Embed Size (px)
Citation preview

BỘ NÔNG NGHIỆP VÀ PHÁT TRIỂN NÔNG THÔN
VIỆN DI TRUYỀN NÔNG NGHIỆP
BÁO CÁO TỔNG HỢP
KẾT QUẢ KHẢO NGHIỆM ĐÁNH GIÁ RỦI RO NGÔ
BIẾN ĐỔI GEN (EVENT TC1507) ĐỐI VỚI ĐA DẠNG
SINH HỌC VÀ MÔI TRƯỜNG SINH THÁI VIỆT NAM
Đơn vị đăng kí: Công ty TNHH Pioneer Hi-Bred Việt Nam
Đơn vị khảo nghiệm: Viện Di truyền Nông nghiệp
Hà nội, tháng 5 năm 2012
Science with Service Delivering Success™

BỘ NÔNG NGHIỆP VÀ PHÁT TRIỂN NÔNG THÔN
VIỆN DI TRUYỀN NÔNG NGHIỆP
BÁO CÁO TỔNG HỢP
KẾT QUẢ KHẢO NGHIỆM ĐÁNH GIÁ RỦI RO NGÔ
BIẾN ĐỔI GEN (EVENT TC1507) ĐỐI VỚI ĐA DẠNG
SINH HỌC VÀ MÔI TRƯỜNG SINH THÁI VIỆT NAM
(Theo Nghị định số 69/2010/NĐ-CP ngày 21 tháng 6 năm 2010 của Chính phủ)
Hà nội, tháng 5 năm 2012

MỤC LỤC
1 THÔNG TIN CHUNG ........................................................................................... 11.1 Đơn vị đăng ký khảo nghiệm ........................................................................... 11.2 Đơn vị thực hiện khảo nghiệm ......................................................................... 11.3 Tên giống cây trồng biến đổi gen khảo nghiệm ............................................... 21.4 Các văn bản pháp lý liên quan .......................................................................... 21.5 Các văn bản kèm theo báo cáo ......................................................................... 3
2 TỔNG QUAN VỀ NGÔ BIẾN ĐỔI GEN EVENT TC1507 ................................ 4
2.1 Thông tin chung về ngô event TC1507 ............................................................ 42.2 Thông tin về sinh vật cho gen .......................................................................... 52.3 Thông tin về sinh vật nhận ............................................................................... 62.4 Phương pháp chuyển nạp gen tạo dòng ngô Event TC1507 ............................ 7
2.4.1 Kích thước, trình tự, chức năng của đoạn gene đưa vào ............................. 82.4.2 Phương pháp xác định, phát hiện gen, đặc trưng của gen ........................... 9
2.5 Đặc tính và hiện trạng sử dụng ngô event TC1507 ........................................ 112.5.1 Protein CRY1F kháng côn trùng, sâu hại .................................................. 112.5.2 Protein PAT kháng glufosinate-ammonium trong đánh dấu chọn lọc ...... 152.5.3 Thông tin liên quan đến biểu hiện tính trạng của gen chuyển nạp vào ngô
chuyển gen TC1507 ................................................................................... 152.5.4 Thông tin khác biệt của dòng ngô chuyển gene TC1507 so với cây bố mẹ
…………………………………………………………………………..162.5.5 Phương pháp phát hiện cây ngô chuyển gene TC1507 ............................. 172.5.6 Thông tin về việc thương mại hoá (phóng thích và sử dụng) ngô chuyển
gene TC1507 trên thế giới. ........................................................................ 172.5.7 Mô tả nguy cơ, khả năng xảy ra nguy cơ để xác định rủi ro có thể xảy ra
khi phóng thích sinh vật chuyển gene ....................................................... 19 3 Xác ĐỊNH VẤN ĐỀ CẦN KHẢO NGHIỆM TẠI VIỆT NAM ......................... 27
3.1 Thông tin về sản xuất và sử dụng cây ngô ..................................................... 273.2 Tình hình sử dụng cây trồng BĐG tại Đông Nam Á. .................................... 283.3 Cơ sở khoa học nghiên cứu tính an toàn đối với đa dạng sinh học và môi
trường trong trồng ngô TC1507 trên thế giới ................................................ 293.3.1 Nguy cơ trôi gen ngoài môi trường ........................................................... 293.3.2 Nguy cơ trở thành cỏ dại, dich hại ............................................................ 303.3.3 Nguy cơ ảnh hưởng tới sinh vật không chủ đích ....................................... 30
i

3.4 Tiêu chí đánh giá trong khảo nghiệm đồng ruộng tại Việt Nam đối với ngô biến đổi gen event TC1507 ........................................................................... 34
4 MỤC TIÊU, NỘI DUNG, VẬT LIỆU, PHƯƠNG PHÁP KN ............................ 36
4.1 Mục tiêu và nội dung khảo nghiệm ................................................................ 364.1.1 Mục tiêu ..................................................................................................... 364.1.2 Nội dung khảo nghiệm .............................................................................. 37
4.2 Vật liệu và phương pháp nghiên cứu trong KNHC ........................................ 384.2.1 Giống Ngô đăng ký KNHC ....................................................................... 384.2.2 Thời gian, địa điểm KNHC ....................................................................... 384.2.3 Phương pháp KNHC (áp dụng cho cả hai vụ) ........................................... 394.2.4 Ghi nhận thông tin, phân tích và xử lý số liệu ........................................... 49
4.3 Vật liệu, phương pháp KNDR ........................................................................ 514.3.1 Giống đăng ký KNDR ............................................................................... 514.3.2 Thời gian và địa điểm KNDR .................................................................... 524.3.3 Phương pháp KNDR (áp dụng chung cho cả 4 địa điểm) ......................... 534.3.4 Phương pháp phân tích và xử lý số liệu .................................................... 604.3.5 Phương pháp thực hiện chung cho tất cả các nội dung trong KNDR ....... 60
5 KẾT QUẢ KHẢO NGHIỆM NGÔ CHUYỂN GEN EVENT TC1507 ĐỐI VỚI
ĐA DẠNG SINH HỌC VÀ MÔI TRƯỜNG SINH THÁI VIỆT NAM ............. 625.1 Kết quả KNHC ngô event TC1507 (2 vụ lên tiếp) ......................................... 62
5.1.1 Kết quả kiểm tra sự hiện diện của protein Cry1F (giai đoạn V4) ............. 625.1.2 Kết quả đa dạng quần thể NTOs trên ruộng KNHC .................................. 635.1.3 Thành phần các loài động vật chân khớp (côn trùng và nhện) .................. 665.1.4 Kết quả đánh giá sự đa dạng của sinh vật đất ............................................ 775.1.5 Đánh giá sự xuất hiện và gây hại bệnh hại ngô chính trong KNHC ......... 935.1.6 Kết quả đánh giá mức độ gây hại của sâu hại ngô không chủ đích ........... 985.1.7 Kết luận sơ bộ kết quả KNHC (2 vụ lên tiếp) ......................................... 100
5.2 Kết quả KNDR ngô BĐG event TC1507 ..................................................... 1015.2.1 Kiểm tra sự hiện diện của protein Cry1F và protein CP4 EPSPS ........... 1015.2.2 Kết quả đánh giá tính ổn định và tính thích ứng của ngô event TC1507 tại
các vùng sinh thái Việt Nam. .................................................................. 1035.2.3 Kết quả đánh giá đa dạng quần thể côn trùng không chủ đích trên ruộng
KNDR ngô chuyển gene TC1507 tại các vùng sinh thái Việt Nam ........ 1165.2.4 Kết luận sơ bộ kết quả KNDR ................................................................. 173
ii

6 DỮ LIỆU KHÍ HẬU CHÍNH TẠI CÁC ĐỊA ĐIỂM VÀ BIỆN PHÁP QUẢN LÍ KHẢO NGHIỆM ............................................................................................... 175
6.1 Dữ liệu thời tiết khí hậu trong thời gian khảo nghiệm ................................. 1756.1.1 Dữ liệu thời tiết khí hậu trong thời gian KNHC(tại Văn Giang) ............. 1756.1.2 Dữ liệu thời tiết khí hậu trong thời gian KNDR (tại 4 địa điểm) ............ 177
6.2 Quản lý khảo nghiệm (ảnh minh họa trong Phụ lục 3) ................................ 1806.2.1 Quản lý khảo nghiệm trong KNHC ......................................................... 180
6.3 Quản lý rủi ro trong KNDR .......................................................................... 1826.3.1 Vật liệu hạt giống chuyển gene sử dụng trong khảo nghiệm .................. 1826.3.2 Quản lý cách ly ........................................................................................ 1826.3.3 Thu hoạch và quản lý đồng ruộng sau thu hoạch .................................... 1836.3.4 Quản lý và xử lý đối với các trường hợp phát tán ngẫu nhiên ................ 1836.3.5 Quản lý, ghi chép hồ sơ tư liệu ................................................................ 1846.3.6 Tiêu hủy sản phẩm và vật liệu di truyền biến đổi gen ............................. 185
7 KẾT LUẬN VÀ ĐỀ NGHỊ ................................................................................ 193
7.1 Kết luận ........................................................................................................ 1937.2 Đề nghị ......................................................................................................... 195
iii

MỤC LỤC BẢNG
Bảng 2-1: Các yếu tố di truyền có trong plasmid PHP8999 ....................................... 11Bảng 2-2: So sánh hiệu quả của ngô TC1507 và ngô không chuyển gen ................... 14Bảng 2-3: Mức độ biểu hiện protein Cry1F đo được trong các mô ngô TC1507 ...... 16Bảng 2-4: Danh sách các nước cho phép trồng và/hoặc sử dụng ngô TC1507 ......... 18Bảng 3-1: Nghiên cứu phòng TN, nhà lưới, đồng ruộng nhằm tìm hiểu ảnh hưởng của
protein Cry1F đối với NTOs ........................................................................ 33Bảng 4-1: Thời gian tiến hành KNHC ........................................................................ 38Bảng 4-2:Thời gian điều tra thành phần quần thể NTO tại ruộng KNHC .................. 42Bảng 4-3: Thời gian thu mẫu Collembola tại ruộng KNHC ....................................... 46Bảng 4-4: Tên giống và các công thức trong KNDR .................................................. 51Bảng 4-5: Thời gian gieo và thu hoạch tại mỗi địa điểm KNDR ................................ 52Bảng 4-6: Chỉ tiêu đánh giá đặc tính nông sinh học ................................................... 54Bảng 4-7: Thang điểm đánh giá mức độ gây hại của một số loại sâu hại chính ......... 56Bảng 4-8: Các loài động vật chân khớp được theo dõi trong nghiên cứu NTOs ........ 58Bảng 4-9: Các đợt thu mẫu côn trùng ở các GĐST của cây ngô tại 4 KNDR ............ 59Bảng 4-10: Các đợt thu mẫu Collembola ở các GĐST của ngô tại 4 điểm KNDR .... 60Bảng 4-1: Thời gian gieo và thu hoạch ....................................................................... 61Bảng 5-1: Tổng số cá thể, số loài chân khớp bắt gặp trên các giống ngô trong KNHC
...................................................................................................................... 64Bảng 5-2: Chỉ số ưu thế và chỉ số đa dạng loài chân khớp trên các giống ngô trong
KNHC ........................................................................................................... 65Bảng 5-3: Số loài chân khớp ghi nhận được qua điều tra trực tiếp trên ngô KNHC .. 61Bảng 5-4: Tần suất bắt gặp các loài chân khớp qua điều tra trực tiếp trên ngô KNHC
...................................................................................................................... 62Bảng 5-5: Số loài chân khớp vào bẫy dính vàng trên các giống ngô KNHC ............. 66Bảng 5-6:
Thành phần loài chân khớp vào bẫy dính vàng trên các giống ngô KNHC
...................................................................................................................... 67Bảng 5-7: Tần suất bắt gặp các loài chân khớp qua điều tra trực tiếp và bẫy dính vàng
trên ruộng ngô trong KNHC (Văn Giang, 2010 - 2011) .............................. 71Bảng 5-8: Mật độ bọ rùa đỏ Nhật Bản trên các giống ngô (con/cây) trong KNHC .. 74Bảng 5-9: Mật độ bọ xít mù xanh trên các giống ngô (con/cây) trong KNHC ........... 75Bảng 5-10: Mật độ nhện lớn trên các giống ngô (con/cây) trong KNHC ................... 76Bảng 5-11: Mật độ Bọ cứng cánh ngắn trên các giống ngô (con/cây) trong KNHC .. 76Bảng 5-12: Thành phần loài bọ đuôi bật (Collembola) trong đất trồng ngô KNHC .. 79
iv

Bảng 5-13: So sánh một số chỉ số định lượng của bọ đuôi bật trong đất trồng ngô KNHC ........................................................................................................... 81
Bảng 5-14: Đa dạng thành phần loài và phân bố của bọ đuôi bật (Collembola) trong các lô thí nghiệm thu bằng phương pháp pitfall trap trong KNHC .............. 84
Bảng 5-15: So sánh số lượng loài, số cá thể của bọ đuôi bật (Collembola) trên các giống ngô khảo nghiệm theo từng đợt thu mẫu trong KNHC ...................... 87
Bảng 5-16: Giá trị chỉ số đa dạng H’ và chỉ số đồng đều (J’) của bọ đuôi bật (Collembola) trên các giống ngô KNHC ...................................................... 89
Bảng 5-17: Các loài bọ đuôi bật (Collembola) phổ biến (%) trong đất trồng ngô KNHC ........................................................................................................... 91
Bảng 5-18: Các loài bọ đuôi bật (Collembola) ưu thế (%) trong đất ngô KNHC ....... 92Bảng 5-19: Mức độ nhiễm bệnh đốm lá lớn của các giống ngô KNHC ..................... 93Bảng 5-20: Mức độ nhiễm bệnh đốm lá nhỏ trên của các giống ngô KNHC ............. 94Bảng 5-21: Mức độ nhiễm bệnh rỉ sắt của các giống ngô KNHC .............................. 95Bảng 5-22: Mức độ bị bệnh khô vằn của các giồng ngô KNHC ................................ 96Bảng 5-23: Mức độ nhiễm bệnh đốm nâu của các giống ngô KNHC ....................... 97Bảng 5-24: Cấp nhiễm trung bình của rệp muội ngô trên các giống ngô KNHC ....... 98Bảng 5-25: Cấp nhiễm trung bình của nhện đỏ trên các giống ngô KNHC ............... 99Bảng 5-26: Cấp nhiễm trung bình của sâu cắn lá ngô trên các giống ngô KNHC ... 100Bảng 5-27: Tỷ lệ mọc và sức sống cây con của các giống ngô KNDR .................... 106Bảng 5-28: Một số đặc điểm hình thái của các giống ngô KNDR ............................ 107Bảng 5-29: Trạng thái cây và tỷ lệ đổ gẫy của các giống ngô KNDR ...................... 108Bảng 5-30: Năng suất và các yếu tố cấu thành năng suất các giống ngô KNDR ..... 109Bảng 5-31: Mức độ nhiễm bệnh Đốm lá lớn của các giống ngô tại 4 điểm KN ....... 111Bảng 5-32: Mức độ nhiễm bệnh Khô vằn trên các giống ngô tại 4 điểm KN .......... 112Bảng 5-33: Mức độ bị hại bởi một số loại sâu trên các giống ngô tại 4 điểm KN ... 115Bảng 5-34: Mức độ bị hại bởi sâu xám, rệp muội ngô của các giống ngô tại 4 điểm
KN .............................................................................................................. 116Bảng 5-35: Số lượng loài chân khớp phân theo bộ ghi nhận được trên ruộng ngô
KNDR tại 4 địa điểm ................................................................................. 118Bảng 5-36: Thành phần các loài chân khớp bắt gặp trên ruộng ngô KNDR tại 4 địa
điểm ............................................................................................................ 120Bảng 5-37: Thành phần, mức độ hiện diện các loài chân khớp điều tra bằng điều tra
trực tiếp trên các giống ngô KNDR tại 4 địa điểm(vụ Hè – Thu, 2011) .... 128Bảng 5-38: Thành phần, mức độ hiện diện loài chân khớp điều tra bằng bẫy dính
vàng trên các giống ngô KNDR tại 4 địa điểm (vụ Hè - Thu, 2011) .......... 136
v

Bảng 5-39: Thành phần, mức độ hiện diện các loài chân khớp trong điều tra trực tiếp và bẫy dính vàng trên các giống ngô KNDR tại 4 địa điểm (vụ Hè - Thu, 2011) ........................................................................................................... 143
Bảng 5-40: Chỉ số đa dạng và chỉ số ưu thế các loài chân khớp bắt gặp trên các giống ngô KNDR tại 4 địa điểm (vụ Hè -Thu, 2011) ........................................... 150
Bảng 5-41: Chỉ số đa dạng loài sâu hại bắt gặp trên các giống ngô KNDR tại 4 địa điểm ........................................................................................................... 152
Bảng 5-42: Chỉ số đa dạng các loài bọ rùa bắt mồi trên ngô KNDR tại 4 địa điểm 153Bảng 5-43: Thành phần loài và phân bố Collembola trong đất ngô KNDR tại 4 địa
điểm ............................................................................................................ 160Bảng 5-44: Số lượng loài, số cá thể Collembola trên các giống ngô KNDR theo từng
đợt thu mẫu tại 4 địa điểm .......................................................................... 170Bảng 5-45: So sánh một số chỉ số định lượng Collembola trong đất trồng ngô
KNDR tại 4 địa điểm (vụ Hè – Thu, 2011) ................................................ 171Bảng 5-46: Giá trị chỉ số đa dạng H’ và chỉ số đồng đều J’ theo từng đợt điều tra trên
đất trồng ngô KNDR tại 4 địa điểm (vụ Hè – Thu, 2011) .......................... 172Bảng 5-47: Tên các loài Collembola phổ biến và ưu thế trong đất trồng ngô KNDR
tại 4 địa điểm (vụ Hè –Thu, 2011) .............................................................. 175Bảng 5-48: Các loài Collembola phổ biến, ưu thế trong đất ngô KNDR tại 4 địa điểm
.................................................................................................................... 173Bảng 6-1: Nhiệt độ, ẩm độ và cường độ ánh sáng trung bình trong các tháng KNHC
ngô tại Văn Giang (2010 – 2011) ............................................................... 175Bảng 6-2: Nhiệt độ trung bình trong các tháng trong KNDR tại 4 địa điểm (oC) .... 177Bảng 6-3: Ẩm độ trung bình trong các tháng khảo nghiệm tại 4 địa điểm (%) ........ 177Bảng 6-4: Cây trồng vụ trước trên nền đất KNDR ................................................... 179Bảng 6-5: Phương pháp tiêu hủy vật liệu trong khu vực KNDR tại 4 địa điểm KN 186Bảng 6-6: Lịch trình các đoàn thuộc cơ quan quản lý tham gia kiểm tra giám sát .. 191
vi

MỤC LỤC HÌNH
Hình 2-1: Sơ đồ các yếu tố di truyền đoạn DNA thẳng Pmei PHP8999A được chuyển nạp trong qui trình tạo dòng ngô Event TC1507. ......................................... 10
Hình 2-2: Plasmid PHP8999 ....................................................................................... 10Hình 2-3: Mô hình hoạt động của protein Cry gây độc đối với côn trùng .................. 13Hình 2-4: Tình hình sử dụng ngô TC1507 trên thế giới ............................................. 19Hình 3-1: Diện tích trồng ngô tại các khu vực trồng ngô chính của Việt Nam .......... 27Hình 4-1: Sơ đồ bố trí KNHC vụ 1 ............................................................................. 40Hình 4-2:Sơ đồ bố trí KNHC vụ 2 .............................................................................. 40Hình 4-3: QuickStix™ Kit dùng phát hiện nhanh protein Cry1F ............................... 41Hình 4-4: Điểm lấy mẫu trong đánh giá bằng mắt thường ......................................... 43Hình 4-5: Chỉ số bệnh hại trên lá ................................................................................ 49Hình 4-6: Sơ đồ KNDR ............................................................................................... 53Hình 4-7: Các dụng cụ kiểm tra sự hiện diện của protein Cry1F và CP4 EPSPS ...... 53Hình 4-8: Bẫy dính vàng đặt trong ruộng ngô ............................................................ 59Hình 5-1: Kết quả kiểm tra sự hiện diện của protein Cry1F trên các giống ngô ........ 62Hình 5-2: Trung bình tổng số loài bắt gặp trong một kỳ điều tra trên ngô KNHC ..... 64Hình 5-3: Trung bình số loài chân khớp bắt gặp trong một kỳ điều tra trên ngô
KNHC. .......................................................................................................... 64Hình 5-4: Giá trị của chỉ số ưu thế Simpson's D trên các giống ngô KNHC ............. 66Hình 5-5: Chỉ số đa dạng Shannon H' của các giống ngô KNHC .............................. 66Hình 5-6: Kiểm tra protein Cry1F trên các giống ngô trong KNDR ........................ 102Hình 6-1: Độ ẩm, Nhiệt độ và Cường độ ánh sáng trung bình trong các tháng khảo
nghiệm ngô tại Văn Giang .......................................................................... 176Hình 6-2: Nhiệt độ trung bình tại 4 địa điểm KNDR ............................................... 178Hình 6-3: Ẩm độ trung bình trong các tháng tai 4 địa điểm KNDR ......................... 178
vii

Chữ viết tắt sử dụng trong báo cáo
BĐG Biến đổi gen
CNSH Công nghệ Sinh học
Đ/C Đối chứng
ĐT Điều tra
GCT Giống cây trồng
GĐST Giai đoạn sinh trưởng
HTX Hợp tác xã
KN Khảo nghiệm
KNDR Khảo nghiệm diện rộng
KNHC Khảo nghiệm hạn chế
KT Kiểm tra
N/C Nghiên cứu
NTO Sinh vật không chủ đích
SX Sản xuất
TG Thời gian
TT Trung tâm
VST Vùng sinh thái
viii

1
1 THÔNG TIN CHUNG
1.1 Đơn vị đăng ký khảo nghiệm
Công ty TNHH Pioneer Hi-Bred Việt Nam Đại diện: Ông Nguyễn Đức Mẫn, Tổng Giám đốc Người và địa chỉ liên lạc: Phạm Chí Hòa, phụ trách CNSH Địa chỉ: Lầu 11, cao ốc Center Plaza, 17 Lê Duẩn, Quận 1, TP. Hồ Chí Minh. Điện thoại: (+84-8) 3 8251610 Ext. 8005 Fax: (+84-8) 3 8251620 Email: [email protected] Website: www.pioneer.com
1.2 Đơn vị thực hiện khảo nghiệm
Viện Di truyền Nông nghiệp, viện Khoa học Nông nghiệp Việt Nam Đại diện: PGS.TS. Lê Huy Hàm, viện Trưởng
Địa chỉ: Đường Phạm Văn Đồng, Từ Liêm, Hà Nội, Việt Nam Điện thoại: (+84-4) 8386734; Fax: (+84-4) 7543196 E-mail: [email protected] Website: http://www.agi.gov.vn
Phụ trách chính: TS. Phạm Thị Liên -Viện Di truyền Nông nghiệp Các đơn vị tham gia khảo nghiệm:
- Viện Bảo vệ Thực vật - Viện Sinh Thái và Tài nguyên Sinh vật - Trạm thực nghiệm Văn Giang, huyện Văn Giang, tỉnh Hưng Yên (viện
Di truyền Nông nghiệp) - Trại SX Giống cây trồng Vũ Di - TT Giống cây trồng Vĩnh Phúc. - HTX Phong Thịnh, huyện Thanh Chương, tỉnh Nghệ An - Viện Khoa học Kỹ thuật Nông lâm nghiệp Tây Nguyên - TT Nghiên cứu thực nghiệm Hưng Lộc (viện Khoa học Nông nghiệp
Miền Nam).
Các thành viên tham gia khảo nghiệm: danh sách cán bộ tham gia khảo nghiệm được liệt kê tại Phụ lục 2.

2
1.3 Tên giống cây trồng biến đổi gen khảo nghiệm
- Tên thông thường (cây chủ): Ngô, bắp, bẹ - Tên khoa học: Zea mays L. (Maize)
+ Họ: Gramineae + Chi: Zea + Loài: mays (2n=20)
- Tên sự kiện: Event TC1507 (ngô mang gen cry1F, kháng côn trùng bộ Cánh vảy)
- Tên giống nền (Đ/C): giống ngô lai 30Y87 - Tên giống Đ/C tham khảo: Stack TC1507xNK603 (ngô mang gen cry1F kháng
côn trùng bộ cánh vảy và gen cp4 epsps kháng thuốc trừ cỏ glyphosate)
1.4 Các văn bản pháp lý liên quan
Khảo nghiệm ngô chuyển gen Event TC1507 (ngô mang gen cry1F kháng côn trùng cánh vảy) tại Việt Nam được triển khai trên cơ sở đảm bảo tuân thủ chặt chẽ các quy định hiện hành của Việt Nam được liệt kê dưới đây:
- Thông tư 69/2009/TT-BNNPTNT ngày 27/10/2009 về Quy định khảo nghiệm
đánh giá rủi ro đối với đa dạng sinh học và môi trường của giống câ y trồng biến đổi gen.
- Thông tư 72/2009/TT-BNNPTNT ngày 17/11/2009 về việc Ban hành danh mục loài cây trồng biến đổi gen được phép khảo nghiệm đánh giá rủi ro đối với đa dạng sinh học và môi trường cho mục đích làm giống cây trồng ở Việt Nam.
- Quyết định 3392/QĐ/BNN-KHCN ngày 25/11/2009 về việc Thành lập Hội đồng An toàn Sinh học ngành NN&PTNT.
- Quyết định 252/QĐ-BNN-KHCN ngày 29 tháng 1 năm 2010 về việc Chỉ định tổ chức khảo nghiệm đánh giá rủi ro đối với đa dạng sinh học và môi trường của giống cây trông biến đổi gen.
- Nghị định số 69/2010/NĐ-CP ngày 21/6/2010 về an toàn sinh học đối với sinh vật biến đổi gen, mẫu vật di truyền và sản phẩm của sinh vật biến đổi gen.
- Quyết định số 1449/QĐ-BNN-KHCN ngày 31/05/2010 của Bộ NN&PTNT về việc cấp phép khảo nghiệm hạn chế đánh giá rủi ro đối với đa dạng sinh học và môi trường của cây ngô biến đổi gene cho Công ty Pioneer Hi-Bred Việt Nam. Viện Di truyền Nông nghiệp là cơ quan khảo nghiệm. Thuyết minh đề cương

3
kế hoạch khảo nghiệm ngô chuyển gene trong diện hẹp đã được Bộ NN&PTNT phê duyệt.
- Quyết định số 907/QĐ-BNN-KHCN ngày 05 tháng 5 năm 2011 của Bộ NN&PTNT về việc công nhận kết quả khảo nghiệm hạn chế và cấp phép khảo nghiệm diện rộng đánh giá rủi ro đối với đa dạng sinh học và môi trường của giống ngô chuyển gen Event TC1507 - mang gen Cry1F kháng côn trùng cánh vảy, cho công ty TNHH Pioneer Hi-Bred Việt Nam; Viện Di truyền là đơn vị khảo nghiệm. Thuyết minh đề cương kế hoạch khảo nghiệm ngô biến đổi gen trên diện rông (4 vùng sinh thái) đã được Bộ NN&PTNT phê duyệt.
1.5 Các văn bản kèm theo báo cáo
- Phụ lục 1: các văn bản phụ lục liên quan đến cấp phép KNHC; vận chuyển và lưu trữ hạt giống; thực hiện và giám sát khảo nghiệm; xử lý và tiêu hủy n gô BĐG gene TC1507 trong 2 vụ khảo nghiệm hạn chế tại Văn Giang.
- Phụ lục 2: các văn bản phụ lục liên quan đến cấp phép KNDR; vận chuyển và lưu trữ hạt giống; thực hiện và giám sát khảo nghiệm; xử lý và tiêu hủy ngô chuyển gene TC1507 trong khảo nghiệm diện rộng tại 4 địa điểm khảo nghiệm là Vĩnh Phúc, Nghệ An, Đắk Lắk và Đồng Nai.
.- Phụ lục 3: các hình ảnh ghi nhận tập huấn, thực tế đồng ruộng trong KNHC và KNDR tại các điểm khảo nghiệm.

4
2 TỔNG QUAN VỀ NGÔ BIẾN ĐỔI GEN EVENT TC1507
2.1 Thông tin chung về ngô event TC1507
Tên sự kiện Event TC1507
Đơn vị đăng kí C.ty TNHH Pioneer Hi-Bred Việt Nam
Địa chỉ: Lầu 11, cao ốc Center Plaza, 17 Lê Duẩn, Quận 1, TP. Hồ Chí Minh. Điện thoại: (+84-8) 3 8251610 Ext. 8005 Fax: (+84-8) 3 8251620
Sinh vật cho Vi khuẩn Bacillus thuringiensis (Bt) ssp. Aizawai dòng PS811 mang gen cry1F kháng côn trùng bộ Cánh vảy
Vi khuẩn Streptomyces viridochromogenes mang gen chỉ thị pat kháng thuốc trừ cỏ nhóm glufosinate-ammonium
Sinh vật nhận Ngô, bắp, bẹ (Maize/Corn)
Vật liệu bố mẹ Các dòng ngô lai được nghiên cứu và SX bởi C.ty Pioneer Hi-Bred
Giống nền Ngô lai 30Y87
Nguồn gốc xuất sứ Mêxicô, Trung-Nam Mỹ
Miêu tả giống Kháng côn trùng bộ cánh vảy (Lipedoptera)
P.P chuyển nạp gen Súng bắn gen (microprojectile bombardment)

5
2.2 Thông tin về sinh vật cho gen
Ngô chuyển gen e vent 1507 tổng hợp p rotein Cry1F và phosphinothricin acetyltransferase (PAT) được tạo ra bằng phương pháp chuyển đoạn DNA thẳng có mang gen cry1F (có nguồn gốc từ Bacillus thuringiensis (Bt) ssp. aizawai), gen pat (có nguồn gốc từ Streptomyces viridochromogenes) cùng các thành phần điều khiển gen cần thiết khác vào các tế bào phôi (embryos) ngô tự nhiên Hi-II.
Bacillus thuringiensis (Bt) thuộc nhóm Bacillus cereus (Bc), là vi khuẩn gram dương, sinh bào tử và là chủng vi khuẩn được tìm thấy ở hầu hết các loại đất. Bt là chủng vi khuẩn có thể tự tạo protein kết tinh (parasporal crystals) trong quá trình hình thành bào tử và chính khả năng tự tạo crystal này thể hiện sự khác biệt của vi khuẩn Bt so với B. cerus. Các protein crystals thuộc loại delta-endotoxins (Cry toxins) được tổng hợp bởi nhóm gen cry của vi khuẩn và thường có tính chất gây độc với một số loài côn trùng nhất định ( Ibrahim và cs, 2010). Đặc tính trừ sâu của Bt đã được biết đến từ khá lâu và thuốc trừ sâu sinh học Bt lần đầu tiên được thương mại hóa và sử dụng từ năm 1938. Hiện nay, người ta đã tìm thấy và phân loại trên 300 Cry protein khác nhau có độc tính với chủ yếu trên một số nhóm côn trùng bộ cánh vảy (Lepidoptera), bộ cánh cứng (Coleoptera), bộ hai cánh (Diptera) và một số ít trong đó (Cry6) có thể gây độc đối với tuyến trùng (Sanahuja và cs, 2011). Ngô BĐG event TC1507 được chuyển nạp duy nhất một bản gen tổng hợp protein Cry1F. Gen cry1F có nguồn gốc từ vi khuẩn Bacillus thuringiensis (Bt) ssp. aizawai dòng PS811. Tính di truyền cũng như mức độ thể hiện của gen cry1F tổng hợp protein Cry1F trên ngô event TC1507 được chứng minh là luôn ổn định qua các thế hệ. Việc tổng hợp protein Cry1F giúp ngô event TC1507 có thể kháng một số loài côn trùng thuộc bộ cánh vảy (Lepidoptera) ví dụ C. partellus và H. armigera và không phụ thuộc yếu tố mùa vụ.
Streptomyces viridochromogenes là chủng vi khuẩn không gây bệnh, thường được tìm thấy trong đất (Eckes và cs., 1989). Streptomyces viridochromogenes mang gen chỉ thị pat mã hóa tổng hợp phosphinothricin-N-acetyltransferase (PAT) là enzyme ức chế quá trình chuyển hóa tổng hợp Glutamine từ Glutamate dẫn đến ngăn chặn quá trình giải độc ammonia dư thừa và do đó ảnh hưởng tới quá trình quang hợp ở thực vật. Tổng hợp protein PAT giúp ngô event TC1507 kháng thuốc trừ cỏ nhóm glufosinate-ammonium (Glufosinate). Trong sự kiện TC1507, pat được sử dụng làm gene chỉ thị, chỉ đư ợc dùng trong quá trình chọn tạo event TC1507 và giúp có thể chọn chính xác các cây ngô con có mang gene chuyển. Protein PAT đã được Sở kiểm tra Sức khỏe Thực vật và Động vật (APHIS) đánh giá là an toàn và không tác động lên sinh vật không chủ đích gồm cả sinh vật có ích. Đoạn DNA mã hóa tổng hợp

6
protein PAT được chứng minh là không độc hại và protein PAT có cấu trúc không tương đồng với các protein gây độc hại hay dị ứng đã được công bố (OECD, 1999). Ngoài ra, sản phẩm thuốc diệt cỏ glufosinate ở Việt Nam chỉ có một số ít các sản phẩm thương mại là Basta (Bayer Vietnam Ltd.), Fasfix (Công ty CP BVTV Sài Gòn), và Proof (Công ty TNHH Alfa (SaiGon). Do hiệu quả di ệt cỏ thấp nên hầu như đã ngừng bán trên thị trường (theo Bayer Vietnam Ltd.). Như vậy, pat chỉ được sử dụng làm gene chỉ thị, protein PAT đã được APHIS đánh giá an toàn sinh học, và thuốc diệt cỏ glufosinate không phổ biến ở Việt Nam.
Ngoài ra, trong giố ng đối chứng tham khảo, stack TC1507 x NK603, dòng ngô biến đổi gen NK603 tổng hợp protein CP4 EPSPS được tạo ra bằng cách chèn đoạn DNA thẳng có chứa hai đoạn gen (2 copies) mang thông tin di truyền tổng hợp 5 -enolpyruvylshikimate-3-phosphate synthase (gen cp4 epsps) có nguồn gốc từ vi khuẩn Agrobacterium sp. dòng CP4 cùng các thành phần điều khiển gen cần thiết khác.
2.3 Thông tin về sinh vật nhận
- Tên khoa học: Zea mays L. - Tên thông thường: Ngô, bắp, bẹ (Maize/Corn) - Họ: Gramineae - Chi: Zea - Loài: mays (2n=20)
Nguồn gốc: Ngô (Zea mays L.) thuộc chi Zea, là loại cây lương thực phổ biến, được thuần canh tại khu vực Trung-Nam Mỹ, sau đó lan tỏa ra khắp châu Mỹ và các phần còn lại của thế giới. Ngô thuộc bộ Maydae, họ phụ Panicoideae thuộc họ hòa thảo (Gramineae) trong đó bao gồm một số loài hoang dại khác, gọi chung là ngô hoang. Bộ Maydae còn bao gồm một số chi là: Zea và Tripsacum phân bố chủ yếu tại vùng Western Hemisphere, và các chi Coix, Polytoca, Chionachne, Schlerachne, và Trilobachne ở châu Á (CFIA, 1994). Ngô hoang hiện chỉ được tìm thấy trong tự nhiên ở Mê-xi-cô và Gua-te-ma-la. Giống nền: Ngô lai 30Y87 có thời gian sinh trưởng phát triển trung bình 115-120 ngày; có số lá trung bình, bản lá rộng, khả năng quang hợp rất tốt. Chiều cao đóng bắp trung bình, bắp dễ bẻ, lá bi bao kín có khả năng hạn chế mối mọt và thối mốc trên đồng ruộng. Đặc biệt ngô 30Y87 có khả năng chịu hạn rất tốt, sạch bệnh, cho năng suất cao và ổn định (8-12 tấn/hecta).

7
2.4 Phương pháp chuyển nạp gen tạo dòng ngô Event TC1507
Ngô event TC1507 được tạo ra bằng phương pháp bắn gene (Biolistics™ PDS-1000He) vào các tế bào phôi ngô Hi-II. Một đoạn DNA thẳng (6235 bp) kí hiệu PHI8999A (Hình 2-1) mang các gen cry1F, pat và kanr
cùng các thành phần điều khiển gen cần thiết khác được cắt ra từ vector PHI8999 (Hình 2-2) bằng việc sử dụng enzyme PmeI. Các gen này chứa thông tin tổng hợp delta -endotoxin của vi khuẩn Bacillus thuringiensis var. aizawai PS811; phosphinothricin acetyltrasnferase từ nấm Streptomyces viridochromogenes và kháng kháng sinh kanamycin. Protein Cry1F trong ngô chuyển gen TC1507 là protein nhân tạo, có cấu trúc ngắn hơn so với protein trong vi khuẩn Bt tự nhiên. Protein này có khả năng diệt sâu đục thân châu Âu (Ostrinia nubilalis), sâu đục thân châu Á (Ostrinia furnacalis), sâu xám (Spodoptera litura) và giảm đáng kể sự gây hại của các loài sâu này với sự hoạt động của gen cry1F trong ngô BĐG. Sự hoạt động của gen cry1F trong ngô chuyển gen TC1507 được điểu khiển bởi promoter polyubiqutine có tên là ubiZM1và có nguồn gốc từ chính cây ngô (Hình 2-1). Ngô TC1507 được chứng minh là chỉ mang duy nhất một đoạn mã (copy number) gen cry1F, được di truyền và luôn thể hiện ổn định qua các thế hệ.
Ở ngô event TC1507, gen pat, có nguồn gốc từ S. viridochromogenes, mã hóa tổng hợp phosphinothricin-N-acetyltransferase (PAT) và được điểu khiển bởi CaMV 35S promoter. Việc chuyển gen pat ở thực vật giúp cây có thể tổng hợp protein PAT gvà do đó có khả năng kháng thuốc trừ cỏ nhóm glufosinate -ammonium (Glufosinate). Trong sự kiện TC 1507, pat được sử dụng như là gene chỉ thị, giúp trong quá trình chọn lọc để chọn được chính xác các cây ngô con có mang gene chuyển nạp. Protein PAT được biểu hiện ở lá với nồng độ thấp 40.8 pg/µg tổng lượng protein giúp cây ngô con chuyển gene chịu thuốc diệt cỏ chứa glufosinate.
Các kết quả kiểm tra trong phòng thí nghiệm đều khẳng định trên ngô chuyển
gen TC1507 chỉ chứa duy nhất một đoạn AND có mang gen cry1F và gen pat nhưng không mang gen kháng kanamycin. Ngoài ra. không có bất cứ Plasmid hay đoạn DNA nào khác được dùng trong sự kiện chuyển gene TC1507 này.
Sau khi chuyển nạp, các tế bào phôi được chuyển vào môi trường nuôi cấy có
chứa thuốc diệt cỏ nhóm glufosinate-ammonium như chất chọn lọc, cho phép các tế bào phôi mang gen chuyển có thể sống só t và tăng trưởng trong môi trường này. Như mong đợi, phần lớn các tế bào của cây cũ (không mang gen chuyển) đã được loại bỏ trong quá trình nuôi cấy trong môi trường chọn lọc này. Các tế bào phôi sống sót và

8
tạo các mô sẹo khỏe, chịu glufosinate-ammonium, sau đó được tái tạo thành các cây trong nhà kính. Các bước tiếp theo như kiểm tra chuyên sâu hơn bằng phân tích phân tử (PCR, Northern blot, Sounthern blot vv..), các bước chọn lọc ngoài điều kiện đồng ruộng cũng như việc kiểm tra, đanh giá khả năng kháng côn trùng chủ đích ví dụ sâu đục ngô Châu Âu (Ostrinia nubulalis). Ngô event TC1507 cuối cùng được lựa chọn tạo ra mang tính kháng một số loài côn trùng bộ Cánh vảy (Lepidoptera).
2.4.1 Kích thước, trình tự, chức năng của đoạn gene đưa vào
2.4.1.1 Gen cry1F
Gene cry1F đã được cắt ngắn và tối ưu cho biểu hiện trong thực vật dưới sự điều khiển của promoter khiển tổng hợp protein ubiquitin và terminator ORF25PolyA (Bảng 2-1). Protein Cry1F trong giống ngô TC1507 kháng lại côn trùng cánh vảy gây hại bao gồm cả sâu đục ngô Châu Á (Ostrinia furnacalis). Cơ chế hoạt động của các tinh thể protein Cry1F là gây chết ấu trùng của những loài côn trùng mẫn cảm với loại protein nay, do làm tổn thương mô và làm mất khả năng ăn của ấu trùng. Trong quá trình tiêu hóa trong ruột ấu trùng, protein này hòa tan và trong vài trư ờng hợp quá trình thủy giải protein bởi các protease có trong ruột của côn trùng sẽ tạo ra phần có độc tính. Phần này sẽ tiếp xúc đặc hiệu với các thụ quan của đường ruột và dẫn đến phá hủy cân bằng thẩm thấu của các tế bào trong ruột giữa của côn trùng, cuối cùng làm chết ấu trùng (Ibrahim và cs., 2010).
Ở dạng tự nhiên, Cry1F được tạo ra lượng lớn dưới dạng tiền độc tố dài 1174
axít amin. Sau quá trình thủy phân và hòa tan trong đường ruột của ấu trùng côn trùng mẫn cảm, phần độc tố có hoạt tính có chiều dài 600 axít amin nằm ở đầu Nitơ (N-terminal) của protein tiền độc tố được tạo ra. Mặc dù việc cắt chính xác chưa được biết, dựa trên mô phỏng trên máy vi tính và dữ liệu trong phòng thí nghiệm, phần có hoạt tính độc được dự đoán nằm từ axít amin 28 đến 612. Do đó, để tạo được tính kháng côn trùng, đoạn cắt ngắn từ gene cry1F chỉ bao gồm đoạn mã hóa phần mang hoạt tính độc tố được chuyển nạp vào ngô.
2.4.1.2 Gen pat
Gene pat được tối ưu cho biểu hiện trong thực vật chịu sự kiểm soát của 35S promoter và teminator (Bảng 2-1). Chức năng của protein PAT trong ngô TC1507 là chất đánh dấu chọn lọc và cho phép chịu các thuốc diệt cỏ chứa glufosinate-ammonium. Sản phẩm này được đăng ký sử dụng cho ngô chịu glufosinate. Gene pat

9
có nguồn gốc từ vi khuẩn đất Streptomyces viridochromogenes chủng Tü494. Nó mã hóa cho một đoạn polypeptide dài 183 axít amin, và protein PAT hoàn chỉnh ở trạng thái tự nhiên được biết là hai cấu phần protein giống nhau nặng khoảng 43 kDa (Wehrmann et al., 1996).
Hoạt tính của protein PAT đã đư ợc mổ tả chi tiết (OECD, 1999). L-
phosphinothricin hay glufosinate (L-PPT) là thành phần hoạt tính trong thuốc diệt cỏ chứa glufosinate-ammonium. Nó gắn và bất hoạt enzym tổng hợp glutamine trong thực vật dẫn đến ngăn chặn quá trình giải độc ammonia dư thừa, cuối cùng làm cây chết. Hoạt tính của protein PAT (phosphinothricin-N-acetyltransferase) được đặc trưng bời xúc tác sự chuyển hóa L-PPT thành N-acetyl-L-PPT. Dạng không có hoạt tính này sẽ không gắn với enzym tổng hợp glutamin (De Block và cộng sự, 1988). Do đó biểu hiện của PAT trong ngô TC1507 dẫn đến sự chuyển hóa glufosinate hiện diện trong môi trường chọn lọc thành dạng không có hoạt tính, cho phép sự khử độc ammonia trong cây (EPA, 1995 & 1997).
2.4.2 Phương pháp xác định, phát hiện gen, đặc trưng của gen
Sự hiện diện của các gen được chuyển nạp vào ngô TC1507 được nghiên cứu qua nhiều thế hệ để bảo đảm tính ổn định di truyền. Biểu hiện kiểu hình và các phân tích phân tử cho thấy rằng ngô TC1507 mang gene cry1F được thừa hưởng ổn định qua ít nhất 6 thế hệ theo mô hình di truyền của Mendel. Năng suất nông nghiệp của ngô TC1507 được khẳng định giống với ngô không chuyển gen ngoại trừ các thay đổi mong muốn trong việc kháng côn trùng. Kết quả là ngô TC1507 biểu hiện prot ein Cry1F cho phép kháng một số côn trùng cánh vảy có hại.
Các quan sát kiểu hình chỉ ra rằng các chuyển gene này được di truyền như các gene trội theo mô hình phân ly của Mendel. Phương pháp phân tích có liên quan đến việc xịt chất chỉ thị glufosinate-ammonium ở mỗi thế hệ để ghi nhận và loại bỏ các phân ly vô nghĩa (các cây không chứa bản sao của gene chuyển). Dữ liệu của sự phân ly ở các thế hệ tiếp theo được lấy từ các cây thu nhận từ thế hệ F1 dựa trên đặc điểm chịu thuốc diệt cỏ và kháng lại sâu đục ngô Châu Âu. Tất cả các cây được xác định chịu glufosinate-ammonium cũng cho thấy kháng lại sự xâm nhiễm của sâu đục ngô Châu Âu.
Dùng phương pháp PCR, sự hiện diện của cả hai gen cry1F và gene đánh dấu
chọn lọc pat trong dòng ngô TC1507 ở hai cá thể lai r iêng biệt lấy từ thế hệ T1S1 đã khẳng định sự ổn định của các gene này trong bộ gene của ngô (CRL-EU, 2005; Lee

10
và cs., 2004). Các thí nghiệm lai phân tử Southern blot cũng cho kết quả tương tự trên các thế hệ T1S1 và BC4F1 củng cố kết luận rằng việc di truyền này là ổn định qua nhiều thế hệ.
Hình 2-1: Sơ đồ các yếu tố di truyền đoạn DNA thẳng Pmei PHP8999A được
chuyển nạp trong qui trình tạo dòng ngô Event TC1507.
Hình 2-2: Plasmid PHP8999
P H P 8 9 9 9 9504 bp
Cry1F (Trunc)
PAT
nptII
UBIZM 1(2)
CAMV35S (Prom) ORF25PolyA
CAMV35S (Term)
Pme I (21)
Pme I (6256)
Hind III (59)
Hind III (3949)
Hind III (6119)
EcoR I (1488)
EcoR I (4690) EcoR I (4744)
EcoR I (6073)
BamH I (2101)
BamH I (3929)
BamH I (5290) BamH I (5605)
BamH I (6095)
Pst I (101)
Pst I (2087)
Pst I (3031)
Pst I (3945) Pst I (5861)
Pst I (6117)

11
Bảng 2-1: Các yếu tố di truyền có trong plasmid PHP8999
Yếu tố di truyền Kích cỡ (kb)
Chức năng
ubiZM 1(2) 1.98 Promoter điều khiển tổng hợp protein ubiquitin (thêm vùng intron và vùng 5' không mã hóa của gene ubiquitin ngô) từ Zea mays (Christensen và cs, 1992)
cry1F (cắt ngắn) 1.82 Gen cry1F đã cắt ngắn lấy từ Bacillus thuringiensis var. aizawai (đã tối ưu cho biểu hiện trong thực vật)
ORF25PolyA 0.72 Terminator từ Agrobacterium tumefaciens pTi15955
CaMV 35S promoter
0.55 35S promoter từ virus khảm bông cải (Odell và cs, 1985)
pat 0.55 Gene kháng glufosinate (đã tối ưu cho biểu hiện trong thực vật), dựa vào gene mã hóa phosphinothricin acetyltransferase lấy từ S. viridochromogenes (Wohlleben và cs., 1988; Eckes và cs., 1989)
CaMV 35S terminator
0.20 35S terminator từ virus khảm bông cải (Pietrzak và cs, 1986)
2.5 Đặc tính và hiện trạng sử dụng ngô event TC1507
2.5.1 Protein CRY1F kháng côn trùng, sâu hại
Gen cry1F có nguồn gốc từ vi khuẩn Bacilus thuringiensis (Bt) var. aizawai. Bt thuộc nhóm vi khuẩn gram dương, là vi sinh vật mang bào tử được biết đến nhiều nhất về giá trị sử dụng như thuốc trừ dịch hại sinh học. Việc tổng hợp protein Bt ở cây trồng chuyển gen giúp cây trồng có tính chống côn trùng (Ibrahim và cs., 2010; Prieto Samsonov và cs, 1997). Protein Cry1F là loại δ -endotoxin và là loại protein có tính kháng côn trùng cao nhất trong nhóm Bt. Hoạt tính của δ-endotoxin trong protein Cry1F tổng hợp trong ngô event 1507 là đầu tiên chúng có thể dính vào những vị trí nhất định trên mô ruột giữa (midgut) của những loài côn trùng mẫn cảm với loại protein này (de Maagd và cs., 2003; Schnepf và cs., 1998). Tiếp đến, chúng ức chế quá trình trao đổi ion của mô ruột dẫn đến tê liệt, thậm chí gây chết hoàn toàn gây ra bởi nhiễm khuẩn (Bravo và cs., 2007). Protein Cry1F chỉ có tính độc duy nhất đối với

12
sâu non một số loài thuộc bộ cánh vảy (Lepidoptera). δ-endotoxin trong protein Bt không thể bám dính trong bề mặt tế bào niêm mạc ruột của động vật có vú. Do vậy, loại protein nay hoàn toàn không gây độc đối với con người cũng như các loài vật nuôi (Siegel, 2001).
Hoạt tính sinh học của protein CRY1F được kiểm tra trên một loạt các loài côn trùng gây hại trên ngô. Các khảo nghiệm được thiết lập bằng cách cho côn trùng ăn thức ăn nhân tạo đã được xử lý với dịch chứa protein CRY1F sản xuất bởi vi khuẩn. Các đặc tính sinh hóa của protein CRY1F tạo bởi cây trồng biến đổi gen cry1F và protein CRY1F tổng hợp từ vi khuẩn (B.t) được khẳng định là không có sự khác biệt (Evans, 1998). Các loài côn trùng sử dụng trong khảo nghiệm bao gồm sâu đục ngô Châu Âu (Ostrinia nubilalis), sâu ăn lá (Spodoptera frugiperda), sâu ăn trái (Helicoverpa zea), sâu xám (Agrotis ipsilon), sâu đục thân (Elasmopalpus lignosellus), sâu đục mía ( Diatraea saccharalis), sâu đục ngô tây nam ( Diatraea grandiosella), sâu đục rễ ngô phía tây ( Diabrotica virgifera virgifera), rệp lá ngô (Rhopalosiphum maidis), và bọ nhảy lá ngô (Dalbulus maidis) (Evans, 1998; Herman và Korjagin, 1999). Kết quả thể hiện sâu đục thân ngô Châu Âu, sâu đàn, sâu đục trái, và sâu xám mẫn cảm với cả protein CRY1F được tổng hợp từ cây trồng chuyển gen và từ vi khuẩn (Evans, 1998). Ở một nghiên cứu khác, việc sử dụng protein CRY1F từ vi khuẩn diệt được sâu đục thân, sâu đục ngô tây nam, và sâu đục mía, nhưng nó không gây độc đối với sâu đục rễ ngô phía tây, rệp lá ngô và và bọ nhảy lá ngô (Herman and Korjagin, 1999).
Ngoài ra, hiệu quả của ngô event TC1507 chống lại hàng loạt các côn trùng gây hại cũng được kiểm tra và so sánh với các đối chứng là ngô thô ng thường trong các điều kiện đồng ruộng Việt Nam (Bảng 2-2). Tóm lại, các kết quả nghiên cứu đều cho thấy ngô chuyển gen mang event TC1507 tốt hơn giống ngô thông thường (không biến đổi gen) trong việc kháng lại một số loài côn trùng bộ cánh vảy.

13
Hình 2-3: Mô hình hoạt động của protein Cry gây độc đối với côn trùng (Ibrahim và cs., 2010)

14
Bảng 2-2: So sánh hiệu quả của ngô TC1507 và ngô không chuyển gen Ngô
ECB tổn thương lá
(1-9)
ECB tổn thương (inch đường đục) a
ECB tổn thương trái
(1-9)
FAW
a tổn thương
(1-9)
CEW
a tổn thương
(1-9)
SWCB
a tổn thương
(ins đường đục)
BCW tổn thương
(% cut-3 ngày)
SCB tổn thương
(% )b
TC 1507 9 0.4 7.8 8 6 0.4 32 4 Đ/C 2 9 4.3 2 4 6.4 100 90
Ghi chú:
a: Điểm tổn thương được ghi nhận bằng quan sát theo thang điểm 1 - 9 (1: nhạy hoàn toàn; 9: kháng hoàn toàn) b: Sự khác biệt bình phương ít nhất (0.05%) = 9.6 ECB = Sâu đục thân ngô Châu Âu FAW = Sâu đàn CEW = Sâu đục trái SWCB = Sâu đục thân ngô Tây Nam BCW = Sâu xám SCB = Sâu đục mía

15
2.5.2 Protein PAT kháng glufosinate-ammonium trong đánh dấu chọn lọc
Gen pat sử dụng trong quá trình chuyển nạp gen trong việc tạo dòng ngô event TC1507 có nguồn gốc tự nhiên từ vi khuẩn Streptomyces viridochromogenes (Eckes và cs., 1989). S. viridochromogenes sản xuất ra L-phosphinothricyl-L-alanyl-alanine (L-PPT) có tính chất kháng thuốc trừ cỏ không chọn lọc. Qua nhiề u thập kỉ, gen pat được sử dụng để chuyển nạp vào nhiều loại cây trồng biến đổi gen nhằm mục đich tạo tính kháng PPT cũng như dạng tổng hợp của nó, glufosinate-ammonium. Ngô chuyển gen tổng hợp protein PAT bắt đầu được thương mại và trồng tại Mỹ từ năm 199 6. Hoàn toàn không ghi nhận bất kì tác dụng phụ nào đối với sức khỏe con người trong việc sử dụng gen pat trên cây trồng chuyển gen như canola và một số giống ngô (Hérouet và cs., 2005; OECD, 1999a; OECD, 2002a).
Sự biểu hiện của protein PAT trong ngô TC1507 cho phép cây chịu thuốc diệt cỏ có glufosinate-ammonium. Glufosinate-ammonium ức chế enzym tổng hợp glutamine dẫn đến tích tụ ammonia và làm chết tế bào cây. Enzym PAT có tính khử độc glufosinate hay phophinothricin bằng acetyl hóa nó thành chất không có hoạt tính. Các thử nghiệm trên ruộng cho thấy ngô event TC1507 chịu thuốc diệt cỏ chứa glufosinate-ammonium ở các mức độ cao và không có hiện tượng gây độc cho cây. Dòng ngô TC1507 có thể được sử dụng như sản phẩm chịu glufosinate -ammonium, khi các sản phẩm thuốc diệt cỏ như vậy được đăng ký cho sử dụng trên ngô chịu glufosinate.
Ở nhiều nghiên cứu, trong tất các mẫu đối chứng hàm lượng protein CRY1F và PAT đều dưới ngưỡng có thể phát hiện . Những nghiên cứu này cũng cho thấy hàm lượng protein CRY1F tìm thấy ở các mức độ có thể đo được trong tất cả các mẫu mô của ngô event TC1507 được kiểm tra. Thêm vào đó, sự biểu hiện của protein PAT chỉ được ghi nhận ở các mức độ có thể đo được trong các mẫu mô lá ngô event TC1507 trong tất cả các mẫu kiểm tra. Điều này khẳng định protein PAT chỉ được biểu hiện trong dòng ngô TC1507 cho phép cây chống chịu glufosinate-ammonium.
2.5.3 Thông tin liên quan đến biểu hiện tính trạng của gen chuyển nạp vào ngô chuyển gen TC1507
Ngô TC1507 cho kết quả biểu hiện protein Cry1F cho phép kháng lại một số côn trùng cánh vảy nhất định và protein PAT cho phép sử dụng hóa chất diệt cỏ glufosinate-ammonium như chất đánh dấu chọn lọc. Các kết quả của nghiên cứu biểu hiện protein trong dòng ngô lai Bt Cry1F TC1507 được tóm tắt trong Bảng 2-3. Các mức độ biểu hiện của protein Cry1F dưới ngưỡng phát hiện trong tất các mẫu đối

16
chứng. Nghiên cứu này cho thấy biểu hiện của protein Cry1F được tìm thấy ở các mức độ có thể đo đếm được trong tất cả các mẫu mô của ngô TC1507 được kiểm tra.
Sự biểu hiện của protein PAT được tìm thấy trong các mẫu mô lá ở các mức độ
có thể phát hiện đến khoảng 54 ng/mg tổng lượng protein. Các mức độ này đủ cho sức chịu đựng thuốc diệt cỏ chứa glufosinate-ammonium trong toàn bộ cây. Trong các mô khác, mức độ của protein PAT nằm dưới ngưỡng phát hiện (20 ng/mg).
Bảng 2-3: Mức độ biểu hiện protein Cry1F đo được trong các mô ngô TC1507
Mô
Cry1F trung bình (pg/µg protein tổng)
Ðộ lệch chuẩn
Khoảng nhỏ/lớn nhất
Lá 110.9 ± 27.2 56.6 - 148.9 Phấn hoa 135.5 ± 13.5 113.4 - 168.2 Râu bắp 50.3 ± 16.5 26.8 - 79.8 Thân 550 ± 104 355.9 - 737.4 Toàn bộ cây 1063.8 ± 361.7 803.2 - 1572.7 Hạt 89.8 ± 23.3 71.2 - 114.8 Toàn bộ cây héo 714.3 ± 95.5 622.2 - 845.3
2.5.4 Thông tin khác biệt của dòng ngô chuyển gene TC1507 so với cây bố mẹ
Trong thời gian khảo nghiệm trên ruộng qua các mùa vụ ở các địa điểm khác nhau trên thế giới, rất nhiều dữ liệu nông nghiệp (nảy mầm, mức tăng trưởng, các đơn vị nhiệt tích tụ lên nhị và nhụy hoa, sự ngã rạp của thân, chiều cao của cây, chiều dài của trái, mức độ phân bố cuối cùng, thời gian héo của lá, bệnh xuất hiện tình cờ, côn trùng phá hoại, độ ẩm và mật độ của hạt) đã được thu thập và khẳng định sự tương tự về kiểu hình của ngô TC1507 so với ngô thông thường không chuyển gene.
-
- Khả năng phát tán: Các ngô
Không có thay đổi khác biệt trong việc tạo phấn hoa, tạo hạt, khả năng sống sót hoặc khả năng nảy mầm của hạt giống so với ngô không chuyển gene đã quan sát được trong các khảo nghiệm ở ngô TC1507.
lai đã được thuần hóa tới mức các hạt giống không thể được phát tán mà không có sự can thiệp của con người. Các dòng ngô TC1507 cho thấy không có sự khác biệt trong phát tán so với ngô không chuyển gene.

