Embed Size (px)
Citation preview

CAR
VEG
RÍO
TESIS
PARA
Xalapa
RTOGRA
GETACIÓ
CITLALA
S QUE PR
A OBTEN
a, Veracruz,
AFÍA DE L
ÓN-AMB
APA, HU
RESENTA
ER EL GR
, México 20
LA VEGE
IENTE EN
UATUSC
CARLA F
RADO DE
011
ETACIÓN
N LA MI
CO, VERA
LORES LO
E MAESTR
N Y RELA
ICROCU
ACRUZ
OT
RO EN CI
ACIONE
UENCA D
ENCIAS
ES
DEL


AGRADECIMIENTOS Al Consejo Nacional de Ciencia y Tecnología que me otorgó una beca de estudios con número de registro A la División de Posgrado del INECOL AC, por otorgarme beca durante el primer semestre de mis estudios. Al Centro “Las Cañadas”, Ricardo Romero y todo el personal, por brindarnos alojamiento, acceso e información de los predios aledaños, y por su interés en generar conocimiento. A Carlos Montaña Carubelli por asesorarme en este trabajo e impulsar mi formación. A Fabiola López Barrera por las valiosas aportaciones y constante apoyo. A Neptalí Ramírez Marcial, Lorena Soto Pinto y José G. García Franco por sus evaluaciones y comentarios que solidificaron el trabajo. A Claudia Gallardo, Francisco Lorea, Gonzalo Castillo, Carlos Durán, Susana Valencia y Lucio Lozada, por su especial ayuda en la identificación del material biológico. A Vania Jiménez Lobato, Luis M. García Feria, Claudia Gallardo y Julieta Flores por su divertido apoyo en el trabajo de campo. A Nicasio y Eliseo por su guía hacia los fragmentos de bosque y su compartir. A los dueños de los predios por permitirnos el acceso.

A tod@s los que motivaron, participaron, digirieron, esperaron y abrazaron este trabajo.


6
Contenido
LISTA DE TABLAS .............................................................................................. 8
LISTA DE FIGURAS ........................................................................................... 8
RESUMEN ........................................................................................................ 11
1. INTRODUCCIÓN .......................................................................................... 13
1.1 Bosques: pérdida y las estrategias para su conservación y restauración. ..... 13
1.2 Características del Bosque Mesófilo de Montaña (BMM). ............................... 15
1.2.1 El BMM en México .................................................................................... 17
1.2.2 Relación vegetación-ambiente .................................................................. 22
1.3 Restauración y uso sustentable del BMM en la región central de Veracruz. ... 24
OBJETIVO GENERAL ...................................................................................... 26
OBJETIVOS PARTICULARES .......................................................................... 26
2. METODOLOGIA ............................................................................................ 26
2.1 Área de estudio ............................................................................................... 27
2.2 Características ................................................................................................ 28
2.3 Cartografía ...................................................................................................... 28
2.4 Toma de datos en campo ................................................................................ 29
2.5 Análisis de datos ............................................................................................. 33
2.5.1 Valor de Importancia relativa .................................................................... 33
2.5.2 Riqueza de especies ................................................................................. 34
2.5.3 Comparación de la riqueza de especies en bosques y cafetales. ............ 38

7
2.5.4 Suelo ......................................................................................................... 39
2.6 Relaciones Vegetación-Ambiente ................................................................... 39
3. RESULTADOS .............................................................................................. 40
3.1 Uso del suelo ........................................................................................ 40
3.2 Vegetación ............................................................................................ 42
3.3 Diversidad ............................................................................................. 46
3.3.1 Comparación de riquezas entre bosques y cafetales. .................... 52
3.3.2 Comparación de CAE por individuos y por transectos. .................. 52
3.3.3 Índice de Shannon ......................................................................... 52
3.4 Análisis Canónico de Correspondencias .............................................. 53
4. DISCUSIÓN .................................................................................................. 59
4.1 Uso de suelo ................................................................................................... 59
4.2 Florística .......................................................................................................... 61
4.3 Diversidad ....................................................................................................... 65
4.4 Análisis Canónico de Correspondencias ......................................................... 67
5. CONCLUSIONES ....................................................................................... ...68
A P É N D I C E ................................................................................................. 70
LISTADO DE ESPECIES LEÑOSAS ................................................................ 71
6. BIBLIOGRAFÍA ............................................................................................. 75

8
Lista de Tablas
Tabla 1. Parámetros tomados en cada uno de los sitios de muestreo dentro de la
microcuenca del Río Citlalapa……………………………………………………….....31
Tabla 2. Áreas de cada tipo de uso de suelo presente en la microcuenca ……….40
Tabla 3. Número de especies e individuos por forma de vida y procedencia .......44
Tabla 4. Proporción de formas de vida y procedencia ……………………………...44
Tabla 5. Especies nativas compartidas ..……………………………………………. 45
Tabla 6. Datos de riqueza (número de especies) de la vegetación estudiada .....46
Tabla 7. Resultados de riqueza esperada con estimadores no paramétricos .…..47
Tabla 8. Resumen de valores obtenidos en el análisis CCA ………………………55
Tabla 9. Correlación obtenida entre los ejes canónicos y las variables ambientales
consideradas …………………………………………………………………………….55
Tabla 10. Coeficientes de correlación obtenida entre las variables ……………....56
Lista de Figuras Figura 1. Distribución mundial del Bosque Mesófilo de montaña (polígonos verdes,
tomado de UNEP-WCMC 2004) ………………………………………………………16
Figura 2. Distribución de los fragmentos de Bosque Mesófilo de Montaña en
México (Rzedowski, 1996: Challenger, 1998) …...…..………………………………18
Figura 3. Ortofoto de 1995 de la región Huatusco con la delimitación de la
Microcuenca Citlalapa (INEGI) ………………………………………………………...29

9
Figura 4. Ilustración sobre el registro de coberturas ...……………………………..32
Figura 5. Mapa de uso de suelo basado en una foto aérea de 1995 …………….41
Figura 6. Relación de géneros y especies por familia (a) y de individuos por familia
(b) …..……………………………………………………………………………………..43
Figura 7.Especies exclusivas de bosques y cafetales; y compartidas entre los tipos
de vegetación ……………………………………………………………………………45
Figura 8. Relación entre el número de especies y el número de individuos ……..47
Figura 9.CAE de todos los sitios, a) utilizando el estimador Mao Tau con los
intervalos de confianza, b) basadas en estimadores incluyendo especies no
detectadas, singltones y doubletones …………………………....…………………...49
Figura 10. CAE para bosques, a) utilizando el estimador Mao Tau con los
intervalos de confianza, b) con base en estimadores incluyendo especies no
detectadas, singletones y doubletones ………………………………………………50
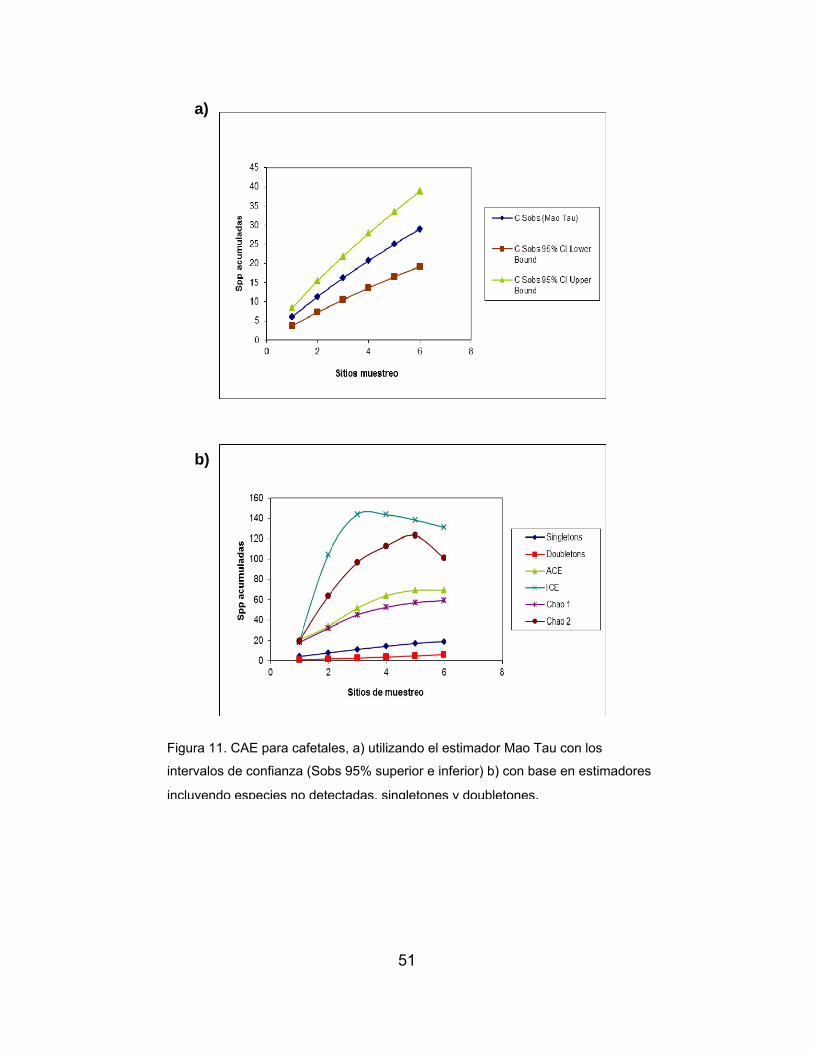
Figura 11. CAE para cafetales, a) utilizando el estimador Mao Tau con los
intervalos de confianza, b) con base en estimadores incluyendo especies no
detectadas, singltones y doubletones ……….……………………………………….51
Figura 12 CAE diferenciando bosques y cafetales a) dimensionando el esfuerzo de
muestreo con individuos y b) dimensionando el esfuerzo de muestreo por
transectos ..………………………………………………………………………………54
Figura 13. CCA de datos florísticos en bosques (diamantes) y cafetales (círculos),
incluyendo variables ambientales de suelo (pH, CE, N, P, C orgánico, arena,
arcilla, limo) de condiciones forestales (área basal total, cobertura, riqueza),
pendiente y altitud ……………………………………..……………………………….57

