H.
Mechanismen der Sedimentbildung in der Fram-Straße, im Arktischen Ozean und in der
Norwegischen See.
167 Seiten, Bremen, 1991.
Schneider, R.
Spätquartäre Produktivitätsänderungen im östlichen Angola-Becken: Reaktion auf Variationen
im Passat-Monsun-Windsystem und in der Advektion des Benguela-Küstenstroms.
198 Seiten, Bremen, 1991.
Hebbeln, D.
Spätquartäre Stratigraphie und Paläozeanographie in der Fram-Straße.
174 Seiten, Bremen, 1991.
Lücke, A.
Umsetzungsprozesse organischer Substanz während der Frühdiagenese in ästuarinen
Sedimenten.
137 Seiten, Bremen, 1991.
Wefer, G., D. Beese, W.H. Berger, U. Blei!, H. Buschhoff, G. Fischer, M. Kalberer, S. Kemle
von Mücke, B. Kemtopf, C. Kothe, D. Lutter, B. Pioch, F. Pototzki, V. Ratmeyer, U. Rosiak,
W. Schmidt, V. Spieß, D. Völker
Bericht und erste Ergebnisse der METEOR-Fahrt M 20/1, Bremen - Abidjan,
18.11.1991- 22.12.1991.
74 Seiten, Bremen, 1992.
Schulz, H.D., D. Beese, M. Breitzke, L. Brück, B. Brügger, A. Dahmke, K. Dehning,
V. Diekamp, B. Donner, I. Ehrhardt, H. Gerlach, M. Giese, R. Glud, R. Gumprecht,
l Gundersen, R. Henning, H. Petermann, M. Richter, J. Sagemann, W. Schmidt, R. Schneider,
M. Segl, U. Wemer, M. Zabel
Bericht und erste Ergebnisse der METEOR-Fahrt M 20/2, Abidjan - Dakar,
27.12.1991 - 3.2.1992.
173 Seiten, Bremen, 1992.
Gingeie, F.
Zur klimaabhängigen Bildung biogener und terrigener Sedimente und ihrer Veränderung
durch die Frühdiagenese im zentralen und östlichen Südatlantik.
202 Seiten, Bremen, 1992.

Nr.27
Nr.28
Nr.29
Nr.30
Nr.31
Nr.32
Nr. 33
Nr.34
Nr.35
Nr.36
Nr.37
Bickert, T.
Rekonstruktion der spätquartären Bodenwasserzirkulation im östlichen Südatlantik über
stabile Isotope benthischer Foraminiferen.
205 Seiten, Bremen, 1992.
Schmidt, H.Der Benguela-Strom im Bereich des Walfisch-Rückens im Spätquartär.
172 Seiten, Bremen, 1992.
Meinecke, G.
Spätquartäre Oberflächenwassertemperaturen im östlichen äquatorialen Atlantik.
181 Seiten, Bremen, 1992.
Bathmann, u., U. Bleil, A. Dahmke, P. Müller, A. Nehrkom, E.-M. Nöthig, M. Olesch,
1. Pätzold, H.D. Schulz, V. Smetacek, V. Spieß, G. Wefer, H. Willems
Bericht des Graduierten Kollegs. Stoff-Flüsse in marinen Geosystemen.
Berichtszeitraum Oktober 1990 - Dezember 1992.
396 Seiten, Bremen, 1992.
Damm,E.
Frühdiagenetische Verteilung von Schwermetallen in Schlicksedimenten der westlichen Ostsee.
115 Seiten, Bremen, 1992.
Antia, E.E.
Sedimentology, Morphodynamics and Facies Association of a mesotidal Barrier Island
Shoreface (Spiekeroog, Southem North Sea).
370 Seiten, Bremen, 1993.
Duinker, 1. und G. Wefer (Hrsg.)
Bericht über den 1. JGOFS-Workshop. 1.12. Dezember 1992 in Bremen.
83 Seiten, Bremen, 1993.
Kasten, S.
Die Verteilung von SChwermetallen in den Sedimenten eines stadtbremischen Hafenbeckens.
103 Seiten, Bremen, 1993.
Spieß, V.
Digitale Sedimentographie. Neue Wege zu einer hochauflösenden Akustostratigraphie.
199 Seiten, Bremen, 1993.
Schinzel, U.
Laborversuche zu frühdiagenetischen Reaktionen von Eisen (III) - Oxidhydraten in
marinen Sedimenten.
189 Seiten, Bremen, 1993.
Sieger, R.
eoTAM - ein Modell zur ModelIierung des Schwermetalltransports in Grundwasserleitern.
56 Seiten, Bremen, 1993.