17
- Khả năng sống sót: Ngô được trồng đã thuần hóa đến mức nó không thể tồn tại bên ngoài môi trường nông nghiệp được quản lý. Việc không thể ngủ đông ngăn chặn hạt giống ngô sống sót từ một mùa vụ này đến mùa vụ kế tiếp. Khi được trồng, sự biểu hiện của protein CRY1F trong ngô TC1507 giúp kháng một số côn trùng cánh vảy có hại và biểu hiện của PAT giúp chịu thuốc diệt cỏ chứa glufosinate-ammonium dùng trong đánh dấu chọn lọc. Các đặc tính sống sót của ngô TC1507 trong môi trường vẫn tương đương với các ngô không chuyển gene. Sự
kháng cự lại côn trùng cánh vảy nhất định không đủ để cho phép sự tồn tại của ngô bên ngoài môi trường nông nghiệp và các chất diệt cỏ phổ rộng glufosinate-ammonium không còn đư ợc sử dụng ở Việt Nam.
2.5.5 Phương pháp phát hiện cây ngô chuyển gene TC1507
Ngoài đồng ruộng, ngô event TC1507 chứa gene pat cho phép kháng lại glufosinate-ammonium nên việc phun các loại thuốc trừ cỏ gốc glufosinate-ammonium là cách đơn giản để xác định cây có phải là TC1507 hay không. Một cách khác để phát hiện TC1507 là dùng kit thử nhanh (que nhúng
, QuickStix™ Kit cho Cry1F - AS 016 cung cấp bởi Envirologix) đặc hiệu cho protein CRY1F. Bằng cách nhúng mặt bên của que thử vào dịch chiết lá sẽ cho kết quả xuất hiện (các) vạch màu. Sự hiện diện của hai vạch chỉ ra sự hiện diện của protein CRY1F trong khi sự hiện diện của một vạch đơn chỉ ra sự vắng mặt của protein CRY1F.
Trong điều kiện phòng thí nghiệm, các phương pháp như
PCR, real time RT-PCR cho phép phát hiện chính xác sự hiện diện của cả hai gen cry1F và gene đánh dấu chọn lọc pat trong dòng ngô TC1507 (CRL-EU, 2005; Lee và cs., 2004). Các thí nghiệm lai phân tử Southern blot cho phép củng cố kết luận sự hiện diện cũng như tính di truyền của các gen này là ổn định qua nhiều thế hệ (IBC No. PION-19-1111).
2.5.6 Thông tin về việc thương mại hoá (phóng thích và sử dụng) ngô chuyển gene TC1507 trên thế giới.
Ngô lai mang gen cry1F kháng côn trùng bộ cánh vảy, mã nhận dạng OECD DAS-Ø15Ø7-1 được thương mại hóa đầu tiên tại Mỹ vào năm 2001 (event TC1507, Herculex I®). Hiện trạng đăng ký sử dụng ngô event TC1507 trên thế giới để dùng như thức ăn chăn nuôi, thực phẩm và trồng trọt trên thế giới được thể hiện trong Bảng 2-4. Ngô event TC1507 được chấp thuận trồng tại Ác-hen-ti-na, Bra-xin, Ca-na-đa, Cô-lôm-bi-a, Nhật, và Mỹ. Từ khi được thương mại hóa ở Mỹ năm 2001, diện tích

18
trồng giống ngô chuyển gen này trên thế giới đã tăng lên đến 5.9 triệu hecta vào năm 2007, dẫn đầu là Mỹ, tiếp đến là Ca-na-đa và Ác-hen-ti-na. Từ năm 2003, ngô event TC1507 cũng được đã chấp thuận cho nhập khẩu để sử dụng trực tiếp làm thực phẩm, thức ăn chăn nuôi hay để chế biến ở Phi -líp-pin, và được trồng từ năm 2008. Các nước khác cho phép nhập khẩu ngô TC1507 là Nhật Bản, Đài Loan, Úc/ Niu- zi-lân, Trung Quốc, các nước Châu Âu, Hàn Quốc, Mê-xi-cô, và Nam Phi.
Bảng 2-4: Danh sách các nước cho phép trồng và/hoặc sử dụng ngô TC1507
Quốc gia Trồng trọt Chứng nhận an toàn
Môi trường Thức ăn/thực phẩm
Argentina 2005 2005 2005 Australia/NZ 2003 Brazil 2008 2008 2008 Canada 2002 2002 2002 China 2004 Colombia 2007 2006 Honduras 2009 2009 El Salvador 2009 European Union 2006 Japan 2005 2002 2002 Korea 2004 Mexico 2003 Philippines 2003 South Africa 2002 United State 2001 2001 2001 Uruguay 2011 2011

19
2.5.7 Mô tả nguy cơ, khả năng xảy ra nguy cơ để xác định rủi ro có thể xảy ra khi phóng thích sinh vật chuyển gene
Event TC1507 mang đoạn ADN chứa gen cry1F là gen chủ đích mã hóa cho protein Cry1F kháng côn trùng cánh phấn và gen chỉ thị pat mã hóa cho protein PAT cho tính trạng chịu thuốc diệt cỏ chứa glufosinate. Vì pat là gen chỉ thị, nó chỉ được dùng trong quá trình chọn lọc tạo event TC1507 và trong chọn lọc hay thử nghiệm để chọn chính xác các cây ngô con chuyển gen TC1507. Protein PAT chỉ phát hiện được biểu hiện ở lá với nồng độ thấp 40.8 pg/µg tổng lượng protein giúp cây ngô con chuyển gen chịu thuốc diệt cỏ chứa glufosinate. Protein PAT đã được Sở kiểm tra Sức khỏe Thực vật và Động vật (
APHIS) đánh giá là an toàn và không tác động lên sinh vật không chủ đích gồm cả sinh vật có ích. Ở sự kiện ngô chuyển gen TC1507, gen cry1F là gen chủ đích mã hóa cho protein Cry1F kháng côn trùng bộ cánh vảy . Như vậy, việc đánh giá rủi ro của ngô chuyển gen TC1507 chỉ liên quan đến gen chủ đích cry1F.
2.5.7.1 Khả năng sống sót ngoài môi trường của ngô TC1507 và tồn tại trong môi trường đất của protein Cry1F
Ngô không phải là loại thực vật tồn tại dai dẳng. Hạt giống ngô chỉ có thể tồn tại trong những điều kiện khí hậu nhất định. Những cây tự mọc dễ bị chết do lạnh, khô
Hình 2-4: Tình hình sử dụng ngô TC1507 trên thế giới

20
hoặc dễ dàng kiểm soát bằng các phương pháp nông học thông thường như canh tác đất và sử dụng thuốc diệt cỏ chọn lọc (Niebur, 1993). Ngô không có khả năng duy trì sinh sản nếu không có sự canh tác của con người và không phải là đối tượng xâm hại môi trường sống tự nhiên (OECD, 2003).
Hạt giống ngô là cơ chế tồn tại duy nhất ngoài môi trường. Ngô là cây trồng hàng năm đã được thuần hóa lâu đời, không có khả năng tự phát tán hạt giống mà không có sự can thiệp của con người. Các điểm khảo nghiệm ngô biến đổi gen luôn được thực hiện trên vùng đất có kiểm soát. Hạt giống ngô không có cơ chế ngủ đông, chúng có thể nảy mầm ngay sau khi thu hoạch khi có điều kiện thích hợp. Do đó hạt của ngô biến đổi gen sau khảo nghiệm được đề nghị sẽ đốt và chôn; thân sẽ được cày chôn lấp và khu vực khảo nghiệm được theo dõi cho ngô mọc tự nhiên ít nhất 2 lần, sau đó hủy và chôn lấp, nếu có.
Việc đưa vào vật liệu di truyền, các cấu trúc, đặc tính mới là kháng côn trùng cánh vảy và chịu thuốc diệt cỏ chứa glufosinate không làm ngô TC1507 trở thành cỏ dại. Không có các đặc tính của cỏ dại được mô tả bởi Baker có liên quan đến kháng hoặc nhạy cảm với côn trùng, và không có lý do để cho rằng protein Cry1F chống côn trùng chủ đích của ngô TC1507 sẽ làm ngô thành cỏ dại.
APHIS kết luận: Ngoài đặc tính kháng một số côn trùng cánh vảy và chịu thuốc diệt cỏ chứa glufosinate, ngô TC1507 có đặc điểm nông học tương tự như ngô lai truyền thống, và không thể hiện những đặc điểm có thể làm tăng tính cỏ dại. Canh tác ngô TC1507 không dẫn đến tăng tính cỏ dại của các ngô trồng khác.
Vì vậy, có thể kết luận rằng nguy cơ tiềm ẩn của ngô TC1507 trở thành cỏ dại và xâm lấn môi trường tự nhiên so với giống ngô nền không chuyển gen là không có.
Trên ngô TC1507, protein Cry1F được tổng hợp với hàm lượng thấp và được giữ lại trong cây. Do đó, cơ chế gây độc hoặc các đường tiếp xúc nhìn chung không liên quan đến protein Cry1F. Theo Halliday (1998) bằng cách xác định hàm lượng tích lũy protein Cry1F trong đất trồng ngô mô phỏng theo điều kiện phòng thí nghiệm, tốc độ phân hủy của protein Cry1F diễn ra nhanh, thời gian tồn tại ngoài môi trường ngắn. Kết quả của nghiên cứu này chỉ ra rằng hoạt tính sinh học của protein Cry1F trong đất suy giảm hơn 20 lần sau 28 ngày. Các ước tính thời gian bán phân hủy (DT50) của Cry1F là 3,13 ngày (Shanahan và cs., 2000; EA của USDA/APHIS, 2001; Shan và cs., 2008).
Một thí nghiệm trong 3 năm đánh giá mức độ tích tụ trong đất của protein Cry1F được tiến hành bằng cách trồng ngô chuyển gen TC1507 trên ba ruộng, cây ngô được

21
chôn ngay trong khu đất canh tác hoặc không canh tác sau mùa vụ mỗi năm. Các mẫu đất được thu thập từ các ruộng thí nghiệm, hàm lượng protein Cry1F trong mỗi mẫu được xác định bằng cách sử dụng phương pháp hấp thụ miễn dịch liên kết enzyme (ELISA). Kết quả không tìm thấy protein Cry1F trong bất kỳ các mẫu đất thu được từ các ruộng ngô CT1507. Kiểm tra khu đất canh tác không thấy có hoạt động sinh học ảnh hưởng bất lợi đối với ấu trùng sâu đục chồi cây thuốc lá
(Heliothis virescens). Các kết quả này chỉ ra rằng mức độ tích lũy protein Cry1F trong đất sau trồng ngô chuyển gen TC1507 trong ba năm liên tục là không đáng kể (Shan và cs., 2008).
2.5.7.2 Khả năng tương tác với các cây trồng cùng loài khác trong hệ sinh thái
Ngô (Zea mays ssp mays) thuộc về giống Zea, trong đó bao gồm một số lo ài hoang dã khác, gọi chung là ngô hoang. Chúng c hỉ được tìm thấy trong tự nhiên ở Mê-xi-cô và Gua-te-ma-la nên việc thử nghiệm ở Việt Nam không có mối đe dọa của việc thụ phấn chéo với các loài hoang dã.
Gen lan truyền từ ngô TC1507 sang các giống ngô khác có thể xảy ra thông qua phát tán phấn hoa trong canh tác mùa vụ. Ngô chủ yếu thụ phấn nhờ gió. Cây tung phấn trong vòng 10-13 ngày, phấn rụng chỉ tồn tại được trong thời gian ngắn, 98% hạt phấn bị rơi xuống trong vòng bán kính 25-50 m (EEA, 2002).
Do đó việc áp dụng các biện pháp cách li không gian (hang rào bảo vệ), và cách li thời gian (lệch thời gian tung phấn) có thể đảm bảo nguy cơ lan truyền gen từ TC1507 sang các loài họ hàng hoang dại tương thích sinh sản hay sang cây ngô khác là không đáng kể.
2.5.7.3 Khả năng ảnh hưởng đối với gia cầm và thủy sản
Với gia cầm: Một bản tóm tắt các giá trị độc tính cấp tính đối tượng chim cút con cho thấy nồng độ gây độc (LC50) là trên 100.000 mg ngô hạt chứa Cry1F/kg khẩu phần ăn (nồng độ cao nhất được thử nghiệm). Điều này tương đương với 10% hoặc 100.000 ppm protein Cry1F trong khẩu phần ăn từ ngô (Shanahan và cs., 2000; EA của USDA/APHIS, 2001).
Với thuỷ sản: Không có bằng chứng cho thấy sự mẫn cảm của các loài thuỷ sản nguy cấp nào với protein Cry1F. Khả năng tiếp xúc với Cry1F qua phấn hoa hoặc các bộ phận cây ngô chứa Cry1F là thấp. Các nghiên cứu độc tính đối với động vật sống

22
dưới nước không xương sống cho thấy Cry1F an toàn đối với cá hay động vật không xương sống. Mức độ hiệu quả (EC50
) đo được trong 48 giờ trong nghiên cứu độc tính cấp tính cho chế độ ăn uống của Daphnia magna là trên 100 mg phấn hoa Cry1F/lít. Mức độ này cao hơn nhiều lần so với nồng độ ước tính là 1,25 μg của phấn hoa Cry1F/lít bay vào ao nước (Shanahan và cs., 2000; EA của USDA/APHIS, 2001).
2.5.7.4 Khả năng ảnh hưởng tới côn trùng chủ đích
Hoạt tính sinh học của protein Cry1F được kiểm tra trên một loạt các côn trùng gây hại trên ngô. Thí nghiệm được tiến hành bằng cách cho côn trùng ăn thức ăn nhân tạo đã được xử lý với dịch chứa protein Cry1F có nguồn gốc từ vi khuẩn Bt. Các đặc tính sinh hóa củ a protein Cry1F sản xuất bởi thực vật và protein Cry1F lấy từ vi khuẩn Bt được khẳng định bởi Evans (1998). Các đối tượng côn trùng thí nghiệm là sâu đục ngô Châu Âu (Ostrinia nubilalis), sâu ăn lá (Spodoptera frugiperda), sâu đục trái (Helicoverpa zea), sâu xám (Agrotis ipsilon), sâu đục thân ( Elasmopalpus lignosellus), sâu đục mía ( Diatraea saccharalis), sâu đục ngô Tây Nam (Diatraea grandiosella), sâu đục rễ ngô phía Tây (Diabrotica virgifera virgifera), rệp ngô (Rhopalosiphum maidis), và bọ nhảy lá ngô (Dalbulus maidis) (Evans, 1998; Herman và Korjagin, 1999). Kết quả cho thấy s âu đục ngô Châu Âu, sâu đàn ( Mythimna separate), sâu đục trái ( Helicoverpa zea), và sâu xám (Agrotis ipsilon) mẫn cảm với cả protein Cry1F thực vật và Cry1F vi khuẩn (Evans, 1998). Ở một nghiên cứu khác sử dụng protein Cry1F tổng hợp từ vi khuẩn, số lượng nhỏ sâu đục thân, sâu đục ngô Tây Nam, và sâu đục mía mẫn cảm với loại protein nay, nhưng nó không gây độc đối với sâu đục rễ ngô phía Tây, rệp ngô và và bọ nhảy lá ngô (Herman and Korjagin, 1999). Kết quả khảo nghiệm ở Phi-lip-pin cho thấy TC1507 kháng sâu đục thân ngô châu Á (Ostrinia furnacalis), sâu xám (Spodoptera litura), sâu đàn (Mythimna separate), sâu đục trái ( Heliothis armigera). Tóm lại, các kết quả cho thấy ngô TC1507 thể hiện tốt hơn các giống ngô thường không BĐG trong việc chống lại các côn trùng cánh vảy.
2.5.7.5 Khả năng tác động lên côn trùng không chủ đích và côn trùng có ích
Côn trùng có ích: Các kết quả chỉ ra rằng với protein Cry1F tổng hợp từ vi khuẩn Bt, nồng độ gây độc 50% (LC50) cấp tính là trên 320 μg Cry1F/g khẩu phần ăn đối với kí sinh bộ Cánh màng (Nasonia vitripennis), và LC50 cấp tính > 480 μg Cry1F/g khẩu phần ăn đối với thiên địch green lacewing (Chrysopa carnea) và bọ rùa cánh cứng (Hippodamia convergens). Những nồng độ này cao hơn nhiều so với ước

23
tính giới hạn trên là 32 μg Cry1F phấn hoa/g khẩu phần ăn xuất phát từ ngô TC1507, chứng tỏ cho thấy ít có nguy cơ gây độc đối với kí sinh, thiên địch trong canh tác ngô TC1507 (Shanahan và cs., 2000; EA của USDA/APHIS, 2001).
Côn trùng không chủ đích: Năm 2005, Jeffrey và cộng sự tiến hành một thí nghiệm trên bướm Yamato-shijimi (có màu xanh nhạt), tên khoa học Pseudozizeeria maha (Kollar) nhằm đánh giá tác động của phấn ngô chứa protein Cry1F của ngô TC1507 được trồng tại Nhật Bản cho thấy: mức độ tiếp xúc dự kiến với phấn hoa chứa Cry1F của loài bướm này là dưới mức gây độc. Tương tự với hầu hết loài nhạy cảm khác đều không phát hiện thấy nguy cơ không đáng kể cho các loài bướm quan tâm khác của Nhật Bản khi phấn hoa chứa Cry1F vượt ra ngoài ruộng ngô hoặc ở mép ruộng.
Với ong mật và động vật chân đốt: Một đơn yêu cầu của Dow-Mycogen để miễn điều hòa ngô chuyển nạp gen cry1F có ghi chi tiết phân tích ảnh hưởng này trong một phụ lục CBI, và đơn yêu cầu tóm tắt nêu ra độc tính cấp tính trong chế độ ăn uống đới với ong mật, LD50
Một nghiên cứu quan sát đồng ruộng 3 năm cho ngô mang gen
> 640 ng Cry1F/ấu trùng (Shanahan và cs., 2000; EA của USDA/APHIS, 2001).
cry1F, sự kiện DAS-Ø15Ø7–1 về các tác động lên động vật chân khớp không chủ đích (Higgins và cs, 2009) được thực hiện tại bốn địa điểm trên các vùng trồng ngô của Mỹ từ năm 2004 đến năm 2006. Tại mỗi địa điểm, các cặp ruộng (0,8 hecta/ruộng) ngô lai thương mại chuyển nạp gen cry1F và đối chứng không chuyển gen đã được trồng. Phương pháp lấy mẫu được sử dụng để theo dõi sinh vật không chủ đích bao gồm quan sát bằng mắt thường, thẻ dính, bẫy pitfall, và bẫy litterbag. Phân tích mức độ đa dạng của động vật chân đốt cho thấy không có khác biệt đáng kể giữa ruộng ngô Bt với ngô Đ/C. Các phân tích thành phần loài, số lượng cá thể từng loài cũng cho thấy không có sự khác biệt đáng kể về mức độ phong phú loài giữa ruộng ngô Bt với ngô Đ/C. Kết quả của những nghiên cứu này cùng với kết quả từ những thí nghiệm trong phòng thí nghiệm trước đó ủng hộ cho giả thuyết ngô chuyển gen Cry1F không gây ra những ảnh hưởng tiêu cực đối với động vật chân đốt không chủ đích trên các ruộng canh tác loại ngô này.
Các sinh vật đất không chủ đích: Một nghiên cứu trong vòng 28 ngày nhằm đánh giá những ảnh hưởng lâu dài của protein Cry1F nguồn gốc từ vi khuẩn đối với bọ đuôi bật Collembola (Shanahan và cs., 2000; EA của USDA/APHIS, 2001). Thí nghiệm được tiến hành với ba cấp độ chất thử nghiệm Cry1F (0,63; 3,1 và 12,5 mg / kg của khẩu phần ăn). Kết quả thí nghiệm cho thấy < 10% chết ở cả lô thí nghiệm và Đ/C. Tỉ lệ sinh sản của bọ đuôi bật không bị ảnh hưởng đáng kể nào sau khi tiếp xúc

24
với chất thử nghiệm. Không ghi nhận bọ chết hay suy giảm số con cháu của bọ đuôi bật sau chúng tiếp xúc với Cry1F trong vòng 28 ngày. Kết quả của nghiên cứu này chỉ ra bọ đuôi bật không bị ảnh hưởng do tiếp xúc lâu dài với Cry1F ở các cấp độ cao hơn hẳn so với thực tế trên ruộng ngô. Dựa trên tính toán trường hợp xấu nhất, tiếp xúc sau thu hoạch dự đoán là 0,350 mg protein Cry1F/kg của toàn bộ vật liệu cây héo hoặc 0,063 mg protein Cry1F/kg đất khô.
Thử nghiệm độc tính cấp tính của protein Cry1F tổng hợp bởi vi khuẩn lên giun đất cũng được tiến hành. Nồng độ gây độc LC50
là > 2,5 mg Cry1F/kg đất khô. Nồng độ này cũng cao hơn đáng kể so với tính toán trường hợp xấu nhất cho tiếp xúc của trùng đất với Cry1F sau thu hoạch (Shanahan và cs., 2000; EA của USDA/APHIS, 2001).
2.5.7.6 Khả năng tác động lên động vật có vú
Các nghiên cứu độc tính được tiến hành để xác định độc tính của protein Cry1F chứng minh protein này có độc tính rất thấp đối với động vật có vú. Trong một nghiên cứu độc tính cấp tính đường miệng ở chuột, các ước tính cấp tính liều độc gây chết 50% (LD50) đưa vào bao tử là > 5.050 mg/kg Cry1F vi khuẩn. Liều lượng này cao gấp 12.190 lần so với ước tính 95% chế độ ăn uống của người có chứa protein Cry1F do tiêu thụ các loại thực phẩm có nguồn gốc từ ngô BĐG.
Trong một nghiên cứu điều kiện phòng thí nghiệm chứng minh protein Cry1F và PAT nhanh chóng và hoàn toàn bị phân hủy trong điều kiện mô phỏng dạ dày có sự hiện diện của pepsin. Điều này cho thấy nguy cơ gây ra các tác động bất lợi đến sức khỏe con người và động vật có vú dù tiếp xúc lâu dài hầu như không tồn tại. Một tìm kiếm cơ sở dữ liệu có liên quan chỉ ra rằng chuỗi axít amin của protein Cry1F và PAT không có tương đồng đáng kể nào với trình tự trong các độc tố hoặc protein gây dị ứng đã biết. Như vậy, Cry1F và PAT rất khó có thể gây nên một phản ứng dị ứng (Shanahan và cs., 2000; EA của USDA/APHIS, 2001; Meyer, 1999).
Nghiên cứu đánh giá hiệu quả dinh dưỡng đối với những con chuột Sprague-Dawley ăn thức ăn SX từ ngô TC1507 và chuột Đ/C cho ăn thức ăn SX từ ngô không biến đổi gen trong khoảng 90 ngày. Không có sự khác biệt đáng kể về mặt độc tính đã quan sát được trong hiệu suất dinh dưỡng, các dấu hiệu lâm sàng và hoạt động thần kinh, nhãn khoa, bệnh học lâm sàng (huyết học, hóa học lâm sàng, đông máu, và nước tiểu), trọng lượng cơ quan, và bệnh lý bất kỳ giữa hai nhóm điều trị. Các kết quả này chứng minh hạt ngô TC1507 là an toàn và tương đồng về dinh dưỡng như hạt ngô không biến đổi gen (MacKenzie và cs., 2007).

25
Trong một thử nghiệm chéo khác, 20 con bò Holstein cho con bú được chia thành 2 nhóm với 2 chế độ ăn uống và cho ăn chế độ ăn có chứa thức ăn ủ chua toàn bộ cây ngô cộng với hạt ngô TC1507 hoặc ngô Đ/C không BĐG. Lượng chất khô, mức độ sản xuất sữa hàng ngày, chất béo, protein, lactose, chất rắn không béo, và tổng số chất rắn trong sữa không khác biệt giữa 2 nhóm bò thí nghiệm. Nhìn chung, nhóm ăn thức ăn từ ngô TC1507 và nhóm Đ/C không khác biệt về các chỉ số tình trạng sức khỏe. Tóm lại, không có sự khác biệt trong sản xuất sữa, thành phần sữa, hay sức khỏe bò được xác định thông qua các phân tích lý hóa học trong máu, và phân tích huyết học giữa bò sữa cho ăn khẩu phần ăn chế biến từ ngô TC1507 và bò sữa cho ăn chế độ ăn uống Đ/C (Faust và cs., 2007).
Tóm lại: từ các dữ liệu thí nghiệm có sẵn liên quan tới protein Cry1F cùng với lịch sử sử dụng an toàn của các sản phẩm Bt nguồn gốc vi khuẩn Bacillus thuringiensis chi aizawai khẳng định sự an toàn của protein kiểm soát côn trùng Cry1F biểu hiện trong thực vật và an toàn của vật liệu di truyền cần thiết cho sản xuất của protein này trong tất cả các nguyên liệu thô.
Protein chỉ thị PAT giúp chịu thuốc diệt cỏ nhóm glufosinate ở ngô chuyển gen TC1507, không gây tác hại hoặc tác động tiêu cực đáng kể đến sinh vật không chủ đích, bao gồm cả sinh vật có lợi, dựa trên dữ liệu cung cấp trong đơn và phân tích của
Protein Cry1F chỉ độc đối với một số loài côn trùng chủ đích đặc thù thuộc bộ cánh vảy (Lepidoptera). Protein Cry1F được đánh giá là an toàn đối với môi trường và đa dạng sinh học.
Sở kiểm tra Sức khỏe Thực vật và Động vật (APHIS) cho dòng ngô chuyển gen đ ược miễn điều hoà trước đó thể hiện PAT. Các ADN mã hóa tổng hợp protein PAT không độc hại và PAT không tương đồng với các protein được biết là chất độc hại hay gây dị ứng (OECD, 1999).
APHIS đã quyết định áp dụng tình trạng miễn điều hòa theo Đi ều 7 CFR phần 340.6 đối với ngô event TC1507 và con cháu của nó cũng như việc canh tác tiếp theo của chúng ở Hoa Kỳ và vùng lãnh thổ thuộc Hoa Kỳ. Cũng không còn có những hạn chế quy định trong việc đăng ký thuốc trừ sâu của RPA cho canh tác của loại ngô này. APHIS đánh giá thông tin từ các tài liệu khoa học cũng như các dữ liệu đặc trưng của ngô event TC1507 và con cháu bắt. Sau quá trình phân tích một cách cẩn trọng, APHIS đưa ra các kết luận sau (EA của USDA/APHIS, 2001):
- Dòng ngô TC1507 thể hiện các đặc tính không gây bệnh cho thực vật. Mặc dù ADN từ các tác nhân gây độc được sử dụng trong chọn tạo giống, loại cây này không bị nhiễm độc bởi các vi sinh vật (Bt), các cây này cũng không là nguy cơ kích thích gây bệnh ở thực vật khác.

26
- Ngô event TC1507 không có khả năng trở thành cỏ dại. Ngô không phải là cỏ dại, và không có lý do đ ể tin rằng các gen được đưa vào có thể biến ngô thành dịch hại, cỏ dại.
- Phát tán gen do lai tạo giữa ngô event TC1507 với ngô hoang dại ở Mỹ và vùng lãnh thổ của nó là rất khó xảy ra. Nguy cơ phát tán gen từ dòng ngô TC1507 vào các loài hoang dã không có khả năng làm tăng nguy cơ trở thành cỏ dại của bất kỳ con cháu, cũng không tác động bất lợi đến đa dạng di truyền của bất kỳ thực vật có liên quan, tương tự như chuyển gen trong chọn tạo giống ngô lai truyền thống.
- Ngô event TC1507 tương đương đáng kể trong thành phần của toàn bộ cây và trong thành phần hạt, chất lượng và đặc điểm khác như đối với ngô không chuyển gen và không có tác động xấu đối với hàng nông sản thô hoặc chế biến.
- Ngô event TC1507 không có tác động xấu đáng kể đến sinh vật không chủ đích, bao gồm cả những sinh vật có lợi cho nông nghiệp, và nó không ảnh hưởng đến các loài bị đe dọa hoặc nguy cơ tuyệt chủng.
Các phân tích trên đây cho thấy kết quả đánh giá rủi ro về sự an toàn của ngô TC1507 đối với môi trường và đa dạng sinh học đã được tiến hành ở nhiều nước, trên nhiều vùng sinh thái khác nhau và đều cho thấy sự an toàn đối với môi trường và đa dạng sinh học.

27
3 XÁC ĐỊNH VẤN ĐỀ CẦN KHẢO NGHIỆM TẠI VIỆT NAM
3.1 Thông tin về sản xuất và sử dụng cây ngô
Tại Việt Nam ngô được trồng phổ biến tại các vùng chính bao gồm các khu vực: Trung du và miền núi phía Bắc; Tây nguyên; Bắc trung bộ và khu vực Duyên hải miền Trung; Đông Nam bộ; Đồng bằng sông Hồng; Đồng bằng sông Cửu Long. Diện tích trồng và tỉ lệ trồng ngô của mỗi vùng được thể hiện trong Hình 2-1. Những năm gần đây, diện tích trồng ngô lai tăng rất ấn tượng. Đến hết năm 2010, diện tích trồng ngô trên cả nước lên đến 1,1 triệu ha, năng suất bình quân 41 tạ/hecta. Tổng sản lượng đạt 4,6 triệu tấn trong đó diện tích trồng ngô lai chiếm hơn 90% tổng diện tích trồng ngô cả nước (nguồn thống kê bộ NN&PTNT,2010; Tổng cục Thống kê, 2010).
Năng suất bình quân ngô của Việt Nam hiện thấp hơn năng suất ngô trung bình của thế giới (51.6 tạ/hecta), Trung Quốc (55,5 tạ/hecta) và thua rất xa so với nước Mỹ (96,5 tạ/hecta). Thực tế là hàng năm Việt Nam vẫn phải nhập khẩu hơn 1 triệu tấn ngô làm thức ăn chăn gia súc (theo Vinanet số ngày 08/2/2012). Hiện nay, nhu cầu ngô sử dụng làm thức ăn cho chăn nuôi là rất lớn. Số liệu của tổng cục Thống kê, giá trị nhập khẩu ngô 12 tháng năm 2011 là 327 triệu USD, khiến cho giá thức ăn chăn nuôi trong nước luôn cao hơn khu vực từ 10-20%. Gía trị nhập khẩu thức ăn gia súc và nguyên liệu (đậu tương, ngô …) trong năm 2011 đạt tới 2.37 tỉ USD. Như vậy nhu cầu về ngô, đặc biệt là trong lĩnh v ực chế biến làm thức ăn chăn nuôi ở Việt Nam hiện nay rất vô cùng lớn.
Hình 3-1: Diện tích trồng ngô tại các khu vực trồng ngô chính của Việt Nam (1,000 ha, 2010-bộ NN&PTNT)

28
3.2 Tình hình sử dụng cây trồng BĐG tại Đông Nam Á.
Phillipines là quốc gia nằm trong khu vực Đông Nam Á, một trong 25 quốc gia mạnh dạn đưa cây trồng công nghệ sinh học vào ứng dụng từ năm 2003. Cho tới 2011, diện tích trồng cây biến đổi gen ở Phillipines đã là 644 nghìn héc ta, tăng 19% so với 2010. Thành tựu đáng kể nhất chính là trồng những giống ngô chuyển gen kháng sâu đục thân và kháng thuốc trừ cỏ. Chính phủ Phillipines đã phê duyệt khảo nghiệm hạn chế ngô biến đổi gen từ năm 1999 trên 600m2
+ Số lượng loài chân khớp (Athropods)
và đã khẳng định được tính kháng của loại ngô này đối với sâu đục thân ngô châu Á, năng su ất vượt trội hơn giống thường tới 60% và cao hơn 41% so với các giống ngô lai. Đến năm 2001, Chính phủ Phillipines đã phê duyệt khảo nghiệm trên diện rộng về hiệu quả sinh học của ngô biến đổi gen tại những vùng trồng ngô chính ở đất nước này. Từ năm 2003, nông dân Phillipines đã chính thức được phép trồng thương mại một số giống cây trồng áp dụng công nghệ sinh học trong đó chủ yếu là ngô chuyển gen. Theo Tiến sĩ Reynaldo V.Ebora: kinh nghiệm trồng ngô chuyển gen ở Phillipines chính là việc chú trọng vào đánh giá rủi ro đối với môi trường và vấn đề hậu thương mại. Các nghiên cứu tập trung vào một số đối tượng chính:
+ Côn trùng cánh vảy (Lepidoptera) + Bọ xít ăn thịt + Bọ rùa Các đối tượng này xuất hiện cả trên ngô thường , ngô lai và ngô chuyển gen tại
tất cả các vùng trồng ngô của Philippines. Kết quả thu được khá khả quan khi ng ô chuyển gen không ảnh hưởng tiêu cực đối với hệ sinh thái, không làm thay đổi thành phần các loài chân khớp, bọ rùa, bọ xít… trên ngô và các khu vực lân cận . Dịch sâu hại không tiếp tục phát sinh do trồng ngô Bt, không có tác động rõ đến các loài chim và các loài sâu hại không chủ đích trên mặt đất, giảm sự xói mòn đất, không có tác động đến độ màu mỡ của đất. Hiệu quả của ngô chuyển gen đến SX nông nghiệp ở Phillipines là: những trang trại trồng ngô biến đổi gen cho năng suất tăng 37%, dẫn đến tăng lợi nhuận khoảng 10.132 peso (4,5 triệu đồng)/hecta; giảm khoảng 60% chi phí thuốc trừ sâu; tăng thu nhập khoảng 1.34 peso/kg sản phẩm.
Bên cạnh Phillipines, một nước khác trong khu vực Đông Nam Á là Myanmar cũng đã chính thức cấp phép trồng thương mại cây trồng biến đổi gen từ 2006-2007 với cây trồng chính là bông biến đổi gen Bt. Tính tới 2011, diện tích trồng Bông biến đổi gen tại Myanmar đạt khoảng 238 nghìn héc ta, chiếm tới 79% tổng diện tích trồng bông của nước này, và tăng 3% so với năm 2010.

29
3.3 Cơ sở khoa học nghiên cứu tính an toàn đối với đa dạng sinh học và môi trường trong trồng ngô TC1507 trên thế giới
Cây trồng chuyển gen hiện đã và đang được nghiên cứu ở nhiều nước trên thế giới. Một số nước với trình độ Khoa học Công nghệ hiện đại đã tạo thành công nhiều giống cây trồng chuyển gen trên các đối tượng như: Bông, Đậu tương, Ngô vv. Các loại giống cây trồng này đã được đưa vào sử dụng ở nhiều nước nước phát triển (Mỹ, Nhật Bản, Ca na đa…) và ở một số nước đang phát triển (Ác hen ti na, Braxin, Me xi cô, Nam Phi, Philippine...).
Năm 2009 (Steven E. Naranjo), sản lượng từ cây trồng sử dụng công nghệ sinh học trong đó Đậu tương chiếm 51% sản lượng toàn cầu, Bôn g chiếm 32% và Ngô chiếm 17%. Cho tới năm 2011 cả thế giới đã có 29 nước trồng 160 triệu hecta cây trồng biến đổi gen (10% tổng diện tích đất trồng trên toàn thế giới), tăng 8% (30 triệu ha) so với năm 2010 (ISAAA Brief 43, 2011). Hơn phần nửa số quốc gia này là các nước đang phát triển, hơn 16.7 triệu nông dân trên thế giới trồng cây áp dụng công nghệ sinh học và gần 90% số này sống tại các quốc gia đang phát triển. Tính từ năm 1996 đến 2010, cây trồng biến đổi gen đã đem lại lợi ích kinh tế cho nông dân sử dụng (trồng) cây trồng chuyển gen trên thế giới ước tính đạt 78.4 tỉ đô la Mỹ, chủ yếu nhờ tăng sản lượng và giảm chi phí sản xuất. Góp phần giảm sử dụng tới 443 nghìn tấn thuốc trừ sâu và giúp giảm sử dụng đất nông nghiệp tới 91 triệu héc ta trên toàn thế giới. Mặc dù, lợi ích của công nghệ mới này đem lại tiềm năng lớn lao cho nền nông nghiệp (tạo ra nhiều lương thực hơn, sử dụng ít diện tích đất, giảm các chi phí về hóa chất trừ sâu) nhưng vấn đề về tính an toàn cho con người và môi trường, cũng như đa dạng si nh học vẫn luôn là vấn đề cần cẩn trọng (Michelle Marvier, 2002). Hiện nay, việc ứng dụng công nghệ mới đang ngày càng trở nên phổ biến và quan trọng đối với toàn cầu.
3.3.1 Nguy cơ trôi gen ngoài môi trường
Ngô tự nhiên được sinh sản trên cơ sở thụ phấn và nảy mầm từ hạt. Ngô TC1507 cũng có cơ chế sinh sản tương tự như ngô tự nhiên.
Kích cỡ hạt phấn ngô lớn (đường kính trung bình 0.1 mm), là một trong những loại hạt phấn có kích cỡ lớn nhất có thể được thụ phấn nhờ gió hạt phấn, do đó ngoài tự nhiên phấn ngô chỉ có khả năng thụ phấn trong khoảng cách khá hạn chế (2-3 m kể từ đầu râu ngô). Các kiểm tra ngoài thực tế cho thấy số lượng hạt phấn giảm mạnh từ 2.3x107 hạt/m2 trong vong bán kính 1 m giảm còn 7.1x103 hạt/m2 (Raynor et al.,

30
1972). Ngô cũng là loại cây thụ phấn cùng loài và tỉ lệ thụ phấn phụ thuộc nhiều vào các yếu tố như gió, ẩm độ, nhiệt độ. Chỉ có 2 loài ngô hoang có thể cho phép thụ phấn khác loài là Tripsacum sp. and Euchlanea sp., tuy nhiên các loài ngô này chỉ có ở Mê -xi-cô, Brasil và Bắc Mỹ (OECD, 2003). Trên thực tế, ngô đã được thuần hóa phục vụ canh tác từ hàng nghìn năm nay, sự nảy mầm của hạt ngô rất dễ bị ảnh hưởng bởi côn trùng, ẩm, nhiệt độ cũng như yêu cầu tính đồng nhất đất cao. Chính bởi tính thuần hóa cao như vậy, ngoà i tự nhiên ngô không thể tồn tại nếu thiếu bàn tay canh tác của con người (CFIA, 1994).
3.3.2 Nguy cơ trở thành cỏ dại, dich hại
Ngoại trừ một số ít nước như Mê-xi-cô, Brasil và Bắc Mỹ, các vùng địa lí khác đều không tồn tại bất cứ loài cỏ dại, ngô hoang nào có họ hàng gần với các giống ngô hiện đang được canh tác (OECD, 2003). Do vậy, mối lo ngại việc lan truyền các gen chuyển nạp cry1F, PAT ra các loài cỏ dại ngoài tự nhiên có thể hoàn toàn loại bỏ. Ngô cũng đã được thuần hóa tới mức hạt ngô không thể tự tách ra từ lõi nếu thiếu yếu tố con người. Ngô TC1507 ngoài đồng ruộng có thể dễ dàng phát hiện bằng cách phun thuốc trừ cỏ gốc glufosinate-ammonium, và sau đó có thể được loại bỏ bằng tay hay biện pháp cơ giới.
3.3.3 Nguy cơ ảnh hưởng tới sinh vật không chủ đích
Nguy cơ ngô TC1507 ảnh hưởng trực tiếp hay gián tiếp tới NTOs và hệ sinh thái đã được đánh giá và ghi nhận bởi Ủy ban An toàn Thực phẩm châu Âu (EFSA, 2005; EFSA, 2010). Chi tiết các thí nghiệm kiểm tra tiến hành trong điều kiện phòng thí nghiệm, nhà lưới cũng như ngoài đồng ruộng được liệt kê tại Bảng 3-1.
- Các loài thụ phấn: hạt phấn ngô có thể bám dính hoặc là thức ăn của một số loài thụ phấn như ong mật. các thí nghiệm sử dụng protein Cry1F làm thức ăn (Bảng 3-1) cho thấy protein này không ảnh hưởng đối vớ i nhộng hay độc đối với ong mật. Các thí nghiệm của Malone and Burgess (2009) sử dụng protein Cry1F cũng như hạt phấn ngô chuyển gen cry1F dung làm thức ăn cho ong mật với liều lượng từ thấp đến cao cũng đưa ra kết luận: không có giống ngô mang gen cry1F thương mại nào ở thời điểm thí nghiệm ảnh hưởng độc đối với ong mật. Kết quả từ 25 nghiên cứu độc lập trong việc kiểm tra ảnh hưởng trực tiếp ảnh hưởng của Cry1F đối với ong mật được tổng hợp trong báo cáo của (Duan và cs., 2008) cũng cho thấy không có bất cứ ảnh hưởng tiêu cực nào từ protein Cry1F đối với tỉ lệ sống sót của ấu trùng cũng như ong trưởng thành. Kết luận từ Ủy ban An toàn Thực Phẩm châu Âu (EFSA GMO Panel) đưa ra là: không

31
có bất cứ bằng chứng nào có thể dẫn tới nghi ngờ ngô TC1507 ảnh hưởng tiêu cực tới quần thể các loài côn trùng thụ phấn, mọi kết quả đều không có sự khác biệt giữa ngô TC1507 và các giống ngô truyền thống.
- Kí sinh và thiên địch: lí do là bởi có tới hàng ngàn loài chân khớp NTOs luôn xuất hiện trên đồng ruộng trồng ngô tại châu Âu, một số loài trong số đó là thiên địch và chúng có thể tiếp xúc trực tiếp/gián tiếp với protein Cry1F trên đồng ngô TC1507. Các loài kí sinh cũng có thể tiếp xuc trực tiếp với protein Cry1F từ phấn hoa, dich nhựa cây hoặc gián tiếp qua các sinh vậ t chủ của chúng. Các nghiên cứu đều không đưa đến bất cứ chứng cứ nào về ngô TC1507 ảnh hưởng tiêu cực đối với quần thể các loài kí sinh, thiên địch (EPA, 2005). Higging và cs. (2009) đã tiến hành thí nghiệm kéo dài trong 3 năm trên ngô TC1507 tại 4 vùng trồng ngô của Mỹ, và tiến hành theo dõi số lượng đáng kể các loài chân khớp không chủ đích bao gồm cả các loài kí sinh, thiên địch và côn trùng hại cỏ. Tiến hành đếm số cây cũng như theo dõi biến động số lượng quần thể từng loài chân khớp bằng cách sử dụng bẫy dính (sticky trap), bẫy hộp (litterbag), bẫy chụp (pitfall trap). Kết quả là không có sự khác biệt thành phần loài cũng như số lượng từng loài chân khớp khảo sát giữa khu trồng ngô TC1507 và ruộng trồng ngô đối chứng.
- Các loài chân khớp NTOs trên mặt đất: trên ngô ở Tây Ban Nha có 4 loài rệp muội hại ngô phổ biến gồm Rhopalosiphum padi, Sitobion avenae, Metopolophium dỉhodum, Macrosiphum euphorbiae. Kết quả điều tra cho thấy mật độ chung của các loài rệp muội trên giống ngô Bt (Compa CB) cao hơn so với trên giống ngô không chuyển gen (Dracma). Trong đó, mật độ loài rệp muội Sitobion avenae cao hơn đáng kể (Eizaguirre et al., 2006; Pons et al., 2005). Một nghiên cứu khác ở cả điều kiện phòng thí nghiệm và ngoài đồng ruộng cũng chỉ ra rằng giống ngô Bt có ảnh hưởng tích cực đối với sự gia tăng mật độ quần thể của rệp muội Rhopalosiphum padi (Lumbierres et al., 2004). Mật độ quần thể rệp muội R. padi trung bình là 2,67 và 1,92 con/3 cây tương ứng trên ngô Bt và ngô đối chứng, không có sự khác biệt ở mức ý nghĩa thống kê (Wold et al., 2001). Nghiên cứu của Rauschen và cs. (2004) lại ghi nhận không có sự khác biệt mật độ quần thể rệp hại lá ngô (Zyginidia scutellaris) trên ngô Bt và ngô không biến đổi gen.
- Vi sinh vật đất: protein cung cấp nguồn nitơ và các bon chính cho các vi sinh vật đất. Hầu hết các loại protein đều bị phân giải nhanh chóng bới các VSV có

32
chưa men protease (Jan et al., 2009), và không hè có bất cứ bằng chứng nào cho thấy protein Cry1F trong ngô TC1507 có những khác biệt với các protein trong tự nhiên (Icoz và Stotzky, 2008). Thậm chí nếu có protein Cry1F thoát ra từ rễ hay phân giải từ các bộ phận thân, lá ngô TC1507, thì chúng cũng sẽ bị phân giải bởi các vi sinh vật đất sau thời gian một đến và tuần. Protein cry được hấp thụ và giữ lại trên bề mặt các thành phần khoáng hữu cơ, hạt sét hay trong các hợp chất mùn, những cơ chế này chỉ có thể làm chậm quá trình chứ không hoàn toàn ngăn cản sự phân giải Cry1F của các VSV ( Crecchio và Stotzky, 1998, 2001; Pagel-Wieder và cs., 2007; Madliger và cs., 2011). Rất nhiều các nghiên cứu đều cho thấy quá trình phân giải protein Cry1F trong đất diến ra nhanh chóng và phổ biến. Nghiên cứu của Shan và cs., 2008 cho thấy sau 3 năm không còn phát hiện Cry1F protein trong đất canh tác ngô TC1507 (độ nhạy: 4.5ng/gr đất khô). Ủy ban An toàn Thực Phẩm châu Âu kết luận không có bất kì nguy hại cũng như không có ảnh hưởng trực tiếp nào từ protein Cry1F đối với môi trường đất (EFSA, 2011).
- Các loài chân khớp NTOs trong đất : các đối tượng như bọ đuôi bật (springtails) hoặc một số loài ve (mites) có đóng gớp vô cùng quan trong trong quá trình phân giải và tái tạo các tàn dư thực vật. Do đó các loài này có thể sử dụng như các loài chỉ thị về chất lượng và thành phần đất. Protein Cry1F sẽ bị phân hủy đáng kể sau một khoảng thời gian ngắn và không thể tích tụ trong đất, các sinh vật đất chỉ có thể tiếp xúc với hàm lượng protein Cry1F tương đối thấp trong các khu ruộng sau khi đã thu hoạch ngô TC1507. Nhìn chung, các nghiên cứu đều cho thấy không có bất cứ ảnh hưởng tiêu cực nào từ protein Cry1F đối với các loài nêu trên (Icoz and Stotzky, 2008).

33
Bảng 3-1: Nghiên cứu phòng TN, nhà lưới, ngoài đồng ruộng nhằm tìm hiểu ảnh hưởng của protein Cry1F đối với NTOs
Nhóm Loài Loại TN Vật liệu Nguồn
Loài thuộc nhóm chân khớp (Arthropods) Bọ đuôi bật Folsomia candida Phòng TN Protein Cry1F EPA, 2005; OECD, 2007 Sâu non bướm Danaus plexippus Phòng TN Protein Cry1F EPA, 2005; OECD, 2007 Bọ ladybird trưởng thành (Bắt mồi)
Hippodamia convergens
Phòng TN Protein Cry1F EPA, 2005; OECD, 2007
Sâu non lacewing (Bắt mồi)
Chrysoperla carnea Phòng TN Protein Cry1F EPA, 2005; OECD, 2007
Ong bắp cày (kí sinh) Nasonia vitripennis Phòng TN Protein Cry1F EPA, 2005; OECD, 2007 Sâu non ong mật (Thụ phấn)
Apis mellifera Phòng TN Protein Cry1F EPA, 2005; OECD, 2007 Đồng ruộng Hạt phấn chứa Cry1F
Rệp nước Daphnia magna Đồng ruộng Hạt phấn chứa Cry1F EPA, 2005; OECD, 2007 Loài không thuộc nhóm chân khớp
Giun đất trưởng thành Eisenia fetida Phòng TN Protein Cry1F EPA, 2005; OECD, 2007 Chim cút non Colinus virginianus Đồng ruộng Hạt phấn chứa Cry1F EPA, 2005; OECD, 2007 Chuột Phòng TN Protein Cry1F EPA, 2005; OECD, 2007
Gà broiler Đồng ruộng Hạt phấn chứa Cry1F EPA, 2005; OECD, 2007
Cá Rainbow trout Onchorhynchus mykiss
Đồng ruộng Hạt phấn chứa Cry1F EPA, 2005; OECD, 2007

34
3.4 Tiêu chí đánh giá trong khảo nghiệm đồng ruộng tại Việt Nam đối với ngô biến đổi gen event TC1507
Ở Việt Nam, Bộ NN&PTNT đã trú trọng đến việc nghiên cứu chuyển gen Bt vào ngô; chuyển gen H5N1
Trong trường hợp yêu cầu khảo nghiệm ngô Bt, Mỹ và Philippines là 2 nước được lựa chọn tham khảo xây dựng kế hoạch, qui trình khảo nghiệm cũng như các tiêu chí đánh giá bởi cả hai nước này đều đã có thời gian dài thương mại hóa và cho phép trồng loại ngô này với diện tích lớn. Mỹ là nước đặc biệt phát triển với một nền khoa học công nghệ tiên tiến; chất lượng cuộc sống, các tiêu chí an toàn sức khỏe, môi trường luôn được chú trọng; các nghiên cứu khoa học chuyên sâu được tiến hành thường xuyên nhằm đáp ứng các đòi hỏi, giải quyết các vấn đề phát sinh trong cuộc sống. Qui trình khảo nghiệm của họ được hầu hết các nước đã và đang ứng dụng cây trồng BĐG trên thế giới tham khảo, học hỏi. Philippines cũng là một nước đã có lịch sử 10 năm canh tác ngô Bt và họ đã thu được thành quả đáng kể từ ứng dụng này. Philippines lại là nước cùng trong khu vực Đông Nam Á, các điều kiện địa lí, sinh
vào Bèo tấm nhằm làm thức ăn cho gia súc, gia cầm; chuyển gen chống sâu xanh vào cây Bông vv... Kết quả là khả quan trong tương lai gần. Hiện tại, Việt Nam đã và đang tiếp tục nghiên cứu nhằm tạo ra giống cây trồng chuyển gen như lúa có hàm lượng vitamin A cao; ngô kháng thuốc trừ cỏ, kháng sâu; đậu tương kháng sâu, chịu hạn; xoan tăng chất lượng gỗ; đu đủ kháng vi rút gây bệnh đốm vòng; bông kháng sâu, chịu hạn vv... Mặc dù lợi ích của công nghệ chuyển gen đem lại tiềm năng lớn lao cho nền nông nghiệp (tạo ra nhiều lương thực hơn, giảm diện tích đất sử dụng, giảm các chi phí về thuốc hóa chất trừ sâu vv…) nhưng vấn đề về tính an toàn cho con người và môi trường, cũng như đa dạng sinh học vẫn c ần được thận trọng xem xét. Trong điều kiện Việt nam, việc ứng dụng cây trồng chuyển gen vào thực tế sản xuất cũng gặp phải những vướng mắc cần giải quyết như: chúng ta đi sau các nước khác một thời gian dài; thiếu kinh nghiệm trong nghiên cứu chuyên sâu và quản lý điều hành; đầu tư thấp. Để khắc phục những hạn chế này, cần tiếp thu một cách có chọn lọc các thành tựu khoa học, công nghệ của thế giới và nhanh chóng ứng dụng vào Việt Nam. Một trong những ứng dụng đó là chính phủ, bộ nghành có liên quan cần sớm cho phép canh tác có chọ n lọc một số giống cây trồng BĐG như ngô Bt, bông Bt kháng sâu, ngô kháng thuốc trừ cỏ tại Việt Nam. Các yêu cầu về qui trình khảo nghiệm, các chỉ tiêu đánh giá, theo dõi cần được xây dựng, chọn lựa một cách khoa học, trên cơ sở tham khảo từ các nước đã có lịch sử trồng những giống cây trồng BĐG này, tính tương đồng về điều kiện sinh thái, trình độ và tập quán canh tác vv…nhằm rút ngắn thời gian, chi phí khảo nghiệm nhưng vẫn đảm bảo tính chính xác, an toàn đối với đa dạng sinh học và môi trường Việt Nam.

35
thái, môi trường cũng như tập quán canh tác của người nông dân khá tương đồng với Việt Nam chúng ta. Ở cả hai nước này đều đòi hỏi phải tiến hành KNHC và KNDR với những qui trình cụ thể nhằm đánh giá, hạn chế rủi ro và đảm bảo ATSH, đa dạng quần thể sinh vật trong khu hệ sinh thái.
Tham khảo qui trình khảo nghiệm từ hai nước nói trên là Mỹ và Philippines , trong khuôn khổ khảo nghiệm đồng ruộng nhằm đánh giá rủi ro ngô BĐG event TC1507 đối với đa dạng sinh học và môi trường sinh thái Việt Nam, hạn chế lặp lại các đánh giá mà kết quả đã được công bố rõ rõ ràng trong khi các chỉ tiêu/ đối tượng theo dõi là không hay rất ít phụ thuộc bởi yếu tố điều kiện sinh thái vùng. Ví dụ các chỉ tiêu đo hàm lượng protein Cry1F trong hạt (chủ yếu phụ thuộc yếu tố giống); thử độc tính cấp tính (acute toxicity), độc tính lâu dài (chronic toxicity) của protein Cry1F; hay y/c kiểm tra tốc độ phân hủy, dư lượng Cry1F trong đất cũng như ảnh hưởng của loại protein này đối với hệ VSV đất là không cần thiết vì các lí luận khoa học cũng như kết quả công bố (mục 3.3.3) là đã rõ ràng và hoàn toàn tin cậy. Bằng việc tuân thủ chặt chẽ các qui định tại Nghị định số 69/2010/NĐ-CP năm 2010 của Chính phủ; dưới sự hướng dẫn và giám sát của Ủy ban ATSH ngành, nội dung và các chỉ tiêu theo dõi được lựa chọn cho mỗi KNHC và KNDR cũng như phương pháp nghiên cứu, theo dõi trong khảo nghiệm đồng ruông đối với ngô event TC1507 được trình bày chi tiết tại mục 4 của báo cáo này.

36
4 MỤC TIÊU, NỘI DUNG, VẬT LIỆU VÀ P.P KHẢO NGHIỆM
4.1 Mục tiêu và nội dung khảo nghiệm
4.1.1 Mục tiêu
Cây trồng chuyển gen là sản phẩm của nền kỹ thuật sinh học hiện đại (kỹ thuật DNA tái tổ hợp). Bằng kỹ thuật này con người có thể tạo ra các giống cây trồng có tính trạng mong muốn mà bằng kỹ thuật tạo giống truyền thống rất khó hoặc không thể tạo nên những giống cây trồng có đặc tính tương tự. Tiến bộ của công nghệ mới, hiện đại đã tạo ra những triển vọng mới đối với nông nghiệp và an ninh lương thực. Song, đây cũng được xem là “vấn đề” hiện gây tranh cãi trên phạm vi toàn cầu . Điều quan trọng là không phải sử dụng đến các loại hóa chất bảo vệ thực vật và năng suất cây trồng tăng đáng kể. Bên cạnh đó, những nghiên cứu về ảnh hưởng của cây trồng chuyển gen đến môi trường và đa dạng sinh học cũng luôn được quan tâm nghiên cứu nhiều hơn (Dale P. và cs., 2002; Harwood J. và cs., 2005; Rosel R. và cs., 2007; Honemamm L. và cs., 2008; Frouz J. và cs., 2008; Powell R. và cs., 2009; Priestley AL. và cs., 2009; Bai Y. và cs., 2010). Ở Việt Nam, hiểu biết về cây trồng chuyển gen còn khá mới mẻ. Nhưng Việt nam là nước mà nền nông nghiệp chiếm 35 - 40% GDP và có tới 75% dân số làm nông nghiệp. Hơn nữa, giá lương thực, thực phẩm ngày càng tăng cao, năng suất cây trồng dần bị hạn chế do nhiều nguyên nhân như biến đổi khí hậu, mất cân bằng sinh thái, dịch bệnh bùng phát liên tục, sử dụng thuốc trừ sâu tràn lan vv... Vì vậy, việ c áp dụng CNSH và sử dụ ng giống cây trồng BĐG, thực phẩm chuyển gen là việc hết sức quan trọng và đáng phải quan tâm. Về mặt chiến lược phát triển KHCN của đất nước, Việt Nam không thể không tiếp cận với công nghệ sinh học phân tử và kỹ thuật chuyển gen nhưng đòi hỏi phải tuân thủ chặt chẽ theo đúng quy trình mà nhà nước Việt Nam ban hành.
Để khẳng định có hay không có tác động của cây trồng chuyển gen đến chất
lượng môi trường đất, đa dạng sinh học và môi trường Việt Nam, bước đầu chính phủ Việt Nam đã cho phép tiến hành trồng khảo nghiệm cây BĐG mà ban đầu là cây Ngô. Xuất phát từ những yêu cầu thực tế trên, được phép của Bộ Nông nghiệp và Phát triển Nông thôn Viện Di truyền Nông nghiệp kết hợp với C.ty TNHH Pioneer - Hi Bred Việt Nam tiến hành khảo nghiệm: “Tác động của ngô chuyển gen event TC1507 đối với đa đạng sinh học và môi trường sinh thái Việt Nam” qua 2 vụ liên tiếp (KNHC) sau đó khảo nghiệm tại 4 vùng sinh thái đại diện 4 vùng trồng ngô chính tại Việt Nam (KNDR)

37
Mục đích: Thu thập số liệu từ thực tế ngoài tự nhiên đồng ruộng nhằm tìm hiểu, chứng minh có hay không có ảnh hưởng của canh tác ngô TC1507 (mang gen cry1F kháng côn trùng cánh vảy) đối với đa dạng sinh học và môi trường trong điều kiện sinh thái Việt Nam.
4.1.2 Nội dung khảo nghiệm
4.1.2.1 Nội dung nghiên cứu trong KNHC
• Đánh giá mức độ ảnh hưởng tới sự đa dạng quần thể NTOs trên thực tế đồng ruộng canh tác ngô TC1507 và Đ/C
• Điều tra thành phần các loài chân khớp (côn trùng và nhện) • Điều tra mật độ các loài chân khớp phổ biến • Đánh giá mức độ đa dạng của sinh vật đất • Đánh giá mức xuất hiện và độ gây hại của một số bệnh, sâu hại ngô NTOs)
trên ruộng canh tác ngô TC1507 và Đ/C
4.1.2.2 Nội dung nghiên cứu trong KNDR
Trên cơ sở các kết quả nghiên cứu, thực tế canh tác và sử dụng sản phẩm ngô Bt, và kết quả khảo nghiệm hạn chế (KNHC) ngô chuyển gen event TC1507 hai vụ liên tiếp tại Văn Giang, Hưng Yên cho thấy: trong điều kiện môi trường Việt Nam cần thiết một lần nữa đánh giá trên diện rộng (tại các vùng sinh thái trồng ngô chính của Việt Nam) để làm rõ thêm ảnh hưởng/tác động nếu có của việc trồng ngô event TC1507 đối với:
+ Các loài bệnh và sâu hại chính không chủ đích + Các loài kí sinh và thiên địch + Các loài sinh vật đất và trong lớp đất mặt + Đánh giá hiệu quả sinh học của ngô TC1507 trong điều kiện áp lực sâu tự
nhiên của Việt Nam.
KNDR được tiến hành theo 3 nội dung chính là:
(1) Kiểm tra sự hiện diện của protein Cry1F (2) Đánh giá tính ổn định và tính thí ch ứng của ngô event TC1507 so với đ ối
chứng tại mỗi vùng sinh thái Việt Nam.
+ Đánh giá các đặc tính nông sinh học và mức độ mẫn cảm với các loại bệnh hại chủ yếu của ngô event TC1507 so với giống đối chứng.

38
+ Đánh giá hiệu quả kháng sâu hại chủ đích (Lepidoptera) của ngô TC1507 ở điều kiện áp lực sâu tự nhiên và tập quán canh tác địa phương tại từng vùng sinh thái Việt Nam
(3) Đánh giá mức độ ảnh hưởng/tác động đối của việc trồng khảo nghiệm ngô event TC1507 với đa dạng quần thể NTOs trong điều kiện vùng sinh thái Việt Nam
+ Đa dạng thành phần loài của quần thể côn trùng NTOs thuộc nhóm ăn thực vật (nhóm chích hút)
+
+ Đa dạng thành phần loài và sự phân bố của quần thể côn trùng thuộc nhóm thụ phấn.
Đa dạng thành phần loài và sự phân bố của quần thể côn trùng NTOs nhóm bắt mồi ăn thịt
+ Đa dạng thành phần loài của bọ đuôi bật (Collembola).
4.2 Vật liệu và phương pháp nghiên cứu trong KNHC
4.2.1 Giống Ngô đăng ký KNHC
- Giống 30Y87H - ngô event TC1507 (mang gen cry1F kháng côn trùng cánh vảy) - Giống 30B80 - đối chứng, không chuyển gen - Giống 30Y87 - giống nền đối chứng, không chuyển gen
4.2.2 Thời gian, địa điểm KNHC
4.2.2.1 Thời gian KNHC ngô event TC1507
Bảng 4-1: Thời gian tiến hành KNHC
Thời gian Ngày gieo Ngày trồng Ngày thu hoạch
Vụ 1 31/7/2010 - 31/10/2010 Vụ 2 27/10/2010
(gieo trong bầu) 3/11/2010 24/3/2011

39
4.2.2.2 Địa điểm KNHC ngô event TC1507
Khảo nghiệm hạn chế được tiến hành trong hai vụ liên tiếp ở cùng một địa điểm tại Trạm thực nghiệm Văn Giang, huyện Văn Giang, tỉnh Hưng Yên.
4.2.3 Phương pháp KNHC (áp dụng cho cả hai vụ)
4.2.3.1 Sơ đồ bố trí thí nghiệm
- Diện tích mỗi công thức là 127,4m2
- Thí nghiệm được bố trí theo khối ngẫu nhiên hoàn toàn (RCBD) với ba (03) lần nhắc lại, mỗi lần nhắc lại gồm ba (03) công thức (Hình 4-2; 4-3). Mỗi công thức thí nghiệm được trồng 13 hàng dài 14m, khoảng cách hàng là 0,7 m, khoảng cách cây là 0,25m. Các công thức khảo nghiệm được thiết kế như sau:
(9,1m x 14m); canh tác theo tập quán ở địa phương.
CT1: 30B80, ngô Đ/C, không phòng trừ sâu, làm cỏ bằng tay. CT2: 30Y87, ngô nền Đ/C, không phòng trừ sâu, làm cỏ bằng tay. CT3: 30Y87H, ngô chuyển gen, không phòng trừ sâu, làm cỏ bằng tay.

40
Hình 4-1: Sơ đồ bố trí KNHC vụ 1
Hình 4-2:Sơ đồ bố trí KNHC vụ 2

41
4.2.3.2 Phương pháp kiểm tra sự hiện diện của protein Cry1F
Sử dụng mẫu lá ngô thu tại ruộng khảo nghiệm theo các công thức khảo nghiệm. Các dụng cụ và hóa chất dùng trong thí nghiệm được mô tả trong Hình 4-1.
Hình 4-3: QuickStix™ Kit dùng phát hiện nhanh protein Cry1F
Ghi chú: 1. Dung dịch đệm QuickStix EB2; 2. Lọ nhỏ giọt 3. Eppendorf 1,5ml; 4. Chày nhựa 5. Que thử nhanh; 6. Hộp đựng que thử nhanh Cách thức thu mẫu và phương thức tiến hành
Tại thời điểm tỉa thưa, xác định sự hiện diện của protein Cry1F tại các ô khảo nghiệm tương ứng nhằm xác định tính chính xác trong bố trí thí nghiệm cũng như tính đồng nhất của hạt giống bằng việc sử dụng QuickStix™ Kit cho cry1F. Thu mẫu lá từ 4 cây ngẫu nhiên tại hai hàng phía trong của mỗi lô thí nghiệm (xem Hình 4-2; 4-3), nghiền nhỏ trong ống eppendorf chứa dung dịch tách chiết DNA. Sử dụng que thử nhanh đối với gen cry1F để xác định sự hiện diện của protein Cry1F. Giai đoạn trước thu hoạch, nếu cây có biểu hiện bị hại bởi sâu đục thân và sâu đục bắp xuất hiện ở các công thức thí nghiệm trồng ngô CT1507, những cây này sẽ được kiểm tra lại bằng que thử của QuickStix™ Kit cho Cry1F.
1 2 3 4 5 6

42
4.2.3.3 Phương pháp đánh giá đa dạng quần thể NTOs
Tiến hành điều tra bằng mắt thường và bẫy dính vàng từ giai đoạn V8-V10, trước khi thụ phấn (VT), thời điểm thụ phấn (R1, khoảng 50% tung phấn), và sau thụ phấn (R2 và R3 hoặc khoảng 2-3 tuần sau 100% tung phấn). Các điểm lấy mẫu được gắn thẻ, đánh số, cắm cột cờ hay ruy-băng để dễ nhận biết cho các lần lấy mẫu sau.
Bảng 4-2:Thời gian điều tra thành phần quần thể NTO tại ruộng KNHC
Đợt điều tra Thời gian điều tra
GĐST Vụ 1 Vụ 2 1 1/9/2010 22/12/2010 Cây có 8 lá (V8) 2 16/9/2010 30/1/2011 Vt 3 28/9/2010 22/2/2011 R1 4 5/10/2010 4/3/2011 R2 5 12/10/2010 15/3/2011 R3 6 22/10/2010 20/3/2011 R6
Ghi chú: Đợt điều tra thứ 6 chỉ điều tra sâu cắn gié (sâu cắn lá ngô) */Giai đoạn tăng trưởng của ngô (Nguồn: How a Corn Plant Develops. Special Report No.48. Iowa State Univ. Cooperative Extension Service. Ames, IA. Jan. 1997.)
V2 - Giai đoạn khi nhìn thấy được cuống lá của lá thứ 2. V4 - Giai đoạn khi nhìn thấy được cuống lá của lá thứ 4.
V6 - Giai đoạn khi nhìn thấy được cuống lá của lá thứ 6. V8 - Giai đoạn khi nhìn thấy được cuống lá của lá thứ 8. V9 - Giai đoạn khi nhìn thấy được cuống lá của lá thứ 9. VT - Giai đoạn khi hoàn toàn có thể nhìn thấy các nhánh cuối cùng của cờ ngô. R1 "Silking" - Giai đoạn khi nhìn thấy được râu ngô (phun râu). R2 "Blister" - Giai đoạn khi hạt có màu trắng bên ngoài và có dạng phồng lên.
Giai đoạn này có thể xảy ra 10-14 ngày sau khi phun râu. R4 "Dough" - Giai đoạn khi các vật chất trong hạt tạo ra ở dạng sáp. Giai đoạn
này có thể xảy ra sớm nhất là 24 ngày sau khi thụ phấn. R6 "Trưởng thành" - Giai đoạn khi thu hoạch. Đây được coi là sự trưởng thành
sinh lý.

