Embed Size (px)
Citation preview

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 1/8
Our reference: JRI 1979 P-authorquery-v8
AUTHOR QUERY FORM
Journal: JRI Please e-mail or fax your responses and any corrections to:
E-mail: [email protected]
Article Number: 1979 Fax: +353 6170 9272
Dear Author,
Please check your proof carefully and mark all corrections at the appropriate place in the proof (e.g., by using on-screen
annotation in the PDF file) or compile them in a separate list.
For correction or revision of any artwork, please consult http://www.elsevier.com/artworkinstructions .
Any queries or remarks that have arisen during the processing of your manuscript are listed below and highlighted by flags in
the proof. Click on the ‘Q’ link to go to the location in the proof.
Location in Query / Remark: click on the Q link to go
article Please insert your reply or correction at the corresponding line in the proof
Reference(s) given here were noted in the reference list but are missing from the text – please position
each reference in the text or delete it from the list.
Q1 Uncited reference: This section comprises reference that occurs in the reference list but not in the body
of the text. Please position each reference in the text or, alternatively, delete it. Any reference not dealt
with will be retained in this section.
Thank you for your assistance.

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 2/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
Journal of Reproductive Immunology xxx (2010) xxx–xxx
Contents lists available at ScienceDirect
Journal of Reproductive Immunology
j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / j r e p r i m m
Decidualization and angiogenesis in early pregnancy: unravelling thefunctions of DC and NK cells
Sandra M. Blois ∗, Burghard F. Klapp, Gabriela Barrientos
Charité Centrum 12 für Innere Medizin und Dermatologie, Reproductive Immunology Research Group, University Medicine of Berlin, Germany
a r t i c l e i n f o
Article history:
Received 1 September 2010
Received in revised form 29 October 2010
Accepted 4 November 2010
Available online xxx
Keywords:
Dendritic cells
NK cells
Decidualization
Decidual angiogenesis
Implantation
a b s t r a c t
Differentiation of endometrial stromal cells and formation of new maternal blood vessels
at the time of embryo implantation are critical for the establishment and maintenance of
gestation. The regulatory functions of decidual leukocytes during early pregnancy, partic-
ularly dendritic cells (DC) and NK cells, may be important not only for the generation of
maternal immunological tolerance but also in the regulation of stromal cell differentiation
and the vascular responses associated with the implantation process. However, the specific
contributions of DC and NK cells duringimplantation arestill difficult to dissect mainly due
to reciprocal regulatory interactions established between them within the decidualizing
microenvironment. The present review article discussescurrent evidence on the regulatory
pathways drivingdecidualization in mice, suggesting thatNK cells promoteuterinevascular
modifications that assist decidual growth but DC directly control stromal cell proliferation,
angiogenesis and the homing and maturation of NK cell precursors in the pregnant uterus.
Thus, successful implantation appears to result from an interplay between cellular compo-
nents of the decidualizing endometrium involving immunoregulatory and pro-angiogenicfunctions of DC and NK cells.
© 2010 Published by Elsevier Ireland Ltd.
1. Introduction
In mice, the implantation of a blastocyst into the uterus
initiates a transformation process, known as decidualiza-
tion, and creation of an extensive vascular network within
thestromal bed, leading to the establishment of pregnancy.
Despite some differences among species, the onset of
implantation always involves complex regulatory mecha-nisms operating on the uterine lining to make it competent
for blastocyst attachment. The steroid hormones, estrogen
and progesterone play a central role during this process,
driving a series of cellular and molecular interactionsin the
∗ Corresponding author at: Charité Centrum 12 für Innere Medizin
und Dermatologie, Reproductive Immunology Research Group, Univer-
sity Medicine of Berlin, BMFZ-Raum 2.0547, Campus Virchow Klinikum,
Augustenburger Platz 1, 13353 Berlin, Germany. Tel.: +4930 450 553791,
fax: +49 30 450 553962.
E-mail address: [email protected](S.M. Blois).
underlying endometrial stroma to form thedecidua, a tran- 28
sient tissue that supports embryo growth and maintains 29
early pregnancy (Dey et al., 2004). 30
Accumulating evidence suggests that immune cells are 31
an important regulatory component within the uterine 32
milieu during early pregnancyin mice. Decidual leukocytes 33
have been extensively characterized with the initial aim of 34
gaining a clear understanding of the regulation of mater- 35
nal tolerance towards fetal alloantigens (Blois et al., 2007), 36
but may also exert regulatory functions during early preg- 37
nancy that go beyond their classical roles as components 38
of the immune system. In particular, recent studies have 39
highlighted the importance of DC (Krey et al., 2008; Plaks 40
et al., 2008) and NK cells (Croy et al., 2003a; Hanna et al., 41
2006) in the regulation of stromal cell differentiation and 42
the vascular responses associated with the implantation 43
process. The intimate contact observed between DC and 44
NK cells, which are often found in the vicinity of decidual 45
blood vessels (Blois et al., 2008; Kammerer et al., 2003), 46
0165-0378/$ – see front matter © 2010 Published by Elsevier Ireland Ltd.doi:10.1016/j.jri.2010.11.002

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 3/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
2 S.M. Blois et al. / Journal of Reproductive Immunology xxx (2010) xxx–xxx
suggests that reciprocal interactions between them may7
also be important in shaping the decidualizing microen-8
vironment during early pregnancy. The first insights have9
been provided by in vitro studies showing that the prolif-0
eration of uterine cells is highly dependent on a synergistic
effect of DC and NKcells(Blois et al., 2008), suggesting that2
such DC-NK cell cross-talk may be important in driving the3
differentiation of the endometrial stroma during implanta-4
tion. In turn, DC appear to play a pivotal role in promoting5
the differentiation of uterine NK (uNK) cells, as highlighted6
by recent in vivo studies describing impaired NK cell func-7
