Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DE OURO PRETO
ESCOLA DE NUTRIÇÃO
PROGRAMA DE PÓS-GRADUAÇÃO EM SAÚDE E NUTRIÇÃO
ÁREA: BIOQUÍMICA E FISIOLOGIA DA NUTRIÇÃO
AVALIAÇÃO DO POTENCIAL ANTIOXIDANTE DA
FARINHA DE BURITI (Mauritia flexuosa) IN VITRO E EM
RATOS DIABÉTICOS
Nara Nunes Lage
Ouro Preto – MG
2014

2
NARA NUNES LAGE
AVALIAÇÃO DO POTENCIAL ANTIOXIDANTE DA FARINHA
DE BURITI (Mauritia flexuosa) IN VITRO E EM RATOS
DIABÉTICOS
Dissertação apresentada ao Programa de Pós-
Graduação em Saúde e Nutrição, como
requisito parcial para obtenção do título de
Mestre em Saúde e Nutrição, área de
concentração: Bioquímica e Fisiologia da
Nutrição.
Orientador (a): Profª. Drª. Maria Lúcia
Pedrosa
Co-orientador: Prof. Dr. Marcelo Eustáquio
Silva
Ouro Preto – MG
2014

3
Catalogação: [email protected]
L174a Lage, Nara Nunes.
Avaliação do potencial antioxidante da farinha de buriti (mauritia flexuosa)
in vitro e em ratos diabéticos [manuscrito] / Nara Nunes Lage. - 2014.
71 f.; il.;color.; grafs.; tab.
Orientadora: Profª Drª. Maria Lúcia Pedrosa.
Coorientador: Prof. Dr. Marcelo Eustáquio Silva.
Dissertação (Mestrado) - Universidade Federal de Ouro Preto. Escola de
Nutrição. Programa de Pós-Graduação em Saúde e Nutrição.
Área de concentração: Bioquímica e Fisiopatologia da Nutrição.
1. Buriti - Teses. 2. Alimentos funcionais - Teses. 3. Diabetes - Teses.
4. Stress (Fisiologia) – Teses. I. Pedrosa, Maria Lúcia. II. Silva, Marcelo
Eustáquio. III. Universidade Federal de Ouro Preto. IV. Título.
CDU: 612.392.7
CDU: 669.162.16

4

5
APOIO FINANCEIRO
Este trabalho foi realizado no Laboratório de Bioquímica Metabólica e no
Laboratório de Nutrição Experimental, com o auxílio financeiro do Conselho Nacional
de Desenvolvimento Científico e Tecnológico (CNPq), da Comissão de Aperfeiçoamento
de Pessoal de Nível Superior (CAPES) e da Fundação de Amparo à Pesquisa do Estado
de Minas Gerais (FAPEMIG).

6
DEDICATÓRIA
À minha mãe Rita e aos meus irmãos
Nádia e Vinícius pelo amor incondicional
e por acreditarem em mim sempre!
E ao meu pai Renato que, mesmo ausente,
sei que está a todo momento me iluminando...
Amo muito vocês!!!

7
AGRADECIMENTO ESPECIAL
Aos meus queridos professores e orientadores Maria Lúcia Pedrosa e Marcelo
Eustáquio Silva, exemplos de seres humanos e profissionais, pelo acolhimento no
grupo de pesquisa; pela paciência em transmitir todo o seu conhecimento; pelo carinho
de todos os dias; pela dedicação e por, desde o início da minha vida acadêmica,
acreditarem no meu potencial. Muito obrigada por todo aprendizado!

8
AGRADECIMENTOS
À Deus por me guiar e me dar forças durante todo o caminho.
À Escola de Nutrição por todo aprendizado e por tornar possível este sonho.
À querida amiga Juliana (Ju) pelo companheirismo constante durante essa minha
trajetória. Para a nossa felicidade o destino nos colocou no mesmo lugar e, mesmo sem
te conhecer, já sabia que a partir daquele momento nos tornaríamos grandes amigas.
Às grandes amigas do Laboratório de Bioquímica Metabólica e Nutrição Experimental
Joyce, Maisa, Renata e Mariana por todo apoio, carinho e dedicação durante o
mestrado. Sem a ajuda de vocês esse momento não seria possível. Muito obrigada!
Aos demais colegas do LBM e Labnex pelos auxílios diários.
As minhas queridas amigas do mestrado por me escutarem sempre e por tornarem os
meus dias mais leves. Sentirei falta de todas vocês!
As minhas irmãs de alma Raíssa e Aninha pela amizade e companheirismo de sempre!
À toda minha família pela torcida e por acreditarem em mim!
Ao Jair e ao Clodoaldo pela atenção dedicada e pelas ajudas no laboratório.
Ao professor Wanderson pelo auxílio durante o experimento.
A todos os professores do Programa de Pós-Graduação em Saúde e Nutrição pelos
ensinamentos.

9
RESUMO
O Buriti (Mauritia flexuosa) é fruto do cerrado e a sua polpa possui quantidades
consideráveis de carotenóides, polifenóis, ácido ascórbico e ácidos graxos mono e poli-
insaturados, apresentando, assim, potencial para ser utilizado na prevenção de doenças
advindas do estresse oxidativo. O diabetes mellitus (DM) é um dos mais importantes
problemas de saúde pública e o desenvolvimento das suas complicações está, entre outros
fatores, intimamente relacionado ao aumento do estresse oxidativo. Portanto, há grande
interesse em compostos e/ou alimentos com capacidade de diminuir espécies reativas de
oxigênio (ROS) nos tecido. Desse modo, o objetivo do nosso estudo consiste em
investigar a capacidade antioxidante da farinha de buriti in vitro e em ratos diabéticos.
A capacidade antioxidante da farinha de buriti foi determinada através do método
do DPPH. A composição centesimal foi realizada segundo métodos da AOAC (1984). A
quantidade de polifenóis foi determinada pelo método Folin-Ciocalteu. Para o ensaio
biológico utilizaram-se 36 ratos Fisher divididos em quatro grupos: Controle (C);
Controle + Buriti (CB); Diabéticos (D); Diabético + Buriti (DB). Os grupos C e CB
receberam dieta padrão (AIN-93M) e os grupos D e DB receberam dieta padrão (AIN-
93M) contendo 2% da farinha de buriti. O diabetes foi induzido no 1º dia do experimento
por injeção intraperitoneal de estreptozotocina (STZ) 35 mg/Kg. No 25º dia do
experimento os ratos foram submetidos ao Teste de Tolerância Oral à Glicose (TTOG).
Após 30 dias de tratamento os animais foram anestesiados com isoflurano e eutanasiados.
Foram realizadas análises séricas de metabólitos, produtos de excreção e enzimas.
Analisaram-se danos oxidativos às biomoléculas através da determinação de proteína
carbonilada e hidroperóxidos lipídicos no fígado, coração, pâncreas e rim. Além disso,
realizaram-se análises histológicas do fígado, coração e pâncreas utilizando a coloração
Hematoxilina e Eosina (H&E). Os resultados foram analisados por ANOVA one way ou
Teste T e Kruskal Wallis ou Mann-Whitney considerando significativos quando p <0,05.
A composição centesimal demonstrou que farinha de buriti possui elevadas
quantidades de lipídios totais e fibra insolúvel. Além disso, apresentou uma concentração
de polifenóis de 444,28 ± 14,76 mg EAG/100g. No ensaio in vitro ela exibiu uma alta
capacidade de neutralização do radical DPPH. Já no experimento in vivo iniciamos
observando, no TTOG, que os grupos D e DB apresentaram níveis maiores de glicose em
todos os tempos comparados aos grupos C e CB. Encontramos, no soro, níveis elevados
de AST, ALT, uréia e níveis reduzidos de albumina, proteínas totais, colesterol total e

10
triacilgliceróis nos grupos D e DB. Com relação aos biomarcadores do estresse oxidativo,
a farinha de buriti reduziu, nos ratos diabéticos, os níveis de TBARS no coração e no rim,
e os níveis de proteínas carboniladas no fígado e no coração. No exame histopatológico
não observamos alterações em nenhum dos tecidos analisados.
Portanto, concluímos que a farinha de buriti apresentou um elevado conteúdo de
polifenóis totais e uma capacidade antioxidante in vitro. Já in vivo, observamos que a
suplementação com farinha de buriti promoveu uma redução dos biomarcadores do
estresse oxidativo TBARS e/ou proteína carbonilada no fígado, coração e rim. Porém,
não foi evidenciado nenhuma alteração a nível celular.
Palavras-chave: Buriti, Diabetes, Estresse Oxidativo, Ratos.

11
ABSTRACT
The Buriti (Mauritia flexuosa) is the result of the Cerrado and its pulp has
considerable amounts of carotenoids, polyphenols, ascorbic acid and fatty acid mono and
poly-unsaturated, thus presenting the potential to be used to prevent diseases from
oxidative stress. Diabetes mellitus (DM) is one of the most important public health
problems and the development of its complications is, among other factors, closely related
to increased oxidative stress. Therefore, there is great interest in compounds and/or foods
with ability to decrease reactive oxygen species (ROS) in tissue. Thus, the aim of our
study is to investigate the antioxidant capacity of Buriti flour in vitro and in diabetic rats.
The antioxidant capacity of Buriti flour was determined by the DPPH method.
The chemical composition was performed according to AOAC (1984). The amount of
polyphenols was determined by Folin-Ciocalteu method. For the biological assay we used
36 Fisher rats divided into four groups: Control (C), control + Buriti (CB); Diabetics (D)
Diabetic + Buriti (DB). C and CB groups received standard diet (AIN-93M) and D and
DB groups received standard diet (AIN-93M) containing 2% of Buriti flour. Diabetes was
induced on day 1 of the experiment by intraperitoneal injection of streptozotocin (STZ)
35 mg / kg. On the 25th day of the experiment the rats were submitted to the Oral Glucose
Tolerance Test (OGTT). After 30 days of treatment the animals were anesthetized with
isoflurane and euthanized. Analysis of serum metabolites, waste products and enzymes
were performed. We analyzed oxidative damage to biomolecules by measuring protein
carbonyl and lipid hydroperoxides in the liver, heart, pancreas and kidney. Furthermore,
histological analyzes were performed liver, heart and pancreas using hematoxylin and
eosin (H&E) staining. The results were analyzed by one-way ANOVA or t test and
Kruskal Wallis or Mann-Whitney considering significant when p < 0.05.
The centesimal composition showed that buriti flour has high amounts of total
lipids and insoluble fiber. Furthermore, submitted a concentration of polyphenols 444.28
± 14.76 mg EAG/100g. In vitro assay it exhibited a high neutralizing capacity DPPH. In
the in vivo experiment we observe initially that the D and DB groups had higher glucose
levels at all times compared to groups C and CB (OGTT). Found in serum increased levels
of AST, ALT, urea and reduced levels of albumin, total protein, total cholesterol and
triglycerides in groups D and DB. With respect to biomarkers of oxidative stress, buriti
flour reduced in diabetic rats, levels of TBARS in the heart and kidney, and levels of

12
protein carbonyls in liver and heart. In histopathological examination did not observe
changes in any of the tissues analyzed.
Therefore, we conclude that the buriti flour showed a high content of total
polyphenols and antioxidant capacity in vitro. Already in vivo, we observed that
supplementation with buriti flour promoted a reduction in biomarkers of oxidative stress
TBARS and/or protein carbonyl in the liver, heart and kidney. However, was not made
any changes at the cellular level.
Already in vivo, we observed that supplementation with Buriti flour promoted a
reduction in biomarkers of oxidative stress TBARS and/or protein carbonyl in the liver,
heart and kidney. However, was not made any changes at the cellular level.
Keywors: Buriti, Diabetes, Oxidative Stress, Rats.

13
LISTA DE TABELAS
Tabela 1: Rendimento do Fruto Buriti ............................................................................ 5
Tabela 2: Composição Química da Polpa de Buriti (%). ................................................ 5
Tabela 3: Composição das partes do fruto Buriti. ........................................................... 6
Tabela 4: Teores de Polifenóis e Ácido Ascórbico presentes na polpa de buriti
expressos em mg/100g de amostra. .................................................................................. 9
Tabela 5: Teor de carotenóides totais e dos principais carotenóides da polpa de buriti.
........................................................................................................................................ 11
Tabela 6: Ações e mecanismos de diversas substâncias antioxidantes. ........................ 18
Tabela 7: Composição das dietas experimentais (g/1000g de dieta). ............................ 26
Tabela 8: Composição Centesimal em 100g da Farinha de Buriti (Mauritia flexuosa). 35
Tabela 9: Avaliação do peso corporal inicial, peso corporal final e glicemia plasmática
dos ratos do grupo controle (C), controle + buriti (CB), diabetes (D) e diabetes + buriti
(DB). ............................................................................................................................... 38
Tabela 10: Peso Relativo do Fígado e Rim, Metabólicos e Enzimas Séricas (uréia,
creatinina, AST e ALT) nos grupos C, CB, D e DB. ..................................................... 38

14
LISTA DE FIGURAS
Figura 1: Palmeira Mauritia flexuosa L. ......................................................................... 3
Figura 2: Frutos do Buriti ................................................................................................ 4
Figura 3: Buriti: a) fruto inteiro e b) corte longitudinal do fruto. ................................... 4
Figura 4: A: Buriti íntegro; B: Buriti sem a casca; C: Parte fibrosa; D: Semente. ......... 4
Figura 5: Comparação dos principais ácidos graxos do óleo de buriti com azeite de
oliva e óleo de canola. ...................................................................................................... 8
Figura 6: Visão geral da relação entre a produção de espécies reativas, o
desenvolvimento de doenças crônicas e do papel dos antioxidantes e da variação
genética sobre a modulação do estresse oxidativo. ........................................................ 15
Figura 7: Estrutura Química da STZ. ............................................................................ 16
Figura 8: Sistema antioxidante enzimático. .................................................................. 19
Figura 9: Sistema Keap1-Nrf2-ARE regulados. ........................................................... 20
Figura 10: Reação do ácido tiobarbitúrico com malondialdeído para produzir um
produto final com coloração detectada na absorbância de 532 nm. ............................... 21
Figura 11: Reação do DNPH com uma proteína carbonilada gerando um produto final
com coloração detectada na absorbância de 370nm. ...................................................... 22
Figura 12: Farinha de buriti adquirida na loja Néctar do Cerrado. ............................... 24
Figura 13: Delineamento amostral e linha do tempo do experimento. ......................... 27
Figura 14: Percentual de inibição do radical DPPH, segundo diferentes concentrações
da farinha de buriti e do antioxidante padrão Trolox, após 30 minutos. ........................ 36
Figura 15:Teste de Tolerância Oral à Glicose ............................................................... 37
Figura 16: Peso do coração e do pâncreas de ratos controle e diabéticos ..................... 39
Figura 17: Níveis de albumina e proteínas totais de ratos controle e diabéticos ......... 39
Figura 18: Níveis de colesterol total e triacilgliceróis de ratos controle e diabéticos ... 40
Figura 19: Atividade das enzimas PON Fenilacetato e PON Paraoxon de ratos controle
e diabéticos ..................................................................................................................... 40
Figura 20: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) e proteínas
carboniladas em tecido hepático de ratos controle e diabéticos ..................................... 41
Figura 21: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) e proteínas
carboniladas em tecido cardíaco de ratos controle e diabéticos ..................................... 42
Figura 22: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) e proteínas
carboniladas em tecido pancreático de ratos controle e diabéticos. ............................... 42

15
Figura 23: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) e proteínas
carboniladas em tecido renal de ratos controle e diabéticos ........................................... 43
Figura 24: Fotomicrografias representativas do fígado................................................. 44
Figura 25: Fotomicrografias representativas do coração. ............................................. 44
Figura 26: Fotomicrografias representativas do pâncreas ............................................. 45
Figura 27: Número de ilhotas pancreáticas e área das ilhotas em ratos controle e
diabéticos. ....................................................................................................................... 45

16
LISTA DE ABREVIATURAS
AGES: Produtos finais de glicação avançada
ARE: Elementos de resposta antioxidante
ALT: Alanina aminotransferase
ANOVA: Análise de variância
ANVISA: Agência Nacional de Vigilância Sanitária
ALT: Alanina aminotransferase
AOAC: Association of Official Agricultural Chemists
AST: Aspartato aminotransferase
C: Grupo de animais que receberam dieta padrão
CB: Grupo de animais que recebeu dieta padrão acrescida de farinha de buriti
CAT: Catalase
D: Grupo de animais diabetéticos que receberam dieta padrão
DB: Grupo de animais diabetéticos que receberam dieta padrão acrescida de farinha de
buriti
DHA: Ácido docosahexaenóico
DM: Diabetes Mellitus
DNPH: 2,4 dinitrofenilhidrazina
DPPH: 2,2-difenil-1picril-hidrazil
EDTA: Ácido etilenodiaminotetracético
EPA: Ácido eicosapentaenoico
ER: Equivalente de Retinol
FDN: Fibra em detergente neutro
GPx: Glutationa peroxidase
GPx 1: glutationa peroxidase 1
GPx 2: Glutationa peroxidase 2
GSH: Glutationa reduzida
GSSG: Glutationa oxidada
GR: Glutationa redutase
HCl: Ácido clorídrico
HO-1: Heme oxigenase 1
Keap1: Kelch-like ECH-associated protein 1
HDL: High density lipoprotein

17
H2O2: Peróxido de hidrogênio
LDL: Low density lipoprotein
MDA: Malondialdeído
NAS: National Academy Sciences
NO: Óxido nítrico
Nrf2: Fator nuclear de transcrição
OMS: Organização Mundial da Saúde
PARP I: Poli ADP-ribose polimerase I
PKC: Proteína quinase C
PON: Enzima Paraoxonase
ROS: Espécies reativas de oxigênio
SOD: Superóxido dismutase
STZ: Estreptozotocina
TBA: Ácido tiobarbitúrico
TBARS: Substâncias reativas ao ácido tiobarbitúrico
TCA: Ácido tricloroacético
TEAC: Capacidade Antioxidante de Equivalente de Trolox
TTOG: Teste de Tolerância Oral a Glicose
Trolox: 6-hidroxi-2,5,7,8-tetrametilcromo-2-ácido carboxílico
TRX: Tiorredoxina
UDPGIcNAc: Uridina difosfato-N-acetil glucosamina
γ-GCS: γ-glutamilcisteína sintetase

18
SUMÁRIO
1. INTRODUÇÃO ...................................................................................................... 1
2. REVISÃO BIBLIOGRÁFICA .............................................................................. 3
2.1. O Buriti: Caracterização e seu Papel como Alimento Funcional ...................... 3
2.1.1. Caracterização Biológica ............................................................................ 3
2.1.2. Composição Química ................................................................................. 5
2.1.3. Buriti como Potencial Alimento Funcional ................................................ 7
2.1.4. Estudos com o fruto buriti in vitro e in vivo ............................................. 11
2.2. Diabetes Mellitus e Estresse Oxidativo ........................................................... 13
2.2.1. Diabetes Experimental Induzida por Estreptozotocina (STZ) .................. 16
2.3. Ações Antioxidantes do Organismo e Marcadores do Estresse Oxidativo ..... 17
2.3.1. Biomarcadores do Estresse Oxidativo ...................................................... 21
3. OBJETIVOS ......................................................................................................... 23
3.1. Objetivo Geral .................................................................................................. 23
3.2. Objetivos Específicos ...................................................................................... 23
4. MATERIAL E MÉTODOS ................................................................................. 24
4.1. Amostra de Buriti ............................................................................................. 24
4.2. Composição Centesimal .................................................................................. 24
4.3. Capacidade Antioxidante do Buriti .................................................................. 25
4.4. Animais ............................................................................................................ 26
4.5. Dieta ................................................................................................................. 26
4.6. Delineamento Experimental ............................................................................ 27
4.7. Teste de Tolerância Oral a Glicose (TTOG).................................................... 28
4.8. Análises Bioquímicas no Soro e Plasma .......................................................... 28
4.8.1. Atividade de Paraoxonase ........................................................................ 29
4.9. Determinação de Marcadores do Estresse Oxidativo em Fragmentos de Tecido
31
4.9.1. Proteína Carbonilada ................................................................................ 31
4.9.2. TBARS – Substâncias Reativas ao Ácido Tiobarbitúrico ........................ 32
4.10. Proteínas Totais ............................................................................................ 33
4.11. Análises Histológicas ................................................................................... 34
4.12. Análise Estatística ........................................................................................ 34
5. RESULTADOS ..................................................................................................... 35
5.1. Composição Centesimal e Fitoquímica da Farinha de Buriti .......................... 35
5.2. Capacidade Antioxidante in vitro contra o Radical DPPH .............................. 35

19
5.3. Teste de Tolerância Oral a Glicose (TTOG).................................................... 36
5.4. Peso Inicial e Final dos Animais e Níveis Plasmáticos de Glicose ................. 37
5.5. Peso dos Órgãos e Níveis Plasmáticos de AST, ALT, Uréia e Creatinina ...... 38
5.6. Metabólitos e Enzimas Séricas ........................................................................ 39
5.7. Biomarcadores do Estresse Oxidativo ............................................................. 41
5.7.1. Fígado ....................................................................................................... 41
5.7.2. Coração ..................................................................................................... 41
5.7.3. Pâncreas .................................................................................................... 42
5.7.4. Rim ........................................................................................................... 43
5.8. Exame Histopatológico do Fígado, Coração e Pâncreas ................................. 43
6. DISCUSSÃO ......................................................................................................... 46
7. CONCLUSÃO ....................................................................................................... 54
8. LIMITAÇÕES DO ESTUDO .............................................................................. 55
9. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................... 56
10. ANEXOS ............................................................................................................... 71

1
1. INTRODUÇÃO
Alterações no perfil alimentar e nutricional, aliado a mudanças econômicas, sociais e
demográficas e suas repercussões na saúde populacional, vêm sendo observadas em
diversos países em desenvolvimento (MONTEIRO, 2001). Recentemente, evidências
mostraram que características qualitativas da dieta são igualmente importantes na
definição do estado de saúde, em particular no que se refere a doenças crônicas da idade
adulta (MONTEIRO et al, 2000a). Assim, torna-se importante a busca de alimentos que
contenham quantidades significativas de nutrientes e que possam ser reconhecidos como
alimentos funcionais, ampliando sua oferta à população.
Não existe uma definição universal para alimentos funcionais. De forma
generalista, são alimentos para os quais pode ser satisfatoriamente demonstrado que eles
afetam beneficamente uma ou mais funções do organismo, além de garantirem efeitos
nutricionais adequados, conduzindo a uma melhoria do estado geral de saúde e bem estar
ou a uma redução do risco de doenças (DIPLOCK et al, 1999).
O que caracteriza um alimento funcional são os compostos bioativos presentes nele,
tais como carotenóides, vitaminas E e C, metabólitos fenólicos e ácidos graxos
poliinsaturados (ANJO, 2004; MORAES e COLLA, 2006). Vários são os mecanismos
pelos quais estes compostos exercem seus efeitos entre eles destaca-se a ação
antioxidante.
O Buriti (Mauritia flexuosa L. f.) é uma palmeira que pertence à família Palmae
ou Arecaceae e a subfamília Lepidocarycideae. No Brasil pertence à flora do cerrado e
pode ser encontrado nos estados do Pará, Amazonas, Maranhão, Piauí, Bahia, Ceará e
Tocantins (SANTOS et al, 2011). Um levantamento da National Academy Sciences
(NAS) das espécies vegetais na Amazônia e com grande potencial sócio-econômico,
concluiu que essa palmácea é uma das mais abundantes na América do Sul, tendo como
produtos potenciais, a polpa e produtos derivados como néctares, doces, bebidas
fermentadas, além do óleo da polpa e da semente, o amido do tronco da árvore e a madeira
para construção e fibra industrial (MANHÃES, 2007).
A polpa de buriti possui quantidades consideráveis de carotenóides, polifenóis e
ácido ascórbico, apresentando, assim, potencial para ser utilizado na prevenção de
doenças advindas do estresse oxidativo. Sua fração lipídica é composta basicamente de
tocoferol e óleos com predominância dos ácidos graxos oléico e palmítico, que ajudam

2
na prevenção de doenças cardiovasculares (MANHÃES, 2007; BARRETO, BENASSI e
MERCADANTE, 2009). Ainda são poucos os estudos que descrevem atividades
funcionais do buriti, mas os já existentes na literatura demonstram seu potencial
fotoprotetivo devido a presença dos carotenóides e sua atividade antibacteriana e
cicatrizante (ZANATTA et al, 2009; BATISTA et al, 2012; YUYAMA et al, 1998,
MANHÃES, 2007).
Neste contexto, este estudo avaliou os efeitos benéficos do buriti especialmente
relacionados à capacidade antioxidante in vitro da farinha de buriti e in vivo através da
avaliação de biomarcadores do estresse oxidativo em tecido hepático, cardíaco,
pancreático e renal em modelo experimental de diabetes, considerando que o
desenvolvimento das complicações do DM está, entre outros fatores, intimamente
relacionado ao aumento do estresse oxidativo (STADLER et al. 2004).

