Embed Size (px)
Citation preview

BIOLOŠKA KEMIJA
Praktikum
Jerka Dumić Sanja Dabelić Olga Gornik
Gordana Maravić Vlahoviček Ruđer Novak
Sandra Šupraha Goreta
Farmaceutsko-biokemijski fakultet Sveučilišta u Zagrebu
Zavod za biokemiju i molekularnu biologiju
Zagreb, 2010.

Urednik
Prof. dr. sc. Jerka Dumić
J. Dumić i sur.,
Biološka kemija Praktikum;
ur. J. Dumić,
izdavač Sveučilište u Zagrebu,
Farmaceutsko-biokemijski fakultet,
Zagreb 2010.
ISBN 978-953-6256-61-7 CIP zapis dostupan u računalnom katalogu
Nacionalne i sveučilišne knjižnice u Zagrebu pod brojem 744598
Umnožavanje, preslike ili pretisak nisu dopušteni bez odobrenja autora.
Recenzenti
Prof. dr. sc. Karmela Barišić
Prof. dr. sc. Jasna Lovrić
Slika na naslovnici: prostorna struktura ljudskog deoksihemoglobina pri rezoluciji 1,74 Å. Slika preuzeta iz proteinske baze podataka RCSB PDB.

BIOLOŠKA KEMIJA — PRAKTIKUM
Vježba 1. Potenciometrijska titracija aminokiselina ............................................................... 1
Vježba 2. Gel-filtracija hemoglobina ....................................................................................... 10
Vježba 3. Pročišćavanje imunoglobulina G iz humanog seruma ....................................... 15
Vježba 4. Enzimska kinetika .................................................................................................... 24
Dodatak: Enzimska kinetika ..................................................................................................... 30
Vježba 5. Ugljikohidrati ............................................................................................................ 39
Vježba 6. Lipidi. ......................................................................................................................... 46
Vježba 7. Termička denaturacija DNA ................................................................................... 51
Sadržaj (sekvenciranje


BIOLOŠKA KEMIJA — PRAKTIKUM
1
Vježba 1.
1.1 Aminokiseline
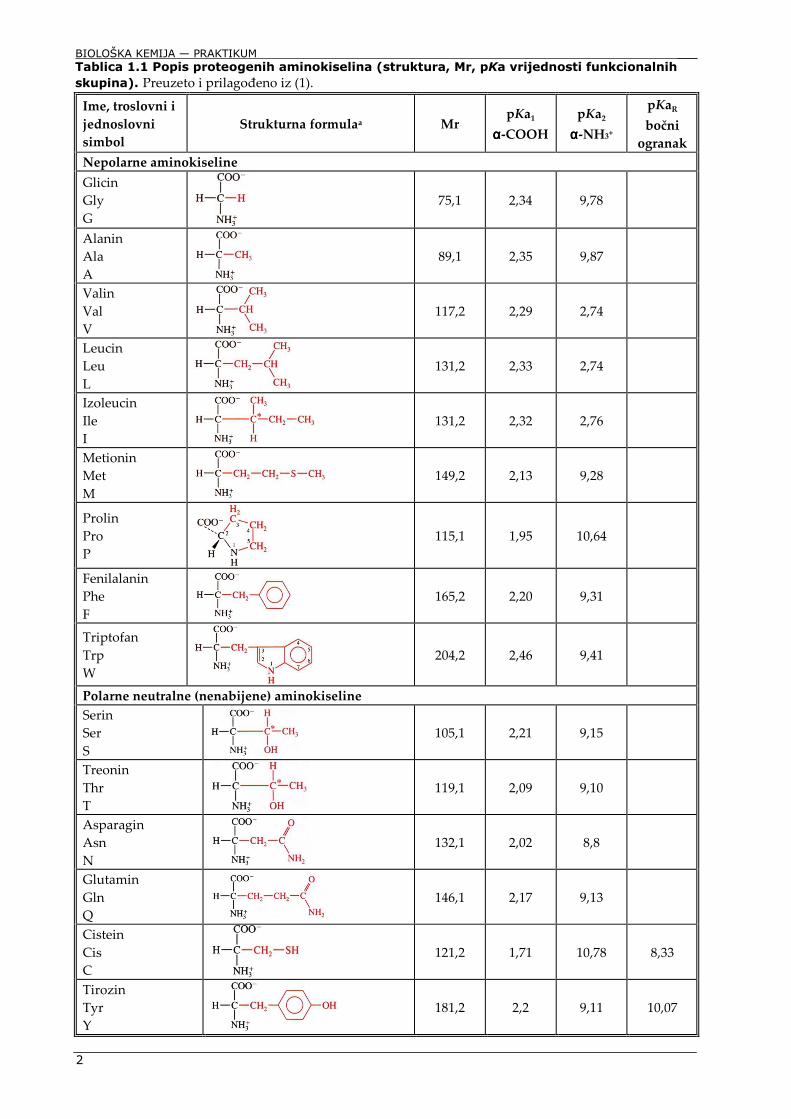
Aminokiseline su α-amino-α-karboksilne kiseline. Na Cα-atom je uz amino (-NH2) i karboksilnu skupinu (-COOH) vezan vodikov atom (-H) te bočni ogranak (-R) (Slika 1.1). Upravo stoga sve aminokiseline osim glicina (kojem je R = H) imaju asimetrični Cα-atom, a u organizmu su prisutne samo lijeve (L) aminokiseline. Bočni ogranci aminokiselina strukturno su raznoliki, od jednostavnih alifatskih struktura (alanin, valin, leucin), aromatskih (fenilalanin, tirozin, triptofan), do cikličkih (prolin, koji ujedno jedini od aminokiselina sadrži sekundarnu amino skupinu), ili pak onih koje u bočnom ogranku sadrže sumpor (metionin, cistein), kisik (treonin, serin, tirozin), dušik (asparagin, glutamin, arginin, histidin) ili karboksilnu skupinu (asparaginska i glutaminska kiselina) (Tablica 1.1). Upravo je strukturna raznolikost aminokiselina ključan preduvjet raznolikosti polipeptida (proteina) koji nastaju povezivanjem aminokiselina peptidnom vezom. Peptidna veza, koja je zapravo amidna veza, nastaje uvijek povezivanjem α-karboksilne skupine jedne s α-amino skupinom druge aminokiseline. Time su polipeptidni lanci kao i sve biološke molekule usmjerene molekule (od N-kraja prve prema C-kraju posljednje amninokiseline u lancu), dok su peptidne veze smjera C-kraj – N-kraj (Slika 1.2). Svi do danas otkriveni proteini izgrađeni su od 20 aminokiselina koje stoga nazivamo proteogenim aminokiselinama.
1.2 Fizikalno-kemijska svojstva aminokiselina
Aminokiseline, a time i proteini kao i neke druge biološke molekule, primjerice nukleinske kiseline, poliprotonske su kiseline. Svaka skupina koja može disocirati (-COOH, -NH3+, =NH2+, -OH, -SH) opisana je konstantom disocijacije Ka, odnosno pKa (-log Ka). Sve aminokiseline u svojoj strukturi imaju najmanje dvije takve skupine: α-amino (pKa ∼2,2) i α-karboksilnu skupinu (pKa ∼9,4). To znači da će pri pH > 3,5 α-karboksilna skupina biti gotovo u potpunosti deprotonirana, dok će α-amino skupina pri pH < 8 biti protonirana, odnosno da su i α-amino i α-karboksilna skupina svih aminokiselina u fiziološkom pH području u potpunosti ionizirane (Slika 1.3).
Potenciometrijska titracija aminokiselina (sekvenciranje
Slika 1.1 Opća strukturna formula
aminokiselina pri pH 1. Preuzeto i prilagođeno iz (1).
Slika 1.2 Stvaranje peptidne veze.
Preuzeto i prilagođeno iz (1).
H3N COOH
R
H
+ Cα
BIOLOŠKA KEMIJA — PRAKTIKUM ___
2
Ime, troslovni i
jednoslovni
simbol
Strukturna formulaa Mr pKa1
αααα-COOH
pKa2
αααα-NH3+
pKaR
bočni
ogranak
Nepolarne aminokiseline
Glicin Gly G
75,1 2,34 9,78
Alanin Ala A
89,1 2,35 9,87
Valin Val V
117,2 2,29 2,74
Leucin Leu L
131,2 2,33 2,74
Izoleucin Ile I
131,2 2,32 2,76
Metionin Met M
149,2 2,13 9,28
Prolin Pro P
115,1 1,95 10,64
Fenilalanin Phe F
165,2 2,20 9,31
Triptofan Trp W
204,2 2,46 9,41
Polarne neutralne (nenabijene) aminokiseline
Serin Ser S
105,1 2,21 9,15
Treonin Thr T
119,1 2,09 9,10
Asparagin Asn N
132,1 2,02 8,8
Glutamin Gln Q
146,1 2,17 9,13
Cistein Cis C
121,2 1,71 10,78 8,33
Tirozin Tyr Y
181,2 2,2 9,11 10,07
Tablica 1.1 Popis proteogenih aminokiselina (struktura, Mr, pKa vrijednosti funkcionalnih
skupina). Preuzeto i prilagođeno iz (1).
BIOLOŠKA KEMIJA — PRAKTIKUM
3
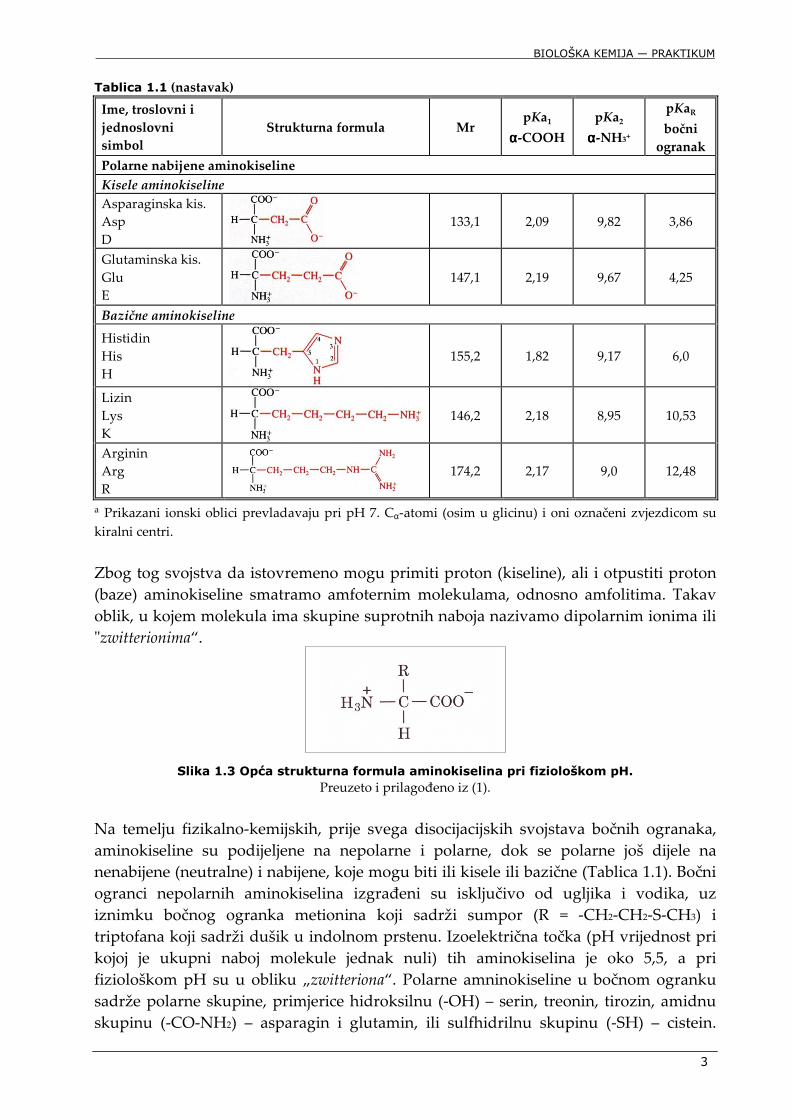
Tablica 1.1 (nastavak) Ime, troslovni i
jednoslovni
simbol
Strukturna formula Mr pKa1
αααα-COOH
pKa2
αααα-NH3+
pKaR
bočni
ogranak
Polarne nabijene aminokiseline
Kisele aminokiseline
Asparaginska kis. Asp D
133,1 2,09 9,82 3,86
Glutaminska kis. Glu E
147,1 2,19 9,67 4,25
Bazične aminokiseline
Histidin His H
155,2 1,82 9,17 6,0
Lizin Lys K
146,2 2,18 8,95 10,53
Arginin Arg R
174,2 2,17 9,0 12,48
a Prikazani ionski oblici prevladavaju pri pH 7. Cα-atomi (osim u glicinu) i oni označeni zvjezdicom su kiralni centri.
Zbog tog svojstva da istovremeno mogu primiti proton (kiseline), ali i otpustiti proton (baze) aminokiseline smatramo amfoternim molekulama, odnosno amfolitima. Takav oblik, u kojem molekula ima skupine suprotnih naboja nazivamo dipolarnim ionima ili "zwitterionima“.
Slika 1.3 Opća strukturna formula aminokiselina pri fiziološkom pH. Preuzeto i prilagođeno iz (1).
Na temelju fizikalno-kemijskih, prije svega disocijacijskih svojstava bočnih ogranaka, aminokiseline su podijeljene na nepolarne i polarne, dok se polarne još dijele na nenabijene (neutralne) i nabijene, koje mogu biti ili kisele ili bazične (Tablica 1.1). Bočni ogranci nepolarnih aminokiselina izgrađeni su isključivo od ugljika i vodika, uz iznimku bočnog ogranka metionina koji sadrži sumpor (R = -CH2-CH2-S-CH3) i triptofana koji sadrži dušik u indolnom prstenu. Izoelektrična točka (pH vrijednost pri kojoj je ukupni naboj molekule jednak nuli) tih aminokiselina je oko 5,5, a pri fiziološkom pH su u obliku „zwitteriona“. Polarne amninokiseline u bočnom ogranku sadrže polarne skupine, primjerice hidroksilnu (-OH) – serin, treonin, tirozin, amidnu skupinu (-CO-NH2) – asparagin i glutamin, ili sulfhidrilnu skupinu (-SH) – cistein.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
4
Skupine u bočnom ogranku dviju od ovih aminokiselina (tirozin i cistein), u lužnatom mediju mogu disocirati (pKa fenilne skupine –C6H4OH tirozina iznosi 10,46, a sulfidrilne skupine –SH cisteina 8,37). Pri fiziološkom su pH ipak sve polarne aminokiseline neutralne (bočni ogranak im je nenabijen), a njihova izoelektrična točka (pI) je oko 5,5. Polarne nabijene aminokiseline podijeljene su na kisele (u bočnom ogranku sadrže karboksilnu skupinu – asparaginska i glutaminska kiselina) i bazične (u bočnom ogranku sadrže ili amino skupinu – lizin, ili guanidino skupinu – arginin, ili imidazolni prsten – histidin) na temelju toga što su im pI vrijednosti u kiselom, odnosno bazičnom području, a pri fiziološkom pH su konjugirane baze, odnosno konjugirane kiseline. Otapanjem izoelektričnog oblika aminokiseline u vodi pH takve otopine imat će pH koji odgovara pI te aminokiseline.
Aminokiseline su, dakle, poliprotonske kiseline, pa ako takvoj molekuli, primjerice lizinu (R = -CH2CH2CH2CH2NH3+), dodajemo kiselinu doći će do protoniranja, odnosno ako dodajemo lužinu, doći će do niza protonskih disocijacija (disocijacija protona aminokiseline lizina prikazana je na slici 1.4.
Slika 1.4 Titracija lizina jakom lužinom.
Preuzeto i prilagođeno iz (2).
2,18 8,95 10,53
14
12
10
8
6
4
2
0
mol NaOH / mol AK
pH
0 1,0 2,0 3,0
pK1
pK2
pK3
pI
AK2+ AK+ AK0 AK-

BIOLOŠKA KEMIJA — PRAKTIKUM
5
U tom se primjeru pKa (-log Ka) vrijednosti (2,18; 8,95 i 10,53) dovoljno razlikuju da titracijska krivulja jasno pokazuje pojedinačne disocijacijske korake. S obzirom na to da pKa odgovara onoj vrijednosti pH pri kojoj je količina disociranog i nedisociranog oblika jednaka, očekujemo da će svaki disocijacijski korak stvoriti svoje pufersko područje.
Nagib titracijske krivulje je minimalan pri dodatku 0,5, 1,5 odnosno 2,5 ekvivalenta baze. Tada je [AK2+] = [AK+], odnosno [AK+] = [AK0], odnosno [AK0] = [AK-]. U tim točkama dodatak baze izaziva najmanju promjenu pH.
Otopina koja se opire promjeni pH poznata je kao puferska otopina. Puferi su prema definiciji smjese slabih kiselina ili baza i pripadajućih im soli. Djelovanje pufera sastoji se, dakle, u sposobnosti A- iona da preuzmu dodane H+ ione i stvore HA te na taj način ublaže promjenu pH. S druge pak strane, prilikom dodatka OH- iona dolazi do preuzimanja protona od strane HA pri čemu nastaju H2O + A- ione, čime se puferska otopina opire promjeni pH.
Puferski kapacitet definira se kao broj molova H+ ili OH- iona koji dodani sustavu smanjuju ili povećavaju pH za jednu jedinicu. Puferski kapaciteti nekog pufera u kiselom i u baznom smjeru mogu biti različiti, a jednaki su samo kada je pH = pKa i tada on ima maksimalnu vrijednost (maksimalni puferski kapacitet). To slijedi izravno iz definicije konstante disocijacije izražene u logaritamskom obliku koji je poznat pod imenom Henderson-Hasselbalchova jednadžba.
Kapacitet pufera ovisi i o njegovoj koncentraciji. Uz isti omjer koncentracija HA i A-,
promjena pH izazvana dodatkom određene količine kiseline ili lužine bit će to manja, što su koncentracije [HA] i [A-] veće, dakle što je puferska otopina koncentriranija. Ako je pKa za disocijaciju protona 6,8, tada će smjesa HA i A- biti dobar pufer oko neutralnog pH.
Kod aminokiselina postupna se ionizacija zbiva na kemijski različitim skupinama. Tako glicin ima karboksilnu skupinu (pKa = 2,34) i amino skupinu (pKa = 9,78). Kod neutralnog pH glicin je u "zwitterionskom" obliku.
+NH3CH2CO2H +NH3CH2CO2– NH2CH2CO2– ukupan naboj +1 0 -1 Titriracijom glicina doći će do disocijacije protona u dva koraka. Glicin je amfoterni elektrolit jer do disocijacije protona dolazi i u kiseloj i u bazičnoj sredini. Očekujemo da će pri određenom pH koncentracija pozitivnih i negativnih naboja biti jednaka. Taj pH poznat je pod imenom izoelektrična točka. Možemo je izračunati, ako znamo pKa1 i pKa2.
2,34
pKa1 pKa2
9,78
"zwitterion"
, odnosno Ka= [H+] [A-]
[HA] pKa + log pH=
[A-]
[HA]

