Embed Size (px)
Citation preview

BILJNA ĆELIJA

ĆELIJSKI CIKLUS
� Ćelijske deobe se odvijaju u meristemima, ćelije se dele svakih 18 - 24h, a deoba traje 4 - 6h.
� Ćelijski ciklus je strogo kontrolisan – faktorima rasta (auksinima, giberelinima, citokininima), kontrola preko ciklina, kinaza, fosforilacijom i defosforilacijom proteina.
� Bez kontrole organizam bi bio bezoblična nefunkcionalna masa.
MITOZA
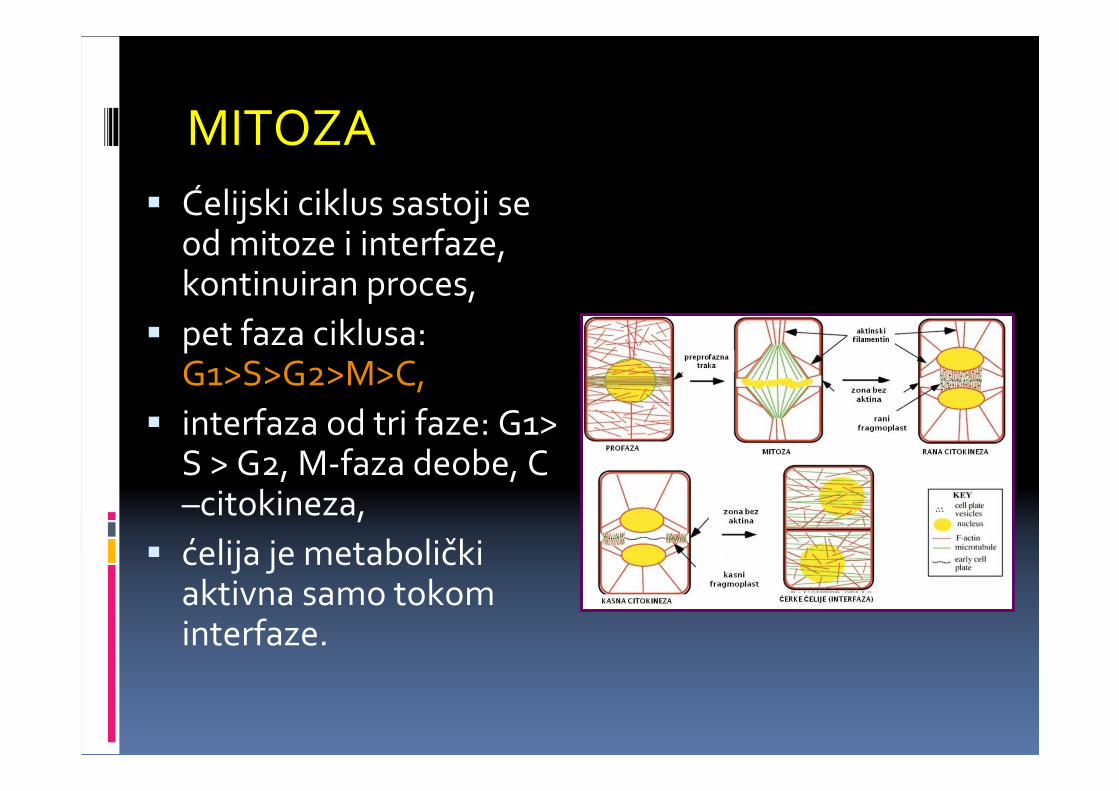
� Ćelijski ciklus sastoji se od mitoze i interfaze,kontinuiran proces,
� pet faza ciklusa: G1>S>G2>M>C,
� interfaza od tri faze: G1> S > G2, M-faza deobe, C –citokineza,
� ćelija je metabolički aktivna samo tokom interfaze.

MITOZA
� G1 – faza između ćelijske deobe i nove replikacije
DNK, ćelija sintetiše proteine, biomolekule.
� U S stadijumu sintetišu se histoni potrebni za
kondezaciju genetičkog materijala, vrši se
replikacija tj. udvajanje molekula DNK.
� G2 odvija se ugrađivanje proteina u hromozome i
akumulacije visokoenergetskih jedinjenja.

MITOZA
� Mitoza: profaza, metafaza, anafaza i telofaza
� Profaza - hromatin se kondezuje, forma hromozoma, sintetiše se proteinski kompleks kinetohor.
� Metafaza - dezintegracija membrane nukleusa, formira se deobno vreteno, kinetohorima se hromozomi vezuju za deobno vreteno, u kretanju hromatida važnu ulogu imaju MAPS proteini (microtubule - associated proteins).

MITOZA
� Anafaza - razdvajanje sestrinskih hromatida, na kraju ove faze svi hromozomi su podeljeni, formira se nukleus, identičan genetički materijal u obe nove ćelije.
� Telofaza – uspostavlja se membrana oko hromozoma, uočava se nukleus i nukleolus, deobno vreteno se dezintegriše, između nukleusa ćerki ćelija formira se fragmoplast sagrađen od mikrotubula koji pozicionira vezikule sa gradivnim materijalom.
� C – faza citokineza, podela citoplazme.

DEOBA ćELIJA BILJAKA
Meristemi
Ravan deobe određuje polarnost embriona,pozicijutkiva i organa.
Kariokineza identična kao kod ćelija životinja.
Citokineza je potpuno drugačija.

DEOBA ćELIJA BILJAKA
Ne poseduju centrozome.
Organozovanje mitotičkog vretena zavisi odmolekularnih motora i pratećih proteinamikrotubula.
� Mikrotubule mitotičkog aparata raspoređenesubmembranski i aktinski filamenti učestvuju u formiraju preprofazne trake koja označavaoblast formiranja ćelijske ploče.

DEOBA ELIJA BILJAKA
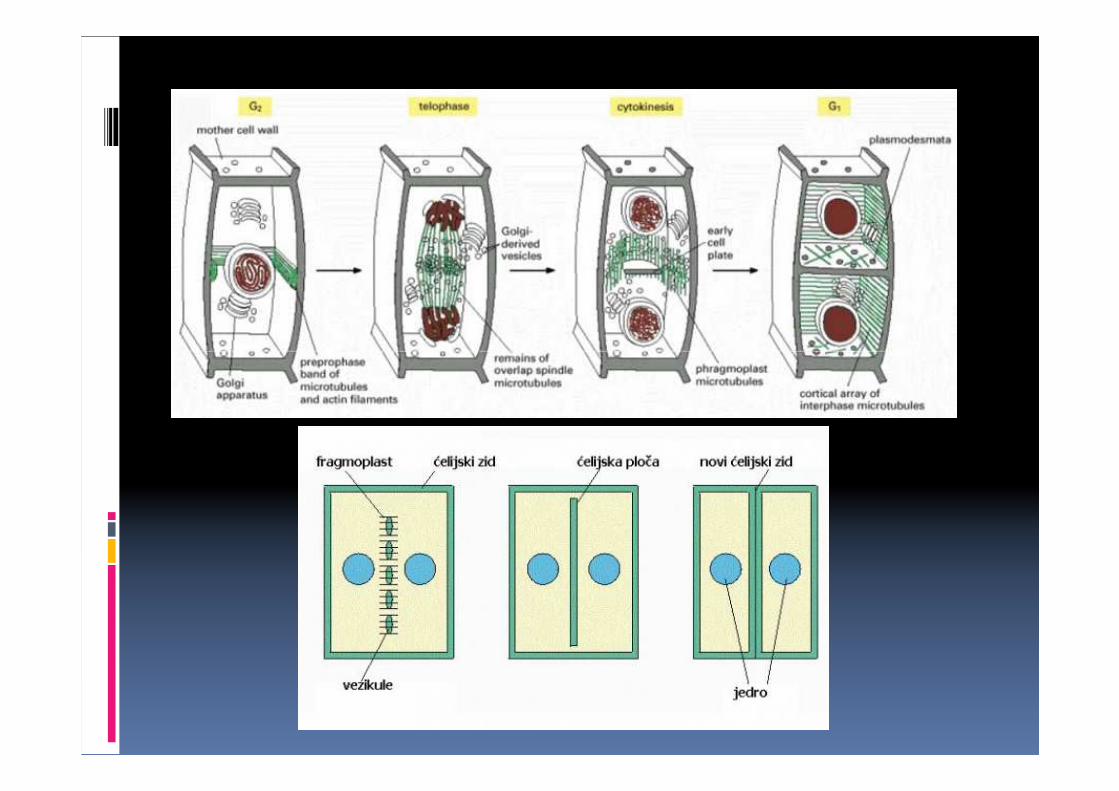
� Ćelijska ploča je membranska struktura u obliku diska koja se formira od vezikula kompleksa Goldži i leži u centru fragmoplasta.
� Fragmoplast specifično uređena citoskeletna struktura koja pomaže formiranje ploče.
� Tubule poreklom od KG i ER i vezikule ispunjene saharidima i glikoproteinima fuzionišu se u centru ekvatorijalne ravni formirajući tubulo-retikularnu membransku strukturu ćelijske ploče.