3
2. REVISÃO BIBLIOGRÁFICA
2.1.O Buriti: Caracterização e seu Papel como Alimento Funcional
2.1.1. Caracterização Botânica
O Buriti (Mauritia flexuosa) é uma palmeira que pertence à família Palmae ou
Arecaceae e a subfamília Lepidocarycideae (Figura 1), pode ser encontrado na América
do Sul, e no Brasil nos estados do Pará, Amazonas, Maranhão, Piauí, Bahia, Ceará e
Tocantins (SANTOS et al, 2011). Esta palmeira tem em torno de 15-20 m de altura e é
típica de regiões que possuem rios e ilhas (DARNET et al, 2011). Essa espécie prospera
mesmo em solos ácidos, pobres em nutrientes e inundados, o que geralmente é
inadequado para a agricultura. A palmeira tem importância estratégica na preservação da
vida selvagem, uma vez que o seu fruto é uma fonte de alimento para pássaros e
mamíferos (GAZEL-FILHO e LIMA, 2001).
É um fruto sazonal, onde sua frutificação em maior escala ocorre nos meses de
dezembro a junho na maioria das regiões (CARNEIRO e CARNEIRO, 2011).
Um levantamento da National Academy Sciences (NAS) das espécies vegetais na
Amazônia e com grande potencial sócio-econômico, concluiu que essa palmácea é uma
das mais abundantes na América do Sul, tendo como produtos potenciais, a polpa e
produtos derivados como néctares, doces, bebidas fermentadas, além do óleo da polpa e
da semente, o amido do tronco da árvore e principalmente a madeira para construção e
fibra industrial para barbante, rede de pesca e rede para dormir (MANHÃES, 2007).
Figura 1: Palmeira Mauritia flexuosa L.

4
Figura 2: Frutos do Buriti
Seu fruto pesa cerca de 75g (DARNET et al, 2011) e, como pode ser visto na
Figura 2, é constituído de uma drupa globoso-alongada de 4-7 cm de comprimento
constituída de epicarpo formado de escamas rombóides de cor castanho-avermelhada,
mesocarpo representado por uma massa espessa de cor alaranjada e endocarpo esponjoso
que envolve a semente (SANTOS et al, 2011).
A Figura 3 mostra em “a” o fruto do Buriti inteiro e em “b” uma secção deste,
podendo ser visualizado as seguintes partes: casca, polpa, parte fibrosa e semente (MELO
et al, 2011) e a Figura 4 mostra essas quatro partes separadamente.
Figura 3: Buriti: a) fruto inteiro e b) corte longitudinal do fruto.
Fonte: MELO et al, 2011.
Figura 4: A: Buriti íntegro; B: Buriti sem a casca; C: Parte fibrosa; D: Semente.
(Arquivo Pessoal)
A B C D
5
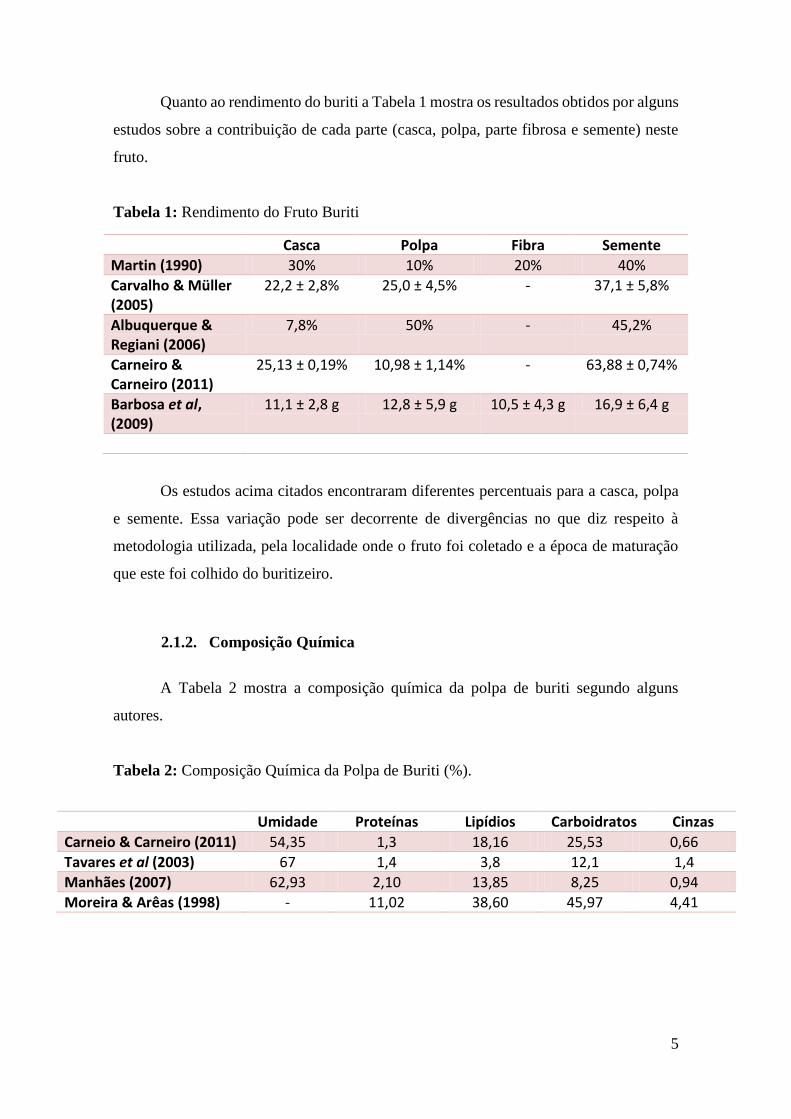
Quanto ao rendimento do buriti a Tabela 1 mostra os resultados obtidos por alguns
estudos sobre a contribuição de cada parte (casca, polpa, parte fibrosa e semente) neste
fruto.
Tabela 1: Rendimento do Fruto Buriti
Casca Polpa Fibra Semente
Martin (1990) 30% 10% 20% 40%
Carvalho & Müller (2005)
22,2 ± 2,8% 25,0 ± 4,5% - 37,1 ± 5,8%
Albuquerque & Regiani (2006)
7,8% 50% - 45,2%
Carneiro & Carneiro (2011)
25,13 ± 0,19% 10,98 ± 1,14% - 63,88 ± 0,74%
Barbosa et al, (2009)
11,1 ± 2,8 g 12,8 ± 5,9 g 10,5 ± 4,3 g 16,9 ± 6,4 g
Os estudos acima citados encontraram diferentes percentuais para a casca, polpa
e semente. Essa variação pode ser decorrente de divergências no que diz respeito à
metodologia utilizada, pela localidade onde o fruto foi coletado e a época de maturação
que este foi colhido do buritizeiro.
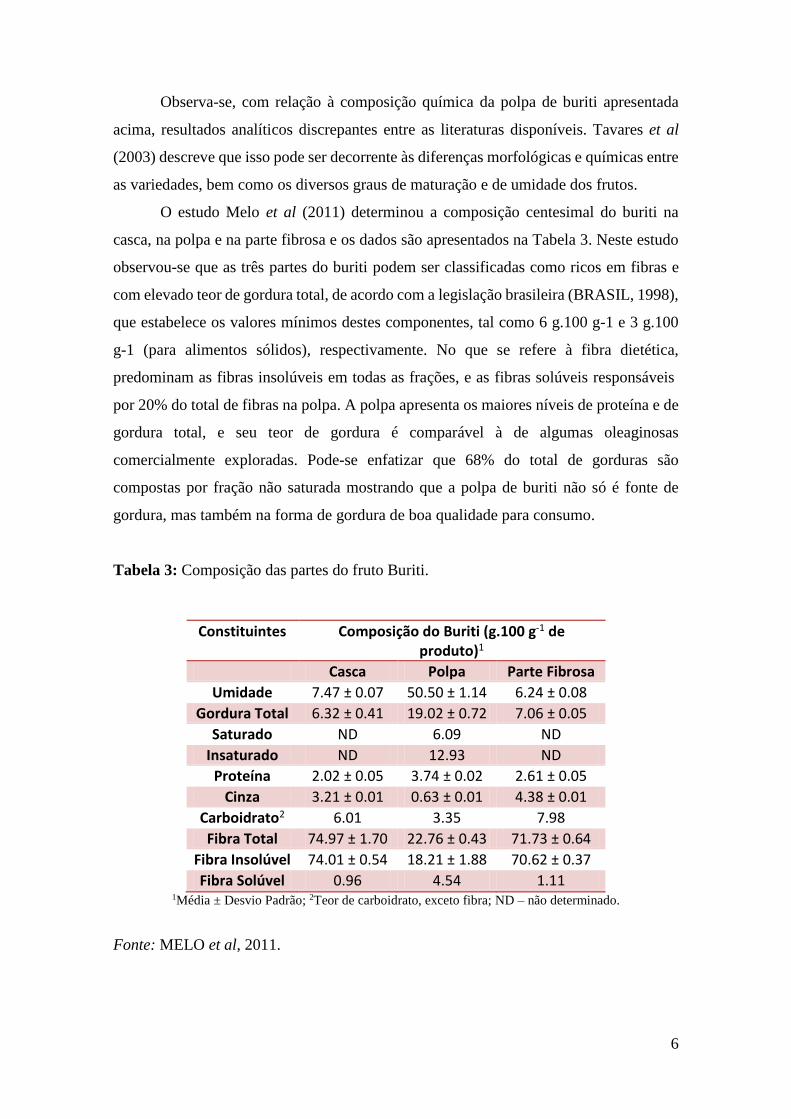
2.1.2. Composição Química
A Tabela 2 mostra a composição química da polpa de buriti segundo alguns
autores.
Tabela 2: Composição Química da Polpa de Buriti (%).
Umidade Proteínas Lipídios Carboidratos Cinzas
Carneio & Carneiro (2011) 54,35 1,3 18,16 25,53 0,66
Tavares et al (2003) 67 1,4 3,8 12,1 1,4
Manhães (2007) 62,93 2,10 13,85 8,25 0,94
Moreira & Arêas (1998) - 11,02 38,60 45,97 4,41
6
Observa-se, com relação à composição química da polpa de buriti apresentada
acima, resultados analíticos discrepantes entre as literaturas disponíveis. Tavares et al
(2003) descreve que isso pode ser decorrente às diferenças morfológicas e químicas entre
as variedades, bem como os diversos graus de maturação e de umidade dos frutos.
O estudo Melo et al (2011) determinou a composição centesimal do buriti na
casca, na polpa e na parte fibrosa e os dados são apresentados na Tabela 3. Neste estudo
observou-se que as três partes do buriti podem ser classificadas como ricos em fibras e
com elevado teor de gordura total, de acordo com a legislação brasileira (BRASIL, 1998),
que estabelece os valores mínimos destes componentes, tal como 6 g.100 g-1 e 3 g.100
g-1 (para alimentos sólidos), respectivamente. No que se refere à fibra dietética,
predominam as fibras insolúveis em todas as frações, e as fibras solúveis responsáveis
por 20% do total de fibras na polpa. A polpa apresenta os maiores níveis de proteína e de
gordura total, e seu teor de gordura é comparável à de algumas oleaginosas
comercialmente exploradas. Pode-se enfatizar que 68% do total de gorduras são
compostas por fração não saturada mostrando que a polpa de buriti não só é fonte de
gordura, mas também na forma de gordura de boa qualidade para consumo.
Tabela 3: Composição das partes do fruto Buriti.
Constituintes Composição do Buriti (g.100 g-1 de produto)1
Casca Polpa Parte Fibrosa
Umidade 7.47 ± 0.07 50.50 ± 1.14 6.24 ± 0.08
Gordura Total 6.32 ± 0.41 19.02 ± 0.72 7.06 ± 0.05
Saturado ND 6.09 ND
Insaturado ND 12.93 ND
Proteína 2.02 ± 0.05 3.74 ± 0.02 2.61 ± 0.05
Cinza 3.21 ± 0.01 0.63 ± 0.01 4.38 ± 0.01
Carboidrato2 6.01 3.35 7.98
Fibra Total 74.97 ± 1.70 22.76 ± 0.43 71.73 ± 0.64
Fibra Insolúvel 74.01 ± 0.54 18.21 ± 1.88 70.62 ± 0.37
Fibra Solúvel 0.96 4.54 1.11 1Média ± Desvio Padrão; 2Teor de carboidrato, exceto fibra; ND – não determinado.
Fonte: MELO et al, 2011.

7
Após a caracterização morfológica do buriti e a descrição dos macronutrientes
presentes neste fruto nos interessa saber quais os componentes bioativos que este contém
e, assim, reportar alguns estudos que mostram o buriti com potencial de ser um alimento
funcional.
2.1.3. Buriti como Potencial Alimento Funcional
A concepção de alimentos funcionais vem mudando no mundo ocidental, devido
ao aumento da associação entre dieta e doenças crônicas demonstrado por vários estudos.
Na década de 80 os japoneses começaram a difundir uma nova concepção de alimentos
através de um programa de governo que tinha como objetivo desenvolver alimentos
saudáveis para uma população que envelhecia e apresentava uma grande expectativa de
vida (ANJO, 2004). Desde então, muitos estudos foram sendo realizados afim de avaliar
os efeitos fisiológicos de vários alimentos e seus constituintes sobre a modulação dos
sistemas fisiológicos humanos (ARAI, 2007). Não existe uma definição universalmente
aceita para os alimentos funcionais. Porém, várias organizações vêm tentando criar uma
definição para essa classe de alimentos. Segundo Health Canada (1998) alimentos
funcionais consistem em “alimentos similares em aparência aos alimentos convencionais,
que são consumidos como parte da dieta habitual e demonstram benefícios fisiológicos
e/ou reduzem o risco de doenças crônicas além de possuírem funções nutricionais
básicas”. Já, no Brasil, a ANVISA (1999) define-os como “alimentos que podem, além
de cumprir com suas funções nutricionais básicas, produzir efeitos metabólicos e ou
fisiológicos e ou efeitos benéficos à saúde, devendo ser seguro para consumo sem
supervisão médica”. Uma outra definição do International Life Sciences Institute of North
América (ILSI, 1999) diz que são alimentos que, em virtude de ter componentes
alimentares bioativos, promovem mais benefícios à saúde, além da nutrição básica. Os
principais compostos bioativos presentes nos alimentos com ação funcional já
comprovada na literatura são os carotenóides, vitamina E e C, metabólitos fenólicos e
ácidos graxos poli-insaturados (ANJO, 2004; MORAES & COLLA, 2006).

8
2.1.3.1.Compostos Bioativos Presentes no Buriti
Ácidos Graxos Poliinsaturados
As famílias de ácidos graxos ômega-3 e ômega-6 consistem de ácidos graxos
poliinsaturados contendo de 18 a 22 carbonos. Os principais ácidos graxos n-3 são o ácido
linolênico, o ácido eicosapentaenoico (EPA) e o ácido docosahexaenóico (DHA),
enquanto os principais n-6 são o ácido linoléico e o ácido araquidônico (SUÁREZ-
MAHECHA et al, 2002). Pesquisas indicam que esses ácidos graxos desempenham
importante papel no organismo humano, uma vez que fazem parte da membrana celular,
têm ações antitrombóticas e anti-inflamatórias, atuando como precursores de
prostaglandinas e leucotrienos. Estão presentes em alimentos como, peixes e óleos
vegetais como a canola, milho, soja, linhaça. O ácido graxo oléico (ômega-9) vem
ganhando destque, uma vez que pode contribuir com a redução dos níveis séricos de
colesterol LDL e triacilgliceróis (MANHÃES, 2007).
O estudo de MANHÃES (2007) comparou a composição do óleo de buriti, em
relação às frações lipídicas, com dois óleos muito utilizados na dieta humana (azeite de
oliva e o óleo de canola) tidos como alimentos saudáveis, e encontrou que o óleo de buriti
possui níveis mais elevados de ácido graxo oléico (n-9) e contém aproximadamente
quatro vezes mais ácido linolênico (n-3) do que o azeite de oliva, como pode ser
visualizado na Figura 5.
Figura 5: Comparação dos principais ácidos graxos do óleo de buriti com azeite de oliva
e óleo de canola.
Fonte: MANHÃES, 2007.

9
Ácido Ascórbico e Polifenóis
O ácido ascórbico, também conhecido como vitamina C, é uma vitamina
hidrossolúvel, sintetizada por plantas e por quase todos os animais, exceto humanos
(PENTEADO, 2003). A vitamina C é essencial à saúde, pois desempenha papel
fundamental no desenvolvimento e regeneração dos músculos, pele, dentes e ossos, na
formação do colágeno, na regulação da temperatura corporal, na produção de diversos
hormônios e no metabolismo em geral (ANDRADE et al, 2002). A vitamina C é um
potente agente redutor capaz de reduzir a maioria das ROS/RNS fisiologicamente
relevantes (HALLIWELL e GUTTERIDGE, 2007), podendo atuar também como
sinergista na regeneração de antioxidantes primários (RAMALHO e JORGE, 2006).
Os polifenóis abrangem o maior grupo dentre os compostos bioativos nos
vegetais, sendo subdivididos em classes, de acordo com a estrutura química de cada
substância. Os principais grupos de polifenóis são os ácidos fenólicos, tendo como
exemplos: o ácido clorogênico, presente no café; os estilbenos, como o resveratrol
presente nas uvas e vinho; as cumarinas, como as furanocumarinas do aipo; as ligninas,
como as lignanas da linhaça; e os flavonóides. As ações fisiológicas desempenhadas pelos
polifenóis já foram relacionadas à prevenção de doenças cardiovasculares,
neurodegenerativas, câncer, entre outras, principalmente em função da sua elevada
capacidade antioxidante (FALLER e FIALHO, 2009).
Com relação aos teores de ácido ascórbico e de polifenóis na polpa de buriti o
estudo de Manhães (2007) encontrou que esta polpa contém teores de vitamina C
próximos aos da laranja e menores dos encontrados na couve. Já para os polifenóis
constatou-se que a polpa de buriti contém maior valor do que o encontrado na literatura
para cenoura e couve (Tabela 4).
Tabela 4: Teores de Polifenóis e Ácido Ascórbico presentes na polpa de buriti expressos
em mg/100g de amostra.
Antioxidantes Polpa de Buriti Laranja Cenoura
Polifenóis 9,47 - 0,171
Ác. Ascórbico 56,90 40-782 -
1CUNHA, 2005.
2SHILLS et al, 2003.
Fonte: MANHÃES, 2007.

10
Carotenóides
Os carotenóides compreendem uma família de compostos naturais, dos quais mais
de 600 variantes estruturais, sendo que as colorações vão desde o amarelo, passando pelo
laranja, até o vermelho intenso. Os mais comumente encontrados nos alimentos vegetais
são o β-caroteno (cenoura), licopeno (tomate), várias xantofilas (milho, manga, mamão e
gema de ovo) e a bixina (aditivo culinário e corante dérmico usado por indígenas
amazônicos, obtido do urucum). Estes compostos tem ação antioxidante, pois possuem a
capacidade de sequestrar oxigênio minimizando os danos causados por radicais livres
(FONTANA et al, 1997).
Na patogênese do DM tipo 2, uma relação entre controle glicêmico, estresse oxidativo
e complicações vasculares tem sido desde há muito tempo reconhecida e os carotenóides
(pró-vitamina A) podem desempenhar um papel na prevenção de DM tipo 2, devido à sua
propriedades antioxidantes (MARTINI et al, 2010). Um estudo observacional que
acompanhou por nove anos uma população idosa saudável quanto aos níveis de glicose
encontrou que o risco de disglicemia foi inferior nos participantes que possuíam maiores
níveis de carotenóides no plasma (AKBARALY et al, 2008). O estudo de Rossoni-Junior
et al (2012) realizado para avaliar os possíveis efeitos do urucum (fonte de carotenóide)
e da suplementação com β-caroteno sobre as espécies reativas de oxigênio e nitrogênio
em ratos diabéticos induzidos com aloxana, mostrou que os neutrófilos de animais
diabéticos produziram significativamente mais espécies reativas de oxigênio que os
animais dos grupos controles, porém a suplementação com β-caroteno e annatto foi capaz
de modular a produção destas espécies.
A Tabela 5 mostra os resultados de dois estudos distintos com relação ao teor de
carotenóides na polpa de buriti. Observa-se que o principal carotenóide desta polpa é o β-
caroteno que possui 100% da atividade pró-vitamina A, sendo que seus níveis foram
superiores aos encontrados na cenoura (MANHÃES, 2007).