43
a) Phương pháp điều tra bằng mắt thường
Điều tra bằng mắt thường được tiến hành theo phương pháp của Viện Bảo vệ thực vật (theo tiêu chuẩn ngành). Cụ thể, trên mỗi ô thí nghiệm chọn các cây ở giữa ô. Các cây điều tra phải nằm gần trung tâm ô khảo nghiệm để trá nh các tác động rìa ngoài (Hình 4-2; 4-3). Điểm lấy mẫu được xác định và ghi vào trong sổ theo dõi khảo nghiệm. Cây điều tra được chọn ngẫu nhiên lần đầu, ghi số treo biển để điều tra các lần tiếp theo cho đến khi kết thúc khảo nghiệm. Năm cây ngô điều tra sẽ được coi là một mẫu đại diện. Tại một điểm quan sát, 3 mẫu đại diện được lấy sao cho các cây không liên tiếp trên 1 hàng sẽ được lựa chọn. Có thể bỏ qua 2-3 cây ở giữa để tránh động đến các cây gần đó trong quá trình đánh giá. Việc quan sát ở mỗi cây được thực hiện như sau: trước tiên, bằng mắt quan sát toàn bộ cây trong mẫu đại diện để ghi nhận số lượng những côn trùng trưởng thành (dễ bay); sau đó, từng cây sẽ được quan sát cẩn thận trên cả hai mặt của lá, vùng bẹ lá, thân cây, bắp ngô và cờ ngô (khi có). Trình tự quan sát tương tự sẽ được thực hiện trên các cây trong mẫu đại diện. Việc quan sát này sẽ được lặp lại tại 3 điểm quan sát cho mỗi ô thí nghiệm. Số lượng cá thể ở từng pha phát triển của mỗi loài chủ yếu (hay đơn vị phân loại chính được quan tâm) được ghi lại cho từng cây điều tra. Các loài chân khớp khác có liên quan cũng được quan sát và ghi lại. Trường hợp khi số lượng cá thể của loài có kích thước cơ thể nhỏ (như rệp muội, nhện nhỏ) quá nhiều, mật độ của chúng có thể được ước tính hoặc phân cấp theo thang cấp quy định chung. Loài ký sinh và trứng bị ký sinh cũng sẽ được ghi lại. Mẫu vật côn trùng các loài chân khớp được đưa về phòng thí nghiệm để giám định tên khoa học. Chỉ tiêu theo dõi: thành phần loài và tần xuất bắt gặp.
Hình 4-4: Điểm lấy mẫu trong đánh giá bằng mắt thường
Các cây được quan sát, lấy mẫu
Điểm quan sát, lấy mẫu
Lô thí nghiệm
Khu vực lấy mẫu

44
b) Phương pháp điều tra bằng bẫy dính vàng
Điều tra bằng bẫy dính vàng theo phương pháp của C.ty Pioneer Hi-Bred International. Cụ thể, 3 bẫy dính (Pherocon AM No Bait traps, Trece Inc., Salinas, USA) được đặt gần trung tâm của mỗi ô thí nghiệm ( Hình 4-2; 4-3). Bẫy dính vàng được gỡ bỏ vào khoảng 7 ngày sau đặt . Thời gian đặt bẫy tùy thuộc vào từng địa điểm, được xác định và ghi vào trong sổ theo dõi khảo nghiệm. Thời gian đặt bẫy dính vàng cho mỗi lần lấy mẫu là giống nhau cho tất cả các công thức.
Mỗi bẫy dính vàng sẽ được gắn với một cột cắm trong đất và đặt giữa các hàng (ảnh Phụ lục 3). Các cột đủ cao (khoảng 1,5 - 2 m so với mặt đất) để có thể điều chỉnh chiều cao bẫy. Các bẫy được đặt cao đến tán lá trong giai đoạn V, và cao ngang bắp khi cây có bắp. Bẫy dính vàng khi thu từ ruộng thí nghiệm được bỏ vào túi zipper và dán nhãn rõ ràng để nhận dạng. Các cột sẽ vẫn để tại chỗ trong thời gian theo dõi thí nghiệm và được gỡ bỏ sau ngày cuối cùng thu bẫy dính vàng. Các bẫy dính vàng thu từ ruộng thí nghiệm về có thể giữ trong tủ lạnh để đánh giá sau. Đếm và ghi lại tất cả các loài chân khớp cần quan tâm dựa trên tính đa dạng của khu hệ chân khớp ở khu vực thí nghiệm. Có thể tiến hành theo dõi bổ sung bằng mắt và bẫy dính vàng khi có sự phát sinh rộ của các loài côn trùng bắt mồi phổ biến. Điểm lấy mẫu bổ sung được ghi vào sổ theo dõi khảo nghiệm.
4.2.3.4 Phương pháp theo dõi mật độ các loài chân khớp phổ biến
Điều tra bằng mắt thường và bẫy dính vàng. Việc điều tra sẽ được tiến hành vào giai đoạn giữa (khoảng V8-V10), trước khi thụ phấn (khoảng VT), trong quá trình thụ phấn (R1, khoảng 50% tung phấn) và sau thụ phấn (R2 và R3 hoặc khoảng 2-3 tuần sau khi 100% tung phấn). Các điểm lấy mẫu sẽ được gắn thẻ và đánh số, cắm cột cờ hay ruy-băng để dễ nhận biết ở các ngày tiếp theo lấy mẫu. Chỉ tiêu theo dõi: mật độ các loài được điều tra cần quan tâm, tỷ lệ nhiễm và chỉ số nhiễm đối với rệp muội.
- Loài có kích thước lớn: đếm số lượng cá thể bắt gặp trên cây điều tra. - Loài có kích thước nhỏ, mật độ dày đặc (rệp muội ngô): đánh giá theo cấp
nhiễm: Cấp 0: không có Cấp 1: rệp muội phân bố từng cá thể rải rác, chưa hình thành quần tụ Cấp 2: rệp muội bắt đầu hình thành một số quần tụ nhỏ Cấp 3: rệp muội có quần tụ nhỏ, bắt đầu hình thành một số quần tụ lớn Cấp 4: rệp muội có nhiều quần tụ lớn Cấp 5: rệp muội có nhiều quần tụ lớn liên kết liền lại thành vùng lớn

45
4.2.3.5 Phương pháp đánh giá đa dạng của quần thể bọ đuôi bật (Collembola)
Phơi nhiễm của bọ đuôi bật (Collembola) với protein Bt là kết quả của việc ăn các phụ phẩm cây trồng như rễ cây, thân lá cây, các hạt phấn rơi xuống đất trong quá trình thụ phấn… (Saxena và cs., 1999; Saxena và cs., 2000). Do đó, các loài bọ đuôi bật Collembola ăn các phần thực vật trên mặt đất được thu thập bằng phương pháp đặt bẫy cốc (Pitfall trap)
4.2.3.5.1 Thu mẫu bọ đuôi bật bằng phương pháp bẫy cốc (Pitfall trap).
Mô tả bẫy cốc: Bẫy cốc gồm 3 bộ phận: nón che mưa, cốc đựng dung dịch và ống trụ cố định (ảnh Phụ lục 3). Nón che mưa: Làm từ tấm mica trong suốt có tác dụng che mưa cho bẫy. Nón có đường kính 12 cm, cao 16-17 cm, được cố định vào nền đất bằng 2 thanh tre 2 bên, cao hơn miệng cốc khoảng 2 cm, trùm ra bên ngoài cốc. Cốc đựng dung dịch: bằng nhựa trong, cứng, đường kính 8 cm, cao 10 cm. Cốc chứa 50 ml dung dịch gồm Ethylene glycol + 95% Ethanol + nước theo tỷ lệ 2:2:1. Cốc được đặt lọt vào bên trong một ống trụ (miệng cốc thấp hơn miệng ống trụ khoảng 3-4 cm), ống này được cố định vào đất. Ống trụ cố định làm từ nhựa cứng, rỗng, gồm một ống nhựa, đườn kính 7,6 cm, dài 8 cm lồng sâu 4 cm vào một măng xông dài 8-9 cm, tạo thành 1 ống trụ dài 12-13 cm.
Thao tác đặt bẫy : Trước tiên chôn ống trụ xuống đất sao cho miệng ống bằng với mặt đất (là vị trí chọn để đặt bẫy cốc). Sau đó cho cốc đựng 50 ml dung dịch bảo quản vào trong ống trụ. Cuối cùng cố định nón che cách phía trên miệng cốc 2 cm bằng 2 thanh tre cắm vào đất.
Vị trí đặt bẫy: Tổng cộng có 3 bẫy pitfall mỗi lô được thiết lập gần trung tâm của từng lô (một bẫy cho mỗi điểm lấy mẫu) ở các giai đoạn sinh trưởng phát triển khác nhau của cây ngô: cây có lá thứ 10-15 (V10-V15); cây có lá thứ 18 đến trổ cờ (V18-VT); cây đóng bắp lụa non (R1); Bắp với các hạt phồng lên, đỉnh hạt nhọn (R2); Bắp với các hạt có sữa non đến hạt có bột, đỉnh hạt phẳng, từ màu trắng đến vàng (R3-R4). Sau 5 ngày đặt sẽ tiến hành thu bẫy (chỉ thu dung dịch đựng trong cốc của bẫy) và dồn 3 cốc của 1 lô TN vào 1 cốc (đây được tính là một mẫu định lượng khi phân tích và xử lý mẫu sau này). Dung dịch của 1 cốc duy nhất này được cho vào hộp nhựa hoặc túi ni lông dày, nút kín cùng với nhãn ghi đầy đủ địa điểm, kí hiệu lô TN, ngày đặt, số lần nhắc lại và được chuyển về phòng TN vào thời gian nhanh nhất có thể. Các cốc sau khi đã trút hết dung dich, được làm sạch và bảo quản sử dụng cho lần sau. Nón che và ống trụ để cố định trong đất, sử dụng cho cả 5 lần đặt mẫu. Như vậy,

46
khi đặt bẫy cốc, số lượng bẫy đặt là: 4 công thức x 3 lần nhắc x 3 bẫy cốc = 36 bẫy cốc. Số lượng bẫy cốc thu được là: 4 công thức x 3 lần nhắc x 1 bẫy cốc = 12 bẫy cốc
Tại phòng thí nghiệm: lọ mẫu được đổ vào phễu lọc sử dụng tờ giấy lọc φ15 để lưu lại mẫu động vật trên tờ giấy lọc. Phần dung dịch chứa mẫu sẽ qua phễu lọc xuống cốc gắn phía dưới phễu. Sau khi lọc xong, mẫu động vật lưu lại trên giấy lọc sẽ được tách riêng các nhóm động vật, tính đếm số lượng cá thể của riêng từng loài với các dụng cụ chuyên dụng, loài được xác định tên khoa học bằng các tài liệu chuyên môn.
Bảng 4-3: Thời gian thu mẫu Collembola tại ruộng KNHC
Đợt thu mẫu
phương pháp thực hiện
Thời gian GĐPT Vụ 1 Vụ 2
Đợt 1
Bẫy cốc 5/9/2010 24/12/2010 Cây có lá thứ 10-15 (V10-V15) Lấy mẫu đất 10/9/2010 29/12/2010
Đợt 2
Bẫy cốc 20/9/2010 10/2/2011 Cây có lá thứ 18 – trỗ cờ (V18-VT) Lấy mẫu đất 25/9/2010 15/2/2011
Đợt 3
Bẫy cốc 2/10/2010 19/2/2011 Cây đóng bắp lụa non (R1) Lấy mẫu đất 6/10/2010 24/2/2011
Đợt 4
Bẫy cốc 11/10/2010 28/2/2011 Bắp với các hạt phồng lên, đỉnh hạt nhọn (R2) Lấy mẫu đất 15/10/2010 3/3/2011
Đợt 5
Bẫy cốc 22/10/2010 7/3/2011 Bắp có hạt sữa - hạt có bột, đỉnh hạt phẳng, từ màu trắng
đến vàng (R3-R4) Lấy mẫu đất 26/10/2010 12/2/2011
4.2.3.5.2 Thu mẫu bọ đuôi bật bằng phương pháp thu mẫu đất (soil core).
Các loài bọ đuôi bật Collembola ăn rễ thực vật được thu thập bằng phương pháp thu mẫu đất (soil core). Theo Ghilarov (1975), số lượng chân khớp bé trong đất đạt từ vài nghìn đến vài chục nghìn cá thể trên một mét vuông. Vì vậy, với một số lượng mẫu ít, khối lượng nhỏ của mẫu đất vẫn có thể đạt được kết quả với độ chính xác đầy đủ và ở thực địa tùy thuộc mục đích nghiên cứu và ở sinh cảnh cụ thể trong một đợt điều tra, chỉ cần thu từ 4-10 mẫu, có khối lượng 125cm3
Tại mỗi lô thí nghiệm: lấy 2 mẫu đất ở lớp bề mặt 0 - 10 cm (kích thước 5x5x10cm trên bề mặt luống, cách gốc cây khoảng 2-4cm) bằng dụng cụ chuyên dụng
là đủ. Trong nghiên cứu này chúng tôi thu mẫu đất định lượng Collembola.

47
(ảnh Phụ lục 3) và thu nhắc lại 3 lần cho một đợt thu mẫu. Như vậy, mỗi lần lấy mẫu sẽ thu tổng số mẫu là: 2 mẫu đ ất x 3 công thức x 3 lần lặp = 18 mẫu. Mẫu đất được cho vào túi nilon kèm theo nhãn (ghi đủ thông tin về địa điểm, ngày thu, ký hiệu ngô, lần nhắc thứ mấy,…) và buộc chặt, chuyển về phòng thí nghiệm.
Tại phòng thí nghiệm: Collembola và các động vậ t không xương sống khác được tách ra khỏi đất bằng phễu Tullgren trong thời gian 7 ngày /đêm ở nhiệt độ phòng thí nghiệm. Phương pháp này cho phép thu thập được số lượng cá thể lớn hơn nhiều khi thu bằng tay và cũng cho phép tập hợp thông tin về mật độ nhiề u hơn (Palacios-Vargas, 1992). Phễu Tullgren có cấu tạo bằng bìa carton cứng cuốn thành cái phễu với đường kính φ20cm, độ cao 15cm -18cm. Ở đáy phễu có gắn một ống nghiệm có đường kính φ1,0cm, dài 4cm bên trong đựng dung dịch định hình cồn 90 o. Mẫu đất mang từ thực địa về, được chuyển vào một cái rây lọc (đường kính rây 18cm, thành rây bằng sắt, cao 5cm, đáy rây là lưới ni lôn với kích thước lỗ lưới 1,5mmx1,5mm). Đặt rây này bên trong phễu carton và đặt trên một giá sắt cố định. Sau 7 ngày thu các ống nghiệm trong đó động vật đất đã chui sâu qua đất, rơi vào ống nghiệm bên dưới và được định hình bằng cồn 90o
Xử lý mẫu vật, bảo quản và định loại: các ống nghiệm có mẫu vật (Collembola) thu được nhờ phễu Tullgren sẽ được lần lượt đổ ra đĩa petri để tính đếm số lượng các nhóm, các dạng loài dưới kính lúp hai mắt (Olympus SZ40) . Để xác định đến loài, tiến hành làm tiêu bản cố định, soi dưới kính hiển vi với độ phóng đại lớn (đến 4000 lần) (Olympus CH2). Định loại với các tài liệu chuyên môn. Các mẫu bọ nhảy không làm tiêu bản, sẽ được cho vào trong ống nghiệm chứa dung dịch bảo quản cồn 90
.
o
4.2.3.5.3 Thu mẫu bọ đuôi bật bằng phương pháp ống hút (suction sample)
. Các ống nghiệm đều được gắn nhãn ghi đầy đủ ngày thu, điểm thu, công thức,…Toàn bộ tiêu bản định loại và các mẫu vật được bảo quản tại phòng Sinh thái Môi trường đất, Viện Sinh thái và Tài nguyên Sinh vật, Viện Khoa học và Công nghệ Việt Nam. Danh sách loài Collembola được sắp xếp theo hệ thống cây chủng loại phát sinh dựa theo hệ thống phân loại của Moen và Ellis, 1984. Các loài trong một giống được sắp xếp theo vần a, b, c. Định tên loài theo tài liệu của Nguyễn Trí Tiến, 1995; Jan Stach (1965), Yoshii Ryozo (1982-1983); Hermann Gisin (1960); Loui Deharveng et Anne Bedos (1995).
Đây là phương pháp nhằm thu b ổ sung thêm về thành phần loài bọ đuôi bật Collembola có ở khu vực nghiên cứu mà không thu được bởi các phương pháp trên. Các mẫu vật Collembola sau khi được hút vào trong ống hút sẽ được phân tích định

48
tên loài ở phòng thí nghiệm để bổ sung danh sách thành p hần loài ở công thức thí nghiệm.
4.2.3.6 Đánh giá mức độ gây hại của sâu, bệnh hại ngô chính
4.2.3.6.1 Đánh giá mức độ gây hại của sâu hại ngô NTOs
Áp dụng chung cho các CT: chọn 1 đối tượng chích hút (rệp muội, bọ xít, nhện đỏ...) và 1 đối tượng miệng nhai (sâu đục bắp, sâu cắn lá, sâu róm, sâu xám, sâu khoang, bọ cánh cứng ăn lá 4 vệt,...) để đánh giá mức độ gây hại trên đồng ruộng. Việc xác định loài cụ thể sẽ được chuyên gia côn trùng thực hiện trước thời điểm đánh giá 1 tuần. Đánh giá 4 lần: giai đoạn giữa (V8 -V10), sau thụ phấn (R2 và R3 hoặc khoảng 2-3 tuần sau khi 100% tung phấn), và trước khi thu hoạc h. Năm cây sẽ được đánh giá là một mẫu đại diện. Tại một điểm lấy mẫu nhất định, 3 mẫu đại diện sao cho các cây không liên tiếp trên 1 hàng sẽ được lựa chọn. Có thể b ỏ qua 2-3 cây ở giữa để tránh động đến các cây gần đó trong quá trình đánh giá. Các điểm lấy mẫu sẽ được gắn thẻ và đánh số, cắm cột cờ hay ruy-băng để dễ nhận biết ở các ngày tiếp theo lấy mẫu. Đánh giá theo thang điểm từ 1-9 (Guthrie và cs, 1960).
4.2.3.6.2 Đánh giá mức độ xuất hiện, gây hại của bệnh hại ngô chính.
Những bệnh hại ngô phổ biến cần quan tâm để đánh giá: - Bệnh đốm lá lớn (Helminthoprium turcicum) - Bệnh đốm lá nhỏ (Helminthoprium maydis) - Bệnh rỉ sắt (Puccinia maydis) - Bệnh đốm nâu (Physoderma maydis) - Bệnh khô vằn (Rhizoctonia solani)
Các loại bệnh trên được đánh giá vào giai đoạn 80 - 85 ngày sau gieo, tỷ lệ bệnh và mức độ nhiễm bệnh được đánh giá theo thang điểm từ 1-9 (phương pháp của Lugod và cs.,2008).
Lấu mẫu: Năm cây sẽ được đánh giá là một mẫu đại diện. Tại một điểm lấy mẫu nhất định, 3 mẫu đại diện sao cho các cây không liên tiếp trên 1 hàng sẽ được lựa chọn (Hình 4-4). Có thể bỏ qua 2 -3 cây ở giữa để tránh động đến các cây gần đó trong quá trình đánh giá. Các mẫu bệnh được xác định trên cơ sở điều tra thực địa tại ruộng khảo nghiệm và ghi nhận tỷ lệ bệnh, chỉ số bệnh. Chỉ số bệnh hại trên lá, thân được đánh giá theo CIMMYT với thang cấp từ 1-9:
Cấp 0: không có vết bệnh

49
Cấp 1: vết bệnh chiếm 1- 10% diện tích bộ phận bị hại Cấp 3: vết bệnh chiếm 11-25% diện tích bộ phận bị hại Cấp 5: vết bệnh chiếm 26-50% diện tích bộ phận bị hại Cấp 7: vết bệnh chiếm 51-75% diện tích bộ phận bị hại Cấp 9: vết bệnh chiếm trên 75% diện tích bộ phận bị hại
Hình 4-5: Chỉ số bệnh hại trên lá. (A) Cấp 0; (B) Cấp 9; (C) Cấp 1; (D) Cấp7.
Tỷ lệ bệnh tính theo công thức: Cây bị bệnh Tỷ lệ bệnh (%) = -------------------------- x 100 Tổng số cây điều tra
Các vật liệu phục vụ cho việc nghiên cứu bao gồm: hệ thống lọc mẫu đất (rây lọc, phễu lọc,v.v….); dụng cụ tách mẫu, phân tích mẫu và làm tiêu bản như đĩa petri, lam kính, lamen và các hóa chất thường dùng trong nghiên cứu động vật đất; kính lúp Olympus SZ40; kính hiển vi Olympus CH2; ngoài ra, một số loại dụng cụ khác như lọ đựng mẫu, túi nilon, hộp lấy mẫu đất, bút, sổ ghi chép, … cũng được sử dụng để phục vụ cho việc nghiên cứu.
4.2.4 Ghi nhận thông tin, phân tích và xử lý số liệu
4.2.4.1 Ghi nhận thông tin, dữ liệu
Cán bộ tham gia chịu trách nhiệm duy trì và giữ cẩn thận Sổ ghi chú khảo nghiệm. Sổ được ghi đầy đủ, chính xác bằng bút chì. Tất cả hoạt động bao gồm các đánh giá, ngày giờ đặt bẫy, ngày giờ quan sát, các lưu ý chung khi quan sát, các kết
A B C D

50
luận của thí nghiệm,… được ghi chép lại. Thông tin, dữ liệu được ghi theo mẫu phụ lục 6.1, 6.3, 6.4 (xem A.5.) của thông tư 69/ 2009/TT-BNNPTNT về Quy định khảo nghiệm đánh giá rủi ro đối với đa dạng sinh học và môi trường của giống cây trồng biến đổi gen.
4.2.4.2 Phân tích và xử lý số liệu
Thống kê và tính toán theo tài liệu của Gorny M. và L. Grum (1993), kết hợp sử dụng phần mềm Primer 5; phần mềm Excel 2003, phân tích phương sai (ANOVA) với độ tin cậy 95%; sử dụng test Kolmogorov-Smirnov (Richard L. Boyce, 2005) để kiểm định giá trị của chỉ số đa dạng H’. Các chỉ số phân tích:
- Số lượng loài: được tính bằng tổng số loài có mặt trong điểm thí nghiệm ở tất cả các lần thu mẫu.
- Mật độ cá thể trung bình: số lượng cá thể trung bình có ở tất cả các cốc của điểm thí nghiệm (với phương pháp bẫy cốc) hay số lượng cá thể trung bình có ở tất cả các lần thu mẫu của điểm thí nghiệm (con/m2
- Chỉ số đa dạng Shannon-Weaver (H’): được sử dụng để tính sự đa dạng loài hay số lượng loài trong quần xã và tính đồng đều về sự phong phú cá thể của các loài trong quần xã. Chỉ số được tính theo công thức:
) (với phương pháp thu mẫu đất).
Nni
NniH
s
i∑−
−=1
ln'
Trong đó: s: số lượng loài ni: số lượng cá thể của loài i N: tổng số lượng cá thể trong toàn bộ mẫu Giá trị của H’ dao động trong khoảng 0 - ∞. Chỉ số đa dạng của quần xã phụ
thuộc vào hai yếu tố là số lượng loài và tính đồng đều về sự phong phú của các loài trong quần xã. Một khu vực có số lượng loài hoặc số cá thể nhiều chưa hẳn nơi đó có tính đa dạng cao. Chỉ số đa dạng, ở một khía cạnh nào đó cho biết tính đa dạng của quần xã và là một chỉ tiêu có thể đánh giá được tính đa dạng về khu hệ động vật của một khu vực.
- Chỉ số đồng đều (J’) hay chỉ số Pielou: được tính theo công thức:
)ln(''
SHJ =
Trong đó: S: tổng số loài; H’: Chỉ số đa dạng Shannon-Weaver Giá trị của J’ dao động trong khoảng từ 0 đến 1; Giá trị của J’ lớn nhất khi tất
cả các loài trong quần xã có số lượng cá thể bằng nhau hay nói cách khác độ phong phú của các loài trong quần xã như nhau.

51
- Loài ưu thế: là những loài có giá trị chỉ số ưu thế bằng hoặc lớn h ơn 5% (Maria Sterzynska, 1990). Chỉ số ưu thế được tính theo công thức:
100xnn
D a=
Trong đó: na
n: tổng số cá thể của toàn bộ mẫu theo sinh cảnh hay địa điểm. : số lượng cá thể của loài a
- Loài phổ biến: là những loài có giá trị chỉ số thường gặp từ 50% -100% . Chỉ số thường gặp được tính theo công thức:
100xNNaC =
Trong đó: Na: số lượng mẫu thu có chứa loài a N: tổng số lượng mẫu của sinh cảnh nghiên cứu
Tổng số 60 mẫu bẫy cốc và 72 mẫu đất đã được thu thập và phân tích. Mẫu vật hiện được bảo quản tại phòng Sinh thái Môi trường đất, Viện Sinh thái và Tài nguyên sinh vật, Viện Khoa học và Công nghệ Việt Nam, 18 Hoàng Quốc Việt, Cầu Giấy, Hà Nội.
4.3 Vật liệu, phương pháp KNDR
4.3.1 Giống đăng ký KNDR
Trên cơ sở các kết luận thu được từ KNHC và quy địn h công nhận c ây trồng BĐG của Việt Nam (đăng ký và công nhận theo Event). Các giống được lựa chọn cho KNDR được thể hiện trong Bảng 4-4.
Bảng 4-4: Tên giống và các công thức trong KNDR
CT Ký hiệu trong báo cáo
Kí hiệu giống
Ghi chú*
1 G1 30Y87H Giống chuyển gene mang event TC1507 kháng côn trùng cánh vảy (Lepidoptera)
2 G2 30Y87HR Giống chuyển gene mang stack TC1507xNK603 kháng côn trùng cánh vảy (Lepidoptera) và kháng thuốc trừ cỏ glyphosate; đối chứng
3 G3 30Y87 Giống nền không chuyển gene; đối chứng * Nguồn: Bộ phận sản xuất và vườn ươm giống của Tập đoàn Pioneer Hi-Bred Intl.

52
4.3.2 Thời gian và địa điểm KNDR
4.3.2.1 Thời gian KNDR ngô event TC1507
Bảng 4-5 Thời gian gieo và thu hoạch tại mỗi địa điểm KNDR
TT Địa điểm khảo nghiệm TG gieo hạt TG thu hoạch
1 Trại SX GCT Vĩnh Phúc 07/5/2011 17-18/5/2011 2 HTX Phong Thịnh - Thanh Chương,
Nghệ An 13/5/2011 21-22/5/2011
3 Nông trường Thống nhất, KrongPak, Đắk Lắk
17/5/2011 26-27/5/2011
4 TT NC Thực nghiệm Hưng Lộc - Đồng Nai.
20/5/2011 30-31/5/2011
4.3.2.2 Địa điểm KNDR ngô event TC1507
+ Trại SX giống cây trồng Vũ Di, Khu 1, thị trấn Vĩnh Tường, Vĩnh Phúc (diện tích: 1ha ) + Xã Phong Thịnh, huyện Thanh Chương, tỉnh Nghệ An (diện tích: 1ha ) + Nông trường cà phê Thắng Lợi, huyện Krông Pak, Đắk Lắk (diện tích :1,2ha) + Trung tâm Nghiên cứu Thực nghiệm Nông nghiệp Hưng Lộc, xã Hưng Thịnh, huyện Trảng Bom, tỉnh Đồng Nai (diện tích: 1ha )
Lý do chọn điểm khảo nghiệm:
Theo thống kê hiện trạng SX ngô của Việt Nam (số liệu thống kê bộ NN&PTNT, 2010), các vùng trồng ngô chính trên cả nước hiện nay là (1) khu vực Trung du và miền núi phía Bắc (Sơn La, Hà Giang…, tổng DT: 460 nghìn hecta); (2) khu vực Tây Nguyên (Đắk Lắk, Gia Lai…, tổng DT: 236.6 nghìn hecta ); (3) khu vực Bắc Trung bộ (Thanh Hóa, Nghệ An …; tổng DT: 213.6 nghìn hecta ); (4) khu vực Đồng bằng sông Hồng (Vĩnh Phúc, Hà Nội …, tổng DT: 97.6 nghìn hecta); và (5) Đông Nam Bộ (các tỉnh Đồng Nai, Bà Rịa- Vũng Tàu…, tổng DT: 81.3 nghìn hecta). Như vậy, ngoại trừ khu vực Trung du và miền núi phía Bắc, mỗi địa điểm khảo nghiệm diện rộng được lựa chọn tương ứng đại diện cho mỗi vùng trồng ngô trọng điểm của Việt Nam hiện nay.

53
4.3.3 Phương pháp KNDR (áp dụng chung cho cả 4 địa điểm)
4.3.3.1 Sơ đồ bố trí KNDR
Mỗi công thức thí nghiệm đánh giá ảnh hưởng đến môi trường và đa dạng sinh học được triển khai trên diện tích 2.025m2
Hình 4-6. Sơ đồ KNDR (áp dụng chung cả 4 vùng sinh thái Việt Nam)
(45m x 45m) và canh tác theo hướn g dẫn của Cục Trồng trọt - Bộ Nông nghiệp và Phát triển Nông thôn.
4.3.3.2 Phương pháp kiểm tra sự hiện diện của protein Cry1F và CP4 EPSPS
Xác định sự hiện diện (nếu có) của các protein chỉ thị tại các ô khảo nghiệm tương ứng nhằm xác định tính chính xác trong bố trí thí nghiệm đồng ruộng cũng như tính đồng nhất của hạt giống bằng việc sử dụng QuickStix™ Kit cho Cry1F và QuickStix™ Kit cho Roundup Ready (chỉ thị sự hiện diện của gen cp4 epsps kháng thuốc trừ cỏ glyphosate). Cách thức thu mẫu và phương thức tiến hành tương tự như trình bày mục 4.2.3.2 trong KNHC.
Hình 4-7. Các dụng cụ kiểm tra sự hiện diện của protein Cry1F và CP4 EPSPS
1 2 3 4 5 6 7 8

54
Ghi chú:
1. Dung dịch đệm QuickStix EB2 5. Que thử nhanh protein Cry1F 2. Lọ nhỏ giọt 6. Hộp đựng que thử nhanh protein Cry1F 3. Ống eppendorf 1,5ml 7. Hộp đựng que thử nhanh Roundup Ready 4. Chày nhựa 8. Que thử nhanh Roundup Ready (chỉ thị gen kháng
thuốc trừ cỏ glyphosate) 4.3.3.3 Phương pháp đánh giá tính ổn định và tính thích ứng của ngô event
TC1507 tại các vùng sinh thái Việt Nam
Thực hiện thông qua việc đánh giá các đặc tính nông sinh học và mức độ mẫn cảm /kháng với các loại sâu bệnh chủ yếu
4.3.3.3.1
của các giống khảo nghiệm
Tính ổn định
Thu thập số liệu (ví dụ như đo chiều cao cây, chiều cao bắp,…), ghi nhận giá trị đo lường ghi lại từ cây khỏe, đại diện trong mỗi ô được lựa chọn một cách khách quan. Tất cả các CT thí nghiệm được đánh giá đặc tính nông học trong cùng một ngày.
Bảng 4-6: Chỉ tiêu đánh giá đặc tính nông sinh học
T.T Chỉ tiêu đánh giá GĐ đánh giá Giá trị ghi nhận
1 Số cây mọc/tỷ lệ mọc V2-V4 Số cây thực tế mọc trong mỗi ô hay tỷ lệ % cây mọc trong mỗi ô
2
Sức sống cây con V2-V4 Ước tính sức sống TB của cây trong mỗi ô theo thang điểm 1 -9 (1:cây ngắn, lá nhỏ, mỏng; 9:cây cao, lá to, khỏe)
3
Thời gian phun râu TG 50% phun râu
Ghi nhận ngày khoảng 50% cây phun râu
4
Thời gian tung phấn TG 50% tung phấn
Ghi nhận ngày khoảng 50% cây tung phấn
5 Chiều cao cây R4 Chiều cao cây từ mặt đất đến đỉnh cờ 6 Chiều cao đóng bắp R4 Chiều cao tính từ mặt đất đến mắt đóng của
bắp chính 7 Trạng thái cây R5 Đánh giá sức khỏe của cây theo th ang điểm
1-9 (1:không có mô thấy màu xanh; 5: 50%

55
T.T Chỉ tiêu đánh giá GĐ đánh giá Giá trị ghi nhận
mô lá còn xanh; 9: rất xanh, khoảng 90% mô lá còn xanh
8 Tỷ lệ đổ gẫy R6 % cây trong mỗi ô bị đổ gẫy bên dưới bắp chính
9 Tỷ lệ bật gốc R6 % cây nghiêng khoảng 300 so với mặt đất 10 Số cây thu hoạch R6 Ghi nhận số cây thực tế còn lại trong các ô
(không tính cây non) 11 Năng suất thực thu R6 Thu bắp của 2 hàng giữa mỗi ô
*/ Công thức tính năng suất thực thu (tấn/ ha):
)(10000(%)15100(%)100
)()( 2
0
3
22
0
1)/(tan mA
PP
mSkgPNS ha ×
−−
××=�
P1
A: Trọng lượng bắp tươi thu hoạch của mỗi ô
0
S: Ẩm độ hạt khi thu hoạch.
0
P: Diên tích ô thu hoạch (chiều dài hàng × độ rộng hàng × số hàng).
2
P: Trọng lượng hạt của mẫu
3
: Trọng lượng bắp tươi của mẫu
4.3.3.3.2 Mức độ mẫn cảm /kháng với các loại sâu bệnh chính
+ Tính mẫn/kháng bệnh Đốm lá lớn (
Helminthosporium turcicum): phương pháp đanh giá được thực hiện tương tự KNHC
+ Tính mẫn/kháng
+ Sâu khoang (Spodoptera litura),
sâu hại ngô trên đồng ruộng: Đánh giá mức độ gây hại trên đồng ruộng do lây nhiễm tự nhiên của các loài:
+ Sâu đục thân Châu Á (Ostrinia furnacalis), + Sâu xám (Agrotis ipsilon), + Sâu đục thân mình hồng (Sesamia inferens). + Rệp muội ngô (Rhopalosiphum maidis)
Việc xác định loài cụ thể cần khảo sát, đánh giá do các chuyên gia côn trùng học quyết định, trên cơ sở thực tế tình hình sâu hại tại địa điểm khảo nghiệm , tùy theo mức độ gây hại tự nhiên nhiều hay ít của sâu hại trước thời điểm đánh giá 1 tuần.
Sâu khoang (Spodoptera litura): Mức độ gây hại của sâu khoang được đánh giá trên 100 cây, tại 5 điểm ngẫu nhiên ở mỗi ô khảo nghiệm (Hình 4-4) ở giai đoạn

56
V4 - V6. Tỷ lệ hại do sâu khoang gây ra được đánh giá theo thang điểm của Guthrie và cs., (1960).
Bảng 4-7: Thang điểm đánh giá mức độ gây hại của một số loại sâu hại chính
Thang điểm*
Mô tả
0 Không có biểu hiện bị gây hại 1 Số lượng ít vết cắn với kích thước bằng đầu kim 2 Số lượng ít vết cắn lỗ tròn nhỏ trên một vài lá 3 Số vết cắn lỗ tròn nhỏ phổ biến trên một vài lá 4 Một số lá với các vết cắn tròn nhỏ và một số vết cắn dài 5 Một số lá với vết cắn dài khoảng 1,3-2,5 cm 6 Một số lá với vết cắn dài hơn 2.5cm 7 Vết cắn dài, xuất hiện trên 1/2 số lá 8 Vết cắn dài, xuất hiện trên 2/3 số lá 9 Hầu hết các lá có các vết cắn kéo dài
*/ Thang điểm đánh giá: theo thang điểm từ 1-9 (Guthrie và cs, 1960).
Sâu đục thân ngô (Ostrinia furnacalis) và sâu đục thân mình hồng (Sesamia inferens): Tỷ lệ gây hại của sâu đục thân được đánh giá bằng cách đếm số cây có biểu hiện bị hại điển hình do sâu đục thân ngô gây ra trên 100 cây tại 5 điểm ngẫu nhiên ở mỗi ô khảo nghiệm tại thời điểm V10-V15. Thang điểm đánh giá sự gây hại của sâu đục thân trên lá ngô được thể hiện trong Bảng 4-7. Ngoài ra, mức độ gây hại của sâu đục thân ngô còn được đánh giá qua các thông số sau tại thời điểm thu hoạch: Lỗ đục: vết đục và số lỗ đục gây ra bởi sâu đục thân trên thân ngô, phía trên vị trí đóng bắp trên 100 cây tại 5 điểm ngẫu nhiên trong mỗi ô khảo nghiệm tại thời điểm thu hoạch.
Số lượng và chiều dài đường đục trên thân gây ra bởi sâu đục thân ngô: Số liệu về số các đường đục được đánh giá tính từ lóng thân thứ 3 phía trên vị trí đóng bắp cao nhất tới gốc của thân ngô được ghi nhận từ 100 cây ngẫu nhiên tại 5 điểm trên mỗi ô khảo nghiệm tại thời điểm thu hoạch. Việc đo đếm được tiến hành dựa trên tổng chiều dài của các rãnh đục với chiều dài tối thiểu là 0.5cm bao gồm các lỗ đục xuyên ngang qua thân, loại trừ các lỗ đục nhìn thấy nhưng chỉ ở phía bên ngoài thân ngô.
Sâu xám (Agrotis ipsilon): Mức độ tổn thương do sâu xám gây ra được đánh
giá vào giai đoạn V2-V5. Số lượng cây bị cắt trong mỗi ô thí nghiệm sẽ được ghi nhận.

57
4.3.3.4 Phương pháp đánh giá đa dạng quần thể NTOs trong KNDR ngô event
TC1507 tại các VST của Việt nam
a) Đa dạng quần thể côn trùng : Được đánh giá bằng phương pháp quan sát kết hợp bẫy dính vàng bao gồm:
+ Sự đa dạng về thành phần loài của quần thể côn trùng thuộc nhóm ăn thực vật (sâu hại nhóm chích hút),
+
+ Sự đa dạng về thành phần loài và sự phân bố của quần thể côn trùng thuộc nhóm thụ phấn.
Sự đa dạng về thành phần loài và sự phân bố của quần thể côn trùng thuộc nhóm bắt mồi ăn thịt,
+ Đánh giá đa dạng và thành phần loài của Bọ đuôi bật Collembola.
b) Mật độ động vật chân đốt/đợt điều tra: Điều tra (đếm) định kỳ trong suốt mùa vụ, tập trung xung quanh giai đoạn tung phấn. Trọng tâm chính là thu mẫu đánh giá các loài chỉ thị trên ruộng thuộc các nhóm: động vật ăn thịt, ký sinh, động vật đất, và động vật ăn thực vật (Bảng 4-8). Dữ liệu phải được thu nhận tại thời gian tương tự trong ngày cho mỗi giai đoạn quan sát với mỗi phương pháp theo dõi.

58
Bảng 4-8: Các loài động vật chân khớp được theo dõi trong nghiên cứu NTO.
1: Shanahan và cs., 2000; EA c ủa USDA/APHIS, 2001, đánh giá tại Mỹ
2: Higgins và cs, 2009, đánh giá thực hiện trong 3 năm tại Mỹ.
4.3.3.4.1 Phương pháp điều tra thu thập số liệu
a) Phương pháp áp dụng cho quan sát và đặt bẫy
Các công thức thí nghiệm được bố trí ngẫu nhiên trong các ô thí nghiệm. Các ô này được đánh dấu để tham khảo trong suốt qua trình thu thập dữ liệu. Việc thu thập dữ liệu trong các ô thí nghiệm tiến hành ở các điểm gần trung tâm của “khu vực thu mẫu” bằng cách loại bỏ 10 m vùng bên ngoài nhằm giảm thiểu hiệu ứng rìa. Điểm thu mẫu có thể cố định trong suốt vụ. Các điểm quan sát được chọn ngẫu nhiên và được ghi nhận trong sổ theo dõi khảo nghiệm.
- Số liệu về đặc điểm nông sinh học và hiệu quả sinh học: được thu thập tại 3 điểm, cụ thể:
+ Mỗi điểm lấy mẫu tiến hành đánh giá trên 10 cây ngẫu nhiên, + Mỗi ô thí nghiệm đánh giá với 4 điểm lấy mẫu. Do vậy, số cây điều tra/công
thức thí nghiệm là: 4 điểm x 10 cây = 40 cây.
- Số liệu điều tra đa dạng thành phần loài côn trùng: được thu thập tại 3 điểm, cụ thể: mỗi điểm lấy mẫu tiến hành đánh giá quan sát trên 15 cây ngẫu nhiên; đặt 3 bẫy dính; lấy 5 mẫu đất
*/ Phương pháp bẫy dính
Chỉ tiêu đánh giá trong KNHC
Chỉ tiêu đã được đánh giá trên thế giới
Nội dung đánh giá trong KNDR
Bọ đuôi bật (Collembola)
Bọ đuôi bật (Collembola) Động vật chân khớp ăn thành phần trong đất
1
Bọ rùa (Coleoptera; Coccinellidae)
Bọ rùa cánh cứng (Coleoptera; Coccinellidae
Quần thể côn trùng thuộc nhóm bắt mồi ăn thịt )
Rệp (Hemiptera; Aphidae)
N/A
N/A
Quần thể côn trùng thuộc nhóm ăn thực vật (nhóm chích hút)
Ong mật (Hymenoptera; Apidae)
Quần thể côn trùng thuộc nhóm thụ phấn 2

59
Sử dụng bẫy dính vàng (Pherocon AM No Bait traps, Trece Inc., Salinas, CA, USA). Việc điều tra được tiến hành ở các giai đoạn sớm (khoảng V4-V6), giai đoạn giữa (khoảng V8-V10), trước khi thụ phấn ( VT), trong quá trình thụ phấn (R1, khoảng 50% tung phấn), và sau thụ phấn (R2 - R3 hoặc 2-3 tuần sau khi 100% cây đã tung phấn). Các điểm lấy mẫu đều được gắn thẻ và đánh số, cắm cột cờ hay ruy-băng để dễ nhận biết cho lấy mẫu tiếp theo. Phương pháp đặt bẫy tương tự như KNHC. Thời gian theo dõi tại Bảng 4-9.
Hình 4-8: Bẫy dính vàng đặt trong ruộng ngô
Bảng 4-9: Các đợt thu mẫu côn trùng ở các GĐST của cây ngô tại 4 KNDR
(tháng 6 - 8, 2011)
Đợt thu V. Phúc N. An Đ. Lắc Đ. Nai GĐST
1 17/5 25/5 10/6 18/6 V4-V6 2 5/6 7/6 24/6 30/6 V8-V10 3 17/6 20/6 13/7 18/7 VT 4 8/7 5/7 20/7 30/7 R1 5 18/7 21/7 28/7 10/7 R3
Mẫu bổ sung cho quan sát bằng mắt và bẫy dính có thể được tiến hành trong thời điểm phổ biến côn trùng săn mồi. Điểm lấy mẫu bổ sung sẽ được ghi vào sổ ghi chú khảo nghiệm.
b) Thu mẫu Collembola
Mẫu đất thu nhằm định lượng Collembola được thực hiện theo tài liệu hướng dẫn của Ghilarov, 1975. Tại mỗi ô thí nghiệm, lấy 5 mẫu đất ở lớp bề mặt 0 – 10cm,

60
kích thước 5x5x10cm trên bề mặt l uống, cách gốc cây khoảng 2-4 cm bằng dụng cụ chuyên dụng. Việc thu mẫu được nhắc lại 3 lần tại các khoảng thời gian (bảng 4-10). Tại mỗi đợt, tổng số mẫu sẽ thu là: 5 mẫu đất x 3 công thức x 4 điểm/công thức = 60 mẫu/ 1 điểm khảo nghiệm. Mẫu đất được cho vào túi ni lông có kèm nhãn (ghi đầu đủ thông tin về địa điểm, ngày th u, ký hiệu giống ngô, vv ), buộc c hặt và chuyển về phòng thí nghiệm.
Bảng 4-10: Các đợt thu mẫu Collembola ở các GĐST của ngô tại 4 điểm KNDR
(tháng 6 - 8, 2011)
Đợt thu V. Phúc N. An Đ. Lắc Đ. Nai GĐST
1 8/ 6 12/ 6 17/ 6 17/ 6 Giai đoạn trước thụ phấn
2 3/ 7 4/ 7 15/ 7 26/ 7 Trong quá trình thụ phấn. 3 18/ 7 22/ 7 29/ 7 12/8 Sau thụ phấn 2-3 tuần
c) Tại phòng thí nghiệm: Phương pháp thực hiện trong phòng thí nghiệm được tiện hành hoàn toàn tương tự như các phương pháp đã trình bày trong KNHC
4.3.4 Phương pháp phân tích và xử lý số liệu
- Xử lý số liệu các chỉ tiêu nông sinh học: So sánh số liệu trung bình của các giống, trong cùng một điểm và trong cả 4 điểm bằng chương trình IRRISTAR 5.0.
- Xử lý số liệu các chỉ tiêu quần thể sinh vật không chủ đích : Phương pháp xử lý giống như phương pháp xử lý trong khảo nghiệm hạn chế
4.3.5 Phương pháp thực hiện chung cho tất cả các nội dung trong KNDR
4.3.5.1 Phương pháp gieo trồng, chăm sóc
Gieo trồng: Đảm bảo ẩm độ đất đầy đủ trước khi gieo trồng. Gieo hạt bằng tay, theo hàng (hàng cách hàng 0,70 m; cây cách cây 0,25 m), mỗi hốc gieo 2 hạt, đảm bảo độ đồng đều về độ sâu lấp đất, khoảng cách và mật độ. Số hạt giống sử dụng cho khảo nghiệm sẽ được gieo trồng hết, nếu thừa sẽ được tiêu hủy hoặc bảo quản theo đúng quy định an toàn sinh học theo thông tư 69/2009/TT-BNNPTNT.

61
Bảng 4-11: Thời gian gieo và thu hoạch
Địa điểm khảo nghiệm TG gieo hạt TG thu hoạch
Trại SX GCT Vĩnh Phúc 07/5/2011 17-18/5/2011 HTX Phong Thịnh, Thanh Chương, Nghệ An
13/5/2011 21-22/5/2011
Nông trường Thống nhất, KrongPak, Đắk Lắk
17/5/2011 26-27/5/2011
TT NC Thực nghiệm Hưng Lộc, Đồng Nai.
20/5/2011 30-31/5/2011
- Kiểm soát mật độ, tỉa thưa: Khi 50% số cây trong mỗi ô khảo nghiệm đạt giai đoạn V2-V4 (khoảng 12-15 ngày sau trồng). Các ô thí nghiệm sẽ được tỉa thưa đảm bảo đúng mật độ và khoảng cách thí nghiệm bằng cách nhổ bỏ những cây yếu. Việc thu gom những cây đã nhổ và tiêu hủy được thực hiện theo đúng hướng dẫn trong thông tư 69/2009/TT-BNNPTNT. Việc kiểm soát số lượng cây trong các ô khảo nghiệm là 1 cây/hốc (cả cây chuyển gen và cây không chuyển gen) và đảm bảo tính đồng đều số cây/hàng trong mỗi ô thí nghiệm.
- Chăm sóc, bón phân, tưới nước, ghi dữ liệu nhiệt độ, ẩm độ tại các điểm khảo nghiệm: Các dữ liệu về thời tiết được thu thập trực tiếp ngay tại ruộng khảo nghiệm bằng máy đo nhiệt độ, độ ẩm đặt tại ruộng hoặc được thu thập từ các trạm khí tượng thủy văn của từng vùng để có dữ liệu đại diện tại mỗi địa điểm khảo nghiệm. Lượng mưa hàng ngày và nhiệt độ không khí hàng ngày tối thiểu /tối đa (độ C) sẽ được ghi lại vào lúc 8 giờ và 14h mỗi ngày trong suốt thời gian khảo nghiệm (từ ngày trồng đến ngày thu mẫu cuối cùng)
- Vành đai bảo vệ: Được thiết lập bằng cách trồng 7 hàng ngô giống 30Y87 xung quanh ruộng khảo nghiệm.
.

62
5 KẾT QUẢ KHẢO NGHIỆM NGÔ CHUYỂN GEN EVENT TC1507 ĐỐI VỚI ĐA DẠNG SINH HỌC VÀ MÔI TRƯỜNG SINH THÁI VIỆT NAM
5.1 Kết quả KNHC ngô event TC1507 (2 vụ lên tiếp)
5.1.1 Kết quả kiểm tra sự hiện diện của protein Cry1F (giai đoạn V4)
Kết quả kiểm tra sự có mặt của protein Cry1F trên các giống ngô khả o nghiệm được thể hiện ở Hình 5-1. Ngô chuyển gen 30Y87H cho kết quả dương tính với protein Cry1F (xuất hiện 2 vạch trên que thử nhanh). Hai giống Đ/C không chuyển gen là 30Y87, 30B80 và que đối chứng (chỉ thử với dịch đệm QuickStix EB2) cho kết quả âm tính với protein Cry1F (xuất hiện 1 vạch trên que thử nhanh). Kết quả thu được cũng hoàn toàn giống nhau ở 3 lần lặp lại, cho cả hai vụ khảo nghiệm. Điều đó thể hiện việc bố trí thí nghiệm ruộng KNHC chế ngô chuyển gen hoàn toàn chính xác và hạt giống thử nghiệm là đồng nhất.
Hình 5-1: Kết quả kiểm tra sự hiện diện của protein Cry1F trên các giống ngô.
(A) Vụ 1 và (B) Vụ 2
A
B

63
• Ghi chú I, II, III: Lần nhắc lại 30Y87H: ngô chuyển gen kháng sâu 30Y87: ngô không chuyển gen 30B80: ngô không chuyển gen Buffer: mẫu đối chứng với dung dịch đệm
5.1.2 Kết quả đa dạng quần thể NTOs trên ruộng KNHC
Công thức tính đa dạng loài của Odum (1971), Colwell (2000) cho phép tính các chỉ tiêu liên quan đến tính đa dạng loài của tập hợp chân khớp trong ruộng ngô khảo nghiệm. Trung bình tổng số cá thể chân khớp bắt gặp trong một kỳ điều tra (độ phong phú) trên các giống ngô khảo nghiệm, vụ trồng khảo nghiệm không giống nhau và không phụ thuộc vào yếu tố mang gen cry1F.
Vụ 1: chỉ tiêu này đạt cao nhất ở giống ngô nền không chuyển gen 30Y87 (191,33 cá thể). Giống ngô 30Y87H (chuyển gen) có trung bình tổng số cá thể chân khớp bắt gặp trong một kỳ điều tra đạt thấp nhất trong các giống ngô khảo nghiệm (49,47 cá thể).
Vụ 2: tổng số cá thể cao nhất ở giống ngô chuyển gen 30Y87H (354,13 cá thể), thấp nhất ở giống ngô 30B80 ( 269,20 cá thể).
So sánh số loài bắt gặp trong một kỳ điều tra trong 2 vụ trồng cho thấy: Vụ1: giống ngô không chuyển gen 30Y87 có giá trị cao nhất về số loài bắt gặp
(20,2 loài), trị số này thấp nhất ở giống 30Y87H (18,7 loài). Ở vụ 2: số loài bắt gặp cao nhất ở giống 30Y87H (13,53 loài), trị số này thấp nhất ở giống 30Y87 (12,87 loài). Tuy nhiên sự chênh lệch này không đáng kể. Như vậy, những s ự khác nhau về số loài bắt gặp trung bình trong một kỳ điều tra giữa giống ngô chuyển gen với các giống ngô không chuyển gen trong cả 2 vụ là không đáng kể (Bảng 5-1, Hình 5-2; 5-3). Kết quả, số loài chân khớp bắt gặp trong một kỳ điều tra ở các giống ngô chuyển gen cry1F thấp hơn giống ngô nền không chuyển gen là hoàn toàn hợp lý, vì trên giống ngô mang gen cry1F không thuận lợi cho sự phát triển của một số loài sâu hại, đặc biệt các loài sâu hại chủ đích (Pilcher et al., 2005). Mặt khác, khi giảm đáng kể mật độ quần thể của sâu hại chủ đích trên cây chuyển gen (sự thiếu vắng sâu hại/mật độ sâu hại thấp) sẽ dẫn đến giảm quần thể của các loài thiên địch chuyên tính của các sâu hại chủ đích và không hấp dẫn các loài thiên địch đa thực đến cư trú, sinh sống trên các giống ngô chuyển gen. Cả hai lý do này đã tạo nên tình huống số loài chân khớp bắt gặp trong một kỳ điều tra trên giống ngô chuyển gen có thể thấp hơn so với giống ngô nền.

64
1.00
3.00
5.00
7.00
9.00
11.00
13.00
15.00
30B80 30Y87 30Y87H
Giống ngô thí nghiệm
S ố lo
ài t
rung b
ình/k
ỳ đ
iều tra
Tổng số loài
0
50
100
150
200
250
300
350
400
30B80 30Y87 30Y87H Giống ngô thí nghiệm
T ổng s
ố c
á thể b
ắt gặp/k
ỳ đ
iều tra
Tổng số cá thể
Bảng 5-1:Tổng số cá thể và số loài chân khớp bắt gặp trên các giống ngô trong
KNHC (Văn Giang, 2010 - 2011)
Giống
Tổng số cá thể bắt gặp/kỳ điều tra (cá thể)
Số loài bắt gặp/kỳ điều tra (loài)
Vụ 1 Vụ 2 Vụ 1 Vụ 2
30B80 159,73 ± 31,61 269,20± 28,88 19,9 ± 2,48 13,30 ± 0,24 30Y87 191,33 ± 38,18 305,80± 38,05 20,2 ± 1,69 12,87 ± 0,57
30Y87H 149,47 ± 23,36 354,13± 6,96 18,7 ± 1,81 13,53 ± 0,44
Hình 5-2: Trung bình tổng số loài bắt gặp trong một kỳ điều tra trên ngô KNHC
(Văn Giang, 2010 -2011). Vụ 1 (hình trái), vụ 2 (hình phải).
Hình 5-3: Trung bình số loài chân khớp bắt gặp trong một kỳ điều tra trên ngô KNHC (Văn Giang, 2010-2011). Vụ 1 (hình trái), vụ 2 (hình phải).

65
Sự đa dạng của quần thể sinh vật không chủ đích (NTO) còn được thể hiện bằng
chỉ số ưu thế và chỉ số đa dạng. Kết quả này thể hiện ở Bảng 5-2 và Hình 5-4. Các giá trị của chỉ số ưu thế Simpson's D đều thấp và biến động từ 0,1392 ở giống ngô 30Y87 đến 0,1576 ở giống ngô 30Y87H (vụ 1); 0,3233 ở giống ngô 30Y87H đến 0,2729 ở giống 30B80 (vụ 2). Điều này chứng tỏ trong sinh quần các giống ngô thí nghiệm, không có loài chân khớp nào phát triển với số lượng trội hơn, chiếm ưu thế hơn để gây mất cân bằng sinh thái. Tuy nhiên, giá trị của chỉ số ưu thế Simpson's D trên giống ngô chuyển gen có xu hướng cao hơn (tương ứng) so với trên giống ngô nền không chuyển gen (Hình 5-4). Sự khác nhau về giá trị chỉ số ưu thế Simpson's D của các giống ngô thí nghiệm là không đáng kể. Trên giống ngô chuyển gen 30Y87H có chỉ số ưu thế Simpson's D cao hơn, đồng thời có chỉ số đa dạng Shannon H' thấp hơn so với trên giống ngô nền không chuyển gen 30Y87 và 30B80 (trong cả 2 vụ trồng khảo nghiệm). Như vậy, c ác giá trị của chỉ số ưu thế Simpson's D hoàn toàn lôgíc, tương ứng phù hợp với giá trị của chỉ số đa dạng Shannon H'.
Khi xét về chỉ số đa dạng Shannon H' (một chỉ tiêu quan trọng nhất của đa dạng sinh học ở cấp độ loài) cho thấy các giá trị của chỉ số đa dạng Shannon H' đều đạt khá cao và đạt > 4 (vụ 1) ; > 3 (vụ 2). Xử lý thống kê giá trị trung bình của chỉ số đa dạng Shannon H’ của các giống ngô khảo nghiệm cho thấy giá trị chỉ số đa dạng Shannon H' của các giống ngô đều nằm trong khoảng từ 4,6258 đến 4,7238 (vụ 1) và 3,0016 đến 3,2517 (vụ 2). Sự khác biệt về chỉ số đa dạng Shannon H' giữa các giống ngô thí nghiệm trong cùng 1 vụ là không có ý nghĩa. Do điều kiện thời tiết 2 vụ trồng ngô có khác nhau dẫn đến chỉ số H’ cũng khác nhau, nhưng sự khác nhau về chỉ số đa dạng H’ ở các giống ngô khảo nghiệm trong 2 vụ không lớn.
Bảng 5-2: Chỉ số ưu thế và chỉ số đa dạng loài chân khớp trên các giống ngô
trong KNHC (Văn Giang, 2010 - 2011)
Giống
Chỉ số ưu thế Simpson's D Chỉ số đa dạng Shannon H' Vụ 1 Vụ 2 Vụ 1 Vụ 2
CT1 0,1458 ± ,0125 0,2729 ± 0,0191 4,6812 ± 0,0758 3,2517 a ± 0,0717 CT2 0,1392 ± 0,0108 0,3219 ± 0,0069 4,7238 ± 0,0445 3,0016 b ± 0,0258 CT3 0,1576 ± 0,0157 0,3233 ± 0,1161 4,6258 ± 0,0573 3,0321 b ± 0,0554
CV(%) 3.99 3.53 LSD. 05 0.35 0.31
Ghi chú: CT1: 30B80 CT2: 30Y87 CT3: 30Y87H

66
Shannon H'
2.00
2.20
2.40
2.60
2.80
3.00
3.20
3.40
30B80 30Y87 30Y87H
Giống ngô thí nghiệm
Ch
ỉ ố đ
d S
h H
'
Shannon H'
4.00
4.10
4.20
4.30
4.40
4.50
4.60
4.70
4.80
30B80 30Y87 30Y87HGiống ngô thí nghiệm
Ch
ỉ ố
đ d
Sh
H'
Shannon H'
Hình 5-4: Giá trị của chỉ số ưu thế Simpson's D trên các giống ngô KNHC (Văn
Giang, 2010-2011). Vụ 1 (hình trái), vụ 2 (hình phải).
Hình 5-5: Chỉ số đa dạng Shannon H' của các giống ngô KNHC (Văn Giang, 2010-2011). Vụ 1 (hình trái), vụ 2 (hình phải).
5.1.3 Thành phần các loài động vật chân khớp (côn trùng và nhện)
5.1.3.1 Thành phần các loài chân khớp ghi nhận bằng điều tra trực tiếp
Qua điều tra, đã bắt gặp được tổng số 42 loài chân khớp (vụ 1) và 34 loài (vụ 2) trên các giống ngô khảo nghiệm . Các loài chân khớp bắt gặp thuộc 28 họ (vụ 1); 27 họ (vụ 2), trong 11 bộ của lớp côn trùng và lớp hình nhện. Trong đó, chúng tập trung nhiều nhất ở bộ Cánh đều: 9 loài (vụ 1); 7 loài (vụ 2) thuộc 4 họ. Các bộ Cánh cứng,

67
bộ Nhện lớn có số loài đã bắt gặp được tương ứng là 8 và 7 loài (vụ 1); 5 và 7 loài (vụ 2). Các bộ khác còn lại có số loài bắt gặp được ít hơn, mỗi bộ mới chỉ ghi nhận được từ 1 đến 4 loài (Bảng 5-3).
Số lượng loài chân khớp bắt gặp được trên các giống ngô nền không chuyển gen và giống ngô chuyển gen tương tự nhau: giống ngô nền 30B80 có 29 loài (vụ 1) và 28 loài (vụ 2), giống 30Y87 có 29 loài (vụ 1) và 30 loài (vụ 2), giống ngô chuyển gen 30Y87H có 32 loài (vụ 1) và 30 loài (vụ 2).
Như vậy, số lượng loài chân khớp bắt gặp được trên các giống ngô nền không chuyển gen và giống ngô chuyển gen tương tự nhau. Tổng số loài chân khớp bắt gặp được trong vụ 1 nhiều hơn so với vụ 2, nhưng có loài ở vụ 1 nhiều, vụ 2 ít hơn (các loài trong bộ Cánh cứng) và ngược lại (các loài trong bộ Nhện đỏ). So sánh tổng số loài trong 11 bộ ở các giống ngô khảo nghiệm trong 2 vụ không thay đổi.
Tần suất bắt gặp các loài chân khớp trên các giống ngô khảo nghiệm không
giống nhau. Vụ 1: hầu hết các loài chân khớp (khoảng 28/42 loài) bắt gặp ở mức rất ít đến ít. Có 5 loài bắt gặp ở mức độ nhiều là rệp muội ngô (Rhopalosiphum maidis), rệp muội bông (Aphis gossypii), bọ xít mù xanh (Cyrtorhinus lividipennis), bọ rùa Nhật Bản (Propylea japonica) và nhện lớn (Clubiona sp.). Có 5-6 loài bắt gặp ở mức độ trung bình là rầy xanh lá mạ (Empoasca flavescens), rầy ngực đỏ, rầy lưng trắng (Sogatella furcifera), rầy nâu (Nilaparvata lugens), dòi đục lá (Liriomyza sp.), nhện bụng nhọn (Argyrodes sp.). Riêng loài sâu đục thân ngô (Ostrinia furnacalis) bắt gặp mức độ nhiều trên các giống ngô nền không chuyển gen 30B80, 30Y87 và mức độ ít trên giống chuyển gen 30Y87H (Bảng 5-4). Vụ 2: kết quả tương tụ như vụ 1.
Trong các loài chân khớp đã ghi nhận trên ruộng ngô khảo nghiệm chỉ có 15 loài là sâu hại cây ngô (những loài c ó dấu sao (*) trong Bảng 5-4); 19 loài là thiên địch của sâu hại ngô; số loài còn lại khác là những loài trung tính hoặc trú ngụ ngẫu nhiên. Như vậy thành phần sâu hại trên tất cả các giống ngô khảo nghiệm đều nghèo nàn. Loài sâu hại rất phổ biến trên cây ngô (như sâu xám Agrotis ipsilon Hufn.) chưa bắt gặp trong ruộng khảo nghiệm). Kết quả các loài chân khớp ghi nhận giữa các công thức và giữa 2 thời vụ trồng khảo nghiệm là tương đối bằng nhau. Nguyên nhân có thể do cây ngô được khảo nghiệm trên diện tích nhỏ và bị cách ly bới nhiều loài cây trồng khác cây ngô.

