Biológiai, Környezet- és Élelmiszertudományi Doktori
Bizottság
64
BIOKÉMIA
BEVEZETÉS
Eredetileg az élő szervezetekben előforduló szerves molekulák
vizsgálata a szerves kémia feladata volt. Később a
szénvegyületekkel kapcsolatos ismeretek olyan szövevényesek lettek,
és annyi részfeladat várt megoldásra, hogy az eredeti célkitűzés
feledésbe merült. Évtizedek múlva új tudományág alakult ki, amely
az élettelen természet, azon belül is a szerves kémia és az élő
természet, azaz a biológia között teremtett kapcsolatot. Ennek az
új tudománynak, a biokémiának lett feladata az élő szervezetben
előforduló biomolekulák vizsgálata, a lejátszódó vegyi folyamatok
feltárása, és az egyes folyamatok közötti bonyolult szabályozási
rendszer megismerése. A biokémia vizsgált folyamatai inkább a
szerves kémiával mutatnak hasonlóságot, módszereiben, amelyek
gyakran statisztikai módszerek, azonban inkább a biológiával rokon.
A biokémia azokat a lebontási és felépítési folyamatokat tárgyalja,
amelyek során az élő szervezetet alkotó biomolekulák állandóan
átalakulnak. Ezek az átalakulási folyamatok szolgáltatják azt az
energiát is, amelynek befektetése lehetővé teszi az energetikailag
instabil, a környezettel egyensúlyban nem lévő élő szervezetek
fennmaradását.
Az anyagcsere folyamatok, azaz a biomolekulák lebomlása és
felépülése az élő szervezet alapegységeiben, a sejtekben játszódnak
le. Minden élőlényre, sőt az egyes élőlények különböző szöveteire
más és más sejttípus jellemző, egyes speciális célsejtek
felépítésétől eltekintve mégis elmondhatjuk, hogy egy sejt
különböző sejtorganellumokat tartalmaz, amelyeket speciális
felépítésű hártyák, úgynevezett membránok választanak el egymástól.
A sejtfalak felépítésében is nagy szerepet játszanak a membránok. A
korábban bemutatott membránok speciális felépítésű, meghatározott
biomolekulákat tartalmazó szerkezeti elemek, amelyeknek nem csak az
egyes szerkezeti egységek elkülönítésében, hanem azok
kapcsolattartásában is jelentős szerepet játszanak. A sejtek
szerkezetét illetően az élő szervezetek két nagy csoportba
oszthatók, az alacsonyabb fejlettségi fokon álló, elkülönített
sejtmaggal nem rendelkező prokariótákra és a fejlettebb, sejtmaggal
rendelkező eukariótákra. A mikroorganizmusok a prokarióták közé
tartoznak. A prokarióták és eukarióták anyagcseréje egyes
folyamatok lefutásában, különösen a nukleinsav anyagcserében,
némileg eltér egymástól.
A sejt néhány jellemző szervezeti egysége külön említést
érdemel. A sejtet a citoplazma, másnéven citoszol tölti ki. Az
anyagcsere szempontjából igen fontos a mitokondrium, a sejtlégzés
szintere, amelynek kettős membránja van. A membránok közül a belső
nagyobb, ezért annak a mitokondrium belső tere, a mátrix tér felé
kitüremkedései vannak. A fotoszintetizáló sejtek szintesteinek, a
kloroplasztiszoknak felépítése némi rokonságot mutat a
mitokondriuméval. A sejt fontos része a riboszóma, amely a
fehérjeszintézis szintere. Az eukarótákban a sejtmag elkülönítve
tartalmazza az öröklődést biztosító dezoxiribonukleinsav (DNS)
tartalmat, a prokariótákban nincs ilyen elkülönülés.
Az élő szervezetben lejátszódó anyagcsere folyamatok kölcsönösen
visszahatnak egymásra, különböző szervezettségi szinten egymást
szabályozzák. E szabályozási rendszerben nagy szerepet játszanak
egyes, kiválasztott feladatot végző molekulák (ezek a hormonok),
amelyek az egyéb szabályozási mechanizmusokkal együtt lehetővé
teszik az élő szervezet harmónikus működését.
Az élő és élettelen világ közötti különbség nem csak a
molekulákat felépítő elemek minőségében és az egyes folyamatok
kölcsönhatásában van. Az élettelen világban vannak olyan szervezeti
egységek, ilyenek például a kőzetek, amelyek létezésének nincs
meghatározott funkciója. Az élő szervezetekben minden szervezeti
egységnek meg van a funkciója, a szabályozási mechanizmusok éppen
ezeket a funkciókat hangolják össze.
ANYAGCSERE FOLYAMATOK
Az anyagcserefolyamatok lebonyolítói, az enzimek
Az anyagcsere folyamatok energetikai háttere
Az élő szervezetben lejátszódó kémiai folyamatokban is
érvényesek a fizikai kémia törvényei. Az élő szervezetek
anyagcseréje, a metabolizmus folytonos kémiai átalakulások
sorozata. A lebontó (a katabolizmusban résztvevő) és
bioszintetikus, azaz felépítő (az anabolizmusban résztvevő)
folyamatok nem azonos úton és nem teljesen azonos enzimkészlettel
működnek, ez teszi lehetővé, hogy a környezettel egyszerű kémiai
egyensúlyban nem lévő élőlény állandóan fenntartsa szervezeti
állandóságát (homeosztázis). Az élő szervezet az egyensúly
szempontjából nyitott rendszer, állandósága valójában stacionárius
(steady state) állapot, ezért a környezettel állandó energia- és
anyag-kicserélődésben van. A felvett energia (autotróf
élőlényeknél, például a növényeknél az esetek túlnyomó többségében
a napenergia, a heterotróf élőlényeknél a tápanyagul szolgáló
biomolekulák lebontásából származó energia) fedezi az élő szervezet
fennmaradásának energiaszükségletét.
Az autotróf szervezetek az energián kívül a környezetből csak
vizet, szén-dioxidot és szervetlen sókat vesznek fel, testük
biomolekuláit maguk építik fel. A heterotróf élőlények
fennmaradásához szerves molekulák felvételére is szükség van,
ezeket esszenciális szerves molekuláknak nevezzük. Az élő
szervezeteket alkotó biomolekulák állandóan lebomlanak és helyükre
új biomolekulák szintetizálódnak. Minden szervezet valamennyi
sejtje, sőt valamennyi molekulája meghatározott periodicitással
kicserélődik. Az élő szervezet a környezettől felvett energia egy
részét makroerg kötések formájában tárolni képes.
Egyes energiaigényes szintézisek úgy mennek végbe, hogy a
reakcióval párhuzamosan egy vagy több makroerg kötés hidrolizálódik
és a felszabaduló energia fedezi a szintézis energiaigényét.
Ilyenkor a szubsztrát és a makroerg kötést viselő molekula
(általában ATP) közös, makroerg kötést tartalmazó intermediert
képez, ennek bomlása során nem csak a termék keletkezik, hanem az
ATP hidrolíziséből származó molekulák is megjelennek.
Az élet molekuláris szintű törvényei
Az anyagcsere folyamatok, közöttük az enzimek működése is, a
biomolekulák különböző kölcsönhatásain, illeszkedésén alapul. A
biomolekulák kölcsönhatásaival foglalkoznak az élet molekuláris
szintű törvényei.
A bioaffinitás törvénye szerint minden biomolekula legalább egy
másik biomolekulával olyan asszociátumot, komplexet tud képezni,
amelyben a komponensek aránya, illeszkedése és térbeli
elrendeződése pontosan definiálható. A bioaffinitás törvényének
tesz eleget az enzim-szubsztrát komplexek (ES) szerkezete is. A
komplexen belül a másodlagos kötések jelenléte dominál, de
előfordulnak másodlagosan kialakuló kovalens kötések is.
A biokatalízis törvénye az enzimek működésével foglalkozik. Az
enzimek is biomolekulák (mint korábban megállapítottuk, néhány
kivételtől eltekintve fehérjék), működésük reakció-specifikus (csak
adott reakciót katalizálnak), és szubsztrát-specifikus (csak egy
biomolekulát, esetleg annak néhány analógját képesek átalakítani),
fajtájuk a sejtben gyakorlatilag annyi, ahány fajta kémiai folyamat
a sejtben végbemegy. Ezért mondhatjuk azt, hogy a sejt
enzimkészlete megszabja a sejt valamennyi tulajdonságát.
A bioreguláció törvénye az enzimek katalizálta folyamatok
összehangolásának folyamatával foglalkozik. Az anyagcsere
folyamatokban meghatározó fontosságú enzimek (kulcsenzimek) és
bizonyos, úgynevezett szabályozó molekulák között a bioaffinitás
törvénye szerint kapcsolat jöhet létre. Ez a kötődés megváltoztatja
az enzim katalitikus aktivitását: aktivátor molekula esetében az
enzimműködés fokozódik, ha inhibítor (gátló) molekula kapcsolódik,
az enzim aktivitása lecsökken, esetleg teljesen megszünik. A
bioregulációt végrehajtó biomolekulák anyagcseréje ugyancsak
szabályozható enzimek segítségével bonyolódik. Ez a bonyolult
szabályozási rendszer hangolja össze az anyagcsere folyamatait,
teszi lehetővé az élőlény bizonyosfokú alkalmazkodását az egyre
változó környezet hatásaihoz.
ENZIMKATALIZÁLT REAKCIÓK
Az enzimek között egyszerű és összetett fehérjék is vannak,
gyakran több alegységet tartalmaznak, tehát negyedleges
szerkezettel bírnak. Általában globulárisak és felületüknek csak
egy kis hányada vesz aktívan részt az enzimreakcióban - ezt a részt
aktív helynek vagy aktív centrumnak nevezzük. A peptidlánc aktív
helyet alkotó aminosav egységei nem feltétlenül szomszédosak a
fehérjelánc szekvenciájában. A fehérje gombolyagban a primer
szerkezetben távolabb lévő aminosav egységek is térben közel
kerülhetnek egymáshoz, sőt másodlagos kötésekkel egymás mellé is
rögzülhetnek. A fehérje aktív helyen kívüli, tehát nagyobb része
elsősorban az aktív hely térszerkezetét (konformációját)
biztosítja.
Az aktív helyet felépítő aminosav egységek oldalláncai
különleges tulajdonságokat vehetnek fel, pédául ha a szerin
alkoholos hidroxilcsoportja egy térben közeli hisztidin egyik
gyűrűnitrogénjével hidrogén kötésbe léphet, ennek következtében
inkább alkoholát anionként reagál, savas tulajdonságúvá válik. Az
aktív hely működésében gyakran szerepet játszik a cisztein SH
csoportja is, amely sokszor kovalens kötést alakít ki a
szubsztráttal, ilyenkor kovalens katalízisről beszélünk.
Az aktív helynek két fontos pontja van. A szubsztrátkötő hely,
amelyhez a szubsztrát pontosan illeszkedik, és amelyhez az esetek
többségében másodrendű kötésekkel kapcsolódik (ez okozza az enzimek
szubsztrát-specificitását, és magyarázza azt, hogy az ES komplex
kialakulása kis aktiválási energiát igényel). A katalitikus hely,
amely az enzimhez kötött szubsztrát átalakítását végzi (ennek
működése felelős az enzim reakció-specifikus voltáért, és az ES
komplex továbbalakításának kis energiaigényéért). Más
reakciópartnert, például koenzimet is igénylő reakcióknál a koenzim
is az aktív helyhez kötődik a szubsztráthoz képest a reakció
szempontjából kedvező konformációban, általában ugyancsak
másodlagos kötésekkel.
A koenzim közreműködését igénylő, de történetesen azt nem
tartalmazó enzimet apoenzimnek, az enzim-koenzim komplexet
holoenzimnek nevezzük. Korábban már említettük, hogy amennyiben a
szükséges reakciópartner az enzimhez kovalensen kötődik, koenzim
helyett prosztetikus csoportról beszélünk. Az enzimek katalizálta
reakciókhoz egyes esetekben különleges funkciójú anyagok,
kofaktorok is szükségesek. A kofaktorok gyakran valamilyen
töltéssel rendelkező részecskék, például fém ionok, amelyek sokszor
az enzim aktív konformációját biztosítják, bár lehetséges, hogy
részt vesznek magában az enzimreakcióban is. Vannak azonban a
szubsztráthoz hasonló felépítésű kofaktorok is. Ez utóbbi típus
akkor szükséges, ha az enzimreakció célja a molekula egyik részének
áthelyezése, de a molekularészlet nem tud átugrani, ezért szükséges
a kofaktor, amelyben két ilyen molekularészlet van, az egyik az
eredeti, a másik az átalakulás utáni helyen. A kofaktor
közvetítésével sorozatos átadások után a szubsztrátból kofaktor
lesz, a kofaktorból termék, végül valamennyi szubsztrát molekula
átalakul egy kivételével, amelyből kofaktor lesz.