BIOLOŠKA KEMIJA — PRAKTIKUM ___
6
]COCHNH[
]][HCOCH[NH
H]COCHNH[
]][HCOCHNH[ =aa
]COCHNH[
]][HCOCH[NH = a
H]COCHNH[
]][HCOCHNH[ = a
-223
+-222
223
+-223
+
21
-223
+-222
2223
+-223
+
1
++
++
×× KK
KK
U izoelektričnoj točki [+NH3CH2CO2H] = [NH2CH2CO2-], pa je
IKK
KK
KK
p = 2
)ap + a(p = pH
][H log = a log + a log
][Ha a
21
2+21
221
+=×
pH izoelektrične točke obično označavamo simbolom pI. pI za glicin je prema tome (2,34 + 9,78) / 2 = 6,06.
Ako molekula ima više od dvije ionizacijske skupine, disocijacija protona odvija se u više koraka. Izoelektrična točka nalazit će se u onom području pH, u kojem je zwitterionski oblik molekule dominantna vrsta. pI će kao i u prethodnom primjeru biti jednak onom pH kod kojeg je koncentracija molekularnog oblika s jednim pozitivnim nabojem jednaka koncentraciji molekulske vrste s jednim negativnim nabojem. Kod glutaminske će kiseline, primjerice, pI biti 3,23, odnosno (pKa1 + pKaR) / 2. Udio dvostruko negativnog molekularnog oblika pri tom je pH zanemariv. To se može lako dokazati (učinite to za vježbu!). Općenito možemo reći da je pI jednak aritmetičkoj sredini pKa vrijednosti onih disocijacijskih reakcija u kojima izravno sudjeluje zwitterionski oblik.
1.3 Zadaća
Titrirajte otopinu izoelektrične aminokiseline kiselinom, a potom i bazom. Na temelju krivulje dobivene potenciometrijskom titracijom uzorka odredite o kojoj se aminokiselini radi. Iz grafa očitajte pKa vrijednosti, pI vrijednost te ekvivalente dodane kiseline, odnosno lužine na osnovi kojih ćete izračunati Mr nepoznate aminokiseline iz uzorka.
N H3CH 2 CO2-
NH2CH2CO2- + H
NH3 C H2 C O2 H+NH3 C H2 C O2
- H+ +
+
+
+

BIOLOŠKA KEMIJA — PRAKTIKUM
7
1.4 Reagencije
• 0,2 M NaOH u dest. vodi • 0,2 M HCl u dest. vodi • otopina izoelektrične aminokiseline (3 g/L destilirane vode)
1.5 Pribor
• pH-metar • kombinirana elektroda • magnetska miješalica • magnet • bireta za HCl (25 mL) • bireta za NaOH (25 mL) • stalak za birete s nastavkom • pipete • čaša od 100 mL • odmjerne tikvice • staničevina • milimetarski papir
1.6 Načelo postupka
Dodatkom jake kiseline ili lužine u otopinu izoelektrične aminokiseline dolazi do protoniranja, odnosno deprotoniranja njenih funkcionalnih skupina, što se očituje kao promjena pH otopine aminokiseline.
1.7 Postupak
• U čašu od 100 mL dodajte 40 mL uzorka (otopina izoelektrične aminokiseline, 3 g/L) te u nju stavite magnet. Postavite čašu na magnetsku miješalicu tako da se magnet lagano okreće.
• Kombiniranu elektrodu prethodno uključenog i baždarenog pH-metra izvadite iz zasićene otopine KCl, isperite je destiliranom vodom, lagano obrišite staničevinom i uronite u čašu s uzorkom. Pazite da elektrodu uronite dovoljno duboko u otopinu, ali tako da je magnet ne može udariti. Napomena: poželjno je elektrodu pH-metra čim prije uroniti u otopinu, odnosno čim kraće vrijeme ostavljati na zraku.
• Napunite biretu od 25 mL 0,2 M otopinom NaOH te je postavite tako da možete titrirati uzorak u koji je uronjena elektroda u čaši na miješalici.
• Titrirajte sadržaj u čaši 0,2 M otopinom NaOH iz birete. Lužinu dodajte u malim obrocima (0,1-0,2 mL), kako bi grafički prikaz titracije sadržavao što više eksperimentalnih točaka. Nakon svakog dodatka lužine pričekajte da se pH ustali, očitajte ga i zabilježite.
• Po završetku titracije izvadite elektrodu i magnet iz čaše, a uzorak koji ste titrirali bacite. Elektrodu isperite vodom i obrišite te je potom uronite u čistu čašu u koju ste stavili novih 40 mL uzorka i magnet i postavili je na miješalicu.
• Na već opisani način titrirajte uzorak 0,2 M otopinom HCl.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
8
• Skicirajte titracijsku krivulju te je interpretirajte na temelju uputstava koja slijede: Početni pH otopine (prije dodatka kiseline ili lužine) nepoznate aminokiseline u izoelektričnom obliku odgovara njenoj izoelektričnoj točki (pI). Postupnim dodatkom malih volumena kiseline ili lužine dolazi do promjene pH otopine što se mjeri pH-metrom. Unošenjem podataka o dodanom volumenu kiseline ili lužine (na os apscisu) i izmjerenom pH (na os ordinatu) moguće je skicirati titracijsku krivulju aminokiseline. Platoi na krivulji odgovarat će puferskom području (oko pKa) pojedine funkcionalne skupine, pa će se tako na titracijskoj krivulji aminokiseline s 2 funkcionalne skupine (koje u bočnom ogranku nemaju skupinu koja može disocirati) razaznati 2 platoa, dok će se na titracijskoj krivulji aminokiseline s 3 funkcionalne skupine (koje u bočnom ogranku imaju skupinu koja može disocirati) razaznati 3 platoa. Volumen dodane kiseline ili lužine potreban da se pojedina funkcionalna skupina prevede iz deprotoniranog oblika u protonirani ili obrnuto sadrži količinu (molove) H+ ili OH- iona koja je ekvivalentna količini (molovima) te funkcionalne skupine, odnosno same aminokiseline (pH = točka ekvivalencije, T.E.). Ukoliko na krivulji odredimo volumen dodane kiseline potreban da se pojedina funkcionalna skupina prevede iz deprotoniranog oblika u oblik kada je 50% skupine još uvijek deprotonirano, a 50% protonirano (pH = pKa), taj volumen sadrži polovinu količine (molova) te funkcionalne skupine, odnosno same aminokiseline. Isto vrijedi i za dodatak lužine samo što tada u potpunosti protoniran oblik funkcionalne skupine prevodimo u oblik kada je 50% skupine deprotonirano, a 50% još uvijek protonirano, pa pH opet odgovara pKa). Na temelju određene količine (broja molova) i poznate mase nepoznate aminokiseline koja je otopljena u volumenu uzetom za titraciju moguće je izračunati njenu molekulsku masu. Na temelju podataka o broju funkcionalnih skupina, pKa vrijednosti i molekulske mase može se odrediti o kojoj se aminokiselini radi.
1.8 Zadaci
1. Pri kojem je pH puferski kapacitet dva puta veći u lužnatom nego u kiselom smjeru?
2. Glicin se često upotrebljava kao pufer. Komercijalno ga možemo nabaviti u obliku glicin hidroklorida, izoelektričnog glicina ili natrijevog glicinata.
glicin hidroklorid (AK+) izoelektrični glicin (AK0) natrijev glicinat (AK-)
Cl- H3 N
+C
COOH
H
H H 3N+
C
COO-
H
H
H2N C
CO O - Na
+
H
H

BIOLOŠKA KEMIJA — PRAKTIKUM
9
Izračunajte ionsku jakost 0,05 M glicinskog pufera pri pH 9,78. Pretpostavite a) da je pufer priređen titracijom izoelektričnog glicina otopinom NaOH b) da je pufer priređen titracijom glicin hidroklorida otopinom NaOH.
3. Skicirajte titracijsku krivulju glutamilserilglutamilvalina. 4. Izračunajte konačni pH otopine ako ste u 500 mL 0,5 M acetatnog pufera pH 5
dodali 1 mL 0,5 M HCl (zanemarite promjenu volumena) (pKa = 4,7). 5. Izračunajte konačni pH otopine ako ste u 250 mL 0,02 M Tris/Cl pufera pH 8,1
dodali 0,1 mL 1 M HCl (zanemarite promjenu volumena) (pKa = 8,1). 6. U svrhu pročišćavanja određenog proteina, potreban vam je 0,1 M glicinski pufer
pH = 9,2. Nažalost, ponestalo vam je glicina u zalihama, ali ste uspjeli pronaći dvije 0,1 M puferske otopine glicina, jednu s pH = 9,0 i drugu s pH = 9,5. Koji volumen svake od ovih dviju otopina morate pomiješati kako biste dobili 100 ml željenog pufera?
1.9 Literatura
1. Voet, D. and Voet, J. (1995) Biochemistry. New York: John Wiley & Sons Inc. 2. Garrett, R.H. and Grisham, C.M. (1999) Biochemistry. Canada: Brooks/Cole
Cengage Learning.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
10
Vježba 2.
2.1 Gel-filtracija
Jedna od osnovnih zadaća preparativne i analitičke biokemije je odjeljivanje i identifikacija bioloških molekula. Za odjeljivanje se često koriste kromatografske metode kod kojih se smjesa tvari otopljena u pokretnoj fazi (koja može biti tekućina ili plin) razdvaja na sastavnice propuštanjem kroz stacionarnu fazu (najčešće čvrsti polimer). Tekućinska kromatografija u koloni podrazumijeva pokretnu fazu (otapalo) i stacionarnu fazu u staklenoj, plastičnoj ili metalnoj koloni. Punjenje kolone, odnosno zrnca kromatografskog materijala, mogu biti inertna ili mogu stupati u interakciju s analitom. Kod kromatografije ionske izmjene, kromatografije obrnute faze i afinitetne kromatografije pojedine komponente ostavaruju interakcije sa zrncima gela ovisno o svom naboju, polarnosti ili afinitetu za ligand te se tako odjeljuju od ostalih komponenti iz uzorka. Suprotno tome, kolona za gel-filtraciju sazdana je od inertnih zrnaca koja ne stupaju u interakciju s analitom, odnosno ne vežu se, ne reagiraju s uzorkom, niti nose električni naboj. Ovdje se razdvajanje temelji na različitoj brzini putovanja molekula ovisno o njihovoj veličini.
U osnovnoj izvedbi, aparaturu za kromatografiju čini samo kolona punjena kromatografskim materijalom. Uzorak se nanosi na kolonu koja se potom ispire puferom, a sadržaj i koncentracija pojedine tvari u sakupljenim se frakcijama određuje naknadno. Između zrnaca gela nalazi se intersticijska tekućina, a unutar njih prostor ispunjen gel-tekućinom. Kod gel-filtracije velike molekule ne mogu prodrijeti kroz pore i ući u zrnca gela pa brzo putuju intersticijskom tekućinom, dok male molekule prodiru u zrnca usporavajući svoje putovanje (Slika 2.1).
Slika 2.1 Prikaz molekularnog prosijavanja tijekom gel-filtracije.
Gel-filtracija hemoglobina
male molekule
velike molekule
porozna zrnca gela (stacionarna faza)

BIOLOŠKA KEMIJA — PRAKTIKUM
11
Ukupni volumen tekućine u koloni jednak je zbroju volumena pufera izvan i pufera unutar zrnaca gela. Volumen dostupan pojedinoj molekuli obrnuto je razmjeran njenoj veličini, odnosno vrlo velike molekule putuju kroz pufer izvan zrnaca gela i prve izlaze iz kolone, a za njima kasnije izlaze manje molekule koje putuju i kroz pufer izvan zrnaca gela i kroz pufer unutar zrnaca. Količina pufera potrebna da neka molekula izađe iz kolone naziva se volumen ispiranja. Raspon masa molekula koje je moguće odijeliti određenim kromatografskim materijalom za gel-filtraciju određen je veličinom njegovih pora. No pored veličine pora, njegovo je važno svojstvo i veličina zrnaca gela (krupna, srednja, sitna i vrlo sitna); što su zrnca krupnija pakiranje kromatografskog materijala je rjeđe pa je brzina protoka veća, a time je separacija uzorka slabija. Stoga se krupna zrnca primjenjuju za velike preparacije kod kojih je brzina važnija od razlučivanja, sitna zrnca za većinu preparativnih postupaka, a vrlo sitna zrnca koriste se u analitičke svrhe kada je potrebno maksimalno razlučivanje.
Zrnca za gel-filtraciju su umreženi polimeri, a razlikujemo tri osnovna tipa: dekstran, agarozu i poliakrilamid. Dekstran je polisaharid sastavljen od monomera glukoze. Sama zrnca kao i pore u njima mogu biti različite veličine. Agaroza je linearni polimer D-galaktoze i 3,6-anhidro-L-galaktoze umrežen vodikovim vezama. Pore na zrncima su mnogo veće nego kod dekstrana, što je korisno za odjeljivanje velikih proteina ili dugih linearnih molekula (DNA, RNA). Poliakrilamidne gelove čine umreženi akrilamid i N,N-metilenbisakrilamid. Po svojstvima pri odjeljivanju slični su dekstranu, no dostupni su u širem rasponu veličine pora.
2.2 Hem i hemoglobin
Hemoglobin (Hb) je izgrađen od proteinskog i neproteinskog dijela. Proteinski dio hemoglobina sastoji se od četiri polipeptidna lanca povezana nekovalentnim interakcijama (kvarterna struktura), a na svaki globinski lanac vezana je po jedna molekula hema koji veže kisik. Hemoglobin A, najzastupljeniji hemoglobin u krvi odraslih osoba, sastoji se od dva α-lanca i dva β-lanca (α2β2). U krvi odraslih osoba postoji i manje zastupljen hemoglobin A2 (2 α-lanca i 2 δ-lanca, α2δ2), dok su tijekom intrauterinog života prisutni embrijski (2 α-lanca i 2 ε-lanca, α2ε2) i fetalni oblici (2 α-lanca i 2 γ-lanca, α2γ2). Molekula hemoglobina gotovo je kuglasta, a četiri proteinska lanca tetraedarski su raspoređena.
Svaka podjedinica hemoglobina sadrži po jedan hem, prostetičku skupinu smještenu u procjepu blizu površine molekule. To je specifična, neproteinska komponenta neophodna za biološku aktivnost hemoglobina. Hem je izgrađen od organskog dijela, protoporfirina IX, u čijem je središtu atom željeza koordinativnim vezama povezan sa četiri atoma dušika iz pirolovih prstena protoporfirina (Slika 2.2). Kako je atom željeza heksakoordiniran u hemu, peto koordinativno mjesto uvijek popunjava elektronski par dušika iz histidinskog ogranka polipeptidnog lanca (His93), dok šesto mjesto može biti prazno ili popunjeno. Hemoglobin, kod kojeg je šesto koordinativno mjesto prazno, naziva se deoksihemoglobin, tamno crveni je oblik Hb, prisutan u venskoj krvi. Kada se putem šestog koordinativnog mjesta veže kisik (oksigenacija) nastaje svijetlo crveni oksihemoglobin (HbO) koji nalazimo u arterijskoj krvi. Ukoliko se hemoglobinsko željezo oksidira iz Fe2+ oblika u trovalentni oblik, Fe3+, nastaje methemoglobin koji je

BIOLOŠKA KEMIJA — PRAKTIKUM ___
12
smeđe boje. Ovaj oblik ima veliki afinitet vezanja negativnih iona, ali i neutralnih molekula s izraženim dipolnim momentom, pa umjesto kisika veže vodu.
Slika 2.2. Graña hemoglobina. A) Tetramerna struktura hemoglobina; B) Aktivno mjesto hemoglobina: Fe-atom leži u ravnini u kojoj su uspostavljene četiri koordinativne veze s atomima porfirinskog dušika. Preostale dvije koordinativne veze povezuju ogranak histidina i molekularni kisik; C) Struktura Fe-protoporfirina IX. 2.2.1 Priprema otopine deoksihemoglobina
Krv se izvadi venepunkcijom u vacutainer-epruvetu koja sadrži antikoagulans. Vacutainer-epruvete mogu biti različitih volumena i namjena. Epruvete koje sadrže K2EDTA ili K3EDTA kao antikoagulans standardno su označene ljubičastim čepom. Prilikom vađenja krvi zbog razlike u tlakovima u epruvetu ulazi točno određeni volumen krvi tako da je konačna koncentracija EDTA 1,8 mg/mL krvi. Nakon vađenja, krv se centrifugira 10 minuta pri 600g i 4°C kako bi se razdvojili eritrociti (donji crveni sloj) od plazme (gornji prozirno žuti sloj). Između navedena dva sloja čiji su volumeni približno jednaki nalazi se vrlo tanak sloj leukocita i tromobocita (bijele boje, poznat pod engleskim nazivom buffy coat). Plazma i leukociti s tromobocitima se otpipetiraju i bace. Eritrociti se potom tri puta isperu fiziološkom otopinom. Ispiranje se provodi tako da se eritrocitima doda jednaki volumen fiziološke otopine, epruveta se lagano promućka i ponovno centrifugira 10 minuta pri 600g i 4°C. Supernatant se baci i postupak pranja se ponovi još dva puta. Potom se eritrocitima doda 7 volumena hipotonične otopine, odnosno destilirane vode. Ubrzo nakon miješanja dolazi do hemolize. Nakon centrifugiranja 10 minuta pri 13000g i 4°C na dnu epruvete se nalazi debris stanica, a supernatant koji predstavlja uzorak otopine deoksihemoglobina se otpipetira i pohrani na -20°C do daljnje uporabe.
2.3 Zadaća
Priredite otopinu methemoglobina dodatkom kalijevog fericijanida u otopinu hemoglobina. Gel-filtracijom razdvojite smjesu feri/ferocijanida i methemoglobina. Protein će daljnjim prolazom kroz gel reagirati s prethodno unijetim Fe(II) reagensom uslijed čega će se reducirati u deoksihemoglobin, a zatim oksigenirati otopljenim
A) B) C)