61
Bảng 5-3: Số loài chân khớp ghi nhận được qua điều tra trực tiếp trên ngô KNHC (Văn Giang, 2010-2011)
TT Bộ Tổng số Giống ngô khảo nghiệm
Loài Họ 30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 Cánh thẳng (Orthoptera) 2 1 1 1 1 1 1 0 0 0
2 Cánh da (Dermapter) 2 1 1 1 1 1 1 1 1 1
3 Cánh đều (Homoptera) 9 7 4 4 8 5 8 5 9 7
4 Cánh nửa cứng (Hemiptera) 4 2 4 2 3 2 3 2 3 2
5 Cánh tơ (Thysanoptera) 1 1 1 1 1 1 1 1 1 1
6 Cánh cứng (Coleoptera) 8 5 4 3 4 4 5 5 6 4
7 Cánh vảy (Lepidoptera) 4 1 2 1 2 0 3 1 1 0
8 Hai cánh (Diptera ) 3 2 3 2 2 2 3 2 2 2
9 Cánh màng (Hymenoptera) 1 1 1 1 0 1 0 1 1 1
10 Nhện nhỏ (Acarina) 1 6 1 6 1 6 1 6 1 6
11 Nhện lớn bắt mồi (Araneae) 7 7 6 6 6 5 6 6 7 6
Tổng số 42 34 28 27 29 28 29 30 32 30

62
Bảng 5-4: Tần suất bắt gặp các loài chân khớp qua điều tra trực tiếp trên các giống ngô KNHC (Văn Giang, 2010 - 2011)
TT Loài Mức độ xuất hiện trên các giống Ngô
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
Sâu hại chủ đích 1 Sâu đục thân ngô (Ostrinia furnacalis Guen)* + + + - + + + - - - 2 Sâu ăn lá ngô Mythimna separata (Walk.)* - ++ - ++ 0 ++
Sâu hại không chủ đích 0 1 Châu chấu voi (Chondracris rosea rosea)* - - - 0 0 0 2 Châu chấu lúa Oxya sp.* - 0 - +++ - 0 3 Bọ phấn trắng (unidentified)* + +++ - +++ + +++ 4 Bọ trĩ (unidentified)* 0 +++ 0 0 0 +++ 5 Rầy điện quang (Recilia dorsalis Mots.) 0 0 0 +++ - - 6 Rệp muội ngô (Rhopalosiphum maidis Fitch)* + + + +++ + + + 0 + + + +++ 7 Rệp muội bông (Aphis gossypii Glover)* + + + 0 + + + 0 + + + 0 8 Sâu xanh (Helicoverpa armigera Hunb.)* 0 0 - 0 0 0 9 Sâu cuốn lá ngô (Cnaphalocrocis medinalis)* 0 0 - 0 + 0 Các loài thiên địch 1 Bọ đuôi kìm (Anisolabis annulipes Lucas) + - + - +++ - 2 Bọ phấn trắng (Bemisia tabaci Genn.) 0 +++ 0 - 0 +++ 3 Bọ rùa Nhật Bản (Propylea japonica Thunb.) + + + - + + + - + + + - 4 Bọ rùa 6 chấm (Menochilus sexmaculatus Fabr.) 0 - - 0 - - 5 Bọ rùa đỏ (Micraspis discolor Fabr.) 0 0 - - - 0
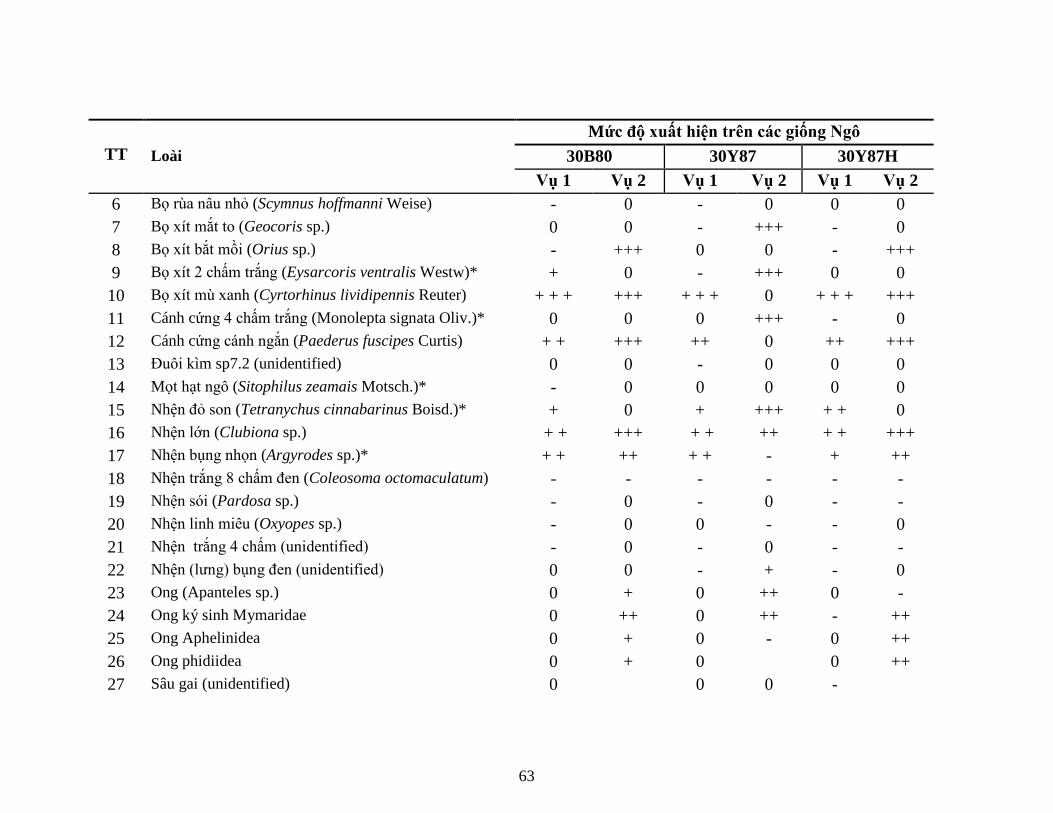
63
TT Loài
Mức độ xuất hiện trên các giống Ngô 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2 6 Bọ rùa nâu nhỏ (Scymnus hoffmanni Weise) - 0 - 0 0 0 7 Bọ xít mắt to (Geocoris sp.) 0 0 - +++ - 0 8 Bọ xít bắt mồi (Orius sp.) - +++ 0 0 - +++ 9 Bọ xít 2 chấm trắng (Eysarcoris ventralis Westw)* + 0 - +++ 0 0 10 Bọ xít mù xanh (Cyrtorhinus lividipennis Reuter) + + + +++ + + + 0 + + + +++ 11 Cánh cứng 4 chấm trắng (Monolepta signata Oliv.)* 0 0 0 +++ - 0 12 Cánh cứng cánh ngắn (Paederus fuscipes Curtis) + + +++ ++ 0 ++ +++ 13 Đuôi kìm sp7.2 (unidentified) 0 0 - 0 0 0 14 Mọt hạt ngô (Sitophilus zeamais Motsch.)* - 0 0 0 0 0 15 Nhện đỏ son (Tetranychus cinnabarinus Boisd.)* + 0 + +++ + + 0 16 Nhện lớn (Clubiona sp.) + + +++ + + ++ + + +++ 17 Nhện bụng nhọn (Argyrodes sp.)* + + ++ + + - + ++ 18 Nhện trắng 8 chấm đen (Coleosoma octomaculatum) - - - - - - 19 Nhện sói (Pardosa sp.) - 0 - 0 - - 20 Nhện linh miêu (Oxyopes sp.) - 0 0 - - 0 21 Nhện trắng 4 chấm (unidentified) - 0 - 0 - - 22 Nhện (lưng) bụng đen (unidentified) 0 0 - + - 0 23 Ong (Apanteles sp.) 0 + 0 ++ 0 - 24 Ong ký sinh Mymaridae 0 ++ 0 ++ - ++ 25 Ong Aphelinidea 0 + 0 - 0 ++ 26 Ong phidiidea 0 + 0 0 ++ 27 Sâu gai (unidentified) 0 0 0 -

64
TT Loài
Mức độ xuất hiện trên các giống Ngô 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2 28 Ruồi ký sinh sâu đục thân Tachinidae - 0 - 0 0 0 29 Ruồi đục lá (Liriomyza sp.) + + 0 + + + + + 0 30 Ruồi ăn rệp (Ischiodon scutellaris Fabr.) 0 - - - 31 Trathala sp. 0 - 0 0 - Loài trung tính và chưa rõ ý nghĩa - 1 Rầy xanh lá mạ (Empoasca flavescens Fabr.) + + + + + 0 + + + 2 Rầy lưng trắng (Sogatella furcifera Horv.) + 0 + + 0 + + 0 3 Rầy nâu nhỏ (Laodelphax striatellus Fall.) + 0 - - + 0 4 Rầy ngực đỏ (unidentified) + + - + + - + + - 5 Rầy nâu (Nilaparvata lugens Stal) + + + + + - + + - 6 Ruồi (Sepedon sp.) 0 - 0 32 0 -
Tổng số loài 29 28 32 30
Mức độ xuất hiện: - : rất ít với tần suất bắt gặp dưới 5% +: ít với tần suất bắt gặp từ 5% đến 25% + +: trung bình với tần suất bắt gặp từ 26 % đến 50% + + +: nhiều với tần suất bắt gặp >50%

65
5.1.3.2 Thành phần loài chân khớp ghi nhận bằng bẫy dính vàng
Song song với việc điều tra trực tiếp, đã tiến hành đặt bẫy dính vàng (ngày đặt
bẫy dính vàng trùng với ngày điều tra của các đợt), kết quả điều tra được ghi trong bảng 4-5. Đã bắt gặp được 40 loài chân khớp (vụ 1) và 26 loài (vụ 2), thuộc 28 họ của 8 bộ côn trùng, nhện lớn (vụ 1) và 26 họ của 9 bộ (vụ 2). Vụ 1: bộ Cánh màng có số loài vào bẫy dính vàng nhiều nhất (11 loài), vụ 2 bộ Cánh đều vào bẫy dính vàng nhiều nhất (7 loài). Tiếp theo là bộ cánh đều (vụ 1: 9 loài) và bộ cánh màng (vụ 2: 6 loài). Các bộ còn lại khác có số loài đã vào bẫy dính vàng biến động từ 1 đến 6 loài (vụ 1); 5 loài (vụ 2) .
Số loài chân khớp vào bẫy dính vàng treo ở các giống ngô khảo nghiệm không giống nhau, trên hai giống ngô nền (30B80, 30Y87) có số loài chân khớp nhiều hơn trên giống ngô chuyển gen (30Y87H). Tuy vậy, sự khác biệt này không lớn và biến động từ 32 loài trên giống ngô chuyển gen 30Y87H, đến 33-34 loài trên giống ngô nền 30Y87, 30B80 (vụ 1). Ở vụ 2, từ 19 loài trên giống 30B80 đến 22-24 loài trên giống 30Y80H và 30Y80.
Những loài có kích thước lớn (châu chấu voi, châu chấu lúa,...) không thu nhận được trên bẫy dính vàng. Một số loài vừa quan sát thấy trên cây ngô khi điều tra trực tiếp vừa vào bẫy dính vàng (rầy lưng trắng Sogatella furcifera, rệp muội ngô Rhopalosiphum maidis, bọ rùa Nhật Bản Propylea japonica, cánh cứng ngắn Paederus fuscipes,...). Tuy nhiên, có nhiều loài có kích thước nhỏ khó nhìn thấy bằng mắt thường khi điều tra trực tiếp (ong ký sinh thuộc họ Aphenlinidae, Eucyrticidae, Eulophidae, Mymaridae, Scelionidae). Hầu như không thấy cá thể trưởng thành của các loài thuộc bộ cánh vảy vào bẫy dính vàng (Bảng 5-5).

66
Bảng 5-5: Số loài chân khớp vào bẫy dính vàng trên các giống ngô KNHC (Văn Giang, 2010-2011)
TT
Bộ
Tổng số Số loài trên bẫy dính vàng Loài Họ 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 Cánh da Dermapter 1 1 1 1 - 0 1 1 - 1 2 Cánh đều Homoptera 9 7 4 7 8 5 8 5 8 7 3 Cánh nửa cứng Hemiptera 3 2 3 2 3 1 3 2 3 1
4 Cánh tơ Thysanoptera 1 1 1 1 1 1 1 1 1 1 5 Coleoptera-Cánh cứng 5 4 2 2 4 3 4 4 5 3 6 Cánh màng Hymenoptera 11 6 9 5 9 6 8 6 7 5 7 Cánh vảy Lepidoptera 0 1 0 1 0 2 0 2 0 2 8 Hai cánh Diptera 4 1 3 1 4 0 4 1 3 0 9 Nhện lớn bắt mồi Araneae 6 3 5 3 5 2 4 3 2 3
Tổng số 40 26 28 20 34 19 33 24 29 22

67
Bảng 5-6: Thành phần loài chân khớp vào bẫy dính vàng trên các giống ngô KNHC (Văn Giang, 2010 - 2011)
TT
Loài
30B80 30Y87 30Y87H VỤ 1 VỤ 2 VỤ 1 VỤ 2 VỤ 1 VỤ 2
Các loài sâu hại 1 Bọ phấn trắng Bemisia tabaci Genn. * 0 X 0 X 0 X 2 Bọ trĩ (unidentified) * X X X 3 Rầy nâu nhỏ Laodelphax striatellus (Fall.)* X 0 X 0 X 0 4 Rầy lưng trắng Sogata furcifera Horv* X 0 X 0 X X 5 Rầy điện quang Recilia dorsalis (Mots.) * 0 0 X 0 X X 6 Rầy xanh đuôi đen Nephotettix nigropictus (Stal) X 0 0 0 0 0 7 Rệp muội ngô Rhopalosiphum maidis Fitch * X X X X X X 8 Sâu ăn lá ngô Mythimna separata (Walk.) * 0 0 0 X 0 0
Các loài thiên địch 1 Cánh cứng ngắn Paederus fuscipes Curtis X 0 X 0 X 0 2 Bọ đuôi kìm Anisolabis annulipes (Lucas) 0 0 X X 0 X 3 Bọ phấn (unidentified) X X X 4 Bọ rùa 6 chấm Menochilus sexmaculatus (Fabr.) X X X X X X 5 Bọ rùa nhật bản Propylea japonica (Thunb.) X X X X X X 6 Bọ rùa nâu nhỏ Scymnus hoffmanni Weise 0 0 0 0 X 0 7 Bọ rùa đỏ Micraspis discolor (Fabr.) X 0 X 0 X 0 8 Bọ trĩ (unidentified) * X X X X X X 9 Bọ xít bắt mồi Orius sp. X 0 X X X 0 10 Bọ xít mắt to Geocoris sp. X 0 X 0 X 0 11 Bọ xít mù xanh Cyrtorhinus lividipennis Reuter X X X X X X

68
TT
Loài
30B80 30Y87 30Y87H VỤ 1 VỤ 2 VỤ 1 VỤ 2 VỤ 1 VỤ 2
12 Nhện lưới Araneus sp. X X X X X 0 13 Nhện lớn hàm to (unidentified) X X X X X X 14 Nhện bụng đen (unidentified) X 0 X 0 0 0 15 Nhện 4 chấm (unidentified) X 0 0 0 0 0 16 Nhện sói Pardosa sp. X 0 0 0 0 0 17 Nhện linh miêu Oxyopes sp. 0 X X X 0 X 18 Ong Apanteles sp. 0 X 0 X 0 X 19 Ong Trathala sp. 0 X 0 X 0 0 20 Ong kén đèn lồng (Trathala) X 0 0 0 0 0 21 Ong kén trắng Apanteles sp. X 0 X 0 X 0 22 Ong Braconidae (unidentified) X 0 X 0 X 0 23 Ong ký sinh rệp muội Lysiphlebia sp. X X X X X X 24 Ong Aphenlinidae (unidentified) X X X X X X 25 Ong Eucyrticidae (unidentified) X 0 0 0 0 0 26 Ong Eulophidae (unidentified) 0 0 X 0 0 0 27 Ong Mymaridae 1 (unidentified) X 0 X 0 X 0 28 Ong Mymaridae 2 (unidentified) X X X 0 X 0 29 Ong ks1 (unidentified) X X X X X X 30 Ruồi Agromizidae (unidentified) X 0 X 0 X 0 31 Ruồi đục lá Liriomyza sp. X 0 X 0 0 0 32 Ruồi ký sinh sâu đục thân Tachinidae X 0 X 0 X 0 Loài trung tính 1 Rầy nâu Nilaparvata lugens Stal X X X X X X
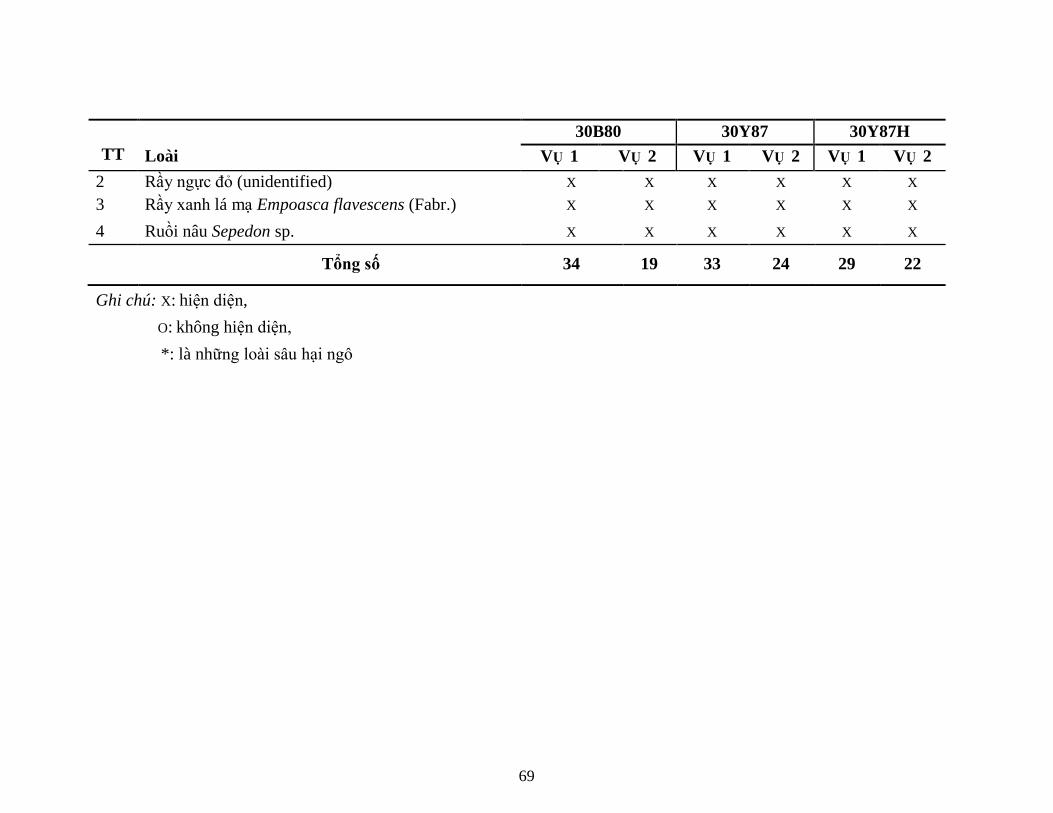
69
TT
Loài
30B80 30Y87 30Y87H VỤ 1 VỤ 2 VỤ 1 VỤ 2 VỤ 1 VỤ 2
2 Rầy ngực đỏ (unidentified) X X X X X X 3 Rầy xanh lá mạ Empoasca flavescens (Fabr.) X X X X X X 4 Ruồi nâu Sepedon sp. X X X X X X
Tổng số 34 19 33 24 29 22
Ghi chú: X: hiện diện, O: không hiện diện, *: là những loài sâu hại ngô

70
5.1.3.3 Thành phần loài chân khớp bắt gặp bằng 2 phương pháp (điều tra trực tiếp
và bẫy dính vàng)
Bằng cả hai phương pháp điều tra trực tiếp và bẫy dính màu vàng đã ghi nhận
trên ruộng ngô khảo nghiệm có 57 loài chân khớp. Chúng thuộc 36 họ của 11 bộ c ôn trùng và nhện. Trong đó, bộ cánh màng có số loài phát hiện được nhiều nhất (11 loài), sau đó là các bộ cánh đều với 10 loài và bộ nhện lớn 9 loài, bộ cánh cứng 8 loài. Những bộ còn lại khác có số loài đã ghi nhận được ít hơn, biến động từ 1 đến 5 loài (Bảng 5-7).
Số lượng loài đã ghi nhận được bằng cả hai phương pháp trên lần lượt các giống 30B80, 30Y87 và 30Y87H gần tương tự nhau là 56, 47 và 44 loài (vụ 1); 28, 30 và 30 (vụ 2) trên giống ngô chuyển gen 30Y87H có số loài ghi nhận được ít hơn 2 g iống nền (vụ 1) nhưng lại nhiều hơn và bằng 2 giống nền ở vụ thứ 2. Tuy nhiên sự khá c nhau này không nhiều ( Bảng 5-7). Trong các loài chân khớp đã ghi nhận được trên ruộng ngô khảo nghiệm, có 15 loài sâu hại cây ngô (những loài có dấu sao (*) trong bảng 4-7), 32 loài là thiên địch và 7 loài trung tính hoặc trú ngụ ngẫu nhiên (vụ 1). Vụ 2, có 6 loài sâu hại cây ngô, 16 loài là thiên địch và 4 loài trung tính hoặc trú ngụ ngẫu nhiên.
Như vậy, số loài ghi nhận được bằng cả 2 phương pháp (điều tra trực tiếp và bẫy dính vàng) trên các giống ngô khảo nghiệm trong cùng một vụ tương đương nhau. Nhưng số liệu này ở vụ 2 thấp hơn vụ 1, do điều kiện thời tiết khí hậu ở vụ 2 (rét đậm – ghi trong phần phụ lục) bất thuận cho một số loài côn trùng hoạt động

71
Bảng 5-7: Tần suất bắt gặp các loài chân khớp qua điều tra trực tiếp và bẫy dính vàng trên ruộng ngô trong KNHC (Văn Giang, 2010 - 2011)
TT
Loài 30B80 30Y87 30Y87H
VỤ 1 VỤ 2 VỤ 1 VỤ 2 VỤ 1 VỤ 2
Các loài sâu hại chủ đích 1 Sâu đục thân ngô Ostrinia furnacalis Guen.* - - - - 0 - 2 Sâu ăn lá ngô Mythimna separata (Walk.)* 0 ++ 0 ++ 0 ++ 3 Sâu xanh Helicoverpa armigera (Hunb.) - 0 0 0 0 0 4 Sâu cuốn lá ngô Cnaphalocrocis medinalis Gueen.* 0 0 - 0 0 0 Các loài sâu hại không chủ đích
1 Châu chấu voi Chondracris rosea rosea (De Geer) - - 0 0 0 0 2 Rầy lưng trắng Sogatella furcifera (Horv.) ++ 0 ++ 0 ++ 0 3 Rầy nâu nhỏ Laodelphax striatellus (Fall.) + - + - + - 4 Rệp muội ngô Rhopalosiphum maidis Fitch* +++ +++ +++ +++ +++ +++ 5 Rệp muội bông Aphis gossypii Glover* ++ ++ ++ ++ ++ + 6 Bọ phấn trắng Bemisia tabaci Genn.* ++ ++ ++ + ++ + 7 Bọ xít hai chấm trắng Eysarcoris ventralis Westw. * + 0 + 0 + 0 8 Bọ trĩ (unidentified) * + +++ + +++ + +++ 9 Cánh cứng 4 chấm trắng Monolepta signata Oliv. 0 0 0 0 - 0 10 Sâu gai (unidentified) 0 0 0 0 + 0 11 Mọt hạt ngô Sitophilus zeamais Motsch.* + 0 + 0 + 0 12 Nhện đỏ son Tetranychus cinnabarinus Boisd.* + 0 + 0 + 0 13 Ruồi đục lá Liriomyza sp. + 0 + 0 + 0
Các loài thiên địch

72
TT
Loài
30B80 30Y87 30Y87H VỤ 1 VỤ 2 VỤ 1 VỤ 2 VỤ 1 VỤ 2
1 Bọ đuôi kìm Anisolabis annulipes (Lucas) + - + - + - 2 Đuôi kìm sp7.2 (unidentified) 0 0 - 0 0 0 3 Bọ xít mù xanh Cyrtorhinus lividipennis Reuter +++ ++ +++ ++ +++ ++ 4 Bọ xít mắt to Geocoris sp. +++ 0 +++ 0 +++ 0 5 Bọ xít bắt mồi Orius sp. +++ +++ +++ +++ +++ +++ 6 Cánh cứng cánh ngắn Paederus fuscipes Curtis +++ +++ +++ +++ +++ +++ 7 Bọ rùa Nhật Bản Propylea japonica (Thunb.) +++ - +++ - +++ - 8 Bọ rùa 6 chấm Menochilus sexmaculatus (Fabr.) ++ - ++ - ++ - 9 Bọ rùa đỏ Micraspis discolor (Fabr.) ++ 0 ++ 0 ++ 0 10 Bọ rùa nâu nhỏ Scymnus hoffmanni Weise ++ 0 ++ - ++ 0 11 Ruồi ký sinh sâu đục thân Tachinidae - 0 - 0 - 0 12 Ruồi ăn rệp Ischiodon scutellaris Fabr. 0 0 - 0 - 0 13 Ong (Trathala) + 0 0 0 0 0 14 Ong kén trắng Apanteles sp. + 0 + 0 + 0 15 Ong Braconidae (unidentified) + 0 + 0 + 0 16 Ong ký sinh rệp muội Lysiphlebia sp. + 0 + 0 + 0 17 Ong Aphenlinidae (unidentified) + 0 + 0 + 0 18 Ong Eucyrticidae (unidentified) + 0 0 0 0 0 19 Ong Eulophidae (unidentified) 0 0 + 0 0 0 20 Ong Mymaridae 1 (unidentified) + ++ + ++ + ++ 21 Ong Mymaridae 2 (unidentified) + + + + + + 22 Ong Scelionidae (unidentified) 0 - 0 - 0 0 23 Ong ks1 (unidentified) + ++ + ++ + ++

73
TT
Loài
30B80 30Y87 30Y87H VỤ 1 VỤ 2 VỤ 1 VỤ 2 VỤ 1 VỤ 2
24 Nhện lưới Araneus sp. + - + 0 + 0 25 Nhện lớn Clubiona sp. +++ +++ +++ +++ +++ +++ 26 Nhện bụng nhọn Argyrodes sp. - ++ + ++ - ++ 27 Nhện trắng 8 chấm đen Coleosoma octomaculatum
(Boes. Et Strand) + - + - - -
28 Nhện sói Pardosa sp. + + - - - - 29 Nhện linh miêu Oxyopes sp. ++ + ++ - ++ - 30 Nhện trắng 4 chấm (unidentified) + 0 + - + - 31 Nhện (lưng) bụng đen (unidentified) - 0 - 0 - 0 32 Nhện lớn hàm to (unidentified) + 0 + 0 + 0
Loài trung tính và chưa rõ ý nghĩa 1 Rầy xanh lá mạ Empoasca flavescens (Fabr.) ++ + ++ - ++ + 2 Rầy xanh đuôi đen Nephotettix nigropictus (Stal) + 0 0 0 0 0 3 Rầy điện quang Recilia dorsalis (Mots.) 0 0 + 0 + 0 4 Rầy ngực đỏ (unidentified) + - + - + - 5 Rầy nâu Nilaparvata lugens Stal + + + - + - 6 Ruồi Agromizidae (unidentified) + 0 + 0 + 0 7 Ruồi nâu Sepedon sp. + - + - + - Tổng 56 28 47 30 44 30
Mức độ xuất hiện: - : rất ít với tần suất bắt gặp dưới 5% +: ít với tần suất bắt gặp từ 5% đến 25% + +: trung bình với tần suất bắt gặp từ 26 % đến 50% + + +: nhiều với tần suất bắt gặp >50%

74
5.1.3.4 Mật độ các loài chân khớp phổ biến
Bằng phương pháp điều tra trực tiếp đã ghi nhận được 18 loài thiên địch (vụ 1 ), 11 loài (vụ 2) của sâu hại trong tổng số 29 loài chân khớp (vụ 1), 20 loài (vụ 2) (Bảng 5-3). Trong đó, các loài thiên địch phổ biến bao gồm bọ rùa Nhật Bản (Propylea japonica (Thunb.)), bọ xít mù xanh (Cyrtorhinus lividipennis Reuter), nhện lớn (Clubiona sp), Bọ cánh cứng cánh ngắn (Paederus fuscipes)
a) Bọ rùa Nhật Bản (Propylea japonica)
Bọ rùa đỏ Nhật Bản là loài thiên địch phổ biến trên các giống ngô khảo nghiệm.
Vụ 1: bắt gặp trong tất cả các kỳ điều tra, trên cả 3 giống ngô khảo nghiệm. Tuy nhiên, mật độ bọ rùa Nhật Bản trên 3 giống ngô khảo nghiệm đạt không cao, biến động từ 0,02 con/cây ở giống ngô 30Y87 đến 0,92 con/cây ở giống ngô 30B80. Mật độ của loài này có xu hướng gia tăng theo thời gian sinh trưởng phát triển của cây ngô. Mật độ quần thể của nó đạt cao nhất trên các giống ngô quan sát được vào ngày 12/10/2010 (giai đoạn R 3
Vụ 2: Giống 30B80, chỉ bắt gặp ở lần điều tra thứ nhất. Giống 30Y87 bắt gặp ở lần điều tra thứ 1,2 và 5. Giống chuyển gen 30Y87H bắt gặp ở lần điều tra thứ nhất và thứ 2. Như vậy, mật độ của bọ rùa đỏ Nhật Bản trên các giống ngô khảo nghiệm trong cùng 1 vụ tương tự nhau, vụ 1 nhiều hơn vụ 2 (Bảng 5-8).
) và biến động từ 0,73 con/cây ở giống ngô không chuyển gen 30Y87 đến 0.88 con/cây ở giống ngô chuyển gen 30Y87H.
Bảng 5-8: Mật độ bọ rùa đỏ Nhật Bản trên các giống ngô (con/cây) trong KNHC
(Văn Giang, 2010 - 2011) Đợt điều
tra 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 0,05 ± 0,03 0,05±0,00 0,02 ± 0,02 0,02±0,02 0,05 ± 0,02 0,02±0,02 2 0,37 ± 0,14 0,00 0,13 ± 0,04 0,02±0,02 0,13 ± 0,03 0,02±0,02 3 0,18 ± 0,09 0 0,18 ± 0,09 0 0,17 ± 0,07 0 4 0,20 ± 0,06 0 0,03 ± 0,03 0 0,18 ± 0,08 0 5 0,92 ± 0,44 0,00 0,73 ± 0,36 0,02±0,02 0,88 ± 0,26 0,00

75
b) Bọ xít mù xanh Cyrtorhinus lividipennis Bọ xít mù xanh là một trong số ít các loài thiên địch được coi là phổ biến trên
ruộng ngô khảo nghiệm, nhưng không phải đều bắt gặp trong tất cả các kỳ điều tra. Kỳ điều tra đầu tiên (giai đoạn V8, vụ 1), kỳ điều tra thứ 3, 4 và 5 (vụ 2) không bắt gặp bọ xít mù xanh trong tất cả các điểm điều tra. Tại những điểm điều tra bắt gặp bọ xít mù xanh với mật độ không cao. Mật độ của bọ xít mù xanh trên các giống ngô khảo nghiệm trong cùng 1 vụ tương tự nhau, vụ 2 ít hơn vụ 1 (Bảng 5-9).
Bảng 5-9: Mật độ bọ xít mù xanh trên các giống ngô (con/cây) trong KNHC
(Văn Giang, 2010 -2011)
Đợt điều tra
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 0 0,03 ± 0,03 0 0,02 ± 0,02 0 0 2 0,02 ± 0,02 0,53 ± 0,29 0 0,52 ± 0,26 0 0,58 ± 0,35 3 0,28 ± 0,12 0 0,15 ± 0,09 0 0,13 ± 0,06 0 4 0 0 0 0 0,10 ± 0,04 0 5 0,27 ± 0,06 0 0,23 ± 0.07 0 0,25 ± 0,09 0
c) Nhện lớn (Clubiona sp.)
Nhện lớn (Clubiona sp.) là loài nhện lớn bắt mồi phổ biến, bắt gặp trong hầu hết ở các kỳ điều tra trên 3 các giống ngô khảo nghiệm. Tuy vậy, mật độ của nhện lớn Clubiona sp. trên ngô khảo nghiệm không cao.
Vụ 1: Mật độ của nhện lớn (Clubiona sp.) có xu hướng giảm theo thời gian sinh trưởng phát triển của cây ngô, Mật độ ở kỳ điều tra thứ 1 (giai đoạn V 8) đạt cao nhất và biến động trong khoảng 0,27 - 0,43 con/cây. Đến kỳ điều tra cuối cùng (giai đoạn R3
Vụ 2: giống 30B80, mật độ nhện lớn đạt từ 0,33 – 0,58 con/cây. Giống 30Y87, mật độ đạt từ 0,22 – 0,73 con/cây. Giống 30Y87H, mật độ từ 0,15 – 0,70 con/cây. Trên cả 3 giống, mật độ nhện lớn tăng giảm trong các lần điều tra không theo quy luật.
), mật độ của loài này giảm rõ ràng và chỉ biến động trong khoảng 0,02 - 0,13 con/cây.
Như vậy, mật độ nhện lớn trên các giống ngô khảo nghiệm khác nhau, tương tự nhau ở các lần điều tra. Ởvụ 2, mật độ nhện lớn cao hơn so với vụ 1 (Bảng 5-10).

76
Bảng 5-10: Mật độ nhện lớn trên các giống ngô (con/cây) trong KNHC
(Văn Giang, 2010-2011)
Đợt điều tra
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 0,27 ± 0,04 0,47 ± 0,24 0,32 ± 0,15 0,73 ± 0,37 0,43 ± 0,16 0,70 ± 0,08 2 0 0,45 ± 0,29 0,07 ± 0,04 0,40 ± 0,21 0,30 ± 0,00 0,15 ± 0,15 3 0,03 ± 0,02 0,55 ± 0,35 0,12 ± 0,06 0,45 ± 0,35 0,08 ± 0,03 0,33 ± 0,23 4 0,08 ± 0,08 0,58 ± 0,39 0,05 ± 0,05 0,22 ± 0,07 0,17 ± 0,12 0,17 ± 0,14 5 0,08 ± 0,04 0,33 ± 0,17 0,03 ± 0,02 0,30 ± 0,15 0,13 ± 0,13 0,18 ± 0,16
d) Bọ cánh cứng cánh ngắn (Paederus fuscipes )
Bọ cánh cứng cánh ngắn (Paederus fuscipes) là loài thiên địch xuất hiện trên các giống ngô khảo nghiệm với mật độ cao nhất trong các loài bắt mồi. Mật độ của nó đạt khá cao trên các giống ngô khảo nghiệm.
Vụ 1: mật độ biến động từ 0,07 đến 2,92 con/cây trên giống ngô 30B80; từ 0 – 1,92 con/cây (30Y80) và từ 0,10 đến 2,43 con/cây trên giống ngô chuyển gen 30Y87H.
Vụ 2: mật độ bọ cánh cứng ngắn đạt từ 1,13 – 1,97 con/cây (30B80); từ 0,8 -2,03 con/cây (30Y80) và 0,77 – 2,25 con/cây.
Vậy, trong cùng một vụ trồng, mật độ bọ cánh cứng cánh ngắn trên các giống ngô khảo nghiệm gần tương tự nhau. 2 vụ trồng khác nhau mật độ chúng khác nhau, cuối vụ 1 mật độ tăng cao hơn đầu vụ và tăng tương đối ổn định trong vụ 2 (Bảng 5-11). Bảng 5-11: Mật độ Bọ cứng cánh ngắn trên các giống ngô (con/cây) trong KNHC
(Văn Giang, 2010-2011)
Đợt điều tra
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 0,07 ± 0,07 1,30 ± 0,38 0,08 ± 0,04 1,85 ± 0,53 0,35 ± 0,03 1,28 ± 0,15 2 0,50 ± 0,31 1,13 ± 0,38 0 0,80 ± 0,23 0,10 ± 0,03 0,77 ± 0,19 3 0,18 ± 0,07 1,97 ± 0,16 0,28 ± 0,18 2,03 ± 0,64 0,20 ± 0,05 1,82 ± 0,19 4 0,43 ± 0,19 1,95 ± 0,19 0,42 ± 0,30 1,75 ± 0,31 0,25 ± 0,18 2,08 ± 0,16 5 2,92 ± 0,63 1,70 ± 0,03 1,92 ± 1,09 1,92 ± 0,13 2,43 ± 0,10 2,25 ± 0,08

77
5.1.4 Kết quả đánh giá sự đa dạng của sinh vật đất
5.1.4.1 Đánh giá đa dạng sinh vật đất bằng phương pháp thu mẫu đất
Đã thực hiện 3 đợt thực địa thu mẫu định lượng bọ đuôi bật (Collembola). Kết quả phân tích về thành phần loài, phân bố của bọ đuôi bật (Collembola) được trình bày ở Bảng 5-12. Trên đất trồng ngô chuyển gen và đất trồng ngô không chuyển gen (đối chứng) đã ghi nhận được 34 loài thuộc 19 giống của 10 họ. Gần 1/2 số loài tập trung ở họ Entomobryidae (18/35 loài, chiếm 51,43% tổng số loài). Giống Lepidocyrtus có nhiều loài nhất (10/35 loài, chiếm 28,57% tổng số loài). Các giống còn lại chỉ có từ 1 – 3 loài
Trong thành phần loài bọ đuôi bật Collembola có nhiều loài thuộc nhóm sống ở trong đất (Acherontiella sabina, Mesaphorura krasbaueri, Proisotoma submuscicola, Megalothorax minimus, Deuterosminthurus sp.1, Deuterosminthurus sp.2, … , một vài loài sống ở bên dưới lớp thảm, trên mặt đất: một số loài thuộc họ Entomobryidae (Entomobrya lanuginosa, Lepidocyrtus (L.) concolourus, L. (Ascocyrtus) sp.4
Trong số 34 loài bọ đuôi bật Collembola nêu trên, có 20 loài có mặt cả ở lô trồng ngô đối chứng và lô trồng ngô chuyển gen, trong đó có 6 loài là gặp phổ biến, bao gồm: Isotomurus punctiferus, Entomobrya lanuginosa, Cyphoderus javanus, Sminthurides bothrium, Isotomurus prasinus, Lepidocyrtus (Asc.) sp.
, …). Có ít loài sống ở trong và bên trên lớp thảm như Brachystomella parvula, Homidia glassa, Cyphoderus javanus,… .
4. Chỉ gặp ở đất trồng ngô đối chứng có 6 loài: Xenylla humicola, Ceratophysella denticulata, Entomobrya muscorum, Homidia subcingula, Willowsia mesothoraxa, Lepidocyrtus (Asc.) dahlii, Megalothorax minimus. Trong số 6 loài này, 4 loài Xenylla humicola, Ceratophysella denticulata, Homidia subcingula, Megalothorax minimus là tương đối phổ biến ở đất trồng màu, đất vườn quanh nhà. Còn Entomobrya muscorum, Willowsia mesothoraxa ít gặp ở đất sản xuất nông nghiệp, hơn nữa số lượng của chúng rất ít (Entomobrya muscorum và Willowsia mesothoraxa mỗi loài chỉ có 1 cá thể), vì vậy đây chỉ là trường hợp ngẫu nhiên bắt gặp. Với đất trồng ngô chuyển gen, cũng có 8 loài không gặp ở đất trồng ngô đối chứng như: Acherontiellina sabina, Friesea sublimis, Proisotoma submuscicola, Lepidocyrtus (L.) simsim, Lepidocyrtus (L.) sp.5, Lepidocyrtus (Acr.) transiens, Dicranocentrus indicus, Salina sp.14 – đây cũng là những loài khá phổ biến trong các sinh cảnh đất nông nghiệp, trảng cỏ, vườn quanh nhà (Nguyễn Thị Thu Anh và cs., 2000; 2008; 2009; 2010; Nguyễn Trí Tiến và

78
cs., 2009). Chúng không bắt gặp ở đất trồng ngô đối chứng hay đất trồng ngô chuyển gen chỉ có thể do số lượng mẫu hoặc thời gian nghiên cứu còn ít, không phải do giống cây trồng gây nên.
So sánh thành phần loài bọ đuôi bật (Collembola )giữa đất trồng ngô đối chứng và đất trồng ngô chuyển gen không thấy có sự sai khác ngoài phần phân tích nêu trên về các loài bọ đuôi bật (Collembola) phân bố đơn lẻ, chỉ có ở 1 trong 2 nền đất khảo nghiệm.
5.1.4.2 Một số chỉ số định lượng của bọ đuôi bật Collembola ở đất trồng KNHC ngô
chuyển gen (thu bằng phương pháp lấy mẫu đất)
Phân tích so sánh giá trị một số chỉ số định lượng của Collembola được trình bày ở Bảng 4-13. Kết quả phân tích cho thấy: Mật độ loài bọ đuôi bật giữa 2 nền đất trồng ngô đối chứng và ngô chuyển gen không có sự sai khác. Điều này cũng giống với kết quả nghiên cứu của các tác giả Mohammad A. Al-Deeb (2003). Đất trồng ngô chuyển gen chỉ hơn kém 1 loài so với đất trồng ngô đối chứng.
- Giá trị của chỉ số mật độ, chỉ số đa dạng H’ và chỉ số đồng đều J’ ở đất trồng ngô chuyển gen lúc tăng, lúc giảm so với đất trồng ngô đối chứng. Với giống ngô chuyển gen 30Y87H có giá trị mật độ trung bình (nghìn con/m 2
- Số lượng loài Collembola giữa 2 nền đất trồng ngô đối chứng và ngô biến đổi gen không có sự sai khác. Kết quả này giống với kết quả nghiên cứu của tác giả Mohammad A. Al-Deeb (2003).
) cao hơn chút ít so với ngô đối chứng cùng giống 30Y87, nhưng giá trị chỉ số đa dạng H’ và chỉ số đồng đều J’ lại thấp hơn. Tuy nhiên, qua kết quả tính thống kê, sự tăng giảm của các chỉ số này giữa ngô đối chứng và ngô chuyển gen, giữa vụ 1 và vụ 2 là không đáng kể.
So sánh thống kê các chỉ số định lượng bọ đuôi bật Collembola giữa 2 nền đất, trong Bảng 4-13 cho thấy không có sự sai khác có ý nghĩa về độ phong phú (F4,07=0,11; P>0,05) và độ đa dạng loài bọ đuôi bật Collembola giữa đất trồng ngô chuyển gen và đất trồng ngô không chuyển gen (D0,34=0,08; p>0,05) ở Trạm thực nghiệm Văn Giang, Hưng Yên.

79
Bảng 5-12: Thành phần loài bọ đuôi bật (Collembola) trong đất trồng ngô KNHC (Văn Giang, 2010 – 2011)
TT
Loài
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
Hypogastruridae 1 Ceratophysella denticulata (Bagnall, 1941) 5-dt 2 2 0 dt 1 2 Xenylla humicola (Fabricius, 1780) 23 1- dt 187 16 Onychiuridae
3 Mesaphorura krausbaueri (Borner, 1901) 2 1 0 0 1 0 Neanuridae
4 Brachystomella parvula (Schaffer, 1896) 13 23 0 140 4-dt 75 5 Friesea sublimis (Tullberg, 1871) 16-dt 4 6 Pseudachorutes subcrasus (Tullberg, 1871) 0 1 2 2 0 1 Isotomidae
7 Isotomurus prasinus (Reuter, 1891) 52-dt 60 91-dt 25 15-dt 38 8 Isotomurus punctiferus (Yosii, 1963) 59-dt 1 9 Folsomina onychiurina (Denis, 1931) 0 0 0 2 0 0 10 Proisotoma submuscicola (Nguyen,1995) 0 0 0 2 0 0
Entomobryidae 11 Dicranocentrus indicus (Bonet, 1930) 22 0 0 0 47 0 12 Entomobrya lanuginosa (Nicolet, 1841) 1 7 23 3 0 3 13 Entomobrya muscorum (Nicolet, 1841) 0 0 0 0 6 0 14 Entomobrya sp. 8 2 1 1 0 22 0 15 Homidia glassa (Nguyen, 2001) 2 14 7 26 2 29 16 Homidia socia (Denis, 1929) 1 0 4 0 1-dt 0

80
TT
Loài
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
17 Homidia subcingula (Denis, 1948) 0 0 1-dt 0 0 0 18 Lepidocyrtus (L.) sp. 4 5 0 0 1 4 2 19 Lepidocyrtus (Cyn.) sp. 0 1 3 10 1 0 1 20 Lepidocyrtus (Acr.) heterolepis (Yosii, 1959) 1 0 5 0 1 0 21 Lepidocyrtus (Acr.) transiens (Yosii, 1982) 0 0 0 1 4 2 22 Lepidocyrtus (Acr.) sp. 0 7 0 3 0 0 0 23 Lepidocyrtus (Asc.) cf. aseanus (Yosii, 1982) 0 0 1 0 1 0 24 Lepidocyrtus (Asc.) concolourus (Nguyen, 2001) 5-dt 0 12-dt 1 4-dt 0 25 Lepidocyrtus (Asc.) dahlii (Schaffer, 1898) 0 0 1 0 0 0 26 Lepidocyrtus (Asc.) sp. 4 4 17 27 27 Willowsia mesothoraxa (Nguyen, 2001) 1 0 0 0 0 0
Cyphoderidae 28 Cyphoderus javanus (Borner, 1906) 23-dt 68-dt 52-dt
Paronellidae 29 Salina sp. 0 14 0 1-dt 0 1-dt 0 30 Megalothorax minimus (Willem, 1900) 4 0 4 0 0 0 31 Sminthurides aquaticus (Bourlet, 1842) 8 0 4 0 7 0 32 Sminthurides bothrium (Nguyen, 2001) 106 0 39 0 69 0
Bourletiellidae 33 Deuterosminthurus pallipes (Bourlet, 1843) 5 0 1 0 8 0 34 Deuterosminthurus sp. 1-dt 1 0 0 0 5-dt 0
Chú thích: dt - loài thu được bằng phương pháp ống hút

81
Bảng 5-13: So sánh một số chỉ số định lượng của bọ đuôi bật (Collembola) trong đất trồng ngô KNHC (Văn Giang, 2010 – 2011)
Chỉ số 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
Số loài 22 17 22 16 23 16
Mật độ (nghìn con/m2 9,13±2,61 ) 11,47±2,89 8,24±2,84 15,25±5,33 10,96±2,80 10,19±2,22
Chỉ số ĐD H’ 2,025 1,97 2,297 1,91 2,026 2,00
Chỉ số ĐĐ J’ 0,655 0,69 0,743 0,69 0,646 0,72
Chú thích: ĐD: đa dạng; ĐĐ: đồng đều

82
5.1.4.3 Đánh giá sự đa dạng sinh vật đất bằng phương pháp bẫy cốc (pitfall trap)
Thành phần loài và phân bố của bọ đuôi bật Collembola trong các lô thí nghiệm trồng ngô khảo nghiệm (thu bằng phương pháp pitfall trap)
Đã ghi nhận được 34 loài bọ đuôi bật Collembola thuộc 16 giống, 9 họ phân bố trong khu vực trồng ngô khảo nghiệm ở Văn Giang, Hưng Yên (Bảng 5-14). Trong số đó, 27 loài đã xác định tên khoa học, 7 loài còn ở dạng sp. (chỉ mới xác định đến chi). Số loài tập trung ở họ Entomobryidae (14 loài, chiếm 41,18% tổng số loài). 8 họ còn lại chỉ có từ 1 đến 4 loài. Giống Lepidocyrtus có số loài nhiều nhất (10 loài, chiếm 71,43% số loài của họ và chiếm 41,18% tổng số loài). 15 giống còn lại có từ 1-4 loài.
Phần lớn các loài bọ đuôi bật Collembola trong danh sách (Bảng 5-14) thuộc nhóm sống ở lớp thảm và ở dưới lớp thảm - trên mặt đất như: Xenylla humicola (Hypogastruridae), Brachystomella parvula; Friesea sublimis (Neanuridae); một số loài thuộc giống Lepidocyrtus: Lepidocyrtus (Acr.) heterolepis, L. (Acr.) transiens, L. (Asc.) concolourus, L. (Asc.) sp.4,… (Entomobryidae); Cyphoderus javanus (Cyphoderidae); một số loài thuộc giống Salina: Salina celebensis, Salina okinawana, Salina pictura, Salina sp.14; một số loài Symphypleonid: Sminthurides bothrium, Deuterosminthurus pallipes, Deuterosminthurus sp.1, Deuterosminthurus sp.2
Trong số 34 loài bọ đuôi bật Collembola, có tới 25 loài (chiếm 73,53% tổng số loài) có mặt ở cả lô thí nghiệm trồng ngô đối chứng và ngô chuyển gen. Chỉ có 8 loài là có mặt trong các lô thí nghiệm trồng hoặc ngô chuyển gen hoặc ngô đối chứng (không chuyển gen), cụ thể: chỉ có mặt ở lô trồng ngô không chuyển gen – Friesea sublimis (1 cá thể ở ngô 30Y87), Cryptopygus thermophilus (3 cá thể, trong đó 2 cá thể ở ngô 30Y87 và 1 cá thể ở ngô 30B80), Lepidocyrtus (Asc.) cf. aseanus (5 cá thể, trong đó 2 cá thể ở ngô 30Y87 và 3 cá thể ở ngô 30B80), Willowsia mesothoraxa (1 cá thể ở ngô 30B80)), Salina pictura (3 cá thể, trong đó 1 cá thể ở ngô 30Y87 và 2 cá thể ở ngô 30B80); chỉ có mặt ở lô trồng ngô chuyển gen, gồm 3 loài: Lepidocyrtys (L.) sp.
,….
5 (1 cá thể ở ngô 30Y87H). Trong 8 loài Bọ đuôi bật Collembola nêu trên, Lepidocyrtus (Asc.) cf. aseanus, Lepidocyrtus (Asc.) dahlii, Salina pictura thường sống trong lớp thảm và gặp ở những sinh cảnh mở. Chúng gặp khá phổ biến trong các dạng sinh cảnh khác nhau ở những địa phương khác nhau của Việt Nam (ở VQG Xuân Sơn, Phú Thọ; khu BTTN Thượng Tiến, Kim Bôi, Hòa Bình; đất canh tác Gia Lâm, Hà Nội; khu vực phía Bắc các tỉnh Na Rì, Sơn La,…; Cryptopygus thermophilus là loài sống trong các tầng nông sâu của đất – đây cũng là loài phân bố khá rộng ở Việt Nam, có mặt ở hầu khắp các loại hình sinh cảnh, từ sinh cảnh rừng tự nhiên, sinh cảnh trảng cỏ cây bụi, đất đô thị, đến sinh cảnh đất trồng màu (Nguyễn Thị Thu Anh và cs., 2000; 2005; 2008; 2009; 2010; Nguyễn Trí Tiến và cs., 2008; 2009). Như vậy,

83
8 loài nêu trên không phải là những loài hiếm, hay đặc biệt về mặt sinh thái mà chúng chỉ là những loài ngẫu nhiên thu được với số lượng rất ít (từ 1 đến 3 cá thể) ở khu vực trồng khảo nghiệm ngô chuyển gen. Điều này cho thấy thành phần loài bọ đuôi bật Collembola ở các lô thí nghiệm trồng các giống ngô đối chứng (không chuyển gen) và chuyển gen không có sự khác biệt nhau.

84
Bảng 5-14: Đa dạng thành phần loài và phân bố của bọ đuôi bật (Collembola) trong các lô thí nghiệm thu bằng phương pháp pitfall trap trong KNHC (Văn Giang, 2010-2011)
TT Loài 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
Hypogastruridae 1 Xenylla humicola (Fabricius, 1780) 831 174 576 489 1828 172
Neanuridae 2 Brachystomella parvula (Schaffer, 1896) 2344 126 2198 1322 2881 558 3 Friesea sublimis MacNamara, 1921 0 0 1 0 0 0 4 Pseudachorutella asigilata (Borner, 1901) 44 10 44 23 27 10 Isotomidae 5 Cryptopygus thermophilus (Axelson, 1900) 1 2 0 0 0 6 Isotomurus prasinus (Reuter, 1891) 2554 266 1653 235 2379 221 7 Isotomurus punctiferus Yosii, 1963 775 629 613 203 511 371 Entomobryidae 8 Entomobrya lanuginosa (Nicolet, 1841) 961 16 922 40 1101 31 9 Entomobrya sp. 10 2 2 7 7 3 1
10 Homidia glassa Nguyen, 2001 263 64 254 76 280 76 11 Lepidocyrtus (L.) lanuginosus (Gmelin, 1788) 14 1 17 0 19 0 12 Lepidocyrtus (L. ) simsim Yosii, 1982 12 0 3 3 2 2 13 Lepidocyrtus (L.) sp. 0 5 0 0 0 1 0 14 Lepidocyrtus (Cyn.) sp. 1614 1 63 6561 80 2475 87 15 Lepidocyrtus (Acr.) heterolepis Yosii, 1959 6 0 2 0 8 2 16 Lepidocyrtus (Acr.) transiens Yosii, 1982 98 0 97 1 96 2

85
17 Lepidocyrtus (Asc.) cf. aseanus Yosii, 1982 3 1 2 0 0 0 18 Lepidocyrtus (Asc.) concolourus Nguyen, 2001 47 1 51 0 56 0 19 Lepidocyrtus (Asc.) dahlii Schaffer, 1898 0 1 0 20 Lepidocyrtus (Asc.) sp. 393 4 7 399 4 438 11 21 Willowsia mesothorax Nguyen, 2001 1 0 0 0 0 0
Cyphoderidae 22 Cyphoderus javanus Borner, 1906 209 4 148 2 203 0
Paronellidae 23 Salina celebensis (Schaffer, 1898) 1 0 1 0 18 0 24 Salina okinawana Yosii, 1983 10 0 0 0 8 0 25 Salina pictura Nguyen, 2001 2 0 1 0 0 0 26 Salina sp. 45 14 0 78 0 50 0
Sminthurididae 27 Sminthurides aquaticus (Bourlet, 1842) 0 579 0 315 0 451 28 Sminthurides bothrium Nguyen, 2001 1411 571 1589 371 1440 541 29 Sphaeridia pumilis (Krausbauer, 1898) 1 0 0 0 4 0 30 Sphaeridia zaheri Yosii, 1966 1 0 0 0 1 0
Bourletiellidae 31 Deuterosminthurus pallipes (Bourlet, 1843) 12 1 3 0 28 0 32 Deuterosminthurus sp. 34 1 1 43 0 58 1 33 Deuterosminthurus sp. 24 2 0 16 0 0 0
Sminthuridae 34 Sphyrotheca macrochaeta Nguyen, 1995 39 0 19 0 2 0

86
So sánh giá trị của một số chỉ số định lượng của Bọ đuôi bật (Collembola) ở các giống ngô trồng khảo nghiệm (thu bằng phương pháp pitfall trap)
Phân tích một số chỉ số định lượng của bọ đuôi bật (Collembola) như: số lượng loài, số lượng cá thể; chỉ số đa dạng Shannon- Weaver (H’); chỉ số đồ ng đều Pielou (J’) (Bảng 5-15, Bảng 5-16) cho thấy:
Kết quả vụ 1: Số lượng loài thu được của các lô khảo nghiệm dao động từ 16 loài đến 30 loài tùy từng lần thu mẫu. Ở lô trồng ngô đối chứng, số loài dao động quanh 14 loài ở lần thu thứ 4 (ở 30Y87H) và thứ 5 (ở 30Y87). Ở các lô trồng ngô không chuyển gen, số loài dao động từ 14 loài (ở 30Y87, lần thu mẫu thứ 5) đến 24 loài (ở 30B80, lần thu thứ hai) (Bảng 5-15). Sự khác biệt về số lượng loài thể hiện ngay ở một lô trồng ngô thí nghiệm (với các lần thu mẫu hoặc giữa các lô trồng ngô cùng là ngô chuyển gen hay cùng là ngô đối chứng không chuyển gen). Điều này có nghĩa sự khác biệt về số lượng loài bọ đuôi bật (Collembola) giữa các công thức trồng khảo nghiệm hạn chế ngô chuyển gen không phụ thuộc vào giống ngô trồng mà có thể phụ thuộc vào các yếu tố ngoại sinh khác như thời tiết, đặc điểm cư trú hoặc đặc điểm dinh dưỡng, đặc điểm di cư của bản thân động vật sống ở khu vực nghiên cứu.
Kết quả vụ 2: Tất cả các loài thu được ở trong khảo nghiệm vụ 2 là những loài đều đã thu được trong khảo nghiệm ở vụ 1. Phần lớn các loài (Collembola) thu được trong danh sách (Bảng 5-15) thuộc nhóm sống ở lớp thảm và ở dưới lớp thảm - trên mặt đất như: Xenylla humicola (Hypogastruridae), Brachystomella parvula; Pseudachorutella assigilata (Neanuridae); một số loài thuộc giống Lepidocyrtus: Lepidocyrtus (Acr.) heterolepis, L. (Acr.) transiens, L. (Asc.) concolourus, L. (Asc.) sp.4,… (Entomobryidae); Cyphoderus javanus (Cyphoderidae); một số loài thuộc Symphypleona: Sminthurides bothrium, Deuterosminthurus pallipes, Deuterosminthurus sp.1
Trong số 21 loài Collembola, có tới 15 loài (chiếm 71,43% tổng số loài) có mặt ở cả lô khảo nghiệm trồng ngô đối chứng và ngô biến đổi gen. Chỉ có 6 loài là có mặt trong các lô khảo nghiệm trồng hoặc ngô biến đổi gen hoặc ngô đối chứng (không biến đổi gen), cụ thể: chỉ có mặt ở lô trồng ngô không biến đổi gen, gồm 5 loài – Lepidocyrtus (L.) cintus, Lepidocyrtus (Asc.) concolourus, Deuterosminthurus pallipes (30B80), Lepidocyrtus (Asc.) dahlii (30Y87), Cyphoderus javanus (30B80; 30Y87); chỉ có mặt ở lô trồng ngô biến đổi gen, gồm 1 loài: Lepidocyrtys (Acr.) heterolepis. Mặc dù, 6 loài nêu trên chỉ có mặt trong các lô thí nghiệm trồng hoặc ngô biến đổi gen hoặc ngô đối chứng (không biến đổi gen) nhưng đây không phải là những loài đặc trưng cho lô khảo nghiệm trồng ngô biến đổi gen hay ngô không biến đổi gen. Trong khảo nghiệm ở vụ 1, những loài này đều có mặt ở cả lô trồng ngô biến
,…

87
đổi gen và ngô không biến đổi gen. Vì vậy, việc không thu được trong lô trồng ngô biến đổi gen hay lô đối chứng không biến đổi gen chỉ là ngẫu nhiên không thu được loài; hơn nữa đây là những loài có thể gặp nhiều ở những sinh cảnh mở, đất nông nghiệp. Chúng gặp khá phổ biến trong các dạng sinh cảnh khác nhau ở những địa phương khác nhau của Việt Nam (ở VQG Xuân Sơn, Phú Thọ; khu BTTN Thượng Tiến, Kim Bôi, Hòa Bình; đất canh tác Gia Lâm, Hà Nội; khu vực phía Bắc các tỉnh Na Rì, Sơn La,…; (Nguyễn Thị Thu Anh và cs., 2000; 20 08; 2009; 2010; Nguyễn Trí Tiến và cs., 2008; 2009).
Như vậy, về thành phần loài và phân bố của Collembola ở đất trồng ngô biến đổi gen và ngô không biến đổi gen ở k hu vực trồng KNHC ngô biến đổi gen của Trạm thực nghiệm Văn Giang, Hưng Yên không có gì khác biệt. Bảng 5-15: So sánh số lượng loài, số cá thể của bọ đuôi bật (Collembola) trên các
giống ngô khảo nghiệm theo từng đợt thu mẫu trong KNHC (Văn Giang, 2010-2011)
Đợt thu
mẫu Chỉ số
Ngô Đ/C Ngô BĐG 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 Số loài 17 13 18 11 18 15 Số cá thể 2464 624 2702 1247 2904 322
2 Số loài 24 12 22 10 23 10 Số cá thể 1957 566 2072 359 1958 657
3 Số loài 20 11 15 12 15 11 Số cá thể 1766 675 6793 653 3198 867
4 Số loài 17 8 16 12 14 12 Số cá thể 4465 317 2695 348 4227 395
5 Số loài 15 11 14 12 15 11 Số cá thể 1108 355 1038 565 1630 274
Chung (cả 5 lần)
Số loài 30 16 27 16 26 17 Số cá thể 11760 2537 15300 3172 13917 2515

88
- Số lượng cá thể (độ phong phú):
Vụ 1: Tính chung sau 5 lần thu mẫu (bẫy cốc) của các lô trồng khảo nghiệm ngô chuyển gen ở Văn Giang, Hưng Yên (Bảng 5-15): số lượng bọ đuôi bật (Collembola) đạt giá trị cao nhất ở lô trồng ngô không chuyển gen (15300 cá thể, trung bình 1020 cá thể/ cốc), thấp hơn ở lô trồng ngô chuyển gen ( 13917 cá thể, trung bình đạt 893-928 cá thể/ cốc) và thấp nhất cũng ở lô trồng ngô không chuyển gen (11760 cá thể, trung bình đạt 784 cá thể/cốc). Kết quả này ch ứng tỏ độ phong phú của bọ đuôi bật (Collembola) ở các lô thí nghiệm trồng ngô chuyển gen có độ dao động khá lớn.
Vụ 2: số lượng Collembola ở lô trồng ngô không biến đổi gen đạt giá trị từ thấp nhất (2515 cá thể) đến giá trị cao nhất (3172 cá thể); số lượng cá thể Collembola ở lô trồng ngô biến đổi gen thu được (2537 cá thể) thấp hơn so với ngô không biến đổi gen cùng giống và cao hơn không đáng kể so với ngô không biến đổi gen khác giống. Nhìn chung, giá trị về số lượng cá thể Collembola ở lô trồng ngô biến đổi gen nằm trong khoảng giá trị của giống ngô đối chứng không biến đổi gen và xu thế tăng giảm giá trị số lượng cá thể từ các lô trồng ngô đối chứng sang lô trồng ngô biến đổi gen, ở các lần thu mẫu cũng như chung cả 5 lần thu mẫu, không làm cho các lô trồng ngô đối chứng khác biệt với lô trồng ngô biến đổi gen về số lượng cá thể của Collembola ở các giống ngô khảo nghiệm (F3,88
- Chỉ số đa dạng H’ và chỉ số đồng đều J’: Kết quả phân tích sự thay đổi giá trị chỉ số đa dạng Shannon-Weaver (H’) và chỉ số đồng đều J’ trình bày ở Bảng 5-16 cho thấy: sau 5 lần thu mẫu, giá trị của chỉ số đa dạng H’ và chỉ số đồng đều (J’) giữa các lô trồng ngô chuyển gen với lô trồng ngô không chuyển gen tương đương nhau, không thấy có sự khác biệt, cụ thể như sau:
=0,37; P>0,05). Điều này chứng tỏ sự biến động số lượng cá thể ở ngô biến đổi gen so với ngô đối chứng không bắt nguồn từ nguyên nhân do ngô biến đổi gen gây ra hay nói cách khác là k hông phải ảnh hưởng do protein Cry1F gây ra. Kết luận này cũng giống với các nghiên cứu của các tác giả Al-Deeb và cs., 2003; Candolfi và cs., 2004; Ahmad và cs., 2005; Bitzer và cs., 2005, đó là protein Cry1F không ảnh hưởng đến mức độ phong phú của động vật chân khớp bé (Collembola) ở đất.
Vụ 1: + Giá trị H’: lô đối chứng 1,89 – 2,22, còn lô trồng ngô chuyển gen 2,17 + Giá trị J’ của ngô đối chứng thấp hơn chút ít so với J’ của ngô chuyển gen.
Vụ 2: + Giá trị H’: lô đối chứng 1,81- 1,88, lô trồng ngô chuyển gen 1,99

89
+ Giá trị J’: xu hướng tăng giảm giá trị của J’ giữa ngô đối chứng không chuyển gen và ngô chuyển gen tương tự như với giá trị của chỉ số đa dạng H’.
Phân tích chi tiết sự thay đổi các giá trị của chỉ số đa dạng, chỉ số đồng đều trong từng lần thu mẫu (Bảng 5-16) cũng cho kết quả tương tự. Đó là: các giá trị H’ hay J’ của ngô chuyển gen 30Y87H cao hơn so với ngô đối chứng 30Y87 và 30B80, nhưng các giá trị chỉ số này vẫn nằm trong khoảng dao động của ngô đối chứng.
Bảng 5-16: Giá trị chỉ số đa dạng H’ và chỉ số đồng đều (J’) của bọ đuôi bật
(Collembola) trên các giống ngô KNHC (Văn Giang, 2010 -2011)
Đợt thu mẫu Chỉ số
Ngô Đ/C Ngô BĐG 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 H’ 2,00 1,77 2,00 1,05 1,86 1,34 J’ 0,71 0,65 0,69 0,44 0,64 0,52
2 H’ 1,94 1,58 1,88 1,86 2,02 1,67 J’ 0,61 0,69 0,61 0,81 0,65 0,67
3 H’ 1,87 1,61 0,77 1,67 1,67 1,86 J’ 0,62 0,67 0,28 0,67 0,62 0,77
4 H’ 1,75 1,55 1,70 1,57 1,64 1,57 J’ 0,62 0,62 0,61 0,75 0,62 0,75
5 H’ 1,98 1,82 1,92 1,87 1,78 1,98 J’ 0,73 0,76 0,73 0,75 0,66 0,82
Chung (cả 5 lần)
H’ 2,22 1,88 1,89 1,81 2,17 1,99 J’ 0,65 0,66 0,57 0,65 0,66 0,72
5.1.4.4 Kết quả phân tích các loài bọ đuôi bật (Collembola) phổ biến và ưu thế
Loài ưu thế là loài có số lượng cá thể riêng chiếm ≥ 50 % tổng cá thể của lô khảo nghiệm (Maria Sterzynska, 1990). Ở mỗi sinh cảnh có một tập hợp loài ưu thế đặc trưng và tập hợp này thay đổi ở các sinh cảnh khác nhau. Sự thay đổi các loài ưu thế phản ánh sự thay đổi điều kiện môi trường sống. Trong điều kiện sống tối ưu, mang tính chất tự nhiên, thông thường các loài trong quần xã có giá trị số lượng cá thể tương đối đồng đều. Giá trị số lượng loài ưu thế không vượt trội hẳn so với các loài khác. Ngược lại, khi điều kiện môi trường thay đổi, tác động trực tiếp đến từng cá thể,

90
sinh vật phải tự điều chỉnh để thích ứng với điều kiện sống mới dẫn đến kết quả một vài loài bị diệt vong, một vài loài khác phát triển đột biến làm thay đổi tập hợp ưu thế trong quần xã. Trên cơ sở thay đổi ấy, người ta có thể phán đoán quá trình cũng như chiều hướng của sự thay đổi điều kiện môi trường. Kết quả phân tích các loài bọ đuôi bật phổ biến và ưu thế được ghi trong Bảng 5-17 và Bảng 5-18. Cả khu vực trồng ngô khảo nghiệm ở Văn Giang, Hưng Yên trong 2 vụ đã ghi nhận được 16 loài bọ đuôi bật (Collembola) phổ biến và 7 loài bọ đuôi bật (Collembola) ưu thế.
Vụ 1: trong số 14 loài bọ đuôi bật (Collembola) phổ biến có 12 loài phổ biến ở cả 3 công thức khảo nghiệm với độ phổ biến từ 53,33% đến 100% (Bảng 5-17). 2 loài bọ đuôi bật còn lại chỉ phổ biến ở 2 công thức của ngô không chuyển gen (Pseudachorutella assigilata và Salina sp.14). Cũng trong 14 loài Bọ đuôi bật (Collembola) phổ biến trên, đồng thời có 6 loài là loài ưu thế ở 3- 4 lô thí nghiệm trồng ngô khảo nghiệm với độ ưu thế từ 6,03% (ở giống ngô 30Y87) đến 42,88% (cũng ở giống ngô 30Y87), bao gồm: Lepidocyrtus (Cyn.) sp.1
Vụ 2: đã ghi nhận được 12 loài phổ biến. Trong số này có 6 loài Collembola là ưu thế ở tất cả các lô khảo nghiệm trồng ngô biến đổi gen và không biến đổi gen (Bảng 5-17), đó là Brachystomella parvula, Isotomurus prasinus, Isotomurus punctiferus, Sminthurides aquaticus, Sminthurides bothrium, Xenylla humicola, vv, Không có loài nào ưu thế riêng cho mỗi lô trồng thí nghiệm. Như vậy, thành phần loài phổ biến và ưu thế giữa lô đất trồng ngô biến đổi gen và không biến đổi gen trong 2 vụ không có sự khác biệt đáng kể.
, Brachystomella parvula, Sminthurides bothrium, Entomobrya lanuginosa, Isotomurus prasinus, Xenylla humicola. Như vậy, những loài Bọ đuôi bật (Collembola) phổ biến và ưu thế ở lô trồng ngô chuyển gen cũng là các loài Bọ đuôi bật (Collembola) ưu thế và phổ biến ở các lô trồng ngô đối chứng, không có sự sai khác.

