Embed Size (px)
Citation preview

Aus dem Institut für Pathologie
der Universität zu Lübeck
Direktor: Prof. Dr. med. Alfred Christian Feller
Charakterisierung von angioimmunoblastischen T-Zell -Lymphomen
auf morphologischer, immunhistochemischer und mole kularer Ebene
mit Schwerpunkt auf dem microRNA-Expressionsprofil
Inauguraldissertation
zur
Erlangung der Doktorwürde
der Universität Lübeck
-Aus der Sektion Medizin-
vorgelegt von
Katharina Reddemann
aus Bonn
Lübeck 2015


1. Berichterstatter: Prof. Dr. med. Christoph Thorns
2. Berichterstatter: Prof. Dr. rer. nat. Heinz Fehrenbach
Tag der mündlichen Prüfung: 14.09.2015
Zum Druck genehmigt. Lübeck, den 14.09.2015
Promotionskommission der Sektion Medizin


I
Inhaltsverzeichnis Abbildungsverzeichnis ............................. ................................................................ IV
Tabellenverzeichnis ............................... .................................................................... V
Abkürzungsverzeichnis ............................. ............................................................... VI
1. Einleitung ........................................ ............................................................. 1
1.1. Grundlagen ................................................................................................... 1
1.1.1. Lymphome .................................................................................................... 1
1.1.1.1. Periphere T-Zell Lymphome .......................................................................... 2
1.1.1.2. Angioimmunoblastische T-Zell-Lymphome .................................................... 4
1.1.2. microRNA .................................................................................................... 12
1.1.2.1. microRNA-Physiologie ................................................................................ 12
1.1.2.2. microRNA-Dysregulation ............................................................................. 15
1.2. Fragestellung ...............................................................................................17
2. Material und Methoden ............................. .................................................18
2.1. Material ........................................................................................................18
2.2. Methoden .....................................................................................................18
2.2.1. Studiengruppe ............................................................................................. 18
2.2.2. Patientenproben .......................................................................................... 18
2.2.3. Histopathologische Methoden ..................................................................... 19
2.2.3.1. Herstellung des Tissue Microarrays ............................................................ 19
2.2.3.2. Herstellung von Paraffin-Schnittpräparaten ................................................. 20
2.2.3.3. Histochemische Färbungen von formalinfixierten, paraffineingebetteten
Gewebsschnitten ......................................................................................... 20
2.2.3.4. Immunhistochemische Färbung von formalinfixierten, paraffineingebetteten
Gewebsschnitten ......................................................................................... 21
2.2.4. Molekularbiologische Methoden .................................................................. 23
2.2.4.1. Nachweis von Epstein-Barr-Virus kodierter RNA (EBER) in formalinfixierten,
paraffineingebetteten Gewebsschnitten mittels chromogener In-situ-
Hybridisierung ............................................................................................. 23
2.2.4.2. RNA-Isolierung aus paraffineingebettetem Gewebe .................................... 23
2.2.4.3. Quantifizierung von RNA in wässriger Lösung ............................................. 23
2.2.4.4. Relative Quantifizierung der microRNA-Expression mittels RT-qPCR ......... 24
2.2.4.5. Reverse Transkription (RT) – Polymerase- Ketten-Reaktion (RT-PCR) ....... 24
2.2.4.6. Quantitative Real Time- Polymerase-Kettenreaktion (qPCR) ...................... 25

II
2.2.5. Statistische Analysen der RT-qPCR- Ergebnisse ........................................ 27
2.2.5.1. Präprozessierung ....................................................................................... 28
2.2.5.2. Quantilnormalisierung ................................................................................. 28
2.2.5.3. Fall-Kontroll-Analyse ................................................................................... 30
3. Ergebnisse ........................................ ......................................................... 31
3.1. Charakteristika der Patienten/innen ............................................................. 31
3.2. Ergebnisse der histopathologischen Analysen ............................................ 31
3.2.1. Ergebnisse der Analyse der Gewebsschnitte hinsichtlich des histologischen
Musters und der Proliferation der hochendothelialen Venolen und der FDC-
Netzwerke ................................................................................................... 31
3.2.2. Ergebnisse der immunhistochemischen Analysen anhand des tissue
microarray (TMA) ........................................................................................ 36
3.3. Ergebnisse der EBER-ISH .......................................................................... 41
3.4. Ergebnisse der microRNA-Quantifizierung .................................................. 42
4. Diskussion ........................................ ......................................................... 44
4.1. Morphologie - Die Proliferation der Venolen als frühes Merkmal von AITL .. 44
4.2. Immunhistochemie ...................................................................................... 45
4.2.1. Stathmin als relevanter und möglicherweise CD10 überlegener Marker in
AITL ............................................................................................................ 45
4.2.2. LMO2 wird nicht von AITL-Zellen exprimiert ................................................ 46
4.3. MicroRNA-Expression ................................................................................. 47
4.3.1. Mögliche Oncomir in AITL ........................................................................... 47
4.3.2. Mögliche Tumor suppressor-miR in AITL .................................................... 54
4.3.3. Vorsicht bei der Auswahl der endogenen Kontrolle und
Normalisierungsmethode in AITL ................................................................ 56
4.3.4. MicroRNA und Subgruppen ......................................................................... 57
4.4. Limitationen der Arbeit und Ausblick............................................................ 59
5. Zusammenfassung ................................... ................................................. 62
6. Literaturverzeichnis .............................. .................................................... 64

III
7. Anhang ............................................ ............................................................81
7.1. Material ........................................................................................................81
7.1.1. Geräte ......................................................................................................... 81
7.1.2. Verbrauchsmaterialien ................................................................................ 82
7.1.3. Reagenzien und Chemikalien ...................................................................... 83
7.1.4. Kits 84
7.1.5. Antikörper .................................................................................................... 84
7.1.6. Primer ......................................................................................................... 85
7.1.7. Software ...................................................................................................... 85
7.1.8. In silico Ressourcen .................................................................................... 85
7.2. Ergänzende Tabellen und Abbildungen ........................................................86
8. Danksagung ........................................ ........................................................97
9. Lebenslauf ........................................ ..........................................................98
10. Erklärung ......................................... ......................................................... 100

IV
Abbildungsverzeichnis
Abbildung 1 Biosynthese und Wirkungsweise von microRNAs ............................ 14
Abbildung 2 Anordnung der Gewebsstanzen im tissue microarray ...................... 20
Abbildung 3 Prinzip der RT-qPCR ....................................................................... 27
Abbildung 4 Histomorphologisches Muster der AITL (n=30) ................................ 32
Abbildung 5 Histomorphologisches Bild eines AITL ............................................ 33
Abbildung 7 Bewertung der Expansion der FDC-Netzwerke ............................... 35
Abbildung 8 Bewertung der Proliferation der hochendothelialen Venolen............ 35
Abbildung 9 Durchschnittswerte des Scorings..................................................... 36
Abbildung 10 Immunhistochemische Färbung mit CD3- Antikörper ...................... 38
Abbildung 11 Immunhistochemische Färbung mit CD20- Antikörper ..................... 38
Abbildung 12 Immunhistochemische Färbung mit Stathmin- Antikörper ................ 39
Abbildung 13 Immunhistochemische Färbung mit CD10- Antikörper ..................... 39
Abbildung 14 Immunhistochemische Färbung mit PD1- Antikörper ...................... 40
Abbildung 15 Färbeverhalten der neoplastischen T-Zellen in AITL für die Antikörper
gegen Stathmin, PD1 und CD10 ..................................................... 40
Abbildung 16 In-situ Hybridisierung für EBER ....................................................... 41
Abbildung 17 Boxplot der CT-Werte pro Fall und Platte nach Präprozessierung ... 29
Abbildung 18 Boxplots der CT-Werte pro Fall und Platte nach
Quantilnormalisierung ...................................................................... 29
Abbildung 19 Boxplots der CT-Werte der signifikant aberrant exprimierten
microRNAs nach ansteigenden adjustierten p-Werten ..................... 43
Abbildung 20 TaqMan Human MicroRNA Array A ................................................. 88
Abbildung 21 TaqMan Human MicroRNA Array B ................................................. 89
Abbildung 22 Boxplot der CT-Werte pro Platte ...................................................... 90

V
Tabellenverzeichnis
Tabelle 1 Periphere T-Zell Lymphome nach aktueller WHO-Klassifikation .......... 3
Tabelle 2 Nomenklatur der microRNAs ..............................................................15
Tabelle 3 Histochemische Färbungen ................................................................21
Tabelle 4 Antikörper (mit Verdünnungen und Vorbehandlungen) .......................22
Tabelle 5 Master Mix für die RT-PCR (Ansatz für eine Probe) ...........................24
Tabelle 6 Thermocycler Programm (Reverse Transkription) ..............................25
Tabelle 7 Ansatz für die Real-Time PCR ............................................................26
Tabelle 8 Graduierung der Proliferation der hochendothelialen Venolen und der
FDC-Netzwerke mit Zuordnung der Score-Werte ...............................32
Tabelle 9 Grenzwerte der immunhistochemischen Färbungen ........................ 37
Tabelle 10 Signifikant aberrant exprimierte RNAs in der Fall-Kontroll-Analyse .....42
Tabelle 11 Geräte ................................................................................................82
Tabelle 12 Verbrauchsmaterialien ........................................................................82
Tabelle 13 Reagenzien und Chemikalien .............................................................84
Tabelle 14 Kits .....................................................................................................84
Tabelle 15 Antikörper ...........................................................................................84
Tabelle 16 Primer .................................................................................................85
Tabelle 17 Software .............................................................................................85
Tabelle 18 In-silico Resourchen ...........................................................................85
Tabelle 19 Patienten-Charakteristika und Ergebnisse der Graduierungen ...........86
Tabelle 20 Ergebnisse der immunhistochemischen Auswertung anhand des tissue
microarrays .........................................................................................87
Tabelle 21 Charakteristika der Kontrollgruppe......................................................87
Tabelle 22 Ergebnisse der Präprozessierung .......................................................90
Tabelle 23 CT-Werte der Lymphom- und Referenz-Fälle .....................................92
Tabelle 24 Top 10 microRNAs für EBER ..............................................................93
Tabelle 25 Top 10 microRNAs für Stathmin .........................................................93
Tabelle 26 Top 10 microRNAs für CD10 ..............................................................94
Tabelle 27 Top 10 microRNAs für PD1 ................................................................94
Tabelle 28 Top 10 microRNAs für die Proliferation der HEV ................................95
Tabelle 29 Top 10 der microRNAs für die Expansion der FDC-Netzwerke ...........95
Tabelle 30 Top 10 der microRNAs für das histomorphologische Muster ..............96

VI
Abkürzungsverzeichnis
Abkürzung Erklärung
ACID Activation induced cell death
ACVBP Doxorubicin, Cyclophosphamid,
Vindesine Bleomycin, Prednison
Ago Argonaut-Protein
AITL Angioimmunoblastisches T-Zell-Lymphom
ALCL Anaplastic large cell lymphoma
ALK Anaplastic lymphoma kinase
AML Akute myeloische Leukämie
Bcl B-cell lymphoma
CD Cluster of differentiation
cDNA Complementary DNA
CGH Comparative genomic hybridization
CHOP Cyclophosphamid, Hydroxydaunorubicin,
Vincristin, Prednison
CP Crossing point
CT Threshold cycle
CTLA4 Cytotoxic T-Lymphocyte Antigen 4
CXC-L/R C-X-C motif chemokin/receptor
DGCR8 DiGeorge Critical Region Gene 8
DNA Desoxyribonucleic Acid
DNMT3A DNA(cytosine-5)-methyltransferase 3A
EBER Epstein-Barr-Virus codierte RNA
EBV Epstein-Barr-Virus
ERK Extracellular-signal regulated kinase
EGFR Epidermal Growth Factor Receptor
FAME Functional assignment of microRNA via
enrichment analysis
Fc Crystallisable Fragment
FDC Follicular dendritic cell
GELA Groupe d’Etudes des lymphomes de
l’Adulte
GSEA Gene set enrichement analysis
VII
GTP Guanosintriphosphat
HCC Hepatocellular carcinoma
HE Hematoxilin Eosin
HIV Human inmmunodeficiency Virus
HRP Horseradish Peroxidase
HTLV Human T-lymphotropic virus
ICOS Inducible t-cell co-stimulator
IDH2 Isocitratdehydrogenase
IGFR1 Insulin-like Growth Factor Receptor
IL Interleukin
IPI Internationaler Prognose Index
K-RAS Kirsten rat sarcoma
KSHV Kaposi-sarcoma Herpes Virus
LDH Laktatdehydrogenase
LMO2 LIM domain only 2
LMP1 Latent membrane protein
MALT Mucosa associated lymphatic tissue
mBACOD Methotrexat, Bleomycin, Doxorubicin,
Cyclophosphamid, Vincristin,
Dexamethason
MDS Myelodysplastisches Syndrom
Min. Minute
MIQE Minimum information for publication of
quantitative Real-time PCR Experiments
miRNA/miR microRNA
mM Milli molar
mRNA Messenger RNA
MTOC Mikrotubulus organizaing center
MYC Myelocytomatose-Gen
nm Nanometer
ng Nanogramm
NFkappaB Nuclear factor kappa light chain enhancer
of activated B-cells
NK Natural killer
NOS Not otherwise specified
NF1 Neurofibromin 1

VIII
NPM Nukleophosmin
Pri-miRNA Primary microRNA
P53 Tumor protein 53
PAS Periodic Schiff Acid
PCR Polymerase Chain Reaction
PD1 Programmed cell death
PIAI Prognosis in AITL
PIT Prognosis Index for T-cell lymphomas
PNEN Pancreatic neuroendocrine neoplasia
PTCL Peripheral t-cell lymphoma
qPCR Quantitative Real-Time PCR
Ras Rat sarcoma (proto-Onkogen)
RISC RNA-induced silencing complex
RNU44 Nucleoläre RNA 44
RT Reverse Transkription
SAP Slam-associated protein
Sec. Sekunde
SEER Surveillance, Epidemiology and End-
Results
siRNA Silencing RNA
SIRT1 Sirtuin-1
snoRNA Small nucleolar RNA
SOX2 Sex-determining region Y-box 2
STAT Signal Transducers and Activators of
Transkription
TCR T-cell receptor
TET2 Ten-eleven Translocation 2 (-Gen)
Tfh Follikuläre T-Helfer Zelle
TNF Tumor necrosis factor
TNM Tumor, Nodus, Mestastasis
(Klassifikation)
TMA Tissue microarray
TRAF TNF receptor associated factor
VEGF Vascular endothelial growth factor
VEGFR1/KDR Vascular endothelial growth factor
receptor 1/

IX
WHO World Health Organization
z.B. Zum Beispiel
°C Grad Celsius
µl Mikroliter
∆ Delta
% Prozent


1
1. Einleitung
1.1. Grundlagen
1.1.1. Lymphome
In der aktuellen Klassifikation der Weltgesundheitsorganisation (WHO) für Tumore der
hämatopoetischen und lymphatischen Gewebe definieren Jaffe et al. Lymphome als
klonale Tumore reifer oder unreifer B-Zellen, T-Zellen oder natürlicher Killer (NK)-
Zellen in verschiedenen Stadien der Differenzierung (148). Ihre Klassifikation basiert
auf der Zusammenschau aller verfügbaren Informationen, die diese Erkrankungen
charakterisieren, von Epidemiologie, geographischer Variation und klinischen
Parametern bis Gewebslokalisation, Histomorphologie, Immunphänotyp und mole-
kularem Profil (56). Morphologie und Immunphänotyp sind in der Praxis ausreichend
für die Diagnose der meisten lymphatischen Neoplasien. Jedoch ist kein antigener
Marker spezifisch für eine Neoplasie, sodass die Kombination aus Morphologie und
eines Panels mehrerer Marker für eine korrekte Diagnose notwendig sind (148).
Traditionell werden Hodgkin-Lymphome von sogenannten Non-Hodgkin-Lymphomen
unterschieden (148).
Hodgkin-Lymphome wurden im 19. Jahrhundert nach ihrem Entdecker Thomas
Hodgkin benannt (136). Heutzutage werden darunter das klassische Hodgkin-Lymph-
om und das noduläre lymphozytenprädominante Hodgkin-Lymphom zusammengefasst,
die in der Mehrzahl der Fälle von B-Zellen ausgehen (5).
Non-Hodgkin-Lymphome umfassen alle weiteren Lymphome der B- und T/NK-Zell-
Reihe, wobei B-Zell Lymphome sehr viel häufiger sind als T-Zell-Lymphome (148).
Laut Bericht „Krebs in Deutschland 2009/2010“ der deutschen Krebsregister erkrankten
im Jahr 2010 insgesamt 8590 Männer und 7640 Frauen in Deutschland an einem Non-
Hodgkin-Lymphom. Für 2014 wird nach Prognose der Krebsregister mit der
Neuerkrankung von 9400 Männern und 7900 Frauen gerechnet.
Im Vergleich zu anderen Krebserkrankungen, wie z.B. Brust- oder Lungenkrebs, sind
maligne Lymphome bei Erwachsenen jedoch eher selten. Im Jahr 2010 standen in
Deutschland Non-Hodgkin-Lymphome bei Männern auf Rang 9, bei Frauen auf Rang 8
der Krebsneuerkrankungsliste. Im Mittel waren Männer bei Diagnosestellung 69,
Frauen 71 Jahre alt (1). Etwa 6000 Menschen versterben jährlich in Deutschland an
dieser Erkrankung (1). Eindeutige Risikofaktoren für Non-Hodgkin-Lymphome lassen
sich nur selten nachweisen (1). Immunschwäche (z.B. HIV-vermittelt) sowie seltene
Autoimmunerkrankungen (wie Hashimoto-Thyroiditis) gehen allerdings mit einem
erhöhten Risiko einher. Auch Viren und andere Krankheitserreger tragen zur Ent-

2
stehung einiger Lymphome bei (1, 148): So gilt zum Beispiel der Zusammenhang
zwischen einer Epstein-Barr-Virus (EBV) – Infektion und dem vorwiegend in Afrika
auftretenden Burkitt-Lymphom als gesichert.
1.1.1.1. Periphere T-Zell Lymphome
Periphere T-Zell Lymphome (PTCL) umfassen alle Neoplasien, die sich von post-
thymischen T-Lymphozyten oder reifen natürlichen Killer (NK)-Zellen ableiten (51).
Es gibt zwei Klassen reifer CD3+ T-Zellen, die anhand der Heterodimere des T-Zell-
Rezeptors eingeteilt werden: alpha-beta und gamma-delta. Alpha-beta T-Zellen
umfassen CD4+ (zumeist Helfer) und CD8+ (zumeist zytotoxische) Subpopulationen.
Sie sind Teil des adaptiven Immunsystems und finden sich zumeist im peripheren Blut
und den peripheren lymphatischen Organen, wie Lymphknoten (68). Gamma-delta T-
Zellen (CD4-CD8-, CD4-CD8+) machen unter 5% der T-Zellen aus. Nebst NK-Zellen
gehören sie zum angeborenen Immunsystem und finden sich vor allem in Haut,
Schleimhäuten und roter Milzpulpa (51, 68).
Periphere T-Zell Lymphome sind selten und stellen etwa 15% der Non-Hodgkin-
Lymphome dar. Epidemiologisch zeigen sie wichtige geographische Variationen,
teilweise überlappend mit Endemiegebieten spezieller viraler Infektionen (159). So ist
zum Beispiel in Endemiegebieten des humanen T-lymphotropischen Virus-1 (HTLV-1)
in Japan und der Karibik, das Risiko für die adulte T-Zell Leukämie durch das Virus
erhöht (68).
Gemäß der Weltgesundheitsorganisation (WHO) werden zur Zeit 23 Entitäten
hinsichtlich Morphologie, Immunphänotyp, molekularem Profil, klinischer Parameter
und dem vermuteten normalen zellulären Pendant in der Gruppe der PTCL
unterschieden. Ein Hauptkriterium zur Klassifizierung ist der Manifestationsort der
Erkrankung: So kommen kutane, nodale und extranodale sowie leukämisch
verlaufende periphere T-Zell Lymphome vor (vgl. Tab 1).

3
PTCL Entität Häufig -
keit (%)°
Postulierte
Ursprungsszelle
Phänotyp
Dissiminiert/Leukämisch
Prolymphozytische T-Zell-Leukämie Tαß Nicht-zytotoxisch
Großzellige granulierte lymphozytische T-Zell Leukämie Tαß (seltener Tγδ) Zytotoxisch (a)
Chronische lymphoproliferative Störung der NK-Zellen* NK Zytotoxisch (a)
Systemische EBV-positive T-Zell Lymphoproliferation der
Kindheit
Tαß Zytotoxisch (a)
Adulte(s) T-Zell Leukämie/Lymphom 9,6 Tαß T regulatorisch
Extranodal
Extranodales NK-/T-Zell Lymphom (nasaler Typ) 10,4 NK (seltener Tαß
oder Tγδ)
Zytotoxisch (a)
Enteropathie-assoziiertes T-Zell Lymphom 4,7 IIL, Tαß (seltener
Tγδ)
Zytotoxisch (a)
Hepatosplenisches T-Zell Lymphom 1,4 Tγδ (seltener Tαß) Zytotoxisch (na)
Kutan
Mycosis fungoides Tαß (meist CD4) Nicht-zytotoxisch
Sézary Syndrom Tαß (meist CD4) Nicht-zytotoxisch
Primäre kutane CD30-positive T-Zell Lymphoproliferation Tαß (meist CD4)
Primäres kutanes großzelliges anaplastisches Lymphom 1,7 Tαß (CD4) Zytotoxisch (a)
Lymphomatoide Papulose Tαß (CD4) Zytotoxisch (a)
Subkutanes Pannikulitis-ähnliches T-Zell Lymphom 0,9 Tαß (CD8) Zytotoxisch (a)
Primäres kutanes γδ T-Zell Lymphom Tγδ Zytotoxisch (a)
Primäres kutanes CD8+ aggressives epidermotropisches
cytotoxisches T-Zell Lymphom*
Tαß (CD8) Zytotoxisch (a)
Primäres kutanes CD4+ kleinzelliges/mittelgroßzelliges T-
Zell Lymphom*
Tαß (CD4, Tfh) Zytotoxisch (a)
Hidroa vacciniformis-ähnliches Lymphom Tαß (selten NK) Zytotoxisch
Nodal
Peripheres T-Zell Lymphom, nicht anderweitig spezifiziert
(NOS)
25,9 Tαß (CD4>CD8),
selten Tγδ
Variabel, Tfh,
zytotoxisch (a)
Angioimmunoblastisches T-Zell Lymphom 18,5 Tαß (CD4, Tfh) Tfh
Großzelliges anaplastisches Lymphom, ALK-positiv 6,6 Tαß (Th2?) Zytotoxisch (a)
Großzelliges anaplastisches Lymphom, ALK-negativ* 5,5 Tαß (Th2?) Zytotoxisch (a)
Tabelle 1 Periphere T-Zell Lymphome nach aktueller WHO-Klassifikation
Modifiziert nach (51), *provisorische Entitäten, °Statistik basierend auf pathologisch-anatomischen Registrierungen mit Unterrepräsentanz der leukämischen und kutanen Entitäten, Abkürzungen (nicht im Verzeichnis): a=aktiviert, na=nicht aktiviert

4
Die variable Morphologie und das Fehlen definierter diagnostischer Marker für die
meisten der Subgruppen macht die Diagnose und Klassifizierung der Erkrankungen
sehr anspruchsvoll und behindert ihre Erforschung (67). Zusätzlich zur Rarität,
Diversität und Heterogenität der PTCL, die das Zusammenstellen einer großen
Patientenkohorte erschweren, enthalten die Tumorproben zumeist eine Vielzahl
reaktiver T-Zellen (35). Außerdem sind nur wenige Zelllinien und gute Tiermodelle
verfügbar, sodass das Verständnis der Pathobiologie der PTCL weit hinter dem Wissen
zu malignen B-Zell Neoplasien zurückbleibt (35).
Mit den momentan geläufigen immunhistochemischen und molekularen Markern sind
30 bis 50% der peripheren T-Zell-Lymphom-Fälle bisher nicht klassifizierbar und
werden als PTCL, not otherwise specified, (PTCL- NOS) kategorisiert (67).
Mit Ausnahme der Translokation t(2;5)(p23;q35) in ALK-positiven anaplastischen
großzelligen Lymphomen, sind bisher wenige spezifische diagnostische genetische
Veränderungen für die Subgruppen der peripheren T-Zell-Lymphome berichtet worden.
Mithilfe von Gen-Expressionanalysen konnten reproduzierbare genetische Signaturen
einiger PTCL-Subgruppen gezeigt werden, die zur Klassifizierung der T-Zell
Lymphome beitragen (6). So gibt es vermehrt Hinweise dafür, dass die Ursprungs-
zellpopulation, ein wichtiger Faktor für die Biologie der PTCL sind (51).
1.1.1.2. Angioimmunoblastische T-Zell-Lymphome Die Erkrankung, die wir heute als angioimmunoblastisches T-Zell Lymphom (AITL)
bezeichnen, wurde in den 1970er Jahren unter anderem von Frizzera et al. und
Lennert erstmals beschrieben (47, 87).
Das angioimmunoblastische T-Zell Lymphom ist mit 18,5% eines der zwei häufigsten
peripheren T-Zell Lymphome weltweit (159). Geographisch finden sich
angioimmunoblastische T-Zell Lymphome häufiger in Europa (29%) als in Nordamerika
(16%) oder Asien (18%) (135, 159). Bislang sind jedoch die Ätiologie, Risikofaktoren
oder eine ethnische Prädisposition nicht identifiziert worden (106).
Die Erkrankung betrifft zumeist ältere Erwachsene mit einem mittleren Alter von 69
Jahren (Spanne von 10 bis 96 Jahren) bei Diagnosestellung. Je nach Studie sind
Männer und Frauen entweder gleich häufig betroffen oder aber die Inzidenz wird für
Männner als etwas höher angegeben (39, 165).
Die Symptomatik des angioimmunoblastischen T-Zell Lymphoms geht weniger auf
direkte Komplikationen des Tumorwachstums als auf ein dereguliertes und
geschwächtes Immunsystem zurück (35).

5
Häufig manifestiert sich die Erkrankung mir einer generalisierten Lymphadenopathie,
oft assoziiert mit B-Symptomen (Fieber, Gewichtsverlust und Nachtschweiß) (39). Viele
der Patienten zeigen zudem eine Hepatosplenomeglie. In bis zu 50% der Fälle
kommen Hauteffloreszenzen, oft in Verbindung mit Juckreiz, vor (36). Seltener wurden
unter anderem arthritische Beschwerden, Pleuraergüsse und Aszites sowie
neurologische, pulmonale und gastrointestinale Symptome beschrieben (36, 108).
Häufige laborchemische Befunde sind Autoimmunphänomene, wie hämolytische
Anämie (Coombs-positiv) oder der Nachweis von Autoantikörpern (Kälteagglutinine,
Rheumafaktoren, Anti-nukleäre Antikörper, Anti-Smooth muscle Antikörper) im Blut (43,
106). Außerdem finden sich häufig eine Thrombozytämie und Lymphopenie sowie
erhöhte Laktatdehydrogenase (LDH)- Werte, eine Eosinophilie, eine erhöhte
Blutsenkungsgeschwindigkeit oder auch eine polyklonale Hypergammaglobulinämie
(106). Eine Beteiligung des Knochenmarks (in bis zu 70% der Patienten vorhanden)
signalisiert eine höhere Tumorzelllast und korreliert mit einer höheren Inzidenz von B-
Symptomen, Hepatosplenomegalie und zirkulierenden Tumorzellen (29).
In über 80% der Fälle wird die Diagnose erst in einem fortgeschrittenen Stadium (Ann
Arbor Stadien III oder IV) gestellt (36, 43).
Die Prognose des AITL ist insgesamt schlecht, die mittlere Überlebenszeit beträgt nur
etwa 3 Jahre, unabhängig von der Therapie (106). Jedoch verläuft die Erkrankung
nicht immer frühzeitig letal, was eine 5-Jahres-Überlebenszeit von 33% belegt (108).
Die meisten Patienten sterben nicht aufgrund der Tumorlast, sondern an infektiösen
Komplikationen bei fortgeschrittener Immundefizienz (39).
Der Internationale Prognose Index (IPI) und der Prognose Index für PTCL (PIT) haben
sich für das AITL als nur eingeschränkt aussagekräftig erwiesen (106).
Anders jedoch der 2013 von Federico et al. vorgeschlagene, vereinfachte „prognosis in
AITL“ - Index (PIAI), der 5 Faktoren berücksichtigt: Alter > 60 Jahre, Performance
status (als Messwert der Lebensqualität) > 2, Befall extranodaler Organe > 1,
Vorhandensein von B-Symptomen und Thrombozytenwerte < 150 000/µl. Im Rahmen
des „International Peripheral T-Cell Lymphoma Projects“ konnten so anhand von 243
Fällen eine low-risk-Gruppe (0-1 Faktor erfüllt) mit einer 5-Jahresüberlebenszeit von
44% und eine high-risk-Gruppe (2 – 5 Faktoren erfüllt) mit einer 5-Jahres-
überlebensrate von 24% unterschieden werden (43).
In einer multivarianten Analyse von 157 AITL-Fällen, behandelt nach den Protokollen
der Groupe d’Etude des Lymphomes de l’Adulte (GELA), konnten nur männliches

6
Geschlecht, mediastinale Lymphadenopathie und Anämie als nachteilig in Bezug auf
das Gesamtüberleben nachgewiesen werden (108).
Xu et al., die eine große Populations-basierte Studie unter Verwendung der
„Surveillence, Epidemiology und End-Results (SEER)“-Datenbank durchführten,
konnten anhand von 1207 Patienten unter anderem die 5-Jahresüberlebensrate von
33% bestätigen. In multivarianter Analyse identifizierten sie Alter > 70 Jahre,
männliches Geschlecht und forgeschrittenes Krankheitsstadium als negative prädiktive
Faktoren für Gesamtüberleben und Erkrankungsspezifisches Überleben. Außerdem
konnte gezeigt werden, dass in den letzten 20 Jahren keine Verbesserung des
Überlebens von Patienten mit AITL erzielt werden konnte (165).
Die Primärlokalisation des AITL ist der Lymphknoten, zusätzlich können auch Milz,
Leber, Haut und Knochenmark betroffen sein (39).
Histologische Charakteristika des AITL
Charakteristischerweise ist die Lymphknotenarchitektur ganz oder teilweise durch ein
diffuses polymorphes Infiltrat zerstört, das sich in erster Linie interfollikulär ausbreitet
(39). Gewöhnlich sind die peripheren kortikalen Sinus erhalten, aber eine Infiltration in
das perinodale Gewebe kommt vor (39). Die neoplastischen Zellen sind mittelgroß und
weisen reichlich helles Zytoplasma, distinkte Zellmembranen und zytologische Atypien
auf (149). Oft formen sie kleine Gruppen um Follikel und hochendotheliale Venolen.
Untermischt sind reichlich kleine Lymphozyten, Histiozyten oder epitheloide Zellen,
Immunoblasten, Eosinophile und Plasmazellen (35).
Zwei weitere typische Befunde sind eine prominente Neovaskularisation, hervorgerufen
durch eine deutliche Proliferation hochendothelialer Venolen sowie perivaskulär
proliferierte follikuläre dendritische Zellen (FDCs) (35, 106).
Desweiteren kommen in über 95% der Patienten vermehrt Epstein-Barr-Virus (EBV-)
positive aber auch -negative B-Zellen vor, die teilweise ein Immunoblasten-ähnliches
Aussehen zeigen, teilweise aber auch Reed-Sternberg-Zellen imitieren (39, 111). Der
B-Zell Reichtum, der nicht das klinische Outcome zu beeinflussen scheint, korreliert mit
dem Nachweis der B-Zell-Klonalität (49, 51). In bis zu einem Drittel der Fälle kommen
klonale und oligoklonale B-Zell-Expansionen vor (35). Ein Teil der Patienten entwickelt
hieraus ein zumeist EBV-positives diffuses großzelliges B-Zell Lymphom (51).
In der Praxis kann durch den B-Zell-Reichtum die Abgrenzung des AITL zum
klassischen Hodgkin-Lymphom oder T-Zell- und Histiozyten-reichen großzelligen B-
Zell-Lymphom schwer fallen.