10
Figura 14. CCA de especies leñosas (árboles y arbustos), incluyendo variables
ambientales de suelo (pH, CE, N, P, C org, arena, arcilla, limo) de condiciones
forestales (Área basal total, cobertura, riqueza), pendiente y altitud ……………...58

11
RESUMEN
La deforestación es una de las principales causas de la pérdida de biodiversidad, siendo
el Bosque Mésofilo de Montaña (BMM) uno de los ecosistemas más afectados a nivel
mundial. En el estado de Veracruz el BMM se encuentra altamente fragmentado e
inmerso en un mosaico agroforestal por el constante cambio de uso de suelo (Williams-
Linera 2007). En el municipio de Huatusco se ha implementado como estrategia de
conservación la “servidumbre ecológica” en el predio “Las Cañadas” inmerso en la
microcuenca del río Citlalapa, lo cual ha propiciado la integración comunitaria para
realizar proyectos de restauración. En este estudio se analizó la estructura y diversidad de
la vegetación leñosa de la microcuenca, así como su relación con variables ambientales;
esto con el fin de impulsar una planificación adecuada y con información de base en la
restauración ecológica a nivel local.
Se generó un mapa de uso de suelo, con base en ortofotos de 1995, en el que se observó
que el 52 % está ocupado por vegetación boscosa, del cual 26% son cafetales con
sombra inducida o rústica y sólo el 26% de la vegetación boscosa permanece sin
intervención de manejo, la cual en su mayoría es vegetación secundaria. Se muestreó un
total de 22 sitios, de los cuales 6 son cafetales.
Se registró un total de 158 especies, correspondientes a 107 géneros y 49 familias. El
64% de las especies fueron árboles y el 88% son nativas. Sólo 12 especies se comparten
entre bosques y cafetales, de las cuales 7 son árboles nativos y su abundancia y forma de
crecimiento permite recomendarlas para estudios explotarios de restauración en la zona:
Buddleja americana, Clethra macrophylla, Quercus lancifolia, Quercus sapotifolia,
Quercus sartorii, Quercus xalapensis, Trema micrantha. La especie con el mayor valor de
importancia relativa fue Palicourea padifolia (21.29%), característica de este tipo de
bosque en etapa sucesional secundaria. Se observó poca invasión de especies leñosas
exóticas en bosques (9%), aunque en su mayoría estos son bosques con especies
pioneras o secundarias, lo que indica intervención, abandono y un estado de recuperación
en algunos de ellos.
La mayoría de las especies (95) están poco representadas en ambos tipos de vegetación,
es decir por sólo uno o dos individuos, y sólo 19 mostraron una abundancia ≥ 10
individuos, indicando un gran recambio de especies (diversidad β), estructura típica del
BMM. La riqueza total de vegetación leñosa en bosques es mayor (p<0.05) en bosques
que en cafetales. En el CCA se observó una alta correlación entre especies y ambiente,

12
explicando con 2 ejes el 97% de la varianza acumulada. La estructura de los bosques
estuvo asociada positivamente al contenido de Carbono, Nitrógeno, conductividad
eléctrica, cobertura del dosel y la pendiente, mientras que la de los cafetales se asoció
negativamente con el contenido de arcilla y fósforo. Las especies asociadas con los
cafetales son Inga sp., Psidium guajava, Trema micrantha, Quercus xalapensis y Clethra
macrophylla, las tres últimas son especies nativas utilizadas para la sombra del sistema
agroforestal. Los bosques conservados en la microcuenca ya solo se encuentran
asociados a pendientes pronunciadas y de difícil acceso. Se discuten las implicaciones de
los resultados en términos de manejo, conservación y restauración.

13
1. INTRODUCCIÓN
1.1 Bosques: pérdida y las estrategias para su conservación y restauración.
En los últimos años se ha observado una acelerada pérdida de la biodiversidad
mundial. Una de las principales causas es la destrucción de áreas boscosas
debidas a la deforestación (Myers et al. 2000, Lambin et al. 2001) cuya tasa anual
se estima en 0.18%, lo que corresponde a la pérdida de 7.3 millones de hectáreas
de áreas boscosas por año (FAO 2007). Las causas principales son el cambio de
uso de suelo y el mal manejo de los recursos, como la transformación de bosques
a cultivos y potreros en respuesta a una creciente presión demográfica y al mal
manejo y aprovechamiento de los recursos (Aguilar et al. 2000).
Este alto índice en la destrucción de bosques no sólo conlleva a la disminución de
cubierta vegetal, sino que tiene una serie de consecuencias negativas ambientales
y por ende afecta el bienestar social. La erosión del suelo es una de ellas, pues la
pérdida de cobertura vegetal incentiva la pérdida de material edáfico por viento y
agua con la consecuente desestabilización de la dinámica biogeoquímica de los
distintos horizontes edáficos (Pimentel y Kounang 1998). Esto provoca cambios
físico-químicos que resultan tanto en una disminución de la oferta de nutrientes
edáficos como en un incremento en los deslizamientos de laderas desnudas,
pérdida de material del suelo y contaminación de arroyos y ríos, lo que a su vez