DEOBA ćELIJA BILJAKA
� Ćelijska ploča se širi ka plazminoj membrani majke ćelije fuzioniše se sa njom formirajući ćelijske membrane ćerki ćelija i primarnu lamelu između njih.
� Aktivnošću celulozo sintetaze počinje sinteza i depozicija primarnog ćelijskog zida.
� U pojedina okca ćelijske ploče umeću se tubule endoplazminog retikuluma stvarajući komunikacione veze-plazmodezme.

BILJNA ĆELIJA

središnja lamela
primarnićelijskizid
ćelijskamembrana
pektin
celuloznifibrili
hemiceluloza
ĆELIJSKI ZID
Srednja lamela-pektini Primarni zidSekundarni zid

PRIMARNI ĆELIJSKI ZID
- 1- 3 mm, elastičan,
- celuloze, hemiceluloze, pektina i glikoproteina
(celulozni fibrili povezuju se molekulma hemiceluloze, pektin kao želatin ispunjava šupljine između fibrila i lamela, hidratiše zid i omogućava dopremanje hranljivihmaterija iz spoljašnje sredine)
- mehanička snaga,
- oblik ćelije,
- reguliše širenje ćelije
- unutarćelijski transport,
- zaštita.

SEKUNDARNI ĆELIJSKI ZID
- Formira se poslezavršenog rasta ćelija,
- jamice, (S1, S2 i S3),
- sadrži 25% lignina i celulozu.
ĆELIJA 1
ĆELIJA 2primarni ćelijski zid
slojevi sekundarnog ćelijskog zida
središnjalamela
PLAZMODEZME
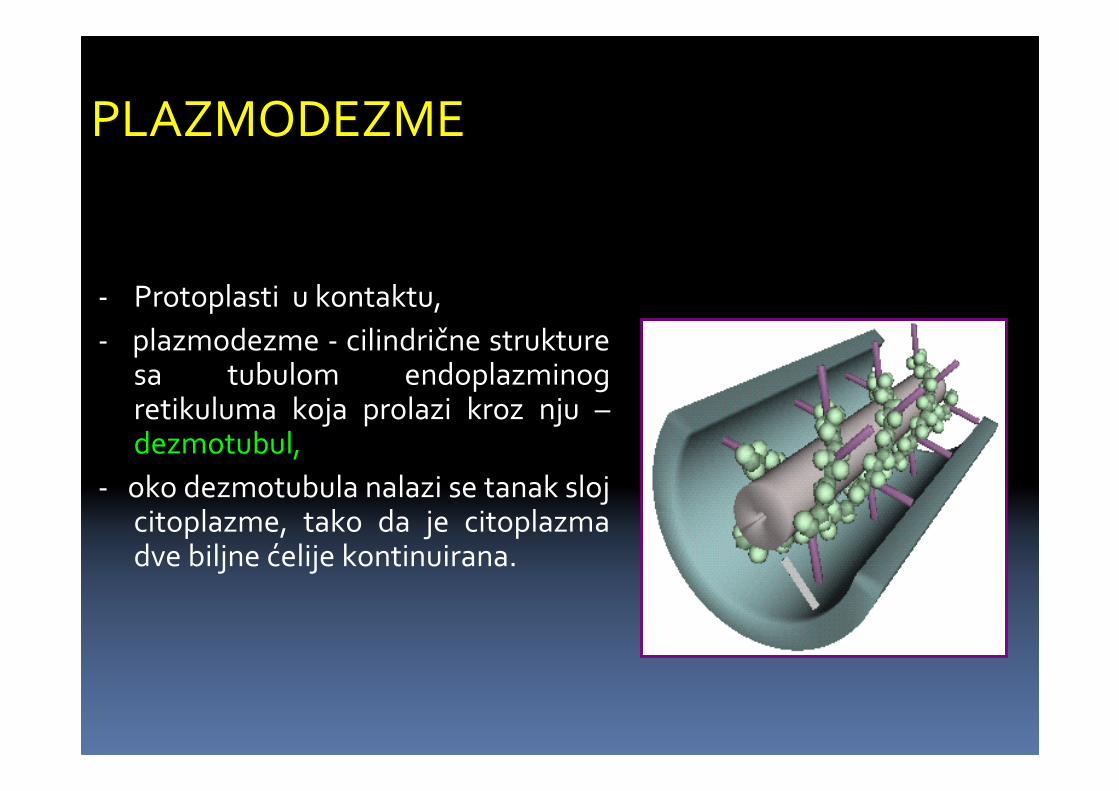
- Protoplasti u kontaktu,
- plazmodezme - cilindrične strukturesa tubulom endoplazminogretikuluma koja prolazi kroz nju –dezmotubul,
- oko dezmotubula nalazi se tanak slojcitoplazme, tako da je citoplazmadve biljne ćelije kontinuirana.

PLAZMODEZME
- obrazovanje jamica počinje sa mesta na komese u primarnom zidu nalazi najveći broj otvoraza prolaz plazmodezmskih vrpci,
- kroz plazmodezme se mogu kretati nutrienti, joni, manji molekuli, električni i hemijskisignali.
središnja
lamela
primarni
ćelijski
zid
endoplazmin
retikulum
dezmotubula

ćelijskizid
ćelijskamembran
anulus
dezmozubula
endoplazmin retikulum
plazmodezma
ĆELIJA 1 ĆELIJA 2ĆELIJA 1 ĆELIJA 2
endoplazmin
retikulum
ćelijski
zid
ćelijskamembrana
anulus
dezmozubula
ćelijska
membranaĆELIJA 2
ĆELIJA 1
PLAZMODEZME

ĆELIJSKA MEMBRANA
- Fosfolipidni dvoslojproteini, ugljeni hidrati
- polupropustljiva,

ĆELIJSKA MEMBRANA
- obrazovanja invaginacija,
- koordinacija sinteze celuloznih mikrofilamenata
ćelijskog zida,
- prevođenje hormonalnih i okolnih signala za
kontrolu ćelije i diferencijaciju.
CITOPLAZMA
- želatinozna,
- 70-80% vode.

VAKUOLA
� Meristemske ćelije – veći broj sitnijih vakuola
� Diferencirane ćelije – jedna velika vakuola
� Tonoplast
� Uloga:
� održavanje turgorovog pritiska,
� reguliše pH citoplazme
� skladište proteina (proteaze),
� enzimi, produkti degradacije i sinteze,
� pigmenti, toksične substance, organski i neorganski nutritienti, jedinjenja koja štite od UV zračenja.

� Hemijskih jedinjenja - (ugljeni hidrati, proteini, lipidi i nukleinske kiseline) prisutni su i
� alkaloidi, flavonoidi, organske kiseline, fenolna jedinjenja, biljni hormoni, vitamini ...
� specifične uloge u biljnoj ćeliji,obavljanje različitih funkcija, kao i prilagođenost u stalno promenljivim uslovima životne sredine.

ĆELIJSKE ORGANELE
� Dvomembranske organele (plastidi, mitohondrije, jedro)
� Jednomembranske (endoplazmin retikulum, goldžiaparat, lizozomi, sferozomi, mikrozomi).
� Nemembranske (ribozomi, mikrotubule).

JEDRO
- membrana,
- nukleoplazma,
- jedarce,
genetske informacijesadržane u DNK
kontroliše aktivnostićelije.