tions in association with decreased levelsof IL-15 and IL-128
in DC depleted implantationsites(Karsten et al., 2009; Krey9
et al., 2008).0
The decidua serves a variety of functions including
the production of growth factors and cytokines to sustain2
embryo development, the regulation of maternal immune3
responses and the control of trophoblast invasion into the4
uterus. The timely coordination of the endometrial changes5
occurring during decidualization is thus critical for the6
establishment of pregnancy, and the elucidation of the cel-7
lular and molecular mechanisms involved may greatly aid8
the design of new therapeutic approaches to pregnancy9
failure. While the morphological changes occurring in the0
endometrialstroma and the hormonal pathways regulating
them are well described aspects that define the decidual-2
ization process, the above evidence points to an important3
role played by DC and NK cells that needs to be further4
explored. Thus, the present review article integrates our5
recent studies on uterine DC physiology with a discussion6
of current knowledge on regulatory pathways operating7
at the endometrium during early pregnancy. Particular8
emphasis will be given to thecontributions of DC to stromal9
cell proliferation and differentiation as the most promi-0
nent feature of the decidualization process, as well as their
interplay with the NK cell subset in promoting vascular2
remodeling and angiogenesis associated with it.3
2. Implantation and decidualization4
Implantation is the process by which the blastocyst5
assumes a fixed position andestablishes an intimate physi-6
ological relationship with the uterine endometrium. Rather7
than a single event, implantation can be seen as a series8
of three stages (apposition, attachment and penetration)9
forming a continuum (Schlafke and Enders, 1975). Appo-0
sition denotes the first intimate contact of the blastocyst
with the endometrium, which in mice can be evidenced as2
a generalized stromal edema that precedes the closure of 3
theuterine lumen at gestationday (Gd)4.5. This is followed4
by increased vascular permeability at the implantation site5
and apoptosis of the epithelial cells during the attachment6
reaction, resulting in a closer contact and the subsequent7
penetration through the endometrial stroma in a regulated8
manner (Dey et al., 2004).9
Blastocyst attachment always occurs in the antime-0
sometrial side of the mouse uterus and triggers the
proliferation of the stromal cells adjacent to the implan-2
tation site, initiating a unique cell differentiation process3
known as decidualization. By Gd 5.5, the stromal cells4
immediately surrounding the embryo differentiate to form5
a primary decidual zone (PDZ) while cell proliferation con- 106
tinues towards the mesometrial compartment, eventually 107
establishing a secondary decidual zone (SDZ) representa- 108
tive of the terminal differentiation of the decidual tissue at 109
Gd 7.5 (Dey et al., 2004). Though the normal stimulus for 110
decidualization is the attachment reaction, certain agents 111
(i.e., oil) can induce a similar process (resulting in decid- 112
uoma formation) when applied locally to pseudopregnant 113
or hormonally treated mice (Hirabayashi et al., 1999; Paria 114
et al., 2001). 115
Regardlessof the initiatingstimulusinvolved, the decid- 116
ualization process is always associated with extensive 117
proliferation and differentiation in the stromal compart- 118
ment, rendering large decidual cells often with binucleated 119
or polyploid status. This is the result of an atypical cell 120
cycle driving DNA replication without cell division, which 121
is tightly regulated through a complex interplay of cyclins, 122
cyclin-dependent kinases (CDKs) and their inhibitors as 123
the classical mediators controlling cell cycle progression 124
in mammalian cells. In mice, cyclin D3 expression at the 125
implantation sitehas been associatedwith the proliferation 126
of decidualizing stromal cells, probably through functional 127
association with cdk4 (Das et al., 1999; Tan et al., 2002). 128
Later on, persistent cyclin D3 expression concomitant with 129
a switch from cdk4 to cdk6 expression and the induction 130
of the p21 inhibitor is thought to drive the terminal differ- 131
entiation of the endometrial stroma. The involvement of 132
cell cycle regulatory proteins in this process is further sup- 133
ported by the finding that the expression of several of these 134
molecules (i.e., cyclin D3 and p21) in the decidua depends 135
on IL-11, a cytokine that has been found to be indispens- 136
able for normal decidualization (Li et al., 2008; Robb et al., 137
1998). 138
It is still difficult to determine the physiological role 139
played by stromal cell polyploidy during the decidualiza- 140
tion process. One of the many functions of the decidua 141
is to support embryo growth during early pregnancy and 142
in this context, polyploidy may increase the biosynthetic 143
capacity of decidual cells by increasing the number of gene 144
copies for transcription. Polyploidization eventually leads 145
to apoptosis of the differentiating cells and may therefore 146
be implicated in the transient nature of the decidual tis- 147
sue, limiting the life span of these cells to allow placental 148
expansion and development (Dey et al., 2004). 149
2.1. Uterine NK cells and decidualization: what comes 150
first? 151
Uterine NK cells are probably the best studied leukocyte 152
population in the mouse and human decidua. In contrast to 153
the human endometrium, where these cells emerge post- 154
ovulation, mature NK cells do not appear in the mouse 155
uterus prior to implantation (King, 2000). Decidual IL-15 156
drivesthe maturation of NK cell precursors into large, gran- 157
ulated cells that gradually accumulate in the mesometrial 158
compartment and start producing detectable levels of IFN 159
at Gd 6.5 (Ashkar et al., 2003; Croy et al., 2003b). 160
Although they differ in the dynamics of their recruit- 161
ment, human and mouse uNK cells are the most abundant 162
leukocyte subset at the time of decidualization and are 163
therefore likely to influence this process. Indeed, uNK cell 164

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 4/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
S.M. Blois et al. / Journal of Reproductive Immunology xxx (2010) xxx–xxx 3
deficiency in mice has often been associated with decid-
ual growth defects including hypocellularity, necrosis and
edema (Ashkar et al., 2000, 2003; Guimond et al., 1997).
