Embed Size (px)
DESCRIPTION
mo ta lieu phacp chua ung thu
Citation preview

CÁC LOẠI THUỐC MỚI ĐẦY HỨA HẸN CHO VIỆC CHỮA TRỊ CÁC TẾ BÀO UNG THƯ VÀ DI CĂN
Các tính chất độc đáo của các tế bào ung thư và di căn cho phép sự tự đổi mới cao và tiềm năng biến đổi khác nhau (bao gồm mức biểu hiện cao của các yếu tố kháng apoptosis, sự vận chuyển thuốc, và sửa chữa DNA và các enzyme giải độc) có thể được kết hợp với tính kháng của chúng đối với các liệu pháp lâm sàng chữa trị ung thư hiện tại và tái phát bệnh. Việc loại bỏ các tế bào ung thư và di căn bằng LPĐPT góp phần vào sự tăng trưởng của chúng, kháng điều trị, do đó, là mối quan tâm điều trị lâu dài.Những liệu pháp mới này sẽ góp phần cải thiện hiệu quả việc điều trị ung thư, di căn,tái phát và gây tử vong.
GIỚI THIỆU
Tiến bộ lớn đối với việc xác định các mục tiêu điều trị mới trong các tế bào ung thư trong những năm gần đây đã dẫn đến việc khám phá và phát triển của các loại thuốc chống ung thư mới. Những loại thuốc chống ung thư hứa hẹn sẽ cải thiện hiệu quả phương pháp điều trị lâm sàng bằng cách phẫu thuật cắt bỏ, liệu pháp kháng nội tiết tố, xạ trị và / hoặc hóa trị liệu. Đặc biệt, sự phát triển của các tế bào bị ung thư xâm lấn sang các tế bào bình thường hoặc di căn thường liên quan với tính kháng với các phương pháp điều trị thông thường hiện nay, tái phát bệnh, và cái chết của bệnh nhân ung thư trong một thời gian ngắn. Do đó, phân tử đích của các sản phẩm gây ung thư khác nhau trong các tế bào ung thư có liên quan đến sự tiến triển ung thư sơ cấp và di căn sang các mô và các cơ quan khác có thể tạo thành nhiều phương pháp tiếp cận đầy hứa hẹn cho điều trị hơn so với đơn trị liệu cho việc phát triển các phương pháp điều trị kết hợp mới chống lại bệnh ung thư tích cực và tái phát.
Quan trọng hơn, các bằng chứng thực nghiệm đã cho thấy sự tích lũy các biến đổi gen và / hoặc biểu mô trong các mô nơi chứa các tế bào gốc trưởng thành hoặc các thế hệ con cháu của chúng có thể dẫn đến biến đổi thành ác tính từ các tế bào ung thư gốc, còn được gọi là ' tế bào khởi nguyên' hoặc 'các tế bào ung thư khởi xướng'. Dựa vào các tế bào ung thư gốc này người ta có thể biết rõ hơn về chất sinh ung thư, nếu lượng bạch cầu hoặc các tế bào ung thư khởi xướng khối u cao có thể cung cấp các chức năng quan trọng cho sự phát triển các tế bào ung thư sơ cấp bằng cách tạo ra với số lượng lớn của các tế bào khối u biệt hóa. Để hỗ trợ cho giả thuyết này, người ta đã chứng minh rằng việc cảm ứng biến đổi gen hoặc biểu mô trong các tế bào gốc trưởng thành hoặc con cháu của chúng có thể dẫn đến sự biến đổi ác tính, sự phát triển bệnh bạch cầu hoặc khối u trong cơ thể động vật. Những kết cấu di truyền bao gồm các đột biến đặc biệt của gen ức chế khối u (p53, p16 INK4A , retinoblastoma hoặc phosphatase, và sự mất đoạn trên nhiễm sắc thể số 10, hoặc PTEN), kích hoạt các đột biến trong các sản phẩm gây ung thư như RAS, sắp xếp lại nhiễm sắc thể tạo ra các protein dung hợp gây ung thư , hoặc cho phép các tín hiệu dẫn truyền khác nhau hoạt động như hedgehog, thụ thể yếu tố sinh trưởng biểu bì
1

(EGFR), HER2, Wnt / β-catenin, và Notch. Đặc biệt, sự tích lũy các đoạn ngắn telomere và các đột biến, duy trì động lực oxy hóa, viêm mãn tính, và xơ hóa xảy ra trong quá trình lão hóa có thể dẫn đến một sự bất ổn định di truyền và biến đổi ác tính của các tế bào chưa trưởng thành.
Hình 1Đề xuất mô hình của giai đoan đầu ung thư , phát triển, di căn và điều trị tính kháng
qua trung gian các tế bào ung thư và di căn ban đầu. Mô hình này cho thấy sự biến đổi của tế bào gốc trưởng thành thành các tế bào gốc ung thư ác tính, cũng như giúp chúng ta biết được thế nào là “tế bào gốc ung thư khởi xướng”, mà có lẽ chúng được tăng cường thông qua di truyền trong các tế bào trưởng thành này và làm thay đổi vị trí của chúng , bao gồm các tế bào mô đệm đã được hoạt hóa. Việc phân chia đối xứng hoặc
2
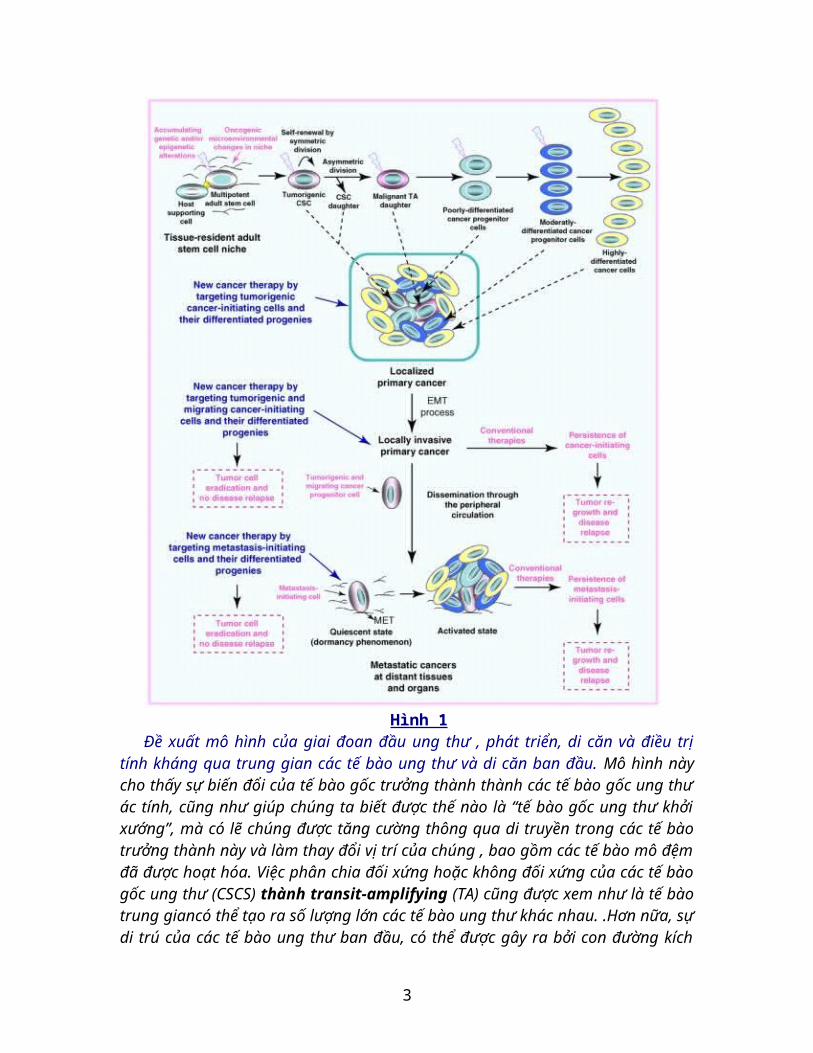