Az enzimes katalízis kinetikai vizsgálatával Michaelis és
Menten, később Briggs és Haldane foglalkoztak. Nevükhöz fűződik az
enzimreakciók egymást követő, több lépéses, azaz konszekutív
jellegének felismerése, és annak megállapítása, hogy míg az
enzim-szubsztrát (ES) komplex képződése egyensúlyi, megfordítható
(reverzibilis) folyamat, az enzim-szubsztrát komplex szétesése
termékké (product, P) és enzimmé gyakorlatilag megfordíthatatlan
(irreverzibilis). A reakciókinetika szabályainak értelmében a
reakció sebessége (v) arányos a reakciópartnerek koncentrációjának
szorzatával, arányossági tényező a sebességi állandó (k). Ennek
megfelelően
A reakció kezdetén a szubsztrát koncentrációja nagy, ezért a
sebesség (kezdeti sebesség, v0) nagy, idővel a szubsztrát
mennyisége csökken, végül elfogy, ezért a termék keletkezése lassan
megáll, a termék koncentrációja az idő függvényében tehát hiperbola
lefutású görbét ad.
Enzimkinetikai diagrammok
Ha a kezdeti sebességet a szubsztrát koncentráció függvényében
ábrázoljuk, ugyancsak hiperbola lefutású függvényt kapunk. A
kezdeti sebesség kezdetben egyenletesen (lineárisan) nő a
szubsztrát mennyiségének növelésével, mert újabb és újabb ES
komplexek keletkeznek. A szubsztrát koncentráció egy pontján már
valamennyi enzim molekula ES formában van (Et = ES), ennél tovább
emelve a szubsztrát koncentrációját a kezdeti sebesség növekedése
már egyre lassabb, egy határérték, a maximális sebesség (Vmax) felé
tart, amelyet elvben csak végtelenül nagy szubsztrát koncentráció
esetében érhet el. Az enzimreakcióra jellemző másik érték a
Michaelis állandó (KM), amely az a szubsztrát koncentráció,
amelynél a kezdeti sebesség az elérhető maximális sebesség fele (KM
= Vmax/2).
A valóságot legjobban megközelítő elképzelés szerint az
enzimreakcióban egy stacionárius állapot ("steady state") alakul
ki, az ES keletkezésének és elbomlásának sebessége közelítően
azonos, az ES komplex koncentrációja sokáig állandó marad
A v0 és a szubsztrát koncentráció összefüggését bemutató
összefüggés az enzimkinetika alapegyenlete, a Michaelis-Menten
egyenlet. Megjegyezzük, hogy Michaelis és Menten eredetileg egy
egyszerűbb modellre írta fel az összefüggést. Megközelítésükben az
ES komplex visszaalakulása enzimmé és szubsztráttá sokkal gyorsabb
volt, mint a termék kialakulása (k-1k2), ez a "gyors egyensúly"
("rapid equilibrium") modell, szemben a "steady state" modellel,
ahol k-1k2). Michaelis és Menten eredeti összefüggésében a
Michaelis állandó (KM) helyett KS, az ES komplex disszociációs
állandója szerepelt. A jelenlegi megközelítésben a "rapid
equilibrium" modell a "steady state" modell szélsőséges esete.
A KM és Vmax állandók kísérletes meghatározása nehéz. A fenti
egyenlet kettős reciprok ábrázolása (Lineweaver-Burk-féle lineáris
ábrázolás) segítségével azonban grafikusan könnyen
meghatározhatók.
E két állandó jól jellemzi az enzimreakció két egymást követő
lépését. A KM a szubsztrátkötő hely, a Vmax a katalitikus hely
állapotára vonatkozóan nyújt információt. Minden olyan behatás,
amely megzavarja az ES komplex kialakulását, tehát a szubsztrátkötő
hely működését, emeli a KM értékét. Minden olyan behatás, amely
megzavarja a katalitikus hely működését, csökkenti a Vmax értékét.
Ilyen módon az enzimreakció paramétereinek hatását kinetikai
mérésekkel vizsgálni lehet. Az enzimek működését fehérje jellegük
is jelentősen befolyásolja. Minden olyan behatás, amely
veszélyezteti az enzim natív (aktív) konformációját, csökkenti az
enzim aktivitását.
Az enzimaktivitás közvetlen, reverzibilis gátlása
Az enzimek katalitikus aktivitásának befolyásolásában nagy
szerepet játszanak különböző szerves molekulák (esetleg ionok),
amelyek az enzimek működését aktiválják vagy gátolják (aktivátorok
és inhibítorok). Az enzimaktivitást módosító hatások reverzibilisek
vagy irreverzibilisek lehetnek. Az anyagcsere szabályozása
szempontjából a reverzibilis gátlásoknak van nagyobb jelentősége. A
Michaelis-Menten kinetikájú enzimeknél három fontosabb gátlás
típust különböztetünk meg: a kompetitív, nemkompetitív és vegyes
gátlást. A három típus különböző mechanizmussal gátolja az enzimek
működését, ezért enzimkinetikai vizsgálatokkal, a KM, illetve Vmax
értékekre gyakorolt hatás alapján megkülönböztethető.
A kompetitív módon ható inhibítor molekulák szerkezete a
szubsztrátéval analóg. Két hasonló szerkezet verseng a
szubsztrátkötő helyért, az ES komplex mellett EI komplex is
keletkezik. Az inhibítorral képződött EI nem képes továbbalakulni.
Értelemszerűen a kompetitív gátlószerek jelenlétében az enzimekre
jellemző KM értéke megnő, a Vmax változatlan marad.
A nemkompetitív módon gátló molekula nem akadályozza meg az ES
komplex képződését, hatását a katalitikus helyen fejti ki,
megakadályozza az ES komplex tovább alakulását. Értelemszerűen
jelenlétében az enzimre jellemző Vmax értéke csökken, a KM
változatlan marad.
A vegyes típusú gátlószerek mind a szubsztrátkötő hely, mind a
katalitikus hely működését akadályozzák. Az inhibítor molekulák nem
csak a szabad enzimen képesek megkötődni, hanem - bár kisebb
affinitással - az ES komplexen is, sőt az EI komplex is képes a
szubsztrátot megkötni. Az ESI és EIS komplexek disszociációs
állandója nem azonos. Ilyen gátlószerek jelenlétében az enzimre
jellemző mindkét állandó értéke megváltozik, a KM nő, a Vmax pedig
csökken.
Az anyagcsere folyamatok szabályozása szempontjából kiemelkedő
fontossága van az allosztérikus szabályozásnak. Ilyenkor hosszabb
anyagcsere folyamatok végterméke a folyamat első enzimére
allosztérikus gátlószerként hat, ez megakadályozza, hogy a
szóbanforgó anyag feldúsuljon. Ezt a szabályozást negatív
visszacsatolásnak (feed back) nevezzük. Az allosztérikus gátlásra
alkalmas enzimek legalább két alegységet tartalmaznak, tehát
negyedleges szerkezettel rendelkeznek. A szabályozó anyag a
szabályozó (reguláló) centrumon kötödik meg, amely nem az aktív
centrumot tartalmazó, hanem a másik alegységben van.
Az allosztérikus szabályozás összehangolt (concerned)
mechanizmussal megy végbe, amelyet Monod tárt fel. Az alegységeknek
két lehetséges konformációja van, az aktív (lazított, relaxed - R)
és inaktív (feszített, tensed - T) konformáció. A két alegység
egyszerre csak azonos konformációban lehet: RR vagy TT. A két forma
egyensúlyban van egymással 1:10000 arányban a T forma javára. A
szubsztrát molekula az aktív helyet tartalmazó alegységnek csak R
konformációjú alakjával képes reagálni. Az allosztérikus
aktiválószer hatására a reguláló centrumot tartalmazó alegység R
konformációjú, így a másik, a katalitikus alegység is R
konformációjú lesz, az ES komplex kialakulása meggyorsul. Az
allosztérikus gátlószerek kötődésekor a reguláló centrumot
tartalmazó alegység a T konformációban rögzül, ez lehetetlenné
teszi, hogy a szubsztrát molekulák az ugyancsak T állású második
centrumon megkötődjenek. Például a szervezetben a maga ATP szint
meggátolja a glikolízis foszfofruktokináz enzimét.
A fentiek értelmében könnyen értelmezhető a hőmérséklet
befolyása az enzimek működésére. A reakciókinetika szabályai
szerint egy energia befektetését igénylő reakció hőmérsékletének 10
oC-kal való emelése a reakciósebességet megkétszerezi. Az enzimek
viszont fehérjék, ezért a hőmérséklet emelése fokozza a denaturáció
valószínüségét. A hőmérséklet emelése ezért két folyamat, egy
aktiváló és egy dezaktiváló folyamat összegződése miatt a kezdeti
sebesség változásában optimumot eredményez. Ez az optimum
látszólagos, mert két ellentétes folyamat eredője, ezért minél
rövidebb ideig tesszük ki az enzimet a hőhatásnak, az optimum annál
magasabb hőmérséklet felé tolód
Az enzimreakciók pH függésének értelmezéséhez jobban meg kell
ismernünk az aktív centrum szubsztrátkötő helyének és katalitikus
helyének szerkezetét. E kitüntetett helyekre jellemző, hogy
működésükben két-két, protont leadni és felvenni képes, általában
savas (csak néhány esetben bázikus) csoport játszik szerepet, ezek
egyike az enzim működése közben disszociált állapotban van, ez a
savasabb, pK értéke kisebb, a másik csoport pedig disszociálatlan
állapotú, ez a kevésbé savas csoport, pK értéke magasabb.
Matematikai összefüggések nélkül is belátható az, hogy olyan erősen
savas körülmények között, amikor a savasabb csoport disszociációja
is visszaszorul, illetve olyan erősen bázikus körülmények között,
amikor a gyengébben savas csoport is disszociálódik, sem a
szubsztrátkötő hely, sem a katalitikus hely nem képes dolgozni.
Az enzimek osztályozása
Az enzimek osztályokba sorolását a hetvenes években az
International Union of Biochemistry (IUB) nemzetközi bizottsága
fogadta el, és látta el az egyes osztályokat sorszámokkal. Az
enzimeket az enzim katalógus (EC) szerint négyjegyű számmal
jellemezzük: az első szám az enzim osztályt, a második az
alosztályt, a harmadik az alosztályon belüli csoportosítást, a
negyedik szám ezen a kisebb alegységen belül az enzim egyéni számát
jelenti.
Az első osztályba az oxidoreduktázok tartoznak, amelyek a
szubsztrát molekulákat oxidálják, illetve redukálják (közben egy
másik molekula - a koenzim -redukálódik, illetve oxidálódik). A
reakció kémiailag cserebomlás, a hidrogének vagy elektronok
áthelyezése. A reakció mechanizmusának megfelelően változik a
közreműködő koenzim, amelynek típusait a koenzimeknél tárgyaljuk.
Az enzim elnevezése mindig a "dehidrogenáz" irányból történik,
mivel a reakciók megfordíthatók.
A második osztályba a transzferázok tartoznak, amelyek
szubsztitúciós rekkciókat katalizálnak. Gyakori egy foszfátcsoport
áthelyezése egy hidroxilcsoportra. Ha a foszfátcsoport ATP
molekulából származik, akkor az enzimet kináznak nevezzük. A
továbbítandó csoport lehet acilcsoport, egy vagy két szénatomot
tartalmazó, úgynevezett C1 vagy C2 komponens (fragmens), illetve
két vagy három szénatomos cukor részlet is. Ez utóbbi reakciókat a
transzketolázok, illetve transzaldolázok katalizálják.
A harmadik osztályba a hidrolázok tartoznak. Közreműködésükkel a
szubsztrát molekula úgy hasad el víz hatására, hogy a víz
alkotórészei, a proton és a hidroxid anion a kialakuló két rész
végére hidrogénként és hidroxilcsoportként épülnek be.