Besides recent in vitro studies showing that uNK cell-
derived factors can influence the gene expression profile
of human endometrial fibroblasts (Germeyer et al., 2009),
evidence to support a direct effect exerted by uNK cells
on stromal cell differentiation is still largely elusive. In
mice, the major defects associated with a lack of uNK cell
signaling during early pregnancy are in fact observed in
the decidual vasculature, showing undilated spiral arteries
that retain their smooth muscle coats throughout gestation
(Ashkar et al., 2000). Thus, a possible scenario is that rather
than directly affecting the differentiation of stromal cells,
the main contribution of uNK cells may be to induce vascu-
lar changes that ensure an adequate blood flow to sustain
tissue growth during the decidualization process.
Vascular modifications induced by uNK cells may gain
particular importance as gestation advances in promoting
placental development. Indeed, implantation in mice pro-
gresses normally in theabsence of uNKcellsand fetal loss is
not detected prior to Gd 10.5 in association with an abnor-
mal architecture of the placental tissues (Guimond et al.,
1997). Gene knock-out modelshave recently demonstrated
that many molecules originally associated with infertil-
ity due to implantation and decidualization failure (i.e.,
IL-11 and homeobox A10) can also affect the recruitment
and maturation of NK cells in the uterus (Ain et al., 2004;
Rahmanet al., 2006). In thiscontext, the earlyaccumulation
of uNK cells should not be seen as a mechanism promot-
ing decidualization but as one of the many endometrial
changes that occur during this process to support normal
pregnancy progression.
The above evidence strongly supports an association
between NK cell functions and the endometrial changes
occurring during early pregnancy, but raises many ques-
tions as to their specific contributions to the different
aspects involved in thisprocess. While vascular remodeling
and development within the decidual bed is most likely to
be regulated by uNK cell derived signals, their direct effects
on endometrial stromal cells are less evident but cannot be
precluded. Therefore, ongoing research in our lab aims at
characterizing uNK cell effects on stromal cell differentia-
tion to provide further insights into the input of this subset
on endometrial decidualization during the early stages of
pregnancy.
2.2. Uterine conventional DC: upstream regulators of
stromal cell differentiation
Resembling other mucosal surfaces, the decidua is
enriched with MHC class II+ antigen presenting cells rep-
resenting more than 10% of the leukocyte population (Blois
et al., 2004; Kammerer, 2005). The most prominentof these
are CD11c+ DC, which have been extensively character-
ized in mice and humans in terms of their potential to
induce immune tolerance during pregnancy (Blois et al.,
2007; Kammerer et al., 2008). DC become most abundant
in the mouse uterus at Gd 5.5, where this early accumula-
tion has been linked to the transient inflammatory milieu
characteristic of the implantation window (Blois et al.,
Fig.1. Schematic representation showinga putativenetworkof DCNKcell
interactions during the early stages of pregnancy. DC directly modulate
the proliferation and differentiation of stromal cells and the angiogenic
expansionof the endometrialvascularbed to sustaintissue growth during
decidualization. DC-derived IL-15 would promote the recruitment andmaturation of NK cell precursors that in turn cooperate in the regulation
of vascular development and remodeling through the production of pro-
angiogenic (e.g. VEGF) and vasoactive mediators.
2004). Besides theseclassicalimmunoregulatory functions, 224
recent studies suggest that DC are a pivotal component 225
of the embryo-maternal dialogue driving implantation by 226
directly influencing uterine receptivityand decidualization 227
(Fig. 1; Krey et al., 2008; Plaks et al., 2008). These stud- 228
ies demonstrated that the absence of DC from the mouse 229
uterus during the peri-implantation period results in early 230
pregnancy failure due to impairments in the decidualiza- 231
tion process including defective stromal cell proliferation, 232
abnormal differentiation of the mesometrial and antime- 233
sometrial compartments and reduced vascularization of 234
the implantation sites. Importantly, the defects associated 235
with DC depletion were also observed upon the induction 236
of deciduoma and in syngeneic pregnancies (Plaks et al., 237
2008), pointing to a key role of DC as promoters of stro- 238
mal cell differentiation that functions independently from 239
tolerance induction. 240
Experiments in vitro involving the co-culture of tro- 241
phoblasts with DC, NK cells and uterine cells isolated from 242
murine allogeneic pregnancies showed that trophoblasts 243
boosted the proliferation rate of uterine cells only when 244
cultured in the presence of both DC and NK cells (Blois 245
et al., 2008), providing the first insights into a role played 246