BIOLOŠKA KEMIJA — PRAKTIKUM
13
molekularnim kisikom. Postupna zbivanja tijekom prolaza proteinske frakcije kroz kolonu pratit će karakteristične promjene boje.
2.4 Reagencije
• 0,02 M natrijev fosfatni pufer, pH 7
• 80 mM FeSO4 u 0,2 M natrijevom fosfatnom puferu, pH 7
• 80 mM EDTA u 0,2 M natrijevom fosfatnom puferu, pH 7 • uzorak (otopina deoksihemoglobina) u dest. vodi • kalijev fericijanid • Sephadex G-25
2.5 Pribor
• staklena kolona, 10 cm, φ=1 cm • gumeni nastavak za kolonu sa stezaljkom • parafilm • stalak sa stezaljkom • staklena čaša od 100 mL • staklena boca • Pasteurova pipeta • plastične epruvete od 1,5 mL • plastične epruvete od 15 mL • stalak za epruvete • špatula • automatske pipete • nastavci za automatske pipete • boce štrcalice
2.6 Načelo postupka
Gel-filtracijom se smeđi methemoglobin razdvaja od puno manjih iona kalija, fericijanida i ferocijanida. Unutar kolone u koju je prethodno unijet reducens u potpunosti se odvija redukcija methemoglobina u deoksihemoglobin. Daljnjim prolaskom deoksihemoglobina kroz kolonu ispunjenu puferom deoksihemoglobin se oksigenira u oksihemoglobin.
2.7 Postupak
• Stezaljkom podesite protok pufera kroz staklenu kromatografsku kolonu u staklenu čašu na 30 kapi u minuti. Prekinite istjecanje kada se razina pufera spusti do površine zrnaca gela.
• Uzorak (otopinu deoksihemoglobina) razrijedite u epruveti puferom u volumnom omjeru 1+4 (70 μL krvi + 280 μL pufera). Dodajte sasvim mali kristal fericijanida i protresite epruvetu.
• U drugoj epruveti pomiješajte 0,1 mL otopine FeSO4 i 0,1 mL otopine EDTA. • Kako biste dobili oštre vrpce slojeva u koloni nužno je pažljivo raditi i prilikom
nanošenja uzoraka na kolonu ne uzmutiti površinski sloj gela. Na kolonu prvo nanesite 0,2 mL svježe priređene smjese FeSO4/EDTA u fosfatnom puferu,

BIOLOŠKA KEMIJA — PRAKTIKUM ___
14
polagano otpuštajte stezaljku da sav dodani volumen uđe u kolonu te stezaljku ponovno pritegnite.
• Na kolonu nanesite oko 1 mL (25 kapi) pufera te otpuštajući stezaljku omogućite da sav pufer uđe u kolonu.
• Potom na kolonu nanesite 350 μL razrijeđene krvi te otpuštanjem stezaljke omogućite da sav uzorak uđe u kolonu.
• Nastavite na kolonu dodavati pufer i ostvarite kontinuirani protok (oko 30 kapi u minuti) kroz kolonu tako da otpustite stezaljku. Pazite da kolona nikada ne ostane bez nadsloja pufera!
• Pratite promjene boje slojeva u koloni. U epruvete od 1,5 mL sakupljajte frakcije svijetlo-crvene odnosno žute boje prilikom njihovog izlaska iz kolone.
2.8 Zadaci
1. Usporedite gel filtraciju i dijalizu. Navedite prednosti i mane pojedine tehnike odjeljivanja molekula.
2. Obilježili ste velik segment DNA radioaktivnim nukleotidima. Koji ćete kromatografski materijal upotrijebiti za razdvajanje molekula DNA od slobodnih nukleotida?
3. Ima li razlike između oksidacije i oksigenacije hemoglobina? Objasnite! 4. Zašto se kisik labilno veže za prostetičku skupinu hemoglobina?
2.9 Literatura
1. Wood, E.J. (1989) Practical Biochemistry for Colleges. Pergamon Press. 2. Dixon, H.B.F. and McIntosh, R. (1967) Reduction of Methaemoglobin in
Haemoglobin Samples using Gel Filtration for Continuous Removal of Reaction Products. Nature 213: 399-400.
3. Voet, D. and Voet, J.g. (1995) Biochemistry. John Wiley & Sonc Inc. 4. Garrett, R.H. and Grisham, C.M. (1999) Biochemistry. Brooks Cole. 5. http://www.aw-bc.com/mathews/ch07/heme.htm

BIOLOŠKA KEMIJA — PRAKTIKUM
15
Vježba 3.
3.1 Humani serum i imunoglobulin G (IgG)
Krv izvađena venepunkcijom bez dodatka antikoagulansa zgrušava se stajanjem na zraku posredstvom sustava za koagulaciju. Centrifugiranjem se ugrušak odvaja od supernatanta koji nazivamo krvnim serumom. Serum je, dakle, dio krvi bez čimbenika zgrušavanja, fibrinogena i krvnih stanica; možemo ga smatrati puferiranom otopinom koja sadrži ione, organske tvari (urea, kreatinin, bilirubin), biološke makromolekule (proteini, lipoproteini, DNA). Proteini čine velik dio suhe tvari seruma (60-80 g/L).
Značajan dio proteinske frakcije seruma čine imunoglobulini (7-16 g/L), koji djeluju kao obrambeni proteini, pa ih nazivamo i protutijela. Stvara ih imunosni sustav kralježnjaka kao odgovor na strane tvari (antigene) koje mogu prodrijeti u organizam. Protutijela prepoznaju specifičan antigen, vežu se na njega i posreduju u aktivaciji imunosnog sustava. Humani serum sadrži pet klasa imunoglobulina (IgA, IgD, IgE, IgG i IgM), među kojima je najzastupljeniji IgG.
Imunoglobulini su heterodimeri koji se sastoje od dva laka i dva teška lanca. Teški su lanci specifični za pojedinu klasu protutijela (α – IgA, δ – IgD, ε – IgE, γ – IgG, μ – IgM). Lanci su međusobno povezani disulfidnim mostovima između prostorno bliskih cisteinskih ostataka. Na molekuli funkcionalno razlikujemo konstantni i varijabilni dio (Slika 3.1). Varijabilni dio specifično prepoznaje i veže antigen, dok konstantni dio posreduje u aktivaciji stanica imunosnog sustava. Kombiniranjem različitih amino-kiselina u varijabilnom dijelu, imunosni sustav posredstvom protutijela može prepoznati milijune različitih stranih molekula u organizmu. 3.2 Metode pročišćavanja proteina
Proces pročišćavanja proteina rijetko se odvija u samo jednom koraku. Najčešće se radi o nizu postupaka, a svakim od njih odjeljuju se pojedine vrste molekula određenom učinkovitošću. Pravilnim odabirom metoda pročišćavanja i njihovim primjerenim slijedom, može se iz smjese proteina izdvojiti točno određeni protein.
3.2.1 Taloženje (isoljavanje) amonijevim sulfatom
Taloženje amonijevim sulfatom omogućava razdvajanje proteina na temelju njihove različite topljivosti pri određenoj koncentraciji amonijevog sulfata u otopini. Metoda se temelji na činjenici da u vodenoj otopini molekule vode koje okružuju proteinske
Pročišćavanje imunoglobulina G iz humanog seruma
Slika 3.1 Shematski prikaz molekule. Ig.
vari
jabi
lni
dio
kons
tant
ni
dio
laki lanac
teški lanac

BIOLOŠKA KEMIJA — PRAKTIKUM ___
16
molekule poprimaju uređenu strukturu. Premda točan mehanizam nije poznat, smatra se da povećanjem koncentracije amonijevog sulfata u otopini dolazi do kompeticije između amonijevih i sulfatnih iona i proteina za slobodne molekule vode što dovodi do agregacije i precipitacije proteina.
Možemo reći da amonijev sulfat oduzima vodu otopljenim proteinima te oni stoga stvaraju nakupine i talože se. Za odvajanje je ključna empirijski utvrđena činjenica da se određeni protein taloži u uskom rasponu koncentracija amonijevog sulfata. Na slici 3.2 prikazana su dva proteina s različitim omjerom hidrofilnih i hidrofobnih skupina na površini molekule. Jasno je da će se protein s lijeve strane slike taložiti pri manjoj koncentraciji amonijevog sulfata (jer ima manje vodi izloženih hidrofilnih skupina, odnosno okružen je s manje molekula vode). Istaložene proteine moguće je centri-fugiranjem razdvojiti od proteina zaostalih u otopini.
3.2.2 Kromatografija ionske izmjene
Kromatografija ionske izmjene provodi se na koloni ionskog izmjenjivača koja se sastoji od čestica gela s kovalentno vezanim nabijenim skupinama. Funkcionalne skupine kationskih izmjenjivača negativno su nabijene, pa elektrostatskim silama vežu katione otopljene u mobilnoj fazi (Slika 3.3 A). Suprotno tome, anionski izmjenjivač s pozitivno nabijenim funkcionalnim skupinama veže anione iz mobilne faze (Slika 3.3 B). U ovoj se vježbi koristi anionski izmjenjivač trimetilaminoetan koji je jaki kation (otporan na promjene pH od 2-14).
A) B)
Slika 3.3 Shematski prikaz zrnaca gela A) kationskog i B) anionskog izmjenjivača.
3.3 SDS-poliakrilamidna gel-elektroforeza
Elektroforeza je metoda razdvajanja molekula pod utjecajem električnog polja na temelju razlika u njihovoj masi i naboju. Različiti proteini imaju različit ukupni naboj koji potječe od bočnih ogranaka aminokiselina na površini molekule proteina. U SDS-poliakrilamidnoj gel elektroforezi, u gelu i u puferu za nanošenje uzoraka, prisutan je anionski detergent natrijev dodecil-sulfat (SDS, od engl. sodium dodecyl sulphate). SDS se
Slika 3.2 Dva proteina s različitim omjerom hidrofilnih (crno) i hidrofobnih (bijelo) skupina.
0
0

BIOLOŠKA KEMIJA — PRAKTIKUM
17
veže za proteine u uzorku, maskira njihov površinski naboj i oni postaju negativno nabijeni. Količina negativno nabijenog detergenta koja će se vezati na protein razmjerna je njegovoj veličini (1,4 g SDS/g proteina) pa putovanje proteina u električnom polju u ovoj vrsti elektroforeze ovisi isključivo o njegovoj molekulskoj masi (s obzirom na to da je omjer naboja i mase konstantan).
Proteine razdvajamo diskontinuiranom elektroforezom uz uporabu sabijajućeg i razdvajajućeg gela koji se međusobno razlikuju u pH i gustoći (Slika 3.4). Proteini se prolaskom kroz sabijajući gel (3-5%) pri pH 6,8 koncentriraju u usku vrpcu te svi zajedno ulaze u razdvajajući gel (10-15%) u kojem je pH 8,8. Brzina putovanja u razdvajajućem gelu uvjetovana je učinkom molekulskog prosijavanja, odnosno manji proteini putuju brže, a veći sporije. Uzorke pripremamo dodatkom pufera za nanošenje uzoraka koji sadrži SDS, β-merkaptoetanol, glicerol i boju bromfenol plavo u puferu pH 6,8. β-merkaptoetanol kida disulfidne mostove, SDS denaturira proteine uz kuhanje pri 95°C, glicerol povećava gustoću uzorka te se oni tako lakše nanose u jažice, a boja bromfenol plavo služi za praćenje tijeka elektroforeze, ali ne boja proteine.
Nakon provedene elektroforeze, proteine vizualiziramo bojanjem gela bojom koja specifično boja proteine, Coomassie blue® R-250, otopljenoj u metanolu i octenoj kiselini. Višak boje s gela uklanjamo ispiranjem u otopini octene kiseline. 3.4 Odreñivanje prisutnosti i koncentracije proteina mjerenjem apsorbancije
Proteini apsorbiraju svjetlost valne duljine 280 nm zbog prisutnosti aromatskih prstenova na bočnim ograncima aminokiselina fenilalanina, triptofana ili tirozina u svojoj strukturi. Temeljem Beer-Lambertovog zakona (A = ε x l x c, pri čemu je ε molarni ekstinkcijski koeficijent, l duljina puta zrake svjetlosti kroz uzorak, a c koncentracija tvari koja apsorbira) može se zaključiti da je odnos vrijednosti apsorbancije i koncentracije proteina linearan. Međutim, linearan odnos vrijedi samo unutar određenog raspona koncentracija proteina. Osim toga, valja uočiti da se mjerenjem apsorbancije može odrediti koncentracija otopine čistog proteina, a ne smjese proteina, jer se proteini razlikuju po molarnom ekstinkcijskom koeficijentu. Za određivanje koncentracije određenog proteina mjerenjem apsorbancije potrebno je stoga ili poznavati njegov molarni ekstinkcijski koeficijent ili konstruirati baždarni pravac koji odražava odnos poznatih koncentracija tog proteina i apsorbancija.
Slika 3.4 Ureñaj za diskontinuiranu SDS-
PAGE. Slika preuzeta i modificirana iz (6).

BIOLOŠKA KEMIJA — PRAKTIKUM ___
18
3.5 Zadaća
Iz uzorka humanog seruma izolirajte imunoglobulin G. Provjerite čistoću pročišćenog proteina SDS-poliakrilamidnom gel elektroforezom te odredite koncentraciju izoliranog proteina. 3.6 Reagencije
• 3,5 M amonijev sulfat u dest. vodi • 10 mM natrijev fosfatni pufer, pH 6,8 • 2 M NaCl u dest. vodi • pufer za nanošenje uzoraka: 187,5 mM Tris/HCl pH 6,8; 6% SDS, 20% glicerol, 5%
β-merkaptoetanol, 0,1% bromfenol plavo u dest. vodi • 1% agaroza u dest. vodi • gel za sabijanje: 30% akrilamid/0,8% bisakrilamid, 1,0 M Tris HCl, pH 6,8, 10%
SDS, 10% amonijev persulfat u dest. vodi • TEMED • gel za razdvajanje: 30% akrilamid/0,8% bisakrilamid, 1,5 M Tris HCl, pH 8,8, 10%
SDS, 10% amonijev persulfat u dest. vodi • pufer za elektroforezu: 192 mM glicin, 3,5 mM SDS, 25 mM Tris HCl, pH 8,3, u
dest. vodi • otopina za bojanje gela (0,1% Coomassie blue® R-250, 45% metanol, 10% ledena
octena kiselina u dest. vodi) • otopina za odbojavanje gela: (10% octena kiselina u dest. vodi) • komercijalno pribavljena smjesa proteina poznatih molekulskih masa • komercijalno pribavljen imunoglobulin G
3.7 Pribor
• automatske pipete • nastavci za automatske pipete • plastične epruvete od 1,5 i 2 mL • stalak za epruvete • stalak sa stezaljkom • staklene boce • flomaster za označavanje • stolna centrifuga • plastične kolone, 10 cm, φ = 1 cm • gel Sephadex G-25 • Fractogel® TMAE-650 (anionski izmjenjivač trimetilaminoetan) • plastične kapaljke • peristaltička pumpa • plastične cijevi s odgovarajućim priključcima • čaše od 100 mL • termostatirana tresilica • mješač Vortex • digestor