không đối xứng của các tế bào gốc ung thư (CSCS) thành transit-amplifying (TA) cũng được xem như là tế bào trung giancó thể tạo ra số lượng lớn các tế bào ung thư khác nhau. .Hơn nữa, sự di trú của các tế bào ung thư ban đầu, có thể được gây ra bởi con đường kích hoạt các tín hiệu gây ung thư khác nhau trong suốt quá trình chuyển đổi giữa biểu mô-trung mô (EMT). Sự xâm lấn của các tế bào ung thư khởi xướng trong nội mô có thể dẫn đến việc chúng trở nên phổ biến ở phạm vi ngoại mô và đến các mô hay cơ quan xa hơn. Bên cạnh đó, khả năng di trú của các tế bào di căn khởi xướng có thể bị mất thông qua quá trình chuyển đổi biểu mô-trung mô (MET) tại các vùng khối u sơ cấp. Hiện tượng tiềm ẩn của các tế bào di căn và sự kích hoạt chúng gắn liền với sự hình thành khối u thứ cấp dưới các điều kiện đặc biệt tại các vùng xa hơn cũng đã được minh họa. Phương pháp điều trị ung thư mới, trong đó sử dụng phân tử đích của tế bào ung thư và di căn khởi xướng để chống lại sự tiến triển ung thư và di căn ở các mô và các cơ quan xa, cũng được chỉ ra.
Quan trọng hơn, các tiểu quần thể của các tế bào ung thư chưa trưởng thành với đặc tính giống như tế bào gốc đã được phân lập từ các mô ác tính sơ khai của bệnh nhân ung thư và thành lập dòng tế bào ung thư. Các loại ung thư có chứa một quần thể của các tế bào ung thư khởi nguyên, trong đó có ung thư bạch cầu, u lympho, sarcomas, u ác tính, u não, và một loạt các bệnh ung thư biểu mô bao gồm cả đầu, da và cổ, tuyến giáp, ung thư phổi, cổ tử cung, thận, gan, thực quản , tiêu hóa, đại tràng, bàng quang, tuyến tụy, tuyến tiền liệt, vú, và ung thư buồng trứng. Các tế bào ung thư khởi nguyên cho thấy một số tế bào gốc giống như các maker bao gồm telomerase, aldehyde dehydrogenase (ALDH), CD133, CD44, CXC chemokine receptor 4 (CXCR4), KIT, chất vận chuyển ATP-binding cassette (ABC), và các nhân tố phiên mã như Oct-3/4, Nanog, và SOX2. Các tế bào bạch cầu hoặc các tế bào gốc UT có thể gia tăng (in vitro và in vivo) dẫn đến làm gia tăng tổng khối lượng của các tế bào ung thư khác biệt mang đặc điểm hình thái phức tạp và kiểu hình không đồng nhất của các khối u nguyên thủy của bệnh nhân (Hình 1).
Ngoài ra, sự di căn của các tế bào gốc ung thư trong suốt quá trình EMT với những thay đổi trong mô đệm cũng có thể dẫn đến sự xâm lấn của chúng từ khối u ban đầu, phổ biến thông qua tuần hoàn ngoại vi ( Hình 1 ). Kết quả là, các khối u và các tế bào ung thư di căn đã được phát hiện tại các vùng trong các khối u ban đầu, cũng như đã được phân lập từ máu ngoại vi. Bên cạnh đó, các tế bào ung thư chưa trưởng thành với các đặc tính giống tế bào gốc, còn được gọi là “ các tế bào di căn khởi xướng“, đã được phát hiện và phân lập từ các mẫu khối u thứ cấp từ các bệnh nhân ung thư và các dòng tế bào ung thư di căn. Mặc dù các tế bào ung thư và di căn nói chung đại diện cho các quần thể tế bào khối u nhưng quan trọng là cần lưu ý rằng: số lượng, kiểu hình và các chức năng của chúng có thể khác nhau trong quá trình tiến triển ung thư, sau khi điều trị, và sau khi bệnh tái phát. Hơn nữa, số lượng các tế bào ung thư khởi nguyên với các đặc tính giống tế bào gốc có thể phụ thuộc vào các phương pháp được sử dụng để phân lập chúng (in vitro và in vivo). Đặc biệt, một tỷ lệ lớn hơn của các tế bào ung thư khởi nguyên có thể được phát hiện trong tổng khối lượng tế bào ung thư sau khi cảm ứng bởi chương trình
3

EMT, cũng như sau khi điều trị ung thư khởi đầu, có thể dẫn đến một loại bỏ đồng thời hàng loạt với số lượng lớn các tế bào ung thư có đặc tính giống với tế bào gốc có khả năng kháng với các phương pháp điều trị ung thư thông thường. Ngoài ra, các tế bào di căn ban đầu được phát hiện trong các tế bào ung thư di căn có thể trải qua quá trình MET và đòi hỏi các đặc điểm kiểu hình và chức năng đặc biệt tại vi môi của chúng phổ biến ở các vị trí di căn nhất định. Do đó, các tế bào di căn ban đầu có thể cho thấy các kiểu hình và chức năng khác nhau và phản ứng khác nhau với các phương pháp điều trị hiện nay.