by DC in driving decidualization in cooperation with NK 247
cells. However, the interactions between DC and NK cells 248
underlying this requirement for both cell subsets in the 249
promotion of stromal cell proliferation await further char- 250
acterization. One possibility is thatthere exists a synergistic 251
effect involving direct actions on stromal cells mediated by 252
both DC and NK cells. Alternatively, this observation could 253
reflect a hierarchical event in which DC directly promote 254
stromal cell growth and differentiation during decidual- 255
ization and their physical interactions with NK cells within 256
the uterus would be involved in the regulation of this pro- 257
cess. In this regard, the selective depletion of DC in vivo 258
has also been associated with impairments in the recruit- 259

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 5/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
4 S.M. Blois et al. / Journal of Reproductive Immunology xxx (2010) xxx–xxx
mentandmaturationofuNKcells(Karsten et al., 2009; Krey0
et al., 2008), which could represent a self-directed con-
trol mechanism induced by DC during the decidualization2
process.3
Uterine DC do not differ from those found in other4
tissues in being a highly heterogeneous cell popula-5
tion (Kammerer et al., 2008). However, evidence on6
the contribution of different decidual DC subsets to the7
establishment of pregnancy remains largely elusive. The8
administration of diphtheria toxin (DT) to CD11c-DTR9
transgenic mice results in the selective elimination of 0
conventional DC (CD11chigh DC) ( Jung et al., 2002), but
spares plasmacytoid DC (pDC) because of their low lev-2
els of CD11c expression. Therefore, it can be assumed3
that the regulatory functions ascribed to DC during mouse4
endometrial decidualization do not rely on cells belong-5
ing to the plasmacytoid subset. Indeed, we corroborated6
this result using the 120G8 monoclonal antibody, which7
reacts with bone marrow stromal Ag-2 (BST-2) expressed8
by pDC, as an alternative approach to deplete this subset9
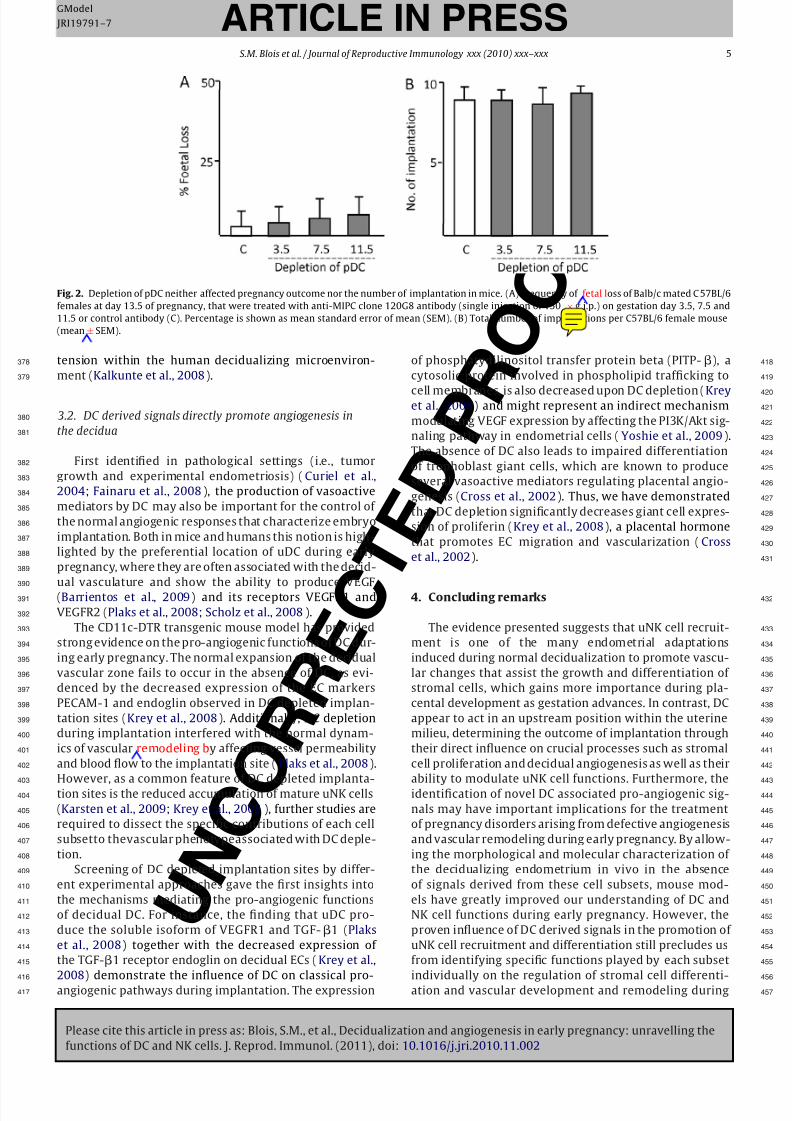
in vivo (Asselin-Paturel et al., 2003). We chose to assess0
not only the role of pDC during implantation (Gd 3.5),
but also in peri-implantation (Gd 7.5) and late pregnancy2
(Gd 11.5). As shown in Fig. 2, depletion of pDC neither3
affected pregnancy outcome nor the number of implanta-4
tion sites,suggesting that pDC arenot involved in anyof the5
mentioned processes during gestation in mice. However,6
it is important to note that pDC numbers were gradually7
restored after 48 h during the antibody-mediated deple-8
tion approach. Thus, the recently generated blood dendritic9
cell antigen-2 (BDCA-2)-DTR transgenic mouse (Swiecki0
and Colonna, 2010) represents a unique tool to further
investigate the role of pDC during gestation as repeated2
administration of DT does not appear to be lethal in this3
model, allowing long term depletion of these cells with4
high efficacy and specificity. In addition,further studies will5
be required to dissect the role of individual CD4+ or CD8+6
CD11chigh populations in implantation outcome as well as7
the underlying mechanisms involved.8
3. Angiogenesis and implantation9
The establishment of pregnancy is highly dependent on0
theproper coordinationof several vascular processesat the
maternal–fetal interface to ensure an adequate blood flow2
in response to the increasing metabolic demands of the3
embryo. Implantation and decidualization occur together4
with an extensive remodeling of the endometrial vascula-5
ture, during which vascular expansion through the process6
of angiogenesis plays a central role (Torry et al., 2007).7
Angiogenesis is a rare phenomenon in most adult tis-8
sues, where a predominance of angiostatic signals renders9
vascular endothelial cells (ECs) quiescent (Distler et al.,0
2003). One of the few exceptions is the female repro-
ductive tract, in which a fine balance between pro- and2
antiangiogenic signals determines EC activation as part3
of a physiological response during the reproductive cycle4
and pregnancy. The pivotal importance of this process for5
reproduction is demonstrated by the finding that a sin-6
gle dose of anti-angiogenic compounds (i.e., AGM-1470)7
applied either before or during implantation disrupts8
decidualization and placental development in mice, result- 319
ing in embryo resorption (Klauber et al., 1997). Decidual 320
angiogenesis however, differs from stromal decidualiza- 321
tion in depending on the presence of the embryo (Ma et al., 322
1997), which highlights the importance of a coordinated 323
action of signals derived both from the maternal and fetal 324
compartments in the control of vascular expansion during 325
early pregnancy. 326
3.1. Uterine NK cells support vascular remodeling and 327
angiogenesis during early pregnancy 328
Angiogenesis involves the expansion of pre-existing 329
vessels through proliferation of ECs and their subsequent 330
migration and differentiation to form new capillary tubes 331