14
destruye el hábitat de organismos tanto terrestres como de agua dulce (Hajabbasi
et al. 1997).
Uno de los ecosistemas altamente amenazados y con la tasa de deforestación
más alta de los bosques tropicales es el Bosque Mesófilo de Montaña (BMM) o
Bosque de Niebla, cuya tasa de pérdida anual se estima en 1.1 % (Doumenge et
al. 1995, Hamilton 1995, Aldrich y Hostettler 2000). En 1974, Persson reportó que
este ecosistema ocupaba 500 mil Km2 (Bruijnzeel 2004), mientras que en un
reporte más reciente se considera que potencialmente ocupa un área de 380 mil
Km2 (Bubb et al. 2004). Con base en estas cifras, se infiere que se han perdido
120 mil Km2 (24%) de estos bosques en 36 años, los cuales solo ocupan el 37%
del área boscosa global. Aunado a esto, el BMM es un ecosistema muy
vulnerable, ya que se reporta una baja tasa de migración como consecuencia de
lentos procesos de crecimiento y maduración, lo que a su vez se refleja en una
lenta recuperación y baja competencia con especies invasoras (Foster 2001); su
distribución es altamente fragmentada y limitada por los factores microambientales
en los que se desarrolla, donde por lo general existen barreras físicas (ríos,
caminos, cultivos) que impiden su expansión (Bush et al. 2004). Por ello se
reconoce como uno de los ecosistemas más amenazados a nivel mundial (Bubb et
al. 2004)
Es por esto que se han generado iniciativas de conservación y restauración de
este ecosistema, para lo cual es necesario contar con la información base sobre la
estructura y composición florística de las zonas boscosas que permita elaborar

15
estrategias para su manejo y desacelerar así la pérdida de masas forestales que
se registra actualmente.
1.2 Características del Bosque Mesófilo de Montaña (BMM).
El Bosque Mesófilo de Montaña (BMM), también conocido como Bosque de
Niebla, se caracteriza por ser un ecosistema con una alta humedad atmosférica
todo el año. Se ubica en laderas montañosas ubicadas a barlovento de zonas
marinas donde el aire cargado de humedad es obligado a ascender por las laderas
provocando el enfriamiento y la consecuente condensación de humedad y la
formación de nubes bajas y neblina (Hamilton 1995). El rango altitudinal de su
distribución en tierra adentro va de 2000 a 3500 msnm, mientras que en zonas
insulares o de costa se puede encontrar entre 500 y 1500 msnm dependiendo de
las condiciones de humedad. Se ha reportado que la precipitación anual presente
en estos ecosistemas varía entre 500 y 6000 mm. Como se mencionó
anteriormente el BMM ocupa aproximadamente 380 mil Km2 que apenas
alcanzan el 0.26% de la superficie terrestre (Bubb et al. 2004) (Fig. 1).

16
Figura 1. Distribución mundial del Bosque Mesófilo de montaña (polígonos verdes). (Tomado de
UNEP-WCMC 2004)
Un aspecto de gran importancia del BMM se refiere a los servicios ambientales
que provee, como son la captación de agua, el enriquecimiento del suelo, el
control de la erosión y deslaves, así como el secuestro de carbono (Williams-
Linera 2002, Manson 2004). Dentro de sus características biológicas está la
extraordinaria diversidad de helechos, plantas epífitas y musgos; en general existe
una alta diversidad de especies de animales, vegetales y hongos pues las
características topográficas donde se asientan son muy heterogéneas y los
factores micro-ambientales varían considerablemente. Otro de los atributos
reportados es que se presenta un gran recambio de especies entre estos bosques,
es decir, que la riqueza de especies entre sitios es poco compartida, lo que refleja
una alta diversidad β (Williams-Linera 2007). Esto último se traduce como
complementariedad entre sitios de bosque en un paisaje, lo que resulta de
consideración básica para cualquier propuesta de conservación o restauración.

17
1.2.1 El BMM en México
El BMM representa uno de los ecosistemas más diversos del país. Su riqueza se
cuenta en aproximadamente 3,000 especies vegetales – 10% de la flora de
México - en poco menos del 1% del territorio nacional, es decir florísticamente es
el más diverso por unidad de área (Fig. 2; Rzedowski 1978, Challenger 1998,
(Toledo-Aceves et al. 2011). Se estima que alrededor de 750 especies vegetales
son endémicas. La composición de especies del BMM es muy variable incluso
entre laderas adyacentes. Las familias de plantas leñosas más comunes son
Compositae, Rubiaceae, Melastomataceae, Leguminosae y Lauraceae. También
existen géneros muy característicos como Liquidambar, Quercus, Clethra, Alnus,
Prunus, Saurauia, Palicourea, Miconia, y Eugenia (Rzedowski 1978).
En Veracruz, como en todo México, la composición de la vegetación del BMM es
el resultado de la confluencia de especies con origen tanto neotropical - en su
mayoría andinas - como templado, que migraron luego de la formación del puente
terrestre centroamericano (Gentry 1982). Entre las especies neotropicales
reportadas se encuentran árboles como Zanthoxylum spp. (subtropical), Turpinia
insignis (tropical), Cornus disciflora (subtropical) y Eugenia spp (tropical) y los
arbustos Miconia glaberrima, Palicourea padifolia, Eugenia xalapensis y Ocotea
psychotrioides. Las especies templadas comunes a estos bosques son
principalmente arbóreas como Juglans pyriformis (templado), Podocarpus
matudae (templado), Fagus mexicana (templado), Carpinus sp, Fagus sp,

18
Liquidambar styraciflua y Quercus sp, (Williams-Linera 2007, Garcia-Franco et al.
2008).
Figura 2. Distribución de los fragmentos de Bosque Mesófilo de Montaña (polígonos negros) en
México (Rzedowski, 1996: Challenger, 1998).
En el caso de la fauna también se ha reportado que la diversidad de vertebrados
es alta así como sus niveles de endemismo (Flores-Villela y Gerez 1994)
especialmente en el caso de los mamíferos que alcanzan la mayor riqueza en este
tipo de ecosistemas (Fa y Morales 1993, Challenger 1998). El BMM contiene 39%
de reptiles, 33% de anfibios, 71% de aves y 53% de los mamíferos del país
(Flores-Villela y Gerez 1988, Toledo y Ordoñez 1993, Challenger 1998, Williams-
Linera 2002).
Sin embargo, en México se estima que se ha perdido más del 70% del área
original cubierta por este ecosistema, de la restante sólo el 47.6% se considera
bosque conservado, y se estima una tasa de 1.5 % de pérdida anual (FAO 2007).

19
Las principales conversiones se deben a la ganadería extensiva, el cultivo del
café, el desarrollo urbano y la extracción selectiva para construcción y leña. Esta
última es considerada una de las causas mayores de la degradación,
principalmente en el modo de acarreo hormiga (Toledo-Aceves 2010).
El 3.7 % de la superficie del estado de Veracruz corresponde a este tipo de
ecosistema, pero el cambio de uso de suelo ha disminuido su área
considerablemente (Flores-Villela y Gerez 1988, Dirzo y García 1992). Aun así se
reporta que la zona montañosa ocupa el 11.6% de la superficie del estado
(Williams-Linera, 2002).
La composición de especies del BMM es muy variable incluso entre laderas
adyacentes lo que estimula la existencia de diversos estudios florísticos que
informan sobre la riqueza de algunos fragmentos en México principalmente en los
estados de Hidalgo, Puebla, Oaxaca.
Los estudios florísticos sobre BMM en México se orientan principalmente a la
elaboración de listados (Soto-Pinto et al. 2001, Cartujano et al. 2002) o al estudio
de las relaciones entre provincias biogeográficas (Luna-Vega et al. 1999). Por otro
lado existen aquéllos cuyo objetivo es obtener información sobre la autoecología
de algunas especies y otros aspectos de ecología funcional, que permitan elaborar
estrategias de conservación y manejo locales (Ramírez-Marcial 2001, Ramírez-
Marcial et al. 2001, Pedraza and Williams-Linera 2003, Ramírez-Marcial et al.
2005, Muñiz-Castro et al. 2006).
En Veracruz ha predominado el enfoque ecológico, informando sobre su fenología,
diversidad y riqueza específica, relación con agro sistemas, la presencia de

