Embed Size (px)
Citation preview

Badanie funkcji genu

Funkcję genu można zbadać różnymi sposobami
Strategia „ukierunkowanej” analizy funkcji genu
1. wprowadzeniemutacji w genie
2. nadekspresjagenu
3. potranskrypcyjnewyciszanie genu(antysensowneRNA oraz RNAi)
GEN FENOTYP
„Przypadkowa” analizy funkcji genu
FENOTYPMUTACJA GEN

1. Inaktywacja (mutacja) genu poprzez homologiczną rekombinację jest podstawą analizy jego funkcji
1. „Kopia” genu, który chcemy wyłączyć czyli odpowiedni fragment
DNA sklonowany w wektorze plazmidowym w komórkach E. coli
2. Przebudowanie sklonowanego genu in vitro – „konstrukt do mutagenezy”
3. Przebudowana kopia genu (konstrukt do mutagenezy) jest dostarczana na wektorze do komórek bakterii X. xxxx
Początek i koniec genu, który chcemy wyłączyć
Bakteria E. coli
Bakteria X. xxxxGen, którego funkcję chcemy ustalić
Gen kodujący oporność na antybiotyk np. kan(kaseta oporności)
4. Homologiczna rekombinacja przerywa gen docelowy – inaktywuje go przez wstawienie kasety kan do genomu bakterii X. xxxx

Jak uzyskać mysz - z „nokautem” genowym ??????
Problemy:
•Diploidalny genom
•Myszy nie da się wyhodować na płytce z selekcją na antybiotyk
•I wiele innych....

1. Infekcja wczesnego zarodka zrekombinowanym wektorem pochodzenia wirusowego
Wektor retrowirusowy
Komórki wczesnego mysiego zarodka

2) Mikroiniekcja in vitroDNA do jednego z przedjądrzy zapłodnionej komórki jajowej przed pierwszym jej podziałem

3) Zastosowanie zarodkowych komórek macierzystych ES (embrionic stem) – totipotencjalnych tzn. takich które nie są przypisane do
żadnego szlaku rozwojowego i mogą dać początek każdemu rodzajowi zróżnicowanych komórek. Takie komórki poddaje się modyfikacji in vitro (w kontrolowanych warunkach). Genetycznie zmienione (po homologicznej rekombinacji in vitro) komórki ES wprowadza się do zarodka myszy
1. Etap: pozyskanie komórek ES i hodowla in vitro
komórki węzła zarodkowego są zdolne do różnicowania się we wszystkie typy komórek

•neo, gen kodujący enzym, który inaktywuje antybiotyk neomycynę oraz jego pochodne, np. G418, który jest toksyczny dla komórek ssaczych;
•tk, gen kodujący kinazę tymidynową – enzym, który fosforyluje analogi nukleozydów np. gancyklowir. Polimeraza DNA może wbudować taki analog nukleotydu do nowosyntetyzowanej nici DNA co prowadzi do zaburzenia procesu replikacji DNA. Dlatego komórki zawierające gen tk, giną po podaniu gancyklowiru (są wrażliwe na gancyklowir).
2. Etap: przygotowanie konstruktu do mutagenezy in vitro na bazie „kopii” docelowego genu sklonowanego w wektorze w E. coli
Tylko pewien procent wprowadzonych konstruktów włącza się, w wyniku homologicznej rekombinacji w komórkach zarodkowych, we właściwej pozycji genomowej. Dlatego konstrukt do mutegenezy wyposaża się w dodatkowe elementy genetyczne
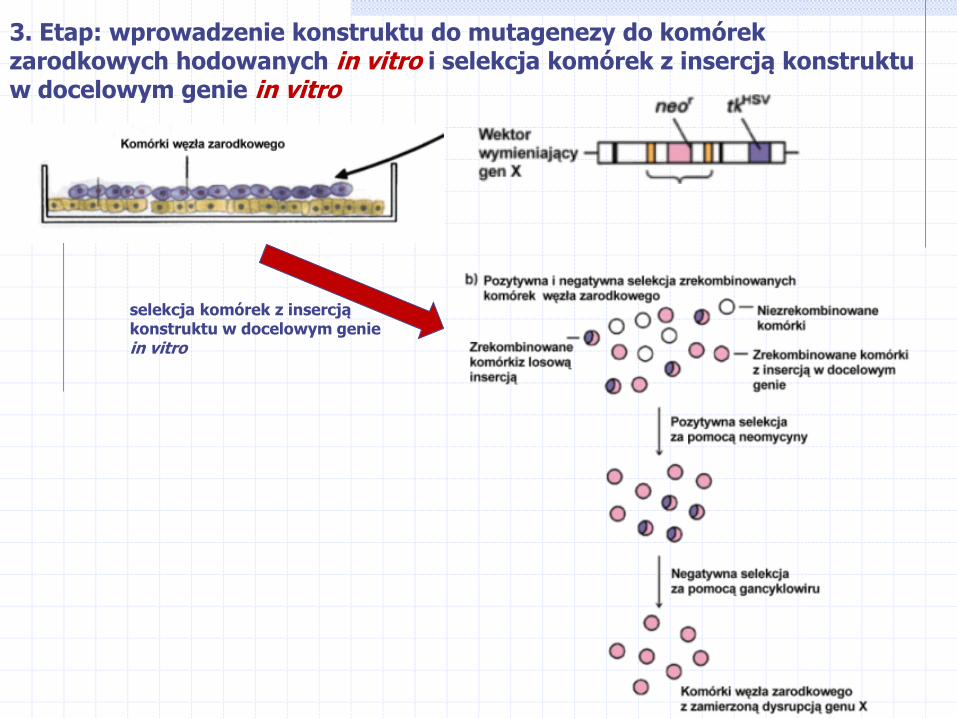
3. Etap: wprowadzenie konstruktu do mutagenezy do komórek zarodkowych hodowanych in vitro i selekcja komórek z insercją konstruktu w docelowym genie in vitro
selekcja komórek z insercją konstruktu w docelowym genie in vitro

