Embed Size (px)
Citation preview

2
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
AVALIAÇÃO DE METODOLOGIAS DE ANÁLISE
ESTATÍSTICA E GEOESTATÍSTICA PARA ESTUDAR O
PROGRESSO ESPACIAL DE Colletotrichum truncatum EM
SEMENTES DE SOJA
RAFAEL NOETZOLD
C U I A B Á – MT/2011

3
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
AVALIAÇÃO DE METODOLOGIAS DE ANÁLISE
ESTATÍSTICA E GEOESTATÍSTICA PARA ESTUDAR O
PROGRESSO ESPACIAL DE Colletotrichum truncatum EM
SEMENTES DE SOJA
RAFAEL NOETZOLD
Engenheiro Agrônomo
Orientador: Prof°. Pós-Doc. MARCELO DE CARVALHO ALVES
Dissertação apresentada à Faculdade de Agronomia e Medicina Veterinária da Universidade Federal de Mato Grosso, para obtenção do título de Mestre em Agricultura Tropical.
C U I A B Á – MT/2011

3
FICHA CATALOGRÁFICA
Dados Internacionais para Catalogação na Publicação (CIP)
Bibliotecária: Patrícia Jaeger / CRB1-1736.
N772a Noetzold, Rafael
Avaliação de metodologias de análise estatística e geoestatística para estudar o
progresso espacial de Colletotrichum truncatum em sementes de soja/ Rafael
Noetzold - - Cuiabá: Universidade Federal de Mato Grosso - UFMT, 2011.
94f.:il.
Dissertação apresentada a Faculdade de Agronomia e Medicina Veterinária da Universidade Federal de Mato Grosso como parte dos requisitos para obtenção ao Título de Mestre em Agricultura Tropical.
Orientador: Prof.º Dr.-Doc. Marcelo de Carvalho Alves.
1. Sementes de Soja. 2. Colletotrichum truncatum. 3. Semivariograma. 4.
Krigagem. 5. Fungo. 6. Inoculação I. Título. II. Noetzold, Rafael. III. UFMT.
CDU: 631.53.027

4
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
CERTIFICADO DE APROVAÇÃO
Título: AVALIAÇÃO DE METODOLOGIAS DE ANÁLISE
ESTATÍSTICA E GEOESTATÍSTICA PARA ESTUDAR O
PROGRESSO ESPACIAL DE Colletotrichum truncatum EM
SEMENTES DE SOJA
Autor: RAFAEL NOETZOLD
Orientador: Prof°. Dr°. MARCELO DE CARVALHO ALVES
Aprovada em 16 de Fevereiro de 2011.

5
DECICATÓRIA
A DEUS, por me sustentar a cada dia, e ter me dado a oportunidade de
concluir o mestrado em Agricultura Tropical.
Aos meus pais, Anna B. Noetzold e Leone da S. Noetzold e meus irmãos
Marcio Noetzold e Ariel Noetzold pela educação, apoio, carinho, incentivo
durante o decorrer de meus estudos.

6
AGRADECIMENTOS
A coordenação de aperfeiçoamento de pessoal de nível superior
(Capes), por me conceder a bolsa de mestrado durante dois anos.
Aos funcionários da Universidade Federal de Mato Grosso – Faculdade
de Agronomia e Medicina Veterinária (Famev), por colaborarem na minha
formação de mestre.
Ao professor doutor, Marcelo de Carvalho Alves pela orientação, por me
ajudar, por me ensinar a pesquisar.
Ao professor doutor, Daniel Cassetari Neto pelo apoio e contribuição na
realização desta pesquisa. Agradeço também ao professor doutor Marcio
Willian Roque, pela amizade e apoio no decorrer do mestrado.
A todos os amigos da Famev, especialmente a turma de mestrado 2009,
pelo companheirismo, paciência, atenção, compreensão durante estes dois
anos.
Ao Centro Universitário de Várzea Grande (Univag) por ter cedido a área
experimental, especialmente à professora mestre, Andréia Quixabeira Machado
e funcionários da área experimental pelo auxílio na condução da pesquisa.
A toda equipe do Laboratório de Sensoriamento Remoto e
Geoinformação (Sergeo), pela ajuda na elaboração e condução da pesquisa.
A Syngenta, em especial ao mestrando, Volmir Cella, pelo incentivo a
pesquisa e pela doação de fungicida e inseticida.

5
SUMÁRIO
Página
AVALIAÇÃO DE METODOLOGIAS DE ANÁLISE ESTATÍSTICA E
GEOESTATÍSTICA PARA ESTUDAR O PROGRESSO ESPACIAL DE
Colletotrichum truncatum EM SEMENTES DE SOJA .................................... 6
1. INTRODUÇÃO ...................................................................................... 10
1.1 Importância da Semente .................................................................. 11
1.2 Arranjo Espacial de Doenças de Plantas ......................................... 14
1.3 Aplicação da Geoestatística ............................................................ 16
1.4 Referências Bibliográficas ............................................................... 18
2. ANÁLISE ESPACIAL DO ARRANJO DE Colletotrichum truncatum EM
SEMENTES E PLANTAS DE SOJA ............................................................ 23
2.1 Introdução ........................................................................................... 25
2.2 Material e Métodos ............................................................................. 26
2.2.1 Análise de dados .......................................................................... 34
2.3 Resultados e Discussão ..................................................................... 37
2.4 Conclusão ........................................................................................... 57
2.5 Referências Bibliográficas .................................................................. 57
3. ANÁLISE GEOESTATÍSTICA APLICADA À VARIABILIDADE ESPACIAL
DE Colletotrichum truncatum EM SEMENTES DE SOJA ............................ 62
3.1 Introdução ........................................................................................... 64
3.2 Material e Métodos ............................................................................. 65
3.2.1 Análise de dados .......................................................................... 71
3.3 Resultados e Discussão ..................................................................... 74
3.4 Conclusão ........................................................................................... 90
3.5 Referências Bibliográficas .................................................................. 90
4. CONSIDERAÇÕES FINAIS .................................................................. 94

6
AVALIAÇÃO DE METODOLOGIAS DE ANÁLISE ESTATÍSTICA E
GEOESTATÍSTICA PARA ESTUDAR O PROGRESSO ESPACIAL DE
Colletotrichum truncatum EM SEMENTES DE SOJA
RESUMO – Com o aumento da área cultivada de soja, há necessidade de
estudar a dinâmica de patógenos como o Colletotrichum truncatum, agente
etiológico da antracnose, pois a ocorrência dessa doença pode reduzir a
produtividade, causando, sob condições favoráveis, perda total da produção.
No sentido de colaborar para o sucesso da sojicultura, a geoestatística
fornece metodologias de análise para descrever a variação espacial de C.
truncatum. Assim, objetivou-se avaliar a distribuição espacial de níveis de C.
truncatum transmitido por sementes de soja por meio de técnicas estatísticas
e geoestatísticas. O experimento foi constituído de 3 parcelas de 9,90 x
10,00 m, onde sementes inoculadas com o fungo foram semeadas na região
central das parcelas, correspondendo aos níveis 0,8, 1,6 e 2,4% de
sementes inoculadas. Uma grade amostral com 112 pontos foi demarcada
em cada parcela para avaliar a incidência do patógeno nos folíolos e nas
sementes. O índice de dispersão, função K de Ripley e mapas de
distribuição de incidência foram utilizados para determinar o padrão espacial
dos folíolos de soja com base nos dados de incidência do patógeno pelo
método da câmara úmida. Após a análise sanitária das sementes pelo
método de incubação em papel de filtro com restrição hídrica, utilizou-se a
análise estatística de sequências ordinárias, índice de dispersão, função K
de Ripley e mapas de distribuição do fungo para avaliar o padrão espacial de
C. truncatum nas sementes. Modelos e métodos de semivariogramas foram
avaliados para descrever a estrutura e magnitude de dependência espacial
da incidência de C. truncatum nas sementes. A krigagem ordinária foi
utilizada para gerar mapas de variabilidade espacial do patógeno nas
sementes. O padrão espacial da transmissibilidade de C. truncatum para as
sementes foi agregado para a parcela com 0,8% de sementes inoculadas na
análise „doublet‟, e padrão aleatório para as parcelas com 1,6 % e 2,4% de
sementes de soja inoculadas com C. truncatum, para as análises „run‟ e

7
„doublet‟, respectivamente. Pelas análises do índice de dispersão o padrão
espacial predominante foi o aleatório. Pelos resultados obtidos pela função K
de Ripley, o padrão espacial predominante da incidência de C. truncatum
nos folíolos e sementes foi agregado para as parcelas com 0,8, 1,6 % e
2,4% de sementes de soja inoculadas com este patógeno. Com relação a
análise geoestatística, constatou-se por meio dos mapas de distribuição de
C. truncatum, padrão espacial agregado aleatório na primeira avaliação de
incidência dos folíolos de soja para os inóculos testados. Por meio da
validação cruzada, foi possível determinar a melhor aplicação de
semivariogramas esféricos isotrópicos para caracterizar a estrutura de
dependência espacial do fungo nas parcelas. Com o método da máxima
verossimilhança restrita foi obtido o melhor ajuste de modelos de
semivariogramas. Constatou-se alcance crescente da incidência do fungo
nas sementes com o aumento da porcentagem de sementes inoculadas nas
parcelas, fato também constatado nos mapas krigados. Foi possível
determinar que houve transmissão do fungo de sementes infectadas para
plantas e de plantas infectadas para semente, ao longo do ciclo de produção
da soja.
Palavras-chave: Semivariograma, krigagem, padrão espacial, fungo,
inoculação.
___________________
* Comitê de Orientação: Marcelo de Carvalho Alves – UFMT (Orientador); Daniel Cassetari
Neto – UFMT (Co-orientador); Marcio Willian Roque – UFMT (Co-orientador)

8
EVALUATION OF STATISTICAL AND GEOSTATISTICS ANALYSIS
METHODS TO STUDY SPACE PROGRESS OF Colletotrichum truncatum
IN SOYBEAN SEEDS
ABSTRACT – Due to the increase in soybean acreage, it‟s necessary to
study the dynamics of pathogens such as Colletotrichum truncatum, causal
agent of anthracnose, since the occurrence of this disease can reduce
productivity and cause under favorable conditions to it total loss of
production. In order to contribute with the success of the soybean crop,
geostatistics provides analysis methodologies to describe the spatial
variation of C. truncatum. Thus, the aim was to evaluate the spatial
distribution of levels of C. truncatum transmitted by soybean seeds through
statistical and geostatistical techniques. The experiment consisted of three
plots of 9.90 x 10.00 m, where seeds inoculated with the fungus were sown
in the central portions, corresponding to levels 0.8, 1.6 and 2.4% of
inoculated seeds. A sampling grid with 112 points was demarcated in each
plot to assess the incidence of infection in leaves and seeds. The dispersion
index Ripley's K function and incidence distribution maps were used to
determine the spatial pattern of soybean leaflets based on the incidence data
of pathogens by the method of wet chamber. After the seed health analysis
by the method of incubation on filter paper with water restriction, it was used
the statistical analysis of ordinary sequences, dispersion index, Ripley's K
function and distribution maps of the fungus to evaluate the spatial pattern of
C. truncatum in the seeds. Semivariogram models and methods were
evaluated to describe the structure and magnitude of spatial dependence of
incidence of C. truncatum in the seeds. The ordinary kriging was used to
create maps of spatial variability of the pathogen on the seeds. The spatial
pattern of transmissibility of C. truncatum to seeds was aggregated for the
parcel with 0.8% of inoculated seeds in the 'doublet' analysis and random
pattern to the plots with 1.6% and 2.4% of soybean inoculated with C.
truncatum, for 'run' and 'doublet' analysis respectively. According to the
dispersion index analysis, the predominant spatial pattern was the random

9
one. From the results obtained by Ripley's K function, the prevalent spatial
pattern in the incidence of C. truncatum in the leaflets and seeds was
aggregated to the plots with 0.8, 1.6% and 2.4% of soybean seeds inoculated
with this pathogen. With regard to the geostatistical analysis, the distribution
maps of C. truncatum showed the occurrence of random aggregated spatial
pattern at the first evaluation of the incidence of soybean leaflets to the
inocula tested. Through cross-validation it was possible to determine the best
application of isotropic spherical semivariograms to characterize the spatial
dependence structure of the fungus in the plots. With the restricted maximum
likelihood method we obtained the best fitting semivariogram models. It was
found that the pathogen incidence got higher on seeds as there was an
increase in the percentage of seeds inoculated on the plots, which was also
seen on the kriged maps. It was found that there was transmission of the
fungus from infected seeds to plants and from infected plants to seed
throughout the cycle of soybean production.
Keywords: Semivariogram, kriging, cross validation, spatial pattern.
____________________
* Advising Committe: Marcelo de Carvalho Alves – UFMT (Adviser); Daniel Cassetari Neto –
UFMT (Co-adviser); Marcio Willian Roque – UFMT (Co-adviser)

10
1. INTRODUÇÃO
A aquisição de sementes sadias é considerada uma etapa primordial
na cadeia produtiva da soja e de acordo com IMEA (2010) representou
7,24% do custo de produção de soja transgênica ha-1 na região de Campo
Verde - MT na safra 2010/2011. A semente é considerada um dos principais
veículos para introdução de doenças em novas áreas e o aumento do
inóculo em áreas já infestadas, com o passar dos anos, pode determinar
danos econômicos em detrimento ao aumento de patógenos.
Entre os agentes patogênicos que podem estar associado às
sementes de plantas, destacam-se os fungos como o maior grupo
(Machado, 1988). Na cultura da soja o fungo Colletotrichum truncatum
(Schw.) é o agente causal da antracnose, considerada uma das principais
doenças na região do cerrado brasileiro (Yorinori, 1997; Embrapa, 2003;
Almeida et al., 2005; Embrapa, 2010). A visualização e a identificação dos
sintomas e sinais desse patógeno nem sempre é possível por inspeção
visual, sendo necessário efetuar análises laboratoriais como a câmara úmida
e o teste de sanidade de sementes.
Nos últimos anos, as condições ambientais como temperatura e
umidade relativa do ar elevadas, associada ao aumento da monocultura da
soja, foram fatores fundamentais para a ocorrência da antracnose no
cerrado, acarretando redução na produtividade em determinadas áreas de
cultivo, onde a doença não foi manejada corretamente.
Atualmente não existe disponível no mercado, cultivares de soja com
resistência completa a antracnose para a região dos cerrados. Os cultivares

11
comercializados são suscetíveis ou moderadamente tolerantes a antracnose,
por este motivo, pesquisas com cultivares comerciais são necessárias a fim
de se conhecer a distribuição espacial e o progresso da doença durante o
ciclo da cultura da soja.
Conforme pesquisas de Campbell e Madden (1990) e Alves et al.,
(2006), existe forte estrutura de dependência espacial entre plantas durante
o progresso de doenças, em função da origem e da disseminação do
inóculo, principalmente quando existem reboleiras. Portanto, cada valor
carrega consigo forte interferência dos valores de sua vizinhança, de
maneira a ilustrar a continuidade espacial (Isaaks e Srivastava, 1989).
A geoestatística tem sido empregada para analisar a distribuição
espacial de epidemias de doenças de plantas, principalmente em parcelas
experimentais, ou em grandes áreas de lavouras (Chellemi et al. 1988; Stein
et al. 1994). Pesquisas feitas estudando a distribuição espacial de fungos
(Jaime-Garcia et al., 2001; Wu et al., 2001, Luo et al. 2001; Musoli et al.
2008) apresentam várias publicações, porém para doenças consideradas
epidêmicas para regiões de clima tropical, como o caso da antracnose da
soja, seu agente etiológico e sua transmissibilidade semente-semente ainda
não apresentam informações específicas.
O presente trabalho, considerou a hipótese que a antracnose da soja
apresenta padrão espacial agregado, ou seja, a sua distribuição ocorre em
reboleiras e que a quantidade de inóculo pode determinar variação espacial
no padrão de transmissibilidade do patógeno da semente para a semente.
Dessa forma, objetivou-se caracterizar a dinâmica e a estrutura de
variabilidade espacial de níveis de C. truncatum e avaliar a
transmissibilidade do patógeno da semente para semente de soja nas
condições de campo.
1.1 Importância da Semente
A agricultura moderna demanda que as sementes apresentam
características genéticas, com qualidade física, fisiológica e sanitária.
Todavia, a semente é um meio de levar ao agricultor todo o potencial de um

12
cultivar (Peske et al., 2004). As sementes contaminadas com fungos
fitopatogênicos são fontes de inóculo para o desenvolvimento da doença no
campo, desde que ocorra a interação entre hospedeiro suscetível, patógeno
agressivo e ambiente favorável.
A semente é uma das vias de introdução, disseminação e transmissão
da antracnose da soja na lavoura. Essa doença causa aumento do custo de
produção e ou redução de produtividade. Conforme Talamini et al. (2002), as
sementes são consideradas eficientes meios de disseminação de
fitopatógenos e, dentre esses, destacam-se as espécies pertencentes ao
gênero Colletotrichum.
A transmissão do patógeno é influenciada pela quantidade e
localização do inóculo nas sementes (Tanaka e Machado,1985 e Barba et
al., 2002). Conforme Machado (1988), a presença de Colletotrichum nas
sementes pode estar no interior das mesmas, seja nas camadas externas,
ou no embrião.
Para reduzir a disseminação de patógenos via sementes, o melhor
método é o uso de sementes livres de contaminações ou dentro de padrões
de tolerância estabelecidos para a cultura. Dessa maneira, do ponto de vista
sanitário, a semente ideal seria aquela livre de qualquer microorganismo
indesejável. Entretanto, isso nem sempre é possível, uma vez que a
qualidade sanitária das sementes é altamente influenciada pelas condições
climáticas sob as quais foram produzidas e armazenadas (Goulart, 1997).
Com a presença de C. truncatum em lavouras de soja, a transmissão
semente-planta e ou semente-semente é influenciada principalmente pelas
condições ambientais. Yorinori (1997) verificou que em anos chuvosos, a
antracnose pode causar perda total da produção, com alta redução do
número de vagens.
A antracnose ocorre em todas as fases de desenvolvimento da cultura
da soja, desde a fase de plântula até a fase inicial de formação das vagens,
constituindo-se em um dos mais importantes problemas da cultura nos
Cerrados (Yorinori, 1997).

13
A antracnose pode causar morte de plântulas, necrose dos pecíolos e
manchas nas folhas, hastes e vagens. Quando o inóculo é oriundo de restos
de cultura e sementes infectadas, pode causar necrose nos cotilédones, que
pode se estender para o hipocótilo, causando tombamento de pré e pós
emergência. O fungo infecta ramos laterais, pecíolos e vagens em qualquer
estádio de formação, podendo causar queda total das vagens ou
deterioração das sementes em colheita retardada. As vagens infectadas nos
estádios R3-R4 adquirem coloração castalho-escura a negra e ficam
retorcidas. Em períodos de alta umidade, as partes infectadas ficam
recobertas por pontuações negras, as quais são as frutificações do fungo. As
sementes apresentam manchas deprimidas, de coloração castanho-escuras
(Almeida et al., 2005).
Quando não se conhece o progresso de uma doença em uma
determinada região, as pesquisas são realizadas basicamente de duas
maneiras: efetuando-se o monitoramento periódico em lavouras onde o
patógeno em estudo está presente, ou por meio da inoculação do patógeno
em sementes, as quais são semeadas em condições de campo, e o
monitoramento é realizado até o final do ciclo.
Várias pesquisas foram feitas com o objetivo de conhecer o melhor
método para a infecção das sementes de soja. Entre elas, Machado et al.
(2001) relataram que para fungos muito agressivos como o C. dematium var.
truncata, 48 horas de exposição das sementes ao inóculo é suficiente para
obter quase 100% de infecção. Galli et al. (2005) verificaram que o tempo de
incubação de 40 horas para o mesmo patógeno é suficiente para obtenção
da totalidade das sementes infectadas.
Contudo, existe a carência de estudos realizados com a finalidade de
avaliar o progresso espaço-temporal de C. truncatum em condições de
campo. A caracterização do padrão espacial da doença e do patógeno pode
gerar informações essenciais na elaboração de estratégias de manejo de
doenças de plantas.

14
1.2 Arranjo Espacial de Doenças de Plantas
O padrão espacial de doença é definido como arranjo ou
posicionamento das plantas doentes, umas em relação às outras (Gilligan,
1983). Os patógenos de plantas possuem distintos agentes de dispersão
como o vento, a água, vetores, material propagativo contaminado, animais
domésticos e silvestres, o homem com calçados, ferramentas, máquinas e
implementos agrícolas. Conforme Gilligan (1982), o padrão espacial de uma
doença depende não só da forma de dispersão do patógeno, como também
da arquitetura das plantas hospedeiras e do arranjo destas no campo.
Basicamente a distribuição espacial de uma doença no campo pode
apresentar três tipos de padrões: regular, casualizado (aleatório) e agregado
(agrupado) (Bergamin Filho et al. 2002).
O padrão espacial aleatório relaciona-se geralmente aos patógenos
dispersados pelo vento, podendo atingir longas distâncias em curto período
de tempo, enquanto patógenos veiculados por respingos de chuva
dispersam-se lentamente ao redor dos focos primários. No entanto, em
condições naturais, a maioria dos patógenos e doenças possuem mais de
um mecanismo de dispersão (Bergamin Filho et al., 2002).
O arranjo espacial de plantas doentes pode mudar ao longo da
epidemia. Considerando-se epidemias como o aumento da intensidade de
doenças em tempo e espaço, é importante associar a dinâmica espacial aos
estudos temporais, bem como enfatizar o arranjo espacial ao planejar
ensaios de campo (Maffia e Mizubuti, 2005).
Segundo Madden et al. (1982), várias técnicas têm sido utilizadas
para analisar e caracterizar o arranjo espacial de plantas doentes. A análise
de „ordinary runs‟, se baseia no uso de dados binários de incidência da
doença (ausência/presença), e é o método preferido por epidemiologistas
para estudar a presença ou não de agregação de indivíduos doentes dentro
e entre linhas em uma população de plantas (Medeiros, 2010). A análise
„doublet‟ também é utilizada para analisar o padrão espacial e emprega
também dados binários de incidência de doença.