A negyedik osztályba tartozó liázok (szintázok) addíciós és
eliminációs reakciókat katalizálnak.
Az ötödik osztályba tartozó izomerázok a szubsztrát molekula
izomer átalakulását katalizálják tautomerizáció segítségével
A hatodik osztályba tartozó ligázok (szintetázok) nagy pozitív
szabad entalpia változással járó reakciók végbemenetelét teszik
lehetővé azzal, hogy egyidejűleg egy ATP vagy más NTP egyik vagy
mindkét makroerg foszforsavanhidrid kötése hidrolitikusan felhasad,
a felszabaduló energia fedezi az irreverzibilis, endoterm reakció
energiaszükségletét.
Az enzimek működését az enzim aktivitásával jellemezzük. Az
aktivitás mérésénél azt határozzuk meg, hogy időegység alatt az
enzim egységnyi mennyisége (1 mg fehérje vagy 1 ml enzim oldat)
mennyi szubsztrátot alakít át. A gyakorlatban két egységet
különböztetnek meg. A standard enzim-egység (standard unit, SU) a
szubsztrát mennyiségét molban adja meg, időegységnek percet számít.
Az SI rendszer a katal egységet ismeri el, ez az átalakult
szubsztrát mennyiségét molban, az időt másodpercben adja meg. Jól
jellemzi az enzim aktivitását az átviteli szám is, amely az enzim
molekula által 1 másodperc alatt átalakított szubsztrát molekulák
számát adja meg.
Membrántranszport folyamatok
A membránok az élő szervezetek sejtjeit vagy sejtrészleteit
(organellumait) úgy választják el egymástól, hogy a membránba
ékelődött és azon áthatoló (integráns) fehérjék útján bizonyos
korlátozott kapcsolatot is lehetővé tesznek. A membránok, amelyek
lipideken és fehérjéken kívül szénhidrátokat is tartalmazhatnak
kötött állapotban, jelentős szerepet játszanak a biokémiai
(anyagcsere) folyamatokban. A felületaktív vegyületek irányított
adszorpciójának segítségével felépülő membránok belső, hidrofób
részében apoláris molekulák, például apoláris (zsírokban oldódó)
vitaminok tárolhatók. A lecitinek az állati sejtek membránjának
felépítésében vesznek részt, a növényekben ezt a feladatok
felületaktív összetett fehérjék végzik.
A membránokon keresztül csak kicsi, valamint nem nagyméretű,
erősen apoláris molekulák képesek akadály nélkül áthaladni. Más
molekulák áthaladását az áthatoló (intergráns) fehérjék segítik
elő. Ennek különböző módozatai vannak.
Az oldott anyagok a membránon keresztül hordozó (karrier)
molekulák segítségével vagy anélkül juthatnak át, az esetek
túlnyomó többségében a passzív diffúzió szabályai szerint, tehát a
magasabb koncentrációjú hely irányából a hígabb oldat felé.
A hordozóval végrehajtott passzív diffúzió legegyszerűbb formája
a könnyített (facilitált) diffúzió, amelyben a hordozó megkönnyíti
a szubsztrát átjutását a koncentrációesés irányába.
Az egyirányú egyidejű transzport (szimport, kotranszport)
folyamatában a hordozó membrán-fehérje azonos oldalon egyszerre
kétféle szubsztrátot köt meg és továbbít a membrán túloldalára,
például a nátrium ion és a glükóz így jut be egyidőben a sejtbe. A
passzív diffúzió szabályai ekkor is érvényben vannak, de a két
anyag koncentrációjának összege a mérvadó - ez esetben a sejten
belüli alacsony Na+ koncentráció teszi lehetővé, hogy a glükóz a
magasabb belső koncentráció ellenére is bejusson.
Az ellentétes irányú egyidejű transzport (antiport) is passzív
transzport, esetében a két szubsztrát egyszerre kötődik meg a
membrán két oldalán, majd a fehérje konformáció változása
következtében a két anyag helyet cserél. Az antiport folyamatokat
kivitelező karrier fehérjéket transzlokázoknak nevezzük.
Míg a karrierrel végrehajtott membrán-transzport folyamatok
külön energia befektetése nélkül csak a passzív diffúzió szabályai
szerint játszódhatnak le, az aktív transzportban a szubsztrát
átvitele koncentráció gradiens ellenében megy végbe, ez energia
befektetését igényli. Ebben az esetben többnyire antiport játszódik
le, amelyet egy makroerg kötés hidrolízisének hatására végbementő
konformációváltozás idéz elő.
Az anyagcserefolymatok fajtái
Az anyagcsere folyamatok (metabolizmus) során az élő szervezetek
biomolekulái, illetve a szervezetbe kerülő tápanyagok molekulái a
levegő oxigénjének felhasználásával oxidatív úton lebomlanak
(katabolizmus), ennek során egyrészt energia szabadul fel, amely az
előző fejezetben ismertetett módon az élő szervezetnek, ezek közül
is elsősorban a heterotróf élőlényeknek, mint energetikailag
nyitott rendszereknek fenntartásához szükséges, másrészt
keletkeznek azok az intermedierek, amelyek a szervezet anyagainak
felépítéséhez, a bioszintetikus folyamatokhoz (anabolizmus)
szükségesek. A lebontó folyamatok legvégén a biomolekulák
végeredményben szén-dioxiddá és vízzé oxidálódnak ("égnek el").
Az autotróf szervezetekben (pl. növények) a szervezet
fenntartásához szükséges energiát a fényenergia szolgáltatja (bár
vannak kémiai energiát felhasználó mikroorganizmusok is), a
felépítő folyamatok egyrészt a fotoszintézis (széndioxid és víz
beépítés), másrészt ammónia beépítés segítségével mennek végbe.
Egyes mikroorganizmusoknál a nitrogén forrás a nitrogén gáz, mások
lebontó folyamataikhoz nem használnak fel oxigént, hanem
erjesztéses folyamatokkal működnek.
Az anyagcsere folyamatok vázlata nem tudja érzékeltetni azt a
sokrétegű szabályozási hálózatot, amely az anyagcsere folyamatokat
összehangolja, így teszi lehetővé az élő szervezet harmónikus
működését.
Lebontó folyamatok, katabolizmus
Az anyagcsere folyamatok tárgyalását a lebontási folyamatokkal
kezdjük. Először végig kísérjük a tápanyagok (szénhidrátok, lipidek
és fehérjék) lebomlását addig az intermedierig (acetil-koenzim-A),
amely a lebontó folyamatok közös, befejező szakaszához vezet el,
majd részleteiben vizsgáljuk a közös lebontó folyamatokat (a
citrátkört és a terminális oxidációt). Ezután áttekintjük e három
biomolekula típus bioszintézisét, végül a fehérjeszintézissel
kapcsolatban foglalkozunk a nukleinsavak anyagcseréjével is.
Az élő szervezetbe kerülő tápanyagok, valamint a nukleinsavak
olyan összetett molekulák (glikozidok, savamidok és észterek),
amelyekből hidrolízissel szabadulnak fel azok az egyszerű
biomolekulák (cukrok, aminosavak, zsírsavak stb.), amelyek az
oxidatív lebontási folyamatok kiindulási anyagai. E biopolimerek
lebontását hidroláz enzimek végzik. A hidrolázoknak két alapvető
típusát különböztetjük meg. Az exo-hidrolázok a hidrolízist a
polimerlánc végén kezdik meg, onnan egy vagy két egységet
tartalmazó részeket hasítanak le. Képesek a láncvégeket
megkülönböztetni, szükség esetén specifikusak a kötések térállására
és a kötésben részt vevő komponensek minőségére is. Az
endo-hidrolázok a lánc közben hasítanak fel kötéseket. Olyan
biopolimerekben, amelyekben a felépítő egységek egymástól
különböznek (pl. fehérjék), meghatározott egységek közötti
kötéseket hidrolizálnak. Az azonos (legfeljebb néhány féle)
egységekből felépülő biopolimerekben az endo-hidrolázok
véletlenszerű (random) helyeken hasítanak.
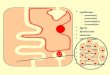
A mitokondrium felépítése
A biomolekulák hidrolízise, majd a hidrolizált termékek oxidatív
lebomlásának első lépései a citoszolban mennek végbe. A további,
oxidatív lebontó folyamatok szintere a mitokondrium, amelynek
kettős membránja van. A membránok közül a belső memrán felülete
nagyobb, ezért annak a mitokondrium belső tere, a mátrix tér felé
kitüremkedései vannak, ezeket krisztáknak nevezik. A külső membrán
és a belső membrán közötti teret intermembrán (közti) térnek
nevezik. A mitokondrium membránjain az anyagok a membrántranszport
szabályainak megfelelően juthatnak át, bár a külső membránon egyes
kis molekulák könnyen átjuthatnak.. Egyes esetekben nem csak a
koncentráció viszonyok a meghatározók, az áthaladás csak bizonyos
molekulák számára megengedett. Ezek a szabályok a belső membrán
esetében szigorúbbak, mint a külső membránnál. A további, oxidatív
lebontó folyamatok a mátrix térben zajlanak le, kivéve a végső
oxidációt, a terminális oxidációt, amelynek a szintere a
mitokondrium belső membránja.
A mitokondrium membránjai
A szénhidrátok lebontásának egyéni szakasza az
acetil-koenzim-A-ig
A poliszacharidok glikozidos kötéseit hidrolizáló enzimeket
glikozidázoknak nevezzük. Az exo-glikozidázok általában a nem
redukáló láncvégről kezdik a hasítást. A keményítő glikozidázai: az
α-amiláz endo-glíkozidáz, a β-amiláz exo-glikozidáz, a nem redukáló
láncvégről maltóz egységeket hasít le. A maltózt a maltáz bontja
két glükózra. A cellulózt bontó cellulázokat csak egyes
mikroorganizmusok termelik. A cellobiózt a cellobiáz, a tejcukrot a
β-galaktozidáz, a szacharózt az invertáz hidrolizálja.
Az élő szervezetekben lezajló szénhidrát anyagcserének
kulcsvegyülete a glükóz, amelyből első lépésben glükóz-6-foszfát,
majd belőle fruktóz-6-foszfát keletkezik. Egyes poliszacharid bontó
enzimek, a foszforilázok meghatározott poliszacharidokról (például
a glikogénről) egy molekula szervetlen foszfát segítségével
közvetlenül glükóz-1-foszfátot hasítanak le, amely a
glükóz-foszfát-mutáz (transzferáz) enzim segítségével, a
glükóz-1,6-biszfoszfát kofaktor közreműködésével alakul
glükóz-6-foszfáttá.
Mivel a szénhidrátok elsősorban a glükózon keresztül bomlanak
le, a tárpanyagban előforduló egyéb cukrok valamilyen származékuk,
gyakran nukleozid-difoszfáttal képzett, aktivált formájukban
alakulnak a megfelelő glükóz-származékká, ilyan például a galaktóz.
A cukrok glikozidos hidroxilcsoportjainak foszfát-észtereiből
nukleozid-trifoszfátokkal képzett, makroerg savanhidrid kötést
tartalmazó NDP-cukor származékainak a poliszacharidok
bioszintézisénél van nagy jelentősége. E származékok keletkezésekor
a kihasadó pirofoszfát hidrolízisekor felszabaduló energia fedezi a
reakció energiaigényét. A mannóz-6-foszfát a
mannóz-foszfát-izomeráz segítségével reverzibilisen képes
glükóz-6-foszfáttá alakulni. Az aszkorbinsav (C-vitamin) a
glükuronsavból keletkezik több lépésben.
Glikolízis
A glükóz anaerob (levegő távollétében lezajló) és aerob (a
levegő oxigénjét felhasználó) lebomlásának első, a piruvátig tartó
közös, a citoplazmában lejátszódó szakaszát glikolízisnek nevezzük.
Megjegyezzük, hogy a legtöbb biokémiai szakkönyv a glükóz teljes
anaerob lebontását hívja glikolízisnek, de a folyamatot ezek is
csak a piruvátig tárgyalják érdemben. Véleményünk szerint a piruvát
az anyagcsere folyamatok egyik fontos elágazási pontja, ezért
indokolt a keletkezését szolgáló folyamatot külön egységnek
tekinteni.