Uzyskujemy celowe „uszkodzenie” (knock-out) genu i uzyskanie tzw. myszy transgenicznych z nokautem genowym, dzięki któremu możliwe staje się zbadanie funkcji takiego genu
4. Etap: iniekcja wyselekcjonowanych in vitro komórek zarodkowych z nokautem genowym i uzyskanie potomstwa z nokautem genowym we wszystkich komórkach ciała

Nokaut genowy w konkretnej tkance lub nadrządzie – zastosowanie
systemu Cre-lox
• Cre miejscowo specyficzna
rekombinaza
• działa na miejsca loxP (potworzone
sekwencje długości 34 pz).
• Dlaczego wycinanie DNA za pomocą Cre/loxP może być użyteczne w
komórkach eukariotycznych?
• System Cre/loxP pochodzi z bakteriofaga P1, a sekwencje docelowe typu loxP
rozpoznawane przez rekombinazę Cre nie występują np. w roślinach czy u zwierząt
• sekwencje loxP mogą być „sztucznie” wstawiane do genomów zwierząt/roślin w
celu precyzyjnego „oznaczenia” określonych sekwencji DNA przeznaczonych do
wycinania, bez obawy że przy okazji może dojść do usunięcia jakiejś istotnej część
genomu

Jak za pomocą systemu Cre/loxP otrzymać mysz, z nokautem genu
w określonej tkance/narządzie?
Do uzyskania obydwu linii transgenicznych myszy (z genem cre pod kontrolą tkankowo-specyficznego promotora oraz z miejscami loxPotaczającym docelowy gen) opiera się na homologicznej rekombinacji, która przeprowadza się in vitro w zarodkowych komórkach macierzystych

Strategia „ukierunkowanej” analizy funkcji genu
1. wprowadzeniemutacji w genie
2. nadekspresjagenu
3. potranskrypcyjnewyciszanie genu(antysensowneRNA oraz RNAi)
GEN FENOTYP

Inaktywacja genu z użyciem tzw. „programowanych nukleaz”
• Metody specyficznych modyfikacji genomów, określane jako „edycja genomu” wykorzystują tzw. „programowane” nukleazy
• System CRISPR-Cas (clusteredregularly interspaced short palindromicrepeats-CRISPR associated), w którym główną rolę odgrywa jedna z tzw. programowanych nukleaz - Cas9
• System wykorzystuje elementy mechanizmu „nabytej odporności” („pamięci immunologicznej”) bakterii i archeonów na infekcję fagami i transformację obcym materiałem genetycznym
• Istnieje kilka typów opisanych systemów CRISPR-Cas, w inżynierii genetycznej zastosowanie znalazły narzędzia oparte na systemie CRISPR typu II.