(Distler et al., 2003). A prior requirement to EC proliferation 332
is an increased vascular permeability and their detachment 333
from smooth muscle cells and extracellular matrix com- 334
ponents. While not representing angiogenesis per se, the 335
initial stages of spiralartery remodelingas observed during 336
mouse and human early pregnancy also involve the abla- 337
tion of the endothelium and smooth muscle of the arterial 338
walls as well as the reorganization of extracellular matrix 339
components. This process appears to be mainly mediated 340
by uNK cellderivedIFN, asthiscytokine has been found to 341
amelioratethe decidual phenotype observed in the absence 342
of uNK cells (Ashkar et al., 2000). Besides impeding an ade- 343
quate blood flow to the implantation site the undilated 344
vessels observed in uNK cell deficient mice would also 345
render EC unable to normally respond to pro-angiogenic 346
signals within the decidualizing microenvironment. In this 347
context, the functions ascribed to NK cells in the remodel- 348
ing of spiral arteries could also be pivotal in the onset of 349
angiogenic responses in the decidua. 350
In several mammalian species, mature uNK cells rep- 351
resent themselves a source of several cytokines and 352
vasoactive mediators which may direct angiogenesis dur- 353
ing implantation and placental development (Leonard 354
et al., 2006; Li et al., 2001). For instance, in vitro and 355
in vivo studies have shown that both human and mouse 356
uNK cells respond to IL-15 by up-regulating the expres- 357
sion of vascular endothelial growth factor (VEGF) isoforms 358
and placenta growth factor (PLGF), two related proteins 359
with potent pro-angiogenic effects (Leonard et al., 2006; 360
Hanna et al., 2003). Interestingly, VEGF-C secretion by 361
human uNK cells has been associated with a self-directed 362
mechanism controlling the cytotoxic activity of these cells 363
(Kalkunte et al., 2009), providing a link between their 364
pro-angiogenic and immunoregulatory functions within 365
the uterine microenvironment. While similar mechanisms 366
have yet to be explored during mouse pregnancy, it is con- 367
ceivable that the high mobility conferred to uNK cells by 368
the unique repertoire of receptors allowing them to recog- 369
nize and respond to trophoblast-derived signals may not 370
only be important for their role as components of innate 371
immunity, but would also give them the ability to guide 372
maternal angiogenesis through VEGF production (Hanna 373
et al., 2003). It is nevertheless worthmentioning thatrather 374
than antigen recognition, the regulation of uNK cell VEGF 375
expression and their interaction with ECs appears to be 376
more dependent on the local cytokine milieu and oxygen 377
8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 6/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
S.M. Blois et al. / Journal of Reproductive Immunology xxx (2010) xxx–xxx 5
Fig. 2. Depletion of pDC neither affected pregnancy outcome nor the number of implantation in mice. (A) Frequency of fetal loss of Balb/c mated C 57BL/6
females at day 13.5 of pregnancy, that were treated with anti-MIPC clone 120G8 antibody (single injection of 150 × g i.p.) on gestation day 3.5, 7.5 and
11.5 or control antibody (C). Percentage is shown as mean standard error of mean (SEM). (B) Total number of implantations per C57BL/6 female mouse
(mean± SEM).
tension within the human decidualizing microenviron-
ment (Kalkunte et al., 2008).
3.2. DC derived signals directly promote angiogenesis in
the decidua
First identified in pathological settings (i.e., tumor
growth and experimental endometriosis) (Curiel et al.,
2004; Fainaru et al., 2008), the production of vasoactive
mediators by DC may also be important for the control of
the normal angiogenic responses that characterize embryo
implantation. Both in mice and humans this notion is high-
lighted by the preferential location of uDC during early
pregnancy, where they are often associated with the decid-
ual vasculature and show the ability to produce VEGF
(Barrientos et al., 2009) and its receptors VEGFR1 and
VEGFR2 (Plaks et al., 2008; Scholz et al., 2008).
The CD11c-DTR transgenic mouse model has provided
strong evidence on the pro-angiogenic functions of DC dur-
ing early pregnancy. The normal expansion of the decidual
vascular zone fails to occur in the absence of DC, as evi-
denced by the decreased expression of the EC markers
PECAM-1 and endoglin observed in DC depleted implan-
tation sites (Krey et al., 2008). Additionally, DC depletion
during implantation interfered with the normal dynam-
ics of vascular remodeling by affecting vessel permeability
and blood flow to the implantation site (Plaks et al., 2008).
However, as a common feature of DC depleted implanta-
tion sites is the reduced accumulation of mature uNK cells
(Karsten et al., 2009; Krey et al., 2008), further studies are
required to dissect the specific contributions of each cell
subsetto thevascular phenotypeassociated with DC deple-
tion.
