Embed Size (px)
Citation preview

Korean Society for Biotechnology and Bioengineering Journal 32(3): 179-186 (2017)http://dx.doi.org/10.7841/ksbbj.2017.32.3.179 ISSN 1225-7117 / eISSN 2288-8268
시판 쌈장에서 분리한 용혈성 Bacillus cereus의 동정 및 특성 조사
김동민1, 박상국2, 오계헌2*
Identification and Characterization of Hemolytic Bacillus cereus
Isolated from Commercial Ssam-jang
Dong-Min Kim1, Sang-Kook Park2, and Kye-Heon Oh2*
Received: 26 May 2017 / Revised: 21 July 2017 / Accepted: 26 July 2017
© 2017 The Korean Society for Biotechnology and Bioengineering
Abstract: This study was undertaken to identify and charac-
terize hemolytic Bacillus cereus isolated from commercial
ssam-jang. The physiological and biochemical properties of
isolate were first examined. Using the BIOLOG system, the
isolate was identified and assigned to B. cereus MH-2. Phylo-
genetic tree of MH-2 was plotted based on 16S rRNA sequ-
ence comparisons. Hemolytic activity was observed around
wells of sheep blood agar plates seeded with MH-2 cultures;
the zone of hemolysis gradually increased with increasing
incubation time of the cultures. Zymographic analysis esti-
mated the molecular weight of the presumed hemolysis-caus-
ing molecule to be about 30 kDa. Survival rates of MH-2 cells
decreased with increasing NaCl concentrations in the media.
The stress shock proteins (e.g., DnaK and GroEL) induced by
NaCl were reduced in proportion to the NaCl concentration
and exposure period to B. cereus MH-2. Analysis of SDS-
PAGE and Western blot revealed that the stress shock pro-
teins, 70-kDa DnaK and 60-kDa GroEL were decreased pro-
portionate to the NaCl concentrations as well as exposure per-
iod in exponentially growing cultures. Scanning electron mic-
roscopy demonstrated the presence of perforations and irregu-
lar rod forms with wrinkled surfaces in cells treated with NaCl.
Keywords: Ssam-jang, Hemolysis, Sodium chloride, Bacillus
cereus MH-2
1. INTRODUCTION
쌈장은 고추장, 된장 등과 함께 우리나라의 고유 전통발효 식
품으로 자리 잡아왔으며, 최근 외식문화의 발달로 쌈장을 비
롯한 다양한 장류의 소비량은 증가추세에 있다. 이에 맞추어
일부 식품회사들에서는 장류연구소를 설립하여 다양한 장
류를 개발하고 여러 가지 측면에서의 연구를 수행하고 있다
[1]. 최근 연구에 따르면 쌈장의 원료인 된장의 기능성으로
항산화, 항암, 혈당강하 및 혈전 용해효과 등 다양한 기능성
이 보고되고 있어 기능성 된장 및 된장을 원료로 하는 쌈장
에 대한 관심이 높아지고 있다 [2]. 그러나 쌈장의 제조공정
에서 유입될 수 있는 Bacillus cereus는 식중독 유발세균으로
알려져 있어 건강상의 문제를 일으킬 수 있다.
B. cereus는 물, 토양, 공기 등의 자연계에 널리 분포하고 있
으며 장류뿐만 아니라 농산물, 곡류 및 곡류가공품, 과실 및
채소류, 즉석조리식품, 식육 및 식육가공품, 유제품, 생식제
품 등에서 많이 분리되고 있다 [3]. B. cereus는 그람양성의 호
기성 세균으로서 내생포자를 형성하는 것으로 알려져 있으
며, 식품가공에서 열처리 후에도 조리된 식품을 실온에 보관
할 경우 내생포자가 발아, 증식하여 독소를 생산하여 식중독
을 유발시키는 것으로 알려져 있다. 또한 B. cereus는 Staphy-
lococcus, Escherichia, Vibrio, Salmonella 등과 함께 식중독 세
1東京大学 生命工學科1Department of Biotechnology, The University of Tokyo, 1-1-1 Yayoi,Bunkyo-ku, Tokyo 113-8657, Japan
2순천향대학교 생명시스템학과2Department of Life Science and Biotechnology, Soonchunhyang Uni-versity, Chung-Nam 31538, KoreaTel: +82-41-530-1353, Fax: +82-41-530-1350e-mail: [email protected]
Research Paper

180 Korean Society for Biotechnology and Bioengineering Journal 32(3): 179-186 (2017)
균으로 보고되고 있다 [4].
B. cereus에 의한 식중독은 구토형과 설사형 식중독으로 구
분되어지며, 구토형 식중독은 용혈독소인 hemolysin BL (Hbl)
에 의해 설사형 식중독은 cereulide에 의해 발생된다고 보고
되었다. 특히 일부 B. cereus 균주가 생성하는 설사형 독소인
Hbl은 binding component B, lytic proteins L1, L2 등으로 구성
되어 있으며, 각각 hblA, hblD, hblC 유전자에 의해 제어되며,
피부 괴사 및 혈관 침투 활성을 가지는 것으로 보고되어, he-
molysin이 용혈성 B. cereus에서 주요 독소로 인식되고 있다 [4].
장류와 젓갈류는 예로부터 보존성을 증진시키기 위하여 고
농도의 식염을 사용하여왔다. 그러나 이들 식품은 염분함량
이 평균 30% 정도로, 염도가 2% 정도인 김치에 비해 매우 높
으며 [5], 현재 WHO에서는 1일 나트륨 섭취량을 2,000 mg 이
하로 권고하고 있으나, 우리나라 국민의 하루 평균 나트륨 섭
취량은 이러한 권고치의 2~3배를 초과하는 것으로 보고되었
다 [6]. 이와 같은 과다한 소금의 섭취는 다양한 만성질환의
발병 가능성을 높이는 위험 요인이 되는 것으로 보고되면서,
최근에는 소비자들이 고염도 식품에 대한 소비를 기피하여
그 수요가 점차 감소하고 있다 [7].