7
Es wurden drei sich überlagernde histomorphologische Muster des angio-
immunoblastischen T-Zell Lymphoms beschrieben (8, 39):
Muster 1 (20% der Patienten) mit weitestgehend erhaltener Lymphknotenarchitektur:
Man sieht hyperplastische, verwaschene B-Zell-Follikel und einen expandierten
Paracortex mit einem polymorphen Infiltrat und einem prominentem Gefäßnetz.
Muster 2 (30% der Patienten) ist charakterisiert durch einen Verlust der normalen
Architektur: Vereinzelt depletierte B-Zell-Follikel mit konzentrisch angeordneten,
teilweise auch gering expandierten FDC-Netzwerken kommen vor. Der übrige Lymph-
knoten wird von einem polymorphen Infiltrat mit steigender Anzahl lymphoider Blasten
eingenommen. Die Gefäßproliferation ist prominent.
Muster 3 (50% der Patienten) mit vollständig zerstörter Lymphknotenarchitektur:
B-Zell-Follikel sind verschwunden, FDC-Netzwerke und Gefäße sind unregelmäßig
proliferiert und ein polymorphes Infiltrat aus verschiedenen Zelltypen herrscht vor.
Diese Muster sind assoziiert mit einer ansteigenden Anzahl neoplastischer Zellen und
man geht davon aus, dass sie fortschreitende Stadien der Erkrankung darstellen (35).
Tan et al. konnten hierzu in einer Meta-Analyse aus eigenen Fällen und von Attygalle
bereits beschriebenen (insgesamt 56) Fällen zeigen, dass Muster 1 tatsächlich einen
Überlebensvorteil im Gegensatz zu Muster 2 und 3 zeigt (152).
Das neoplastische T-Zell-Infiltrat wird oft durch den reaktiven Hintergrund maskiert,
und macht mit 5 bis 30% häufig nur einen Bruchteil des gesamten Zellbildes aus (35,
106, 163, 179). Dominante klonale T-Zellen, von denen man annimmt, dass sie die
neoplastische Population darstellen, sind in 70% der angioimmunoblastischen T-Zell
Lymphome mit konventionellen molekularbiologischen Techniken detektierbar (23).
Jedoch können teilweise simultan auch expandierende B-Zell Klone nachgewiesen
werden (23).
Molekulare Pathogenese
Als normales Pendant der neoplastischen T-Zellen in angioimmunoblastischen T-Zell-
Lymphomen wurden anhand von Gen-Expressionsanalysen follikuläre T-Helfer-(Tfh-)
Zellen postuliert (37, 123). Diese sind eine seltene Untergruppe der alpha-beta T-
Helfer-Zellen. Sie sind hauptsächlich in Keimzentren lokalisiert und interagieren mit
den Keimzentrums-B-Zellen, um das Überleben der B-Zellen, den Immunglobulin
Klassen-Wechsel und die somatische Hypermutation zu fördern und um schließlich
Plasmazellen und Memory-B-Zellen zu weichen (51). Die Tfh-Differenzierung ist ab-
hängig von dem Transkriptionsfaktor bcl6.

8
Die Funktion der Tfh-Zellen korreliert mit ihrem spezifischen Sekretionsprofil
(Expression von IL-2 und CXCL13 = Chemokin, entscheidend für B-Zell-Rekrutierung
und B-Zell-Aktivierung), dem spezifischen Immunphänotyp (Expression von CXCR5=
Rezeptor für CXCL13, notwendig für die Lokalisation der Tfh-Zellen im Keimzentrum)
und Ko-stimulierenden Molekülen wie ICOS, PD1, CD28 und CD40L, die die Tfh-B-
Zell-Interaktion fördern (51).
AITL ist der Prototyp der sich von Tfh-Zellen-ableitenden Neoplasien (37, 66).
Die Herkunft des AITL von Tfh-Zellen erklärt viele der pathobiologischen Eigenschaften
des Lymphoms, z.B. die Expansion der B-Zellen, die Proliferation der FDCs und die
Autoimmun-Effekte (51). Hier gilt CXCL13 als möglicher Mediator mit Schlüsselfunktion
(49). Zusätzlich können normale Tfh-Zellen T-Zell Antworten supprimieren und man
nimmt an, dass die neoplastische T-Zell Population so zu der defekten T-Zell Antwort
in AITL beiträgt (51).
Dass eines der häufigsten T-Zell Lymphome sich von einer der seltensten T-Zell
Subgruppen ableitet, ist bemerkenswert (37, 124) . Es lässt ein erhöhtes Risiko der
Tfh-Zellen für eine maligne Transformation annehmen (124). Anderseits wird die enge
Interaktion mit B-Lymphozyten und FDCs diskutiert und es ist vorstellbar, dass unter
physiologischen Bedingungen der neoplastische Klon von diesen Zellgruppen und dem
entstehenden Mikromilieu Unterstützung erhält (124).
Die neoplastische Transformation der Tfh-Zellen ist bisher nicht verstanden.
In zytogenetischen Analysen wurden verschiedene klonale Aberrationen gefunden –
darunter in bis zu 90% der Fälle Trisomien der Chromosomen 3, 5 und 21, Hinzu-
gewinn von Chromosom X und Verlust von 6q (149). Neuere Studien zeigten in etwa
50%, 30-40% bzw. 10% der jeweils untersuchten AITL-Fälle wiederkehrende
Punktmutationen in den Genen TET2, IDH2 und DNMT3A, die für Enzyme – verant-
wortlich für DNA-Methylierung und epigenetische Transkriptionskontrolle - kodieren.
TET2-Mutationen sind mit einem fortgeschrittenerem Krankheitsstadium, einem hohen
Internationalen Prognostischen Index (IPI) – Wert und einem kürzeren progressions-
freien Überleben assoziiert (21, 30, 85).
Odejide et al. untersuchten mittels next-generation sequencing über 200 Gene, in
denen Mutationen für hematologische Neoplasien bekannt sind. Hierbei zeigte sich,
dass die Gesamtheit der häufigen Genmutationen in AITL eher der myeloider
Neoplasien als der anderer Lymphome ähnelt (113).
Gen-Expressionsanalysen, deren Ergebnisse von Iqbal et al. 2010 veröffentlicht
wurden, haben gezeigt, dass die Gen-Signatur von Tfh-Zellen in AITL signifikant
angereichert ist (66). So konnte neben der Überexpression von Schlüsselgenen der

9
neoplastischen T-Zellen, CTLA4 und ICOS, auch die normaler Tfh-Zellen, CXCL13 und
CXCR5, in AITL nachgewiesen werden (66). Allerdings fanden sich nach molekularen
Klassifikationskriterien zudem auch drei weitere prominente Signaturen in AITL: Die
von B-Zellen und follikulären dendritischen Zellen, die in der Umgebung der
neoplastischen T-Zellen angereichert sind, sowie von Zytokinen, die teilweise von der
stromalen Komponente des Mikromilieus im AITL abhängig sind (66). So ließ sich zum
Beispiel eine höhere Expression von Genen nachweisen, die mit germinalen B-Zellen
assoziiert sind, von denen man – wie bei CD10 und bcl6 - jedoch auch weiß, dass sie
von den neoplastischen T-Zellen exprimiert werden können. Mittels Gene Set
Enrichment Analysis (GSEA) konnte zudem eine signifikante Anreicherung von
Molekülen gezeigt werden, die bestimmten Signalwegen zugeordnet werden können:
Zum Beispiel dem NFkappaB-Signalweg: AITL-Fälle zeigten im Vergleich zu nicht
spezifizierten PTCL eine Aktivierung dieser Signalkaskade; zweitens immun-
suppressiven Wirkungspfaden: insbesondere Gensignaturen, assoziiert mit dem TGF-
beta-Signalweg zeigten sich angereichert; drittens der IL-6- Signalwirkung: Interleukin
6 erfüllt diverse Funktionen und ist unter anderem in die Proliferation von Tfh-Zellen
sowie von EBV-infizierten B-Zellen involviert.
In weiterführenden Studien stellten Iqbal et al. 2014 für AITL insbesondere die Gene
der Signatur heraus, die Assoziationen mit Angiogenese und vaskulärer Endothel-
funktion sowie Zellmigration zeigten (67).
Insbesondere VEGF scheint hier involviert zu sein (124, 126). So konnte gezeigt
werden, dass VEGF nicht nur von bystander-Zellen, sondern auch von den
neoplastischen Zellen in AITL produziert wird (124). Zusätzlich konnten Piccaluga et al.
demonstrieren, dass AITL-Zellen den Rezeptor VEGFR1/KDR exprimieren. Dies führte
zu der Hypothese, dass die VEGF-Produktion nicht nur die Angiogenese sondern auch
das Tumorwachstum über einen autokrinen/parakrinen Loop stimulieren kann und dass
AITL möglicherweise sensitiv für die anti-angiogenetischen Medikamente Thalidomid
und Bevicizumab sein könnten (124, 126, 131).
Die Dichte der Mikrogefäße in Lymphknoten von Patienten mit AITL stellt außerdem
einen negativen prädiktiven Faktor im Hinblick auf progressionsfreies und Gesamt-
Überleben dar (176).
Zusammengefasst spiegeln die molekularen Klassifikationskriterien und die Signalweg-
Signaturen von vielen peripheren T-Zell Lymphomen das Mikromilieu der jeweiligen
Lymphom-Subgruppe wider (50, 67). Dies wurde herausragend an AITL demonstriert
(67). Gleichzeitig konnte auch gezeigt werden, dass insbesondere das Zytokin-Milieu in

10
AITL mit seinem enormen Repertoire aus Chemokinen und Rezeptoren ein komplexes
immunologisches Netzwerk darstellt (67). Außerdem weisen die Studien von Iqbal et al.
stark darauf hin, dass das Mikromilieu signifikant mit der Prognose in AITL verknüpft ist
(67).
Im Vergleich zu B-Zell Lymphomen ist das immunphänotypische Profil von T-Zell
Lymphomen im Hinblick auf die Subklassifizierung sehr viel weniger spezifisch und
hilfreich (148). Nichtsdestotrotz können in Konkordanz mit dem Genexpressionsprofil
des AITL einige Marker in der immunhistochemischen Diagnostik zur Identifizierung
der neoplastischen T-Zell-Population und weiterer Charakteristika im Lymphknoten-
gewebe genutzt werden.
So exprimieren die neoplastischen T-Zellen in AITL typischerweise die Pan-T-Zell-
Antigene CD3, CD4 und CD5 (149). Ein abbaranter Verlust oder eine verminderte
Expression von einzelnen T-Zell-Antigenen können vorkommen (36, 49).
Wie beschrieben, wurde das AITL als eine Neoplasie der follikulären T-Helfer Zellen
erkannt. Dementsprechend können die neoplastischen T-Zellen im AITL immun-
phänotypische Eigenschaften normaler Tfh-Zellen aufweisen: In über 90% der Fälle
exprimieren sie CXCL-13, ein Chemokin, das den B-Zellen den Eintritt in die
Lymphfollikel ermöglicht, und programmed cell- death (PD-1), ein Mitglied der CD28
Rezeptor Familie, das die zelluläre Immunanwort reguliert (77, 171). Desweiteren
konnten Attygalle et al. und Ree et al. zeigen, dass zwei Marker von Keimzentrums-B-
Zellen, CD10 und bcl6, ebenfalls von der Mehrheit der neoplastischen T-Zellen
exprimiert werden (8, 133). Das induzierbare T-Zell ko-stimulierende Molekül (ICOS)
und das Adapter-Molekül SLAM-associated protein (SAP) wurden ebenfalls als
nützliche Marker zur Identifizierung der neoplastischen T-Zellen in T-Zell Lymphomen,
die sich von Tfh-Zellen ableiten, erfolgreich herangezogen (49, 102).
Besonders hilfreich im Hinblick auf die Diagnosestellung AITL ist zudem die immun-
histochemische Darstellung der follikulären dendritischen Zell- (FDC) Netzwerke mit
den Primärantikörpern gegen CD21 und CD23 um die charakteristische Expansion
sichtbar zu machen (22).
Ein standardisiertes Therapie-Regime wurde bislang für Patienten mit AITL nicht
etabliert (165). Momentan wird eine Anthrazyklin-basierte Chemotherapie als
Erstlinien-Therapie betrachtet (106). Durch die intensive Chemotherapie werden in 50-
70% der Patienten zunächst komplette Remissionen erzielt, jedoch hält der Effekt
meist nur kurzzeitig an und die Mehrzahl der Patienten erleidet im Verlauf ein Rezidiv
(106, 131).

11
In Bezug auf das Gesamtüberleben konnte weder ein Unterschied zwischen
verschiedenen Anthrazyklin-basierten Regimes (ACVBP, CHOP, mBACOD) noch
zwischen Anthrazyklin-basierter und nicht-anthrazyklin-basierter Chemotherapie belegt
werden (108, 159).
Insgesamt bleibt die Entwicklung molekular basierter therapeutischer Strategien für T-
Zell-Lymphome insbesondere im Vergleich zu denen für B-Zell-Lymphome insuffizient
(62), dennoch wurden einige Medikamente experimentell mit Erfolg getestet:
Thalidomid zum Beispiel ist ein immunmodulatives Agens, das den Tumor Nekrose
Faktor (TNF), den vaskulären endothelialen Wachstumsfaktor (VEGF), IL-6 und
NFkappaB inhibiert. Es wurde erfolgreich eingesetzt und konnte klinisch und radio-
logisch messbare Remissionen der Erkrankung erzielen (106). Basierend auf diesen
Erkenntnissen wurde eine zurzeit (Mai 2014) laufende Studie zu einer Kombinations-
therapie aus CHOP und Lenalidomid initiiert (NCT01553786).
Nachdem eine Phase II-Studie gezeigt hat, dass 16% der refraktär an AITL erkrankten
Patienten auf eine Therapie mit Romedepsin, einem Histon-Deazetylase-Inhibitor,
ansprechen (125), wird zurzeit in einer Phase III-Studie die Langzeiteffizienz dieser
Therapiefom untersucht (NCT0196002). Vielversprechende Ergebisse konnten durch
eine Phase-II-Studie unter Einschluss von 25 AITL-Patienten mit gutem Performance-
Status mit der Kombination aus Rituximab und CHOP erzielt werden. Rituximab ist ein
monoklonaler CD20-Antikörper, der sich gegen die B-Zellen im Microenvironment der
Lymphomzellen richtet. Die Remissionsrate (ORR) lag insgesamt bei 80%, jedoch
zeigte sich kein Einfluss auf das Gesamtüberleben der Patienten (38).
Weitere experimentelle Studien unter anderem mit Alemtuzumab (CD52-Antikörper),
Pralatrexat (Antifolat) Bevacizumab (VEGF-Antikörper) und Bortezomib (Proteasom-
inhibitor) wurden durchgeführt und lassen teilweise ein gutes Ansprechen erkennen
(48, 95). Jedoch ist die Biologie des Angioimmunoblastischen T-Zell Lymphoms nicht
ausreichend gut verstanden und es fehlen größere Studien sowie Langzeit-
beobachtungen um eine Signifikanz dieser Ergebnisse abzusehen.

12
1.1.2. microRNA
1.1.2.1. microRNA-Physiologie
MicroRNAs sind eine Klasse kleiner, endogen exprimierter, nicht proteincodierender
RNAs (173), die die Genexpression, insbesondere durch posttranskriptionelles Gene-
Silencing, regulieren können.
MicroRNAs und ihre assoziierten Proteine gehören zu den häufigeren Ribonukleo-
proteinkomplexen in einer Zelle (9) und ihr Einfluss ist allgegenwärtig (88).
Nichtsdestotrotz wurde man erst in den neunziger Jahren durch Erforschung der
Entwicklungsbiologie des Wurms Caenorhabditis elegans auf sie aufmerksam (80, 82,
83). MicroRNAs sind, anders als siRNAs, durch genomische DNA codiert. Bisher geht
man davon aus, dass etwa 1 bis 4 % des humanen Genoms für microRNA codieren (9,
10, 88) und dass etwa 60% der mRNA in Säugetieren durch sie reguliert werden (88).
Inzwischen sind in der Online-Datenbank „miRbase“ (http://www.mirbase.org/, v21,
Juni 2014) 35828 reife microRNA, identifiziert in 223 Spezies, zusammengetragen
worden (76), von denen 1872 im humanen Genom lokalisiert sind. Es wurden bislang
mehr als 4500 reife microRNAs im menschlichen Organismus gefunden (62).
Reife microRNAs sind einzelsträngig und bestehen aus etwa 22 Nukleotiden (~22 nt)
(9). Sie sind dadurch charakterisiert, dass ihre Vorstufen kurze Haarnadel-Strukturen
ausbilden und spezifische Reifungsprozesse durchlaufen, die definierten kanonischen
und nicht-kanonischen biogenetischen Wegen folgen (24, 169). Die Biogenese der
tierischen microRNAs verläuft mehrschrittig auf dem sogenannten kanonischen Weg:
Ein microRNA-Gen wird meist durch die Polymerase II transkribiert (10). Es resultiert
ein bis mehrere hundert Nukleotide langes, schlaufenförmiges Primärtranskript
(Primary MicroRNA, pri-miRNA), das noch im Zellkern durch einen Komplex aus einem
RNAse III-ähnlichen Enzym (Drosha) und DGCR8 (DiGeorge Critical Region Gene 8)
in eine 70 bis 100 Nukleotide lange Precursor-microRNA (pre-miRNA) gespalten wird.
Diese weist eine Haarnadel (Hairpin)-Struktur auf und gelangt via Exportin 5 in das
Zytoplasma.
Nach dem Modell von Maniataki et al. nimmt im menschlichen Organismus ein
Enzymkomplex, der unter anderem aus einer weiteren RNAse III, genannt Dicer, und
einem Argonaut 2 (Ago2) –Protein besteht, die pre-miRNA auf (100) . Dicer spaltet die
pre-miRNA in eine 19 bis 22 Nukleotide lange doppelsträngige microRNA (9, 24).
Offenbar simultan kommt es ATP-unabhängig zur Entwindung und Singularisierung
des microRNA-Duplexes (100). Es entstehen zwei Einzelstränge, ein 3´- Strang und

13
ein 5´-Strang, von denen meist nur einer als reife microRNA biologisch aktiv bleibt (61)
und auf das Ago2-Protein geladen wird (100). Nach Erkenntnissen von Maniataki et al.
dissoziiert Dicer von dem Enzymkomplex und das entstehende microRibonukleoprotein
(miRNP) führt zur Formung eines RNA-induced silencing-Komplexes (RISC) aus reifer
microRNA und Proteinen der Argonaut-Familie als Kernstrukturen (100).
Über komplementäre Basenpaarung kann die im RISC inkorporierte microRNA an die
3’untranslatierte Region (3´UTR) einer mRNA binden und in Abhängigkeit vom Grad
ihrer Komplementarität entweder eine Repression der Translation oder aber die
Zerschneidung dieser Ziel-mRNA bewirken (9). In Tieren reicht die Komplementarität
zu einer kurzen Sequenz aus etwa 7 Nukleotiden der microRNA (sogenannte seed-
region, meist Nukleotide 2-7) aus, um die produktive Translation der daran bindenden
mRNA erfolgreich zu unterdrücken (10). Das Phänomen der Zerschneidung der Ziel-
mRNA kommt im tierischen Organismus selten vor und setzt die perfekte (vollständige)
Komplementarität von miRNA und mRNA voraus (88). Rechnerisch und experimentell
kann eine einzige microRNA potentiell tausende mRNA-Moleküle als Zielstrukturen
erkennen (116). Ebenso kann eine Ziel-mRNA durch mehrere verschiedene
microRNAs reguliert werden (89).
Experimente unter anderem von Leung et al. haben gezeigt, dass die Proteine der
Argonaut-Familie sich zumeist in sogenannten „processing-bodies“, kurz P-bodies,
innerhalb des Zytoplasmas einer eukaryoten Zelle anreichern. In diesen um-
schriebenen Protein-Aggregaten wird die unterdrückte mRNA fest- und von der
Translationsmaschinerie ferngehalten sowie der RNA-Abbau vermittelt (88).

14
Abbildung 1 Biosynthese und Wirkungsweise von micro RNAs
kanonischer Weg, modifiziert nach (65)
Kürzlich wurden eine Reihe weiterer alternativer Mechanismen der microRNA-
Synthese, sogenannte nicht-kanonische Wege, beschrieben, die sowohl DROSHA- als
auch DICER-unabhängig sein können (u.a. zusammengefasst in (24) und (169)).
Grundsätzlich darf nicht vergessen werden, dass microRNAs nur Transkripte be-
einflussen können, die in einem bestimmten entwicklungsbiologischen oder patho-
logischen Kontext induziert wurden; sie können nicht a priori die Genexpression
herbeiführen (116). Dies legt nahe, dass die Zielstrukturen der microRNA
höchstwahrscheinlich gewebsspezifisch sind und dass Ergebnisse von microRNA-
Expressionsstudien unter Einbeziehung des Ursprungsgewebes interpretiert werden
müssen (116).
DNA
Pol II
Pri -miR
Pre-miR
Pre-miR miR:miR* - Duplex Dicer
Helicase
RISC
miR
miR
mRNA
DGCR8 Drosha
miR*

15
Abkürzung (Beispiel) Erklärung
Pri-miR Primary microRNA
Pre-miR Precursor microRNA
miR-1 Mature, einsträngige Form
miR-1* Komplementärer Strang, der degradiert
wird
miR-17-5p 5’-Arm Beide Stränge stellen die mature Form
dar, keiner wird bevorzugt degradiert miR-17-3p 3’-Arm
miR-20a (13q31.3) Paraloge mit 1-2 Basen
Sequenzunterschieden miR-20b (Xq26.2)
Hsa-miR-1 Homo
sapiens
microRNA mit gleicher Nummer können
unterschiedliche Sequenzen in
verschiedenen Spezies haben Mmu-MiR-1 Mus
musculus
Tabelle 2 Nomenklatur der microRNAs
Modifiziert nach (61)
1.1.2.2. microRNA-Dysregulation
Das Interesse dysregulierte microRNA-Profile in Erkrankungen, insbesondere Krebs,
zu identifizieren, ist groß (116). Eine abnormale microRNA-Expression wurde für viele
Tumorentitäten, einschließlich maligner Lymphome, beschrieben (einen Überblick gibt
hierzu (173), bezüglich Lymphome (41, 62, 151)). Für T-Zell-Lymphome/Leukämien
haben Ikeda und Tagawa die neuesten Erkenntnisse bezüglich microRNAs kürzlich
zusammengefasst und veröffentlicht (vgl. hierzu (62)). Angioimmunoblastische T-Zell-
Lymphome wurden darin nicht berücksichtigt. Auch gibt es nur wenige Tumore, für die
ein direkter kausaler Zusammenhang zwischen microRNA-Dysregulation und
Tumorentwicklung festgestellt werden konnte (116).
Diejenigen microRNAs, deren Expression in Tumoren erhöht ist, werden als mögliche
Onkogene betrachtet und „oncomirs“ genannt (173). Ein prominentes Beispiel ist miR-
155, die in verschiedenen B-Zell-Lymphomen überexprimiert wird (139). MicroRNAs,
deren Expression in Krebszellen vermindert ist, gelten als mögliche Tumorsuppressor-
microRNAs. So zum Beispiel die miR-34-Familie, die ein Zielgen von p53 darstellt und
in zahlreichen Geweben bei Überexpression direkt oder indirekt verschiedene
Onkogene hemmen und den Zelltod eines Tumors begünstigen sowie die

16
Metastasierung verhindern kann (4). Konsequenterweise ist ein miR-34-Analogon die
erste microRNA, die die erste Phase einer klinischen Studie erreicht hat (4).
Außerdem können microRNA-Profile bei der Subklassifizierung von Lymphomen
hilfreich sein. Im Hinblick auf B-Zell-Lymphome konnten Roehle et al. vier microRNAs
identifizieren, die mit einer Genauigkeit von 98% zwischen diffusem großzelligem B-
Zell-Lymphom und follikulärem Lymphom sowie reaktivem Lymphknoten unterscheiden
können (134). Bezüglich T-Zell-Lymphomen konnten zum Beispiel Liu et al. eine
Signatur aus 11 microRNAs ermitteln, die ALK-negative großzellige anaplastische
Lymphome (ALCL) von anderen nodalen PTCL differenziert.

17
1.2. Fragestellung
Ziel der vorliegenden explorativ angelegten Arbeit ist es, das äußerst seltene
angioimmunoblastische T-Zell-Lymphom anhand von 30 Fällen auf morphologischer,
immunhistochemischer und molekularer Ebene näher zu charakterisieren. Der
Schwerpunkt der Arbeit liegt hierbei auf der erstmaligen Erstellung eines microRNA-
Profils der Erkrankung und dessen Vergleich mit dem des nicht-neoplastischen
Kontraparts, Lymphadenitis mit T-Zonen-Hyperplasie. Aufgrund ihrer histologischen
und immunphänotypischen Überschneidungen (77), erscheinen die derartig reaktiv
veränderten Lymphknoten im Hinblick auf Gewebsspezifität, Zellzusammensetzung
und nicht zuletzt aufgrund ihrer Verfügbarkeit als Vergleichskollektiv sehr gut geeignet.
Desweiteren soll die sinnvolle Ergänzung des immunhistochemischen Marker-Panels
für die Routinediagnostik angioimmunoblastischer T-Zell-Lymphome durch Antikörper
gegen Stathmin und LMO2, ein bekanntes Oncoprotein und einen Transkriptionsfaktor,
geprüft werden.
Außerdem geht es um die Erfassung und isolierte Betrachtung bekannter morpho-
logischer Charakteristika als Ausdruck des Mikromilieus in angioimmunoblastischen T-
Zell-Lymphomen, deren Zuordnung zu bereits etablierten histomorphologischen
Mustern und ihren Einfluss auf molekulare Befunde.
Weiter entfernte Ziele sind natürlich, zum Verständnis der Pathophysiologie des
Lymphoms beizutragen und mögliche Biomarker zu identifizieren, um so die
Entwicklung molekular basierter Therapie-Strategien zu fördern und die seit 20 Jahren
stagnierend schlechte Prognose des AITL zu verbessern.