Trong quá trình điều trị, người ta cũng đã chứng minh rằng khả năng kháng lại của các tế bào ung thư và di căn ban đầu đối với các liệu pháp lâm sàng hiện nay có thể giúp chúng cố thủ tại khối u sơ cấp hoặc thứ cấp khi bắt đầu điều trị, và theo đó, gây ra tái phát bệnh ung thư. Do đó, các phân tử đích của những tế bào ung thư và di căn chưa trưởng thành cũng phải được xem xét để khắc phục tính kháng điều trị. Về vấn đề này, chúng tôi tóm tắt các nghiên cứu gần đây để xác định các phân tử đích mới trong các tế bào ung thư và di căn ban đầu. Các thông tin được cung cấp sẽ giúp chúng ta trong việc đưa ra một chiến lược điều trị mới cho việc loại bỏ khối lượng tế bào khối u, bao gồm các tế bào ung thư và di căn khởi xướng, và cải thiện các liệu pháp lâm sàng hiện tại chống lại ung thư di căn, tái phát, và gây chết.
Kiểu hình nội tại và kiểu hình thu được của các tế bào ung thư và di căn ban đầu liên quan đến khả năng chống lại các phương pháp điều trị ung thư hiện nay
Nhiều thử nghiệm đã chứng minh được rằng các tế bào ung thư và di căn có thể đề kháng với một số loại thuốc hóa trị liệu hoặc xạ trị vào lúc bắt đầu điều trị và có được một kiểu hình nâng cao khả năng kháng thuốc (MDR) với sự phát triển của bệnh ung thư hoặc sau khi điều trị ban đầu. Một số thuộc tính nội tại của các tế bào ung thư và di căn phổ biến với những bản sao của chúng, các tế bào gốc trưởng thành cư trú trong mô có thể đóng góp cho tính kháng của chúng với cách phương pháp điều trị lâm sàng, và tái phát bệnh hiện nay. Đặc biệt, các tế bào ung thư và di căn chưa trưởng thành, cũng như như các tế bào gốc trưởng thành, có thể tồn tại ở những nơi có hoạt động trao đổi chất yếu. Do đó, chúng có thể phân hóa chậm và mang tính kháng cao hơn các thế hệ sau của chúng để chống lại các tia bức xạ hoặc các loại thuốc gây độc cho tế bào. Thông qua trạng thái hoạt động của các tế bào ung thư và di căn có thể giải thích cho hiện tượng ngủ lâu dài liên quan với sự cư trú của chúng tại các khối ung thư sơ cấp mà không có dấu hiệu lâm sàng hoặc mô bệnh học của di căn rõ ràng. Tuy nhiên, những thay đổi trong vi trường của các tế bào ung thư và di căn khởi xướng dẫn đến việc kích hoạt lại con đường tín hiệu mitogenic có thể gây ra sự gia tăng của chúng và tái phát triển khối u và tái phát bệnh (Hình 1).
Ngoài ra, mức độ biểu hiện cao của các chất vận chuyển thuốc, sửa chữa DNA, và các enzym giải độc trong các tế bào ung thư và di căn có lẽ là đáng tin cậy một phần, cho khả năng chống lại bức xạ ion hóa và các loại thuốc hóa trị liệu nhất định. Cụ thể hơn, các chất vận chuyển ABC đóng ở màng plasmic hoặc endolysosomal có thể bảo vệ các tế bào ung thư chưa trưởng thành khỏi các tác động gây độc tế bào gây ra bởi các loại thuốc
4

hóa trị liệu và theo cách này, góp phần vào kiểu hình MDR của chúng ( Hình 2 ). Trong thực tế, các bơm ABC này có thể chủ động loại bỏ các tác nhân gây độc tế bào ra khỏi tế bào bằng cách sử dụng năng lượng thủy phân ATP, và do đó làm giảm tích lũy thuốc trong tế bào ( Hình 2 ). Về vấn đề này, sự biểu hiện cao của bơm ABC trong tế bào ung thư gốc / tổ tiên là cơ sở của phương pháp loại trừ thuốc nhuộm Hoechst (Hoechst dye), rất hữu ích để cô lập một phần nhỏ tế bào được xem như là “quần thể phụ” (SP), biểu thị khả năng cao để loại bỏ thuốc nhuộm Hoechst 33.342 từ khối tế bào ung thư. Người ta đã quan sát thấy một nhóm nhỏ các tế bào SP phân lập từ các khối u của bệnh nhân hoặc các dòng tế bào ung thư khác nhau cho thấy có các đặc tính giống như tế bào gốc, bơm ABC và các yếu tố chống apoptosis hiện diện ở mức độ cao (in vivo) và khả năng kháng các loại thuốc hóa trị liệu hoặc liệu pháp chiếu xạ cao hơn so với các tế bào không phải là SP. Ví dụ, các tế bào ung thư gan HuH7 SP biểu hiện khả năng kháng thuốc cao (MDR1/ABCB1) mã hóa P-glycoprotein, protein kháng ung thư vú (BCRP-1/ABCG2) và CEACAM6 biểu thị sức đề kháng cao với doxorubicin.
Một quần thể tế bào SP được phát hiện trong các tế bào u nguyên bào thần kinh (neuroblastoma) từ 15 trong số 23 bệnh nhân (65%), trong đó có mức biểu hiện cao của các chất vận chuyển BCRP/ABCG2 và ABCA3. Điều quan trọng là cần lưu ý rằng, giống như các phương pháp làm giàu và cô lập các tiểu quần thể nhỏ của các tế bào ung thư gốc khác - bao gồm cả tế bào hoạt hóa huỳnh quang, việc sử dụng các kháng thể đặc biệt chống lại một hoặc một số maker trên bề mặt tế bào gốc, và phân lập cytometric của các tế bào dựa trên sự hoạt động của enzim ALDH, dựa trên thử nghiệm của Aldehfluor – phương pháp loại bỏ thuốc nhuộm Hoechst có thể có những hạn chế nhất định. Thật vậy, cần phải lưu ý rằng số lượng của các tế bào ung thư được phát hiện trong quần thể tế bào SP so với phần không phải SP có thể bị ảnh hưởng đáng kể bởi các điều kiện thí nghiệm, bao gồm cả nguồn gốc của các mô ác tính của bệnh nhân (không được điều trị, điều trị, và các bệnh nhân tái phát), và điều kiện nuôi cấy được sử dụng. Hơn nữa, đã có những báo cáo rằng thuốc nhuộm Hoechst có thể gây độc cho một số loại tế bào và các quần thể tế bào SP có lẽ không phát hiện được trong một số tế bào lành tính hoặc ác tính.