11
Tabela 5: Teor de carotenóides totais e dos principais carotenóides da polpa de buriti.
Carotenóides Polpa Buriti (µg/100g)
Polpa Buriti1 (µg/100g)
Cenoura (µg/100g)
α-Caroteno 1480 ± 0,09 8010 0
β-Caroteno 13710 ± 0,75 35980 54001
Carotenóides Totais
23360 ± 0,98 - -
1GODOY & RODRIGUES – AMAYA, 1994. 2MANGELS et al, 1993.
Fonte: MANHÃES, 2007.
Yuyama et al (1998) encontrou na farinha de buriti um teor de carotenóides de
16,71 µg/g (fração alfa) e 110,46 µg/g (fração beta), perfazendo um total de 1976
ER/100g de vitamina A.
Um estudo caso-controle foi realizado em típicas regiões semi-áridas do Rio
Grande do Norte, as cidades de João Câmara, Parelhas e Jardim do Seridó, objetivando
avaliar a eficácia do doce de buriti no tratamento e prevenção da xeroftalmia em 44
crianças de 12 anos, através da suplementação diária com uma quantidade correspondente
a 134 µg de equivalentes de retinol (12 g do doce de buriti) durante 20 dias. Os resultados
desse estudo mostraram que o buriti é uma ótima fonte de vitamina A e que este pode
reverter os sinais clínicos da xeroftalmia, além de restaurar reservas hepáticas dessa
vitamina. Desse modo, estes resultados sugerem a possível utilização do buriti em
programas de combate a hipovitaminose A em países onde o fruto encontra-se disponível
ou têm potencial para cultivo (MARIATH et al, 1989).
2.1.4. Estudos com o fruto buriti in vitro e in vivo
Zanatta et al (2008) avaliaram os possíveis efeitos citotóxicos de cremes e loções
produzidas com óleo de Buriti (Mauritia flexuosa) em queratinócitos e fibroblastos
através do método de liberação vermelho neutro (ensaio NRR) e encontraram que as
emulsões com óleo de buriti apresentaram baixa citotoxicidade para as células em
concentrações elevadas e com a adição de vitamina E aumentou a viabilidade celular.
Outro estudo realizado pelos mesmos autores em 2010 para avaliar o potencial
fotoprotetor de emulsões formuladas com óleo de buriti contra a radiação UV em

12
queratinócitos e fibroblastos concluíram que as emulsões de óleo de buriti poderiam ser
consideradas potenciais veículos para o transporte de antioxidantes e, também, ser
utilizadas como adjuvantes em formulações de proteção solar.
O estudo de Batista et al (2012) avaliou a atividade antibacteriana in vitro,
utilizando cinco bactérias patogênicas incluindo espécies gram-positivas e espécies gram-
negativas mediante o uso do método de difusão em Agar, e cicatrizante do óleo de buriti
(M. flexuosa) em feridas realizadas em 40 ratos da linhagem Wistar, Os resultados obtidos
neste estudo mostraram, em relação à avaliação da atividade antibacteriana, que houve
inibição do crescimento bacteriano em quatro dos cinco patógenos testados. Quanto à
atividade cicatrizante avaliada in vivo, o óleo do fruto de buriti mostrou-se eficiente no
processo cicatricial de feridas cutâneas em ratos Wistar.
Um estudo experimental conduzido em 28 dias utilizando 48 ratos machos da
linhagem Wistar, para determinar a biodisponibilidade dos carotenoides do buriti in vivo,
seguindo o seguinte delineamento experimental: Grupo Deficiente (ração completa à base
de caseína sem adição de vitamina A na mistura vitamínica), Grupo Controle 1200 (ração
completa à base de caseína com adição de 1200 ER/Kg oriunda do palmitato de retinila),
Grupo Controle 2400 (ração completa à base de caseína com adição de 2400 ER/Kg
oriunda do palmitato de retinila), Grupo Buriti 1200 (ração completa à base de caseína
com adição de 1200 ER/Kg oriunda da farinha de buriti) e Grupo Buriti 2400 (ração
completa à base de caseína com adição de 2400 ER/Kg oriunda da farinha de buriti). Este
estudo encontrou que as reservas hepáticas de vitamina A foram significativamente
maiores nos grupos de animais que receberam dieta suplementada com buriti quando
comparado com os grupos Controle, demonstrando ser o buriti uma fonte de pró-vitamina
A altamente biodisponível (YUYAMA et al, 1998).
Como pôde ser visto, o buriti contém quantidades significativas de compostos
bioativos, confirmando que este fruto tem potencial antioxidante e já existente na
literatura estudos que demonstram seu potencial fotoprotetivo devido a presença dos
carotenóides e sua atividade antibacteriana e cicatrizante. Então, tendo em vista que o
DM é uma doença na qual provoca alterações metabólicas que culminam no aumento da
produção de ROS, qual seria o papel do buriti dentro deste contexto?

13
2.2.Diabetes Mellitus e Estresse Oxidativo
O diabetes mellitus (DM) consiste em uma desordem metabólica de múltiplas
etiologias e que se caracteriza por um estado de hiperglicemia crônica, decorrente da
anormalidade na secreção e/ou ação da insulina, com consequentes distúrbios no
metabolismo de carboidratos, lipídios e proteínas. É um dos mais importantes problemas
de saúde pública mundial e sua prevalência vem crescendo a cada ano. A Organização
Mundial da Saúde (OMS) prevê que até o ano 2025, 300 milhões de pessoas apresentarão
a doença. Neste contexto o Brasil se encontra entre os 10 países com maior número de
casos de diabetes no mundo (WILD et al. 2004).
A classificação do DM baseia-se na sua etiologia, eliminando os termos diabetes
melito insulinodependente e não-insulinodependente. Desse modo, tem-se que o DM do
tipo 1 resulta primariamente da destruição das células beta pancreáticas e tem tendência
à cetoacidose. Inclui casos decorrentes de doença auto-imune e aqueles nos quais a causa
da destruição das células beta não é conhecida. Corresponde a 5% a 10% do total de casos.
A forma rapidamente progressiva é comumente observada em crianças e adolescentes,
porém pode ocorrer também em adultos. Já o DM do tipo 2 resulta, em geral, de graus
variáveis de resistência à insulina e deficiência relativa de secreção de insulina. A maioria
dos pacientes tem excesso de peso e a cetoacidose ocorre apenas em situações especiais,
como durante infecções graves. O diagnóstico, na maioria dos casos, é feito a partir dos
40 anos de idade, embora possa ocorrer mais cedo, mais raramente em adolescentes.
Abrange 85% a 90% do total de casos (SOCIEDADE BRASILEIRA DE DIABETES,
2002).
A progressão da doença resulta em alterações patológicas e complicações que
envolvem diversos tecidos tais como, fígado, rim, retina, sistema cardiovascular e sistema
nervoso central e periférico (FAJANS et al. 1997). Tais complicações comprometem a
produtividade, qualidade de vida e sobrevida dos indivíduos, além de envolver altos
custos no seu tratamento.
O desenvolvimento das complicações da DM está, entre outros fatores,
intimamente relacionado ao aumento do estresse oxidativo. Alterações nos sistemas de
defesa antioxidante endógeno associadas ao aumento na produção de espécies reativas de
oxigênio, desempenham um papel fundamental no desenvolvimento do dano tecidual
inicial observado no diabetes, que posteriormente evolui para as complicações tardias da
doença (STADLER et al. 2003).

14
A produção de radicais livres consiste em um processo contínuo e fisiológico,
cumprindo funções biológicas relevantes. Nos processos metabólicos, esses radicais
atuam como mediadores para a transferência de elétrons nas várias reações bioquímicas.
A produção contínua de radicais livres durante os processos metabólicos é balanceada por
mecanismos de defesa antioxidante (FERREIRA e MATSUBARA, 1997; SHAMI e
MOREIRA, 2004). Assim, o processo de estresse oxidativo decorre da existência de um
desequilíbrio entre a produção de ROS e a capacidade antioxidante endógena, em favor
da geração excessiva de radicais livres ou em detrimento da velocidade de remoção
desses. O excedente de radicais livres gerado por esse desequilíbrio promove à oxidação
de biomoléculas com consequente perda de suas funções biológicas e/ou desequilíbrio
homeostático, cuja manifestação é o dano oxidativo potencial contra células e tecidos
(HALLIWELL e WHITEMAN, 2004).
Fatores externos e internos podem causar danos moleculares e resultar em estresse
oxidativo devido ao aumento na produção de espécies reativas de oxigênio (ROS) ou de
espécies reativas de nitrogênio (RNS). Esse aumento também pode alterar a expressão
gênica, levando a um quadro inflamatório que resulta em maior produção dessas espécies
reativas. Dessa forma, a inflamação aliada ao estresse oxidativo contribuem para o
desenvolvimento de doenças crônicas como câncer, doenças cardivasculares e diabetes
mellitus tipo 2, as quais também contribuem para o aumento na produção de ROS e RNS.
Os antioxidantes endógenos e da dieta trabalham juntos para minimizar esse quadro e,
consequentemente, o desenvolvimento de dano celular. Além disso, esse sistema de
defesa endógena e exógena é influenciado pela variação genética de cada indivíduo
(COSTA et al, 2012) (Figura 6).

15
Figura 6: Visão geral da relação entre a produção de espécies reativas, o
desenvolvimento de doenças crônicas e do papel dos antioxidantes e da variação genética
sobre a modulação do estresse oxidativo.
Fonte: COSTA et al, 2012
Muitas fontes estão relacionadas ao aumento do estresse oxidativo no diabetes,
tais como enzimáticas, não enzimáticas e vias mitocondriais. A hiperglicemia pode
estimular diretamente a produção de ROS através do aumento do fluxo de elétrons pela
cadeia respiratória. Cerca de 1 a 3% dos elétrons podem “escapar” da cadeia de transporte
e gerar o ânion superóxido, sob condições de hiperglicemia este processo é acentuado
(VALKO et al. 2006). As fontes não enzimáticas estão relacionadas a reações
bioquímicas de oxidação da glicose por vias secundárias a via glicolítica.
Uma via secundária que pode ocorrer no diabetes é a via de formação dos produtos
finais de glicação avançada (AGES). Os AGES são formados através do processo de
glicação, no qual a glicose, em excesso, reage com grupos amino livres de proteínas.
Existem três mecanismos de toxicidade dos AGES: interação com o receptor, deposição
no tecido e glicação in situ. Desse modo, eles podem desencadear nas células uma
resposta pró-inflamatória, pró-fibrótica e pró-coagulante capazes de danificar os tecidos
(HEIDLAND et al, 2001, DAROUX et al, 2010).
A hiperglicemia intracelular pode, também, favorecer a conversão enzimática da
glicose em sorbitol, através da via dos polióis, reduzindo as concentrações celulares de
NADPH e glutationa reduzida. Ou pode aumentar os níveis de diacilglicerol (DAG) e
ativar proteína quinase C (PKC) (KOYA & KING, 1998). Estudos demonstram que uma
hiperglicemia intracelular aumenta a quantidade de DAG em culturas de células
microvasculares, na retina e em glomérulos renais de animais diabéticos. E o aumento da

16
síntese de novo DAG está relacionado com a ativação de PKC, levando a consequências
patológicas, como por exemplo, estimulação de vias pró-inflamatórias por meio do NF-
κB e aumento da produção de ROS pela ativação da NADPH oxidase (BROWNLEE,
2001).
Portanto, medidas que reduzem esse aumento na produção de ROS, provocados
pela hiperglicemia no diabetes, têm ganhado destaque no meio científico. E uma
alternativa que tem sido bastante investigada é com relação aos efeitos provocados por
um alimento com potencial antioxidante sobre um modelo experimental de diabetes.
2.2.1. Diabetes Experimental Induzida por Estreptozotocina (STZ)
A estreptozotocina (STZ) é uma glicosamina-nitrosureia (Figura 7) que nas células
do pâncreas é convertida em metabólitos, um dos quais o óxido nítrico (NO). Ela é
captada pelas células pancreáticas que contêm transportadores de glicose GLUT-2.
Células produtoras de insulina que não expressam esse transportador são resistentes a
STZ (ELSNER et al, 2000). Vários são os mecanismos propostos para a citotoxicidade
produzida pela STZ. Um deles propõe que o NO reage com o oxigênio para produzir
N2O3 e outros óxidos de nitrogênio, que danificam o DNA e proteínas. Lesões no DNA
ativam a enzima de reparo chamada poli ADP-ribose polimerase I (PARP I), que utiliza
grandes quantidades de NAD+ como substrato e, eventualmente, o ATP, inibindo a síntese
e secreção de insulina (SZKUDELSKI, 2001).
Figura 7: Estrutura Química da STZ.

17
Alguns estudos realizados in vivo, por nosso grupo de pesquisa, comprovam o estresse
oxidativo gerado pela indução de diabetes por STZ. O estudo de Silva et al (2011) avaliou
os efeitos da estreptozotocina sobre os marcadores de estresse oxidativo em hamsteres e
observou maiores concentrações de substâncias reativas ao ácido tiobarbitúrico (TBARS)
no fígado e nos rins. Além disso, apresentou significativo aumento da concentração de
glutationa no fígado e menores atividades da paraoxonase e do superóxido dismutase.
Outro estudo de Guerra et al (2011) que avaliou o possível efeito protetor da polpa de
açaí sobre a produção de ROS por neutrófilos e sobre o sistema de defesa antioxidante
hepático em ratos controle e diabéticos induzidos por STZ mostrou que os ratos diabéticos
apresentaram redução na expressão de mRNA que codificam para Zn-superóxido
dismutase (Zn-SOD), glutationa peroxidase (GPx) e γ-glutamilcisteína sintetase (γ-GCS),
e aumento dos níveis de TBARS e proteínas carboniladas quando comparado aos ratos
controle. O estudo de Sampaio (2012), em ratos, também observou um aumento na
geração de EROS por neutrófilos no grupo diabético em relação ao grupo controle.
2.3.Ações Antioxidantes do Organismo e Marcadores do Estresse Oxidativo
Antioxidante, segundo uma definição ampla, é “qualquer substância que, presente
em baixas concentrações quando comparada a do substrato oxidável, atrasa ou inibe a
oxidação deste substrato de maneira eficaz” (SIES & STAHL, 1995). Ou seja, um
antioxidante consiste em uma molécula capaz de inibir a oxidação de outras moléculas.
E eles podem ser classificados em enzimáticos e não enzimáticos, como pode ser
visualizado na Tabela 6 (BARBOSA et al, 2010).

18
Tabela 6: Ações e mecanismos de diversas substâncias antioxidantes.
Antioxidantes Ação
Não enzimáticos (de origem dietética)
Vitamina A (β-caroteno) Proteção contra a oxidação de lipídeos e DNA
Vitamina C (ácido ascórbico) Inibição das EROs (agente reductor). Estimula o
poder antioxidante da vitamina E e selênio.
Proteção contra danos causados pela LDL-ox
Vitamina E (α-tocoferol) Proteção contra a peroxidação de ácidos graxos
insaturados da membrana celular e das LDL.
Converte O2• e H2O2 em formas menos reativas
Cu, Zn, Mn, Se Cofatores das enzimas antioxidantes SOD-Cu/Zn,
SOD-Mn e GSH-Pox
Outros carotenoides (licopeno) Proteção contra a oxidação de lipídeos, LDL,
proteínas e DNA. Sequestra e inativa os radicais
livres
Fitoquímicos (resveratrol, catequinas,
quercetinas, ácidos fenólicos e outros)
Proteção contra a oxidação de lipídeos e DNA
Enzimáticos
Superóxido Dismutase SOD-Cu/Zn (citoplasma), SOD-Mn (mitocôndria).
Catalisa a conversão do radical superóxido (O2•)
em peróxido de hidrogênio (H2O2)
Catase Catalisa a conversão de H2O2 em O2 e H2O
Glutationa Peroxidase Catalisa a redução do H2O2 a H2O
Fonte: BARBOSA et al, 2010
Sistema Enzimático
O sistema de defesa enzimático inclui as enzimas Superóxido Dismutase (SOD),
Catalase (CAT) e Glutationa Peroxidase (GPx) (Figura 8). A enzima SOD converte
ânions superóxido a peróxido de hidrogênio (H2O2). Este pode, então, ser rapidamente
removido pela GPx ou pela CAT, enzimas que agem com o mesmo propósito, o de
impedir o acúmulo de H2O2. A ação da GPx depende da manutenção do ciclo redox da
glutationa, por meio do controle da relação entre glutationa reduzida (GSH) e oxidada
(GSSG). A atividade dessas enzimas, na maioria das vezes, depende da participação de
cofatores enzimáticos, especialmente antioxidantes de origem dietética. Esses cofatores
podem se diferenciar de acordo com os compartimentos celulares de ação das enzimas. A
SOD pode ser encontrada sob duas formas: no citoplasma, é dependente de cobre e zinco
(SOD-Cu/Zn), enquanto na mitocôndria, necessita do manganês como cofator (SOD-
Mn). A GPx também existe sob duas formas: dependente e independente de selênio e
pode apresentar-se no citoplasma ou na mitocôndria (FERREIRA et al, 1997;

19
SCHNEIDER et al, 2004). A catalase é uma hemeproteína citoplasmática e é encontrada
no sangue, medula óssea, mucosas, rim e fígado. Sua atividade é dependente de NADPH
(SCOTT, 1991).
Figura 8: Sistema antioxidante enzimático.
O estresse oxidativo ativa uma resposta específica que inclui aumento da
expressão de moléculas antioxidantes endógenas. A transcrição da maioria dos genes
antioxidantes é regulada através do fator nuclear de transcrição (transcription factor
nuclear factor- E2–related factor -Nrf2) e elementos de resposta antioxidante (ARE) na
região promotora destes genes (MOI et al, 1994). Nrf2 normalmente está sequestrado no
citoplasma por “Kelch-like ECH-associated protein 1” - Keap1, uma proteína citossólica
ancorada à actina que facilita a degradação de Nrf2 pela via ubiquitina – proteossoma
(NGUYEN et al, 2003). Com o estresse oxidativo, Nrf2 dissocia de Keap1 e se desloca
para o núcleo, onde induz a transcrição de Genes “ARE-regulados”, incluindo ɣ-
glutamilcisteina sintetase (ɣ-GCS) (WILD et al, 1999) heme oxigenase 1 (HO-1) (1),
peroxirredoxina (PRX) (ISHI et al, 2000), glutationa peroxidase 2 (GPx2) (BANNING
et a,l 2005.) e tiorredoxina (TRX) (Figura 9).

20
Figura 9: Sistema Keap1-Nrf2-ARE regulados.
Fonte: GOLDRING, 2006.
Outra enzima que possui capacidade antioxidante é a arilesterase/paraoxonase
(PON). Ela está presente no soro humano e consiste numa enzima que catalisa a hidrólise
de organofosfatos, como por exemplo o paraoxon. Muitos desses organofosfatos são
neurotóxicos e amplamente encontrados na alimentação cotidiana. Estudos têm
demonstrado uma relação inversa entre a PON, aterosclerose e doença coronariana. Isso
devido ao fato de que essa enzima circula no plasma associada com a lipoproteína HDL,
contribuindo para o efeito protetor dessa lipoproteína contra a oxidação por LDL
(DURRINGTON et al, 2001). Em experimentos com HDL e PON, Aviram et al (2000),
determinaram que tanto a HDL associada a PON quanto a PON purificada são potentes
inibidores da oxidação por LDL. Seus experimentos implicaram num papel peroxidativo
para PON devido à sua hidrólise ativa de H2O2, uma das maiores espécies reativas de
oxigênio produzida durante o estresse oxidativo.
Sistema Não-Enzimático
O sistema de defesa não-enzimático inclui, especialmente, os compostos
antioxidantes de origem dietética, entre os quais se destacam: vitaminas, minerais e
compostos fenólicos. O ácido ascórbico (vitamina C), o α-tocoferol e β-caroteno,
precursores das vitaminas E e A, respectivamente, são compostos vitamínicos
potencialmente antioxidantes. Outros carotenóides sem atividade de vitamina A, como
licopeno, luteína e zeaxantina, também o são. Também são encontrados, frequentemente,

21
como antioxidantes os compostos fenólicos, tais como os flavonóides. Quanto aos
minerais destacam-se o zinco, cobre, selênio e magnésio que atuam como cofatores de
enzimas antioxidantes (BIANCHI e ANTUNES, 1999; BARBOSA et al, 2010).
2.3.1. Biomarcadores do Estresse Oxidativo
A quantificação das concentrações das enzimas e/ou moléculas antioxidantes e a
observação dos efeitos dos radicais livres sobre as moléculas celulares são meios indiretos
de avaliação do estresse oxidativo.
A maioria dos marcadores de dano oxidativo utilizado reflete o ataque dos radicais
livres a ácidos graxos poli-insaturados, com via clássica peroxidação lipídica,
hidroperóxidos e produtos finais de malondialdeído. A peroxidação lipídica é iniciada
pela reação de uma espécie reativa com um ácido graxo insaturado e propagada por
radicais peroxilas. Isso resulta na formação de hidroperóxidos lipídicos e aldeídos, tais
como o malondialdeído, 4-hidroxinonenal e isoprostanos, que podem ser detectados em
amostras biológicas e utilizados para se avaliar o estresse oxidativo. Uma das técnicas
mais utilizadas para se avaliar a oxidação de lipídios é através da determinação da
concentração de malondialdeído (MDA), o mais abundante aldeído resultante de
peroxidação lipídica e sua reação com o ácido tiobarbitúrico (TBA) forma um produto
que pode ser detectado espectrofotometricamente (Figura 10) (PUNCHARD e KELLY,
1996).
Figura 10: Reação do ácido tiobarbitúrico com malondialdeído para produzir um produto
final com coloração detectada na absorbância de 532 nm.
Proteínas também podem sofrer danos oxidativos. Como consequência, a modificação
oxidativa das proteínas pode ocorrer numa variedade de processos fisiológicos. Embora,
por vezes, a distinção é arbitrária, estas modificações podem ser primárias ou secundárias.
As modificações primárias ocorrem por reações catalisadas por metal; oxidação mediada

22
por radiação e na oxidação por ozônio ou óxidos de nitrogênio. Modificações secundárias
ocorrem quando as proteínas são modificadas por moléculas geradas pela oxidação de
outras moléculas. Um exemplo importante é a modificação covalente de proteínas por
hidroxinonenal produzido por oxidação de lipídios. Os grupos carbonilados podem ser
introduzidos em proteínas por qualquer uma destas reações, e o aparecimento desses
grupos evidencia modificação oxidativa das proteínas. Desse modo, a quantificação do
conteúdo de carbonilas consiste em um método muito utilizado para determinar oxidação
protéica e baseia-se na reação dos grupos carbonilados com 2,4-dinitrofenilhidrazina
(DNPH) (PUNCHARD e KELLY, 1996).
Figura 11: Reação do DNPH com uma proteína carbonilada gerando um produto final
com coloração detectada na absorbância de 370nm.
Considerando o papel de ROS no desenvolvimento das complicações do diabetes
e que o buriti contém componentes que podem controlar os efeitos do estresse oxidativo,
esse estudo foi realizado para avaliar se este fruto possui capacidade antioxidante in vitro
e se in vivo consegue reduzir danos oxidativos às biomoléculas em diferentes tecidos de
ratos diabéticos.

23
3. OBJETIVOS
3.1.Objetivo Geral
Avaliar o potencial antioxidante da farinha de buriti in vitro e em tecidos de ratos
diabéticos STZ-induzidos.
3.2.Objetivos Específicos
Este trabalho teve como intuito avaliar na farinha de buriti:
I. À composição centesimal e fitoquímica;
II. À capacidade antioxidante in vitro.
E avaliar no modelo in vivo:
III. O perfil glicêmico dos animais durante o experimento;
IV. O perfil sérico de alguns parâmetros bioquímicos;
V. Danos oxidativos às biomoléculas (proteína e lipídeo) no fígado, coração,
pâncreas e rim;
VI. À histologia dos tecidos cardíaco, pancreático e do fígado.