18
2. Material und Methoden
2.1. Material
Die verwendeten Gerätetypen, Verbrauchsmaterialien, Reagenzien und Chemikalien
sowie eine Auflistung der Kits, Antikörper, Primer, Software und in silico Resourcen
finden sich in übersichtlicher, alphabetisch geordneter, tabellarischer Form im Anhang
(Tabellen 11-17).
2.2. Methoden
2.2.1. Studiengruppe Die Studie basiert auf dem Vergleich von zwei Patientenkollektiven. Dreißig Fälle
angioimmunoblastischer T-Zell Lymphome (malignes Lymphom) wurden einer Kontroll-
gruppe von zehn Lymphadenitiden mit T-Zonen-Hyperplasie (benigne entzündliche
Veränderungen), dem vermuteten nicht-neoplastischen Kontrapart, gegenübergestellt.
Pro Patient wurde jeweils eine Einsendung berücksichtigt und rekrutiert. Alle ein-
gesetzten Proben entstammen dem Untersuchungsgut des Referenzzentrums für
Lymphomdiagnostik am Institut für Pathologie des Universitätsklinikums Schleswig-
Holstein, Direktor Prof. Dr. med. A.C. Feller, aus den Jahren 2010 bis 2012.
Mithilfe der NEXUS Datenbank wurden unter Eingabe der Suchbegriffe „9705/3“ (ICD-
O Code für das AITL) bzw. „Lymphadenitis [AND] T-Zonen-Dysplasie“ entsprechende
Fälle ermittelt. Die initialen Diagnosen wurden von langjährig erfahrenen Fachärzten
für Pathologie des Instituts gestellt. Das jeweilige Schnitt- und Blockmaterial wurde aus
den Archiven des Instituts extrahiert und anhand der routinemäßigen diagnostischen
Schnittpräparate reevaluiert. Die Diagnosen wurden im Kontext der aktuellen WHO
Klassifikation (2008) von drei Untersuchern (Prof. Dr. med. A.C. Feller, Prof. Dr. med.
Ch. Thorns, K.Reddemann) im Konsensusverfahren validiert. Es folgte die Auswahl
von dreißig Fällen angioimmunoblastischer T-Zell-Lymphome und zehn Lymph-
adenitiden, die hinsichtlich der histomorphologischen Charakteristika der jeweiligen
Erkrankung die höchste Repräsentanz zeigten.
2.2.2. Patientenproben
Alle Proben waren humane, formalinfixierte, paraffineingebettete Lymphknoten, die
verschiedenen Lokalisationen (zervikal, axillär, inguinal, nuchal, thorakal, abdominell)
entstammten. Beiden Gruppen gemeinsam war die Voraussetzung eines genügend

19
großen und ausreichend beurteilbaren Lymphknotenanteils, frei von Manifestationen
anderer morphologisch fassbarer Erkrankungen. Alle Proben samt zugehöriger Daten
wurden nach der Rekrutierung für die Studie einer umgehenden Anonymisierung im
Sinne des genehmigten Ethikantrages der Ethikkommission der Universität zu Lübeck
unterzogen (Aktenzeichen: 13-179A).
2.2.3. Histopathologische Methoden
2.2.3.1. Herstellung des Tissue Microarrays Ein Tissue Microarray (TMA) ist ein Paraffinblock, in den mehrere ausgestanzte
Gewebszylinder unterschiedlicher Herkunft eingebracht wurden. So war es möglich
diese verschiedenen Proben gleichzeitig unter gleichen Bedingungen mittels immun-
histochemischer Verfahren oder in-situ-Hybridisierung zu analysieren.
Zunächst wurden die hinsichtlich ihrer Repräsentanz, Dicke und Gewebs-
beschaffenheit am besten geeigneten Gewebe-Paraffinblöcke der 40 oben
beschriebenen Lymphknotenproben ausgewählt. Um einen Überblick über die aktuelle
Zellzusammensetzung in den ausgewählten Blöcken zu erhalten, wurden davon
Hämatoxylin-Eosin-gefärbte Schnittpräparate hergestellt, von zwei Untersuchern
begutachtet und ein repräsentatives Gewebsareal für die Positionierung der
Stanzzylinderentnahme ausgewählt (Prof. Dr. med. Ch. Thorns, K.Reddemann).
Die Herstellung des TMAs erfolgte mit dem Tissue-Tek® Tissue Microarray System
(Sakura Finetek, USA) nach Angaben des Herstellers.
Aufgrund der Heterogenität des untersuchten Gewebes wurde der Durchmesser der
Gewebsstanzen mit jeweils 2 mm relativ groß gewählt. Zur Orientierung auf dem
späteren Schnittpräparat wurde das TMA in Subarrays unterteilt und eine
asymmetrische Anordnung der Proben gewählt:

20
Abbildung 2 Anordnung der Gewebsstanzen im tissue m icroarray
(1-30 Lymphom-Fälle, K1-K10 Fälle der Referenzgruppe)
2.2.3.2. Herstellung von Paraffin-Schnittpräparaten
Zur Anfertigung von Paraffinschnitten der Gewebsblöckchen wurde ein Rotations-
mikrotom der Firma Leica verwendet. Die Schnittdicke betrug 1,5 Mikrometer.
2.2.3.3. Histochemische Färbungen von formalinfixie rten, paraffineingebetteten Gewebsschnitten
Histochemische Färbungen zählen zu den klassischen Techniken der Histologie und
sind die Basis der morphologischen Diagnostik. Verschiedene Gewebs- und
Zellkomponenten werden durch sie sichtbar und unterscheidbar. Sie basieren auf
physikalischen Prozessen, wie der Diffusion oder Elektroadsorption, und chemischen
Reaktionen, wie der Ionenbindung. Aufgrund der Interaktion zwischen
Gewebebestandteil und eingesetztem Reagenz entsteht ein Farbstoff.
Das unterschiedliche Färbeverhalten verschiedener Gewebskomponenten ergänzt so
das histologische Erscheinungsbild.
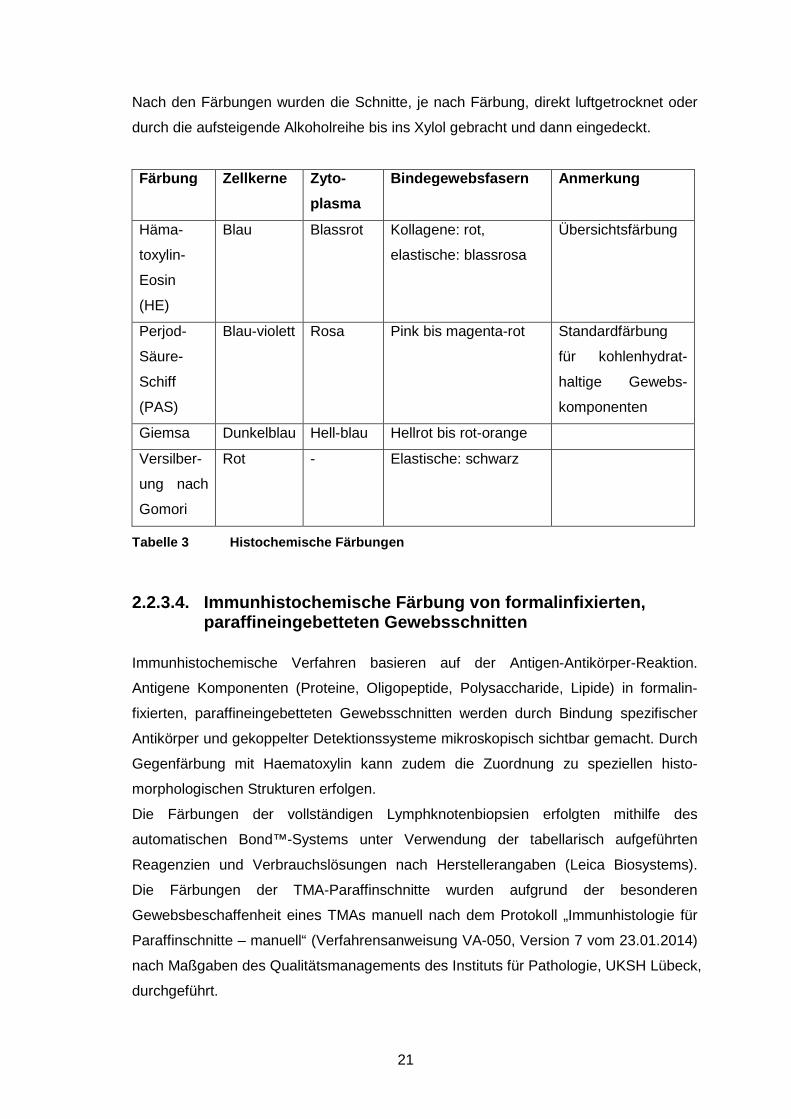
Folgende histochemische Färbungen wurden in der vorliegenden Arbeit berücksichtigt:
Die Hämatoxylin-Eosin-Färbung, die Perjod-Säure-Schiff-Reaktion, die Giemsa-
Färbung und die Versilberung nach Gomori.
Die Färbungen wurden entweder manuell oder automatisch (ST 5020, Leica) nach
akkreditiertem Standard des Qualitätsmanagements des Instituts für Pathologie, UKSH
Lübeck, durchgeführt.
Vor den Färbungen wurden die Schnitte in Xylol entparaffiniert und durch die
absteigende Alkoholreihe wieder in ein wässriges Milieu überführt.
21
Nach den Färbungen wurden die Schnitte, je nach Färbung, direkt luftgetrocknet oder
durch die aufsteigende Alkoholreihe bis ins Xylol gebracht und dann eingedeckt.
Färbung Zellkerne Zyto -
plasma
Bindegewebsfasern Anmerkung
Häma-
toxylin-
Eosin
(HE)
Blau Blassrot Kollagene: rot,
elastische: blassrosa
Übersichtsfärbung
Perjod-
Säure-
Schiff
(PAS)
Blau-violett Rosa Pink bis magenta-rot Standardfärbung
für kohlenhydrat-
haltige Gewebs-
komponenten
Giemsa Dunkelblau Hell-blau Hellrot bis rot-orange
Versilber-
ung nach
Gomori
Rot - Elastische: schwarz
Tabelle 3 Histochemische Färbungen
2.2.3.4. Immunhistochemische Färbung von formalinfi xierten, paraffineingebetteten Gewebsschnitten
Immunhistochemische Verfahren basieren auf der Antigen-Antikörper-Reaktion.
Antigene Komponenten (Proteine, Oligopeptide, Polysaccharide, Lipide) in formalin-
fixierten, paraffineingebetteten Gewebsschnitten werden durch Bindung spezifischer
Antikörper und gekoppelter Detektionssysteme mikroskopisch sichtbar gemacht. Durch
Gegenfärbung mit Haematoxylin kann zudem die Zuordnung zu speziellen histo-
morphologischen Strukturen erfolgen.
Die Färbungen der vollständigen Lymphknotenbiopsien erfolgten mithilfe des
automatischen Bond™-Systems unter Verwendung der tabellarisch aufgeführten
Reagenzien und Verbrauchslösungen nach Herstellerangaben (Leica Biosystems).
Die Färbungen der TMA-Paraffinschnitte wurden aufgrund der besonderen
Gewebsbeschaffenheit eines TMAs manuell nach dem Protokoll „Immunhistologie für
Paraffinschnitte – manuell“ (Verfahrensanweisung VA-050, Version 7 vom 23.01.2014)
nach Maßgaben des Qualitätsmanagements des Instituts für Pathologie, UKSH Lübeck,
durchgeführt.

22
In einem ersten Schritt wurden mithilfe geeigneter pH-Lösungen (Novocastra Epitope
Retrieval Solutions, Novocastra) unter Hitzeinduktion in einem Dampfgarer, Dampftopf
oder in der Mikrowelle die Epitope in den Gewebsschnitten freigelegt (Epitop-/Antigen-
demaskierung).
Es folgte die indirekte immunhistochemische Färbung.
Hierbei bindet ein unkonjugierter Primär-Antikörper an das Antigen. Danach wird ein
zweiter Antikörper, der gegen das Fc-Fragment des Primär-Antikörpers (jetzt als
Antigen wirkend) gerichtet ist, aufgetragen. Dieser Sekundärantikörper ist mit einem
Enzym gekoppelt, das im Folgenden den sichtbaren Farbumschlag durch Präzipitation
eines zugesetzen Chromogens katalysiert.
Als Primärantikörper kamen hier monoklonale Maus- und Kaninchenantikörper zur
Anwendung, die gegen folgende Zielstrukturen gerichtet waren: CD3, CD10, CD20,
CD21, CD23, LMO2, PD1 und Stathmin (s. Tab. 3).
Antikörper Hersteller Klon Verdünnung Antigen -
demaskierung
CD3 Lab Vision SP7 1:100 Dampfgarer (pH 9,0)
CD10 Novocastra C56C6 1:30 Dampfgarer (pH 9,0)
CD20 Lab Vision L26 1:500 Dampfgarer (pH 9,0)
CD21 Cellmarque 2G9 1:50 EDTA
CD23 Thermo
Scientific
SP23 1:100 EDTA
LMO2 Cellmarque SP51 1:100 EDTA
PD1 Cellmarque MRQ-22 1:100 EDTA
Stathmin DCS SP49 1:100 Citratpuffer (pH 6,0)
Tabelle 4 Antikörper (mit Verdünnungen und Vorbehan dlungen)
Als Sekundärantikörper wurde ein Ziegenantikörper eingesetzt, der mit einer
Meerrettich-Peroxidase (engl. Horseradish-Peroxidase, abgekürzt HRP) konjugiert ist
(Poly-HRP-Anti-mouse/rabbit/rat IgG, ImmunoLogic). Als Chromogen wurde das
ursprünglich fast farblose Diaminobenzidin (Bright-DAB, ImmunoLogic) zugesetzt,
welches im Beisein von Peroxidase einen Farbumschlag in einen braun-schwarzen
Niederschlag am Ort des Zielantigens zeigt.
Die Aktivität unspezifischer Gewebsperoxidasen wurde in einem Zwischenschritt durch
Inkubation mit 6%iger Wasserstoffperoxidlösung unterdrückt.

23
2.2.4. Molekularbiologische Methoden
2.2.4.1. Nachweis von Epstein-Barr-Virus kodierter RNA (EBER) in formalinfixierten , paraffineingebetteten Gewebsschnitten mittels chromogener In-situ-Hybridisierung
Grundlage dieser Methode ist die Bindung einer synthetischen Oligonukleotidsonde
mittels Basenpaarung (Hybridisierung) an einer nachzuweisenden Nukleinsäure im
Gewebe (in-situ).
Der qualitative Nachweis von Epstein-Barr-Virus kodierter RNA (EBER) wurde hier
mittels einer Fluorescein-konjugierten Oligonukleotidsonde (EBER Probe, Bond ISH
Probe, Leica Biosystems) unter Verwendung des Bond™ Enzyme Pretreatment Kits,
des Bond™ Polymer Refine Detection-Systems und eines Gebrauchsfertigen Bond™
Anti-Fluorescein Antibodys (Leica Biosystems) mit Hilfe des automatischen Bond-
Systems nach Herstellerangaben geführt.
2.2.4.2. RNA-Isolierung aus paraffineingebettetem G ewebe Im Rahmen der RNA-Extraktion wurde das RecoverAll™ Total Nucleic Acid Isolation
Kit (Applied Biosystems, P/N 1975M Rev. C) entsprechend den Herstelleranweisungen
verwendet. Es wurden 20 Gewebsschnitte à 5 Mikrometer pro Probe eingesetzt. Die
Proben wurden zuerst mittels Inkubation und Waschung in Xylol und Ethanol
entparaffiniert. Dann erfolgte der Proteaseverdau. Der dritte Schritt war die Isolierung
und Aufreinigung der RNA mittels eines Glasfaser-Filtersystems in einem Mikroliter-
Reagenzgefäß inklusive einer filtergebundenen Nukleasebehandlung.
Die RNA wurde danach bei -80°C gelagert. Die maximale Lagerungsdauer betrug drei
Monate.
2.2.4.3. Quantifizierung von RNA in wässriger Lösung Die Konzentrationsbestimmung der RNA in wässriger Lösung erfolgte über Messung
ihrer Absorption bei einer Wellenlänge von 260 nm unter Verwendung des NanoDrop-
Spektrophotometers.

24
2.2.4.4. Relative Quantifizierung der microRNA-Expr ession mittels RT-qPCR
Die relative Quantifizierung der microRNA-Expression erfolgte unter Anwendung eines
zweistufigen Versuchaufbaus. Zuerst erfolgte die reverse Transkription (RT), dann
wurde die quantitative Real-time PCR (qPCR) angeschlossen.
2.2.4.5. Reverse Transkription (RT) – Polymerase- K etten-Reaktion (RT-PCR)
Im ersten Schritt erfolgte die reverse Transkription, bei der die cDNA unter
Verwendung der jeweiligen Megaplex RT-Primer (microRNA-spezifische stem-loop
Primer), human pool A v2.1 und B v3.0 (Applied Biosystems), dem TaqMan®
MicroRNA Reverse Transcription Kits (Applied Biosystems) sowie der zuvor isolierten
RNA synthetisiert wurde. Es wurden stets 350 ng RNA pro Probe verwendet, sodass
auf eine Präamplifizierung verzichtet werden konnte. Alle Reagenzien wurden gemäß
des Megaplex™ Pools Protocol (Applied Biosystems, P/N 4399721 Rev C) eingesetzt.
Die Zusammensetzung des Master Mix lautete wie folgt:
Komponente Volumen
Aqua dest 0,2 µl
10x Puffer 0,8 µl
dNTP mix (100 mM total) 0,2 µl
MgCl2 (25 mM) 0,9 µl
Multiscribe™ RT Enzym (50 U/µl) 1,5 µl
RNase Inhibitor (20 U/µl) 0,1 µl
Tabelle 5 Master Mix für die RT-PCR (Ansatz für ein e Probe)
Dann erfolgte die Zugabe von jeweils 0,8 µl Megaplex RT Primer (entweder Human
pool A v2.1 oder B v3.0) und der zuvor isolierten RNA-Probe (350 ng in 3 µl Aqua dest),
sodass sich ein Gesamtvolumen von 7,5 µl für den Ansatz ergab.
Nach fünfminütiger Inkubation auf Eis folgte die reverse Transkription in einem
Thermocycler (Biometra) unter den nachfolgend genannten Bedingungen (40 Zyklen).

25
Temperatur Zeit Schritt
16°C 2 min. Zyklus (40 Zyklen)
42°C 1 min.
50°C 1 sec.
85°C 5 min. Halten
4°C ∞ Pause
Tabelle 6 Thermocycler Programm (Reverse Transkript ion)
Die so entstandene cDNA wurde bei -20°C verwahrt und innerhalb von sieben Tagen
weiter verarbeitet.
2.2.4.6. Quantitative Real Time- Polymerase-Kettenr eaktion (qPCR) Im zweiten Schritt erfolgte die quantitative Real-time-Polymerase-Kettenreaktion
(qPCR): Die Vervielfältigung der zuvor synthetisierten cDNA mittels Polymerase-
Kettenreaktion und die gleichzeitige Messung der Menge des entstehenden Amplifikats
in Echtzeit. Die quantitative Detektion der Amplifikate erfolgte über die Bestimmung
von Fluoreszenzintensitäten. Hierzu kamen Oligonukleotidsonden (TaqMan-Sonden)
zum Einsatz, die an einem Ende mit einem Reporterfluoreszenzfarbstoff und am
anderen Ende mit einem sogenannten Quencher markiert waren. Aus der räumlichen
Nähe von Reporter und Quencher ergab sich zunächst eine vollständige Suppression
der Fluoreszenz.
Die Sonden hybridisierten spezifisch an bestimmte DNA-Abschnitte. Sie wurden so
gewählt, dass ihre Zielsequenzen in einem DNA-Abschnitt lagen, der durch ein
spezifisches Primerpaar amplifiziert wurde.
Wenn im Verlauf der PCR die verwendete Taq-Polymerase mittels ihrer 5’-3’-Exo-
nukleaseaktivität bei der Gegenstrangsynthese das 5’-Ende der Sonde hydrolytisch
abspaltete, kam es zur Trennung von Reporter und Quencher. Dieser Schritt resultierte
in einer steigenden Fluoreszenzintensität definierter Wellenlänge des angeregten
Reporterfluoreszenzfarbstoffes, welche zum Ende jedes Zyklus detektiert wurde. Stieg
die Intensität des spezifischen Fluoreszenzsignals in einer Probe über das un-
spezifische Hintergrundrauschen und erreichte ein definiertes Fluoreszenzniveau
(Baseline) war der sogenannte Crossing point (CP) oder auch der threshold cycle (Ct)
erreicht. Ab diesem Zeitpunkt verlief die Reaktion nicht mehr linear, sondern
exponentiell. Ab hier war die gemessene Fluoreszenzintensität proportional zu der
Anzahl der Amplifikate und somit zur der Ausgangsmenge der cDNA.

26
Hieraus ergab sich das Maß für die Quantifizierung der Startmenge: Die Anzahl der
PCR-Zyklen die nötig war, um ein definiertes Fluoreszenzniveau zu erreichen.
Die Quantifizierung der MicroRNAs wurde als quantitative RT-PCR basierte Array-
Methode (qRCR-array) unter Verwendung des TaqMan® Universal PCR Master Mix
No AmpErase® UNG und der TaqMan® Array Human MircoRNA A+B cards (alles von
Applied Biosystems) mit Hilfe des 7900HT Real-Time PCR-Systems (Applied
Biosystems) durchgeführt. Das System-Set besteht aus zwei 384-well Reaktionsplatten,
die jeweils mit TaqMan® MicroRNA-Assays (Primern etc.) vorgeladen sind. Drei
endogene und eine negative Kontrolle sind jeweils in den Karten inkludiert, um die
spätere Normalisierung der Daten zu vereinfachen. Mit dem Set konnten in jeweils
zwei qPCR-Läufen 760 microRNAs pro Probe analysiert werden.
Alle Reagenzien wurden gemäß des Megaplex™ Pools Protocol (Applied Biosystems,
P/N 4399721 Rev C) eingesetzt. Für jeden Real-Time-PCR-Lauf wurden hierzu die
Reagenzien in folgender Menge zusammengefügt (siehe Tabelle 6):
Kompon ente Volumen
Universal PCR Master Mix 400µl
Megaplex RT-Produkt 5,3µl
Nukleasefreies Wasser 395µl
Tabelle 7 Ansatz für die Real-Time PCR
Mit dem so entstandenen Gesamtvolumen von 800µl wurde jeweils eine Karte beladen.
Die weitere Prozessierung der Karten erfolgte nach Angaben des Herstellers (Applied
Biosystems TaqMan® Array User Bulletin, P/N 4371129 Rev. B).
Pro Reaktionsplatte wurde ein qPCR-Lauf mit 45 Zyklen durchgeführt.

27
Abbildung 3 Prinzip der RT-qPCR
Modifiziert nach www.appliedbiosystems.com (Zugriffsdatum 23.06.2014)
2.2.5. Statistische Analysen der RT-qPCR- Ergebniss e Zur Darstellung der Resultate der qPCR wurden die automatisch generierten SDS-
Dateien (SDS-Software v2.3, Applied Biosystems) in eine Arbeitsmappe des RQ-
Managers (RQ-manager 1.2.1, Applied Biosystems) transferiert. Nach Empfehlungen
des System-Herstellers Applied Biosystems wurden die Studiendaten unter
Verwendung der automatischen Baseline und einem Threshold Cycle von 0,2
analysiert. Die ermittelten Ct-Werte wurden für die statistischen Analysen in eine Excel-
Tabelle exportiert.
Die statistischen Analysen erfolgten in Zusammenarbeit mit Damian Gola und Dr. Arne
Schillert vom Institut für Medizinische Biometrie und Statistik, Universität Lübeck,
Lübeck.
miR Stem-loop Primer
1. RT-PCR
2. qPCR
forward Primer
reverse Primer TaqMan®-Sonde

28
2.2.5.1. Präprozessierung Insgesamt wurden pro Patientenprobe 760 microRNAs (jeweils 380 auf Platten A und
B) untersucht.
In Anlehnung an verschiedene Publikationen, u.a. von He et al. und Vasilescu et al.,
wurden diejenigen Ct-Werte als fehlend (missing value) betrachtet, die einen Wert über
44 zeigten (57, 157). Platten A und B wurden daraufhin unabhängig voneinander einer
Präprozessierung unterzogen: Um relevante microRNAs für die weitere Analyse und
Diskussion herauszufiltern, wurden in einem ersten Schritt diejenigen microRNAs
exkludiert, die in über 50% der Fälle fehlende Ct-Werte zeigten. Für die Platte A
blieben so 151, für die Platte B 82 microRNAs erhalten. Daraufhin wurden alle Proben
ausgeschlossen, bei denen mehr als 45% der Ct-Werte für diese microRNAs fehlten.
Für die Platte A wurden so 4, für die Platte B 6 Lymphom-Proben von weiteren
Analysen ausgeschlossen. In einem dritten Schritt wurden erneut microRNAs gesucht,
die in den erhaltenen Fällen noch über 50% fehlende Ct-Werte zeigten. Hier wurden
keine weiteren microRNAs ausgeschlossen.
2.2.5.2. Quantilnormalisierung
Die vergleichende Analyse von Experimenten, die –wie hier- multiple Arrays involvieren,
erfordert zudem eine Normalisierung der Daten, da z.B. während der Proben-
präparation und -prozessierung nicht-biologische Variationen auftreten können (15).
Die Normalisierung der Datensätze erfolgte hier mithilfe der Quantilnormalisierung. Die
Grundidee dieser Methode ist einfach: Die Histogramme aller Arrays sehen gleich aus.
D.h. nicht nur das 50%-Quantil (= der Median), sondern alle Quantile sollen für alle
Arrays in etwa gleich sein. Nachteil ist, dass in einzelnen Arrays differentiell exprimierte
MicroRNAs am unteren und oberen Ende der Messskala angeglichen werden.

29
Abbildung 4 Boxplot der CT-Werte pro Fall und Platt e nach Präprozessierung
Freundlicherweise zur Verfügung gestellt von Gola und Schillert
Abbildung 5 Boxplots der CT-Werte pro Fall und Plat te nach Quantilnormalisierung
Freundlicherweise zur Verfügung gestellt von Gola und Schillert

30
2.2.5.3. Fall-Kontroll-Analyse
Um Unterschiede in der microRNA-Expression von Lymphom-Gruppe zu Kontroll-
Gruppe herauszuarbeiten wurde der Wilcoxon-Mann-Whitney-Test angewandt. Zur
Kontrolle der Irrtumswahrscheinlichkeit kam die Bonferroni-Holm-Prozedur zum Einsatz.
Das globale Signifikanz-Level wurde mit α (G) = 0,05 gewählt. Das lokale Signifikanz-
Level wurde wie folgt berechnet:
Formel 1 Berechnung des lokalen Signifikanzlevels
Für den Vergleich von mehr als zwei Gruppen im Rahmen der Subgruppenanalyse
kam der Kruskal-Wallis-Test zum Einsatz.
Die Durchführung der genannten statistischen Tests erfolgte mit R, Version 3.0.2
(2013-09-05).

31
3. Ergebnisse
3.1. Charakteristika der Patienten/innen Die dreißig Patienten der Gruppe angioimmunoblastischer T-Zell-Lymphome waren bei
Lymphknotenextirpation im Mittel 69,5 Jahre alt (43 bis 90 Jahre). Die Lymphknoten
stammen von 13 Frauen und 17 Männern.
Das Profil der Patienten in der Studiengruppe entspricht somit im Hinblick auf Alter und
Geschlecht in etwa dem in bisher veröffentlichten Studien (36, 43, 165).
Weitere klinische Angaben zu den Patienten lagen nicht vor.
Die zehn Patienten der Kontrollgruppe waren im Schnitt 36,6 Jahre alt, darunter 6
Frauen und 4 Männer.
3.2. Ergebnisse der histopathologischen Analysen
3.2.1. Ergebnisse der Analyse der Gewebsschnitte hi nsichtlich des histologischen Musters und der Proliferation der hochendothelialen Venolen und der FDC-Netzwerke
Die Auswertung hinsichtlich des histologischen Musters und der Proliferation sowohl
der hochendothelialen Venolen als auch der FDC-Netzwerke erfolgte licht-
mikroskopisch (Axioskop, Zeiss) nach Anwendung der oben aufgeführten histo-
chemischen Färbeverfahren und der immunhistochemischen Darstellung der FDC-
Netzwerke mithilfe der Primärantikörper gegen CD21 und CD23. Um einen für diese
Analyse angemessenen Gewebsabschnitt zu überblicken, wurden hierfür Schnitte der
vollständigen Lymphknotenproben herangezogen. Die Analyse erfolgte im Vier-Augen-
Prinzip durch Herrn Prof. Dr. med. A.C. Feller und Frau K. Reddemann.
Anhand der Analyse der konventionell histochemisch gefärbten Schnittpräparate der
vollständigen Lymphknotenproben wurde das morphologische Muster der angio-
immunoblastischen T-Zell-Lymphome, gemäß den von Attygalle 2002 aufgestellten
Kriterien bestimmt (8). Alle Lymphomfälle konnten einem der drei einleitend
beschriebenen histomorphologischen Muster zugeordnet werden (vgl. Einleitung). Alle
drei Muster wurden durch die ausgesuchten Fälle abgedeckt: Muster 1 wurde in 10%
(3/30), Muster 2 in 47% (14/30) und Muster 3 in 43% (13/30) der Fälle diagnostiziert.

32
Abbildung 6 Histomorphologisches Muster der AITL (n =30)
Die Proliferation der hochendothelialen Venolen, genauso wie die der follikulären
dendritschen Netzwerke findet in der Bestimmung des Musters nach Attygalle bereits
ihre Berücksichtigung. Da zum Beispiel die Neovaskularisation als Antwort auf die
Tumorzellen durch verschiedene Zellen des Mikromilieus verstanden wird und einen
prognostischen Faktor ebenso wie einen therapeutischen Ansatzpunkt darstellt (176),
haben wir beide Charakteristika für weitere Analysen außerdem separat betrachtet. Die
Dichte und Ausbreitung der kleinen Gefäße sowie die Proliferation der follikulären
dendritischen (FDC) Netzwerke wurde hierfür anhand der vollständigen Lymphknoten-
proben semiquantitativ mit einem entsprechend ansteigenden Score von 1 bis 3
bewertet (vgl. Tabelle 7).
Proliferat\Score 1 2 3 Venolen Keine oder gering
vermehrte Gefäß-strukturen, keine vermehrten Aufzweigungen
Mäßig vermehrte Gefäßstrukturen, vermehrte Gefäß-aufzweigungen
Stark vermehrte Gefäßstrukturen, multiple Gefäß-aufzweigungen
FDC-Netzwerke Keine Proliferation über die Follikel hinaus
Geringe bis mäßige Proliferation über die Follikel hinaus
Ausgedehnte, unregelmäßige Proliferation über die Follikel hinaus
Tabelle 8 Graduierung der Proliferation der hochend othelialen Venolen und der FDC-Netzwerke mit Zuordnung der Score-Werte

33
Hierbei wurde für die Gefäßproliferation in 40% der Fälle der Score von 2 vergeben, in
37% bzw. 23% (7/30) wurde ein Punktwert von 3 bzw. 1 zugeteilt.
Bei der vom Muster unabhängigen Betrachtung der FDC-Netzwerke wurde in jeweils
28,5 % der Wert 1 bzw. 3 vergeben. In 43% der Fälle ein Score von 2. Für zwei
Lymphomproben waren die Färbungen aus technischen Gründen nicht adäquat
auswertbar. Die Ergebnisse sind in Tabelle 21 (s. Anhang) dargestellt.
Die durchschnittlichen Punktwerte sowohl für die Proliferation der Venolen als auch für
die Ausdehnung der FDC-Netzwerke stiegen von Muster 1 bis Muster 3 kontinuierlich
an. Die Proliferation der Venolen wurde tendenziell in Mustern 1 und 2 mit einem
durchschnittlich höheren Score bewertet als die Expansion der FDC-Netzwerke.
Abbildung 7 Histomorphologisches Bild eines AITL
Fall 6, HE, Originalvergrößerung: 100x, Histomorphologisches Muster Score 3, Vaskularität Score 2

34
Abbildung 8 Beispiele der histomorphologischen Must er nach Attygalle et al. sowie
der Graduierung der Proliferation der hochendotheli alen Venole n und der FDC-Netzwerke.
A: Muster 1, B: Muster 2, C: Muster 3; HE, Originalvergrößerung: 100x; D: Venolen Score 1, E: Venolen Score 2, F: Venolen Score 3; Gomori Versilber-ung, Original-vergrößerung: 100x; G: FDC Score 1, H: FDC Score 2, I: FDC Score 3; CD23, Originalvergrößerung: 100x. A,D,G: Fall-Nr. 29, B,E,H: Fall-Nr. 11, C,F,I: Fall-Nr. 17.