5

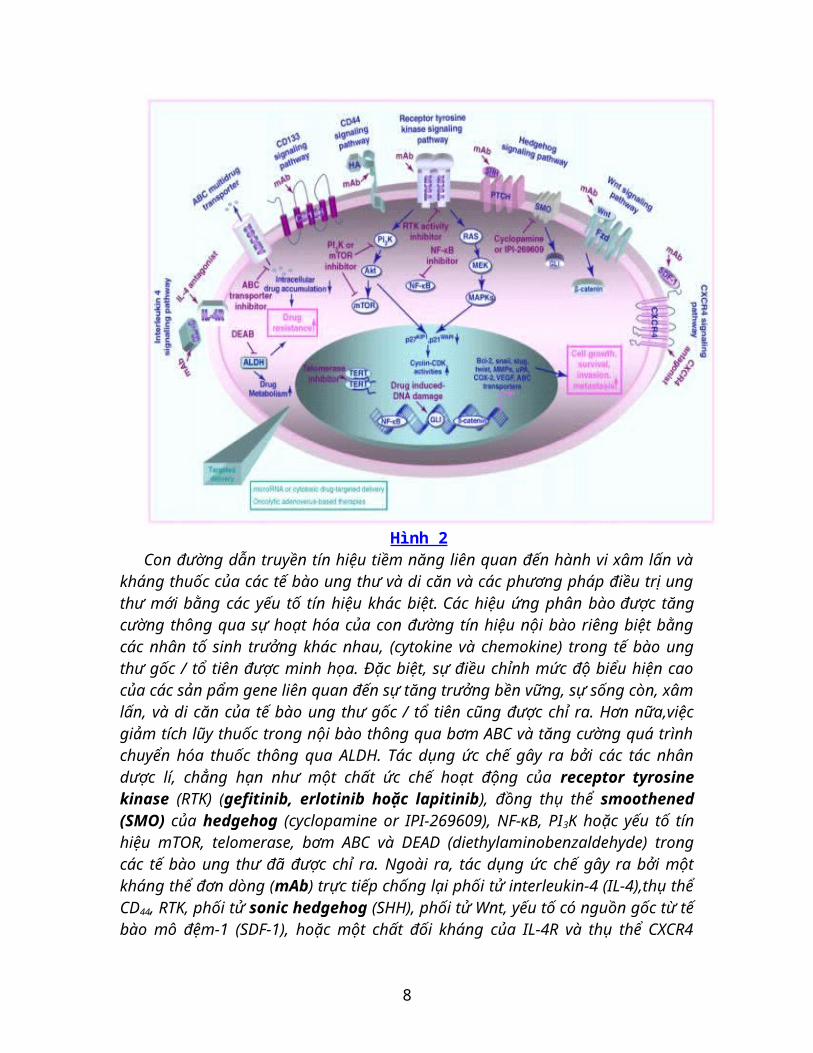
Hình 2Con đường dẫn truyền tín hiệu tiềm năng liên quan đến hành vi xâm lấn và kháng
thuốc của các tế bào ung thư và di căn và các phương pháp điều trị ung thư mới bằng các yếu tố tín hiệu khác biệt. Các hiệu ứng phân bào được tăng cường thông qua sự hoạt hóa của con đường tín hiệu nội bào riêng biệt bằng các nhân tố sinh trưởng khác nhau, (cytokine và chemokine) trong tế bào ung thư gốc / tổ tiên được minh họa. Đặc biệt, sự điều chỉnh mức độ biểu hiện cao của các sản pẩm gene liên quan đến sự tăng trưởng bền vững, sự sống còn, xâm lấn, và di căn của tế bào ung thư gốc / tổ tiên cũng được chỉ ra. Hơn nữa,việc giảm tích lũy thuốc trong nội bào thông qua bơm ABC và tăng cường quá trình chuyển hóa thuốc thông qua ALDH. Tác dụng ức chế gây ra bởi các tác nhân dược lí, chẳng hạn như một chất ức chế hoạt động của receptor tyrosine kinase (RTK) (gefitinib, erlotinib hoặc lapitinib), đồng thụ thể smoothened (SMO) của hedgehog (cyclopamine or IPI-269609), NF-κB, PI3K hoặc yếu tố tín hiệu mTOR, telomerase, bơm ABC và DEAD (diethylaminobenzaldehyde) trong các tế bào ung thư đã được chỉ ra. Ngoài ra, tác dụng ức chế gây ra bởi một kháng thể đơn dòng (mAb) trực tiếp chống lại phối tử interleukin-4 (IL-4),thụ thể CD44, RTK, phối tử sonic hedgehog (SHH), phối tử Wnt, yếu tố có nguồn gốc từ tế bào mô đệm-1 (SDF-1), hoặc một chất đối kháng của IL-4R và thụ thể CXCR4 được chỉ định. Viết tắt: COX-2: cylooxygenase-2, MMPs: matrix metalloproteinase, VEGF: yếu tố tăng trưởng nội mô mạch máu, uPA: urokinase plaminogen activator.
6

Ngoài ra, mức độ biểu hiện cao của O6-methylguanine DNA methyltransferase trong các tế bào ung thư và di căn có thể bảo vệ chúng chống lại các tác dụng gây độc tế bào gây ra bởi các tác nhân methylating và chloroethylating [carmustine, 1,3-bis (2-chloroethyl) -1 - nitrosourea, methyl methanesulfonate và temozolomide] bằng cách sửa chữa nhóm alkyl ở vị trí -O6 của guanine trong DNA. Hơn nữa,các tế bào ung thư gốc / tổ tiên được tìm thấy trong bệnh bạch cầu, võng mạc, phổi, đầu và cổ, gan, tụy, đại trực tràng, ung thư vú, và ung thư tuyến tiền liệt cũng cho thấy mức độ biểu hiện cao của ALDH. ALDHs là một nhóm các enzym có vai trò quan trọng trong quá trình trao đổi chất và giải độc tế bào bằng cách xúc tác quá trình oxy hóa (khử) andehit thành axit cacboxylic. Do đó, ALDHs có thể góp phần vào việc giải độc của một loạt các hợp chất, bằng cách chuyển vị trí nhóm akyl. Về vấn đề này, đã có những báo cáo chắc chắn rằng isoenzymes ALDH (bao gồm cả ALDH1 trong tế bào chất),có mặt trong một số tế bào gốc và thế hệ của chúng, có thể đóng một vai trò rất quan trọng trong sinh tổng hợp acid retinoic (RA) và quy định về số phận của tế bào gốc trong quá trình biệt hóa.