24
4. MATERIAL E MÉTODOS
4.1.Amostra de Buriti
No experimento utilizou-se a farinha de buriti (Mauritia flexuosa L.), um produto
artesanal brasileiro, produzida pela loja Néctar do Cerrado, Montes Claros, Minas Gerais.
Ela consiste de um produto natural, derivada da polpa desidratada de buriti e não possui
conservantes (Figura 12).
Figura 12: Farinha de buriti adquirida na loja Néctar do Cerrado.
4.2.Composição Centesimal
As determinações de umidade residual (105ºC), proteínas, lipídeos, cinzas e fibra
na farinha de buriti foram realizadas em triplicatas de amostras, segundo métodos da
AOAC (1984). As análises foram realizadas no Laboratório de Bromatologia do
Departamento de Alimentos da Escola de Nutrição, Universidade Federal de Ouro Preto
(UFOP).
Na determinação de umidade utilizou-se o método de secagem em estufa à
temperatura de 105ºC. As cinzas totais foram determinadas após as amostras serem
submetidas à incineração em mufla (500-600ºC). Para determinação das proteínas
utilizou-se o Método de Kjeldahl, ao qual consiste na determinação através de N total. Os
lipídeos foram determinados pela extração com solvente à quente, onde o extrator
utilizado foi do tipo Soxhlet. Para as fibras foi utilizado o método de determinação de
fibra em detergente neutro - aparelho digestor para fibra Tecnal/TE-149. Este método
consiste em um processo rápido para a determinação da fibra total em alimentos. A fibra
em detergente neutro (FDN) é constituída por celulose, hemicelulose e lignina. Por fim,
os carboidratos foram determinados por diferença (AOAC, 1995) (Anexo I).

25
4.3.Capacidade Antioxidante do Buriti
A capacidade antioxidante da farinha de buriti foi determinada através do método
do DPPH (2,2-difenil-1picril-hidrazil) (Brand-Williams et al, 1995). Esse método baseia-
se na transferência de elétrons de um composto antioxidante para o radical DPPH em
solução de metanol.
As determinações foram realizadas adicionando em tubos de ensaio, 3,9 mL de
solução de DPPH 60 µM, dissolvidos em metanol 80%, e 100 µL de solução com farinha
de buriti em diferentes concentrações (0,01; 0,02 e 0,05 g/mL) e de água destilada
(amostra controle).
Utilizou-se Trolox (6-hidroxi-2,5,7,8-tetrametilcromo-2-ácido carboxílico) como
antioxidante de referência. Do mesmo modo, adicionou-se em tubos de ensaio 3,9 mL de
solução de DPPH e 100 µL de solução de Trolox em diferentes concentrações (100, 200
e 400 µM) em metanol 80%.
Posteriormente, cada tubo foi homogeneizado e mantido na ausência de luz por
30 minutos à temperatura ambiente. A atividade foi determinada através da redução da
absorbância do radical DPPH a 515nm; água destilada foi utilizada como branco e o
percentual de inibição foi determinado de acordo com a equação:
% inibição do radical DPPH = (A controle 515 – A amostra 515 / A controle 515) x 100
Onde:
A controle 515: é a absorbância da solução de DPPH mais água destilada (sem a presença de
antioxidantes), obtida a 515nm.
A amostra 515: é a absorbância da solução de DPPH mais a solução com farinha de buriti ou
Trolox nas diferentes concentrações (amostra), obtida a 515nm.

26
4.4.Animais
Foram utilizados ratos Fischer fêmeas, com aproximadamente 80 dias de idade e
pesando cerca de 190 ± 2,0g provenientes do Laboratório de Nutrição Experimental da
Universidade Federal de Ouro Preto (UFOP). O protocolo relativo ao uso dos animais foi
aprovado pela Comissão de Ética no Uso de Animais (CEUA) - UFOP (Protocolo:
201148) (Anexo II).
4.5.Dieta
Os animais receberam dieta padrão AIN-93M (REEVES et al, 1993), ou dieta
padrão acrescida de 2% da farinha de buriti conforme apresentado na Tabela 7.
As dietas foram confeccionadas no Laboratório de Nutrição Experimental da
Escola de Nutrição, UFOP, acondicionadas em sacos plásticos e armazenadas em freezer
a -20ºC.
Tabela 7: Composição das dietas experimentais (g/1000g de dieta).
Componentes Dietas
Padrão Padrão + 2% Buriti
Amido de Milho 620,7 600,7
Caseína 140 140
Sacarose 100 100
Celulose 50 50
Mist. Minerais1 35 35
Mist. Vitaminas2 10 10
Colina 2,5 2,5
Cisteína 1,8 1,8
Óleo de Soja 40 40
Farinha de Buriti - 20 ¹ - Mistura de Minerais (expresso em g/kg da mistura): NaCl - 74/KI - 0,01/Citrato Tripotássico - 28/CaCO3 -
357/MnCO3 - 0,63/Citrato de Ferro - 6,06/ MgO - 24/ K2SO4 -46,6/ KH2PO4 - 250 / ZnCO3 - 1,65/CuCO3 -
0,3/Na2SeO4 - 0,01/(NH4)6MoO24 . 4 H2O - 0,00795.
² - Mistura de Vitaminas (expresso em g/kg da mistura): Niacina - 3/ Pantotenato de Cálcio - 1,6/Piridoxina HCl -
0,7/Tiamina HCl - 0,6/Riboflavina - 0,6/Ácido Fólico - 0,2/Biotina - 0,02/Cianocobalamina - 2,5/Vitamina E (500 IU/g)
- 15/Vitamina A (500.000 IU/g) - 0,8/Vitamina D (400.000 IU/g) - 0,25/Vitamina K - 0,075/Sacarose q.s.p. 1Kg.
27
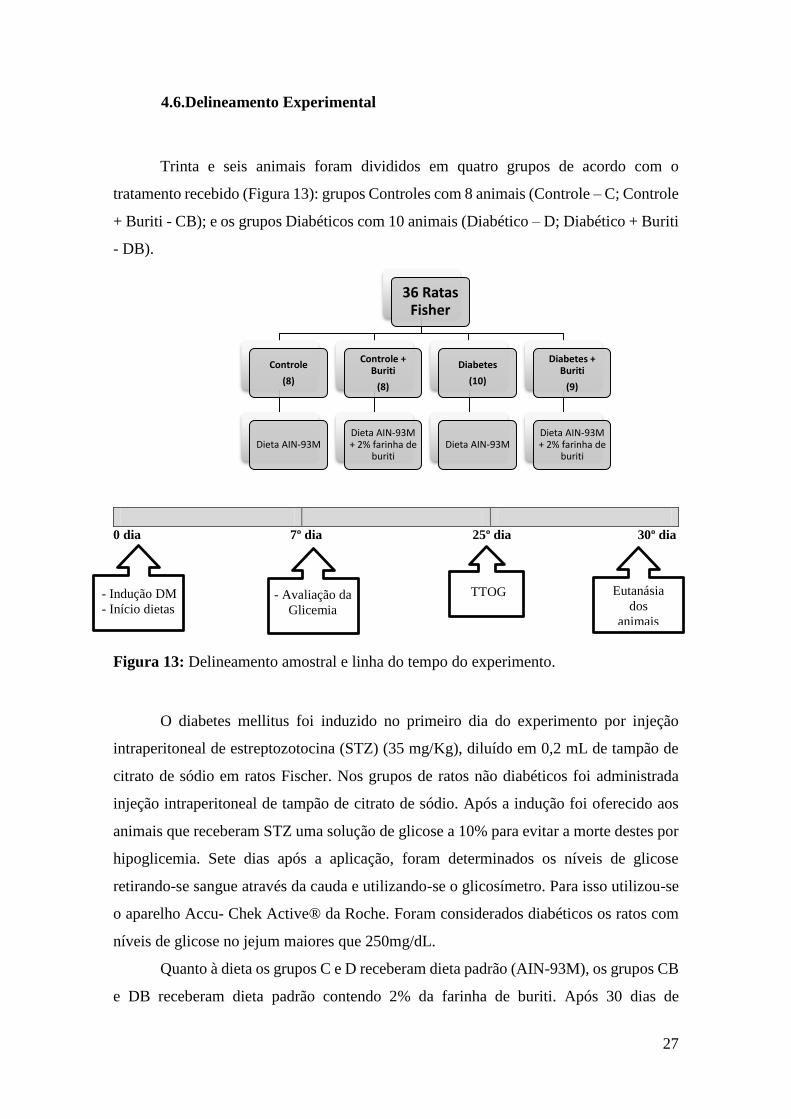
4.6.Delineamento Experimental
Trinta e seis animais foram divididos em quatro grupos de acordo com o
tratamento recebido (Figura 13): grupos Controles com 8 animais (Controle – C; Controle
+ Buriti - CB); e os grupos Diabéticos com 10 animais (Diabético – D; Diabético + Buriti
- DB).
0 dia 7º dia 25º dia 30º dia
Figura 13: Delineamento amostral e linha do tempo do experimento.
O diabetes mellitus foi induzido no primeiro dia do experimento por injeção
intraperitoneal de estreptozotocina (STZ) (35 mg/Kg), diluído em 0,2 mL de tampão de
citrato de sódio em ratos Fischer. Nos grupos de ratos não diabéticos foi administrada
injeção intraperitoneal de tampão de citrato de sódio. Após a indução foi oferecido aos
animais que receberam STZ uma solução de glicose a 10% para evitar a morte destes por
hipoglicemia. Sete dias após a aplicação, foram determinados os níveis de glicose
retirando-se sangue através da cauda e utilizando-se o glicosímetro. Para isso utilizou-se
o aparelho Accu- Chek Active® da Roche. Foram considerados diabéticos os ratos com
níveis de glicose no jejum maiores que 250mg/dL.
Quanto à dieta os grupos C e D receberam dieta padrão (AIN-93M), os grupos CB
e DB receberam dieta padrão contendo 2% da farinha de buriti. Após 30 dias de
36 Ratas Fisher
Controle
(8)
Dieta AIN-93M
Controle + Buriti
(8)
Dieta AIN-93M + 2% farinha de
buriti
Diabetes
(10)
Dieta AIN-93M
Diabetes + Buriti
(9)
Dieta AIN-93M + 2% farinha de
buriti
- Indução DM
- Início dietas
- Avaliação da
Glicemia
TTOG
Eutanásia
dos
animais

28
tratamento os animais foram deixados em jejum por 12 horas, anestesiados com
isoflurano e eutanasiados. O sangue foi recolhido para obtenção do soro e plasma e os
órgãos: fígado, pâncreas, coração e rins foram coletados imersos em nitrogênio líquido e
armazenados em freezer - 80oC para análises posteriores sendo que um fragmento do
fígado, coração e pâncreas foi armazenado em formol tamponado para realização de
análises histológicas.
Durante a realização do experimento, os animais foram mantidos em ambiente
arejado, com controle de temperatura, umidade e ventilação, e receberam água ad libitum.
4.7.Teste de Tolerância Oral a Glicose (TTOG)
O TTOG foi realizado no 25º dia do experimento. Para isso, os animais foram
deixados em jejum por 12 horas e, posteriormente, receberam 2,5g de glicose por Kg de
peso corporal por gavagem. Para tal foi utilizada uma solução de glicose 25% ajustando
o volume injetado para o peso do animal. A glicose plasmática foi medida antes (tempo
0) e após 30, 60 e 120 minutos, utilizando glicosímetro digital (Accu- Chek Active® da
Roche).
4.8.Análises Bioquímicas no Soro e Plasma
As dosagens bioquímicas - colesterol total, triacilglicerol, uréia, creatinina,
alanina aminotransferase (ALT), aspartato aminotransferase (AST), glicose, proteínas
totais e albumina - foram realizadas utilizando-se kits comerciais Labtest Diagnóstica S.A
de acordo com as instruções do fabricante. Segue, abaixo, os nomes dos kits com seus
respectivos números de referência:
Colesterol Liquiform – Ref. 76 Glicose Liquiform - Ref. 133
Triglicérides Liquiform - Ref. 87 Proteínas Torais - Ref. 99
Uréia CE - Ref. 27 Albumina - Ref. 19
Creatinina - Ref. 35
Transaminase Pirúvica - Ref. 53
Transaminase Oxalacética - Ref. 52

29
4.8.1. Atividade de Paraoxonase
Paraoxonase – Atividade Arilesterásica
A determinação da atividade arilesterásica da enzima paraoxonase, usando o
fenilacetato como substrato, tem como base a velocidade de hidrólise do fenilacetato
(ECKERSON et al, 1983), e foi determinada conforme descrito por Beltowski et al
(2002).
Procedimento da dosagem:
Em tubo de ensaio adicionaram-se 2mL de tampão Tris-HCl, 9mM; pH = 8,0
contendo 0,9mmol/L de cloreto de cálcio e 5μL de soro. O conteúdo do tubo foi
homogeneizado em vórtex e incubado em banho-maria durante dois minutos. Após esse
procedimento, adicionaram-se 0,5mL da solução Tris - fenilacetato (1μL de fenilacetato
para cada 1500μL de Tris-HCl 9mM; pH = 8,0). Depois de exatamente 3 minutos, a
absorbância foi determinada a 270nm. O espectrofotômetro foi zerado com o branco que
continha todos os reagentes acima, exceto o soro.
Cálculo:
A definição para atividade enzimática foi feita de acordo com Beltowski et al
(2002), onde 1U de paraoxonase é equivalente à hidrólise de 1mmol de fenilacetato (ε =
1310 L.mol-1.cm-1) por minuto. Assim, a atividade arilesterásica da enzima é:
U/mL = Absorbância
ε ×b ×
1000
Tempo ×Volume do Soro
Onde:
ε = Coeficiente de extinção molar;
b = Caminho óptico, igual a 10 mm;
Tempo = 3 minutos.

30
Paraoxonase – Atividade Paraoxonase
A determinação da atividade da enzima paraoxonase, usando paraoxon como
substrato, tem como base a velocidade de hidrólise deste, com a liberação do
paranitrofenol (ECKERSON et al, 1983), e foi determinada conforme descrito por
Beltowski (2002).
Procedimento da dosagem:
Primeiramente, preparou-se uma solução de paraoxon que contém 9mL de tampão
glicina/NaOH 50mM; pH 10,5 contendo CaCl2 0,9mM e 2μL de paraoxon.
Posteriormente, em tubo de polipropileno adicionaram-se 780μL dessa solução e 20μL
de soro. Homogeneizou-se a solução e a leitura das absorbâncias das amostras foi
efetuada no espectrofotômetro a 412nm, exatamente a cada minuto, por 3 minutos. O
branco (tubo com 780μL da solução preparada inicialmente e 20μL de água) foi utilizado
para calibrar o aparelho.
Cálculo:
Segundo Beltowski et al (2002) 1U da enzima paraoxonase é equivalente à
hidrólise de 1mmol de paraoxon (ε = 18290 L.mol-1.cm-1) por minuto. A atividade
paraoxonase da enzima paraoxonase é:
U/mL = Absorbância
ε ×b ×
1000
Tempo ×Volume do Soro
A absorbância utilizada nessa expressão foi o delta obtido das absorbâncias final e
inicial (absorbância final – absorbância inicial / 2).

31
4.9.Determinação de Marcadores do Estresse Oxidativo em Fragmentos de
Tecido
4.9.1. Proteína Carbonilada
Proteína carbonilada é um marcador da oxidação de proteína por ROS. A oxidação
de proteínas por ROS leva a formação de derivados carbonilados. Estes podem ser
mensurados utilizando o 2,4-dinitrofenilhidrazina (DNPH). O DNPH reage com grupos
carbonilados gerando a hidrazona correspondente, a qual pode ser analisada
espectrofotometricamente. A determinação da concentração de proteína carbonilada foi
realizada conforme descrito por Levine et al. (1994) em fragmentos de tecido hepático,
cardíaco, pancreático e renal.
Procedimento da dosagem:
Inicialmente, um fragmento de tecido hepático, cardíaco, pancreático e renal foi
homogeinizado em 2 mL de tampão fosfato 50 mM (pH 6,7) e em seguida centrifugado
a 10.000g por 15 minutos à 4ºC. 500 µL do sobrenadante foi transferido para dois tubos
de polipropileno identificados como Amostra (A) e Controle (C). A cada tubo foi
adicionado igual volume de ácido tricloroacético (TCA) 10% e após centrifugação, a
5000g por 10 minutos a 4ºC, o sobrenadante foi descartado e, em seguida, foram
adicionados ao tubo A 500 µL de DNPH 10 mM, em HCl 2M, e ao tubo C 500 µL de
HCl 2M. Ambos foram mantidos no escuro à temperatura ambiente por um período de 30
minutos e a cada 15 minutos misturados vigorosamente. No passo seguinte adicionaram-
se 500 µL de TCA 10% em cada tubo. Logo após, os tubos foram centrifugados a 5000g
por 10 minutos a 4ºC e os sobrenadantes descartados. Em seguida, os precipitados em
ambos os tubos foram lavados com 1 mL da mistura etanol/acetato de etila, na proporção
de 1:1, misturando no vórtex e novamente centrifugados conforme descrito na etapa
anterior e o sobrenadante novamente descartado. Este último passo foi repetido por duas
vezes. Ao final do processo de lavagem, adicionou-se em ambos os tubos 1 mL de SDS
6% e, após agitação, centrifugou-se à 10000g por 10 minutos à 4ºC. Finalmente, as
absorbâncias dos sobrenadantes foram determinadas a 370nm. Os resultados foram
expressos em nmol de DNPH incorporado/mg de proteína.

32
Cálculo:
Para determinar a concentração de proteína carbonilada utilizou-se a equação de
Lambert Beer:
A= C x b x ε
Onde:
A = Absorbância da Amostra (tubo A) - Absorbância do Controle (tubo C).
C = Concentração
b=Caminho óptico
ε=Coeficiente de extinção molar:
O conteúdo de proteína carbonilada foi calculado usando o coeficiente de extinção
molar do DNPH (22000 M -1 cm -1) e expresso por nmol de proteína carbonilada formada
por mg de proteína.
4.9.2. TBARS – Substâncias Reativas ao Ácido Tiobarbitúrico
Para se determinar a peroxidação lipídica foi utilizado o ensaio de substâncias
reativas ao ácido tiobarbitúrico (TBA) utilizando o método de Buege & Aust (1978).
Primeiramente, um fragmento de tecido hepático, cardíaco, pancreático e renal foi
homogeneizado com 1 mL de tampão Tris HCl 20 mM (pH 7,4), e em seguida
centrifugado por 10 minutos a 4ºC. O sobrenadante foi retirado e usado como amostra.
Então, 500 µL do sobrenadante do homogenato foram misturados com TCA (28% p/v em
HCl 0,25 M), TBA (1% em ácido acético 0,25M) e BHT (125mM em etanol), aquecido
por 15min a 95ºC e em seguida colocado em um banho de gelo. O precipitado foi
removido por centrifugação a 13.000g por 10 minutos a 4ºC, e a absorbância do
sobrenadante determinada a 535nm. Utilizou-se água destilada como branco.

33
Cálculo:
Para determinar a concentração de TBARS utilizou-se a equação de Lambert
Beer:
A= C x b x ε
Onde:
A = Absorbância da amostra
C = Concentração
b=Caminho óptico
ε=Coeficiente de extinção molar
Os níveis de TBARS foram calculados utilizando o coeficiente de extinção molar
do MDA (154000 M-1cm-1).
4.10. Proteínas Totais
A concentração de proteínas totais foi determinada de acordo com o método de
Lowry et al (1951). O princípio do método baseia-se na redução do reagente de Folin
Ciocalteau, ao reagir com aminoácidos aromáticos, catalisada por íons cobre, em meio
alcalino, formando uma coloração azul.
Inicialmente foram preparadas as soluções de trabalho conforme descrito abaixo:
Reagente A: foram dissolvidos 0,25g de sulfato de cobre e 0,5 de citrato de sódio em 100
mL de água destilada.
Reagente B: foram dissolvidos 5g de carbonato de sódio e 1g de hidróxido de sódio em
250 mL de água destilada.
Reagente C: em 50 mL do reagente B foram adicionados 1 mL do reagente A.
Reagente D: em1 mL de reagente de Folin foram adicionados 1 mL de água destilada.
Para a realização do ensaio, 10 µL do sobrenadante do homogenato de tecido
hepático, cardíaco, pancreático e renal foram diluídos em 90 µL de água destilada.

34
Em tubos de polipropileno foram adicionados 100 µL da amostra diluída ou
soluções padrão de albumina e água destilada (branco). Em seguida foram adicionados 1
mL do reagente C. Os tubos foram agitados em vórtex e incubados a temperatura
ambiente por 15 minutos. Posteriormente, foram adicionados 100 µL do reagente D em
todos os tubos, misturou-se vigorosamente e após 30 minutos de incubação no escuro as
absorbâncias das amostras relativas ao branco foram determinadas a 660 nm. Diluições
seriadas de uma solução de concentração conhecida de albumina bovina sérica foram
utilizadas para a construção da curva de calibração. Após análise de regressão linear, foi
determinada a equação da reta, sendo utilizada para determinar a concentração de
proteínas totais no homogenato dos tecidos.
4.11. Análises Histológicas
Ao final do experimento, o menor lobo do fígado e um fragmento do coração e
pâncreas dos animais foram removidos e fixados em formol tamponado a 4%.
Para efetuar a análise histológica, inicialmente, os tecidos foram cortados
transversalmente e processados em série decrescente de álcoois e, posteriormente, foram
molhados em parafina. Secções parafinadas de aproximadamente quatro micrômetros de
espessura foram obtidos em micrótomo, semi-automático, montados em lâminas de vidro
previamente limpas e desengorduradas. Utilizou-se a técnica de coloração Hematoxilina
& Eosina (H&E) para identificam possíveis alterações teciduais. As fotomicrografias
foram obtidas em microscópio Leica DM5000 acoplado à câmera digital situado no
Laboratório Multiusuários do NUPEB.
4.12. Análise Estatística
Os dados foram submetidos à análise de normalidade pelo teste de Kolmogorov–
Smirnov. Dados com distribuição normal foram submetidos à análise de variância com
post hoc teste de Tukey, ou teste T. Os demais dados foram submetidos ao teste não
paramétrico Kruskal Wallis com post hoc teste de Dunns, ou Mann-Whitney. Utilizou-se
o software GraphPad Prism versão 5.00 para Windows (San Diego, California, USA).
Diferenças foram consideradas significativas quando p 0.05.