15
Amaral Mello (2007), estudando o fitoplasma associado ao
enfezamento do repolho e utilizando seqüências ordinárias, concluiu que o
padrão espacial da doença foi agregado.
A análise de sequências ordinárias permite a constatação de
agrupamento dentro de linhas de plantio ou em sentido perpendicular a elas.
Assim, é possível relacionar (indiretamente) o padrão espacial encontrado à
práticas de manejo adotadas nos locais estudados por meio de técnicas
estatísticas (Laranjeira, 2002).
O índice de dispersão (D) é outro método aplicado em estudos
epidemiológicos para indicar o padrão espacial. Quando D<1 o padrão é
regular, quando D=1 o padrão é aleatório e quando D>1 o padrão é
considerado agregado (Upton e Flingleton, 1985). Conforme Bergamin Filho
et al. (2002), o teste do Chi-quadrado, pode ser utilizado para confirmar
objetivamente essas indicações e, este índice é válido para dados de
contagem (exemplo: número de lesões por planta ou o número de insetos
por planta).
Na análise por sub-região é definida uma área a partir de determinado
número de linhas e números de plantas na área amostral denominado de
quadrat. A análise por quadrats é utilizada para definir o padrão espacial da
doença ou do patógeno. De acordo com Krainski e Ribeiro Junior (2006),
esta análise pode ser implementada com o índice de dispersão binomial,
índice de dispersão Poisson e o modelo beta-binomial. Os mesmos autores
afirmam que quando as dimensões da matriz de dados não for múltipla das
dimensões do quadrat, os dados excedentes são desconsiderados da
análise. Se faltar alguma planta para completar um quadrat dentro da área
amostral, este também é desconsiderado da análise.
Para Madden e Hughes (1995) os índices de dispersão de modo
geral, baseados nas premissas da distribuição de Poisson, não devem ser
usados para dados de incidência (especialmente para incidências > 20%),
sendo que para esse tipo de dado (binário), a melhor aproximação de uma
condição de aleatoriedade é dada pela distribuição binomial.

16
Apesar do índice D ser muito utilizado, este índice de dispersão tem
como limitação não levar em consideração a posição relativa de cada
medida (Nicot et al., 1984).
Além destes métodos, a análise de padrões pontuais também está
sendo empregada para investigar o padrão espacial de doenças de plantas.
A metodologia consiste em analisar dados disponíveis em um determinado
espaço e por meio de métodos e modelagem, procura verificar se os eventos
observados apresentam comportamento sistemático, como por exemplo,
agregação, regularidade ou aleatoriedade (Spósito, 2003).
A função K de Ripley e a distância mínima são exemplos em análises
de padrões pontuais, sendo considerado por Ripley (1981), um bom
indicador de estruturas espaciais. Ela é uma análise de distribuição espacial
de dados pontuais de segunda ordem com ampla utilização em ecologia
(Haase, 1995). A distância mínima é definida como a distância entre uma
planta doente e a planta doente mais próxima, de acordo com Krainski et al.
(2011), é aceitável assumir que quanto menor a média das distâncias
mínima das plantas doentes, dada uma incidência, mais agregado é o
padrão espacial.
Nesse sentido pesquisas realizadas por Spósito et al. (2007),
estudaram o padrão espacial da mancha preta dos citrus por meio do índice
de dispersão, função K de Ripley e distância mínima média, e os resultados
das três análises indicaram padrão agregado entre as plantas doentes,
independentemente da incidência da doença.
1.3 Aplicação da Geoestatística
O conhecimento e o emprego da ferramenta geoestatística está sendo
utilizado em áreas do setor agrícola, entre elas destacam-se a elaboração de
mapas de produtividade, mapeamento de atributos físicos e químicos de
solos e mais recentemente no mapeamento de pragas e doenças de plantas.
A teoria fundamental da geoestatística é a esperança de que, na
média, as amostras próximas no tempo e no espaço sejam mais similares
entre si do que as que estiverem distantes (Issaks e Srivastava, 1989).

17
As técnicas geoestatísticas podem ser usadas para descrever e
modelizar padrões espaciais (variografia), para predizer valores em locais
não amostrados (krigagem), para obter a incerteza associada a um valor
estimado em locais não amostrados (variância de krigagem) e para otimizar
malhas de amostragem (Andriotti, 2003).
A escolha de um modelo teórico do semivariograma é um dos
aspectos mais importantes das aplicações das variáveis regionalizadas e
pode ser uma das maiores fontes de ambigüidade e polêmicas nessas
aplicações (Vieira, 2000). Entre os diversos modelos de semivariogramas os
mais utilizados são o esférico, o exponencial e o gaussiano.
Para escolher o melhor modelo e método de ajuste do
semivariograma, critérios de validação são empregados. De acordo com
Faraco et al. (2008), são utilizados: Informação de Akaike, de Filliben, de
validação cruzada e o máximo valor do logaritmo da função verossimilhança.
De acordo com Cressie (1993), validação cruzada tem sido um meio
popular de avaliar a estimativa estatística e previsão desde os artigos de
Stone (1974) e Geisser (1975). A idéia básica é a de eliminar alguns dos
dados e utilizar os dados restantes para a predição das observações que foi
excluída. Então, o erro de previsão pode ser inferido a partir dos valores
preditos menos os obsevados. A repetição da técnica para os dados permite
analisar o erro do sistema.
Conforme Cressie (1993), na comparação de estimadores, a exclusão
de observações para melhorar a inferência de um parâmetro estimado foi
chamado jackkninfing por Tukey (1958). Jackknifing foi proposto por
Quenouille (1949), para reduzir o viés no estimador original. Envolve uma
exclusão sistemática de cada dado (ou, mais geralmente, os subconjuntos
dos dados) (Cressie, 1993).
Conforme Isaaks e Srivastava (1989), a validação cruzada é a técnica
de avaliação de erros de estimativa que permite comparar valores preditos
com os amostrados.
A comparação entre modelos de semivariogramas e métodos de
ajuste pode ser efetuada através do erro médio e erro reduzido, fornecendo

18
dados precisos e acurados, em função da importância do resultado dos
coeficientes de qualidade, consequentemente contribui para a escolha do
modelo e método que melhor represente a variável em estudo.
Salienta-se que a validação cruzada é um método de seleção do
modelo de semivariograma, que permite escolher o modelo matemático que
melhor descreve a dependência espacial dos pontos amostrais da variável
em estudo, em função da distância. Para Hernández e Emeri (2009), a idéia
da validação cruzada consiste em validar a aptidão do modelo ajustado de
semivariograma associado à incerteza do atributo não amostrado.
1.4 Referências Bibliográficas
ALMEIDA, A.M.R. FERREIRA, L.P.; YORINORI, J.T. et al. Doenças da soja (Glycine Max). In: KIMATI, H. et al. In: Kimat et al. Manual de fitopatologia: Doenças das plantas cultivadas. 4ª Ed. Vol. 2, São Paulo: Agronômica Ceres, 2005, p.571-588. ALVES, M.C.; POZZA, E.A.; MACHADO, J.C.; et al. Geoestatística como metodologia para estudar a dinâmica espaço-temporal de doenças associadas a Colletotrichum spp. transmitidos por sementes. Fitopatologia Brasileira, v.31, p.557-563, 2006. AMARAL MELLO, A.P.O. Identificação molecular de fitoplasmas associados ao enfezamento do repolho e análise epidemiológica da doença. 2007. 64f. Tese (Doutorado). Escola Superior de Agricultura Luiz de Queiroz, Piracicaba – SP, 2007. ANDRIOTTI, J.L.S. Fundamentos de Estatística e Geoestatística. São Leopoldo. 2003, 165p. BARBA, J.T.; REIS, E.M.; FORCELINI, C.A. Efeito da temperatura e de fungicida na transmissão de Bipolaris sorokiniana da semente para plântulas de cevada. Fitopatologia Brasileira, v.27, n.5, p.500-507, 2002. BERGAMIN FILHO, A. HAU, B.; AMORIM, L. et al. Análise espacial de epidemias. Revisão Anual de Patologia de Plantas, Passo Fundo, v.10, 2002, p.155-218. CAMPBELL, C.L.; MADDEN, L.V. Introduction to plant disease epidemiology. New York: J. Wiley, 1990, 532p.

19
CHELLEMI, D.O.; ROHRBACH, K.G.; YOST, R.S.; SONODA, R.M. Analysis of the spatial pattern of plant pathogens and diseased plants using geostatistics. Phytopathology, v.8, p.221-226, 1988. CRESSIE, N. Statistics for spatial data. New York: John Wiley, 1993, 900p. EMBRAPA. Tecnologias de produção de soja região central do Brasil. Londrina: EMBRAPA Soja (Sistema de Produção n.1), 2003, 32p. EMBRAPA. Tecnologias de produção de soja região central do Brasil 2011. Londrina: Embrapa Soja: Embrapa Cerrados: Embrapa Agropecuária Oeste (Sistemas de Produção n.14), 2010, 255p. FARACO, M.A.; URIBE-OPAZO, M.A.; SILVA, E. A.A.; JOHANN, J. A.; BORSSOI, J. A. Seleção de modelos de variabilidade espacial para elaboração de mapas temáticos de atributos físicos do solo e produtividade da soja. Revista Brasileira de Ciência do Solo, v.32, p.463-476, 2008. GALLI, J.A. et al. Efeito de Colletotrichum dematium var. truncata e Cercospora kikuchii na germinação de sementes de soja. Revista Brasileira de Sementes. v.27, p.182-187, 2005. GEISSER, S. The predictive sample reuse method with applications. Journal of the American Statistical Association, v.70, p.320-328, 1975. GILLIGAN, N.A. Statistical analysis of the spatial pattern of Botrytis fabae on Vicia faba: a methological study. Transactions of the British Mycology Society, v.79, p.193-200, 1982. GILLIGAN, C.A. Modeling of soilborne pathogens. Annual Review of Phytopathology, v.21, p.45-64, 1983. GOULART, A.C.P. Tratamento de Sementes de Soja com Fungicidas. Dourados: EMBRAPA –CPAO. (Circular Técnica, 6), 30p, 1997. KRAINSKI, E.T.; RIBEIRO JUNIOR, P.J. Análise por quadrat counts em R . (2006). Disponível em: http://www.leg.ufpr.br/Rcitrus/quadrats/quadrats.pdf. Acesso em: 17 mar. 2011.
KRAINSKI, E.T.; RIBEIRO JUNIOR, P.J.; ANDRADE NETO, P.R.; BASSANEZI, R.B. Um ambiente para monitoramento da morte súbita dos citrus. Disponível em: http://www.dpi.inpe.br/geoinfo/geoinfo2005/papers/P72.PDF. Acesso em: 17 mar. 2011.

20
HAASE, P. Spatial pattern analysis in ecology based on Ripley's K-function: introduction and methods of edge correction. Journal of Vegetation Science, v.6, p. 575-582, 1995. HERNÁNDEZ, J; EMERY, X. A geostatistical approach to optimize sampling designs for local forest inventories. Published by NRC Research Press. Can. J. For. Res. v.39, p.1465–1474, 2009. IMEA - Instituto Mato-grossense de Economia Agropecuária. Custo de Produção Soja - Safra 10/11. Mato Grosso. Base Setembro/10. Disponível em http: http://<www.imea.com.br/upload/publicacoes/arquivos/10_09_CPS.pdf>. Acesso em: 19 fev. 2011.
ISAAKS, E.H.; SRIVASTAVA, R.M. An introduction to applied geostatistics. New York, Oxford University Press, 1989, 561p. JAIME-GARCIA, R.; ORUM, T.V.; FELIX-GASTELUM, R.; TRINIDAD-CORREA, R.; VANETTEN, H.D.; NELSON, M.R. Spatial analysis of Phytophthora infestans genotypes and late blight severity on tomato and potato in the del fuerte valley using geostatistics and geographic information systems. Phytopathology, v.91, p.156-165, 2001. LARANJEIRA, F. F. Epidemiologia da clorose variegada dos citros no Estado de São Paulo. 2002. 158f. Tese (Doutorado em Fitopatologia), Escola Superior de Agricultura Luiz de Queiroz, Piracicaba – SP, 2002. LUO, Y.; CHONG, S.K.; MYERS, O. Spatio-temporal analysis of soybean root colonization by Fusarium solani f. sp. glycines in fields. Plant Disease. v.85, p.303-310, 2001. MACHADO, J.C. Patologia de sementes: fundamentos e aplicações. Ciências Agrárias nos Trópicos Brasileiros. Brasília: MECESAL-FAEPE, 1988, 106p. MACHADO, J.C.; OLIVEIRA, J.A; VIEIRA, M.G.G.C.; ALVES, M.C. Inoculação artificial de sementes de soja por fungos, utilizando solução de manitol. Revista Brasileira de Sementes, v.23, p.95-101, 2001. MADDEN, L.V. HUGHES, G. Plant disease incidence: distributions, heterogeneity, and temporal analysis. Annual Review of Phytopathology, v.33, p. 529-564, 1995. MADDEN, L.V.; LOUIE, R., ABT; J.J., KNOKE, J.K. Evaluation of tests for randomness of infected plants. Phytopathology, St. Paul, v.72, p.195-198, 1982.

21
MAFFIA, L.A.; MIZUBUTI, E.S.G. Epidemiologia de doenças radiculares. In: MICHEREFF, S.J.; DOMINGOS, E.G.T.; ANDRADE, M.M. Ecologia e manejo de patógenos radiculares em solos tropicais. Recife: UFRPE, Imprensa Universitária, 2005, p.207-246. MEDEIROS, R.D. Análise do arranjo espacial e determinação da estrutura de focos da resinose coqueiro (Cocos nucifera L.) na região de Neópolis, Sergipe. 2010. 40f. Dissertação (Mestrado em Agroecossiste-mas). Universidade Federal de Sergipe, Sergipe – SE, 2010. MUSOLI C.P.; PINARD F.; CHARRIER A.; KANGIRE A.; TEN HOOPEN G.M.; KABOLE C.; OGWANG J.; BIEYSSE D.; CILAS C. Spatial and temporal analysis of coffee wilt disease caused by Fusarium xylarioides in Coffea canephora. European journal of plant pathology, v.4, p.451-460, 2008. NICOT, P.C.; ROUSE, D.I.; YANDELL, B.S. Comparison of statistical methods for studying patterns of soilbome plant pathogens in the field. Phytopathology, v.74, p. 1399-1402, 1984. PESKE, S.T., SCHUCH, L.O.B., BARROS, A.C.S.A.; Produção de arroz irrigado, Pelotas: Universidade Federal de Pelotas. Editora Universitária, 2004. QUENOUILLE, M. Approsimate tests of correlation in time series. Journal of the Royal Statistics Society B, v.11, p.68-84, 1949. RIPLEY, B.D. Spatial statistic. New York: John Wiley, 1981. 272p. SPÓSITO, M.B. Dinâmica temporal e espacial da mancha preta (Guignardia citricarpa) e quantificação dos danos causados à cultura dos citros. 2004. 112p. Tese (Doutorado em Agronomia) – Universidade de São Paulo, Piracicaba - SP, 2003.
SPÓSITO, M.B.; AMORIM, L.; RIBEIRO JUNIOR, P.J.; BASSANEZI, R.B.; KRAINSKI, E.T. Spatial Pattern of Trees Affected by Black Spot in Citrus Groves in Brazil. Plant Disease, v. 91, p. 36-40, 2007. STEIN, A.; KOCKS, C.G.; ZADOKS, J.C.; FRINKING, H.D.; RUISSEN, M.A.; MYERS, D. E. A geostatistical analysis of the spatiotemporal development of the downy mildew epidemics in cabbage. Phytopathology, v.84, p.1227-1239, 1994. STONE, M. Cross-validatory choice and assessment of statistical predictions. Journal of the Royal Statistics Society B, v.36, p.111-133, 1974.

22
TANAKA, M.A.S.; MACHADO, J.C. Patologia de Sementes. Informe Agropecuário, v.11, p.40-46, 1985.
TALAMINI, V.; POZZA, E.A.; MACHADO, J.C.; et al. Epidemiologia de doenças associadas a Colletotrichum spp. Transmitidas por sementes. Revisão Anual de Patologia de Plantas, v. 10, 2001, p. 219-248. TUKEY, J.W. Bias and confidence in not-quite large samples (abstracts). Annals of Mathematical Statistics, v.29, p.614, 1958. UPTON, G. e FLINGLETON, B. Spatial data analysis by example. Point pattern and quantitative data. Chichester: Wiley, 1985. 409p. VIEIRA, S.R. Geoestatística em estudos de variabilidade espacial do solo. In: NOVAIS, R.F. de; ALVAREZ V., V.H.; SCHAEFER, C.E.G.R. (Ed.). Tópicos em Ciência do Solo. Viçosa: Sociedade Brasileira de Ciência do Solo, v.1, 2000, p.1- 54.
YORINORI, J.T. In: DO VALE, F.X.R.; ZAMBOLIM, L. Controle de doenças de plantas. v.2. Grandes Culturas. UFV, Viçosa-MG, p.953-1023, 1997. WU, B.M.; VAN BRUGGEN, A.H.C.; SUBBARAO, K.V.; PENNINGS, G.G.H. Spatial Analysis of Lettuce Downy Mildew Using Geostatistics and Geographic Information Systems. Phytopathology, v.91, p.134-142, 2001.

23
2. ANÁLISE ESPACIAL DO ARRANJO DE Colletotrichum truncatum EM
SEMENTES E PLANTAS DE SOJA
RESUMO - O conhecimento do padrão espacial de patógenos de plantas é
uma informação fundamental para o manejo de doenças transmitidas por
sementes na cultura da soja. Objetivou-se avaliar o padrão espacial de
Colletotrichum truncatum em sementes e plantas de soja. O experimento foi
conduzido na área experimental do UNIVAG, constituído por 3 parcelas de
9,90 x 10,00 m, espaçadas 0,45 m entre linhas. A fonte de inóculo foi
semeada na região central das parcelas. Efetuou-se avaliações de
incidência do patógeno em folíolos de soja. Após a colheita e a secagem,
efetuou-se a análise sanitária das sementes pelo método „blotter test‟,
correspondente a 112 pontos demarcados em cada parcela. O padrão
espacial do patógeno foi obtido pela transmissibilidade semente-semente do
patógeno, nas linhas perpendicu- lares a linha de semeadura por meio das
análises de seqüências ordinárias ‘run‟ e „doublet‟. O índice de dispersão,
função K de Ripley e mapas de distribuição de incidência foram utilizados
para determinar o padrão espacial dos folíolos e sementes de soja. O padrão
espacial da transmissibilidade de C. truncatum para as sementes foi
agregado para a parcela com 0,8% de sementes inoculadas na análise
„doublet‟, e padrão aleatório para as parcelas com 1,6 % e 2,4% de
sementes de soja inoculadas com C. truncatum, tanto para análise „run‟
como „doublet‟. A distância mínima média de uma planta para outra planta
com a presença do patógeno diminuiu com o passar do tempo e com o
aumento do inóculo. Pelas análises do índice de dispersão e função K de
Ripley o padrão espacial predominante foi o aleatório e agregado
respectivamente. Constatou-se por meio dos mapas de distribuição de C.
truncatum, padrão espacial agregado aleatório na primeira avaliação de
incidência dos folíolos de soja para os inóculos testados.
Palavras-chave: Fonte de inóculo, seqüências ordinárias, função K de
Ripley, padrão agregado.

24
2. SPATIAL ARRANGEMENT ANALYSIS OF Colletotrichum truncatum
IN SOYBEAN PLANTS AND SEEDS
ABSTRACT - The knowledge of the spatial pattern of plant pathogens is
fundamental information for the management of seed-borne diseases in
soybean. The objective was to evaluate the spatial pattern of Colletotrichum
truncatum in soybean plants and seeds. The experiment was conducted at
the UNIVAG‟s experimental area, consisting of three plots of 9.90 x 10.00 m,
with 0.45 m distance between rows. The source of inoculum was sown in the
central portions of the plots. It was conducted assessments of incidence of
the pathogen in soybean leaflets. After harvesting and drying, it was
conducted the seed health analysis by 'blotter test' method corresponding to
112 points demarcated on each plot. The spatial pattern of the pathogen was
obtained from seed to seed transmissibility of the pathogen in perpendicular
lines to the sowing line through the ordinary sequence analysis 'run' and
'doublet'. The dispersion index, Ripley's K function and distribution maps of
incidence were used to determine the spatial distribution of leaflets and
soybean seeds. The spatial pattern of transmissibility of C. truncatum for
seeds was aggegated to the parcel with 0.8% of inoculated seeds in the
'doublet' analysis and random pattern to the plots with 1.6% and 2.4% of
soybean seeds inoculated with C. truncatum for both 'run' and 'doublet'
analysis. The minimum average distance from one plant to another plant with
the presence of the pathogen decreased over time and with the increase of
the inoculum. According to the analysis of the dispersion index and Ripley's
K function, the predominant spatial patterns were the random and
aggregated ones respectively. The distribution maps of C. truncatum showed
the occurrence of random aggregated spatial pattern at the first evaluation of
the incidence of soybean leaflets to the inocula tested.
Keywords: Source of inoculum, ordinary runs, Ripley‟s K function,
aggregated pattern.