Sinh tổng hợp RA từ vitamin A, còn được gọi là retinol, bao gồm hai bước oxy hóa tuần tự là quá trình oxy hóa của retinol thành retinaldehyde, tiếp theo đó là chuyển đổi retinaldehyde thành RA (xúc tác bởi NADP + ALDHs, bao gồm ALDH1). Mặc dù ALDHs đóng vai trò rất quan trọng trong sinh tổng hợp RA ở một số tế bào gốc nhất định của người, bao gồm các tế bào gốc tạo máu và tế bào gốc thần kinh, một vài nghiên cứu đã tìm hiểu mối quan hệ giữa mức độ biểu hiện ALDHs và sinh tổng hợp RA trong các tế bào ung thư với khả năng khác thường, bao gồm cả tế bào gốc ung thư trong suốt quá trình phát triển của bệnh. Các kết quả từ một nghiên cứu gần đây đã chỉ ra rằng sự ức chế hoạt động enzyme ALDH bằng cách sử dụng một tác nhân ức chế đặc hiệu là diethylaminobenzaldehyde trong các dòng tế bào ung thư vú thúc đẩy hình thành khối u hình cầu. Ngược lại, việc điều trị các tế bào ung thư vú với chất ngoại sinh all-trans RA dẫn đến sự biệt hóa tế bào và ức chế khả năng hình thành khối u hình cầu.
Những nghiên cứu này cho thấy rằng sự kích hoạt tín hiệu RA, có thể được gây ra thông qua isoenzymes ALDH trong các tế bào UT khởi xướng (bao gồm cả tế bào ung thư vú khởi xướng), có thể làm giảm số lượng quần thể tế bào chưa trưởng thành. Ngược lại, sự ức chế ALDH1 trong quần thể tế bào tuyến tụy MiaPaCa-2 bằng các siRNA đã làm giảm khả năng xâm lấn của chúng. Điều này cho thấy khả năng khác của ALDHs, bao gồm ALDH1 là thúc đẩy sự di trú của các tế bào ung thư tuyến tụy. Quan trọng hơn, các khuyết điểm trong con đường tín hiệu RA, chẳng hạn như methyl hóa RA thụ thể β2, và ức chế phản hồi của ALDH gây ra ở nồng độ RA cao trong nội bào có thể xảy ra trong một số dòng tế bào ung thư (bao gồm cả ung thư tế bào vú) và cản trở khả năng biệt hóa của RA. Hơn nữa, chỉ có một số isoenzymes ALDH có thể xúc tác cho sự chuyển đổi của vitamin A thành RA, do đó, mức độ biểu hiện của chúng ở các tế bào UT gốc có thể là một yếu tố quyết định để kiểm soát quá trình sinh ung thư. Trong tương lai, cần có nhiều hơn nữa các nghiên cứu về sự liên quan giữa mức độ biểu hiện của các isoenzyme ALDH khác nhau, sinh tổng hợp RA, và chức năng cụ thể của chúng trong tế bào ung thư
7
khởi xướng so với các tế bào bình thường trong sự phát triển của các loại bệnh ung thư khác nhau, cũng như ý nghĩa của chúng trong việc kháng lại các liệu pháp chữa ung thư hiện tại và tái phát bệnh.
Các đặc điểm kiểu hình khác có thể đóng góp cho sự tồn tại và kháng điều trị của những tế bào chưa trưởng thành bao gồm kích hoạt hoạt động của những con đường tín hiệu đa dạng và mức độ biểu hiện cao của chúng về các yếu tố chống apoptosis như Bcl-2, survivin, NF-κB và quá trình EMT liên quan đến phân tử, bao gồm cả snail, slug, và twist. Những yếu tố tín hiệu có thể các tế bào chưa trưởng thành có lợi thế sống sót trong điều kiện stress và tình trạng thiếu oxy phổ biến trong các khối u sơ cấp. Với triển vọng như thế, chúng tôi giới thiệu LP chữa bệnh mới để khắc phục tính kháng điều trị và diệt trừ số lượng tế bào khối u (bao gồm các tế bào ung thư và di căn khởi xướng).
Liệu pháp nhắm trúng đích phân tử trong điều trị ung thư, những tế bào di căn khởi xướng và quá trình biệt hóa của chúng
Đích phân tử của những chỉ thị sinh học đặc trưng và sự loại bỏ các yếu tố dẫn truyền đã gián tiếp điều chỉnh các sự kiện biến đổi diễn ra ở bệnh UT và những tb di căn khởi xướng trong suốt quá trình diễn biến của bệnh. Và đặc biệt hơn, trong quá trình EMT và sự di căn ở những vị trí xa nhau đã đưa đến một chiến lược chữa trị mới đầy hứa hẹn nhằm cải tiến các liệu pháp chữa UT hiện nay.
Thật thú vị, vài dữ liệu gần đây đã chỉ ra rằng tb gốc như những maker (gồm cả protein CD133 và CD44 ) có thể hình thành được vô số loại tb gốc UT (biểu hiện ác tính) và do đó, có khả năng chữa bệnh UT bằng các ĐPT.
Chẳng hạn, 1 KTĐD (mAb) anti- CD133 (kháng CD133 ) kháng lại epitope 1 (AC133/W6B3C1) hoặc epitope 2 (AC141) cảm ứng đến tác dụng gây độc tố ở những tb khối u FEMX-I (in vitro) và làm giảm khả năng di căn của chúng (trong cơ thể chuột).
(Epitope: nhân tố quyết định kháng nguyên)Trong cùng một cách, việc dùng các phân tử nhỏ oligosaccharide hyaluronan có
thể cạnh tranh hiệu quả với phối tử hyaluronan tự nhiên khi gắn kết với thụ thể CD44, cũng làm ức chế sự hình thành những u thần kinh đệm cột sống sau khi cấy ghép một phần tb SP từ dòng tb u thần kinh đệm C6 (ở chuột).
Hơn nữa, việc xử lí những tb CD90+ với tb gốc và một phần tb CD90− từ dòng tb gan MHCC-97L với một mAb anti- CD44 đã cảm ứng cho quá trình apoptosis (sự chết tb theo lập trình) (in vitro), trái lại, chỉ có phần tb CD90+từ dòng tb PLC là nhạy cảm với cách xử lý này.
Quan trọng hơn, mAb anti-CD44 cũng ngăn chặn sự hình thành khối u cục bộ và toàn cơ thể bởi phần nhỏ tb CD90+ từ dòng tb MHCC-97L dưới da hoặc cấy vào trong gan của chuột.
Mặc dù việc chữa UT bằng LPĐPT có tính khả thi lớn thông qua các tb gốc (biểu hiện cho những tb UT khởi xướng), nhưng khả năng gây độc của những tác nhân này trên các mô bình thường- nơi cư trú của những tb gốc trưởng thành/con cũng cần được xem xét trước khi đưa vào các thử nghiệm lâm sàng.