A glikolízis első, hexokináz (transzferáz) katalizálta lépését,
amelyben a glükózon egy ATP foszfát-észter kötést alakít ki és
glükóz-6-foszfát keletkezik, sokan a glükóz aktiválásának tekintik.
A megfordíthatatlan reakcióban keletkező glükóz-6-foszfát nem
tartalmaz makroerg kötést vagy nagyon könnyen reagáló csoportot,
ezért nem tekinthető "aktív" származéknak. Kétségtelen azonban,
hogy a molekula poláris jellege a foszforilálással nő, ionos
másodlagos kötésekkel enzimekkel könnyebben képez enzim-szubsztrát
komplexet. A hexokináznak egyéb hexózok, így a fruktóz is
szubsztrátja lehet, de a fruktóz affinitása az enzimhez a
glükózénak csak töredéke, ezért a fruktóz lebomlása során csak
később kapcsolódik be a glikolízisbe.
A glükóz-6-foszfátból a glükóz-foszfát-izomeráz (régebbi nevén
foszfo-glükóz-izomeráz) hatására kettős tautomer átrendeződéssel
fruktóz-6-foszfát keletkezik, amelynek 1-es szénatomján, a primer
hidroxilcsoporton ismét egy ATP alakít ki foszfát-észter kötést,
ugyancsak megfordíthatatlan reakcióban a foszfo-frukto-kináz (PFK)
enzim (ezt is helyesebb lenne fruktóz-foszfát-kináznak nevezni)
hatására. A PFK a glikolízis sebesség meghatározó (kulcs) enzime,
fruktóz-2,6-biszfoszfáttal jelentős mértékben aktiválható, ezen
kívül számos anyagcsere termék befolyásolja aktivitását (pl. ATP,
AMP, citrát stb). A glikolízis hat szénatomos (C6) szakaszának
utolsó lépésében az aldoláz (liáz) a fruktóz-1,6-biszfoszfátot két
trióz-3-foszfáttá, dihidroxi-aceton-foszfáttá (DHAP) és
glicerinaldehid-3-foszfáttá hasítja (a kialkuló kettős kötés az
utóbbi karbonilcsoportjában van).
A glikolízis hat szénatomos (C6) szakasza
A két trióz-foszfát a trióz-foszfát-izomeráz segítségével
egyensúlyi elegyet alkot. A glicerinaldehid-3-foszfát a
glicerinaldehid-3-foszfát-dehidrogenáz (oxidoreduktáz) hatására egy
szervetlen foszfát beépítésével a makroerg kötést tartalmazó
glicerinsav-1,3-biszfoszfáttá oxidálódik, a folyamat NAD koenzimmel
működik, tehát (NADH+H) keletkezik. Ez a glikolízis egyik energia
termelő lépése.
A glikolízis három szénatomos (C3) szakasza és a piruvát
továbbalakulási lehetőségei
A glicerinsav-1,3-biszfoszfát makroerg kötésének energiája a
glicerát-foszfát-kináz (transzferáz) hatására egy ADP-ATP
átalakulásba épül be. A glicerinsav-3-foszfátból a
glicerinsav-2,3-biszfoszfát kofaktor közreműködésével a
glicerát-foszfát-mutáz (transzferáz) hatására glicerinsav-2-foszfát
keletkezik, amely az enoláz (liáz) hatására vizet veszít és a
glikolízis második energia termelő lépésében a makroerg enol-észter
kötést tartalmazó foszfoenolpiruvát (PEP) keletkezik.
A glicerát-foszfát-mutáz katalizálta reakció
A glikolízis három szénatomos (C3) szakaszának utolsó lépésében,
energetikailag megfordíthatatlan folyamatban a piruvát-kináz
(transzferáz) hatására a makroerg kötés energiája egy ADP-ATP
átalakulásba épül be, így keletkezik a piruvát. Megjegyezzük, hogy
a glikolízis enzimei és intermedierjei régóta ismertek, több nevük
is használatban van, a karboxilát-foszfát nevek helyett a
foszfo-karboxilát nevek is legalább annyira elterjedtek.
A glükóz anaerob (oxigént nem igénylő) lebomlásának további
lépései is a citoplazmában játszódnak le. A tejsavas erjedésben a
piruvát a glikolízisben keletkező redukált koenzimet felhasználva
L-tejsavvá (laktáttá) redukálódik a laktát-dehidrogenáz
(oxidoreduktáz) hatására. Így keletkezik az izommunka kezdeti
szakaszában tejsav.
Az alkoholos erjedésben a piruvát a piruvát-dekarboxiláz
hatására acetaldehiddé alakul, a transzfer koenzim, mint a
dekarboxilázoknál általában, a TPP. Az acetaldehid a glikolízisben
keletkező redukált koenzim felhasználásával, az
alkohol-dehidrogenáz hatására etilalkohollá (etanollá) redukálódik.
A szeszes italok alkoholja ezen folyamat megfordításával bomlik
le.
A borhamisítási botrányok hívták fel a figyelmet az egyébként
fagyállóként közismert etilén-glikol mérgező tulajdonságaira. Az
etilén-glikol a szervezetben az alkohol-dehidrogenáz hatására
először glikolaldehiddé, majd további enzimreakciókban
végeredményben a mérgező és vesekárosító oxálsavvá oxidálódik.
Az etilén-glikol mérgezésben igen hatásos ellenszer az etanol,
mert az alkohol-dehidrogenáz kompetitív gátlószere. Mivel a
rendkívül mérgező metanol (faszesz) is analóg úton oxidálódik
formaldehiddé, majd hangyasavvá, az etanol itt is ellenszer
lehet.
Egyéb egyszerű cukrok bekapcsolódása a lebontási
folyamatokba
A fruktóz lebontása
Mint említettük, a fruktóz affinitása a hexokinázhoz kicsi,
ezért lebomlása kezdetben eltér a glükózétól. Először a fruktokináz
(transzferáz) hatására egy ATP fruktóz-1-foszfátot alakít ki, amely
a frukóz-1-foszfát-aldoláz (liáz) hatására DHAP-tá és
glicerinaldehiddé hasad szét. A DHAP bekapcsolódik a glikolízisbe.
A glicerinaldehidet a trióz-kináz (transzferáz) egy ATP
segítségével glicerinaldehid-3-foszfáttá foszforilálja, amely
ugyancsak bekapcsolódhat a glikolízisbe. Alacsonyabbrendű
élőlényekben a glükóz és fruktóz reverzibilisen egymásba
alakítható.
Mivel a szénhidrátok elsősorban a glükózon keresztül bomlanak
le, a tárpanyagban előforduló egyéb cukrok valamilyen származékuk,
gyakran nukleozid-difoszfáttal képzett, aktivált formájukban
alakulnak a megfelelő glükóz-származékká, ilyan például a galaktóz.
A cukrok glikozidos hidroxilcsoportjainak foszfát-észtereiből
nukleozid-trifoszfátokkal képzett, makroerg savanhidrid kötést
tartalmazó NDP-cukor származékainak a poliszacharidok
bioszintézisénél van nagy jelentősége. E származékok keletkezésekor
a kihasadó pirofoszfát hidrolízisekor felszabaduló energia fedezi a
reakció energiaigényét. A mannóz-6-foszfát a
mannóz-foszfát-izomeráz segítségével reverzibilisen képes
glükóz-6-foszfáttá alakulni. Az aszkorbinsav (C-vitamin) a
glükuronsavból keletkezik több lépésben.
A piruvát-dehidrogenáz katalizálta reakció
A piruvát számára a mitokondrium külső és belső membránja
mindkét irányban egyaránt átjárható, ezért aerob körülmények között
a piruvát a mitokondriumban oxidálódik tovább, szén-dioxid
kihasadásával "aktív acetsavvá", a makroerg tiol-észter kötést
tartalmazó acetil-koenzim-A-vá alakul.
A piruvát oxidatív dekarboxileződését a piruvát-dehidrogenáz
multienzim komplex katalizálja, amely három enzim alegységet, három
koenzimet (TPP, Koenzim-A, NAD) és két prosztetikus csoportot
(liponsav, FAD) tartalmaz. A dekarboxiláz alegység a piruvát két
szénatomos egységét szén-dioxid vesztés közben a TPP
tiazolgyűrűjének már koenzimeknél bemutatott 2-es szénatomjához
köti. A folyamatot egy proton lehasadása, majd a piruvát
oxocsoportja oxigénjéhez kapcsolódása teszi teljessé. Az "aktív
acetaldehid" igen reakcióképes "acetálos" hidroxilcsoportja a
transzacetiláz alegység hatására oxidálódik, közben a liponsav
prosztetikus csoport redukálódik és egyben a képződő
acetilcsoporttal elreagál, S-acetil-dihidro-liponsav keletkezik. A
liponsavat gyakran liponsavamidnak nevezik, mert prosztetikus
csoportként az enzim egyik lizinjének oldalláncvégi
aminocsoportjához kapcsolódik savamid kötéssel. Az acetilcsoport
további útját a multienzim komplex harmadik alegysége, a
dihidro-lipoil-dehidrogenáz irányítja. A dihidro-liponsavról az
acetilcsoport egy koenzim-A láncvégi tiolcsoportjára kerül, így
alakul ki az acetil-koenzim-A. A dihidro-liponsav FAD prosztetikus
csoport közvetítésével NAD segítségével oxidálódik vissza.
A makroerg tiolészter csoportot tartalmazó acetil-koenzim-A a
kiindulási anyaga az egyik közös lebontó folyamatnak, a
citrátkörnek, amely működése során a szénatomok széndioxiddá
oxidálódnak
A lipidek lebontásának egyéni szakasza az
acetil-koenzim-A-ig
A lipidek lebontási folyamatainál meg kell különböztetnünk az
összetett és egyszerű lipidek lebomlását. Az összetett lipidek
lebontása a citoszolban kezdődik az észtercsoport lipázok
katalizálta hidrolízisével (a foszfolipideket hidrolizáló enzimeket
foszfolipázoknak nevezzük). A triglicerideken bemutatva a
hidrolízis három lépésben zajlik le, először az egyik láncvégi
észtercsoport bomlik el, majd tetszőleges sorrendben hasad le a
másik két zsírsav. A zsírok fokozott lebontására az emberi
szervezetet az adrenalin hormon készteti, amely egy bonyolult
reakciósor (kaszkád rendszer) beindításával, foszforilálás útján
aktiválja a lipáz enzimeket. A hidrolízis termékek közül a
glicerolt először egy kináz segítségével egy ATP foszforilálja, az
így "aktivált" glicerol-3-foszfátot (ismertebb, és a biokémiában
gyakrabban használt néven α-glicerol-foszfátot) az
α-glicerol-foszfát-dehidrogenáz enzim DHAP-tá oxidálja, amely
bekapcsolódik a glikolízisbe.
A zsírsavak általában a 3-as (β) szénatomon kiinduló lebontási
folyamattal, a β-oxidáxióval bomlanak le. A trigliceridek
hidrolízise során a citoszolban keletkező zsírsavaknak be kell
jutniuk a mitokondriumba, a β-oxidáció szinterére. A bejutás egy
speciálisan poláris molekulához, a karnitinhez kapcsolódva,
karnitin ellenében antiport membrán-transzport folyamatban,
transzlokáz közreműködésével történik.
A citoszolban a zsírsavak a karbonsavakra általánosan jellemző
módon egy ligáz (acil-koenzim-A-szintetáz) közreműködésével
aktiválódnak: először egy ATP molekula segítségével a makroerg
savanhidrid kötést tartalmazó acil-AMP keletkezik, amely a
koenzim-A-val helyettesítési reakcióba lép, makroerg tiol-észter
kötést tartalmazó acil-koenzim-A és AMP keletkezik. A reakció
energiaigényét a leszakadó pirofoszfát hidrolízise biztosítja.
A zsírsavak aktív származéka, az acil-koenzim-A a citoszolbeli
karnitin-acil-transzferáz hatására képes egy karnitint megacilezni,
majd a mitokondriumban egy koenzim-A segítségével a mitokondriális
karnitin-acil-transzferáz közreműködésével az acil-koenzim-A
regenerálódik, a karnitin visszatér a citoszolba.
A zsírsavak β-oxidációja
Az acil-koenzim-A β-oxidációjának lépései a zsírsav 3-as
metiléncsoportján egy karbonilcsoport kiépítését célozzák egy
kettős kötés oxidatív kialakításán, vízaddíción és a
β-hidroxilcsoport oxidációján keresztül.