35
29%
42%
29%
Score 1
Score 2
Score 3
Abbildung 9 Bewertung der Expansion der FDC-Netzwer ke
(nach Scores, prozentual, n=28)
23%
40%
37%
Score 1
Score 2
Score 3
Abbildung 10 Bewertung der Proliferation der hochen dothelialen Venolen
(nach Scores, prozentual, n=30)
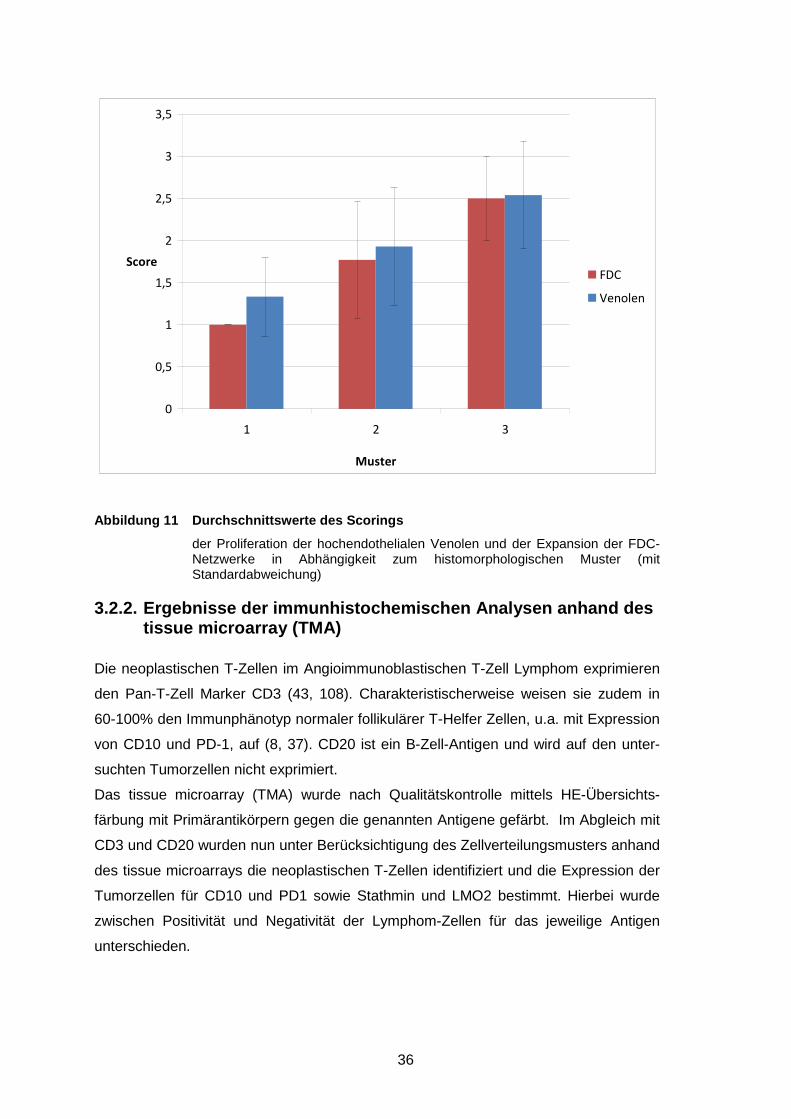
36
0
0,5
1
1,5
2
2,5
3
3,5
1 2 3
Muster
Score
FDC
Venolen
Abbildung 11 Durchschnittswerte des Scorings
der Proliferation der hochendothelialen Venolen und der Expansion der FDC-Netzwerke in Abhängigkeit zum histomorphologischen Muster (mit Standardabweichung)
3.2.2. Ergebnisse der immunhistochemischen Analysen anhand des tissue microarray (TMA)
Die neoplastischen T-Zellen im Angioimmunoblastischen T-Zell Lymphom exprimieren
den Pan-T-Zell Marker CD3 (43, 108). Charakteristischerweise weisen sie zudem in
60-100% den Immunphänotyp normaler follikulärer T-Helfer Zellen, u.a. mit Expression
von CD10 und PD-1, auf (8, 37). CD20 ist ein B-Zell-Antigen und wird auf den unter-
suchten Tumorzellen nicht exprimiert.
Das tissue microarray (TMA) wurde nach Qualitätskontrolle mittels HE-Übersichts-
färbung mit Primärantikörpern gegen die genannten Antigene gefärbt. Im Abgleich mit
CD3 und CD20 wurden nun unter Berücksichtigung des Zellverteilungsmusters anhand
des tissue microarrays die neoplastischen T-Zellen identifiziert und die Expression der
Tumorzellen für CD10 und PD1 sowie Stathmin und LMO2 bestimmt. Hierbei wurde
zwischen Positivität und Negativität der Lymphom-Zellen für das jeweilige Antigen
unterschieden.

37
Die Auswertung erfolgte lichtmikroskopisch (Axioskop, Zeiss) im Konsensusverfahren
und Vier-Augen-Prinzip (J. Knief, K.Reddemann). Die Kontrolle erfolgte durch Herrn
Prof. Dr. med. Ch. Thorns im Konsensusverfahren.
Um eine gewisse Vergleichbarkeit mit anderen Publikationen zu gewährleisten, wurden
bereits für diese Antigene und diese Lymphom-Entität beschriebene Grenzwerte
berücksichtigt (Tabelle 8).
Antigen/Ziel Grenzwert (Pos itivität
liegt vor, wenn….)
Referenz
CD10 > 5% des CD3-positiven T-
Zell-Infiltrats positiv sind
(8, 39)
LMO2 > 30% der Lymphomzellen
positiv sind
(110)
PD1 > 20% der Lymphomzellen
positiv sind
(171)
Stathmin > 40% der Lymphomzellen
positiv sind
(101)
Tabelle 9 Grenzwerte der immunhistochemischen Färbu ngen
CD10 wurde in unserer Fallauswahl nur in 13% der Fälle auf den Tumorzellen
nachgewiesen. Für PD1 dagegen konnte die hohe Expressionsrate bestätigt werden:
77% (85% bei 23 von 27 positiv, bei 3 nicht auswertbaren Fällen).
Wir konnten außerdem zeigen, dass die Tumorzellen in 67% der untersuchten Fälle
Stathmin exprimieren (71% bei 20/28 positiven und 2 nicht auswertbaren Fällen).
LMO2 wird, bei vier aufgrund von Schnittartefakten nicht verwertbaren Fällen, in
keinem der untersuchten Fälle auf den neoplastischen T-Zellen exprimiert.
In nur 10% der Fälle wurden die Tumorzellen als positiv für alle drei beschrieben
Marker bewertet. Die größten Überschneidungen zeigten sich bei der Positivität für
PD1 und Stathmin (50%). In 17% bzw. 7% wurden die atypischen CD3+T-Zellen allein
durch den Marker PD1 bzw. Stathmin markiert. In nur einem Fall lag ausschließlich
CD10 auf den atypischen CD3+T-Zellen vor. Ebenfalls in nur einem Fall ließ sich
keiner der Marker verlässlich auf der postulierten Tumorzellpopulation nachweisen.
Die Ausfallquote der nicht-beurteilbaren Stanzen unter Berücksichtigung der TMAs für
Stathmin, CD10 und PD1 belief sich insgesamt auf 10%, vergleichbar mit Angaben in
den Leitlinien von Ilyas et al. (64).
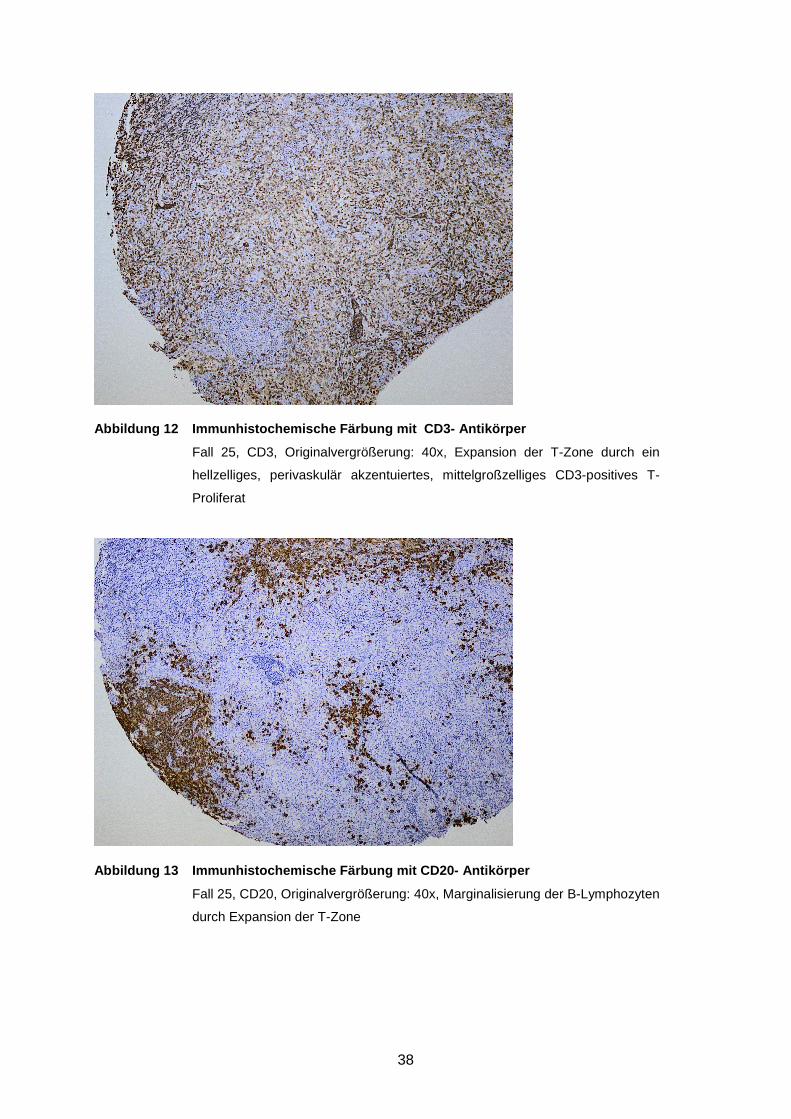
38
Abbildung 12 Immunhistochemische Färbung mit CD3- Antikörper
Fall 25, CD3, Originalvergrößerung: 40x, Expansion der T-Zone durch ein
hellzelliges, perivaskulär akzentuiertes, mittelgroßzelliges CD3-positives T-
Proliferat
Abbildung 13 Immunhistochemische Färbung mit CD20- Antikörper
Fall 25, CD20, Originalvergrößerung: 40x, Marginalisierung der B-Lymphozyten
durch Expansion der T-Zone

39
Abbildung 14 Immunhistochemische Färbung mit Stathm in- Antikörper
Fall 20, Stathmin, Originalvergrößerung:100x, Markierung des perivaskulär ak-
zentuierten, mittelgroßzelligen, atypischen T-Zell-Proliferats mit Stathmin.
Abbildung 15 Immunhistochemische Färbung mit CD10- Antikörper
Fall 25, CD10, Originalvergrößerung:100x, Markierung des perivaskulär akzen-
tuierten, mittelgroßzelligen, atypischen T-Zell-Proliferats mit CD10.
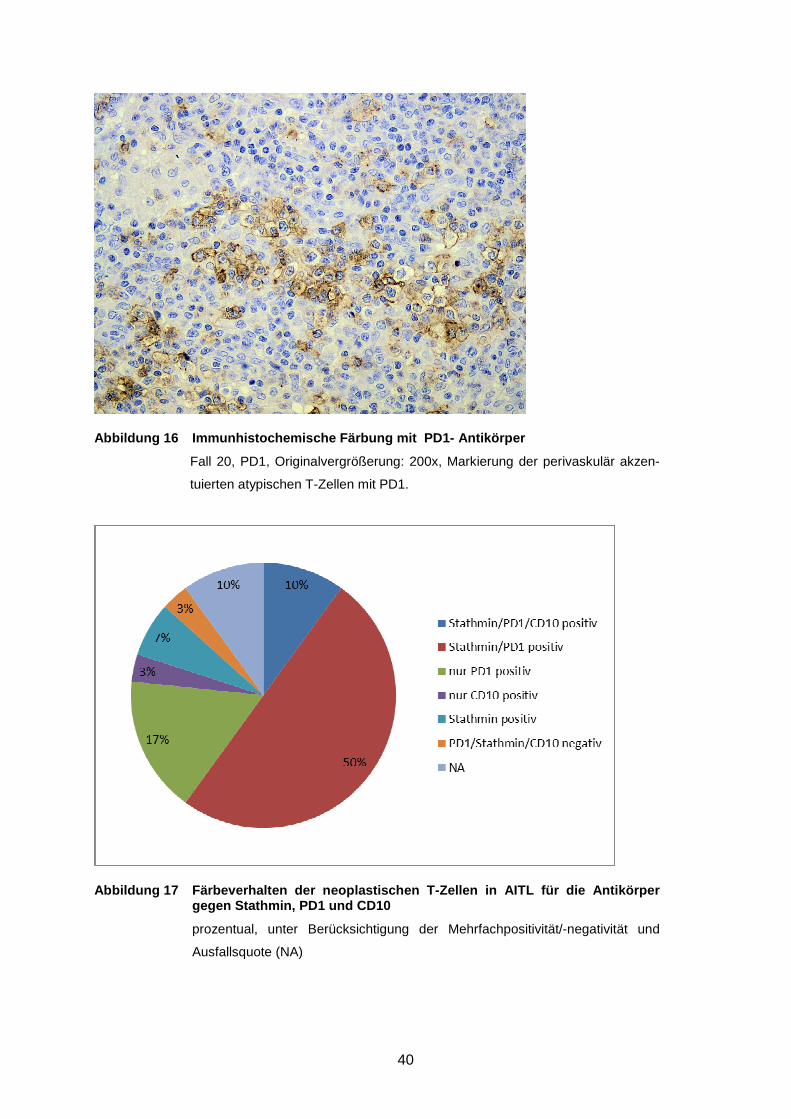
40
Abbildung 16 Immunhistochemische Färbung mit PD1- Antikörper
Fall 20, PD1, Originalvergrößerung: 200x, Markierung der perivaskulär akzen-
tuierten atypischen T-Zellen mit PD1.
Abbildung 17 Färbeverhalten der neoplastischen T-Ze llen in AITL für die Antikörper gegen Stathmin, PD1 und CD10
prozentual, unter Berücksichtigung der Mehrfachpositivität/-negativität und
Ausfallsquote (NA)

41
3.3. Ergebnisse der EBER-ISH Anhand der In-situ-Hybridisierung für EBER wurde mittels des tissue microarrays die
An- bzw. Abwesenheit von EBV-positiven B-Blasten im Begleitinfiltrat des Lymphoms
ermittelt.
Hierbei wurden diejenigen Fälle als positiv bewertet, die 50 und mehr positive EBV-
positive Blasten pro low power field (hier definiert 1 mm Objektfelddurchmesser)
aufwiesen.
42% (11/26) der Fälle konnten so als positiv bewertet werden. Vier Fälle waren
aufgrund von Schnittartefakten des tissue microarrays nicht adäquat auswertbar.
Auch ein Fall der Kontrollgruppe wurde so als EBV-positiv identifiziert.
Abbildung 18 In-situ Hybridisierung für EBER
Fall 5, EBER, Ausschnitt eines Low-power-fields, Originalvergrößerung:200x,

42
3.4. Ergebnisse der microRNA-Quantifizierung
In Konkordanz mit verschiedenen Leitlinien für Experimente mit tissue microarrays und
qPCR wurden die statistischen Analysen von Experten, Damian Gola und Arne
Schillert aus dem Institut für Biometrie und Statistik, Universität zu Lübeck,
durchgeführt (20, 64).
Die Nullhypothese lautete: Es gibt keinen Unterschied in der microRNA-Expression
zwischen der Fall- und der Kontroll-Gruppe.
Für sieben microRNAs konnte auf dem nach Bonferroni-Holm lokal adjustierten
Signifikanzlevel die Null-Hypothese zurückgewiesen werden. Alle microRNAs sind für
den Menschen beschrieben, im Folgenden wird somit auf den expliziten Zusatz
„hsa“ für die microRNAs verzichtet.
Vier microRNAs, wiesen in den untersuchten Lymphomfällen ein signifikant niedrigeres
Expressionslevel auf als die Fälle der nicht-neoplastischen Kontrollgruppe: miR-140-3p,
let-7g, miR-30b und miR-664.
Drei microRNAs zeigten dagegen eine signifikant höhere Expression in der AITL-
Gruppe: miR-146a, miR-193b und miR-34a.
Außerdem konnte die Nullhypothese auch für RNU44, eine kleine nukleoläre RNA
(snoRNA), verneint werden. RNU44 zeigte sich in der Lymphom-Gruppe signifikant
runterreguliert.
microRNA Medianer
∆CT
fold change
(Veränderungen
um das n-fache)
p-Wert Adjustierter
p-Wert
Konfidenz-
intervall
miR-140-3p -1,51 0,35 0,000000527 0,000123 [-2,31, -1,06]
Let-7g -1,41 0,38 0,00001 0,00254 [-2,19, -0,90]
miR-146a 1,27 2,42 0,0000266 0,00615 [0,61, 1,48]
miR-193b 0,91 1,88 0,0000307 0,00706 [0,66, 1,56]
miR-30b -0,93 0,53 0,0000543 0,0124 [-1,70, -0,43]
miR-664 -0,55 0,68 0,0000913 0,0207 [-1,08, -0,33]
miR-34a 2,45 5,45 0,000102 0,0231 [1,35, 3,36]
RNU44 -1,32 0,4 0,0000478 0,011 [-2,65, -0,75]
Tabelle 10 Signifikant aberrant exprimierte RNAs in der Fall-Kontroll-Analyse
Mit p-Werten, adjustiert nach der Bonferroni-Holm-Prozedur, und Konfidenz-
intervallen

43
Abbildung 19 Boxplots der CT-Werte der signifikant aberrant exprimierten microRNAs nach ansteigenden adjustierten p-Werten
Freundlicherweise zur Verfügung gestellt von Gola und Schillert Bezogen auf die Subgruppen, die sich durch die Positivität bzw. Negativität der
neoplastischen T-Zellen für CD10, PD1 und Stathmin, sowie durch die An-bzw. Ab-
wesenheit EBV-positiver B-Blasten ergaben, konnte für keine microRNA die Null-
hypothese auf dem nach Bonferroni-Holm adjustierten Signifikanzlevel verworfen
werden. Auch im Vergleich von drei Gruppen unter Anwendung des Kruskal-Wallis-
Tests, fand sich kein signifikanter Unterschied in der microRNA-Expression in Bezug
auf die Gefäßproliferation, die FDC-Expansion oder das histomorphologische Muster.
Statistisch war somit insgesamt kein Unterschied der microRNA-Expression in den
verschiedenen Gruppen messbar.
Verzichtet man auf die Adjustierung und sortiert man aberrant exprimerte microRNAs
pro Subgruppe nach aufsteigendem p-Wert, ergeben sich 7 Tabellen, deren Top Ten
im Anhang zu finden sind (Tabellen 26-32).
Eine Klassifikation der microRNA-Expression anhand des histologischen Musters oder
des Gefäß- bzw. FDC-Scores war aufgrund des unvollständigen Datensatzes, der sich
durch die Präprozessierung ergab, nicht sinnvoll möglich.

44
4. Diskussion
4.1. Morphologie - Die Proliferation der Venolen al s frühes Merkmal von AITL
Das morphologische Profil angioimmunoblastischer T-Zell-Lymphome ist zwar charak-
teristisch, jedoch keineswegs spezifisch für die Erkrankung und zeigt Überschnei-
dungen zu anderen Neoplasien oder reaktiven Lymphknotenveränderungen (104). Man
kann davon ausgehen, dass gerade Frühformen histologisch oft nicht erkannt werden
und deswegen die meisten Fälle erst in fortgeschrittenem Stadium diagnostiziert
werden. Letztendlich kann auch die schlechte Prognose der Erkrankung zum Teil
darauf zurückgeführt werden.
Setzt man voraus, dass die von Attygalle et al. beschriebenen histomorphologischen
Muster Progressionsstadien der Erkrankung darstellen, konnte anhand der hier
untersuchten und klassifizierten dreißig AITL-Fälle ein positiv korrelierender Trend
sowohl der Proliferation der Gefäße als auch der FDC-Netzwerke mit ansteigendem
Grad der Architekurstörungen bestätigt werden.
Zudem ließ sich anhand der Durchschnittswerte des semiquantitativen Scorings
feststellen, dass tendenziell die Proliferation der Gefäße in früheren Krankheitsstadien
(Muster 1 und 2) früher höher bewertet wurde als die Expansion der FDC-Netzwerke.
In der Praxis könnte somit das Merkmal der Gefäßproliferation im Frühstadium den
Weg für die richtige Diagnose bereits bahnen, auch wenn die FDC-Expansion noch
nicht erkennbar ist.
Diese Beobachtung könnte auch auf eine sequentielle Krankheitsgenese hindeuten.
Vergleichsweise vermuten Pileri et al., dass die VEGF Deregulation, die auch für das
kutane T-Zell-Lymphom Mycosis fungoides beschrieben wurde, in dieser Entität ein
frühes Ereignis darstellt (126). Die Induktion der Angiogenese durch eine abnorme
VEGF-Expression sowohl der neoplastischen als auch der nicht-neoplastischen
Bystander-Zellen könnte in AITL ebenfalls ein erstes Ereignis sein, das die Krankheits-
entstehung und das Tumorwachstum erst begünstigt.

45
4.2. Immunhistochemie
4.2.1. Stathmin als relevanter und möglicherweise C D10 überlegener Marker in AITL
Stathmin, oder auch Onkoprotein 18, wurde ursprünglich als Onkoprotein in T-Zell-
Leukämie-Zellen charakterisiert (103). Die Stathmin-Protein Familie ist hochgradig kon-
serviert. Durch Bindung an freie Tubulinheterodimere im Zytoplasma sind sie Regu-
latoren des dynamischen Netzwerks von Mikrotubuli-Filamenten und des Mikrotubulus-
Organisationszentrums (MTOC) in einer Zelle (31). Filbert et al. konnten zeigen, dass
Stathmin in T-Zellen nach Aktivierung über den T-Zell-Rezeptor ERK-abhängig phos-
phoryliert wird und so das Mikrotubulus –Wachstum verstärkt (44). Verma et al. fanden
heraus, dass Stathmin unter Einflussnahme des Transkriptionsfaktors STAT3 die
Mikrotubulus-Dynamik in migierenden T-Zellen reguliert, was auch die Zelladhäsion an
hochendotheliale Venolen mitinvolviert (158).
Belletti und Baldassarre berichten, dass eine hohe Stathmin-Expression das An-
sprechen maligner Tumore auf Chemotherapeutika, deren Zielstrukturen das Mikro-
tubulus-Netzwerk darstellen (z.B. Vincristin, Bestandteil des CHOP-Therapie-Regimes),
negativ beeinflussen kann (13).
Die Diagnosestellung von Lymphomen ist anspruchsvoll und erfordert zumeist ein
Panel aus mehreren immunhistochemischen Markern. Gerade im Hinblick auf die Sub-
klassifizierung von T-Zell-Lymphomen ist die Identifikation hilfreicher, wenn möglich
sogar spezifischer Marker von großem Interesse, da ihre bisherige Anzahl weit hinter
denen der B-Zell-Lymphome zurücksteht (148).
Anhand von 55 Fällen zeigten Marafioti et al., dass Stathmin auch in neoplastischen
Zellen von T-Zell-Lymphomen exprimiert wird (101). Darunter zeigten sich auch 6 von
8 Fälle von angioimmunoblastischen T-Zell-Lymphomen positiv für Stathmin.
Stathmin könnte somit in der Praxis als immunhistochemischer Marker insbesondere
für angioimmunoblastische T-Zell-Lymphome interessant sein. Weiterhin wäre die
Korrelation zwischen Therapieansprechen und Positivität der Tumorzellen für Stathmin
möglicherweise ein Ansatzpunkt für weitere Forschungsvorhaben. Gründe, die Beo-
bachtung von Marafioti et al. mit einer größeren Fallzahl zu verifizieren und mit bereits
etablierten Markern (PD1 und CD10) für AITL zu vergleichen.
Die Positivität der neoplastischen T-Zellen für Stathmin in den untersuchten Lymph-
knotenproben, befallen von AITL, konnte in 71% (20 von 28 Fällen) bestätigt werden.
Prozentual ist das Ergebnis mit dem von Marafioti et al. (75%) vergleichbar. Stärkste
Überschneidungen zeigte die Stathmin-Expression mit PD-1, das in 85% der Fälle von
den Tumorzellen exprimiert wurde.

46
CD10 ist eine Zellmembrangebundene Metallopeptidase, die insbesondere von B-
Lymphozyten der follikulären Keimzentren exprimiert wird (39). Die Rate der CD10-
positiven AITL-Fälle blieb mit 13% weit hinter den in der Literatur angegebenen 71 bis
90% zurück (39, 104, 108). Anzumerken ist jedoch, dass die CD10 –Positivität der
Tumorzellen in AITL bereits oft als heterogen (nur auf einem Teil der Tumorzellen und
in variabler Intensität) beschrieben und vereinzelt auch kontrovers diskutiert wurde (36,
154). Hierbei wird ein Nachteil der tissue microarray Methode erkennbar: Der
Durchmesser der Gewebsstanzen ist oft nicht groß genug, um bei heterogener
Antigenverteilung im Gewebe sicher eine Positivität auszuschließen. Dieser Pro-
blematik wurde versucht durch sorgfältige Fall- und Arealauswahl entgegenzusteuern.
Außerdem wurde mit 2 mm der Durchmesser der Stanzen relativ groß gewählt. Ein
weiterer Grund der niedrigen Rate CD10-positiver AITL-Fälle in der vorliegenden
Studie ist die subjektive Wahrnehmung des Untersuchers. Gerade für CD10 ist bei
spärlichen internen Kontrollen (Stroma-Fibroblasten und Granulozyten) und eventuell
zusätzlich suboptimalen Färbungen eine Bewertung als schwierig beschrieben worden
(34). Hier jedoch zeigten sich die Vorteile des TMA: Es konnte Kontrollgewebe mit
verbaut werden und durch Komprimierung der Fälle auf einen einzigen Schnitt erfolgte
die sorgfältige manuelle Färbung für alle Proben unter gleichen Bedingungen. Und
durch die Auswertung im Konsensusverfahren im Vier-Augen-Prinzip sowie ledigliche
Dichotomie der Merkmale sollte der Interobserver-Variabilität ausreichend Rechnung
getragen worden sein.
Nichtdestotrotz sind semiquantitative immunhistochemische Analysen von Lymphomen
immer kritisch zu bewerten (vgl hierzu auch (34)). Eine Doppelpositivität für Stathmin
und CD10 mit Negativität für PD1 war nicht nachweisbar.
In 2 Fällen konnte ausschließlich Stathmin die neoplastische T-Zell-Population an-
färben, sowohl CD10 als auch PD1 wurden hier als negativ bewertet.
Daran ist ein möglicher Informationszugewinn durch Stathmin im Antikörper-Panel zur
Diagnosesicherung eines AITL abzulesen.
Die Färbungen bleiben mit verlässlichen Doppelfärbungen zu validieren.
4.2.2. LMO2 wird nicht von AITL-Zellen exprimiert
LMO2 (LIM domain only 2) ist ein Cystein-reicher Transkriptionsfaktor, der eine
tragende Rolle in der Hämatopoese spielt (110, 119). In Patienten mit akuter
lymphoblastischer T-Zell-Leukämie wird LMO2 durch die chromosomalen Trans-
lokationen t(7;11)(q35;p13) oder t(11;14)(p13;q11) aktiviert (110). Die LMO2-Ex-
pression ist außerdem eng mit dem Keimzentrums-phänotyp von B-Lymphozyten

47
verbunden (2). Mittlerweile ist ein monoklonaler Kaninchen-Antikörper für immunhisto-
chemische Färbungen in Paraffin-eingebetten Gewebe verfügbar, der in der Routine-
Diagnostik eingesetzt werden kann (3).
Dyhdalo et al. konnten zeigten, dass LMO2 in Ergänzung zu den Markern CD10 und
bcl6 die Bestimmung des Keimzentrum-Phänotyps in follikulären Lymphomen verein-
facht und bei der Abgrenzung zu Marginalzonenlymphomen helfen kann (40). Im
Hinblick auf normale T-Zellen dokumentieretn Natkunam et al., dass in normalem
lymphatischem Gewebe LMO2 weder auf CD3+-T-Zellen innerhalb noch außerhalb der
Keimzentren nachgewiesen werden konnte (110). Für PTCL allgemein ergaben Unter-
suchungen von Agostinelli et al., dass nur 9 von 237 Fälle LMO2-positive Elemente
zeigten (3).
Da der Immunphänotyp der neoplastischen T-Zellen in angioimmunoblastischen T-Zell-
Lymphomen oft Überscheidungen mit dem Phänotyp von Keimzentrums B-Zellen zeigt
und bisher AITL nicht im Hinblick auf ihre LMO2-Expression untersucht worden sind,
wurde dies hier nachgeholt. Vorteil des verwendeten Markers wäre seine strikte
nukleäre Anfärbung der positiven Zellen. Hier jedoch zeigte sich in keinem der dreißig
untersuchten AITL-Fälle ein mit dem Verteilungsmuster der neoplastischen CD3+T-
Zellen übereinstimmendes Muster für LMO2. Außerdem erschwerte die Positivität der
zahlreichen Endothelien die Auswertung, sodass LMO2 für die Identifikation der
neoplastischen T-Zellen in AITL ungeeignet erscheint.
4.3. MicroRNA-Expression
Die Interpretation und Diskussion der Ergebnisse der microRNA-Expressionsanalyse
erfolgte im Abgleich mit Publikationen aus der Online-Datenbank „pubmed“. Es wurden
Studien zu der Rolle von microRNAs sowohl in Krebserkrankungen und Lymphomen
allgemein als auch in der T-Zell-Entwicklung und -Maturation berücksichtigt, um so
Rückschlüsse auf Erkrankungsassoziierte Signalwege und Gene zu ziehen. Da in den
meisten Fällen die Ziel-Strukturen einer microRNA unbekannt bleiben, stellt der
Nachweis einer microRNA-Dysregulation im Vergleich zum Normalgewebe einen
ersten Schritt in der Charakterisierung physiologischer und pathologischer Prozesse
dar (14).
4.3.1. Mögliche Oncomir in AITL
Diejenigen microRNAs, deren Expression im Vergleich zu nicht-neoplastischem
Normalgewebe erhöht ist, kommen als mögliche Onkogene/ „Oncomirs“ in Betracht. Es
konnte gezeigt werden, dass sie die Tumorentwicklung begünstigen, indem sie Tumor-