91
Bảng 5-17: Các loài bọ đuôi bật (Collembola) phổ biến (%) trong đất trồng ngô
KNHC (Văn Giang, 2010 – 2011)
TT Loài 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 Lepidocyrtus (Cyn.) sp. 100 1 100 100 100 100 100 2 Brachystomella parvula 100 100 100 100 100 100 3 Sminthurides aquaticus 100 100 100 100 100 100 4 Sminthurides bothrium 0 100 0 100 0 100 5 Entomobrya lanuginosa 93.33 80 100 80 93.33 80 6 Cyphoderus javanus 93.33 80 100 0 86.67 60 7 Xenylla humicola 93.33 80 93.33 100 100 100 8 Homidia glassa 93.33 100 93.33 100 100 100 9 Isotomurus punctiferus 86.67 100 93.33 100 86.67 100 10 Lepidocyrtus (Asc.) sp. 86.67 4 60 93.33 0 93.33 0 11 Lepidocyrtus (Cyn.) sp. 0 1 100 0 100 0 100 12 Lepidocyrtus (Asc.)
concolourus 73.33 0 86.67 0 93.33 0
13 Isotomurus prasinus 80 100 80 100 80 100 14 Lepidocyrtus (Acr.)
transiens 86.67 80 80 0 93.33 60
15 Pseudachorutella asigilata 53.33 0 53.33 0 0 0 16 Salina sp. 0 14 0 53.33 0 0 0

92
Bảng 5-18: Các loài bọ đuôi bật (Collembola) ưu thế (%) trong đất ngô KNHC (Văn Giang, 2010 – 2011)
TT Loài 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 Lepidocyrtus (Cyn.) sp. 13.72 1 42.88 17.78 2 Brachystomella parvula 19.93 5.01 14.37 41.68 20.70 21.99 3 Sminthurides aquaticus 0 23.02 0 9.93 17.78 4 Sminthurides bothrium 11.99 22.70 10.38 11.70 10.35 21.32 5 Entomobrya lanuginosa 8.17 0 6.03 0 7.91 0 6 Cyphoderus javanus 0 0 0 0 0 0 7 Xenylla humicola 7.07 6.92 0 15.42 13.14 6.78 8 Homidia glassa 0 0 0 0 0 0 9 Isotomurus punctiferus 6.59 25.01 0 6.40 0 14.62 10 Isotomurus prasinus 21.72 10.58 10.80 7.91 17.09 8.71
Như vậy, kết quả phân tích sau 2 vụ trồng khảo nghiệm hạn chế ngô chuyển gen (30Y87H) của công ty Pioneer ở Trạm Thực nghiệm Văn Giang, Hưng Yên, bước đầu như sau: Dù thu mẫu bằng phương pháp nào lấy mẫu đất (soil core) hay bẫy cốc (pitfall trap) cũng đều cho thấy:
- Ngô chuyển gen không ảnh hưởng đến bọ đuôi bật Collembola, cụ thể không thấy có sự sai khác có ý nghĩa về thành phần loài, số lượng loài và mức độ phong phú của bọ đuôi bật Collembola giữa các giống ngô trồng khảo nghiệm (ngô đối chứng và ngô chuyển gen).
- Tính chung cho cả các lần t hu mẫu định tính và định lượng bọ đuôi bật Collembola, số lượng cá thể, giá trị của các chỉ số đa dạng H’ và chỉ số đồng đều J’ của ngô chuyển gen tương đương nhau hay nói cách khác các chỉ số này ở ngô chuyển gen đều nằm trong khoảng dao động của các giá trị này ở ngô đối chứng.
- Tập hợp bọ đuôi bật Collembola ưu thế giữa hai lô đất trồng ngô chuyển gen và không chuyển gen có nhiều điểm chung và không có nhiều sự sai khác.

93
5.1.5 Đánh giá sự xuất hiện và gây hại của các bệnh hại ngô chính trong KNHC
Vào thời điểm 85 ngày sau gieo đã tiến hành đánh giá mức độ bị nhiễm các bệnh
chính của các giống ngô thí nghiệm. Vào thời điểm tiến hành đánh giá này, các giống ngô trong thí nghiệm bị các bệnh đốm lá lớn (Helminthoprium turcicum), bệnh đốm lá nhỏ (Helminthoprium maydis), bệnh gỉ sắt ( Puccinia maydis), bệnh khô vằn (Rhizoctonia solani) và bệnh đốm nâu ( Physoderma maydis). Tỷ lệ và chỉ số bệnh trên các giống ngô khảo nghiệm trong 2 vụ trồng cụ thể như sau:
5.1.5.1 Bệnh đốm lá lớn (Helminthoprium turcicum)
Bệnh đốm lá lớn là một trong số các bệnh khá phổ biến trên cây ngô ở điều kiện sinh thái Việt Nam, song tùy từng điều kiện sinh cảnh cụ thể mà tỷ lệ và chỉ số bệnh khác nhau. Kết quả theo dõi tỷ lệ, chỉ số bệnh đốm lá lớn trên các giống ngô khảo nghiệm hạn chế được ghi ở Bảng 5-19.
Vụ 1: Cả 3 giống ngô khảo nghiệm không bị bệnh đốm lá lớn. Vụ 2: Bệnh đốm lá lớn xuất hiện và gây hại rất nhẹ trên các giống ngô khảo
nghiệm. Bệnh có xu hướng gây hại mạnh hơn trên giống ngô chuyển gen với tỷ lệ hại lên tới 20% trong khi giống nền tỷ lệ hại là 6,67%. Tuy nhiên sự sai khác này vẫn nằm trong giới hạn của phân lớp (cùng phân lớp a), cho nên sự sai khác là không có ý nghĩa.
So sánh các chỉ số trên ở vụ 1 với vụ 2, thì có sự khác biệt lớn. Tuy nhiên, sự khác biệt này do điều kiện thời tiết gây nên chứ không phái do giống có Protein Bt gây nên. Cuối năm 2010, đầu năm 2011 có nhiều đợt rét đậm kéo dài, nhiệt độ xuống < 150
C, ẩm độ không khí tăng, điều kiện này không thích hợp cho cây ngô sinh trưởng, phát triển nhưng lại là điều kiện thuận lợi để cho nấm bệnh đốm lá lớn phát triển mạnh.
Bảng 5-19: Mức độ nhiễm bệnh đốm lá lớn của các giống ngô KNHC (Văn Giang, 2010-2011)
Tên giống Tỷ lệ bệnh (%) Chỉ số bênh (%)
Vụ 1 Vụ 2 Vụ 1 Vụ 2 30B80 0 3a 0 15a 30Y87 0 6,8a 0 2a 30Y87H 0 20a 0 5,3a

94
5.1.5.2 Bệnh đốm lá nhỏ (Helminthoprium maydis)
Bệnh đốm lá nhỏ là bệnh khá phổ biến trên tất cả các khu vực trồng ngô trong đó có Việt Nam. Kết quả theo dõi bệnh đốm lá nhỏ trong khảo nghiệm hạn chế được ghi trong Bảng 5-20.
Vụ 1: Bệnh phát sinh gây hại khá nặng trên tất cả các giống ngô khảo nghiệm. Tỷ lệ cây ngô bị bệnh đốm lá nhỏ đã quan sát được trên các giống biến động từ 76,7% ở giống 30B80, 30Y87 đến 88,3% ở giống 30Y87H. Mức độ bị bệnh quan sát được phổ biến ở cấp bệnh 1 và cấp bệnh 3. Tỷ lệ cây ngô bị nhiễm ở cấp bệnh 1 biến động trong khoảng 20,0 - 48,33%; cây ngô bị nhiễm ở cấp bệnh 3 là chủ yế u và với tỷ lệ biến động trong khoảng 28,33 - 66,67%; cây ngô bị nhiễm cấp bệnh cao nhất là cấp bệnh 7 với tỷ lệ rất thấp và chỉ là 1,67% . Chỉ số bệnh trung bình của bệnh đốm lá nhỏ trên các giống ngô thí nghiệm đạt không thấp, biến động từ 14,81% ở giống 30B80 đến 25, 37% ở giống 30Y87.
Vụ 2: Tỷ lệ bệnh đốm lá nhỏ dao động từ 93,34%-98,3%, chỉ số bệnh dao động từ 23,33%-40,67% trên cả giống nền và giống chuyển gen. Trong 3 giống thí nghiệm chỉ có giống 30B80 có chỉ số bệnh là thấp nhất (23,33%) và sai khác có ý nghĩa với giống không chuyển gen 30Y80, nhưng không sai khác so với giống chuyển gen 30Y87H còn lại. Chỉ số này giữa giống nền (30Y87) và giống chuyển gen (30Y87H) sai khác nhau là không nhiều (47,67%và 40,67%) và không có ý nghĩa.
Như vậy, giữa các giống ngô chuyển gen và không chuyển gen không có sự khác biệt về mức độ bị bệnh đốm lá nhỏ.
Bảng 5-20: Mức độ nhiễm bệnh đốm lá nhỏ trên của các giống ngô KNHC
(Văn Giang, 2010 -2011)
Tên giống Tỷ lệ bệnh (%) Chỉ số bênh (%)
Vụ 1 Vụ 2 Vụ 1 Vụ 2 30B80 76,70 a 93,34a 14,81 a 23,33 a 30Y87 76,70 a 98,3a 21,85 ab 47,67b 30Y87H 88,30 a 96,6a 25,37 b 40,67ab CV (%) 17,58 7,95 25,54 31,83 LSD 27,58 .05 15,26 10,86 23,69
Ghi chú: Giá trị trung bình trong cùng một cột mang chữ cái khác nhau thì sai khác có ý nghĩa thống kê (p < 0.05).

95
5.1.5.3 Bệnh gỉ sắt (Puccinia maydis)
Kết quả theo dõi bệnh gỉ sắt phát sinh và gây hại trên các giống ngô khảo nghiệm được ghi trong Bảng 5-21.
Vụ 1: tỷ lệ cây ngô bị bệnh gỉ sắt đạt rất cao, từ 91,7% ở giống 30Y87H đến 100% ở giống 30B80. Mức độ bị bệnh quan sát được ở tất cả các cấp bệnh, nhưng chủ yếu bị nhiễm bệnh gỉ sắt từ cấp 5 và cấp 7. Tỷ lệ cây ngô bị nhiễm ở cấp bệnh 5 (cấp bệnh phổ biến hơn cả) biến động trong khoảng 36,67 - 50,0%; số cây ngô bị nhiễm cấp bệnh cao nhất là cấp bệnh 9 với tỷ lệ là 0 - 16,67%. Chỉ số bệnh trung bình của bệnh gỉ sắt trên các giống ngô thí nghiệm đạt khá cao, biến động từ 49,81% ở giống 30Y87H đến 70,0 % ở giống 30B80. Nhưng sự khác biệt có ý nghĩa thống kê chỉ quan sát thấy ở giống ngô chuyển gen (30Y87H) so với giống ngô không chuyển gen (30B80). Mức độ bị bệnh gỉ sắt của các giống ngô chuyển gen (30Y87H) và giống ngô nền tương ứng (30Y87) là tương tự nhau.
Vụ 2: Bệnh gỉ sắt không xuất hiện và gây hại trên tất cả các giống ngô khảo nghiệm.
Như vậy, tỷ lệ và chỉ số bệnh không bị ảnh hưởng bởi giống ngô chuyển gen hay không chuyển gen, sự khác biệt này chỉ do điều kiện thời tiết gây nên.
Bảng 5-21: Mức độ nhiễm bệnh rỉ sắt của các giống ngô KNHC
(Văn Giang, 2010-2011)
Tên giống Tỷ lệ bệnh (%) Chỉ số bênh (%)
Vụ 1 Vụ 2 Vụ 1 Vụ 2 30B80 100 a 0 70,00a 0 30Y87 96,70 ab 0 53,33 ab 0 30Y87H 91,70 b 0 49,81 b 0 CV (%) 4,20 16,36 LSD 7,68 .05 18,82
Ghi chú: Giá trị trung bình trong cùng một cột mang chữ cái khác nhau thì sai khác có ý nghĩa thống kê (p < 0.05)

96
5.1.5.4 Bệnh khô vằn (Rhizoctonia solani)
Bệnh khô vằn là bệnh gây hại nhiều trên cây ngô ở Việt Nam. Tuy nhiên, tùy điều kiện thời tiết cụ thể mà mức độ bị bệnh khác nhau. Kết quả theo dõi mức độ bị bệnh khô vằn trên các giống ngô khảo nghiệm được ghi trong Bảng 5-22.
Vụ 1: Tỷ lệ cây ngô bị bệnh khô vằn đã quan sát được trên các giống khảo nghiệm đạt không cao, biến động từ 36,7% ở giống 30B80 đến 40,0% ở các giống 30Y87 và 30Y87H. Mức độ bị bệnh quan sát được ở tất cả các cấp bệnh, nhưng phổ biến là ở cấp bệnh 1 và cấp bệnh 3. Tỷ lệ cây ngô bị nhiễm ở cấp bệnh 1 và cấp bệnh 3 biến động trong khoảng 10,0 - 23,3%; số cây ngô bị nhiễm bệnh cao nhất là cấp bệnh 9 với tỷ lệ là 1,67 - 6,67%. Chỉ số bệnh trung bình của bệnh khô vằn đạt ở mức thấp, biến động từ 8,15% ở giống 30B80 đến 17,78% ở giống 30Y87. Chỉ số bệnh trung bình của các giống ngô biến đổi gen và giống ngô nền đã quan sát được tương tự nhau.
Vụ 2: Bệnh khô vằn không xuất hiện trên tất cả các giống ngô khảo nghiệm. Qua kết quả theo dõi cho thấy, bệnh khô vằn gây hại trên ngô không chịu ảnh
hưởng yếu tố giống ngô chuyển gen hay không chuyển gen, mà chỉ phụ thuộc bởi điều kiện thời tiết, khí hậu.
Bảng 5-22: Mức độ bị bệnh khô vằn của các giồng ngô KNHC
(Văn Giang, 2010-2011)
Tên giống Tỷ lệ bệnh (%) Chỉ số bênh (%)
Vụ 1 Vụ 2 Vụ 1 Vụ 2 30B80 36,70b 0 8,15a 0 30Y87 40,00 b 0 17,78 a 0 30Y87H 40,00 b 0 13,33 a 0 CV (%) 15,82 44,92 LSD 13,03 .05 12,40
Ghi chú: Giá trị trung bình trong cùng một cột mang chữ cái khác nhau thì sai khác có ý nghĩa thống kê (p < 0.05)

97
5.1.5.5 Bệnh đốm nâu (Physoderma maydis)
Bệnh đốm nâu là một trong 5 loại bệnh hại cây ngô phổ biến ở Việt Nam. Kết quả theo dõi được ghi trong Bảng 5-23.
Vụ 1: Tỷ lệ cây ngô bị bệnh đốm nâu đã quan sát được trên các giống thí nghiệm đều đạt rất cao, ở mức 100%. Mức độ bị bệnh quan sát được chủ yếu ở cấp bệnh cao nhất là cấp bệnh 9 với tỷ lệ từ 78,33% ở giống 30Y87 đến 86,67% ở giống 30B80 và giống 30Y87H. Chỉ số bệnh trung bình đã quan sát được đạt ở mức rất cao, từ 95,19% ở giống 30Y87 đến 97,04% ở giống 30B80 và giống 30Y87H. Mức độ nhiễm bệnh đốm nâu của giống ngô biến đổi gen và giống ngô nền không khác biệt ở mức có ý nghĩa thống kê
Vụ 2: tỷ lệ và chỉ số bệnh trên các giống ngô chuyển gen hay không chuyển gen tương đương nhau và dao động trong khoảng 41,67% ( 30B80) - 58,67% (30Y87).
Như vậy, qua 2 vụ khảo nghiệm hạn chế ngô chuyển gen liên tiếp tại Trạm thực nghiệm văn Giang, Hưng Yên cho thấy ngô chuyển gen hay không chuyển gen không ảnh hưởng đến tỷ lệ và chỉ số bệnh. Thời vụ khác nhau ảnh hưởng đến tỷ lệ và chỉ số các bệnh hại ngô chính. Kết quả này trùng với kết quả của một số tác giả khảo nghiệm về cây trồng biến đổi gen ở một số nước trên thế giới (Zwahlen C., Nentwig W., Bigler F., Hilbeck A., 2000).
Bảng 5-23: Mức độ nhiễm bệnh đốm nâu của các giống ngô KNHC
(Văn Giang, 2010 – 2011)
Tên giống Tỷ lệ bệnh (%) Chỉ số bênh (%)
Vụ 1 Vụ 2 Vụ 1 Vụ 2
30B80 100 a 81,67a 97,04 a 41,67a 30Y87 100 a 100a 95,19 a 58,67a 30Y87H 100 a 88,33a 97,04 a 44,33a CV (%) 0,0 21,68 4,25 35,01 LSD 0,0 .05 38,98 7,71 33,73
Ghi chú: Giá trị trung bình trong cùng một cột mang chữ cái khác nhau thì sai khác có ý nghĩa thống kê (p < 0.05)

98
5.1.6 Kết quả đánh giá mức độ gây hại của sâu hại ngô không chủ đích
Để đánh giá mức độ gây hại của sâu hại ngô NTOs trên các giống ngô khảo nghiệm. Chọn 2 loài chích hút (rệp muội ngô Rhopalosiphum maidis, nhện đỏ son Tetranychus cinnabarinus ) và 1 loài miệng nhai (sâu cắn lá ngô Mythimna separata Walk).
5.1.6.1 Rệp muội ngô (Rhopalosiphum maidis)
Rệp muội ngô (Rhopalosiphum maidis) đã ghi nhận được trong hầu hết các kỳ điều tra trên tất cả các giống ngô khảo nghiệm.
Vụ 1: cấp nhiễm trung bình rệp muội hại ngô có xu hướng gia tăng theo thời gian sinh trưởng phát triển của cây ngô. Ở kỳ điều tra đầu tiên (giai đoạn V8) đạt rất thấp và biến động trong khoảng 0,07 - 0,35. Chỉ tiêu này trong kỳ điều tra cuối cùng (giai đoạn R3
Vụ 2: cấp nhiễm trung bình dao động từ 0,03 – 0,47 và không theo quy luật. ) đạt khá cao từ 1,08 đến 1,6.
Tuy nhiên, cấp nhiễm trung bình rệp hại ngô có thay đổi theo từng kỳ điều tra, nhưng các giống ngô khác nhau (chuyển gen hay không chuyển gen) không ảnh hưởng đến cấp nhiễm rệp muội. Vụ trồng khác nhau có ảnh hưởng đến cấp nhiễm này, vụ 2 cấp nhiễm nhẹ hơn vụ 1.
Bảng 5-24: Cấp nhiễm trung bình của rệp muội ngô trên các giống ngô KNHC
(Văn Giang, 2010 -2011)
Đợt điều tra
30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 0,07 ± 0,07 0,03 ± 0,03 0,08 ± 0,04 0,32 ± 0,02 0,35 ± 0,03 0,25 ± 0,08 2 0,50 ± 0,31 0,23 ± 0,09 0 0,47 ± 0,23 0,10 ± 0,03 0,37 ± 0,08 3 0,18 ± 0,07 0,07 ± 0,04 0,28 ± 0,18 0,08 ± 0,06 0,20 ± 0,05 0,03 ± 0,02 4 0,43 ± 0,19 0,07 ± 0,04 0,42 ± 0,30 0,17 ± 0,03 0,25 ± 0,18 0,13 ± 0,08 5 1,60 ± 0,45 0,08 ± 0,08 1,08 ± 0,48 0,53 ± 0,19 1,08 ± 0,11 0,43 ± 0,09
5.1.6.2 Nhện đỏ son (Tetranychus cinnabarinus)
Nhện đỏ son (Tetranychus cinnabarinus) trên ngô thí nghiệm bắt đầu ghi nhận được bằng mắt thường từ kỳ điều tra giai đoạn R1. Cấp nhiễm trung bình của nhện đỏ

99
trên các giống ngô khảo nghiệm đạt không cao, biến động từ 0,28 trên giống ngô 30Y87 đến 2,9 trên giống ngô 30B80. Cấp nhiễm trung bình của nhện đỏ trên các giống ngô khảo nghiệm cao nhất quan sát được đạt từ 1,70 trên giống ngô 30Y87H đến 2,9 trên giống ngô 30B80. Mức độ nhiễm nhện đỏ son của giống ngô chuyển gen và giống ngô nền tương ứng là tương tự nhau (vụ 1, Bảng 5-25). Vụ 2 trên tất cả 3 giống khảo nghiệm, nhện đỏ son không xuất hiện. Như vậy, giống ngô chuyển gen hay không chuyển gen không ảnh hưởng tới mức độ gây hại của nhện đỏ son.
Bảng 5-25: Cấp nhiễm trung bình của nhện đỏ trên các giống ngô KNHC
(Văn Giang 2010 – 2011)
Đợt điều tra 30B80 30Y87 30Y87H
Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
3 (R1) 0,62 ± 0,09 0 0,58 ± 0,04 0 0,58 ± 0,0 0 4 (R2) 2,9 ± 0,38 0 1,72 ± 0,39 0 1,70 ± 0,31 0 5 (R3) 0,42± 0,004 0 0,28± 0,001 0 0,32 ± 0,003 0
5.1.6.3 Sâu cắn lá ngô (Mythimna separata)
Sâu cắn lá ngô là loài sâu hại ngô phổ biến, nhưng phát sinh cục bộ và thường có mật độ khá cao trong vụ ngô đông và vụ ngô xuân ở vùng đồng bằng sông Hồng. Kết quả điều tra sâu cắn lá ngô trong khảo nghiệm hạn chế ngô chuyển gen được ghi trong Bảng 5-26.
Vụ 1: trong cả 5 kỳ điều tra, tại những điểm điều tra bắt gặp sâu cắn lá ngô thường chỉ bắt gặp sâu non tuổi nhỏ trên công thức đối chứng, không bắt gặp sâu cắn lá ngô trên giống ngô chuyển gen. Do đó, loài sâu hại này hầ u như không gây hại cho các giống ngô khảo nghiêm. Cấp hại trung bình do sâu cắn lá ngô gây ra ghi nhận được trong lần điều tra cuối vào ngày 22/10/2010 (giai đoạn R6) đạt rất thấp và chỉ là 0,02.
Vụ 2: mật độ của loài sâu hại này đạt thấp, nên cấp hại do nó gây ra cũng rất thấp trong cả 5 kỳ điều tra. Cấp hại trung bình do sâu cắn lá ngô gây ra ghi nhận được trong lần điều tra cuối vào ngày 20/3/2011 (giai đoạn R6) đạt cao nhất cũng chỉ là 1,03 ở giống ngô biến đổi gen 30Y87H và là 1,68 ở giống ngô nền 30B80.
Kết quả theo dõi 3 loài sâu hại ngô không chủ đích (rệp muội ngô Rhopalosiphum maidis, nhện đỏ son Tetranychus cinnabarinus và sâu cắn lá ngô Mythimna separata Walk) trên các giống ngô khảo nghiệm (2 giống không chuyển

100
gen 30B80; 30Y87 và 1 giống chuyển gen 30Y87H) trong 2 thời vụ liên tiếp cho thấy: các giống ngô khảo nghiệm không ảnh hưởng đến cấp hại trung bình của 3 loài sâu hại không chủ đích chính. Chỉ có thời vụ ảnh hưởng tới sự xuất hiện và gây hại của 3 loài sâu hại đó.
Bảng 5-26: Cấp nhiễm trung bình của sâu cắn lá ngô trên các giống ngô KNHC
(Văn Giang, 2010 - 2011)
Đợt điều tra
30B80 30Y87 30Y87H Vụ 1 Vụ 2 Vụ 1 Vụ 2 Vụ 1 Vụ 2
1 0,02 ± 0,02 1,05 ± 0,69 0 0 0 0 2 0 0,02 ± 0,02 0 0,05 ± 0,03 0 0,02 ± 0,02 3 0 0 0 0 0 0 4 0 0,73 ± 0,17 0 0,32 ± 0,27 0 0,50 ± 0,15 5 0 1,68 ± 0,33 0 1,08 ± 0,17 0 1,03 ± 0,22
5.1.7 Kết luận sơ bộ kết quả KNHC (2 vụ lên tiếp)
Khảo nghiệm hạn chế được thực hiện trong 2 vụ liên tiếp tại trạm Thực nghiệm Văn Giang (2010-2011) do viện Di truyền Nông nghiệp thực hiện nhằm đánh giá rủi ro của việc trồng ngô chuyển gen TC1507. Kết luận sơ bộ được tóm tắt như sau:
1. Kết quả kiểm tra sự hiện diện của protein Cry1F chứng tỏ việc bố trí thí nghiệm ruộng khảo nghiệm hạn chế ngô chuyển gen hoàn toàn chính xác và hạt giống thí nghiệm là đồng nhất.
2. Đánh giá chỉ số đa dạng quần thể NTO cho thấy: chỉ số đa dạng Shannon H' ở các giống ngô khảo nghiệm đều đạt giá trị khá cao. Không có sự khác biệt về chỉ số đa dạng Shannon H' giữa các giống ngô khảo nghiệm. Mưcc độ phong phú trên các giống ngô thí nghiệm là tương đương và không phụ thuộc vào yếu tố mang gen cry1F. Các giá trị chỉ số ưu thế Simpson's D đều rất thấp và không khác biệt.
3. Cả 2 phương pháp điều tra trực tiếp và sử dụng bẫy dính vàng trong khu khảo nghiệm đã thu thập được 56 loài chân khớp thuộc 36 họ thuộc 11 bộ của lớp côn trùng và lớp hình nhện, trong đó có 15 loài là sâu hại ngô. Số lượng loài đã ghi nhận trên ngô chuyển gen và không chuyển gen không có sự khác biệt.

101
4. Điều tra trực tiếp trên ruộng ngô khảo nghiệm đã phát hiện được 19 loài thiên địch của sâu hại nói chung và sâu hại ngô nói riên g. Mật độ của các loài thiên địch này trên giống ngô chuyển gen và giống ngô không chuyển gen là tương tự nhau.
5. Việc canh tác ngô chuyển gen tổng hợp protein Cryp1F không ảnh hưởng đến quần bọ đuôi bật Collembola. Cụ thể không có sự sai khác về thành phần loài, số lượng loài và mức độ phong phú của Collembola giữa các giống ngô trồng khảo nghiệm (ngô đối chứng và ngô biến đổi gen).
6. Bệnh hại chủ yếu trên ngô là các bệnh: Đốm lá nhỏ, Gỉ sắt, bệnh Đốm nâu và Khô vằn. Tỷ lệ bệnh và chỉ số bệnh trên các giống ngô chuyển gen và không chuyển gen tương tự nhau.
7. Trong điều kiện khảo nghiệm, thành phần sâu hại ngô nghèo nàn và mật độ rất thấp. Không có loài sâu hại nào gây tác hại đáng kể đến sinh trưởng, phát triển của các giống ngô thí nghiệm. Tuy nhiên, giống ngô chuyển gen (30Y87H) thể hiện tính kháng sâu đục thân ngô rõ ràng hơn so với các giông ngô không chuyển gen.
5.2 Kết quả KNDR ngô BĐG event TC1507
5.2.1 Kiểm tra sự hiện diện của protein Cry1F và protein CP4 EPSPS
Kết quả Hình 5-6 tại 4 địa điểm khảo nghiệm là Vĩnh Phúc, Nghệ An, Đắk Lắk và Đồng Nai nhận thấy: (1) Giống chuyển gen 30Y87H cho kết quả dương tính với protein Cry1F và âm tính với protein CP4 EPSPS, biểu hiện ở việc xuất hiện 2 vạch trên que thử nhan h gen Cry1F, nhưng chỉ xuất hiện 1 vạch trên que thử nhanh gen chống chịu thuốc diệt cỏ glyphosate. (2) Giống chuyển gen 30Y87HR cho kết quả dương tính với cả hai loại protein Cry1F và protein CP4 EPSPS, biểu hiện ở việc xuất hiện 2 vạch trên cả hai loại que thử. (3) Giống ngô nền không chuyển gen 30Y87 và kết quả trên que thử đối chứng (chỉ thử với dung dịch đệm QuickStix EB2) đều cho kết quả âm tính với cả hai loại protein Cry1F và protein CP4 EPSPS, biểu hiện ở việc xuất hiện 1 vạch trên cả hai loại que thử nhanh. Kết quả thu được hoàn toàn tương tự nhau với tất cả các mẫu thử và ở cả 4 địa điểm khảo nghiệm. Như vậy, việc bố trí các công thức khảo nghiệm là chính xác và các giống tham gia khảo nghiệm cũng chính là những giống đã được cấp phép khảo nghiệm trên diện rộng.

102
(A) (B)
(C) (D)
Hình 5-6: Kiểm tra sự hiện diện của protein Cry1F trên các giống ngô trong
KNDR. (A) Vĩnh Phúc, (B) Nghệ An, (C) Đắk Lắk, (D) Đồng Nai
Ghi chú: 87: giống nền không chuyển gene 30Y87 H: giống chuyển gene 30Y87H mang event TC1507 cho tính trạng kháng
sâu cánh vảy (Lepidoptera) HR: giống chuyển gene 30Y87HR, giống đối chứng tham khảo mang
stack TC1507xNK603 cho tính trạng kháng sâu cánh vảy (Lepidoptera) và chịu thuốc trừ cỏ glyphosate
Que thử màu xanh tím: kiểm tra sự có mặt của protein Cry1F

103
Que thử màu đỏ: kiểm tra sự có mặt của protein CP4EPSPS (chịu thuốc trừ cỏ glyphosate)
Buffer: mẫu đối chứng với dung dịch đệm
5.2.2 Kết quả đánh giá tính ổn định và tính thích ứng của ngô event TC1507 tại các vùng sinh thái Việt Nam.
5.2.2.1 Kết quả đánh giá tính ổn định của ngô event TC1507
Để chứng minh có hay không ngô chuyển gen event TC1507 nguy cơ trở thành cỏ dại, dịch hại thông qua việc đánh giá các đặc tính hình thái, sinh trưởng, phát triển và mức độ mẫn cảm với các loại bệnh hại chủ yếu của ngô chuyển gene event TC1507 so với đối chứng không chuyển gene. Kết quả được thể hiện trong Bảng 5-27 đến Bảng 5-30.
Số liệu Bảng 5-27 cho thấy: t ỷ lệ mọc trung bình của của các giống khảo
nghiệm tại 4 điểm khảo nghiệm là 91.85%. Việc so sánh tỷ lệ mọc của 3 giống trong cùng một địa điểm khảo nghiệm cho thấy: (1) Tại Vĩnh Phúc, tỷ lệ mọc của giống đối chứng tham khảo 30Y87 HR (85.83%) cao hơn so với 30Y87H (80.53%) nhưng tương đương giống nền 30Y87 (81.18) ở mức tin cậy 95%. (2) Tại Nghệ An và Đắk Lắk, tỷ lệ mọc của cả 3 giống như nhau ở mức sác xuất 95% (Nghệ An: 92.33 - 95.65 %; Đắk Lắk: 96.90 - 97.50%). (3) Tại Đồng Nai, tỷ lệ mọc của 2 giống chuyển gen đều cao hơn so với giống nền ( lần lượt là 97.70; 96.50 so với 91.00%). So sánh tỷ lệ mọc của 3 giống ở 4 địa điểm cho thấy: Tại Vĩnh Phúc tỷ lệ mọc của 3 giống là thấp nhất, lần lượt 80.53; 85.83 và 81.18%. Tỷ lệ mọc cao nhất ghi nhận được tại Đắk Lắk, lần lượt 97.50; 97.20 và 97.70%. So sánh tỷ lệ mọc của từng giống tại 4 địa điểm khảo nghiệm cho kết quả: giống 30Y87H mang gen cry1F có tỷ lệ mọc thấp nhất tại Vĩnh Phúc (80.53%) và cao nhất tại Đồng Nai (97.70%); giống 30Y87HR (kháng côn trùng cánh vảy và kháng thuốc trừ cỏ glyphosate) có tỷ lệ mọc thấ p nhất tại Vĩnh Phúc (85.83% ) và cao nhất tại Đắk Lắk (97.50%). Đánh giá sức sống cây con cũng cho kết quả tương tự. Sức sống cây con ở cá c điểm khảo nghiệm tương tốt và đồng đều. Duy nhất tại Vĩnh Phúc do ảnh hưởng của thời tiết dẫn đến sức sống cây con giống 30Y87 là thấp nhất. Như vậy, so sánh tỷ lệ mọc, sức sống cây con của 3 giống ở cùng địa điểm và ở cả 4 địa điểm cho thấy không có sự khác biệt đáng kể (mức tin cậy 95%).

104
Theo dõi thời gian phun râu, thời gian tung phấn, chiều cao cây và chiều cao đóng bắp của 3 giống khảo nghiệm kết quả được thể hiện trong Bảng 5-28. Thời gian phun râu trung bình của 3 giống khảo nghiệm tại 4 địa điểm là 52.54 ngày. Thời gian phun râu dài nhất ghi nhận được tại Vĩnh Phúc, lần lượt 53.50; 54.25 và 55.10 ngày đối với các giống G1, G2 và G3. Chỉ tiêu này tại Nghệ An, Đắk Lắk và Đồng Nai cho kết quả tương tự nhau (từ 51.53 đến 52.30 ngày). So sánh thời gian phun râu của 3 giống ở cùng địa điểm khảo nghiệm cũng cho kết quả tương tự nhau. Thời gian tung phấn trung bình của cả 3 giống khảo nghiệm tại 4 địa điểm trung bình là 52.15 ngày. Các giống khác nhau trồng trong cùng một địa điểm và tính chung ở cả 4 địa điểm khảo nghiệm đều không có sự khác biệt đang kể về thời gian tung phấn (mức tin cậy 95%).
So sánh chiều cao cây giữa các giống khảo nghiệm cũng cho kết quả tương tự. Trong cùng mỗi điểm và trong cả 4 địa điểm khảo nghiệm, chiều cao cây sấp xỉ nhau (mức sác xuất 95%). Cao cây trung bình đạt 216.64 cm . Tại Vĩnh Phúc, chiều cao cây của 3 giống G1, G2 và G3 lần lượt là 224.40; 217.50 và 219.80 (cm). Tại Nghệ an, giá trị này lần lượt 220.50; 221,98 và 219.15 (cm). Tại Đắk Lắk là 238.50; 233.80 240.10 (cm). Tại Đồng Nai là197.30; 189.90 và 197.90 (cm). Chiều cao đóng bắp trung bình của 3 giống tại 4 điểm là 113.94 cm. So sánh chiều cao đóng bắp của 3 giống tại cùng 1 địa điểm hay tính chung cho cả 4 địa điểm khảo nghiệm đều cho kết quả không chênh lệch đáng kể.
Như vậy, thời gian phun râu, thời gian tung phấn, chiều cao cây và chiều cao đóng bắp của 3 giống ngô khảo nghiệm tại từng địa điểm hay tại cả 4 địa điểm khảo nghiệm là tương đương nhau (p < 0.05).
Theo dõi trạng thái cây, tỷ lệ đổ gẫy và tỷ lệ bật gốc của 3 giống ngô khảo
nghiệm. Kết quả được ghi nhận trong Bảng 5-29. Trạng thái cây trung bình của 3 giống ở 4 điểm đạt 8.1 điểm. So sánh trạng thái cây hay sức sống cây của 3 giống ở mỗi địa điểm khảo nghiệm cho giá trị tương tự nhau (p < 0.05). Tại các địa điểm khảo nghiệm khác nhau, trạng thái cây của các giống là khác nhau. Lí giải kết quả này, do các địa điểm khác nhau chịu ảnh hưởng của điều kiện thời tiết khác nhau, chênh lệch nhiệt độ ngày đêm khác nhau. Tại Đắk Lắk và Vĩnh Phúc khi thu hoạch cây có bộ lá còn xanh (cả 3 giống khảo nghiệm có giá trị này là 9 điểm). Tại 2 địa điểm còn lại, Nghệ An và Đồng Nai, trị số này đạt từ 7 đến 8 điểm. Tỷ lệ đổ gẫy ghi nhận tại 3 điểm khảo nghiệm (Nghệ an, Đắk Lắk và Đồng Nai) là rất thấp, chỉ từ 0.28 – 0.72 (%). Các giống khác nhau, tỷ lệ gẫy đổ không khác biệt đáng kể. Tuy nhiên, tại Vĩnh Phúc do ảnh hưởng của lốc xoáy (ghi nhận ngày 17/7/2011) dẫn đến tỷ lệ gẫy đổ tương đối cao với c ả 3 giống khảo nghiệm. Cụ

105
thể, giống G1 là 22.80%; giống G2 là 41.50% và thấp nhất là giống G3 với 9.40% cây bị gẫy đổ. Tỷ lệ bật gốc của các giống khảo nghiệm là không đáng kể và không sai khác khi so sánh ở cùng một địa điểm hay giữa 4 địa điểm khảo nghiệm. Như vậy, đánh giá về trạng thái cây, tỷ lệ gẫy đổ và tỷ lệ bật gốc của các giống cho kết quả hoàn toàn tương đồng (nếu không chịu ảnh hưởng của điều kiện thời tiết bất thường, ví dụ tại Vĩnh Phúc).
Năng suất và các yếu tố cấu thành năng suất của 3 g iống ngô khảo nghiệm
được thể hiện trong Bảng 5-30. Số cây thu hoạch trung bình của 3 giống tính chung tại 4 điểm khảo nghiệm là 2620.3 cây. Trong đó, số liệu lần lượt của các giống G1, G2, G3 tại Vĩnh Phúc lần lượt là 1839.5; 1962.5 và 1843.8 cây. Tại Nghệ An là 2273; 2094 và 2023 cây. Tại Đắk Lắk là 3336; 3042 và 3494. Tại Đồng Nai là 3265; 3233 và3060. Số cây thu hoạch giữa 3 giống tại cùng một địa điểm là tương tự nhau, nhưng số cây thu hoạch của từng giống giữa các điểm khảo nghiệm khác nhau có sự chênh lệch đáng kể. Vĩnh Phúc có số cây thu hoạch thấp nhất, tiếp sau là Nghệ An. Lí do bởi trong thời gian khảo nghiệm, tại 2 điểm này bị ảnh hưởng của cơn bão số 2 (ngày 28/6/2011) và lốc xoáy (ngày 17/7/2011). Đắk Lắk và Đồng Nai cho số cây thu hoạch cao hơn và tương đương nhau ở mức tin cậy 95%.
So sánh tỷ lệ hạt/bắp của các giống ở cùng một điểm và giữa 4 điểm khảo nghiệm cho thấy:Tỷ lệ hạt/bắp trung bình của 3 giống tính chung cả 4 điểm là 77.28. Tại Vĩnh Phúc và Nghệ An, 2 giố ng chuyển gen (G1 và G2) có tỷ lệ hạt/bắp cao hơn so với đối chứng. Tuy nhiên, tại Đắk Lắk và Đồng Nai, tỷ lệ này của 2 giống chuyển gen là không cao hơn so với giống nền.
Năng suất thực thu trung bình của 3 giống là 7.40 tấn /hecta. Trong đó năng suất cao nhất thu được Đắk Lắk G1 (9.91 tấn /hecta); G2 (10.50 tấn /hecta) và G3 (10.33 tấ n/hecta). Tại Vĩnh Phúc năng suất của các giống G1 , G2, G3 lần lượt là 6.14; 6.01 và 5.26 tấn /hecta. Tương tự tại Nghệ An là 6.73; 5.58 và5.20 tấn /hecta. Như vậy số liệu ghi nhận tại 2 điểm khảo nghiệm phía Bắc thấp hơn so với 2 điểm phía Nam. Li do bởi ảnh hưởng nhiều của điều kiện thời tiết không thuận.
Như vậy, kết quả đánh giá về các đặc tính nông sinh học của 3 giống tham gia khảo nghiệm cho thấy chúng có đặc điểm tương đồng nhau. Tuy nhiên, tại n hững điểm có áp lực sâu tự nhiên lớn năng suất thực thu của các giống chuyển gen cao hơn hẳn so với giống đối chứng không chuyển gen.

106
Bảng 5-27: Tỷ lệ mọc và sức sống cây con của các giống ngô KNDR
Điểm KN Giống KN Tỷ lệ mọc (%)
Sức sống cây con (điểm 1-9)
Vĩnh Phúc (D1)
*
G1 80.53 7.75 G2 85.83 7.75 G3 81.18 4.00
Nghệ An (D2)
G1 95.65 8.33 G2 92.33 8.48 G3 92.43 8.30
Đắk Lắk (D3)
G1 96.90 9.00 G2 97.50 9.00 G3 97.20 9.00
Đồng Nai (D4)
G1 97.70 7.90
G2 G3
96.50 91.00
7.90 7.60
Trung bình 91.85 7.92 CV (%) 7.30 9.30
LSD.05 (D) 5.60 2.09 LSD.05 (G) 4.80 0.53
LSD.05 (DxG) 9.60 1.06
Ghi chú: G1: giống ngô 30Y87H (mang gen kháng côn trùng cảnh vảy) G2: giống ngô 39Y87HR (Stack mang gen kháng côn trùng cánh vảy và
kháng thuốc trừ cỏ glyphosate) G3: giống ngô 30Y87 - giống ngô nền, đối chứng */
Tại Vĩnh Phúc sau khi gieo bị mưa lớn dẫn đến tỷ lệ mọc của cả 3 giống khảo nghiệm không cao.

107
Bảng 5-28: Một số đặc điểm hình thái của các giống ngô KNDR
Điểm KN
Giống KN
T.G phun râu (ngày sau trồng)
T.G tung phấn (ngày sau trồng)
Cao cây (cm)
Chiều cao đóng bắp (cm)
Vĩnh Phúc (D1)
G1 53.50 52.00 224.40 120.30 G2 54.25 52.00 217.50 112.70 G3 55.10 52.75 219.80 109.10
Nghệ An (D2)
G1 51.53 51.95 220.50 121,90 G2 51.48 51.98 221.98 121.58 G3 51.40 51.73 219.15 121.70
Đắk Lắk (D3)
G1 52.30 52.70 238.50 121.10 G2 52.20 52.60 233.80 119.00 G3 52.30 52.50 240.10 121.20
Đồng Nai (D4)
G1 51.90 51.98 197.30 104.90 G2 51.80 51.98 189.90 101.90 G3 51.90 52.10 197.90 102.30
Trung bình 52.54 52.15 216.64 113.94 CV % 2.00 1.20 3.50 7.00
LSD.05 (D) 1.28 1.44 14.93 13.27 LSD.05 (G) 0.75 0.44 5.44 6.50
LSD.05 (DxG) 1.51 0.88 10.88 12.99
Ghi chú: G1: giống ngô 30Y87H (mang gen kháng côn trùng cảnh vảy) G2: giống ngô 39Y87HR (Stack mang gen kháng côn trùng cánh vảy và
kháng thuốc trừ cỏ glyphosate) G3: giống ngô 30Y87 - giống ngô nền, đối chứng

108
Bảng 5-29: Trạng thái cây và tỷ lệ đổ gẫy của các giống ngô KNDR
Điểm KN Giống KN Trạng thái cây (điểm 1-9)
Tỷ lệ đổ gẫy (%)
Tỷ lệ bật gốc (%)
Vĩnh Phúc (D1)
G1 9.00 22.8 0.09 G2 9.00 41.5 0.12 G3 9.00 9.40 0.09
Nghệ An (D2)
G1 7.50 0.48 0 G2 8.00 0.54 0 G3 7.00 0.72 0
Đắk Lắk (D3)
G1 9.00 0.27 0.02 G2 9.00 0.25 0.02 G3 9.00 0.28 0.02
Đồng Nai (D4)
G1 7.00 0.27 0.05 G2 7.00 0.26 0.06 G3 7.00 0.28 0.08
Trung bình 8.10 6.40 0.05 CV% 5.40
LSD.05 (D) 1.60 LSD.05 (G) 0.51
LSD.05 (DxG) 0.62
Ghi chú: Đánh giá trạng thái
G1: giống ngô 30Y87H (mang gen kháng côn trùng cảnh vảy)
cây theo thang điểm 1-9 (1:không có mô màu xanh; 5: 50% mô lá còn xanh; 9: rất xanh, khoảng 90% mô lá còn xanh)
G2: giống ngô 39Y87HR (Stack mang gen kháng côn trùng cánh vảy và kháng thuốc trừ cỏ glyphosate)
G3: giống ngô 30Y87 - giống ngô nền, đối chứng.

109
Bảng 5-30: Năng suất và các yếu tố cấu thành năng suất các giống ngô KNDR
Điểm KN Giống KN Số cây thu hoạch (cây)
Tỷ lệ hạt/bắp (%)
Năng suất thực thu (tấn/hecta)
Vĩnh Phúc (D1)
G1 *
1839.50 76.25 6.14 G2 1962.50 78.00 6.01 G3 1843.80 74.75 5.26
Nghệ An (D2)
G1 **
2273.00 80.53 6.73 G2 2093.75 78.73 5.85 G3 2023.00 74.28 5.20
Đắk Lắk (D3)
G1 3336.00 74.58 9.91 G2 3043.00 76.15 10.50 G3 3493.50 75.12 10.33
Đồng Nai (D4)
G1 3265.00 77.28 7.70 G2 3233.80 77.35 7.80 G3 3060.00 77.28 7.40
Trung bình 2620.30 76.69 7.40 CV% 11.20 3.30 6.30
LSD.05 (D) 241.22 4.10 0.39 LSD.05 (G) 496.74 1.80 0.34
LSD.05 (DxG) 421.38 3.61 0.67
Ghi chú: G1: giống ngô 30Y87H (mang gen kháng côn trùng cảnh vảy) G2: giống ngô 39Y87HR (Stack mang gen kháng côn trùng cánh vảy và
kháng thuốc trừ cỏ glyphosate) G3: giống ngô 30Y87 - giống ngô nền, đối chứng */ Điểm khảo nghiệm bị ảnh hưởng bởi cơn lốc xoáy ngày 17/7/2011. **/
Điểm khảo nghiệm bị ảnh hưởng bởi cơn bão số 2 ngày 28/6/2011.
5.2.2.2 Kết quả đánh giá tính thích ứng của ngô TC1507 so với đối chứng tại các
vùng sinh thái Việt Nam.
Kết quả đánh giá tính thích ứng (tính mẫn cảm/tính kháng) của ngô TC1507 và các giống đối chứng đối với một số loại sâu bệnh hại chủ yếu tại 4 địa điểm khảo nghiệm được thể hiện trong các Bảng 5-31 đến Bảng 5-34.

110
5.2.2.2.1 Bệnh Đốm lá lớn (Helminthosporium turcicum)
Đốm lá lớn là một trong số các bệnh khá phổ biến trên cây ngô trong điều kiện sinh thái Việt Nam, song tùy từng điều kiện sinh cảnh cụ thể mà mức độ gây hại của bệnh khác nhau. Số liệu Bảng 5-31 thể hiện mức độ bị bệnh Đốm lá lớn trên các giống ngô tại 4 điểm khảo nghiệm. So sánh mức độ bị hại bởi bệnh Đốm lá lớn ở từng điểm khảo nghiệm cho thấy:
Điểm Vĩnh Phúc, bệnh hại ở mức độ nhẹ, cấp hại trung bình từ 1,68 đến 2,24 điểm hay vết bệnh chỉ chiếm 1 – 10 % diện tích lá. M ức độ bị nhiễm bệnh trên cả 3 giống khảo nghiệm là tương đương nhau (mức sác xuất 95%).
Điểm Nghệ An, mức độ bị hại bởi bệnh Đốm lá lớn trên các giống ngô khảo nghiệm cũng cho kết quả tương tự nhau. Cấp độ bệnh hại ghi nhận được đều ở mức trên 2 điểm và dưới 3 điểm.
Điểm Đắk Lắk, cấp hại ghi nhận được là từ 3.00 – 3.25 có nghĩa khoảng từ 11 – 25% diện tích lá ngô bị hại. Các giống ngô khảo nghiệm khác nhau, nhưng cấp hại như nhau ở mức sác xuất 95%.
Điểm Đồng Nai, cấp hại ghi nhận được là cao nhất trong cả 4 điểm khảo nghiệm, từ 4.25 – 4.75 điểm hay khoảng dưới 26% diện tích lá ngô bị hại. Tuy nhiên không có sự khác biệt đáng kể giữa các giống ngô khảo nghiệm về cấp độ nhiễm bênh.
Như vậy, so sánh giá trị trung bình cấp hại của các giống ngô tại 4 địa điểm khảo nghiệm cho thấy: ngô khảo nghiệm tại Vĩnh Phúc bị bệnh nhẹ nhất, tiếp đến là Nghệ An, Đắk Lắk và nặng nhất là 3 giống ngô khảo nghiệm tại Đồng Nai. Tuy nhiên mức độ nhiễm bệnh Đốm lá lớn của cả 3 giống ngô ở những địa điểm khảo nghiệm đều chỉ dưới cấp 5, tức là vết bệnh chỉ chiếm dưới 26% diện tích lá. Nói cách khác, tại cũng địa điểm khảo nghiệm giữa các giống khảo nghiệm khác nhau, giống chuyển gen và giống không chuyển gen, không có sự khác biệt đáng kể về mức độ nhiễm bệnh Đốm lá lớn. Tuy nhiên, tại các địa điểm trồng ngô khác nhau thì mức độ bệnh hại là khác nhau.

111
Bảng 5-31: Mức độ nhiễm bệnh Đốm lá lớn của các giống ngô tại 4 điểm KN
(vụ Hè – Thu, 2011)
Điểm khảo nghiệm
Giống khảo nghiệm
Cấp bệnh hại (1-9)*
CV (%)
LSD.05
Vĩnh Phúc
G1 1.68 25.74 0.72 G2 1.84
G3 2.24
Nghệ An G1 2.68
16.64 0.87 G2 2.75 G3 2.39
Đắk Lắk
G1 3,25
15.34 0.69 G2 3,00 G3 3,25
Đồng Nai
G1 4.25 10.76 0.85 G2 4.54
G3 4.75
Ghi chú:* cấp 0: không có vết bệnh; cấp 1: vết bệnh chiếm 1 – 10% diện tích bộ phận bị hại; cấp 3: vết bệnh chiếm 11 – 25% diện tích bộ phận bị hại ; cấp 5: vết bệnh chiếm 26 – 50% diện tích bộ phận bị hại; cấp 7: vết bệnh chiếm 51 – 75% diện tích bộ phận bị hại; cấp 9: vết bệnh chiếm trên 75% diện tích bộ phận bị hại.
5.2.2.2.2 Bệnh khô vằn (Rhizoctonia solani)
Khô vằn cũng là một trong những loại bệnh hại đáng kể trên ngô trồng ở Việt Nam. Tuy nhiên, tùy điều kiện thời tiết cụ thể mà mức độ là bệnh khác nhau. Kết quả theo dõi bệnh Khô vằn trên 3 giống khảo nghiệm (30Y87H; 30Y87HR và 30Y87) được thể hiện tại Bảng 5-32. So sánh mức độ hại do bệnh khô vằn trên 3 giống ngô khảo nghiệm tại mỗi điểm và cả 4 điểm khảo nghiệm cho thấy trong vụ Hè - Thu năm 2011 tại 4 vùng trồng ngô chính (Vĩnh Phúc, Nghệ An, Đắk Lắc và Đồng Nai) mức độ hại bởi bệnh khô vằn trên các giống ngô khảo nghiệm đều rất nhẹ, từ 0 đến 0.19 hay hầu như không phát hiện thấy vết bệnh Khô vằn trên lá, thân ở cả 3 giống ngô khảo nghiệm.

112
Bảng 5-32: Mức độ nhiễm bệnh Khô vằn trên các giống ngô tại 4 điểm KN
(vụ Hè – Thu, 2011)
Điểm khảo nghiệm
Giống khảo nghiệm
Cấp bệnh hại (1-9)*
CV (%)
LSD.05
Vĩnh Phúc
G1 0.13 23.07 0.25 G2 0.07
G3 0.00
Nghệ An G1 0.16
3.10 0.34 G2 0.19 G3 0.12
Đắk Lắk
G1 0.07 4.21 0.27 G2 0.09
G3 0.12
Đồng Nai G1 0.14
2.16 0.31 G2 0.12 G3 0.17
5.2.2.2.3 Mức độ mẫn/kháng với một số loại sâu hại chính
+ Sâu khoang (Spodoptera)* + Sâu đục thân châu Á (Ostrinia furnacalis)* + Sâu xám (Agrotis ipsilon)* + Sâu đục thân mình hồng ( Sesamia inferens)*
Trong điều kiện áp lực sâu tự nhiên, sâu khoang, sâu đục thân châu Á, sâu xám và sâu đục thân mình hồng là những loại sâ u hại phổ biến trên cây ngô trồng tại các vùng trồng ngô chính của Việt Nam. Tuy nhiên, trong vụ Hè – Thu năm 2011 áp lực sâu tự nhiên tại 2 điểm (Vĩnh Phúc, Nghệ An) trong số 4 điểm khảo nghiệm là tương đối lớn, đặc biệt đối với sâu đục thân và sâu xám. Tại 2 điểm còn lại (Đắk Lắk, Đồng Nai) hầu như không ghi nhận có áp lực sâu tự nhiên. Kết quả ghi nhận mức độ hại của một số loại sâu hại tự nhiên được thể hiện trong Bảng 5-33 và Bảng 5-34.
Sâu khoang: tại Vĩnh Phúc và Nghệ An cả 3 giống ngô khảo nghiệm đều bị sâu khoang gây hại nhẹ, tuy nhiên mức độ bị hại do sâu khoang giữa các giống không khác biệt đáng kể. Tại 2 điểm khảo nghiệm còn lạ i là Đắk Lắk và Đồng Nai đều không có sự gây hại của loại sâu này trên cả 3 giống ngô khảo nghiệm.

113
Sâu đục thâ n châu Á: Giống ngô 30Y87 (G3) là giống không chuyển gen kháng sâu. Tại Vĩnh Phúc giống này bị hại đáng kể bởi sâu đục thân châu Á, điểm hại trung bình ghi nhận được là 6.67, tức là vết thương kéo dài xuất hiện trên ½ số lá. Giống 30Y87H (G1) và 30Y87HR (G2) là các giống ngô chuyển gen kháng côn trùng cánh vảy. Tại Vĩnh Phúc các giống này hầu như không bị sâu đục thân phá hại. Tại 3 điểm khảo nghiệm còn lại là Nghệ An, Đắk Lắk, Đồng Nai hầu như đều không ghi nhận sự gây hại của loại sâu này trên cả 3 giống khảo nghiệm.
Sâu đục thân mình hồng: Tại cả 4 địa điểm khảo nghiệm hầu như không ghi nhận sự xuất hiện và gây hại của loại sâu này trên giống cả 3 giống ngô (cả giống chuyển gen và giống không chuyển gen).
Sâu xám: thuộc họ ngài đêm (Noctuidae), bộ cánh vảy (Lepidoptera). Sâu xám là loại sâu đa thực, có thể phá hại nhiều loại cây trồng khác nhau trên thế giới. Ở nước ta sâu xám phá hại trên câ y ngô, cà chua, đậu, bông vải và các loại họ bầu bí… Trên cây ngô, Sâu tuổi 1- 3 ăn lá ngô non hoặc gặm xung quanh thân ngô. Tuổi 4 trở đi sâu phá hại mạnh, cắn đứt ngang thân ngô non kéo xuống đất. Sâu tuổi 6 mỗi đêm có thể cắn đứt 3 - 4 cây ngô non. Khi cây ngô có 7 - 8 lá, thân cây đã cứng, sâu th ường đục vào thân gần sát gốc ăn phần non mềm ở giữa làm thân cây ngô bị héo và chết. Ruộng ngô bị sâu xám gây hại trông mất khoảng lỗ chỗ, mật độ cây giảm, thiệt hại về năng suất. Sâu xám thường hại nặng trên ngô trồng trên đất cát pha và đất thịt nhẹ. Kết quả đánh giá mức độ bị hại của sâu xám trên 3 giống ngô tại 4 điểm khảo nghiệm được thể hiện ở Bảng 5-34. Trong khảo nghiệm diện rộng, tại Vũ Di (Vĩnh Phúc) sự gây hại của sâu xám (Agrotis ipsilon) được theo dõi đánh giá ngày 17/5/2011 khi cây ngô ở giai đoạn sinh trưởng V2. Tại thời điểm đánh giá, trên tất cả các công thức khảo nghiệm đều không ghi nhận sự xuất hiện gây hại, làm khuyết mật độ cây con do sâu xám gây ra. Tại Nghệ An, sâu xám xuất hiện và gây h ại trên cả 3 giống khảo nghiệm nhưng ở mức độ khác nhau. Trên 2 giống chuyển gen mật độ trung bình 7.50 – 9.00 con/100 cây. Trên giống đối chứng không chuyển gen ghi nhận trung bình cao hơn, đạt 12,5 con/100 cây. Tại Đắk Lắk và Đồng Nai mật độ sâu xám ghi nhận được là không đáng kể trên cả 3 giống tham gia khảo nghiệm, trung bình từ 0 – 0,03 con/100 cây.
Rệp muội ngô (Rhopalosiphum maidis) cũng đã ghi nhận được trên tất cả các giống ngô thí nghiệm tại 4 địa điểm và trong hầu hết các kỳ điều tra, nhất là vào giai đoạn cây ngô bắt đầu trỗ cờ cho đến giai đoạn chín sữa . Tuy vậy, mật độ quần thể cũng như cấp hại của rệp muội ngô ghi nhận là rất thấp. Cấp nhiễm trung bình của rệp muội ngô luôn nhỏ hơn 1 (biến động từ 0,16 đến 0,24) và không có sự khác biệt về

114
cấp nhiễm trung bình giữa các giống ngô chuyển gen và giống ngô không chuyển gen (Bảng 5-34).
Như vậy, tại các điểm trồng khảo nghiệm có áp lực sâu tự nhiên lớn thì 2 giống ngô chuyển gen đã thể hiện được tính kháng sâu đục thân (Vĩnh Phúc). Với sâu xám (tại Nghệ An), giống ngô chuyển gen cũng bị hại nhưng ở mức độ nhẹ hơn. Áp lực sâu xám rất lớn có thể giải thích là vì điểm khảo nghiệm tại Nghệ An là bãi đất phù sa ven sông, thành phần cơ giới đất nhẹ. Cây trồng vụ trước cũng là cây ngô, sau đó chặt bỏ cả khu vực cách ly với bán kính 200 m. Tuy nhiên, khi so sánh năng suất giữa các giống khảo nghiệm tại Nghệ An thì mức độ gây hại do sâu xám không ảnh hưởng đến yếu tố năng xuất. Tóm lại, trên cơ sở số liệu khảo nghiệm thu được cho thấy: các đặc tính nông sinh học của 3 giống tham gia khảo nghiệm là hoàn toàn tương tựu nhau. Tại những điểm có áp lực sâu tự nhiên lớn (ví dụ sâu đục thân), năng suất của các giống chuyển gen cao hơn hẳn so với giống đối chứng không chuyển gen. Ngoài ra, khả năng mẫm/kháng một số sâu bệnh hại chính của 3 giống không có sự khác biệt. Kết luận: tại các điểm trồng khảo nghiệm có áp lực sâu tự nhiên lớn thì 2 giống ngô chuyển gen thể hiện được tính kháng đặc biệt là kháng sâu đục thân.

115
Bảng 5-33: Mức độ bị hại bởi một số loại sâu trên các giống ngô tại 4 điểm KN (vụ Hè – Thu, 2011)
Ghi chú: Thang điểm đánh giá: thang điểm từ 1-9 (Guthrie và cs, 1960). Điểm 0: Hoàn toàn không bị gây hại; 1: Không có biểu hiện gây hại hay những vết cắn với kích thước bằng đầu kim; 2: Số lượng ít vết cắn lỗ tròn nhỏ trên một vài lá; 3: Số vết cắn lỗ tròn nhỏ phổ biến trên một vài lá; 4: Một số lá với các vết cắn tròn nhỏ và một số vết thương dài (hình chữ nhật); 5: Một số lá với vết thương dài khoảng 1,3 đến 2,5 cm; 6: Một số lá với vết thương dài hơn 2.5cm; 7: Vết thương kéo dài xuất hiện trên 1/2 số lá; 8: Vết thương kéo dài xuất hiện trên 2/3 số lá; 9: Hầu hết các lá có các vết thương kéo dài
Điểm khảo nghiệm
Giống khảo nghiệm
Sâu khoang (1-9)*
Sâu đục thân châu Á (1-9)*
Sâu đục thân mình hồng (1-9)*
Vĩnh Phúc
G1 0.03 0.12 0 G2 0.04 0.03 0 G3 0.31 6.67 0
Nghệ An
G1 0.44 0.10 0 G2 0.20 0.03 0 G3 1.30 0.15 0
Đắk Lắk
G1 0 0.05 0 G2 0 0.05 0 G3 0 0.05 0
Đồng Nai
G1 0 0.03 0 G2 0 0.03 0.05 G3 0 0.04 0.11

116
Bảng 5-34: Mức độ bị hại bởi sâu xám, rệp muội ngô của các giống ngô tại 4
điểm khảo nghiệm (vụ Hè – Thu, 2011)
Điểm khảo nghiệm
Giống khảo nghiệm
Sâu xám* (con/100 cây)
Cấp nhiễm rệp muội trung bình
Vĩnh Phúc
G1 0 0.23 a ± 0.01 G2 0 0.16 a ± 0.03 G3 0 0.18 a ± 0.03
LSD.05 0.08
Nghệ An G1 9.00 0.13 a ± 0.05 G2 7.50 0.24 a ± 0.10 G3 12.5 0.22 a ± 0.07
LSD.05 0.26
Đắk Lắk G1 0 0.17 a ± 0.03 G2 0.01 0.19 a ± 0.01 G3 0.03 0.18 a ± 0.02
LSD.05 0.09
Đồng Nai G1 0 0.21 a ± 0.02 G2 0 0.19 a ± 0.02 G3 0.03 0.20 a ± 0.01
LSD.05 0.07
5.2.3 Kết quả đánh giá đa dạng quần thể côn trùng không chủ đích trên ruộng KNDR ngô chuyển gene TC1507 tại các vùng sinh thái Việt Nam
5.2.3.1 Kết quả phân loại theo bộ, họ và loài chân khớp tại 4 địa điểm KNDR
Thành phần loài chân khớp (côn trùng, nhện) được điều tra trực tiếp và bằng sử dụng phương pháp đặt bẫy dính vàng trên ruộng ngô thí nghiệm vào 5 giai đoạn sinh trưởng phát triển chính của cây ngô (Giai đoạn V6, V10, Vt, R1 và R6 ). Số liệu Bảng 5-35 ghi nhận tổng hợp số lượng bộ, họ và loài chân khớp bằng cả 2 phương pháp trên ruộng ngô thí nghiệm diện rộng tại 4 điểm khảo nghiệm.

117
Tại Vũ Di (Vĩnh Phúc), phát hiện được tổng số 86 loài chân khớp , thuộc 54 họ, 13 bộ của lớp côn trùng và lớp hình nhện. Trong đó, chúng tập trung nhiều nhất ở bộ nhện lớn bắt mồi (Araneae, 19 loài) và bộ cánh cứng (Coleoptera, 18 loài). Các bộ cánh đều (Homoptera), cánh nửa (Hemiptera) và cánh vảy (Lepidoptera) có số loài đã phát hiện được tương ứng là 11, 9 và 8 loài. Các bộ khác còn lại có số loài phát hiện được ít hơn, mỗi bộ mới chỉ ghi nhận được từ 1 đến 4 loài (Bảng 5-35).
Tại Phong Thịnh (Thanh Chương, Nghệ An) tổng số loài chân khớp phát hiện được là 83 loài thuộc 49 họ, 11 bộ của lớp côn trùng và lớp hình nhện. Trong đó, bộ cánh cứng (Coleoptera) có số loài đã phát hiện được nhiều nhất (19 loài). Tiếp đến là bộ nhện lớn bắt mồi (Araneae, 14 loài) và bộ cánh vảy (Lepidoptera, 12 loài). Các bộ cánh đều (Homoptera), bộ cánh nửa (Hemiptera) và bộ cánh màng (Hymenoptera) có số loài ghi nhận được tương ứng là 11, 9 và 8 loài. Các bộ khác còn lại có số loài phát hiện được ít hơn, mỗi bộ mới chỉ ghi nhận được từ 1 đến 3 loài (Bảng 5-35).
Tại Nông trường Thống Nhất (Đắk Lắk) tổng số loài chân khớp ghi nhận là 46 loài, thuộc 33 họ, 11 bộ của lớp côn trùng và lớp hình nhện. Trong đó, bộ cánh cứng Coleoptera có số loài được ghi nhận nhiều nhất (9 loài). Bộ nhện lớn bắt mồi (Araneae, 8 loài). Bộ cánh vảy (Lepidoptera, 6 loài). Các bộ cánh đều (Homoptera), bộ cánh nửa (Hemiptera) và bộ cánh màng (Hymenoptera) có số loài đã phát hiện được tương ứng là 5, 6 và 6 loài. Các bộ khác còn lại có số loài phát hiện được ít hơn, mỗi bộ mới chỉ ghi nhận được từ 1 đến 3 loài (Bảng 5-35).
Tại Hưng Lộc (Đồng Nai) phát hiện được tổng số 66 loài chân khớp thuộc 43 họ, 11 bộ của lớp côn trùng và lớp hình nhện. Trong đó, bộ cánh cứng (Coleoptera, 18 loài), bộ nhện lớn bắt mồi (Araneae, 10 loài) bộ cánh vảy (Lepidoptera 8 loài). Các bộ cánh đều (Homoptera), bộ cánh nửa (Hemiptera) và bộ cánh màng (Hymenoptera) có số loài đã phát hiện được tương ứng là 7, 8 và 6 loài. Các bộ khác còn lại có số loài phát hiện được ít hơn, mỗi bộ mới chỉ ghi nhận được từ 1 đến 4 loài (Bảng 5-35).