JEDRO
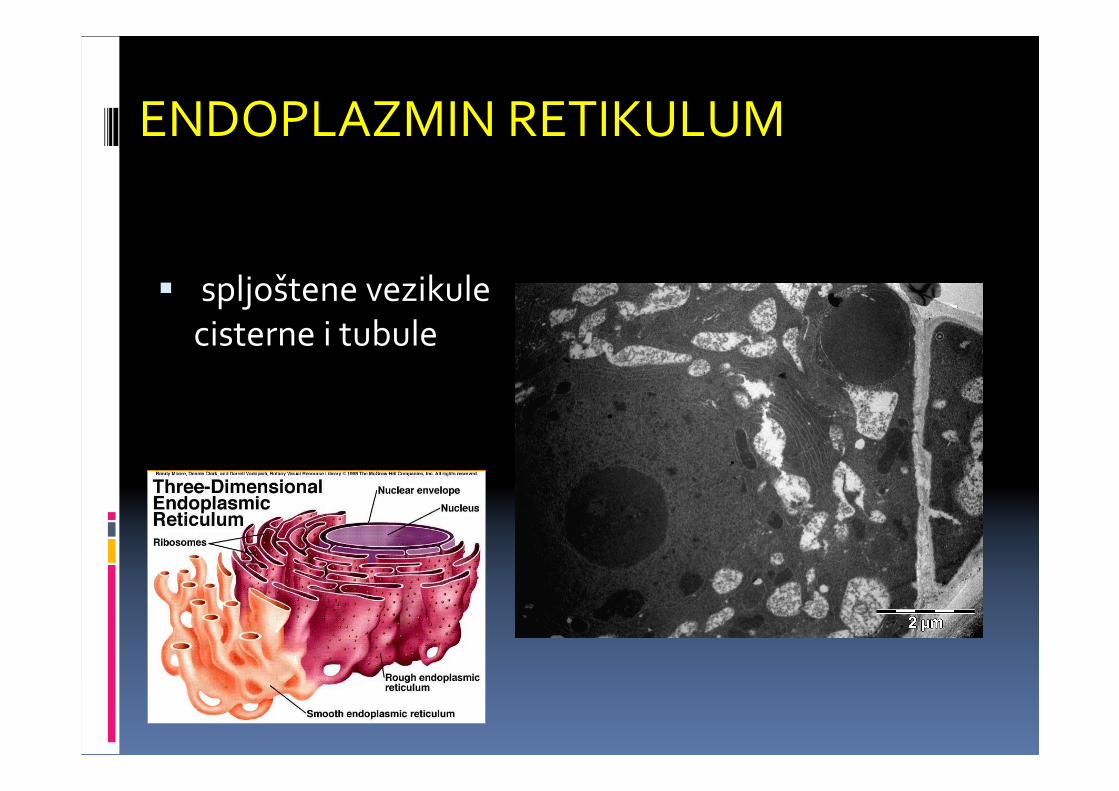
ENDOPLAZMIN RETIKULUM
� spljoštene vezikulecisterne i tubule
ENDOPLAZMIN RETIKULUM
sinteza proteina,lipida,
transport,
razmena supstanci,
enzimski procesi.
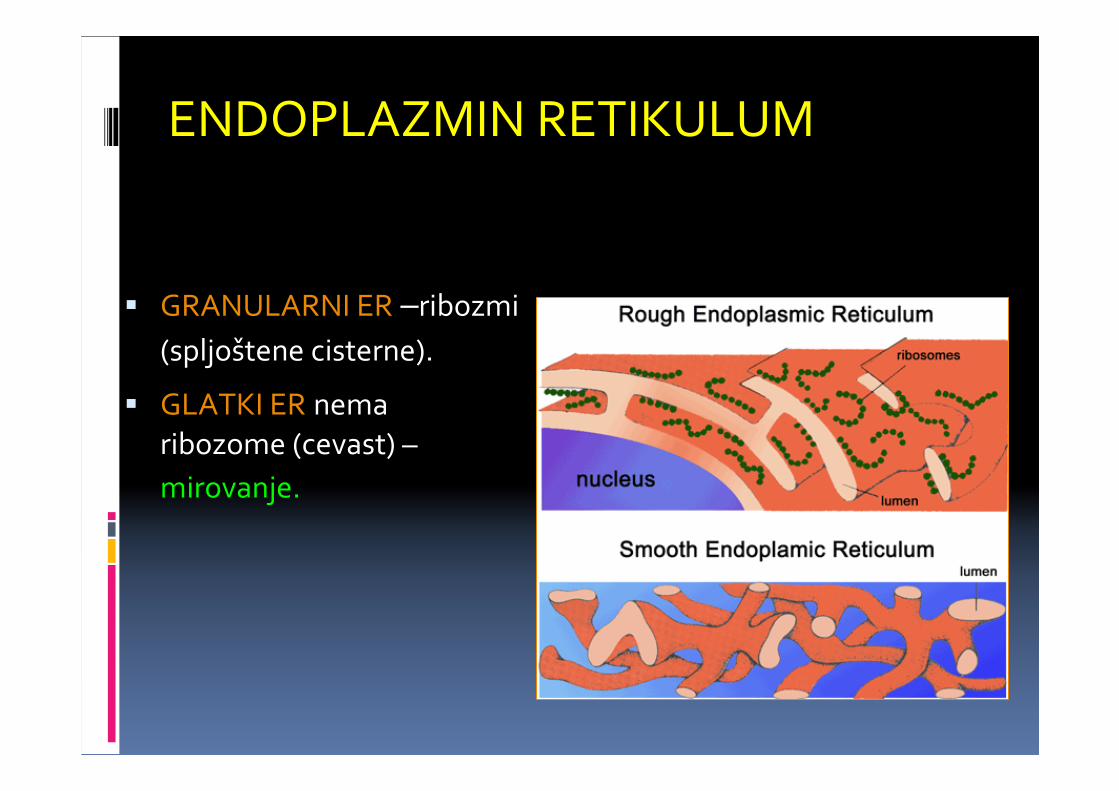
ENDOPLAZMIN RETIKULUM
� GRANULARNI ER –ribozmi
(spljoštene cisterne).
� GLATKI ER nemaribozome (cevast) –
mirovanje.

RIBOZOMI
� Nastaju u nukleolusu, subjedinice
� spajaju se u citoplazmi.
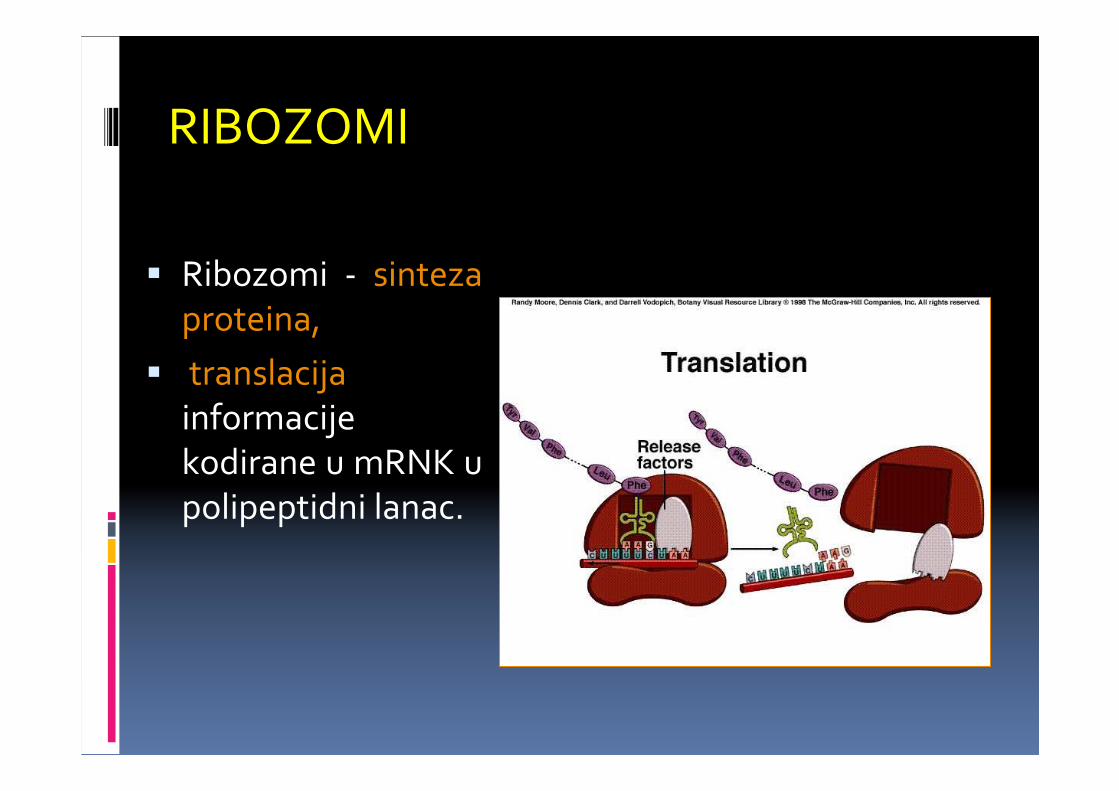
RIBOZOMI
� Ribozomi - sintezaproteina,
� translacijainformacijekodirane u mRNK u polipeptidni lanac.