소득수준 향상으로 인한 건강지향적인 식품을 선호하는
소비자들이 증가하고 있어 여러 식품기업들에서는 저염도
의 식품을 개발하는데 관심이 집중되고 있으며, 특히 장류와
젓갈 등의 고염도의 식품에서 식품보존을 위한 저염도의 기
준을 정하는 것이 주목받고 있다. 그러나 제조공정에서 다양
한 식중독 세균이 유입될 수 있으며, 특히 내생포자를 형성
하며 효율적인 멸균법이 잘 알려져 있지 않은 B. cereus는 일
정수준의 염도에서 생존이 가능한 것으로 보고되고 있어 큰
문제가 되고 있다 [8].
본 연구에서는 시판되고 있는 쌈장에서 용혈성 Bacillus
cereus MH-2를 분리하여 형태학적 및 생리화학적 특성을 조
사하였으며, BIOLOG 분석 시스템을 사용하여 분리세균을
동정하였다. 16S rRNA 염기서열 분석을 통해 분리세균의 분
자유전학적 계통수를 작성하였으며, 분리세균의 용혈활성
의 특성을 조사하기 위하여 zymography를 실시하였다. 또한
MH-2의 NaCl에 노출에 따른 세포반응을 조사하기 위하여
생존율 조사와 NaCl에 의해 유도되는 스트레스 충격단백질
인 DnaK와 GroEL의 변화를 SDS-PAGE와 Western blotting
을 통하여 조사하였다. 또한 NaCl에 노출된 세포의 외부형
태를 주사전자현미경으로 관찰하였다.
2. MATERIALS AND METHODS
2.1. 균주의 확보 및 배양조건
시판되고 있는 쌈장에서 농화배양기법을 통하여 용혈성을
가지는 Bacillus cereus를 분리하였다. 시료 5 g을 채취하여
100 mL의 멸균된 생리식염수 (physiological saline)이 담긴 플
라스크에 넣고 혼탁한 후, 상등액 100 L를 5% sheep blood
agar (Difco Co., MO, USA)에 도말하고, 30oC의 배양기에서
24시간 동안 배양하였다. 균주의 생육과정에서 용혈활성을
나타내어 집락주변에 투명환 (clear zone)을 형성하는 균주를
선별하였으며, 선별된 균주를 5% sheep blood agar에서 3회에
걸친 도말평판법을 통한 순수배양으로 최종적으로 세균을
분리하였다. 분리세균을 LB (Luria-Bertani, Difco Co., Sparks,
NV, USA) 액체 배지에 접종하고 진탕배양기(30oC, 160 rpm)
에서 배양을 유지하며 본 실험에 이용하였다.
2.2. 분리세균의 형태학적 관찰 및 생화학적 특성 조사
분리세균을 LB 고체평판배지에 도말하여 단일 집락의 형태
를 관찰하고, 그람염색을 실시하여 위상차현미경을 이용하
여 분리세균의 형태학적 특성을 조사하였다. 또한 분리세균
에 대한 여러 가지 생화학적 특성을 조사하기 위하여 포도당
이용 여부, 녹말과 젤라틴 이용 여부, citrate 이용 여부, 인돌
(indole) 생성 유무, methyl red-voges proskauer (MR-VP) 시험,
catalase와 oxidase 활성 시험, klingler iron agar (KIA) 시험, 리
트머스 우유 (litmus milk) 시험 등을 실시하였다.
2.3. 분리세균의 동정
분리세균의 동정을 위해 GP2 Micro Plate을 사용하여 다양
한 탄소원 이용능력에 근거한 특성을 조사하였다. 순수 배양
된 단일 집락을 5% sheep blood agar에 도말한 후, 30oC에서
24시간 배양하였다. 배양된 균주를 생리식염수에 현탁하여
GP2 Micro Plate에 접종한 후, BIOLOG automated Micro-
Station instrument을 이용하여 각 분리균주의 탄수화물 이용
여부를 조사하여 균주를 동정하였다.
2.4. 16S rRNA 염기서열 분석 및 계통수 작성
분리세균의 유전학적 계통수 (phylogenetic tree)를 작성하고
자, 16S rRNA에 대하여 PCR (polymerase chain reaction)을 실
시하였다. 8F primer (5′-AGAGTTTGATCCTGGCTCAG-3′)와
1541R primer (5′-AAGGAGGTGATCCAGCCGCA-3′)를 사용
하여 16S rRNA 유전자를 증폭하였으며, PCR Premix (Gen-
DEPOT, USA)를 이용하여 PCR을 수행하였다. 증폭된 부분
적인 염기서열을 ABI373 Automated sequencer (Foster City,
CA, USA)를 이용하여 서열을 결정하였으며, BLAST data-
base (NCBI, Bethesda, MD, USA)를 통하여 분석하였다. 분석
된 16S rRNA 염기서열은 Clustal X software (http://www.
clustal.org)를 이용하여 정리하였으며 MEGA4 package (Bio-
design Institute, Tempe, AZ, USA)를 이용하여 계통수를 작성
하였다.