Nr.38
Nr.39
Nr.40
Nr.41
Nr.42
Nr.43
Nr.44
Nr.45
Willems, H. (Ed.)
Geoscientific Investigations in the Tethyan Himalayas.
183 Seiten, Bremen, 1993.
Harner, K.
Entwicklung von Laborversuchen als Grundlage rur die Modellierung des Transportverhaltens
von Arsenat, Blei, Cadmium und Kupfer in wassergesättigten Säulen.
147 Seiten, Bremen, 1993.
Sieger, R.
Modellierung des Stoffiransports in porösen Medien unter Ankopplung kinetisch gesteuerter
Sorptions- und Redoxprozesse sowie thermischer Gleichgewichte.
158 Seiten, Bremen, 1993.
Thießen, W.
Magnetische Eigenschaften von Sedimenten des östlichen SüdatIantiks und ihre
paläozeanographische Relevanz.
170 Seiten, Bremen, 1993.
Spieß, v., A Abelmann, T. Bickert, 1. Brehme, R. Cordes, AP. Cavalcanti de C. Laier,
K. Dehning, T. v.Dobeneck, B. Donner, 1. Ehrhardt, M. Giese, 1. Grigel, R. Haese, W. Haie,
S. Hinrichs, S. Kasten, C. Knaack, W.-T. Ochsenhirt, H. Petermann, R. Rapp, M. Richter,
1. Rogers, A Schmidt, M. Scholz, F. Skowronek, M. Teixeira de Oliveira, M. Zabel
Report and preliminary results ofMETEOR-Cruise M 23/1, Kapstadt - Rio de Janeiro,
4.2.1993 - 25.2.1993.
Bremen, 1994 (im Druck).
Bleil, u., A Ayres Neto, D. Beese, M. Breitzke, K. Dehning, V. Diekarnp, T. v.Dobeneck,
A Figueiredo, M. Pimentel Esteves, M. Giese, R. Glud, 1. Grigel, J. Gundersen, R. Haese,
S. Hinrichs, S. Kasten, G. Meinecke, S. Mulitza, H. Petermann, R. Petschik, R. Rapp,
M. Richter, C. RühIemann, M. Scholz, K. Wallmann, M. Zabel
Report and preliminary results ofMETEOR-Cruise M 23/2, Rio de Janeiro - Recife,
27.2.1993 - 19.3.1993
133 Seiten, Bremen, 1994.
Wefer, G., D. Beese, W.H. Berger, K. Buhlmann, H. Buschhoff, M. Cepek, V. Diekamp,
G. Fischer, E. Holmes, S. Kemle-von Mücke, B. Kemtopf, C.B. Lange, S. Mulitza,
W.-T. Ochsenhirt, R. Plugge, V. Ratmeyer, C. Rühlemann, W. Schmidt, M. Schwarze,
C. Wallmann, M. Zabel
Report and preliminary results ofMETEOR-Cruise M 23/3, Recife - Las Palmas,
21.3.1993 - 12.4.1993
71 Seiten, Bremen, 1994.
Giese, M. und G. Wefer (Hrsg.)
Bericht über den 2. JGOFS-Workshop. 18../19. November 1993 in Bremen.
93 Seiten, Bremen, 1994.

Nr. 46
Nr.47
Nr. 48
Nr.49
Nr.50
Nr.51
Nr.52
Nr.53
Nr.54
Nr.55
Balzer, W., M. Bleckwehl, H. Buschhoff, G. Fischer, F.G. Palma, M. Kalberer, U. Kuller,
V. Ratmeyer, U. Rosiak, D. Schneider, A. Zimmermann
Report and preliminary results ofMETEOR-Cruise M 22/1, Hamburg - Recife,
22.9. - 21.10.1992.24 Seiten, Bremen, 1994.
Stax, R.
Zyklische Sedimentation von organischem Kohlenstoff in der Japan See: Anzeiger für
Änderungen von Paläoozeanographie und Paläoklima im Spätkänozoikum.
150 Seiten, Bremen, 1994.
Skowronek, F.
Frühdiagenetische Stoff-Flüsse gelöster Schwermetalle an der Oberfläche von Sedimenten
des Weser Ästuares.
107 Seiten, Bremen, 1994 (im Druck).
Dersch-Hansmann, M.
Zur Klimaentwicklung in Ostasien während der letzten 5 Millionen Jahre:
Terrigener Sedimenteintrag in die Japan See (ODP Ausfahrt 128).
149 Seiten, Bremen, 1994 (im Druck).
Zabel, M.
Frühdiagenetische Stoff-Flüsse in Oberflächen-Sedimenten des äquatorialen und
östlichen Südatlantik.
129 Seiten, Bremen, 1994 (in Vorbereitung).
Bleil, U. et al.
Bremen, 1994 (in Vorbereitung).
Symposium
The South AtIantic: Present and Past Circulation.
Bremen, Germany, 15 - 19 August 1994.
Abstracts
167 Seiten, Bremen, 1994.
Kretzmann, U.B.
57Fe-Mössbauer-Spektroskopie an Sedimenten
Möglichkeiten und Grenzen.
183 Seiten, Bremen, 1994 (im Druck).
Bachmann, M.
Die Karbonatrampe von Organya im oberen Oberapt und unteren Unteralb (NE-Spanien,
Provo Lerida): Fazies, Zyklo- und Sequenzstratigraphie.
147 Seiten, Bremen, 1994 (im Druck).
Kemle-von Mücke, S.
Oberflächenwasserstruktur und -zirkulation des SüdostatIantiks im Spätquartär.
151 Seiten, Bremen, 1994 (im Druck).





