BIOLOŠKA KEMIJA — PRAKTIKUM
19
• češalj i razmaknice za elektroforezu • stakla za elektroforezu • štipaljke • kadica za elektroforezu • žice s priključcima • izvor istosmjerne struje (ispravljač, eng. power supply) • kvarcna kiveta • staničevina • boce štrcalice • UV/VIS spektrofotometar
3.8 Načelo postupka
Dio proteina humanog seruma, među kojima i IgG, taloži se pri određenoj koncentraciji amonijevog sulfata (2,37 M), a talog se odvaja od topljivih proteina centrifugiranjem. Nakon otapanja istaloženih proteina u fosfatnom puferu zaostali se amonijev sulfat odvaja od proteinske frakcije gel-filtracijom. Na kolonu anionskog izmjenjivača pri određenim uvjetima (pH 6,8) vežu se svi ostali proteini osim IgG, a isti se s kolone ispiru povećanjem ionske jakosti. Prisutnost proteina u sakupljenim frakcijama utvrđuje se na temelju porasta apsorbancije mjerene pri valnoj duljini od 280 nm. Prisutnost i čistoća IgG u pojedinim frakcijama provjerava se elektroforetskim razdvajanjem proteina sakupljenih frakcija u poliakrilamidnom gelu u denaturirajućim uvjetima, pri čemu se polipeptidni lanci razdvajaju na temelju razlike u svojoj masi te vizualiziraju bojom koja se veže na proteine. Koncentracija čistog IgG u pojedinim frakcijama određuje se na temelju apsorbancije tih frakcija i apsorbancija otopina poznatih koncentracija IgG (uz konstruiranje baždarnog pravca) pri valnoj duljini 280 nm.
3.9 Postupak
3.9.1 Isoljavanje amonijevim sulfatom
• U 200 μL humanog seruma dodajte 400 μL 3,5 M otopine amonijevog sulfata, tako da njegova konačna koncentracija bude 2,3 M. Začepite epruvetu i promiješajte uzorak.
• Uzorak centrifugirajte 5 minuta pri 5400g te potom supernatant pipetom odbacite u čašu za otpad.
• Preostali talog resuspendirajte u početnom volumenu (200 μL) 10 mM fosfatnog pufera pH 6,8.
3.9.2 Odsoljavanje gel-filtracijom (princip metode objašnjen je u Vježbi 2.)
• Iz kolone za gel-filtraciju ispustite višak fosfatnog pufera u čašu za otpad. Nadsloj pufera mora biti u razini površinskoj sloja gela.
• Uzorak (resuspendirani talog nakon taloženja amonijevim sulfatom) nanesite na kolonu za gel-filtraciju i istovremeno u epruvetu od 1,5 mL počnite sakupljati tekućinu koja istječe. Kada uzorak uđe u kolonu na vrh kolone dodajte još pufera. Kolona nikad ne smije ostati bez nadsloja tekućine.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
20
• Nakon što sakupite prvih 1 mL tekućine koja istječe iz kolone, tu frakciju bacite, a sljedeća 2 mL koja istječu iz kolone nastavite sakupljati u epruvetu od 2 mL. Taj uzorak sačuvajte za sljedeći korak pročišćavanja (anionsku izmjenu).
• Kako bi kolonu za gel-filtraciju isprali od amonijevog sulfata, propustite kroz nju 30 mL pufera koji po izlasku s kolone sakupljajte u čašu za otpad i potom bacite.
3.9.3 Kromatografija ionske izmjene
• Označite i priredite na stalku 20 epruveta volumena 1,5 mL za sakupljanje uzoraka. Pokrenite peristaltičku pumpu i namjestite brzinu protoka pufera na 0,5 mL/min (oznaka 5 na pumpi Pharmacia P-3). Ispustite pufer iznad zrnaca gela u čašu za otpad.
• Nanesite uzorak na kolonu anionskog izmjenjivača i odmah počnite sakupljati uzorke eluensa od po 1 mL. Sakupite 10 frakcija, a po potrebi dodajte još pufera u kolonu.
• Kolonu zatim isperite s 10 mL 2 M NaCl uz protok od 0,8 mL/min (oznaka 8 na peristaltičkoj pumpi) te sakupite 10 frakcija volumena 1 mL.
• Uravnotežite kolonu ispiranjem s 30 mL 10 mM fosfatnog pufera koji nakon izlaska iz kolone sakupljajte u čaši za otpad.
3.9.4 Odreñivanje prisutnosti proteina mjerenjem apsorbancije pri λ = 280 nm
• Kvarcnu kivetu u koju ste ulili 10 mM fosfatni pufer stavite u spektrofotometar te apsorbanciju pri λ = 280 nm postavite na vrijednost 0.
• Ispraznite kivetu, isperite je destiliranom vodom i protresite kako biste iz nje uklonili svu tekućinu.
• U kivetu ulijte pojedinu frakciju sakupljenu tijekom anionske izmjene te očitajte apsorbanciju te frakcije pri λ = 280 nm. (Postupak mjerenja ponovite na isti način za sve frakcije.)
• Prikažite dobivene rezultate grafički na milimetarskom papiru nanoseći na apscisu broj sakupljene frakcije, a na ordinatu A280.
3.9.5 Elektroforeza proteina u diskontinuiranom denaturirajućem poliakrilamidnom gelu
• Razrijedite uzorak seruma 50 puta te pomiješajte 10 µL seruma s 5 µL pufera za nanošenje uzoraka (187,5 mM Tris/HCl pH 6,8; 6% SDS, 20% glicerol, 5% β-merkaptoetanol, 0,1% bromfenol plavo).
• Pomiješajte 2 µL supernatanta nakon taloženja proteina seruma amonijevim sulfatom s 8 µL vode i 5 µL pufera za nanošenje uzoraka.
• Frakciju proteina sakupljenu nakon gel-filtracije seruma razrijedite 10 puta te pomiješajte 10 µL razrijeđene frakcije s puferom za nanošenje uzoraka u omjeru 2:1.
• Pomiješajte 3 µL pročišćene frakcije IgG sa 7 µL vode i 5 µL pufera za nanošenje uzoraka.
• Pomiješajte 10 µL pufera koji je s kolone izlazio nakon imunoglobulina G, a prije proteina, s puferom za nanošenje uzoraka u omjeru 2:1.
BIOLOŠKA KEMIJA — PRAKTIKUM
21
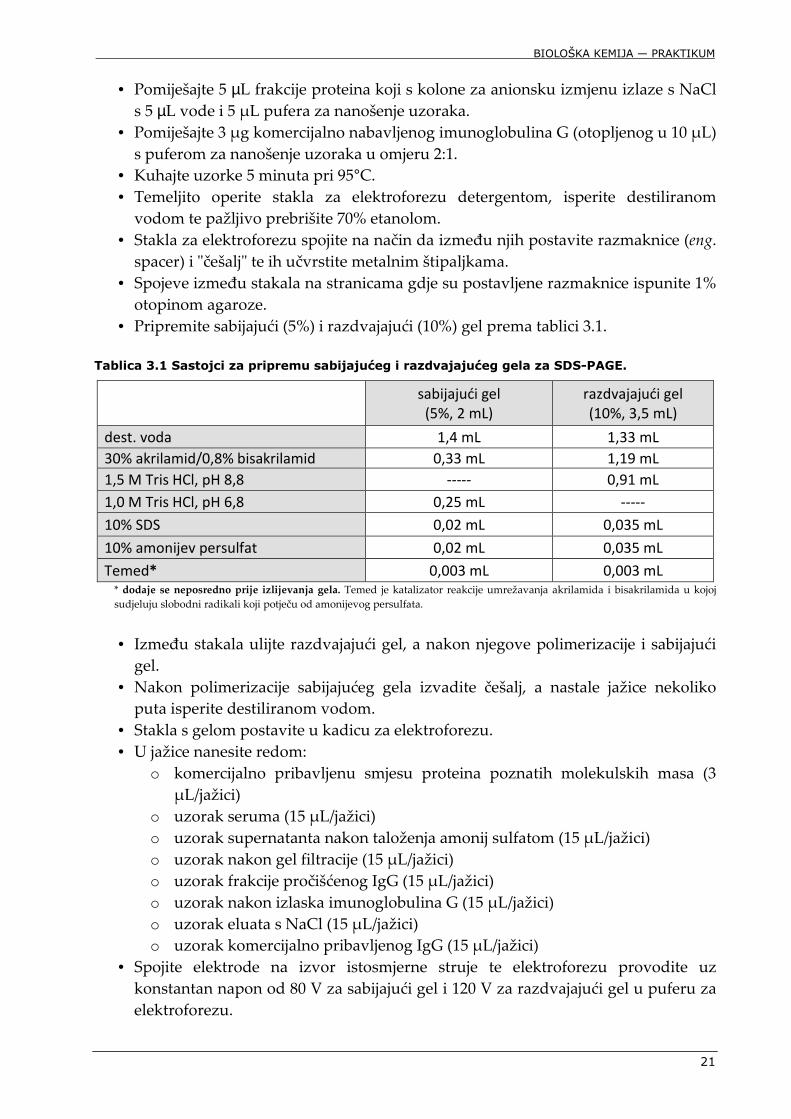
• Pomiješajte 5 µL frakcije proteina koji s kolone za anionsku izmjenu izlaze s NaCl s 5 µL vode i 5 μL pufera za nanošenje uzoraka.
• Pomiješajte 3 μg komercijalno nabavljenog imunoglobulina G (otopljenog u 10 μL) s puferom za nanošenje uzoraka u omjeru 2:1.
• Kuhajte uzorke 5 minuta pri 95°C. • Temeljito operite stakla za elektroforezu detergentom, isperite destiliranom
vodom te pažljivo prebrišite 70% etanolom. • Stakla za elektroforezu spojite na način da između njih postavite razmaknice (eng.
spacer) i "češalj" te ih učvrstite metalnim štipaljkama. • Spojeve između stakala na stranicama gdje su postavljene razmaknice ispunite 1%
otopinom agaroze. • Pripremite sabijajući (5%) i razdvajajući (10%) gel prema tablici 3.1.
Tablica 3.1 Sastojci za pripremu sabijajućeg i razdvajajućeg gela za SDS-PAGE.
sabijajući gel
(5%, 2 mL)
razdvajajući gel
(10%, 3,5 mL)
dest. voda 1,4 mL 1,33 mL
30% akrilamid/0,8% bisakrilamid 0,33 mL 1,19 mL
1,5 M Tris HCl, pH 8,8 ----- 0,91 mL
1,0 M Tris HCl, pH 6,8 0,25 mL -----
10% SDS 0,02 mL 0,035 mL
10% amonijev persulfat 0,02 mL 0,035 mL
Temed* 0,003 mL 0,003 mL * dodaje se neposredno prije izlijevanja gela. Temed je katalizator reakcije umrežavanja akrilamida i bisakrilamida u kojoj sudjeluju slobodni radikali koji potječu od amonijevog persulfata. • Između stakala ulijte razdvajajući gel, a nakon njegove polimerizacije i sabijajući
gel. • Nakon polimerizacije sabijajućeg gela izvadite češalj, a nastale jažice nekoliko
puta isperite destiliranom vodom. • Stakla s gelom postavite u kadicu za elektroforezu. • U jažice nanesite redom:
o komercijalno pribavljenu smjesu proteina poznatih molekulskih masa (3 μL/jažici)
o uzorak seruma (15 μL/jažici) o uzorak supernatanta nakon taloženja amonij sulfatom (15 μL/jažici) o uzorak nakon gel filtracije (15 μL/jažici) o uzorak frakcije pročišćenog IgG (15 μL/jažici) o uzorak nakon izlaska imunoglobulina G (15 μL/jažici) o uzorak eluata s NaCl (15 μL/jažici) o uzorak komercijalno pribavljenog IgG (15 μL/jažici)
• Spojite elektrode na izvor istosmjerne struje te elektroforezu provodite uz konstantan napon od 80 V za sabijajući gel i 120 V za razdvajajući gel u puferu za elektroforezu.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
22
• Kada boja iz uzorka (bromfenol plavo) dosegne donji rub razdvajajućeg gela prekinite tok električne struje.
• Razdvojite stakla, uronite gel u otopinu za bojanje te sve zajedno zagrijavajte 45 sekundi pri maksimalnoj jakosti u mikrovalnoj pećnici. Potom ostavite gel u otopini za bojanje na zibaljci tijekom sljedećih 15 minuta.
• Izlijte otopinu za bojanje iz kadice u kojoj se nalazi gel te kadicu s gelom kratko isperite destiliranom vodom. Potom u nju dodajte otopinu za odbojavanje koju zatim zagrijte tijekom 45 sekundi u mikrovalnoj pećnici pri najvećoj jakosti. Nastavite odbojavanje gela tijekom sljedećih 15 minuta na zibaljci.
• Utvrdite u kojim se frakcijama nalazi IgG i je li pročišćen od ostalih proteina seruma.
3.7.6 Odreñivanje koncentracije pročišćenog imunoglobulina G
Očekivane vrijednosti koncentracije IgG u sakupljenim frakcijama kreću se od 0,1 mg/mL do 2 mg/mL pa se stoga priređuju standardne otopine IgG koncentracija: 0,02; 0,06; 0,10; 0,40; 0,80; 1,20; 1,60; i 2,00 mg/mL. Otopine se priređuju u duplikatu razrjeđivanjem matične otopine IgG koncentracije 2 mg/mL u 10 mM fosfatnom puferu. Kao referentna vrijednost uzima se prosječna izmjerena vrijednost A280 za pojedinu koncentraciju IgG.
• Nacrtajte na milimetarskom papiru baždarni pravac nanoseći na apscisu koncentracije IgG iz tablice 3.1, a na ordinatu pripadajuće prosječne vrijednosti A280.
• Pomoću baždarnog pravca odredite koncentraciju IgG u nepoznatom uzorku (u frakciji koja sadrži najviše IgG).
Tablica 3.2 Eksperimentalno utvrñene vrijednosti apsorbancije različitih koncentracija IgG pri svjetlosti valne duljine 280 nm.
konačna konc.
IgG
V (2 mg/mL
otopine IgG)
V (10 mM
fosfatnog pufera) A1 A2 A
0 mg/mL 0 mL 1 mL 0,002 -0,002 0,000
0,02 mg/mL 0,01 mL 0,99 mL 0,022 0,026 0,024
0,06 mg/mL 0,03 mL 0,97 mL 0,063 0,062 0,063
0,10 mg/mL 0,05 mL 0,95 mL 0,104 0,129 0,117
0,40 mg/mL 0,20 mL 0,80 mL 0,440 0,442 0,441
0,80 mg/mL 0,40 mL 0,60 mL 0,899 0,889 0,894
1,20 mg/mL 0,60 mL 0,40 mL 1,351 1,359 1,355
1,60 mg/mL 0,80 mL 0,20 mL 1,849 1,853 1,851
2,00 mg/mL 1,00 mL 0,00 mL 2,291 2,296 2,294
3.8 Zadaci
1. Koja je razlika između krvnog seruma i krvne plazme? 2. Koja je funkcija varijabilnog dijela, a koja konstantnog dijela imunoglobulina G? 3. Koji dio proteinske strukture apsorbira svjetlost valne duljine 280 nm? 4. SDS-poliakrilamidnom elektroforezom analizirali ste smjesu proteina i nakon
bojanja gela bojom Coomassie na gelu vidite tri vrpce. Možete li na osnovu toga sa

BIOLOŠKA KEMIJA — PRAKTIKUM
23
sigurnošću reći je li uzorak sadržavao tri različita proteina? Objasnite svoj odgovor!
5. Protein A je homodimer s molekulskom masom monomera 42 kDa, protein B je heterodimer koji se sastoji od polipeptida molekulske mase 20 kDa i 30 kDa, dok je protein C monomeran protein molekulske mase 14 000 Da. Nacrtajte shematski prikaz SDS-PAGE gela u kojem je svaki protein pušten kao zasebni uzorak. Ne zaboravite obilježiti anodu i katodu te molekulske mase pojedinih proteinskih vrpci.
3.9 Literatura
1. Gornik, I., Maravić, G., Dumić, J., Flögel, M. and Lauc, G. (1999) Fucosylation of IgG Heavy Chains is Increased in Rheumatoid Arthritis. Clinical Biochemistry. 32: 605-608.
2. Aurer, I., Lauc, G., Dumić, J., Rendić, D., Matišić, D., Miloš, M., Heffer-Lauc, M., Flögel, M. and Labar, B. (2007) Aberrant Glycosylation of Igg Heavy Chain in Multiple Myeloma. Coll Antropol. 1: 247-251.
3. http://www.ucl.ac.uk/~ucbcdab/enzpur/amso4.htm 4. Deyl Z. ed. (1983). Electrophoresis: A survey of techniques and applications.
Elsevier Scientific, Amsterdam/New York. 5. Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature. 227: 680-685. 6. Alberts, B. et al. (2002) Molecular Biology of the Cell. Garland Science 7. Shapiro AL, Viñuela E, Maizel JV Jr. (September 1967). Molecular weight
estimation of polypeptide chains by electrophoresis in SDS-polyacrylamide gels. Biochem Biophys Res Commun. 28 (5): 815–820.
8. Voet, D. and Voet, J. (1995) Biochemistry. New York: John Wiley & Sons Inc.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
24
Vježba 4.
4.1. Klasifikacija enzima
Nazivlje enzima najčešće potječe od imena supstrata ili vrste kemijske reakcije koju kataliziraju, uz nastavak – aza. Tako je primjerice katekol supstrat enzima katekolaze, a proteaze su enzimi koji kataliziraju razgradnju proteina. Prema IUBMB nomenklaturi (nomenklaturi Međunarodnog udruženja za biokemiju i molekularnu biologiju, engl. International Union of Biochemistry and Molecular Biology, IUBMB) enzimi se označavaju brojevima EC. Svaki je enzim opisan slijedom četiri broja, pri čemu prvi broj klasificira enzime u jednu od 6 osnovnih skupina koje su formirane ovisno o mehanizmu reakcije koju kataliziraju (Tablica 4.1).
Tablica 4.1 Klasifikacija enzima.
Oznaka Razred Vrsta reakcije
EC 1 Oksidoreduktaze Oksido-redukcija
EC 2 Transferaze Prijenos funkcionalne skupine
EC 3 Hidrolaze Hidroliza
EC 4 Liaze Odstranjivanje skupine i tvorba dvostruke veze
EC 5 Izomeraze Izomerizacija
EC 6 Ligaze Tvorba kovalentne veze u sprezi s hidrolizom ATP
4.2. Katekolaza
U oksidoreduktaze ubrajamo i katekolazu (drugim imenom katekol-oksidazu; tirozinazu odnosno 1,2-dihidroksibenzen: kisik oksidoreduktazu, EC 1.10.3.1), enzim široko rasprostranjen u biljkama. Katekolaza katalizira reakciju katekola i kisika u kojoj se katekol oksidira u benzokinon (Slika 4.1). U stanicama se nalazi u vezikulama i dolazi u doticaj s katekolom i kisikom nakon oštećenja stanice. Smeđa boja koja nastaje kada se, primjerice, voće razreže i stoji na zraku potječe od nastalog benzokinona. Benzokinon inhibira rast mikroorganizama te tako štiti oštećenu biljku od truljenja.
Slika 4.1 Reakcija katekola s kisikom katalizirana katekolazom.
Supstrati za katekolazu mogu biti i drugi dihidroksibenzeni, čijom oksidacijom nastaju visokoreaktivni kinoni, koji potom u neenzimskoj reakciji daju pigment melanin (Slika 4.2).
Enzimska kinetika (sekvenciranje
katekol
(1,2 – dihidroksibenzen)
katekolaza
2
+ O2
benzokinon
(crven)
2
+ 2 H2O

BIOLOŠKA KEMIJA — PRAKTIKUM
25
Slika 4.2 Reakcija dopamina s kisikom katalizirana katekolazom.
4.3. Kinetika reakcije katekolaze
Osnovni principi enzimske kinetike objašnjeni su u zasebnom odjeljku (Dodatak – enzimska kinetika). Prilikom praćenja enzimske kinetike u in vitro uvjetima uobičajeno je da se u reakcijskoj smjesi nalaze enzim i supstrat u puferiranom vodenom mediju. Tijek enzimske reakcije koju katalizira katekolaza može se pratiti spektrofotometrijski – tijekom enzimske reakcije bezbojni se supstrat katekol pretvara u smeđi produkt benzokinon, koji apsorbira svjetlost valne duljine 540 nm. Apsorbancija otopine proporcionalna je koncentraciji tvari koja apsorbira, što je poznato kao Beer-Lambertov zakon (A = ε x l x c, pri čemu je ε molarni ekstinkcijski koeficijent, l duljina puta zrake svjetlosti kroz uzorak, a c koncentracija tvari koja apsorbira). Početna brzina enzimske reakcije stoga se može izraziti pomoću vrijednosti asporbancije:
vo= d[P]/dt = dA/dt
Katekolaza koja se koristi za ovu vježbu nije čisti protein otopljen u puferu, već se kao izvor enzima koristi ekstrakt krumpira. Ekstrakt krumpira također apsorbira svjetlost valne duljine 540 nm, no s obzirom na to da se njegova koncentracija u određenoj reakcijskoj smjesi tijekom vremena ne mijenja, ta je apsorbancija konstantna tijekom enzimske reakcije. Kako bi izmjerena apsorbancija potjecala isključivo od nastalog produkta, kao slijepa proba koristi se smjesa ekstrakta krumpira i pufera koja ne sadrži supstrat i u kojoj je koncentracija ekstrakta krumpira jednaka onoj u uzorku. Prilikom izvođenja pokusa na umu valja imati da enzimska reakcija, koju pratimo spektrofotometrijski, počinje u trenutku kada se u reakcijskoj smjesi istovremeno nađu enzim i supstrat. Stoga je takvu, u potpunosti priređenu reakcijsku smjesu potrebno odmah premjestiti u kivetu i započeti mjerenje.
4.4. Zadaća
Priredite reakcijske smjese s istim koncentracijama enzima i različitim koncentracijama supstrata i odredite apsorbancije tako priređenih smjesa u tridesetosekundnim intervalima tijekom prve tri minute enzimske reakcije. Zaključite kada je enzimska reakcija u ustaljenom, a kada u post-ustaljenom stanju. Izračunajte vrijednosti početne brzine enzimske reakcije pri različitim koncentracijama supstrata.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
26
Nacrtajte Michaelis-Menteničin graf ovisnosti početne brzine o koncentraciji supstrata, odredite maksimalnu početnu brzinu enzimske reakcije pri zadanoj koncentraciji enzima (Vm), Michaelisovu konstantu za katekolazu u navedenim reakcijskim uvjetima (Km) te zaključite kako koncentracija supstrata utječe na početnu brzinu enzimske reakcije.
Priredite reakcijske smjese s različitim koncentracijama enzima i istim koncentracijama supstrata te odredite apsorbancije tako priređenih smjesa u tridesetosekundnim intervalima tijekom prvih devedeset sekundi enzimske reakcije. Izračunajte vrijednosti početne brzine enzimske reakcije pri različitim koncentracijama enzima. Nacrtajte graf ovisnosti početne brzine enzimske reakcije o koncentraciji enzima i zaključite kako koncentracija enzima utječe na početnu brzinu enzimske reakcije. 4.5. Reagencije
• 10% ekstrakt krumpira u fosfatnom puferu, pH 7,0 (držati na ledu u tamnom) • 20 mM katekol u dest. vodi • 50 mM natrijev fosfatni pufer, pH 7,0
4.6. Pribor
• staklene epruvete • plastične epruvete od 15 mL • stalak za epruvete • staklene čaše • aluminijska folija • automatske pipete • nastavci za automatske pipete • staklena kiveta • staničevina • staklene boce • boce štrcaljke • štoperica • VIS spektrofotometar
4.7. Načelo postupka
U enzimskoj reakciji katekolaze s katekolom nastaje smeđi produkt bezokinon. Koncentracija nastalog produkta tijekom određenog vremena može se odrediti izravnim mjerenjem apsorbancije istog. Na temelju rezultata mjerenja može se zaključiti tijekom kojeg vremenskog perioda traje ustaljeno stanje te izračunati vrijednosti početne brzine enzimske reakcije, Vm i Km kao i ispitati kako početna brzina enzimske reakcije ovisi o koncentraciji supstrata i koncentraciji enzima.