RIBOZOMI
� Slobodni ribozomi - proteinipotrebni u citoplazmi,
� vezani za RER - proteiniugrađuju se u ćelijskemembrane ili eksportuju izćelije,
� ribozomi u mitohondrijama
i hloroplastima - interne specifične proteine.

GOLDŽIJEV KOMPLEKS
Spljoštene cisterneproširene na krajevima i vezikule.
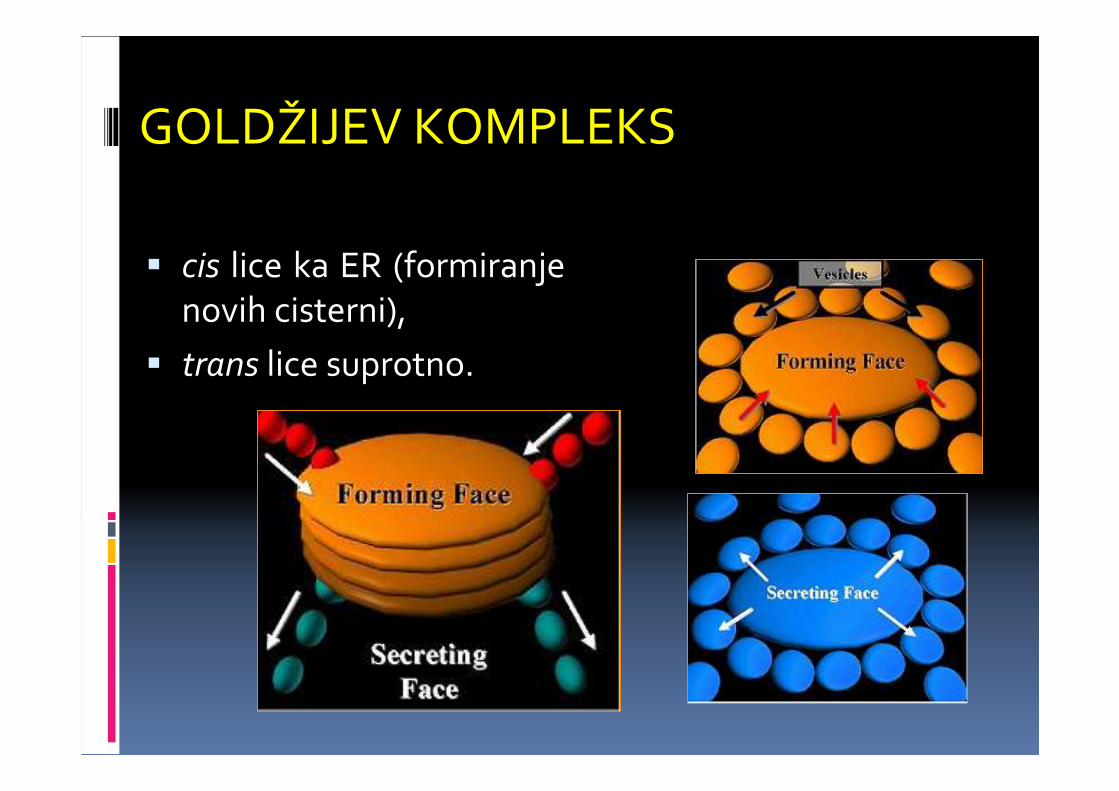
GOLDŽIJEV KOMPLEKS
� cis lice ka ER (formiranjenovih cisterni),
� trans lice suprotno.

GOLDŽIJEV KOMPLEKS
Sinteza,
polimerizacija ugljenihhidrata,
sekreciji sluzi,
transport
(glikoproteina,polisaharida).

MITOHONDRIJE
� Lipoproteini,� fosfolipidi,� RNK,� ribozomi 70-S, � cirkularna DNK.
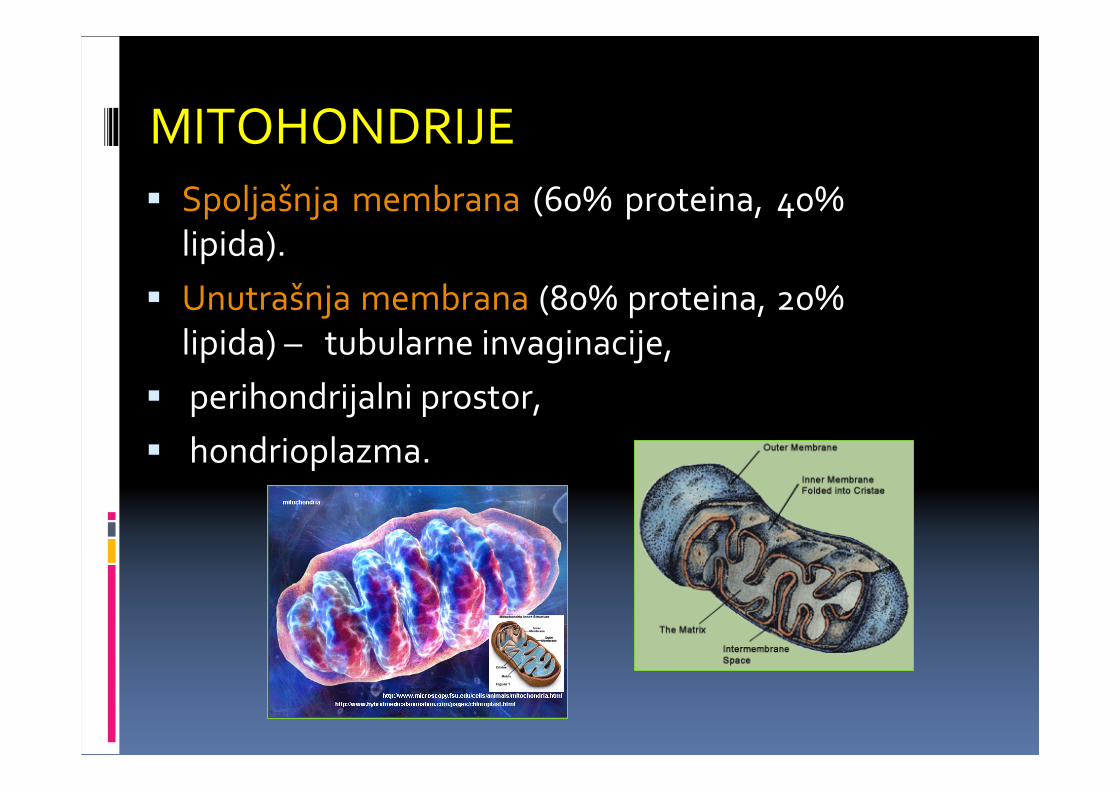
MITOHONDRIJE� Spoljašnja membrana (60% proteina, 40%
lipida).
� Unutrašnja membrana (80% proteina, 20% lipida) – tubularne invaginacije,
� perihondrijalni prostor,
� hondrioplazma.

MITOHONDRIJE
� Autonomne organele,
� autoreprodukcija –deoba (septe ili konstrikcija).
� Pupljenje.

MITOHONDRIJE
� Aerobna ćelijskarespiracija,
� (u prisustvu kiseonika –oksidativnafosforilacija),
konverzijakonverzija nutrijenatanutrijenatau ATPu ATP..