25
2.1 Introdução
O padrão espacial de uma doença é definido como arranjo ou
posicionamento das plantas doentes, umas em relação às outras (Gilligan,
1983), e é dividido basicamente em regular, aleatório ou casualizado e
agregado ou agrupado. Conhecer o padrão espacial do patógeno pode gerar
informações essenciais na elaboração de estratégias de manejo integrado
de doenças.
O conhecimento das características de uma epidemia possibilita uma
visão ampla da estrutura e do processo do patossistema, ademais,
informações podem ser extraídas nas análises espaço-temporal dos dados,
possibilitando entender a forma de como ocorre a dispersão do patógeno no
campo, o que resulta, em padrões espaciais de doença (Campbell e
Madden, 1990).
Para obtenção de informações a respeito da distribuição espacial de
fitopatógenos em culturas agrícolas, fatores como localização do inóculo
inicial na lavoura, relevo do terreno, direção do vento, temperatura do ar,
umidade relativa do ar e duração do molhamento foliar, são importantes.
Na cultura da soja, a presença do agente etiológico Colletotrichum
truncatum não resulta necessariamente em antracnose, pois o patógeno
pode permanecer latente se as condições ambientais não forem favoráveis.
A semente infectada com C. truncatum é considerada como um dos
principais meios de introdução e ou aumento de ocorrência da antracnose
em lavouras de soja.

26
Existem vários métodos para descrever o padrão espacial de
doenças, com o intuito de estudar a dinâmica de distribuição e localização
de plantas doentes em lavouras. Entre esses métodos, destacando-se os
testes „run‟ e „doublet‟ (Bergamin Filho et al. 2004; Ferreira et al., 2009),
índice de dispersão (D) e função K de Ripley ( Spósito et al., 2007). Esses
testes também podem ser também utilizados para detectar o padrão espacial
de fitopatógenos.
Conforme ressaltou Hughes et al., (1997), a principal razão para se
estudar padrões espaciais de doenças no campo é expressar o processo de
dispersão do patógeno, a fim de se conhecer as características da
distribuição do patógeno na lavoura, e com isso, estabelecer estratégias
para controlar a doença.
O presente trabalho teve por objetivo avaliar o padrão espacial de
Colletotrichum truncatum em sementes e plantas de soja em parcelas
experimentais.
2.2 Material e Métodos
O experimento foi conduzido durante o período de agosto de 2009 à
abril de 2010, na área experimental limitada pelas coordenadas 8270210 S e
596532 W e altitude de 180m, localizada no Centro Universitário de Várzea
Grande – UNIVAG na cidade de Várzea Grande – MT. A pesquisa em
campo foi constituída por 3 parcelas de 9,9x10m cada.
O isolado de C. truncatum foi obtido de sementes de soja
submetidas à análise de rotina („blotter test‟) no Laboratório de Fitopatologia
do UNIVAG, da safra 2009/2010. O inóculo do patógeno foi multiplicado em
meio de cultura BDA (200 g de batata, 20 g de dextrose e 20 g de Agar). O
cultivo foi realizado em placa de Petri de 9 cm de diâmetro, por 7 dias a
temperatura de 20 (± 2°C) em regime alternado de 12 h luz / 12 h escuro.
Após esse período, 130 sementes de soja do cultivar TMG 131 RR,
suscetível ao fungo C truncatum e livres de patógenos foram distribuídas na
placa de Petri contendo a colônia fúngica em crescimento ativo sobre o BDA,
disposta em uma única camada ocupando toda a superfície da mesma, por

27
um período de 48 h. As sementes foram retiradas da placa e colocadas para
secar sobre papel toalha em temperatura ambiente, por 2 horas (h).
O cultivar TMG 131 RR foi escolhido em função das seguintes
vantagens: facilidade do manejo das plantas daninhas, resistência aos
nematóides de galhas das raças 1 e 3, menor tempo de permanência no
campo favorecendo menor pressão da ferrugem asiática e por ser cultivado
no estado de Mato Grosso desde a região sul (baixa altitude) até a região do
médio norte (Fundação Mato Grosso, 2010).
Em agosto de 2009 foram coletados amostras de solo na área
experimental a fim de determinar a textura e características químicas do solo
(Tabela 1).
Tabela 1. Análise química e textural do solo da área experimental.
pH P K Ca Mg Areia Silte Argila
Água CaCl2 Mg dm-3 Cmolc dm-3 % 6,8 6,0 27,7 44 1,3 0,7 85,7 4,0 10,3
Zn Cu Fe Mn B S
mg dm-3 0,7 1,0 194 40,3 0,22 6,4
Já em setembro de 2009 foram semeadas 4 linhas de milho híbrido
AG 7088 ao redor das 3 parcelas , distanciadas a 2 m das mesmas em todas
as direções (norte, sul, leste, oeste) com espaçamento entre linhas de 0,50
m e população de 60000 plantas ha-1 (Figura 1). A adubação foi de acordo
com os níveis de nutrientes no solo (Tabela 1) juntamente com a
recomendação de Alves et al. (1999), sendo aplicado 20 kg de N, 100 kg
P2O5 e 80 kg K2O. Os demais tratos culturais estão descritos na tabela 2.
O preparo convencional do solo foi realizado duas vezes com grade
aradora e uma vez com grade niveladora. Após a abertura manual dos
sulcos com auxílio de enxada, o adubo foi distribuído ao lado e abaixo das
sementes, conforme recomendação da Embrapa Soja (2008) (Tabela 2), em
seguida houve a incorporação do adubo, e as sementes foram distribuídas
em profundidade de 2 a 3 cm.

28
Figura 1. Croqui da área experimental. Parcelas com 0,8% (A), 1,6% (B) e
2,4% (C) de sementes inoculadas com C. truncatum.
29
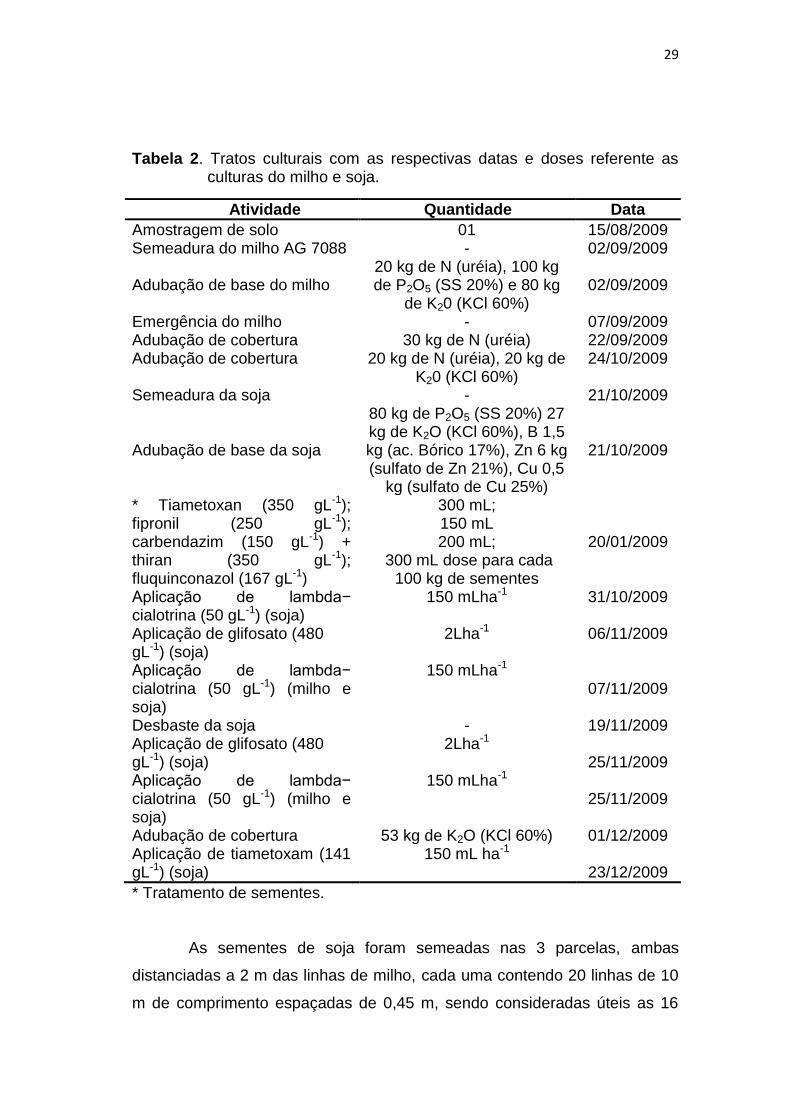
Tabela 2. Tratos culturais com as respectivas datas e doses referente as culturas do milho e soja.
Atividade Quantidade Data
Amostragem de solo 01 15/08/2009 Semeadura do milho AG 7088 - 02/09/2009 Adubação de base do milho
20 kg de N (uréia), 100 kg de P2O5 (SS 20%) e 80 kg
de K20 (KCl 60%)
02/09/2009
Emergência do milho - 07/09/2009 Adubação de cobertura 30 kg de N (uréia) 22/09/2009 Adubação de cobertura 20 kg de N (uréia), 20 kg de
K20 (KCl 60%) 24/10/2009
Semeadura da soja - 21/10/2009 Adubação de base da soja
80 kg de P2O5 (SS 20%) 27 kg de K2O (KCl 60%), B 1,5 kg (ac. Bórico 17%), Zn 6 kg (sulfato de Zn 21%), Cu 0,5
kg (sulfato de Cu 25%)
21/10/2009
* Tiametoxan (350 gL-1); fipronil (250 gL-1); carbendazim (150 gL-1) + thiran (350 gL-1); fluquinconazol (167 gL-1)
300 mL; 150 mL 200 mL;
300 mL dose para cada 100 kg de sementes
20/01/2009
Aplicação de lambda− cialotrina (50 gL-1) (soja)
150 mLha-1 31/10/2009
Aplicação de glifosato (480 gL-1) (soja)
2Lha-1 06/11/2009
Aplicação de lambda− cialotrina (50 gL-1) (milho e soja)
150 mLha-1 07/11/2009
Desbaste da soja - 19/11/2009 Aplicação de glifosato (480 gL-1) (soja)
2Lha-1 25/11/2009
Aplicação de lambda− cialotrina (50 gL-1) (milho e soja)
150 mLha-1 25/11/2009
Adubação de cobertura 53 kg de K2O (KCl 60%) 01/12/2009 Aplicação de tiametoxam (141 gL-1) (soja)
150 mL ha-1 23/12/2009
* Tratamento de sementes.
As sementes de soja foram semeadas nas 3 parcelas, ambas
distanciadas a 2 m das linhas de milho, cada uma contendo 20 linhas de 10
m de comprimento espaçadas de 0,45 m, sendo consideradas úteis as 16

30
linhas centrais. Foram semeadas 16,5 à 20 sementes de soja por metro
linear no sentido norte-sul. Em cada parcela experimental foram semeadas
quantidades diferentes de sementes inoculadas com C. truncatum, para
obtenção de níveis iniciais de incidência do patógeno de 0,8%, 1,6% e 2,4%.
Assim 20 sementes foram semeadas em 1 linha da parcela com 0,8% de
incidência, 40 sementes foram distribuídas em 2 linhas da parcela com 1,6%
de incidência e 60 sementes foram semeadas em 3 linhas da parcela com
2,4% de incidência. Todas as sementes foram distribuídas no centro de cada
parcela, onde cada linha correspondeu a 1 metro (m) linear (Figura 1).
As sementes não inoculadas com o patógeno foram tratadas com
fungicidas e inseticidas recomendados pelo MAPA (Tabela 2) e
posteriormente foi aplicado Bradyrhizobium elkanni na dosagem de 600
g250-1 kg de sementes (Biomax Premium, 7,2 x 109), 6 mL de água kg-1 de
semente na concentração de 10% de açúcar, em seguidas as sementes
foram secas na sombra em temperatura ambiente, por 1 h.
O desbaste das plantas foi realizado aos 23 dias após a emergência
da soja, deixando aproximadamente 15 plantas por m linear. Esse desbaste
não foi efetuado nas linhas contendo as sementes inoculadas com o
patógeno.
As pragas e plantas indesejáveis foram controladas com produtos
fitossanitários registrados no MAPA, ao longo do ciclo das culturas de milho
e soja (Tabela 2). Na ausência de precipitação pluvial e havendo a
necessidade de umidade no solo foi realizada irrigação por aspersão com
pivô central, no início da manhã ou ao final da tarde.
Utilizando-se GPS Topcon Hiper Lite configurado em datum WGS
84, zona UTM 21 Sul, modo RTK, foi demarcada em cada parcela uma
grade com 112 pontos amostrais (Figura 2), sendo cada ponto representado
por duas plantas, uma para avaliação da incidência da antracnose no campo
e a outra para a coleta de folíolos para avaliar a incidência do patógeno em
laboratório.

31
Figura 2. Malhas amostrais da área experimental. Parcelas com 0,8% (A),
1,6% (B) e 2,4% (C) de sementes inoculadas com C. truncatum.

32
Foram coletados semanalmente 112 folíolos do baixeiro das plantas
dos respectivos pontos demarcados (Tabela 3), os quais foram
armazenados em sacos plásticos, identificados e encaminhados ao
laboratório de fitopatologia da UFMT. No laboratório os folíolos foram
analisados pelo método de câmara úmida. Primeiramente, os folíolos foram
submetidos a desinfestação superficial com a lavagem em água corrente,
em seguida preparou-se as placas de Petri esterilizadas colocando 2 folhas
de papel filtro nas mesmas e umedecendo-as com água destilada para a
acomodação de cada folíolo de soja na placa previamente identificada. As
placas foram incubadas em regime alternado de 12 h luz / 12 h escuro com
temperatura de 22ºC (± 2ºC). E as leituras da presença ou ausência de C.
truncatum foram realizadas aos 7, 14 e 21 dias após a incubação, com
auxílio de microscópio estereoscópio.
Tabela 3. Datas de coletas de folíolos de soja para incubação por meio do
método da câmara úmida.
Atividade Data
1ª Coleta de folíolos da parcela com 2,4% de C. truncatum 23/12/2009 1ª Coleta de folíolos da parcela com 1,6% de C. truncatum 07/01/2010 1ª Coleta de folíolos da parcela com 0,8% de C. truncatum 14/01/2010 2ª Coleta de folíolos da parcela com 2,4% de C. truncatum 22/01/2010 2ª Coleta de folíolos da parcela com 1,6% de C. truncatum 29/01/2010 2ª Coleta de folíolos da parcela com 0,8% de C. truncatum 05/02/2010 3ª Coleta de folíolos da parcela com 2,4% de C. truncatum 12/02/2010 3ª Coleta de folíolos da parcela com 1,6% de C. truncatum 18/02/2010
No campo realizou-se o monitoramento semanal da incidência da
antracnose nos pontos amostrais. Os primeiros sintomas da antracnose
foram visualizados apenas nos cotilédones das plântulas da região do
inóculo (linhas centrais de cada parcela), por meio de manchas deprimidas
com os bordos salientes, de coloração castanho-escuras e queda dos
cotilédones.
Após a visualização de sintomas dessa doença em pontos amostrais
fora da região do inóculo foram efetuadas 7 avaliações de incidência nas
plantas referentes aos 336 (112 em cada parcela) pontos demarcados

33
(Tabela 4). Contabilizando o número de folhas sadias e doentes de cada
ponto amostral.
Tabela 4. Datas de avaliações de incidência da antracnose no campo com
suas respectivas datas.
Atividade Data
1ª Avaliação de incidência 29/12/2009 2ª Avaliação de incidência 05/01/2010 3ª Avaliação de incidência 12/01/2010 4ª Avaliação de incidência 19/01/2010 5ª Avaliação de incidência 26/01/2010 6ª Avaliação de incidência 02/02/2010 7ª Avaliação de incidência 16/02/2010
Ao final do ciclo da soja realizou-se a colheita em cada ponto
amostral e em seguida efetuou-se a debulha manual. Após a secagem das
sementes à sombra, elas foram homogeneizadas e submetidas ao teste de
sanidade „blotter test‟ com restrição hídrica considerando a metodologia de
Brasil (2009), no Laboratório de Fitopatologia da UFMT.
Na análise sanidade, foram semeadas 25 sementes por placa com
substrato umedecido com manitol a – 0,6 MPa e ágar a 0,2%, para cada
ponto amostral. As placas de Petri com as sementes permaneceram na sala
de crescimento com temperatura de 22ºC (± 2ºC), e fotoperíodo de 12 h luz
e 12 h escuro. A leitura de incidência de C. truncatum nas sementes foi
realizada aos 4 e 7 dias após a incubação, com auxílio de microscópio
estereoscópio e quando necessário utilizou-se o microscópio óptico, sendo
retiradas as sementes com a presença do patógeno na primeira leitura.
Durante o ciclo da soja foi realizado o monitoramento dos seguintes
elementos meteorológicos: temperatura do ar (°C), umidade relativa (%),
radiação solar (w m-2), duração do molhamento foliar (h), intensidade do
molhamento foliar (kpa), precipitação (mm) e direção do vento, por meio de
sensores instalados em uma estação meteorológica automática (Watch Dog
2000 Series Weather Stations, USA). Essa estação foi instalada entre a
parcela com 1,6% e 2,4% de sementes inoculadas com C. truncatum e, os

34
dados foram armazenados a cada 30 minutos. Três sensores de
molhamento foliar foram fixados no tripé da estação a 0,3 m do solo com
inclinação de 45° (um em cada pé), os demais sensores foram instalados à 2
m de altura.
2.2.1 Análise de dados
Com os dados da câmara úmida das folhas efetuou-se o cálculo da
área abaixo da curva do progresso da incidência (AACPI), conforme
Campbell e Madden (1990).
1
11 )(
2
n
i
iiii tt
yyAACPI
Em que iy é proporção da doença na i-ésima observação; it é tempo em dias
na i-ésima observação e n é número total de avaliações.
De posse dos dados meteorológicos efetuou-se a média diária de
cada variável climática dos respectivos 48 valores de cada dia, exceto para a
radiação solar global a qual a média foi obtida com 24 valores de w.m-2,
entre às 6:00 até as 18:00 h.
O padrão espacial da transmissibilidade de níveis de C. truncatum
em sementes de soja foi analisado com os resultados obtidos nos pontos
amostrais por meio das análises de sequências „doublet‟ (Bergamin Filho et
al., 2004) e „run‟ (Campbell e Madden, 1990), utilizando dados binários
referente ao teste de sanidade das sementes.
Na análise „run‟ foram atribuídas na direção perpendicular da linha
de semeadura, para as sequências de sementes sadias o valor zero (0) e
para as sementes infectadas e contaminadas com C. truncatum o valor um
(1). Com estas informações, contabilizou-se o número de „runs‟ observado,
em seguida foi calculado o número esperado de „runs‟ E(R)=1+2m(N-m)/N e
o seu desvio padrão s(R)=2m(N-m)[2m(N-m)–N]/[N2(N-1)]¹/². Em que: R:
número de „runs‟; m: número de plantas doentes; N: número de plantas

35
avaliadas na linha. Efetuou-se um teste de normalidade pela equação:
ZR=[R+0,5-E(R)]/s(R).
Foi considerado padrão espacial agregado quando o número
observado de ‟runs‟ apresentou valores menores que o esperado. Para ZR
classificou-se como agregação, quando o seu valor foi < -1,64 (P=0,05)
(Gibbons, 1976).
Na análise de „doublet‟ foram caracterizadas na direção
perpendicular das linhas de semeadura a seqüência de sementes sadias (0)
e sementes associadas (1), com estas informações, contabilizou-se o
número de „doublets‟ observado, posteriormente foi calculado o número
esperado de „doublets‟ E(D)=m(m-1)/N e sua variância σ2(D)=[m(m-1)[N(N-
1)+(2N(m-2)+N(m-2)(m-3)-(N-1)m(m-1)]/N2(N-1)]. Em que: D: número de
„doublet‟; m: número de plantas doentes; N: número de plantas na linha. Foi
determinado também o s(D) desvio-padrão. Calculou-se um teste de
normalidade pela equação: ZD=[D+0,5-E(D)]/s(D).
Considerou-se padrão espacial agregado quando o valor observado
apresentou valores maiores que o esperado e o valor de ZD > 1,64 (P=0,05).
A distribuição espacial padrão do C. truncatum nos folíolos e
sementes de soja nas parcelas amostrais foi avaliada pela análise do índice
de dispersão binomial (D) (Madden e Hughes, 1995), pela função K de
Ripley para análise espacial pontual padrão (Ripley, 1981), e o teste de
Monte Carlo para determinar a distância média mínima entre plantas com o
patógeno. As áreas avaliadas foram divididas em quadrats de 2x2, 2x3, 2x4,
2x7, 3x 2, 3x3, 3x 4, 3x7, 4x2, 4x3, 4x 4, 4x7 plantas para as análises do
índice de dispersão (D). A incidência de C. truncatum nos folíolos e
sementes de soja foi estimada por: P = ΣXi/nN, onde Xi é o número de
plantas com incidência de C. truncatum em folíolo ou sementes de soja no i-
ésimo quadrat, n é o número de plantas por quadrat e N é o número total de
quadrats (Madden e Hughes, 1995). A partir destes dados, calculou-se a
variância binomial (Vbin) foi determinada por Vbin = p(1-p)/n e a variância
observada (Vobs) foi calculada por (Hughes et al., 1996)

36
)1²(/)²(1
NnnpxVi
N
iobs . O índice D foi obtido pela equação D = Vobs/
Vbin (Gottwald et al., 1996). Para cada avaliação o valor de D foi aplicado o
teste do x² (chi-quadrado) a 5% de significância. Valores de D iguais a 1
indicam padrão distribuição aleatória e quando os valores de D
significativamente maiores que 1 indicam distribuição agregada (Madden e
Hughes, 1995).
A análise por quadrats foi implementada com o índice de dispersão
binomial, pois a distribuição de Poisson deve ser aplicada somente para
dados binários com a freqüência de doença menor que 20% (Bergamin Filho
et al., 2007).
A análise de padrões pontuais foi efetuada pela função K (h) de
Ripley, esta análise adota estratégia diferente para evitar a escolha subjetiva
dos tamanhos dos quadrats e verifica a densidade de plantas doentes em
cada parcela, considerando a variância de todas as distâncias radiais (h)
possíveis a partir de todas as plantas sintomáticas. A função K de Ripley foi
calculada pela fórmula K (h) = E -1, em que é a densidade dos eventos ou
o número médio de plantas com folíolos ou sementes com incidência de C.
truncatum por unidade de área, e E é o número de plantas com folíolos ou
sementes com incidência de C. truncatum dentro de uma unidade de área
definida pelo a distância h, a partir de uma planta doente escolhida
aleatoriamente. A função K (h) foi estimada por Ripley (1981), Spóstio et al.,
(2007):
n
EjEiIhK
n
i
n
j h1 1
1 ),()( para i # j e h > 0 (1)
Em que Ih é o indicador da função e n é o número de eventos. Ih é 1 se estiver dentro Ej a distância h do Ei, se não está dentro Ej a distância h do Ei, então Ih é 0.
Quando ocorre independência, o pradrão é aleatório no espaço e K
(h) = πh2. Para padrão agregado espera-se que K πh2>(h); e para padrão
regular, K πh2<(h).