48
suppressorgene und/oder Gene, die die Zelldifferenzierung und die Apoptose ver-
mitteln, hemmen (173). Grundsätzlich werden solche Krebs-Phänotypen als
schwerwiegender betrachtet, die von einer Überexpression, einem „Gain-of-function“,
ihrer Gene verursacht werden (116). Durch Hemmung von Genen, die normalerweise
in einem Zelltyp nicht unterdrückt werden, kann „microRNA Gain-of-function“ zu
aberranten Funktionen führen. Der Verlust der microRNA-Funktion (loss of function)
würde demnach nur Zellen betreffen, in denen das Ziel-Gen und die microRNA gleich-
zeitig exprimiert sind (116).
Die microRNAs miR-146a, miR-193b und miR-34a zeigten in den Lymphknoten der
untersuchten Fälle angioimmunoblastischer T-Zell-Lymphome im Vergleich zu den
Fällen der reaktiven Lymphadenitis eine signifikant höhere Expression. Sie kommen
somit für AITL als potentielle „Oncomirs“ in Betracht. Die höchste Signifikanz zeigt
darunter miR-146a (adjustierter p-Wert 0,000123). Den höchsten Fold change weist
miR-34a auf; sie wird über 5 mal höher in AITL als in reaktiv veränderten Lymphknoten
exprimiert.
MiR-146a wird laut „miRBase“ auf Chromosom 5 codiert. Nach Informationen der
Online-Datenbank „miRWalk“ sind für diese microRNA 689 Ziel-Gene validiert, 551
rechnerisch vorhergesagt worden.
MiR-146a ist ein Modulator des Immunsystems und spielt sowohl in myeloischen, als
auch lymphozytären Zellen eine Rolle (32). Ihre Expression kann in myeloischen Zellen
durch mikrobielle Komponenten oder proinflammatorische Zytokine induziert werden.
Zudem gehört sie zu der microRNA-Signatur normaler menschlicher T-Zellen (94). In
naiven T-Zellen ist ihre Expression niedrig, in T-Gedächtniszellen jedoch sehr hoch
(78). MiR-146a ist ein wichtiger Feedback-Regulator des NF-kappaB-Signalweges (11).
Durch zwei Bindungsstellen für NFkappaB im Promoter von miR-146a wird ihre
Expression im Zuge der Aktivierung der T-Zell-Rezeptor (TCR)-Signalkaskade
NFkappaB-abhängig hochreguliert (11, 128). Ziele von miR-146a sind wiederum der
TNF Rezeptor-assoziierte Faktor 6 (TRAF6) und die IL-1 Rezeptor-assoziierte Kinase 1
(IRAK1), die in einem negativen Feedback-Loop die Intensität und die Dauer des
NFkappaB-Signals kontrollieren (11, 150). In humanen T-Zellen kann eine miR-146a-
Überexpression die Produktion von IL-2 hemmen (143). In humanen B-Zellen kann die
miR-146a-Expression NFkappaB-abhängig durch das Epstein-Barr Virus latent
membrane protein 1 (LMP1) induziert werden (107, 143). Yang et al. stellten fest, dass
eine miR-146a-Defizienz eine Überstimulation der T-Zellen durch die TCR-
Signalkaskade, und somit eine ungehemmte T-Zell-vermittelte Entzündungsreaktion

49
hervorrufen kann (170). Weitere Loss-of-function-Studien an Mäusen zeigten, dass
miR-146a eine wichtige Rolle in der Entwicklung von Autoimmunerkrankungen spielen
kann. So führt der Verlust von miR-146a zu einer Hochregulation des Signal-
Transducer and Activator of Transcription 1 (STAT1) in regulatorischen T-Zellen, was
zu einer fatalen IFNgamma-vermittelten Autoimmunität führt (97). Auch bei Menschen,
die an den Autoimmunerkrankungen Lupus erythematodes und rheumatoider Arthritis
leiden, zeigt sich eine Dysregulation von miR-146a. Interessanterweise scheinen
gerade diese beiden Erkrankungen eng mit einer Dysregulation follikulärer T-Helfer-
Zellen, den postulierten Ursprungszellen des AITL, assoziiert zu sein (118). In
mononukleären Blutzellen von Lupus-Patienten ist die miR-146a-Expression herunter-
reguliert und zeigt eine negative Korrelation mit der Erkrankungsschwere (143). Im
Unterschied dazu ist in CD4+T-Zellen von Patienten mit rheumatoider Arthritis miR-
146a häufig hochreguliert und die Expression zeigt eine enge positive Korrelation mit
erhöhten TNFalpha-Leveln in diesen Patienten (89). Li et al. konnten in diesem Kontext
zeigen, dass eine Überexpression von miR-146a in vitro die Apoptose von T-Zellen
hemmt (89).
Auch in einigen Malignomen konnte eine aberrante miR-146a-Expression gezeigt
werden. In soliden Tumoren, unter anderem in anaplastischen Schilddrüsen-
karzinomen, Glioblastomen und Plattenepithelkarzinomen der Cervix uteri ließ sich bei
Vergleich mit dem entsprechenden Normalgewebe ebenfalls eine erhöhte Expression
von miR-146a nachweisen (81, 114, 161). In Karzinomen des Pankreas, Magens und
der Prostata zeigte sich die miR-146a-Expression jeweils im Vergleich zu dem
normalen Kontrapart herunterreguliert (74, 90, 92).
In EBV-positiven Burkitt-Lymphom-Zelllinien wurde eine erhöhte miR-146a-Expression
beschrieben (155). Peveling-Oberhag konnte nachweisen, dass diese microRNA in
splenischen Marginalzonen-lymphomen ebenfalls erhöht ist (122). Anhand von 90
Patientenproben haben Zhong et al. zudem demonstriert, dass niedrige miR-146a-
Level bei de-novo aufgetretenen diffusen großzelligen B-Zell-Lymphomen mit höheren
Remissionsraten, besserem Gesamtüberleben und längerem progressionsfreiem
Überleben assoziiert sind (177).
In Bezug auf T-Zell-Lymphome fanden Curtale et al. anhand der humanen Jurkat-T-
Zell-Lymphom-Zelllinie heraus, dass miR-146a in aktivierten T-Zellen über Modulation
des Activation-induced cell-death – (ACID)-Moleküls die Apoptose verhindern kann
(32).
Studien von Paik et al. an Gewebsproben zeigten, dass NK/T-Zell Lymphome des
oberen aerodigestiven Trakts, die eine niedrige miR-146a-Expression aufweisen eine
schlechtere Prognose haben als solche mit hoher miR-146a-Expression. In vitro-

50
Analysen mit NK/T-Zelllinien ließen zudem erkennen, dass eine miR-146a-Über-
expression die Zellproliferation hemmt, die Apoptose induziert und die Chemo-
sensitivität verstärkt, indem durch gezielte TRAF6-Modulation der NFkappaB-Weg
inhibiert wird (115).
Auch ist eine geringe Expression von miR-146a ein hervorstechendes Merkmal von
ALK-positiven anaplastischen großzelligen Lymphomen (ALK(+)ALCL) und zudem ein
Kriterium (von 11), mit dem sich ALK-negative ALCL von PTCL-NOS und AITL gezielt
abgrenzen lassen (94). Desweiteren konnten Tomita et al. zeigen, dass das humane
lymphotrope Virus 1 (HTLV1) in T-Zell-Linien miR-146a NFkappaB-abhängig hoch-
reguliert und somit potentiell ein therapeutisches Ziel in adulten T-Zell-Leukämien
darstellt (155).
In AITL wären folgende Ursachen für eine erhöhte miR-146a-Expression denkbar: A)
die Anzahl der T-Zellen in den Lymphknotenproben der angioimmunoblastischen T-
Zell-Lymphome ist durchschnittlich höher als in denen der reaktiven Lymphadenitis,
sodass die microRNA-Signatur normaler CD3+T-Zellen inklusive miR-146a als
überdurchschnittlich wahrgenommen wird. Durch sorgfältige Fall- und Probenauswahl
sowie Eingrenzung der Kontrollgruppe auf Lymphadenitiden mit T-Zonen-Hyperplasie
wurde jedoch versucht, dieser Variablen entgegenzusteuern. B) Ähnlich wie in rheuma-
toider Arthritis könnte die Expression von miR-146a durch pro-inflammatorische
Zytokine (z.B. IL-6) induziert worden sein und somit das entzündliche Mikromilieu im
Lymphknoten widerspiegeln. Zu Bedenken ist jedoch, dass das hier verwendete nicht-
neoplastische Vergleichsgewebe so gewählt wurde, dass ein ähnliches entzündliches
Zellbild wie in AITL vorherherrscht. C) Die EBV-Last der B-Blasten in AITL könnte eine
messbar erhöhte miR-146a-Expression in der Gesamtgewebsprobe bewirkt haben.
Jedoch zeigt die Subgruppenanalyse, dass es keinen signifikanten Unterschied
zwischen EBER-positiven und EBER-negativen Fällen im Hinblick auf die microRNA-
Expression gibt. Auch wenn die Adjustierung des p-Wert vernachlässigt wird, ist keine
Tendenz für einen möglichen Zusammenhang statistisch belegbar. D) Eine vermehrte
Aktivierung des NFkappaB-Weges aus tumoreigener Ursache könnte zur vermehrten
Transkription des miR-146a-Gens führen. Gleichzeitig müsste man davon ausgehen,
dass der negative Rückkopplungsmechanismus über TRAF6 und IRAK defekt ist. E)
Eine Trisomie von Chromosom 5, deren häufiges Vorkommen in AITL zytogenetische
Untersuchungen belegt haben (149), könnte aufgrund des Gen-Dosis-Effekts zu einer
verstärkten Expression der microRNA führen. Vergleichsweise konnten in Lympho-
zyten von Kindern mit Down-Syndrom (Trisomie 21) dysregulierte microRNAs mit Gen-
locus auf Chromosom 21 nachgewiesen werden, die möglicherweise zu den Immun-

51
defekten in dieser Erkrankung beitragen können (166). Tatsächlich ist miR-146a eine
von zwei microRNAs (neben miR-15a/16) deren Herunterregulation direkt auf eine
chromosomale Aberration zurückgeführt werden kann (116): Für myelodysplastische
Syndrome, die mit einer 5q-Deletion einhergehen, ist eine verminderte Expression von
miR-146a belegt worden (146).
MiR-34a wird ganz überwiegend als Tumorsuppressor-microRNA beschrieben. Sie
wird auf Chromosom 1p36 codiert und gehört zu einer Familie von microRNAs (miR-
34a, -b und –c), die ein direktes Ziel des Tumorsuppressors p53 darstellen (59). Ihre
Hochregulation kann die Apoptose und den Zell-Zyklus-Arrest induzieren (25, 57).
MiR-34a hat unter anderem SIRT-1 zum Ziel, eine NAD-abhängige Deacetylase, die
viele pro-apoptotische Proteine hemmt (18). Durch Repression von SIRT1 wird miR-
34a Teil eines positiven Rückkopplungsweges, der die weitere p53-Aktivierung zur
Folge hat (59, 167). Interessant ist zudem, dass Kauppinen et al. berichten, dass auch
der NFkappaB-Signalweg durch miR-34a die SIRT1-Aktivität herunterregulieren kann.
So können NFkappaB-vermittelte Entzündungsreaktionen stimuliert und der Energie-
stoffwechsel verändert werden (72). Außerdem scheint eine hohe Expression von miR-
34a (durch Hemmung der Diacylglycerolkinase ζ) die TCR-abhängige T-Zell-Akti-
vierung zu verstärken (142). Auch verbindet miR-34a das p53-Netzwerk mit bcl2,
ebenfalls einem anti-apoptotischen Faktor, der eine Ziel-Struktur von miR-34a darstellt
(58). In Tumorzellen kann die miR-34-vermittelte Apoptose durch Inaktivierung von p53
und/oder der miR-34-Gene supprimiert sein (59).
Niedrige miR-34a-Level sind zum Beispiel ein Charakteristikum von MYC-induzierten
Lymphomen, z.B. Burkitt-Lymphomen, da MYC nachweislich die miR-34a-Expression
supprimieren kann (145). He et al. zeigten zudem, dass die miR-34a-Expression in
MALT-Lymphomen und diffusen großzelligen B-Zell-Lymphomen des Magens im
Vergleich zu normalem Magengewebe und mononukleären Blutzellen vermindert ist.
Die verminderte miR-34a-Expression, verbunden u.a. mit einer hohen bcl2-Expression,
war zudem mit einem schlechten Gesamtüberleben in diesen Entitäten verbunden (58).
Allerdings ist in der microRNA-Signatur von splenischen Marginalzonenlymphomen
miR-34a eine der abweichend am höchsten exprimierten microRNAs (7).
Korrespondierend dazu liegen Hinweise dafür vor, dass die Funktion von miR-34a
stark vom zellulären Kontext, der Expression ihrer Ziel-Moleküle und Stoffwechsel-
prozessen abhängt (59). Chang et al. konnten außerdem zeigen, dass die miR-34a-
induzierte Apoptose zumindest teilweise die Präsenz des p53-Wildtyps voraussetzt
(25).

52
Dass hier die als Tumorsuppressor bekannte miR-34a im Tumor stärker exprimiert wird
als im nicht-neoplastischen Kontrapart könnte dafür sprechen, dass die pro-apop-
totischen Funktionen der microRNA hier nicht im Fokus stehen oder aber nicht greifen
können. Ein Hinweis wäre hierfür zum Beispiel auch bcl2, eine postulierte Zielstruktur
von miR-34a, die bekanntermaßen in AITL ebenfalls überexprimiert wird und als
Oncogen gilt (66, 71). Allerdings war die bcl2-Expression nicht Gegenstand der vor-
liegenden Studie und ist für das Fallkollektiv nicht ermittelt worden. Eine vermehrte
Detektion von p53, ein Indikator für die Mutation des p53-Gens, konnte für periphere T-
Zell-Lymphome gezeigt und mit einer schlechteren Prognose in Verbindung gebracht
werden (71). Vielleicht stellt somit miR-34a das Bindeglied zwischen bcl2 und p53
sowie den Grund der Deregulation multipler Apoptosewege in peripheren T-Zell-
Lymphomen dar (vgl. hierzu auch (71)).
Zumindest findet sich auch in ALK-negativen und –positiven anaplastischen groß-
zelligen Lymphomen eine erhöhte miR-34a-Expression als Teil einer 7 microRNAs
umfassenden Schnittmenge zwischen diesen beiden Entitäten (94).
Andererseits könnte wiederum im Rahmen des entzündlichen Hintergrundes in AITL,
möglicherweise auch NFkappaB-vermittelt, die Expression induziert worden sein.
So haben Forte et al. gezeigt, dass miR-34a stark p53-unabhängig über NFkappaB-
Aktivierung von dem EBV- latent membran protein 1 induziert werden kann und dass in
vielen EBV- und Kaposi-Sarkom-assoziierten-Virus (KSHV)-infizierten B-Zell-
Lymphom-Zell-Linien miR-34a eine wachstumsvermittelnde, onkogene Rolle einnimmt
(45).
MiR-193b wird auf Chromosom 16p11.13 codiert (86).
Die in-silico-Recherche in der Datenbank pubmed ergab insgesamt nur 59 Treffer im
Vergleich zu fast 600 Treffern für jeweils miR-146a und miR-34a (Stand Juni 2014).
Sehr viele der veröffentlichten Studien beziehen sich auf Experimente mit Tumorzell-
Linien, was nicht automatisch bedeutet, dass das ermittelte Ergebnis der Wirklichkeit in
vivo entspricht. Liu C et al. stellten zum Beispiel fest, dass microRNA-Experimente
anhand von Zelllinien und Fällen von ALCL oft nicht positiv korrelieren, was vermuten
lässt, dass die Regulation von microRNAs in vivo sehr komplex ist, dass verschiedene
Stromakomponenten die Variabilität des microRNA-Profils beeinflussen und es
spezifische Unterschiede zwischen den Zelllinien gibt (94)
In vitro konnte miR-193b als möglicher Tumorsuppressor in Prostatakarzinomen (132),
in akuter myeloischer Leukämie (AML) (42), in Pankreas-, Magen und Ovarial-
karzinomzellen (63, 109, 178) identifiziert werden. Ebenfalls in vitro zeigten Lenarduzzi
et al., dass miR-193b in Zell-Linien von Plattenepithelkarzinomen des Kopfes und

53
Halses überexprimiert ist (86). Als eine Zielstruktur von miR-193b konnten sie Neuro-
fibromin1 (NF1), eine GTPase, experimentell sichern. Sie postulierten, dass Tumore,
die miR-193b überexprimieren und damit NF1 herunterregulieren, konstitutiv den Ras-
Signalweg aktivieren können. In T-Zellen ist die T-Zell-Rezeptor- induzierte Aktivierung
des Ras-Signalweges wichtig für die Entwicklung, Proliferation und die Differenzierung
(79). Abnormale Ras-Expression, -Aktivierung oder –Signale in T-Zellen spielen eine
wichtige Rolle für die Entwicklung von Autoimmunerkrankungen (105), was miR-193b
im Hinblick auf das klinische Erscheinungsbild in AITL, das ebenfalls durch
Autoimmun-Effekte bestimmt wird, in den Fokus rücken könnte.
Analysen an Paraffin-Material von Nicht-kleinzelligen Lungenkarzinomen zeigten, dass
miR-193b hier ebenfalls niedrigere Level aufweist als im verglichenen Normalgewebe
(60). Thorns et al. fanden dagegen heraus, dass miR-193b im Gewebe neuro-
endokriner Neoplasien des Pankreas (PNEN) im Vergleich zu pankreatischen Insel-
zellen sowie im Serum dieser Patienten erhöht ist und bezeichnen miR-193b als viel-
versprechenden Biomarker (153). Ein interessanter Punkt ist zudem, dass miR-193b
Stathmin zum Ziel haben kann. Über Herunterregulation von Stathmin durch miR-193b
in Melanom-Zelllinien konnte die Migration und die Proliferation der Krebszellen
gehemmt werden (26). In unseren Fällen angioimmunoblastischer T-Zell-Lymphome
konnte eine Expression von Stathmin in etwa 70% der Fälle auf den neoplastischen T-
Zellen nachgewiesen werden. Ein signifikanter Unterschied von Stathmin-positiven zu
–negativen Fällen konnte in der Subgruppenanalyse bezüglich des microRNA-Profils
jedoch nicht ermittelt werden. Experimente, die den für Krebs-Zelllinien beschriebenen
Zusammenhang zwischen miR-193b und Stathmin in vivo verifizieren, wären hierzu
wünschenswert.
Eine Überexpression von miR-193b in angioimmunoblastischen T-Zell-Lymphomen
bringt somit den RAS-Signalweg und Stathmin als mögliche deregulierte Mechanismen
in dieser Erkrankung ins Spiel.

54
4.3.2. Mögliche Tumor suppressor-miR in AITL
In vielen Neoplasien wurde eine verminderte Expression von spezifischen microRNAs
gefunden. Diese microRNAs kommen als mögliche Tumorsuppressoren in Betracht
(116).
Grundsätzlich erscheint ein Funktionsverlust (loss-of-function) von Genen in der Onko-
genese weniger schwerwiegend als ein Funktionshinzugewinn (gain-of-function).
Ebenso gilt der Funktionsverlust als stärker altersabhängig (116). Wie microRNAs
herunterreguliert werden, konnte bisher nur für wenige spezifische Krebserkrankungen
belegt werden (s.o. 5q-Deletion in MDS).
Die vorliegende Studie ergab, dass die Expression von vier microRNAs, nämlich miR-
30b, miR-140-3p, let-7g und miR-664, in den Fällen angioimmunoblastischer T-Zell-
Lymphome signifikant niedriger ist, als in den verglichenen Proben von reaktiver
Lymphadenitis.
Was ist zu diesen microRNAs bereits bekannt, was im Hinblick auf angioimmuno-
blastische T-Zell-Lymphome relevant sein könnte?
Im Überblick gibt es für die vier zu diskutierenden microRNAs vergleichsweise sehr viel
weniger publiziertes Material als für die bereits beschriebenen möglichen Oncomirs.
Am interessantesten erscheint miR-30b . Diese microRNA wird auf Chromosom 8
codiert (www.mirbase.org). Insbesondere in kolorektalen Karzinomen wurde miR-30b
sowohl in vitro als auch in vivo als möglicher Tumorsuppressor identifiziert. Liao et al.
konnten zeigen, dass ihre Expression im Karzinom signifikant geringer ist, als in
vergleichbarem Normalgewebe. Auch war ein niedrigeres miR-30b-Level eng mit einer
schlechteren Differenzierung, einem höheren TNM-Stadium und einer schlechteren
Prognose des Tumors verknüpft (91). Auch zeigte sich miR-30b in Lebermetastasen
kolorektaler Karzinome im Vergleich zu den Primärtumoren herunterreguliert (175).
Hier allerdings bleibt die mutmaßliche Gewebsspezifität des microRNA-Profils zu
berücksichtigen. Die Überexpression von miR-30b in vitro wirkte sich zudem hemmend
auf die Zellmigration und Invasion aus und induzierte die Apoptose der
Kolonkarzinomzellen (91, 175).
Bridge et al. belegten experimentell, dass die Expression von Delta-like 4 (DLL4),
einem membranständigen Liganden des Notch-Signalweges, von miR-30b post-
transkriptionell gehemmt werden kann (17). DLL4 spielt eine wichtige Rolle in der
pathologischen Angiogenese und ist in Tumoren überexprimiert, oft in Assoziation mit
Markern der Entzündung und der Angiogenese (120, 121). In Kaposi-Sarkom-Herpes-
Virus-infizierten lymphatischen Endothelzellen wurde die simultane Hochregulation von

55
DLL4 und Suppression von miR-30b beschrieben (17). Weitere Beobachtungen der
Gruppe um Bridge enthüllten, dass DLL4 und miR-30 insbesondere in Tumortypen mit
mittelhoher und hoher Vaskularität negativ korreliert sind und dass die miR-30-Familie
indirekt in vivo den VEGFA-Signalweg kontrollieren kann (17).
Die Recherche bestätigt somit miR-30b auch in anderen Geweben als möglichen
Tumorsuppressor und stellt eine prognostische Relevanz der microRNA in Aussicht.
Außerdem weist die Suppression von miR-30b in den AITL-Fällen auf den angio-
genetischen Marker DLL4 hin, dessen Expression und Funktion in AITL im Hinblick auf
die erhöhte Neovaskularisation in befallenen Lymphknoten bedeutsam sein könnte.
Auch miR-140-3p kommt als tumorsupprimierender Faktor in AITL in Betracht, da ihre
Expression in diesen Lymphknoten signifikant im Vergleich zur nicht-neoplastischen
Kontrollgruppe herunterreguliert ist. Auch in Osteosarkom-, Kolon- (144), Mamma-
(174) und nicht kleinzelligem Lungenkarzinom-Zell-Linien (172) wurden erniedrigte
Level von miR-140-3p beobachtet. In Geweben von Basalzellkarzinomen und kutanen
Plattenepithelkarzinomen (137, 138) sowie nicht-kleinzelligen Bronchialkarzinomen
(172) fanden sich die Level von miR-140-3p ebenfalls erniedrigt. In Kaposi-Sarkomen
konnten O’Hara et al. dagegen zeigen, dass miR-140-Level progressiv mit dem Grad
der Transformation steigen und höhere, aggressivere Stadien der Erkrankung mit
höheren miR-140-Leveln assoziiert sind (112).
Als mögliche Ziele von miR-140 wurden in diesen Studien SOX2, ein Tran-
skriptionsfaktor in embryonalen Stammzellen, und Insulin-like Growth Factor Receptor
Typ I (IGF1R) identifiziert.
Interessant ist, dass diesen beiden Zielstrukturen, im Hinblick auf periphere T-Zell-
Lymphome, bisher nur in Zelllinien ALK-positiver anaplastischer großzelliger
Lymphome eine potentielle onkogene Rolle in Verbindung mit dem entitätseigenen
Fusionsprotein NPM-ALK zukommt (53, 141).
Die microRNA let-7g wird auf Chromosom 3 codiert (www.mirbase.org). In Publika-
tionen, gelistet in der online-Datenbank pubmed, wird sie ganz überwiegend als
möglicher Tumorsuppressor geführt. Eine verringerte Expression von let-7g ging
sowohl bei Brustkrebs-Patienten als auch bei Menschen mit Hepatozellulärem
Karzinom (HCC) mit einer höheren Metastasierungsrate und einem schlechteren
Gesamtüberleben einher (69, 129). Eine Hochregulation von let-7g in Magenkarzi-
nomen war mit einer besseren Chemosensitivität des Tumors assoziiert und macht so
diese microRNA zu einem potentiell nützlichen Biomarker mit dem man die individuelle
Krebstherapie steuern kann (73).

56
Wegweisend könnte auch das Verhalten von let-7g in nicht-kleinzelligem
Lungenkarzinom sein: In Adenokarzinomen der Lunge war let-7g insgesamt herunter-
reguliert, am stärksten jedoch in der Gruppe, die weder EGFR- noch K-RAS-
Mutationen zeigte, was auf die Korrelation von let-7g - und microRNAs generell – mit
dem Mutationsstatus in Malignomen aufmerksam werden lässt (33).
Zuletzt bleibt miR-664 zu erwähnen. Sie wird auf Chromosom 1 codiert
(www.mirbase.org) und tritt bisher in Onlinedatenbanken kaum in Erscheinung. Bisher
validierte Ziele der microRNA (vier an der Zahl) sind nur auf der Plattform
„miRWalk“ abrufbar und werden nahezu ausschließlich in einer einzigen Publikation
genannt, die Vorhofflimmern in Hunden und Menschen zum Thema hat (98). In
vereinzelten Studien konnte die Hochregulation von miR-664 mit einer schlechteren
Prognose im Hepatozellulären Karzinom und mit der Proliferation von Prolactinomen
der Hypophyse in Verbindung gebracht werden (28, 168). Wang et al. zeigten dagegen,
dass miR-664 in metastasierten papillären Schilddrüsenkarzinomen höher exprimiert
wird als in nicht-metastasierten Karzinomen gleicher Art (162).
Zu erwähnen ist, dass miR-664 die einzige der signifikant aberrant exprimierten
microRNAs ist, die mit Array B der RT-qPCR detektiert worden ist. Mit diesem Array
wurden vom Hersteller die erst kürzlich entdeckten microRNAs berücksichtigt (27). Das
heißt auch, dass auf diese microRNA in älteren Studien eventuell noch gar nicht
getestet werden konnte, da die microRNA noch nicht bekannt und ein entsprechender
Primer nicht verfügbar war. Weitere Untersuchungen, auch bereits auf ihr microRNA-
Profil hin analysierter Tumorentitäten, mit aktualisierten Primersets könnte den hier
entstandenen Diskussionsschwerpunkt auf microRNAs des Arrays A verschieben.
4.3.3. Vorsicht bei der Auswahl der endogenen Kontr olle und Normalisierungsmethode in AITL
RNU44 ist eine kleine nukleoläre, nicht protein-codierene RNA (snoRNA) mit etwa 70
Nukleotiden Länge. Zusammen mit den weiteren sn/snoRNAs RNU48, U47 und
RNU6B wird sie von Applied Biosystems als endogene Kontrolle für die Normalisierung
von Daten, erhoben mit TaqMan® MicroRNA Assays, mittels der ∆CT-Methode
empfohlen, da sie in zahlreichen Normalgeweben konstitutiv und wenig variabel
exprimiert wird (164).
Auch in der vorliegenden Studie wurde ihre Expression automatisch bei Verwendung
der TaqMan® Array Human MircoRNA A+B cards mitbestimmt.