PLASTIDI
PROPLASTIDetioplast
hromoplasthloroplast leukoplast
amiloplast elioplast proteinoplast
statolit
PLASTIDI

HLOROPLASTI
spoljašnja i unutrašnja membrana hloroplasta
tilakoidistrome
stroma
tilakoidigrane
tilakoidistrome
stroma
granum
spoljašnja membrana hloroplasta
unutrašnja membrana hloroplasta
stroma
granum
tilakoidistrome
tilakoidi grane
intermembranski prostor
tilakoidistrome
tilakoidi granelumen tilakoida
granum
tilakoidi grane
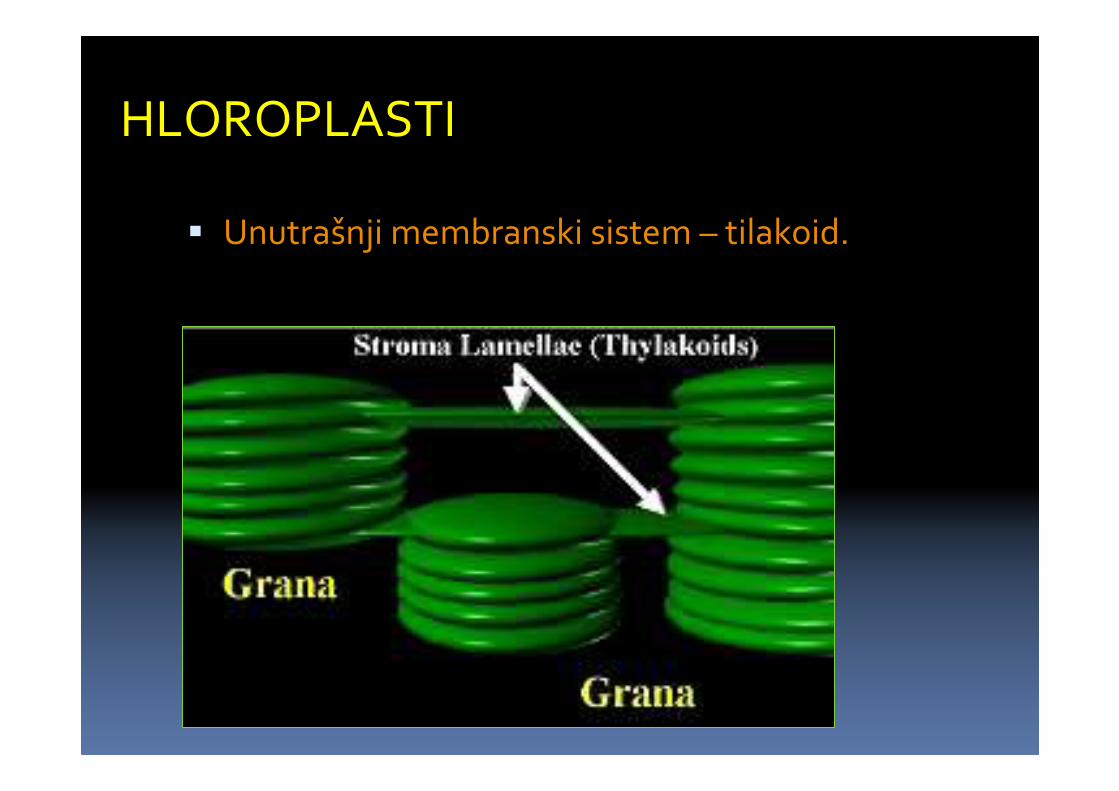
HLOROPLASTI
� Unutrašnji membranski sistem – tilakoid.

HLOROPLASTI
� Tilakoidi strome (duži) i tilakoidi grana (kraći)
� kraći naslagani tilakoidi obrazuju agregate –granum-e.
� Proteini 60%,
� lipidi 20%,
� hlorofil 4,3%,
� RNK 1,7-3,5%,
� DNK 0,5%,
� polisaharidi.

� Hlorofili pripadaju klasi jedinjenja poznati kao porfirini. To su zeleni pigmenti koji učestvuju u fotosintezi, a ulaze u sastav tilakoidnih membrana u hloroplastu.
� Za hlorofile je karakteristično da imaju atom magnezijuma u centru prstena, kao i peti ciklopentanonski prsten, koji se naknadno formira.

HLOROPLASTI
� Hlorofil apsorbuje svetlost, ekscitacija-svetlosna se pretvara u hemijsku energiju – za ugradnju i redukciju neorganskog ugljen dioksida - sinteza organskih molekula i oslobađanje kiseonika.
FOTOSINTEZA

PLASTIDI – etioplast
Klijanje semena u mraku, prolamelarno telose na svetlostirazvija u tilakoide.
LEUKOPLASTI
� Fotosintetički neaktivni, lamelarna građa,
� na svetlosti prelaze u hloroplaste.
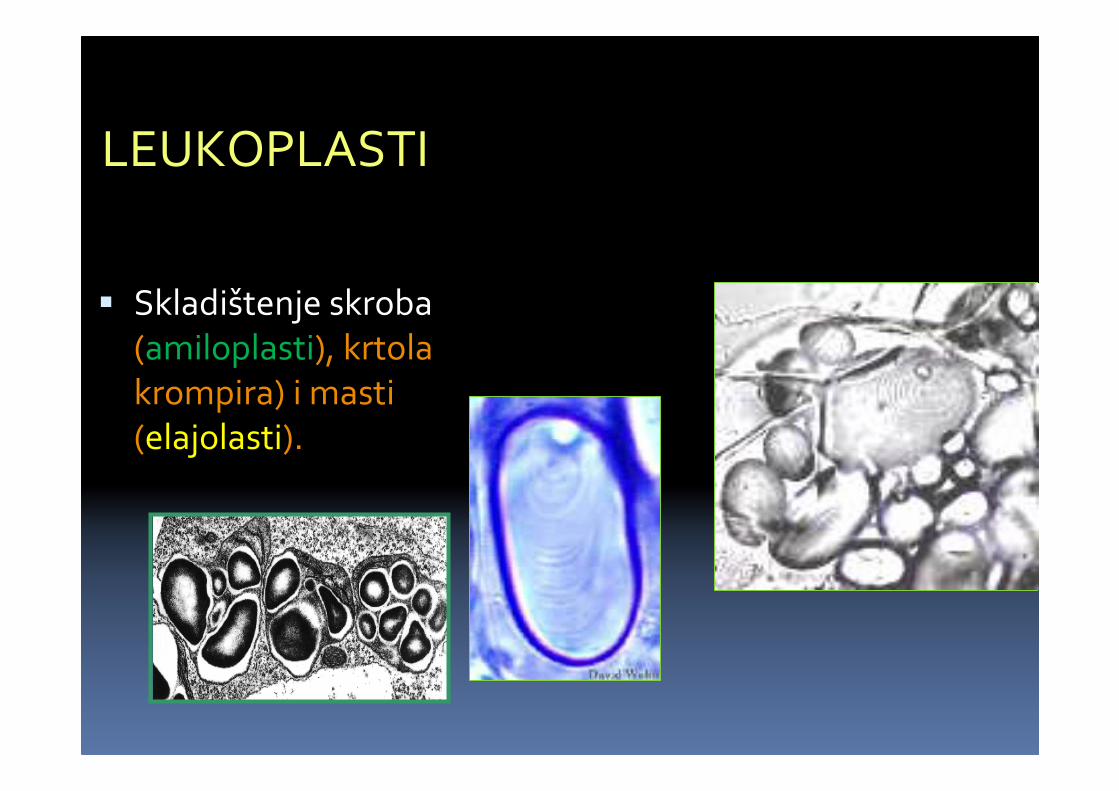
LEUKOPLASTI
� Skladištenje skroba(amiloplasti), krtola krompira) i masti (elajolasti).

ELAJOPLAST
Tilakoidi skoro nepostoje, lipidna telašcau stromi.

HROMOPLASTI
� Fotosintetički neaktivni,
� postaju od leukoplasta ili hloroplasta,
� karoteni, ksantofili, karotenoidi.