37
A normalização da função K de Ripley (L (h)) facilita a interpretação
gráfica (expressão 2), uma vez que o gráfico da função é uma linha
horizontal sob aleatoriedade espacial; valores positivos indicam atração
espacial (agregação) e os valores negativos indicam repulsão ou
regularidade (Kenkel, 1998, Spósito et al., 2007).
L(h) = h
h
hK
²
)(
(2)
As simulações dos envelopes obtidos por Monte Carlo foram
construídas para funções K ou L a fim de testar a hipótese de aleatoriedade
espacial. Sendo plotado os valores de L no eixo das ordenadas e os valores
da distância no eixo das abcissas. Conforme Kenkel (1998), os valores
situados dentro do envelope representam as regiões em que a hipótese de
aleatoriedade espacial não está descartada.
A função K de Ripley foi calculada com distância de 3,35 m,
representando aproximadamente a metade da menor distância de cada
parcela amostral. Kenkel (1998) afirma que quando o gráfico gerado pela
função K de Ripley for uma linha horizontal sob aleatoriedade espacial e
apresentar valores positivos, indicou-se agregação, a presença de valores
negativos indicam repulsão ou regularidade (Kenkel, 1988).
As análises de quadrats counts, distância mínima média, função K
de Ripley e os mapas de distribuição da incidência de C. truncatum em
folíolos e nas sementes de soja foram realizadas no software R. Sendo que
os dados foram avaliados com adaptação do método na análise da função K
de Ripley e seu respectivo envelope utilizando o pacote “splancs” do
software R (Rowlingson e Diggle, 1993).
2.3 Resultados e Discussão
No decorrer do desenvolvimento da cultura da soja verificou-se a
influência do patógeno sobre o hospedeiro, pois ocorreu menor
desenvolvimento vegetativo nas plantas provenientes de sementes

38
inoculadas por C. truncatum, quando comparado com as demais plantas
(Figura 3 A e B). Goulart (2007) e Cassetari Neto e Machado (2009),
observaram que plântulas originadas de sementes infectadas apresentam
necrose dos cotilédones, que pode se estender para o hipocótilo, causando
tombamento ou o estabelecimento de plantas pouco vigorosas.
A antracnose influenciou também em distúrbios fisiológicos visíveis
nas plantas, principalmente, no final do ciclo, em função da indução das
plantas à retenção foliar, hastes e vagens verdes, as quais apresentaram
maturação desuniforme entre as plantas e nas próprias plantas, além de
contribuir para redução e ou não enchimento no número de vagens (Figura 3
C, D, E e F).
Figura 3. Desenvolvimento vegetativo das plantas de soja provenientes de
sementes inoculadas com C. truncatum (A) e plantas próximas a fonte no inóculo (B) aos 45 dias após a semeadura. Distúrbios fisiológicos causados por C. truncatum (C, D, E e F) aos 134 dias após a semeadura.
As condições do tempo foram favoráveis, principalmente em função
da distribuição de chuvas durante o ciclo da soja (Figura 4), contribuindo
com o processo de colonização e disseminação do patógeno e
consequentemente, com a manifestação da doença, apresentando aumento
crescente da antracnose ao longo do tempo.

39
Observou-se que a média diária da irradiação solar global oscilou,
apresentando valor mínimo de 124,96, máximo de 712,13 e média de 472,75
w m-2 entre as avaliações de incidência da antracnose no campo, período
compreendido dos 69 aos 118 dias após a semeadura da soja (Figura 4 D).
Este resultado contribuiu para a dispersão do C. truncatum, pois conforme
Vale et al. (2004), a irradiação solar interferiu diretamente na fotossíntese do
hospedeiro, na evapotranspiração e na sobrevivência do patógeno.
Com relação a média diária da duração e intensidade do
molhamento foliar, os valores medidos foram de 8 horas e vinte minutos e
7,26 kpa, respectivamente, durante o ciclo da cultura da soja (Figura 4 E e 4
F).
As condições climáticas favoreceram a infecção, colonização e
disseminação da antracnose, principalmente a partir da visualização dos
sintomas, pois o hospedeiro e o patógeno possivelmente foram influenciados
pelo microclima favorável. Conforme Almeida et al. (2005), a maior
intensidade da antracnose nos cerrados pode ser atribuída à elevada
precipitação e às altas temperaturas.
Durante o ciclo da soja, a média diária da temperatura do ar foi de
28,39°C variando de 24,19°C a 31,82°C (Figura 4 A), a média diária da
umidade relativa do ar foi 76,11%, com média diária mínima de 59,22% e
média diária máxima de 96,66% (Figura 4 B). A precipitação associada com
a irrigação durante esse período foi 1563,10 mm (Figura 4 C).
De acordo com Vale et al. (2004), a germinação de esporos e a
infecção pela maioria dos fungos dependem da combinação da temperatura
e duração da umidade relativa do ar próxima à saturação, ou da duração do
molhamento foliar. Por outro lado, o crescimento da lesão é dependente
principalmente da temperatura.
Estudos realizados por Hamawaki et al. (2002), afirmam que as
variedades de soja cultivadas em temperaturas, com média anual acima de
30 ºC, e chuvas no período da maturação à colheita, foram predispostas a
infecções por C. dematium var. truncata.

40
Tem
epar
tua
Méd
ia D
iári
a (°
C)
15
20
25
30
35
40
Nov/09 Dez/09 Jan/10 Fev/10 Mar/10
Ch
uva
+ i
rrig
ação
(m
m)
0
20
40
60
80
100
120
140
160
Nov/09 Dez/09 Jan/10 Fev/10 Mar/10
Rad
iaçã
o S
ola
r M
édia
diá
ria
(W/m
²)
0
100
200
300
400
500
600
700
800
Nov/09 Dez/09 Jan/10 Fev/10 Mar/10
Inte
nsi
dad
e M
olh
amen
to F
oli
ar D
iári
o (
Kpa)
0
2
4
6
8
10
12
14
16
Nov/09 Dez/09 Jan/10 Fev/10 Mar/10
Intensidade Molhamento Foliar (kpa)
Dura
ção M
olh
amen
to F
oli
ar (
h)
0
5
10
15
20
25
30
Nov/09 Dez/09 Jan/10 Fev/10 Mar/10
Figura 4. Médias diárias de temperatura do ar (A), umidade relativa do ar
(B), somatória do acumulado da precipitação mais irrigação diária (C), radiação solar global (D), intensidade (E) e duração do molhamento foliar (F), de outubro de 2009 a março de 2010.
Observou-se que as sementes inoculadas com o fungo,
apresentaram seus primeiros sintomas nos cotilédones, porém após a queda
dos mesmos, o patógeno permaneceu em estágio de latência (planta) e
saprofítico (solo). Sendo que os sintomas da doença foram observados
novamente de maneira visual a partir do final do florescimento tanto nos
pontos de inóculo quanto em alguns pontos amostrais das parcelas. De
E
D
A B
F
C
E

41
acordo com Yorinori et al. (2010), à medida que a planta se desenvolve, o
fungo associa-se às células da epiderme, mantendo-se em estado latente
sem causar danos aparente e, dependendo do cultivar, das condições
climáticas e dos níveis de fertilidade do solo, a doença pode se manifestar
com maior ou menor severidade.
A parcela contendo 1,6% de sementes inoculadas com C. truncatum
apresentou nas avaliações realizadas, exceto na primeira e última avaliação,
maior média de incidência de antracnose no campo quando comparado aos
demais níveis de inóculo (Figura 5), porém apresentou menor média da
transmissibilidade semente-semente (Tabela 5). Isso ocorreu,
provavelmente, em função da maior queda de folhas e ou dos folíolos das
plantas provenientes da parcela contendo 2,4% de sementes inoculadas
com C. truncatum, pois nas avaliações de incidência em campo foram
contabilizadas apenas as folhas completas.
Figura 5. Valores médios da incidência da antracnose no campo, em
parcelas semeadas com 0,8, 1,6 e 2,4% de sementes inoculadas com C. truncatum.

42
Observou-se nos níveis 0,8 e 1,6% de sementes inoculadas, valores
próximos entre os dados binários, entretanto o nível 2,4% de sementes
inoculadas apresentou menor valor no coeficiente de variação (Tabela 5),
pois apenas nas sementes de 10 (8,93%) dos 112 pontos amostrais não
foram detectadas a presença de C. truncatum (Figura 6 L).
Tabela 5. Estatística descritiva dos dados binários referente a transmissibilidade de C. truncatum para sementes de soja em parcelas semeadas com 0,8, 1,6 e 2,4% de sementes inoculadas com C. truncatum.
Índices Estatísticos
Total de sementes inoculadas com C. truncatum
0,8% 1,6% 2,4%
Média 0,85 0,76 0,91 Desvio padrão 0,36 0,43 0,28 Variância 0,13 0,18 0,08 CV% 42,49 56,65 31,45
Com relação a incidência do patógeno nos folíolos, as parcelas
amostrais apresentaram incidência crescente de C. truncatum nos folíolos de
soja, principalmente em função da porcentagem do inóculo inicial e da época
de coleta dos folíolos. Na parcela com 0,8% de sementes de soja inoculadas
com o patógeno, as incidências foram 49,11% e 57,14% (Figura 6 A e B). Na
parcela com 1,6% de sementes inoculadas com o fungo, as incidências
foram 22,32%, 54,46% e 73,21% (Figura 6 C, D e E). E na parcela com 2,4%
de sementes inoculadas com C. truncatum, as incidências foram 17,86%,
47,32% e 49,11% (Figura 6 F, G e H).

43
Figura 6. Mapas de distribuição da incidência de C. truncatum em folíolos,
em cada planta avaliada das parcelas com 0,8% (A, B), 1,6% (C, D, E) e 2,4% (F, G, H) de sementes inoculadas, com suas respectivas datas de coletas. Incidência do patógeno nas sementes para os inóculos 0,8, 1,6 e 2,4% respectivamente (I, J, L).
44
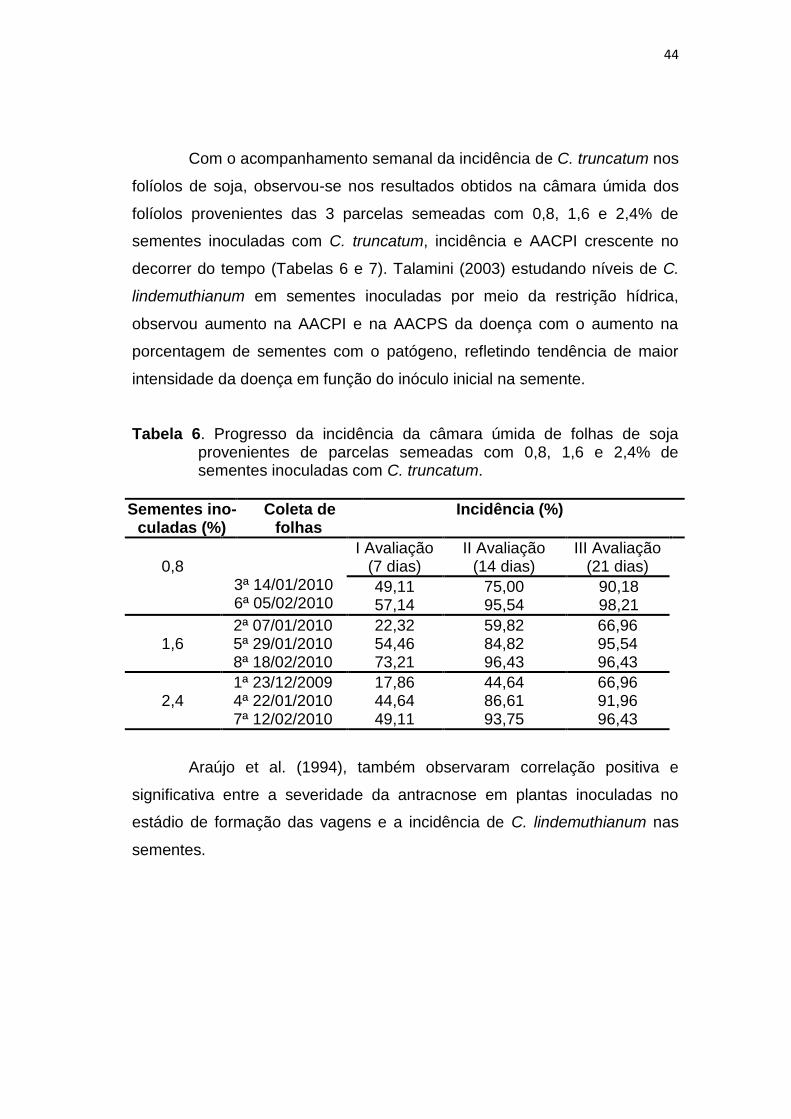
Com o acompanhamento semanal da incidência de C. truncatum nos
folíolos de soja, observou-se nos resultados obtidos na câmara úmida dos
folíolos provenientes das 3 parcelas semeadas com 0,8, 1,6 e 2,4% de
sementes inoculadas com C. truncatum, incidência e AACPI crescente no
decorrer do tempo (Tabelas 6 e 7). Talamini (2003) estudando níveis de C.
lindemuthianum em sementes inoculadas por meio da restrição hídrica,
observou aumento na AACPI e na AACPS da doença com o aumento na
porcentagem de sementes com o patógeno, refletindo tendência de maior
intensidade da doença em função do inóculo inicial na semente.
Tabela 6. Progresso da incidência da câmara úmida de folhas de soja provenientes de parcelas semeadas com 0,8, 1,6 e 2,4% de sementes inoculadas com C. truncatum.
Sementes ino-culadas (%)
Coleta de folhas
Incidência (%)
0,8
3ª 14/01/2010 6ª 05/02/2010
I Avaliação (7 dias)
II Avaliação (14 dias)
III Avaliação (21 dias)
49,11 75,00 90,18 57,14 95,54 98,21
1,6
2ª 07/01/2010 5ª 29/01/2010 8ª 18/02/2010
22,32 59,82 66,96 54,46 84,82 95,54 73,21 96,43 96,43
2,4
1ª 23/12/2009 4ª 22/01/2010 7ª 12/02/2010
17,86 44,64 66,96 44,64 86,61 91,96 49,11 93,75 96,43
Araújo et al. (1994), também observaram correlação positiva e
significativa entre a severidade da antracnose em plantas inoculadas no
estádio de formação das vagens e a incidência de C. lindemuthianum nas
sementes.

45
Tabela 7. Valores da área abaixo da curva do progresso da incidência
(AACPI) do patógeno para parcelas semeadas com 0,8%, 1,6% e 2,4% de sementes inoculadas com C. truncatum.
Data final da avaliação
Porcentagem de sementes com C. truncatum
0,8% 1,6% 2,4%
AACPI
13/01/2010 - - 609,38 28/01/2010 - 731,25 - 04/02/2010 1012,50 - - 12/02/2010 - - 1084,38 19/02/2010 - 1118,75 - 26/02/2010 1212,50 - - 05/03/2010 - - 1165,63 11/03/2010 - 1268,75 -
Com relação às análises de seqüências ordinárias ‘runs‟, foi
detectada 85,71, 100 e 100% nas linhas de semeadura para as parcelas
com 0,8, 1,6 e 2,4% de sementes inoculadas, respectivamente,
caracterizando padrão espacial aleatório de C. truncatum nas sementes de
soja (Tabela 8). Os inóculos testados não apresentaram valores
significativos de ZR (P=0,05), demonstrando a existência de aleatoriedade de
C. truncatum nas sementes de soja. Conforme Andrade e Michereff (2000),
„ordinary runs‟ é uma análise unidimensional e apenas examina a estrutura
dos dados dentro e entre linhas.
Pelas análises de seqüências ordinárias „doublet‟, o padrão aleatório
na direção perpendicular das linhas de semeadura representou 57,14%,
85,71% e 71,43% respectivamente para as parcelas com 0,8%, 1,6% e 2,4%
de sementes inoculadas. Na média de todas as linhas de cada parcela
constatou-se padrão agregado apenas para o inóculo 0,8% (Tabela 9).
Observou-se que o arranjo espacial do patógeno pela análise „run‟,
forneceu informações semelhantes às da análise de „doublet‟, exceto para a
parcela com 0,8% de sementes inoculadas.

46
Tabela 8. Análise de arranjo espacial da transmissibilidade em parcelas com 0,8, 1,6 e 2,4% de sementes de soja inoculadas com C. truncatum, analisado por seqüências ordinárias „runs‟.
Sementes inoculadas com 0,8 % Teste ‘run’ P=0,05
Linha N m R E(R) σ2(R) s(R) ZR Decisão
1 16 15 2 2,88 0,11 0,33 -1,13 casual. 2 16 11 7 7,88 2,69 1,64 -0,23 casual.
3 16 12 2 7 2 1,41 -3,18 agreg.
4 16 16 1 1 0 0,00 0,00 casual.
5 16 15 3 2,88 0,11 0,33 1,89 casual.
6 16 13 5 5,88 1,26 1,12 -0,33 casual.
7 16 13 7 5,88 1,26 1,12 1,45 casual.
Total 112 95 *3,86 *4,77 7,43 5,96 *-0,22 casual.
Sementes inoculadas com 1,6 %
Linha N m R E(R) σ2(R) s(R) ZR Decisão
1 16 13 6 5,88 1,26 1,12 0,56 casual. 2 16 9 9 8,88 3,61 1,90 0,33 casual.
3 16 14 5 4,50 0,58 0,76 1,31 casual.
4 16 16 1 1 0,00 0,00 0,00 casual.
5 16 15 3 2,88 0,11 0,33 1,89 casual.
6 16 9 7 8,88 3,61 1,90 -0,72 casual.
7 16 9 6 8,88 3,61 1,90 -1,25 casual.
Total 112 85 *5,29 *5,84 12,78 7,92 *0,30 casual.
Sementes inoculadas com 2,4 %
Linha N m R E(R) σ2(R) s(R) ZR Decisão
1 16 16 1 1 0,00 0,00 0,00 casual.
2 16 15 3 2,88 0,11 0,33 1,89 casual.
3 16 16 5 1 0,00 0,00 0,00 casual.
4 16 16 1 1 0,00 0,00 0,00 casual.
5 16 14 3 4,5 0,58 0,76 -1,31 casual.
6 16 13 7 5,88 1,26 1,12 1,45 casual.
7 16 12 6 7 2,00 1,41 -0,35 casual.
Total 112 102 *3,71 *3,32 3,95 3,63 *0,24 casual.
*Valores médios; N: nº plantas; m: plantas doentes; R: „runs‟; E(R): „runs‟ esperados; σ2: variância; s: desvio-padrão; casual.: casualizado; agreg.: agregado.
Outros estudos aplicando esses testes como o de Ferreira et al.
(2009), verificaram para a mancha manteigosa em cafeeiro arranjo espacial
aleatório em 100% das linhas de plantio pelas análises de seqüências
ordinárias „runs‟ e predominância do arranjo espacial aleatório, em 62,5%

47
das linhas de plantio, com relação às análises de seqüências ordinárias
„doublet‟, sendo este padrão mais representativo para esclarecer a dispersão
espacial da doença.
Tabela 9. Análise de arranjo espacial da transmissibilidade em parcelas com 0,8, 1,6 e 2,4% de sementes de soja inoculadas com C. truncatum, analisado por seqüências ordinárias „doublet‟.
Sementes inoculadas com 0,8% Teste de ‘doublet’ P=0,05
Linha N m D E(D) σ2(D) s(D) ZD Decisão
1 16 15 14 13,13 0,11 0,33 4,16 agreg. 2 16 11 8 6,88 0,86 0,93 1,75 agreg. 3 16 12 11 8,25 0,69 0,83 3,92 agreg. 4 16 16 15 15 0,00 0,00 0,00 casual. 5 16 15 13 13,13 0,11 0,33 1,13 casual. 6 16 13 10 9,75 0,49 0,70 1,07 casual. 7 16 13 9 9,75 0,49 0,69 -0,36 casual.
Total 112 95 *11,43 *10,84 2,74 3,81 *1,67 agreg.
Sementes inoculadas com 1,6 %
Linha N m D E(D) σ2(D) s(D) ZD Decisão
1 16 13 10 9,75 0,49 0,70 1,07 casual. 2 16 9 5 4,50 1,05 1,03 0,98 casual. 3 16 14 11 11,38 0,28 0,53 0,23 casual. 4 16 16 15 15,00 0,00 0,00 0,00 casual. 5 16 15 13 13,13 0,11 0,33 1,13 casual. 6 16 9 5 4,50 1,05 1,03 0,98 casual. 7 16 9 6 4,50 1,05 1,03 1,95 agreg.
Total 112 85 *9,29 *8,96 4,03 4,64 *0,91 casual.
Sementes inoculadas com 2,4%
Linha N m D E(D) σ2(D) s(D) ZD Decisão
1 16 16 15 15 0,00 0,00 0,00 casual. 2 16 15 13 13,13 0,11 0,33 1,13 casual. 3 16 16 15 15 0,00 0,00 0,00 casual. 4 16 16 15 15 0,00 0,00 0,00 casual. 5 16 14 12 11,38 0,28 0,53 2,11 agreg. 6 16 13 11 9,75 0,49 0,70 2,51 agreg. 7 16 12 8 8,25 0,69 0,83 0,30 casual.
Total 16 102 *12,71 *12,50 1,57 2,39 *0,86 casual.
*Valores médios; N: nº plantas; m: plantas doentes; D: „doublet‟; E(D): „doublet‟ esperados; σ2: variância; s: desvio-padrão; casual.: casualizado; agreg.: agregado.