A páratlan szénatomszámú zsírsavak lebontása
A koenzim-A tiolcsoportjának nukleofil támadása a kialakított
3-as oxocsoporton a tioláz enzim katalizálta tiolízis. Az ezt
megelőző, fent említett három lépés erős hasonlóságot mutat a
később ismertetett citrátkör utolsó három lépésével. A β-oxidáció
végén egy acetil-koenzim-A és egy két szénatommal rövidebb
zsírsavval acilezett koenzim-A keletkezik, az utóbbi -oxidációval
addig bomlik tovább, ameddig valamennyi zsírsav részlet
acetil-koenzim-A-vá alakul. A páratlan szénatommal rendelkező
zsírsavak lebontásánál az utolsó tiolízis után propionil-koenzim-A
marad vissza, amely karboxileződés és alkil-vándorlás után
szukcinil-koenzim-A formájában kapcsolódik be a citrátkörbe.
A csírázó növényekben egy alternatív lebomlás (α-oxidáció)
páratlan szénatomos zsírsavak, valamint zsíralkoholok keletkezését
teszi lehetővé egy peroxidáz enzim és hidrogén-peroxid
segítségével. A folyamathoz aktiválásra nincs szükség.
A zsírsavak α-oxidációja
A foszfolipidek valamennyi észtercsoportját különböző
foszfolipáz enzimek hidrolizálják le. Az egyszerű lipidek a
β-oxidációhoz hasonlító oxidatív folyamatokkal bomlanak le
acetil-koenzim-A molekulává.
A fehérjék, majd az aminosavak lebontásának egyedi útja
valamelyik közös intermedierig
Az élő szervezetbe kerülő fehérjék a citoplazmában először
proteázok (peptidázok) segítségével aminosavakká hidrolizálódnak.
Az exo-peptidázok közül az amino-peptidázok a hidrolízist az
N-terminálison kezdik, a C-terminálist a karboxi-peptidázok
hasítják. A karboxi-peptidáz-B a láncvégi bázisos aminosavakat
(Lys, Arg) hasítja le, a karboxi-peptidáz-A mindegyik láncvégi
aminosavat lehasítja, kivéve a bázisos oldalláncúakat. Az
endo-peptidázok meghatározott aminosavak közötti peptidkötéseket
hidrolizálnak. A pepszin savas körülmények között aktív, olyan
peptidkötéseket bont, amelyek amino komponense aromás aminosav
(tehát az aromás aminosavak előtti kötést bontja), ilyenek a Phe,
Tyr, His. Lúgos körülmények között dolgozik a tripszin és az
-kimotripszin. Az előbbi bázisos aminosav (Lys, Arg) utáni, az
utóbbi pedig aromás aminosavak (Phe, Tyr, Trp) utáni (tehát
karbonil részletét tartalmazó) peptidkötéseket hidrolizál. A
proteázok egy része aktív centrumuk katalitikus helyén nem két
karboxilcsoportot viselnek, hanem a savasabb csoportként különleges
aktivitású szerint (szerin-proteázok, pl. tripszin, kimotripszin)
vagy ciszteint (cisztein-proteázok) tartalmaznak. A
fém-proteázokban aktív helyen speciálisan kötött fém ion segíti elő
a mellette rögzített peptidkötés hidrolitikus hasadását (pl. a
karboxi-peptidáz-A-ban a Zn2+).
A proteázok saját környezetük fehérjéire is veszélyesek
lennének, ezért általában inaktív állapotban, proenzimek
(zimogének) formájában vannak. A zimogénekből megfelelő peptidázok
kémiai, általában hidrolitikus átalakításával alakulnak ki az aktív
enzimek. A pepszin zimogénje a pepszinogén, a tripsziné a
tripszinogén stb.
A citoplazmában az aminosavak egy része helyben aktiválódik és
közvetlenül a fehérjeszintézisben kerül, másik része négy lépéses
folyamatban bomlik le. Az aminosavból először az aminocsoport
távozik és α-keto-karbonsav keletkezik (transzaminálás), a folyamat
további lépései: oxidatív dezaminálás, ammónia kiürítés és az
oldalláncok oxidatív lebontása.
Az L-α-aminoavak lebomlása acetil-koenzim-A-ig vagy a citrátkör
valamelyik intermedierjéig
Transzaminálás
A citoplazmában lejátszódó transzaminálásban az aminosavak
aminocsoportjukat a transzaminázok közreműködésével,
aldimin-ketimin átrendeződéssel, α-keto-glutársavnak adják át, azt
glutaminsavvá változtatják, maguk közben α-keto-karbonsavakká
alakulnak. A transzaminázok koenzimje a piridoxál-foszfát (PAL),
amely kondenzációs reakcióban (valódi vízkilépés), aldimin képzés
közben veszi át az aminocsoportot, majd tautomer átrendeződéssel
ketiminné alakul, ez hidrolizálódik piridoxamin-foszfáttá (PAM) és
α-keto-karbonsavvá. A PAM az előbbivel analóg, de fordított módon
adja át aminocsoportját egy α-keto-glutársavnak és ketimin-aldimin
átrendeződés, valamint hidrolízis után így regenerálódik a PAL, míg
az aminocsoport glutaminsavba épülve bomlik tovább. A
transzaminálás végén valamennyi aminosav aminocsoportjából
glutaminsav képződik.
A transzaminálás első intermedierje, a piridoxál-foszfáttal
képzett aldimin nem csak ketiminné tud átalakulni. Az aldiminek
szén-dioxid vesztéssel jelentős biológiai hatású bázisokká (biogén
aminokká) alakulnak. A ciszteinből származtatható ciszteamin a
koenzim-A része, a hisztidinből származtatható hisztamin az emberi
szervezetben az allergiás folyamatokban és a gyomorpanaszoknál
játszik fontos szerepet. Az aminosavakból képzett aldiminekben az
aminosav oldallánca is lehasadhat. Példa erre a szerin-glicin
átalakulás, amelyet a hidroxi-metil-transzferáz katalizál. A
szerinből származó hidroxi-metil oldallánc (C1) komponens, a
tetrahidro-folsav (THF) 5-ös nitrogénjére kerül. Az aldiminek
valamennyi átalakulásának első lépése mindig egy proton lehasadása.
A biogén aminok több képviselőjét a biomolekulák tárgyalásakor
mutattuk be.
Oxidatív dezaminálás
A glutaminsav az aminosavak lebontásának kulcsvegyülete a
citoplazmában maradva oxidatív dezaminálással bomlik tovább.
Aminocsoportját ammónia formájában veszti el (ez a sejtben, a vizes
oldatban természetesen NH4 formában van), a glutarát-dehidrogenáz
enzim és NAD koenzim közreműködésével könnyen hidrolizálódó
imino-sav intermedieren keresztül regenerálódik a transzamináláshoz
az α-keto-glutarát. A növényekben egyes α-keto-karbonsavakból
ammónia segítségével képzett iminosavon keresztül közvetlenül
redukcióval képezhető aminosav. Ezt a felépítő folyamatot reduktív
aminálásnak nevezzük. A koenzim ilyenkor természetesen a
(NADPH+H).
Az ammónía-liázok működése
Az aminosavakból ammónía-liázok hatására telítetlen sav és
ammónia keletkezik. Így keletkezik például a növényekben a
fenilalaninból a transz-fahéjsav.
A glutamin kialakulása és bomlása
Csak egysejtűek, egyes halak és rákok képesek az ammónia
közvetlen kiürítésére. A magasabbrendű állati szervezeteket az
ammónia mérgezi. A glutamát oxidatív dezaminálása után felszabaduló
ammónia végleges kiürítése a karbamid (urea) ciklusban, karbamid
formájában történik, de az ammónia szervezeten belüli szállítására
is szükség van. Az ammónia a mitokondriumban a glutamin-szintetáz
(ligáz) hatására a glutaminsav -karboxilcsoportjába savamidként,
glutamint alkotva épül be, az ATPADP átalakulással képzett,
makroerg savanhidrid kötést tartalmazó intermedier a
-glutamil-foszfát. Analóg módon keletkezhet aszparagin is az
aszparaginsavból. Az ammónia a szervezetben glutamin formában
szállítható. A célhelyen a glutaminból az ammónia a glutamináz
(hidroláz) segítségével regenerálható. A növényekben a glutamin és
az aszparagin szintézise az ammónia beépítésének egyik
alternatívája.
A glutamát alternatív keletkezése
A glutamin és glutaminsav más reakcióban is kapcsolatba
kerülhet. Bár ez inkább felépítő folyamat, kapcsolatrendszere miatt
itt tárgyaljuk. A glutamin és az α-keto-glutarát reduktív
folyamatban, (NADPH+H) koenzim és a glutamát-szintáz
közreműködésével két glutamáttá alakul.
A karbamid (urea) ciklus
Az ammónia különböző nitrogén-származékok, az esetek túlnyomó
többségében karbamid (urea) formájában ürül ki az élő
szervezetekből. Az ammónia a karmamid ciklusban (Krebs-féle urea
ciklus) alakul át karbamiddá. A karbamid ciklus a mitokondriumban
kezdődik, mivel ott szabadul fel a glutaminból az ammónia, amelyből
szén-dioxid segítségével, két makroerg kötés energiájának
felhasználásával (2ATD2ADP), két lépésben, makroerg kötést
tartalmazó intermedier, karbamoil-foszfát (egyes szakkönyvek
karbamil-foszfátnak nevezik) keletkezik. A karbamoil-foszfát egy
ritka aminosavval, a karmabid ciklus első tagjával, az ornitinnal
citrullinná alakul az ornitin-transzkarbamoiláz (transzferáz)
hatására. A citrullin kijut a citoszolba, ott egy aszparaginsavval
két makroerg kötés hasadását igénylő reakcióban
(ATPAMP+PPi,valamint PPi+H2O2Pi), az arginino-szukcinát-szintetáz
(ligáz) közreműködésével arginino-szukcináttá alakul, amely az
arginino-szukcinát-liáz hatására argininra és fumársavra esik szét.
A fumársav intermedierként felhasználódik, kapcsolatot teremtve a
citrátkörrel, elősegítve az aszparaginsav regenerálását. Az arginin
hidrolitikusan (argináz) ornitinná alakul vissza, valamint
izokarbamid keletkezik, amely tautomer átalakulással a végtermék
karbamiddá alakul és kiürül a szervezetből.
Az α-keto-karbonsavak oxidatív lebomlása
Az aminosavak transzaminálása után visszamaradó
α-keto-karbonsavak egy része a citrátkör vagy a glikolízis egyik
intermedierje. A többi oldallánc oxidatív lebomlása során ugyancsak
a fenti intermedierek keletkeznek. Piruváttá bomlik le az Ala, Thr,
Gly, Ser és a Cys oldallánca, aceto-acetil-koenzim-A-vá bomlik a
Phe, Tyr, Trp, Leu és a Lys oldallánca, α keto-glutaráttá bomlik a
Glu, Gln, His, Arg és a Pro oldallánca. A Met, Val és Ile
oldalláncából szukcinil-koenzim-A képződik, az Asp és Asn
oldallánca oxál-acetátot ad. E folyamatok részletes ismertetésére
itt sajnos nincs lehetőség.
A tápanyagok lebontásából származó közös intermedier, az
acetil-koenzim-A további sorsa
A különböző tápanyagokból származó acetil-koenzim-A a
mitokondriumban oxidatív lebontási folyamatokban szén-dioxiddá és
vízzé alakul ("ég el"), közben jelentős energia keletkezik, ennek
egy része a szervezet későbbi energia szükségletének kielégítéséig
ADPATP átalakulással makroerg kötések formájában tárolódik. A
mitokondriumban lejátszódó közös lebontási folyamatnak azt a
szakaszát, amelyben az eredetileg a tápanyagból származó
széntartalomból szén-dioxid keletkezik és a tápanyagból származó
hidrogének redukált koenzimek formájában kerülnek tovább,
citrátkörnek nevezzük. Azt a folyamatot, amelyben a koenzimek
regenerálódnak, a tápanyagból származó hidrogének a levegő
oxigénjével vizet alkotnak, azaz az oxigén redukálódik
(elektron-transzport), közben ADP-ből és szervetlen foszfátból ATP
keletkezik (oxidatív foszforiláció), terminális oxidációnak
nevezzük.