• Mikroorganizmy włączają do swojego genomu (loci CRISPR) fragmenty obcego DNA (protospacer), co umożliwia im w przyszłości szybkie rozpoznanie i zniszczenie obcego DNA (np. fagowego).
• Długość włączanych sekwencji różni się i wynosi 21-72 nukleotydów; najczęściej są to fragmenty 32-38-nukleotydowe
• Każdej integracji towarzyszy duplikacja końcowej sekwencji powtórzonej tak, aby została zachowana architektura: powtórzenie proste-sekwencja rozdzielająca-powtórzenie proste (długość powtórzeń prostych także jest gatunkowo swoista i wynosi 24-47 nt, zwykle 28-37 nt)
• W integracji tych sekwencji do genomu biorą udział dwie endonukleazy: Cas1 oraz Cas2
System CRISPR-Cas – zasada działania

• Przy ponownym „kontakcie” z obcym DNA moduł kodujący
crRNA ulega ekspresji – powstaje pierwotny transkrypt
pre-crRNA, obejmujący wszystkie sekwencje rozdzielające
i powtórzenia proste
• Dojrzewanie pre-crRNA wymaga obecności transaktywującego RNA (tracrRNA). tracrRNAhybrydyzuje z sekwencjami odpowiadającymi powtórzeniom prostym w pre-crRNA
• Dupleks taki jest trawiony przez bakteryjną RNazę III w obecności nukleazy Cas9, tworząc dojrzałe cząsteczki crRNA
• Dojrzałe cząsteczki crRNA rozpoznają (na zasadzie komplementarności) obce DNA, które ulega trawieniu przez obecną w kompleksie nukleazę Cas9
crRNA rozpoznaje obce DNA, które ulega degradacji przez Cas9

• W systemach będących narzędziem do precyzyjnej modyfikacji genomowego DNA komórek eukariotycznych do wprowadzenia zmiany w pożądanym miejscu genomu niezbędne elementy systemu są dostarczone komórce „z zewnątrz”. Ich ekspresja (zazwyczaj przejściowa), powoduje wprowadzenie stałych zmian w docelowym genomie
• Systemy CRISPR-Cas używane jako narzędzia są uproszczone przez zastosowanie chimerycznego RNA (połączenie crRNA z tracrRNA) określanego mianem sgRNA (single-guide RNA )
• Fragment crRNA w obrębie sgRNA nakierowuje kompleks na docelowe miejsce w genomie, w którym dochodzi do trawienia DNA
• CRISPR-Cas typu II wykorzystuje małe cząsteczki RNA (crRNA i tracrRNA) do precyzyjnego nakierowania nukleazy efektorowej – Cas9 – na konkretne miejsce w DNA (jest to sekwencja komplementarna do crRNA)

System wykorzystujący nukleazę Cas9 może być użyty do:
• wprowadzania trwałych zmian w genomie (np. typu nokaut) – system CRISPR-Cas9 pozwala na samo przecięcie nici DNA, jednak po połączeniu go z mechanizmami naprawy DNA możliwe jest nie tylko wycięcie określonego segmentu DNA ale i wstawienie w przecięte miejsce dostarczonego z zewnątrz elementu genetycznego
• aktywacji bądź wyciszenia ekspresji wybranych genów (narzędzia oparte na systemie CRISPR-Cas znalazły zastosowanie przy tworzeniu zwierzęcych i komórkowych modeli wielu chorób, np. konkretnych typów nowotworów)
Edycja genomu wykorzystująca programowane nukleazy może znaleźć szerokie zastosowanie m.in. w medycynie np. do walki z chorobami genetycznymi
Jednoczesne powstanie dwóch pęknięć w obu niciach DNA na jednym chromosomie (DSB – double strandbrake) może prowadzić do pojawienia się mutacji typu indel (efekt naprawy przecięcia DNA) w obu docelowych loci albo do delecji bądź insercji obcego fragmentu DNA (nokaut genu)
mutacje typu indel duża delecja insercja obcego DNA

Strategia „ukierunkowanej” analizy funkcji genu
1. wprowadzeniemutacji w genie
2. nadekspresjagenu
3. potranskrypcyjnewyciszanie genu(antysensowneRNA oraz RNAi)
GEN FENOTYP

2. Nadekspresja genu - celem jest ustalenie czy znacząco zwiększona
ekspresja badanego genu wpłynie na fenotyp organizmu np. myszy transgenicznej.
Transgeniczna mysz do której komórek wprowadza się cDNAbadanego genu sklonowany w wektorze pod kontrolą silnego promotora, który spowoduje ekspresję cDNA np. w określonych tkankach