48
Em análise de seqüências ordinárias para descrição do arranjo
espacial do vira-cabeça do fumo (Groundnut ringspot virus, GRSV), Silva et
al. (2001), observaram também a predominância do arranjo aleatório de
plantas doentes e baixo porcentual de agregação nas linhas de plantios.
Estes autores concluíram que os arranjos espaciais de plantas doentes são
influenciados pela interação de vários fatores, incluindo o ambiente e,
possíveis vetores.
Embora as descrições estatísticas não propiciem, por si próprias,
uma explanação dos mecanismos responsáveis pelos arranjos espaciais
(Campbell e Madden, 1990), arranjos agregados de plantas doentes indicam
que houve disseminação planta a planta ou que o inóculo estava
espacialmente agregado (Campbell et al., 1984).
Quanto à disseminação, arranjos agregados são mais
caracteristicamente associados com fontes de inóculo próximas, ou mesmo,
dentro de populações do hospedeiro (Burdon, 1987).
Notou-se visualmente de forma semelhante nas análises „run‟ e
„doublet‟, que o padrão espacial da transmissibilidade semente para semente
de C. truncatum não ficou definido com clareza, este resultado é explicado
pelas condições climáticas favoráveis durante o ciclo da soja juntamente
com a agressividade do patógeno, favorecendo ao agrupamento
praticamente contínuo da incidência de C. truncatum, especialmente na
parcela com 2,4% de sementes inoculadas (Figura 6 I, J e L).
Os resultados obtidos pelas análises „run‟ e „doublet‟, neste caso não
foram suficientes para determinar o padrão espacial do C. truncatum nas
sementes de soja, pois ambos os métodos apresentam a limitação por
considerarem apenas os dados binários, além de não levar em consideração
a localização espacial de cada ponto amostral avaliado. Porém existem
outros métodos para obter o padrão espacial, como análise espacial utilizada
por Jaime-Garcia et al. (2001), Alves et al. (2006), Leal et al. (2010) que
representaram o padrão espacial, e a dependência espacial da doença de
planta em estudo.

49
Ressalta-se que a presença de C. truncatum no hospedeiro não
resultou necessariamente em antracnose, pois o patógeno pode permanecer
latente e se as condições ambientais não forem favoráveis, não há processo
de penetração e colonização do hospedeiro e consequentemente
manifestação da doença. Klingelfuss e Yorinori (2001) verificaram nas
avaliações da infecção latente, em laboratório, pela análise dos tecidos das
folhas e hastes de soja, a ocorrência de C. truncatum e Cercospora kikuchii,
mesmo não havendo sintomas no campo.
A presença do patógeno nos folíolos de soja nos pontos amostrais,
foi considerada como planta doente, onde observou-se que a distância
mínima média de uma planta para outra planta ambas doentes, diminuiu
com o passar do tempo e com o aumento do inóculo. Nas parcelas com
1,6% e 2,4% de sementes inoculadas com C. truncatum na primeira
avaliação de incidência, observou-se que a distância mínima média foi de
1,02 e 1,05 m e na terceira avaliação, foi de 0,50 e 0,63 m, confirmando o
aumento da distribuição da doença no intervalo de 42 e 51 dias (Figura 7 B e
7 C).
Para a distância mínima média das sementes com a incidência do
patógeno nos pontos amostrais, observou-se diferença menor que 0,02 m
entre a parcela com 0,8 e 2,4% de sementes inoculadas com C. truncatum
(Figura 7 D). Essas informações contribuem para o entendimento do
progresso da antracnose, pois o patógeno presente nas folhas aumentou
rapidamente, especialmente no intervalo entre a primeira e segunda
avaliação da coleta de folíolos, para as parcelas de soja semeadas com
0,8%, 1,6% e 2,4% de sementes inoculadas com C. truncatum, observou-se
também na última avaliação de incidência do patógeno 98,21%, 96,43% e
96,43% respectivamente nas parcelas contendo 0,8%, 1,6% e 2,4% de
sementes inoculadas com o fungo (Tabela 6). Consequentemente todas as
plantas avaliadas no campo apresentaram sintomas da antracnose a partir
de 76, 83 e 97 dias após a semeadura, para as parcelas com 1,6%, 2,4% e
0,8% de sementes inoculadas respectivamente.

50
Figura 7. Distância mínima média de plantas de soja com folíolos com
incidência de C. truncatum. Parcela com 0,8% (A), parcela com 1,6% (B), parcela com 2,4% de sementes de soja inoculadas com o patógeno (C) e, distância mínima média dos pontos amostrais com a presença de C. truncatum nas sementes colhidas nos inóculos avaliados 0,8%, 1,6% e 2,4% (D), respectivamente.

51
Pelos dados obtidos pelo índice de dispersão binomial (D),
observou-se predominantemente padrão espacial aleatório de C. truncatum
nos folíolos. Em 47,73% dos quadrats testados para os três níveis de inóculo
avaliados, representaram valor de D maior que 1, porém não foram
significativos ao nível de 5%. Exceto na primeira avaliação, para os quadrats
3x2 e 4x2, para a parcela contendo 0,8% e, na terceira avaliação, para o
quadrat 4x7 na parcela 1,6% de sementes inoculadas com o patógeno,
observou-se padrão espacial agregado, e padrão espacial regular nos
quadrats 3x3 e 4x4 ambos na terceira avaliação dos folíolos da parcela com
2,4% de sementes inoculadas com C. truncatum. Já para as sementes,
ocorreu padrão espacial agregado apenas no quadtat 4x7 da parcela com
0,8% (Tabelas 10, 11 e 12).
Pesquisas realizadas por Ferreira et al. (2009), observaram que o
índice de dispersão não foi satisfatório para a descrição do padrão espacial
da mancha manteigosa. Laranjeira et al. (2004), estudaram o arranjo
espacial das plantas com a clorose variegada dos citros e, com aplicação do
índice D, não foi observado um padrão com explicação biológica em um dos
talhões avaliados, esses autores relataram que não ocorreu tendência clara
para a elucidação do padrão espacial da doença na maioria das avaliações
e, que os resultados variaram em função do tamanho do quadrat.

52
Tabela 10. Incidência e padrão espacial de C. truncatum em folíolos e sementes de soja em combinações de quadrat com diferentes tamanhos para a parcela com 0,8% de sementes de soja inoculadas com C. truncatum.
Câmara úmida folha Semente 21/01/10 12/02/10
Quadrata N° qua- dratsb
P D P D P D
2x2 24 50,00 1,04 ns 59,38 1,07 ns 84,38 1,12 ns 2x3 16 50,00 0,98 ns 59,38 0,73 ns 84,38 1,09 ns 2x4 8 43,75 0,73 ns 64,06 0,99 ns 84,38 1,02 ns 2x7 8 49,11 0,77 ns 57,14 1,50 ns 84,82 1,50 ns 3x2 15 52,22 1,80 * 60,00 1,17 ns 85,56 1,52 ns 3x3 10 52,22 1,59 ns 60,00 0,95 ns 85,56 1,01 ns 3x4 5 45,00 1,28 ns 63,33 1,54 ns 86,67 0,58 ns 3x7 5 50,48 2,15 ns 58,09 1,70 ns 85,71 2,14 ns 4x2 12 50,00 1,82* 59,38 1,33 ns 84,38 1,57 ns 4x3 8 50,00 1,71 ns 59,38 0,64 ns 84,38 1,52 ns 4x4 4 43,75 1,35 ns 64,06 1,15 ns 84,38 1,42 ns 4x7 4 49,17 1,56 ns 57,14 1,07 ns 84,82 2,84*
a Tamanho do quadrat (número de linhas de semeadura x número de plantas na linha),b Quantidade de quadrats construída dentro da área, ns = padrão aleatório e * = padrão agregado.
Tabela 11. Incidência e padrão espacial de C. truncatum em folíolos e sementes de soja em combinações de quadrat com diferentes tamanhos para a parcela com 1,6% de sementes de soja inoculadas com C. truncatum.
Câmara úmida dos folíolos Sementes 14/01/10 05/02/10 25/02/10
Qua-drata
N°qua-dratsb
P D P D P D P D
2x2 24 23,96 0,89 ns 51,04 0,91 ns 72,92 0,98 ns 79,17 1,14 ns 2x3 16 23,96 0,48 ns 51,04 0,84 ns 72,92 0,89 ns 79,17 0,74 ns 2x4 8 26,56 0,63 ns 53,13 0,83 ns 70,31 1,02 ns 81,25 0,47 ns 2x7 8 22,32 0,52 ns 54,46 0,98 ns 73,21 1,22 ns 75,89 1,22 ns 3x2 15 23,33 0,91 ns 52,22 1,13 ns 74,44 0,99 ns 80,00 0,77 ns 3x3 10 23,33 0,75 ns 52,22 0,40 ns 74,44 0,92 ns 80,00 0,43 ns 3x4 5 25,00 0,67 ns 55,00 1,28 ns 73,33 1,36 ns 83,33 0,30 ns 3x7 5 21,91 0,64 ns 55,24 0,83 ns 75,24 1,59 ns 77,14 0,73 ns 4x2 12 23,96 1,06 ns 51,04 1,13 ns 72,92 1,48 ns 79,17 1,15 ns 4x3 8 23,96 0,71 ns 51,04 0,99 ns 72,92 1,42 ns 79,17 0,72 ns 4x4 4 26,56 0,72 ns 53,13 0,92 ns 70,31 1,87 ns 81,25 0,27 ns 4x7 4 22,32 0,88 ns 54,46 1,86 ns 73,21 2,73* 75,89 0,35 ns

53
a Tamanho do quadrat (número de linhas de semeadura x número de plantas na linha),b Quantidade de quadrats construída dentro da área, ns = padrão aleatório e * = padrão agregado.
Tabela 12. Incidência e padrão espacial de C. truncatum em folíolos e sementes de soja em combinações de quadrat com diferentes tamanhos para a parcela com 2,4% de sementes de soja inoculadas com C. truncatum.
Câmara úmida dos folíolos Sementes 30/12/09 29/01/10 19/02/10
Qua-drata
N°qua- dratsb
P D P D P D P D
2x2 24 19,79 0,96 ns 45,83 0,76 ns 48,96 0,99 ns 93,75 1,21 ns 2x3 16 19,79 1,43 ns 45,83 0,76 ns 48,96 0,66 ns 93,75 1,09 ns 2x4 8 20,31 0,87 ns 48,44 1,92 ns 48,74 0,78 ns 98,44 1,02 ns 2x7 8 17,86 1,53 ns 47,32 1,39 ns 49,11 1,34 ns 91,07 1,19 ns 3x2 15 21,11 0,78 ns 46,67 0,59 ns 51,11 0,81 ns 94,44 1,66 ns 3x3 10 21,11 1,25 ns 46,67 0,48 ns 51,11 0,22 R 94,44 1,53 ns 3x4 5 21,67 0,64 ns 51,67 0,90 ns 50,00 0,50 ns 98,33 1,07 ns 3x7 5 19,05 1,08 ns 48,57 1,28 ns 51,43 0,71 ns 91,43 0,73 ns 4x2 12 19,79 1,07 ns 45,83 0,76 ns 48,96 1,04 ns 93,75 0,97 ns 4x3 8 19,79 1,64 ns 45,83 0,96 ns 48,96 0,82 ns 93,75 0,71 ns 4x4 4 20,31 1,13 ns 48,44 2,06 ns 48,44 0,06 R 98,44 1,02 ns 4x7 4 17,86 1,95 ns 47,32 1,37 ns 49,11 2,23 ns 91,07 0,44 ns
a Tamanho do quadrat (número de linhas de semeadura x número de plantas na linha),b Quantidade de quadrats construída dentro da área, ns = padrão aleatório, * = padrão agregado e R = padrão regular.
A caracterização do padrão espacial da doença e do patógeno pode
gerar informações essências na elaboração de estratégias de manejo das
doenças de plantas. Conforme Bergamin Filho et al. (2002), a maioria dos
patógenos e doenças possuem mais de um mecanismo de dispersão, esta
característica aliada aos dados da incidência do patógeno a cada 1,30 m na
linha de semeadura pode explicar o comportamento do C. truncatum, a qual
apresentou distribuição agrupada aleatória na primeira avaliação dos
folíolos, pois houve formação de reboleiras casualizadas, com o passar do
tempo ocorreu aumento do progresso da incidência, e consequentemente a
distribuição do patógeno presente nas sementes ocorreu em mais de 83
pontos dos 112 pontos analisados em todos os inóculos avaliados,

54
apresentado padrão espacial aleatório, predominante para os testes „run‟,
„doublet‟ e índice D.
Por meio dos mapas de distribuição do C. truncatum nas sementes,
observou-se 84,82%, 75,89% e 91,07% de incidência do patógeno para as
parcelas 0,8%, 1,6% e 2,4% de sementes inoculadas respectivamente, com
ocorrência praticamente continua nas proximidades das fontes de inóculo
(Figura 6 I, J e L), comprovando que a disseminação do patógeno ocorreu
principalmente à curtas distâncias.
Com o acompanhamento semanal da incidência de C. truncatum nos
folíolos das folhas de soja, observou-se com os resultados obtidos na
câmara úmida das 3 parcelas semeadas com 0,8, 1,6 e 2,4% de sementes
inoculadas com o patógeno, incidência crescente no decorrer do tempo.
Pelas informações geradas pelas estimativas k (h) e dos envelopes
de confiança para a presença de C. truncatum nos folíolos ou nas sementes
de soja, ocorreu padrão espacial diferenciado, quando comparado com os
demais métodos testados. A vantagem da função K de Ripley quando o
padrão for agregado, em relação aos métodos „run‟, „doublet‟, índice D,
utilizados para detectar o padrão espacial, é que esta função apresentou a
distância inicial a partir de 1,5 m com ocorrência de agrupamento na primeira
avaliação de folíolos da parcela com 2,4% de sementes inoculadas com C.
truncatum. A função K de Ripley emprega a variância dos dados em distintos
raios a partir de cada planta com a presença de C. truncatum seja em folíolo
ou sementes de soja.
Com bases nas informações obtidas pela função K de Ripley e pelo
envelope de Monte Carlo (onde as linhas pontilhadas do envelope de
confiança representam 95% de aleatoriedade espacial), observou-se padrão
espacial agregado significativo para todas as épocas de avaliação dos
folíolos de soja e das sementes colhidas, exceto para a primeira avaliação
de incidência de folíolos da parcela com 1,6% de sementes inoculadas com
C. truncatum (Figura 8).
De forma semelhante Spósito et al. (2007) estudando o padrão
espacial da mancha preta dos citros em pomares comerciais, verificou que

55
apenas os resultados obtidos usando a função K de Ripley indicaram padrão
espacial agregado para todos os pomares e épocas avaliadas. Pesquisas
feitas por Liu et al. (2007), também utilizaram simulação de Monte Carlo para
caracterizar os padrões de mortalidade espaço-temporal de doença em
florestas e obtiveram agregação das árvores doentes.
A função K de Ripley além de analisar o padrão espacial das
doenças de plantas, tem sido aplicada também para detectar o padrão
espacial de espécies de plantas em florestas. Kelly e Meentemeyer (2002)
estudaram a dinâmica espaço-temporal da doença causada por
Phytophthora ramorum em espécie florestal, e utilizaram além de técnicas de
sensoriamento remoto e sistema de informação geográfica, a função K de
Ripley para ajudar a quantificar a dimensão e extensão da aglomeração
espacial da doença. Silva et al. (2008), analisaram a estrutura horizontal
espacial da espécie Eremanthus erythropapus (Candeia), por meio da
função K de Ripley, observaram que as plantas adultas apresentaram um
padrão agregado passando para o aleatório.
A partir da fonte inicial de inóculo pré estabelecida, foi possível notar
melhor aplicação da função K de Ripley para obtenção do padrão espacial
agregado de C. truncatum nos folíolos e sementes, pois neste método foi
possível informar a distância a ser estudada. Assim a função gerou
informações dos dados referentes ao raio dessa distância, e obteve como
resultado, à distância a partir da qual ocorreu agregação, além das
vantagens já citadas. Consequentemente essas informações contribuíram
para corroborar que a disseminação de C. truncatum no campo ocorreu a
curtas distâncias, sendo favorecida pela água proveniente de chuva,
irrigação ou orvalho, e direção predominante do vento.
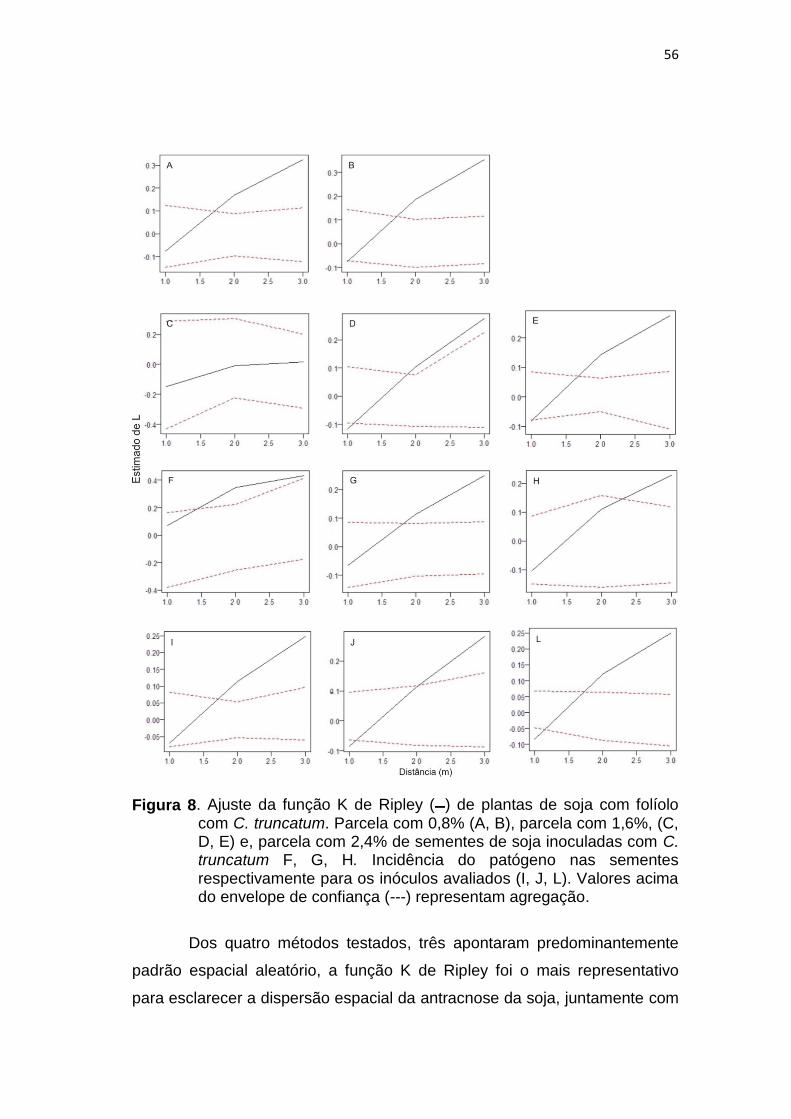
56
Figura 8. Ajuste da função K de Ripley ( ) de plantas de soja com folíolo
com C. truncatum. Parcela com 0,8% (A, B), parcela com 1,6%, (C, D, E) e, parcela com 2,4% de sementes de soja inoculadas com C. truncatum F, G, H. Incidência do patógeno nas sementes respectivamente para os inóculos avaliados (I, J, L). Valores acima do envelope de confiança (---) representam agregação.
Dos quatro métodos testados, três apontaram predominantemente
padrão espacial aleatório, a função K de Ripley foi o mais representativo
para esclarecer a dispersão espacial da antracnose da soja, juntamente com

57
a visualização dos mapas de distribuição do patógeno, observou-se padrão
espacial agregado aleatório de C. truncatum nos folíolos analisados na
primeira avaliação principalmente para as parcelas inoculadas com 1,6 e
2,4% de sementes com C. truncatum (Figura 6 C e F), sendo que em todas
as avaliações a presença do patógeno foi constatada na região da fonte do
inóculo. Notou-se também que com o passar do tempo as plantas vizinhas
foram contaminadas pelo fungo, comprovando a agressividade do patógeno
em estudo.
2.4 Conclusão
O padrão espacial da transmissibilidade de C. truncatum para as
sementes foi agregado para a parcela com 0,8% de sementes inoculadas na
análise „doublet‟, e padrão aleatório para as parcelas com 1,6 % e 2,4% de
sementes de soja inoculadas com C. truncatum, tanto para análise „run‟
como „doublet‟.
A distância mínima média de uma planta doente para outra planta
doente diminuiu com o passar do tempo e com o aumento do inóculo.
Pelas análises do índice de dispersão, o padrão espacial da
antracnose foi predominante aleatório.
Pelos resultados obtidos pela função K de Ripley, o padrão espacial
predominante da incidência de C. truncatum nos folíolos e sementes foi
agregado para as parcelas com 0,8, 1,6 % e 2,4% de sementes de soja
inoculadas com este patógeno.
Constatou-se por meio dos mapas de distribuição de C. truncatum,
padrão espacial agregado aleatório na primeira avaliação de incidência dos
folíolos de soja para os inóculos testados.
2.5 Referências Bibliográficas
ALMEIDA, A.M.R. FERREIRA, L.P.; YORINORI, J.T. et al. Doenças da soja (Glycine Max). In: KIMATI, H. et al. In: Kimat et al. Manual de fitopatologia: Doenças das plantas cultivadas. 4ª Ed. Vol. 2, São Paulo: Agronômica Ceres, 2005, p.571-588.