2.5. 분리세균 B. cereus MH-2의 농축 배양 상등액 제조
분리세균 MH-2의 용혈활성이 생장기간 동안 생성되는 독소
에 의해 나타나는 것인지 확인하기 위하여 배양 상등액을 5%
sheep blood agar에 주입하여 투명환 (clear zone)의 생성여부
를 관찰하였다. 먼저 분리세균을 LB 액체 배지에 접종하여
30oC에서 배양하였다. 배양기간 중 용혈활성의 변화를 확인
하기 위하여, 배양기간에 따른 배양 상등액을 15 L씩 주입

용혈성 Bacillus cereus의 동정 및 특성 조사 181
하여 최대 용혈활성을 조사하였다. 48시간 동안 배양된 배양
액을 4,000 rpm에서 30분간 원심분리하여 균체를 제거하고,
상등액만 취하여 공극 0.45 m인 syringe filter로 여과하여
cell debris를 제거한 후 동결 건조하였다. 건조된 시료를 멸균
된 증류수에 녹여 20배로 농축하였다.
2.6. Zymography
분리세균의 용혈환성을 가지는 단백질의 분자량을 측정하기
위하여 변형된 Leber 등 [9]의 방법으로 zymography를 실시
하였다. Zymography를 위한 시료는 20배로 농축된 배양상등
액을 사용하였으며, zymography의 수행조건은 Kleiner 등 [10]
의 조건에 따라 실시하였다. 전기영동 완료 후 gel은 Triton
X-100에 담구어 4oC에서 2시간동안 교반하여 gel에 남아있는
SDS를 제거하였다. SDS를 제거한 gel을 증류수로 3회 세척
한 후, 5% sheep blood agar에 부착하여 37oC에서 24시간 배
양하여 용혈활성을 확인하였다. 용혈활성이 일어나는 부분
의 분자량을 확인하기 위하여 용혈현상이 일어난 위치를 표
준 마커와 비교하여 분자량의 크기를 확인하였다.
2.7. NaCl에 노출된 B. cereus MH-2의 생존율 측정
NaCl (sodium chloride) 노출에 따른 분리세균 MH-2의 생존
율을 알아보기 위하여 분리세균을 LB 액체 배지에 배양하였
다. 대수생장기를 거치면서 파장 600 nm에서 혼탁도가 0.8일
때, 배양액을 10분간 원심분리 (4oC, 13,000 rpm)하여 얻어진
균체 PBS buffer로 3회 세척하고, 다시 동일조건 하에 원심분
리를 실시하여 얻어진 균체를 다양한 농도 (0~10%)의 NaCl
에 노출시켰다. 1시간 간격으로 LB 고체 평판배지에 100 μL
씩 평판 도말하여 30oC에서 24시간 배양한 후, 형성된 집락
을 계수하여 NaCl 농도와 노출시간에 따른 분리세균의 생존
율을 각각 조사하였다.
2.8. 스트레스 충격단백질 (SSPs) 발현 조사
NaCl에 노출된 분리세균 MH-2의 스트레스 충격단백질 (SSPs)
의 발현 변화를 조사하기 위하여 SDS-PAGE와 Western blot-
ting을 통하여 SSPs의 변화를 조사하였다. 분리세균을 다양
한 농도의 NaCl에 노출시킨 후 균체를 회수하고 PBS buffer
로 3회 세척한 후 시료를 준비하였다. 단백질 추출은 기존에
기술된 방법 [11]에 따라 단백질을 추출하였으며, Bradford 방
법 [12]으로 각 시료들을 정량하였다. 추출한 단백질을 Kim
등 [11]의 방법을 따라 SDS-PAGE를 실시하였으며, SSPs (예,
DnaK, GroEL)의 발현을 조사하기 위하여 전기영동한 gel을
이용하여 western blotting을 수행하였다. Western blotting에 사
용한 1차 항체는 anti-DnaK와 anti-GroEL (StressGen Biotech-
nologies Corp.)을 사용하였고, 2차 항체는 anti-mouse IgG
HRP conjugate (Promega)를 사용하였다. 반응 검출을 위하여
Western blot detection system (GenDEPOT, Barker, TX, USA)
을 사용하여 X-ray film (AGFA, Mortsel, Belgium)에 현상한
후, 스트레스 충격단백질의 발현을 비교 분석하였다.
2.9. NaCl 노출에 따른 세포 외부형태 관찰
NaCl에 노출된 분리세균의 외부형태 변화를 주사전자 현미
경을 통하여 관찰하였다. 세균을 LB 액체배지에 접종하여
12시간 동안 배양시킨 후, PBS로 3회 세척하여 균체를 준비
하였다. 준비한 균체를 10%의 NaCl에 4시간 동안 노출시킨
후, 배양액을 원심 분리하여 균체를 회수하고, 여과지에 부착
하여 고정 및 탈수한 후, hexamethyldisilazane (HMDS) 원액
에 10분간 반응시켜 공기 중에서 건조하였다. 건조시킨 mem-
brane filter를 slide glass 위에 부착하여 sputter coater (IB-3,
Giko Engineering Co., Tokyo, Japan)를 사용하여 gold coating
한 후, 주사전자현미경으로 관찰하였다.
3. RESULTS AND DISCUSSION
3.1. 분리세균의 분리, 형태학적 관찰, 특성조사
시판되고 있는 쌈장으로부터 균주를 분리하여 5% sheep
blood agar에서 투명대를 형성하는 용혈성 Bacillus cereus를
분리하여 본 실험에 이용하였다. 분리세균을 LB 고체평판배
지에 도말하여 배양된 집락은 크림색을 나타내었으며, 그람
염색 후 위상차 현미경으로 관찰한 결과, 그람양성의 간균으
로 관찰되었다. 분리세균에 대하여 다양한 생화학적 특성조
사를 실시하였다. Glucose, starch, gelatin 이용여부는 모두 양
성으로, 그리고 indole 형성여부에 있어서 음성으로 각각 나
타났다. Methyl red 시험은 양성이었으며, voges-proskauer 시
험은 음성으로 조사되었다. KIA 배지에서 조사한 disulfhyd-
rase에 의한 H2S 형성과 oxidase 존재여부는 모두 음성으로 나
타내었다. Litmus milk 시험에서는 단백질을 분해여 산성으
로 나타났으며, 펩톤화 (peptonization)는 이루어지지 않았다.