Screening of DC depleted implantation sites by differ-
ent experimental approaches gave the first insights into
the mechanisms mediating the pro-angiogenic functions
of decidual DC. For instance, the finding that uDC pro-
duce the soluble isoform of VEGFR1 and TGF-1 (Plaks
et al., 2008) together with the decreased expression of
the TGF-1 receptor endoglin on decidual ECs (Krey et al.,
2008) demonstrate the influence of DC on classical pro-
angiogenic pathways during implantation. The expression
of phosphatydilinositol transfer protein beta (PITP-), a 418
cytosolic protein involved in phospholipid trafficking to 419
cell membranes, is also decreased upon DC depletion (Krey 420
et al., 2008) and might represent an indirect mechanism 421
modulating VEGF expression by affecting the PI3K/Akt sig- 422
naling pathway in endometrial cells (Yoshie et al., 2009). 423
The absence of DC also leads to impaired differentiation 424
of trophoblast giant cells, which are known to produce 425
several vasoactive mediators regulating placental angio- 426
genesis (Cross et al., 2002). Thus, we have demonstrated 427
that DC depletion significantly decreases giant cell expres- 428
sion of proliferin (Krey et al., 2008), a placental hormone 429
that promotes EC migration and vascularization (Cross 430
et al., 2002). 431
4. Concluding remarks 432
The evidence presented suggests that uNK cell recruit- 433
ment is one of the many endometrial adaptations 434
induced during normal decidualization to promote vascu- 435
lar changes that assist the growth and differentiation of 436
stromal cells, which gains more importance during pla- 437
cental development as gestation advances. In contrast, DC 438
appear to act in an upstream position within the uterine 439
milieu, determining the outcome of implantation through 440
their direct influence on crucial processes such as stromal 441
cell proliferation and decidual angiogenesis as well as their 442
ability to modulate uNK cell functions. Furthermore, the 443
identification of novel DC associated pro-angiogenic sig- 444
nals may have important implications for the treatment 445
of pregnancy disorders arising from defective angiogenesis 446
and vascular remodeling during early pregnancy. By allow- 447
ing the morphological and molecular characterization of 448
the decidualizing endometrium in vivo in the absence 449
of signals derived from these cell subsets, mouse mod- 450
els have greatly improved our understanding of DC and 451
NK cell functions during early pregnancy. However, the 452
proven influence of DC derived signals in the promotion of 453
uNK cell recruitment and differentiation still precludes us 454
from identifying specific functions played by each subset 455
individually on the regulation of stromal cell differenti- 456
ation and vascular development and remodeling during 457

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 7/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
6 S.M. Blois et al. / Journal of Reproductive Immunology xxx (2010) xxx–xxx
the decidualization process. In this regard, experimental8
approaches combining genetic engineering technologies9
and pharmacological methods allowing the simultaneous0
depletion of DC and NK cells in vivo from the pregnant
uterus would provide further insights on individual and2
cooperative effects mediated by these cells on endome-3
trial stromal cells and the vascular endothelium, greatly4
improving our understanding of the regulatory pathways5
involved in endometrial decidualization.6
Uncited referenceQ17
Sharkey and Smith (2003).8
Acknowledgements9
We apologize to the many authors whose excellent0
papers could not be cited in this review for space limita-
tions. G.B. received a doctoral scholarship from the Charité2
Stiftung. The work discussed in this review was supported3
by Habilitation training grant from the Charité, Sonnenfeld4
Stiftung, Deutsche Forschungsgemeinschaft (BL 1115/1-1)5
and Fritz Thyssen Stiftung (Az. 10.10.2.125) to S.M.B.6
References7
Ain, R., Trinh, M.L., Soares, M.J., 2004. Interleukin-11 signaling is required8
forthe differentiation of naturalkillercellsat thematernal–fetal inter-9
face. Dev. Dyn. 231, 700–708.0
Ashkar, A.A., Black, G.P., Wei, Q., He, H., Liang, L., Head, J.R., et al., 2003.Assessment of requirements for IL-15 and IFN regulatory factors2
in uterine NK cell differentiation and function during pregnancy. J.3
Immunol. 171, 2937–2944.4
Ashkar, A.A., Di Santo, J.P., Croy, B.A., 2000. Interferon gamma contributes5
to initiation of uterine vascular modification, decidual integrity, and6
uterine natural killer cell maturation during normal murine preg-7
nancy. J. Exp. Med. 192, 259–270.8
Asselin-Paturel, C., Brizard, G., Pin, J.J., Briere, F., Trinchieri, G., 2003.9