20
especies indicadoras, su estado de conservación y de fragmentación, así como de
los servicios ambientales que provee como es la captación de agua entre otros
(Pedraza y Williams-Linera 2003, Alvarez-Aquino et al. 2004, López-Gómez 2004,
Pineda et al. 2005, Muñiz-Castro et al. 2006, Manson et al. 2008, Martínez et al.
2009). Recientemente han resurgido los estudios florísticos, pero aún existen
fragmentos carentes de información local (Garcia-Franco et al. 2008). Otros más
tratan sobre la conservación en áreas con manejo, como los agrosistemas de café
(Romero-Alvarado et al. 2002, Harvey et al. 2008).
La gran heterogeneidad en la composición florística y la abundancia de especies
raras (Hartshorn 2002, Williams-Linera et al. 2005) inducen la necesidad de
realizar análisis a nivel regional (cuencas o microcuencas) para identificar
aquellos remanentes prioritarios para la conservación. Diversos estudios de
manejo y restauración de bosques han coincidido en que caracterizar la
vegetación y relacionarla con variables ambientales resulta indispensable para
implementar una planificación adecuada en su rehabilitación biótica y abiótica
(Palmer et al. 1997, Finegan y Delgado 2000, Temperton 2004, Palmer et al. 2005,
Young et al. 2005).
Los remanentes de BMM en Veracruz están inmersos en mosaicos agroforestales
(Williams-Linera 2002, Guerrero y Equihua 2005), generando un paisaje regional
con una configuración propia tanto estructural como funcional (Palik et al. 2000,
López-Gómez 2004). De hecho, el estado de deterioro del BMM es tal, que las
áreas conservadas están reducidas a zonas de cañadas y laderas escarpadas, es
decir, áreas de difícil acceso (Cartujano et al. 2002). Este estado relictual

21
magnifica la importancia de conocer el grado de fragmentación y los tipos de uso
de suelo y vegetación circundante de las áreas con BMM del estado, con el cual
se pueden reconocer zonas prioritarias y vegetación potencial para estrategias de
recuperación del ecosistema.
El paisaje actual del área que originalmente ocupaba el BMM corresponde a un
paisaje agroforestal heterogéneo con fragmentos de BMM, incrustados en una
matriz de cultivos de subsistencia, pastizales, matorrales, cafetales y acahuales.
El cultivo del café es uno de los sistemas productivos predominantes en el centro
del estado, lo que ha encauzado a realizar estudios sobre su impacto en la
biodiversidad local, en los cuales se han identificado sistemas tradicionales menos
agresivos e incluso definidos como reservorios de la misma. Según la clasificación
reportada en Manson et al (2008), los cafetales rústicos son aquéllos con menor
impacto ecológico y se caracterizan por presentar una cobertura de al menos el
60%, árboles grandes, de los cuales no se pretende obtener algún
aprovechamiento y con más de un metro de diámetro, y una composición rica en
epífitas. En cambio en un policultivo se presentan árboles de los cuales se busca
un beneficio además de la sombra, que generalmente son exóticos pero generan
gran sombra y hábitat para epífitas.
El BMM en Veracruz se reporta en un clima templado húmedo con lluvias en
verano y una precipitación anual de entre 1500 y 2000 mm, sustentado por suelos
andosoles y luvisoles, lo cual coincide con las características de la zona y lo
reportado en descripciones de este tipo de bosque (Rzedowsky 1978, Challenger
1998, Williams-Linera 2007, www.inegi.gob.mx)

22
Los estudios sobre florística y diversidad de los BMM reportan una riqueza
florística de 60 a 78 especies de árboles y de 24 hasta 63 especies de arbustos
(Ramírez-Marcial 2001, Williams-Linera 2002, Ponce-Vargas et al. 2006) y
destacan una alta diversidad beta y complementariedad entre fragmentos de este
bosque (Williams-Linera 2002, Williams-Linera et al. 2005). Esos estudios han
empleado los índices paramétricos de Shannon o Simpson (Magurran 1988) para
medir la diversidad alfa. El índice de Shannon toma en cuenta no sólo el número
de especies sin su frecuencia, mientras que el índice de Simpson pondera la
dominancia de las especies, es decir, sobrevalora las especies más abundantes
en detrimento de la riqueza total. Sin embargo la utilidad de estos índices depende
tanto del número de individuos muestreados, como del área muestreada (Chazdon
et al. 1998). Para solucionar este problema se han desarrollado métodos que
estiman la riqueza de especies a través de la construcción de Curvas de
Acumulación de Especies (CAE) que son insensibles al tamaño y orden de
muestreo así como a la distribución de las especies (Chazdon et al. 1998) los
cuales se decidió utilizar en este estudio debido a la alta fragmentación de la zona.
1.2.2 Relación vegetación-ambiente
El BMM constituye un mosaico de comunidades vegetales que, debido a la
accidentada topografía, están estrechamente relacionadas con las características
ambientales del sitio donde se encuentren. Diversos autores han mencionado la
importancia de los factores edáficos en la organización de las comunidades
(Montaña 1990, Montaña y Greig-Smith1990, Valverde et al. 1996, Cruz-Ruggiero

23
et al. 2002, Nadkarni et al. 2002, Bautista-Cruz y Del Castillo 2005). El suelo en el
BMM se caracteriza por ser muy húmedo todo el año y presentar un alto contenido
de materia orgánica en los horizontes superiores (Hamilton 1995). En general se
describen como suelos ácidos (pH 4-6), someros o profundos, de rojizos a
negruzcos y de arenosos a arcillosos (Challenger 2003).
Se ha reportado que el desmonte de estos bosques para cultivo y su subsecuente
abandono por escasez de fertilidad, altera las propiedades del suelo, aun siendo
repoblado por vegetación secundaria. Mientras se asocia acidez y materia
orgánica con bosques maduros, se considera que la textura es una de las
propiedades del suelo que no cambia por el manejo del mismo; pero cuando la
deforestación es severa, se pueden eliminar horizontes y aflorar materiales
arcillosos (Bautista-Cruz et al. 2005).
En la región de Veracruz los suelos son derivados del vulcanismo (andosoles) que
retienen un alto porcentaje de humedad por los minerales alófanos que contienen,
de color pardo-oscuros y porosos, y tienen un alto contenido de materia orgánica,
pero con relativamente baja disponibilidad de nutrientes para las plantas (Geissert
e Ibañez 2008), sin embargo en esta región no existe un estudio que relacione
estas características con la vegetación existente.
Los diversos contextos de deforestación, condición relictual y alta heterogeneidad
entre comunidades que definen al BMM actualmente, provocan la necesidad de
caracterizar los fragmentos locales con elementos clave como son las variables
edáficas, la ubicación y estructura que forjen criterios argumentados para su
conservación y manejo.

24
1.3 Restauración y uso sustentable del BMM en la región central de Veracruz.
Existen estudios de vegetación y distribución espacial, en el centro del estado de
Veracruz, que reflejan la gran reducción y deterioro de fragmentos de bosque
mesófilo (Muñoz-Villers y López-Blanco2008), por lo que es necesario diseñar
estrategias de conservación, manejo y recuperación de éstos. Dentro de las
estrategias de conservación y restauración de estos fragmentos se pueden
considerar la protección oficial, la restauración del paisaje con corredores
biológicos, y el pago a los propietarios por preservar dichos fragmentos. En tales
esquemas surge el programa de “Pago por Servicios Ambientales” (PSA), el cual
se define como “una clase emergente de proyectos de desarrollo sustentable que
encuentran su sentido en la valoración económica de los recursos naturales y la
biodiversidad” (Burstein et al. 2002). Esto permite la aplicación de estrategias de
conservación y restauración de ecosistemas bajo un esquema que mantenga la
productividad local, nombrando “servidumbres ecológicas” a las zonas
conservadas que se encuentran bajo un plan de protección integral.
Una servidumbre ecológica es un contrato entre al menos dos propietarios, en
virtud del cual, uno o los dos limitan voluntariamente los usos o intensidad de
desarrollo sobre sus terrenos, con el objeto de conservar sus atributos naturales o
culturales. La principal ventaja es que la propiedad se mantiene en manos
privadas, los términos son flexibles y el período puede ser desde 10 años o hasta
la perpetuidad. Finalmente, si existen futuros propietarios están obligados a
mantener dicho contrato (Chacón y Castro 1998).