3.2. 분리세균의 동정
분리세균을 동정하기 위하여 BIOLOG 시스템을 이용하였으
며, 그람양성 세균 동정에 이용되는 GP2 Micro Plate에 분리
세균을 접종하여 나타난 다양한 탄소원 이용 여부에 근거하
여 분리세균을 동정하였다. 분리세균은 MicroLog database
software를 이용한 결과분석을 통하여 Bacillus cereus로 동정
되었으며, Bacillus cereus MH-2로 명명되었다 (Table 1).
3.3. 16S rRNA 염기서열 분석 및 계통수 작성
분리세균 MH-2의 유전학적 계통수 작성을 위하여 16S rRNA
에 대하여 PCR을 실시하였다. 1,138 bp의 부분적인 염기서열
을 증폭하였으며, PCR Premix (GenDEPOT, USA)를 이용하
여 PCR을 수행하였다. 변성 (94oC, 30초), 냉각 (60oC, 30초),
신장 (72oC, 45초)의 세 단계를 35회 반복한 후, 72oC에서 15
분간 유지하였다. 증폭된 염기서열데이터는 NCBI의 Blast를
이용하여 유전적 상동성을 비교하였다. 그 결과, Bacillus
cereus와 99%의 유전적 상동성을 보여주었다. 이를 바탕으로
다른 Bacillus cereus 균주들과의 유연관계를 Fig. 1에 제시하
였다.

182 Korean Society for Biotechnology and Bioengineering Journal 32(3): 179-186 (2017)
Table 1. Physiological and biochemical characterization of Bacillus cereus MH-2 using the BIOLOG analysis system
Physiological & biochemical tests
Water α-Methyl-D-galactoside L-Malic acid α-Cyclodextrin β-Methyl-D-galactoside Methyl pyruvate β-Cyclodextrin 3-Methylglucose Mono-methyl succinate Dextrin α-Methyl-D-glucoside Propionic acid Glycogen β-Methyl-D-glucoside Pyruvic acid Inulin α-Methyl-D-mannoside Succinamic acid Mannan Palatinose Succinic acid Tween 40 D-Psicose N-Acetyl-L-glutamic acid Tween 80 D-Raffinose L-Alaninamide N-Acetyl-D-glucosamine L-Rhamnose D-Alanine N-Acetyl-D-mannosamine D-Ribose L-Alanine Amygdalin Salicin L-Alanyl-glycine L-Arabinose Sedoheptulosan L-Asparagine D-Arabitol D-Sorbitol L-Glutamic acid Arbutin Stachyose Glycyl-L-glutamic acid Cellobiose Sucrose L-Pyroglutamic acid D-Fructose D-Tagatose L-Serine L-Fucose D-Trehalose Putrescine D-Galactose Turanose 2,3-Butanediol D-Galacturonic acid Xylitol Glycerol Gentiobiose D-xylose Adenosine D-Gluconic acid Acetic aicd 2-Deoxy adenosine α-D-Glucose α-Hydroxybutyric acid Inosine m-Inositol β-Hydroxybutyric acid Thymidine α-D-Lactose γ-Hydroxybutyric acid Uridine Lactulose p-Hydroxy-phenyl acetic acid Adenosine-5'-monophosphate Maltose α-Ketoglutaric acid Thymidine-5'-monophosphate Maltotriose α-Ketovaleric acid Uridine-5'-monophosphate D-Mannitol Lactamide D-Fructose-6-phosphate D-Mannose D-Lactic acid-methyl ester α-D-Glucose-1-phosphate D-Melezitose L-Lactic acid D-Glucose-6-phosphate D-Melibiose D-Malic acid D-L-α-Glycerol phosphate
+: Positive reaction, : Negative reaction.
Fig. 1. Phylogenetic analysis of B. cereus MH-2 (●) related Bacillus cereus group based on 16S rRNA sequence comparisons. Bar, 0.005
substitutions per nucleotide position.