Về vấn đề này, việc thay đổi mục tiêu của các tác nhân dược lý có thể tạo ra các LP chữa bệnh thay thế để ngăn ngừa việc gây độc tb trên toàn hệ thống.
8

Ngoài ra, trong số các phân tử đích thường được thay đổi trong các tb UT và di căn khởi xướng, có các yếu tố tăng trưởng, yếu tố dẫn truyền tín hiệu (cytokine và chemokine) đóng vai trò quan trọng trong việc điều hòa các qt tự tái tạo, biệt hóa và sự di trú của các tb gốc trưởng thành/nhỏ ở các mô bình thường.
Các yếu tố tăng trưởng, con đường dẫn truyền tín hiệu (cytokine and chemokine) trong LPĐPT
Một vài nghiên cứu chỉ ra rằng việc chuyển biến bệnh UT sang ác tính và những tb di căn có liên quan đến hoạt động của enzyme telomerase, các yếu tố tăng trưởng, cytokine và chemokine thông qua các thụ thể của chúng trong qt diễn biến của bệnh.Các yếu tố này bao gồm: hedgehog, EGFR, HER2, Wnt/β-catenin, Notch, yếu tố tăng trưởng-β (TGF-β)/TGFRβ, interleukin (IL)-4/IL-4Rα, IL-6/IL-6R, IL-8/IL-8 receptor CXCR1, BMI-1, yếu tố tb gốc/KIT, phức hợp chất nền ngoại bào/integrin, và yếu tố có nguồn gốc từ mô đệm tb -1 (SDF-1)/CXCR4.
Đặc biệt, các telomerase đích đã ngăn chặn được các con đường hình thành khối u bằng cách sử dụng mAb, các oligonucleotide đối mã, siRNA, hoặc các tác nhân ức chế đặc hiệu, dẫn đến sự ức chế tăng trưởng, chết tb theo lập trình và làm giảm sự lự lây lan các tb UT (in vitro và vivo)
Ví dụ, việc sử dụng chức ức chế đặc hiệu của smoothened (SMO) co-recepter, cyclopamine hoặc IPI-269609, cũng như các yếu tố phiên mã siRNA GLI-1 của hedgehog cascade, được xem là có khả năng làm giảm một lượng lớn tb UT khởi xướng và mầm mống của chúng, đồng thời ức chế sự tăng trưởng của khối u (in vitro và vivo).
Hơn nữa, việc xử lý cyclopamine đã cải thiện được tác dụng gây độc tố và sự di căn nhờ các loại thuốc hóa trị trên các tb UT u thần kinh đệm, tiền liệt tuyến và tuyến tụy.
Tương tự như thế, chất ức chế đặc hiệu của EGFR tyrosine kinase là gefitinib và erlotinib gây ra tác dụng chống tăng sinh và gây độc tế bào trên các tế bào khối u khởi xướng EGFR + / CD133 + từ 5 bệnh nhân với gliomablastomas (GBM tics). Trong đó có 2 TH với Akt cao là không nhạy cảm với cả 2 loại thuốc, hoặc chỉ nhạy cảm khi nồng độ của erlotinib cao.
Như vậy, việc sử dụng kết hợp chất ức chế của EGFR và Akt có thể có hiệu quả hơn khi chỉ dùng một loại ở bệnh nhân GBM.
Trong một thử nghiệm khác, 40 phụ nữ bị UT vú được điều trị kết hợp giữa erbB2 cùng với oral dual EGFR cùng với sử dụng riêng chất ức chế erbB2 tyrosine kinase laptinib trong 6 tuần, cùng với LP hóa trị chuẩn và mAb trastuzumab anti-erbB2 cũng làm thoái hóa khối u ở 63% bệnh nhân.
Những dữ liệu này chỉ ra rằng việc điều trị chắc hẳn có hiệu quả làm giảm lượng lớn tb UT vú khởi xướng CD44+/CD24low . Phân tử đích của IL-4 chứa cytokine có thể bảo vệ các tb gốc UT biểu bì ruột kết khỏi cái chết lập trình bằng cách sử dụng chất đối kháng (antagonist) IL-4Rα hoặc kháng thể trung hòa (neutralizing antibody) anti-IL-4 cũng nhạy cảm với các tb khối u khởi xướng và tác dụng chống khối u nhờ các loại thuốc hóa trị chuẩn.
Việc sử dụng các tác nhân dược lý có thể can thiệp vào sự lây lan, duy trì việc ht khối u và sự di trú của các tb gốc UT đến bề mặt màng trong và phía trong chất nền. Ngay cả tủy xương (bone marrow) cũng có khả năng điều trị UT.
9

Cụ thể hơn, việc khóa các hệ thống gradient chemoattractant như SDF-1/CXCR4 có thể dẫn đến sự triệt bỏ các khối u, các tb UT khởi xướng di trú và di căn, do đó, ngăn ngừa được việc lan truyền sự di căn, ht khối u và tái phát bệnh.
Cùng với đó, việc xử lý các tb UT tiền liệt tuyến CD133+/CXCR4+ từ khối u ban đầu cùng với mAb anti-CXCR4 cũng triệt bỏ qt di trú nhờ protein ngoại sinh SDF-1 (exogenous protein) (in vitro).
Tương tự, việc xử lý dòng tb UT tuyến tụy di căn ở người L3.6 pl chứa quần thể tb UT CD133 + / CXCR4 +cùng với mAb cũng làm giảm khả năng di căn của chúng (in vivo).Tóm lại, LPĐPT được thực hiện nhằm chống lại sự xâm lấn của các tế bào ung thư khởi xướng cũng như ảm ứng các gen trong quá trình EMT và điều này liên quan đến sự sống còn của các tb UT và kháng điều trị.
Quá trình EMT trong liệu pháp nhắm trúng đích
Trong suốt giai đoạn đầu của bệnh ung thư, từ những tb ung thư khởi xướng và mầm mống của chúng, ngày càng có nhiều kiểu hình ác tính được biểu hiện. Và đặc biệt hơn nữa, quá trình EMT có thể giúp chúng trong việc di cư và lan truyền sang những vị trí xa hơn.
Nói chung, chương trình EMT thường phá vỡ sự tương tác giữa tb và tb với nhau, tổ chức lại các phức hợp ở ngoại bào, và các tb ung thư có lợi thế sống sót cao hơn.
Cụ thể hơn, quá trình EMT có thể dẫn đến ức chế các phân tử bám dính như E-cadherin, cùng với đó là tăng cường biểu hiện các gen trung mô như vimentin, β-catenin, snail, slug, và twist. Điều này đóng vai trò quan trọng trong việc chống lại các liệu pháp trị ung thư và tái phát bệnh.