25
En el municipio de Huatusco, centro de Veracruz, se nombró en 1995 la primera
servidumbre ecológica en México en el predio “Las Cañadas”, el cual tiene una
extensión de 406 ha de las cuales 367 están destinadas como área de
conservación. Ésta última se subdivide en áreas de conservación absoluta,
amortiguamiento y recuperación, donde hay zonas acahualadas en regeneración,
zonas de reforestación, fragmentos de bosque maduro y corredores riparios
(Romero 2006).
Todo el municipio presenta un grave deterioro ecológico, por lo que es necesario
implementar proyectos de restauración y planes de manejo. Además, algunos
propietarios están dispuestos a realizar manejo sustentable de sus propiedades
pero necesitan información de base para tal fin.
Sin embargo antes de arrancar una propuesta de restauración es necesario
caracterizar la vegetación y su distribución en la microcuenca. González-Espinosa
et al (2004) comentan que el éxito de los programas de reforestación reposa
frecuentemente en la calidad del conocimiento biológico y ambiental de los sitios a
reforestar, ya que esto determina el número e identidad de las especies que se
usarán para obtener una reforestación exitosa.
En este contexto se pretende generar información local específica que consolide
la conservación de especies nativas con las propuestas de compensación ante la
pérdida de cobertura boscosa.

26
OBJETIVO GENERAL Generar información biológica de base que sirva como plataforma para
implementar proyectos de restauración ecológica de BMM en la microcuenca del
río Citlalapa; evidenciando la escasa presencia de fragmentos conservados así
como el potencial florístico nativo y microambiental.
OBJETIVOS PARTICULARES 1 – Elaborar la cartografía de los usos del suelo y determinar el área ocupada por
vegetación boscosa remanente.
2 - Evaluar la diversidad y estructura florística de las formaciones vegetales
leñosas en fragmentos boscosos.
3 - Analizar las relaciones vegetación-microambiente e identificar asociaciones
potenciales para la restauración.
2. METODOLOGIA
El procedimiento se compone de los siguientes pasos: 1) generación de un mapa
de uso de suelo y corroboración en campo, 2) muestreo de vegetación leñosa en
dos tipos de fragmentos forestales con diferencias en su cobertura (bosques y
cafetales) y toma de datos microambientales, 3) determinación de ejemplares y
descripción de la estructura florística por cada tipo de vegetación 4) cálculo de
índices de diversidad y curvas de acumulación de especies por tipo de vegetación

27
y 5) análisis de correspondencia entre especies y las variables microambientales
(CANOCO).
2.1 Área de estudio
Antiguamente la región del municipio de Huatusco estaba dominada por BMM
(Córdova-Santamaría 2005), sin embargo, las actividades agrícolas, la industria y
el consiguiente aumento en la población humana han modificado el paisaje
natural, implicando importantes cambios en el uso del suelo como es la forestación
con especies exóticas, la expansión de potreros por la explotación agrícola y
ganadera, la constante extracción de leña, tanto a nivel doméstico (“acarreo
hormiga”) como industrial y la creciente urbanización. Esto disminuyó
considerablemente el área ocupada por la vegetación original (Romero 2006). Las
áreas agrícolas y pecuarias son muy extensas y están dedicadas principalmente
al cultivo del café y el ganado, lo cual ha transformado el paisaje de bosque
original. Por ejemplo, Muñoz-Villers y López-Blanco (2008) reportan que en la
cuenca de La Antigua en la zona central de Veracruz, entre los años de 1990 y
2003, se perdió un 34% de la superficie de BMM y aumentó principalmente el área
de pastizal cultivado y de sistemas agroforestales. En la zona se encontraron
fragmentos boscosos que eran cafetales de sombra del tipo rústico, una mezcla de
éste con policultivos, y monocultivos con cuatro o cinco especies de árboles.

28
2.2 Características
El BMM en Veracruz se reporta en un clima templado húmedo con lluvias en
verano y una precipitación anual de entre 1500 y 2000 mm, sustentado por suelos
andosoles y luvisoles, lo cual coincide con las características de la zona y lo
reportado en descripciones de este tipo de bosque (Rzedowski 1978, Challenger
1998, Williams 2007, www.inegi.gob.mx)
El área de estudio está ubicada entre las longitudes 96° 55’ 47’’ W y 97° 02’ 19’’ W
y las latitudes 19° 8’ 57’’ N y 19° 12’ 42’’ N, y forma parte de la Sierra Madre
Oriental. La extensión de la microcuenca es de 4352 ha y su gradiente altitudinal
va de 1077 a 1868 msnm.
El tipo de suelo es principalmente luvisol (rojo y arcilloso) y andosol (derivado de
cenizas volcánicas y de color negro; (Rosas et al. 2008). El clima es templado-
húmedo-regular, con una temperatura media de 19º C y una precipitación media
anual de 1825 mm. Tiene lluvias abundantes en verano, a principios de otoño y
con menor intensidad en invierno (http://www.huatusco.gob.mx).
2.3 Cartografía
Con base en dos ortofotos de 1995 blanco y negro, que integran la zona de la
microcuenca del río Citlalapa (Fig. 3), se hizo una clasificación de usos de suelo
con el fin de reconocer la distribución de las áreas con cobertura arbórea densa, y
seleccionar los sitios de muestro de vegetación y suelo. Para esto se utilizó el
programa Arcview 3.2 por medio de clasificación no supervisada que define

29
grupos de clases espectrales homogéneas. Posteriormente se hizo una
comparación con padrones de cafetales existentes (SAGARPA), con lo que se
pudo separar claramente bosques de cafetales. Finalmente se visitó el área para
verificar la representatividad de la clasificación zonal.
Figura 3. Ortofoto de 1995 de la región Huatusco con la delimitación de la Microcuenca Citlalapa
(INEGI).
2.4 Toma de datos en campo
De acuerdo con las áreas cuya respuesta espectral se interpretó como vegetación
boscosa, con GPS se verificó en campo con 50 puntos la clasificación asignada.
Con base en dicho recorrido, se seleccionaron los sitios para el muestreo de

30
vegetación (bosques maduros, bosques secundarios y cafetales tanto rústicos
como policultivos), limitados a la continuidad de caminos y a la accesibilidad de los
propietarios.
Se muestrearon un total de 22 sitios de los cuales 10 eran bosques maduros (BM),
6 bosques secundarios (BS) y 6 cafetales (C). Se consideró BM aquel que
tuviera mayor cobertura de vegetación, individuos longevos (DAP alto)
abundantes, tamaño del fragmento (designación subjetiva) y BS aquel que tuviera
mayor apertura del dosel, con abundancia de individuos jóvenes (DAP bajo). Por
su parte el Cafetal se reconoció por la distribución regular de individuos y mayor
dominancia de Coffea arabica. El muestreo se realizó en tres visitas durante los
meses de octubre y noviembre del 2006 y febrero del 2007.
En cada sitio se registraron los parámetros ambientales que se muestran en la
Tabla 1. Para el muestreo de vegetación se utilizó el método recomendado por
Gentry (1988) esto es un transecto con área, modificado a 50 x 3 m (Figura 4)
(Gentry 1988), cuya orientación se determinó al azar. Se registró la presencia de
arbustos y de árboles cuyo diámetro a una altura del pecho de 1.30 cm (DAP)
fuese mayor a 2.5 cm. De cada individuo se registró el DAP (cm), altura (m) y,
cuando fue necesario se colectó material para la identificación taxonómica del
individuo.

31
Tabla 1. Parámetros tomados en cada uno de los sitios de muestreo dentro de la microcuenca del
Río Citlalapa.
Parámetro Características
Numérico Unidad/Instrumento
Coordenadas UTM/GPS Vista 3m
Altitud MSNM/GPS Vista 3m
Pendiente
Grados (°) inclinación /Clinómetro (PM-5/360 PC de
Suunto
Cobertura (%) Densiómetro semi-esférico cóncavo
Suelo Varios (ver detalles en análisis de datos)
Nominal Clasificación
Tipo de vegetación Bosque maduro (BM), bosque secundario (BS), Cafetal
( C)
Sumersión Inundable (I) o no (NI)
Posición topográfica Baja (b), media pendiente (mp), alta pendiente (ap)
Superficie del suelo Mantillo (m), bioderma (bd)
En cada transecto se definieron cuatro puntos equidistantes cada 10 m, y en cada
uno de esos puntos se registraron 4 datos de cobertura, uno por cada punto
cardinal (N, S, E y O), lo que da un total de 16 registros por transecto. Para el
registro de la cobertura se utilizó un densiómetro, donde, por el método de cuatro
puntos centrales por cuadro (Lemmon, 1957), se contó el número de puntos
sombreados por el dosel y el total se divide por el total de puntos que tiene el
densiómetro (96), y finalmente se multiplica por el factor 1.04 (Fig. 4). Las 16
mediciones tomadas en cada transecto se promediaron para obtener el porcentaje
de cobertura por sitio.