118
Bảng 5-35: Số lượng loài chân khớp phân theo bộ ghi nhận được trên ruộng ngô KNDR tại 4 địa điểm (vụ Hè - Thu, 2011)
Tên các bộ chân khớp Địa điểm khảo nghiệm
Tên Việt Nam Tên Khoa học Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai Loài Họ Loài Họ Loài Họ Loài Họ
Cánh thẳng Orthoptera 4 2 4 2 2 1 2 2 Cánh da Dermaptera 1 1 1 1 1 1 1 1 Cánh đều Homoptera 11 4 11 4 5 2 7 4 Cánh nửa Hemiptera 9 6 9 4 6 2 8 5 Cánh tơ Thysanoptera 1 1 1 1 1 1 1 1 Cánh cứng Coleoptera 18 8 19 7 9 5 18 7 Cánh mạch Neuroptera 1 1 1 1 1 1 1 1 Cánh vảy Lepidoptera 8 4 12 6 3 3 8 3 Cánh màng Hymeniptera 9 9 8 8 7 6 6 6 Hai cánh Diptera 3 3 3 3 3 3 4 3 Nhện lớn bắt mồi Araneae 19 13 14 12 8 8 10 10 Chuồn chuồn Odonata 1 1 0 0 0 0 0 0 Nhện nhỏ Acarina 1 1 0 0 0 0 0 0
Tổng số 86 54 83 49 46 33 66 43
Bảng 5-36 thể hiện tên Việt Nam và tên khoa học các loài châp khớp bắt gặp trên ruộng khảo nghiệm diện rộng tại 4 địa điểm bằng phương pháp điều tra trực tiếp và phương pháp sử dụng bẫy dính vàng. Các loài chân khớp đ iều tra được phân thành 5 nhóm bao gồm (1) n hóm sâu hại (được chia thành 2 nhóm nhỏ là sâu hại chủ đích và sâu hại không chủ đích; (2) nhóm thiên địch; (3) nhóm côn trùng ký sinh; (4) nhóm côn trùng thụ phấn và (4) nhóm côn trùng chưa rõ ý nghĩa kinh tế.
Tại Vĩnh Phúc điều tra ghi nhận 29 loài thuộc nhóm 1; 31 loài thuộc nhóm 2; 9 loài thuộc nhóm 3; 1 loài thuộc nhóm 4 và 16 loài thuộc nhóm 5.
Tại Nghệ An ghi nhận được 28 loài thuộc nhóm 1; 28 loài thuộc nhóm 2; 8 loài thuộc nhóm 3; 1 loài thuộc nhóm 4 và 18 loài thuộc nhóm 5.
Tại Đắk Lắk ghi nhận được 13 loài thuộc nhóm 1; 18 loài thuộc nhóm 2; 4 loài thuộc nhóm 3; 3 loài thuộc nhóm 4 và 7 loài thuộc nhóm 5.
Tại Nghệ An ghi nhận được 19 loài thuộc nhóm 1; 26 loài thuộc nhóm 2; 4 loài thuộc nhóm 3; 1 loài thuộc nhóm 4 và 10 loài thuộc nhóm 5.

119
Như vậy, tại Vĩnh Phúc và Nghệ An có đa dạng thành phần loài lớn nhất. Thành phần loài nghèo nàn nhất được ghi nhận tại Đắk Lắk
.

120
Bảng 5-36: Thành phần các loài chân khớp bắt gặp trên ruộng ngô KNDR tại 4 địa điểm
(vụ Hè – Thu, 2011)
TT Tên Việt Nam Tên khoa học Điểm khảo nghiệm
V. Phúc N. An Đ. Lăk Đ. Nai
1. Nhóm sâu hại (côn trùng ăn thực vật) 1.1 Sâu hại chủ đích
1 Sâu xám Agrotis ipsilon Hufn 0 + 0 0 2 Sâu xanh Helicorverpa armigera (Hubner) + + + + 3 Sâu cắn gié Mythimna separata (Walk.) + + + + 4 Sâu đục thân Ostrinia furnacalis Guen. + + + + 5 Sâu khoang Spodoptera litura (Farb.) + + + + 6 Sâu keo Spodoptera sp. 0 + 0 0 Tổng 1.1 4 6 4 4 1.2 Sâu hại không chủ đích
1 Châu chấu voi Chondracris rosea rosea (De Geer) + + 0 + 2 Châu chấu mía Hieroglyphus tonkinensis + 0 0 + 3 Châu chấu cánh ngắn Pseudoxya diminuta (Walk) + + 0 + 4 Rầy trắng lớn C0fana spectra (Distant) + + 0 0 5 Rầy xanh lá mạ Empoasca flavescens (Fabr.) + + 0 + 6 Rầy nâu nhỏ Laodelphals striatellus (Fallen) + 0 0 0 7 Rầy lưng trắng Sogatella furcifera (Horv.) + + 0 + 8 Rệp muội bông Aphis gossypii Glover + 0 0 + 9 Rệp muội ngô Rhopalosiphum maidis Fitch + + + + 10 Bọ xít gai vai Cletus sp. + + 0 0 11 Bọ xít 2 chấm trắng lớn Eysarcoris guttiger (Thunb.) + + + 0

121
TT Tên Việt Nam Tên khoa học Điểm khảo nghiệm
V. Phúc N. An Đ. Lăk Đ. Nai 12 Bọ xít xanh Nezara viridula (Linnaeus) + + 0 0 13 Bọ xít dài Leptocorisa oratorius (Fabbricius) 0 + + 0 14 Bọ xít đen Scotinophara lurida (Burmeister) + + 0 0 15 Bọ trĩ Thrips sp. + + + + 16 Mọt cà phê Araecerus fasciculatus (de Geer) + 0 + 0 17 Bọ nhảy đen Chaetocnema basalis Baly + + 0 + 18 Sâu gai ngô Dicladispa armigera (Oliver) + + 0 0 19 Bọ lá 4 vệt Monolepta signata (Oliver) + + + + 20 Mọt ngô đầu dài Sitophilus zeamais Motsch. + + + + 21 Sâu cuốn lá Cnaphalocrocis medinalis (Guen) + + + + 22 Sâu róm chỉ đỏ Euproctis scintillans (Walk) + + + + 23 Cuốn lá loại lớn Pelopidas conjuncta (Her.-Schaf) + 0 0 0 24 Sâu róm lúa Psalis securis Hubner + + + + 25 Ruồi đục lá Liriomyza sp. + + + + 26 Nhện đỏ son Tetranychus cinnabarinus Boisd. + 0 0 0 27 Câu cấu xanh lớn Hypomeces squamosus Fabricius 0 + 0 0 28 Sâu đo (unidentified) 0 + 0 0 29 Dế chó Gryllus testaceus Walker 0 0 + 0 30 Bọ cắt lá Deporaus marginatus P. 0 0 + 0 Tổng 1.2 25 22 9 15 Tổng nhóm 1 29 28 13 19 2. Nhóm thiên địch (côn trùng sử dụng sâu hại làm thức ăn)
1 Chuồn chuồn kim Aciagrion sp. + 0 + + 2 Dế bắt mồi Oecanthus sp. + + 0 0 3 Bọ đuôi kìm Anisolabis annulipes (Lucas) + + + +

122
TT Tên Việt Nam Tên khoa học Điểm khảo nghiệm
V. Phúc N. An Đ. Lăk Đ. Nai 4 Bọ xít mù xanh Cyrtorhinus lividipennis Reuter + + 0 + 5 Bọ xít bắt mồi mắt lồi Geocoris sp. + + 0 0 6 Bọ xít bắt mồi Orius sp. 1 + + + + 7 Bọ xít bắt mồi Geocoris sp. 2 + 0 + + 8 Bọ rùa 8 chấm Leis dimidiate + + 0 + 9 Bọ rùa 2 mảnh đỏ Lemnia biplagiata (Swar.) + + + + 10 Bọ rùa 6 chấm Menochilus sexmaculatus (Fabr.) + + + + 11 Bọ rùa 4 chấm Scymnus frontalis quadrimaculatus Herbst 0 0 + 0 12 Bọ xít hoa Eocanthecona furcellata W. 0 0 + 0 13 Bọ rùa Nhật Bản Propylea japonica (Thunb.) + + + + 14 Bọ rùa Scymnus hoffmanni Weise + + 0 + 15 Cánh cứng cánh ngắn Paederus fuscipes Curtis + + 0 + 16 Bọ 3 khoang Ophionea indica (Thumberg) + + 0 + 17 Bọ mắt vàng Chrysopa sp. + + 0 + 18 Nhện lưới Araneus inustus 1 + 0 + + 19 Nhện lưới Araneus sp. 2 + + 0 + 20 Nhện lớn bụng nhọn Argyrodes sp. + + + 0 21 Nhện lớn Clubiona sp. 1 + + 0 + 22 Nhện lớn Cyclosa sp. 2 0 + 0 0 23 Nhện trắng 8 chấm đen Coleosoma octomaculatum + + 0 + 24 Nhện linh miêu Oxyopes sp. + + + + 25 Nhện sói Pardosa sp. + + 0 + 26 Nhện lớn Runcinia sp. 3 + + + 0 27 Nhện chân dài hàm to Tetragnatha spp. + + 0 + 28 Nhện nhảy Salticidae (unidentified) + + 0 +

123
TT Tên Việt Nam Tên khoa học Điểm khảo nghiệm
V. Phúc N. An Đ. Lăk Đ. Nai 29 Nhện nâu lớn (unidentified) + + 0 + 30 Nhện lớn đa giác (unidentified) + + + + 31 Nhện lớn 4 chấm (unidentified) + + + + 32 Nhện bụng nhọn Argyrodes sp. + + 0 + 33 Nhện lớn bạc (unidentified) + + 0 + 34 Nhện tổng số (unidentified) + 0 0 0 35 Nhện Lycosa Lycosa pseudoannulata 0 0 + 0 36 Nhện lùn Atypena formosana 0 0 + 0 37 Kiến 3 khoang Ophionea nigrofassciata 0 0 + 0 Tổng nhóm 2 31 28 18 26 3. Nhóm côn trùng ký sinh
1 Ong Apanteles sp. 1 + + 0 + 2 Ong2 Charops bicolor (Szep.) + + 0 3 Ong3 Aphenlinidae + + 0 0 4 Ong ký sinh rệp muội Aphidiidae + 0 + 0 5 Ong Mymaridae (unidentified) 4 + + + 0 6 Ong5 Scelionidae (unidentified) + + 0 0 7 Ong ký sinh KS1 (unidentified) 6 + + + + 8 Ong ký sinh sâu non sâu
đục thân (unidentified) + + 0 +
9 Ruồi ký sinh Tachinidae + + + + Tổng nhóm 3 9 8 4 4 4. Nhóm côn trùng thụ phấn
1 Ong mật Apis spp. + + + +

124
TT Tên Việt Nam Tên khoa học Điểm khảo nghiệm
V. Phúc N. An Đ. Lăk Đ. Nai 2 Bướm muồng Catopsilia pomona Fabr. 0 0 + 0 3 Bướm phượng đen Papilio polytes Linnaeus 0 0 + 0 Tổng nhóm 4 1 1 3 1 5. Nhóm côn trùng chưa rõ ý nghĩa kinh tế
1 Rầy xanh lá mạ Empoasca flavescens (Fabr.) + + + 0 2 Rầy xanh đuôi đen Nephotettix nigropictus (Stal) + + 0 0 3 Rầy điện quang Recilia dorsalis Motschulsky + + 0 + 4 Rầy ngực đỏ Thaia subrufa (Motsch.) + + 0 + 5 Rầy nâu Nilaparvata lugen Stal. + + 0 + 6 Bọ phấn trắng Bemisia tabaci Genn. + + 0 0 7 Bọ xít 2 đùi cong mập Physomerus grossipes + + 0 + 8 Bọ bầu vàng Aulacophora femoralis Weise + + + + 9 Bọ nhảy sọc cong Phyllotreta striolata Farb. + + 0 0 10 Ba na xanh Taiwania circumdata Herbst. + 0 0 0 11 Bọ vòi voi (unidentified) + 0 0 0 12 Cánh cứng sp1 (unidentified) + + 0 + 13 Cánh cứng dài (unidentified) + + 0 0 14 Ruồi Sepedon sp. + + + + 15 Bọ xít Bengal Pygomenida bengalensis Westw. 0 + 0 0 16 Bọ xít 2 vai nhọn Cletus sp. + + + + 17 Bọ xít đen Scotinophara lurida (Burmeister) + + 0 + 18 Bọ xít dài (unidentified) 0 + 0 + 19 Câu cấu nâu nhỏ (unidentified) 0 + 0 0 20 Ruồi Sepedol sp. 0 + 0 0

125
TT Tên Việt Nam Tên khoa học Điểm khảo nghiệm
V. Phúc N. An Đ. Lăk Đ. Nai Tổng nhóm 5 16 18 7 10
Tổng số loài 86 83 46 66
Ghi chú: + :bắt gặp; 0: không bắt gặp

126
5.2.3.2 Kết quả điều tra thành phần các loài châp khớp bằng phương pháp trực tiếp trên các giống ngô KNDR.
Thành phần loài chân khớp phát hiện được bằng phương pháp điều tra trực tiếp trên ruộng ngô khảo nghiệm diện rộng vụ Hè – Thu năm 2011 được thể hiện trong Bảng 5-37. Phân tích số liệu cho thấy:
Tại Vĩnh Phúc, - Thành phần loài: tổng số loài chân khớp phát hiện bằng điều tra trực tiếp được
trên 3 giống ngô khảo nghiệm (30Y87H – chuyển gen, 30Y87HR – chuyển gen tham khảo và 30Y87 – đối chứng) tương tự nhau, lần lượt là 59, 61 và 53 (loài). Trong đó nhóm sâu hại chủ đích là 4, 4 và 3 (loài), sâu hại không chủ đích 23, 23 và 22 (loài). Nhóm thiên địch là 25, 25 và 23 (loài), nhóm côn trùng ký sinh là 2, 3 và 3 (loài), nhóm côn trùng thụ phấn là 1, 1 và 1 (loài), loài côn trùng chưa rõ ý nghĩa kinh tế là 8, 9 và 4 (loài).
- Mức độ xuất hiện (hay tần suất bắt gặp): hầu hết các loài bắt gặp ở tấn suất (-) <5%, riêng sâu đục thân ngô trên giống đối chứng không chuyển gen (30Y87) tần suất bắt gặp (+++)>50%, tần suất bắt gặp sâu đục thân trên 2 giống ngô chuyển gen rất ít (hầu như không đáng kể). Tần suất bắt gặp Bọ xít đen, bọ trĩ, bọ xít bắt mồi1, cánh cứng ngắn, nhện lưới1
Như vậy, ngô chuyển gen hay không chuyển gen trồng tại Vĩnh Phúc (áp lực sâu tự nhiên lớn) không ảnh hưởng đến số loài chân khớp và mức độ (tần suất bắt gặp) ngoài trừ đối với sâu đực thân, giống ngô chuyển gen thể hiện được tính kháng rất lớn.
và nhện tổng số trên c ả 3 giống ngô khảo nghiệm rất cao (+++)>50%.
Tại Nghệ An, - Thành phần loài: tổng số loài chân khớp phát hiện bằng điều tra trực tiếp được
trên 3 giống ngô khảo nghiệm (30Y87H – chuyển gen, 30Y87HR – chuyển gen tham khảo và 30Y87 – đối chứng) tương tự nhau, lần lượt là 58, 65 và 59 (loài). Trong đó nhóm sâu hại chủ đích là 5, 6 và 6 (loài), sâu hại không chủ đích 23, 24 và 23 (loài). Nhóm thiên địch là 23, 26 và 24 (loài), nhóm côn trùng ký sinh là 2, 3 và 1 (loài), nhóm côn trùng thụ phấn là 1, 1 và 1 (loài), loài côn trùng chưa rõ ý nghĩa kinh tế là 9, 11 và 10 (loài).
- Mức độ xuất hiện (hay tần suất bắt gặp): đa số các loài bắt gặp ở tấn suất (-) <5%, riêng bọ xít bắt mồi, nhện linh miêu tần suất bắt gặp trung bình (++) 26 – 50%, tần suất bắt gặp bọ rùa Nhật bản, cánh cứng ngắn và nhện lưới 1 trên cả 3 giống ngô khảo nghiệm rất cao (+++)>50%.

127
Tại Đắk Lắk, - Thành phần loài: t ổng số loài chân khớp phát hiện bằng điều tra trực tiếp
được trên 3 giống ngô khảo nghiệm (30Y87H – chuyển gen, 30Y87HR – chuyển gen tham khảo và 30Y87 – đối chứng) tương tự nhau, lần lượt là 47, 44 và 43 (loài). Trong đó nhóm sâu hại chủ đích là 3, 3 và 4 (loài), sâu hại không chủ đích 14, 13 và 13 (loài). Nhóm thiên địch là 18, 17 và 17 (loài), nhóm côn trùng ký sinh là 4, 3 và 3 (loài), nhóm côn trùng thụ phấn là 3, 3 và 3 (loài), loài côn trùng chưa rõ ý nghĩa kinh tế là 5, 5 và 3 (loài). Hầu hết thành phần loài tại điểm Đắk Lắk thấp hơn so với 3 điểm khác.
- Mức độ xuất hiện (hay tần suất bắt gặp): đa số các loài bắt gặp ở tấn suất (-) <5%, riêng 2 loài bọ rùa Nhật Bản và nhện lưới 1
Tại Đồng Nai,
tần suất bắt gặp trung bình (++) 26 – 50% trên cả 3 giống ngô khảo nghiệm.
- Thành phần loài: tổng số loài chân khớp phát hiện bằng điều tra trực tiếp được trên 3 giống ngô khảo nghiệm (30Y87H – chuyển gen, 30Y87HR – chuyển gen tham khảo và 30Y87 – đối chứng) tương tự nhau, lần lượt là 57, 55 và 54 (loài). Trong đó nhóm sâu hại chủ đích là 3, 3 và 4 (loài), sâu hại không chủ đích 14, 15 và 15 (loài). Nhóm thiên địch là 26, 25 và 23 (loài), n hóm côn trùng ký sinh là 4, 4 và 3 (loài), nhóm côn trùng thụ phấn là 1, 1 và 1 (loài), loài côn trùng chưa rõ ý nghĩa kinh tế là 9, 7 và 8 (loài).
- Mức độ xuất hiện (hay tần suất bắt gặp): đa số các loài bắt gặp ở tấn suất (-) <5%, riêng 2 loài (bọ rùa Nhật Bản và nhện lưới 1
Qua phân tích số liệu cho thấy, giống ngô chuyển gen hay không chuyển gen không ảnh hưởng đến thành phần loài và tần suất bắt gặp các loài chân khớp trong vụ khảo nghiệm diện rộng tại 4 địa điểm.
) tần suất bắt gặp trung bình (++) 26 – 50% trên cả 3 giống ngô khảo nghiệm (giống chuyển gen, giống chuyển gen tham khảo và giống đối chứng).
Trong 4 địa điểm khảo nghiệm, điểm Đắk Lắk có số loài và tần suất bắt gặp các loài trên cả 3 giống ngô thấp nhất. 3 điểm khác có số loài trên các g iống tương tự nhau hay mức độ phong phú về loài tương đương nhau.

128
Bảng 5-37: Thành phần, mức độ hiện diện các loài chân khớp điều tra bằng điều tra trực tiếp trên các giống ngô KNDR tại 4 địa điểm(vụ Hè – Thu, 2011)
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
1. Nhóm sâu hại (côn trùng ăn thực vật) 1.1. Sâu hại chủ đích 1 Sâu xám - - - - - - 0 0 0 0 0 0 2 Sâu xanh 0 0 0 - - - - - - - - - 3 Sâu cắn gié - - 0 - - - - - - - - - 4 Sâu đục thân - - +++ - - - 0 0 - 0 0 - 5 Sâu khoang - - - - - - - - - - - - 6 Sâu keo 0 0 0 0 - - 0 0 0 0 0 0 Tổng nhóm 1.1 4 4 3 5 6 6 3 3 4 3 3 4 1.2. Sâu hại không chủ đích 1 Châu chấu voi - - - - - - 0 0 0 - - - 2 Châu chấu mía - - - 0 0 0 0 0 0 - - - 3 Châu chấu cánh ngắn - - - - - - 0 0 0 - - - 4 Rầy trắng lớn - - 0 0 - - 0 0 0 0 0 0 5 Rầy xanh lá mạ 0 0 0 0 0 0 0 0 0 - - - 6 Rầy nâu nhỏ 0 0 - 0 0 0 0 0 0 0 0 0 7 Rầy lưng trắng - - - - - - 0 0 0 0 - - 8 Rệp muội bông - - - 0 0 0 0 0 0 - - - 9 Rệp muội ngô - - - - - - - - - - - - 10 Bọ xít gai vai - - - - - - 0 0 0 0 0 0

129
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
11 Bọ xít 2 chấm trắng lớn - - - - - - - - - 0 0 0 12 Bọ xít xanh - - - - - - 0 0 0 0 0 0 13 Bọ xít dài 0 0 0 - - - - - - 0 0 0 14 Bọ xít đen +++ +++ +++ - - 0 0 0 0 0 0 0 15 Bọ trĩ +++ +++ +++ + + + - - - - - - 16 Mọt cà phê 0 0 0 0 0 0 - - - 0 0 0 17 Bọ nhảy đen - 0 - - - - 0 0 0 - - - 18 Sâu gai ngô 0 0 0 - - 0 0 0 0 0 0 0 19 Bọ lá 4 vệt - - - - - - - - - - - - 20 Mọt ngô đầu dài 0 - - 0 0 0 - - - - - - 21 Sâu cuốn lá nhỏ - - - - 0 - - - - - - - 22 Sâu róm chỉ đỏ - 0 - 0 0 - - - - - - - 23 Cuốn lá loại lớn 0 0 0 0 0 0 0 0 0 0 0 0 24 Sâu róm lúa - - - - - 0 - - - - - - 25 Ruồi đục lá - - - - - - - - - - - - 26 Nhện đỏ son - - - 0 0 0 0 0 0 0 0 0 27 Câu cấu xanh lớn 0 0 0 - - - 0 0 0 0 0 0 28 Sâu đo 0 0 0 - - - 0 0 0 0 0 0 29 Dế chó 0 0 0 0 0 0 - - - 0 0 0 30 Bọ cắt lá 0 0 0 0 0 0 - - - 0 0 0 Tổng nhóm 1.2 19 19 19 18 18 17 14 13 13 14 15 15 Tổng nhóm I 23 23 22 23 24 23 17 16 17 17 18 19

130
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
2. Nhóm thiên địch (côn trùng sử dụng sâu hại làm thức ăn) 1 Chuồn chuồn kim - 0 0 0 0 0 - - - - - - 2 Dế bắt mồi - - - - - - 0 0 0 0 0 0 3 Bọ đuôi kìm - - - - - - - - - - - - 4 Bọ xít mù xanh - - - - + - 0 0 0 - - - 5 Bọ xít bắt mồi mắt lồi - - - 0 - - 0 0 0 0 0 0 6 Bọ xít bắt mồi +++ 1 +++ +++ + + + + + + + + + + + + 7 Bọ xít bắt mồi 0 2 0 0 0 0 0 - - - - - - 8 Bọ rùa 8 chấm 0 - 0 0 - - 0 0 0 - - - 9 Bọ rùa 2 mảnh đỏ - - - - - - - - - - - - 10 Bọ rùa 6 chấm - - - - - 0 - - - - - - 11 Bọ rùa 4 chấm 0 0 0 0 0 0 - - - 0 0 0 12 Bọ xít hoa 0 0 0 0 0 0 - - - 0 0 0 13 Bọ rùa Nhật Bản + - - +++ +++ +++ ++ ++ ++ ++ + ++ 14 Bọ rùa - - - - - - 0 0 0 - - - 15 Cánh cứng cánh ngắn +++ +++ +++ +++ +++ +++ 0 0 0 - 0 - 16 Bọ 3 khoang - - - - - - 0 0 0 0 - 0 17 Bọ mắt vàng 0 - - - - - 0 0 0 - - - 18 Nhện lưới +++ 1 +++ +++ +++ +++ +++ ++ ++ ++ ++ ++ ++ 19 Nhện lưới - 2 - - 0 - - 0 0 0 - - 0 20 Nhện lớn bụng nhọn + + + - - - 0 0 0 - - - 21 Nhện lớn - 1 - - - - - 0 0 0 - - -

131
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
22 Nhện lớn 0 2 0 0 - - - 0 0 0 0 0 0 23 Nhện trắng 8 chấm đen - - - - - - 0 0 0 - 0 - 24 Nhện linh miêu - - - + ++ + - - - - - - 25 Nhện sói - - - - - - 0 0 0 - - 0 26 Nhện lớn - 3 - 0 - - - - + + 0 0 0 27 Nhện chân dài hàm to - - - - - - 0 0 0 - - - 28 Nhện nhảy - - - - - - 0 0 0 - - - 29 Nhện nâu lớn 0 0 0 + + + 0 0 0 - - - 30 Nhện lớn đa giác - - 0 - - 0 - - - - - 0 31 Nhện lớn 4 chấm 0 0 0 0 0 0 - - - - - - 32 Nhện bụng nhọn 0 0 0 0 0 0 0 0 0 - - - 33 Nhện lớn bạc - - - 0 0 0 0 0 0 - - - 34 Nhện tổng số +++ +++ +++ 0 0 0 0 0 0 0 0 0 35 Nhện Lycosa - 0 - 0 0 0 - - - 0 0 0 36 Nhện lùn 0 0 0 0 0 0 - - - 0 0 0 37 Kiến 3 khoang 0 0 0 0 0 0 - - - 0 0 0 Tổng nhóm 2 25 25 23 23 26 24 18 17 17 26 25 23 3. Nhóm côn trùng ký sinh 1 Ong 0 1 0 - 0 - 0 0 0 0 - - - 2 Ong2 0 0 0 0 0 0 0 0 0 0 0 0 3 Ong3 0 0 0 0 0 0 0 0 0 0 0 0 4 Ong ký sinh rệp muội 0 0 0 0 0 0 - - - 0 0 0

132
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
5 Ong 0 4 0 0 0 0 0 - - - 0 0 0 6 Ong5 0 0 0 0 0 0 0 0 0 0 0 0 7 Ong ký sinh - 6 - - 0 0 0 - - - - - - 8 Ong ký sinh sâu non sâu
đục thân - - - - - 0 0 0 0 - - -
9 Ruồi ký sinh 0 - 0 - - - - 0 - - - 0 Tổng nhóm 3 2 3 3 2 3 1 4 3 3 4 4 3 4. Nhóm côn trùng thụ phấn 1 Ong mật - - - + + + + + + + + + 2 Bướm muồng 0 0 0 0 0 0 - - - 0 0 0 3 Bướm phượng đen 0 0 0 0 0 0 - - - 0 0 0 Tổng nhóm 4 1 1 1 1 1 1 3 3 3 1 1 1 5. Loài côn trùng chưa rõ ý nghĩa kinh tế 1 Rầy xanh lá mạ - - - 0 - - - - 0 0 0 0 2 Rầy xanh đuôi đen 0 - 0 - - - 0 0 0 0 0 0 3 Rầy điện quang - - 0 - - 0 0 0 0 - - - 4 Rầy ngực đỏ - - - - - - 0 0 0 - - - 5 Rầy nâu - - 0 - - - 0 0 0 - - - 6 Bọ phấn trắng 0 0 0 0 0 0 0 0 0 0 0 0 7 Bọ xít 2 đùi cong mập 0 0 - 0 0 0 0 0 0 + + - 8 Bọ bầu vàng 0 0 0 0 - - - - - 0 - 0 9 Bọ nhảy sọc cong - 0 - - - - 0 0 0 0 0 0

133
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
10 Ba na xanh 0 0 0 0 0 0 0 0 0 0 0 0 11 Bọ vòi voi 0 - 0 0 0 0 0 0 0 0 0 0 12 Cánh cứng sp1 - - 0 0 0 0 0 0 0 - 0 - 13 Cánh cứng dài - - 0 - - - - - - - - 0 14 Ruồi - - 0 - - - - 0 - 0 - 15 Bọ xít Bengal 0 0 0 - - - 0 0 0 0 0 0 16 Bọ xít 2 vai nhọn 0 0 0 0 0 0 - - - - - - 17 Bọ xít đen 0 0 0 0 0 0 0 0 0 - 0 - 18 Bọ xít dài 0 0 0 0 0 0 0 0 0 0 0 0 19 Câu cấu nâu nhỏ 0 0 0 - - - 0 0 0 0 0 0 20 Ruồi 0 0 0 - - - 0 0 0 0 0 0
Tổng nhóm 4 8 9 4 9 11 10 5 5 3 9 7 8 Tổng số loài 59 61 53 58 65 59 47 44 43 57 55 54
Ghi chú: G1: giống 30Y87H; G2:30Y87HR; G3: 30Y87 Mức độ xuất hiện: (0): Không xuất hiện; ( -) : Rất ít với tần suất bắt gặp < 5%;
(+) : Ít với tần suất bắt gặp: 6 – 25%; (++) : Trung bình với tần suất bắt gặp 26 – 50%; (+++): Nhiều với tần suất bắt gặp >50%

134
5.2.3.3 Kết quả điều tra thành phần các loài châp khớp bằng bẫy dính vàng trên các giống ngô KNDR.
Những loài chân khớp ghi nhận được bằng bẫy dính vàng trên ruộng ngô khảo nghiệm diện rộng vụ Hè-Thu năm 2011 tại 4 điểm được ghi trong Bảng 5-38.
Vĩnh Phúc, về thành phần loài, tổng số loài chân khớp phát hiện bằng bẫy dính vàng trên 3 giống ngô khảo nghiệm (30Y87H – chuyển gen, 30Y87HR – chuyển gen tham khảo và 30Y87 – đối chứng), lần lượt là 38, 43 và 48 loài. Các loài trong nhóm sâu hại chủ đích không vào bẫy dính vàng. Các loài thuộc nhóm sâu hại không chủ đích lần lượt là 7, 8 và 10 loài. Tương tự thành phần loài thuộc nhóm thiên địch là 12, 14 và 16 loài; nhóm côn trùng ký sinh là 9, 9 và 9 loài; nhóm côn trùng thụ phấn chỉ ghi nhận được 1 loài trên cả 3 giống khảo nghiệm; loài côn trùng chưa rõ ý nghĩa kinh tế là 9, 12 và 12loài. Mức độ xuất hiện (hay tần suất bắt gặp), tất cả các loài đều bắt gặp ở tấn suất rất ít (-)/<5%, đến ít (+)/5-26% trên cả 3 giống ngô khảo nghiệm.
Nghệ An, về thành phần loài, tổng số loài chân khớp phát hiện bằng bẫy dính vàng trên 3 giống ngô khảo nghiệm (30Y87H; 30Y87HR; và 30Y87) lần lượt là 45, 43 và 39 loài. Chỉ ghi nhận 1 loài duy nhất (sâu xanh) trong nhóm sâu hại chủ đích vào bẫy dính vàng trên 2 giống ngô chuyển gen. S âu hại không chủ đích ghi nhận được lần lượt là 9, 9 và 8 loài. Nhóm thiên địch là 13, 13 và 11 loài; nhóm côn trùng ký sinh là 9, 9 và 9 loài; nhóm côn trùng thụ phấn cũng chỉ ghi nhận được 1 loài trên cả 3 giống khảo nghiệm. Nhóm côn trùng chưa rõ ý nghĩa kinh tế là 12, 10 và 10 loài. Mức độ xuất hiện (hay tần suất bắt gặp) cũng tương tự như ở Nghệ An, tất cả các loài đều bắt gặp ở tấn suất rất ít (-)/<5%, đến ít (+)/ 5-26% trên cả 3 giống ngô khảo nghiệm.
Đắk Lắk, về thành phần loài, tổng số loài chân khớp phát hiện bằng bẫy dính vàng trên 3 giống ngô khảo nghiệm (30Y87H ; 30Y87HR; và 30Y87) lần lượt là 32, 34 và 34 loài. Trong đó, nhóm sâu hại chủ đích vào bẫy dính vàng trên 3 giống ngô 1, 2 và 3 loài. Nhóm sâu hại không chủ đích lần lượt 10, 11 và 11 loài. Nhóm thiên địch là 13, 14 và 14 loài; nhóm côn trùng ký sinh là 3, 3 và 3(loài). Nhóm côn trùng thụ phấn và nhóm côn trùng chưa rõ ý nghĩa kinh tế đều ghi nhận được 3 loài trên mỗi giống ngô khảo nghiệm. Về mức độ xuất hiện (hay tần suất bắt gặp), tất cả các loài bắt gặp ở tấn suất rất ít (-)/<5%, đến ít (+)/5-26% trên cả 3 giống ngô khảo nghiệm.
Đồng Nai, tổng số loài chân khớp phát hiện bằng bẫy dính vàng trên 3 giống ngô khảo nghiệm (30Y87H; 30Y87HR; và 30Y87) lần lượt là 43, 42 và 48 loài. Trong đó, nhóm sâu hại chủ đích vào bẫy dính vàng trên 3 giống ngô là 1, 1 và 3 loài. Nhóm sâu hại không chủ đích lần lượt 8, 9 và 10 loài. Nhóm thiên địch là 21, 21 và 22 loài , nhóm côn trùng ký sinh là 4, 2 và 4loài. Nhóm côn trùng thụ phấn và nhóm côn trùng

135
chưa rõ ý nghĩa kinh tế ghi nhận lần lượt là 1 và 8 loài đối với cả 3 giống ngô khảo nghiệm. Mức độ xuất hiện (hay tần suất bắt gặp) cũng tương tự các địa điểm khảo nghiệm khác, các loài đều bắt gặp ở tấn suất rất ít (-)/<5%, đến ít (+)/5-26% trên cả 3 giống ngô khảo nghiệm.
Như vậy, số lượng loài chân khớp phát hiện được bằng bẫy dính vàng trên 3 giống ngô khảo nghiện là tương đương nhau. Một số ít loài xuất hiện trên các giống ngô chuyển gen cao hơn, nhưng cũng có một số loài trên giống ngô đối chứng cao hơn. Việc so sánh số loài chân khớp bắt gặp trên bẫy dính vàng ở các điểm cho thấy điểm Đăk Lawk có số lượng loài thấp hơn so với 3 địa điểm còn lại. Số lượng loài vào bẫy dính vàng trên 3 giống tại 4 địa điểm khảo nghiệm diện rộng vụ Hè - Thu năm 2011 đều nhiều hơn so với trên giống ngô tương ứng trong khảo nghiệm hạn chế vụ thu và vụ đông năm 2010 tại Văn Giang (24-33 loài trên giống 30Y87 và 22-29 loài trên giống 30Y87H)
.

136
Bảng 5-38: Thành phần, mức độ hiện diện loài chân khớp điều tra bằng bẫy dính vàng trên các giống ngô KNDR tại 4 địa điểm (vụ Hè - Thu, 2011)
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
1. Nhóm sâu hại (côn trùng ăn thực vật) 1.1. Sâu hại chủ đích 1 Sâu xanh 0 0 0 + + 0 0 0 - 0 0 0 2 Sâu cắn gié 0 0 0 0 0 0 - - 0 - - - 3 Sâu đục thân 0 0 0 0 0 0 0 0 - 0 0 - 4 Sâu khoang 0 0 0 0 0 0 0 - - 0 0 - Tổng nhóm 1.1 0 0 0 1 1 0 1 2 3 1 1 3 1.2. Sâu hại không chủ đích 1 Rầy trắng lớn + + + + + + 0 0 0 0 0 0 2 Rầy xanh lá mạ 0 0 0 0 0 0 0 0 0 - - - 3 Rầy lưng trắng + + + + + + 0 0 0 - - - 4 Rệp muội bông 0 0 0 0 0 0 - - - 0 - 0 5 Rệp muội ngô + + + + + + - + - + - - 6 Bọ xít 2 chấm trắng lớn 0 0 0 0 0 0 - - - 0 0 0 7 Bọ xít xanh 0 0 0 0 0 + 0 0 0 0 0 0 8 Bọ xít dài 0 0 0 0 0 0 0 - 0 0 0 0 9 Bọ trĩ + + + + + 0 - 0 - - - - 10 Mọt cà phê 0 0 0 0 0 0 - - - 0 0 0 11 Bọ nhảy đen + + + + + + 0 0 0 - - - 12 Sâu gai ngô 0 0 + 0 0 0 0 0 0 0 0 0

137
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
13 Bọ lá 4 vệt + + + + + + - - 0 0 - - 14 Mọt ngô đầu dài 0 + + + + + + + + - - - 15 Sâu cuốn lá nhỏ 0 0 0 0 0 0 + + + - 0 - 16 Sâu róm chỉ đỏ 0 0 0 0 0 0 0 0 0 0 - - 17 Cuốn lá loại lớn 0 0 + + + + 0 0 0 0 0 0 18 Ruồi đục lá + + + + + 0 + - - - 0 + Tổng nhóm 1.2 7 8 10 9 9 8 9 9 8 8 9 10 Tổng nhóm 1 7 8 10 10 10 8 10 11 11 9 10 13 2. Nhóm thiên địch (côn trùng sử dụng sâu hại làm thức ăn) 1 Chuồn chuồn kim 0 0 0 0 0 0 - - - - - - 2 Dế bắt mồi 0 + + + + + 0 0 0 0 0 0 3 Bọ đuôi kìm 0 0 0 0 + 0 - + - + - - 4 Bọ xít mù xanh + 0 + + + + 0 0 0 0 0 0 5 Bọ xít bắt mồi mắt lồi + + + 0 + 0 0 0 0 0 0 0 6 Bọ xít bắt mồi + 1 + + + + + + - + - - - 7 Bọ rùa 8 chấm 0 + + + + + 0 0 0 + + + 8 Bọ rùa 2 mảnh đỏ 0 0 0 0 + 0 - - - - - - 9 Bọ rùa 6 chấm + + + + + + + + + + + + 10 Bọ rùa 4 chấm 0 0 0 0 0 0 - - - 0 0 0 11 Bọ rùa Nhật Bản + + + + + + + + + + - - 12 Bọ rùa + + + + + + 0 0 0 - - - 13 Cánh cứng cánh ngắn + + + + + + 0 0 0 - - -

138
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
14 Bọ 3 khoang + + + + + + 0 0 0 - - - 15 Bọ mắt vàng 0 0 0 0 0 0 0 0 0 - - - 16 Nhện lưới + 1 + + + 0 + + + + 0 - + 17 Nhện lưới 0 2 0 0 0 0 0 0 0 0 - + + 18 Nhện lớn bụng nhọn 0 + 0 0 0 0 + + + 0 0 0 19 Nhện lớn 1 0 0 + 0 0 0 + - - 20 Nhện lớn 0 2 0 0 0 0 0 0 0 0 0 0 0 21 Nhện trắng 8 chấm đen 0 0 0 0 0 0 0 0 0 - - - 22 Nhện linh miêu 0 + + + 0 0 + + + - + - 23 Nhện sói + + + 0 0 0 0 0 0 - - - 24 Nhện lớn 0 3 0 + 0 0 0 0 0 0 0 0 0 27 Nhện chân dài hàm to + 0 + 0 0 0 0 0 0 + + + 25 Nhện nhảy 0 0 0 + 0 0 0 0 0 - - - 26 Nhện nâu lớn + + + + + + 0 0 0 + + + 27 Nhện lớn đa giác 0 0 0 0 0 0 - - - - 0 - 28 Nhện bụng nhọn 0 0 0 0 0 0 0 0 0 - - - 29 Nhện Lycosa 0 0 0 0 0 0 0 - - 0 0 0 30 Nhện lùn 0 0 0 0 0 0 - - - 0 0 0 31 Kiến 3 khoang 0 0 0 0 0 0 + + + 0 0 0 Tổng nhóm 2 12 14 16 13 13 11 13 14 14 21 21 22 3. Nhóm côn trùng ký sinh 1 Ong + 1 + + + + + 0 0 0 + 0 +

139
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
2 Ong2 + + + + + + 0 0 0 0 0 0 3 Ong3 + + + + + + 0 0 0 0 0 0 4 Ong ký sinh rệp muội + + + + + + 0 0 0 0 0 0 5 Ong + 4 + + + + + + + + 0 0 0 6 Ong5 + + + + + + 0 0 0 0 0 0 7 Ong ký sinh + 6 + + + + + + + + + + + 8 Ong ký sinh sâu non sâu đục
thân + + + + + + 0 0 0 + + +
9 Ruồi ký sinh + + + + + + - - - - 0 - Tổng nhóm III 9 9 9 9 9 9 3 3 3 4 2 4 4. Nhóm côn trùng thụ phấn 1 Ong mật + + + + + + + + + + + + 2 Bướm muồng 0 0 0 0 0 0 + + + 0 0 0 3 Bướm phượng đen 0 0 0 0 0 0 + + + 0 0 0 Tổng nhóm IV 1 1 1 1 1 1 3 3 3 1 1 1 5. Nhóm loài côn trùng chưa rõ ý nghĩa kinh tế 1 Rầy xanh lá mạ + + + + + + + + + 0 0 0 2 Rầy xanh đuôi đen + + + + + + 0 0 0 0 0 0 3 Rầy điện quang + + 0 + + + 0 0 0 + + + 4 Rầy ngực đỏ + + + + + + 0 0 0 - + - 5 Rầy nâu + + + + + + 0 0 0 + + + 6 Bọ phấn trắng + + + + + 0 0 0 0 0 0 0

140
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
7 Bọ xít 2 đùi cong mập 0 0 0 0 0 0 0 0 0 0 0 0 8 Bọ bầu vàng 0 0 + + 0 + + + - + - - 9 Bọ nhảy sọc cong + + + + + + 0 0 0 0 0 0 10 Ba na xanh 0 + + 0 0 0 0 0 0 0 0 0 11 Bọ vòi voi 0 + + 0 0 0 0 0 0 0 0 0 12 Cánh cứng sp1 + + + 0 0 0 0 0 0 + + + 13 Cánh cứng dài 0 + + + + + 0 0 0 0 0 0 14 Ruồi + + + 0 0 0 + - + - - - 15 Bọ xít Bengal 0 0 0 + 0 0 0 0 0 0 0 0 17 Bọ xít đen 0 0 0 0 0 0 0 0 0 + + + 18 Bọ xít dài 0 0 0 0 0 0 0 0 0 - - - 19 Câu cấu nâu nhỏ 0 0 0 + + + 0 0 0 0 0 0 20 Ruồi 0 0 0 + + + 0 0 0 0 0 0
Tổng nhóm V 9 12 12 12 10 10 3 3 3 8 8 8 Tổng số loài 38 43 48 45 43 39 32 34 34 43 42 48
Ghi chú: G1: Giống 30Y87H; G2:30Y87HR; G3: 30Y87 Mức độ xuất hiện: (0): Không xuất hiện; ( -) : Rất ít với tần suất bắt gặp < 5%; (+) : Ít với tần suất bắt gặp: 6 – 25%; (++) : Trung bình với tần suất bắt gặp 26 – 50%; (+++): Nhiều với tần suất bắt gặp >50%

141
5.2.3.4 Kết quả điều tra thành phần các loài châp khớp bằng điều tra trực tiếp và
bẫy dính vàng trên các giống ngô KNDR
Trên đồng ruộng, một số loài chân khớp vừa có thể quan sát trực tiếp được trên trên cây ngô vừa vào bẫy dính vàng như (ví dụ rầy lưng trắng (Sogatella furcifera), rệp muội ngô (Rhopalosiphum maidis), bọ trĩ (Thrips sp.), bọ nhảy đen nhỏ (Chaetocnema basalis), bọ lá 4 vệt (Monolepta signata), bọ rùa Nhật Bản (Propylea japonica), cánh cứng ngắn (Paederus fuscipes, vv…). Tuy nhiên, trong thực tế có nhiều loài có kích thước nhỏ khó có thể phát hiện bằng mắt thường khi điều tra trực tiếp, nhưng đã thu được bằng bẫy dính vàng như (ví dụ các loài ong ký sinh thuộc họ Aphenlinidae, Aphidiidae, Mymaridae, Scelionidae). Kết quả điều tra các loài chân khớp ghi nhận được bằng cả 2 phương pháp (quan sát trực tiếp và bẫy dính vàng) được thể hiện trong Bảng 5-39.
Tại Vũ Di (Vĩnh Phúc) đã phát hiện được tổng số 86 loài chân khớp . Số lượng loài chân khớp phát hiện được b ằng cả hai phương pháp trên ngô khảo nghiệm diện rộng vụ Hè-Thu năm 2011 tại Vũ Di trên mỗi giống đều cao hơn so với trên từng giống ngô đó trong khảo nghiệm hạn chế trong vụ thu năm 2010 tại Văn Giang (47 loài trên giống 30Y87 và 44 loài trên giống 30Y87H). Số lượng loài ghi nhận được bằng cả hai phương pháp trên cả hai giống ngô biến đổi gen gần như tương tự nhau, lần lượt là 71 và 68 loài. Trong khi đó, trên giống ngô nền không biến đổi gen , 30Y87, số loài chân khớp ghi nhận được 78 loài, nhiều hơn đôi chút so với các giống ngô chuyển gen . Trên các giống ngô khảo nghiệm d iện rộng tại Vĩnh Phúc có thành phần sâu hại và thiên địch nghèo nàn hơn so với thành phần loài sâu hại và thiên địch trên cây ngô đã công bố ở miền Bắc trước đây (Nguyễn Quý Hùng và nnk., 1978; Phạm Văn Lầm, 1996; Viện Bảo vệ thực vật 1976). Có những loài sâu hại rất phổ biến trên cây ngô như sâu xám (Agrotis ipsilon Hufn) nhưng lại không bắt gặp trong thí nghiệm này. Nguyên nhân có thể do khảo nghiệm diện rộng lần này được bắt đầu muộn hơn đôi chút (ngày gieo hạt 7/5/2011) so với trồng ngô thông thường tại đây.
Tại Nghệ An, tổng số các loài chân khớp ghi nhận được trên 3 giống ngô khảo nghiệm (30Y87H-chuyển gen; 30Y87HR-chuyển gen tham khảo; và 30Y87-đối chứng) lần lượt là 75, 76 và 73 loài. Trong đó, nhóm sâu hại chủ đích là 6 loài và tương tự nhau trên cả 3 giống. Nhóm sâu hại không chủ đích lần lượt là 26, 26 và 25 loài. Nhóm thiên địch ghi nhận được là 26, 27 và 25 (loài). Các nhóm côn trùng ký sinh, nhóm côn trùng thụ phấn và nhóm côn trùng chưa rõ ý nghĩa kinh tế đều có số loại ghi nhân được tương đương nhau trên cả 3 giống ngô khảo nghiệm và lần lượt là là 9, 1 và 13 loài đối với từng nhóm.

142
Tại Đắk Lắk, đã ghi nhận được tổng số các loài chân khớp trên 3 giống ngô khảo nghiệm lần lượt là 40, 39 và 42 loài. Trong đó nhóm sâu hại chủ đích là 1, 2 và 4 loài và sâu hại không chủ đích là 14, 14 và 17 loài. Nhóm thiên địch ghi nhận được lần lượt là 15, 14 và 15 loài. Nhóm côn trùng ký sinh và nhóm côn trùng thụ phấn ghi nhận số lượng loài trên cả 3 giống khảo nghiệm là như nhau, lần lượt là 4 và 3 loài. Nhóm côn trùng chưa rõ ý nghĩa kinh tế cũng tương tự nhau trên 3 giống ngô khảo nghiệm, lần lượt là 4, 4 và 3 loài. Thành phần loài chân khớp thu được tại Đắk Lắk nghèo hơn so với thành phần các loài chân khớp thu được tại 3 địa điểm còn lại (Vính Phúc, Nghệ An và Đồng Nai).
Tại Đồng Nai, tổng số các loài chân khớp ghi nhận được bằng 2 phương pháp đối với 3 giống khảo nghiệm là 55, 56 và 54 loài. Trong đó, sâu hại chủ đích là 3, 2 và 4 loài; sâu hại không chủ đích 16, 15 và 20 loài. Nhóm thiên địch lần lượt là 23, 26 và 21 loài. Nhóm côn trùng chưa rõ ý nghĩa kinh tế là 10, 10 và 7 loài. Số loài trên cả 3 giống khảo nghiệm là như nhau đối với cả 2 nhóm còn lại, nhóm côn trùng ký sinh và nhóm côn trùng thụ phấn, lần lượt là 5và 1 loài.
Như vậy, có thể kết luận là thành phần các loài chân khớp bắt gặp bằng cả 2 phương pháp điều tra trên 3 giống ngô khảo nghiệm là không có sự khác biệt đáng kể. Nói cách khác là giữa các giống ngô chuyển gen và giống đối chứng không chuyển gen đều không có sự khác biệt về thành phần các loài chân khớp dù là điều tra bằng phương pháp quan sát trực tiếp hay sử dung bẫy dính vàng. Các giống ngô chuyển gen hoặc hoàn toàn không ảnh hưởng đến thành phần loài chân khớp.

143
Bảng 5-39: Thành phần, mức độ hiện diện các loài chân khớp trong điều tra trực tiếp và bẫy dính vàng trên các giống ngô KNDR tại 4 địa điểm (vụ Hè - Thu, 2011)
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
1. Nhóm sâu hại (côn trùng ăn thực vật) 1.1. Sâu hại chủ đích 1 Sâu xám 0 0 0 - - - 0 0 0 0 0 0 2 Sâu xanh - - - - - - - - - - - - 3 Sâu cắn gié - - 0 - - - 0 - - - 0 - 4 Sâu đục thân - - +++ - - - 0 0 - 0 - - 5 Sâu khoang 0 0 - - - + 0 0 - - 0 - 6 Sâu keo 0 0 0 - + ++ 0 0 0 0 0 0 Tổng nhóm 1.1 3 3 3 6 6 6 1 2 4 3 2 4 1.2. Sâu hại không chủ đích 1 Châu chấu voi - - - - - - 0 0 0 - - - 2 Châu chấu mía - - - 0 0 0 0 0 0 + + + 3 Châu chấu cánh ngắn - - - - - - 0 0 0 - 0 - 4 Rầy trắng lớn - - - - - - 0 0 0 0 0 0 5 Rầy xanh lá mạ - 0 - 0 0 0 0 0 0 + + + 6 Rầy nâu nhỏ - 0 0 0 0 0 0 0 0 0 0 0 7 Rầy lưng trắng ++ +++ +++ - + + 0 0 0 - - - 8 Rệp muội bông - - - 0 0 0 0 0 0 + + + 9 Rệp muội ngô + + + - + - - - - + + + 10 Bọ xít gai vai - - - - - - 0 0 0 0 0 0

144
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
11 Bọ xít 2 chấm trắng lớn - - - - - - + + + 0 0 0 12 Bọ xít xanh - - - - - - 0 0 0 0 0 0 13 Bọ xít dài 0 0 0 - - - - - - 0 0 0 14 Bọ xít đen +++ +++ +++ - - - 0 0 0 0 0 0 15 Bọ trĩ ++ ++ + + ++ + + + + - - + 16 Mọt cà phê - - - 0 0 0 + + + 0 0 0 17 Bọ nhảy đen - - - +++ +++ +++ 0 0 0 - 0 - 18 Sâu gai ngô 0 0 - - - 0 0 0 0 0 0 0 19 Bọ lá 4 vệt - - - + ++ + - 0 - 0 - - 20 Mọt ngô đầu dài 0 0 - - - - + + + - - - 21 Sâu cuốn lá nhỏ - - - - - - - 0 - - - - 22 Sâu róm chỉ đỏ 0 0 - 0 0 - - - - - 0 - 23 Cuốn lá loại lớn - 0 - 0 0 0 - - 0 0 - - 24 Sâu róm lúa - - - - - 0 0 - - 0 - - 25 Ruồi đục lá - - - - - - - - - - - - 26 Nhện đỏ son - - - 0 0 0 0 0 0 0 0 0 27 Câu cấu xanh lớn 0 0 0 - - - 0 0 0 0 0 0 28 Sâu đo 0 0 0 - - - 0 0 0 0 0 0 29 Dế chó 0 0 0 0 0 0 - - - 0 0 0 30 Bọ cắt lá 0 0 0 0 0 0 - - - 0 0 0 Tổng nhóm 1.2 21 19 23 20 20 19 13 12 13 13 13 16 Tổng nhóm I 24 22 29 26 26 25 14 14 17 16 15 20

145
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
2. Nhóm thiên địch (côn trùng sử dụng sâu hại làm thức ăn) 1 Chuồn chuồn kim - 0 0 0 0 0 + + + + + + 2 Dế bắt mồi - - - - + - 0 0 0 0 0 0 3 Bọ đuôi kìm - - - - - - - - - + - - 4 Bọ xít mù xanh - - - - ++ - 0 0 0 - 0 0 5 Bọ xít bắt mồi mắt lồi - - - - - - 0 0 0 0 0 0 6 Bọ xít bắt mồi +++ 1 +++ +++ +++ +++ +++ ++ ++ ++ +++ ++ ++ 7 Bọ xít bắt mồi 0 2 0 - 0 0 0 - - - - - - 8 Bọ rùa 8 chấm - - - - - - 0 0 0 + + 0 9 Bọ rùa 2 mảnh đỏ - - - - - - - 0 - 0 - 0 10 Bọ rùa 6 chấm - - - - - - + - - - + - 11 Bọ rùa đen nhỏ 0 0 0 - - - 0 0 0 0 - 0 12 Bọ xít hoa 0 0 0 0 0 0 - - - 0 0 0 13 Bọ rùa Nhật Bản + + + +++ +++ +++ ++ ++ + ++ + ++ 14 Bọ rùa - - - - - - 0 0 0 - - - 15 Cánh cứng cánh ngắn +++ +++ +++ +++ +++ +++ 0 0 0 + 0 0 16 Bọ 3 khoang - - - - + - 0 0 0 - - - 17 Bọ mắt vàng 0 - - - - - 0 0 0 + + + 18 Nhện lưới +++ 1 +++ +++ +++ +++ +++ 0 0 0 ++ ++ ++ 19 Nhện lưới +++ 2 + ++ - - - 0 0 0 - - - 20 Nhện lớn bụng nhọn + + + - - - + + + 0 0 0 21 Nhện lớn - 1 - - - - - 0 0 0 0 - -

146
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
22 Nhện lớn - 2 - - - - 0 0 0 0 - - 0 23 Nhện trắng 8 chấm đen + + + - - - 0 0 0 - - - 24 Nhện linh miêu - - - + ++ + + + + - - - 25 Nhện sói 0 0 0 - - - 0 0 0 0 - - 26 Nhện lớn 0 3 0 0 - - - - - 0 0 0 0 27 Nhện chân dài hàm to - - - - - - 0 0 0 - - - 28 Nhện nhảy - - - - - - 0 0 0 - - 0 29 Nhện nâu lớn +++ +++ +++ ++ +++ ++ 0 0 0 - - - 30 Nhện lớn đa giác - - 0 0 0 0 + + + ++ ++ ++ 31 Nhện lớn 4 chấm - 0 - 0 0 0 + ++ + - - - 32 Nhện bụng nhọn 0 0 0 0 0 0 0 0 0 0 - - 33 Nhện lớn bạc 0 - - 0 - 0 0 0 0 - + + 34 Nhện tổng số - - - 0 0 0 0 0 0 0 0 0 35 Nhện Lycosa 0 0 0 0 0 0 - - - 0 0 0 36 Nhện lùn 0 0 0 0 0 0 - - + 0 0 0 37 Kiến 3 khoang 0 0 0 0 0 0 + + - 0 0 0 Tổng nhóm II 26 26 27 26 27 25 15 14 15 23 26 21 3. Nhóm côn trùng ký sinh 1 Ong - 1 - - - - - 0 0 0 - - - 2 Ong2 - - - + + + 0 0 0 0 0 0 3 Ong3 - - - - - - 0 0 0 0 0 0 4 Ong ký sinh rệp muội - - - - - - - - - 0 0 0

147
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
5 Ong + 4 + + - - - + + - 0 0 0 6 Ong5 - - - - - - + + + - - - 7 Ong ký sinh +++ 6 +++ +++ - - - 0 0 0 + - - 8 Ong KS sâu non sâu đục thân - - - - - - 0 0 0 - - - 9 Ruồi ký sinh ++ + +++ - - - - - - + - - Tổng nhóm 3 9 9 9 9 9 9 4 4 4 5 5 5 4. Nhóm côn trùng thụ phấn 1 Ong mật - - - + + + + + + - + - 2 Bướm muồng 0 0 0 0 0 0 - - - 0 0 0 3 Bướm phượng đen 0 0 0 0 0 0 + - - 0 0 0 Tổng nhóm 4 1 1 1 1 1 1 3 3 3 1 1 1 5. Nhóm loài côn trùng chưa rõ ý nghĩa kinh tế 1 Rầy xanh lá mạ ++ + ++ - - - + - - 0 0 0 2 Rầy xanh đuôi đen - - - - - - 0 0 0 0 0 0 3 Rầy điện quang - - 0 - - - 0 0 0 - - 0 4 Rầy ngực đỏ +++ +++ +++ +++ +++ +++ 0 0 0 - + - 5 Rầy nâu - - - - - - 0 0 0 - - 0 6 Bọ phấn trắng +++ +++ +++ - - - 0 0 0 0 0 0 7 Bọ xít 2 đùi cong mập 0 0 - 0 0 0 0 0 0 - - - 8 Bọ bầu vàng 0 0 - - - - + + + - + - 9 Bọ nhảy sọc cong - 0 - - - - 0 0 0 0 0 0 10 Ba na xanh 0 0 - 0 0 0 0 0 0 0 0 0

148
TT Tên loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
11 Bọ vòi voi - - 0 0 0 0 0 0 0 0 0 0 12 Cánh cứng sp1 - - - - - - 0 0 0 + + - 13 Cánh cứng dài - - - - - - 0 0 0 0 0 0 14 Ruồi + + ++ 0 0 0 + + ++ - - ++ 15 Bọ xít Bengal 0 0 0 - - - 0 0 0 0 0 0 16 Bọ xít 2 vai nhọn 0 0 0 0 0 0 + - 0 - - - 17 Bọ xít đen 0 0 0 0 0 0 0 0 0 - + + 18 Bọ xít dài 0 0 0 0 0 0 0 0 0 - - 0 19 Câu cấu nâu nhỏ 0 0 0 - - - 0 0 0 0 0 20 Ruồi 0 0 0 - - - 0 0 0 0 0 0
Tổng nhóm V 11 10 12 13 13 13 4 4 3 10 10 7
Tổng số loài 71 68 78 75 76 73 40 39 42 55 56 54
Ghi chú: G1: giống 30Y87H; G2:30Y87HR; G3: 30Y87 Mức độ xuất hiện: (0): Không xuất hiện; ( -) : Rất ít với tần suất bắt gặp < 5%; (+) : Ít với tần suất bắt gặp: 6 – 25%; (++) : Trung bình với tần suất bắt gặp 26 – 50%; (+++): Nhiều với tần suất bắt gặp >50%

149
5.2.3.5 Kết quả đánh giá sự đa dạng sinh học ở cấp độ loài của tập hợp chân khớp
trên ruộng ngô KNDR
Sử dụng công thức của Odum (1971), Colwell (2000) để tính toán một số chỉ tiêu chung về sự đa dạng sinh học ở cấp độ loài của tập hợp chân khớp trong ruông ngô khảo nghiệm, kết quả thể hiện trong Bảng 5-40.
Tại Vĩnh Phúc, tổng số cá thể/lần điều tra trên 3 giống ngô (30Y87H-chuyển gen; 30Y87HR-chuyển gen tham khảo và 30Y87-đối chứng) lần lượt là 464.9; 467.9 và 609.3 cá thể. Trị số này trên giống ngô đối chứng là lớn hơn nhiều so với 2 giống ngô chuyển gen , lí do bởi giống đối chứng bị sâu đục thân gây hại rất nhiều. Tuy nhiên, tổng số loài/lần điều tra trên 3 giống là tương đương nhau, đều trong khoảng từ 29.8 – 30.1 loài. Chỉ số ưu thế Simpson's D trên các giống ngô khảo nghiệm gần tương tự nhau và rất thấp, chỉ từ 0.14 -0.18. Chỉ số đa dạng H’ đạt từ 4.55- 4.85.
Tại Nghệ An, tổng số cá thể/lần điều tra trên 3 giống ngô lần lượt là 407; 394 và 469.5 cá thể . Không có sự khác biệt đáng kể về tổng số loài/lần điều tra trên 3 giống, từ 29.41- 32.47 loài. Chỉ số ưu thế Simpson's D trên các giống ngô khảo nghiệm gần tương tự nhau và rất thấp, từ 0.15 -0.17. Chỉ số đa dạng H’ đạt từ 4.96 - 4.13.
Tại Đắk Lắk, tổng số cá thể/lần điều tra trên 3 giống ngô lần lượt là 206.7; 213.2 và 231.2 cá thể. Tổng số loài/lần điều tra trên 3 giống tương đương nhau, đạt từ 19.41- 22.37 loài. Chỉ số ưu thế Simpson's D trên các giống ngô khảo nghiệm gần tương tự nhau và rất thấp, từ 0.10 -0.12. Chỉ số đa dạng H’ đạt từ 4.01 - 4.76.
Tại Đắk Lắk, tổng số cá thể/lần điều tra trên 3 giống ngô chuyển gen, chuyển gen tham khảo và đối chứng lần lượt là 296.6; 313.2 và 331.3 cá thể. Tổng số loài/lần điều tra trên 3 giống là tương đương nhau và trong khoảng từ 19.22- 21.39 loài. Chỉ số ưu thế Simpson's D trên các giống ngô khảo nghiệm gần tương tự nhau và rất thấp, từ 0.11 -0.13. Chỉ số đa dạng H’ đạt từ 4.51 - 4.76.
Phân tích số liệu trên cho thấy: trong khảo nghi ệm ở Vĩnh Phúc, giống ngô chuyển gen 30Y87H có chỉ số ưu thế Simpson's D thấp nhất (0,14) và có chỉ số đa dạng Shannon H' cao nhất (4,85). Kết quả tương tự trong các khảo nghiệm ở Nghệ An, Đắk Lắk và Đồng Nai. Các giá trị chỉ số ưu thế Simpson's D hoàn toàn lôgíc, tương ứng phù hợp với giá trị của chỉ số đa dạng Shannon H'. Kết quả này chứng tỏ trong sinh quần các giống ngô khảo nghiệm không có loài chân khớp nào phát triển với số lượng nổi trội hơn, chiếm ưu thế hơn để có thể gây mất cân bằng sinh thái. Giá trị chỉ số ưu thế Simpson's D của các giống ngô thí nghiệm khác nhau ở mức rất nhỏ, không có ý nghĩa thống kê.