MIKROZOMI (MIKROTELA)
� Poreklo od cisterni Er-a,
� u blizini hloroplasta i mitohondrija.
� Peroksizomi i glioksizomi.

MIKROZOMI (MIKROTELA)
� Jednoslojnamembrana i enzimi.
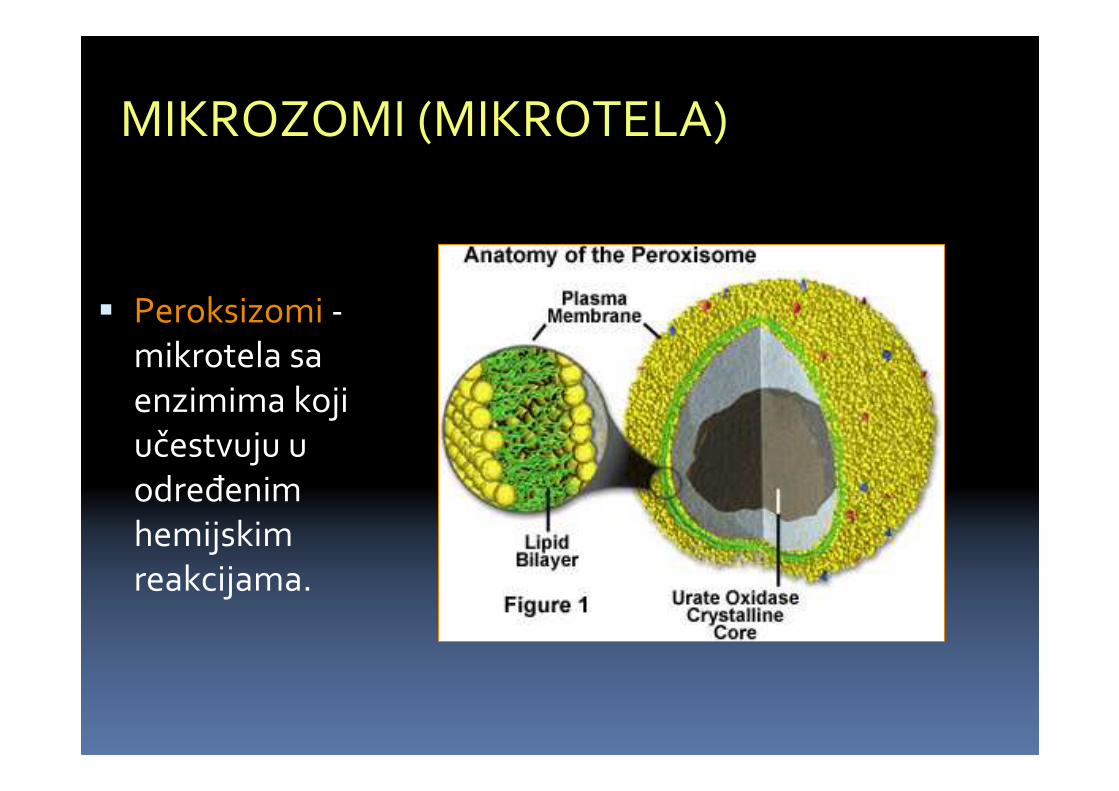
MIKROZOMI (MIKROTELA)
� Peroksizomi -mikrotela saenzimima kojiučestvuju u određenimhemijskimreakcijama.

MIKROZOMI (MIKROTELA)
� Glioksizomi – u semenima sarezervama masti, sadrže uratnuoksidazu i enzimeglioksilatnog ciklusa.– Konverzija masti u šećere,(glukoneogeneza) zarast embriona.

LIZOZOMI
� Sferična telašca obavijenajednoslojnomlipoproteinskommembranom stvaraju se ER-u.
� Hidrolitički enzimikatalizuju razgradnju svihosnovnih vrsta organskihmolekula.
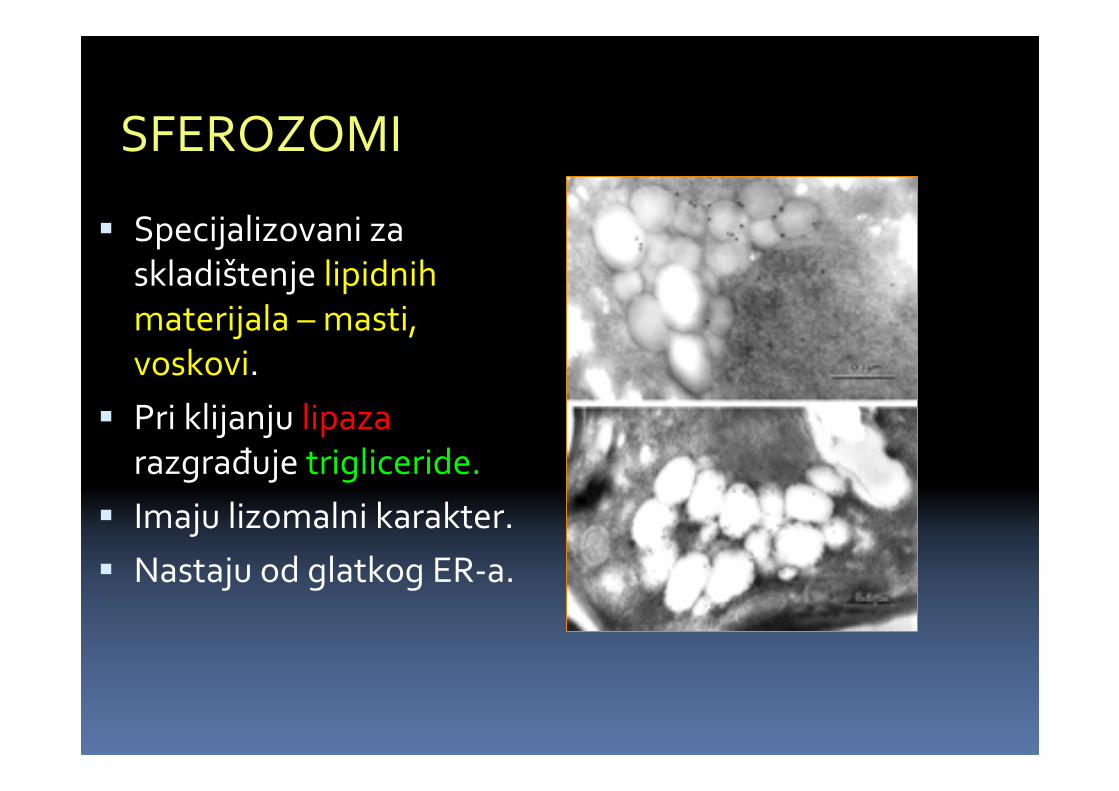
SFEROZOMI
� Specijalizovani zaskladištenje lipidnihmaterijala – masti, voskovi.
� Pri klijanju lipazarazgrađuje trigliceride.
� Imaju lizomalni karakter.
� Nastaju od glatkog ER-a.