Citrátkör (Szent-Györgyi - Krebs ciklus)
A mitokondriumban lejátszódó citrátkör ciklus jellegét Krebs
fedezte fel, mechanizmusának tisztázásában Szent-Györgyi Albert
jelentős szerepet játszott, ezért a kutatásáért ő is kapott Nobel
díjat. A ciklusban a szén-dioxid és a redukált koenzimek
keletkezésén kívül szubsztrát szinten egy makroerg kötés is kiépül
(GDPGTP átalakulás). Az acetil-koenzim-A és a citrátkör tagjai
számos bioszintetikus folyamatnak kiindulási anyagai, ezért a
citrátkört az anyagcsere gyűjtőhelyének ("gyűjtőmedencéjének")
tekintjük.
Az acetil-koenzim-A először az oxál-acetáttal citromsavat képez.
Az acetil-koenzim-A-ból a citrát-szintázzal (liáz) képzett ES
komplexben az enzim aktív helyének hisztidin oldalláncával
kialakított hidrogén-kötés hatására enol-szerkezet alakul ki,
amelynek láncvégi metilén-csoportja nukleofil támadást intéz az
oxál-acetát karbonilcsoportja ellen, majd a koenzim-A
hidrolitikusan lehasad.
A citrát vízvesztéssel (akonitsav) és vízaddícióval az akonitáz
(liáz) hatására izocitráttá alakul, amelynek hidroxilcsoportját az
izocitrát-dehidrogenáz a NAD koenzim segítségével ketonná oxidálja.
Az oxál-borostyánkősav azonnal dekarboxileződik α-keto-glutaráttá.
Az α-keto-glutarát egy multienzim komplex
(α-keto-glutarát-dehidrogenáz) segítségével, koenzim-A belépésével
oxidatív dekarboxileződésen esik át, makroerg tiol-észter kötést
tartalmazó szukcinil-koenzim-A keletkezik. Ez a folyamat a piruvát
oxidatív dekarboxileződéséhez hasonló módon megy végbe, az enzim a
piruvát-dehidrogenáz enzimmel analóg felépítésű (TPP, liponsav,
FAD, NAD, koenzim-A). Az intermedier makroerg kötésének energiája
egy GDPGTP átalakulásba épül be, miközben egy ligáz, a
szukcinil-koenzim-A-szintetáz (szukcinát-tiokináz) hatására a
molekula felhasad, szukcinát (borostyánkősav) és koenzim-A
keletkezik. A következő három lépés analóg a zsírsavak
β-oxidációjának megfelelő lépéseivel: a szukcinát fumaráttá
oxidálódik a szukcinát-dehidrogenáz hatására (koenzim: FAD), ezt
követi a vízaddíció (az enzim a fumaráz - liáz), majd a keletkezett
L-malát (almasav) oxidációja oxál-acetáttá (az enzim a
malát-dehidrogenáz, a koenzim ismét NAD).
Megjegyezzük, hogy az áttekinthetőség kedvéért az ábrázolásban
logikai bukfencet követtünk el, ugyanis az intermediereket a
szokott formában rajzoltuk. Így nem látszik, hogy az intermedierek
az eredeti acetilcsoport felől kötődnek az enzimekhez, a
dekarboxileződési folyamatokban keletkező szén-dioxidok az
oxál-acetátból származnak, az acetilcsoport két szene a regenerált
oxál-acetátba épül be. Ezt az ellentmondást úgy oldottuk fel, hogy
az oxál-acetát mintkét ábrázolását felrajzoltuk.
CH3–COSKoA+3NAD+FAD+GDP+Pi 2CO2+3(NADH+H)+FADH2+GTP
A citrátköt anyagmérlege
A citrátkör enzimei nagy enzim komplexekben a mitokondrium belső
falához tapadva működnek. A citrátkör szabályozásában az ATP/ADP
arány meghatározó szerepet játszik. Az ATP ugyanis a citrát-szintáz
(régi, helytelen nevén citrát-szintetáz) speciális (allosztérikus)
gátlószere, egyes intermedierek is a körfolyamat különböző enzimeit
allosztérikusan gátolják.
Az anyagcsere lebontási folyamatai bonyolult szabályozás alatt
állnak. A glikolízis és a citrátkör visszacsatolásos (feed back)
szabályozás alatt van. A glikolízis első lépésének inhibítora a
termék glükóz-6-foszfát. Mint már jeleztük, a PFK allosztérikus
aktivátora a fruktóz-2,6-biszfoszfát és az AMP. Ugyanezt a
folyamatot a citrát és az ATP is gátolja. A PEPpiruvát átalakulást
a zsírsavak gátolják. A piruvát-dehidrogenáz működését, az
acetil-koenzim-A keletkezését az ATP, a (NADH+H) és maga az
acetil-koenzim-A gátolja.
Negatív visszacsatolás
Nem közvetlenül az enzimek aktív helyére hatnak, mégis
közvetlenül befolyásolják az enzim aktivitását azok a vegyületek,
amelyek az allosztérikus szabályozásban vesznek részt. Az
anyagcsere folyamatok szabályozása szempontjából kiemelkedő
fontossága van az allosztérikus szabályozásnak. Ilyenkor hosszabb
anyagcsere folyamatok végterméke a folyamat első enzimére
allosztérikus gátlószerként hat, ez megakadályozza, hogy a
szóbanforgó anyag feldúsuljon. Ezt a szabályozást negatív
visszacsatolásnak (feed back) nevezzük. Az allosztérikus gátlásra
alkalmas enzimek legalább két alegységet tartalmaznak, tehát
negyedleges szerkezettel rendelkeznek. A szabályozó anyag a
szabályozó (reguláló) centrumon kötödik meg, amely nem az aktív
centrumot tartalmazó, hanem a másik alegységben van. Gyakran a
szubsztrát molekulák maguk is szabályozó hatásúak. Elvben
természetesen lehetséges pozitív visszacsatolás is, de ez nem igen
fordul elő az élő világban.
Az allosztérikus szabályozás összehangolt (concerned)
mechanizmussal megy végbe, amelyet Monod tárt fel. Az alegységeknek
két lehetséges konformációja van, az aktív (lazított, relaxed - R)
és inaktív (feszített, tensed - T) konformáció. A két alegység
egyszerre csak azonos konformációban lehet: RR vagy TT. A két forma
egyensúlyban van egymással 1:10000 arányban a T forma javára. A
szubsztrát molekula az aktív helyet tartalmazó alegységnek csak R
konformációjú alakjával képes reagálni. Az allosztérikus
aktiválószer hatására a reguláló centrumot tartalmazó alegység R
konformációjú, így a másik, a katalitikus alegység is R
konformációjú lesz, az ES komplex kialakulása meggyorsul. Az
allosztérikus gátlószerek kötődésekor a reguláló centrumot
tartalmazó alegység a T konformációban rögzül, ez lehetetlenné
teszi, hogy a szubsztrát molekulák az ugyancsak T állású második
centrumon megkötődjenek.
Terminális oxidáció
A tápanyagok közös lebontási folyamatának az a része, amelyben a
redukált koenzimekről a tápanyagokból származó hidrogének az
oxigénre kerülnek, közben a koenzimek visszoxidálódnak és az oxigén
vízzé redukálódik (elektron-transzport részfolyamat), a
tápanyagokból származó energia pedig makroerg kötésekbe épülve
jelenik meg, ADP-ből és szervetlen foszfátból ATP szintetizálódik
(oxidatív foszforiláció részfolyamat). Megjegyezzük, hogy a két
részfolyamat nevét gyakran a teljes folyamat szinonímájaként
használják. A terminális oxidáció helye a mitokondrium belső
membránja, amelyről már korábban megemlítettük, hogy lévén a külső
membránnál nagyobb, a belső tér (mátrix tér) felé kitüremkedései
vannak, ezeket krisztáknak nevezik. Emlékeztetőül:
A mitokondrium membránjai
A mitokondrium belső membránjában rögzítve helyezkednek el az
eletron-transzport nagyobb enzim komplexei (NADH-Q-reduktáz,
citokróm-reduktáz, citokróm-oxidáz), valamint a belső membránban
szabadon mozdulhatnak el a kisebb egységek, a Koenzim-Q és a
citokróm-c. Ugyancsak a mitokondrium belső membránjában van az
oxidatív foszforiláció enzimje, az FoF1-ATP-áz. A hidrofób Fo
alegység a proton csatorna, aktív transzporttal H transzportot tesz
lehetővé a mitokondrium mátrix teréből a belső membránon keresztül
a közti térbe. Az ADPATP átalakulást katalizáló F1 alegység a belső
membránnak a mátrix tér felé eső oldalán mintegy kidudorodásokat
képez. A belső membránban több transzlokáz működik, az egyik úgy
enged be egy ADP molekulát, hogy antiport folyamatban ATP
molekulára cseréli.
Az elektron-transzportban a redukált koenzimekről a hidrogének
(a folyamat elején együtt az elektronok és protonok, később térben
elválasztva) az egyik enzim komplexről a másikra kerülnek, a
részfolyamat végén a protonok, az elektronok és a levegőből
származó, és a magasabbrendű élőlényeknél a hemoglobin segítségével
hemoglobin-oxigén komplex formájában a vérkeringéssel a sejtbe
vitt, oxigénre kerülnek, azaz vizet alkotnak. Az
elektron-transzportban az elektronok (és a velük vagy tőlük
elválasztva a protonok) a negatívabb standard redoxpotenciálú
helyről a pozitívabb standard redoxpotenciálú hely felé tartanak.
Azoknál az átmeneteknél, amelyeknél a standard redoxpotenciál
különbség nagy, az elektron átvitel nagy negatív szabad entalpia
változással jár, következik be az ATP szintézis. A standard redox
potenciál értékeket zárójelben jelezzük.
Az elektron-transzport kiindulási anyaga a (NADH+H) (-0,32V),
innen a hidrogének (hidrid anion a koenzimről és proton a mátrix
térből) a NADH-Q-reduktáz (-0,30V) enzim komplexre jutnak. A
NADH-Q-reduktáz FMN prosztetikus csoportot és vas-kén fehérjéket
tartalmaz. Ez utóbbiak szabályos ("cluster") elrendeződésű, a
fehérjében lévő cisztein oldalláncban kovalensen és vashoz ionosan
kötött kenet tartalmaznak (ez utóbbi kén-tartalom savanyításra
kénhidrogén formájában távozik, ezt záptojás szag jelzi).
Ezután a hidrogének (az elektronok és protonok még mindig
együtt, kinonhidrokinon átalakulással) a koenzim-Q-ra (ubikinon)
(+0,04V) kerülnek. A koenzim-Q molekulák szabadon úsznak a
membránban. így viszik a hidrogéneket a következő enzim komplexhez,
a citokróm-reduktázhoz (), közben hidrokinon-származékból
(ubikinol) visszaoxidálódnak ubikinonná. A koenzim-Q aránylag nagy
mozgékonysága miatt a mitokondrium belső membránjának külső
oldaláról is át tud venni hidrogéneket, ezért a NADH-Q-reduktáznál
pozitívabb standard redoxpotenciálú FADH2 és FMNH2, amely itt
kapcsolódik be az elektron-transzportba, akkor is át tudja adni
neki a hidrogénjeit, ha történetesen a közti térben van.. A
továbbiakban a hidrogénekből származó elektronok és protonok útja
szétválik. A protonok a mátrix térbe, illetve az oxidatív
foszforiláció során a két membrán közötti térbe kerülnek, az
elektronok pedig egyenként vándorolnak tovább a hemet tartalmazó
citokrómok Fe3+Fe2+ átalakulásainak segítségével. A
citokróm-reduktáz komplex több citokróm fajtát tartalmaz: az
elektronok a citokróm-b566-ra (+0,07V) onnan a citokróm-b562-re
kerülnek, innen hurokszerű átmenettel, bonyolult visszacsatolással
vas-kén fehérjék következnek, majd a citokróm c1 (+0,23V), amely az
enzim komplex utolsó állomása. Ezután a membránhoz lazán kötött,
tehát szabadabban mozgó citokróm-c komplex következik, erről
kerülnek egyesével az elektronok a lánc utolsó enzim komplexére, a
citokróm-oxidázra, amelyben szintén több citokróm típus van, az
utolsó, a citokróm-aa3 (+0,55V) két réz iont is tartalmaz. A
citokróm-aa3-ról jutnak át az elektronok a mátrix térből származó
protonokkal együtt az oxigénre (+0,82V), így alakulnak ki a H2O
molekulák.