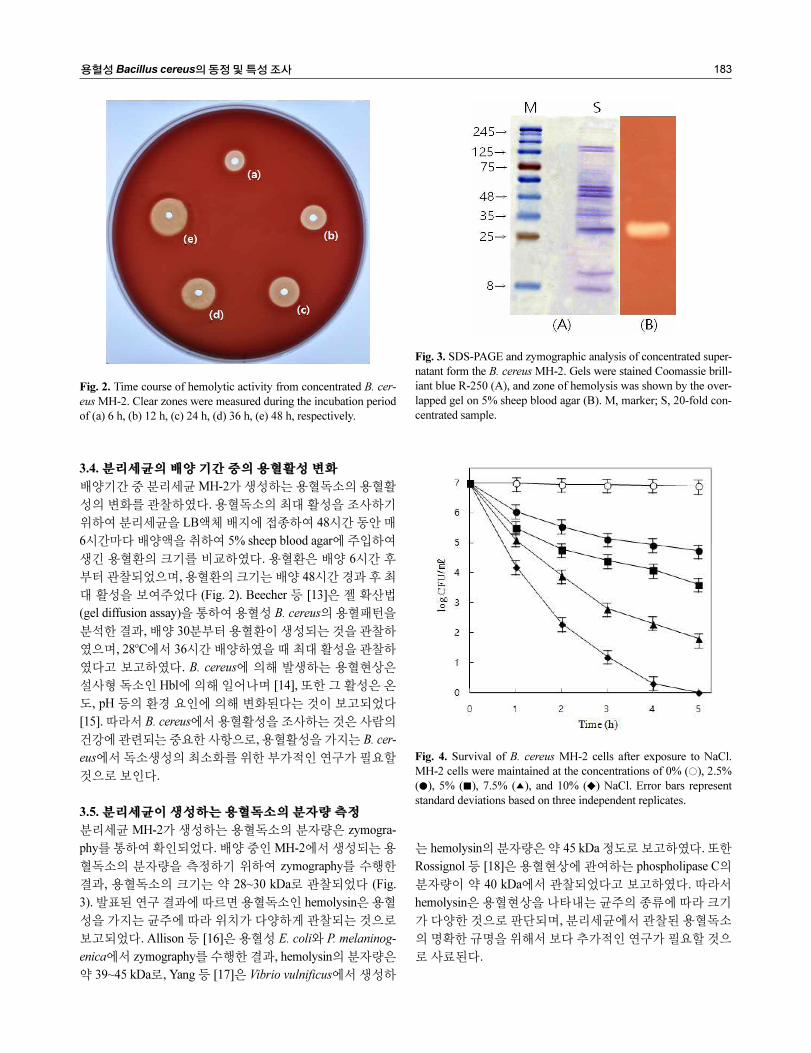
용혈성 Bacillus cereus의 동정 및 특성 조사 183
3.4. 분리세균의 배양 기간 중의 용혈활성 변화
배양기간 중 분리세균 MH-2가 생성하는 용혈독소의 용혈활
성의 변화를 관찰하였다. 용혈독소의 최대 활성을 조사하기
위하여 분리세균을 LB액체 배지에 접종하여 48시간 동안 매
6시간마다 배양액을 취하여 5% sheep blood agar에 주입하여
생긴 용혈환의 크기를 비교하였다. 용혈환은 배양 6시간 후
부터 관찰되었으며, 용혈환의 크기는 배양 48시간 경과 후 최
대 활성을 보여주었다 (Fig. 2). Beecher 등 [13]은 젤 확산법
(gel diffusion assay)을 통하여 용혈성 B. cereus의 용혈패턴을
분석한 결과, 배양 30분부터 용혈환이 생성되는 것을 관찰하
였으며, 28oC에서 36시간 배양하였을 때 최대 활성을 관찰하
였다고 보고하였다. B. cereus에 의해 발생하는 용혈현상은
설사형 독소인 Hbl에 의해 일어나며 [14], 또한 그 활성은 온
도, pH 등의 환경 요인에 의해 변화된다는 것이 보고되었다
[15]. 따라서 B. cereus에서 용혈활성을 조사하는 것은 사람의
건강에 관련되는 중요한 사항으로, 용혈활성을 가지는 B. cer-
eus에서 독소생성의 최소화를 위한 부가적인 연구가 필요할
것으로 보인다.
3.5. 분리세균이 생성하는 용혈독소의 분자량 측정
분리세균 MH-2가 생성하는 용혈독소의 분자량은 zymogra-
phy를 통하여 확인되었다. 배양 중인 MH-2에서 생성되는 용
혈독소의 분자량을 측정하기 위하여 zymography를 수행한
결과, 용혈독소의 크기는 약 28~30 kDa로 관찰되었다 (Fig.
3). 발표된 연구 결과에 따르면 용혈독소인 hemolysin은 용혈
성을 가지는 균주에 따라 위치가 다양하게 관찰되는 것으로
보고되었다. Allison 등 [16]은 용혈성 E. coli와 P. melaninog-
enica에서 zymography를 수행한 결과, hemolysin의 분자량은
약 39~45 kDa로, Yang 등 [17]은 Vibrio vulnificus에서 생성하
는 hemolysin의 분자량은 약 45 kDa 정도로 보고하였다. 또한
Rossignol 등 [18]은 용혈현상에 관여하는 phospholipase C의
분자량이 약 40 kDa에서 관찰되었다고 보고하였다. 따라서
hemolysin은 용혈현상을 나타내는 균주의 종류에 따라 크기
가 다양한 것으로 판단되며, 분리세균에서 관찰된 용혈독소
의 명확한 규명을 위해서 보다 추가적인 연구가 필요할 것으
로 사료된다.
Fig. 2. Time course of hemolytic activity from concentrated B. cer-
eus MH-2. Clear zones were measured during the incubation period
of (a) 6 h, (b) 12 h, (c) 24 h, (d) 36 h, (e) 48 h, respectively.
Fig. 3. SDS-PAGE and zymographic analysis of concentrated super-
natant form the B. cereus MH-2. Gels were stained Coomassie brill-
iant blue R-250 (A), and zone of hemolysis was shown by the over-
lapped gel on 5% sheep blood agar (B). M, marker; S, 20-fold con-
centrated sample.