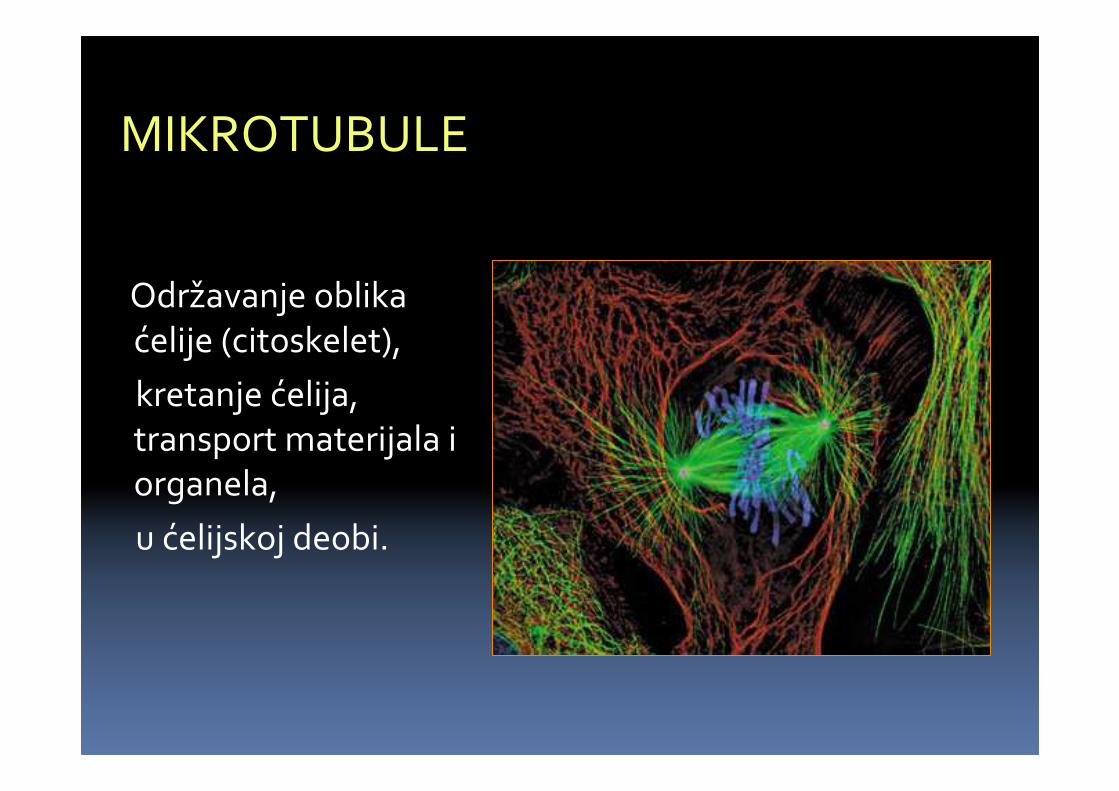
MIKROTUBULE
U aktivnimparenhimskim i meristemskimćelijama.
� Cilindričnestrukture u svimeukariotskimćelijama, sarazličitimfunkcijama.
Arabidopsis sp.

MIKROTUBULE
� Najbolje se uočavaju u deobnom vretenu – ( odmikrotubula i odgovarajućihproteina).
MIKROTUBULE
Održavanje oblikaćelije (citoskelet),
kretanje ćelija, transport materijala i organela,
u ćelijskoj deobi.

MIKROFILAMENTI
� Filament grade dva paralelnalanca koja se uvijaju.
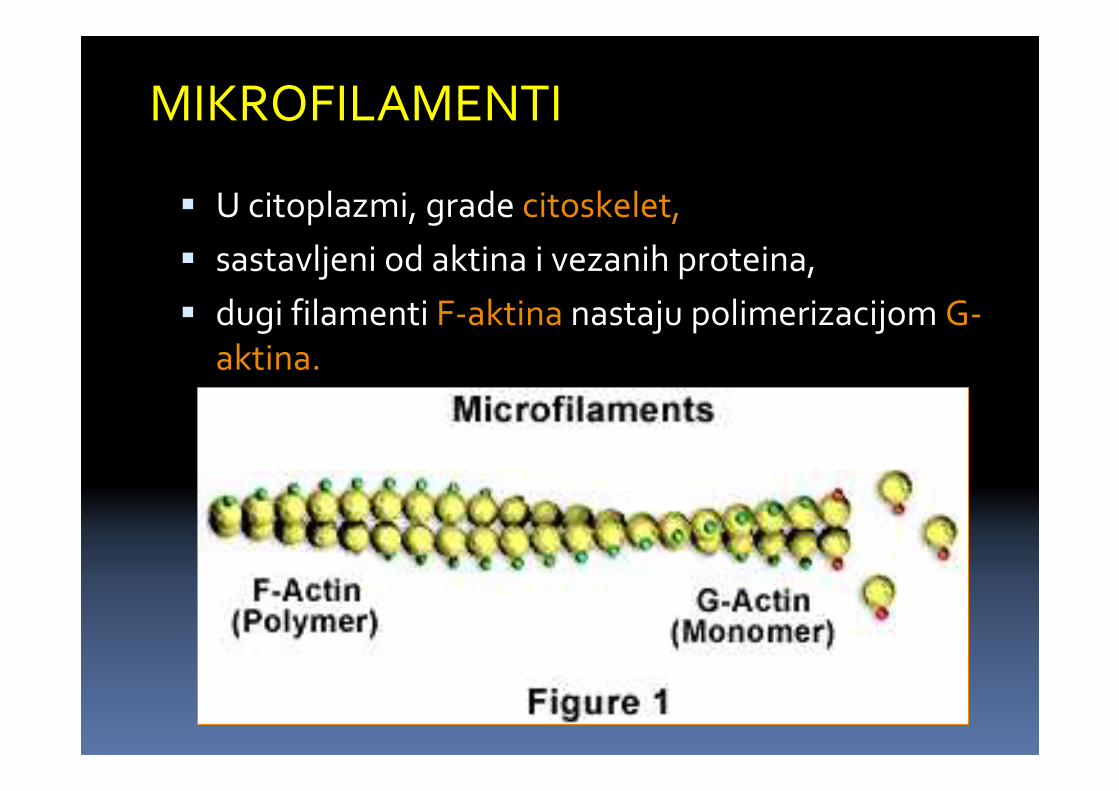
MIKROFILAMENTI
� U citoplazmi, grade citoskelet,
� sastavljeni od aktina i vezanih proteina,
� dugi filamenti F-aktina nastaju polimerizacijom G-aktina.
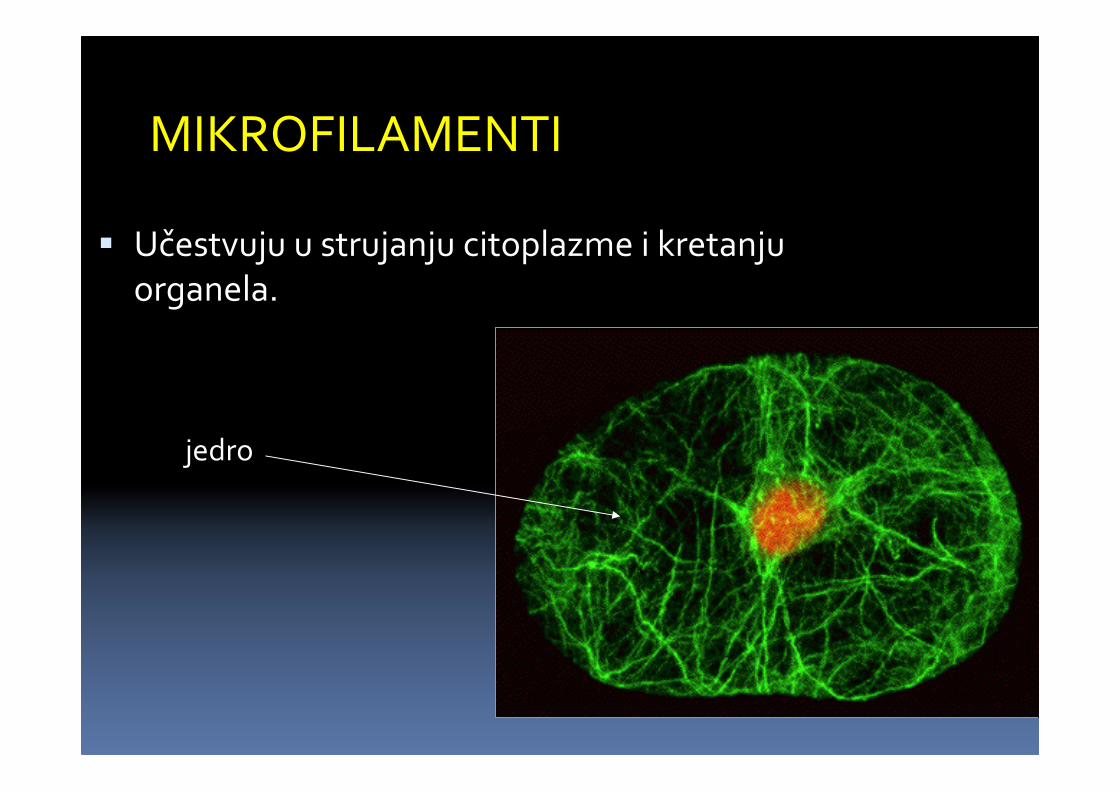
MIKROFILAMENTI
� Učestvuju u strujanju citoplazme i kretanjuorganela.
jedro

PROGRAMIRANA ĆELIJSKA SMRT
Stalan broj ćelija (ćelijska homeostaza) u organizmu održava se zahvaljujući dinamičkoj ravnoteži dva suprotna procesa: ćelijskoj deobi i ćelijskoj smrti.
Kao što svaka ćelija poseduje program nastanka odnosno ćelijsku deobu, tako poseduje i program ćelijskog umiranja.