32
Figura 4. Ilustración sobre el registro de coberturas.
En el diagrama superior se ilustra un transecto y los cuatro puntos equidistantes
en los que se tomaron tanto los registros de cobertura por cada punto cardinal,
como la muestra de suelo a 10 cm de profundidad.
Adicionalmente en cada uno de estos cuatro puntos (10, 20, 30 y 40 m) se tomó
una muestra de suelo a 10 cm de profundidad. Las 4 muestras de suelo de cada
transecto se mezclaron para obtener una muestra compuesta de suelo por sitio.
____________________ ________ 50 m ___________________________
3m
Transecto con 4 puntos cada 10 m
Método de 4 puntos en densiómetro
+NS EO

33
2.5 Análisis de datos
2.5.1 Valor de Importancia relativa
De cada especie se obtuvo material vegetativo y/o reproductivo y posteriormente
se realizó la determinación de los ejemplares colectados con ayuda del personal
del Herbario “XAL” del Instituto de Ecología, AC.
Se obtuvo el área basal (cm2) de cada individuo utilizando la fórmula,
Para cada sitio se calculó el área basal promedio (m2/ha), la cobertura promedio y
la densidad (ind/m2). Se determinó el valor de importancia relativa (VIR) por
especie para toda la microcuenca, a partir de la siguiente fórmula (Mueller-
Dombois and Ellenberg 1974):
VIR= densidad relativa (dr)*100 + frecuencia relativa (fr)*100 + dominancia relativa
(dor)*100/3
donde, dr= núm. de individuos de la especie/ núm. total de individuos, fr= núm.
sitios en los que estuvo presente la especie/total de sitios y dor= área basal total
de la especie/ área basal total (todos los individuos) (Mostacedo and Fredericksen
2000).

34
2.5.2 Riqueza de especies
Se construyeron matrices de presencia/ausencia (incidencia) y de número de
individuos (abundancia) de las especies en los transectos. Con ellas se
construyeron Curvas de Acumulación de Especies (CAE) mediante métodos de
rarefacción usando el programa EstimateS (Colwell 2000a) teniendo en cuenta:
a) Datos de presencia/ausencia (incidencia) y solo las especies observadas en el
muestreo:
Sobs, según Colwell et al (2004), si se tienen H muestras y sj es el número
de especies encontrado en precisamente j muestras,
,
y donde s0 es el número de especies que existen en esa comunidad y que
no se han podido detectar, es decir que se han detectado en precisamente
0 muestras, la riqueza total es:
Los intervalos de confianza del 95% se calcularon por las fórmulas dadas
por Colwell (Op. Cit.).

35
b) Datos de presencia/ausencia (incidencia) teniendo en cuenta tanto las
especies detectadas como las no detectadas:
Método Chao 2, estimadas sobre la base de la incidencia de las especies
menos frecuentes (que aparecen en una y dos muestras), singletones
(representadas por un solo individuo) y doubletones (representadas por dos
individuos):
Sobs es el número de especies observado en todo el muestreo.
Qi = Número de especies que se presentan en exactamente j muestras. Q1 es
el número de especies “únicas” (es decir, especies presentes en una sola
muestra) y Q2 es el número de especies “duplicadas” (es decir, especies
presentes en solo dos muestras) (Chazdon et al 1998).
c) Teniendo en cuenta la presencia de ambas especies corregidas por una
estimación de la amplitud del muestreo:
Método ICE, estima sobre la proporción de individuos que pertenecen a
especies “infrecuentes” (i.e. que están en menos de 10 muestras) pero
excluyendo a las que están en un solo muestreo:
Sfreq = número de especies frecuentes (encontradas en más de 10
muestras)
Sinfr = número de especies infrecuentes (encontradas 10 o menos muestras)

36
Cice = Estimador de amplitud de cobertura del muestreo de incidencia
γice = Coeficiente de variación estimado del Qi de las especies infrecuentes
Qi = Número de especies que se presentan en exactamente j muestras (Q1
es la frecuencias de “únicas” y Q2 es la frecuencia de “duplicadas”).
El estimador del “sample coverage” es
(Chao and Lee 1992, Chao et al. 1993, Chazdon et al. 1998)
d) Datos de abundancia teniendo en cuenta tanto las especies detectadas como
las no detectadas en el muestreo (estimadas sobre la base de las frecuencia
de las especies raras):
Método Chao 1 (Chao 1984):
Donde
Fi es el número de especies que tienen exactamente i individuos cuando se
agrupan todos las muestras (F1 es la frecuencia de “singletones” y F2 es la
frecuencia de “doubletones”.
Actualmente se usa la fórmula corregida

37
e) Datos de abundancia de ambos tipos de especies corregidos por un estimador
de la amplitud del muestreo:
Método ACE (Chao et al. 1993)
El estimador de amplitud de muestreo (“sample coverage estimator”) basado
en abundancia es:
donde
es decir, el estimador de la amplitud del muestreo es la proporción de todos los
individuos que pertenecen a especies raras (i.e. que tienen menos de 10
individuos cuando se agrupan todos las muestras).
El estimador de riqueza de especies ACE es
donde

38
2.5.3 Comparación de la riqueza de especies en bosques y cafetales.
Aunque el objetivo original era realizar la descripción de la riqueza de la
vegetación de la microcuenca como unidad, debido a la intensidad del muestreo
limitada, se exploraron los datos por tipos de vegetación (bosques y cafetales).
La riqueza (número de especies) de bosques y cafetales se comparó con una
prueba Z de dos colas con distribución Poisson (Zar 1996). Se realizaron dos
gráficas que comparan las curvas de acumulación de especies en bosques y
cafetales, una donde el eje de las abscisas representa los individuos acumulados,
es decir, dimensionada en individuos, lo que compara riqueza de especies, y otra
donde en el eje de las abscisas están los transectos acumulados, o dimensionada
en transectos, que compara densidad promedio de especies por área (Colwell et
al. 2004).
Se obtuvo el índice de Shannon para cada sitio y posteriormente se calculó el
promedio de éstos para bosques y para cafetales.
Para comparar ambas medias, se aplicó una prueba Anova de una vía, asumiendo
una distribución normal como propiedad del índice de Shannon calculado para
varias muestras (Magurran, 1988).

39
2.5.4 Suelo
Se realizaron análisis físicos y químicos del suelo. Se determinó la textura (arcilla,
limo y arena) por el método Bouyoucos (Gee y Bauder 1986). Se calculó el pH por
medio de un potenciómetro, para la conductividad eléctrica (CE) se tomó un
extracto de pasta saturada y se midió directamente con un conductímetro (el cual
mediante dos electrodos incluidos en una sola sonda, produce unas pequeñas
corrientes entre éstos, que miden el paso de la corriente del agua mediante un
voltímetro de gran precisión); se calculó el contenido de N por el método Kjeldhal
(Yuen y Pollard1953). Se determinó el porcentaje de carbono por el método
Walkley-Black, para la materia orgánica se siguió el método de pérdida de materia
orgánica por ignición (Davies 1974) y el fósforo se cuantificó por el procedimiento
de Bray y Kurtz (Bray y Kurtz1945). Todos los análisis se realizaron en el
Laboratorio de Análisis Químicos de Suelos, Agua y Plantas del Instituto de
Ecología, AC.
2.6 Relaciones Vegetación-Ambiente
Se realizó un análisis de correspondencia canónica (CCA) donde se incluyeron los
datos ambientales (altitud, pendiente, cobertura, características de los suelos) y
los florísticos de presencia/ausencia (en este caso se usaron solo las especies
que estaban en dos o más sitios). Dicha ordenación se realizó con el programa
Canoco para Windows 4.5 (Braak y Šmilauer 2002).
En el texto las medias se presentan con un error estándar, a menos que otra cosa
sea indicada.