57
Allerdings kam nach Beratungen mit Experten der Statistik bei der Auswertung nicht
die ∆CT-Methode, sondern die Quantilnormalisierung zum Einsatz und RNU44 ergab
sich als signifikant geringer exprimierte snoRNA in den AITL-Fällen im Vergleich zu der
nicht-neoplastischen Kontrollgruppe. Diese Beobachtung weist auf die generelle
Problematik der Verwendung von endogenen Kontrollen und die Normalisierungs-
methode in Zusammenhang mit microRNA Arrays hin.
Grundsätzlich sind alle Normalisierungsmethoden besser als gar keine (127). Eine
Normalisierung soll Abweichungen durch nicht-biologische Einflüsse reduzieren und ist
für Experimente mit multiplen Arrays notwendig (15).
Valleron et al. jedoch konnten zeigen, dass snoRNAs in benignen und malignen T-Zell-
Populationen unterschiedlich exprimiert werden und spezifische Signaturen sogar
prognostische Relevanz haben (156). Auch Gee et al. fanden heraus, dass ins-
besondere für niedrige Level von RNU44 in Plattenepithelkarzinomen des Kopfes und
Halses sowie Mammakarzinomen eine starke Korrelation mit einer schlechten
Prognose besteht (52). Sie beobachteten weiterhin, dass die Verwendung der
snoRNAs als Referenzgene in der microRNA-Expressionsanalyse Verzerrungen und
Tendenzen hervorrufen können, zeigen jedoch keine Alternativen auf (52). Anders Qin
et al., die die Entwicklung besserer Normalisierungsmethoden in der microRNA Array-
Analyse anmahnen und insbesondere die Vorteile der Quantilnormalisierung nennen
(130).
4.3.4. MicroRNA und Subgruppen
Die Analyse der a posteriori definierten und sich überschneidenden Subgruppen ist
schwierig zu interpretieren und bei der kleinen Fallzahl von hochexplorativem
Charakter, egal ob für multiples Testen adjustiert wurde oder nicht.
Wichtig erscheint festzuhalten, dass auf lokalem nach Holm adjustiertem Signifikanz-
level in der Subgruppenanalyse die Nullhypothese für keine microRNA zurückgewiesen
werden konnte. Dies betrifft sowohl die Subgruppen, die sich nur auf die neoplastische
T-Zell-Population beziehen (Stathmin-, PD1-, CD10-Positivität/Negativität), als auch
diejenigen, die Merkmale des Mikromilieus widerspiegeln (Anwesenheit EBV-positiver
B-Blasten, Proliferation der Gefäße, Expansion der FDC, histomorphologisches
Muster).
Letztendlich war das grobe Ziel der Subgruppenanalyse herauszufinden, ob die
microRNA-Expression in der Lymphknotenprobe sich einem oder mehreren dieser
Merkmale unterwirft. Die Erwartung war z.B., dass es einen signifikanten Unterschied
in der microRNA-Expression geben würde, in Abhängigkeit davon, ob in der Probe

58
EBV-positive B-Blasten vermehrt sind oder nicht, da einige microRNAs (z.B. miR-146a)
durch das Virus induziert werden können. Auch lag es nahe anzunehmen, dass sich
die microRNA-Level mit zunehmener Architekturstörung des Lymphknotens
(klassifiziert anhand des histomorphologischen Musters) verändern würden. Leider war
jedoch eine Klassifikation aufgrund der geringen Gruppengrößen und des inkompletten
Datensatzes nicht sinnvoll möglich.
Dass das Mikromilieu keine Auswirkung auf die microRNA-Expression haben soll, wie
es die statistische Auswertung hier möglicherweise suggeriert, ist schwer vorstellbar.
Um jedoch aus dem mircoRNA-Profil ähnlich wie aus dem mRNA-Profil der AITL, die
Signaturen der einzelnen Zell-Komponenten und Signalwege, die das Lymphom
charakterisieren, herauszufiltern, bedarf es größer angelegter Studien und sehr viel
mehr Wissen um die einzelnen microRNAs.
Ein nicht zu übersehender Punkt ist auch, dass sich die Kontroll-und Testgruppe in der
Alterszusammensetzung unterscheiden. Die Patienten der Kontrollgruppe sind im
Schnitt 32,9 Jahre jünger als die Patienten des AITL-Kollektivs (36,6 Jahre versus 69,5
Jahre).
Das Alter der Patienten ist bei Beurteilung von Lymphknotenvergrößerungen, die
letztendlich zu einer Lymphknotenbiopsie führen können, von entscheidender
Bedeutung. Bei Kindern sind die meisten Lymphadenopathien gutartig; wohingegen bei
Erwachsenen die Wahrscheinlichkeit für ein Malignom mit dem Alter steigt (54, 84).
Reaktive lymphatische Hyperplasien sind zudem bei älteren Menschen („elderly“)
aufgrund der Unterdrückung der humoralen Immunantwort im Alter ungewöhnlich (70).
In die vorliegende Studie wurden deswegen nur Lymphknoten
Erwachsener,„adulter“ und „älterer“ Personen, einbezogen (vgl. (99)).
Das Gruppenmatching erfolgte mit Fokus auf dem histologischen Bild. Als
definierendes Merkmal der Kontrollgruppe wurde die Diagnose Lymphadenitis mit T-
Zonen Hyperplasie gewählt, da deren histologisches Bild, Zellzusammensetzung und
Struktur, stark dem des angioimmunoblastischen T-Zell Lymphoms (Muster 1) ähnelt
und sogar ein Kontinuum nicht ausgeschlossen ist (46, 75). Die Probenauswahl
konzentrierte sich entsprechend auf die Krankheitsentität und Organspezifität
(Lymphknoten). In einem zweiten Schritt wurde dann die Verfügbarkeit, Qualität und
Repräsentanz des Materials evaluiert.
Somit ist letztendlich der Altersunterschied der Fall- und Kontrollgruppe das Resultat
und der Kompromiss aus Biologie und der Probenverfügbarkeit.
Natürlich sind auch die Ergebnisse der Studie vor dem Hintergrund des
Altersunterschieds kritisch zu bewerten. Die Gruppengröße war zu klein als dass eine

59
Subgruppenanalyse hinsichtlich des Alters Sinn gemacht hätte. Das Alter hätte
beginnend bei der Probenauswahl stärker angeglichen werden müssen. Dies war
jedoch aus o.g. Gründen nicht möglich.
Biologisch kann Altern als Zeit-abhängige Minderung der physiologischen Funktion
eines Organismus beschrieben werden (96). Multiple Faktoren sowohl auf dem
molekularen als auch zellulären Level tragen zu dem Verlust der physiologischen
Integrität mit dem Alter bei (z.B. Telomer-Verkürzung, epigenetische Veränderungen,
Signalwege, die den Alterungsprozess regulieren, verminderte Mitochondriale Funktion,
vermindertes regeneratives Potential) (93).
Das Altern betrifft alle Komponenten des Immunsystems auf unterschiedlichen Ebenen
(16)- auch microRNAs (55, 93). Allerdings ist das alternde Immunsystem nicht nur
durch die erwähnten zellulären und molekularen Veränderungen charakterisiert,
sondern auch von der Antigenexposition, die ein Individuum zeit seines Lebens erfährt
(93). Der Einfluss des Alterns auf das microRNA-Expressionsprofil, noch dazu unter
Berücksichtigung der Organspezifität, erscheint dadurch vielfältig.
Gerade im Hinblick auf Lymphknoten, die im Laufe des Lebens auf eine Vielzahl von
Antigenen und Allergenen (u.a. Bakterien, Viren, Chemikalien, Umweltgifte, Drogen,
Medikamente) reagieren (117) sowie aufgrund der Tatsache, dass wir immer nur eine
Momentaufnahme der Veränderungen untersuchen, muss zusätzlich bedacht werden,
dass durch das Altersmatching lediglich eine Art Pseudosicherheit erzeugt worden
wäre. So haben unterschiedliche medizinische Studien gezeigt, dass microRNAs
sensitiv sind für Umwelteinflüsse, wie Luftverschmutzung, Zigarettenrauch,
Nanopartikel und Chemikalien (zusammengefasst in (160)).
4.4. Limitationen der Arbeit und Ausblick
Die Erforschung angioimmunoblastischer T-Zell-Lymphome ist aufgrund ihrer Rarität
und Komplexität sowie der Heterogenität ihres Erscheinungsbildes anspruchsvoll.
Im Rahmen einer experimentellen Promotionsarbeit aus dem Fachbereich der Medizin
stößt man dadurch leicht an Grenzen. Zum Beispiel ist die Anzahl der untersuchten
Fälle mit insgesamt 40 paraffineingebetteten Lymphknotenproben eher klein, sodass
statistisch ermittelte Aussagen und die Übertragbarkeit auf die gesamte Lymphom-
entität nur unter Vorbehalt möglich sind. Ebenfalls wäre die Ergänzung und Korrelation
der erhobenen Daten mit weiteren klinischen Angaben insbesondere zum Verlauf der
Erkrankung wünschenswert gewesen. Auch im Hinblick auf die Auswahl der nicht-
neoplastischen Kontrollgewebe wäre hierdurch möglicherweise ein multifaktorielles

60
matching möglich geworden, was jedoch eventuell lediglich eine Art „Pseudo-
sicherheit“ erzeugt hätte.
Die in-silico Recherche in der Datenbank Pubmed ergibt bei Eintrag der Suchbegriffe
„AITL [AND] microRNA“ bisher (Stand Juni 2014) einen einzigen Treffer: Eine
Veröffentlichung von Liu et al. in der Zeitschrift „blood“ aus dem Jahr 2013 über die
microRNA-Signatur von anaplastischen großzelligen Lymphomen (ALCL) in Abgrenz-
ung zu weiteren PTCL-Subgruppen, die auch 9 AITL-Fälle berücksichtigt (94).
Legt man die Erkenntnisse von Liu et al. zugrunde, wurden bereits 5 der 7 microRNAs
(miR-146a, miR-34a, miR-193b, miR-30b, let-7g), deren Expression wir für AITL als
signifikant aberrant ermittelt haben, nicht nur als wichtige Bestandteile der CD3+T-Zell-
Signatur, sondern auch der anderer nodaler peripherer T-Zell-Lymphome beschrieben
(94). Desweiteren stellt eine sechste microRNA unseres Profils (miR-140-3p) den
komplementären Strang zu miR-140-5p dar, die als eine der am stärksten exprimierten
microRNAs in der Schnittmenge von normalen CD3+T-Zellen und ALK-negativen
ALCLs in der Studie von Liu et al. genannt wird. Hierzu ist interessant zu wissen, dass
viele microRNAs einer Familie dieselbe seed sequence einer Ziel-mRNA erkennen
können (116).
Natürlich sind die beiden Studien nur unter großem Vorbehalt vergleichbar. Und auch
die weiteren oben aufgeführten Diskussionspunkte bezüglich der microRNA-
Expression der vorliegenden Arbeit müssen unter dieser Prämisse gesehen werden.
Die Ursachen für Variabilität im Hinblick auf RT-qPCR-basierten Studien sind gut
bekannt und erstrecken sich vom Studien-Design über technische bis hin zu bio-
logischen Variablen (19). Auch die Daten-Analyse insbesondere die Normalisierung
der erhobenen Daten ist bisher nicht vereinheitlicht worden (19, 140). Becker et al.
meinen sogar, dass es kaum möglich ist, die wahre Menge einer microRNA exakt in
einer biologischen Probe mit den existierenden Methoden zu messen (12).
Mit Veröffentlichung der MIQE-Guidelines durch Bustin et al. 2009, die eine Checkliste
beinhalten, welche minimalen Informationen für die Publikation von Ergebnissen von
RT-qPCR-Experimenten notwenig sind, ist jedoch ein Leitfaden verfügbar, der die
Seriösität, Integrität und Transparenz der Resultate sichern soll (20).
Außerdem gilt die von uns und von Liu et al. verwendete Methode der quantitativen
RT-PCR (TaqMan) als Goldstandard im Hinblick auf die microRNA-Messung (140). Sie
wird zudem als Validierungsmethode von Mircoarray-Daten eingesetzt (140). Unter den
RT-qPCR-Methoden wurde für die vorliegende Arbeit - und auch von Liu et al. - die
stem-loop RT-qPCR basierend auf TaqMan Arrays (kommerziell erhältlich über Applied
Biosystems) verwendet, die die hohe Sensititivität der stem-loop RT-PCR mit der

61
Möglichkeit, die Expression einer großen Anzahl von microRNAs zu messen, in einem
einzigen Experiment kombiniert (27). Chen et al. konnten für diese Methode eine hohe
Reproduzierbarkeit und eine sehr viel höhere Sensitivität und Spezifität im Vergleich zu
Microarrays feststellen (27). Auch bei dem Vergleich von Reverse-Transkriptase-Reak-
tionen und qPCR-Arrays, die an verschiedenen Tagen durchgeführt wurden, zeigte
sich keine signifikante Variation der Ergebnisse (27). Vor diesem Hintergrund können
die beschriebenen Überschneidungen unserer Daten mit denen von Liu et al. vorsichtig
als ein Zeichen der Validität unserer Ergebnisse interpretiert werden.
Das bunte Zellbild in den Lymphknoten mit AITL, wo der Anteil der neoplastischen T-
Zellen schwankt und sehr gering sein kann, wirft zudem die Frage auf, wie ein
microRNA-Profil bei Heterogenität des Gewebes zu werten ist. Und ob die Mikro-
dissektion der neoplastischen T-Zellen aus dem Gewebe im Hinblick auf das Ver-
ständnis der Lymphomentstehung die bessere Wahl gewesen wäre. Gleichzeitig
müsste jedoch gerade in der Erforschung peripherer T-Zell-Lymphome, in denen das
Mikromilieu aus nicht-neoplastischem Begleitinfiltrat und Stroma in Wechselwirkung mit
den Tumorzellen als „emerging theme“ eine große Rolle zu spielen scheint (50),
besondere Berücksichtigung finden. Hier wären Klassifikationsverfahren, wie sie bei
Gen-Expressionsanalysen von AITL bereits zum Einsatz gekommen sind und die die
Signatur der verschiedenen Komponenten der Lymphomerkrankung erkennen konnten,
hilfreich (vgl. hierzu (66)).
An Grenzen stieß diese Arbeit auch bei der Suche nach eventuellen Zielen der
aberrant exprimierten microRNAs. Hier wurde die Recherche lediglich auf
Publikationen in der Datenbank Pubmed beschränkt. Natürlich wurden auch ver-
schiedene online- Plattformen wie „miRWalk“ konsultiert, die validierte und durch ver-
schiedene Algorithmen vorhergesagte Zielgene für microRNAs auflisten. Aufgrund der
Unmenge an möglichen Zielstrukturen, kamen wir jedoch schnell zu der Einsicht, dass
eine sinnvolle Untersuchung auf Zielstrukturen der microRNA nur in Korrelation mit
Genexpressionsanalysen und den damit verbundenen Klassifikationsverfahren möglich
ist. Eine zukunftsträchtige Methode wäre zum Beispiel die von Suzuki et al. kürzlich
entwickelte GSEA-FAME (GFA)-Analyse (Gene set enrichment analysis and functional
assignment of microRNAs via enrichment analysis) (147).
Ebenfalls von Interesse könnte die Kopplung von microRNA-Expressionsuntersuch-
ungen mit CGH-Analysen sein, um eventuellen zytogenetischen Veränderungen als
Ursache einer microRNA-Deregulation auf die Spur zu kommen.
Der Charakter der vorliegenden Studie ist demnach als hochexplorativ anzusehen.

62
5. Zusammenfassung
Ziel der vorliegenden explorativ angelegten Arbeit war die nähere Charakterisierung
des seltenen, aggressiven und nach wie vor prognostisch ungünstigen angioimmuno-
blastischen T-Zell-Lymphoms im Hinblick auf die Morphologie, den immunhisto-
chemischen Phänotyp der neoplastischen T-Zellen und das molekulare Profil.
Hierbei kamen sowohl praxisnahe Methoden, wie die immunhistochemische Gewebs-
färbung, als auch bisher nur in der Forschung angewendete Methoden, wie die quanti-
tative Real-time PCR für microRNA, zum Einsatz.
Entsprechend sind die gewonnenen Ergebnisse teils praxisnah, teils als zukunfts-
weisend für weitere Forschungsvorhaben anzusehen.
Der Schwerpunkt lag hier auf der erstmaligen Erstellung eines isolierten microRNA-
Profils der Lymphomerkrankung und dem Vergleich mit dem des nicht-neoplastischen
Kontraparts anhand von insgesamt vierzig formalinfixierten und in Paraffin-einge-
betteten Lymphknotenproben (davon 30 AITL-Fälle und 10 Fälle von Lymphadenitiden
mit T-Zonen Hyperplasie als nicht-neoplastische Kontrolle).
Unter Berücksichtigung der aufgezeigten Limitationen der Arbeit, konnten so miR-146a,
miR-34 und miR-193b als mögliche Oncomirs sowie miR-30b, miR-140-3p, let-7g und
miR-664 als mögliche tumorsuppressive microRNAs in AITL identifiziert werden. In
Abgleich mit Ergebnissen der Recherche in der Online-Datenbank pubmed, verweisen
diese aberrant exprimierten microRNAs insbesondere auf den NFkappaB- sowie den
Ras-Signalweg als mögliche deregulierte Wirkungspfade in der Erkrankung.
Ebenso machen sie auf die Rolle viraler Infektionen (namentlich EBV und KSHV) als
möglicherweise onkogene Faktoren in AITL aufmerksam, da einige der beschrieben
microRNAs von den Viren nachweislich induzierbar sind. Darüber hinaus rückt durch
die Untersuchungen auch VEGF in Verbindung mit dem angiogenetischen Marker
DLL4 in den Fokus für weitere Untersuchungen. Zusätzlich finden sich Parallelen und
Überschneidungen zu microRNA-Profilen der Autoimmunerkrankungen systemischer
Lupus erythmatodes und rheumatoider Arthritis sowie zu denen von weiteren
peripheren T-Zell-Lymphomen.
Die ebenfalls signifikante Herunterregulation von RNU44, einer vielfach verwendeten
endogenen Kontrolle in microRNA-Expressionsanalysen, regt zudem die Debatte um
die Gültigkeit ihrer Verwendung im Allgemeinen an und könnte auf einen Vorteil der
Quantilnormalisierung hinweisen.
Die hier vorgenommenen Analysen zur Darstellung eines Zusammenhangs zwischen
microRNA-Profil und verschiedenen histomorphologischen und immunhistochemischen

63
Charakteristika, sowohl der neoplastischen T-Zell-Population als auch des Mikromilieus,
erbrachten überraschenderweise keine signifikante Korrelation.
Dies warf die grundsätzliche Frage auf, ob heterogene Gewebsproben, die wie das
AITL im Lymphknoten mit einem bunten nicht-neoplastischen Begleitinfiltrat und einer
ausgeprägten Stromakomponente einhergehen, überhaupt für eine microRNA-
Expressionsanalyse geeignet sind. Eine Lösung dieser Problematik wäre hier die
Kopplung an Gen-Expressionsanalysen und die einhergehenden Klassifikations-
verfahren, um das Mikromilieu auch auf microRNA-Ebene, insbesondere in hetero-
genen Tumoren wie peripheren T-Zell-Lymphomen, nicht unberücksichtigt zu lassen.
Abschließend empfahl sich in unseren praxisnahen Analysen der Antikörper gegen
Stathmin als sinnvolle Ergänzung im Panel der immunhistochemischen Marker zur
Detektion und Charakterisierung der neoplastischen T-Zell-Population in AITL.

64
6. Literaturverzeichnis
1 (Hrsg.) RK-IHudGdeKiDeV: Krebs in Deutschland 2009/2010. 116-119, 2013.
2 Agostinelli C, Hartmann S, Klapper W, Korkolopoulou P, Righi S, Marafioti T,
Piccaluga PP, Patsouris E, Hansmann ML, Lennert K, and Pileri SA: Peripheral
T cell lymphomas with follicular T helper phenotype: a new basket or a distinct
entity? Revising Karl Lennert's personal archive. Histopathology 59: 679-691,
2011.
3 Agostinelli C, Paterson JC, Gupta R, Righi S, Sandri F, Piccaluga PP, Bacci F,
Sabattini E, Pileri SA, and Marafioti T: Detection of LIM domain only 2 (LMO2)
in normal human tissues and haematopoietic and non-haematopoietic tumours
using a newly developed rabbit monoclonal antibody. Histopathology 61: 33-46,
2012.
4 Agostini M and Knight RA: miR-34: from bench to bedside. Oncotarget 5: 872-
881, 2014.
5 Anagnostopoulos I HM, Fransilla K, Harris M, Harris NL, Jaffe ES, Han J, van
Krieken JM, Poppema S, Marafioti T, Franklin J, Sextro M, Diehl V, Stein H:
European Task Force on Lymphoma project on lymphocyte prdominant
Hodgkin disease: histologic and immunologic analysis of submitted cases
reveals 2 types of Hodgkin disease with a nodular growth pattern and abundant
lymphocytes. Blood: 1889-1899, 2000.
6 Armitage JO: The aggressive peripheral T-cell lymphomas: 2012 update on
diagnosis, risk stratification, and management. American journal of hematology
87: 511-519, 2012.
7 Arribas AJ, Gomez-Abad C, Sanchez-Beato M, Martinez N, Dilisio L, Casado F,
Cruz MA, Algara P, Piris MA, and Mollejo M: Splenic marginal zone lymphoma:
comprehensive analysis of gene expression and miRNA profiling. Modern
pathology : an official journal of the United States and Canadian Academy of
Pathology, Inc 26: 889-901, 2013.
8 Attygalle A, Al-Jehani R, Diss TC, Munson P, Liu H, Du MQ, Isaacson PG, and
Dogan A: Neoplastic T cells in angioimmunoblastic T-cell lymphoma express
CD10. Blood 99: 627-633, 2002.
9 Bartel DP: MicroRNAs: genomics, biogenesis, mechanism, and function. Cell
116: 281-297, 2004.
10 Bartel DP: MicroRNAs: target recognition and regulatory functions. Cell 136:
215-233, 2009.

65
11 Baumjohann D and Ansel KM: MicroRNA-mediated regulation of T helper cell
differentiation and plasticity. Nature reviews Immunology 13: 666-678, 2013.
12 Becker C, Hammerle-Fickinger A, Riedmaier I, and Pfaffl MW: mRNA and
microRNA quality control for RT-qPCR analysis. Methods (San Diego, Calif) 50:
237-243, 2010.
13 Belletti B and Baldassarre G: Stathmin: a protein with many tasks. New
biomarker and potential target in cancer. Expert opinion on therapeutic targets
15: 1249-1266, 2011.
14 Benes V and Castoldi M: Expression profiling of microRNA using real-time
quantitative PCR, how to use it and what is available. Methods (San Diego,
Calif) 50: 244-249, 2010.
15 Bolstad BM, Irizarry RA, Astrand M, and Speed TP: A comparison of
normalization methods for high density oligonucleotide array data based on
variance and bias. Bioinformatics (Oxford, England) 19: 185-193, 2003.
16 Boraschi D, Aguado MT, Dutel C, Goronzy J, Louis J, Grubeck-Loebenstein B,
Rappuoli R, and Del Giudice G: The gracefully aging immune system. Science
translational medicine 5: 185ps188, 2013.
17 Bridge G, Monteiro R, Henderson S, Emuss V, Lagos D, Georgopoulou D,
Patient R, and Boshoff C: The microRNA-30 family targets DLL4 to modulate
endothelial cell behavior during angiogenesis. Blood 120: 5063-5072, 2012.
18 Brooks CL and Gu W: How does SIRT1 affect metabolism, senescence and
cancer? Nature reviews Cancer 9: 123-128, 2009.
19 Bustin SA: Why the need for qPCR publication guidelines?--The case for MIQE.
Methods (San Diego, Calif) 50: 217-226, 2010.
20 Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R,
Nolan T, Pfaffl MW, Shipley GL, Vandesompele J, and Wittwer CT: The MIQE
Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR
Experiments. Clinical chemistry 55: 611-622, 2009.
21 Cairns RA, Iqbal J, Lemonnier F, Kucuk C, de Leval L, Jais JP, Parrens M,
Martin A, Xerri L, Brousset P, Chan LC, Chan WC, Gaulard P, and Mak TW:
IDH2 mutations are frequent in angioimmunoblastic T-cell lymphoma. Blood 119:
1901-1903, 2012.
22 Carbone A and Gloghini A: Follicular dendritic cell pattern in early lymphomas
involving follicles. Advances in anatomic pathology 21: 260-269, 2014.
23 Carbone A, Gloghini A, Cabras A, and Elia G: The Germinal centre-derived
lymphomas seen through their cellular microenvironment. British journal of
haematology 145: 468-480, 2009.

66
24 Castellano L and Stebbing J: Deep sequencing of small RNAs identifies
canonical and non-canonical miRNA and endogenous siRNAs in mammalian
somatic tissues. Nucleic acids research 41: 3339-3351, 2013.
25 Chang TC, Wentzel EA, Kent OA, Ramachandran K, Mullendore M, Lee KH,
Feldmann G, Yamakuchi M, Ferlito M, Lowenstein CJ, Arking DE, Beer MA,
Maitra A, and Mendell JT: Transactivation of miR-34a by p53 broadly influences
gene expression and promotes apoptosis. Molecular cell 26: 745-752, 2007.
26 Chen J, Abi-Daoud M, Wang A, Yang X, Zhang X, Feilotter HE, and Tron VA:
Stathmin 1 is a potential novel oncogene in melanoma. Oncogene 32: 1330-
1337, 2013.
27 Chen Y, Gelfond JA, McManus LM, and Shireman PK: Reproducibility of
quantitative RT-PCR array in miRNA expression profiling and comparison with
microarray analysis. BMC genomics 10: 407, 2009.
28 Chen YX, Li Q, Wang CD, Su ZP, Li WQ, Chen XB, and Wu ZB: [Differential
expression analysis of prolactinoma-related microRNAs]. Zhonghua yi xue za
zhi 92: 320-323, 2012.
29 Cho YU, Chi HS, Park CJ, Jang S, Seo EJ, and Huh J: Distinct features of
angioimmunoblastic T-cell lymphoma with bone marrow involvement. American
journal of clinical pathology 131: 640-646, 2009.
30 Couronne L, Bastard C, and Bernard OA: TET2 and DNMT3A mutations in
human T-cell lymphoma. The New England journal of medicine 366: 95-96,
2012.
31 Curmi PA, Gavet O, Charbaut E, Ozon S, Lachkar-Colmerauer S, Manceau V,
Siavoshian S, Maucuer A, and Sobel A: Stathmin and its phosphoprotein family:
general properties, biochemical and functional interaction with tubulin. Cell
structure and function 24: 345-357, 1999.
32 Curtale G, Citarella F, Carissimi C, Goldoni M, Carucci N, Fulci V, Franceschini
D, Meloni F, Barnaba V, and Macino G: An emerging player in the adaptive
immune response: microRNA-146a is a modulator of IL-2 expression and
activation-induced cell death in T lymphocytes. Blood 115: 265-273, 2010.
33 Dacic S, Kelly L, Shuai Y, and Nikiforova MN: miRNA expression profiling of
lung adenocarcinomas: correlation with mutational status. Modern pathology :
an official journal of the United States and Canadian Academy of Pathology, Inc
23: 1577-1582, 2010.
34 de Jong D, Xie W, Rosenwald A, Chhanabhai M, Gaulard P, Klapper W, Lee A,
Sander B, Thorns C, Campo E, Molina T, Hagenbeek A, Horning S, Lister A,
Raemaekers J, Salles G, Gascoyne RD, and Weller E: Immunohistochemical

67
prognostic markers in diffuse large B-cell lymphoma: validation of tissue
microarray as a prerequisite for broad clinical applications (a study from the
Lunenburg Lymphoma Biomarker Consortium). Journal of clinical pathology 62:
128-138, 2009.
35 de Leval L and Gaulard P: Pathobiology and molecular profiling of peripheral T-
cell lymphomas. Hematology / the Education Program of the American Society
of Hematology American Society of Hematology Education Program: 272-279,
2008.
36 de Leval L, Gisselbrecht C, and Gaulard P: Advances in the understanding and
management of angioimmunoblastic T-cell lymphoma. British journal of
haematology 148: 673-689, 2010.
37 de Leval L, Rickman DS, Thielen C, Reynies A, Huang YL, Delsol G, Lamant L,
Leroy K, Briere J, Molina T, Berger F, Gisselbrecht C, Xerri L, and Gaulard P:
The gene expression profile of nodal peripheral T-cell lymphoma demonstrates
a molecular link between angioimmunoblastic T-cell lymphoma (AITL) and
follicular helper T (TFH) cells. Blood 109: 4952-4963, 2007.
38 Delfau-Larue MH, de Leval L, Joly B, Plonquet A, Challine D, Parrens M,
Delmer A, Salles G, Morschhauser F, Delarue R, Brice P, Bouabdallah R,
Casasnovas O, Tilly H, Gaulard P, and Haioun C: Targeting intratumoral B cells
with rituximab in addition to CHOP in angioimmunoblastic T-cell lymphoma. A
clinicobiological study of the GELA. Haematologica 97: 1594-1602, 2012.
39 Dogan A, Attygalle AD, and Kyriakou C: Angioimmunoblastic T-cell lymphoma.
British journal of haematology 121: 681-691, 2003.
40 Dyhdalo KS, Lanigan C, Tubbs RR, and Cook JR: Immunoarchitectural patterns
of germinal center antigens including LMO2 assist in the differential diagnosis of
marginal zone lymphoma vs follicular lymphoma. American journal of clinical
pathology 140: 149-154, 2013.
41 Fabbri M, Croce CM, and Calin GA: MicroRNAs in the ontogeny of leukemias
and lymphomas. Leukemia & lymphoma 50: 160-170, 2009.
42 Fan LP, Huang HB, Shen JZ, Fu HY, and Zhou HR: [MicroRNA-193b
expression in newly diagnosed leukemia patients and its significance].
Zhongguo shi yan xue ye xue za zhi / Zhongguo bing li sheng li xue hui =
Journal of experimental hematology / Chinese Association of Pathophysiology
21: 843-846, 2013.
43 Federico M, Rudiger T, Bellei M, Nathwani BN, Luminari S, Coiffier B, Harris NL,
Jaffe ES, Pileri SA, Savage KJ, Weisenburger DD, Armitage JO, Mounier N, and
Vose JM: Clinicopathologic characteristics of angioimmunoblastic T-cell

68
lymphoma: analysis of the international peripheral T-cell lymphoma project.
Journal of clinical oncology : official journal of the American Society of Clinical
Oncology 31: 240-246, 2013.
44 Filbert EL, Le Borgne M, Lin J, Heuser JE, and Shaw AS: Stathmin regulates
microtubule dynamics and microtubule organizing center polarization in
activated T cells. Journal of immunology 188: 5421-5427, 2012.
45 Forte E, Salinas RE, Chang C, Zhou T, Linnstaedt SD, Gottwein E, Jacobs C,
Jima D, Li QJ, Dave SS, and Luftig MA: The Epstein-Barr virus (EBV)-induced
tumor suppressor microRNA MiR-34a is growth promoting in EBV-infected B
cells. Journal of virology 86: 6889-6898, 2012.
46 Frizzera G, Kaneko Y, and Sakurai M: Angioimmunoblastic lymphadenopathy
and related disorders: a retrospective look in search of definitions. Leukemia 3:
1-5, 1989.
47 Frizzera G, Moran EM, and Rappaport H: Angio-immunoblastic
lymphadenopathy with dysproteinaemia. Lancet 1: 1070-1073, 1974.
48 Gallamini A, Zaja F, Patti C, Billio A, Specchia MR, Tucci A, Levis A, Manna A,
Secondo V, Rigacci L, Pinto A, Iannitto E, Zoli V, Torchio P, Pileri S, and Tarella
C: Alemtuzumab (Campath-1H) and CHOP chemotherapy as first-line treatment
of peripheral T-cell lymphoma: results of a GITIL (Gruppo Italiano Terapie
Innovative nei Linfomi) prospective multicenter trial. Blood 110: 2316-2323, 2007.
49 Gaulard P and de Leval L: Follicular helper T cells: implications in neoplastic
hematopathology. Seminars in diagnostic pathology 28: 202-213, 2011.
50 Gaulard P and de Leval L: The microenvironment in T-cell lymphomas:
emerging themes. Seminars in cancer biology 24: 49-60, 2014.
51 Gaulard P and de Leval L: Pathology of peripheral T-cell lymphomas: where do
we stand? Seminars in hematology 51: 5-16, 2014.
52 Gee HE, Buffa FM, Camps C, Ramachandran A, Leek R, Taylor M, Patil M,
Sheldon H, Betts G, Homer J, West C, Ragoussis J, and Harris AL: The small-
nucleolar RNAs commonly used for microRNA normalisation correlate with
tumour pathology and prognosis. British journal of cancer 104: 1168-1177, 2011.
53 Gelebart P, Hegazy SA, Wang P, Bone KM, Anand M, Sharon D, Hitt M,
Pearson JD, Ingham RJ, Ma Y, and Lai R: Aberrant expression and biological
significance of Sox2, an embryonic stem cell transcriptional factor, in ALK-
positive anaplastic large cell lymphoma. Blood cancer journal 2: e82, 2012.
54 Habermann TM and Steensma DP: Lymphadenopathy. Mayo Clinic proceedings
75: 723-732, 2000.