BIOLOŠKA KEMIJA — PRAKTIKUM
27
4.8. Postupak
4.8.1. Tijek enzimske reakcije i učinak koncentracije supstrata na početnu brzinu enzimske reakcije
• Označite epruvete i u njih precizno ispipetirajte odgovorajuće volumene pufera, vode i katekola (navedene u tablici 4.2).
Tablica 4.2 Sastav reakcijskih smjesa.
oznaka
epruvete
volumen (izražen u mL)
pufer, pH 7,0 H20 katekol
S 2 2,5 -
1 2 2,3 0,2
2 2 2,0 0,5
3 2 1,7 0,8
4 2 1,5 1,0
5 2 1,0 1,5
6 2 0,5 2,0
• Prva epruveta (označena S) ne sadrži katekol te služi kao slijepa proba. Dodajte u prvu epruvetu 0,5 mL ekstrakta krumpira, začepite epruvetu i promućkajte je te sadržaj epruvete prelijte u kivetu. Stavite kivetu u spektrofotometar te apsorbanciju pri 540 nm namjestite na vrijednost 0.
• Ispraznite kivetu, isperite je destiliranom vodom i protresite kako biste iz nje uklonili svu tekućinu.
• Dodajte u sljedeću epruvetu (1) 0,5 mL katekola i istovremeno uključite štopericu. Začepite epruvetu i promućkajte je, potom njezin sadržaj prelijte u kivetu koju umetnite u spektrofotometar te očitavajte apsorbancije svakih 30 sekundi tijekom 3 minute. Rezultate upisujte u tablicu 4.3.
• Na isti gore opisani način izmjerite apsorbancije ostalih uzoraka. Napomena: nije potrebno ponovno koristiti slijepu probu.
Tablica 4.3 Vrijednosti apsorbancije tijekom enzimske reakcije uz konstantnu koncentraciju enzima katekolaze.
oznaka
epruvete ckatekol (mM)
vrijeme u minutama
0,5 1,0 1,5 2,0 2,5 3,0
1 0,8
2 2,0 3 3,2 4 4,0 5 6,0 6 8,0
• Prikažite rezultate grafički nanoseći na apscisu vrijeme (t) u minutama, a na ordinatu A540. Zaključite tijekom kojeg vremenskog raspona traje ustaljeno stanje.
• Iz vrijednosti apsorbancija koje se odnose isključivo na ustaljeno stanje izračunajte početnu brzinu enzimske reakcije pri određenoj koncentraciji supstrata. Početnu brzinu enzimske reakcije izrazite kao prosječnu promjenu apsorbancije u minuti. Kako je vrlo moguće da eksperimentalno dobivene
BIOLOŠKA KEMIJA — PRAKTIKUM ___
28
vrijednosti apsorbancije u ustaljenom stanju malo odstupaju od pravca kojim se može opisati ovisnost apsorbancije o vremenu, prosječnu promjenu apsorbancije u minuti izračunajte prema formuli:
• Nacrtajte Michaelis-Menteničin graf i Lineweaver-Burkov graf, očitajte vrijednosti Vm i Km i zaključite kako koncentracija supstrata utječe na početnu brzinu enzimske reakcije.
4.8.2. Učinak koncentracije enzima na početnu brzinu enzimske reakcije
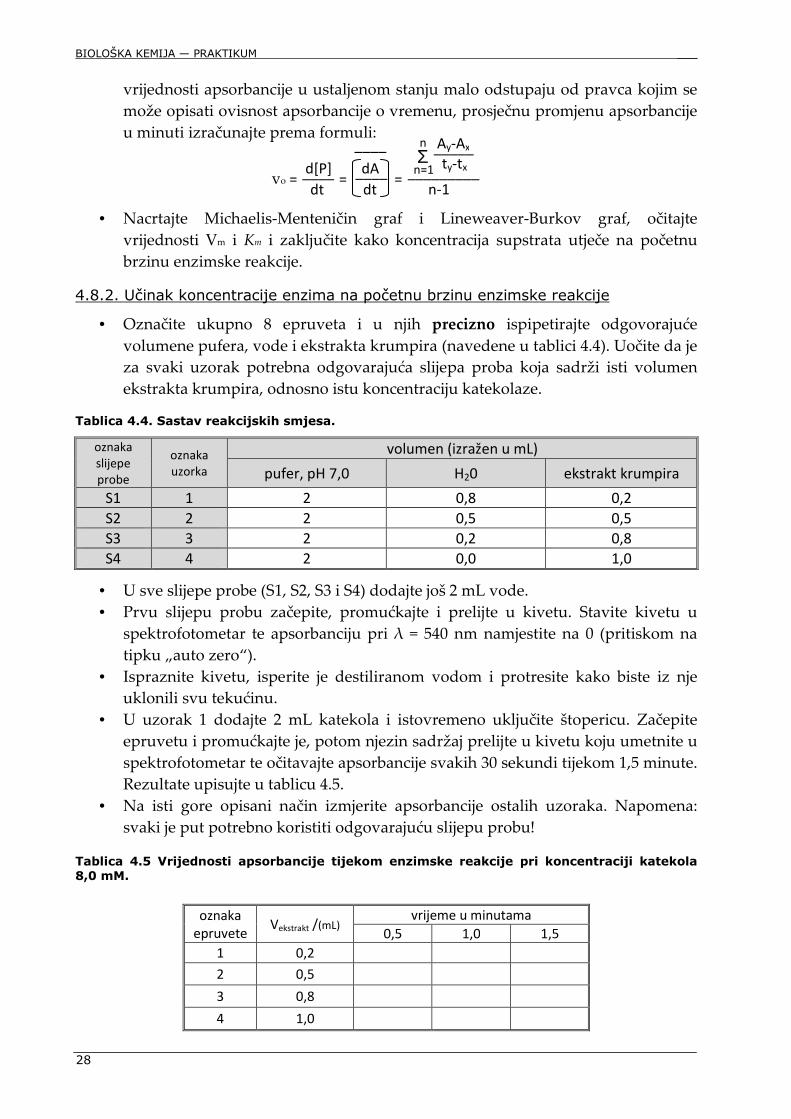
• Označite ukupno 8 epruveta i u njih precizno ispipetirajte odgovorajuće volumene pufera, vode i ekstrakta krumpira (navedene u tablici 4.4). Uočite da je za svaki uzorak potrebna odgovarajuća slijepa proba koja sadrži isti volumen ekstrakta krumpira, odnosno istu koncentraciju katekolaze.
Tablica 4.4. Sastav reakcijskih smjesa.
oznaka
slijepe
probe
oznaka
uzorka
volumen (izražen u mL)
pufer, pH 7,0 H20 ekstrakt krumpira
S1 1 2 0,8 0,2
S2 2 2 0,5 0,5
S3 3 2 0,2 0,8
S4 4 2 0,0 1,0
• U sve slijepe probe (S1, S2, S3 i S4) dodajte još 2 mL vode. • Prvu slijepu probu začepite, promućkajte i prelijte u kivetu. Stavite kivetu u
spektrofotometar te apsorbanciju pri λ = 540 nm namjestite na 0 (pritiskom na tipku „auto zero“).
• Ispraznite kivetu, isperite je destiliranom vodom i protresite kako biste iz nje uklonili svu tekućinu.
• U uzorak 1 dodajte 2 mL katekola i istovremeno uključite štopericu. Začepite epruvetu i promućkajte je, potom njezin sadržaj prelijte u kivetu koju umetnite u spektrofotometar te očitavajte apsorbancije svakih 30 sekundi tijekom 1,5 minute. Rezultate upisujte u tablicu 4.5.
• Na isti gore opisani način izmjerite apsorbancije ostalih uzoraka. Napomena: svaki je put potrebno koristiti odgovarajuću slijepu probu!
Tablica 4.5 Vrijednosti apsorbancije tijekom enzimske reakcije pri koncentraciji katekola 8,0 mM.
oznaka
epruvete Vekstrakt /(mL)
vrijeme u minutama
0,5 1,0 1,5
1 0,2
2 0,5 3 0,8 4 1,0
____
d[P] =
dt ____ = vo ____ dA
dt
Ay-Ax
n-1
_____
= n=1 Σ
_________ ty-tx
n

BIOLOŠKA KEMIJA — PRAKTIKUM
29
• Iz vrijednosti apsorbancija izračunajte početnu brzinu enzimske reakcije pri određenoj koncentraciji katekolaze. Početnu brzinu enzimske reakcije izrazite kao prosječnu promjenu apsorbancije u minuti.
• Nacrtajte graf ovisnosti početne brzine enzimske reakcije o koncentraciji enzima i zaključite o kakvoj je ovisnosti riječ i zašto.
4.9. Zadaci
1. Možete li metodom opisanom u ovoj vježbi pratiti što se događa s koncentracijom produkta tijekom predustaljenog stanja? Zašto?
2. Zašto ekstrakt krumpira tijekom ove vježbe držite na ledu? 3. Koje su osnovne osobine ustaljenog stanja enzimske reakcije koja prati Michaelis-
Menteničinu kinetiku? 4. Kako biste podesili reakcijske uvjete ako ste u pokusu A izmjerili određenu
maksimalnu brzinu studirane enzimske reakcije, a u pokusu B želite da izmjerena maksimalna brzina bude 2 puta veća od brzine u pokusu A?
4.10. Literatura
1. Voet, D. and Voet, J. (1995) Biochemistry. New York: John Wiley & Sons Inc. 2. Wood, E.J. (1989) Practical Biochemistry for Colleges. Oxford: Pergamon Press. 3. Price, N.C., Dwex, R.A., Ratcliffe, R.G. and Wormald, M.R. (2005) Principles and
Problems in Physical Chemistry for Biochemists. Oxford: Oxford University Press.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
30
Dodatak:
Svaku kemijsku reakciju pokreće negativna razlika Gibbsove energije između produkata reakcije i reaktanata. Tijekom reakcije ta se energetska razlika smanjuje. Sustav ulazi u ravnotežu kad je razlika Gibbsovih energija jednaka nuli. Pozitivna energetska razlika pokreće povratnu reakciju. Oslobađanje energije bitno je za spontanost reakcije. Međutim, ne teku sve energetski povoljne reakcije spontano i brzo. Postoje vrlo spore reakcije usprkos velikom energetskom potencijalu. Njih koči energetska prepreka koju treba prijeći na putu do produkta reakcije: treba aktivirati reaktante. U kemiji se potrebna energija aktivacije najčešće namakne povišenjem temperature. U organizmu, koji može živjeti u vrlo ograničenom temperaturnom rasponu, spore se reakcije ubrzavaju pomoću enzima, vrlo specifičnih i moćnih bioloških katalizatora. Enzimska je reakcija složenijeg mehanizma od nekatalizirane reakcije, ali teče brže, uz smanjenu energiju aktivacije (Slika 1). Katalitička moć enzima je vrlo različita, a najveća brzina enzimske reakcije ograničena je difuzijom (k = 10-9Ms-1).
Slika 1. Shematski prikaz promjene Gibbsove energije (∆∆∆∆G) nekatalizirane i enzimski katalizirane reakcije S P.
Početkom 20. stoljeća Michaelis i Menten pretpostavili su da se enzimska reakcija može odviti samo ako enzim E i supstrat S stvore funkcionalni kompleks ES.
E + S ES P + E
Kompleks ES zatim može dati produkt i slobodni enzim ili može u povratnoj reakciji disocirati na reaktante. Enzim ne mijenja položaj ravnoteže S P, on samo ubrzava reakciju pretvorbe supstrata u produkt do uspostavljanja ravnoteže.
Brzina nastajanja produkta ovisi o katalitičkim svojstvima i koncentraciji enzima te koncentraciji supstrata. Zbog načina vezanja supstrata, prirode katalitičke reakcije i dinamičnosti proteinske strukture, tijek i brzina reakcije vrlo su osjetljivi na reakcijske
Enzimska kinetika (sekvenciranje
#
ΔG0#(ES→→→→P#)
S
P
ΔG0#(S→→→→S#)