3. Potranksrypcyjne wyciszanie genu
•antysensowne RNA (aRNA)
•interferencja RNA (RNAi)
Strategia „ukierunkowanej” analizy funkcji genu
1. wprowadzeniemutacji w genie
2. nadekspresjagenu
3. potranskrypcyjnewyciszanie genu(antysensowneRNA oraz RNAi)
GEN FENOTYP
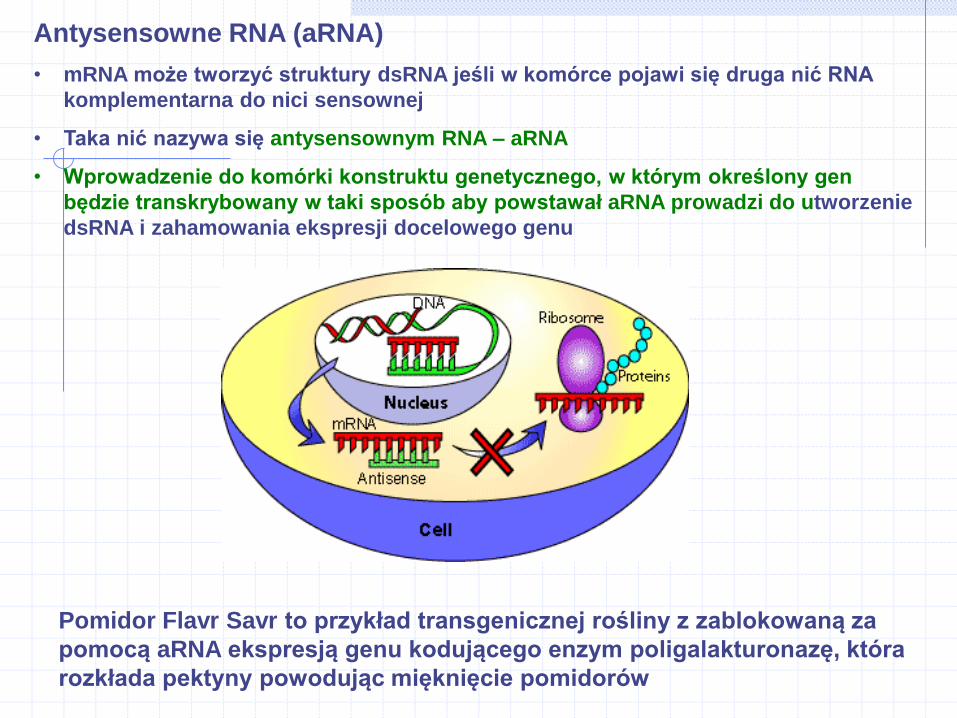
Antysensowne RNA (aRNA)
• mRNA może tworzyć struktury dsRNA jeśli w komórce pojawi się druga nić RNA
komplementarna do nici sensownej
• Taka nić nazywa się antysensownym RNA – aRNA
• Wprowadzenie do komórki konstruktu genetycznego, w którym określony gen
będzie transkrybowany w taki sposób aby powstawał aRNA prowadzi do utworzenie
dsRNA i zahamowania ekspresji docelowego genu
Pomidor Flavr Savr to przykład transgenicznej rośliny z zablokowaną za
pomocą aRNA ekspresją genu kodującego enzym poligalakturonazę, która
rozkłada pektyny powodując mięknięcie pomidorów

Interferencja RNA (RNAi) • 1990 Napoli i wsp. przeprowadzili doświadczenie którego celem było uzyskanie odmiany
petunii z kwiatami o bardziej intensywnej purpurowej barwie
• W tym celu do komórek roślinnych wprowadzono dodatkowe kopie genu (cDNA)
syntazy chalonowej odpowiedzialne za syntezę fioletowego barwnika kwiatów
• W efekcie zamiast ciemniejszych kwiatów uzyskano odmiany pstrokate lub zupełnie białe.
Wprowadzenie dodatkowego genu nie prowadziło do wydajniejszej syntezy barwnika, ale spowodowało zahamowanie wytwarzania barwnika kwiatu
• Pomiar w komórce roślinnej petunii poziomu mRNA genu, którego dodatkową kopię dodano wykazał, że jego poziom znacząco spadł, a obserwowane zjawisko miało tendencję do rozprzestrzeniania się na inne komórki rośliny
Zjawisko to nazwano kosupresją - zahamowanie
ekspresji dotyczyło zarówno „normalny” gen
kodujący purpurową barwę, jak również jego
wprowadzonych dodatkowych kopii

W 1998 r. Fire i wsp. scharakteryzowali na poziomie molekularnym potranskrypcyjny mechanizm regulujący poziom mRNA w komórkach nicienia C. elegans
• Opisano zjawisko zostało nazwane interferencją RNA (RNA interference – RNAi)
• polegało ono na wyciszaniu ekspresji docelowego genu przez krótkie dwuniciowe RNA którego jedna nić była komplementarna do mRNA konkretnego genu
• Potranskrypcyjne wyciszanie zachodziło na zasadzie degradacji mRNA określonego genu