69
55 Hackl M, Brunner S, Fortschegger K, Schreiner C, Micutkova L, Muck C,
Laschober GT, Lepperdinger G, Sampson N, Berger P, Herndler-Brandstetter D,
Wieser M, Kuhnel H, Strasser A, Rinnerthaler M, Breitenbach M, Mildner M,
Eckhart L, Tschachler E, Trost A, Bauer JW, Papak C, Trajanoski Z, Scheideler
M, Grillari-Voglauer R, Grubeck-Loebenstein B, Jansen-Durr P, and Grillari J:
miR-17, miR-19b, miR-20a, and miR-106a are down-regulated in human aging.
Aging cell 9: 291-296, 2010.
56 Harris NL, Jaffe ES, Stein H, Banks PM, Chan JK, Cleary ML, Delsol G, De
Wolf-Peeters C, Falini B, Gatter KC, and et al.: A revised European-American
classification of lymphoid neoplasms: a proposal from the International
Lymphoma Study Group. Blood 84: 1361-1392, 1994.
57 He L, He X, Lim LP, de Stanchina E, Xuan Z, Liang Y, Xue W, Zender L,
Magnus J, Ridzon D, Jackson AL, Linsley PS, Chen C, Lowe SW, Cleary MA,
and Hannon GJ: A microRNA component of the p53 tumour suppressor network.
Nature 447: 1130-1134, 2007.
58 He M, Gao L, Zhang S, Tao L, Wang J, Yang J, and Zhu M: Prognostic
significance of miR-34a and its target proteins of FOXP1, p53, and BCL2 in
gastric MALT lymphoma and DLBCL. Gastric cancer : official journal of the
International Gastric Cancer Association and the Japanese Gastric Cancer
Association 2013.
59 Hermeking H: The miR-34 family in cancer and apoptosis. Cell death and
differentiation 17: 193-199, 2010.
60 Hu H, Li S, Liu J, and Ni B: MicroRNA-193b modulates proliferation, migration,
and invasion of non-small cell lung cancer cells. Acta biochimica et biophysica
Sinica 44: 424-430, 2012.
61 Hussein K: [Pathobiology of the microRNA system]. Der Pathologe 33: 70-78,
2012.
62 Ikeda S and Tagawa H: Dysregulation of microRNAs and their association in the
pathogenesis of T-cell lymphoma/leukemias. International journal of hematology
99: 542-552, 2014.
63 Ikeda Y, Tanji E, Makino N, Kawata S, and Furukawa T: MicroRNAs associated
with mitogen-activated protein kinase in human pancreatic cancer. Molecular
cancer research : MCR 10: 259-269, 2012.
64 Ilyas M, Grabsch H, Ellis IO, Womack C, Brown R, Berney D, Fennell D, Salto-
Tellez M, Jenkins M, Landberg G, Byers R, Treanor D, Harrison D, Green AR,
Ball G, and Hamilton P: Guidelines and considerations for conducting
experiments using tissue microarrays. Histopathology 62: 827-839, 2013.

70
65 Iorio MV CC: MicroRNAs in cancer: small molecules with huge impact. Journal
of clinical oncology : official journal of the American Society of Clinical Oncology:
5848-5856, 2009.
66 Iqbal J, Weisenburger DD, Greiner TC, Vose JM, McKeithan T, Kucuk C, Geng
H, Deffenbacher K, Smith L, Dybkaer K, Nakamura S, Seto M, Delabie J, Berger
F, Loong F, Au WY, Ko YH, Sng I, Armitage JO, and Chan WC: Molecular
signatures to improve diagnosis in peripheral T-cell lymphoma and
prognostication in angioimmunoblastic T-cell lymphoma. Blood 115: 1026-1036,
2010.
67 Iqbal J, Wright G, Wang C, Rosenwald A, Gascoyne RD, Weisenburger DD,
Greiner TC, Smith L, Guo S, Wilcox RA, Teh BT, Lim ST, Tan SY, Rimsza LM,
Jaffe ES, Campo E, Martinez A, Delabie J, Braziel RM, Cook JR, Tubbs RR, Ott
G, Geissinger E, Gaulard P, Piccaluga PP, Pileri SA, Au WY, Nakamura S, Seto
M, Berger F, de Leval L, Connors JM, Armitage J, Vose J, Chan WC, and Staudt
LM: Gene expression signatures delineate biological and prognostic subgroups
in peripheral T-cell lymphoma. Blood 123: 2915-2923, 2014.
68 Jaffe ES: Pathobiology of peripheral T-cell lymphomas. Hematology / the
Education Program of the American Society of Hematology American Society of
Hematology Education Program: 317-322, 2006.
69 Ji J and Wang XW: A Yin-Yang balancing act of the lin28/let-7 link in
tumorigenesis. Journal of hepatology 53: 974-975, 2010.
70 Johnson JT and Myers EN: Evaluation of cervical masses in the elderly.
Geriatrics 38: 99-106, 1983.
71 Jung JT, Kim DH, Kwak EK, Kim JG, Park TI, Sohn SK, Do YR, Kwon KY, Song
HS, Park EH, and Lee KB: Clinical role of Bcl-2, Bax, or p53 overexpression in
peripheral T-cell lymphomas. Annals of hematology 85: 575-581, 2006.
72 Kauppinen A, Suuronen T, Ojala J, Kaarniranta K, and Salminen A: Antagonistic
crosstalk between NF-kappaB and SIRT1 in the regulation of inflammation and
metabolic disorders. Cellular signalling 25: 1939-1948, 2013.
73 Kim CH, Kim HK, Rettig RL, Kim J, Lee ET, Aprelikova O, Choi IJ, Munroe DJ,
and Green JE: miRNA signature associated with outcome of gastric cancer
patients following chemotherapy. BMC medical genomics 4: 79, 2011.
74 Kogo R, Mimori K, Tanaka F, Komune S, and Mori M: Clinical significance of
miR-146a in gastric cancer cases. Clinical cancer research : an official journal of
the American Association for Cancer Research 17: 4277-4284, 2011.
75 Kojima M, Nakamura S, Oyama T, Motoori T, Itoh H, Yoshida K, Suchi T, and
Masawa N: Autoimmune disease-associated lymphadenopathy with histological

71
appearance of T-zone dysplasia with hyperplastic follicles. A clinicopathological
analysis of nine cases. Pathology, research and practice 197: 237-244, 2001.
76 Kozomara A and Griffiths-Jones S: miRBase: annotating high confidence
microRNAs using deep sequencing data. Nucleic acids research 42: D68-73,
2014.
77 Krishnan C, Warnke RA, Arber DA, and Natkunam Y: PD-1 expression in T-cell
lymphomas and reactive lymphoid entities: potential overlap in staining patterns
between lymphoma and viral lymphadenitis. The American journal of surgical
pathology 34: 178-189, 2010.
78 Labbaye C and Testa U: The emerging role of MIR-146A in the control of
hematopoiesis, immune function and cancer. Journal of hematology & oncology
5: 13, 2012.
79 Lapinski PE and King PD: Regulation of Ras signal transduction during T cell
development and activation. American journal of clinical and experimental
immunology 1: 147-153, 2012.
80 Lau NC, Lim LP, Weinstein EG, and Bartel DP: An abundant class of tiny RNAs
with probable regulatory roles in Caenorhabditis elegans. Science (New York,
NY) 294: 858-862, 2001.
81 Lavon I, Zrihan D, Granit A, Einstein O, Fainstein N, Cohen MA, Cohen MA,
Zelikovitch B, Shoshan Y, Spektor S, Reubinoff BE, Felig Y, Gerlitz O, Ben-Hur
T, Smith Y, and Siegal T: Gliomas display a microRNA expression profile
reminiscent of neural precursor cells. Neuro-oncology 12: 422-433, 2010.
82 Lee RC and Ambros V: An extensive class of small RNAs in Caenorhabditis
elegans. Science (New York, NY) 294: 862-864, 2001.
83 Lee RC, Feinbaum RL, and Ambros V: The C. elegans heterochronic gene lin-4
encodes small RNAs with antisense complementarity to lin-14. Cell 75: 843-854,
1993.
84 Lee Y, Terry R, and Lukes RJ: Lymph node biopsy for diagnosis: a statistical
study. Journal of surgical oncology 14: 53-60, 1980.
85 Lemonnier F, Couronne L, Parrens M, Jais JP, Travert M, Lamant L, Tournillac
O, Rousset T, Fabiani B, Cairns RA, Mak T, Bastard C, Bernard OA, de Leval L,
and Gaulard P: Recurrent TET2 mutations in peripheral T-cell lymphomas
correlate with TFH-like features and adverse clinical parameters. Blood 120:
1466-1469, 2012.
86 Lenarduzzi M, Hui AB, Alajez NM, Shi W, Williams J, Yue S, O'Sullivan B, and
Liu FF: MicroRNA-193b enhances tumor progression via down regulation of
neurofibromin 1. PloS one 8: e53765, 2013.

72
87 Lennert K: [Nature, prognosis and nomenclature of angioimmunoblastic
(lymphadenopathy (lymphogranulomatosis X or T-zone lymphoma)]. Deutsche
medizinische Wochenschrift (1946) 104: 1246-1247, 1979.
88 Leung AK and Sharp PA: Quantifying Argonaute proteins in and out of GW/P-
bodies: implications in microRNA activities. Advances in experimental medicine
and biology 768: 165-182, 2013.
89 Li J, Wan Y, Guo Q, Zou L, Zhang J, Fang Y, Zhang J, Zhang J, Fu X, Liu H, Lu
L, and Wu Y: Altered microRNA expression profile with miR-146a upregulation in
CD4+ T cells from patients with rheumatoid arthritis. Arthritis research & therapy
12: R81, 2010.
90 Li L, Chen XP, and Li YJ: MicroRNA-146a and human disease. Scandinavian
journal of immunology 71: 227-231, 2010.
91 Liao WT, Ye YP, Zhang NJ, Li TT, Wang SY, Cui YM, Qi L, Wu P, Jiao HL, Xie
YJ, Zhang C, Wang JX, and Ding YQ: MicroRNA-30b functions as a tumour
suppressor in human colorectal cancer by targeting KRAS, PIK3CD and BCL2.
The Journal of pathology 232: 415-427, 2014.
92 Lin SL, Chiang A, Chang D, and Ying SY: Loss of mir-146a function in hormone-
refractory prostate cancer. RNA (New York, NY) 14: 417-424, 2008.
93 Linterman MA: How T follicular helper cells and the germinal centre response
change with age. Immunology and cell biology 92: 72-79, 2014.
94 Liu C, Iqbal J, Teruya-Feldstein J, Shen Y, Dabrowska MJ, Dybkaer K, Lim MS,
Piva R, Barreca A, Pellegrino E, Spaccarotella E, Lachel CM, Kucuk C, Jiang
CS, Hu X, Bhagavathi S, Greiner TC, Weisenburger DD, Aoun P, Perkins SL,
McKeithan TW, Inghirami G, and Chan WC: MicroRNA expression profiling
identifies molecular signatures associated with anaplastic large cell lymphoma.
Blood 122: 2083-2092, 2013.
95 Liu S, Yuan Z, Zhang C, Fu W, and Hou J: Bortezomib-based treatment for
relapsed and refractory angioimmunoblastic T-cell lymphoma: Case report and
literature review. Oncology letters 4: 262-264, 2012.
96 Lopez-Otin C, Blasco MA, Partridge L, Serrano M, and Kroemer G: The
hallmarks of aging. Cell 153: 1194-1217, 2013.
97 Lu LF, Boldin MP, Chaudhry A, Lin LL, Taganov KD, Hanada T, Yoshimura A,
Baltimore D, and Rudensky AY: Function of miR-146a in controlling Treg cell-
mediated regulation of Th1 responses. Cell 142: 914-929, 2010.
98 Lu Y, Zhang Y, Wang N, Pan Z, Gao X, Zhang F, Zhang Y, Shan H, Luo X, Bai
Y, Sun L, Song W, Xu C, Wang Z, and Yang B: MicroRNA-328 contributes to

73
adverse electrical remodeling in atrial fibrillation. Circulation 122: 2378-2387,
2010.
99 Luscieti P, Hubschmid T, Cottier H, Hess MW, and Sobin LH: Human lymph
node morphology as a function of age and site. Journal of clinical pathology 33:
454-461, 1980.
100 Maniataki E and Mourelatos Z: A human, ATP-independent, RISC assembly
machine fueled by pre-miRNA. Genes & development 19: 2979-2990, 2005.
101 Marafioti T, Copie-Bergman C, Calaminici M, Paterson JC, Shende VH, Liu H,
Baia M, Ramsay AD, Agostinelli C, Briere J, Clear A, Du MQ, Piccaluga PP,
Masir N, Nacheva EP, Sujobert P, Shanmugam K, Grogan TM, Brooks SP,
Khwaja A, Ardeshna K, Townsend W, Pileri SA, Haioun C, Linch D, Gribben JG,
Gaulard P, and Isaacson PG: Another look at follicular lymphoma:
immunophenotypic and molecular analyses identify distinct follicular lymphoma
subgroups. Histopathology 62: 860-875, 2013.
102 Marafioti T, Paterson JC, Ballabio E, Chott A, Natkunam Y, Rodriguez-Justo M,
Plonquet A, Rodriguez-Pinilla SM, Klapper W, Hansmann ML, Pileri SA,
Isaacson PG, Stein H, Piris MA, Mason DY, and Gaulard P: The inducible T-cell
co-stimulator molecule is expressed on subsets of T cells and is a new marker of
lymphomas of T follicular helper cell-derivation. Haematologica 95: 432-439,
2010.
103 Melhem RF, Zhu XX, Hailat N, Strahler JR, and Hanash SM: Characterization of
the gene for a proliferation-related phosphoprotein (oncoprotein 18) expressed
in high amounts in acute leukemia. The Journal of biological chemistry 266:
17747-17753, 1991.
104 Merchant SH, Amin MB, and Viswanatha DS: Morphologic and
immunophenotypic analysis of angioimmunoblastic T-cell lymphoma: Emphasis
on phenotypic aberrancies for early diagnosis. American journal of clinical
pathology 126: 29-38, 2006.
105 Mor A, Philips MR, and Pillinger MH: The role of Ras signaling in lupus T
lymphocytes: biology and pathogenesis. Clinical immunology (Orlando, Fla) 125:
215-223, 2007.
106 Mosalpuria K, Bociek RG, and Vose JM: Angioimmunoblastic T-cell lymphoma
management. Seminars in hematology 51: 52-58, 2014.
107 Motsch N, Pfuhl T, Mrazek J, Barth S, and Grasser FA: Epstein-Barr virus-
encoded latent membrane protein 1 (LMP1) induces the expression of the
cellular microRNA miR-146a. RNA biology 4: 131-137, 2007.

74
108 Mourad N, Mounier N, Briere J, Raffoux E, Delmer A, Feller A, Meijer CJ, Emile
JF, Bouabdallah R, Bosly A, Diebold J, Haioun C, Coiffier B, Gisselbrecht C, and
Gaulard P: Clinical, biologic, and pathologic features in 157 patients with
angioimmunoblastic T-cell lymphoma treated within the Groupe d'Etude des
Lymphomes de l'Adulte (GELA) trials. Blood 111: 4463-4470, 2008.
109 Nakano H, Yamada Y, Miyazawa T, and Yoshida T: Gain-of-function microRNA
screens identify miR-193a regulating proliferation and apoptosis in epithelial
ovarian cancer cells. International journal of oncology 42: 1875-1882, 2013.
110 Natkunam Y, Zhao S, Mason DY, Chen J, Taidi B, Jones M, Hammer AS,
Hamilton Dutoit S, Lossos IS, and Levy R: The oncoprotein LMO2 is expressed
in normal germinal-center B cells and in human B-cell lymphomas. Blood 109:
1636-1642, 2007.
111 Nicolae A, Pittaluga S, Venkataraman G, Vijnovich-Baron A, Xi L, Raffeld M, and
Jaffe ES: Peripheral T-cell lymphomas of follicular T-helper cell derivation with
Hodgkin/Reed-Sternberg cells of B-cell lineage: both EBV-positive and EBV-
negative variants exist. The American journal of surgical pathology 37: 816-826,
2013.
112 O'Hara AJ, Chugh P, Wang L, Netto EM, Luz E, Harrington WJ, Dezube BJ,
Damania B, and Dittmer DP: Pre-micro RNA signatures delineate stages of
endothelial cell transformation in Kaposi sarcoma. PLoS pathogens 5: e1000389,
2009.
113 Odejide O, Weigert O, Lane AA, Toscano D, Lunning MA, Kopp N, Kim S, van
Bodegom D, Bolla S, Schatz JH, Teruya-Feldstein J, Hochberg E, Louissaint A,
Dorfman D, Stevenson K, Rodig SJ, Piccaluga PP, Jacobsen E, Pileri SA, Harris
NL, Ferrero S, Inghirami G, Horwitz SM, and Weinstock DM: A targeted
mutational landscape of angioimmunoblastic T-cell lymphoma. Blood 123: 1293-
1296, 2014.
114 Pacifico F, Crescenzi E, Mellone S, Iannetti A, Porrino N, Liguoro D, Moscato F,
Grieco M, Formisano S, and Leonardi A: Nuclear factor-{kappa}B contributes to
anaplastic thyroid carcinomas through up-regulation of miR-146a. The Journal of
clinical endocrinology and metabolism 95: 1421-1430, 2010.
115 Paik JH, Jang JY, Jeon YK, Kim WY, Kim TM, Heo DS, and Kim CW:
MicroRNA-146a downregulates NFkappaB activity via targeting TRAF6 and
functions as a tumor suppressor having strong prognostic implications in NK/T
cell lymphoma. Clinical cancer research : an official journal of the American
Association for Cancer Research 17: 4761-4771, 2011.

75
116 Palanichamy JK and Rao DS: miRNA dysregulation in cancer: towards a
mechanistic understanding. Frontiers in genetics 5: 54, 2014.
117 Pangalis GA, Vassilakopoulos TP, Boussiotis VA, and Fessas P: Clinical
approach to lymphadenopathy. Seminars in oncology 20: 570-582, 1993.
118 Park HJ, Kim DH, Lim SH, Kim WJ, Youn J, Choi YS, and Choi JM: Insights into
the Role of Follicular Helper T Cells in Autoimmunity. Immune network 14: 21-29,
2014.
119 Patel JL, Pournazari P, Haggstrom SJ, Kosari F, Shabani-Rad MT, Natkunam Y,
and Mansoor A: LMO2 (LIM domain only 2) is expressed in a subset of acute
myeloid leukaemia and correlates with normal karyotype. Histopathology 64:
226-233, 2014.
120 Patel NS, Dobbie MS, Rochester M, Steers G, Poulsom R, Le Monnier K,
Cranston DW, Li JL, and Harris AL: Up-regulation of endothelial delta-like 4
expression correlates with vessel maturation in bladder cancer. Clinical cancer
research : an official journal of the American Association for Cancer Research
12: 4836-4844, 2006.
121 Patel NS, Li JL, Generali D, Poulsom R, Cranston DW, and Harris AL: Up-
regulation of delta-like 4 ligand in human tumor vasculature and the role of basal
expression in endothelial cell function. Cancer research 65: 8690-8697, 2005.
122 Peveling-Oberhag J, Crisman G, Schmidt A, Doring C, Lucioni M, Arcaini L,
Rattotti S, Hartmann S, Piiper A, Hofmann WP, Paulli M, Kuppers R, Zeuzem S,
and Hansmann ML: Dysregulation of global microRNA expression in splenic
marginal zone lymphoma and influence of chronic hepatitis C virus infection.
Leukemia 26: 1654-1662, 2012.
123 Piccaluga PP, Agostinelli C, Califano A, Carbone A, Fantoni L, Ferrari S,
Gazzola A, Gloghini A, Righi S, Rossi M, Tagliafico E, Zinzani PL, Zupo S,
Baccarani M, and Pileri SA: Gene expression analysis of angioimmunoblastic
lymphoma indicates derivation from T follicular helper cells and vascular
endothelial growth factor deregulation. Cancer research 67: 10703-10710, 2007.
124 Piccaluga PP, Tabanelli V, and Pileri SA: Molecular genetics of peripheral T-cell
lymphomas. International journal of hematology 99: 219-226, 2014.
125 Piekarz RL, Frye R, Prince HM, Kirschbaum MH, Zain J, Allen SL, Jaffe ES, Ling
A, Turner M, Peer CJ, Figg WD, Steinberg SM, Smith S, Joske D, Lewis I,
Hutchins L, Craig M, Fojo AT, Wright JJ, and Bates SE: Phase 2 trial of
romidepsin in patients with peripheral T-cell lymphoma. Blood 117: 5827-5834,
2011.

76
126 Pileri SA and Piccaluga PP: New molecular insights into peripheral T cell
lymphomas. The Journal of clinical investigation 122: 3448-3455, 2012.
127 Pradervand S, Weber J, Thomas J, Bueno M, Wirapati P, Lefort K, Dotto GP,
and Harshman K: Impact of normalization on miRNA microarray expression
profiling. RNA (New York, NY) 15: 493-501, 2009.
128 Punj V, Matta H, Schamus S, Tamewitz A, Anyang B, and Chaudhary PM:
Kaposi's sarcoma-associated herpesvirus-encoded viral FLICE inhibitory protein
(vFLIP) K13 suppresses CXCR4 expression by upregulating miR-146a.
Oncogene 29: 1835-1844, 2010.
129 Qian P, Zuo Z, Wu Z, Meng X, Li G, Wu Z, Zhang W, Tan S, Pandey V, Yao Y,
Wang P, Zhao L, Wang J, Wu Q, Song E, Lobie PE, Yin Z, and Zhu T: Pivotal
role of reduced let-7g expression in breast cancer invasion and metastasis.
Cancer research 71: 6463-6474, 2011.
130 Qin LX, Tuschl T, and Singer S: An Empirical Evaluation of Normalization
Methods for MicroRNA Arrays in a Liposarcoma Study. Cancer informatics 12:
83-101, 2013.
131 Ramasamy K, Lim Z, Pagliuca A, Salisbury JR, Mufti GJ, and Devereux S:
Successful treatment of refractory angioimmunoblastic T-cell lymphoma with
thalidomide and dexamethasone. Haematologica 91: ECR44, 2006.
132 Rauhala HE, Jalava SE, Isotalo J, Bracken H, Lehmusvaara S, Tammela TL,
Oja H, and Visakorpi T: miR-193b is an epigenetically regulated putative tumor
suppressor in prostate cancer. International journal of cancer Journal
international du cancer 127: 1363-1372, 2010.
133 Ree HJ, Kadin ME, Kikuchi M, Ko YH, Suzumiya J, and Go JH: Bcl-6 expression
in reactive follicular hyperplasia, follicular lymphoma, and angioimmunoblastic T-
cell lymphoma with hyperplastic germinal centers: heterogeneity of intrafollicular
T-cells and their altered distribution in the pathogenesis of angioimmunoblastic
T-cell lymphoma. Human pathology 30: 403-411, 1999.
134 Roehle A, Hoefig KP, Repsilber D, Thorns C, Ziepert M, Wesche KO, Thiere M,
Loeffler M, Klapper W, Pfreundschuh M, Matolcsy A, Bernd HW, Reiniger L,
Merz H, and Feller AC: MicroRNA signatures characterize diffuse large B-cell
lymphomas and follicular lymphomas. British journal of haematology 142: 732-
744, 2008.
135 Rudiger T, Weisenburger DD, Anderson JR, Armitage JO, Diebold J,
MacLennan KA, Nathwani BN, Ullrich F, and Muller-Hermelink HK: Peripheral T-
cell lymphoma (excluding anaplastic large-cell lymphoma): results from the Non-

77
Hodgkin's Lymphoma Classification Project. Annals of oncology : official journal
of the European Society for Medical Oncology / ESMO 13: 140-149, 2002.
136 S W: Enlargement of the lymphatic glands and spleen (or, Hodgkin's disease)
with remarks. Guy's Hosp Rep 11:56 1856.
137 Sand M, Skrygan M, Georgas D, Sand D, Hahn SA, Gambichler T, Altmeyer P,
and Bechara FG: Microarray analysis of microRNA expression in cutaneous
squamous cell carcinoma. Journal of dermatological science 68: 119-126, 2012.
138 Sand M, Skrygan M, Sand D, Georgas D, Hahn SA, Gambichler T, Altmeyer P,
and Bechara FG: Expression of microRNAs in basal cell carcinoma. The British
journal of dermatology 167: 847-855, 2012.
139 Sandhu SK, Croce CM, and Garzon R: Micro-RNA Expression and Function in
Lymphomas. Advances in hematology 2011: 347137, 2011.
140 Sato F, Tsuchiya S, Terasawa K, and Tsujimoto G: Intra-platform repeatability
and inter-platform comparability of microRNA microarray technology. PloS one 4:
e5540, 2009.
141 Shi P, Lai R, Lin Q, Iqbal AS, Young LC, Kwak LW, Ford RJ, and Amin HM: IGF-
IR tyrosine kinase interacts with NPM-ALK oncogene to induce survival of T-cell
ALK+ anaplastic large-cell lymphoma cells. Blood 114: 360-370, 2009.
142 Shin J, Xie D, and Zhong XP: MicroRNA-34a enhances T cell activation by
targeting diacylglycerol kinase zeta. PloS one 8: e77983, 2013.
143 So AY, Zhao JL, and Baltimore D: The Yin and Yang of microRNAs: leukemia
and immunity. Immunological reviews 253: 129-145, 2013.
144 Song B, Wang Y, Xi Y, Kudo K, Bruheim S, Botchkina GI, Gavin E, Wan Y,
Formentini A, Kornmann M, Fodstad O, and Ju J: Mechanism of
chemoresistance mediated by miR-140 in human osteosarcoma and colon
cancer cells. Oncogene 28: 4065-4074, 2009.
145 Sotillo E, Laver T, Mellert H, Schelter JM, Cleary MA, McMahon S, and Thomas-
Tikhonenko A: Myc overexpression brings out unexpected antiapoptotic effects
of miR-34a. Oncogene 30: 2587-2594, 2011.
146 Starczynowski DT, Kuchenbauer F, Argiropoulos B, Sung S, Morin R, Muranyi A,
Hirst M, Hogge D, Marra M, Wells RA, Buckstein R, Lam W, Humphries RK, and
Karsan A: Identification of miR-145 and miR-146a as mediators of the 5q-
syndrome phenotype. Nature medicine 16: 49-58, 2010.
147 Suzuki HI, Mihira H, Watabe T, Sugimoto K, and Miyazono K: Widespread
inference of weighted microRNA-mediated gene regulation in cancer
transcriptome analysis. Nucleic acids research 41: e62, 2013.

78
148 Swerdlow SH CE, Harris NL, Jaffe ES, Pileri SA, Stein H, Thiele J, Vardiman JW
(Editors), IARC: WHO classification of Tumours of Haematopoetic and Lymphoid
Tissues. 158-166, 2008.
149 Swerdlow SH CE, Harris NL, Jaffe ES, Pileri SA, Stein H, Thiele J, Vardiman JW
(Editors), IARC: WHO Classification of Tumours of Haematopoietic and
Lymphoid Tissues, Fourth Edition. 309-311, 2008.
150 Taganov KD, Boldin MP, Chang KJ, and Baltimore D: NF-kappaB-dependent
induction of microRNA miR-146, an inhibitor targeted to signaling proteins of
innate immune responses. Proceedings of the National Academy of Sciences of
the United States of America 103: 12481-12486, 2006.
151 Tagawa H, Ikeda S, and Sawada K: Role of microRNA in the pathogenesis of
malignant lymphoma. Cancer science 104: 801-809, 2013.
152 Tan LH, Tan SY, Tang T, Lim ST, Tan D, Lim LC, Kam GL, Loh TP, Tao M, and
Koay ES: Angioimmunoblastic T-cell lymphoma with hyperplastic germinal
centres (pattern 1) shows superior survival to patterns 2 and 3: a meta-analysis
of 56 cases. Histopathology 60: 570-585, 2012.
153 Thorns C, Schurmann C, Gebauer N, Wallaschofski H, Kumpers C, Bernard V,
Feller AC, Keck T, Habermann JK, Begum N, Lehnert H, and Brabant G: Global
microRNA profiling of pancreatic neuroendocrine neoplasias. Anticancer
research 34: 2249-2254, 2014.
154 Tokunaga T, Shimada K, Yamamoto K, Chihara D, Ichihashi T, Oshima R,
Tanimoto M, Iwasaki T, Isoda A, Sakai A, Kobayashi H, Kitamura K, Matsue K,
Taniwaki M, Tamashima S, Saburi Y, Masunari T, Naoe T, Nakamura S, and
Kinoshita T: Retrospective analysis of prognostic factors for angioimmunoblastic
T-cell lymphoma: a multicenter cooperative study in Japan. Blood 119: 2837-
2843, 2012.
155 Tomita M, Tanaka Y, and Mori N: MicroRNA miR-146a is induced by HTLV-1 tax
and increases the growth of HTLV-1-infected T-cells. International journal of
cancer Journal international du cancer 130: 2300-2309, 2012.
156 Valleron W, Ysebaert L, Berquet L, Fataccioli V, Quelen C, Martin A, Parrens M,
Lamant L, de Leval L, Gisselbrecht C, Gaulard P, and Brousset P: Small
nucleolar RNA expression profiling identifies potential prognostic markers in
peripheral T-cell lymphoma. Blood 120: 3997-4005, 2012.
157 Vasilescu C, Rossi S, Shimizu M, Tudor S, Veronese A, Ferracin M, Nicoloso
MS, Barbarotto E, Popa M, Stanciulea O, Fernandez MH, Tulbure D, Bueso-
Ramos CE, Negrini M, and Calin GA: MicroRNA fingerprints identify miR-150 as
a plasma prognostic marker in patients with sepsis. PloS one 4: e7405, 2009.