Mouse strain differences in plasmacytoiddendritic cellfrequency and0
function revealed by a novel monoclonal antibody. J. Immunol. 171,6466–6477.2
Barrientos, G., Tirado-Gonzalez, I., Klapp, B.F.,Karimi, K., Arck, P.C., Garcia,3
M.G.,et al.,2009. Theimpactof dendriticcellson angiogenic responses4
at the fetal–maternal interface. J. Reprod. Immunol. 83, 85–94.5
Blois, S.M.,AlbaSoto,C.D., Tometten, M.,Klapp,B.F., Margni,R.A., Arck, P.C.,6
2004. Lineage, maturity, and phenotype of uterine murine dendritic7
cells throughout gestation indicate a protective role in maintaining8
pregnancy. Biol. Reprod. 70, 1018–1023.9
Blois, S.M., Barrientos, G., Garcia, M.G., Orsal, A.S., Tometten, M., Cordo-0
Russo, R.I.,et al., 2008.Interactionbetweendendritic cells and naturalkiller cells during pregnancy in mice. J. Mol. Med. 86, 837–852.2
Blois, S.M., Kammerer, U., Alba Soto, C., Tometten, M.C., Shaikly, V., Bar-3
rientos, G., et al., 2007. Dendritic cells: key to fetal tolerance? Biol.4
Reprod. 77, 590–598.5
Cross, J.C., Hemberger, M., Lu, Y., Nozaki, T., Whiteley, K., Masutani, M.,6
et al., 2002. Trophoblast functions, angiogenesis and remodeling of 7
the maternal vasculature in the placenta. Mol. Cell. Endocrinol. 187,8
207–212.9
Croy, B.A., He,H., Esadeg, S., Wei, Q.,McCartney,D., Zhang,J., et al., 2003a.0
Uterine natural killer cells: insights into their cellular and molecularbiology from mouse modelling. Reproduction 126, 149–160.2
Croy, B.A., Esadeg, S., Chantakru, S., van den Heuvel, M., Paffaro, V.A., He,3
H., et al.,2003b. Update on pathwaysregulating the activation of uter-4
ine Natural Killer cells. Theirinteractions withdecidual spiral arteries5
and homing of their precursors to the uterus. J. Reprod. Immunol. 59,6
175–191.7
Curiel, T.J., Cheng, P., Mottram,P., Alvarez, X., Moons, L., Evdemon-Hogan,8
M., et al.,2004. Dendriticcell subsets differentiallyregulateangiogen-9
esis in human ovarian cancer. Cancer Res. 64, 5535–5538.0
Das, S.K., Lim, H.,Paria, B.C., Dey, S.K., 1999. CyclinD3 in themouse uterusis associated withthe decidualization process during earlypregnancy.2
J. Mol. Endocrinol. 22, 91–101.
Dey, S.K., Lim, H., Das, S.K., Reese, J., Paria, B.C., Daikoku, T., et al., 2004. 523
Molecular cues to implantation. Endocr. Rev. 25, 341–373. 524
Distler, J.H.,Hirth, A., Kurowska-Stolarska, M., Gay, R.E., Gay,S., Distler, O., 525
2003. Angiogenic and angiostatic factors in the molecular control of 526
angiogenesis. Q. J. Nucl. Med. 47, 149–161. 527
Fainaru, O., Adini, A., Benny, O., Adini, I., Short, S., Bazinet, L., et al., 2008. 528
Dendritic cells support angiogenesis and promote lesion growth in a 529
murine model of endometriosis. FASEB J. 22, 522–529. 530
Germeyer, A., Sharkey, A.M., Prasadajudio, M., Sherwin, R., Moffett, A., 531
Bieback, K.,et al., 2009. Paracrineeffectsof uterine leucocyteson gene 532
expression of human uterine stromal fibroblasts. Mol. Hum. Reprod. 533
15, 39–48. 534
Guimond, M.J., Luross, J.A., Wang, B., Terhorst, C., Danial, S., Croy, B.A., 535
1997. Absence of natural killercells during murinepregnancy is asso- 536
ciated withreproductive compromise in TgE26mice. Biol. Reprod. 56, 537
169–179. 538
Hanna, J., Goldman-Wohl, D., Hamani, Y., Avraham, I., Greenfield, C., 539
Natanson-Yaron, S., et al., 2006. Decidual NK cells regulate keydevel- 540
opmental processesat the human fetal–maternal interface. Nat. Med. 541
12, 1065–1074. 542
Hanna,J., Wald, O., Goldman-Wohl,D., Prus, D.,Markel, G., Gazit,R., et al., 543
2003.CXCL12expressionby invasivetrophoblasts induces thespecific 544
migration of CD16-human natural killer cells. Blood 102, 1569–1577. 545
Hirabayashi, H., Sato, T., Kohno, S., Tanaka, M., Kobayashi, S., Ohta, Y., 546
et al., 1999. Apoptotic cell death in artificially induced deciduoma of 547
pseudopregnant mice. Anat. Rec. 254, 205–213. 548
Jung, S., Unutmaz, D., Wong, P., Sano, G., De los Santos, K., Sparwasser, 549
T., et al., 2002. In vivo depletion of CD11c(+) dendritic cells abro- 550
gatesprimingof CD8(+) T cells by exogenouscell-associated antigens. 551
Immunity 17, 211–220. 552
Kalkunte,S., Chichester, C.O.,Gotsch, F., Sentman,C.L., Romero,R., Sharma, 553
S., 2008. Evolution of non-cytotoxic uterine natural killer cells. Am. J. 554
Reprod. Immunol. 59, 425–432. 555
Kalkunte, S.S., Mselle, T.F., Norris, W.E., Wira, C.R., Sentman, C.L., Sharma, 556
S., 2009. Vascular endothelial growth factor C facilitates immune tol- 557
erance and endovascular activity of human uterine NK cells at the 558
maternal–fetal interface. J. Immunol. 182 (7), 4085–4092. 559
Kammerer, U., 2005. Antigen-presenting cells in the decidua. Chem. 560
Immunol. Allergy 89, 96–104. 561
Kammerer, U., Eggert, A.O., Kapp, M., McLellan, A.D., Geijtenbeek, T.B., 562
Dietl, J., et al., 2003. Unique appearance of proliferating antigen- 563