Az a kívánatos, hogy egy oxigén molekulára egyidőben kötődjék
négy elektron és négy proton, ugyanis ellenkező esetben mérgező
peroxid és szuperoxid gyökök (mivel párosítatlan elektronjuk
negatív töltést kölcsönöz nekik, ionoknak is nevezhetjük őket)
keletkeznek. Ennek érdekében a citokróm-oxidáz, illetve azon belül
a citokróm-aa3 csak akkor engedi el az oxigént (O2), ha mind a négy
elektron rajta van és a protonok is hozzákapcsolódtak, tehát a
komplexről kész víz molekulák hasadnak le. E folyamatban játszanak
jelentős szerepet a citokróm-aa3-hoz kapcsolódó Cu2+ ionok.
A toxikus melléktermékek keletkezésének lehetőségét két
védőenzim is csökkenti. A szuperoxid-dizmutáz a szuperoxid
gyök-ionokat oxigén segítségével peroxid ionokká alakítja. A
peroxid gyök-ionok protonokkal hidrogén-peroxidot képeznek, amely a
kataláz segítségével oxigénné és vízzé alakul. A két védőenzim nem
csak a mitokondriumban található, előfordulásuk univerzális,
feladatuk mindenhol a fenti méregtelenítés.
Szuperoxid ion keletkezése: O2 + e O2 Ө
Peroxid anionkeletkezése: O2 + 2e O22 Ө
Hidrogén-peroxid keletkezése: 2 H + O22Ө H2O2
A szuperoxid anion eliminálása a szuperoxid-dizmutáz hatására: 2
O2Ө O22Ө + O2
A hidrogén-peroxid eliminálása kataláz hatására: 2 H2O2 2H2O +
O2
A mérgező gyökök eliminálásának folyamat
Az elektron-transzport-lánc egyes enzimei közötti standard
redoxpotenciál különbségek okozta szabad entalpia változás három
esetben nagy negatív érték, a NADH-Q-reduktáz és a Koenzim-Q, a
citokróm-reduktáz (benne is a citokróm-b) és a citokróm-c, valamint
a citokróm-aa3 és az oxigén között. Ezeknél az átmeneteknél
felszabaduló energia épül be az oxidatív foszforilációban egy-egy
ATP egy-egy makroerg kötésébe. Mivel a FADH2 és a FMNH2 a korábban
ismertetett okokból csak a Koenzim-Q-nál kapcsolódik bele a
transzport folyamatba, oxidációjakor csak két ADPATP átalakulás
következik be.
Az oxidatív foszforiláció során az elektron-transzportban
felszabaduló energia makroerg kötésekbe épül. A folyamat
mechanizmusával kapcsolatban számos elméletet dolgoztak ki, ezek
közül a Mitchell-féle kemiozmózisos kapcsolás elmélet közelíti meg
legjobban a valóságot, bár egyes részletek még ma is
tisztázatlanok. Mitchell feltételezése szerint az
elektron-transzport megfelelő helyeinél felszabaduló energia az
FoF1-ATP-áz Fo egységének konformációváltozásának segítségével
protonokat pumpál ki a mitokondrium mátrix teréből a közti térbe. A
protonpumpa működése következtében proton koncentráció gradiens
(különbség) jön létre a belső membrán két oldala között, ez az
ozmotikus energia az ATP szintézis hajtóereje.
A citoszolban képződött (NADPH+H) elsődleges feladata az, hogy a
reduktív bioszintetikus folyamatokhoz elektronokat biztosítson.
Szükség esetén a (NADPH+H) az elektron-transzportba is tud
hidrogéneket juttatni, a NADPH-dehidrogenáz enzim
közreműködésével.
(NADPH+H) + NAD NADP + (NADH+H)
A NADPH-dehidrogenáz katalizálta reakció
Mitchell eredeti elképzelése szerint minden nagyobb negatív
szabad entalpia vátozáskor egy-egy proton, az újabb kísérletek
szerint három-három proton pumpálódik ki a közti térbe. A membrán
két oldala között kialakuló proton koncentráció különbség hatására
az FoF1-ATP-áz F1 alegysége katalizálja az ADPATP átalakulást a
belső membránban. Az ADP + Pi = ATP + H2O reakció során a víz eddig
pontosan nem tisztázott körülmények között úgy keletkezik, hogy a
membrán két oldala között a proton koncentráció különbség
kiegyenlítődik. Ez utóbbi folyamatot, mivel meghatározott térbeli
irányba mutat, vektoriális reakciónak nevezzük. Mivel a redukált
koenzimből csak két proton származhat, a kipumpált protonok nagy
többsége a víz disszociációjából származik. A mátrix térben
visszamaradó hidroxid anionok negatív töltése a potenciál
különbséget még hangsúlyozottabbá teszi.
A Mitchell-féle kemiozmózisos kapcsolás elméletre több
bizonyíték létezik. A legszemléletesebb bizonyíték az
elektron-transzport és az oxidatív foszforiláció szétkapcsolása
úgynevezett szétkapcsoló anyagokkal. Ha ugyanis egy molekula más
mechanizmussal képes a fennálló töltéseloszlással ellentétes
töltéseket a membrán túloldalára juttatni, a töltések
kiegyenlítődése az elektron transzportját nem akadályozza, de az
ATP szintézisét lehetetlenné teszi. Ilyen szétkapcsoló anyagok
egyes antibiotikumok, amelyek megváltoztatják a membránok
átjárhatósági tulajdonságait (például az oligomicin, amelyről az Fo
alegység a nevét kapta, és az úgynevezett ionofór antibiotikumok,
amelyek specifikus komplexet képeznek egyes kationokkal - például a
valinomicin a kálium ionokkal - és ezeket átviszik a mitokondrium
belső membránján). Olyan erősen apoláris, protonra és anionra
disszociálódni képes molekulák, amelyek számára a membrán mind
disszociálatlan, mind disszociált állapotban átjárható, ugyancsak
szétkapcsolók lehetnek. Ilyen vegyületek az éppen ezért rendkívül
mérgező dinitro-fenolok, amelyeket a növényvédelemben rovarölőként
(inszekticidként) használnak.
Az anyagcsere lebontó folyamatainak egymás közötti
kapcsolatai
Az anyagcsere lebontó és felépítő (bioszintetikus) folyamatai,
valamint az egyes lebontó folyamatok között a kapcsolatok bonyolult
rendszere alakult ki. Az élő szervezeteknek a kapcsolatok kölcsönös
szabályozásán kívül ez arra is lehetőséget ad, hogy egyik vagy
másik folyamat ideiglenes vagy állandó korlátozásakor a megfelelő
intermediert vagy végterméket alternatív úton hozza létre. A
szénhidrátok, lipidek és fehérjék lebontási folyamatainak
ismertetésénél többször utaltunk a fennálló kapcsolatokra, például
a lipidek lebontásából származó glicerol és a glikolízis
összefüggésére, illetve a transzaminálás, a karbamid ciklus és a
citrátkör összefonódására. A kapcsolatok rendszerét a
bioszintetikus folyamatok ismertetése teszi majd teljessé. Itt most
olyan folyamatokat mutatunk be, amelyek nélkül a tápanyagok teljes
lebontása nem lenne lehetséges.
Anaplerotikus (feltöltő) reakciók
Ahhoz, hogy a citrátkör zökkenőmentesen működjék, az
acetil-koenzim-A molekulákon kívül a mitokondriumban megfelelő
oxál-acetát koncentráció szükséges. Ha nincs jelen elegendő
oxál-acetát, a felhalmozódott acetil-koenzim-A-ból nem kívánatos
melléktermékek, úgynevezett ketontestek keletkeznek (aceto-acetát,
azaz acetecetsav, abból dekarboxileződéssel aceton, redukcióval
3-hidroxi-butirát). A ketontestek számára a mitokondrium membránjai
átjárhatók. Ez az állapot akkor következik be, ha a szervezet túl
kevés glükózt és túl sok zsíradékot bont le, ezért felszaporodik az
acetil-koenzim-A. Egyes anyagcsere-betegségekben (pl.
cukorbetegség) is felszaporodhatnak a ketontestek, ez okozza a
cukorbetegek jellegzetes illatú (acetonos) lehelletét. A
cukorbetegségben azért halmozódnak fel a ketontestek, mert a glükóz
anyagcsere zavara miatt az oxál-acetát egy része glükóz
bioszintézisre fordítódik.
Ketontestek képződése
Megjegyezzük, hogy a legújabb kutatások szerint az emberi
szívizomban fordított folyamat játszódhat le, az acetecetsav az
acetil-koenzim-A prekurzora is lehet, így energiaforrásként
szerepelhet.
Normális körülmények között a szervezet gondoskodik arról, hogy
a citrátkör zavartalan működéséhez elegendő oxál-acetát álljon
rendelkezésre. Az oxál-acetát készletet (szintet) biztosító
reakciókat nevezzük anaplerotikus reakcióknak. Az oxál-acetát
koncentráció fenntartása többszörös biztosítással, különböző
enzimek segítségével történik.
Az állati szervezetekben a legelterjedtebb út a mitokondriumban
a piruvát szén-dioxid megkötése (karboxilezése) a
piruvát-karboxiláz hatására. A koenzim a biotin, ennek megfelelően
az irreverzibilis reakció végbemeneteléhez makroerg kötés hasadása
(ATPADP), az enzim működéséhez mangán ionok jelenléte
szükséges.
A citoszolban a PEP karboxileződhet. A
foszfo-enol-piruvát-karboxi-kináz által katalizált reakció
megfordítható, mert a PEP makroerg enol-észter kötésének energiája
egy újabb makroerg kötésbe (GDPGTP) épül be. Az enzim működéséhez
magnézium ionok jelenléte szükséges. Ezeknek a folyamatoknak a
glükóz egyik bioszintézis útjában is nagy jelentősége van. A
citoszolban keletkezett oxál-acetát nem tud a mitokondriumba
bejutni, ezért maláttá redukálódva jut át a membránon és a
mitokondriumban regenerálódik.
A piruvátból két lépésben reduktív úton is keletkezhet
oxál-acetát maláton keresztül a "malát" enzim
(dekarboxiláló-malát-dehidrogenáz) segítségével. Ez a reakció egy
nagyobb körfolyamat része, amely lehetővé teszi, hogy az
acetil-Koenzim-A a mitokondriumból a citoplazmába jusson, közben a
citoszolban a (NADH+H)-ból a felépítő folyamatokhoz szükséges
(NADPH+H) keletkezzék.
Megjegyezzük, hogy oxál-acetátot eredményez az aszparaginsav
transzaminálása is az aminosavak lebontásánál ismertett módon.
Redox ingák
A citoszolban a glikolízis során a glicerinaldehid-3-foszfát
dehidrogénezésekor glükóz molekulánként két redukált koenzim
(NADH+H) keletkezik. Mivel a redoxi koenzimek számára a
mitokondrium membránjai átjárhatatlanok, a (NADH+H)
visszaoxidálásáról a citoszolban kell gondoskodni. Két lehetséges
megoldást a glükóz anaerob lebontásánál ismertettünk (alkoholos és
tejsavas erjedés), Az erjedéses folyamatok azonban energianyereség
szempontjából nem kedvezőek, ugyanakkor a mitokondriumban a
terminális oxidációban két redukált koenzim hat makroerg kötés
szintézisét jelentené. Két enzimrendszer lehetővé teszi azt, hogy a
redukált koenzimekről legalább a hidrogének bejussanak a
mitokondriumba, és ott hasonló koenzimekre kerülve beléphessenek a
terminális oxidációba.
Ezekben a folyamatokban a citoszolban képződött (NADH+H) olyan,
a citoszolban elérhető mennyiségben előforduló intermediert
redukál, amely számára a mitokondrium membránja átjárható. A
mitokondriumban a redukált intermedier visszaoxidálódik, a
hidrogének egy mitokondriális redoxi koenzimre, majd a terminális
oxidációba kerülnek, a visszaoxidálódott intermedier pedig
egyszerűbb-bonyolultabb módon visszakerül a citoszolba Az állandó
visszatérés miatt elvileg egyetlen hordozó molekula elegendő lenne
a hidrogének átvitelére. Ezt szemléletesen fejezi ki az angol
"shuttle" kifejezés, ami (szövőgépi) vetélőt jelent. Az általunk
használt "inga" szó is analóg jelentésű, ezért használjuk annak
ellenére, hogy a magyar szaknyelv csak elvétve használja. Ezt a
folyamatot gyakran extramitokondriálisan képződő (NADH+H)
oxidációjának nevezik.