58
ALVES, M.C.; POZZA, E.A.; MACHADO, J.C.; et al. Geoestatística como metodologia para estudar a dinâmica espaço-temporal de doenças associadas a Colletotrichum spp. transmitidos por sementes. Fitopatologia Brasileira, v.31, p.557-563, 2006. ALVES, V.M.C.; VASCONCELLOS, C.A.; FREIRE, F.M. et al. Milho. In: Ribeiro, A.C.; GUIMARÃES, P.T.G.; ALVARES V.V.H. Comissão de Fertilidade do Solo do Estado de Minas Gerais Recomendações para o uso de corretivos e fertilizantes em Minas Gerais - 5ª Aproximação – Viçosa, MG, 1999, p. 314-316. ANDRADE, D.E.G.T.; MICHEREFF, S.J. Incidência da murcha-de-fusário do tomateiro no Agreste de Pernambuco e determinação do tamanho da amostra para quantificação da doença. Fitopatologia Brasileira, v.25, p.36-41, 2000. ARAÚJO, E.; ZAMBOLIN, L.; VALE, F.X.R.; VIEIRA, C. Correlação entre a severidade da antracnose em vagens de feijoeiro e a transmissão de Colletotrichum lindemuthianum pelas sementes. Turrialba, San José, v.44, p.255-260, 1994. BERGAMIN FILHO, A.; HAU, B.; AMORIM, L.; LARANJEIRA, F. F. Análise espacial e temporal de epidemias de patógenos de solo. In: POLTRONIERI, L. S.; VERZIGNASSI, J. R. Fitossanidade na Amazônia: inovações tecnológicas. Belém, PA: Embrapa Amazônia Oriental, 2007. p.17-36. BERGAMIN FILHO, A.; HAU, B.; AMORIM, L.; JESUS JUNIOR, W.C. de. Análise espacial de epidemias. In: VALE, F.X.R. do; JESUS JUNIOR, W. C. de; ZAMBOLIM, L. (Eds.). Epidemiologia aplicada ao manejo de doenças de plantas. Belo Horizonte: Perfil, 2004, p.195-240. BERGAMIN FILHO, A. HAU, B.; AMORIM, L. et al. Análise espacial de epidemias. Revisão Anual de Patologia de Plantas, Passo Fundo, v.10, 2002, p.155-218. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Manual de Análise Sanitária de Sementes. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. – Brasília: Mapa/ACS, 2009, 200p. BURDON, J.J. Diseases and Plant Population Biology. Cambridge. Cambridge University Press, 1987.
CASSETARI NETO, D.; MACHADO, A.Q. Atenção redobrada. Revista Cultivar – Grandes Culturas. Ano XI, n.119, p.16-20, 2009.

59
CAMPBELL, C.L.; JACOBI, W.R.; POWELL, N.T.; MAIN, C.E. Analysis of disease progression and the randomness of occurrence of infected plants during tobacco black shank. Phytopathology, v.74, p.230-235, 1984. CAMPBELL, C.L.; MADDEN, L.V. Introduction to plant disease epidemiology. New York: J. Wiley, 1990, 532p. EMBRAPA SOJA. Tecnologias de Produção de Soja - Região Central do Brasil, 2009/2010. Londrina: Embrapa Soja: Embrapa Cerrados: Embrapa Agropecuária Oeste, 2008, 262p. FERREIRA, J.B.; ABREU, M.S.; PEREIRA, I.S. Análise da dinâmica, estrutura de focos e arranjo espacial da mancha manteigosa em campo. Ciência e Agrotecnologia, v.33, p.24-30, 2009. Fundação MT. Boletim de pesquisa de soja 2010. Rondonópolis, n°14, 2010, 418p. GIBBONS, J.D. Nonparametric methods for quantitative analysis. New York: Holt, Rinehart & Winston, 463p, 1976. GILLIGAN, C.A. Modeling of soilborne pathogens. Annual Review of Phytopathology, v.21, p.45-64, 1983. GOTTWALD, T.R.; CAMBRA, M.; MORENO, J.M.; CAMARASA, E.; PIQUER, J. Spatial and temporal analysis of citrus tristeza vírus in esatern Spain. Phytopathology, v.86, p.45-55, 1996. GOULART, A.C.P. Semente blindada. Revista Cultivar – Grandes Culturas. (Caderno técnico). Ano IX, n.98, p.08-11, 2007. HAMAWAKI, O.T.; JULIATTI, F.C.; GOMES, G.M.; RODRIGUES, F.A.; SANTOS, V.L.M. Avaliação da qualidade fisiológica e sanitária de sementes de genótipos de soja do ciclo precoce/médio em Uberlândia, Minas Gerais. Fitopatologia Brasileira, v.27, p.201-205, 2002. HUGHES, G. MADDEN, L.V.; MUNKVOLD, G.P. Cluster sampling for disease incidence data. Phytopatology, v.86, p. 132-137, 1996. HUGHES, G.; McROBERTS, N; MADDEN, L.V; NELSON, S.C. Validating mathematical models of plant-disease progress in space and time. IMA Journal of Mathematics Applied in Medicine and Biology, v.14, p.85-112. 1997. JAIME-GARCIA, R.; ORUM, T.V.; FELIX-GASTELUM, R.; TRINIDAD-CORREA, R.; VANETTEN, H.D.; NELSON, M.R. Spatial analysis of Phytophthora infestans genotypes and late blight severity on tomato and

60
potato in the del fuerte valley using geostatistics and geographic information systems. Phytopathology, v.91, p.1156-1165, 2001. KELLY, M., MEENTEMEYER, R.K. Landscape dynamics of the spread of Sudden Oak Death. Photogrammetric Engineering & Remote Sensing. v.68. p.1001–1009, 2002. KENKEL, N.C. Pattern of self-thinning in Jack Pine: testing the random mortality hypothesis. Ecology, v.69, p.1017-1024, 1988. KLINGELFUSS, L.H.; YORINORI, J.T. Infecção latente de Colletotrichum truncatum e Cercospora kikuchii em soja. Fitopatologia Brasileira, v.26, p.201-205, 2001. LARANJEIRA, F.F.; BERGAMIM FILHO, A.; AMORIM, L.; GOTTWALD, T.R. Dinâmica espacial da clorose variegada dos citros em três regiões do estado de São Paulo. Fitopatologia Brasileira, Brasília, v.29, p.56-65, 2004. LEAL, R.M.; BARBOSA, J.C.; COSTA, M.G.; YAMAMOTO, P.T.; DRAGONE, J. Distribuição espacial de Huanglongbing (Greening) em citros utilizando geoestatística. Revista Brasileira de Fruticultura, v.32, p.1-8, 2010.
MADDEN, L.V. HUGHES, G. Plant disease incidence: distributions, heterogeneity, and temporal analysis. Annual Review of Phytopathology, v.33, p. 529-564, 1995.
RIPLEY, B.D. Spatial statistic. New York: John Wiley, 1981. 272p. ROWLINGSON, B.; DIGLE, P. Splancs: spatial point pattern analysis code in S-Plus. Computers and Geoscience, v.19, p. 627-655, 1993. SILVA, M.A.; MELLO, J.M.; SCOLFORO, J.R.S.; CZANCK JÚNIOR, L.; ANDRADE, I.S.; OLIVEIRA, A.D. Análise da distribuição espacial da candeia (Eremanthus erythropappus (DC.) MacLeish) sujeita ao sistema de manejo porta-sementes. Cerne (UFLA), v.14, p.311-316, 2008. SILVA, J.N.; MICHEREFF, S.J.; PIO-RIBEIRO, G.; ANDRADE, G.P. Arranjo espacial do vira-cabeça do fumo em Arapiraca, Estado de Alagoas. Fitopatologia Brasileira, v.26, p.721-725, 2001. SPÓSITO, M.B.; AMORIM, L.; RIBEIRO JUNIOR, P.J.; BASSANEZI, R.B.; KRAINSKI, E.T. Spatial Pattern of Trees Affected by Black Spot in Citrus Groves in Brazil. Plant Disease, v.91, p. 36-40, 2007. TALAMINI, V. Progresso espacial e temporal da antracnose a partir de diferentes níveis de inóculo inicial em sementes de feijoeiro. 2003. 144f.

61
Tese (Doutorado em Fitopatologia) - Universidade Federal de Lavras, Lavras – MG, 2003. VALE, F.X.R.; ZAMBOLIM, L.; COSTA, L.C. et al. Influência do clima no desenvolvimento da doenças de plantas. In: VALE, F.X.R. do; JESUS JUNIOR, W.C. de; ZAMBOLIM, L. (Eds.). Epidemiologia aplicada ao manejo de doenças de plantas. Belo Horizonte: Perffil Editora, 2004, p.47-88. YORINORI, J.T.; YUYAMA, M.M.; SIQUERI, F.V. Doenças da soja. In: Fundação MT. Boletim de Pesquisa de Soja 2010. (Boletim n° 14). Rondonópolis, 2010, p. 218-274.

62
3. ANÁLISE GEOESTATÍSTICA APLICADA À VARIABILIDADE
ESPACIAL DE Colletotrichum truncatum EM SEMENTES DE SOJA
RESUMO - Objetivou-se com este trabalho identificar o melhor modelo
geoestatístico para caracterizar a variabilidade espacial da transmissibilidade
de sementes de soja inoculadas com C. truncatum. O experimento foi
conduzido em condições de campo na safra 2009/10, em 3 parcelas de 9,9 x
10m, com sementes inoculadas com C. truncatum. O inóculo correspondeu a
0,8, 1,6 e 2,4% do total semeado. Foram demarcadas com GPS 3 malhas
amostrais com 112 pontos distanciados a 1,3 m na linha em cada parcela.
Após a colheita e a secagem, efetuou-se a análise sanitária das sementes
pelo método blotter test, referente aos 336 pontos demarcados. De posse
dos dados, 5 modelos de semivariogramas foram ajustados utilizando os
métodos mínimos quadrados ordinários (OLS), mínimos quadrados
ponderados (WLS), máxima verrosimilhança (ML) e máxima verrosimilhança
restrita (REML). A validação cruzada foi empregada para escolha final do
modelo e método do semivariograma. Os mapas de krigagem ilustraram a
transmissibilidade da semente para a semente. Verificou-se estrutura de
dependência espacial da transmissibilidade de C. truncatum via semente. O
melhor modelo de semivariograma foi o esférico e o melhor ajuste foi obtido
pelo método da máxima verossimilhança restrita. Houve alcance de 0,95,
4,03 e 7,05 m para as parcelas com 0,8, 1,6 e 2,4% de sementes inoculadas
respectivamente. O incremento médio diário da dependência espacial foi de
0,65, 2,76 e 4,83 cm para as parcelas 0,8%, 1,6% e 2,4% de sementes
inoculadas, respectivamente.
Palavras - chave: Variação espacial, semivariograma esférico, máxima
verossimilhança restrita.

63
3. GEOSTATISTICAL ANALYSIS APPLIED TO THE SPATIAL
VARIABILITY OF Colletotrichum truncatum IN SOYBEAN SEEDS
ABSTRACT – The aim of this work was to identify the best geostatistical
model to characterize the variability in the transmissibility of soybean
inoculated with C. truncatum. The experiment was conducted under field
conditions in 2009/10 in three plots of 9.9 x 10m with seeds inoculated with
C. truncatum. The inoculum corresponded to 0.8, 1.6 and 2.4% of total sown.
GPS was used to demarcate 3 sampling grids with 112 points spaced 1.3 m
on the line in each plot. After harvesting and drying, it was conducted the
seed health analysis by the blotter test method, referring to the 336 marked
points. By having the data, five models of semivariograms were adjusted
using ordinary least squares (OLS), weighted least squares (WLS), maximum
likelihood (ML) and restricted maximum likelihood (REML) methods. Cross-
validation was used to make the final choice of the semivariogram model and
method. The kriging maps illustrated the transmissibility from seed to seed.
There was spatial dependence structure of the transmissibility of C.
truncatum by seed. The best semivariogram model is the spherical one and
the best fitting was obtained by restricted maximum likelihood. There was a
range of 0.95, 4.03 and 7.05 m for plots measuring 0.8, 1.6 and 2.4% of
inoculated seeds respectively. The average daily increment of spatial
dependence was 0.65, 2.76 and 4.83 cm for plots 0.8%, 1.6% and 2.4% of
inoculated seeds respectively.
Key words: Spatial varation, spherical semivariogram, restricted maximum
likelihood.

64
3.1 Introdução
O Brasil é o segundo maior produtor mundial de soja (Embrapa,
2010), e o estado de Mato Grosso é o maior produtor nacional de soja
(Conab, 2010). Para que a produtividade estabilize ou aumente, é
necessário utilizar dentre outros fatores, sementes sadias com qualidade
fitossanitária satisfatória, pois a semente se constitui em insumo básico para
o sucesso da agricultura. Conforme Talamini et al. (2002), as sementes são
consideradas eficientes meios de disseminação de fitopatógenos e, dentre
esses, destacam-se as espécies pertencentes ao gênero Colletotrichum.
Alves et al. (2006), verificaram aplicação satisfatória da
geoestatística no estudo do progresso espacial e temporal da antracnose do
feijoeiro, tendo em vista ao caráter agregado do inóculo e da doença no
agroecossistema detectado por métodos estatísticos clássicos (Pinto, 2001;
Talamini et al., 2002).
Conforme Vieira (2000) a geoestatística tem por objetivo caracterizar
a variabilidade espacial dos atributos do solo e das plantas e fazer
estimativa, utilizando o princípio da variabilidade espacial, a fim de se
identificar inter relações destes atributos no espaço e no tempo, além de
permitir estudar padrões de amostragem adequada.
No caso da soja, o conhecimento da variabilidade espacial de C.
truncatum encontrado nos campos de produção de sementes, constitui-se
fator fundamental no progresso da antracnose.
Se o inóculo primário, transportado via-semente encontrar ambiente
favorável e o hospedeiro for suscetível, pode-se estabelecer focos iniciais de

65
infecção. Posteriormente, poderá ocorrer o progresso da doença no espaço
e no tempo, a partir da fonte de inóculo secundário, produzido nas lesões
das plantas infectadas (Campbell e Madden, 1990; Tanaka e Menten, 1992).
Após a queda dos cotilédones e prostamento de pecíolos com sintomas da
antracnose em soja, forma-se o inóculo policíclico. Dependendo das
condições climáticas pode haver o progresso da epidemia na lavoura,
ocorrendo assim, a transmissão semente-planta e da planta-semente.
Objetivou-se nesta pesquisa, caracterizar a estrutura de variação
espacial da variabilidade espacial da transmissibilidade de sementes de soja
inoculadas com C. truncatum e determinar o melhor modelo e método de
semivariograma que represente a transmissibilidade de C. truncatum
semente-semente.
3.2 Material e Métodos
O experimento foi conduzido durante o período de agosto de 2009 à
abril de 2010, na área experimental limitada pelas coordenadas 8270210 S e
596532 W e altitude de 180m, localizada no Centro Universitário de Várzea
Grande – UNIVAG na cidade de Várzea Grande – MT. A pesquisa em
campo foi constituída por 3 parcelas de 9,9x10m cada.
O isolado de C. truncatum foi obtido de sementes de soja
submetidas à análise de rotina („blotter test‟) no Laboratório de Fitopatologia
do UNIVAG, da safra 2009/2010. O inóculo do patógeno foi multiplicado em
meio de cultura BDA (200 g de batata, 20 g de dextrose e 20 g de Agar). O
cultivo foi realizado em placa de Petri de 9 cm de diâmetro, por 7 dias a
temperatura de 20 ± (2°C) em regime alternado de 12 h luz / 12 h escuro.
Após esse período, 130 sementes de soja do cultivar TMG 131 RR,
suscetível ao fungo C truncatum e livres de patógenos foram distribuídas na
placa de Petri contendo a colônia fúngica em crescimento ativo sobre o BDA,
disposta em uma única camada ocupando toda a superfície da mesma, por
um período de 48 h. As sementes foram retiradas da placa e colocadas para
secar sobre papel toalha em temperatura ambiente, por 2 h.

66
O cultivar TMG 131 RR foi escolhido em função das seguintes
vantagens: facilidade do manejo das plantas daninhas, resistência aos
nematóides de galhas das raças 1 e 3, menor tempo de permanência no
campo favorecendo menor pressão da ferrugem asiática e por ser cultivado
no estado de Mato Grosso desde a região sul (baixa altitude) até a região do
médio norte (Fundação Mato Grosso, 2010).
Em agosto de 2009 foram coletados amostras de solo na área
experimental a fim de determinar a textura e características químicas do solo
(Tabela 1).
Tabela 1. Análise química e textural do solo da área experimental.
pH P K Ca Mg Areia Silte Argila
Água CaCl2 Mg dm-3 Cmolc dm-3 % 6,8 6,0 27,7 44 1,3 0,7 85,7 4,0 10,3
Zn Cu Fe Mn B S
mg dm-3 0,7 1,0 194 40,3 0,22 6,4
Já em setembro de 2009 foram semeadas 4 linhas de milho híbrido
AG 7088 ao redor das 3 parcelas , distanciadas a 2 m das mesmas em todos
as direções (norte, sul, leste, oeste) com espaçamento entre linhas de 0,50
m e população de 60000 plantas ha-1 (Figura 1) . A adubação foi de acordo
com os níveis de nutrientes no solo (Tabela 1) juntamente com a
recomendação de Alves et al. (1999), sendo aplicado 20 kg de N, 100 kg
P2O5 e 80 kg K2O. Os demais tratos culturais estão descritos na tabela 2.
O preparo convencional do solo foi realizado duas vezes com grade
aradora e uma vez com grade niveladora. Após a abertura manual dos
sulcos com auxílio de enxada, o adubo foi distribuído ao lado e abaixo das
sementes, conforme recomendação da Embrapa Soja (2008) (Tabela 2).
Após a incorporação do adubo, as sementes foram distribuídas em
profundidade de 2 a 3 cm.

67
Figura 1. Croqui da área experimental. Parcelas com 0,8% (A), 1,6% (B) e
2,4% (C) de sementes inoculadas com C. truncatum.