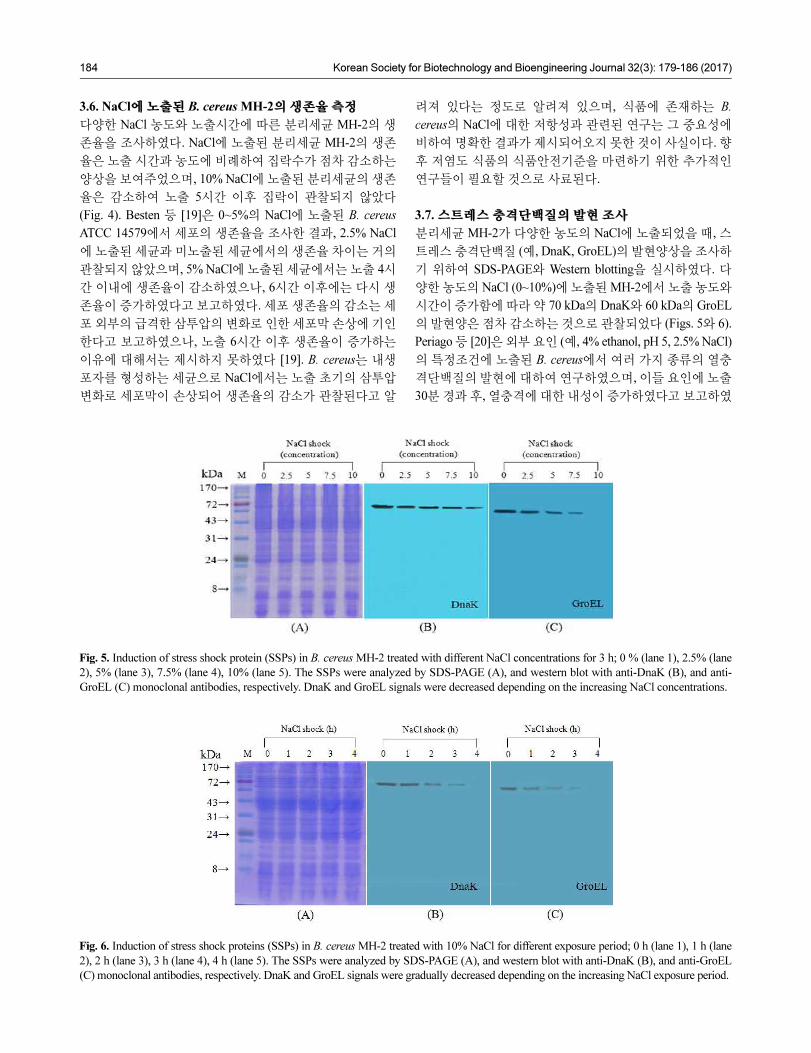
Fig. 4. Survival of B. cereus MH-2 cells after exposure to NaCl.
MH-2 cells were maintained at the concentrations of 0% (○), 2.5%
(●), 5% (■), 7.5% (▲), and 10% (◆) NaCl. Error bars represent
standard deviations based on three independent replicates.
184 Korean Society for Biotechnology and Bioengineering Journal 32(3): 179-186 (2017)
3.6. NaCl에 노출된 B. cereus MH-2의 생존율 측정
다양한 NaCl 농도와 노출시간에 따른 분리세균 MH-2의 생
존율을 조사하였다. NaCl에 노출된 분리세균 MH-2의 생존
율은 노출 시간과 농도에 비례하여 집락수가 점차 감소하는
양상을 보여주었으며, 10% NaCl에 노출된 분리세균의 생존
율은 감소하여 노출 5시간 이후 집락이 관찰되지 않았다
(Fig. 4). Besten 등 [19]은 0~5%의 NaCl에 노출된 B. cereus
ATCC 14579에서 세포의 생존율을 조사한 결과, 2.5% NaCl
에 노출된 세균과 미노출된 세균에서의 생존율 차이는 거의
관찰되지 않았으며, 5% NaCl에 노출된 세균에서는 노출 4시
간 이내에 생존율이 감소하였으나, 6시간 이후에는 다시 생
존율이 증가하였다고 보고하였다. 세포 생존율의 감소는 세
포 외부의 급격한 삼투압의 변화로 인한 세포막 손상에 기인
한다고 보고하였으나, 노출 6시간 이후 생존율이 증가하는
이유에 대해서는 제시하지 못하였다 [19]. B. cereus는 내생
포자를 형성하는 세균으로 NaCl에서는 노출 초기의 삼투압
변화로 세포막이 손상되어 생존율의 감소가 관찰된다고 알
려져 있다는 정도로 알려져 있으며, 식품에 존재하는 B.
cereus의 NaCl에 대한 저항성과 관련된 연구는 그 중요성에
비하여 명확한 결과가 제시되어오지 못한 것이 사실이다. 향
후 저염도 식품의 식품안전기준을 마련하기 위한 추가적인
연구들이 필요할 것으로 사료된다.
3.7. 스트레스 충격단백질의 발현 조사
분리세균 MH-2가 다양한 농도의 NaCl에 노출되었을 때, 스
트레스 충격단백질 (예, DnaK, GroEL)의 발현양상을 조사하
기 위하여 SDS-PAGE와 Western blotting을 실시하였다. 다
양한 농도의 NaCl (0~10%)에 노출된 MH-2에서 노출 농도와
시간이 증가함에 따라 약 70 kDa의 DnaK와 60 kDa의 GroEL
의 발현양은 점차 감소하는 것으로 관찰되었다 (Figs. 5와 6).
Periago 등 [20]은 외부 요인 (예, 4% ethanol, pH 5, 2.5% NaCl)
의 특정조건에 노출된 B. cereus에서 여러 가지 종류의 열충
격단백질의 발현에 대하여 연구하였으며, 이들 요인에 노출
30분 경과 후, 열충격에 대한 내성이 증가하였다고 보고하였
Fig. 5. Induction of stress shock protein (SSPs) in B. cereus MH-2 treated with different NaCl concentrations for 3 h; 0 % (lane 1), 2.5% (lane
2), 5% (lane 3), 7.5% (lane 4), 10% (lane 5). The SSPs were analyzed by SDS-PAGE (A), and western blot with anti-DnaK (B), and anti-
GroEL (C) monoclonal antibodies, respectively. DnaK and GroEL signals were decreased depending on the increasing NaCl concentrations.
Fig. 6. Induction of stress shock proteins (SSPs) in B. cereus MH-2 treated with 10% NaCl for different exposure period; 0 h (lane 1), 1 h (lane
2), 2 h (lane 3), 3 h (lane 4), 4 h (lane 5). The SSPs were analyzed by SDS-PAGE (A), and western blot with anti-DnaK (B), and anti-GroEL
(C) monoclonal antibodies, respectively. DnaK and GroEL signals were gradually decreased depending on the increasing NaCl exposure period.