150
Bảng 5-40: Chỉ số đa dạng và chỉ số ưu thế các loài chân khớp bắt gặp trên các giống ngô KNDR tại 4 địa điểm (vụ Hè -Thu, 2011)
Điểm KN Giống KN
Tổng số cá thể/lần ĐT
Tổng số loài/lần ĐT
Chỉ số ưu thế Simson’D
Chỉ số đa dạng Shannon H’
Vĩnh Phúc
G1 464,85±36,26 29,80±0,76 0,18±0,03 4,85a±0,34 G2 467,05±24,68 30,10±0,69 0,15±0,02 4,81a±0,28 G3 609,30±53,44 30,00±2,32 0,14±0,04 4,55a±0,34
Nghệ An
G1 406,98±30,16 30,40±0,98 0,15±0,01 4,96a±0,14 G2 393,96±19,47 29,41±3,46 0,17±0,01 5.13a±0,11 G3 469,50±11,67 32,47±7,35 0,16±0,01 5.01a±0,13
Đắk Lắk
G1 206.68±20,10 20,20±0,96 0,10±0,02 4,16a±0,11 G2 213.17±19,07 19,41±2,66 0,13±0,01 4.13a±0,13 G3 231.21±10,27 22,37±5,35 0,11±0,03 4.01a±0,15
Đồng Nai
G1 G2 G3
296.58±20,11 313.17±19,17 331.26±10,28
19,22±0,91 20,43±1,67 21,39±2,35
0,11±0,03 0,13±0,02 0,12±0,04
4,76a±0,13 4.51a±0,11 4.61a±0,14
5.2.3.6 Kết quả đánh giá đa dạng sinh học ở cấp độ loài của một số nhóm sâu hại
phổ biến và loài bọ rùa bắt mồi trên các giống ngô KNDR.
Kết quả đánh giá đa dạng ở cấp độ loài của nhóm sâu hại và nhóm bọ rùa bắt mồi được thể hiện tại Bảng 5-41 và 5-42. Trong KNDR tại Vĩnh Phúc, trên ruộng ngô thí nghiệm đã ghi nhận được 29 loài sâu hại ngô, gồm 4 loài sâu hại chủ đích và 2 5 loài sâu hại không chủ đích . Tổng số cá thể của nhóm các loài sâu hại ngô bắt gặp trong một kỳ điều tra trên những giống ngô khảo nghiệm không giống nhau. Chỉ tiêu này đạt cao nhất ở giống ngô nền không biến đổi gen (30Y87), trung bình 233.65 cá thể/kỳ điều tra. Các giống ngô biến đổi gen 30Y87H và 30Y87HR có trung bình tổng số cá thể của nhóm các loài sâu hại ngô bắt gặp trong một kỳ điều tra thấp hơn và gần tương tự nhau, tương ứng là 90.20 và 80.60 cá thể/kỳ điều tra (Bảng 5-41). Tại Phong Thịnh (Nghệ An), chỉ tiêu trung bình tổng số cá thể của nhóm các loài sâu hại ngô bắt gặp trong một kỳ điều tra trên những giống ngô khảo nghiệm là tương tự nhau và it khác biệt hơn so với ở Vũ Di (Vĩnh Phúc). Tuy nhiên, chỉ tiêu này trên giống ngô nền không chuyển gen (30Y87) vẫn có xu hướng cao hơn so với các giống ngô chuyển gen. Giá trị cao nhất trong toàn bộ khảo nghiệm và chung cho cả 3 giông khảo nghiệm

151
là 215,75 cá thể/kỳ điều tra. Các giống ngô chuyển gen 30Y87H và 30Y87HR có trung bình tổng số cá thể của nhóm các loài sâu hại ngô bắt gặp trong một kỳ điều tra đạt tương ứng là 163,22 và 184,77 cá thể/kỳ điều tra (Bảng 5-41). Như vậy, chỉ tiêu tổng số cá thể của nhóm các loài sâu hại ngô bắt gặp trong một kỳ điều tra trong KNDR ở Vĩnh Phúc có sự khác biệt có ý nghĩa giữa các giống ngô chuyển gen và giống ngô không chuyển gen. Trong khi đó, khảo nghiệm ở Nghệ An chỉ tiêu này khác biệt ở mức không có ý nghĩa thống kê. Tại Đắk Lắk và Đồng Nai, tổng số cá thể của nhóm sâu hại ngô bắt gặp trong một kỳ điều tra trên cả 3 giống là tương đương nhau. Tuy nhiên, tại Đắk Lắk trị số tổng số cá thể nhóm sâu hại ngô là thấp nhất và thấp hơn đáng kể so với 3 điểm khảo nghiệm còn lại.
Trên các giống ngô khảo nghiệm đã bắt gặp 5 loài bọ rùa hoạt động bắt mồi
trên ruộng ngô, bao gồm Bọ rùa Nhật Bản, Bọ rùa 6 chấm, Bọ rùa hofmani, bọ rùa 8 chấm và bọ rùa 2 mảnh đỏ. Trong số chúng Bọ rùa Nhật Bản có tần suất bắt gặp và mật độ cao hơn cả trên tất cả các giống ngô khảo nghiệm. Trong 4 điểm khảo nghiệm, tại Nghệ An tần suất bắt gặp bọ rùa nhiều hơn so với 3 địa điểm khảo nghiệm khác, lần lượt đạt 42.17, 43.92 và 51.31con trên các giống chuyển gen, chuyển gen tham khảo và đối chứng. Tuy nhiên, nếu chỉ xét tại từng điểm thì giữa các giống khảo nghiệm là không có sự khác biệt ý nghĩa (độ tin cậy 95%). Chỉ số đa dạng H’của nhóm bọ rùa đạt không cao. S ự khác nhau của chỉ số H’ giữa các giống ngô khảo nghiệm không có ý nghĩa thống kê. Số loài bọ rùa bắt gặp/kỳ điều tra tại Vĩnh Phúc, Nghệ An và Đồng Nai tương tự như nhau, riêng ở Đắk Lắk chỉ tiêu này thấp nhất.Như vậy, tuy có sự sai khác về số cá thể, số loài bọ rùa/ lần điều tra nhưng sự sai khác đó ở mức cho phép và không có ý nghĩa thống kê. Kết luận, giống ngô chuyển gen 30Y87H và 30Y87HR không ảnh hưởng đến sự phân bố và tần suất bắt gặp các loài bọ rùa bắt mồi trên ruộng ngô trong KNDR tại 4 địa điểm.

152
Bảng 5-41: Chỉ số đa dạng các loài sâu hại bắt gặp trên các giống ngô KNDR tại 4 địa điểm (vụ Hè –Thu, 2011)
Điểm khảo nghiệm
Giống khảo nghiệm
Tổng số cá thể/lần điều tra
Tổng số loài/lần điều tra
Chỉ số Shannon H’
Vĩnh Phúc
G1 90.20b 6.90±12.60 a 2.67±0.69 a±0.36 G2 80.60b 6.75±14.68 a 2.72±0.17 a±0.39 G3 233.65a 7.15±28.13 a 1.64±0.36 b±0.36
LSD.05 63.06 1.46 0.39 Nghệ An
G1 163.22a 8.23±18.50 ab 2.68±0.19 a±0.08 G2 184.77a 8.00±21.00 b 2.53±0.70 a±0.17 G3 215.75a 9.67±19.55 a 2.63±0.34 a±0.17
LSD.05 67.94 1.60 0.47 Đắk Lắk
G1 50.10a 3.90 ±13.61 a 1.69±0.69 a±0.35 G2 40.70a 3.75 ±14.67 a 1.73±0.17 a±0.33 G3 59.61a 4.15±28.13 a 1.84±0.36 b±0.31
LSD.05 13.01 1.40 0.35 Đồng Nai
G1 G2 G3
LSD.05
90.29a
80.60 ±12.60 a
93.65 14.68
a
23.01 ±28.13
5.95a
6.05±061
a
6.15±0.19
a
1.12 ±0.38
2.07a
2.12±0.31
a
1.94±0.35
a
0.28 ±0.33

153
Bảng 5-42: Chỉ số đa dạng các loài bọ rùa bắt mồi trên các giống ngô KNDR tại 4 địa điểm (vụ Hè –Thu, 2011)
Điểm khảo nghiệm
Giống khảo nghiệm
Tổng số cá thể/lần điều tra
Tổng số loài/lần điều tra
Chỉ số Shannon H’
Vĩnh Phúc
G1 14.00a 2.35±1.87 a 1.19 ±0,13 a±0.16 G2 10.95a 2.80 ±1.35 a 1.54 ±1.06 a±0.04 G3 9.85a 2.25± 0.26 a 1,27± 0.12 a ±0.10
LSD.05 4.39 0.34 0.39
Nghệ An G1 42.17a 2.62±7.24 a 0.46 ±0,14 a±0.28 G2 43.92a 3.07 ±3.17 a 0.56 ±0.24 a±0.33 G3 51.30a 2.65± 7.93 a 0.52± 0.15 a ±0.29
LSD.05 20.68 0.58 0.15
Đắk Lắk G1 7.05a 1.75± 1.89 a 1.01 ±0,14 a±0.15 G2 6.45a 1.40 ±1.75 a 1.14 ±0.82 a±0.07 G3 5.75a 1.45± 0.96 a 1,17± 0.22 a ±0.10
LSD.05 3.30 0.54 0.67
Đồng Nai G1 G2 G3
LSD.05
12.19a
13.82±5.14
a
11.70 ±3.94
a
9.88 ± 5.91
2.02a
2.17 ±0,12
a
2.45 ±0.21
a
0.61 ± 0.19
0.87a
0.85±0.21
a
0.71±0.35
a
0.15 ±0.25
5.2.3.7 Thành phần loài và phân bố của Collembola trong các lô thí nghiệm trồng
ngô KNDR
Nói đến chất lượng môi trường đất (sức khỏe của đất) người ta thường quan tâm đến các thành phần sinh học trong đất vì có liên quan trực tiếp đến số lượng và mức độ đa dạng của các động vật không xương sống trong đất. Các động vật không xương sống này được coi như những thành phần sống của đất và là những sinh vật chỉ thị cho chất lượng đất (Blair và cs., 1996). Trong số các động vật không xương sống ở đất, Collembola được chú ý đến như một nhóm sinh vật chỉ thị vì vai trò to lớn của chúng trong các chu trình dinh dưỡng và phân hủy, đóng vai trò quan trọng trong mạng lưới thức ăn trên cạn (Frampton, 1997, Moore và cs., 1988). Phần lớn các động vật chân khớp bé, trong đó có Collembola thường xuất hiện ở lớp đất mặt 0-10cm và chúng chiếm 90-95% về số lượng trong các mẫu thu động vật đất.

154
Kết quả phân tích thành phần loài và phân bố của Collembola trên ruộng ngô trong KNDR tại 4 địa điểm được trình bày ở Bảng 5-43.
Tại Vĩnh Phúc: đất trồng 3 giống ngô khảo nghiệm (30Y87H; 30Y87HR và 30Y87) đã ghi nhận được 51 loài Collembola thuộc 30 giống, 13 họ. Trong số đó, 41 loài đã xác định tên khoa học, 10 loài còn ở dạng sp. (chỉ mới xác định đến tên giống). Số loài tập trung nhiều nhất ở 2 họ, họ Entomobryidae (20 loài, chiếm 39,22% tổng số loài) và họ Isotomidae (10 loài, chiếm 19,23% tổng số loài). Đây cũng là hai họ thường bắt gặp với số lượng loài lớn trong các sinh cảnh (rừng, vườn quanh nhà, trảng cỏ, cây bụi) thu được ở Việt Nam. Trong số11 họ còn lại chỉ có từ 1 đến 5 loài. Trong số các họ Collembola ghi nhận được, g iống Lepidocyrtus có số loài nhiều nhất (12 loài, chiếm 60 % số loài của họ và chiếm 23,53% tổng số loài). 29 giống còn lại có từ 1-3 loài/giống.
Phần lớn các loài Collembola ghi nhận được tại Vĩnh Phúc ( Bảng 5-43) bao gồm hỗn hợp các loài (1) sống chủ yếu ở đất (Tullbergia sp.1; một số loài thuộc họ Isotomidae (Folsomides exiguus, Folsomina onychiurina, Isotomiella minor, Isotomodes pseudoproductus), Megalothorax minimus); (2) nhóm sống ở dưới lớp thảm - trên mặt đất : Xenylla humicola (Hypogastruridae), đa số l oài thuộc họ Entomobryidae (Ẹntomobrya lanuginosa, Entomobrya muscorum,...), họ Bourletiellidae (Deuterosminthurus pallipes, Deuterosminthurus sp.1,...), họ Sminthuridae (Sphyrotheca macrochaeta),vv…; (3) nhóm sống ở bên trên lớp thảm (các loài thuộc giống Lepidocyrtus: Lepidocyrtus (Acr.) heterolepis, L. (Acr.) transiens, L. (Asc.) concolourus, L. (Asc.) sp., vv; (4) loài thuộc giống Seira (Seira camgiangensis); Sinella coeca; Rambutsinella honchongensis (Entomobryidae); Cyphoderus javanus (Cyphoderidae); loài thuộc giống Salina (Salina sp.14
Trong tổng số 51 loài Collembola, có tới 28 loài (chiếm 53,85% tổng số loài) có mặt ở các lô thí nghiệm trồng 3 giống ngô trong KNDR. Có tới 39 loài (chiếm 75,00% tổng số loài) có mặt trong lô trồng của 2/3 giống ngô: hoặc ngô đối chứng và ngô chuyển gen, hoặc ngô đối chứng và ngô chuyển gen tham khảo, hoặc ngô chuyển gen và ngô chuyển gen tham khảo. Chỉ có 12 loài có mặt tro ng các lô thí nghiệm trồng một giống ngô nhất định, cụ thể: chỉ có mặt ở lô trồng ngô đối chứng - Folsomides americanus (1 cá thể), Lepidocyrtus (Acr.) heterolepis (2 cá thể); chỉ có mặt ở lô trồng ngô chuyển gen, gồm 4 loài: Isotomurus f. prasinus (1 cá thể), Rambutsinella honchongensis (1 cá thể), Lepidocyrtus (Asc.) sp.
);
1 (1 cá thể), Salina sp.14 (3 cá thể). Chỉ có mặt ở lô trồng ngô chuyển gen tham khảo, gồm 6 loài: Mesaphorura krausbaueri (1 cá thể), Pseudachorutella assigilata (1 cá thể), Entomobrya sp.2 (7 cá thể), Pseudosinella octopunctata (1 cá thể), Lepidocyrtus

155
(Acr.) segamanus (1 cá thể), Lepidocyrtus (Asc.) cf. aseanus (1 cá thể). 12 loài nêu trên chỉ là các loài gặp ngẫu nhiên hoặc ở nền đất này, hoặc ở nền đất kia vì số lượng của chúng rất ít; hơn nữa chúng có thể gặp khá phổ biến trong các dạng sinh cảnh khác nhau (trên đất nông nghiệp: đất trồng màu, vườn quanh nhà,...) ở những địa phương khác nhau của Việt Nam (ở Hà Trung (Thanh Hoá), Gia Lâm (Hà Nội), Yên Thuỷ, Cao Phong (Hoà Bình), Đông Mai (Hưng Yên), Hiệp Hoà (Bắc Giang), VQG Xuân Sơn, Phú Thọ; khu BTTN Thượng Tiến, Kim Bôi, Hòa Bình; khu vực phía Tây các tỉnh từ Quảng Trị đến Quảng Nam,… (Nguyễn Thị Thu Anh và cs., 2000, 2008, 2009; Phạm Đức Tiến và cs., 2007; Nguyễn Trí Tiến và cs., 2007, 2009). Như vậy, cả 12 loài nêu trên không phải là những loài hi ếm, hay đặc biệt về mặt sinh thái mà chúng chỉ là những loài ngẫu nhiên thu được với số lượng rất ít (từ 1 đến 7 cá thể) ở khu vực KNDR ngô chuyển gen. Điều này cho thấy thành phần loài và phân bố của Collembola ở đất trồng các giống ngô đối chứng, chuyển gen và chuyển gen tham khảo (stack) ở Trại sản xuất giống cây trồng Vũ Di, Vĩnh Phúc không có khác biệt.
Tại Nghệ An: Ở địa điểm khảo nghiệm này do thời tiết mưa nhiều vào thời gian thu của đợt cuối (đợt thu mẫu thứ 3), ¼ diện tích các giống bị ngập và hỏng, nên số lượng mẫu thu của đợt thu mẫu thứ 3 là: 5 mẫu đất x 3 công thức x 3 điểm thu= 45 mẫu. Như vậy, tổng số mẫu thu được trong 3 đợt thực địa ở điểm khảo nghiệm diện rộng là 165 mẫu đất định lượng. Kết quả đã ghi nhận được 45 loài Collembola thuộc 26 giống, 10 họ phân bố trong khu vực trồng khảo nghiệm diện rộng ngô chuyển gen ở HTX Phong Thịnh, Thanh Chương, Nghệ An (Bảng 5-43). Trong số đó, 33 loài đã xác định tên khoa học, 12 loài còn ở dạng sp.. Số loài tập trung chủ yếu ở 2 họ, trong đó gặp nhiều loài nhất ở họ Entomobryidae (21 loài, chiếm 46,67% tổng số loài), tiếp đến là họ Isotomidae (7 loài, chiếm 15,56% tổng số loài). 8 họ còn lại chỉ có từ 1 đến 4 loài. Giống Lepidocyrtus có số loài nhiều nhất (10 loài, chiếm 47,62% số loài của họ và chiếm 22,22% tổng số loài). 25 giống còn lại có từ 1-4 loài.
Khu hệ Collembola ở đây bao gồm hỗn hợp (1) các loài sống chủ yếu ở đất (các loài thuộc họ Isotomidae (Folsomides exiguus, Folsomina onychiurina, Isotomodes pseudoproductus,..), Megalothorax minimus; (2) các loài sống ở thảm - đất (đại diện của Neanuridae (Brachystomella parvula); đa số loài của họ Entomobryidae (Entomobrya lanuginosa, Sinella coeca,..) và (3) các loài sống bên trên lớp thảm (Xenylla humicola, Cỵphoderus javanus, Homidia glasa, Willow sia sp.1,...), một số loài Symphypleonid: Sminthurides bothrium, Deuterosminthurus pallipes, Deuterosminthurus sp.1, Deuterosminthurus sp.2, vv.

156
Trong số 45 loài Collembola, có tới 23 loài (chiếm 51,11% tổng số loài) có mặt ở các lô thí nghiệm trồng cả 3 giống ngô, trong đó có 4 loài bắt gặp phổ biến, bao gồm: Deuterosminthurus sp.1, Cyphoderus javanus, Sminthurides aquaticus, Deuterosminthurus pallipes. Có 34 loài (chiếm 75,56% tổng số loài) có mặt trong đất trồng 2/3 giống ngô: hoặc ngô đối chứng và ngô chuyển gen, hoặc ngô đối chứng và ngô chuyển gen đối chứng tham khảo, hoặc ngô chuyển gen và ngô chuyển gen đối chứng tham khảo. Chỉ có 11 loài là có mặt trong các lô thí nghiệm trồng một giống ngô nhất định, cụ thể: chỉ có mặt ở lô trồng ngô đối chứng - Mesaphorura krausbaueri (2 cá thể), Isotomiella minor (1 cá thể), Isotomodes pseudoproductus (1 cá thể), Rambutsinella honchongensis (1 cá thể) , Lepidocyrtus (Asc) aseanus (1 cá thể), Sinella coeca (1 cá thể); chỉ có mặt ở lô trồng ngô chuyển gen, gồm 2 loà i: Xenylla humicola (6 cá thể), Tullbergia sp.1
Tại Đắk Lắk, ghi nhận được 31 loài Collembola thuộc 21 giống, 10 họ phân bố trong khu vực KNDR ngô chuyển gen (Bảng 5-43). Trong số đó, 26 loài đã xác định tên khoa học, 5 loài chỉ mới xác định đến giống . Số loài tập trung chủ yếu ở 3 họ, trong đó họ Entomobryidae có số loài nhiều nhất ( 11 loài, chiếm 35,48% tổn g số loài), tiếp đến là họ Isotomidae (6 loài, chiếm 19,35% tổng số loài) và họ Sminthurididae (5 loài, chiếm 16,13% tổng số loài). 7 họ còn lại chỉ có từ 1 đến 2 loài, chiếm tỷ lệ rất nhỏ trong tổng số loài thu được. Có 3 giống có số loài nhiều nhất
(2 cá thể); chỉ có mặt ở lô trồng ngô chuyển gen đối chứng tham khảo (stack), gồm 3 loài: Brachystomella parvula (1 cá thể), Pseudosinella immaculata (1 cá thể), Lepidocyrtus (L.) cyaneus (2 cá thể). Trong số 11 loài chỉ có mặt trong các lô trồng thí nghiệm một giống ngô nhất định, Rambutsinella honchongensis là loài ít gặp ở đất sản xuất nông nghiệp, hơn nữa số lượng cá thể thu được của loài này rất ít (chỉ duy nhất 1 cá thể). V ì vậy đây chỉ là trường hợp ngẫu nhiên bắt gặp, còn các loài còn lại là những loài khá phổ biến trong các sinh cảnh đất nông nghiệp, trảng cỏ cây bụi, vườn quanh nhà,... (Nguyễn Thị Thu Anh và cs., 2000; 2008; 2009; 2010; Nguyễn Trí Tiến và cs., 2008; 2009). Mặt khác, khi so sánh kết quả về thành phần loài thu được ở các lô trồng ngô khảo nghiệm cùng giống (30Y87H, 30Y87) trồng khảo nghiệm hạn chế tại Văn Giang (Hưng Yên) và trồng khảo nghiệm diện rộng tại các địa phương khác như Vĩnh Phúc, Buôn Mê Thuột, Đồng Nai thì những loài này vẫn có mặt. Như vậy, cả 11 loài này đều xuất hiện hoặc ở đất trồng ngô chuyển gen hoặc ở đất trồng ngô không chuyển gen và đất trồng ngô chuyển gen tham khảo (stack). Nếu chúng vắng mặt hay có mặt ở nền đất trồng ngô đối chứng không chuyển gen hay đất trồng ngô chuyển gen và đất trồng ngô chuyển gen đối chứng tham khảo (stack) chỉ có thể là ngẫu nhiên, hoặc do số lượng mẫu hoặc thời gian nghiên cứu còn ít chứ không phải do yếu tố giống cây trồng gây nên.

157
là giống Homidia, giống Lepidocyrtus và giống Sminthurides (cùng có 3 loài, chiếm 9,68% tổng số loài). 18 giống còn lại có từ 1-2 loài.
Phần lớn các loài Collembola trong danh sách (Bảng 5-43) thuộc hỗn hợp các nhóm sống ở: (1) trong đất như một số loài thuộc họ Isotomidae (Folsomina onychiurina, Isotomiella minor; loài Tullbergia sp.1 (Onychiuridae); (2) trong lớp thảm và ở dưới lớp thảm - trên mặt đất , như một số loài thuộc giống Lepidocyrtus: Lepidocyrtus (Acr.) lanuginosus, L. (Asc.) concolourus, L. (Asc.) sp.4, Homidia glasa, Homidia subcingula, Entomobrya lanuginosa…(Entomobryidae); Cyphoderus javanus (Cyphoderidae); (3) loài thuộc giống Salina: Salina celebensis; (4) một số loài Symphypleonid: Sminthurides bothrium, Deuterosminthurus sp.1, Deuterosminthurus sp.2
Trong số 31 loài Collembola, có tới 16 loài ( 51,61% tổng số loài) có mặt ở các lô thí nghiệm trồng 3 nhóm ngô (ngô đối chứng, ngô chuyển gen gen và ngô chuyển gen tham khảo ). Có tới 20 loài (chiếm 64,52% tổng số loài) có mặt trong it nhất 2 nhóm ngô (hoặc ngô đối chứng và ngô chuyển gen tham khảo hoặc ngô đối chứng và ngô chuyển gen, hoặc ngô chuyển gen và ngô chuyển gen tham khảo). Có 11 loài có mặt trong các lô thí nghiệm trồng một nhóm ngô nhất định, cụ thể: chỉ có mặt ở lô trồng ngô đối chứng - Protaphorura sp.
,….
1 (1 cá thể), Lepidocyrtus (L.) lanuginosus (1 cá thể), Salina celebensis (1 cá thể) , Deuterosminthurus sp.1 (2 cá thể); Chỉ có mặt ở lô trồng ngô chuyển gen, gồm 3 loài: Pseudosinella immaculata (1 cá thể), Lepidocyrtus (Asc.) sp.4
Tất cả 11 loài Collembola (họ Entomobryidae) nêu trên đều là những loài gặp khá phổ biến trong các dạng sinh cảnh khác nhau, từ sinh cảnh rừng tự nhiên (VQG Xuân Sơn) đến sinh cảnh đất trồng màu (Bắc Giang, Hải Dương) và ở những địa phương khác nhau của Việt Nam ( VQG Xuân Sơn, Phú Thọ; khu BTTN Thượng Tiến, Kim Bôi, Hòa Bình; đất canh tác Gia Lâm, Hà Nội; khu vực phía Tây các tỉnh từ Quảng Trị đến Quảng Nam, Hưng Yên vv… (Nguyễn Thị Thu Anh và cs., 2000; 2008; 2009; 2010 Nguyễn Trí Tiến và cs., 2008; 2009). Như vậy, 11 loài này đều không phải là những loài hiếm, hay đặc biệt về mặt sinh thái mà chúng chỉ là những loài ngẫu nhiên thu được với số lượng rất ít (từ 1 đến 5 cá thể) trong khu vực trồng KNDR ngô chuyển gen. Điều này cho thấy thành phần loài Collembola ở các lô thí nghiệm trồng các giống ngô đối chứng, ngô chuyển gen và ngô chuyển gen tham khảo không có sự khác biệt nhau.
(1 cá thể), Calvatomina antena (1 cá thể); Chỉ có mặt ở lô trồng ngô chuyển gen tham khảo, gồm 4 loài: Isotomurus f. prasinus (1 cá thể), Isotomurus punctiferus (1 cá thể), Entomobrya lanuginosa (1 cá thể) và Seira camgiangensis (1 cá thể).

158
Tại Đồng Nai, ghi nhận được 43 loài Collembola thuộc 24 giống, 11 họ phân bố trong khu vực trồ ng ngô khảo nghiệm ở Trung tâm Nghiên cứu T hực nghiệm Nông nghiệp, Hưng Thịnh, Trảng Bom, Đồng Nai (Bảng 5-43). Trong số đó, 33 loài đã xác định tên khoa học, 10 loài còn ở dạng sp. (chỉ mới xác định đến giống). Tương tự như các địa điểm khảo nghiệm khac, số loài tập trung nhiều nhất ở họ Entomobryidae (17 loài, chiếm 39,53% tổng số loài), tiếp đến là họ Isotomidae (8 loài, chiếm 18,60% tổng số loài). 8 họ còn lại chỉ có từ 1 đến 5 loài. Giống Lepidocyrtus có số loài nhiều nhất (10 loài, chiếm 58,82% số loài của họ Entomobryidae và chiếm 23,26% tổng số loài); 23 giống còn lại có từ 1-4 loài.
Xét về đặc điểm sinh thái của các loài Collembola thu được trong khu KNDR cho thấy (Bảng 5-43) các loài Collembola bao gồm hỗn hợp các loài sống chủ yếu ở (1) đất: Tullbergia sp.1 (Onychiuridae), Acherontiella sabina (Hypogastruridae), một số loài của họ Isotomidae (Folsomides exiguus, Folsomina onychiurina, Isotomiella minor), Megalothorax minimus; (2) Các loài sống ở dưới lớp thảm - trên mặt đất như: Xenylla humicola (Hypogastruridae), đa số loài của họ Entomobryidae (Ẹntomobrya lanuginosa, Entomobrya muscorum,...), họ Bourletiellidae (Deuterosminthurus pallipes, Deuterosminthurus sp.1,...); (3) Các loài sống ở bên trên lớp thảm (các loài thuộc giống Lepidocyrtus: Lepidocyrtus (Acr.) heterolepis, L. (Acr.) transiens, L. (Asc.) concolourus, L. (Asc.) sp.4
Trong tổng số 43 loài Collembola thu thập được, có tới 21 loài (48,84% tổng số loài) có mặt ở các lô thí nghiệm trồng 3 nhóm ngô (ngô đối chứng, ngô chuyển gen và ngô chuyển gen tham khảo). 31 loài (72,09% tổng số loài) có mặt trong 2 đến 3 nhóm ngô (hoặc ngô đối chứng và ngô chuyển gen, hoặc ngô đối chứng và ngô chuyển gen tham khảo, hoặc ngô chuyển gen và ngô chuyển gen tham khảo). Chỉ có 12 loài là có mặt trong các lô thí nghiệm trồng một giống ngô nhất định, cụ thể: chỉ có mặt ở lô trồng ngô đối chứng – Protaphorura yodai (2 cá thể), Lepidocyrtus (L.) sp.
,…; (4) loài thuộc giống Seira ( Seira camgiangensis), Sinella coeca, Homidia glassa, (Entomobryidae); Cyphoderus javanus (Cyphoderidae);
5 (1 cá thể), Megalothorax minimus (1 cá thể), Arrhopalites sp.1 (1 cá thể); chỉ có mặt ở lô trồng ngô chuyển gen, gồm 5 loài: Pseudosinella assigilata (1 cá thể), Homidia sauteri f. sinensis (2 cá thể), Sminthurides parvulus (2 cá thể), Deuterosminthurus sp.2 (6 cá thể), Deuterosminthurus sp.3 (1 cá thể). Chỉ có mặt ở lô trồng ngô chuyển gen tham khảo, gồm 3 loài: Acherontiella sabina (1 cá thể), Isotomodes pseudoproductus (1 cá thể), Lepidocyrtus (L.) cyaneus (2 cá thể). Trong số 12 loài này, Megalothorax minimus, Homidia sauteri f. sinensis là những loài gặp khá phổ biến ở đất nông nghiệp (đất trồng màu, vườn quanh nhà,…). 10 loài còn lại chỉ là các loài gặp ngẫu nhiên hoặc ở nền đất này, hoặc ở nền đất k ia vì số lượng của chúng rất ít. H ơn nữa

159
chúng có thể gặp khá phổ biến trong các dạng sinh cảnh khác nhau, ở những địa phương khác nhau của Việt Nam (ở Hà Trung (Thanh Hoá), Gia Lâm (Hà Nội), Yên Thuỷ, Cao Phong (Hoà Bình), Đông Mai (Hưng Yên), Hiệp Hoà (Bắc Giang), VQG Xuân Sơn, Phú Thọ; khu BTTN Thượng Tiến, Kim Bôi, Hòa Bình; khu vực phía Tây các tỉnh từ Quảng Trị đến Quảng Nam,… (Nguyễn Thị Thu Anh và cs., 2000, 2008, 2009; Phạm Đức Tiến và cs., 2007; Nguyễn Trí Tiến và cs., 2007, 2009). Như vậy, 12 loài nêu trên không phải là những loài hiếm, hay đặc biệt về mặt sinh thái mà chúng chỉ là những loài ngẫu nhiên thu được với số lượng rất ít (từ 1 đến 6 cá thể) ở khu vực trồng khảo nghiệm diện rộng ngô chuyển gen. Điều này cho thấy thành phần loài và phân bố của Collembola ở đất trồng các giống ngô đối chứng, chuyển gen và chuyển gen tham khảo (stack) ở Trung tâm N ghiên cứu Thực nghiệm Nông nghiệ p Hưng Lộc, Đồng Nai không có sự khác biệt.

160
Bảng 5-43: Thành phần loài và phân bố của Collembola trong đất trồng ngô KNDR tại 4 địa điểm
(vụ Hè –Thu, 2011)
TT Thành phần loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
I Hypogastruridae
1 Xenylla humicola (Fabricius, 1780) + + + + 0 0 0 0 0 + + +
2 Acherontiella sabina Bonet, 1945 0 0 0 + 0 + 0 0 0 0 + 0
II Onychiuridae 1 Protaphorura yodai (Yosii, 1966) 0 0 0 0 0 0 0 0 0 0 0 + 2 Protaphorura sp. 0 1 0 0 + + + 0 0 + 0 0 0 3 Mesaphorura krausbaueri Borner, 1901 0 0 0 0 0 + 0 0 0 0 0 0 4 Protaphorura tamdaona Nguyen, 1995 0 + + 0 0 0 0 0 0 0 0 0 5 Mesaphorura krausbaueri Borner, 1901 0 + 0 0 0 0 0 0 0 0 0 0 6 Tullbergia sp. + 1 0 + + 0 0 0 + + + + +
III Neanuridae 1 Brachystomella parvula (Schaffer, 1896) + + 0 0 + 0 0 0 0 0 0 0 2 Pseudachorutella assigilata (Borner, 1901) 0 + 0 0 0 0 0 0 0 + 0 0
IV Isotomidae 1 Proisotoma tenella (Reuter, 1895) 0 0 0 0 0 0 0 0 0 + + + 2 Proisotoma submuscicola Nguyen, 1995 + + + 0 + + + + + 0 0 0 3 Cryptopygus thermophilus (Axelson, 1900) + + + + + + + + +

161
TT Thành phần loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
0 0 0 4 Folsomides americanus Denis, 1931 0 0 + 0 0 0 0 0 0 0 0 0 5 Folsomides exiguus Folsom, 1932 + + + + + 0 0 0 0 0 + + 6 Folsomina onychiurina Denis, 1931 + + + + + + + + + + + + 7 Isotomiella minor (Schaffer, 1896) + + + 0 0 + 0 + + + + 0 8 Isotomodes pseudoproductus Stach, 1965 0 + + 0 0 + 0 0 0 0 + 0 9 Isotomurus palustris (Muller, 1776) + + + + + + 0 0 0 + + + 10 Isotomurus f. prasinus (Reuter, 1891) + 0 0 0 0 0 0 + 0 0 0 0 11 Isotomurus punctiferus Yosii, 1963 + + + + + + 0 + 0 + + +
V Entomobryidae 1 Entomobrya muscorum (Nicolet, 1841) + + + + + + 0 0 0 0 + + 2 Entomobrya lanuginosa (Nicolet, 1841) + + + + + + 0 + 0 + + + 3 Entomobrya sp. 0 2 + 0 + + + 0 0 0 + + 0 4 Homidia glassa Nguyen, 2001 + + + + + + + + 0 + + + 5 Pseudosinella octopunctata Borner, 1901 0 + 0 0 0 0 0 0 0 0 0 0 6 Homidia sauteri f. sinensis Denis, 1929 0 0 0 + + 0 + + + + 0 0 7 Homidia subcingula Denis, 1948 0 0 0 0 0 0 + + + 0 0 0 8 Pseudosinella immaculata (Lie-Petterson) 0 0 0 0 + 0 + 0 0 0 0 0 9 Rambutsinella honchongensis (Deharveng et Bedos) + 0 0 0 0 + 0 0 0 0 0 0 10 Willowsia sp. 0 1 0 0 + + + 0 0 0 0 0 0 11 Lepidocyrtus (L.) cyaneus Tullberg, 1871 + + + 0 + 0 0 0 0 0 + 0 12 Lepidocyrtus (L.) lanuginosus (Gmelin, 1788) + + + + + 0 0 0 + + + 0

162
TT Thành phần loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
13 Lepidocyrtus (L.) sp. 0 5 0 0 0 0 0 0 0 0 0 0 + 14 Lepidocyrtus (Cyn.) sp. + 1 + + + 0 + 0 0 0 0 + + 15 Lepidocyrtus (Acr.) heterolepis Yosii, 1959 0 0 + 0 0 0 0 0 0 0 + + 16 Lepidocyrtus (Acr.) segamanus Yosii, 1982 0 + 0 0 0 0 0 0 0 0 0 0 17 Lepidocyrtus (Acr.) transiens Yosii, 1982 0 + + + + + 0 0 0 + + +
18 Lepidocyrtus (Asc.) cf. aseanus Yosii, 1982 0 + 0 0
0
+
0 0 0 0 0 0
19 Lepidocyrtus (Asc.) concolourus Nguyen, 2001 + + + + + + + 0 + + + + 20 Lepidocyrtus (Asc.) dahlii Schaffer, 1898 + + + + + 0 0 0 0 0 + + 21 Lepidocyrtus (Asc.) sp. + 1 0 0 + + + 0 0 0 0 0 0 22 Lepidocyrtus (Asc.) sp. + 2 + + + 0 + 0 0 0 + + + 23 Lepidocyrtus (Asc.) sp. + 4 + + + + 0 + 0 0 + + + 24 Seira camgiangensis Nguyen, 2001 + + + 0 0 0 0 + 0 + 0 + 25 Sinella coeca (Schott, 1896) + 0 + 0 0 + + + + + + 0 26 Sinella pseudomonoculata Nguyen, 1995 0 0 0 + + + + + + 0 0 0 27 Seira oligomacrochaeta Nguyen, 2001 + + +
VI Cyphoderidae 1 Cyphoderus javanus Borner, 1906 + + + + + + + + + + + +
VII Paronellidae 1 Salina sp. + 14 0 0 0 0 0 0 0 0 0 0 0 2 Salina celebensis (Schaffer, 1898) 0 0 0 0 0 0 0 0 + 0 0 0

163
TT Thành phần loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
VIII Neelidae 1 Megalothorax minimus Willem, 1900 0 + + 0 + + 0 0 0 0 0 +
IX Sminthurididae 1 Sminthurides aquaticus (Bourlet, 1842) + + + + + + + + + + + + 2 Sminthurides bothrium Nguyen, 2001 + + + + + + + + + + + + 3 Sminthurides parvulus (Krausbauer, 1898) + 0 + + + + + + + + 0 0 4 Sphaeridia pumilis (Krausbauer, 1898) + + + 0 0 0 + + + + + + 5 Sphaeridia zaheri Yosii, 1966 + + + 0 0 0 + + + + + +
X Arrhopalitidae 1 Arrhopalites sp. + 1 + + 0 0 0 0 0 0 0 0 + 2 Collophora mysticiosa Yosii, 1966 + + 0 0 0 0 + + + 0 0 0
XI Katiannidae 1 Sminthurinus victorius Nguyen, 2001 + + 0 0 0 0 + + + + + +
XII Bourletiellidae 1 Deuterosminthurus pallipes (Bourlet, 1843) + + + + + + 0 0 0 + + + 2 Deuterosminthurus sp. + 1 + + + + + 0 + 0 + + + 3 Deuterosminthurus sp. + 2 + + + + + + + + + 0 0 4 Deuterosminthurus sp. 0 3 0 0 + + + 0 0 0 + 0 0
XIII Sminthuridae 1 Nesminthurus sp. 0 1 0 0 + 0 + 0 0 0 0 0 0

164
TT Thành phần loài Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
2 Sphyrotheca macrochaeta Nguyen, 1995 + + 0 + + + 0 0 0 0 0 0
XIV Dicyrtomidae 1 Calvatomina antena (Nguyen, 1995) 0 0 0 0 0 0 + 0 0 0 0 0
Tổng số loài 39 42 37 34 33 35 21 24 22 31 33 31

165
5.2.3.8 Một số chỉ tiêu định lượng Collembola ở đất trồng ngô KNDR tại 4 địa điểm
Kết quả phân tích một số chỉ số định lượng Collembola như: số lượng loài, số lượng cá thể (Bảng 5-44); mật độ trung bình (con/m2
), các chỉ số đa dạng Shannon - Weiner (H’), chỉ số đồng đều Pielou (J’) (Bảng 5-45) cho thấy:
a) Số lượng loài
Tại Vĩnh Phúc tính chung sau 3 đợt thu mẫu định lượng Collembola, số lượng loài dao động từ 37 loài ( ngô đối chứng) đến 39 - 42 loài (ngô chuyển gen và ngô chuyển gen tham khảo (stack)). Tại Nghệ An số lượng loài dao động từ 35 loài (ngô đối chứng), 33-34 loài ở ngô chuyển gen và ngô chuyển gen tham khảo (stack). Tại Đắk Lắk số lượng loài dao động từ 22 loài (ngô đối chứng) và 21-24 loài ở ngô chuyển gen và ngô chuyển gen tham khảo (stack). Và tại Đồng Nai, sau 3 đợt thu mẫu định lượng Collembola, số lượng loài dao động từ 31 loài (ngô đối chứng), 31-33 loài ở ngô chuyển gen và ngô chuyển gen tham khảo (stack). Như vậy, số lượng loài Collembola phân bố trong các lô thí nghiệm trồng các giống ngô chuyển gen mặc dù cao hơn đôi chút so với ngô đối chứng, nhưng là không đáng kể, không có sự khác biệt có ý nghĩa về số lượng loài Collembola trên ruộng trồng ngô chuyển gen và ruộng trồng ngô đối chứng (không chuyển gen) về mặt thống kê.
Phân tích thành phần loài Collembola ở đất trồng KNDR ngô chuyển gen cho
thấy giữa hai nền đất đối chứng (ngô không chuyển gen) và đất thí nghiệm (ngô chuyển gen và stack chuyển gen tham khảo) có chung 28 loài. Chúng chỉ khác nhau ở 12 loài, trong đó nền đất trồng ngô chuyển gen vắng mặt loài Folsomides americanus và Lepidocyrtus (Acr.) heterolepis, nhưng lại có thêm 10 loài khác không có trên đất trồng ngô đối chứng là: Mesaphorura krausbaueri, Pseudachorutella assigilata, Isotomurus prasinus, Entomobrya sp.2, Pseudosinella octopunctata, Rambutsinella honchongensis, Lepidocyrtus (Acr.) segamanus, Lepidocyrtus (Asc.) aseanus, Lepidocyrtus (Asc.) sp.1, Salina sp.14. Xét với từng loài cho thấy: ngoại trừ Folsomides americanus, Lepidocyrtus (Acr.) heterolepis mặc dù không tìm thấy trong đất trồng ngô chuyển gen ở điểm khảo nghiệm Vĩnh Phúc nhưng loài này vẫn có trong đất trồng ngô chuyển gen ở Đồng Nai. 10 loài Collembola vắng mặt ở đất trồng ngô không chuyển gen trong KNDR nhưng lại tìm thấy trên đất trồng giống ngô này ở các địa điểm khác (không thuộc KNDR) như Văn Giang (Hưng Yên), Phong Thịnh (Nghệ An), Hưng Lộc (Đồng Nai). Như vậy có thể khẳng định tất cả các loài Collembola đã đề cập phía trên đều xuất hiện hoặc ở cả đất trồng ngô chuyển gen và đất trồng ngô không chuyển gen dù ở địa phương này hay địa phương khác. Nếu

166
chúng vắng mặt hay tìm thấy ở nền đất trồng ngô chuyển gen hay ngô không chuyển gen ở nơi này hay nơi khac thì chỉ là ngẫu nhiên, hoặc do số lượng mẫu thu còn ít nên chưa gặp.
b) Số cá thể theo từng đợt thu mẫu:Nhìn vào kết quả phân tích chi tiết độ phong
phú của Collembola theo từng đợt thu mẫu Bảng 5-44 cho thấy: Tại Vĩnh Phúc, số lượng cá thể Collembola ở đợt thu thứ nhất cao vượt trội so
với hai đợt thu còn lại. Cụ thể: trên nền đất trồng ngô Đ /C và ngô chuyển gen, số lượng cá thể thu được ở đợt thứ 2 chỉ bằng 1/4 đến 1/8 số lượng cá thể thu ở đợt thu mẫu thứ nhất. Số lượng cá thể thu ở đợt thứ ba bằng 1/3 đến 1/2 số lượng cá thể ở đợt thu mẫu thứ nhất. Điều này có thể giải thích là do khoảng thời gian thu mẫu từ đợt thu mẫu thứ 2 thời tiết mưa nhiều làm cho đất luôn ở trạng thái ướt, đây là nguyên nhân làm giảm một số lượng lớn cá thể các loài Collembola không thích ứng với trạng thái đất ngập nước. Tuy nhiên sự giảm số lượng cá thể Collembola là đồng đều ở các nền đất trồng ngô Đ/C và ngô chuyển gen. Tương tự, tại Nghệ An số lượng cá thể Collembola ở đợt thu thứ nhất cao vượt trội so với hai đợt thu còn lại. Cụ thể đợt 1, số cá thể lần lượt ở các giống chuyển gen, chuyển gen tham khảo và đối chứng là 314, 312 và 308 cá thể; đợt 2, số cá thể lần lượt là 261, 285 và 264; và đợt 3 là 190, 184 và 159 cá thể. Tại Đắk Lắk số lượng cá thể Collembola thu ở một lần thấp nhất là 62 cá thể (ngô 30Y87HR, lần thu thứ nhất) và cao nhất đạt 169 cá thể (ở ngô 30Y87H, lần thu thứ 2 ). Tính chung sau 3 đợt thu mẫu số lượng Collembola thấp nhất ở ngô chuyển gen tham khảo (297 cá thể) và cao nhất ở ngô chuyển gen 30Y87H (357 cá thể). Trong 3 đợt thu mẫu, số lượng Collembola cao nhất thu được là ở đợt thu thứ 2, 451 cá thể, giảm dần ở đợt thu thứ 3 (292 cá thể), thấp nhất ở đợt thu thứ nhất (227 cá thể). Sự sai khác về số lượng cá thể Collembola giữa các đợt thu mẫu thể hiện ngay cùng trong một lô trồng ngô khảo nghiệm hay ngay giữa các giống ngô của cùng một nhóm (chuyển gen, chuyển gen tham khảo hay đối chứng). Tại Đồng Nai số lượng cá thể Collembola ở đợt thu thứ nhất cao hơn đợt thu mẫu thứ 2 nhưng thấp hơn đợt thu mẫu thứ 3. Cụ thể đợt 1, số cá thể lần lượt ở các giống chuyển gen, chuyển gen tham khảo và đối chứng là 450, 212 và 304; đợt 2 số cá thể lần lượt là 205, 181 và 320; đợt 3là 693, 412 và 513 cá thể. So sánh số lượng cá thể Collembola chung cả 3 đợt trên các giống cũng có sự khác nhau. Ở lô đất trồng ngô chuyển gen thu được 1348 cá thể, ngô chuyển gen tham khảo 805 cá thể và ngô đối chứng 1137 cá thể. Như vậy, có sự chênh lệch về số lượng cá thể ở nền đất trồng ngô đối chứng so với nền đất trồng ngô chuyển gen nhưng sự khác biệt này cũng vẫn ở mức không có ý nghĩa trong thống kê. Kết quả này cho thấy độ phong phú loài Collembola ở các lô thí nghiệm trồng ngô trong KNDR có độ dao động khá lớn. Điều này chứng tỏ sự biến động số lượng cá thể

167
ở ngô chuyển gen và ngô chuyển gen tham khảo so với ngô đối chứng không bắt nguồn từ nguyên nhân do ngô chuyển gen gây ra. Nói cách khác protein Cry1F và protein EPSPS không phải là nguyên nhân gây ra sự biến động loài Collembola trong KNDR. Kết luận này cũng hoàn toàn phù hợp với các nghiên cứu của các tác giả Al-Deeb và cs., 2003; Candolfi và cs., 2004; Ahmad và cs., 2005; Bitzer và cs., 2005, khi khẳng định protein Cry1 không ảnh hưởng đến mức độ phong phú của động vật chân khớp bé (Collembola) ở đất.
c) Mật độ trung bình quần thể (con/m2): Tại Vĩnh Phúc, tính chung sau 3 đợt thu
mẫu ở nền đất KNDR ngô chuyển gen TC1507 tại Trại Sản xuất Giống cây trồng Vũ Di (Bảng 5-45): mật độ trung bình của Collembola ở nền đất trồng ngô đối chứng là 10867 cá thể/ m 2; ở nền đất trồng ngô chuyển gen là 10927 cá thể/m2 và ở nền đất trồng ngô chuyển gen tham khảo (stack) là 12787 cá thể/ m2. Kết quả này chứng tỏ mức độ phong phú Collembola ở nền đất trồng ngô trong KNDR có độ dao động khá lớn. Tuy nhiên, sự chênh lệch về số lượng cá thể ở nền đất trồng ngô đối chứng so với nền đất trồng ngô chuyển gen không khác biệt nhiều ( F5,14=0,07, p= 0,95). Tại Nghệ An, mật độ trung bình Collembola ở đất trồng ngô không chuyển gen thấp hơn so với đất trồng ngô chuyển gen và ngô chuyển gen tham khảo. Tuy nhiên, mức độ thấp hơn này không thể hiện lô trồng ngô đối chứng khác biệt với lô thí nghiệm (F3,41=0,06; p= 0,95). Tại Đắk Lắk, mật độ Collembola trung bình (con/m2
) trong đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối chứng lần lượt là 2380, 1980 và 2107. Cuối cùng, tại Đồng Nai, tính chung sau 3 đợt thu mẫu ở các lô trồng KNDR ngô chuyển gen TC1507 tại Trung tâm Nghiên cứu Thực nghiệm Nông nghiệp Hưng Lộc cho thấy số lượng cá thể Collembola đạt giá trị cao nhất ở lô trồng ngô chuyển gen (1348 cá thể), thấp hơn ở lô trồng ngô đối chứng (1137 cá thể) và thấp nhất ở lô trồng ngô chuyển gen tham khảo (805 cá thể). Như vậy, kết quả KNDR tại 4 địa điểm chứng tỏ độ phong phú của Collembola ở các lô thí nghiệm trồng ngô chuyển gen có độ dao động khá lớn. Tuy nhiên, giá trị độ phong phú trên đất trồng ngô chuyển gen (30Y87H,30Y87HR) vẫn chỉ nằm ở khoảng trên và dưới so với giống ngô đối chứng (30Y87). Điều này chứng tỏ sự biến động số lượng cá thể ở ngô chuyển gen và chuyển gen tham khảo so với ngô đối chứng không bắt nguồn từ nguyên nhân do ngô chuyển gen gây ra. Nói cách khác là không phải ảnh hưởng do protein Cry1F gây ra.
d) Các chỉ số đa dạng và chỉ số đồng đều: Kết quả phân tích sự thay đổi giá trị các chỉ số: chỉ số Shannon-Weiner (H’), và chỉ số đồng đều Pielou (J’) trình bày ở Bảng 5-45. Sau 3 đợt thu mẫu, giá trị của chỉ số đa dạng H’ và chỉ số đồng đều (J’) giữa các lô trồng ngô chuyển gen với lô trồng ngô đối chứng và ngô chuyển gen tham

168
khảo (stack) tương đương nhau, không thấy có sự khác biệt (D0,33
+ Với giá trị H’: lô đối chứng có giá trị H’= 2.26, 2.29, 2.27 và 2.28. Lô trồng ngô chuyển gen có giá trị H’= 2.29, 2.30, 2.26 và 2.27. Lô trồng ngô chuyển gen tham khảo (stack): 254, 2.26, 2.30 và 2.30 (lần lượt ở các điểm Vĩnh Phúc, Nghệ An, Đắk Lắk và Đồng Nai.
=0,17, p>0,05). Cụ thể như sau:
+ Giá trị J’ của ngô chuyển gen thấp hơn chút ít so với J’ của ngô đối chứng và ngô chuyển gen tham khảo (stack).
Phân tích chi tiết sự thay đổi các giá trị của chỉ số đa dạng, chỉ số đồng đều trong từng lần thu mẫu (Bảng 5-46). Các chỉ số đa dạng (H’) và chỉ số đồng đều (J’) theo từng đợt thu mẫu:
+ Giá trị H’: Đợt 1: lần lượt ở lô đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối
chứng tại các địa điểm là 1.86, 2.05, 1.89 (Vĩnh Phúc); 2.05, 1.71, và 1.69 (nghệ An), 2.18, 2.05 và 1.84 (Đắk Lắk); 1.41, 1.78 và 1.21 (Đồng Nai).
Đợt 2: lần lượt ở lô đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối chứng tại các địa điểm là 2.85, 2.93, 2.49 (Vĩnh Phúc); 2.05, 1.71, và 1.69 (nghệ An), 2.18, 2.05 và 1.84 (Đắk Lắk); 1.41, 1.78 và 1.21 (Đồng Nai).
Đợt 3: lần lượt ở lô đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối chứng tại các địa điểm là 5.50, 2.74, 2.41 (Vĩnh Phúc); 1.90, 1.93, và 2.34 (nghệ An), 1.96, 2.05 và 2.09 (Đắk Lắk); 2.08, 2.04 và 2.15 (Đồng Nai).
+ Giá trị J’: Đợt 1: lần lượt ở lô đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối
chứng tại các địa điểm là 0.64, 0.60, 0.57 (Vĩnh Phúc); 0.69, 0.54, và 0.57 (nghệ An), 0.88, 0.78 và 0.77 (Đắk Lắk); 0.55, 0.72 và 0.49 (Đồng Nai).
Đợt 2: lần lượt ở lô đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối chứng tại các địa điểm là 0.89, 0.84, 0.97 (Vĩnh Phúc); 0.67, 0.58, và 0.56 (nghệ An), 0.73, 0.81 và 0.74 (Đắk Lắk); 0.59, 0.77 và 0.62 (Đồng Nai).
Đợt 3: lần lượt ở lô đất trồng ngô chuyển gen, chuyển gen tham khảo và ngô đối chứng tại các địa điểm là 0.73, 0.78, 0.74 (Vĩnh Phúc); 0.66, 0.68, và 0.76 (nghệ An), 0.71, 0.72 và 0.79 (Đắk Lắk); 0.67, 0.61 và 0.65 (Đồng Nai).
Sau 3 đợt thu mẫu, giá trị của cả 2 chỉ số, chỉ số đa dạng H’ và chỉ số đồng đều J’, ở các đất trồng ngô chuyển gen có sự tăng giảm nhiều hơn so với đất trồng ngô đối chứng. Cụ thể đất trồng ngô chuyển gen 30Y87H có giá trị chỉ số đa dạng H' thấp hơn chút ít so với đất trồng ngô đối chứng , nhưng giá trị của chỉ số đồng đều J' lại cao hơn. So sánh cũng cho kết quả tương tự theo từng cặp các nhóm ngô (ngô đối chứng

169
với ngô chuyển gen tham khảo và ngô chuyển gen với ngô chuyển gen tham khảo). Tuy nhiên, sự tăng giảm giá trị của các chỉ số này giữa ngô đối chứng với ngô chuyển gen, và với ngô chuyển gen tham khảo là không đáng kể (D0,34=0,22; p>0,05).

170
Bảng 5-44: Số lượng loài, số cá thể Collembola trên các giống ngô KNDR theo từng đợt thu mẫu tại 4 địa điểm
(vụ Hè – Thu, 2011)
Đợt thu mẫu Chỉ số
Địa điểm khảo nghiệm Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai
G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3 Đợt 1 Số loài 18 30 28 20 23 19 12 14 11 13 12 12
Số cá thể 1159 1176 1004 314 312 308 84 62 81 450 212 304 Đợt 2 Số loài 25 32 23 14 15 11 13 15 13 21 16 15
Số cá thể 139 268 173 261 285 264 169 138 144 205 181 320 Đợt 3 Số loài 31 33 26 18 17 22 16 17 14 22 29 28
Số cá thể 341 474 453 190 184 159 104 97 91 693 412 513 Chung 3 đợt Số loài
Số cá thể 39
1639 42
1918 37
1630 34
765 33
781 35
731 21
357 24
297 22
316 31
1348 33
805 31
1137

171
Bảng 5-45: So sánh một số chỉ số định lượng Collembola trong đất trồng ngô KNDR tại 4 địa điểm (vụ Hè – Thu, 2011)
Chỉ số Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
Số loài 39 42 37 34 33 35 21 24 22 31 33 31 Mật độ TB (con/m2 10927 )
± 2.54 12787 ± 2.84
10867 ± 2.24
5564 ± 2.26
5680 ± 2.35
5316 ± 2.6
2380 ± 4.44
1980 ± 3.80
2107 ± 3.38
8987 ± 2.44
5367 ± 1.25
7580 ± 1.16
Chỉ số đa dạng H’ 2.29 2.54 2.26 2.30 2.26 2.29 2.26 2.30 2.27 2.27 2.30 2.28 Chỉ số đồng đều J’ 0.63 0.62 0.68 0,66 0.65 0.64 0.74 0.72 0.73 0.66 0.66 0.66

172
Bảng 5-46: Giá trị chỉ số đa dạng H’ và chỉ số đồng đều J’ theo từng đợt điều tra trên đất trồng ngô KNDR tại 4 địa điểm (vụ Hè – Thu, 2011)
Đợt thu mẫu Chỉ số Địa điểm khảo nghiệm
Vĩnh Phúc Nghệ An Đắk Lắk Đồng Nai G1 G2 G3 G1 G2 G3 G1 G2 G3 G1 G2 G3
Đợt 1 H’ 1.86 2.05 1.89 2.05 1.71 1.69 2.18 2.05 1.84 1.41 1.78 1.21 J’ 0.64 0.60 0.57 0.69 0.54 0.57 0.88 0.78 0.77 0.55 0.72 0.49
Đợt 2 H’ 2.85 2.93 2.49 1.77 1.58 1.34 1.87 2.20 1.90 1.81 2.14 1.69 J’ 0.89 0.84 0.79 0.67 0.58 0.56 0.73 0.81 0.74 0.59 0.77 0.62
Đợt 3 H’ 5.50 2.74 2.41 1.90 1.93 2.34 1.96 2.05 2.09 2.08 2.04 2.15 J’ 0.73 0.78 0.74 0.66 0.68 0.76 0.71 0.72 0.79 0.67 0.61 0.65
Chung 3 đợt H’ 2.29 2.54 2.26 2.30 2.26 2.29 2.26 2.30 2.27 2.27 2.30 2.28 J’ 0.62 0.68 0.63 0.66 0.65 0.64 0.74 0.72 0.73 0.66 0.66 0.66

173
5.2.3.9 Các loài Collembola phổ biến và ưu thế
Loài ưu thế là loài có số lượng cá thể riêng chiếm ≥ 5% tổng cá thể của lô thí nghiệm. Ở mỗi sinh cảnh có một tập hợp loài ưu thế đặc trưng và tập hợp này thay đổi ở các sinh cảnh. Sự thay đổi các loài ưu thế phản ánh sự thay đổi điều kiện môi trường sống. Trong điều kiện sống tối ưu, mang tính chất tự nhiên, thông thường các loài trong quần xã có giá trị số lượng cá thể tương đối đồng đều. Giá trị số lượng loài ưu thế không vượt trội hẳn so với các loài khác. Ngược lại, khi điều kiện môi trường thay đổi, tác động trực tiếp đến từng cá thể, sinh vật phải tự điều chỉnh để thích ứng với điều kiện sống mới dẫn đến kết quả: một vài loài bị diệt vong, một vài loài khác phát triển đột biến làm thay đổi tập hợp ưu thế trong quần xã. Trên cơ sở thay đổi ấy, người ta có thể phán đoán quá trình cũng như chiều hướng diễn thế của sự thay đổi điều kiện môi trường. Loài phổ biến là loài có mặt ở ≥ 50% tổng số mẫu định lượng của lô thí nghiệm (Maria Sterzynska, 1990). Kết quả phân tích các mẫu đất tại 4 địa điểm trồng 3 giống ngô khảo nghiệm, ghi nhận được 8 loài Collenbola phổ biến và ưu thế ( Bảng 5-47) và phân bố của 8 loài đó trên 3 giống ngô trồng ở 4 địa điểm (Bảng 5-48)
Vĩnh Phúc, ghi nhận 5 loài Collembola phổ biến và 6 loài Collembola ưu thế. Trong số 5 loài Collembola phổ biến, có 4 loài Collembola là phổ biến trên cả nền đất trồng ngô đối chứng, chuyển gen và chuyển gen tham khảo. Đó là Isotomurus palustris, Sminthurides aquaticus, Deuterosminthurus pallipes, Cyphoderus javanus, Xenylla humicola, với độ phổ biến từ 50,00% đến 73,33%. Trong thí nghiệm này, Deuterosminthurus sp.1
Nghệ An, ghi nhận được 3 loài Collembola phổ biến và 7 loài Collembola ưu thế. Trong số 3 loài phổ biến có 1 loài phổ biến ở cả lô trồng ngô đối chứng và lô trồng ngô chuyển gen với độ phổ biến từ 45,46% đến 50,91% ( Bảng 5-48). 2 loài Collembola còn lại chỉ phổ biến hoặc ở đất trồng ngô đối chứng ( Sminthurides aquaticus), hoặc ở đất trồng ngô chuyển gen và ngô chuyển gen tham khảo (Cyphoderus javanus). 3 loài Collembola phổ biến trên cùng với Deuterosminthurus pallipes và Deuterosminthurus sp.
chỉ là loài Collembola phổ biến ở nền đất trồng ngô chuyển gen. Cũng trong 5 loài Collembola phổ biến trên, Xenylla humicola đồng thời là loài ưu thế ở cả nền đất trồng ngô đối chứng và ngô chuyển gen, với độ ưu thế từ 5,58% (giống ngô Đ/C) đến 29,08% (cũng ở giống ngô Đ /C). Như vậy, những loài Collembola phổ biến và ưu thế ở lô trồng ngô chuyển gen cũng là các loài Collembola ưu thế và phổ biến ở lô trồng ngô đối chứng, không có sự sai khác giữa hai nền đất nay.
2 là những loài ưu thế ở cả hai nền đất trồng khảo nghiệm ngô với độ ưu thế từ 6,51% ( ngô đối chứng 30Y87) đến 24,93% (cũng ở

174
giống ngô đối chứng 30Y87). Trong 2 loài ưu thế còn lại, Sminthurides bothrium, cùng ưu thế ở đất trồng ngô đối chứng (30Y87) và đất trồng ngô chuyển gen tham khảo (30Y87HR) với độ ưu thế tương ứng 13,99% và 11,4%. Chỉ có loài Isotomurus palustris chỉ ưu thế ở lô trồng ngô chuyển gen (30Y87H) với độ ưu thế 5,08%, tuy nhiên loài này có độ ưu thế không lớn. Như vậy, ngay cả tập hợp các loài Collembola ưu thế trong đất trồng ngô chuyển gen, chuyển gen tham khảo và đối chứng cũng không có sự khác biệt về loài giữa các lô đất khảo nghiệm 3 giống ngô này.
Tại Đắk Lắk, cả khu vực KNDR ngô chuyển gen ở Nông trường Thống Nhất, huyện Krongpak đã ghi nhận được duy nhất 1 loài Collembola phổ biến chung cho các nền đất trồng 3 nhóm ngô khảo nghiệm (30Y87, 30Y87H và 3087HR), với độ phổ biến từ 51,67% (30Y87HR) đến 75,56% (30Y87). Có 7 loài Collembola chiếm ưu thế là Cryptopygus thermophilus, Sminthurides aquaticus, Sminthurinus victorius, Cyphoderus javanus, Sminthurides parvulus, Proisotoma submuscicola, Homidia subcingula. Trong số này, có 4 loài là ưu thế ở cả 3 nền đất trồng ngô chuyển gen, ngô chuyển gen tham khảo và ngô đối chứng và đó là 4 loài được có tên đầu tiên trong số 7 loài nêu ở trên. 3 loài còn lại là Sminthurides parvulus, Proisotoma submuscicola và Homidia subcingula chỉ chiếm ưu thế ở đất trồng ngô chuyển gen và chuyển gen tham khảo. Tuy nhiên, cả 3 loài này với độ ưu thế không lớn, chỉ từ 5,57% đến 12,33%. Như vậy, trên các lô KNDR ngô tại Đắk Lắk không có sự khác biệt về tập hợp loài Collembola phổ biến hay ưu thế ở lô trồng ngô chuyển gen và không chuyển gen.
Đồng Nai, khu vực trồng ngô KNDR tại Trung tâm Nghiên cứu Thực nghiệm Nông nghiệp Hưng Lộc. Ghi nhận duy nhất 1 loài Collembola phổ biến và 7 loài Collembola ưu thế. Loài phổ biến duy nhất ghi nhận là phổ biến ở cả 3 nhóm ngô khảo nghiệm, với độ phổ biến từ 50,0% (30Y87H) đến 51, 67% (30Y87HR và 30Y87). Deuterosminthurus sp.1
là loài ưu thế ở tất cả các lô thí nghiệm trồng ngô khảo nghiệm với độ ưu thế từ 5,28% (30Y87) đến 23,11% (30Y87HR). Duy nhất loài Cyphoderus javanus chỉ ưu thế ở lô trồng ngô chuyển gen tham khảo (30Y87HR) với độ ưu thế không lớn, chỉ 6,96%. Như vậy, những loài Collembola phổ biến và ưu thế ở lô trồng ngô chuyển gen cũng là các loài Collembola ưu thế và phổ biến ở các lô trồng ngô đối chứng và chuyển gen tham khảo.