40
3. RESULTADOS
3.1 Uso del suelo
Según el mapa de uso de suelo (Fig. 5), la mayor parte de la microcuenca está
ocupada por pastizales inducidos (32%) y la superficie de las zonas dedicadas al
cultivo incluyendo cafetales, cereales, zonas de temporal y áreas habitadas
totaliza el 39% del área. Sólo el 26% está ocupado por bosques maduros y
secundarios, aunque, según la observación en el campo, la mayoría son bosques
secundarios (Tabla 2).
Tabla 2. Áreas de cada tipo de uso de suelo presente en la microcuenca.
Tipo Área (ha) Proporción (%)
Cafetales 1125.3 26
Bosques 1127.6 26
Cultivo 421 10
Áreas urbanas 153.4 4
Parcelas en
descanso 148.6 3
Pastizal 1377.2 32
Total 4353.2 100

Figuura 5. Mapa de uso de suelo b
41
basado en la cllasificación de una foto aéreaa de 1995.

42
3.2 Vegetación
En los 22 transectos se registró un total de 158 especies pertenecientes a 107
géneros y 49 familias. Las familias con mayor número de especies fueron
Asteraceae, Fabaceae, Lauraceae y Rubiaceae, las cuales tuvieron entre 10 y 18
especies representadas (Fig. 6). En la relación individuo-familia (Fig. 4b), la familia
Rubiaceae resulta sobresaliente con un total de 130 individuos, lo que representa
un 17% del total, en cambio, la familia Asteraceae está menos representada en
abundancia a pesar de ser la familia con mayor número de especies.
En la tabla 3 se presentan los números de especies e individuos, por forma de
vida y procedencia (nativa o exótica) y en la tabla 4 la proporción correspondiente
de cada forma de vida con respecto al inventario registrado.
El promedio de área basal de la microcuenca resultó de 52.32± 5.96 m2/ha, para
bosques fue de 49.53 ± 7.73 m2/ha y para cafetales de 59.78 ± 7.33 m2/ha donde
no se encontró diferencia significativa (t= -0.75, gl=20, p>0.05) entre ellos.
La densidad promedio resultó de 2,269 ± 278 ind/ha. Para comparar bosques y
cafetales sólo se tomaron los individuos arbóreos con un DAP ≥ 25 cm, debido a
que en los cafetales se ha eliminado el sotobosque como consecuencia de
manejo. En los bosques se obtuvo una densidad de 2,862 ± 249 ind/ha y en los
cafetales de 688 ± 37 ind/ha, no se encontró diferencia significativa entre estos
valores (t=-1.64 gl=20, p>0.05)

43
Géneros y especies por familia
0 5 10 15 20
Tiliaceae
Piperaceae
Myrsinaceae
Solanaceae
Verbenaceae
Myrtaceae
Fagaceae
Rubiaceae
Lauraceae
Fabaceae
Asteraceae
Gen x familia
Spp x familia
Individuos por familia
0 20 40 60 80 100 120 140
Hammamelidaceae
Loganiaceae
Solanaceae
Myrsinaceae
Ulmaceae
Fabaceae
Asteraceae
Clethraceae
Myrtaceae
Fagaceae
Lauraceae
Rubiaceae
Fam
ilias
Individuos
a)
b)
Figura 6. Relación de géneros y especies por familia (a) y de
individuos por familia (b).

44
Tabla 3. Número de especies e individuos por forma de vida y procedencia.
Procedencia*
Bosques Cafetales General
Form
a de
vid
a
Nativa Exótica TOTAL Nativa Exótica TOTAL Nativa Exótica TOTAL
Spp Ind Spp Ind Spp Ind Spp Ind Spp Ind Spp Ind Spp Ind Spp Ind Spp Ind
Árbol 76 396 10 17 86 413 17 52 10 12 27 64 87 452 16 28 101 480
Arbusto 53 265 2 3 55 268 2 4 0 0 2 4 53 264 2 5 57 269
TOTAL 129 661 12 20 141 681 18 56 10 12 29 68 140 716 18 33 158 749
*Los totales no son la suma de los tipos pues hay 8 especies nativas compartidas (7 árboles y 1 arbusto) y 4
especies exóticas compartidas (sólo árboles).
Tabla 4. Proporción de formas de vida y procedencia
Porcentaje especies
(%)
Form
a de
vid
a Nativa Exótica TOTAL
Árbol 55 10 65
Arbusto 34 1 35
TOTAL 89 11 100
El 92% de las especies de bosque resultaron exclusivas de este tipo de
vegetación, mientras que los cafetales tuvieron un 59% de especies exclusivas
(Fig. 7)

45
Figura 7.Especies exclusivas de bosques y cafetales; y compartidas entre los tipos de vegetación.
De las 12 especies compartidas, 8 son nativas las cuales están entre las más
abundantes (Tabla 5).
Tabla 5. Especies nativas compartidas
Especie /morfoespecie Abundancia
Desconocida sp 01 18
Clethra macrophylla 34
Quercus lancifolia 18
Quercus sapotifolia 21
Quercus sartorii 7
Quercus xalapensis 3
Buddleja americana 16
Trema micrantha 24
Las especies con los mayores valores de importancia relativa (VIR) para la
microcuenca fueron Palicourea padifolia (21.29%), Quercus sapotifolia (14.32%),

46
Quercus insignis (13.33%), Trema micrantha (13.31%), Quercus lancifolia
(12.67%), Clethra macrophylla. (12.62%), Turpinia insignis (11.23), Beilschmedia
mexicana (10.5%) y Cinammomum effusum (10.09%); ver Apéndice). El 62% de
las especies tuvo un VIR menor a dos. Los transectos realizados en bosques
registraron una cobertura del dosel de 91.14 ±1.12% y los realizados en cafetales
80.11 ± 3.06%. El promedio para los 22 transectos fue 88± 1.55% de cobertura.
3.3 Diversidad
En la siguiente tabla (Tabla 6) se muestran el número de especies obtenidas en
los muestreos de la vegetación tanto para el total de la microcuenca como por los
tipos de vegetación: Bosques y Cafetales. Sin embargo los resultados observados
difieren de los estimados (Tabla 7) a los observados.
Tabla 6. Datos de riqueza (número de especies) de la vegetación estudiada.
Total Bosques Cafetales Compartidas
Individuos 749 681 68
Transectos 22 16 6
Riqueza 158 141 29 12
Singletones 65 55 19 0
Doubletones 30 26 6 2
3 o más
individuos 63 60 4 10

47
Tabla 7. Resultados de riqueza esperada con estimadores no paramétricos.
Tipo Vegetación Bosque Café
Sobs (Mao Tau) 141 29
Singletons Mean 55 19
Doubletons Mean 26 6
ACE Mean 203.48 69.1
ICE Mean 304.68 131
Chao 1 Mean 199.17 59.08
Chao 2 Mean 295.88 101
Shannon Mean 4.2 2.84
Simpson Mean 33.74 10.33
La distribución de abundancias en números reales con relación al esfuerzo de
muestreo medido en individuos refleja la gran cantidad de especies representadas
con uno o dos individuos (singletones y doubletones) y la escasez de especies
muy abundantes, como se muestra en la Fig. 8.
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8 9 10 11 12 16 18 19 21 24 26 34 89
Núm indivi
Espe
cies
Figura 8. Relación entre el número de especies y el número de individuos.