68
Tabela 2. Tratos culturais com as respectivas datas e doses referente as culturas do milho e soja.
Atividade Quantidade Data
Amostragem de solo 01 15/08/2009 Semeadura do milho AG 7088 - 02/09/2009 Adubação de base do milho
20 kg de N (uréia), 100 kg de P2O5 (SS 20%) e 80 kg
de K20 (KCl 60%)
02/09/2009
Emergência do milho - 07/09/2009 Adubação de cobertura 30 kg de N (uréia) 22/09/2009 Adubação de cobertura 20 kg de N (uréia), 20 kg de
K20 (KCl 60%) 24/10/2009
Semeadura da soja - 21/10/2009 Adubação de base da soja
80 kg de P2O5 (SS 20%) 27 kg de K2O (KCl 60%), B 1,5 kg (ac. Bórico 17%), Zn 6 kg (sulfato de Zn 21%), Cu 0,5
kg (sulfato de Cu 25%)
21/10/2009
* Tiametoxan (350 gL-1); fipronil (250 gL-1); carbendazim (150 gL-1) + thiran (350 gL-1); fluquinconazol (167 gL-1)
300 mL; 150 mL 200 mL;
300 mL dose para cada 100 kg de sementes
20/01/2009
Aplicação de lambda− cialotrina (50 gL-1) (soja)
150 mLha-1 31/10/2009
Aplicação de glifosato (480 gL-1) (soja)
2Lha-1 06/11/2009
Aplicação de lambda− cialotrina (50 gL-1) (milho e soja)
150 mLha-1 07/11/2009
Desbaste da soja - 19/11/2009 Aplicação de glifosato (480 gL-1) (soja)
2Lha-1 25/11/2009
Aplicação de lambda− cialotrina (50 gL-1) (milho e soja)
150 mLha-1 25/11/2009
Adubação de cobertura 53 kg de K2O (KCl 60%) 01/12/2009 Aplicação de tiametoxam (141 gL-1) (soja)
150 mL ha-1 23/12/2009
* Tratamento de sementes.
As sementes de soja foram semeadas nas 3 parcelas, ambas
distanciadas a 2 m das linhas de milho, cada uma contendo 20 linhas de 10
m de comprimento espaçadas de 0,45 m, sendo consideradas úteis as 16

69
linhas centrais. Foram semeadas 16,5 à 20 sementes de soja por metro
linear no sentido norte-sul. Em cada parcela experimental foram semeadas
quantidades diferentes de sementes inoculadas com C. truncatum, para
obtenção de níveis iniciais de incidência do patógeno de 0,8%, 1,6% e 2,4%.
Assim 20 sementes foram semeadas em 1 linha da parcela com 0,8% de
incidência, 40 sementes foram distribuídas em 2 linhas da parcela com 1,6%
de incidência e 60 sementes foram semeadas em 3 linhas da parcela com
2,4% de incidência. Todas as sementes foram distribuídas no centro de cada
parcela, onde cada linha correspondeu a 1 metro (m) linear (Figura 1).
As sementes não inoculadas com o patógeno foram tratadas com
fungicidas e inseticidas recomendados pelo MAPA (Tabela 2) e
posteriormente foi aplicado Bradyrhizobium elkanni na dosagem de 600
g250-1 kg de sementes (Biomax Premium, 7,2 x 109), 6 mL de água kg-1 de
semente na concentração de 10% de açúcar, em seguidas as sementes
foram secas à sombra em temperatura ambiente, por 1 h.
O desbaste das plantas foi realizado aos 23 dias após a emergência
da soja, deixando aproximadamente 15 plantas por m linear. Esse desbaste
não foi efetuado nas linhas contendo as sementes inoculadas com o
patógeno.
As pragas e plantas indesejáveis foram controladas com produtos
fitossanitários registrados no MAPA, ao longo do ciclo das culturas de milho
e soja (Tabela 2). Na ausência de precipitação pluvial e havendo a
necessidade de umidade no solo foi realizada irrigação por aspersão com
pivô central, no início da manhã ou ao final da tarde.
Utilizando-se GPS Topcon Hiper Lite configurado em datum WGS
84, zona UTM 21 Sul, modo RTK, foi demarcada em cada parcela uma
grade com 112 pontos amostrais (Figura 2), sendo cada ponto representado
por duas plantas, uma para avaliação da incidência da antracnose no campo
e a outra para a coleta de folíolos para avaliar a incidência do patógeno em
laboratório.

70
Figura 2. Malhas amostrais da área experimental. Parcelas com 0,8% (A),
1,6% (B) e 2,4% (C) de sementes inoculadas com C. truncatum.

71
Ao final do ciclo da soja realizou-se a colheita em cada ponto
amostral e em seguida efetuou-se a debulha manual. Após a secagem das
sementes à sombra, elas foram homogeneizadas e submetidas ao teste de
sanidade „blotter test‟ com restrição hídrica considerando a metodologia de
Brasil (2009), no Laboratório de Fitopatologia da UFMT.
Na análise sanidade, foram semeadas 25 sementes por placa com
substrato umedecido com manitol a – 0,6 MPa e ágar a 0,2%, para cada
ponto amostral. As placas de Petri com as sementes permaneceram na sala
de crescimento com temperatura de 22ºC (± 2ºC), e fotoperíodo de 12 h luz
e 12 h escuro. A leitura de incidência de C. truncatum nas sementes foi
realizada aos 4 e 7 dias após a incubação, com auxílio de microscópio
estereoscópio e quando necessário utilizou-se o microscópio óptico, sendo
retiradas as sementes com a presença do patógeno na primeira leitura.
Em seguida, os dados foram tabulados, e com a soma das leituras,
foram transformados em porcentagem.
3.2.1 Análise de dados
Na estatística descritiva não espacial, foi calculado os valores da
curtose, assimetria, máximo, mínimo, média, desvio padrão e coeficiente de
variação. Na estatística descritiva espacial, efetuou-se a análise de quantis.
A partir da análise exploratória dos dados, a geoestatística foi
utilizada para estudar a variabilidade espacial de C. truncatum, bem como
para escolher o modelo de semivariograma e método de ajuste que
descreveu melhor a variabilidade dos dados.
A dependência espacial da transmissibilidade da antracnose nas
sementes foi analisada por meio de ajustes de semivariogramas, com base
na pressuposição de estacionariedade da hipótese intrínseca. O
semivariograma experimental foi estimado de acordo com o estimador
robusto, descrito pela equação 1, conforme Diggle e Ribeiro Junior (2007).
γ(h) = 42/1 ])}()({
1
1[ ii xzhxz
i
Nh
Nh/ 0,914 +
Nh
988,0 (1)

72
Em que γ(h) corresponde a semivariância estimada e N(h) aos números de
pares de valores das observações )()( ii xzhxz separados pela distância
h. A transformação da raiz quadrada das diferenças é apresentada como tendo momentos parecidos com aqueles da distribuição normal e o denominador da equação utilizada para correção de tendências nos dados.
Os modelos teóricos esféricos, exponenciais, gaussianos, gneiting e
matern de semivariogramas isotrópicos utilizados foram definidos de acordo
com Diggle e Ribeiro Junior (2007), como apresentado nas equações de 2 a
6.
● Modelo Esférico:
γ (h) = {1 – 1,5 a
a
h
a
h)(5,0 } se h < a (2)
0, caso contrário
● Modelo Exponencial:
γ (h) = exp (-h / a) (3)
● Modelo Gaussiano:
γ (h) = exp {- (h2/a2)} (4)
● Modelo Gneiting:
C (h) = (1 + 8sh + 25 (sh)2 + 32 (sh)3) (1 - sh)8 1[0,1] (sh)
Em que, s = 0,301187465825 (5)
● Modelo Matern:
γ (h) = )()()(2
11 a
hK
a
h
kk
k
k (6)
k = 1
Em que a é o alcance e h, a distância.
Verificou-se o grau de dependência espacial (GDE) da
transmissibilidade do C. truncatum pela relação (c0/c0+c) x 100,

73
classificando-se como forte, com valor até 25, moderada de 25 a 75 e fraca
acima de 75 (Cambardella et al., 1994).
Após o cálculo das semivariâncias, foram ajustados aos
semivariogramas experimentais, os modelos testados pelo método dos
mínimos quadrados ordinários (OLS), conforme Diggle e Ribeiro Junior
(2007).
Posteriormente, os semivariogramas foram ajustados aos dados
pelo método dos mínimos quadrados ponderados (WLS), máxima
verossimilhança (ML) e máxima verossimilhança restrita (REML), de acordo
com Diggle e Ribeiro Junior (2007).
Em seguida, foi realizada a interpolação por krigagem para mapear a
transmissibilidade de níveis de C. truncatum em sementes de soja.
O valor médio de z em um ponto ^
z xi foi estimado de acordo com
Burrough e McDonnell (1998), conforme a equação 7.
)(.)(^
iii xzxxz (7)
Em que, )(^
ixz é o valor estimado da transmissibilidade do fungo para as
sementes no ponto, ix é o vetor de pesos determinado pelo ajuste do
semivariograma, e, )( ixz é o valor da variável representada por vizinhos de
um ponto z .
O método da validação cruzada foi empregado para escolha do
modelo de semivariograma teórico (Cressie, 1993; Goovaerts, 1997). Em
seguida efetuou-se a comparação entre os modelos, em cada método de
ajuste, por meio dos coeficientes de qualidade do erro médio (ME) (Equação
8) e quadrado médio do desvio padrão (MSDR) (Equação 9).
ME = )}()({1 ^
1 i
N
i i xzxzN
(8)
MSDR = )(
)}()({12
2^
1i
iiN
i x
xzxz
N (9)

74
Em que )( ixz é o valor observado no ponto )( ix ; )(^
ixz é o valor estimado pela
krigagem para o ponto )( ix e σ )( ix é a variância da krigagem.
Para o modelo do variograma ser considerado satisfatório, o valor do
MSDR deve ser próximo ou igual a 1 (Webster e Oliver, 2007a, b), e o valor
de EM próximo ou igual a 0 (Faraco et al., 2008).
Na análise exploratória não espacial, foi calculado a média, valor
máximo, valor mínimo, coeficiente de variação (CV%), curtose e desvio
padrão. Efetuou-se também, a análise de quantis e as demais análises
geoestatísticas no „software‟ R com os pacotes geoR, geoRglm e splancs
(Diggle e Ribeiro Junior, 2007).
Para testar a significância de dependência espacial dos dados,
utilizou-se a técnica de envelope proposta por Diggle e Ribeiro Junior (2007),
que consiste num recurso de simulação baseado em um teste gerado por
simulação de Monte Carlo, de forma que, com base na hipótese de ausência
de correlação espacial, a simulação delimita uma região onde os pontos do
semivariograma posicionam-se dentro ou fora do envelope. Conforme
Guedes et al. (2008), a ocorrência de pelo menos 1 ponto do
semivariograma fora dos limites de variação do envelope, indicou que os
dados apresentaram dependência espacial.
3.3 Resultados e Discussão
O método de inoculação do fungo nas sementes foi eficiente, pois a
maioria das sementes germinaram e apresentaram os primeiros sintomas
nos cotilédones com manchas deprimidas de coloração castanho-escuras
(Figura 3). Após a queda dos cotilédones, as plantas não apresentaram
sintomas ou sinais da doença e do patógeno respectivamente até o final do
florescimento, período em que observou-se sintomas em folhas, hastes de
soja e com o desenvolvimento das plantas os sintomas foram observados
também nas vagens, consequentemente o fungo infectou as demais plantas
com o passar do tempo.

75
Figura 3. Sintomas característicos da antracnose oriundo das sementes
inoculadas com C. truncatum.
Com base na análise exploratória não espacial dos dados, observou-
se que quanto maior o inóculo primário, menor foram os valores de
assimetria e coeficiente de variação (Tabela 3). O elevado valor do
coeficiente de variação entre 64 e 91%, foi justificado pelos diferentes
valores encontrados nos pontos amostrais, em que os maiores valores
encontraram-se praticamente ao redor das linhas onde foram semeadas 40
e 60 sementes de soja inoculadas com C. truncatum, correspondendo
respectivamente a 1,6 e 2,4% do total de sementes semeadas, conforme
ilustrado nos mapas de quantis (Figura 4), sendo que os dados da parcela
com maior inóculo apresentaram maior média, valor máximo e desvio
padrão. A parcela onde foram semeadas 20 sementes inoculadas com C.
truncatum (0,8% do total de sementes), apresentou valores de
transmissibilidade acima de 28% na extremidade norte da parcela (Figura 4),
por esta razão, apresentou o maior coeficiente de variação (Tabela 3). Leal
et al. (2010) avaliando a distribuição espacial e a expansão da
Huanglongbing (greening) em citrus com metodologia geoestatística,

76
também observou elevados valores de coeficiente de variação entre 85,95 a
313,05% em 36 talhões avaliados.
Tabela 3. Estatística descritiva referente a transmissibilidade em parcelas com 0,8, 1,6 e 2,4% de sementes de soja inoculadas com C. truncatum.
Índices Estatísticos
Sementes inoculadas
0,8% 1,6% 2,4%
Curtose 0,50 0,69 - 0,89 Assimetria 1,05 0,85 0,16 Mínimo 0,00 0,00 0,00 Média 17,29 11,68 35,12 Máximo 64,00 44,00 92,00 Desvio padrão 15,73 9,90 22,75 CV% 91,00 84,79 64,78
A estatística clássica não permitiu quantificar a estrutura e
magnitude de dependência espacial entre as amostras em estudo. Portanto,
utilizou-se a geoestatística para avaliar a variabilidade espacial do fungo nas
parcelas.
Com os mapas de quantis, verificou-se que a distribuição da
transmissibilidade semente-semente dos níveis do patógeno em estudo, teve
resultado semelhante especialmente para as parcelas contendo 1,6 e 2,4%
de sementes inoculadas, em que os maiores valores foram encontrados na
região das linhas com sementes inoculadas e nas suas proximidades (Figura
4).
A B
F E D

77
Figura 4. Mapas de quantis da transmissibilidade semente-semente de C. truncatum nas parcelas com 0,8% (A), 1,6% (B) e 2,4% (C) de sementes inoculadas.
A
B
C

78
A figura 5, ilustra o semivariograma experimental omnidirecional
para cada porcentagem de sementes inoculadas com C. truncatum, com os
respectivos envelopes. Com base na simulação de Monte Carlo, observou-
se que pelo menos um ponto da semivariância esteve fora dos limites de
variações do envelope, indicando que os dados apresentaram dependência
espacial. Contudo, constatou-se também que o maior número de pontos fora
do envelope sob condições da parcela com 2,4% de sementes inoculadas
com C. truncatum (Figura 5).
O grau de dependência espacial foi classificado como moderado a
forte, para todos os modelos e métodos testados, sendo 56,67% dos valores
classificados como moderado e 43,33% como forte estrutura de
dependência espacial. Observou-se que todos os modelos avaliados
ajustado ao método ML apresentaram os maiores valores de dependência
espacial da transmissibilidade de C. truncatum semente-semente (Tabela 4).
Verificou-se que os menores valores do efeito pepita foram
encontrados nos modelos gneiting e gaussiano ajustado ao método OLS
para a parcela com 0,8%, modelos esférico e exponencial ajustado ao
método OLS e WLS para a parcela com 1,6% e, modelos esférico e
exponencial ajustados ao método WLS para a parcela com 2,4% de
sementes de soja inoculadas com o patógeno. Não ocorreu nenhum efeito
pepita puro para os modelos e métodos de ajuste testados (Tabela 4).
Conforme Valeriano e Santos (2002), dados com efeito pepita grande (em
relação ao patamar atingido) indicam pequena dependência espacial e
requerem maior cuidado para sua espacialização e conseqüente
mapeamento.
A dependência espacial da transmissibilidade de C. truncatum para
as sementes, foi constatada por meio de variografia. Outro ponto observado
foi que os semivariogramas gneitings, esféricos, exponenciais, gaussianos e
materns ajustados ao método OLS e modelos exponenciais e materns
ajustados ao método WLS apresentaram alcance com valor crescente, com
o aumento do inóculo, comprovando a agressividade do patógeno. Para os
semivariogramas esféricos, observou-se alcances variando de 1,36 a 8,83 m

79
respectivamente para as parcelas 0,8 e 2,4% de sementes inoculadas
(Tabela 4), indicando a distância de dependência espacial da
transmissibilidade do patógeno para as sementes.
O mesmo comportamento foi obtido para os modelos exponenciais e
esféricos ajustados ao método REML, onde observou-se alcance de 0,95 a
7,05 m, para os semivariogramas esféricos. Sendo que a partir da
semivariância estabilizada do respectivo alcance, não houve mais
dependência espacial (Tabela 4). Já para os semivariogramas exponenciais,
obteve-se alcance crescente quando se utilizou o REML (Tabela 4), ou seja,
quanto maior a fonte de inóculo primário, maior foi a dependência espacial
da transmissibilidade da semente para a semente.
Com esses resultados, percebeu-se o progresso e a agressividade
do patógeno se propagar no espaço e no tempo. Alves et al. (2006),
observaram alcance prático de até 3,15 m aos 61 dias após a semeadura,
para a severidade da antracnose do feijoeiro em parcelas com inóculo de
0,5%, além de apresentar forte grau de dependência espacial.
A superioridade do ML e REML em relação ao OLS e WLS, é em
função da estimativa dos parâmetros do banco de dados originais e não
apenas aos pontos do variograma experimental, ou seja, os
semivariogramas foram ajustados aos dados originais e não aos pontos do
semivariograma. Conforme Mello et al. (2005), ao se aplicar o método da
máxima verossimilhança para um mesmo conjunto de dados, serão sempre
obtidos os mesmos parâmetros, eliminando assim, as fontes de
subjetividade. O mesmo autor afirma também, que na condição de
normalidade, este método proporcionará o melhor conjunto de parâmetros
para um dado conjunto de dados. De acordo com Silva (2006), o estimador
de máxima verossimilhança possui propriedades tais como: ser
assintoticamente não tendencioso, conferir estimativa de variância mínima,
ser consistente e invariante.

80
Figura 5. Envelope de Monte Carlo aplicado aos semivariogramas
experimentais omnidirecionais, nas parcelas contendo 0,8% (A), 1,6% (B) e 2,4% (C) de sementes inoculadas com C. truncatum.
A
B
C
Sem
ivari
ância
Distância (m)
Distância (m)
Distância (m)
Sem
ivari
ância
S
em
ivari
ância

81
Tabela 4. Parâmetros e coeficiente de semivariogramas e métodos ajustados, referente a transmissibilidade em parcelas com 0,8, 1,6 e 2,4% de sementes de soja inoculadas com C. truncatum.
Sementes inoculadas
(%)
Modelo Método Efeito pepita (C0)
Patamar (C0+C)
Alcance (a) (m)
Alcance prático
GDE
0,8 gneiting OLS 0,00 175,93 0,23 0,38 0,00 0,8 esférico OLS 65,53 180,88 1,36 1,36 36,23 0,8 exponencial OLS 85,32 194,83 1,22 3,66 43,79 0,8 gaussiano OLS 0,00 175,97 0,23 0,39 0,00 0,8 matern OLS 98,05 191,68 0,78 3,13 51,15 0,8 gneiting WLS 133,87 196,19 2,38 4,06 68,24 0,8 esférico WLS 119,76 195,13 4,62 4,62 61,38 0,8 exponencial WLS 106,78 208,53 2,23 6,67 51,21 0,8 gaussiano WLS 134,01 196,70 2,41 4,18 68,13 0,8 matern WLS 121,84 203,91 1,44 5,77 59,75 0,8 gneiting ML 196,30 324,10 7,13 12,17 60,57 0,8 esférico ML 182,70 281,04 11,28 11,28 65,01 0,8 exponencial ML 178,70 286,00 5,63 16,87 62,48 0,8 gaussiano ML 196,50 327,50 7,33 12,69 60,00 0,8 matern ML 189,20 302,20 4,11 16,43 62,61 0,8 gneiting REML 196,20 738,80 12,61 21,52 26,56 0,8 esférico REML 74,21 248,41 0,95 0,95 29,87 0,8 exponencial REML 71,84 251,84 0,66 1,96 28,53 0,8 gaussiano REML 142,23 296,75 6,52 11,29 47,93 0,8 matern REML 192,20 6066,20 51,89 207,50 3,17 1,6 gneiting OLS 21,90 130,59 2,32 3,96 16,77
1,6 esférico OLS 0,00 126,34 4,41 4,41 0,00
1,6 exponencial OLS 0,00 201,60 4,01 12,02 0,00
1,6 gaussiano OLS 22,22 131,86 2,37 4,09 16,85
1,6 matern OLS 10,67 156,27 1,69 6,74 6,83
1,6 gneiting WLS 18,16 129,37 2,22 3,78 14,04
1,6 esférico WLS 0,00 125,69 4,29 4,29 0,00
1,6 exponencial WLS 0,00 191,46 3,67 10,98 0,00
1,6 gaussiano WLS 18,66 130,60 2,26 3,91 14,29
1,6 matern WLS 3,71 150,50 1,50 6,00 2,47
1,6 gneiting ML 36,08 104,23 2,02 3,45 34,62
1,6 esférico ML 17,84 110,89 4,00 4,00 16,09
1,6 exponencial ML 7,69 98,09 1,35 4,03 7,84
1,6 gaussiano ML 36,49 103,81 2,05 3,55 35,15
1,6 matern ML 20,72 97,52 0,91 3,64 21,25
1,6 gneiting REML 36,20 113,40 2,12 3,61 31,92
1,6 esférico REML 17,12 115,25 4,03 4,03 14,85
1,6 exponencial REML 9,38 109,88 1,64 4,92 8,54
1,6 gaussiano REML 36,70 114,52 2,18 3,77 32,05

82
Sementes inoculadas
(%)
Modelo Método Efeito pepita (C0)
Patamar (C0+C)
Alcance (a) (m)
Alcance prático
GDE
1,6 matern REML 21,83 106,36 1,03 4,11 20,52
2,4 gneiting OLS 207,38 753,90 3,95 6,75 27,51 2,4 esférico OLS 148,61 767,22 8,83 8,83 19,37 2,4 exponencial OLS 113,64 932,24 5,34 15,99 12,19 2,4 gaussiano OLS 207,07 819,64 3,99 6,90 25,26 2,4 matern OLS 169,04 836,68 2,75 10,98 20,20 2,4 gneiting WLS 193,46 726,60 3,57 6,10 26,63 2,4 esférico WLS 86,76 717,13 6,98 6,98 12,10 2,4 exponencial WLS 41,82 871,22 4,07 12,18 4,80 2,4 gaussiano WLS 194,10 730,41 3,62 6,26 26,57 2,4 matern WLS 128,46 806,05 2,35 9,41 15,94 2,4 gneiting ML 231,10 492,00 2,78 4,74 46,97 2,4 esférico ML 204,60 459,40 5,85 5,85 44,54 2,4 exponencial ML 198,80 546,10 4,30 12,87 36,40 2,4 gaussiano ML 231,00 492,93 2,80 4,84 46,86 2,4 matern ML 220,50 543,60 2,37 9,47 40,56 2,4 gneiting REML 231,90 532,70 2,91 4,97 43,53 2,4 esférico REML 204,20 517,10 7,05 7,05 39,49 2,4 exponencial REML 202,40 967,90 10,62 31,83 20,91 2,4 gaussiano REML 231,00 533,30 2,93 5,08 43,32 2,4 matern REML 222,50 694,90 3,11 12,44 32,02
GDE = grau de dependência espacial
De acordo com Maestre (2008), o método de ajuste de modelos de
semivariogramas por OLS tem se mostrado insensível ao ajuste, mas esse
método é de implementação mais fácil.
Os valores do efeito pepita e do grau de dependência não foram
suficientes para representar o melhor modelo e método de ajuste testado,
porém, a validação cruzada permitiu por meio dos coeficientes de qualidade
a escolha da melhor combinação para a representação variográfica dos
dados. Os melhores coeficientes de qualidade, foram observados nos
métodos ML e REML (Tabela 5), porém, no ML, houve vários alcances que
apresentaram distâncias maiores que a área experimental (Tabela 4).
Comparando os modelos e métodos de ajuste de semivariogramas,
ficou evidente que o modelo isotrópico esférico ajustado ao método REML
apresentou para as parcelas com 0,8, 1,6 e 2,4% de sementes inoculadas