175
Bảng 5-47: Tên các loài Collembola phổ biến và ưu thế trong đất trồng ngô KNDR tại 4 địa điểm (vụ Hè –Thu, 2011)
Ký hiệu loài Tên loài Loài 1 Isotomurus palustris Loài 2 Sminthurides aquaticus Loài 3 Deuterosminthurus pallipes Loài 4 Deuterosminthurus sp.Loài 5
1
Cyphoderus javanus Loài 6 Xenylla humicola Loài 7 Proisotoma tenella Loài 8 Cryptopygus thermophilus Loài 9 Seira oligomacrochaeta

173
Bảng 5-48: Các loài Collembola phổ biến và ưu thế trong đất trồng ngô KNDR tại 4 địa điểm (vụ Hè –Thu, 2011)
Điểm Giống UT/PB (%) Loài 1 Loài 2 Loài 3 Loài 4 Loài 5 Loài 6 Loài 7 Loài 8 Loài 9
Vĩnh Phúc
G1 UT 19.70 25.20 12.60 11.10 6.10 10.60 PB 53.30 71.70 60.00 63.30 50.00
G2 UT 23.10 19.29 11.11 13.87 PB 55.00 73.33 65.00 50.00 51.67
G3 UT 20.12 29.08 13.07 9.51 5.58 6.69 PB 63.33 70.00 55.00 65.00
Nghệ An
G1 UT 5.08 22.70 19.90 21.10 9.83 8.62 PB 50.91 50.91
G2 UT 22.00 23.30 16.90 7.58 10.80 PB 24.93 47.27 43.64
G3 UT 43.64 16.34 6.51 7.76 PB
Đắk Lắk
G1 UT 18.38 5.57 6.13 6.96 37.05 PB 58.33
G2 UT 18.90 7.40 6.30 12.33 23.01 PB 51.67
G3 UT 21.40 6.39 6.71 32.30 PB 75.56
Đồng Nai
G1 UT 11.28 18.84 16.39 8.23 20.85 8.38 PB 50.00
G2 UT 9.57 16.16 13.91 10.68 6.96 16.15 9.44 PB 51.67

174
Điểm Giống UT/PB (%) Loài 1 Loài 2 Loài 3 Loài 4 Loài 5 Loài 6 Loài 7 Loài 8 Loài 9
G3 UT 11.61 19.17 12.84 5.28 19.96 15.57 PB 51.67

173
Kết quả sau một vụ trồng khảo nghiệm diện rộng các ngô chuyển gen (30Y87H,
30Y87HR) và ngô đối chứng không chuyển gen (30Y87) ở 4 địa điểm khảo nghiệm đại diện 4 vùng trồng ngô chính tại Việt nam (Vĩnh Phúc, Nghệ An, Buôn Ma Thuột, Đồng Nai) nhận thấy:
- Ngô chuyển gen chứ a protein Cry1F không làm ảnh hưởng đến quần thể Collembola trong đất trồng ngô. Cụ thể không thấy có sự sai khác có ý nghĩa về thành phần loài, phân bố, số lượng loài và mức độ phong phú loài Collembola giữa các giống ngô trồng khảo nghiệm (ngô đối chứng; ngô chuyển gen và ngô chuyển gen tham khảo). Kết luận, ngô chuyển gen tuy có chứa protein Cry1F (B.t) nhưng không độc hay không gây ảnh hưởng đến quần thể Collembola, hoặc lượng protein Cry1F không có hoặc có nhưng với lượng rất thấp trong đất trồng khảo nghiệm, không gây chết Collembola.
- Không phát hiện thấy ảnh hưởng nào đến các hoạt động sinh học của Collembola trong đất. Các kết quả nghiên cứu trên cũng phù hợp kết quả từ một số nghiên cứu khác trên thế giới, đó là cây trồng chuyển gen cry1 không ảnh hưởng đến các loài Collembola trong đất trồng cây chuyển gen này. Tác giả Yu và cs. (1997) ghi nhận độc tố của Cryp1Ab và Cryp1Ac không ảnh hưởng đến Folsomina candida Willem (Collembola: Isotomidae); Sims và Martin (1997) khẳng định không có ảnh hưởng độc nào của Cryp1Ab, Cry1Ac, Cry2A và Cry3A đến hai loài Collembola là Folsomia candida Willem và Xenylla grisea Axelson.
- Tính chung cả 3 đợt thu mẫu định lượng Collembola, số lượng cá thể, mật độ trung bình (con/m2
- Tập hợp Collembola ưu thế giữa các lô đất trồng ngô đối chứng, ngô chuyển gen và chuyển gen tham khảo (stack) có nhiều điểm chung và không có nhiều sự sai khác.
), giá trị của các chỉ số đa dạng H’ và chỉ số đồng đều J’ của lô khảo nghiệm ngô chuyển gen, ngô chuyển gen tham khảo và ngô đối chứng không có sự khác biệt.
5.2.4 Kết luận sơ bộ kết quả KNDR
Kết quả thu được từ khảo nghiệm diện rộng ngô chuyển gen event TC1507 tại
4 địa điểm đại diện 4 vùng trồng ngô chính của Việt nam cho thấy:

174
- Kết quả kiểm tra protein Cry1F khẳng định việc bố trí các công thức khảo nghiệm là hoàn toàn chính xác, giống tham gia khảo nghiệm đúng là những giống đã được cấp phép KNDR.
- Kết quả đánh giá các đặc tính nông sinh học của 3 giống ngô KNDR cho thấy chúng có đặc điểm hoàn toàn giống nhau. Tại những điểm có áp lực sâu tự nhiên lớn, các giống chuyển gen cho năng suất cao hơn hẳn so với giống đối chứng không chuyển gen.
- Khả năng mẫm/kháng một số sâu bệnh hại chính của 3 giống ngô trong KNDR là không khác nhau. Tại các điểm trồng khảo nghiệm có áp lực sâu tự nhiên lớn 2 giống ngô chuyển gen đã thể hiện được tính kháng cao đặc biệt là kháng sâu đục thân.
- Số lượng loài chân khớp phát hiện được trên giống ngô nền (30Y87) và 2 giống ngô chuyển gen (30Y87H, 30Y87HR) là không có sự khác biệt.
- Chỉ số ưu thế Simpson’s D trên cả 3 giống khảo nghiệm là tương đương nhau. Chỉ số này trên cả 4 điểm khảo nghiệm đều rất thấp và có sự khác biệt nhưng ở mức không có ý nghĩa thống kê.
- Chỉ số đa dạng ShannonH’ ở các giống ngô khảo nghiệm đều đạt giá trị khá cao và không khác biệt (p < 0.05). Các giá trị ghi nhận được của chỉ số đa dạng H’ và chỉ số ưu thế D trên những giống ngô KNDR hoàn toàn lôgic. Kết quả này chứng tỏ trong sinh quần các giống ngô khảo nghiệm không có loài chân khớp nào phát triển với số lượng nổi trội, chiếm ưu thế hơn các loài khác trong sinh quần để có thể gây mất cân bằng sinh thái.
- Ngô chuyển gen tuy có chứa protein Cry1F (Bt) nhưng không độc , không ảnh hưởng đến Collembola đã thu mẫu; hoặc lượng protein Cry1F không có hoặc có trong đất trồng khảo nghiệm là rất thấp, không đủ gây chết Collembola. Kết luận ngô chuyển gen event TC1507 không ảnh hưởng đến Collembola là dựa trên cơ sở không có sự khác biệt về thành phần loài, phân bố, số lượng loài và mức độ phong phú loài Collembola trong các nền đất trồng các giống ngô KNDR.
- Số lượng cá thể, mật độ trung bình (con/m2
- Tập hợp Collembola phổ biến và ưu thế giữa các lô đất trồng ngô đối chứng, ngô chuyển gen và chuyển gen tham khảo (stack) có nhiều điểm chung và không có nhiều sự khác biệt.
), giá trị của các chỉ số đa dạng H’ và chỉ số đồng đều J’ của quần thể Collembola trong các nền đất trồng các giống ngô chuyển gen, ngô chuyển gen tham khảo (stack) và ngô đối chứng không có sự khác biệt ở mức thống kê có ý nghĩa.

175
6 DỮ LIỆU KHÍ HẬU CHÍNH TẠI CÁC ĐỊA ĐIỂM VÀ BIỆN PHÁP QUẢN LÍ KHẢO NGHIỆM
6.1 Dữ liệu thời tiết khí hậu trong thời gian khảo nghiệm
6.1.1 Dữ liệu thời tiết khí hậu trong thời gian KNHC(tại Văn Giang)
Các thông số chính về khí hậu ghi nhận trong thời gian KNHC (2 vụ khảo nghiệm liên tiếp tại Văn Giang) được thể hiện trong Bảng 6-1 và Hình 6-1. Số liệu cho thấy: trong vụ 1 (từ 31/7 – 31/10/2010) độ ẩm trung bình, nhiệt độ trung bình và cường độ ánh sáng trung bình cao hơn hẳn so với vụ thứ 2 (gieo hạt từ 27/10/2010 – thu hoạch 24/3/2011). Điều kiện khí hậu tác động đến các chỉ tiêu đo đạc nhưng nhìn chung các dữ liệu thu được về xuất hiện của côn trùng không chủ đích, thiên địch; xuất hiện và mức độ gây hại của sâu hại và bệnh chính; cũng như về mức độ đa dạng sinh học trên cả ngô chuyển gene event TC1507 và ngô đối chứng là tương đương nhau.
Bảng 6-1:Nhiệt độ, ẩm độ và cường độ ánh sáng trung bình trong các tháng KNHC ngô tại Văn Giang (2010 – 2011)
Thời gian
Độ ẩm (%) Nhiệt độ (o Cường độ AS (lux) C) Sáng (8h) Chiều (14h) Sáng (8h) Chiều (14h) Sáng (8h) Chiều (14h)
7/2010 76.5 76.6 34.9 35.6 65103.2 81464.5 8/2010 77.6 77.9 33.4 34.5 82590.0 88460.0 9/2010 81.9 81.6 32.8 33.8 85420.0 84620.0
10/2010 61.7 61.5 23.1 23.2 46556.7 48614.2 11/2010 60.5 60.4 22.5 22.6 47751.9 49826.1 12/2010 46.2 46.6 20.2 20.2 48645.2 49383.9 1/2011 44.9 44.6 13.5 13.5 11495.7 11488.0 2/2011 80.2 80.7 19.5 19.6 8418.2 8413.2 3/2011 83.0 83.5 21.8 21.4 8474.4 8420.4

176
Hình 6-1: Độ ẩm, Nhiệt độ và Cường độ ánh sáng trung bình trong các tháng
khảo nghiệm ngô tại Văn Giang

177
6.1.2 Dữ liệu thời tiết khí hậu trong thời gian KNDR (tại 4 địa điểm)
Trong thời gian KNDR các số liệu thời tiết chính như nhiệt đọ, ẩm độ tại ruộng khảo nghiệm đều được thu thập và ghi chép lại. Kết quả ghi trong Bảng 6-2 và Bảng 6-3; Hình 6-2 và Hình 6-3.
Bảng 6-2: Nhiệt độ trung bình trong các tháng trong KNDR tại 4 địa điểm (oC)
Địa điểm TG theo doĩ
Nhiệt độ TB các tháng khảo nghiệm Tháng 5 Tháng 6 Tháng 7 Tháng 8 Ghi chú
Vĩnh Phúc 8 h 32.82 30.87 32.68 32.05 Ảnh hưởng bởi lốc xoáy 17/7/2011 14h 35.53 33.37 35.77 35.64
Nghệ An 8 h 32.82 30.87 32.68 32.05 Ảnh hưởng bởi bão số 2, ngập chết ¼ diện tích KN 14h 35.53 33.37 35.77 35.64
Đắk Lắk 8 h 23.59 23.46 22.96 22.76 14h 29.53 28.01 28.03 28.00
Đồng Nai 8 h 27.00 26.72 26.98 27.72 14h 29.18 28.68 28.03 28.62
Bảng 6-3: Ẩm độ trung bình trong các tháng khảo nghiệm tại 4 địa điểm (%)
Địa điểm TG theo dõi
Ẩm độ TB các tháng khảo nghiệm Tháng 5 Tháng 6 Tháng 7 Tháng 8 Ghi chú
Vĩnh Phúc 8 h 65.00 60.57 58.16 58.36 14h 59.82 54.97 53.58 55.05
Nghệ An 8 h 65.00 60.57 58.16 58.31 14h 59.82 54.97 53.58 55.05
Đăk Lăc 8 h 78.27 75.46 78.29 77.67 14h 75.33 73.45 75.12 74.23
Đồng Nai 8 h 73.82 75.70 75.81 74.90 14h 75.55 73.37 71.84 70.45

178
0
5
10
15
20
25
30
35
40
Sáng Chiều Sáng Chiều Sáng Chiều Sáng Chiều
Vĩnh Phúc Nghệ An Đăk Lắc Đồng Nai
Nhiệt độ trung bình trong các tháng khảo nghiệm tại 4 điểm
Tháng 5Tháng 6Tháng 7Tháng 8
0
10
20
30
40
50
60
70
80
Sáng Chiều Sáng Chiều Sáng Chiều Sáng Chiều
Vĩnh Phúc Nghệ An Đăk Lắc Đồng Nai
Tháng 5Tháng 6Tháng 7Tháng 8
Hình 6-2: Nhiệt độ trung bình tại 4 địa điểm KNDR
Hình 6-3: Ẩm độ trung bình trong các tháng tai 4 địa điểm KNDR

179
Bảng 6-4: Cây trồng vụ trước trên nền đất KNDR
Địa điểm khảo nghiệm Cây trồng vụ trước Loại đất
Trại Sản xuất Giống cây trồng Vũ Di, Vĩnh Phúc
Đậu tương - vụ Xuân Hè
Đất thịt nặng
HTX Phong Thịnh, Thanh Chương, Nghệ An
Ngô - vụ Xuân Hè Đất phù sa ven sông Nam
Nông trường Thống Nhất, Krong Pak, Đắk Lắk
Ngô - mùa mưa năm 2010
Đất đỏ bazan
TT Nghiên cứu thực nghiệm Nông nghiệp Hưng Lộc, Trảng Bom, Đồng Nai
Sắn năm 2010 Đất đỏ
- Tại Vĩnh Phúc: đất đã trồng đầu tương tại giai đoạn cây có quả xanh, giải tỏa
sau đó trồng ngô KNDR. - Tại Nghệ An: đất đã trồng ngô vụ Xuân – Hè, giải tỏa diện tích trồng khảo
nghiệm và khu vực xung quanh bán kính 200 m nhằm đảm bảo cách ly không gian. - Tại Đắk Lắk: mùa mưa năm 2010 trồng ngô. Sau thu hoạch ngô, để đất trống
qua mùa khô. 17/5/2011 trồng ngô KNDR (mùa mưa 2011) - Tại Đồng nai: mùa mưa năm 2010 trồng sắn. Sau thu hoạch sắn, để đất trống
qua mùa khô. 20/5/2011 trồng ngô KNDR (mùa mưa 2011).
Như vậy, cây trồng trên nền đất trước KNDR ngô chuyển gen event TC1507 từng địa điểm khảo nghiệm là có khác nhau. Thêm vào đó, điều kiện thời tiết khí hậu của từng vùng cũng khác nhau. Các yếu tố này, đặc biệt là yếu tố thời tiết có thể phần nào ảnh hưởng đến kết quả chung khi so sánh kết quả khảo nghiệm giữa các vụ, các vùng sinh thái khác nhau.

180
6.2 Quản lý khảo nghiệm (ảnh minh họa trong Phụ lục 3)
6.2.1 Quản lý khảo nghiệm trong KNHC
- Quản lý vận chuyển hạt giống: tất cả các hạt giống chuyển gen đều được cung cấp bởi Công ty TNHH Pioneer Hi-Bred Việt nam. Việc nhập khẩu, vận chuyển đều tuân thủ các yêu cầu về ghi nhãn, chỉ rõ người chịu trách nhiệm, địa chỉ và số điện thoại liên lạc. Những hạt giống này được chuyển tới Viện Di truyền Nông nghiệp tuân thủ mọi quy định về vận chuyển vật liệu biến đổi gen. Số hạt gieo thừa được bảo quản trong tủ có khóa của công ty TNHH Pioneer Hi-Bred Việt Nam (Phụ lục 1 mục 6.2 và lưu trong hồ sơ). - Địa điểm khảo nghiệm: có hàng rào để ngăn chặn sự t hâm nhập của các loài động vật và người không phận sự (hình ảnh khu khảo nghiệm trong Phụ lục 3). Có chòi canh gác 24/24 h; 7 ngày/tuần đảm bảo những người không phận sự không được ra vào khu khảo nghiệm.
- Vành đai bảo vệ: Trồng 4 hàng ngô 30Y87 gi ống nền (Đ/C không chuyển gen) xung quanh ruộng khảo nghiệm.
- Cách ly về thời gian và không gian: Địa điểm khảo nghiệm được khảo sát trước khi gieo, đảm bảo tuân thủ sự cách ly về thời gian trước hoặc sau ngày gieo tối thiểu là 25 ngày so với các ruộng ngô trồng xung quanh và với bán kính cách ly tối thiểu 200 mét.
- Biện pháp phòng tránh rủi ro cho cán bộ tham gia khảo nghiệm:
+ Trang bị bảo hộ lao động như ủng, găng tay, khẩu trang và quần áo bảo hộ. + Tập huấn kỹ thuật về việc tuân thủ quản lý rủi ro theo đúng thông thư 69 của
Bộ NN&PTNT và nghị đinh 69 của Chính phủ trước mỗi khảo nghiệm (trong phòng) và trong thời gian khảo nghiệm (ngoài đồng ruộng) (Phụ lục 3).
- Gieo trồng: Gieo bằng tay, theo hàng, mỗi hốc gieo 2 hạt. Đảm bảo độ đồng đều về độ sâu lấp đất, khoảng cách và mật độ. Số hạt giống sử dụng cho KNHC được gieo trồng hết, hạt thừa được tiêu hủy ngay (đào hố chôn ngay trong khu khảo nghiệm) . Tiến hành tiêu hủy theo đúng quy định An toàn Sinh học theo thông tư 69/2009/TT-BNNPTNT. - Kiểm soát mật độ: Khi 50% số cây đạt giai đoạn V2-V4 (khoảng 12-15 ngày sau trồng). Các lô thí nghiệm được tỉa thưa nhằm đảm bảo đúng mật độ, khoảng cách thí nghiệm, bằng cách nhổ bỏ những cây yếu. Thu gom những cây đã nhổ và tiêu hủy

181
theo hướng dẫn trong thông tư 69/2009/TT-BNNPTNT ngày 27 tháng 10 năm 2009 của Bộ NN&PTNT. Việc kiểm soát mật độ nhằm đảm bảo 1 cây/hốc (với tất cả các giống KNHC). - Chăm sóc, bón phân, tưới nước: Các yếu tố phi thí nghiệm được tiến hành đồng đều trên tất cả các giống, trong lô khảo nghiệm, cho cả hai vụ KNHC. - Thu hoạch và quản lý đồng ruộng sau thu hoạch: Khảo nghiệm được thu hoạch khi cây ngô chín sinh lý (khoảng 100 - 110 ngày tính từ khi gieo). Sau khi thu toàn bộ ngô bắp, đo đếm các chỉ tiêu nông học. Toàn bộ bắp ngô, bao gồm cả ngô Đ/C và ngô bắp của hàng bảo vệ, được tiêu hủy bằng luộc chín và chôn sâu. Tất cả thân, rễ cây và các vật liệu khác (bao gồm cả hàng bảo vệ ) được đốt tại ruộng. Mọi hoạt động liên quan đến thu hoạch đều được giám sát bởi đại diện cơ quan quản lý nhà nước, Hội đồng An toàn Sinh học và được lưu trong hồ sơ khảo nghiệm theo đúng quy định tại Phụ lục 6.3 của Thông tư 69/2009/TT-Bộ NNPTNT (biên bản kèm trong Phụ lục 2).
Ruộng khảo nghiệm được xử lý bằng cách cày ngay sau khi thu hoạch và được tưới cho đủ ẩm nhằm thúc đẩy sự nảy mầm và sinh trưởng của các ngô mọc tự do. Quá trình theo dõi ruộng sau khảo nghiệm được tiến hành 7 -10 ngày /lần. Tất cả các cây ngô mọc lên tìm thấy trong địa điểm KNHC được đếm, nhổ bỏ và tiêu hủy. Quy trình theo dõi này được nhắc lại cho đến khi không tìm thấy bất kỳ cây ngô mọc tự do. Toàn bộ quá trình theo dõi và quản lý sau thu hoạch được lưu trong hồ sơ khảo nghiệm (Phụ lục 2) theo mẫu quy định tại Phụ lục 6.4 của Thông tư 69/2009/TT-Bộ NNPTNT.
- Lưu trữ và quản lý hồ sơ khảo nghiệm: Tất cả các hoạt động diễn ra trong quá trình KNHC, từ khi triển khai nhập khẩu giống đến khi kết thúc khảo nghiệm đều tuân thủ theo Giấy phép khảo nghiệm của Bộ NN&PTNT. Đồng thời được lưu trong hồ sơ khảo nghiệm theo các mẫu quy định tại Thông tư 69/2009/TT-Bộ NNPTNT về khảo nghiệm đánh giá rủi ro cây trồng biến đổi gen, và Nghị định 69/2010/NĐ-CP về an toàn sinh học đối với sinh vật biến đổi gen, mẫu vật di truyền và sản phẩm của sinh vật biến đổi gen. - Các thực hành quản lý canh tác khác: Tuân theo hướng dẫ n đánh giá khảo nghiệm của Cục Trồng trọt – Bộ NN&PTNN.

182
6.3 Quản lý rủi ro trong KNDR
6.3.1 Vật liệu hạt giống chuyển gene sử dụng trong khảo nghiệm
Vật liệu hạt giống chuyển gen sử dụng trong khảo nghiệm: - 30Y87H (TC1507; ngô Bt kháng sâu hại bộ cánh vảy) - 30Y87HR, Đ/C tham khảo (stack TC1507xNK603; ngô Bt kháng sâu hại bộ
cánh vảy và kháng thuốc trừ cỏ glyphosate)
Vật liệu hạt giống không chuyển gen sử dụng trong khảo nghiệm: đối chứng không chuyển gen (giống nền 30Y87)
Tất cả các hạt giống chuyển gen đều được cung cấp bởi Công ty TNHH Pioneer Hi-Bred Việt Nam. Việc nhập khẩu, vận chuyển đều tuân thủ các yêu cầu về ghi nhãn chỉ rõ người chịu trách nhiệm, địa chỉ và số điện thoại liên lạc (Phụ lục 2). Những hạt giống này được chuyển cho Viện Di truyền Nông nghiệp sau đó đưa đi các điểm khảo nghiệm. Việc nhập khẩu, vận chuyển đều tuân thủ theo các quy định về vận chuyển vật liệu biến đổi gen. Số hạt giống gieo thừa được tiêu hủy tại ruộng khảo nghiệm bằng cách đào hố sâu 50cm và chôn lấp ngay.
6.3.2 Quản lý cách ly
a) Quản lý cách ly sinh sản đối với các ruộng ngô khác xung quanh khu vực khảo nghiệm được duy trì bằng một trong các phương pháp sau:
- Cách ly không gian: trong khoảng cách 200m bán kính xung quanh ruộng khảo nghiệm không được trồng ngô trước hoặc sau 25 ngày tiến hành gieo khảo giống khảo nghiệm.
- Cách ly thời gian: thời gian gieo hạt sẽ được triển khai trước hoặc sau các ruộng ngô xung quanh điểm khảo nghiệm tối thiểu 25 ngày để đảm bảo thời gian trỗ cờ, phun râu của ruộng khảo nghiệm không bị trùng với thời gian trỗ cờ, phun râu trên các ruộng ngô truyền thống xung quanh.
a) Quản lý cách ly chống xâm nhập của các đối tượng không tham gia vào khu khảo nghiệm: làm hàng rảo bảo vệ và lưới để phòng tránh xâm nhập của gia súc, chim, chuột vv. Ngoài ra còn có chòi canh và bảo vệ 24/24 giờ/ngày để bảo đảm không có sự xâm nhập của các đối tượng không mong muốn vào khu vực khảo nghiệm.
b) Quản lý cách ly chống thất thoát vật liệu khảo nghiệm: Ruộng khảo nghiệm có

183
rào lưới bảo vệ nghiêm ngặt và có các rãnh thoát nước , lưới ngăn mắt lưới nhỏ 2mm phòng tránh thất thoát vật liệu ra môi trường. Cây con sau tỉa thưa được chôn ngay trong khu khảo nghiệm (tương tự trong KNHC).
c) Tập huấn cho cán bộ và những người tham gia khảo nghiệm:
+ Trang bị bảo hộ lao động như ủng, găng tay, khẩu trang và quần áo bảo hộ. + Tập huấn kỹ thuật về việc tuân thủ quản lý rủi ro theo đúng thông thư 69 của
Bộ Nông nghiệp và nghị đinh 69 của Chính phủ trước khi khảo nghiệm (trong phòng) và trong thời gian KNDR (ngoài đồng ruộng).
6.3.3 Thu hoạch và quản lý đồng ruộng sau thu hoạch
- Trước khi thu hoạch, Viện Di truyền Nông nghiệp thông báo đến người chịu trách nhiệm theo dõi, giám sát của Bộ Nông nghiệp và Phát triển Nông thôn, Sở Nông nghiệp, Sở tài nguyên Môi trường của các địa phương tham gia khảo nghiệm về thời gian thu hoạch. Khi thu hoạch luôn có sự giám sát và hiện diện của cán bộ nghiên cứu Viện Di truyền Nông nghiệp; đại diện giám sát Bộ Nông nghiệp; các sở Tài nguyên môi trường và Sở Nông nghiệp của các địa phương và nhân viên có trách nhiệm đối với khảo nghiệm đánh giá rủi ro của đơn vị đăng ký khảo nghiệm.
- Thu hoạch được tiến hành từng ô một. Sau khi đã thu hoạch to àn bộ các cây ở hàng bảo vệ, ô thu hoạch được ghi nhận và thu thập số liệu như phần phương pháp thí nghiệm đã mô tả.
- Viện Di truyền Nông nghiệp và các đơn vị phối hợp khảo nghiệm có trách nhiệm kiểm tra kĩ để đảm bảo không còn bất cứ cây ngô chuyển gene cũng như cây trồng cùng loài còn tồn tại trong thí nghiệm trước khi rời địa điểm khảo nghiệm.
- Thành viên tham gia, thời gian bắt đầu và kết thúc thu hoạch được ghi chi tiết trong sổ theo dõi và các phụ lục theo qui định.
- Khảo nghiệm được thu hoạch khi cây ngô chín sinh lý, khoảng 95-100 ngày tính từ khi gieo. Toàn bộ sản phẩm ngô thu hoạch tại ruộng khảo nghiệm sẽ được cam kết sử dụng theo đúng quy định trong giấy phép khảo nghiệm.
6.3.4 Quản lý và xử lý đối với các trường hợp phát tán ngẫu nhiên
- Loại bỏ hoàn toàn cây/vật liệu chuyển gen trong quá trình vệ sinh khu khảo nghiệm.
- Tất cả các thiết bị dùng cho hạt giống hay dùng ở nơi khảo nghiệm được lau chùi sạch sẽ trước khi đưa vào sử dụng ở nơi tiến hành khảo nghiệm, bao gồm hạt giống và các vật liệu cây trồng có thể có trước khi tiến hành khảo nghiệm. Tất cả các

184
thiết bị sử dụng trong khảo nghiệm và thu hoạch được vệ sinh sạch sẽ trước khi đưa đến nơi khảo nghiệm để loại bỏ các vật liệu cây chuyển gen có thể đem vào hay thất thoát ra ngoài khi vận chuyển. Các phương pháp lau chùi có thể bao gồm làm sạch bằng tay, nén khí, hút chân không các hạt còn sót lại hay xịt nước áp lực cao.Vật liệu thu được được tiêu huỷ và ghi nhận trong sổ theo dõi khảo nghiệm.
- Người quản lý khảo nghiệm, giám sát khu khảo nghiệm và vùng đệm xung quanh không để tồn tại các cây trồng không mong muốn trên ruộng khảo nghiệm, bao gồm các cây mọc hoang. Các cây này bị loại bỏ bằng cách đốt trước khi ra hoa.Người quản lý khu khảo nghiệm theo dõi khu vực khảo nghiệm ít nhất 7-10 ngày/lần trong thời gian từ khi thu hoạch đến khi không còn cây ngô nào mọc trên khu đất khảo nghiệm, ghi nhận các cây cần hạn chế mọc trong khu khảo nghiệm và tiêu hủy ngay trong khu vực khảo nghiệm.
- Tất cả vật liệu trong khảo nghiệm đều được tiêu huỷ, kể cả hàng bảo vệ. Ruộng khảo nghiệm phải được cày lại ngay sau khi kết thúc khảo nghiệm với sự kiểm tra, giám sát của những thành viên tham gia thu hoạch.
6.3.5 Quản lý, ghi chép hồ sơ tư liệu
- Hồ sơ tư liệu liên quan đến ngô chuyển gen được bảo mật trong nhóm công tác, tuyệt đối không cung cấp cho bên thứ 3 khi chưa có sự đồng ý của Viện Di truyền Nông nghiệp và đơn vị đăng ký khảo nghiệm là Công ty Pioneer Hi-Bred Việt Nam.
- Hồ sơ tư liệu được quản lí chặt chẽ, giao cho người phụ trách khảo nghiệm chính là TS Phạm Thị Liên – Phó giám đốc Trung tâm T hực nghiệm Sinh học Nông nghiệp Công nghệ cao, Viện Di truyền Nông nghiệp. Tất cả các hoạt động (như ngày, giờ đặt bẫy, quan sát, các lưu ý lúc quan sát, vv) được ghi nhận đúng lúc, chính xác và bằng mực trong suốt quá trình thực hiện thí nghiệm.
- Tất cả công thức thí nghiệm sẽ được ghi ngày trồng, ký tên bởi người trồng. Tất các các ghi nhận hoạt động, phiếu thu nhận thông tin, và các lưu ý lúc quan sát sẽ gửi về người chịu trách nhiệm chính khảo nghiệm khi kết thúc khảo nghiệm. Hồ sơ được cất giữ trong tủ có khoá. Hồ sơ tài liệu bao gồm:
+ Sổ nhật kí khảo nghiệm + Nhật ký ra, vao khu khảo nghiệm + Sổ số liệu thô thu thập trong khi khảo nghiệm + Các phụ lục theo dõi

185
6.3.6 Tiêu hủy sản phẩm và vật liệu di truyền biến đổi gen
Toàn bộ sản phẩm sau thu hoạch bao gồm thân cây ngô và bắp ngô được tiêu huỷ theo đúng quy định hiện hành của Việt Nam. Các biện pháp tiến hành bao gồm: chôn, nghiền nát làm rồi rắc ra ruộng sau đó cầy lấp, vv... (Bảng 5-5; ảnh trong Phụ lục 3). Ruộng khảo nghiệm được quản lý sau thu hoạch theo đúng quy định tại nghị định 69/2010/NĐCP và các qui định hiện hành có liên quan khác.
Tóm lại: Trong quá trình khảo nghiệm đã tuân thủ theo đúng hướng dẫn tại Thông tư 69/2009/TT-BNNPTNT ngày 27/10/2009 về Quy định khảo nghiệm đánh giá rủi ro đối với đa dạng sinh học và môi trường của giống cây trồng biến đổi gen. Nghị định số 69/2010/NĐ-CP ngày 21/6/2010 về an toàn sinh học đối với sinh vật biến đổi gen, mẫu vật di truyền và sản phẩm của sinh vật biến đổi gen.
Chính vì vậy, quá trình khảo nghiệm (cả KNHC và KNDR) không bị bất kì rủi ro đáng tiếc ảnh hưởng đến kết quả khảo nghiệm, đến sức khỏe những người tham gia khảo nghiệm và không có bất cứ vật liệu chuyển gen nào thất thoát ra khỏi khu vực khảo nghiệm mà không được quản lý.

186
Bảng 6-5: Phương pháp tiêu hủy vật liệu trong khu vực KNDR tại 4 địa điểm
khảo nghiệm (vụ Hè – Thu, 2011)
Địa điểm khảo nghiệm Phương pháp tiêu hủy Lượng hạt
tiêu hủy (tấn)
Trại Sản xuất Giống cây trồng Vũ Di, Vĩnh Phúc
- Bắp: nghiền nhỏ, rắc ra ruộng - Cây: đốt cùng với hạt, sau đó cày vùi, lấp - Các vật liệu khác: đốt
6,2
HTX Phong Thinh, huyện Thanh Chương, Nghệ An
- Bắp: trộn với vôi bột, chôn - Cây :đốt - Các vật liệu khác: đốt
5,3
Nông trường Thống Nhất, Krong Pak, Đắk Lắk
- Bắp: nghiền nhỏ, chôn - Cây: đốt cùng với hạt, sau đó cày vùi, lấp - Các vật liệu khác: đốt
10,5
TT Nghiên cứu thực nghiệm Nông nghiệp Hưng Lộc, Trảng Bom, Đồng Nai
- Bắp: trộn với vôi bột, chôn - Cây :đốt - Các vật liệu khác: đốt
7,1

191
Bảng 6-6: Lịch trình các đoàn thuộc cơ quan quản lý tham gia kiểm tra giám sát khảo nghiệm
TT
Nội dung
Thời gian
Địa điểm
Thành phần (tên cụ thể cán bộ tham dự)
Loại hình khảo nghiệm
1. Giám sát khi gieo hạt vụ 1
31/7/2010
Trạm
Văn Giang
Ông: Nguyễn Văn Tuất - CT HĐATSH Hạn chế, vụ 1 Bà: Kiều Anh - Cục BTĐDSH - Bộ TNMT Ông: Nguyễn Văn Cần - phòng TNMT huyện VG
2.
Giám sát thu hoạch và tiêu hủy vật liệu biến đổi gen
31/10/2010
Trạm Văn Giang
Ông: Nguyễn Văn Tuất- CT HĐATSH Ông: Lê Huy Hàm - PCT HĐATSH Bà: Kiều Anh - Cục BTĐDSH - Bộ TNMT Ông: Nguyễn Văn Cần - phòng TNMT huyện VG Ông Phạm Văn Toản - HĐATSH
3.
Giám sát gieo hạt vụ 2
3/11/2010
Trạm Văn Giang
Bà: Kiều Anh - Cục BTĐDSH - Bộ TNMT Hạn chế, vụ 2 Ông: Nguyễn Văn Cần - Phòng TNMT huyện VG
4.
Giám sát thu hoạch và tiêu hủy vật liệu biến đổi gen
24/3/2011
Trạm Văn Giang
Ông Nguyễn Văn Tuất - CT HĐATSH Bà: Kiều Anh - Cục BTĐDSH - Bộ TNMT Ông: Nguyễn Văn Cần - Phòng TNMT huyện VG Ông: Phạm Văn Toản - HĐATSH
5.
Giám sát gieo hạt tại Vĩnh Phúc
7/5/2011
Vĩnh Phúc
Ông: Nguyễn Hữu Khang -Vụ KHCN&MT Bộ NN&PTNT
Diện rộng, 4 vùng sinh Thái
Bà: Kiều Anh - Cục BTĐDSH - Bộ TNMT Ông: Nguyễn Văn Dũng - PGĐ sở NN Vĩnh Phúc Bà Trần Bích Thủy - Sở Tài nguyên MT VP
6.
Giám sát gieo hạt tại Nghệ An
13/5/2011
Nghệ An
Ông: Phạm Văn Toản - HĐATSH Ông: Nguyễn Kim - Sở NN tỉnh Nghệ An Ông: Nguyễn Sơn - Sở TNMT Nghệ An

192
TT
Nội dung
Thời gian
Địa điểm
Thành phần (tên cụ thể cán bộ tham dự)
Loại hình khảo nghiệm
7.
Giám sát gieo hạt tại Đăk lăc
17/5/2011
Đắk Lắk
Bà: Nguyễn Thị Thanh Hương - Sở TNMT Đắk Lắk Ông: Nguyễn Hoàng Anh - Sở NN tỉnh Đắk Lắk
8.
Giám sát gieo hạt tại Đồng Nai
20/5/2011
Đồng Nai
Ông: Dương Hoa Xô - HĐATSH Ông: Trần Hoài Ân - Sở NN tỉnh Đồng Nai Ông: Mai Thanh Lâm - phòng TNMT huyện Trảng Bom
9.
Giám sát thu hoạch và tiêu hủy vật liệu biến đổi gen tại Vĩnh Phúc
13/8/2011
Vĩnh Phúc
Ông: Nguyễn Hữu Khang -Vụ KHCN&MT, Bộ NN&PTNN
Ông: Phạm Văn Toản - HĐATSH Bà: Kiều Anh - Cục BTĐDSH - Bộ TNMT Ông: Nguyễn Văn Dũng - PGĐ sở NN Vĩnh Phúc Bà: Trần Bích Thủy - Sở Tài nguyên MT VP
10.
Giám sát thu hoạch và tiêu hủy vật liệu biến đổi gen tại Nghệ An
22/8/2011
Nghệ An
Ông: Nguyễn Hữu Khang -Vụ KHCN&MT, Bộ NN&PTNN
Ông: Nguyễn Kim - Sở NN tỉnh Nghệ An Ông: Nguyễn Sơn - Sở TNMT Nghệ An
11.
Giám sát thu hoạch và tiêu hủy vật liệu biến đổi gen tại Đắk Lắk
26/8/2011
Đắk Lắk
Ông: Phạm Văn Toản - HĐATSH Ông: Nguyễn Hữu Khang -Vụ KHCN&MT, Bộ
NN&PTNN Bà: Nguyễn Thị Thanh Hương - Sở TNMT Đắk Lắk Ông: Nguyễn Hoàng Anh - Sở NN tỉnh Đắk Lắk
12.
Giám sát thu hoạch và tiêu hủy vật liệu biến đổi gen tại Đồng Nai
29/8/2011
Đồng Nai
Ông: Dương Hoa Xô - HĐATSH Ông: Phạm Văn Toản - HĐATSH Ông: Nguyễn Hữu Khang -Vụ KHCN&MT, Bộ
NN&PTNN Ông: Trần Hoài Ân - Sở NN tỉnh Đồng Nai Ông: Mai Thanh Lâm - Phòng TNMT huyện Trảng Bom

193
7 KẾT LUẬN VÀ ĐỀ NGHỊ
7.1 Kết luận
Trên cơ sở k ết quả khảo nghiệm ngô biến đổi gen event TC1507 trong khảo nghiệm hạn chế 2 vụ liên tiếp tại Văn Giang - Hưng Yên, khảo nghiệm diện rộng ngô chuyển gen event TC1507 được triển khai tại 4 địa điểm khác nhau, đại diện 4 vùng trồng ngô chính tại Việt Nam (Vĩnh Phúc, Nghệ An, Đắk Lắk và Đồng Nai). Kết quả khảo nghiệm cho phép đưa ra những kết luận sau: 1. Toàn bộ quá trình thực hiện KNHC và KNDR được triển khai theo đúng các
qui trình đã được phê duyệt bởi Hội đồng An toàn Sinh học – Bộ Nông nghiệp và Phát triển Nông thôn. Tuân thủ chặt chẽ và đầy đủ các qui định hiện hành về yêu cầu điều kiện khảo nghiệm như địa điểm, thời gian, qui mô khảo nghiệm , đơn vị thực hiện khảo nghiệm vv…
2. Toàn bộ quá trình thực hiện KNHC và KNDR tuyệt đối tuân thủ theo đúng hướng dẫn tại Thông tư 69/2009/TT-BNNPTNT ngày 27/10/2009 về Quy định khảo nghiệm đánh giá rủi ro đối với đa dạng sinh học và môi trường của giống cây trồng biến đổi gen ; Nghị định số 69/2010/NĐ-CP ngày 21/6/2010 Về an toàn sinh học đối với sinh vật biến đổi gen, mẫu vật di truyền và sản phẩm của sinh vật biến đổi gen; và các quy định hiện hành khác có liên quan đối với yêu cầu giám sát, quản lý rủi ro trong khảo ngh iệm giống cây trồng BĐG của Việt Nam
3. Các phương pháp, kĩ thuật thực hiện KNHC và KNDR đảm bảo tính khoa học, minh bạch, chính xác và đều đã được công bố, sử dụng rộng rãi tại Việt Nam và/hoặc trên thế giới. Các số liệ u thu thập từ KNHC và KNDR và công bố trong báo cáo là khách quan, trung thực, chính xác, minh bạch và hoàn toàn đáng tin cậy.
4. Dựa trên các ghi nhận về các chỉ số đa dạng quần thể sinh vật không chủ đích;
các số liệu thành phần, số lượng loài, phân bố và mức độ phong phú bọ đuôi bật Collembola (trong đất và trên lớp đất mặt) ; số liệu về mật độ, sự đa dạng các loài chân khớp đại diện cho các nhóm côn trùng chỉ thị đóng vai trò quan

194
trọng trong chuỗi thức ăn (các loài sâu hại NTOs, các loài bọ rùa ăn thịt vv…) kết luận ngô event TC1507 không gây ảnh hưởng bất lợi tới sinh vật không chủ đích trong điều kiện khảo nghiệm ở Việt Nam.
5. Không có nguy cơ trôi/phát tán gen từ ngô TC1507 sang các sinh vật khác. Lí do: (1) Việt Nam không có các loài ngô hoang để có nguy cơ xảy ra hiện lai khác loài; (2) kết quả âm tính đối với tất cả các mẫu từ lô Đ/C (giống nền) ở tất cả các vụ/vùng đã khảo nghiệm.
6. Trên cơ sở tương đồng về các đặc tính nông sinh học, mức độ mẫn/kháng đối với các loại bệnh, sâu hại không chủ đích chính trên ngô giữa các giống ngô BĐG và ngô đối chứ ng trong KNHC và KNDR. Kết luận ngô event TC1507 không có nguy cơ trở thành cỏ dại hay nguy cơ lan tràn xâm lấn môi trường tự nhiên Việt Nam.
7. Ngô event TC1507 đã được cấp phép sử dụng làm thực phẩm/thức ăn gia súc tại 16 nước; được trồng thương mại tại 7 nước; và được chứng minh an toàn đối với môi trường tại 6 nước khác nhau trên thế giới (Bảng 2-4). Quá trình KNHC và KNDR tại Việt Nam cũng không hề ghi nhận bất cứ dấu hiệu bất thường, hay khác biệt giữa các giống ngô BĐG và ngô Đ/C . Chứng tỏ ngô TC1507 không gây nguy cơ trở thành dịch hại, không là nguyên nhân ảnh hưởng tiêu cực đối với môi trường và các hệ sinh thái ở Việt Nam.

195
7.2 Đề nghị
Từ các kết quả khảo nghiệm hạn chế 2 vụ liên tiếp và khảo nghiệm diện rộng tại
4 địa điểm đại diện 4 vùng sinh thái trồng ngô chính của Việt Nam để đánh giá rủi ro
đối với an toàn sinh học và môi trường của ngô chuyển gen event TC1507, không
nhận thấy có bất kỳ rủi ro và nguy cơ nào ảnh hưởng bất lợi đến đa dạng sinh học
hoặc trở thành cỏ dại hay dịch hại trong điều kiện canh tác tại Việt Nam.
Kính đề nghị Hội đồng An toàn Sinh học – Bộ Nông nghiệp và Phát triển Nông
thôn công nhận kết quả khảo nghiệm ngô chuyển gen event TC1507 an toàn với
đa dạng sinh học và môi trường sinh thái Việt Nam.
Hà nội, ngày 8 tháng 5 năm 2012
Xác nhận của cơ quan khảo nghiệm
Xác nhận của đơn vị đăng kí khảo nghiệm
Phụ trách khảo nghiệm chính
TS. Phạm Thị Liên

196
TÀI LIỆU THAM KHẢO CHÍNH
Tài liệu tham khảo tiếng Anh
1. Ahmad, A., G.E. Wilde, and K.Y. Zhu. 2005. Detectability of coleopteran-specific Cry3Bb1 protein in soil and its effect on nontarget surface and below-ground arthropods. Environ. Entomol. 34:385-394.
2. Al-Deeb A. Mohammad, G. E. Wilde, 2003. Effect of Bt corn expressing the Cryp3Bb1 toxin for corn rootworm control on aboveground nontarget arthropods. Environ. Entomol., 32: 1164-1170.
3. Al-Deeb A. Mohammad, G. E. Wilde, J. M. Blair, T. C. Todd. 2003. Effect of Bt Corn for corn rootworm control on nontarget soil microarthropods and nematodes. Environ. Entomol., 32(4): 859-865
4. Al-Deeb M.A., Wilde G.E., Higgins R.H., 2001. No effect of Bacillus thuringiensis corn and Bacillus thuringiensis on the predator Orius insidiosus (Hemiptera: Anthocoridae). Environmental Entomology 30: 625-629.
5. Babrikova T., Kuzmanova I., 1984. The toxicity of biological preparations based on Bacillus thuringiensis to some stages of Chrysopa septempunctata Wesm., Chrysopa formosa Br. and Chrysopa perla L. Gradinarska i Lozarska Nauka, 21: 55-59.
6. Babrikova T., Kuzmanova I., Lai N.T., 1982. The effect of biological preparations of Bacillus thuringiensis on some stages of the lacewing Chrysopa carnea Steph. Gradinarska i Lozarska Nauka, 19: 40-45.
7. Bai Y. Y., R. H. Yan, G. Y. Ye, F. N. Huang, J. A. Cheng. 2010. Effects of transgenic rice expressing Bacillus thuringiensis Cry1Ab protein on ground-dwelling Collembolan community in postharvest seasons. Environmental Entomology, 39(1): 243-251.
8. Bakonyi G., F. Szira, I. Kiss, I. Villanyi, A. Seres, A. Szekacs. 2006. Preference tests with collembolas on isogenic and Bt-maize. European Journal of Soil Biology, 42: 132-135.
9. Bitzer, R.J., M.E. Rice, C.D. Pilcher, C.L. Pilcher, and W.-K.F. Lam. 2005. Biodiversity and community structure of epedaphic and euedaphic springtails (Collembola) in transgenic rootworm Bt corn. Environ. Entomol. 34:1346-1376.
10. Blair J.M., P. J. Bohlen; D. W. Freckman, 1996. Soil invertebrates as indicators of soil quality. SSSA Special Publication No 49, Soil Science Society, Madison, WI: 283-301.

197
11. Boyce L. Richard, 2005. Life Under Your Feet: Measuring Soil Invertebrate Diversity, Teaching Issues and Experiments in Ecology, Vol. 3: http://tiee.ecoed.net/vol/v3/experiments/soil/abstract.html
12. Bravo A, Gill SS, Soberón M (2007) Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 49: 423-435
13. Brookes G., P. Barfoot, 2006. GM crops: The first ten years – Global socio-economic and environmental impacts. Brief No 36, ISAAA, Ithaca, NY.
14. Candolfi M.P., Brown K., Grimm C., Reber B., Schmidli H., 2004. A faunistic approach to assess potential side-effects of genetically modified Bt-corn om non-target arthropods under field conditions. Biocontrol Sci. and Tech., 14(2): 129-170.
15. CFIA (1994) The Biology of Zea mays (L.) (Maize). Canadian Food Inspection Agency, BIO1994-11
16. Colwell R., 2000. EstimateS: Statistical estimation of specÝe richness and shared species from samples. Version 6. In. User's Guide and application published at: http://viceroy.eeb.uconn.edu/estimates
17. Cortet J., B. S. Griffiths, M. Bohanec, D. Demsar, M. N. Andersen, S. Caul, A. N. E. Birch, C. Pernin, E. Tabone, A. de Vaufleury, X. Ke, P. H. Krogh. 2005. Impact of transgenic VIP3AxCry1Ab Lepidopteran-resistant field corn on the nontarget arthropod community. Environmental Entomology, 34(5): 1267-1291.
18. Cortet J., B. S. Griffiths, M. Bohanec, D. Demsar, M. N. Andersen, S. Caul, A. N. E. Birch, C. Pernin, E. Tabone, A. de Vaufleury, X. Ke, P. H. Krogh. 2007. Evaluation of effects of transgenic Bt maize on microarthropods in a European multi-site experiment. Pedobiologia, 51: 207-218.
19. CRL, EU validation report (2005). Event-specific method for the quantitation of maize line TC1507 using real-time PCR.
20. Dale P. J., B. Clarke, E. M. G. Fontes. 2002. Potential for the environmental impact of transgenic crops. Nature Biotechnology, 20: 567-574.
21. De Maagd RA, Bravo A, Berry C, Crickmore N, Schnepf HE (2003) Structure, Diversity, and Evolution of Protein Toxins from Spore-Forming Entomopathogenic Bacteria. Annual Review of Genetics 37: 409-433
22. Dively G. and R. Rose, 2002. Effects of Bt transgenic and conventional insecticide control strategies on the natural enemy community in sweet corn. In: R. Van Driesche (ed.), First International Symposium on Biological Control

198
of Arthropods. U.S. Department of Agriculture, Forest Service, Morgantown, WV.: 265-274.
23. Dutton A., Klein H., Romeis J., Bigler F., 2002. Uptake of Bt-toxin by herbivors feeding on transgenic maize and consequences for the predator Chrysoperla carnea. Ecological Entomology, 27:441-447.
24. Dutton A., Romeis J., Bigler F., 2003. Assessing the risks of insect resistant transgenic plants on entomophagous arthropods: Bt-maize expressing Cry1A as a case study. Biocontrol, 48: 611-636.
25. Eckes P, Uijtewaal B, Donn G (1989) A synthetic gene confers resistance to the broad spectrum herbicide L-phosphinothricin in plants. Journal of Cellular Bioechemistry Supplement 13D: 334, Abstract M516
26. Effect of transgenic brinjal expressing cry1Ac gene on soil microflora, collembola, nematodes, and earthworms, and assessment of Br protein in soil. 2009. Maharashtra Hybrid Seeds Company Ltd.
27. EFSA, 2005. Opinion of the Scientific Panel on Genetically Modified Organisms on a request from the Commission related to the notification (Reference C/ES/01/01) for the placing on the market of insect-tolerant genetically modified maize 1507, for import, feed and industrial processing and cultivation, under Part C of Directive 2001/18/EC from Pioneer Hi-Bred International/Mycogen Seeds. The EFSA Journal 181, 1-33.
28. EFSA, 2010. Minutes of the 61st plenary meeting of the Scientific Panel on Genetically Modified Organisms held on 20-21 October 2010 in Parma, Italy; request from the European Commission to review scientific studies related to the Testbiotech report on the cultivation of maize 1507, http://www.efsa.europa.eu/en/events/event/gmo101020-m.pdf
29. EPA, 2005. Biopesticides Registration Action Document Bacillus thuringiensis Cry1F Corn, http://www.epa.gov/opp00001/biopesticides/ingredients/tech_docs/brad_006481.pdf
30. Frampton G. K., 1997. The potential of Collembola as indicators of pesticide usage; evidence and methods from the UK arable ecosystem. Pedobiologia, 41: 179-184.
31. Frouz J., D. Elhottova, M. Helingerova, F. Kocourek. 2008. Effect of bt-corn on soil invertebrates, soil microbial community and decomposition rates of corn post-harvest residues under field and laboratory conditions. Journal of Sustainable Agriculture, 32(4): 645-655.

199
32. Gormy C., Grum L., 1993. Methods in Soil Zoology. PWN- Polish Scientife publisher, Warszawa, 518-620.
33. Greenslade P., 1997. Are Collembola useful as indicators of the conservation value of native grasslands?. Pedobiologia, 41: 215-220.
34. Gisin H., 1960. Collembolenfauna Europas. Museum D’histoire Naturelle Geneve: 1-300.
35. Head G., J. B. Surber, J. A. Watson, J. W. Martin, J. J. Duan, 2002. No detection of Cryp1Ac protein in soil after multiple years of transgenic Bt cotton (Bollgard) use. Environ. Entomol., 31: 30-36.
36. Heckmann L.H., B. S. Griffiths, S. Caul, J. Thompson, M. Pusztai-Carey, W. J. Moar, M. N. Andersen, P. H. Krogh. 2006. Consequences for Protaphorura armata (Collembola: Onychiuridae) following exposure to genetically modified Bacillus thuringiensis (Bt) maize and non-Bt maize. Environmental Pollution, 142: 212-216.
37. Hérouet C, Esdaile DJ, Mallyon BA, Debruyne E, Schulz A, Currier T, Hendrickx K, vander Klis R-J, Rouan D (2005) Safety evaluation of the phosphinothricin acetyltransferase proteins encoded by the pat and bar sequences that confer tolerance to glufosinate-ammonium herbicide in transgenic plants. Regulatory Toxicology and Pharmacology 41: 134-149.
38. Hilbeck A., Baumgartner M., Fried P.M., Bigler F., 1998. Effect of transgenic Bacillus thuringiensis corn-fed prey on mortality and development time of immature Chrysoperla carnea (Neuroptera: Chrysopidae). Environmental Entomology 27: 480-487.
39. Hilbeck A., Moar W.J., Pusztai-Carey, Filippini A., Bigler F., 1999. Prey-medialed effects of Cry1Ab toxin and protoxin and Cry2A protoxin on the predator Chrysoperla carnea. Entomol.Exp. Appl. 91: 305-316.
40. Honemann L., C. Zurbrugg, W. Nentwig. 2008. Effects of Bt-corn decomposition on the composition of the soil meso- and macrofauna. Applied Soil Ecology, 40(2): 203-209.
41. Hopkins D. W. and E. G. Gregorich, 2003. Detection and decay of the Bt endotoxin in soil from a field trial with genetically modified maize. Eur. J. Soil. Sci., 54: 793-800.
42. IBC No. PION-19-1111. Application for Commercialization of Genetically Engineered Organisms and Products Thereof
43. James C., 2009. Global status of commercialized biotech/GM crops: 2009. Brief No 41, ISAAA, Ithaca, NY.

200
44. Lee S.H., Park Y.H., Kim J.k., Park K.W., KIM Y.M., (2004). Qualitative PCR Method for Detection of Genetically Modified Maize Lines NK603 and TC1507. Agric. Chem. Biotechnol. 47(4), 185-188.
45. Mertens J., L. Beladjal, F. Janssens, P. Matthys. 2007. Pitfall trapping in flooding habitats: a new technique reveals Archisotoma pulchella (Collembola: Isotomidae) as new to the Belgian fauna. Belg. J. Zool., 137(2): 177-181.
46. Naranjo S. E. 2009. Impacts of Bt crops on non-target invertebrates and insecticide use patterns. http://www.cababstractsplus.org/cabreviews
47. Naranjo S. E., G. Head, G. P. Dively. 2005. Field studies assessing arthropod nontarget effects in Bt transgenic crops. Environmental Entomology, 34(5): 1178-1180.
48. O’Callaghan M., T. R. Glare, E. P. J. Burgess, L. A. Malone. 2005. Effects of plants genetically modified for insect resistance on nontarget organisms. Annu. Rev. Entomol., 50: 271-292.
49. Odum E.P., 1971. Fundamentals of ecology. Third edition, W.B. Saunders Company. Philadelphia, London, Toronto.
50. OECD (1999a) Consensus document on general information concerning the genes and their enzymes that confer tolerance to phosphinothricin herbicide. Organisation for Economic Co-operation and Development, ENV/JM/MONO(99)13.
51. OECD (2002a) Series on Harmonisation of Regulatory Oversight in Biotechnology, No. 25. Module II: Phosphinothricin. Organisation for Economic Co-operation and Development, ENV/JM/MONO(2002)14.
52. OECD, 2007. Consensus Document on the biology of Zea mays subsp. Mays (maize). Series on Harmonisation of Regulatory Oversight in Biotechnology (ENV/JM/MONO(2007)14), No. 42: 1-10, http://www.oecd.org/dataoecd/36/61/46815888.pdf
53. Orr D.B., Landis D.A., 1997. Oviposition of european corn born borer (Lepidoptera: Pyralidae) and impact of natural enemy populations in transgenic versus isogenic corn. Jour. Of Economic Entomology, 90: 905-909.
54. Palm C. J., K. K. Donegan and R. J. Sieddler, 1996. Persistence in soil of transgenic plant produced Bacillus thuringensis var. kurstaki δ-endotoxin. Can. J. Microbiol., 42: 1258-1262.

201
55. Pilcher C. D., M. E. Rice, J. J. Obrycki. 2005. Impact of transgenic Bacillus thuringiensis corn and crop phenology on five nontarget arthropods. Environmental Entomology, 34(5): 1302-1316.
56. Powell R. Jeff, D. J. Levy-Booth, R. H. Gulden, W. L. Asbil, R. G. Campbell, K. E. Dunfield, A. S. Hamill, M. M. Hart, S. Lerat, R. E. Nurse, K. P. Pauls, P. H. Sikkema, C. J. Swanton, J. T. Trevors, J. N. Klironomos. 2009. Effects of genetically modified, herbicide-tolerant crops and their management on soil food web properties and crop litter decomposition. Journal of Applied Ecology, 46: 388-396.
57. Priestley AL., Brownbridge M. 2009. Field trials to evaluate effects of Bt-transgenic silage corn expressing the Cry1Ab insecticidal toxin on non-target soil arthropods in northern New England, USA. Transgenic Res., 18(3): 425-443.
58. Prieto-Samsónov DL, Vázquez-Padrón RI, Ayra-Pardo C, González-Cabrera J, de la Riva GA (1997) Bacillus thuringiensis: from biodiversity to biotechnology. Journal of Industrial Microbiology & Biotechnology 19: 202-219
59. Romeis J., M. Battini, F. Bigler. 2003. Transgenic wheat with enhanced fungal resistance causes no effects on Folsomia candida (Collembola: Isotomidae). Pedobiologia, 47(2): 141-147.
60. Rosel R., G. P. Dively. 2007. Effects of insecticide- treated and Lepidopteran- active Bt transgenic sweet corn on the abundance and diversity of arthropods. Environmental Entomology, 36(5): 1254-1268.
61. Saxena D. and G. Stotzky, 2000. Insecticidal toxin from Bacillus thuringensis is released from roots of transgenic Bt corn in vitro and in situ. FEMS Microbiol. Ecol., 33: 35-39.
62. Saxena D., S. Flores and G. Stotzky, 1999. Insecticidal toxin in root exudates from Bt corn. Nature (Lond.): 402-480.
63. Sayaboc A. S.,R. S. Raros and L. C. Raros,1975. The abundance of predatory and saprophagous acarines associated with decomposing rice stubble with a consideration of the effects of insecticide residues. Philippine Entomologisst, 2: 375-383.
64. Siegel JP (2001) The Mammalian Safety of Bacillus thuringiensis- Based Insecticides. Journal of Invertebrate Pathology 77: 13-21
65. Sims S. R. and Holden L. R., 1996. Insect bioassay for determining soil degradation of Bacillus thuringensis subsp. kurstaki Cry1a(b) protein in corn tissue. Environ. Entomol., 25: 659-664.

202
66. Sims S. R. and J. W. Martin, 1997. Effect of Bacillus thuringensis insecticidal proteins Cryp1Ab, Cry1Ac, Cry2A and Cry3a on Folsomia candida and Xenylla grisea (Insecta: Collembola), Pedobiologia, 41: 412-416.
67. Stach J., 1965. On some Collembola of north Vietnam. Acta. Zoo. Cracoviensia, T.X. No 4: 345-372.
68. Sterzynska Maria, 1990. Fragmenta faunistica, Tom 34(11), Warszawska Drukarnia Naukowa: 262p
69. Straalen Nico M. van, 2002. Assessment of soil contamination – a functional perspective. Biogdegradation, 13: 41-52.
70. Tapp H. and G. Stotzky, 1998. Persistence of the insecticidal toxin from Bacillus thuringensis subsp. Kurstaki in soil. Soil Biol. Biochem., 30: 471-476.
71. Wandeler H., Bahylova J. Nebtwig W., 2002. Comsumption of two Bt and six non-Bt corn varieties by the woodlouse Porcellio scaber. Basic Applied Ecology, 3:357-365.
72. Yu L., Berry RE, Croft BA, 1997. Effects of Bacillus thuringensis toxins in transgenic cotton and potato on Folsomia candida (Collembola: Isotomidae) and Oppia nitens (Acari: Oribatidae). J. Econ. Entomol., 90: 113-118.
73. Zimmer M., Topp W., 2000. Species-specific ultilization of food sources by sympatric woodlice (Isopoda: Oniscidea). Journal of Animal Ecology, 69: 1071-1082.

203
Tài liệu tham khảo tiếng Việt
1. Dương Thị Thanh, Nguyễn Trí Tiến, Nguyễn Thị Thu Anh, 2010. Khu hệ Collembola (Insecta) ở một số đô thị vùng đồng bằng sông Hồng. Tạp chí Công nghệ sinh học 8(3A): 1021-1030.
2. Nguyễn Thị Thu Anh, Nguyễn Trí Tiến, 2000. Nghiên cứu ảnh hưởng của phân hữu cơ vi sinh đến nhóm bọ nhảy (Insecta: Collembola) ở đất chuyên canh rau xã Gia Xuyên, huyện Gia Lộc, Hải Dương. Kỷ yếu HNCTH TQ lần thứ 6. Nxb Nông nghiệp, Hà Nội: 447-454.
3. Nguyễn Thị Thu Anh, Nguyễn Trí Tiến, 2009. Nghiên cứu bọ nhảy (Collembola) ở đất ô nhiễm chì tại Văn Lâm, Hưng Yên. Kỷ yếu HNKHTQ lần thứ 3 về Sinh thái và Tài nguyên sinh vật. Nxb Nông nghiệp, Hà Nội: 1168-1173.
4. Nguyễn Thị Thu Anh, Nguyễn Trí Tiến, Lê Thị Hoa, 2008. Ảnh hưởng của hiệu lực bón kali khác nhau đến một số định lượng của Collembola ở đất trồng màu huyện Gia Lâm, Hà Nội. Kỷ yếu HNCTH TQ lần thứ 6. Nxb Nông nghiệp, Hà Nội: 440-446.
5. Nguyễn Thị Thu Anh, Nguyễn Trí Tiến, Mai Phú Quý, Lê Quốc Doanh, 2005. Đặc điểm cư trú của bọ nhảy (Collembola) ở hệ sinh thái nông nghiệp trên đất dốc miền núi phía Bắc Việt Nam. Hội nghị môi trường toàn quốc lần thứ 5. Nxb Nông nghiệp, Hà Nội: 279-284.
6. Nguyễn Thị Thu Anh, Nguyễn Trí Tiến, Phan Thị Thu Hiền, 2008. Nghiên cứu ảnh hưởng của một số liều lượng bón phân lân đến động vật chân khớp bé ở ruộng trồng lạc huyện Gia Lâm, Hà Nội. Kỷ yếu HNCTH TQ lần thứ 6. Nxb Nông nghiệp, Hà Nội: 432-439
7. Nguyễn Trí Tiến, Nguyễn Thị Bích Ngọc, Thái Trần Bái, 2009. Khu hệ bọ nhảy (Insecta: Collembola) ở khu bảo tồn thiên nhiên Thượng Tiến, huyện Kim Bôi, tỉnh Hòa Bình. Kỷ yếu HNKHTQ lần thứ 3 về Sinh thái và Tài nguyên sinh vật. Nxb Nông nghiệp, Hà Nội: 847-852
8. Nguyễn Trí Tiến, Nguyễn Thị Định, Nguyễn Thị Thu Anh, Nhữ Thị Hoài, 2009. Nghiên cứu ảnh hưởng của phân hữu cơ và kỹ thuật vùi phụ phẩm nông nghiệp trên đất trồng mía đến Collembola (Insecta) ở nông trường Hà Trung, Thanh Hóa. Kỷ yếu HNKHTQ lần thứ 3 về Sinh thái và Tài nguyên sinh vật. Nxb Nông nghiệp, Hà Nội: 1705-1710.
9. Nguyễn Trí Tiến, Nguyễn Văn Quảng, Lê Thị Quyên, 2008. Khu hệ Collembola vườn quốc gia Xuân Sơn. Kỷ yếu HNTQ 2007, nghiên cứu cơ bản trong khoa học sự sống. Nxb Khoa học và Kỹ thuật, Hà Nội: 195-198.

204
10. Phạm Văn Lầm, 2002. Tài nguyên thiên địch của sâu hại: nghiên cứu và ứng dụng. Quyển I. Nxb Nông nghiệp, Hà Nội, 7-57.
11. Phạm Văn Lầm, 2006. Nhện lớn bắt mồi trên đồng lúa Việt Nam. Nxb Nông nghiệp, Hà Nội.
12. Phạm Văn Lầm, Bùi Hải Sơn, Trần Thị Hường, 1993. Nhận xét bước đầu bước đầu về diễn biến số lượng của một số loài ăn thịt rầy nâu. Tạp chí Bảo vệ thực vật, 2: 30-34.
13. Phạm Văn Lầm, Nguyễn Kim Hoa, Trương Thị Lan, 2003. Nhện lớn bắt mồi trên ruộng lúa và yếu tố ảnh hưởng đến số lượng của chúng. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống . Báo cáo Khoa học Hội nghị toàn quốc lần thứ hai, Huế 25-26/7/2003. Nxb Khoa học và Kĩ thuật, Hà Néộ, 646 -649.