BIOLOŠKA KEMIJA — PRAKTIKUM
31
uvjete, kao što su temperatura, pH i ionska jakost te drugi sastojci u reakcijskom mediju (proteini, aktivatori, inhibitori i dr.).
I. Tijek enzimske reakcije
Tijek pretvorbe supstrata u produkt u prisutnosti enzima može se teorijski pratiti bilo kojom indikatorskom metodom koja prati smanjenje koncentracije supstrata ili povećanje koncentracije produkta. Praktički je najčešće moguće mjeriti samo nastajanje produkta, jer je povećanje koncentarcije tvari koja raste od 0 do neke vrijednosti analitički pristupačno, dok je smanjenje velike koncentracije tvari za neku malu vrijednost analitički teško uočljivo. Grafički prikaz tipičnog reakcijskog tijeka dan je na slici 2.
vrijeme
[P] i
[ES
]
PRED-USTALJENOSTANJE
USTALJENOSTANJE
POST-USTALJENOSTANJE
d[P] /dt=v0=konst
d[ES]/dt > 0
d[ES] /dt = 0 d[ES] /dt < 0
[P]
[ES]
Slika 2. Tijek enzimski katalizirane reakcije. Nakon miješanja enzima i supstrata dolazi do stvaranja kompleksa ES. Koncentracija ES raste dok se ne ustali, odnosno dok reakcijski sustav ne uđe u ustaljeno stanje. Ustaljeno se stanje uspostavi vrlo brzo (ms < t < 1s), a kratki period koji mu prethodi nazivamo predustaljeno stanje.
U ustaljenom stanju produkt (P) nastaje stalnom brzinom, koju nazivamo početnom brzinom, a označava se vo. Michaelis-Menteničina kinetika odnosi se na brzine enzimskih reakcija u ustaljenom stanju.
Kad se potrošio osjetni dio supstrata, prestaje ustaljeno stanje i započinje postustaljeno stanje, tijekom kojeg dolazi do postupnog smanjenja koncentracije ES, intenziviranja povratne reakcije, stoga i do postupnog smanjenja brzine nastanka produkta. Koncentracija produkta P raste sve do uspostavljanja ravnoteže, u kojoj su brzina nastajanja produkta i brzina njegove razgradnje izjednačene.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
32
II. Kinetika ustaljenog stanja
Ustaljeno stanje uspostavit će se u uvjetima kada je koncentracija enzima znatno manja od koncentracije supstrata, a trajat će dokle god je koncentracija produkta zanemarljivo mala u odnosu na koncentraciju supstrata. Prema tome shema enzimske reakcije u ustaljenom stanju zanemaruje povratnu reakciju.
E + S ES P + E
Brzina reakcije u ustaljenom stanju ovisna je isključivo o koncentraciji kompleksa ES koja je konstantna, jer su brzine nastanka i razgradnje kompleksa jednake. Tu brzinu nazivamo početnom brzinom enzimske reakcije i označavamo je kao vo.
vo= k2 [ES]
gdje je k2 prava katalitička konstanta (I reda, kcat), pa je brzina nastajanja produkta iz kompleksa ES izravno proporcionalna koncentraciji kompleksa ES
k1 [E][S] = (k-1 + k2) [ES]
Budući da se početna (ukupna) koncentracija enzima ET raspodjeljuje na slobodni enzim E i kompleks ES
[E ]= [ET] - [ES]
Na osnovi gornjih jednadžbi možemo izraziti koncentraciju ES kao [ES] = gdje je Km = (k-1 + k2)/k1.
Konstantu Km nazivamo Michaelisovom konstantom. Slijedi da je početna brzina enzimske reakcije određena izrazom
vo=
Mjerimo li početne brzine enzimske reakcije uz različite početne koncentracije supstrata, opazit ćemo da početna brzina raste s porastom supstrata sve dok ne dosegne neku maksimalnu brzinu, Vm. Maksimalna brzina se postigne kad je sav enzim zasićen supstratom, tako da je koncentracija kompleksa ES jednaka početnoj koncentraciji enzima.
vo = Vm = k2 [ET]
Prema tome početnu brzinu možemo izraziti kao
vo =
Kad je [S] << Km (< 0,01 Km) početna brzina linearno ovisi o početnoj koncentraciji supstrata.
vo = gdje je = k
[ET] [S]
Km + [S]
k2[ET] [S]
Km + [S]
Vm [[[[S]]]]
Km + [[[[S]]]]
Vm [S]
Km
Vm
Km
k2 k-1 k1
BIOLOŠKA KEMIJA — PRAKTIKUM
33
v = d[S]
dt = k[S]
gdje je k konstanta brzine I reda.
Znajući konstantu brzine možemo izračunati količinu nastalog produkta u određenom vremenu iz integriranog oblika v. Kad je [S] >> Km (> 100 Km) reakcija je 0-tog reda. Količina nastalog produkta ne ovisi o S, pa je količina produkta jednaka
[P] = Vm × t
Enzimska kinetika u dijagnostičke svrhe mjeri se upravo u takvim uvjetima. Znamo da je Vm = k2 [ET] pa količina nastalog produkta izravno ovisi o koncentraciji aktivnog enzima.
[P] = k2 [ET] t III. Mjerenje katalitičke aktivnosti i brzine enzimski katalizirane reakcije
Katalitička aktivnost enzima obično se određuje mjerenjem koncentracija nastalog produkta nakon inkubacije smjese poznatih koncentracija supstrata [S] i enzima [E] pri pogodnom pH u određenim vremenskim razdobljima. Najčešće se koncentracija nastalog produkta određuje spektrofotometrijskom metodom koja se temelji na mjerenju promjene apsorbancije. Kako je sposobnost apsorpcije svjetlosti razmjerna koncentraciji tvari koja apsorbira, uz odgovarajući baždarni dijagram (ili poznati molarni ekstinkcijski koeficijent) moguće je odrediti koncentraciju mjerene tvari. Ako optička svojstva produkta enzimske reakcije ne dopuštaju izravno mjerenje odgovarajuće promjene apsorbancije, neophodno je uvesti konsekutivnu "indikatorsku" reakciju koja će nastali produkt prevesti u obojeni produkt indikatorske reakcije. Ovisnost početne brzine o koncentraciji supstrata prikazana je grafički na slici 3.
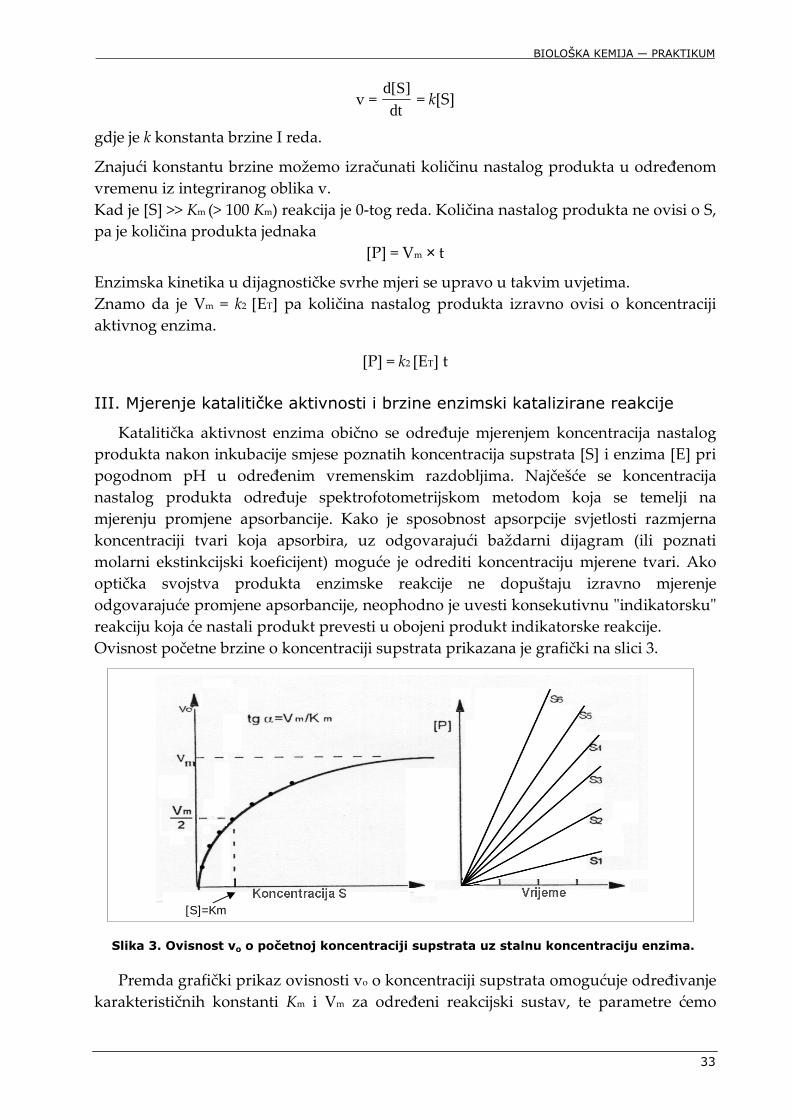
Slika 3. Ovisnost vo o početnoj koncentraciji supstrata uz stalnu koncentraciju enzima.
Premda grafički prikaz ovisnosti vo o koncentraciji supstrata omogućuje određivanje karakterističnih konstanti Km i Vm za određeni reakcijski sustav, te parametre ćemo
[S]=Km

BIOLOŠKA KEMIJA — PRAKTIKUM ___
34
lakše grafički odrediti na osnovi algebarskih transformacija osnovnog izraza za početnu brzinu. Najčešće se koristi Lineweaver-Burkov prikaz (Slika 4.)
Slika 4. Odreñivanje Km i Vm po Lineweaver-Burku.
Aktivnost enzima moguće je izraziti na više načina. U SI-sustavu jedinica enzimske aktivnosti je katal. Katal je definiran kao enzimska aktivnost (količina enzima) koja u jednoj sekundi pretvara 1 mol supstrata u produkt pri optimalnim uvjetima (1 kat = mol L-1s-1). Enzimske aktivnosti izražavaju se obično u mikrokatalima (µkat) i nanokatalima (nkat) što odgovara jedinicama µmol L-1 s-1 i nmol L-1 s-1. Starija jedinica za enzimsku aktivnost je tzv. internacionalna jedinica enzimske aktivnosti, IU. Definirana je kao količina enzima koja katalizira pretvorbu jednog mikromola supstrata u produkt u jednoj minuti (µmol L-1 min-1) pri standardnim (optimalnim) uvjetima. 1IU = 1/60 µkat = 16,67 nkat, odnosno 1kat = 6×107 IU. Srodne mjere enzimske aktivnosti su specifična aktivnost enzima, molarna aktivnost enzima i obrtni broj. Specifična aktivnost enzima izražava se kao kat g-1, a kaže koliko molova produkta nastaje u prisutnosti jednog grama proteina u jedinici vremena (ili IU mg-1, mikromoli supstrata koje 1 mg proteina prevede u produkt u jednoj minuti). Molarna aktivnost enzima (kat mol-1) odgovara broju molova supstrata koje jedan mol enzima prevede u produkt u jedinici vremena. Prometni (obrtni) broj određenog enzima ima jedinicu min-1, a predstavlja broj molova nastalog produkta po molu enzima u minuti. Budući da enzim može imati više aktivnih središta, prometni broj često se iskazuje kao broj molova produkta koji nastaju po molu aktivnih središta u minuti. Katalitička koncentracija enzima određena je brojem katala (ili IU) u litri otopine.
IV. Čimbenici koji utječu na brzinu enzimske reakcije
IV.I Inhibicija enzimske reakcije
Kinetička studija enzimske reakcije pomaže u otkrivanju inhibitora i u prepoznavanju mehanizma njihova djelovanja. Inhibitori imaju veliko kliničko značenje, bilo terapijsko, bilo toksično.
Inhibitore možemo podijeliti u dvije skupine: reverzibilne i ireverzibilne. U prvom slučaju aktivnost se može ponovno uspostaviti odgovarajućim uklanjanjem inhibitora, primjerice dijalizom. Inhibitorno djelovanje reverzibilnog inhibitora ovisi o konstanti
1 /
v o
1 /
V max
-1/K m 1/[S]
nagib (tg α)=K m /Vmax

BIOLOŠKA KEMIJA — PRAKTIKUM
35
inhibicije odnosno disocijacijskoj konstanti kompleksa enzim-inhibitor (Ki). Ireverzibilna inhibicija je neuklonjiva i progresivna.
Razlikujemo tri osnovna tipa reverzibilne inhibicije: kompetitivna (konkurentna), nekompetitivna (nekonkurentna) i akompetitivna inhibicija. Svaki od njih se na karakterističan način odražava u prividnim vrijednostima Km i Vm.
Kompetitivni inhibitor je spoj koji je po svojoj strukturi veoma sličan pravom supstratu i veže se u aktivno središte i tako konkurira supstratu pri vezanju na enzim. On zbog toga povećava prividnu Km vrijednost. Stupanj inhibicije ovisi o relativnom odnosu koncentracija supstrata i inhibitora tako da odgovarajućim povećanjem koncentracije supstrata možemo potpuno potisnuti djelovanje kompetitivnog inhibitora.
E + S ES E + P
E + I EI
[EI]
[I] [E] =
[ES]
[S] [E] = is KK
Ukupna koncentracija enzima jednaka je
i
sst
[I]
[S]
[ES] + [ES] +
[S]
[ES] =][E
K
KK ×
[S]
[S]V = v
mI
mo +K
immI
[I] + 1=K
KK
[S]
[I] + 1
v
v
mI
m
I
o
+×=
KK
K
VmI = Vm
Primjer je kompetitivne inhibicije djelovanje benzojeve kiseline (homologa pravog supstrata - katekola) na katekolazu.
Do nekompetitivne inhibicije dolazi kad reverzibilna inhibicija ovisi isključivo o koncentraciji inhibitora, a ne možemo na nju utjecati promjenom koncentracije supstrata. Nekompetitivni inhibitor ne veže se u samom aktivnom središtu, već njegovo vezanje na enzim izaziva konformacijsku promjenu koja enzim učini neaktivnim, što smanjuje ukupnu koncentraciju aktivnog enzima. Smanjuje se maksimalna brzina, jer je time jedan dio enzima uklonjen iz katalitičkog tijeka, a aktivni udio enzima zadržava početni afinitet za supstrat, tj. Km se u nekompetitivnoj inhibiciji ne mijenja.
Akompetitivna inhibicija nastaje kad se inhibitor ne veže na slobodni enzim nego na neki enzimski oblik koji sam više ne može vezati supstrat, na primjer na kompleks ES.
E + S ES E + P
ES + I ESI

BIOLOŠKA KEMIJA — PRAKTIKUM ___
36
i
mmi
i
m
i
[I]+1
[I]+1
V = v
K
KK
K
=
Mjerenje ovisnosti vo i [S] prikazano u Lineweaver-Burkovom dijagramu jasno predočuje kinetičke razlike različitih tipova inhibicije (Slika 5.).
Slika 5. Grafičko odreñivanje tipa inhibicije po Lineweaver-Burku
(A - neinhibirana reakcija, B - nekompetitivna inhibicija, C - kompetitivna inhibicija, D - akompetitivna inhibicija).
IV.II Utjecaj pH na brzinu enzimske reakcije
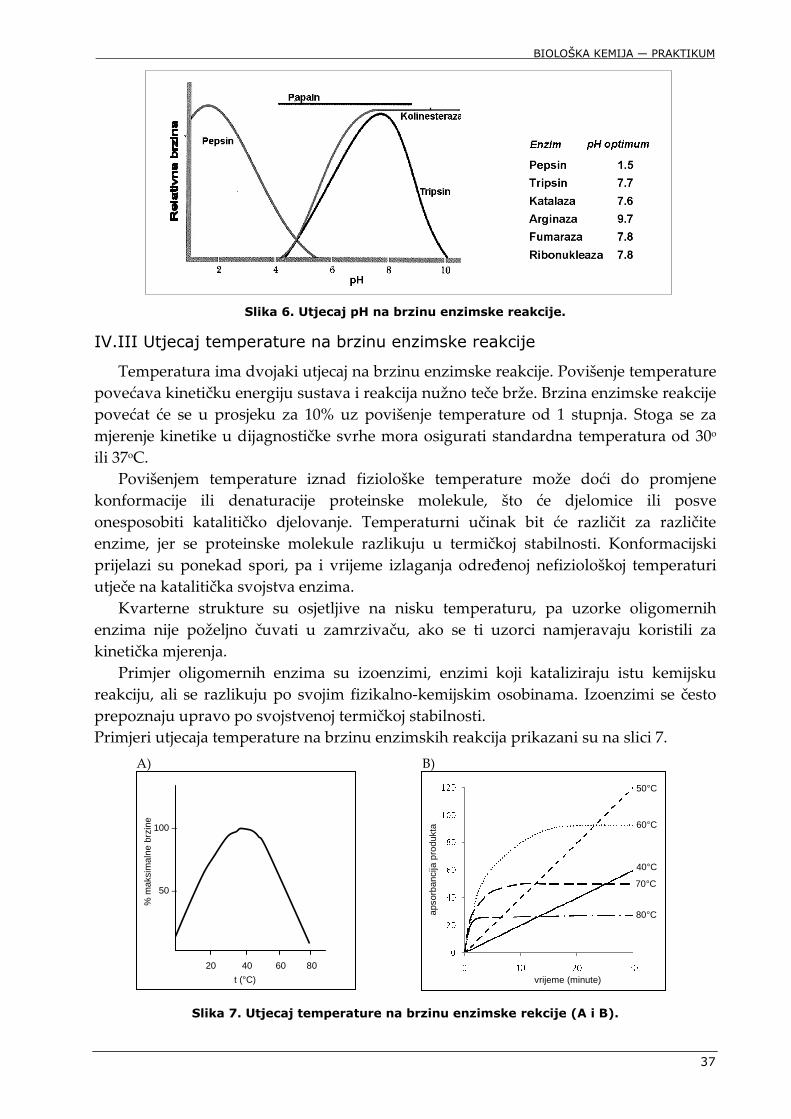
Katalitičko središte enzima sastoji se od katalitičkih i veznih mjesta koja moraju biti u odgovarajućem prostornom razmještaju i ionizacijskom obliku kako bi moglo doći do uspješne katalize. Proteinska struktura enzima vrlo je osjetljiva na kiselost medija, jer ukupna prostorna konformacija ovisi i o slabim nekovalentnim vezama od kojih su mnoge elektrostatske prirode. Razumljivo je da će kiselost i ionska jakost medija značajno i specifično utjecati na katalitička svojstva enzima. Zbog toga mnogi enzimi postižu maksimalnu katalitičku učinkovitost samo u ograničenom području pH. Primjeri učinka pH na enzimsku aktivnost prikazani su na slici 6.
1 / V o
1 / V max
-1/K m 1/[S]
B D
C
A
BIOLOŠKA KEMIJA — PRAKTIKUM
37
Slika 6. Utjecaj pH na brzinu enzimske reakcije.
IV.III Utjecaj temperature na brzinu enzimske reakcije
Temperatura ima dvojaki utjecaj na brzinu enzimske reakcije. Povišenje temperature povećava kinetičku energiju sustava i reakcija nužno teče brže. Brzina enzimske reakcije povećat će se u prosjeku za 10% uz povišenje temperature od 1 stupnja. Stoga se za mjerenje kinetike u dijagnostičke svrhe mora osigurati standardna temperatura od 30o ili 37oC.
Povišenjem temperature iznad fiziološke temperature može doći do promjene konformacije ili denaturacije proteinske molekule, što će djelomice ili posve onesposobiti katalitičko djelovanje. Temperaturni učinak bit će različit za različite enzime, jer se proteinske molekule razlikuju u termičkoj stabilnosti. Konformacijski prijelazi su ponekad spori, pa i vrijeme izlaganja određenoj nefiziološkoj temperaturi utječe na katalitička svojstva enzima.
Kvarterne strukture su osjetljive na nisku temperaturu, pa uzorke oligomernih enzima nije poželjno čuvati u zamrzivaču, ako se ti uzorci namjeravaju koristili za kinetička mjerenja.
Primjer oligomernih enzima su izoenzimi, enzimi koji kataliziraju istu kemijsku reakciju, ali se razlikuju po svojim fizikalno-kemijskim osobinama. Izoenzimi se često prepoznaju upravo po svojstvenoj termičkoj stabilnosti. Primjeri utjecaja temperature na brzinu enzimskih reakcija prikazani su na slici 7.
Slika 7. Utjecaj temperature na brzinu enzimske rekcije (A i B).
% m
aksi
mal
ne b
rzin
e
apso
rban
cija
pro
dukt
a
A) B)
20 40 60 80
t (°C)
100 –
50 –
vrijeme (minute)
40°C
50°C
60°C
70°C
80°C

BIOLOŠKA KEMIJA — PRAKTIKUM ___
38
V. Literatura
1. Voet, D. and Voet, J. (1995) Biochemistry. New York: John Wiley & Sons Inc. 2. Price, N. C., Dwek, R. A., Ratcliffe, R. G., Wormald, M. R. (2001) Principles and
Problems in Physical Chemistry for Biochemists. Oxford: Oxford University Press

BIOLOŠKA KEMIJA — PRAKTIKUM
39
Vježba 5.
5.1 Ugljikohidrati
Jedna od četiri osnovne skupine bioloških makromolekula su ugljikohidrati - polihidroksialdehidi ili polihidroksiketoni opće formule (CnH2n0n)x, pri čemu je n≥3 (te ovisno o tome razlikujemo trioze, tetroze, pentoze, heksoze itd.) dočim x može biti 1 (kod monosaharida), 2 (kod disaharida), 3-10 (kod oligosaharida) te >10 (kod polisaharida). Monosaharidi mogu biti različiti metabolički intermedijari, a gradivne su jedinice već spomenutih di-, oligo- i polisaharida kao i nukleinskih kiselina i glikokonjugata. Reakcijom karbonilne skupine monosaharida i jedne od njegovih hidroksilnih skupina nastaju poluacetali ili poluketali, pri čemu se rastvoreni oblik ugljikohidrata (koji nacrtan u linearnom obliku nazivamo Fischerovim prikazom) zatvara u ciklički oblik (koji prikazujemo Haworthovom projekcijom). Na taj se način stvaraju peteročlani prsteni (furanoze) ili šesteročlani prsteni (piranoze) koji osim ugljika sadrže i jedan atom kisika. Pri tome karbonilni C-atom postaje asimetričan (anomeran), a ovisno o položaju novonastale hidroksilne skupine na anomernom C-atomu razlikujemo α i β anomere. Aldoheksoze većinom stvaraju piranoze (reakcijom aldehidne skupine i hidroksilne skupine na petom C-atomu), a aldotetroze, aldopentoze i ketoheksoze stvaraju furanoze. Na slici 5.1 prikazana je aldoheksoza glukoza, koja se još naziva i grožđani šećer, jedini monosaharid koji se u stanicama nalazi samostalno, a nalazimo je i u krvi i ostalim tjelesnim tekućinama, dok je na slici 5.2 prikazana ketoheksoza fruktoza, koja se naziva i voćni šećer.
Slika 5.1 D-glukoza. A) Linearan oblik – Fischerov prikaz B) Ciklički oblik – Haworthov prikaz.
Ugljikohidrati (sekvenciranje
otvoreni
oblik (<1%) A) B)
stvaranje poluacetala
D-glukofuranoza (<1%) D-glukopiranoza (99%)