35
5. RESULTADOS
5.1.Composição Centesimal e Fitoquímica da Farinha de Buriti
A composição centesimal da farinha de buriti foi determinada e os resultados
encontrados estão descritos na Tabela 8.
Tabela 8: Composição Centesimal em 100g da Farinha de Buriti (Mauritia flexuosa).
Constituintes Composição (g) em 100g Farinha
Umidade 4,60 g
Lipídio 55,66 g
Carboidrato* Níveis insignificantes
Proteína** 4,85 g
Fibra (FDN)*** 38,3 g
Cinzas 2,10 g
As análises foram feitas em triplicata. Os dados são apresentados como a média destas análises.
* Carboidratos = 100- (umidade + proteína + lipídios + cinzas + fibras).
** Fator de conversão do nitrogênio em proteína: 6,25.
*** FDN = fibra detergente neutro.
Quanto ao conteúdo de polifenóis totais a farinha de buriti apresentou 444,28 ±
14,76 mg EAG/100g (mg de equivalentes de ácido gálico).
5.2.Capacidade Antioxidante in vitro contra o Radical DPPH
Para essa análise, utilizaram-se três diferentes concentrações da farinha de buriti
e do antioxidante de referência Trolox, e os resultados foram expressos em percentual de
inibição do radical DPPH. A Figura 14 mostra que a farinha de buriti apresentou alta
capacidade de neutralização do radical DPPH, semelhante ao antioxidante padrão Trolox,
na faixa de 100 a 400 µM após 30 minutos, de maneira dose-dependente.
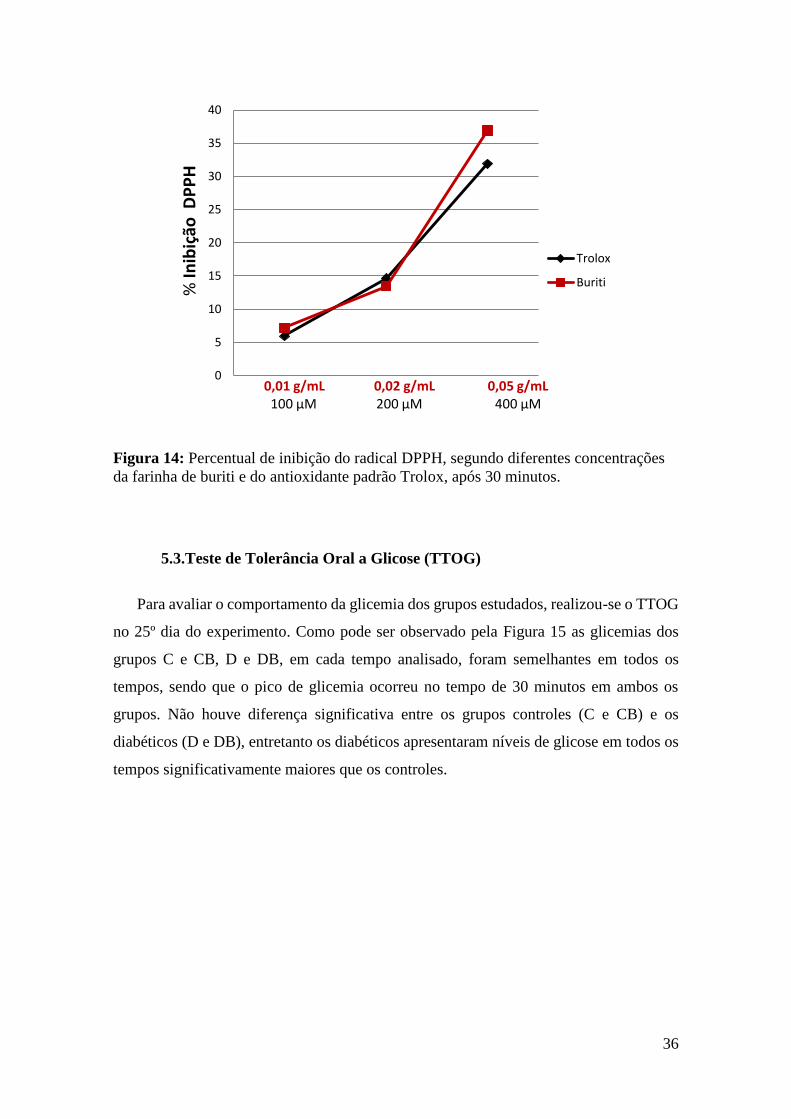
36
Figura 14: Percentual de inibição do radical DPPH, segundo diferentes concentrações
da farinha de buriti e do antioxidante padrão Trolox, após 30 minutos.
5.3.Teste de Tolerância Oral a Glicose (TTOG)
Para avaliar o comportamento da glicemia dos grupos estudados, realizou-se o TTOG
no 25º dia do experimento. Como pode ser observado pela Figura 15 as glicemias dos
grupos C e CB, D e DB, em cada tempo analisado, foram semelhantes em todos os
tempos, sendo que o pico de glicemia ocorreu no tempo de 30 minutos em ambos os
grupos. Não houve diferença significativa entre os grupos controles (C e CB) e os
diabéticos (D e DB), entretanto os diabéticos apresentaram níveis de glicose em todos os
tempos significativamente maiores que os controles.
0
5
10
15
20
25
30
35
40
% In
ibiç
ão D
PP
H
Trolox
Buriti
0,01 g/mL 0,02 g/mL 0,05 g/mL100 µM 200 µM 400 µM
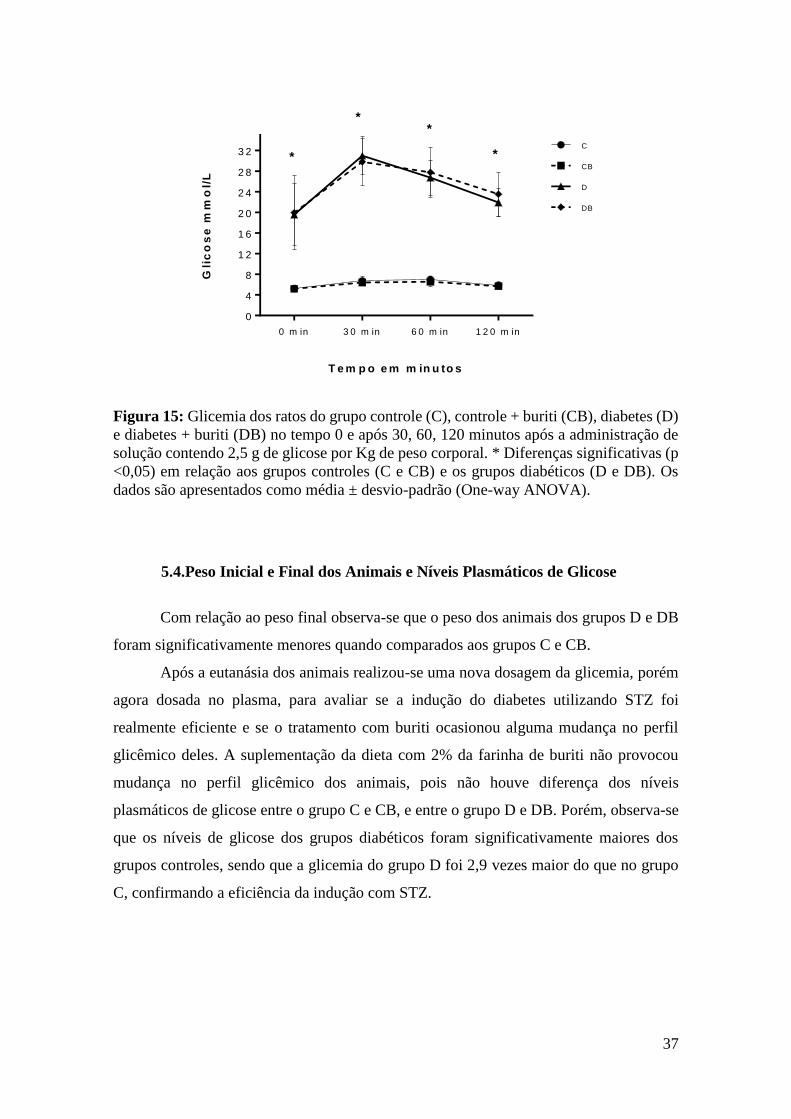
37
T e m p o e m m in u to s
Gli
co
se
mm
ol/
L
0
4
8
1 2
1 6
2 0
2 4
2 8
3 2C
CB
D
DB
0 m in 3 0 m in 6 0 m in 1 2 0 m in
*
**
*
Figura 15: Glicemia dos ratos do grupo controle (C), controle + buriti (CB), diabetes (D)
e diabetes + buriti (DB) no tempo 0 e após 30, 60, 120 minutos após a administração de
solução contendo 2,5 g de glicose por Kg de peso corporal. * Diferenças significativas (p
<0,05) em relação aos grupos controles (C e CB) e os grupos diabéticos (D e DB). Os
dados são apresentados como média ± desvio-padrão (One-way ANOVA).
5.4.Peso Inicial e Final dos Animais e Níveis Plasmáticos de Glicose
Com relação ao peso final observa-se que o peso dos animais dos grupos D e DB
foram significativamente menores quando comparados aos grupos C e CB.
Após a eutanásia dos animais realizou-se uma nova dosagem da glicemia, porém
agora dosada no plasma, para avaliar se a indução do diabetes utilizando STZ foi
realmente eficiente e se o tratamento com buriti ocasionou alguma mudança no perfil
glicêmico deles. A suplementação da dieta com 2% da farinha de buriti não provocou
mudança no perfil glicêmico dos animais, pois não houve diferença dos níveis
plasmáticos de glicose entre o grupo C e CB, e entre o grupo D e DB. Porém, observa-se
que os níveis de glicose dos grupos diabéticos foram significativamente maiores dos
grupos controles, sendo que a glicemia do grupo D foi 2,9 vezes maior do que no grupo
C, confirmando a eficiência da indução com STZ.

38
Tabela 9: Avaliação do peso corporal inicial, peso corporal final e glicemia plasmática
dos ratos do grupo controle (C), controle + buriti (CB), diabetes (D) e diabetes + buriti
(DB).
Variáveis
Grupos Experimentais
Controle Controle + Buriti Diabetes Diabetes + Buriti
Peso Corporal Inicial (g) 192,0 ± 14,91 191,8 ± 13,48 192,5 ± 11,42 192,1 ± 8,66
Peso Corporal Final (g) 214,6 ± 14,86a 216,9 ± 16,38a 154,0 ± 30,63b 160,2 ± 27,56b
Glicose (mmol/L) 7,29 ± 0,58b 7,84 ± 1,47b 21,04 ± 6,10a 22,17 ± 3,51a Os dados são apresentados como média ± desvio padrão (One-way ANOVA). Diferenças significativas
(p<0,05) estão representadas por letras diferentes.
5.5.Peso dos Órgãos e Níveis Plasmáticos de AST, ALT, Uréia e Creatinina
A Tabela 10 mostra o peso relativo do fígado e do rim observado nos animais ao
final do experimento e apresenta os níveis plasmáticos de enzimas e metabólitos séricos
envolvidos no metabolismo dos respectivos órgãos.
Observou-se um aumento no peso relativo tanto do fígado quanto do rim no grupo
D quando comparado aos grupos C e CB. Por outro lado, os animais diabéticos que
receberam dieta contendo 2% de buriti apresentaram uma diminuição significativa no
peso desses órgãos.
Com relação às enzimas hepáticas AST e ALT observou-se um aumento nas
atividades dessas enzimas nos animais dos grupos D e DB, assim como nos níveis de
uréia. Os níveis de creatinina foi estatisticamente igual nos quatro grupos.
Tabela 10: Peso Relativo do Fígado e Rim, Metabólicos e Enzimas Séricas (uréia,
creatinina, AST e ALT) nos grupos C, CB, D e DB.
C CB D DB
Peso Relativo Fígado 3,11 ± 0,32c 3,07 ± 0,27c 4,15 ± 0,38a 3,58 ± 0,13b
AST (U/mL) 25,04 (22,98; 25,04)b 26,46 (23,31; 31,38)b 193,6 (67,57; 250,90)a 232,7 (49,58; 265,70)a
ALT (U/mL) 12,66 (11,95; 17,36)b 14,52 (14,13; 18,23)b 156,3 (69,04; 254,10)a 235,5 (37,36; 266,50)a
Peso Relativo Rim 0,54 ± 0,03c 0,55 ± 0,02c 1,09 ± 0,10a 0,99 ± 0,06b
Uréia (mmol/L) 5,54 ± 0,43b 5,77 ± 0,45b 14,89 ± 2,89a 14,37 ± 4,58a
Creatinina (mmol/L) 71,42 ± 5,83 70,19 ± 5,29 64,87 ± 5,78 71,62 ± 5,07
Os dados são apresentados como média ± desvio padrão (One-way ANOVA) ou apresentados como
mediana e percentis (25%; 75%) (Kruskal Wallis). Diferenças significativas (p<0,05) estão representadas
por letras diferentes.