용혈성 Bacillus cereus의 동정 및 특성 조사 185
다. DnaK와 GroEL은 손상된 단백질을 정상으로 회복시키는
역할을 하는 열충격단백질로서, E. coli에서 처음 발견되어
발현 조사가 이루어졌으나, 최근 전통장류의 제조공정에서
사용되는 B. subtilis에서 뿐만 아니라, 식중독 세균인 Salmo-
nella, Aeromonas 등에서도 외부 환경변화에 의해 발현이 유
도되는 것으로 보고되었다 [20-22]. 본 연구는 Western blot 기
법을 이용하여 B. cereus에서 NaCl의 다양한 농도와 노출시
간에 따른 결과를 보여주는 것으로 스트레스 충격단백질의
변화 패턴을 관찰한 연구결과는 거의 보고된 바 없다.
3.8. NaCl 노출에 따른 세포 외부형태 관찰
분리세균 MH-2가 NaCl에 노출되었을 때 발생되는 세포 외
부형태의 변화를 주사전사현미경을 이용하여 관찰하였다.
NaCl에 미처리된 MH-2세포는 정상적인 둥근 간균으로 보여
주었으나, 10% 농도의 NaCl에 노출된 세균은 세포의 표면이
심하게 손상을 입은 것으로 관찰되었다 (Fig. 7). B. cereus 세
포 외부의 형태적 변화는 녹차카테킨인 EGCG에 노출되었을
경우에 다른 양상으로 일어난다는 것이 보고되었다. Kim 등
[11]은 EGCG에 노출된 B. cereus에서 노출 농도 및 시간이 증
가함에 따라 세포표면이 울퉁불퉁해지고, 특정 부분이 돌출
되는 것을 보고하였으며, Shigemune 등 [23]은 B. cereus JCM
2152 세포가 EGCG에 노출되었을 때 세포 표면이 심하게 주
름이 지고, 손상되는 모습을 관찰하였다. 본 연구에서는 NaCl
에 노출되었을 때 일어난 형태적 변화를 관찰한 것으로 EGCG
노출에서 관찰된 것에 비하여 더욱 손상된 것으로 나타났는
데, 노출 물질의 종류에 따른 손상기작이 다른지에 대한 연구
는 추가적인 연구가 필요할 것으로 사료된다.
최근에 B. cereus에 의한 식중독 사례가 증가하는 추세에 있
어 식중독 세균으로서의 B. cereus가 관심의 대상이 되고 있
다. 특히 본 연구에서 국민들의 기호식품인 쌈장에서 용혈성
B. cereus가 검출된 것은 상온으로 유통되고 있는 장류의 식
품안전성에 대한 적신호로 이를 극복하기 위한 대안이 필요
한 상황이다. 최근 국민건강을 위하여 장류, 젓갈류 등의 저
염화가 이루어지고 있으나, 저염도에서 다양한 식중독세균
의 생존이 지속적으로 보고되고 있다. 따라서 식품안전을 위
하여 저염도의 기준을 정하는 것이 주목받고 있다. 향후 본
연구에서의 결과를 바탕으로 용혈성 B. cereus가 생산하는 용
혈단백질의 특성과 저해 기작의 규명에 대한 연구가 진행되
어야 할 것이다.
4. CONCLUSIONS
본 연구는 시판되는 쌈장에서 용혈성의 Bacillus cereus MH-2
를 분리 동정하고 특성조사를 하기 위하여 수행되었다. 먼저
MH-2의 생리학적 및 생화학적 특성을 조사하였다. 분리 균
주는 BIOLOG 분석시스템을 사용하여 B. cereus MH-2로 동
정하였다. MH-2의 16S rRNA 염기서열 비교분석하여 유전
학적 계통수를 작성하였다. MH-2 배양액으로 채워진 sheep
blood 평판배지의 wells 주변에서 용혈활성이 관찰되었으며,
용혈대는 MH-2 배양액의 배양시간에 따라 서서히 증가하였
다. Zymography 분석을 통해서 용혈현상의 원인이 되는 분자
의 분자량은 약 30 kDa 가량되는 것으로 관찰되었다. MH-2
세포의 생존율은 배지 내에 포함된 NaCl의 농도에 따라 감
소하였다. 스트레스 충격단백질 생성 (예, DnaK과 GroEL)은
NaCl의 농도와 노출시간 증가에 따라 감소되었다. 대수생장
기의 배양에서 70-kDa DnaK와 60-kDa GroEL의 스트레스 충
격단백질은 NaCl 농도 증가와 노출기간이 길어짐에 따라 발
현은 비례하여 감소되는 것이 SDS-PAGE and Western blot 분
석을 통해서 관찰되었다. NaCl에 처리된 세포에서 구멍이 뚫
리고 주름이 있는 불규칙한 형태가 주사전자현미경 분석을
통해서 관찰되었다.
Acknowledgements
본 연구는 순천향대학교의 학술연구지원사업의 일부 지원으
로 수행되었음.
Fig. 7. Scanning electron micrographs of B. cereus MH-2. (A) untreated cells, (B) cells treated with 10% NaCl for 3 h.

186 Korean Society for Biotechnology and Bioengineering Journal 32(3): 179-186 (2017)
REFERENCES
1. Kwon, S. H., K. B. Lee, K. S. Im, S. O. Kim, and K. Y. Park (2006)
Weight reduction and lipid lowering effects of Korean traditional
soybean fermented products. Kor. J. Food Sci. Nutr. 35: 1194-1199.
2. Kwak, C. S., M. S. Lee, and S. C. Park (2007) Higher antioxidant
properties of cheonggukjang, a fermented soybean paste, may be
due to increased aglycone and malonylglycoside isoflavone during
fermentation. Nutr. Res. 27: 719-727.