BIOLOŠKA KEMIJA — PRAKTIKUM ___
40
Slika 5.2 D-fruktoza. Linearan oblik – Fischerov prikaz i ciklički oblik – Haworthov prikaz.
U krutom stanju monosaharidi su u cikličkom obliku, a u vodenim otopinama postoji ravnoteža između otvorenih i cikličkih oblika. Ukoliko hidroksilna skupina koja nastaje tijekom stvaranja poluacetala ili poluketala sudjeluje u stvaranju veze između monosaharida i druge molekule koja može, ali ne mora biti ugljikohidrat (takvu vezu nazivamo glikozidnom vezom), ciklički oblik monosaharida ne može se više rastvoriti u linearan oblik. To zapravo znači da se ne može ponovno spontano stvoriti slobodna karbonilna skupina. Na slici 5.3 prikazana je struktura saharoze, uobičajenog stolnog šećera, koja se sastoji od glukoze i fruktoze međusobno povezanih α1 →β2 vezom (tj. vezom između dvije hidroksilne skupine na anomernim C-atomima koji nastaju nakon ciklizacije linearnih oblika monosaharida) i struktura laktoze, mliječnog šećera, koji se sastoji od galaktoze i glukoze povezanih β(1,4) vezom.
Slika 5.3 Saharoza i laktoza.
Aldehidna skupina ugljikohidrata lako se oksidira, pa aldoze, ako nisu preko
anomernog kisika povezane u glikozidnu vezu, djeluju kao reduktivna sredstva, pri čemu nastaju aldonske kiseline. Različitim vrstama obojenih reakcija može se utvrditi je li određeni ugljikohidrat reduktivan. Te se reakcije temelje na redukciji metalnih iona; Tromerova, Fehlingova i Benedictova reakcija (Slika 5.4) zasnivaju se na redukciji bakrovog (II) sulfata u bakrov (I) oksid koji se taloži u obliku svijetlo crvenog taloga, a Nylanderova reakcija na redukciji bizmutovog nitrata do elementarnog bizmuta, koji se taloži u obliku crnog mulja.
D-fruktoza
(linearni oblik)
D-fruktofuranoza
(Haworthov prikaz)
saharoza laktoza

BIOLOŠKA KEMIJA — PRAKTIKUM
41
R−C −H + 2Cu2+ + 5OH- → R−C −O- + Cu2O + 3H2O
Slika 5.4 Benedictova reakcija. Benedictov reagens sadrži bakrov (II) sulfat u alkalnoj vodenoj otopini natrijevog citrata i natrijevog karbonata. U prisutnosti reduktivnog šećera taloži se svijetlo crveni bakrov (I) oksid i nastaje aldonska kiselina.
Ketoze nije moguće razlikovati od aldoza u reakcijama na reduktivne šećere jer se te reakcije provode u blago lužnatom mediju u kojem se uspostavlja ravnoteža između aldoznih i ketoznih oblika preko enolnih oblika (keto-enolna tautomerija, Slika 5.5).
Slika 5.5 Keto-enolna tautomerija ketoza i aldoza.
Polisaharidi, polimeri više od 10 jedinica monosaharida, sadrže premalo slobodnih
reduktivnih krajeva u usporedbi s velikom količinom vezanih šećera, pa ne pokazuju pozitivne reakcije reduktivnih šećera. U skupinu polisaharida ubraja se u biljnom svijetu široko rasprostranjeni škrob, koji se sastoji većinom od amilopektina (glukoznih jedinica povezanih α(1,4) vezom) i u manjoj mjeri od amiloze (kod koje su molekule glukoze povezane također α(1,4) vezom, ali se razgranatost postiže i α(1,6) vezom). U spiralnu strukturu amiloze može se u reakciji s Lugolovom otopinom (otopinom elementarnog joda u kalijevom jodidu) interkalirati jod čime nastaje plavo-ljubičasto obojenje. Celuloza, polisaharid koji sadrži glukoze povezane β(1,4) vezom, ne boja se jodom.
Prisutnost ugljikohidrata u vodenim otopinama moguće je dokazati Molischovom reakcijom. U kiseloj sredini u kojoj se odvija Molischova reakcija polisaharidi hidroliziraju na monosaharide. U prisutnosti koncentrirane sulfatne kiseline monosaharidi se dehidratiraju stvarajući derivate furfurala (pentoza daje furfural, a heksoza 5-hidroksimetil-furfural). U daljnjim reakcijama s α-naftolom furfurali stvaraju ljubičasto obojene kondenzacijske produkte (Slika 5.6). Prema nijansama obojenja moguće je razlikovati pentoze od heksoza.
C ||||||||
C ||||||||
ketoza enol aldoza

BIOLOŠKA KEMIJA — PRAKTIKUM ___
42
Slika 5.6 Molischova reakcija. Prikazana je A) dehidratacija glukoze u 5-hidroksimetil-furfural i B) konačan produkt reakcije s α-
naftolom.
Ugljikohidrati, za razliku od proteina, ne vežu boju Coomassie Blue G250, što je temelj za Bradfordovu reakciju. Bradfordova je reakcija jedna od nekoliko metoda određivanja prisutnosti, ali i koncentracije proteina u smjesi. U kiseloj se otopini boja Coomassie Blue 250 uglavnom veže na bočne ogranke bazičnih aminokiselina pri čemu se boja otopine mijenja iz zelenkaste u plavu. Spektrofotometrijskim mjerenje apsorbancije (λ = 595 nm) otopine proteina uz Bradfordov reagens može se odrediti koncentracija ukupnih proteina u uzorku (uz konstruiranje baždarnog pravca na temelju apsorbancija otopina proteina poznatih koncentracija).
5.2 Zadaća
Razgradite laktozu u mlijeku djelovanjem enzima laktaze te odredite prisutnost i koncentraciju na taj način nastale glukoze u mlijeku. U različitim uzorcima provjerite prisutnost: škroba (Lugolovom reakcijom), proteina (Bradfordovom reakcijom), ugljikohidrata (Molischovom reakcijom) i reduktivnih ugljikohidrata (Benedictovom reakcijom). 5.3 Reagencije
• laktaza (β-galaktozidaza, 0,1 U/μL) u fiziološkoj otopini • kravlje mlijeko (koje sadrži laktozu) nerazrijeđeno i razrijeđeno 1:10 destiliranom
vodom • 1% otopina škroba u dest. vodi • zasićena otopina saharoze u dest. vodi • zasićena otopina glukoze u dest. vodi • zasićena otopina fruktoze u dest. vodi • otopina goveđeg serumskog albumina (BSA, 1 mg/mL) u dest. vodi • humani serum • 10% ekstrakt krumpira u 40 mM fosfatnom puferu pH 7,0
B)
A)
obojani furfuril-difenilmetan
5-hidroksimetil-furfural

BIOLOŠKA KEMIJA — PRAKTIKUM
43
• otopina DNA iz kravljeg timusa u puferu SSC/10 (15 mM NaCl, 1,5 mM natrijev citrat, pH 7,0 - 7,1) približne koncentracije 15 μg/mL
• suncokretovo ulje • Lugolova otopina (2% I2/ 10% KI u dest. vodi) • Bradfordov reagens (0,05 g/L Coomassie Blue G250 u 5% metanol/8,5% H3PO4 u
dest. vodi) • Benedictov reagens (0,95 M natrijev karbonat, 0,67 M natrijev citrat i 70 mM
bakrov (II) sulfat pentahidrat u dest. vodi) • 5% α-naftol u 96% etanolu • koncentrirana H2SO4
5.4 Pribor
• plastične epruvete od 1,5 mL • stalak za epruvete od 1,5 mL • automatske pipete • nastavci za automatske pipete • testne trake za semikvantitativno određivanje glukoze (primjerice Keto-Diastix,
Bayer) • staničevina • staklene epruvete • stalak za staklene epruvete • kapaljke • vodena kupelj ili čaša i grijač • staklene boce • drvene hvataljke za epruvete
5.5 Načelo postupka
Laktoza iz mlijeka razgrađuje se djelovanjem enzima laktaze na glukozu i galaktozu. Koncentracija glukoze određuje se pomoću komercijalno dostupnih test-traka na kojima se odvijaju dvije enzimske reakcije u slijedu: glukoza-oksidaza katalizira oksidaciju glukoze iz uzorka u glukonsku kiselinu pri čemu se stvara i vodikov peroksid; peroksidaza potom katalizira reakciju vodikovog peroksida s kalij-jodidnim kromogenom pri čemu nastaje smeđe obojani produkt (intenzitet boje ovisi o koncentraciji glukoze u uzorku).
Jod iz Lugolove otopine interkalira se u spiralnu strukturu amiloze koja je sastavni dio škroba pri čemu se boja mijenja iz smeđe u plavo-ljubičastu.
Bočni ogranci bazičnih aminokiselina u proteinima vežu boju Coomassie Blue G250 pri čemu se mijenja boja otopine u plavu, odnosno povećava se apsorbancija pri valnoj duljini od 595 nm.
Koncentrirana sulfatna kiselina uzrokuje hidrolizu složenih ugljikohidrata na monosaharide koji potom stvaraju derivate furfurala. Derivati furfurala s α-naftolom u konačnici stvaraju ljubičasto obojene produkte.
Slobodna aldehidna skupina ugljikohidrata oksidira dvovalente ione bakra pri čemu nastaju aldonske kiseline i narančasti talog bakrovog (I) oksida.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
44
5.6 Postupak
5.6.1 Priprema uzorka mlijeka koje ne sadrži laktozu
• Po 1 mL nerazrijeđenog kravljeg mlijeka otpipetirajte u 2 plastične obilježene epruvete od 1,5 mL.
• U jednu epruvetu dodajte 50 μL otopine laktaze. • Obje epruvete inkubirajte 15 minuta pri sobnoj temperaturi. • U pojedinu epruvetu uronite po jednu testnu traku za određivanje razine
glukoze, odmah je izvadite van i s trake uklonite višak mlijeka pomoću staničevine.
• Nakon 30 sekundi usporedite testno polje za glukozu sa skalom boja na pakiranju testnih traka te zaključite kolika je koncentracija glukoze u uzorcima.
5.6.2 Dokazivanje prisutnosti škroba Lugolovom reakcijom
• 11 plastičnih epruveta obilježite brojevima, stavite na stalak i u njih otpipetirajte redom po 1 mL sljedećih uzoraka: 1) otopine škroba, 2) otopine saharoze, 3) otopine glukoze, 4) otopine fruktoze, 5) otopine proteina BSA, 6) seruma, 7) ekstrakta krumpira, 8) razrijeđenog mlijeka, 9) otopine DNA, 10) vode u koju dodajte komadić staničevine, 11) suncokretovog ulja.
• U svaku pojedinu epruvetu kapnite po 3 kapi Lugolove otopine (~100 μL) i protresite epruvete.
• Zabilježite promjenu boje iz smeđe u plavo-ljubičasto koja označava pozitivnu reakciju.
5.6.3 Dokazivanje prisutnosti proteina Bradfordovom metodom
• 11 plastičnih epruveta obilježite brojevima, stavite na stalak i u njih kapnite 3 kapi (~100 μL) sljedećih uzoraka: 1) otopine škroba, 2) otopine saharoze, 3) otopine glukoze, 4) otopine fruktoze, 5) otopine proteina BSA, 6) seruma, 7) ekstrakta krumpira, 8) razrijeđenog mlijeka, 9) otopine DNA, 10) vode u koju dodajte komadić staničevine, 11) suncokretovog ulja.
• U svaku pojedinu epruvetu dodajte 1 mL Bradfordovog reagensa i protresite epruvete.
• Zabilježite promjenu boje iz zelenkaste u plavu koja označava pozitivnu reakciju.
5.6.4 Dokazivanje prisutnosti ugljikohidrata Molischovom reakcijom
• 11 plastičnih epruveta obilježite brojevima, stavite na stalak i u njih kapnite 6 kapi (~200 μL) sljedećih uzoraka: 1) otopine škroba, 2) otopine saharoze, 3) otopine glukoze, 4) otopine fruktoze, 5) otopine proteina BSA, 6) seruma, 7) ekstrakta krumpira, 8) razrijeđenog mlijeka, 9) otopine DNA, 10) vode u koju dodajte komadić staničevine, 11) suncokretovog ulja.
• U svaku pojedinu epruvetu dodajte 6 kapi (~200 μL) otopine α-naftola i protresite epruvete.
• U svaku pojedinu epruvetu dodajte pažljivo kapaljkom uz stijenku epruvete oko 0,5 mL koncentrirane sulfatne kiseline. Pazite da ne uzmutite smjesu tijekom

BIOLOŠKA KEMIJA — PRAKTIKUM
45
dodavanja koncentrirane sulfatne kiseline. Nakon dodatka sulfatne kiseline nemojte protresati epruvete!
• Nakon 5 minuta utvrdite je li u sredini pojedine epruvete nastao ljubičasti prsten koji upućuje na pozitivnu reakciju.
5.6.5 Dokazivanje prisutnosti reduktivnih ugljikohidrata Benedictovom reakcijom
• 11 staklenih epruveta obilježite brojevima, stavite na stalak i u njih kapnite 10 kapi (~300 μL) sljedećih uzoraka: 1) otopine škroba, 2) otopine saharoze, 3) otopine glukoze, 4) otopine fruktoze, 5) otopine proteina BSA, 6) seruma, 7) ekstrakta krumpira, 8) razrijeđenog mlijeka, 9) otopine DNA, 10) vode u koju dodajte komadić staničevine, 11) suncokretovog ulja.
• U svaku pojedinu epruvetu dodajte 1 mL Benedictovog reagensa i protresite epruvete.
• Epruvete pomoću drvene hvataljke stavite u čašu s vrućom vodom i zagrijavajte do pojave narančastog obojenja (što predstavlja pozitivnu reakciju) ili do razbistravanja otopina (što predstavlja negativnu reakciju).
5.7 Zadaci
1. Koji će od navedenih uzoraka pokazivati pozitivnu reakciju s I) Lugolovom otopinom, II) Bradfordovim reagensom, III) Molischovim reagensom i IV) Benedictovim reagensom i zašto:
a. D-glukozamin b. D-glukarna kiselina c. N-acetil-D-galaktozamin d. albumin e. imunoglobulin G f. maltoza
2. Pretpostavite i objasnite rezultate pokusa u kojem a) škrob, b) celulozu i c) glikogen izlažete djelovanju α-amilaze iz sline uz prisutnost Lugolove otopine.
3. Je li u reakcijama na reduktivne ugljikohidrate moguće razlikovati D-gliceraldehid od dihidroksiacetona? Obrazložite!
5.8 Literatura
1. Voet, D. and Voet, J. (1995) Biochemistry. New York: John Wiley & Sons Inc. 2. Koolman, J. and Roehm, K.H. (2005) Color Atlas of Biochemistry. Stuttgart:
Thieme Verlag. 3. http://www.biochemweb.org/carbohydrates.shtml

BIOLOŠKA KEMIJA — PRAKTIKUM ___
46
Vježba 6.
6.1 Lipidi
Lipidi su biološke makromolekule raznolikih struktura koje, za razliku od proteina, nukleinskih kiselina i polisaharida nisu polimeri. Jedna od njihovih najvažnijih osobina je netopljivost ili vrlo slaba topljivost u vodi. Velika većina lipida su esteri u kojima je kiselinska komponenta dugolančana masna kiselina (jedna ili više), a alkoholni dio može biti trovalentni alkohol glicerol (masti i ulja), dvovalentni dugolančani aminoalkohol sfingozin (sfingolipidi), jednovalentni dugolančani alkohol (voskovi) ili jednovalentni sterolni alkohol (esteri kolesterola). Neki su lipidi izoprenoidnog porijekla: alkani karakterizirani dugim lancima s višestruko konjugiranim dvostrukim vezama (β-karoten) ili izoprenoidi višestruko ciklizirani u steroidne strukture (kolesterol i steroidni hormoni). Neki lipidi sadrže fosfatnu kiselinu (fosfolipidi) ili šećerne strukture (glikolipidi).
Masne su kiseline nerazgranate karboksilne kiseline koje sadrže 4-24 (najčešće 16 do 18) ugljikova atoma. One koje ne sadrže dvostruke veze među ugljikovim atomima nazivaju se zasićenim masnim kiselinama, dočim se one kod kojih postoje takve veze nazivaju nezasićenima (Slika 6.1.). U organizmima se rijetko nalaze u slobodnom obliku; većinom su esterificirane te su kao takve sastavnica membrana stanica i organela, a u velikim se količinama nalaze u tkivima koja služe za pohranu energije.
Slika 6.1 α-linolenska kiselina. Nezasićena masna kiselina koju nalazimo u lanenim sjemenkama. Naziva se još i ω-3 masnom kiselinom jer se prva dvostruka veza nalazi na trećem ugljikovom atomu od kraja (hidrofobni rep). Slika preuzeta s (2).
Unos masnih kiselina povezuje se s razinom kolesterola u krvi i povećanim rizikom
od razvoja srčanih bolesti (zasićene masne kiseline – „loše“, nezasićene masne kiseline – „dobre“). Zasićene masne kiseline uglavnom se nalaze u proizvodima poput mesa i mlijeka, dok se nezasićene masne kiseline najčešće nalaze u biljnim namirnicama i ribi. Stupanj nezasićenosti masnih kiselina u mastima i uljima može se odrediti reakcijama dvostruke veze - adicijom halogena ili oksidacijskim reakcijama, kao što je oksidacija pomoću KMnO4.
Lipidi