39
Já com relação ao peso do coração e do pâncreas observou-se uma diminuição
significativa nos ratos diabéticos (Figura 16).
CC
B DD
B
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
aa
b b
Gra
ma
s
CC
B DD
B
0
1
2
3
Gra
ma
s
a
a
bb
Figura 16: Peso do coração (A) e do pâncreas (B) de ratos controle (C e CB) e diabéticos
(D e DB). Os dados são apresentados como média ± desvio-padrão (One-way ANOVA)
ou mediana e percentins 25 e 75 (Kruskal Wallis). Diferenças significativas (p < 0,05)
estão representadas por letras diferentes. C: Controle; CB: Controle + Buriti; D: Diabetes;
DB: Diabetes + Buriti.
5.6.Metabólitos e Enzimas Séricas
A concentração sérica de albumina e proteínas totais estão apresentadas na Figura
17. Com relação a esses parâmetros bioquímicos observou-se, em ambos, uma diminuição
significativa nos grupos D e DB quando comparados com os grupos controles C e CB.
CC
B DD
B
0
2 0 0
4 0 0
6 0 0
8 0 0
a a
b b
Alb
um
ina
(m
mo
l/L
)
CC
B DD
B
5 0
6 0
7 0
8 0
9 0
1 0 0
a
a
bb
Pro
teín
as
To
tais
(g
/L)
Figura 17: Níveis de albumina (A) e proteínas totais (B) de ratos controle (C e CB) e
diabéticos (D e DB). Os dados são apresentados como mediana e percentins 25 e 75
(Kruskal Wallis). Diferenças significativas (p < 0,05) estão representadas por letras
diferentes. C: Controle; CB: Controle + Buriti; D: Diabetes; DB: Diabetes + Buriti.
A B
A
A B
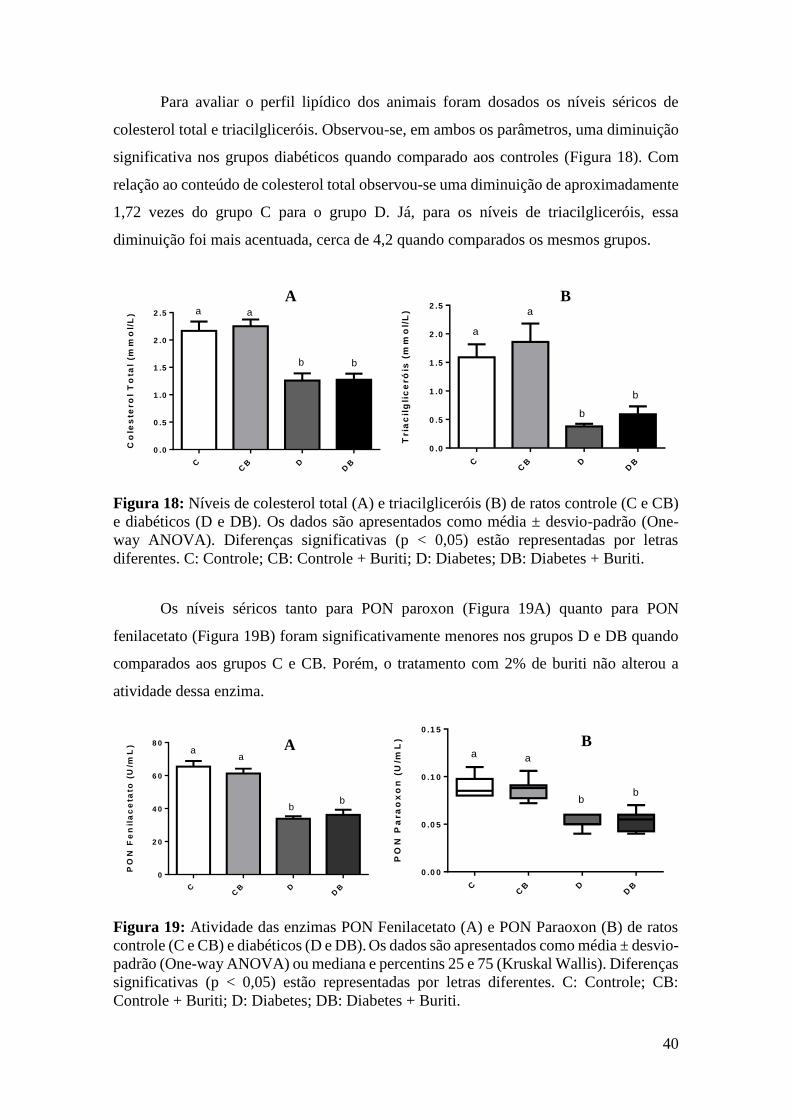
40
Para avaliar o perfil lipídico dos animais foram dosados os níveis séricos de
colesterol total e triacilgliceróis. Observou-se, em ambos os parâmetros, uma diminuição
significativa nos grupos diabéticos quando comparado aos controles (Figura 18). Com
relação ao conteúdo de colesterol total observou-se uma diminuição de aproximadamente
1,72 vezes do grupo C para o grupo D. Já, para os níveis de triacilgliceróis, essa
diminuição foi mais acentuada, cerca de 4,2 quando comparados os mesmos grupos.
CC
B DD
B
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
b b
a a
Co
les
tero
l T
ota
l (m
mo
l/L
)
CC
B DD
B
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
a
a
b
bT
ria
cil
gli
ce
ró
is (
mm
ol/
L)
Figura 18: Níveis de colesterol total (A) e triacilgliceróis (B) de ratos controle (C e CB)
e diabéticos (D e DB). Os dados são apresentados como média ± desvio-padrão (One-
way ANOVA). Diferenças significativas (p < 0,05) estão representadas por letras
diferentes. C: Controle; CB: Controle + Buriti; D: Diabetes; DB: Diabetes + Buriti.
Os níveis séricos tanto para PON paroxon (Figura 19A) quanto para PON
fenilacetato (Figura 19B) foram significativamente menores nos grupos D e DB quando
comparados aos grupos C e CB. Porém, o tratamento com 2% de buriti não alterou a
atividade dessa enzima.
CC
B DD
B
0
2 0
4 0
6 0
8 0a
a
bb
PO
N F
en
ila
ce
tato
(U
/mL
)
CC
B DD
B
0 .0 0
0 .0 5
0 .1 0
0 .1 5
a a
bb
PO
N P
ara
ox
on
(U
/mL
)
Figura 19: Atividade das enzimas PON Fenilacetato (A) e PON Paraoxon (B) de ratos
controle (C e CB) e diabéticos (D e DB). Os dados são apresentados como média ± desvio-
padrão (One-way ANOVA) ou mediana e percentins 25 e 75 (Kruskal Wallis). Diferenças
significativas (p < 0,05) estão representadas por letras diferentes. C: Controle; CB:
Controle + Buriti; D: Diabetes; DB: Diabetes + Buriti.
A B
A B
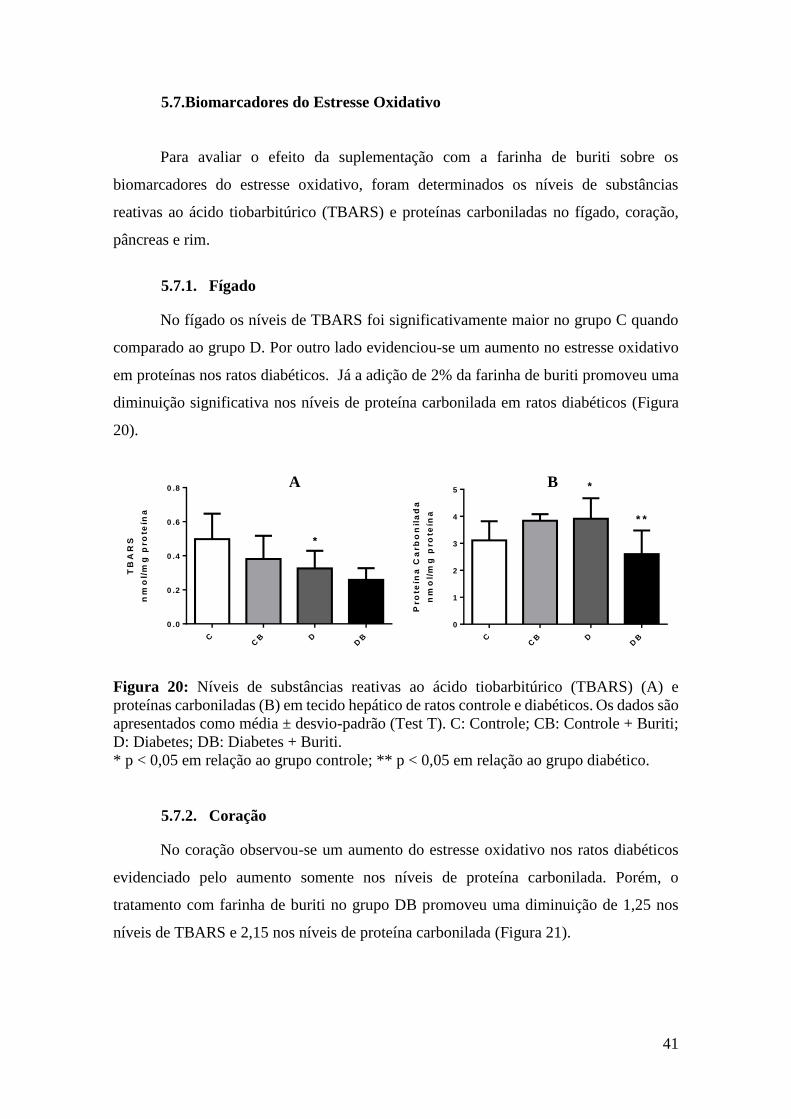
41
5.7.Biomarcadores do Estresse Oxidativo
Para avaliar o efeito da suplementação com a farinha de buriti sobre os
biomarcadores do estresse oxidativo, foram determinados os níveis de substâncias
reativas ao ácido tiobarbitúrico (TBARS) e proteínas carboniladas no fígado, coração,
pâncreas e rim.
5.7.1. Fígado
No fígado os níveis de TBARS foi significativamente maior no grupo C quando
comparado ao grupo D. Por outro lado evidenciou-se um aumento no estresse oxidativo
em proteínas nos ratos diabéticos. Já a adição de 2% da farinha de buriti promoveu uma
diminuição significativa nos níveis de proteína carbonilada em ratos diabéticos (Figura
20).
TB
AR
S
nm
ol/
mg
pro
teín
a
CC
B DD
B
0 .0
0 .2
0 .4
0 .6
0 .8
*
Pro
teín
a C
arb
on
ila
da
nm
ol/
mg
pro
teín
a
CC
B DD
B
0
1
2
3
4
5 *
* *
Figura 20: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) (A) e
proteínas carboniladas (B) em tecido hepático de ratos controle e diabéticos. Os dados são
apresentados como média ± desvio-padrão (Test T). C: Controle; CB: Controle + Buriti;
D: Diabetes; DB: Diabetes + Buriti.
* p < 0,05 em relação ao grupo controle; ** p < 0,05 em relação ao grupo diabético.
5.7.2. Coração
No coração observou-se um aumento do estresse oxidativo nos ratos diabéticos
evidenciado pelo aumento somente nos níveis de proteína carbonilada. Porém, o
tratamento com farinha de buriti no grupo DB promoveu uma diminuição de 1,25 nos
níveis de TBARS e 2,15 nos níveis de proteína carbonilada (Figura 21).
A B
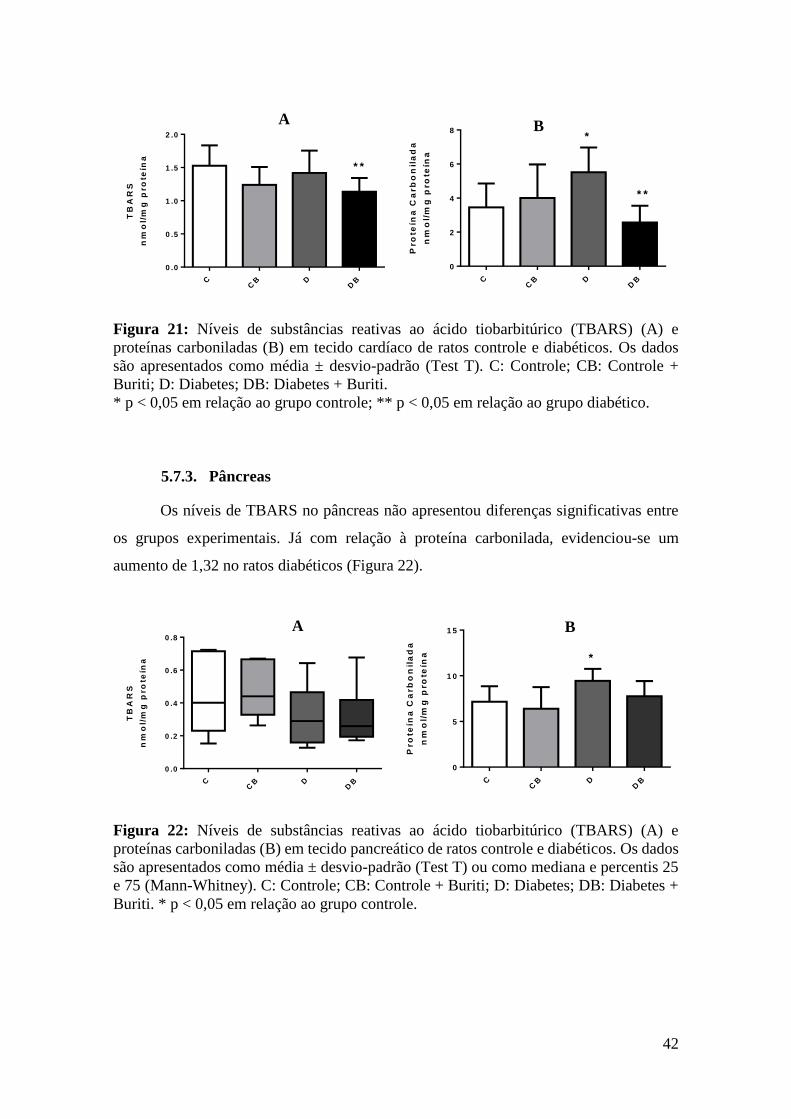
42
TB
AR
S
nm
ol/
mg
pro
teín
a
CC
B DD
B
0 .0
0 .5
1 .0
1 .5
2 .0
* *
Pro
teín
a C
arb
on
ila
da
nm
ol/
mg
pro
teín
a
CC
B DD
B
0
2
4
6
8*
* *
Figura 21: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) (A) e
proteínas carboniladas (B) em tecido cardíaco de ratos controle e diabéticos. Os dados
são apresentados como média ± desvio-padrão (Test T). C: Controle; CB: Controle +
Buriti; D: Diabetes; DB: Diabetes + Buriti.
* p < 0,05 em relação ao grupo controle; ** p < 0,05 em relação ao grupo diabético.
5.7.3. Pâncreas
Os níveis de TBARS no pâncreas não apresentou diferenças significativas entre
os grupos experimentais. Já com relação à proteína carbonilada, evidenciou-se um
aumento de 1,32 no ratos diabéticos (Figura 22).
TB
AR
S
nm
ol/
mg
pro
teín
a
CC
B DD
B
0 .0
0 .2
0 .4
0 .6
0 .8
Pro
teín
a C
arb
on
ila
da
nm
ol/
mg
pro
teín
a
CC
B DD
B
0
5
1 0
1 5
*
Figura 22: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) (A) e
proteínas carboniladas (B) em tecido pancreático de ratos controle e diabéticos. Os dados
são apresentados como média ± desvio-padrão (Test T) ou como mediana e percentis 25
e 75 (Mann-Whitney). C: Controle; CB: Controle + Buriti; D: Diabetes; DB: Diabetes +
Buriti. * p < 0,05 em relação ao grupo controle.
A B
A B
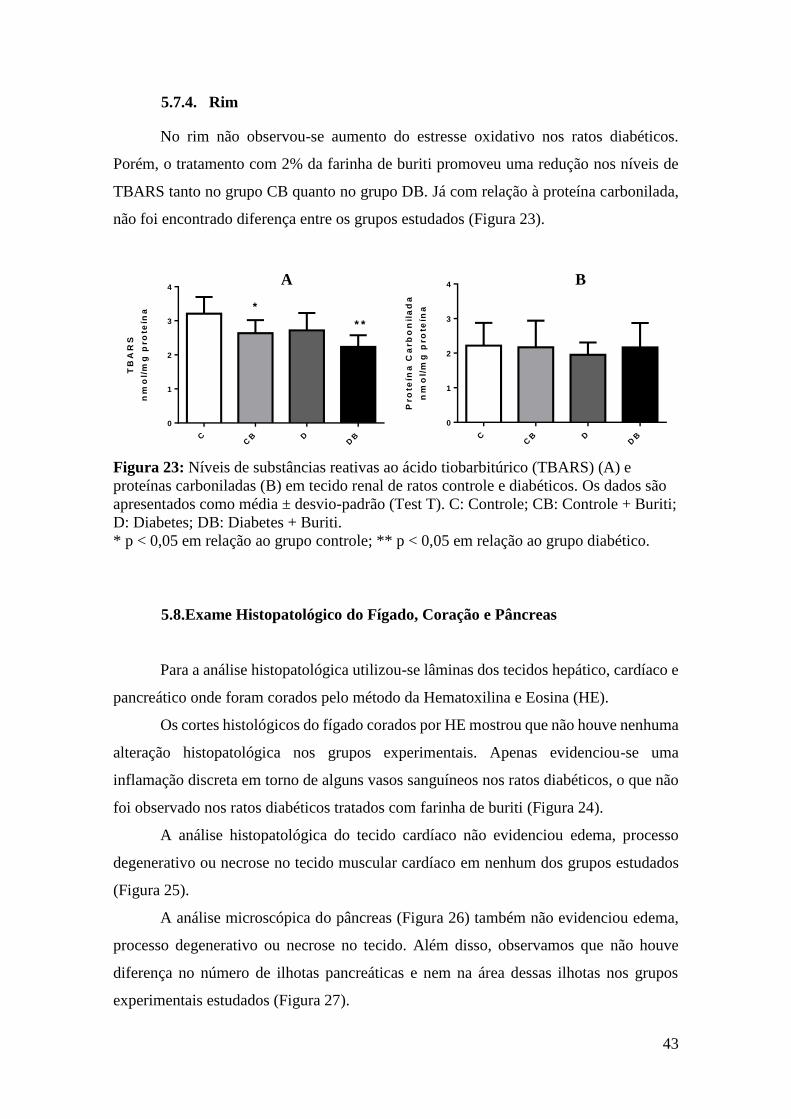
43
5.7.4. Rim
No rim não observou-se aumento do estresse oxidativo nos ratos diabéticos.
Porém, o tratamento com 2% da farinha de buriti promoveu uma redução nos níveis de
TBARS tanto no grupo CB quanto no grupo DB. Já com relação à proteína carbonilada,
não foi encontrado diferença entre os grupos estudados (Figura 23).
TB
AR
S
nm
ol/
mg
pro
teín
a
CC
B DD
B
0
1
2
3
4
*
* *
Pro
teín
a C
arb
on
ila
da
nm
ol/
mg
pro
teín
a
CC
B DD
B
0
1
2
3
4
Figura 23: Níveis de substâncias reativas ao ácido tiobarbitúrico (TBARS) (A) e
proteínas carboniladas (B) em tecido renal de ratos controle e diabéticos. Os dados são
apresentados como média ± desvio-padrão (Test T). C: Controle; CB: Controle + Buriti;
D: Diabetes; DB: Diabetes + Buriti.
* p < 0,05 em relação ao grupo controle; ** p < 0,05 em relação ao grupo diabético.
5.8.Exame Histopatológico do Fígado, Coração e Pâncreas
Para a análise histopatológica utilizou-se lâminas dos tecidos hepático, cardíaco e
pancreático onde foram corados pelo método da Hematoxilina e Eosina (HE).
Os cortes histológicos do fígado corados por HE mostrou que não houve nenhuma
alteração histopatológica nos grupos experimentais. Apenas evidenciou-se uma
inflamação discreta em torno de alguns vasos sanguíneos nos ratos diabéticos, o que não
foi observado nos ratos diabéticos tratados com farinha de buriti (Figura 24).
A análise histopatológica do tecido cardíaco não evidenciou edema, processo
degenerativo ou necrose no tecido muscular cardíaco em nenhum dos grupos estudados
(Figura 25).
A análise microscópica do pâncreas (Figura 26) também não evidenciou edema,
processo degenerativo ou necrose no tecido. Além disso, observamos que não houve
diferença no número de ilhotas pancreáticas e nem na área dessas ilhotas nos grupos
experimentais estudados (Figura 27).
A B
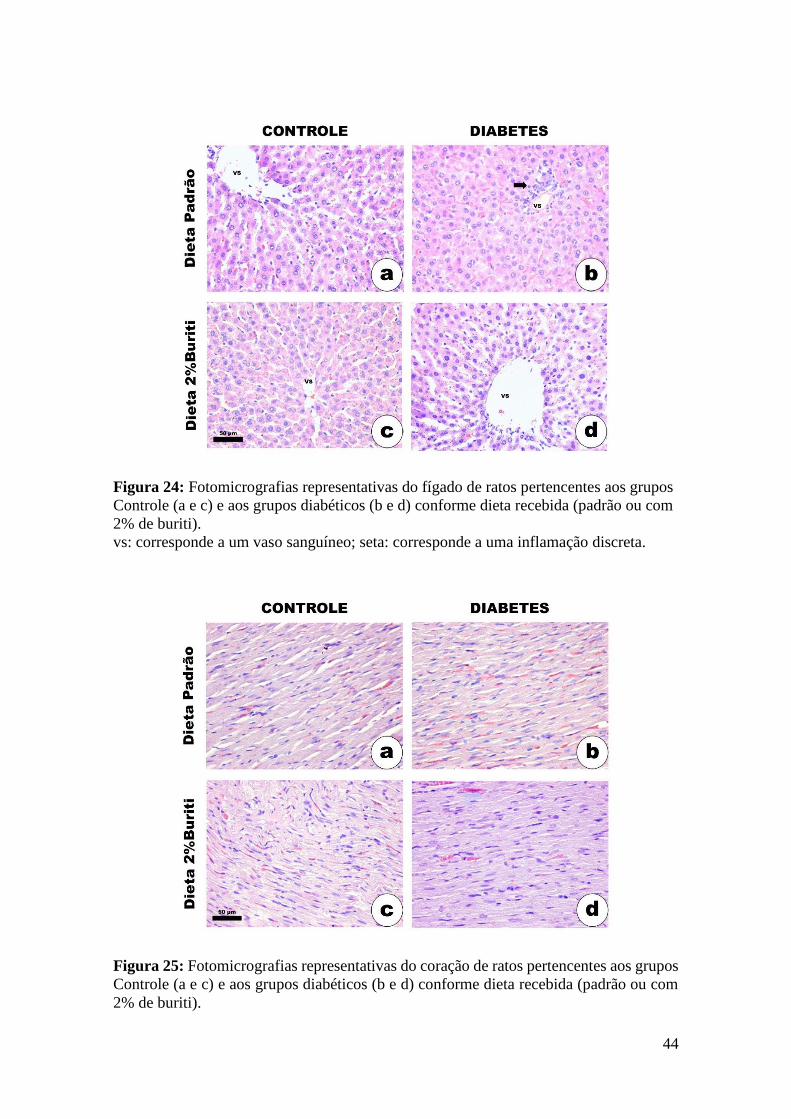
44
Figura 24: Fotomicrografias representativas do fígado de ratos pertencentes aos grupos
Controle (a e c) e aos grupos diabéticos (b e d) conforme dieta recebida (padrão ou com
2% de buriti).
vs: corresponde a um vaso sanguíneo; seta: corresponde a uma inflamação discreta.
Figura 25: Fotomicrografias representativas do coração de ratos pertencentes aos grupos
Controle (a e c) e aos grupos diabéticos (b e d) conforme dieta recebida (padrão ou com
2% de buriti).
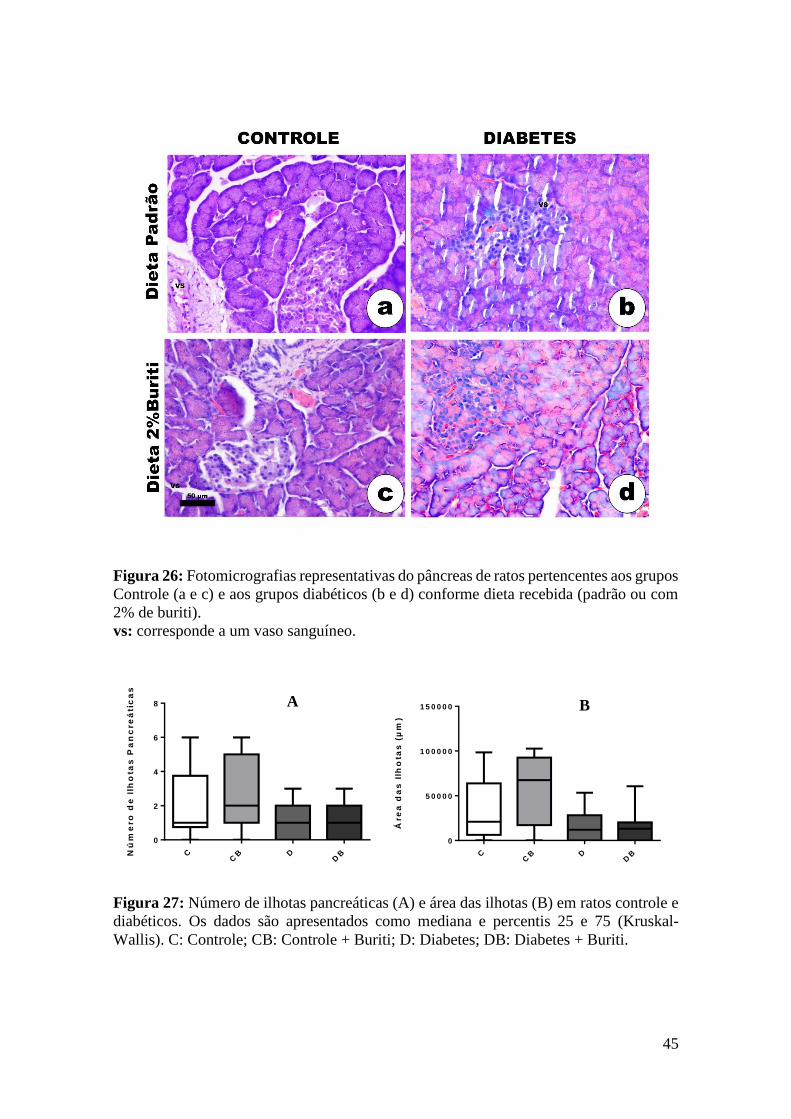
45
Figura 26: Fotomicrografias representativas do pâncreas de ratos pertencentes aos grupos
Controle (a e c) e aos grupos diabéticos (b e d) conforme dieta recebida (padrão ou com
2% de buriti).
vs: corresponde a um vaso sanguíneo.
Nú
me
ro
de
Ilh
ota
s P
an
cre
áti
ca
s
CC
B DD
B
0
2
4
6
8
Áre
a d
as
Ilh
ota
s (
µm
)
CC
B DD
B
0
5 0 0 0 0
1 0 0 0 0 0
1 5 0 0 0 0
Figura 27: Número de ilhotas pancreáticas (A) e área das ilhotas (B) em ratos controle e
diabéticos. Os dados são apresentados como mediana e percentis 25 e 75 (Kruskal-
Wallis). C: Controle; CB: Controle + Buriti; D: Diabetes; DB: Diabetes + Buriti.
A B

46
6. DISCUSSÃO
O diabetes mellitus é uma disfunção metabólica de múltipla etiologia
caracterizada por hiperglicemia crônica resultante da deficiência na secreção de insulina,
ação da insulina ou ambos (TOSCANO, 2004). O aumento da incidência mundial de
diabetes tem sido relacionado às mudanças no estilo de vida e do meio ambiente trazidas
pela industrialização (NARAYAN, 2000). Sabe-se que o DM aumenta a produção de
ROS e que o desenvolvimento das complicações da DM está, entre outros fatores,
intimamente relacionado ao aumento do estresse oxidativo (STADLER et al. 2003).
Desse modo, tem-se um crescente interesse no uso de antioxidantes naturais como
estratégia para reduzir as complicações do diabetes e ampliar a oferta de alimentos da
flora brasileira ricos em antioxidantes e com potencial funcional.
Na literatura há vários estudos com a caracterização físico-química da polpa de
buriti (CARNEIRO & CARNEIRO, 2011; TAVARES et al (2003); MANHÃES, 2007;
MELO et al, 2011). E alguns trabalhos evidenciaram que ela possui quantidades
consideráveis de carotenóides, polifenóis, ácido ascórbico e ácido graxo ômega-9
apresentando, assim, potencial para ser utilizado na prevenção de doenças advindas do
estresse oxidativo, como por exemplo o diabetes (MANHÃES & SABAA-SRUR, 2011;
BARRETO, BENASSI e MERCADANTE, 2009). Porém, ainda não existe estudos
envolvendo o fruto buriti e sua possível ação antioxidante in vivo.
Inicialmente determinamos a composição centesimal da farinha de buriti utilizada
no experimento in vivo. Nela encontramos um maior teor de lipídios e fibras insolúveis
(celulose, hemicelulose e lignina), comparado ao estudo de Melo et al (2011) que
determinou a composição centesimal na polpa fresca. Os valores mais elevados
encontrados desses macronutrientes podem ser decorrentes da perda de umidade
ocasionada pelo processamento térmico que foi necessário para produção da farinha.
Outros estudos também mostraram que a polpa de buriti contém altos teores de lipídios,
principalmente contendo ácidos graxos insaturados (MELO et al, 2011; CARNEIRO &
CARNEIRO, 2011; TAVARES et al; 2003; MANHÃES & SABAA-SRUR, 2011;
MOREIRA & ARÊAS, 1998). Vale lembrar que no conteúdo de lipídios totais incluem
as substâncias solúveis em solvente orgânico, como os carotenóides e vitaminas
lipossolúveis (IAL, 2005).
Várias pesquisas vêm sendo desenvolvidas no intuito de determinar substâncias
bioativas em frutas e hortaliças, de modo a relacionar o consumo in natura com o combate

47
às doenças crônicas. Estudos comprovam os benefícios proporcionados à saúde por
alimentos que contêm carotenóides, vitaminas C e E, compostos fenólicos, ácidos graxos
poli-insaturados e fibras (ANJO, 2004; MORAES e COLLA, 2006).
Na farinha de buriti o conteúdo de polifenóis encontrado foi menor do que os
valores observados na polpa de buriti por Barreto et al (2009) e Canuto et al (2010). Essa
diferença pode ser decorrente do tratamento térmico ao qual a polpa de buriti foi
submetida para a obtenção da farinha. Além dos fatores climáticos e tempo de maturação
do fruto, sabe-se que durante o aquecimento ocorre degradação de antocianinas
pertencente à classe dos polifenóis totais (FALCÃO et al, 2007). Porém, encontramos
valores superiores aos da polpa congelada de açaí (GUERRA et al, 2011). O estudo de
Delfini e Canniatti-Brazaca (2008) constatou que o processo de cocção reduz
consideravelmente o teor de polifenóis no feijão comum.
Os polifenóis constituem um grupo de substâncias largamente distribuídas nos
vegetais e são os antioxidantes mais abundantes da dieta (MANACH et al, 2004). Eles
atuam através de mecanismos de defesa antioxidante não enzimáticos que, em adição aos
efeitos protetores dos antioxidantes endógenos, diminuem o risco do desenvolvimento de
doenças associadas ao acúmulo de radicais livres (POMPELLA, 1997). Dados de outros
autores mostram que o buriti também contém alto teor de carotenóides (MARIATH et al,
1989; YUYAMA et al, 1998; MANHÃES, 2007) micronutriente com atividade
antioxidantede, uma vez que é capaz de sequestrar oxigênio minimizando os danos
causados por radicais livres (FONTANA et al, 1997).
Considerando a presença de polifenóis, carotenóides e ácido ascórbico, utilizamos
o método do DPPH para testar a atividade antioxidante da farinha de buriti. O método se
baseia na capacidade de seus compostos ou conjunto de compostos em “sequestrar” o
radical DPPH. Neste trabalho encontramos que a farinha de buriti possui um alta
capacidade antioxidante, uma vez que mesmo em baixas concentrações foi capaz de
neutralizar o radical DPPH. Um estudo de Ferreira et al (2011) encontrou que óleo de
buriti também possui potencial antioxidante. Em estudo comparativo Barreto et al (2009)
concluiu que a polpa de buriti apresentou o maior nível de carotenóides totais e altos
níveis de ácido ascórbico e polifenóis, porém exibiu atividade antioxidante menor do que
outras frutas tropicais como murici, piquia e tucumã. Os autores sugerem que a baixa
atividade pode ser decorrente da proporção dos compostos bioativos presentes no buriti,
já que o estudo confirmou uma baixa correlação entre carotenóides e a capacidade de
sequestrar radicais livres.