Một vài nghiên cứu chỉ ra rằng liệu pháp chữa bệnh nhắm trúng đích và những yếu tố tín hiệu khác đã góp phần vào sự sống còn của các tb khởi xướng UT để chống lại các tia bức xạ và kiểu hình MDR, đồng thời hạn chế sự tăng trưởng của khối u và sự di căn.
Trong liệu pháp nhắm đích, các phân tử đích thường thay đổi trong quá trình bệnh diễn ra, đó là các tác nhân thực hiện tín hiệu tế bào của chương trình EMT (Cripto-1, tenacin C, snail, slug, và twist), PI3K/Akt/mTOR, NF-kB, bơm ABC, ALDHs, Bcl-2 và surviving, cũng như các yếu tố tín hiệu bãi bỏ apoptotic như ceramide và caspase.
Chẳng hạn như, trong một chiến lược sàng lọc cao nhằm xác định các tác nhân gây độc tế bào, người ta thấy rằng EMT liên kết chặt chẽ với các phân tử và phát hiện ra một hợp chất salinomycin, đây như là một độc tính chọn lọc cho tb ung thư vú khởi xướng. Điều này cho thấy rằng, salinomycin có thể ức chế được sự ht các hạch di căn và sự tăng trưởng của khối u vú (in vivo); làm giảm một lượng lớn tb UT vú CD44+/CD24low và tăng số lượng các tb biểu mô UT vú.
Ngược lại, một loại thuốc hóa trị liệu thường được dùng trong điều trị UT vú là paclitaxel chỉ có tác dụng gây độc tố trên các tb biểu mô UT vú, còn các tb CD44+/CD24low thì lại tăng lên sau qt điều trị.
Những dữ liệu này nhấn mạnh tầm quan trọng của việc sử dụng các loại thuốc hoặc sự kết hợp của các tác nhân gây độc tế bào nhắm vào các tb ung thư gốc có khả năng sinh
10

khối u cao, phát triển phác đồ trị liệu hiệu quả để điều trị và chẩn đoán bệnh nhân ung thư cũng như tái phát bệnh.
Hơn nữa, dofequidar fumarate một phức hợp quinoline có thể ức chế kênh vận chuyển ABC, làm giảm tác dụng của các loại thuốc hóa trị liệu và làm tăng tính nhạy cảm của các tb SP với các tb gốc được tách ra từ nhiều dòng tb UT khác nhau. Do đó, dofequidar fumarate có thể là một tác nhân chữa bệnh tiềm năng cho việc đảo ngược kiểu hình MDR của các tb UT chưa trưởng thành.
Ngoài ra, hoạt động liên tục của các yếu tố dẫn truyền tín hiệu PI3K/Akt/mTOR gây ra bởi các yếu tố tăng trưởng và sự bất hoạt gen của khối u PTEN thông quá việc xóa bỏ hoặc gây đột biến gen là sự kiện thường xuyên biến đổi trong nhiều loại UT ở người.
Vì vậy, việc sử dụng chất ức chế đặc hiệu của Akt (perifosine) và mTOR (rapamycin hoặc HSP90)- những tác nhân dẫn truyền tín hiệu ngược- cung cấp các chức năng rất quan trọng cho sự phát triển bền vững của tế bào, sự sống còn, và di cư, đây cũng là mối quan tâm lớn trong việc giảm sự phát triển khối u và sự lan rộng của các tế bào UT di căn khởi xướng.
Trong thực tế, việc ức chế hoạt động enzyme Akt kinase bằng perifosine đã làm giảm một lượng lớn ALDH1-biểu hiện các tb UT vú khởi xướng và sự phát triển của khối u sơ cấp và thứ cấp.
Hơn nữa, sự ức chế con đường PI3K/Akt đã cảm ứng cho qt xâm lấn của các khối u não bằng cách cấy trực tiếp tb GBM của một bệnh nhân sang cơ thể chuột suy giảm miễn dịch.
Tuy nhiên, theo một báo cáo khác, hợp chất thiên nhiên từ cây bông cải xanh (broccoli) là sulforaphane gây ra hiệu ứng apoptotic trên các tb UT tuyến tụy khởi xướng thông qua sự ức chế của NF-κB gây ra nhờ con đường chống apoptotic, kết quả là các khối u dần hoại tử.
Thực tế là, kênh vận chuyển thuốc ABC có thể đóng vai trò chính trong kiểu hình MDR của các tb gốc UT, người ta đã nghiên cứu một vài tác nhân dược lý để ức chế hoạt động của bơm này. Người ta theo dõi việc điều trị tb SP từ dòng tb UT biểu bì H357 (ở miệng) với chất ức chế kênh vận chuyển ABC là verapamil…. Mitoxantrone…
Mặc dù đã có những kết quả quan trọng nhưng vẫn cần có thêm nhiều nghiên cứu hơn nữa để chứng minh cho hiệu quả của các loại thuốc và nhờ đó có thể trị tận gốc các tb UT
11

Targeted delivery strategies (các cách phân phối thuốc)
Các tác nhân trị liệu được đưa vào trong tb UT và di căn khởi xướng bằng cách tiếp hợp thuốc đến các khối u-kháng thể đặc hiệu, độc tố của thuốc sẽ được gói trong liposome hoặc những chất mang khác như hạt nano, các tb gốc được chuyển gen…Các phương pháp này có thể tăng cường tác dụng của thuốc, nâng cao tính chọn lọc, và tiếp tục phân phối thuốc vào trong các khối u khác, do đó làm giẩm độc tính hệ thống. Hơn nữa, các phương pháp điều trị dựa trên adenovirus oncolytic cũng được xem là có hiệu quả để loại bỏ ung thư và tế bào di căn.
Thú vị hơn, một vài nghiên cứu gần đây chỉ ra rằng các phân tử RNA mạch đơn nhỏ và không mã hóa, có chiều dài khoảng 20-23 Nu (microRNA hay miRNAs) có vai trò kiểm soát nghiêm ngặt trong việc biểu hiện gen, và chúng có thể góp phần giải thích nguyên nhân gây UT.
miRNA có thể tác động đến vùng 3’không mã hóa của các phân tử RNA đích, và nhờ vậy mà ngăn cản quá trình dịch mã của chúng hay đúng hơn là làm suy thoái quá trình phiên mã các phân tử đích. Do đó, miRNA có thể kiểm soát sự biểu hiện của một loạt các protein có thể là mục tiêu tiềm năng trị liệu để điều chỉnh các sản phẩm gen đa dạng mà chúng góp phần vào hành vi ác tính của các tế bào ung thư khởi xướng.