Wyciszanie ekspresji genu poprzez degradację mRNA
• w przypadku degradacji mRNA w komórce obserwuje się obecność
specyficznych pośredników interferencji - są to małe (długości 21-23 pz)
dwuniciowe RNA (dsRNA) tzw. interferujące RNA (siRNA, small interferingRNA)
• Jedna z nici siRNA jest komplementarna do transkryptuwyciszanego genu
Skąd się biorą w komórce cząsteczki siRNA?
• Powstają w wyniku trawienia większych cząsteczek dsRNA, które
zazwyczaj są dla komórki sygnałem „nienormalności”,
• dsRNA może pojawić się w komórce np. w wyniku infekcji wirusem RNA czy
nadmierną ilością kopii genu
• Uważa się, że mechanizm siRNA powstał w komórkach najprawdopodobniej jako system obrony przed dwuniciowymi wirusami RNA (dsRNA)

Jak działa RNAi na poziomie
molekularnym?
• dsRNA jest dla komórki czymś
niezwykłym i sygnałem „nienormalności”.
• Obecna w komórkach rybonukleaza
Dicer, rozpoznaje dsRNA i tnie go małe
fragmenty 21-23 pz– siRNA.
• Następnie powstaje tzw. kompleks
wyciszającego zwany RISC (RNA-
induced silencing complex), który ulega
aktywacji wtedy kiedy siRNA rozdzieli się
na pojedyncze nici,
• Aktywny kompleks RISC wiąże się do
mRNA za pomocą antysensownego RNA.
• Obecna w kompleksie RISC rybonukleaza
Slicer, trawi związane mRNA powodując
wyciszenie ekspresji genu.

•Zjawisko RNAi jest powszechne u komórkach Eukariota (oprócz drożdży),
występuje u pierwotniaków, jamochłonów, muszki owocowej, ryb, płazów, grzybów,
myszy, człowieka, u różnych gatunków roślin niższych i wyższych
•Zjawisko RNAi jest niezwykle specyficzne: dostarczenie do cytoplazmy
komórki dwuniciowych cząsteczek dsRNA odpowiadających mRNA
konkretnego genu powoduje skuteczne obniżenie poziomu ekspresji tego
genu (poprzez degradację jego transkryptu)
RNAi można zastosować:
1. W badaniach podstawowych: do precyzyjnego i ukierunkowanego
blokowania ekspresji genów
2. W praktyce: np. w terapii genowej (m.in. nowotworów – wstrzykiwanie
siRNA komplementarnego do znanych protoonkogenów)

Dostarczanie siRNA do ukierunkowanego wyciszania genów
1. siRNA może być produkowane in vivo
• W wektorze umieszcza się odpowiednio
spreparowane DNA, w którym do nici
sensownego RNA danego genu
wprowadza się krótkie odcinki
antysensowne,
• W wyniku ekspresji w komórkach
docelowych nić sensowna i
antysensowna będą mogły utworzyć
odcinki dwuniciowego RNA,
• dsRNA po przekształceniu w siRNA
prowadzi do zahamowanie ekspresji
konkretnego genu

2. siRNA może być produkowane in vitro i dostarczane do komórki
•przez transkrypcję in vitro małych RNA (siRNA)
• transkrypcja in vitro długich RNA,
które będą wprowadzane do komórki
i przekształcane w siRNA przez rybonukleazę
typu Dicer
• Na drodze syntezy chemicznej siRNA

Potranskrypcyjne wyciszanie jest powszechnym mechanizmem regulacji ekspresji genów w komórkach eukariotycznych
a) degradacji mRNA b) blokowania translacji mRNA
• W przypadku mechanizmu interferencji z translacją mRNA, mediatorami są cząsteczki tzw. mikro-RNA (miRNA), zakodowane w genomie „jak normalne geny”
• Geny kodujące są transkrybowane przez polimerazę RNA II – powstają długie cząsteczki miRNA, które tworzą struktury typu szpilki do włosów, podlegające obróbce podobnej do siRNA
• miRNA wchodzą w skład kompleksów rybonukleoproteinowych specyficznie blokujących translację mRNA
• W odróżnieniu od siRNA, sekwencja miRNA nie musi być w 100% identyczne do sekwencji docelowego mRNA
• Ocenia się, że miRNA biorą udział w regulacji 30% ludzkich genów
