48
Outra questão que tem sido questionada por alguns autores é que o aumento na
ingestão de antioxidantes não necessariamente promoverá uma proteção aos tecidos. Isso
porque a absorção e o transporte de nutrientes são muito bem regulados por processos
homeostáticos e, como tal, o aumento na disponibilidade de um micronutriente não
necessariamente é acompanhado pelo aumento do seu consumo (PUNCHARD &
KELLY, 1996). Portanto, considerando que o β-carotenos possui uma absorção reduzida
nos roedores (RIBAYA-MERCADO et al, 1988; HALLIWELL & GUTTERIDGE,
2007) e que existe uma baixa correlação entre os seus teores e a capacidade de sequestrar
radicais livres, a atividade antioxidante in vitro exibida pela farinha de buriti é,
possivelmente, decorrente do conteúdo de polifenóis. Como já foi dito anteriormente,
ainda não existem, na literatura, estudos envolvendo o buriti e sua possível capacidade
antioxidante in vivo. Portanto avaliamos o potencial do buriti num modelo experimental
que gera estresse oxidativo. Nesse sentido, a indução do diabetes tipo 1 em ratos constitui
um bom modelo experimental para se estudar possíveis efeitos de um alimento com
potencial antioxidante.
Primeiramente, observamos que o modelo experimental proposto foi bem
sucedido, uma vez que o TTOG mostrou que os ratos diabéticos STZ-induzidos
apresentaram uma glicemia significativamente elevada em todos os tempos, o que
também foi evidenciado na glicemia de jejum determinada no plasma dos mesmos
animais. Os nossos resultados corroboram com os estudos de Guerra (2011), Silva et al
(2011), Junior et al (2009), El-Bahr (2013), que também utilizaram STZ para induzir o
DM. Este agente destrói células β pancreáticas através da produção de NO que, por sua
vez, reage com o oxigênio para produzir N2O3 e outros óxidos de nitrogênio, danificando
o DNA e proteínas. Esse processo culmina na inibição da síntese e secreção de insulina
(LENZEN, 2008; DYKE et al, 2010; GUPTA et al, 2004).
Os animais diabéticos apresentaram menor peso corporal ao final do experimento,
o que também foi observado por Guerra et al (2011). Essa perda de peso é característico
do quadro de diabetes e deve-se, principalmente, ao estado catabólico instalado,
decorrente da maior ação dos hormônios contra-regulatórios (NELSON & COX, 2006).
O tratamento com a farinha de buriti não alterou a glicemia e nem o peso final dos animais
diabéticos, sugerindo que a suplementação não foi suficiente para anular ou aliviar os
danos causados às células β pelo STZ.
Ainda assim, demos continuidade ao estudo, pois pacientes diabéticos mesmo
tratados com insulina ou hipoglicemiantes orais apresentam alterações patológicas e

49
complicações que envolvem ação de ROS sobre diversos tecidos tais como, fígado, rim,
retina, sistema cardiovascular e sistema nervoso central e periférico (FAJANS et al.
1997). Nesse estudo observamos um aumento no peso relativo tanto do fígado quanto do
rim nos ratos diabéticos, o que corrobora com os trabalhos de Ayepola et al (2013) e Zou
et al (2014), que também utilizaram o STZ para indução do diabetes em ratos. Isso indica
que o diabetes pode ter ocasionado danos aos órgãos, mas que foram reduzidas
significativamente quando os ratos diabéticos foram tratados com a farinha de buriti.
O fígado desempenha um papel central no metabolismo. Ele sintetiza e distribui
lipídios, glicose, albumina e corpos cetônicos para os outros tecidos via corrente
sanguínea, e converte o excesso de nitrogênio em uréia. As enzimas aminotransferases
AST e ALT estão presentes nos hepatócitos, sendo que qualquer lesão tissular que afete
o parênquima hepático pode provocar um aumento na liberação dessas enzimas para a
corrente sanguínea (HENRY, 1996). Sendo assim, essas enzimas são boas marcadoras de
função hepática e, neste trabalho, verificamos um aumento na atividade de ambas as
enzimas nos ratos diabéticos. Nossos resultados corroboram com os trabalhos de Silva
(2011), Rao et al (2013), Ashraf et al (2013) e Masjedi et al (2013), apesar destes estudos
terem utilizado linhagens direferentes do nosso trabalho e Rao, Ashraf e Masjedi terem
injetado uma dose mais elevada de STZ.
Verificamos, também nos ratos diabéticos, uma diminuição significativa dos
parâmetros bioquímicos de albumina, proteínas totais, triacilgliceróis e colesterol total.
Silva et al (2011) e Ashraf et al (2013) também observaram uma redução no conteúdo de
proteínas totais e Silva (2011) observou redução de albumina e triacilgliceróis. Essas
modificações podem ter ocorrido em virtude da disfunção hepática aparente, ocasionando
um impacto tanto no metabolismo de proteínas como no metabolismo de lipídios. Além
disso, como a insulina estimula a síntese de colesterol e triacilgliceróis no fígado por meio
da ativação das enzimas HMG-CoA redutase e ACC respectivamente, uma redução na
produção desse hormônio poderá levar a uma diminuição nas concentrações sanguíneas
desses metabólitos. No diabates não-controlado também ocorre proteólise, justificando
os níveis diminuidos de proteínas totais e os níveis aumentados de uréia no sangue dos
ratos diabéticos (NELSON & COX, 2006).
Observou-se, também, uma diminuição no peso do pâncreas e do coração nos
grupos diabéticos, fato adicional que justifica o menor peso corporal dos animais destes
grupos. O trabalho de Masjedi et al (2013) também encontrou um peso reduzido para o

50
pâncreas e os trabalhos de Silva (2011) e Han et al (2014) observaram uma diminuição
no peso do coração.
Espécies reativas de oxigênio e espécies reativas de nitrogênio são produzidos
como um resultado fisiológico normal de processos metabólicos e desempenham um
papel importante na sinalização celular, na transcrição gênica e na resposta imune. Porém,
uma produção excessiva ou o acúmulo de espécies reativas pode ter efeito prejudicial,
uma vez que participam de reações de oxi-redução causando danos a macromoléculas,
membranas celulares, DNA e vias de sinalização. Por essa razão, o organismo
desenvolveu um complexo sistema de defesa de modo a evitar danos oxidativos (VALKO
et al, 2007; MCCORD, 2000). A enzima paraoxonase (PON) possui capacidade
antioxidante e circula no plasma associada com a HDL protegendo lipoproteínas da ação
dos radicais livres, principalmente contra a oxidação da LDL (MACKNESS et al, 1997;
FERRÉ et al). Sendo assim, uma redução dos seus níveis sanguíneos pode favorecer
oxidação de LDL e, em consequência, o desenvolvimento de aterosclerose, um quadro
frequente em pacientes diabéticos. Existem dois métodos distintos de se dosar a atividade
da PON. Em nosso trabalho encontramos uma redução significativa na atividade da PON
nos ratos diabéticos, em ambas as dosagens, corroborando com Ghaffari et al (2011) e
Aslan et al (2013). Porém, o tratamento com 2% da farinha de buriti não foi não capaz de
reverter essa situação.
O estresse oxidativo pode ser determinado pela ação de espécies reativas e
biomoléculas e o presente estudo dosou dois importantes biomarcadores no fígado,
coração, pâncreas e rim: TBARS e proteína carbonilada. O primeiro método mensura a
peroxidação lipídica que se baseia na reação do ácido tiobarbitúrico (TBA) com
malondialdeído (MDA), produto da peroxidação lipídica, gerando um produto rosado, o
(TBA)2 – MDA, que pode ser determinado por absorção a 532. Essa determinação é
importante, pois níveis aumentados de certos produtos finais da peroxidação lipídica estão
elevados em muitas doenças humanas, inclusive no diabetes. Assim como acontece com
os lipídios, as proteínas também podem sofrer ataque direto dos radicais livres. Um
método muito utilizado para determinar danos oxidativos em proteínas é o ensaio de
proteínas carboniladas. A oxidação de resíduos amino-ácidos em proteínas, por radicais
livres (incluindo OH•, RO•, ONOO-, HOCC), forma grupos carbonilados que podem
reagir com DNPH gerando um produto final com coloração detectável
espectrofotometricamente (HALLIWELL & GUTTERIDGE, 2007). Estudos

51
comprovam alterações sobre biomarcadores do estresse oxidativo em ratos diabéticos
STZ-induzidos (SILVA et al, 2011; GUERRA, 2011; SAMPAIO, 2012).
O estresse oxidativo apresenta papel específico na patogênese da fibrose e nas
doenças hepáticas. Estudos demonstram um aumento na lipoperoxidação em animais
diabéticos induzidos por estreptozotocina decorrente do aumento no malondealdeído
nesse órgão (GOLDIN et al, 1997; TSUKAMOTO et al, 1993; OHKUWA et al, 1995).
Neste estudo notou-se uma redução nos níveis de TBARS no tecido hepático dos ratos
diabéticos. Esse resultado não corrobora com outros que utilizaram o mesmo modelo
experimental (GUERRA et al, 2011; SILVA, 2011). Embora permaneça a ser esclarecido
nível reduzido de TBARS nos ratos diabéticos pode ter sido decorrente da falta de insulina
provocada pelo modelo, resultando em mudanças no metabolismo de glicose e lipídios.
A falta de glicose nas células promove um aumento no catabolismo de lipídios para
fornecimento de energia. Sendo assim, os ratos diabéticos podem apresentar uma menor
quantidade de lipídios no organismo, diminuindo a possibilidade de peroxidação lipídica.
Com relação à dosagem de proteínas carboniladas encontramos níveis elevados no grupo
diabético, assim como foi verificado por Guerra et al (2011) e Silva (2011), sendo que a
farinha de buriti conseguiu reduzir esses valores. A contradição entre resultados para
TBARS e proteína carbonilada pode ser devido as diferentes velocidades de metabolismo
entre os dois. Produtos da peroxidação lipídica, uma vez formados e livres, são
metabolizados rapidamente. Proteínas oxidadas são degradadas, principalmente pelo
sistema de proteassoma, e sua metabolização parece acontecer mais lentamente
(HALLIWELL & GUTTERIDGE, 2007).
Relatórios do Framingham e de outros estudos têm demonstrado que DM constitui
um forte fator de risco de morbidade e mortalidade cardiovascular (KANNEL & MCGEE,
1979). O estudo de Shao et al (2011) encontrou correlação entre carbonilação de proteínas
e disfunção cardíaca devido à perda da atividade das proteínas que participam da
contração e ou relaxamento do músculo cardíaco. Nossos resultados mostraram que
houve um aumento do estresse oxidativo no tecido cardíaco dos ratos diabéticos, fato
revelado somente pela dosagem de proteína carbonilada. Em contra partida, a farinha de
buriti foi capaz de reduzir tanto os níveis de TBARS quanto os de proteínas carboniladas
nos ratos diabéticos, sugerindo que o buriti teve efeito antioxidante neste órgão para
ambos os parâmetros analisados. Um estudo de Sampaio (2012) que avaliou os efeitos da
suplementação com ferro injetável sobre danos oxidativos em tecidos cardíaco e
pancreático de ratos diabéticos STZ-induzidos, não observou diferença entre os grupos

52
experimentais para a dosagem de TBARS e não encontrou diferença entre o grupo
diabético e o controle para o parâmetro proteínas carboniladas.
O estresse oxidativo induzido por hiperglicemia pode também causar dano celular
ao pâncreas. O estudo de Gutierrez (2012) observou que a lesão associada ao diabetes e
radicais livres promoveu um acúmulo de produtos de peroxidação de lipídios, depleção
de GSH, redução da razão GSH/GPx e sub-regulação de enzimas antioxidantes
importantes não apenas no tecido hepático, mas também no pâncreas. Os resultados do
nosso estudo revelaram que não houve diferença significativa entre os grupos nos níveis
de TBARS no tecido pancreático. Porém, observou-se um aumento nos níveis de
proteínas carboniladas nos ratos diabéticos, sugerindo um possível aumento do estresse
oxidativo neste órgão. O estudo de Sampaio (2012) também avaliou danos oxidativos às
biomoléculas no tecido pancreático e encontrou um aumento nos níveis tanto de TBARS
quanto de proteínas carboniladas nos ratos diabéticos.
A nefropatia diabética é uma dentre as muitas complicações microvasculares no
diabetes, sendo a principal causa do estágio final da disfunção renal. O aumento na
geração de ROS e a diminuição da atividade de enzimas antioxidantes resulta em estresse
oxidativo nos rins de ratos diabéticos STZ-induzidos (LING et al, 2013). Encontramos,
no rim, diferença somente na dosagem de TBARS, onde os ratos tratados com farinha de
buriti apresentaram uma menor peroxidação lipídica em relação aos seus respectivos
grupos controles. Esses resultados mostram que não observamos um aumento do estresse
oxidativo nesse órgão, mas o buriti reduziu a peroxidação lipídica nos grupos tratados. O
trabalho de Schmatz et al (2012), que investigou os efeitos do resveratrol sobre
parâmetros do estresse oxidativo no fígado e rins em ratos diabéticos STZ-induzidos,
encontrou um amento nos níveis de TBARS em todos os grupos diabéticos estudados e o
tratamento com resveratrol foi capaz de reduzir esses níveis.
Após a avaliação do efeito do diabetes e da farinha de buriti sobre biomarcadores
do estresse oxidativo, realizamos exame histopatológico do fígado, coração e pâncreas
com o intuito de analisar, microscopicamente, se houve alguma alteração tecidual nesses
órgãos. A análise histopatológica não evidenciou alterações nesses tecidos, mostrando
que, apesar do estresse oxidativo ter sido confirmado nos ratos diabéticos, não houve
danos ao nível tecidual. O estudo de Silva (2011) encontrou, no fígado, indicativo
morfológico de esteatose microvesicular em hamsters diabéticos. Um estudo de Silva et
al (2011), que avaliou o efeito da STZ sobre o estresse oxidativo em hamsters, observou
uma diminuição significativa na área ocupada pelas ilhotas pancreáticas nos hamsters

53
diabéticos. Já o estudo de Sampaio (2012) evidenciou um aumento do número de células
e focos inflamatórios nos cortes histológicos do coração dos ratos diabéticos, porém, não
encontrou diferença entre o grupo controle e o grupo diabético com relação ao número de
ilhotas pancreáticas. Essas diferenças podem ser explicadas pelos diferentes modelos
utilizados por cada estudo. Silva et al (2011) usou hamsters e Sampaio (2012) considerou
diabéticos ratos Fishers machos com glicemia acima de 320 mg/dL. Enquanto no presente
estudo usamos ratos Fisher fêmeas com glicemia a partir de 250 mg/dL. Essas
divergências podem ter contribuído para as contradições histológicas observadas. Além
disso, Uzun et al (2013) demonstrou em seu trabalho que a imperfeita homeostase redox
é substancialmente mais proeminente nos machos do que nas fêmeas e que as diferenças
observadas entre os gêneros podem ser atribuídas aos processos de envelhecimento,
estado hormonal relacionada ao gênero e mecanismos alterados de homeostase celular.
Portanto, em conjunto, nossos resultados indicam que, apesar da farinha de buriti
não ter promovido mudanças significativas no perfil glicêmico e nos parâmetros
bioquímicos dosados no soro, promoveu redução nos níveis de TBARS e/ou proteínas
carboniladas no fígado, coração e rim, indicando efeito antioxidante in vivo.

54
7. CONCLUSÃO
Neste estudo verificamos que a farinha de buriti possui grande quantidade de
lipídios totais. Encontramos, também, que essa farinha contém um conteúdo elevado de
polifenóis totais e apresentou uma alta capacidade antioxidante in vitro.
In vivo observamos que o modelo experimental proposto de diabetes tipo 1 foi
bem sucedido, o que foi comprovado pelo TTOG e glicemia final dos animais. As
dosagens séricas para transaminases e uréia realizadas no soro juntamente com os dados
de peso relativo do fígado sugerem dano nesse órgão nos ratos diabéticos. Para pâncreas
e coração foram evidenciados redução de seus pesos.
Quanto a dosagem dos biomarcadores do estresse oxidativo no fígado, coração,
pâncreas e rim encontramos resultados diferentes em cada órgão. Porém, em conjunto,
eles demonstram que houve um aumento do estresse oxidativo no fígado, coração e
pâncreas, e que a suplementação com 2% da farinha de buriti foi capaz de reduzir os
níveis de TBARS e/ou proteína carbonilada nestes tecidos, exceto pâncreas. Já análise
histopatológica do fígado, coração e pâncreas não identificou nenhuma alteração tecidual,
provavelmente decorrente do baixo tempo de experimento.
Concluímos, então, que a suplementação com farinha de buriti nos ratos diabéticos
promoveu redução de danos oxidativos a biomoléculas no fígado, coração e rim,
evidenciando uma provável atividade antioxidante in vivo dos componentes bioativos
presentes nesse fruto.

55
8. LIMITAÇÕES DO ESTUDO
As limitações do nosso estudo incluem o curto período de tratamento, imposto
pela gravidade do quadro clínico induzido pelo STZ e a baixa absorção de β-caroteno
observado para ratos (RIBAYA-MERCADO et al, 1988). Entretanto, nosso modelo
permitiu verificar que outros componentes do buriti promovem redução de danos
oxidativos, permitindo sugerir que o consumo da polpa de buriti pode trazer benefícios à
saúde humana e outros modelos experimentais poderão contribuir para melhor elucidar
os efeitos desse fruto sobre a redução de danos oxidativos.

56
9. REFERÊNCIAS BIBLIOGRÁFICAS
AEBI, H. Catalase in vitro. Methods Enzymol., v.105, p. 121-6, 1984.
AKBARALY, T.N.; FONTBONNE, A.; FAVIER, A.; BERR, C. Plasma carotenoids
and onset of dysglycemia in an elderly population: results of the epidemiology of
vascular ageing study. Diabetes Care, v.31, p.1355–1359, 2008.
ALBUQUERQUE, S.R.S.; REGIANI, A.M. Estudo do fruto do buriti (Mauritia
flexuosa) para obtenção de óleo e síntese de biodiesel. In: 29a Reunião Anual da
Sociedade Brasileira de Química. Sociedade Brasileira de Química (SBQ). 2006,
ANJO, D.F.C. Alimentos funcionais em angiologia e cirurgia vascular. J Vasc Br, v.3,
n. 2, p.145-154, 2004.
ARAI. Global view on functional foods: Asian perspectives. British Journal of
Nutrition, v.88, p.139-143, 2007.
ASHRAF, H.; HEIDARI, R.; NEJATI, V. and ILKHANIPOOR, M. Effects of Aqueous
Extract of Berberis integerrima Root on Some Physiological Parameters in
Streptozotocin-Induced Diabetic Rats. Iranian Journal of Pharmaceutical Research,
v.12, n.2, p.425-434, 2013.
ASLAN, M.; OZCAN, F. and KUCUKSAYAN, E. Increased Small Dense LDL and
Decreased Paraoxonase Enzyme Activity Reveals Formation of an Atherogenic Risk
in Streptozotocin-Induced Diabetic Guinea Pigs. Journal of Diabetes Research,
v.2013, 8p., 2013.
ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS (AOAC). Official
methods of analysis of the Association of Official Analytical Chemists. 14 ed.
Arlington, 1141p., 1995.

57
AVIRAM, M.; ROSENBLAT, M.; BISGAIER, C.L.; NEWTON, R.S. et al.
Pararoxonase Inhibits High-density Lipoprotein Oxidation and Preserves its
Functions: A Possible Peroxidative Role for Paraoxonase. J Clin Invest, v.101, n.8,
p.1581-1590, 1998.
AYEPOLA, O.R.; CHEGOU, N.N.; BROOKS, N.L.B. and OGUNTIBEJU, O.O.
Kolaviron, a Garcinia biflavonoid complex ameliorates hyperglycemia-mediated
hepatic injury in rats via suppression of inflammatory responses. BMC
Complementary and Alternative Medicine, v.13:363, 2013. Disponível em:
<http://www.biomedcentral.com/1472-6882/13/363>. Acessado em: 30/01/13.
BARBOSA, R.I.; LIMA, A.D.; JR, M.M. Biometria de frutos do buriti (Mauritia
flexuosa L.f. ‐‐‐ Arecaceae): estimativas de produtividade de polpa e óleo vegetal
em uma área de savana em Roraima. In: Instituto Nacional de Pesquisas da Amazônia
– INPA, Coordenação de Pesquisas em Ecologia – CPEC. Ministério da Ciência e
Tecnologia. 2009, Amazônia. Relatório de Pesquisa... Amazônia: INPA, 2009. p.23.
BANNING, A.; DEUBEL, S.; KLUTH, D.; ZHOU, Z.; BRIGELIUS-FLOHE´, R. The
GI-GPx gene is a target for Nrf2. Mol Cell Biol, v.25, p.4914–4922, 2005.
BARBOSA, K.B.F.; COSTA, N. M. B.; ALFENAS, R.C.G.; DE PAULA, S. O.; MINIM,
V. P. R.; Josefina BRESSAN, J. Estresse oxidativo: conceito, implicações e fatores
modulatórios. Rev. Nutr., Campinas, v.23, n.4, p.629-643, 2010.
BARRETO, G.P.M.; BENASSI, M.T.; MERCADANTE, A. Z. Bioactive Compounds
from Several Tropical Fruits and Correlation by Multivariate Analysis to Free
Radical Scavenger Activity. J. Braz. Chem. Soc., v.20, n.10, 1856-1861, 2009.
BATISTA, J.S.; OLINDA, R.G.; MEDEIROS, V.B.; RODRIGUES, C.M.F.;
OLIVEIRA, A.F.; PAIVA, E.S.; FREITAS, C.I.A.; MEDEIROS, A.C. Atividade
antibacteriana e cicatrizante do óleo de buriti Mauritia flexuosa L. Ciência Rural,
v.42, n.1, p.136-141, 2012.

58
BELTOWSKI, J.; WOJCICKA, G.; JAMROZ, A. Differential effect of 3-hydroxy-3-
methylglutaryl coenzyme A reductase inhibitors on plasma paraoxonase 1 activity
in the rat. Polish Journal of Pharmacology, v. 54, n. 6, p. 661-671, 2002.
BIANCHI, M.L.P.; ANTUNES, L.M.G. Radicais livres e os principais antioxidantes
da dieta. Rev Nutr., v.12, n.12, p.123-30, 1999.
BRAND-WILLIAMS, W.; CUVELIER, M. and BERSET, C. Use of a free radical
method to evaluate antioxidant activity. LWT – Food Science and Technology, v.28,
p.25-30, 1995.
BRASIL. Agência Nacional de Vigilância Sanitária. Portaria nº 27, de 13 de janeiro de
1998. Regulamento Técnico referente à Informação Nutricional Complementar.
Diário Oficial da República Federativa do Brazil, Brasília, DF, 16 jan. 1998.
BROWNLEE, M. Biochemistry and molecular cell biology of diabetic complications.
Nature, v.414, p.813–820, 2001.
BUEGE, J. A.; AUST, S. D. Microsomal lipid peroxidation. Methods in Enzymology,
v. 52, p. 302-310, 1978.
CANUTO, G.A.B; XAVIER, A.A.O.; NEVES, L.C.; BENASSI, M.T. Cracterização
físico-química de polpas de frutos da amazônia e sua correlação com a atividade
anti-radical livre. Rev. Bras. Frutic. Jaboticabal - SP, v.32, n.4, p. 1196-1205, 2010.
CARNEIRO, T. B.; CARNEIRO, J. G. M. Frutos e polpa desidratada Buriti (Mauritia
flexuosa L.): aspectos físicos, químicos e tecnológicos. Revista Verde, v.6, n.2, p.105 –
111, 2011.
CARVALHO, J.E.U. de, MÜLLER C.H. Biometria e Rendimento Percentual de Polpa
de Frutas Nativas da Amazônia. In: Embrapa- Comunicado Técnico 139, 2005.

59
COSTA, L.G; GIORDANO, G.; FURLONG, C.E: Pharmacological and dietary
modulators of paraoxonase 1 (PON1) activity and expression: the hunt goes on.
Biochem Pharmacol, v.81, p.337–344, 2011.
COSTA, L.A.; BADAWI, A.; EL-SOHEMY, A. Nutrigenetics and Modulation of
Oxidative Stress. Ann Nutr Metab, v.60 (suppl 3), p.27–36, 2012.
COUTO, M.A.L.; CANNIATTI-BRAZACA, S.G. Quantificação de vitamina C e
capacidade antioxidante de variedades cítricas. Ciênc. Tecnol. Aliment., v.30(Supl.1),
p.15-19, 2010.
CRAVEIRO, A.A.; CRAVEIRO, A.C. Alimentos Funcionais. A Nova Revolução.
Fortaleza/UFC-PADETEC, p.282, 2003.
DARNET, S.H.; SILVA, L.H.M.; RODRIGUES, A.M.C.; LINS, R.T. Nutritional
composition, fatty acid and tocopherol contents of buriti (Mauritia flexuosa) and
patawa (Oenocarpus bataua) fruit pulp from the Amazon region. Cienc. Tecnol.
Aliment., Campinas, v.31, n.2, p.488-491, 2011.
DAROUX, M.; PRÉVOST, G.; MAILLARD-LEFEBVRE, H.; GAXATTE, C.;
D’AGATI, V.D.; SCHMIDT, A.M.; BOULANGER, E. Advanced glycation end-
products: Implications for diabetic and non-diabetic nephropathies. Diabetes
Metab., v.36, n.1, p.1-10, 2010.
DELFINI, R.A. e CANNIATTI-BRAZACA, S.G. Polifenóis e sua interação com a
digestibilidade e tempo de co cção em feijão comum. Alim. Nutr., v.19, n.4, p.401-407,
2008.
DIPLOCK, A.T.; AGGETT, P.J.; ASHWELL, M.; BORNET, F.; FERN, E.B.;
ROBERFROID, M.B. Scientific Concepts of Functional Foods in Europe: Consensus
Document. Br J Nutr. v.81, p.1-27, 1999.