BIOLOŠKA KEMIJA — PRAKTIKUM
47
6.2 Saponifikacija
Sapuni i detergenti su amfifilne molekule koje u vodenom okružju stvaraju micele – hidrofilni dijelovi molekula okreću se prema vodi, a hidrofobni jedan prema drugome stvarajući unutrašnjost micele. Smanjujući površinsku napetost vode i emulgirajući lipide omogućavaju njihovo otapanje u vodi. Hidrofilni dio detergenta čine anionske, kationske ili neionske polarne skupine, a hidrofilni dio sapuna karboksilna skupina. Hidrofobni dio detergenta može biti raznolike strukture (primjerice linearan ili razgranati ugljikovodični lanac ili sterolni prsten deoksikolata), a kod sapuna je hidrofobni dio zapravo lanac masne kiseline (Slika 6.2). Slika 6.2 Sapuni i detergenti stvaraju micele. A) sapun (natrijev stearat) i B) detergent (natrijev-p-dodecilbenzensulfonat). Na slici A) i B) sivo su osjenčani hidrofobni dijelovi, a plavo zaokruženi i osjenčani hidrofilni dijelovi molekula. C) micela – hidrofobni dijelovi čine unutrašnjost, a hidrofilni vanjsku stranu micele koja je okrenuta prema vodi.
Sapuni su, dakle, soli (većinom natrijeve i kalijeve) masnih kiselina. Nastaju reakcijom saponifikacije koja uključuje hidrolizu triacilglicerola i neutralizaciju masnih kiselina jakom lužinom (Slika 6.3). U pripremi sapuna najčešće se koriste trigliceridi životinjskog porijekla, iako se mogu koristiti i biljne masti, kao što su kokosovo, palmino ili drugo biljno ulje.
Slika 6.3 Saponifikacija. A) Slijed reakcija kojima iz triacilglicerola (estera glicerola i tri masne kiseline) uz dodatak lužine nastaje sapun (sol masnih kiselina) i u konačnici alkohol (glicerol). B) Kemijski prikaz ukupne reakcije saponifikacije kojom iz 1,3-distearil-2-palmitil-glicerata nastaju glicerol, natrijev palmitat i dvije molekule natrijevog stearata, odnosno tri molekule sapuna.
C) A)
B)
A)
monoacilglicerol
sapun sapun lužina lužina
diacilglicerol
sapun + + + triacilglicerol
lužina
glicerol
B)
C17H33
C15H31
C17H33
CH2O
CHO
CH2O
+ 3 NaOH
C
O
C
O
C
O
CH2OH
CHOH
CH2OH
+ 2 C17H33COO-Na
+ + C15H31COO
-Na
+

BIOLOŠKA KEMIJA — PRAKTIKUM ___
48
6.3 Zadaća
Ekstrahirajte lipide iz uzorka čokolade i lanenih sjemenki acetonom te odredite stupanj nezasićenosti ekstrahiranih lipida oksidacijom pomoću KMnO4.
Napravite sapun miješanjem jake lužine i jestivog ulja te ispitajte učinkovitost otapanja ulja tako priređenog sapuna i komercijalno pribavljenog detergenta.
6.4 Reagencije
• 20% otopina NaOH u dest. vodi • biljno ulje • 5 M NaCl u dest. vodi • dest. voda ohlađena na 4°C • detergent • aceton • 2% otopina KMnO4 u dest. vodi • mliječna čokolada • lanene sjemenke
6.5 Pribor
• staklene čaše (10 mL i 400 mL) • magnetska miješalica s grijačem • magnet za miješanje • stakleni štapić • stakleni lijevak • filter papir • Erlenmayerova tikvica • staklene epruvete • vaga • kapaljka
6.6 Načelo postupka
Lipidi su topljivi u nepolarnim otapalima pomoću kojih se mogu ekstrahirati iz smjese.
Stupanj nezasićenosti masne kiseline može se odrediti pomoću KMnO4, koji oksidira dvostruku vezu između ugljikovih atoma masne kiseline.
Reakcijom jake lužine i ulja nastaje sapun. Sapuni i detergenti omogućavaju otapanje lipida u vodi. 6.7 Postupak
6.7.1 Ekstrakcija lipida i odreñivanje stupnja nezasićenosti masnih kiselina
• U tarioniku usitnite čokoladu te 0,5 g tako usitnjene čokolade prebacite u označenu staklenu čašicu.
• U tarioniku usitnite lanene sjemenke te 0,5 g tako usitnjenih sjemenki prebacite u drugu označenu staklenu čašicu.

BIOLOŠKA KEMIJA — PRAKTIKUM
49
• U svaku pojedinu čašu dodajte 5 mL acetona i smjese miješajte staklenim štapićem oko 1 minutu.
• Filtriranjem kroz filter papir postavljen na stakleni lijevak sakupite topljive frakcije tih dvaju smjesa u dvije suhe epruvete.
• U treću suhu epruvetu dodajte volumen acetona jednak onima koji imate u drugim epruvetama. Taj aceton služi kao slijepa proba.
• U sve tri epruvete dodajte 1 kap 2% KMnO4, pratite promjene boje tijekom 10 minuta i objasnite ih.
6.7.2 Saponifikacija
• Pomiješajte 15 mL 20% otopine NaOH i 10 mL suncokretovog ulja u staklenoj čaši od 400 mL.
• Dodajte magnet za miješanje i zagrijavajte na magnetskoj miješalici uz konstantno miješanje. Poželjno je da smjesa lagano kipi, a u slučaju prejakog kipljenja, smanjite zagrijavanje. Zagrijavajte dok ne nastane gusta smjesa (približno 30 minuta).
• Smjesu maknite s grijača i u nju odmah ulijte 150 mL 5 M otopine NaCl. • Miješajte staklenim štapićem nekoliko minuta dok se sapun ne odvoji od stijenki
čaše. • Filtrirajte kroz obični filter papir u Erlenmayerovu tikvicu uz dva dodatna
ispiranja s po 100 mL vode koju ste prethodno ohladili u frižideru. 6.7.3 Osobine sapuna i detergenta
• U tri epruvete ulijte po 5 mL destilirane vode, a u druge tri epruvete po 5 mL tvrde vode (u tu svrhu koristite vodovodnu vodu).
• U drugu i petu epruvetu dodajte malo (vrh špatule) pripremljenog sapuna. • U treću i šestu epruvetu dodajte 100 μL detergenta. • Zagrijavajte svih šest epruveta. Drugu i petu epruvetu povremeno promiješajte
staklenim štapićem. • U svih šest epruveta dodajte po tri kapi ulja. • Promatrajte što se događa te zapišite svoja zapažanja.
6.8 Zadaci
1. Usporedite temperaturu taljenja zasićenih i nezasićenih masnih kiselina. 2. Usporedite maslinovo ulje i maslac obzirom na udio zasićenih i nezasićenih
masnih kiselina. 3. U mrkvi se, između ostalog, nalazi i β-karoten, za koji je karakteristična
izoprenska struktura. Ukoliko naribanu mrkvu stavite u navedena otapala, u kojima će β-karoten biti topljiv i po čemu to možete zaključiti:
a. destilirana voda b. fiziološka otopina c. 50% etanol u destiliranoj vodi d. 96% etanol e. petroleter

BIOLOŠKA KEMIJA — PRAKTIKUM ___
50
f. suncokretovo ulje 4. Podrobno objasnite mehanizam djelovanja sapuna i detergenta, odnosno princip
na kojem se temelji primjena sapuna i detergenata kao sredstava za čišćenje.
6.9 Literatura
1. Voet, D. and Voet J. (1995) Biochemistry. New York: John Wiley & Sons Inc. 2. http://biology.nicerweb.com/med/Alpha-linolenic_acid.gif 3. http://www.cem.msu.edu/~reusch/VirtualText/lipids.htm 4. http://chemistry.about.com/od/cleanerchemistry/a/how-soap-cleans.htm

BIOLOŠKA KEMIJA — PRAKTIKUM
51
Vježba 7.
7.1 Deoksiribonukleinska kiselina
Deoksiribonukleinska kiselina (DNA) je vrlo dugačka, nitasta molekula sastavljena od velikog broja deoksiribonukleotida. Oni su izgrađeni od dušičnih baza te šećera i fosfata. Purinske ili pirimidinske baze u DNA nositelji su genske informacije, dočim deoksiriboze i fosfatne skupine imaju strukturnu ulogu. Okosnica DNA, konstantna duž cijele molekule, sastoji se od deoksiriboza povezanih fosfodiesterskim vezama. Fosfodiesterska veza povezuje 3'-hidroksilnu skupinu jedne deoksiriboze s 5'-hidroksilnom skupinom susjedne. U sastavu molekule DNA nalazimo četiri različite dušične baze: purinske baze adenin (A) i gvanin (G) te pirimidinske baze timin (T) i citozin (C). DNA se sastoji od dva polinukleotidna lanca čije smjerove označavamo 5'-3'. Lanci uvijaju oko zajedničke osi i međusobno su antiparalelni. Baze su smještene u unutrašnjosti uzvojnice, a fosfatne i šećerne jedinice u vanjskom dijelu. Lanci su povezani vodikovim vezama koje stvaraju baze u unutrašnjosti uzvojnice te van der Waalsovim interakcijama među bazama usporedno s osi uzvojnice. Planarne bazne parove čine adenin i timin povezani dvjema vodikovim vezama te gvanin i citozin povezani trima vodikovim vezama. Tu pojavu nazivamo komplementarnošću baznih parova. 7.2 Denaturacija DNA
Ukoliko se vodikove veze među bazama razore uslijed izlaganja povišenoj temperaturi, promjeni pH ili promjeni ionske jakosti, dva se lanca DNA odvajaju te DNA prelazi iz konformacije dvostruke uzvojnice u konformaciju nasumičnog klupka (Slika 7.1). Taj se proces naziva denaturacija.
Slika 7.1 Shematski prikaz denaturacije DNA. Preuzeto i prilagođeno iz (1).
Termička denaturacija DNA (sekvenciranje

BIOLOŠKA KEMIJA — PRAKTIKUM ___
52
Denaturacija je praćena kvalitativnim promjenama fizikalnih svojstava DNA. Primjerice, karakteristična velika viskoznost otopine nativne DNA, koja je posljedica krute strukture dvostruke uzvojnice, izrazito se smanjuje kada DNA prelazi u oblik razmjerno slabo međusobno povezanih jednostrukih lanaca. Prilikom denaturacije događaju se i promjene optičke gustoće. Heterociklički prstenovi nukleotida jako apsorbiraju svjetlost ultraljubičastog područja (s maksimumom blizu 260 nm, što je karakteristično za pojedine baze). Međutim, apsorpcija odgovarajuće smjese slobodnih nukleotida je oko 40% viša od apsorpcije molekule DNA. Ta se pojava naziva hiperkromizam i rezultat je gubitka interakcija među elektronskim sustavima gusto složenih baza u DNA. Svaki poremećaj dvostruke uzvojnice ogleda se kao povećavanje optičke gustoće prema vrijednostima karakterističnim za slobodne baze (Slika 7.2 A)).
A) B)
Slika 7.2 A) Apsorpcijski spektar u UV području nativne i termički denaturirane DNA iz
bakterije Escherichia coli. Denaturacija ne mijenja značajno oblik apsorpcijske krivulje, već samo povećava njezinu vrijednost. B) Krivulja mekšanja DNA. Preuzeto i prilagođeno iz (1).
Hiperkromni pomak se pri određenoj valnoj duljini (najčešće 260 nm) odvija unutar uskog temperaturnog područja. To znači da je denaturacija DNA kooperativni proces pri čemu razaranje dijela strukture destabilizira ostatak i potiče daljnju denaturaciju. Krivulja mekšanja je prikaz ovisnosti apsorbancije otopine DNA o temperaturi (Slika 7.2 B). Iz nje se može očitati temperatura pri kojoj je postignuto 50% denaturacije i koju nazivamo temperaturom mekšanja, Tm.
Stabilnost dvostruke uzvojnice DNA, a time i njezin Tm, ovisi o nekoliko čimbenika kao što su sastav baza DNA, ionska jakost i pH otopine te prisutnost drugih molekula koje se vežu na DNA. Veličina Tm linearno je razmjerna molarnom udjelu baznih parova GC. Tri vodikove veze koje povezuju gvanin i citozin stabilnije su od dviju vodikovih veza između adenina i timina. Tm se povećava za oko 0,4°C ako se udio GC baznih parova poveća 1%. Kada se DNA nalazi u otopini pri približno fiziološkim uvjetima,

BIOLOŠKA KEMIJA — PRAKTIKUM
53
vrijednost Tm je obično unutar područja 85-95°C (DNA koja sadrži 40% GC parova, što je vrijednost svojstvena genomu sisavaca, pri fiziološkim uvjetima ima Tm oko 87°C, dok DNA koja sadrži 60% GC parova, pod istim uvjetima ima Tm oko 95°C.) Stoga je bez intervencije staničnog sustava dvolančana DNA stabilna unutar stanice.
Homogene DNA (primjerice virusne ili plazmidne) koje se sastoje od identičnih kratkih dijelova, denaturiraju se u uskom rasponu promjene temperature. Za razliku od homogenih, heterogene DNA sadrže mnoštvo različitih fragmenata nastalih nasumičnim kidanjem kromosoma. Kako se Tm tih fragmenata razlikuju, temperaturni raspon pri kojem se takva DNA denaturira je širi.
Na temperaturu mekšanja utječu i ionska jakost i vrijednost pH. Međusobno odbijanje negativnih naboja, koje pri fiziološkim uvjetima imaju fosfatne skupine, destabilizira uzvojnicu DNA. Kako je elektrostatsko odbijanje istoimenih naboja ovisno o dielektričnoj konstanti otopine, povećanjem ionske jakosti povećava se dielektrična konstanta te se smanjuje međusobno odbijanje fosfatnih skupina. Uzvojnica DNA se stabilizira, krivulja denaturacije se sužava i Tm raste. Promjena pH koja uzrokuje protoniranje ili deprotoniranje baza narušava vodikove veze među njima te dovodi do denaturacije DNA. Zapravo, bilo koja molekula koja može stvoriti vodikove veze s funkcionalnim skupinama baza u DNA ili omesti slaganje baza jedne iznad druge može pridonijeti denaturaciji uzvojnice. Takve su molekule, primjerice, urea i formamid, ali i proteini koji se vežu na jednolančanu DNA.
Denaturacija DNA je reverzibilan proces. Ukoliko se promijenjeni parametri sustava koji su uzrokovali denaturaciju DNA (primjerice temperatura) vrate na početne vrijednosti, DNA se renaturira. Ako se otopina denaturirane DNA naglo ohladi na temperaturu znatno nižu od Tm, nastala DNA bit će samo djelomično dvolančana poradi toga što komplementarni lanci neće imati dovoljno vremena da se u u otopini susretnu i pravilno spare prije nego što se tako djelomično dvolančana struktura "smrzne" u otopini. Ako se, pak, otopina denaturirane DNA polagano hladi do temperature 20-25°C manje od Tm, bit će dovoljno toplinske energije da se kratkim, pogrešno sparenim dijelovima DNA omogući razdvajanje i ispravno sparivanje te na taj način stvaranje duljih komplementarnih sljedova. Oni se na toj temperaturi više ne mogu razdvojiti te se DNA postupno u cijelosti renaturira, odnosno zauzima nativnu konformaciju. Slično tome, komplementarni lanci RNA i DNA u procesu hibridizacije tvore dvolančane RNA-DNA hibride. Dvostruka uzvojnica odgovarajuće DNA samo je neznatno stabilnija od tako nastalih hibrida.
Denaturacija i renaturacija DNA bitne su za biološku ulogu ove molekule pri procesima replikacije i transkripcije. Tm kao mjera denaturacije DNA važan je čimbenik za mnoge eksperimentalne metode koje uključuju hibridizacije nukleinskih kiselina (primjerice, lančana reakcija polimeraze, eng. polymerase chain reaction - PCR). 7.3 Zadaća
Postupno zagrijavajte uzorak DNA do temperature 92°C i pratite denaturaciju mjerenjem apsorbancije pri valnoj duljini 260 nm. Odredite Tm iz krivulje mekšanja.

BIOLOŠKA KEMIJA — PRAKTIKUM ___
54
7.4 Reagencije
• pufer SSC/10 (15 mM natrijev klorid, 1,5 mM natrijev citrat, u dest. vodi , pH 7,0 - 7,1) – standardni pufer za eksperimente denaturacije
• otopina DNA izolirana iz kravljeg timusa u puferu SSC/10 – uzorak
7.5 Pribor
• plastične epruvetice od 1,5 mL • stalak za epruvete • automatske pipete • nastavci za automatske pipete • vodena kupelj • posuda sa smjesom leda i vode • UV spektrofotometar • kvarcna kiveta
7.6 Načelo postupka
Izlaganjem povišenoj temperaturi DNA se denaturira. Termičku denaturaciju DNA moguće je pratiti spektrofotometrijski, mjereći A260 u temperaturnom rasponu od sobne temperature do temperature potpune denaturacije.
Budući da dostupan spektrofotometar nema termostatiranu kivetu koja bi se mogla postupno zagrijavati, uzorci DNA se zagrijavaju do određene temperature, a zatim naglo hlade u smjesi leda i vode. Pri tome dolazi do renaturacije dijela lanca. Krivulja koja se konstruira iz dobivenih podataka u korelaciji je s krivuljom koja bi se dobila kao rezultat pokusa s termostatiranom kivetom. 7.7 Postupak
• Po 1 mL otopine DNA otpipetirajte u 10 plastičnih obilježenih epruveta. • Epruvete stavite u stalak i uronite u vodenu kupelj. • Kupelj postupno zagrijavajte do 92°C. Kada se dostigne željena temperatura, po
jednu epruvetu izvadite i odmah ohladite u smjesi leda i vode. Predlažu se sljedeće temperature (°C):
25 40 55 65 70 75 80 84 88 92
• U prikladnu tablicu upisujte stvarnu temperaturu pri kojoj je uzorak izvađen. • Nakon što je prikupljen i dobro ohlađen i posljednji uzorak, uzorcima redom
izmjerite A260, na milimetarskom papiru nacrtajte graf ovisnosti apsorbancije o temperaturi te odredite temperaturu mekšanja (budući da se ona razlikuje od vrijednosti koju biste dobili mjerenjem kod stvarnih temperatura, označite je kao Tm0).

BIOLOŠKA KEMIJA — PRAKTIKUM
55
7.8 Zadaci
1. Koje sile združuju dva lanca molekule DNA? 2. U 0,2 M otopini natrijevog klorida Tm DNA je 75°C. U drugoj otopini ista DNA
ima Tm od 100°C. Kakva svojstva ima nepoznata otopina u odnosu na 0,2 M otopinu natrijevog klorida?
7.9 Literatura
1. Voet, D. and Voet, J. (1995) Biochemistry. New York: John Wiley & Sons Inc.





