Az α-glicerol-foszfát inga
Ebben a folyamatban a redukálódó vegyület a
dihidroxi-aceton-foszfát (DHAP), amely a glikolízisben nagy
mennyiségben áll rendelkezésre. A DHAP a citoszolban a (NADH+H)
segítségével glicerol-3 foszfáttá (régiesebb és gyakrabban használt
nevén α-glicerol-foszfáttá redukálódik, amely csak a mitokonrium
küldő membránján képes átjutni, és felhalmazódik a közti térben. A
közti térben az α-glicerol-foszfát a FAD segítségével oxidálódik
vissza DHAP molekulává, amely visszatér a citoszolba. A FADH2 nem
kerül be a mátrix térbe, a belső membrán külső oldalánál lépes
hidrogénjeit a membránban úszó koenzim-Q-nak átadni. Ez a redox
inga látszólag energia veszteséggel működik, mert az eredeti,
citoszolbeli (NADH+H) három makroerg kötés keletkezését indikálná,
a mitokondrium közti terében keletkezett FADH2 csak kettőt jelent.
Ez a folyamat a látszólagos energia veszteség ellenére is kedvező,
mert a FADH2 redukált koenzim a nagyobb mitokondriumbeli (NADH+H)
koncentráció, tehát koncentráció gradiens ellenében is képes a
hidrogéneket a citoszolból a terminális oxidációba juttatni, tehát
aktív membrán transzportnak tekinthető.
A malát-aszpartát inga
A másik redox inga elsősorban a máj és szív sejtjeiben működik.
A malát-aszpartát rendszer nem jár energia veszteséggel, mivel a
mitokondriumban is (NADH+H) keletkezik, ennek megfelelően az
áthaladásra a passzív transzport szabályai érvényesek. Ez az oka
annak, hogy a koncentráció viszonyoktól függően a hidrogén
szállítás iránya megfordulhat (reduktív ekvivalens). Ebben az
ingában a citoszolban oxál-acetát - malát átalakulás történik, és a
malát számára egy transzlokáz segítségével a mitokondrium belső
membránja is átjárható. A mitokondriumban az oxál-acetát
regenerálódik. Mivel azonban számára a mitokondrium belső membránja
átjárhatatlan, kerülőútra kényszerül.
Egy transzaminálási folyamatban az oxál-acetát és egy
glutaminsav aszparaginsavvá és α-keto-glutaráttá alakul. E két
anyag számára transzlokázok segítségével már átjárható a
mitokondrium belső membránja. Az α-keto-glutarát a malát partnere
az antiport membrán-transzport folyamatban. Az aszpartát egy másik
transzlokáz segítségével jut ki, cserepartnere az a glutaminsav,
amely az oxál-acetát mellett a regeneráló citoszolbeli,
transzamináz katalizálta reakcióban keletkezik. A folyamat első
fele megegyezik azzal a transzport folyamattal, amellyel az
anaplerotikus reakciókban az oxál-acetát bejuttatható a
mitokondriumba.
A malát-aszpartát inga csak a csúcsra járatott szervekben (ilyen
a máj és a szív) képes működni, amelyekben a mátrix (NADH+H)
koncentrációja a magas igénybevétel miatt aránylag alacsony.
A glükóz aerob lebontásának anyagmérlege
Egy molekula glükóz lebontása a α-glicerol-foszfát ingával:
C6H12O6 + 6 O2 6 CO2 + 6 H2O, valamint 36 ADP + 36 Pi 36 ATP +
36 H2O
A malát-aszpartát inga közreműködésével a képződő makroerg
kötések száma 38.
Pentóz-foszfát ciklus
Az anyagcserében a lebontó és felépítő folyamatok között
különleges helyet foglal el a citoszolban lejátszódó pentóz-foszfát
ciklus. Természetét tekintve a glükóz alternatív lebontása. Ezt
jelzi másik elnevezése is (a glükóz direkt oxidációja), amely
félrevezető, mert a folyamat nem glükózból, hanem
glükóz-6-foszfátból indul ki. A pentóz-foszfát ciklusban a
glükóz-6-foszfát oxidatív degradáció (lebomlás) következtében
szén-dioxidot veszít és ribulóz-5-foszfáttá, tehát pentóz-foszfáttá
alakul, közben két (NADPH+H) keletkezik. Ez a redukált koenzim a
bioszintetikus (felépítő) folyamatok koenzime, ez különleges
kapcsolatot jelent a lebontó és felépítő anyagcsere folyamatok
között. A ribulóz-5-foszfát transzfer cukor-foszfát átalakítási
módszerekkel ciklusos reakcióban alakul tovább, végeredményben 6
ribulóz-5-foszfátból 5 glükóz-6-foszfát keletkezik. A glükóz
alternatív lebontása energianyerés szempontjából nem különbözik
számottevően a glükóz aerob lebontásától (egy glükóz-6-foszfátból
12 redukált koenzim keletkezik, ez elvben 36 makroerg kötésnek
felel meg). A folyamat jelentősége nem ebben, hanem a keletkezett
intermedierekben és végtermékekben van.
A pentóz-foszfát ciklusban keletkeznek a heterotróf élőlények
reduktív, felépítő folyamataihoz szükséges redukált (NADPH+H)
koenzimek (az autotróf növények ezekhez a fotoszintézisben jutnak).
A pentóz-foszfát ciklusra a növényeknek is szükségük van, mert a
fotoszintézisben kulcs-szerepet játszó ribulóz-1,5-biszfoszfát
kiindulási anyaga, a ribulóz-5-foszfát itt keletkezik. A ciklusban
a ribulóz-5-foszfátból egyéb cukor-foszfátok, így ribóz-5-foszfát
is keletkezik, amely a nukleinsavak egyik felépítő anyaga, tehát a
pentóz-foszfát ciklusra valamennyi élőlénynek szüksége van. Az
emlősökben a glükóz körülbelül 10 %-a bomlik le ezen az úton.
A folyamat bevezető két lépésében keletkezik a két (NADPH+H), a
glükóz-6-foszfát6-foszfo-glukonsav-lakton
(glükóz-6-foszfát-dehidrogenáz), illetve a szén-dioxid fejlődésével
járó glukonsav-6-foszfátribulóz-5-foszfát (Mg2+ ionokat igénylő
glukonsav-6-foszfát-dehidrogenáz) átalakulásban. A
ribulóz-5-foszfát egy izomeráz hatására ribóz-5-foszfáttá, egy
epimeráz hatására pedig xilulóz-5-foszfáttá alakul.
A cukor-foszfátok további átalakulásában két transzferáz típus
működik közre, mindkettő egy ketóz-foszfátból és egy
aldóz-foszfátból úgy készít egy aldóz- és egy ketóz-foszfátot, hogy
az eredeti ketóz-foszfát első két (transzketoláz) vagy három
(transzaldoláz) szénatomos részletét az aldóz-foszfátra viszi, így
az eredeti ketóz-foszfátból rövidebb aldóz-foszfát, az eredeti
aldóz-foszfátból hosszabb ketóz-foszfát keletkezik. A fenti
transzferázok csak olyan átalakulásokat katalizálnak, amelyekben az
eredeti ketóz-foszfát 3-as szénatomjához és a kialakuló
ketóz-foszfát 3-as szénatomjához (transzketoláz), illetve az
átkerülő 3-as szénatomhoz (transzaldoláz) kapcsolódó
hidroxilcsoport L térállású.
A transzketolázok és a transzaldoláz működésének eredményeképen
6-pentóz-5-foszfátból 4 glükóz-6-foszfát és 2
glicerinaldehid-3-foszfát keletkezik. A két utóbbiból a később
bemutatott glükoneogenézis folyamata szerint keletkezik
glükóz-6-foszfát.
A pentóz-foszfát ciklus anyagmérlege
Felépítő (bioszintetikus) folyamatok, anabolizmus
A szénhidrátok és a lipidek felépítő folyamatainak egy része a
lebontó folyamatok fordított irányú lejátszódása néhány lényeges
kulcslépés kivételével.
A szénhidrátokat felépítő folyamatok
A szénhidrátokat felépítő folyamatok közül elsősorban a glükóz
bioszintézise, valamint a poliszacharidok felépülésének elve
érdemel részletezést.
A glükóz bioszintézise (glükoneogenézis)
A glükóz piruvátból, laktátból, vagy a citrátkör
intermedierjeiből kiinduló bioszintézisét nevezzük
glükoneogenézisnek (a glükóz ujjászületésének). Kiindulási anyagul
olyan aminosavak is szolgálhatnak, amelyek transzaminálással a
fenti kiindulási anyagok valamelyik képviselőjévé alakulhatnak. A
glükoneogenézis voltaképen a glikolízis fordított menetét
követi.
A glikolízisnek három olyan lépése van, amely energetikailag nem
megfordítható, ezeket a glikolízis ismertetésénél részletesen
tárgyaltuk. A glükoneogenézis szempontjából az első lépés a
piruvátPEP átalakulás is ezek közé tartozik, ezért csak kerülőúton
mehet végbe. A lépést bonyolultabbá teszi, hogy a piruvát a
mitokondriumban fordul elő nagyobb mennyiségben, a PEP-re pedig a
citoplazmában van szükség. A piruvát a mitokondriumban az
anaplerotikus reakcióknál ismertetett módon szén-dioxidot köthet
meg a piruvát-karboxiláz hatására és oxál-acetáttá alakul. Az
oxál-acetáton keresztül kapcsolódhatnak be a folyamatba a citrátkör
egyéb intermedierjei. Az oxál-acetát ugyancsak az anaplerotikus
reakcióknál részletezett feltételek mellett malát formájában jut ki
a mitokondriumból a citoszolba és alakul PEP molekulává a
foszfo-enol-piruvát-karboxi-kináz hatására. Ez utóbbi reakció most
az anaplerotikus reakcióhoz képest ellentétes irányba megy végbe,
ezért egy makroerg kötés energiáját igényli (GTPGDP).
Végeredményben a piruvátPEP átalakulás két makroerg kötés
felhasadását igényli (ATPADP és GTPGDP), a PEP molekula pedig csak
egy makroerg enol-észter kötéssel rendelkezik, így a folyamat egy
makroerg kötés energiáját emészti fel. A mitokondriumban lejátszódó
lépések a bekeretezett részben láthatók.
Glükoneogenézis folyamata
A glikolízis másik két megfordíthatatlan reakciója a két kináz
katalizálta foszforilálás. A fruktóz-1,6-biszfoszfát és a
glükóz-6-foszfát foszfát-észter kötéseinek hidrolitikus hasadását a
glükóz bioszintézisében két specifikus foszfatáz (hidroláz) végzi.
Ha a glükoneogenézis célja poliszacharidok felépítése, akkor a
glükóz-6-foszfátból a glükóz kialakulása elmarad.
A glükóz bioszintézise a növényekben (fotoszintézis)
Az élő világ szempontjából alapvető fontosságú az a folyamat,
amelyben a napfény energiája fotonok formájában kémiai energiává
képes átalakulni, ez a növényi fotoszintézis. Arra, hogy élő
szervezetek szervetlen anyagokból szerves molekulákat állítsanak
elő, kevés kivételtől eltekintve a fotoszintézis (a glükóz
bioszintézise széndioxidból és vízből), az egyetlen lehetőség. A
fotoszintézis során az elnyelt fényenergia a víz fotolízisét
okozza, közben a levegő szén-dioxidja asszimilálódik, belőle glükóz
szintetizálódik.
6 H2O + 6 CO2 12 h C6H12O6 + 6 O2
A víz és a szén-dioxid átalakulási folyamatai a lebontási
folyamatokkal analóg módon a fotoszintézisben is elkülönülnek. A
fotoszintézis fényszakaszában két fényelnyelő rendszer segítségével
két foton energiájának hatására a víz fotolízist szenved és oxigén
szabadul fel. A vízből származó hidrogének (elektronok és protonok)
egy elektron-tra