60
DU, X.L.; EDELSTEIN, D.; ROSSETTI, L.; FANTUS, I.G.; GOLDBERG, H.;
ZIYADEH, F. et al. Hyperglycemia-induced mitochondrial superoxide
overproduction activates the hexosamine pathway and induces plasminogen
activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc Natl Acad Sci
USA, v.97, n.22, p.12222-6, 2000.
DURRINGTON, P.N.; MACKNESS, B.; MACKNESS, M.I. Paraoxonase and
Atherosclerosis. Arterioscler Thromb vasc Biol, v.21, n.4, p.473-80, 2001.
DYKE, K.V. et al. Oxidative/nitrosative Stresses trigger type I Diabetes: preventable
in streptozotocin rats and detectable in human disease. Ann. N.Y. Acad. Sci., v.1203,
p.138-45, 2010.
ECKERSON, H.W.; WYTE, C.M. and LA DU, B. The human serum
paraoxonase/arylesterase polymorphism. Am.J.Hum.Genet., v.35, p.1126-38, 1983.
EL-BAHR, S.M. Curcumin regulates gene expression of insulin like growth factor,
B-cell CLL/lymphoma 2 and antioxidant enzymes in streptozotocin induced diabetic
rats. BMC Complementary and Alternative Medicine [online], v.13:368, 2013.
Disponível em: < http://www.biomedcentral.com/1472-6882/13/368>. Acessado em:
30/01/14.
ELSNER, M.; GULDBAKKE, B.; TIEDGE, M.; MUNDAY, R.; LENZEN, S. Relative
importance of transport and alkylation for pancreatic beta-cell toxicity of
streptozotocin. Diabetologia. v.43, n.12, p.1528-33, 2000.
ERKKILÄ, A.T., et al. Dietary associates of serum total, LDL, and HDL cholesterol
and triglycerides in patients with coronary heart disease. Preventive Medicine, v. 28,
p.558-565, 1999.
FAJANS, S.S.; CLOUTIER, M.C.; CROWTHER, R.L. Clinical and etiological
heterogeneity idiopathic diabetes mellitus (Banting Memoral Lecture). Diabetes, v.7,
p.1112–1125, 1997.

61
FALLER, A.L.K.; FIALHO, E. Disponibilidade de polifenóis em frutas e hortaliças
consumidas no Brasil. Rev Saúde Pública, v.43, n.2, p.211-8, 2009.
FALCÃO, A.P.; CHAVES, E.S.; KUSKOSKI, E.M.; FETT, R.; FALCÃO, L.D.;
BORDIGNON-LUIZ1, M.T. Índice de polifenóis, antocianinas totais e atividade
antioxidante de um sistema modelo de geléia de uvas. Ciênc. Tecnol. Aliment.,
Campinas, v.27, n.3, p.637-642, 2007.
FERRÉ, N.; CAMPS, J.; FERNÁNDEZ-BALLART, J.; ARIJA, V.; MURPHY, M.M.;
CERUELO, S.; BIARNÉS, E.; VILELLA, E.; TOUS, M. AND JOVEN, J. Regulation
of Serum Paraoxonase Activity by Genetic, Nutritional, and Lifestyle Factors in the
General Population. Clinical Chemistry, v.49, n.9, p.1491–1497, 2003.
FERREIRA, A.L.A.; MATSUBARA, L.S. Radicais livres: conceitos, doenças
relacionadas, sistema de defesa e estresse oxidativo. RAMB., v.43, n.1, p.61-8, 1997.
FERREIRA, B.S.; ALMEIDA, C.G.; FAZA, L.P.; ALMEIDA, A.; DINIZ, C.G.; SILVA,
V.L.; GRAZUL, R.M. and HYARIC, M.L. Comparative Properties of Amazonian Oils
Obtained by Different Extraction Methods. Molecules, v.16, p.5875-5885, 2011.
FONTANA, J.D.; MENDES, S.V.; PERSIKE, D.S.; PERACETTA, L.F.; PASSOS, M.
Carotenóides. Biotecnologia Ciência & Desenvolvimento, p.40-45, 1997.
GAZEL-FILHO, A.B.G.; LIMA, J.A.S. O Buritizeiro (Mauritia flexuosa) e seu
potencial de utilização. Macapá: EMBRAPA, 2001.
GHAFFARI, T.; NOURI, M.; IRANNEJAD, E. AND RASHIDI, M.R. Effect of Vitamin
E and Selenium Supplement on Paraoxonase-1 Activity, Oxidized Low Density
Lipoprotein and Antioxidant Defense in Diabetic Rats. BioImpacts, v.1, n.2, p.121-
128, 2011.
GOLDIN, E.; ARDITE, E.; ELIZALDE, J.I.; ODRIOZOLA, A.; PANÉS, J.; PIQUÉ,
J.M.; FERNANDEZ-CHECA, J.C. Gastric mucosal damage in experimental diabetes
in rats: role of endogenous glutathione. Gastroenterology, v.112, p.855-63, 1997.

62
GOLDRING, C.; KITTERINGHAM, N.; JENKINS, R.; COPPLE, I.; JEANNIN, J.F. and
PARK, B.K. Plasticity in cell defence: access to and reactivity of critical protein
residues and DNA response elements. The Journal of Experimental Biology v.209,
p.2337-2343, 2006.
GUERRA, J.F.C. Polpa de açaí modula a produção de espécies reativas de oxigênio
por neutrófilos e a expressão gênica de enzimas antioxidantes em tecidos hepáticos
de ratos. Ouro Preto: Universidade Federal de Ouro Preto, 2011. 77p. Dissertação
Mestrado.
GUPTA, S.; KATARIA, M.; GUPTA, P.K.; MURGANANDAN, S. and YASHROY,
R.C. Protective role of extracts of neem seeds in diabetes caused by streptozotocin in
rats. Journal of Ethnopharmacology, v.90, p.185–189, 2004.
GUTIERREZ, R.M.P. Effect of the hexane extract of Piper auritum on insulin release
from β-cell andoxidative stress in streptozotocin-induced diabetic rat. Pharmacogn
Mag., v.8, n.32, p.308–313, 2012.
HALLIWELL, B.; WHITEMAN, M. Measuring reactive species and oxidative
damage in vivo and in cell culture: how should you do it and what do the results
mean? Br J Pharmacol., v.142, n.2, p.231-55, 2004.
HALLIWELL, B. and GUTTERIDGE, J. M. C. Free radicals in biology and medicine.
Fourth Edition, United States by Oxford University Press Inc., New York, p.81/
p.174/p.316-36, 2007.
HAN, J.C.; GOO, S.; BARRETT, J.C.; MELLOR, K.; TABERNER, A.J.; LOISELLE,
D.L. The afterload-dependent peak efficiency of the isolated working rat heart is
unaffected by streptozotocin-induced diabetes. Cardiovascular Diabetology, v.13, n.1,
4p., 2014.

63
HEIDLAND, A.; SEBEKOVA, K.; SCHINZEL, R. Advanced glycation end products
and the progressive course of renal disease. Am J Kidney Dis, v.38, p.100-106, 2001.
HENRY, J.B. Clinical Diagnosis and Management by laboratory Methods. 19ª ed.
Saunders Company, 1996.
INSTITUTO ADOLFO LUTZ - IAL. Normas Analíticas do Instituto Adolf Lutz.
Métodos químicos e físicos para análise de alimentos. 4. ed. Brasília: Ministério da Saúde,
1018p., 2005.
INTERNATIONAL LIFE SCIENCE INSTITUTE (ILSI). Safety assessment and
potential health benefits of food componentes based on selected scientific criteria.
Noth America Technical Committee on Food Components for Health Promotion. Crit.
Rev. Food Sci. Nutr., v.39, p.203-316, 1999.
ISHI, T.; ITOH, K.; TAKAHASHI, S.; SATO, H.; YANAGAWA, T.; KATOH, Y.;
BANNAI, S.; YAMAMOTO, M. Transcription factor Nrf2 coordinately regulates a
group of oxidative stress-inducible genes in macrophages. J Biol Chem, v.275,
p.16023–16029, 2000.
JUNIOR, L.A.F.; KRETZMANN, N.A.; WSKI, M.; DIAS, A.S.; MARRONI, N.A.P.
Estresse oxidativo e alterações estruturais pulmonares no diabetes mellitus
experimental. J Bras Pneumol., v.35, n.8, p.788-791, 2009.
KANNEL, W.B.; MCGEE, D.L. Diabetes and glucose tolerance as risk factors for
cardiovascular disease: the Framingham study. Diabetes Care., v.2, p.120–126, 1979.
KOYA, D. and KING, G.L. Protein Kinase C Activation and the Development of
Diabetic Complications. Diabetes, v.47, n.6, p.859-66.
KURUSHIMA, H., et al. Opposite effects on cholesterol metabolism and their
mechanisms induced by dietary oleic acid and palmitic acid in hamsters. Biochimica
et Biophysica Acta. v.1258, p.251-256, 1995.

64
LENZEN, S.; DRINKGERN, J.; TIEDGE, M. Low Antioxidant Enzyme gene
Expression in Pancreatic Islets Compared with Various other Mouse Tissues. Free.
Radic. Biol. Med., v.20, p.463-66, 1996.
LEVINE, R.L. et al. Carbonyl assays for determination of oxidatively modified
proteins. Methods in Enzymoloy, v. 233, p. 346-357, 1994.
LIN HE, M.S. and FERNANDEZ, M.L. Saturated fat and simple carbohydrates
elevate plasma LDL cholesterol concentrations by specific alterations on hepatic
cholesterol metabolism. Nutrition Research. v.18, n.6, p. 1003-1015, 1998.
LING, C.; JINPING, L.; XIA, L.; RENYONG, Y. Ursolic Acid provides kidney
protection in diabetic rats. Curr Ther Res Clin Exp., v.75, p.59-63, 2013.
LOWRY, O.H.; ROSEBROUGH, N.J.; FARR, A.L. and RANDALL, R.J. Protein
measurement with the Folin phenol reagent. J.Biol.Chem., v.193, p.265-75, 1951.
MACKNESS, M. I; ARROL, S.; MACKNESS, B.; DURRINGTON, P.N. Alloenzymes
of paraoxonase and effectiveness of high-density lipoproteins in protecting low-
density lipoprotein against lipid peroxidation. Lancet, v.349, p.851–852, 1997.
MANACH, C., SCALBERT, A.; MORAND, C.; RÉMÉSY, C.; JIMENEZ, L.
Polyphenols: food sources and bioavailability. Am. J. Clin. Nutr.,v.79, p.727-47, 2004.
MANHÃES, L.R.T.; SABAA-SRUR, A.U.O. Centesimal composition and bioactive
compounds in fruits of buriti collected in Pará. Ciênc. Tecnol. Aliment., Campinas,
v.31, n.4, p.856-863, 2011.
MANHÃES, L.R.T. Caracterização da polpa de buriti (Mauritia flexuosa, Mart.):
um potente alimento funcional. Rio de Janeiro: Universidade Federal Rural do Rio de
Janeiro, 2007. 78p. Dissertação Mestrado.

65
MARIATH, J.G.R.; LIMA, M. C.C.; SANTOS, L.M.P. Vitamin A activity of buriti
(Mauritia vinifera Mart) and its effectiveness in the treatment and prevention of
xerophthalmia. Am J Clin Nutr, v.49, p.849-53, 1989.
MARTIN, F.W. Perenial edible fruits of the topics. United Departament of agriculture.
Kansas City, 1990.
MARTINI, L.A.; CATANIA, A.S.; FERREIRA, S.R.G. Role of vitamins and minerals
in prevention and management of type 2 diabetes mellitus. Nutrition Reviews®, v.68,
n.6, p.341–354, 2010.
MASJEDIA, F.; GOLB, A. and DABIRIC, S. Preventive Effect of Garlic (Allium
sativum L.) on Serum Biochemical Factors and Histopathology of Pancreas and
Liver in Streptozotocin-Induced Diabetic Rats. Iranian Journal of Pharmaceutical
Research, v.12, n.3, p.325-338, 2013.
MCCORD, J.M. The evolution of free radicals and oxidative stress. Am J Med, v.108,
p.652–659, 2000.
MELO, W. S.; PENA, R.S.; RODRIGUES, M.C.; SILVA, L.H.M. Hygroscopic
behavior of buriti (Mauritia flexuosa) fruit. Ciênc. Tecnol. Aliment., Campinas, v.31,
n.4, p.935-940, 2011.
MOI, P.; CHAN, K.; ASUNIS, I.; CAO, A.; KAN, Y.W. Isolation of NF-E2-related
factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds
to the tandem NFE2= AP1 repeat of the beta-globin locus control region. Proc Natl
Acad Sci U S A, v.91, p.9926–9930, 1994.
MONTEIRO, C.A.; MONDINI, L.; COSTA, R.L. Mudanças na composição e
adequação nutricional da dieta familiar nas áreas metropolitanas do Brasil (1988-
1996). Revista de Saúde Pública, v.34, p.251-258, 2000a.

66
MONTEIRO, C.A.; CONDE, W.L.; POPKIN, B.M. Independent effects of income and
education on the risk of obesity in Brazil adult population. Journal of Nutririon,
v.131, p.881-886, 2001.
MORAES, F.P.; COLLA, L.M. Alimentos funcionais e nutracêuticos: definições,
legislação e benefícios à saúde. Revista Eletrônica de Farmácia, v.3, n.2, p.99-112,
2006.
MOREIRA, R.S.R.; ARÊAS, J.A. G. Estudo da composição centesimal, perfil de
ácidos graxos e teor de carotenóides da farinha da polpa de buriti (Mauritia vinifera,
Mart.) seca e prensada. In: XVI Congresso Brasileiro de Ciência e Tecnologia de
Alimentos, Rio de Janeiro, 1998.
NELSON, D.L.; COX, M.M. Integração e Regulação do Metabolismo dos Mamíferos.
Lehninger Princípios de Bioquímica. São Paulo: SARVIER, 4 ed., Cap.22, 552-587,
2006.
NGUYEN, T.; SHERRATT, P.J.; HUANG, H.C.; YANG, C.S.; PICKETT, C.B.
Increased protein stability as a mechanism that enhances Nrf2-mediated
transcriptional activation of the antioxidant response element: degradation of Nrf2
by the 26S proteasome. J Biol Chem, v.278, p.4536–4541, 2003.
OHKUMA, T.; SATO, Y. and NAIO, M. Hydroxyl radical formation in diabetic rats
induced by streptozotocin. Life Sciences, v.56, p.1789-98, 1995.
PENTEADO, M.V.C. Vitaminas: Aspectos nutricionais, bioquímicos, clínicos e
analíticos. Barueri, SP: Manole, p.55-56, 2003.
POMPELLA, A. Biochemistry and histochemistry of oxidant stress and lipid
peroxidation. Rev. Nutr., Campinas, v.12, n.2, p.123-130, 1999.
PUNCHARD, N.A. and KELLY, F. J. Free Radicals: A Practical Approach. Oxford
University Press, p.115-31/159-70, 1996.

67
RAMALHO, V.C.R.; JORGE, N. Antioxidantes utilizados em óleos, gorduras e
alimentos gordurosos. Quim. Nova, v.29, n.4, p.755-760, 2006.
RAO, P.V.; MADHAVI, K.; NAIDU, M.D. and GAN, S.H. Rhinacanthus nasutus
Improves the Levels of Liver Carbohydrate, Protein, Glycogen, and Liver Markers
in Streptozotocin-Induced Diabetic Rats. Evidence-Based Complementary and
Alternative Medicine, v.2013, p.1-7, 2013.
REEVES, P.G.; NIELSEN, F.H. & FAHEY, G.C. AIN-93 purified diets for laboratory
rodents: final report of the American Institute of Nutrition and hoc writing
committee on the reformulation of the AIN-76A rodent diet. J.Nutr., v.123, p.1939-
51, 1993.
RIBAYA-MERCADO, J.D; HOLMGREN, S.C.; FOX, J.G. and RUSSELL, R.M.
Dietary β-Carotene Absorption and Metabolism in Ferrets and Rats. American
Institute of Nutrition., p.665-668, 1988.
ROSSONI-JÚNIOR, J.V.; ARAÚJO, G.R.; PÁDUA, B.C.; CHAVES, M.M.;
PEDROSA, M.L.; SILVA, M.E.; COSTA, D.C. Annatto extract and beta-carotene
modulate the production of reactive oxygen species/nitric oxide in neutrophils from
diabetic rats. J. Clin. Biochem. Nutr., v.50, n.3, p.177–183, 2012.
SAMPAIO, A.F.S. Efeitos da Suplementação com Ferro Injetável a Produção de
ERO por Granulócitos e Danos Oxidativos em Tecidos Cardíaco e Pancreático de
Ratos Diabéticos. Ouro Preto: Universidade Federal de Ouro Preto, 2012. 96p.
Dissertação Mestrado.
SANTOS, C.A.; RIBEIRO, R.C.; SILVA, V.C.; SILVA, N.S.; SILVA, B.A.; SILVA, G.
F.; BARROS, B.C.V. Elaboração de biscoito de farinha de buriti (Mauritia flexuosa
L. f ) com e sem adição de aveia (Avena sativa L.). Revista Brasileira de Tecnologia
Agroindustrial, v.05, n.01, p.262-273, 2011.

68
SCHNEIDER, C.D.; OLIVEIRA, A.R. Radicais livres de oxigênio e exercício:
mecanismos de formação e adaptação ao treinamento físico. RBME., v.10, n.10,
p.308-13, 2004.
SCHMATZ, R.; PERREIRA, L.B.; STEFANELLO, N.; MAZZANTI, C.;
SPANEVELLO, R.; GUTIERRES, J. et al. Effects of resveratrol on biomarkers of
oxidative stress and on the activity of delta aminolevulinic acid dehydratase in liver
and kidney of streptozotocin-induced diabetic rats. Biochimie., v.94, n.2, p.374-83,
2012.
SCOTT, M. D.; LUBIN, B. H.; ZUO, L.; KUYPERS, F. A. Erythrocyte defense against
hydrogen peroxide: preeminent importance of catalase. J Lab Clin Med, v.118, p.7-
16, 1991.
SHAO, C.H. et al. Carbonylation contributes to SERCA2a activity loss and diastolic
dysfunction in a rat model of type 1 diabetes. Diabetes, v.60, n.3, p.947-59, 2011.
SHAMI, N.J.I.E.; MOREIRA, E.A.M. Licopeno como agente antioxidante. Rev Nutr.,
v.17, n.2, p.227-36, 2004.
SIES, H. Strategies of antioxidante defence. Review. European Journal of
Biochemistry, v.215, n.2, p.213-219, 1993.
SIES, H., STAHL, W. Vitamins E and C, β-carotene, and other carotenoids as
antioxidants. American Journal of Clinical Nutrition, v.62, n.6, p.1315-1321, 1995.
SILVA, M. Participação e homeostase do diabetes tipo 1 em modelos animais. Ouro
Preto: Universidade de Ouro Preto, 2011. 128p. Tese de Doutorado.
SILVA, M.; LIMA, W. G. L.; SILVA, M. E.; PEDROSA, M. L. Efeito da
estreptozotocina sobre os perfis glicêmico e lipídico e o estresse oxidativo em
hamsters. Arq Bras Endocrinol Metab., v.55, n.1, p.46-53, 2011.

69
SOCIEDADE BRASILEIRA DE DIABETES. Consenso brasileiro sobre diabetes
2002: diagnóstico e classificação do diabetes melito e tratamento do diabetes melito
do tipo 2. Rio de Janeiro: Diagraphic, p.72, 2003.
STADLER, K.; JENEI, V.; BOLCSHAZY, G.; SOMOGYI, A.; JAKUS, J. Increased
nitric oxide levels as an early sign of premature aging in diabetes. Free Radic Biol
Med, v.35, p.1240–51, 2003.
SUÁREZ-MAHECHA, H.; FRANCISCO, A.; BEIRÃO, L.H.; BLOCK, J.M.; SACCOL,
A.; PARDO-CARRASCO, S. Importância de ácidos graxos poliinsaturados presentes
em peixes de cultivo e de ambiente natural para a nutrição humana. Boletim do
Instituto de Pesca, São Paulo, v.28, n.1, p.101-110, 2002.
SZKUDELSKI, T. The Mechanism of Alloxan and Streptozotocin Action in B Cells
of the Rat Pancreas. Physiol. Res., v.50, p.536-546, 2001.
TAVARES, M.; AUED-PIMENTEL S.; LAMARDO, L.C.A.; CAMPOS, N.C.; JORGE,
L.I.F.; GONZALEZ, E. Composição química e estudo anatômico dos frutos de buriti
do Município de Buritizal, Estado de São Paulo. Revista do Instituto Adolfo Lutz, v.62,
n.3, p. 227-232, 2003.
TOSCANO, C.M. As campanhas nacionais para detecção das doenças crônicas não-
transmissíveis: diabetes e hipertensão arterial. Ciência & Saúde Coletiva, v.9, n.4,
p.885-895, 2004.
TSUKAMOTO, H.; KIM, C.M.; LUO, Z.Z.; HORN, W.; SU, L.C.; BRITTENHAM,
G.M. Role of lipid peroxidation in vivo and in vitro models of liver fibrogenesis.
Gastroenterology, v.104, 1012A, 1993.
VALKO, M.; RHODES, C.J.; MONCOL, J.; IZAKOVIC, M.; MAZUR, M. Free
radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol
Interact, v.160, p.1–40. 2006.

70
VALKO, M.; LEIBFRITZ, D.; MONCOL, J.; CRONIN, M.T.D.; MAZUR, M.;
TELSER, J. Free radicals and antioxidants in normal physiological functions and
human disease. Int J Biochem Cell Biol, v.39, p.44–84, 2007.
VIEIRA, D.E.O.; GOMES, M. Efeitos adversos no tratamento da tuberculose:
experiência em serviço ambulatorial de um hospital-escola na cidade de São Paulo.
J Bras Pneumol.; v.34, n.12, p.1049-1055, 2008.
ZANATTA, C.F.; UGARTONDO, V.; MITJANS, M.; ROCHA-FILHO, P.A.;
VINARDELL, M.P. Low cytotoxicity of creams and lotions formulated with Buriti
oil (Mauritia flexuosa) assessed by the neutral red release test. Food and Chemical
Toxicology, v.46, p.2776–2781, 2008.
ZANATTA, C.F.; UGARTONDO, V.; MITJANS, M.; ROCHA-FILHO, P.A.;
VINARDELL, M.P. Photoprotective potential of emulsions formulated with Buriti
oil (Mauritia flexuosa) against UV irradiation on keratinocytes and fibroblasts cell
lines. Food and Chemical Toxicology, v.48, p.70–75, 2010.
WILD, A.C.; MOINOVA, H.R.; MULCAPHY, R.T. Regulation of gamma-
glutamylcysteine synthetase subunit gene expression by the transcription factor
Nrf2. J Biol Chem, v.274, n.33627– 33436, 1999.
WILD, S.; ROGLIC, G.; GREEN, A.; SICREE, R.; KING, H. Global Prevalence of
Diabetes. Diabetes Care, v.27, p.1047-1053, 2004.
YUYAMA, L.K.O.; YONEKURA, L.; AGUIAR, J.P.L.; SOUSA, R.F.S.
Biodisponibilidade dos carotenoides do Buriti (Mauritia flexuosa L.) em ratos. ACTA
Amazônica, v.28, n.4, p.409-415, 1998.

71
10. ANEXOS
Anexo I: Protocolo CEUA - UFOP





