79
158 Verma NK, Dourlat J, Davies AM, Long A, Liu WQ, Garbay C, Kelleher D, and
Volkov Y: STAT3-stathmin interactions control microtubule dynamics in
migrating T-cells. The Journal of biological chemistry 284: 12349-12362, 2009.
159 Vose J, Armitage J, and Weisenburger D: International peripheral T-cell and
natural killer/T-cell lymphoma study: pathology findings and clinical outcomes.
Journal of clinical oncology : official journal of the American Society of Clinical
Oncology 26: 4124-4130, 2008.
160 Vrijens K, Bollati V, and Nawrot TS: MicroRNAs as Potential Signatures of
Environmental Exposure or Effect: A Systematic Review. Environmental health
perspectives 123: 399-411, 2015.
161 Wang X, Tang S, Le SY, Lu R, Rader JS, Meyers C, and Zheng ZM: Aberrant
expression of oncogenic and tumor-suppressive microRNAs in cervical cancer is
required for cancer cell growth. PloS one 3: e2557, 2008.
162 Wang Z, Zhang H, Zhang P, Li J, Shan Z, and Teng W: Upregulation of miR-
2861 and miR-451 expression in papillary thyroid carcinoma with lymph node
metastasis. Medical oncology 30: 577, 2013.
163 Willenbrock K, Renne C, Gaulard P, and Hansmann ML: In angioimmunoblastic
T-cell lymphoma, neoplastic T cells may be a minor cell population. A molecular
single-cell and immunohistochemical study. Virchows Archiv : an international
journal of pathology 446: 15-20, 2005.
164 Wong L LK, Russell I, Chen C: Endogenous controls for real-time quantitation of
miRNA using TaqMan® MicroRNA Assays. . Applied Biosystems Application
Note, Publication 127AP11-01, available at wwwappliedbiosystemscom 2007.
165 Xu B and Liu P: No survival improvement for patients with angioimmunoblastic
T-cell lymphoma over the past two decades: a population-based study of 1207
cases. PloS one 9: e92585, 2014.
166 Xu Y, Li W, Liu X, Chen H, Tan K, Chen Y, Tu Z, and Dai Y: Identification of
dysregulated microRNAs in lymphocytes from children with Down syndrome.
Gene 530: 278-286, 2013.
167 Yamakuchi M and Lowenstein CJ: MiR-34, SIRT1 and p53: the feedback loop.
Cell cycle (Georgetown, Tex) 8: 712-715, 2009.
168 Yang H, Cho ME, Li TW, Peng H, Ko KS, Mato JM, and Lu SC: MicroRNAs
regulate methionine adenosyltransferase 1A expression in hepatocellular
carcinoma. The Journal of clinical investigation 123: 285-298, 2013.
169 Yang JS and Lai EC: Alternative miRNA biogenesis pathways and the
interpretation of core miRNA pathway mutants. Molecular cell 43: 892-903, 2011.

80
170 Yang L, Boldin MP, Yu Y, Liu CS, Ea CK, Ramakrishnan P, Taganov KD, Zhao
JL, and Baltimore D: miR-146a controls the resolution of T cell responses in
mice. The Journal of experimental medicine 209: 1655-1670, 2012.
171 Yu H, Shahsafaei A, and Dorfman DM: Germinal-center T-helper-cell markers
PD-1 and CXCL13 are both expressed by neoplastic cells in angioimmunoblastic
T-cell lymphoma. American journal of clinical pathology 131: 33-41, 2009.
172 Yuan Y, Shen Y, Xue L, and Fan H: miR-140 suppresses tumor growth and
metastasis of non-small cell lung cancer by targeting insulin-like growth factor 1
receptor. PloS one 8: e73604, 2013.
173 Zhang B, Pan X, Cobb GP, and Anderson TA: microRNAs as oncogenes and
tumor suppressors. Developmental biology 302: 1-12, 2007.
174 Zhang Y, Eades G, Yao Y, Li Q, and Zhou Q: Estrogen receptor alpha signaling
regulates breast tumor-initiating cells by down-regulating miR-140 which targets
the transcription factor SOX2. The Journal of biological chemistry 287: 41514-
41522, 2012.
175 Zhao H, Xu Z, Qin H, Gao Z, and Gao L: miR-30b regulates migration and
invasion of human colorectal cancer via SIX1. The Biochemical journal 460: 117-
125, 2014.
176 Zhao S, Zhang L, Zhang M, Yao G, Zhang X, Zhao W, Ren W, Wang J, and
Zhang Q: Angioimmunoblastic T-cell lymphoma: the effect of initial treatment
and microvascular density in 31 patients. Medical oncology 29: 2311-2316, 2012.
177 Zhong H, Xu L, Zhong JH, Xiao F, Liu Q, Huang HH, and Chen FY: Clinical and
prognostic significance of miR-155 and miR-146a expression levels in formalin-
fixed/paraffin-embedded tissue of patients with diffuse large B-cell lymphoma.
Experimental and therapeutic medicine 3: 763-770, 2012.
178 Zhou H, Wang K, Hu Z, and Wen J: TGF-beta1 alters microRNA profile in
human gastric cancer cells. Chinese journal of cancer research = Chung-kuo
yen cheng yen chiu 25: 102-111, 2013.
179 Zhou Y, Attygalle AD, Chuang SS, Diss T, Ye H, Liu H, Hamoudi RA, Munson P,
Bacon CM, Dogan A, and Du MQ: Angioimmunoblastic T-cell lymphoma:
histological progression associates with EBV and HHV6B viral load. British
journal of haematology 138: 44-53, 2007.

81
7. Anhang
7.1. Material
7.1.1. Geräte
Beschreibung Name Hersteller
Abzüge Captair 5000NU Captair
Filtersysteme
Array-Verschlussapparat Low Density Array Sealer Applied Biosystems
Automat für
immunhistochemische
Färbungen und in-Situ-
Hybridisierung
Bond Max Automated
Immunhistochemistry and
in-situ-Hybridisation
System
Menarini
Brutschrank (verschiedene) Memmert
Computer (verschiedene) HP
Dampfgarer Multi Gourmet Braun
Feuchte Kammer k.A. CarlRoth
Gefrierschränke (verschiedene) Bosch, Liebherr
Heizplatten (verschiedene) Gerhardt, Medax,
Digsi-Therm
Kühlplatte Parafreezer Pathotec GmbH
Kühlschränke (verschiedene) Bosch, Liebherr
Magnetrührer Combimag-RCO Ikamag
Markierungsstift Dako Pen Dako
Mehrkanalpipetten Eppendorf Research®
Plus, verschiedene
Größen
Eppendorf
Mikroskop Axioskop Zeiss
Mikroliter-Pipetten Einkanalmikroliterpipetten
Eppendorf Research®
Eppendorf
Minischüttler Minishaker Sarstedt
Multifärber Leica ST5020 Leica
Objektträger (verschiedene) Leica
Paraffin-Ausgießstation TES 99.250 Medite
pH-Meter PH 537 WTW
Tischzentrifugen verschiedene Sarstedt, Beckman,
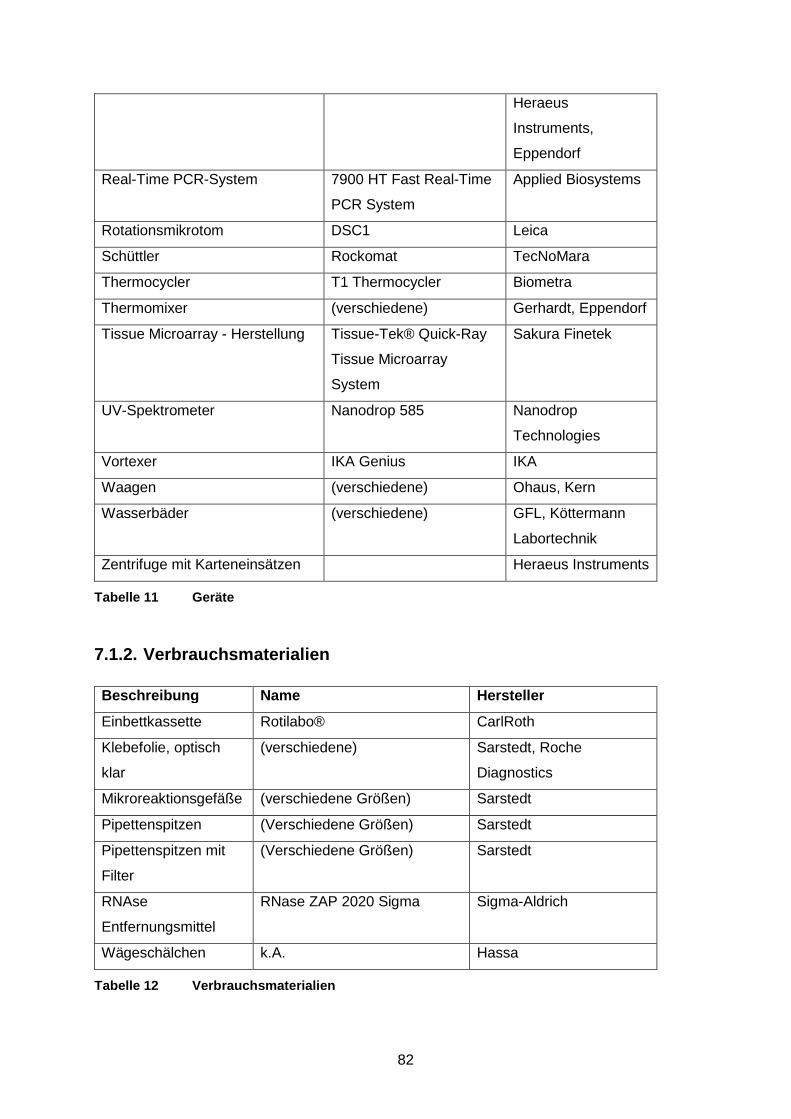
82
Heraeus
Instruments,
Eppendorf
Real-Time PCR-System 7900 HT Fast Real-Time
PCR System
Applied Biosystems
Rotationsmikrotom DSC1 Leica
Schüttler Rockomat TecNoMara
Thermocycler T1 Thermocycler Biometra
Thermomixer (verschiedene) Gerhardt, Eppendorf
Tissue Microarray - Herstellung Tissue-Tek® Quick-Ray
Tissue Microarray
System
Sakura Finetek
UV-Spektrometer Nanodrop 585 Nanodrop
Technologies
Vortexer IKA Genius IKA
Waagen (verschiedene) Ohaus, Kern
Wasserbäder (verschiedene) GFL, Köttermann
Labortechnik
Zentrifuge mit Karteneinsätzen Heraeus Instruments
Tabelle 11 Geräte
7.1.2. Verbrauchsmaterialien Beschreibung Name Hersteller
Einbettkassette Rotilabo® CarlRoth
Klebefolie, optisch
klar
(verschiedene) Sarstedt, Roche
Diagnostics
Mikroreaktionsgefäße (verschiedene Größen) Sarstedt
Pipettenspitzen (Verschiedene Größen) Sarstedt
Pipettenspitzen mit
Filter
(Verschiedene Größen) Sarstedt
RNAse
Entfernungsmittel
RNase ZAP 2020 Sigma Sigma-Aldrich
Wägeschälchen k.A. Hassa
Tabelle 12 Verbrauchsmaterialien

83
7.1.3. Reagenzien und Chemikalien Beschreibung Name Hersteller
Ammoniaklösung 25% Merck
Ammonium-Eisen-III-Sulfat Merck
Antigendemaskierungsreagenzien Novocastra™/Bond™
Epitope Retrieval
Solutions
Novocastra, Leica
Biosystems
Antikörperverdünnungslösung Antibody Diluent DAKO
Anti-Fluorescein-Antikörper Gebrauchsfertige Bond™ Anti-Fluor-escein Antibody 15ml
Leica Biosystems
Aqua dest (hauseigen)
Chromogen Bright DAB Chromogen ImmunoLogic
EBER-Detektionssonde Bond™ ISH Probe
EBER Probe
Leica Biosystems
Eindeckmittel Pertex Medite
Entparaffinierungslösung Novocastra Bond
Dewax Solution
Leica Biosystems
Eosin Eosin, yellowish Waldeck
Ethanol Leica, Büfa
Chemikalien
Formalin Formalin 4,5% und 37% Fluka, Riedl de Haen
Giemsa Stammlösung Giemsas Azur-Eosin -Mehtylenblau-Lösung
Merck
Goldchlorid Merck
Hämatoxylin Haemalaun, sauer nach
Mayer
Waldeck
Kaliumdisulfidlösung Waldeck
Kaliumpermanganat Merck
Kernechtrot Chroma
Methanol Merck
Natriumhydroxid Merck
Natriumthiosulfat Merck
Nukleasefreies Wasser DEPC, treated water Ambion
Oxalsäure Merck

84
Paraffin Medite Pure™ Paraffin Medite
Perjodsäure AnalaR Normapur VWR Chemicals
Proteinase K S 2019 DAKO
Puffer Tris Puffer pH 7,2-7,6 DAKO
Puffer Puffer pH 9,0 Novocastra
Schiffs Reagenz Merck
Silbernitrat Caelo
Waschpuffer Bond™ Waschlösung 10x
Leica Biosystems
Xylol 100% J.T. Baker, Büfa Chemikalien
Tabelle 13 Reagenzien und Chemikalien
7.1.4. Kits Name Hersteller
Bond™ Enzyme Pretreatment Kit Leica Biosystems
Bond™ Polymer Refine Detection Leica Biosystems
Bright DAB ImmunoLogic
Bright Vision + Poly- HRP- Anti Ms/Rb/Rt IgG Biotinfrei ImmunoLogic
Recover All™ Total Nucleic Acid Isolation Ambion
TaqMan® Array Human MircoRNA A+B cards v3.0 Applied Biosystems
TaqMan® Universal PCR Master Mix, No AmpErase®
UNG
Applied Biosystems
Tabelle 14 Kits
7.1.5. Antikörper Antikörper Klon Hersteller
CD3 SP7 Lab Vision
CD10 C56C6 Novocastra
CD20 L26 Lab Vision
CD21 2G9 Cellmarque
CD23 SP23 Thermo Scientific
LMO2 SP51 Cellmarque
PD1 MRQ-22 Cellmarque
Stathmin SP49 DCS
Tabelle 15 Antikörper

85
7.1.6. Primer Name Hersteller
Megaplex RT Primer, human pool A v2.1 Applied Biosystems
Megaplex RT Primer, human pool B v3.0 Applied Biosystems
Tabelle 16 Primer
7.1.7. Software Name Hersteller
Endnote V6..0.1 Thomson Reuters
Excel Microsoft
Nexus Pathologie NEXUS AG
R v3.0.2 r-project.org
RQ-Manager 1.2.1 Applied Biosystems
SDS Software v.2.2.2 Applied Biosystems
Thermocycler Management Software Biometra
Word Microsoft
Tabelle 17 Software
7.1.8. In silico Ressourcen Name URL Provider
mirbase http://www.mirbase.org/ University of Man-
chester, Faculty of Life
Science
miRWalk http://www.umm.uni-
heidelberg.de/apps/zmf/mirwalk/
Ruprecht-Karls-Univer-
sität Heidelberg, Medizi-
nische Fakultät Mann-
heim
Pub med http://www.ncbi.nlm.nih.gov/pubmed/ US National library of
Medicine National Insti-
tutes of Health
Tabelle 18 In-silico Resourchen
86
7.2. Ergänzende Tabellen und Abbildungen
Fall Alter Geschlecht Pattern Venolen CD23/21 (FDC)
1 65 m 2 2 1 2 68 m 2 1 1 3 79 w 2 1 3 4 45 m 2 1 2 5 69 m 3 2 2 6 69 m 3 2 3 7 80 w 2 3 1 8 72 m 3 3 NA 9 72 w 3 2 3
10 73 m 3 3 3 11 43 m 2 2 2 12 79 w 3 3 2 13 73 w 2 2 2 14 57 m 3 3 2 15 69 m 2 2 2 16 72 w 3 3 2 17 90 w 3 3 3 18 78 w 3 3 2 19 71 m 1 1 1 20 77 m 3 2 2 21 79 w 2 3 1 22 75 w 2 2 NA 23 51 w 1 2 1 24 72 m 2 2 1 25 52 w 3 3 3 26 82 w 2 3 3 27 67 m 2 1 2 28 63 m 3 1 3 29 68 m 1 1 1 30 75 m 2 2 2
Tabelle 19 Patienten-Charakteristika und Ergebnisse der Graduierungen
87
Fall-Nr. Stathmin CD10 LMO2 PD1 1 positiv positiv negativ positiv 2 positiv negativ negativ positiv 3 positiv negativ negativ negativ 4 positiv negativ negativ positiv 5 positiv negativ negativ positiv 6 positiv negativ negativ positiv 7 positiv negativ negativ positiv 8 positiv negativ negativ positiv 9 positiv negativ negativ positiv
10 positiv positiv negativ positiv 11 positiv negativ negativ negativ 12 negativ negativ negativ positiv 13 positiv negativ negativ positiv 14 positiv negativ negativ positiv 15 NA negativ NA NA 16 negativ positiv negativ negativ 17 positiv negativ negativ positiv 18 negativ negativ negativ positiv 19 negativ negativ negativ positiv 20 positiv negativ negativ positiv 21 negativ negativ negativ negativ 22 positiv negativ negativ positiv 23 negativ negativ negativ positiv 24 positiv negativ negativ positiv 25 positiv positiv negativ positiv 26 negativ negativ NA NA 27 positiv negativ negativ positiv 28 NA negativ NA NA 29 positiv negativ negativ positiv 30 negativ negativ NA positiv
Tabelle 20 Ergebnisse der immunhistochemischen Ausw ertung anhand des tissue microarrays
Kontrolle Nr. Alter Geschlecht
1 67 m 2 58 m 3 31 w 4 41 m 5 27 w 6 23 w 7 38 w 8 16 m 9 48 w
10 17 w
Tabelle 21 Charakteristika der Kontrollgruppe

88
Abbildung 20 TaqMan Human MicroRNA Array A übernommen von www.appliedbiosystems.com

89
Abbildung 21 TaqMan Human MicroRNA Array B übernommen von www.appliedbiosystems.com

90
Abbildung 22 Boxplot der CT-Werte pro Platte
Freundlicherweise zur Verfügung gestellt von Gola und Schillert
Tabelle 22 Ergebnisse der Präprozessierung
nach verschiedenen Schwellen der prozentualen Ausfallquote (missing fraction, MiF). Freundlicherweise zur Verfügung gestellt von Gola und Schillert

91
microRNA/Probe 1 2 3 K1 4 5 6 K2 7 8 RNU44-001094 37,096 30,981 45 27,147 28,387 32,965 31,94 25,787 32,778 28,283 hsa-miR-146a-000468 25,952 24,357 28,496 45 23,155 45 26,264 23,518 25,639 25,256 hsa-miR-34a-000426 31,683 29,247 30,46 29,208 27,193 29,591 29,202 30,064 31,417 30,931 hsa-miR-193b-002367 31,239 29,448 29,634 28,749 28,289 29,349 29,497 28,009 30,285 28,828 hsa-miR-30b-000602 34,187 31,584 39,349 28,765 29,681 45 32,569 27,431 31,588 29,815 hsa-let-7g-002282 35,282 31,452 35,31 28,315 29,769 34,046 34,112 26,699 31,095 29,499 hsa-miR-140-3p-002234 37,029 34,697 37,821 31,617 32,4 35,973 35,129 30,068 34,139 32,732 hsa-miR-664-002897 34,291 33,135 33,691 31,438 32,018 35,305 34,962 29,609 32,806 32,936
microRNA/Probe 9 K3 10 11 12 K4 13 14 15 K5 RNU44-001094 45 27,204 45 29,867 27,109 26,283 28,535 27,456 30,851 27,668 hsa-miR-146a-000468 30,345 24,18 32,048 25,969 23,012 23,871 25,579 23,948 24,502 25,449 hsa-miR-34a-000426 39,847 31,279 36,686 31,982 25,756 28,783 31,799 26,934 30,152 32,283 hsa-miR-193b-002367 31,286 29,88 33,217 30,121 25,737 27,82 29,433 27,372 29,706 29,9 hsa-miR-30b-000602 45 28,705 45 30,308 26,556 27,981 30,913 28,955 30,081 29,499 hsa-let-7g-002282 45 27,872 45 30,145 26,613 27,922 30,258 29,198 29,871 28,504 hsa-miR-140-3p-002234 45 31,054 45 33,491 29,963 30,239 33,003 31,494 33,046 31,785 hsa-miR-664-002897 35,324 32,642 34,142 30,91 29,036 29,245 30,841 29,619 31,04 31,457

92
microRNA/Probe 16 17 18 K6 19 20 21 K7 22 23 RNU44-001094 33,785 30,556 27,956 25,992 27,488 25,76 30,801 27,387 33,671 27,144 hsa-miR-146a-000468 26,008 25,939 22,977 23,594 24,245 23,498 26,757 24,913 26,338 22,265 hsa-miR-34a-000426 28,521 28,163 26,794 29,511 28,97 27,468 31,088 33,27 30,288 27,154 hsa-miR-193b-002367 28,727 28,846 27,215 28,353 28,494 26,73 29,363 28,716 28,629 27,035 hsa-miR-30b-000602 33,512 32,508 29,099 27,98 29,503 27,386 32,255 28,971 33,918 27,321 hsa-let-7g-002282 35,047 31,996 28,97 27,094 28,681 27,506 32,618 28,367 32,259 27,397 hsa-miR-140-3p-002234 34,663 35,611 31,576 30,268 32,137 30,899 36,093 31,085 38,335 30,971 hsa-miR-664-002897 31 45 29,795 29,488 29,302 29,229 32,236 30,464 33,121 29,252
microRNA/Probe 24 K8 25 26 27 K9 28 29 30 K10 RNU44-001094 26,164 28,353 30,662 28,314 28,042 27,517 27,954 45 32,115 28,248 hsa-miR-146a-000468 22,499 25,84 25,575 24,433 24,848 24,936 25,376 29,973 25,982 25,637 hsa-miR-34a-000426 28,867 33,125 29,972 30,718 29,829 30,126 32,753 35,538 30,856 33,763 hsa-miR-193b-002367 27,822 30,255 29,256 27,923 28,463 28,384 27,954 31,07 28,926 29,901 hsa-miR-30b-000602 28,528 30,092 32,279 29,716 29,569 28,529 30,228 45 33,396 30,109 hsa-let-7g-002282 28,142 28,957 31,067 29,801 29,196 27,714 29,735 36,448 33,211 29,092 hsa-miR-140-3p-002234 30,977 32,096 33,7 32,187 31,944 31,062 32,974 45 35,011 31,306 hsa-miR-664-002897 45 31,925 33,441 31,305 30,108 30,497 32,265 30,982 35,768 31,073
Tabelle 23 CT-Werte der Lymphom- und Referenz-Fälle
für die 8 verschiedenen aberrant exprimierten microRNAs und RNU44

93
Tabelle 24 Top 10 microRNAs für EBER
Nach p-Werten des Wilcoxon-Mann-Whitney-Tests, adjustiert mit der Bonferroni-Holm Prozedur inklusive der nicht-adjustierten 95%-Konfidenzintervalle der Differenz im Median Freundlicherweise zur Verfügung gestellt von Gola und Schillert
Tabelle 25 Top 10 microRNAs für Stathmin
Nach p-Werten des Wilcoxon-Mann-Whitney-Tests, adjustiert mit der
Bonferroni-Holm Prozedur inklusive der nicht-adjustierten 95%-
Konfidenzintervalle der Differenz im Median.
Freundlicherweise zur Verfügung gestellt von Gola und Schillert
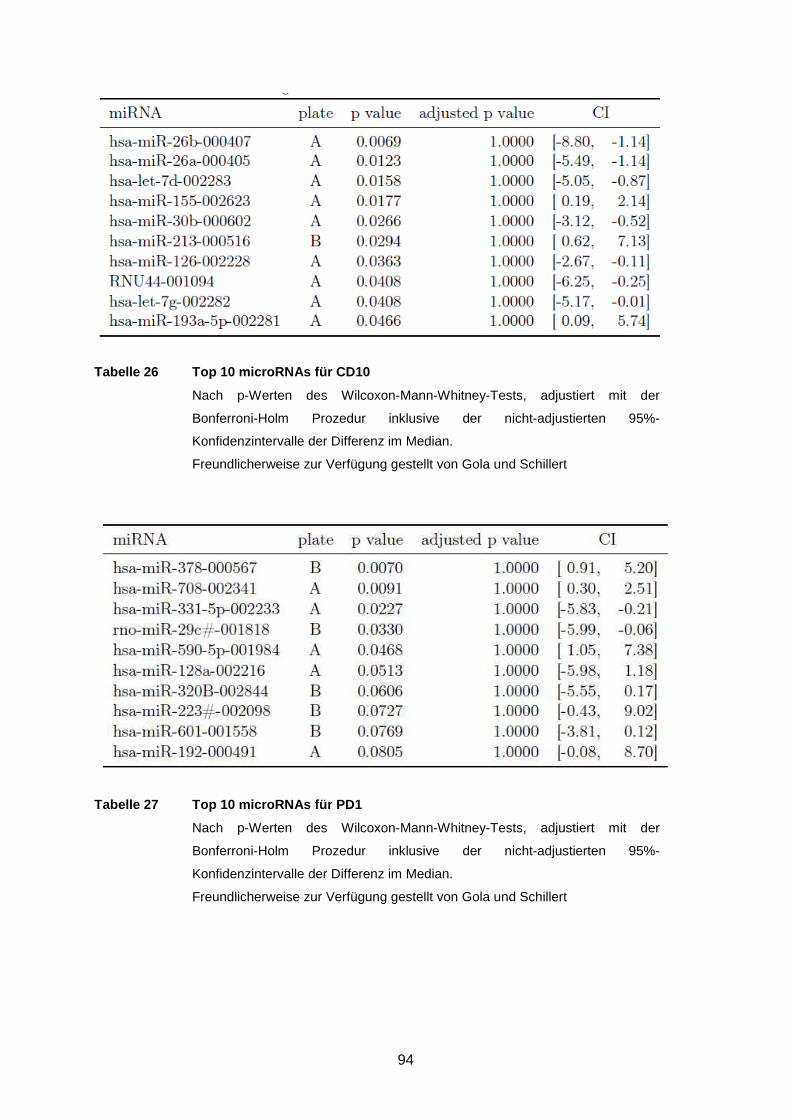
94
Tabelle 26 Top 10 microRNAs für CD10
Nach p-Werten des Wilcoxon-Mann-Whitney-Tests, adjustiert mit der
Bonferroni-Holm Prozedur inklusive der nicht-adjustierten 95%-
Konfidenzintervalle der Differenz im Median.
Freundlicherweise zur Verfügung gestellt von Gola und Schillert
Tabelle 27 Top 10 microRNAs für PD1
Nach p-Werten des Wilcoxon-Mann-Whitney-Tests, adjustiert mit der
Bonferroni-Holm Prozedur inklusive der nicht-adjustierten 95%-
Konfidenzintervalle der Differenz im Median.
Freundlicherweise zur Verfügung gestellt von Gola und Schillert

95
Tabelle 28 Top 10 microRNAs für die Proliferation d er HEV
Nach p-Werten des Kruskal-Wallis-Tests, adjustiert mit der Bonferroni-Holm
Prozedur
Freundlicherweise zur Verfügung gestellt von Gola und Schillert
Tabelle 29 Top 10 der microRNAs für die Expansion d er FDC-Netzwerke
Nach p-Werten des Kruskal-Wallis-Tests, adjustiert mit der Bonferroni-Holm
Prozedur
Freundlicherweise zur Verfügung gestellt von Gola und Schillert

96
Tabelle 30 Top 10 der microRNAs für das histomorpho logische Muster
Nach p-Werten des Kruskal-Wallis-Tests, adjustiert mit der Bonferroni-Holm
Prozedur
Freundlicherweise zur Verfügung gestellt von Gola und Schillert

97
8. Danksagung
Ich danke von ganzem Herzen:
Herrn Prof. Alfred Feller für die Überlassung des Themas.
Herrn Prof. Christoph Thorns für seine stets offene Tür, seinen intelligenten
Pragmatismus und seine aufmunternden Kommentare („Wir sind auf einem guten
Weg...“).
Juliana Knief, mit der ich seit mehr als 4 Jahren durch dick- und dünn gehe.
Dr. Julika Ribbat-Idel für ihre Hilfe bei Computerproblemen und ihre mentale
Unterstützung (Nervennahrung).
Annette Aufseß für ihre großartige technische Unterstützung und ihre Erklärungen.
Damian Gola und Arne Schillert für die professionelle statistische Auswertung.
Dr. Niklas Gebauer für seinen Humor und sein Know-How, das er gerne stets mit mir
teilte.
Den Mitarbeitern des Eck- und Molekular-Labors.
Meinen Eltern und Großeltern, für alles was sie mir mitgegeben haben und immer noch
geben.

98
9. Lebenslauf
Name: Katharina Reddemann
Geburtsdatum: 19. April 1985
Geburtsort: Bonn
Staatsangehörigkeit: Deutsch
Ausbildungsverlauf:
1994 - 2004 St. Adelheid Gymnasium,
Bonn
10/2004 – 11/2010 Studium der Humanmedizin,
Rheinische Friedrich - Wilhelms - Universität Bonn
Praktisches Jahr:
08/2009 - 12/2009 Abteilung für Chirurgie,
Landspítali Universitätshospital Reykjavík, Island
12/2009 - 04/2010 Abteilung für Hals-Nasen-Ohren-Heilkunde und -Chirurgie,
Klinikum Lüdenscheid, NRW
04/2010 - 07/2010 Abteilung für Kardiologie, Klinikum Lüdenscheid, NRW
Anstellungen:
08/2011 – heute Ärztin in Weiterbildung im Fach Pathologie.
Institut für Pathologie, UKSH, Lübeck
Direktor: Prof. Dr. A.C. Feller
Promotion (vorliegend):
Thema: „Charakterisierung von angioimmunoblastischen T-Zell-Lymphomen auf
morphologischer, immunhistochemischer und molekularer Ebene mit Schwerpunkt auf
dem microRNA-Expressionsprofil“ seit August 2013

99
Publikationen:
Auszüge der vorliegenden Arbeit wurden publiziert in:
Reddemann K, Gola D, Schillert A, Knief J, Kuempers C, Ribbat-Idel J, Ber S,
Schemme J, Bernhard V, Gebauer N, Feller AC, Thorns C. Dysregulation of
microRNAs in angioimmunoblastic T-cell lymphoma. Anticancer Res. 2015
Apr;35(4):2055-61.
Knief J, Gebauer N, Bernard V, Schemme J, Reddemann K, Gebauer J, Rades D,
Brabant G, Lehnert H, Feller AC, Thorns C. Oncogenic mutations and chromosomal
aberrations in primary extranodal diffuse large B-cell lymphomas of the thyroid - a
study of 21 cases. J Clin Endocrinol Metab. 2014 Nov 25:jc20143250.

100
10. Erklärung
Gemäß §9 Abs. 1 der Promotionsordnung
Hiermit versichere ich, dass ich die vorliegende Arbeit ohne fremde Hilfe im Institut für
Pathologie unter Betreuung durch Herrn Prof. Dr. med. Christoph Thorns angefertigt
und keine anderen als die in der Arbeit genannten Hilfsmittel benutzt habe.
Ich habe weder anderweitig versucht eine Dissertation einzureichen oder eine
Doktorprüfung durchzuführen, noch habe ich diese Dissertation oder Teile derselben
einer anderen Prüfungskommission vorgelegt.
Katharina Reddemann





