PROGRAMIRANA ĆELIJSKA SMRT
Da bi se biljke pravilno razvijale i preživljavale, programirana ćelijska smrt je važan strateški odgovor na razne unutrašnje i spoljašnje signale, za pravilan razvoj organizma, formiranje specifičnih oblika organa, kao i za odstranjivanje oštećenih ili inficiranih ćelija.
Eukarioti imaju razvijene načine ćelijskog samoubistva koji je poznat kao programirana ćelijska smrt PCD.

PROGRAMIRANA ĆELIJSKA SMRT
Za razliku od životinja koje su u mogućnosti da eliminišu mrtvu ćeliju, kod biljaka to nije moguće zbog odsustva procesa fagocitoze.
Okidači kod većine biljaka za razvoj programirane smrti ćelije su hormoni i signali smrti koji se prenose preko hormonalnih signalnih puteva.
Razvojna smrt ćelije reguliše se unutrašnjim biološko- aktivnim molekulima, i sprovodi se uz pomoć genskih mehanizama signalne transdukcije.

PROGRAMIRANA ĆELIJSKA SMRT
Iako je u svim slučajevima ishod smrt ćelija, morfološke promene koje karakterišu pojedine etape ovih procesa međusobno se razlikuju i zavise od organa u kome se odigravaju.
U nekim slučajevima, prve promene zapažaju se u nivou jedra, u drugim na nivou plastida, ili endoplazminog retikuluma.

PROGRAMIRANA ĆELIJSKA SMRT
Samo dve familije Araceae i Aponogetonace
stvaraju perforacije putem PCD
Protoplast - model

PROGRAMIRANA ĆELIJSKA SMRT
U ksilemu vaskularnog tkiva, korenovoj kapi, tkivu reproduktivnih organa mogu se zapaziti neke karakteristične osobine programirane smrti ćelije.
Tokom ksilogeneze, procesa diferenciranja ksilemskog tkiva, trahejni elementi prolaze kroz nekoliko etapa koje su pod genetičkom kontrolom.
Na početku ksilogeneze meristemske ćelije koje prethode formiranim trahejnim elementima imaju jedro, jedarce, ER, GA, brojne male vakuole i primarni ćelijski zid.

PROGRAMIRANA ĆELIJSKA SMRT
Intenzivni sintetski procesi dovode do njihovog rastenja i izduživanja.
Potom jedra od ovalno-jajolikih postupno postaju relativno nepravilna sa istaknutim invaginacijam na površini.
Diktiozomi se premeštaju u periferni pojas citoplazme i postavljaju uz ćelijsku membranu.

PROGRAMIRANA ĆELIJSKA SMRT
Tokom i po formiranju segmenata sekundarnogćelijskog zida, u citoplazmi ovih ćelija uočavaju se degenerativne promene (odvajanje ribozoma od membrana ER, prskanje tonoplasta i oslobađanje hidrolitičkih enzima koji razlažu ER, GA, mitohondrije i nukleus).
Po okončanju ksilogeneze ostaju samo ćelijski zidovi koji predstavljaju trahejne elemente ksilemskih provodnih sudova.

PROGRAMIRANA ĆELIJSKA SMRT
Sezonske promene - opadanje listova i cvetova,sazrevnje i truljenje plodova primeri su genetički programiranih procesa ćelijskog umiranja, koji u prvim fazama podrazumevaju promene koje se mogu nazvati ćelijskim starenjem.

PROGRAMIRANA ĆELIJSKA SMRT
Prve morfološke promene se uočavaju na nivou plastida, promena orjentacije tilakoida grana i tilakoida strome.
Biohemijska ispitivanja su pokazala da hlorofil, azot i fosfor napuštaju tilakoidne membrane.
Membrana postaje nekontrolisano propustljiva tako da brojni proteini koji odlikuju stromu prelaze u citoplazmu.

PROGRAMIRANA ĆELIJSKA SMRT
Potom dolazi do degradacije lipida, proteina i nukleinskih kiselina. U citoplazmi se zapažaju promene u nivou ER, mitohondrija i jedra.
Tonoplast, koji biljnu ćeliju štiti od delovanja hidrolitičkih enzima prisutnih u vakuolarnom soku, poslednji podleže promenama, a oslobođeni enzimi razlažu ostatke organela.

PROGRAMIRANA ĆELIJSKA SMRT
Morfološke i funkcijske promene koje se tokom procesa starenja i umiranja uočavjau na nivou hloroplasta pokazuju određene sličnosti sa promenama koje odlikuju mitohondrije u ćelijama životinja tokom programiranog ćelijskog umiiranja.
To ukazuje da bi plastidi u biljnim ćelijama mezofila možda mogli imati analognu, regulatornu ulogu u okviru koje bi gubljenje integriteta membrana hloroplasta predstavljalo signal koji pokreće starenje.

PROGRAMIRANA ĆELIJSKA SMRT
Ćelijska smrt može nastupiti i kao rezultat mehaničkih oštećenja, a brojni su i primeri interakcije ostvarenih između patogenih organizama i biljnih ćelija praćeni umiranjem ćelija domaćina.
U mnogim od tih slučajeva ćelijska smrt se može smatrati mehanizmom za koji je data ćelija i tkivo kome pripada programirana i na taj način ona doprinosi odbrani jedinki.

PROGRAMIRANA ĆELIJSKA SMRT
Da bi se izborile sa DNK oštećenjem i oksidativnim stresom biljke su razvile visoko diferencirane popravke i mehanizme rekombinacije kao i redoks regulacije ćelijskog ciklusa.
Biljni ekstrakti - DNK

PROGRAMIRANA ĆELIJSKA SMRT
Pored strukturnih i funkcionlni sličnosti, biljne i životinjske ćelije razlikuju se po najosnovnijim odlukama koje preuzimaju, da li će da se podvrgnu deljenju, diferencijaciji, mirovanju, starenju,zaustavljanju ćelijskog ciklusa ili samoubistvu.

MUTACIJE
� Mutacije – promene u genetskom kodu.
� Podrazumeva zamene nukleotida, umetanja, izostavljanja, duplikacije i translokacije unutar gena, a takođe i krupne promene hromozoma, one su manje-više trajne promene u vrsti, broju, ili rasporedu nukleotida u genetskom materijalu.

MUTACIJE
� PROMENE U BROJU HROMOZOMA (numeričke aberacije), dva tipa:
� Euploidija i Aneuploidija.
� Euploidija – promne u broju celih garnitura hromozoma: monoploidije i poliploidije.
� Monoploidije – diploidan broj hromozoma na haploidan.
� Poliploidije – hromozomska garnitura se nalazi u višku: autopoliploidije (udvajaju se garniture iste vrste) i alopoliploidije (kombinacija genoma različitih ili srodnih vrsta).

MUTACIJE
� Kod biljaka poliploidija se izaziva hibridizacijom(lepši cvetovi, sočniji plodovi.. krompir, kafa, kikiriki).
� Aneuploidija – uvećanje ili smanjenje osnovne
garniture za jedan, ili veći broj hromozoma.

MUTACIJE
� Indukovane - izazvane mutacije – izlaganje ćelije nekom agensu, agens sa sposobnošću da izazove mutaciju je mutagen.
� Fizički agensi (UV zraci, X zraci, gama-zraci, α-čestice...), kratke talasne dužine prodiru duboko, promena genetičkog materijla.
� Hemijski agensi (vodonik-peroksid, azotasta kiselina, analozi baza, boje...), spečavaju replikaciju DNK.

MUTACIJE
� Upotreba indukovanih mutacija
� X- zraci: mutacija u ječmu (čvršća stabljika, gušćiklas, viši prinos),
� gama zraci: crvena pšenica - boja ćilbara, otpornost, patuljasta sorta pirinča,
� šećerna repa – uzgajanje sa 18 na 10 meseci.