presenting cells expressing DC-SIGN (CD209) in the decidua of early 564
human pregnancy. Am. J. Pathol. 162, 887–896. 565
Kammerer, U.,Kruse, A.,Barrientos, G., Arck,P.C., Blois, S.M., 2008. Role of 566
dendriticcells in theregulation of maternal immune responsesto the 567
fetus during mammalian gestation. Immunol. Invest. 37, 499–533. 568
Karsten, C.M., Behrends, J., Wagner, A.K., Fuchs, F., Figge, J., Schmudde, I., 569
et al., 2009. DC within the pregnant mouse uterus influence growth 570
and functional properties of uterine NK cells. Eur. J. Immunol. 39, 571
2203–2214. 572
King, A., 2000. Uterine leukocytes and decidualization. Hum. Reprod. 573
Update 6, 28–36. 574
Klauber, N., Rohan, R.M.,Flynn,E., D’Amato,R.J., 1997.Critical components 575
of the female reproductive pathway are suppressed by the angiogen- 576
esis inhibitor AGM-1470. Nat. Med. 3, 443–446. 577
Krey, G., Frank, P., Shaikly, V., Barrientos, G., Cordo-Russo, R., Ringel, F., 578
et al.,2008.In vivodendriticcell depletion reduces breedingefficiency, 579
affecting implantation and early placental development in mice. J. 580
Mol. Med. 86, 999–1011. 581
Leonard,S., Murrant, C.,Tayade,C., vanden Heuvel, M.,Watering, R.,Croy, 582
B.A., 2006. Mechanisms regulating immune cell contributions to spi- 583
ral artery modification—facts and hypotheses—a review. Placenta 27 584
(Suppl. A), S40–S46. 585
Li, F., Devi, Y.S., Bao, L., Mao, J., Gibori, G., 2008. Involvement of cyclin D3, 586
CDKN1A (p21), and BIRC5 (Survivin) in interleukin 11 stimulation of 587
decidualization in mice. Biol. Reprod. 78, 127–133. 588
Li, X.F., Charnock-Jones, D.S., Zhang, E., Hiby, S., Malik, S., Day, K., et al., 589
2001.Angiogenic growthfactormessengerribonucleicacidsin uterine 590
natural killer cells. J. Clin. Endocrinol. Metab. 86, 1823–1834. 591
Ma,G.T., Roth, M.E., Groskopf, J.C., Tsai, F.Y., Orkin, S.H., Grosveld,F., et al., 592
1997. GATA-2 and GATA-3 regulate trophoblast-specific gene expres- 593
sion in vivo. Development 124, 907–914. 594
Paria, B.C., Ma, W., Tan, J., Raja, S., Das, S.K., Dey, S.K., et al., 2001. Cellular 595
andmolecular responsesof theuterus to embryoimplantationcan be 596
elicited by locally applied growth factors. Proc. Natl. Acad. Sci. U.S.A. 597
98, 1047–1052. 598
Plaks, V., Birnberg, T., Berkutzki, T., Sela, S., BenYashar, A., Kalchenko, 599
V., et al., 2008. Uterine DCs are crucial for decidua formation during 600
embryo implantation in mice. J. Clin. Invest. 118, 3954–3965. 601

8/6/2019 Blois Et Al 2011 Proofs
http://slidepdf.com/reader/full/blois-et-al-2011-proofs 8/8
Please cite this article in press as: Blois, S.M., et al., Decidualization and angiogenesis in early pregnancy: unravelling the
functions of DC and NK cells. J. Reprod. Immunol. (2011), doi:10.1016/j.jri.2010.11.002
ARTICLE IN PRESS
U N C O
R R E C
T E D
P R
O O F
GModel
JRI19791–7
S.M. Blois et al. / Journal of Reproductive Immunology xxx (2010) xxx–xxx 7
Rahman, M.A., Li, M., Li, P., Wang, H., Dey, S.K., Das, S.K., 2006. Hoxa-10deficiency alters region-specific gene expression and perturbs differ-entiation of natural killer cells during decidualization. Dev. Biol. 290,105–117.
Robb,L., Li, R., Hartley,L., Nandurkar, H.H., Koentgen, F., Begley, C.G.,1998.Infertilityin femalemicelackingthe receptorfor interleukin11 is dueto a defective uterine responseto implantation. Nat.Med. 4, 303–308.
Schlafke, S., Enders, A.C., 1975. Cellular basis of interaction between tro-phoblast and uterus at implantation. Biol. Reprod. 12, 41–65.
Scholz, C., Toth, B., Santoso, L., Kuhn, C., Franz, M., Mayr, D., et al., 2008.
Distribution and maturity of dendritic cells in diseases of insufficientplacentation. Am. J. Reprod. Immunol. 60, 238–245.
Sharkey, A.M., Smith, S.K., 2003. The endometrium as a cause of implan-tation failure. Best Pract. Res. Clin. Obstet. Gynaecol. 17, 289–307.
Swiecki, M., Colonna, M., 2010. Unraveling the functions of plasmacytoid 615
dendritic cells during viral infections, autoimmunity, and tolerance. 616
Immunol. Rev. 234, 142–162. 617
Tan,J., Raja,S., Davis, M.K., Tawfik, O.,Dey, S.K.,Das, S.K.,2002.Evidencefor 618
coordinatedinteraction of cyclinD3 withp21 andcdk6 indirecting the 619
development of uterine stromal cell decidualization and polyploidy 620
during implantation. Mech. Dev. 111, 99–113. 621
Torry, D.S., Leavenworth, J., Chang, M., Maheshwari, V., Groesch, K., Ball, 622
E.R.,et al.,2007. Angiogenesisin implantation. J. Assist.Reprod. Genet. 623
24, 303–315. 624
Yoshie, M.,Miyajima, E., Kyo,S., Tamura, K., 2009.Stathmin, a microtubule 625
regulatoryprotein, is associatedwith hypoxia-inducible factor-1alpha 626
levels in human endometrialand endothelial cells. Endocrinology 150, 627
2413–2418. 628





