Sự di chuyển của miRNA-124 hoặc miRNA-137 gây ra sự khác biệt của các tế bào gốc của chuột trưởng thành có nguồn gốc từ oligodendroglioma và tế bào gốc của con người có nguồn gốc từ GBM và kìm hãm sự tăng trưởng của các tế bào GBM.
Quan trọng hơn, việc xử lý các tb UT vú CD44 + / CD24-/low/Lin- phân tách từ một bệnh nhân UT vú bằng LP hóa trị cộng với let-7-lentivirus đã xóa bỏ sự lây lan của chúng cũng như khả năng hình thành khối u (mammosphere) (in vitro và vivo).
Việc tăng cường biểu hiện let-7 kèm theo đó là biểu hiện của H-RAS giảm đi và biến đổi của nhóm AT-hook 2, các phân tử đích có thể làm giảm khả năng tái phát phát bệnh và tăng cường sự biệt hóa của các tb u vú. Bằng cách này, việc cảm ứng biệt hóa các tb UT bằng các tác nhân như RA hoặc các chất tổng hợp tương tự, interferons (IFNs) hoặc chức ức chế enzyme histone deacetylase cũng có thể là một cách điều trị mới đầy hứa hẹn.
Ví dụ như, một nghiên cứu đã báo cáo rằng việc điều trị IFN-α làm giảm đáng kể các phần tế bào SP nhạy cảm với verapamil (verapamil-sensitive SP cell) từ các dòng tế bào ung thư buồng trứng (ovarian cancer).
Đáng chú ý hơn là các LP điều trị ung thư nên được kết hợp với các tác nhân gây độc tế bào nhắm vào các tế bào gốc UT để ngăn ngừa tái phát bệnh.
12

Vi môi trường của các phân tử đích(Molecular targeting of the microenvironment of cancer- and metastasis-initiating
cells and their differentiated progenies)
Vì vi môi trường (microenvironment) chứa các tb gốc UT cũng đóng vai trò tích cực trong việc chuyển sang giai đoạn ác tính từ những khối u đầu tiên và sự di căn sang nhiều vị trí xa nhau nên phân tử đích của các tb mô đệm chủ cũng có khả năng hạn chế sự phát triển của khối u và qt tạo mạch (angiogenic process).
Khi quan sát qt điều trị khối u thần kinh đệm U87 ở chuột với mAb anti-VEGF, bevacizumab, đã làm giảm rõ rệt mật độ vi mạch (microvasculature) và sự tăng trưởng khối u. Tác dụng chống chất sinh UT (anti-carcinogenic) này kèm theo giảm một lượng mạch lien kết với các tb khối u CD133+/nestin+ .
Việc kết hợp giữa chất ức chế chống UT, VEGFR2 antibody DC101 và tác nhân gây độc tb (cyclophosphamide) là có kết quả hơn so với xử lý độc lập trong việc làm giảm số lượng các tb sinh khối u thần kinh mô đệm C6.
Tuy nhiên, một vài nghiên cứu đã chỉ ra rằng sự hồi phục và hợp nhất các tb chưa trưởng thành có nguồn gốc từ BM và các tb gốc trung mô, tb gốc nội mô có thể làm tăng cường phát triển neoplastic và qt neovascularization. Để ủng hộ cho quan điểm này, một nghiên cứu đã tiến hành dùng các tb nội mô có nguồn gốc từ BM …
Ngoài những ảnh hưởng sinh UT gây ra bởi con đường hedgehog ở các tb UT thì các tín hiệu hedgehog có thể góp phần vào việc hình thành nên nhiều bệnh UT biểu bì ở con người bao gồm: UT tuyến tụy, ruột kết, tuyến tiền liệt, vú và buồng trứng.
Đặc biệt hơn, việc khóa các tín hiệu hedgehog bằng chất ức chế SMO IPI-926 đã gây tác dụng chống tạo mạch và cải thiện tác dụng chống khối u của gemcitabine bằng cách phá vỡ desmoplastic stroma trên tb UT tuyến tụy (in vivo).
Hơn nữa, theo một nghiên cứu gần đây chỉ ra rằng việc ức chế tín hiệu hedgehog bằng cyclopamine trong UT tụy cũng gây ức chế qt sinh khối u.
Concluding remarks (Kết luận)
Những nghiên cứu gần đây về các loại thuốc chữa bệnh UT bằng LP đích phân tử tác động lên các tb UT chưa trưởng thành đóng vai trò rất quan trọng trong qt điều trị của bệnh nhân UT. Đặc biệt hơn, để cải tiến các phương pháp trị bệnh UT hiện nay thì việc kết hợp giữa các tác nhân trị liệu là rất quan trọng nhằm chống lại sự di căn và tái phát bệnh sau qt điều trị.
Tuy nhiên, cần có thêm nhiều cuộc điều tra, nghiên cứu hơn nữa để tìm ra chính xác các chỉ thị sinh học đặc biệt, các sản phẩm gen thay thế trong các tb UT và di căn khởi
13

xướng; và những thay đổi xảy ra trong quá trình bệnh khởi phát và di căn tại vi môi trường của chúng. Điều quan trọng hơn nữa là xác định các đặc điểm về kiểu hình và chức năng của các tb có nguồn gốc từ nhiều loại UT khác nhau cũng như xóa bỏ các cơ chế liên quan đến biểu hiện ác tính của chúng.
Ngoài ra, nên tiến hành phân tích, so sánh biểu hiện gen của các tb UT khởi xướng và qt EMT chống lại các mô bình thường, cần làm sáng tỏ những sự kiện biến đổi phân tử xảy ra trong các các tế bào ác tính và hậu quả bệnh lý của chúng. Hơn nữa, việc thành lập các cơ chế phân tử liên quan với sự di căn cần phải được xác định trước; hiện tượng ngủ và sự kích hoạt các tế bào di căn tại các địa điểm xa nhau sau một thời gian dài cũng là mối quan tâm lớn.
Những nghiên cứu này sẽ dẫn đến việc tìm ra các loại thuốc mới và các chiến lược điều trị để khắc phục tính kháng điều trị và ngăn ngừa tái phát triển khối u, di căn, và tái phát bệnh mà không có tác dụng gây độc tế bào trên các tế bào bình thường, bao gồm cả gốc trưởng thành cư trú ở trong mô. LPĐPT có thể kết hợp với nhiều LP khác nhau gây ra tác dụng tốt hơn cho việc điều trị và thậm chí chữa lành cho những bệnh nhân được chẩn đoán bị ung thư di căn, tái phát, và chết trước đó.
14





