3. Park, Y. B., J. B. Kim, S. W. Shin, J. C. Kim, S. H. Cho, B. K. Lee,
J. J. Ahn, and D. H. Oh (2009) Prevalence, genetic diversity, and
antibiotic susceptibility of Bacillus cereus strains isolated from rice
and cereals collected in Korea. J. Food Prot. 72: 612-617.
4. Granum, P. E. and T. Lund (1997) Bacillus cereus and its food poi-
soning toxins. FEMS Microbiol. Lett. 157: 223-228.
5. Do, S. D., Y. M. Lee and H. G. Chang (1993) The study on kinds
and utilities of jeot-kal (fermented fish products). Kor. J. Soc. Food
Sci. 9: 222-229.
6. World Health Organization (2007) Reducing salt intake in popula-
tions. Report of a WHO Forum and Technical Meeting, Geneva,
Switzerland.
7. Choi, S. A. and M. S. Cho (2012) Changes in quality characteris-
tics of eggplant pickles by salt content and drying time during stor-
age. J. Kor. Soc. Food Cult. 27: 211-224.
8. Raevuori, M. and C. Genigeorgis (1975) Effect of pH and sodium
chloride on growth of Bacillus cereus in laboratory media and cer-
tain foods. Appl. Microbiol. 29: 68-73.
9. Leber, T. M. and F. R. Balkwill (1997) Zymography: A single-step
staining method for quantitation of proteolytic activity on substrate
gels. Anal. Biochem. 249: 24-28.
10. Kleiner, D. E. and W. G. Stetlerstevenson (1994). Quantitative zym-
ography: Detection of picogram quantities of gelatinases. Anal.
Biochem. 218: 325-329.
11. Kim, D. M., S. K. Park, and K. H. Oh (2016) Cellular responses
and proteomic analysis of hemolytic Bacillus cereus MH-2 exposed
to epigallocatechin gallate (EGCG). Kor. J. Microbiol. 52: 260-268.
12. Bradford, M. M. (1976) A rapid and sensitive method for the quan-
titation of microgram quantities of protein utilizing the principle of
protein-dye binding. Anal. Biochem. 72: 248-254.
13. Beecher, D. J. and A. C. Wong (1994) Identification of hemolysin
BL-producing Bacillus cereus isolates by a discontinuous hemoly-
tic pattern in blood agar. Appl. Environ. Microbiol. 60: 1646-1651.
14. Beecher, D. J., J. L. Schoeni, and A. C. Wong (1995) Enterotoxic
activity of hemolysin BL from Bacillus cereus. Infect. Immun. 63:
4423-4428.
15. Dietrich, R., C. Fella, S. Strich, and E. Märtlbauer (1999) Produc-
tion and characterization of monoclonal antibodies against the hem-
olysin BL enterotoxin complex produced by Bacillus cereus. Appl.
Environ. Microbiol. 65: 4470-4474.
16. Allison, H. E. and J. D. Hillman (1997) Cloning and characteriza-
tion of a Prevotella melaninogenica hemolysin. Infect. Immun. 65:
2765-2771.
17. Yang, H. O., M. S. Cha, and M. J. Kim (1998) Studies on the hem-
olysin produced by Vibrio vulnificus ys-1. Kor. J. Life Sci. 8: 145-
157.
18. Rossignol, G., A. Merieau, J. Guerillon, W. Veron, O. Lesouhaitier,
M. G. Feuilloley, and N. Orange (2008) Involvement of a phospho-
lipase C in the hemolytic activity of a clinical strain of Pseudomo-
nas fluorescens. BMC Microbiol. 8: 189.
19. den Besten, H. M., C. J. Ingham, van Hylckama Vlieg, J. E., Beer-
thuyzen, M. M., M. H. Zwietering, and T. Abee (2007). Quantita-
tive analysis of population heterogeneity of the adaptive salt stress
response and growth capacity of Bacillus cereus ATCC 14579.
Appl. Environ. Microbiol. 73: 4797-4804.
20. Periago, P. M., W. van Schaik, T. Abee, and J. A. Wouters (2002)
Identification of proteins involved in the heat stress response of
Bacillus cereus ATCC 14579. Appl. Environ. Microbiol. 68: 3486-
3495.
21. Chuang, S. E. and F. R. Blatiber (1993) Characterization of twenty
six new heat shock genes of Escherichia coli. J. Bacteriol. 175:
5242-5252.
22. Kim, D. M. and K. H. Oh (2016) Characterization of hemolytic
Aeromonas sp. MH-8 responding to the green tea catechin, EGCG.
KSBB 31: 228-236.
23. Shigemune, N., M. Nakayama, T. Tsugukuni, J. Hitomi, C. Yoshi-
zawa, Y. Mekada, M. Kurahachi, and T. Miyamoto (2012) The
mechanisms and effect of epigallocatechin gallate (EGCg) on the
germination and proliferation of bacterial spores. Food Control 27:
269-274.

![조선 후기 禮敎적 시선의 변주와 변화* - Hallymtaedong/nonmun/pdf/35/35_ (10).pdf · 2017. 6. 21. · 조선 후기 禮敎적 시선의 변주와 변화 291 다리머리[髲髢]를](https://img.pdfslide.tips/doc/110x75/610d68beb12f3d50560d02df/-e-c-oe-e-e-hallym-taedongnonmunpdf3535.jpg)












![Research Papersoongene.net/nonmun/KSBB145-154.pdf · buffer (pH 6.8)를 첨가하여 흡광도 값을 측정하였다. Tyrosinase inhibitory activity (%) = [(C −T) / C] × 100 C](https://img.pdfslide.tips/doc/110x75/5e31f4860bcdef63e4582823/research-buffer-ph-68e-e-ee-e-e-tyrosinase.jpg)