83
avaliadas, os melhores coeficientes de qualidade, sendo utilizado para a
representação variográfica e para a confecção dos mapas de krigagem
(Tabela 5 e Figuras 6 e 7). Notou-se outras combinações de ajuste entre
modelo e método com coeficientes de qualidades iguais ou muito próximos
dos encontrados no modelo e método escolhido, porém, na maioria destes
casos o valor correspondente do alcance ultrapassou a distância real das
parcelas experimentais (Tabelas 4 e 5). Alves et al. (2011), caracterizaram a
estrutura espacial de pragas do cafeeiro e, obtiveram também melhor
aplicação do semivariograma esférico ajustado pelo método REML.
Tabela 5. Validação cruzada da krigagem e métodos ajustados, referente a transmissibilidade em parcelas com 0,8, 1,6 e 2,4% de sementes de soja inoculadas com C. truncatum.
Sementes inocula- das (%)
Modelo Método ME MSDR
0,8 gneiting OLS 0,01 1,39 0,8 esférico OLS 0,05 1,53 0,8 exponencial OLS 0,01 1,49 0,8 gaussiano OLS 0,01 1,39 0,8 matern OLS 0,01 1,52 0,8 gneiting WLS 0,00 1,41 0,8 esférico WLS 0,00 1,41 0,8 exponencial WLS 0,01 1,44 0,8 gaussiano WLS 0,00 1,41 0,8 matern WLS 0,00 1,42 0,8 gneiting ML 0,00 1,00 0,8 esférico ML 0,00 1,01 0,8 exponencial ML 0,00 1,01 0,8 gaussiano ML - 0,01 1,00 0,8 matern ML 0,00 1,01 0,8 gneiting REML 0,00 1,00 0,8 esférico REML 0,05 1,02 0,8 exponencial REML 0,03 1,10 0,8 gaussiano REML 0,01 1,37 0,8 matern REML 0,00 1,00 1,6 gneiting OLS - 0,01 1,55 1,6 esférico OLS 0,02 1,90 1,6 exponencial OLS 0,02 1,67 1,6 gaussiano OLS 0,00 1,56 1,6 matern OLS 0,01 1,72 1,6 gneiting WLS - 0,01 1,83

84
Sementes inocula- das (%)
Modelo Método ME MSDR
1,6 esférico WLS 0,02 1,86 1,6 exponencial WLS 0,02 1,61 1,6 gaussiano WLS - 0,01 1,83 1,6 matern WLS 0,04 2,69 1,6 gneiting ML - 0,01 0,98 1,6 esférico ML - 0,01 0,98 1,6 exponencial ML - 0,01 0,98 1,6 gaussiano ML - 0,01 0,98 1,6 matern ML - 0,01 0,98 1,6 gneiting REML - 0,01 0,98 1,6 esférico REML 0,00 0,98 1,6 exponencial REML 0,00 0,98 1,6 gaussiano REML - 0,01 0,98 1,6 matern REML 0,00 0,98 2,4 gneiting OLS - 0,02 1,14 2,4 esférico OLS - 0,03 1,19 2,4 exponencial OLS - 0,05 1,33 2,4 gaussiano OLS - 0,05 2,44 2,4 matern OLS - 0,03 0,99 2,4 gneiting WLS - 0,03 1,19 2,4 esférico WLS - 0,04 1,66 2,4 exponencial WLS - 0,06 2,11 2,4 gaussiano WLS - 0,03 1,19 2,4 matern WLS - 0,04 1,54 2,4 gneiting ML - 0,04 0,99 2,4 esférico ML - 0,04 0,99 2,4 exponencial ML - 0,04 0,99 2,4 gaussiano ML - 0,04 0,99 2,4 matern ML - 0,03 0,99 2,4 gneiting REML - 0,04 0,99 2,4 esférico REML - 0,03 0,99 2,4 exponencial REML - 0,04 0,99 2,4 gaussiano REML - 0,04 0,99 2,4 matern REML - 0,04 1,22
ME = erro médio MSDR = quadrado médio do desvio padrão
Charest et al. (2002), estudando a distribuição espacial de Ventura
inaequalis e Dinardo-Miranda e Fracasso (2009) pesquisando a distribuição
espacial de nematóides, observaram melhor aplicação do semivariograma
esférico para caracterizar a variabilidade espacial dos dados. Dinardo-
Miranda et al. (2007), usaram análises geoestatistícas e, estimaram a

85
dependência espacial de mahanarva finbriolata somente após o início da
segunda geração desta praga, sendo observado também, melhor aplicação
do semivariograma esférico. Outros pesquisadores (Trangmar et al., 1985;
Cambardella et al., 1994; Paz et al., 1996; Salviano, 1996), também
relataram que o semivariograma esférico, foi o mais adaptado para
descrever a melhor representação de atributos de solo e de planta.
O alcance encontrado para o modelo e método escolhido foi de 0,95,
4,03 e 7,05 m, para as parcelas com 0,8, 1,6 e 2,4% de sementes
inoculadas (Tabela 4). Na prática, isso caracterizou aumento de
dependência espacial de 0,65, 2,76 e 4,83 cm da incidência do patógeno
nas sementes respectivamente a cada dia, pois o ciclo da cultivar TMG 131
RR foi de146 dias.
A transmissibilidade de C. truncatum na parcela com 0,8% de
sementes inoculadas, apresentou maior ocorrência no centro da parcela e
no lado nordeste. A justificativa provável de a transmissibilidade estar
também localizada na região nordeste, pode ser explicada pela maior
população de plantas nesta área, aliado ao sentido da direção do sistema de
irrigação por pivô central, podendo ter causado injúrias as plantas, tornando-
as predispostas à ocorrência da doença e, consequentemente, maior
transmissibilidade para as sementes. Observou-se que na parcela com 0,8%
de sementes inoculadas, ou seja, menor quantidade de inóculo, houve
aumento na distância entre os focos secundários com maior porcentagem de
transmissibilidade para a semente (Figura 7 A). Essa capacidade de
dispersar pode ser uma característica da sobrevivência do C. truncatum.
Este fato caracterizou o patossitema C. truncatum versus soja, como
descrito por Talamini (2003), uma vez que a antracnose é policíclica.
Resultados semelhantes foram encontrados por Alves et al. (2006),
avaliando a severidade da antracnose em plantas de feijoeiro, constataram
por meio de mapas de krigagem, mecanismo de dispersão da doença com
formação de focos secundários além do foco primário.

86
Figura 6. Modelo de semivariograma esférico ajustado pelo método REML, para as sementes com 0,8% (A); 1,6% (B) e, 2,4%. (C), sementes de soja inoculadas com C. truncatum.
Distância (m)
Sem
ivari
ância
A
B
C
Distância (m)
Distância (m)
Sem
ivari
ância
S
em
ivari
ância

87
Araújo et al. (2006) estudaram a relação entre porcentagens de
sementes de algodoeiro infectadas com C. gossypii var. cephalosporioides e
o progresso da ramulose no campo e, por meio de análise estatística, com o
teste de Tukey, concluíram que a transmissão do patógeno planta-semente
foi maior com aumento do inóculo inicial nas sementes.
A parcela com 1,6% de sementes inoculadas teve maior
transmissibilidade agrupada no centro da parcela e poucos pontos
espalhados aleatoriamente nas proximidades do foco primário (Figura 7 B).
A parcela com 2,4% de sementes inoculadas apresentou maior
transmissibilidade na região central da parcela, espalhando-se mais para o
sentido sudeste, até a bordadura (Figura 7 C). Observou-se por meio dos
mapas de krigagem, que quanto maior foi o inóculo primário, a distribuição
da transmissibilidade semente para semente ocorreu com maior incidência
próxima à fonte do inóculo, representando padrão agregado (Figura 7).
Resultado semelhante foi encontrado por Alves et al. (2006), ao
caracterizar o progresso espaço-temporal da severidade da antracnose do
feijoeiro e da ramulose do algodoeiro e, observaram padrão espacial
agregado dessas epidemias. Musoli et al. (2008), estudaram por meio de
técnicas geoestatísticas a estrutura espaço-temporal da murcha do café
causada por Fusarium xylarioides em Coffea canephora e, observaram
progresso temporal lento e padrão espacial agregado dessa doença.
Almeida et al. (2011), ao estudar a dependência espacial de Phytophthora
nicotianae, por meio do modelo de semivariograma isotrópico esférico
ajustado aos dados, detectaram ausência de correlação espacial do
patógeno entre as amostras avaliadas, sendo caracterizada com alto índice
de infestação do fungo em toda área amostrada, uma vez que a área
experimental foi inoculada em anos anteriores com o mesmo fungo para
realização de outros trabalhos, revelando assim, que a área apresentava
uma distribuição agregada do patógeno.
Ressaltou-se que a direção predominante do vento durante o ciclo
da cultura foi norte. É interessante destacar também, o fato que a partir de
76, 83 e 97 dias após a semeadura, todas as plantas demarcadas

88
apresentavam sintomas da antracnose, para as parcelas com 1,6, 2,4 e
0,8% de sementes inoculadas, respectivamente.
Araújo e Chitarra (2005) observaram em Mato Grosso, aumento na
severidade da ramulose com o aumento dos níveis de incidência de C.
gossypii var. cephalosporioides nas sementes. A partir de 1% de incidência
do patógeno nas sementes, a doença na planta foi mais severa.

89
Figura 7. Mapas de krigagem gerados a partir de modelo esférico ajustado
pelo método REML para as parcelas com 0,8% (A), 1,6% (B) e 2,4% (C), de sementes inoculadas com C. truncatum.
C
B
A

90
3.4 Conclusão
O melhor modelo de semivariograma foi o esférico, ajustado pelo
método da máxima verossimilhança restrita.
Os valores de alcance foram de 0,95, 4,03 e 7,05 m para as parcelas
com 0,8%, 1,6% e 2,4% de sementes inoculadas com C. truncatum
respectivamente.
Quanto maior o inóculo primário, maior foi a transmissibilidade de
semente para semente próximo à fonte de inóculo.
3.5 Referências Bibliográficas
ALMEIDA, F.A.; BARRETO, M.; BARBOSA, J.C.; et al. Distribuição espacial de Phytophthora nicotianae e reação de cultivares de cebola ao fungo. Summa Phytopathologica, v.37, p.334-337, 2011. ALVES, V.M.C.; VASCONCELLOS, C.A.; FREIRE, F.M. et al. Milho. In: Ribeiro, A.C.; GUIMARÃES, P.T.G.; ALVARES V.V.H. Comissão de Fertilidade do Solo do Estado de Minas Gerais Recomendações para o uso de corretivos e fertilizantes em Minas Gerais - 5ª Aproximação – Viçosa, MG, 1999, p. 314-316. ALVES, M.C.; POZZA, E.A.; MACHADO, J.C.; et al. Geoestatística como metodologia para estudar a dinâmica espaço-temporal de doenças associadas a Colletotrichum spp. transmitidos por sementes. Fitopatologia Brasileira, v.31, p.557-563, 2006.
ALVES, M.C.; SILVA, F.M.; MORAES, J.C.; POZZA, E. A.; OLIVEIRA, M.S.; SOUZA, J.C.S.; ALVES, L.S. Geostatistical analysis of the spatial variation of the berry borer and leaf miner in a coffee agroecosystem. Precision Agriculture, v.12, p.18-31, 2011.
ARAÚJO, A.E.; CHITARRA, L.G. Efeito de níveis de inóculo nas sementes sobre o progresso da ramulose do algodoeiro no Mato Grosso. In: XXXVIII Congresso Brasileiro de Fitopatologia, Brasília. Fitopatologia Brasileira. Fortaleza: Sociedade Brasileira de Fitopatologia, v.30, p.S193-S193, 2005.
ARAÚJO, D.V.; ZAMBENEDETTI, E.B.; MACHADO, J.C.; CELANO, F.A.O. Relação entre níveis de inóculo de Colletotrichum gossypii var. cephalosporioides nas sementes e o progresso da ramulose do algodoeiro. Fitopatologia Brasileira, v.31, p.147-151, 2006. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Manual de Análise Sanitária de Sementes. Ministério da Agricultura, Pecuária e

91
Abastecimento. Secretaria de Defesa Agropecuária. – Brasília: Mapa/ACS, 2009, 200p. BURROUGH, P.A.; McDONNELL, R.A. Principles of Geographical Information Systems. 2nd Edition, Oxford University Press, Oxford, 1998, 333p. CAMBARDELLA, C.A.; MOORMAN, T.B.; NOVAK, J.M.; PARKIN, T.B.; KARLEN, D.L.; TURCO, R.F.; KONOPKA, A.E. Field-scale variability of soil properties in Central Iowa soils. Soil Science Society of America Journal, Madison, v.58, p.1501-1511, 1994.
CAMPBELL, C.L.; MADDEN, L.V. Introduction to plant disease epidemiology. New York: J. Wiley, 1990, 532p.
CHAREST, J.; DEWDNEY, M.; PAULITZ, T.; PHILION, V.; CARISSE, O. Spatial Distribution of Venturia inaequalis Airborne Ascospores in Orchards. Phytopathology, v.92, p.769-779, 2002.
CONAB - Companhia Nacional de Abastecimento. Séries Históricas. Disponível em: <http://www.conab.gov.br/conteudos.php?a=1252&t=2&Pagina_objcmsconteudos=2#A_objcmsconteudos>. Acesso em: 13 nov. 2010. CRESSIE, A.G. Statistics for spatial data. New York: Wiley, 1993, 900p.
DIGGLE, P.J.; RIBEIRO JR, P.J. Model-based Geoestatistics. New York, Springer, 2007, 228p. DINARDO-MIRANDA, L.L.; FRACASSO, J.V. Spatial distribution of plant-parasitic nematodes in sugarcane fields. Scientia Agricola, v.66, p.188-194, 2009.
DINARDO-MIRANDA L.L.; VASCONCELOS, A.C.M.; VIEIRA, S.R.; FRACASSO, J.V.; GREGO, C.R. Uso da geoestatística na avaliação da distribuição espacial de Mahanarva fimbriolata em cana-de-açúcar. Bragantia, v.66, p.449-455, 2007. EMBRAPA – Empresa Brasileira de Pesquisa Agropecuária. Soja em números (safra 2008/2009). Disponível em: <http://www.cnpso.embrapa.br/index.php?cod_pai=2&op_page=294>. Acesso em: 16 nov. 2010. EMBRAPA SOJA. Tecnologias de Produção de Soja - Região Central do Brasil, 2009/2010. Londrina: Embrapa Soja: Embrapa Cerrados: Embrapa Agropecuária Oeste, 2008, 262p.

92
FARACO, M.A.; URIBE-OPAZO, M.A.; SILVA, E.A.A. et al. Seleção de modelos de variabilidade espacial para elaboração de mapas temáticos de atributos físicos do solo e produtividade da soja. Revista Brasileira de Ciência do Solo, v.32, p.463-476, 2008.
GOOVAERTS, P. Geostatistics for natural resources evaluation. New York: Oxford University Press, 1997. GUEDES, L.P.C.; URIBE-OPAZO, M.A.; JOHANN, J.A. et al. Anisotropia no estudo da variabilidade espacial de algumas variáveis químicas do solo. Revista Brasileira de Ciência do Solo, Viçosa, v.32, 2008 . Disponível em: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-06832008000600001&lng=pt&nrm=iso>. Acesso em: 18 nov. 2010. MAESTRE, M.R. Estimação de contrastes de médias de tratamentos, de um experimento em blocos ao acaso, utilizando as análises clássica e espacial. Dissertação (Mestrado). – Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, Universidade de São Paulo, Piracicaba, 70p.:Il. 2008. MELLO, J.M.; BATISTA, J.L.F.; RIBEIRO JÚNIOR, P.J.; OLIVEIRA, M.S. Ajuste e seleção de modelos espaciais de semivariograma visando à estimativa volumétrica de Eucalyptus grandis. Scientia Forestalis, v.1, p.25-37, 2005.
MUSOLI, C.P.; PINARD, F.; CHARRIER, A. et al. Spatial and temporal analysis of coffee wilt disease caused by Fusarium xylarioides in Coffea canephora. European Journal of Plant Pathology, v.122, p.451-460, 2008. LEAL, R.M.; BARBOSA, J.C.; COSTA, M.G.; YAMAMOTO, P.T.; DRAGONE, J. Distribuição espacial de Huanglongbing (Greening) em citros utilizando geoestatística. Revista Brasileira de Fruticultura, v.32, p.1-8, 2010.
PAZ, A.; TABOADA, M.T.; GÓMEZ, M.J. Spatial variability in topsoil micronutrients contents in one-hectare cropland plot. Communication in Soil Science and Plant Analysis, v.27, p.479-503, 1996. PINTO, A.C.S., POZZA, E.A., TALAMINI, V., MACHADO, J.C., SALES, N.L.P., GARCIA JÚNIOR, D. e SANTOS, D.M. Análise do padrão espacial e do gradiente da antracnose do feijoeiro em duas épocas de cultivo. Summa Phytopathologica, v. 27, p. 392-398, 2001. SALVIANO, A.A.C. Variabilidade de atributos de solo e de Crotalaria juncea em solo degradado do município de Piracicaba-SP. 1996. 91f. Tese (Doutorado em Agronomia) - Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, Piracicaba - SP, 1996.

93
SILVA, P. M. O. Modelagem do escoamento superficial e da erosão hídrica na sub-bacia hidrográfica do ribeirão Marcela, Alto Rio Grande, MG. 2006. 155f. Tese (Doutorado em Engenharia Agrícola) - Universidade Federal de Lavras, Lavras – MG, 2006. TALAMINI, V.; POZZA, E.A.; MACHADO, J.C.; OLIVEIRA, F.A. Epidemiologia de doenças associadas a Colletotrichum spp. transmitidas por sementes. Revisão Anual de Patologia de Plantas. Passo Fundo, v.10, p. 219-248, 2002. TALAMINI, V. Progresso espacial e temporal da antracnose a partir de diferentes níveis de inóculo inicial em sementes de feijoeiro. 2003. 144f. Tese (Doutorado em Fitopatologia) - Universidade Federal de Lavras, Lavras – MG, 2003. TANAKA, M.A.S.; MENTEN, J.O.M. Relação entre a resistência do algodoeiro à ramulose e a transmissão de Colletotrichum gossypii var. cephalosporiodes pelas sementes. Summa Phytopathologica, Piracicaba, v.18, p. 227-234, 1992. TRANGMAR, B.B.; YOST, R.S.; UEHARA, G. Application of geoestatistic to spatial studies of soil properties. Advance in Agronomy, New York, v.38, p. 45-94, 1985.
VALERIANO, M. M.; SANTOS, A.O. Mapping of plant-soil data with high random variability. Acta Scientiarum, v.24, p.921-928, 2002. VIEIRA, S. R. Geoestatística em estudos de variabilidade espacial do solo. In: NOVAIS, R.F. de; ALVAREZ V.,V.H.; SCHAEFER, C.E.G.R. (Ed.). Tópicos em Ciência do Solo. Viçosa: Sociedade Brasileira de Ciência do Solo, v.1, p.1-54, 2000. WEBSTER, R.; OLIVER, M.A. Sample adequately to estimate variograms of soil properties. Journal of Soil Science, 43, 177–192. 2007a.
WEBSTER, R.; OLIVER, M.A. Geostatistics for environmental scientists. England: Wiley. 2007b.

94
4. CONSIDERAÇÕES FINAIS
Com o avanço da tecnologia muitos dos métodos clássicos de
análises de dados estão sendo substituídos por técnicas geoestatísticas.
As informações obtidas nesta pesquisa, podem ser utilizadas em
estudos futuros, com intuito de enriquecimento no banco de dados em
sistema de previsão e aviso da antracnose da soja, mapas de probabilidades
do progresso espaço-temporal da antracnose e da transmissibilidade do C.
truncatum semente – semente.
Nas condições do presente trabalho, as parcelas com 0,8%, 1,6% e
2,4% de sementes inoculadas com C. truncatum foram caracterizadas com
alcance de 0,95, 4,03 e 7,05 m respectivamente, representando o raio de
influência atribuída a cada ponto amostral. Isso, ilustra a necessidade do
estabelecimento de legislação com padrão de tolerância de C. truncatum em
sementes de soja com base em pesquisas científicas.
Para estudos futuros em lavouras comerciais localizadas em
distintas regiões produtoras de soja, recomenda-se utilizar malhas amostrais
com diferentes distâncias, além do monitoramento das variáveis climáticas
(temperatura do ar, umidade relativa do ar, precipitação, duração do
molhamento foliar, radiação solar e direção do vento).





