48
Las curvas de acumulación de especies (CAE) para todos los transectos de la
microcuenca se muestra en la Fig. 9, mientras las CAEs para bosques y cafetales
se muestran en las Figs. 10 y 11 respectivamente. Los estimadores Chao2 e ICE
se basan en la cantidad de singletones y doubletones existentes, por lo que el pico
en las curvas de estos estimadores indica un elevado número de especies poco
representadas el cual disminuye conforme se aumenta el muestreo (Fig 9b).
A lo largo de las curvas de acumulación es claro que cuando se consideran las
especies poco frecuentes o poco abundantes las estimaciones de la riqueza
aumentan considerablemente.
Tanto ICE como CHAO2 se basan en la incidencia, es decir, solo reflejan la
probabilidad de una riqueza asintótica. Al no establecerse en una meseta, indica
que se está lejos de alcanzar la riqueza con el muestreo realizado. En este caso,
en los cafetales se observó que 17 especies de 28, es decir el 60%, son especies
únicas y además son single- o doubletones de los sitios, lo que refleja una riqueza
estimada mucho mayor a la encontrada en el esfuerzo de muestreo, es decir,
habría que aumentar el área para lograr que la pendiente encuentre la riqueza
asintótica.

49
a)
b)
Figura 9.CAE de todos los sitios, a) utilizando el estimador Mao Tau con los intervalos
de confianza (Sobs 95% superior e inferior) b) basadas en estimadores incluyendo
especies no detectadas, singletones y doubletones.

50
a)
b)
Figura 10. CAE para bosques, a) utilizando el estimador Mao Tau con los intervalos de confianza
(Sobs 95% superior e inferior) b) con base en estimadores incluyendo especies no detectadas,
singletones y doubletones.
51
a)
b)
Figura 11. CAE para cafetales, a) utilizando el estimador Mao Tau con los
intervalos de confianza (Sobs 95% superior e inferior) b) con base en estimadores
incluyendo especies no detectadas, singletones y doubletones.

52
3.3.1 Comparación de riquezas entre bosques y cafetales.
La media de la riqueza de especies de los bosques por transecto fue 16.4 ± 1.07 y
en cafetales de 6.0 ± 0.02 especies
A partir de la ecuación:
Z= | X1 – X2| / √ X1 + X2,
se comparó si existía diferencia significativa entre bosques y cafetales en cuanto a
su riqueza y se obtuvo el valor crítico de 2.20, siendo que Z0.05 (2) = t0.05 (2) ∞ = 1.96,
resulta que hay diferencia significativa con una P < 0.05 entre bosques y cafetales.
3.3.2 Comparación de CAE por individuos y por transectos.
Con respecto a la acumulación de especies en bosques y en cafetales, se observó
que en la escala de individuos (Fig. 12a) los intervalos de confianza se
sobreponen, lo que indica que al dimensionar la riqueza por individuos
acumulados como esfuerzo de muestreo no habría diferencia significativa entre la
riqueza de bosques y cafetales. Sin embargo, en la escala de transectos (Fig. 12b)
esto no sucede e incluso las curvas se separan conforme se aumenta el esfuerzo
de muestreo, lo que indica que la densidad de especies por unidad de área es
mayor en bosques que en cafetales.
3.3.3 Índice de Shannon
El índice de Shannon promedio para bosques fue de 2.41 y el de cafetales de
1.50.

53
En la comparación de medias de los índices de Shannon entre bosques y
cafetales se obtuvo una F1, 20= 17.43, lo que indica que la riqueza entre bosques y
cafetales difiere significativamente (P< 0.000467).
3.4 Análisis Canónico de Correspondencias
En el análisis canónico de correspondencias (Canoco o CCA) se obtuvo una
inercia total de 5.37 (ver Tabla 8). El primer y segundo ejes presentaron valores
altos de dispersión (eigenvalores) de 0.525 y 0.418 respectivamente. Las
correlaciones entre especies y ambientes fueron mayores a 0.95 para los 2 ejes.
Los dos primeros ejes representaron el 29.4% (16.4% y 13.0% respectivamente)
de la varianza acumulada en la relación especies – ambiente. En la correlación de
las variables con los ejes (ver Tabla 9), el eje 1 mostró alta correlación con los
porcentajes de arcilla (+), arena (-) y cobertura (-), con la pendiente (-) y con la
riqueza (-), mientras que el eje 2 se relacionó con el pH (+) y el AB (-).
Existen variables ambientales altamente correlacionadas entre sí, tanto positiva
como negativamente (ver Tabla 10). Se excluyó la materia orgánica, pues tenía

54
una correlación de 1 con el porcentaje de carbono orgánico, lo que hacía
redundante su inclusión.
Curvas de acumulación por riqueza
0
20
40
60
80
100
120
140
160
180
0 100 200 300 400 500 600 700 800
Individuos
Espe
cies
B Sobs (Mao Tau)B Sobs 95% CI Lower BoundB Sobs 95% CI Upper BoundC Sobs (Mao Tau) C Sobs 95% CI Upper Bound C Sobs 95% CI Upper Bound
Curvas de acumulación por densidad
0
20
40
60
80
100
120
140
160
180
0 2 4 6 8 10 12 14 16 18
Sitios
Espe
cies
B Sobs (Mao Tau)B Sobs 95% CI Lower BoundB Sobs 95% CI Upper BoundC Sobs (Mao Tau) C Sobs 95% CI Lower Bound C Sobs 95% CI Upper Bound
a)
b)
Fig. 12 CAE diferenciando bosques y cafetales a) dimensionando el esfuerzo
de muestreo con individuos y b) dimensionando el esfuerzo de muestreo por
transectos.

55
Se encontró relación positiva del C orgánico, el porcentaje de arena, el N total y
C.E., que a su vez están negativamente relacionados con la arcilla y el P.
Tabla 8. Resumen de valores obtenidos en el análisis CCA.
EJES 1 2 3 4 Inercia Total
Eigenvalores 0.52 0.41 0.394 0.341 5.369Correlación spp - ambiente 0.97 0.944 0.985 0.978
% Varianza acumuladade spp 9.6 17.3 24.7 31
de relación spp-ambiente 16.4 29.4 41.9 52.7Suma de eigenvalores 5.369
Suma eigenvalores canónicos 3.163
Tabla 9. Correlación obtenida entre los ejes canónicos y las variables ambientales consideradas*.
SPEC AX1
SPEC AX2
SPEC AX3
SPEC AX4
ENVI AX1
ENVI AX2
ENVI AX3
ENVI AX4
SPEC AX1 1
SPEC AX2 0.054 1
SPEC AX3 -0.001 0.018 1
SPEC AX4 -0.023 -0.044 -0.011 1
ENVI AX1 0.97 0 0 0 1
ENVI AX2 0 0.944 0 0 0 1
ENVI AX3 0 0 0.985 0 0 0 1
ENVI AX4 0 0 0 0.978 0 0 0 1
pH H2O 1 -0.194 0.711 0.247 0.165 -0.2 0.753 0.251 0.169
C.E. -0.399 0.381 -0.21 0.155 -0.412 0.403 -0.214 0.158
P Bray 0.475 0.112 0.16 0.051 0.49 0.119 0.162 0.053
. C Org -0.472 -0.113 -0.237 0.294 -0.486 -0.12 -0.24 0.301
. N tota -0.334 0.139 -0.464 0.258 -0.344 0.147 -0.471 0.264
arcilla 0.599 0.166 0.294 -0.07 0.617 0.176 0.298 -0.072
limo -0.001 0.167 -0.225 -0.198 -0.001 0.177 -0.228 -0.203
arena -0.666 -0.254 -0.234 0.159 -0.687 -0.27 -0.238 0.163
Cobertur -0.56 0.073 0.188 -0.384 -0.577 0.078 0.191 -0.393
Pendient -0.642 0.177 -0.007 0.13 -0.661 0.187 -0.007 0.133
Altitud -0.073 0.213 -0.593 0.398 -0.076 0.226 -0.602 0.407
AB TOT -0.15 -0.688 0.046 -0.051 -0.154 -0.729 0.047 -0.052
Riqueza -0.633 0.208 -0.322 -0.487 -0.653 0.22 -0.327 -0.498

56
*Se excluyó la Materia Orgánica, con el fin de disminuir la dispersión ya que tenía los mismos valores del C
orgánico.
Tabla 10. Coeficientes de correlación obtenida entre las variables.
pH H2O 1 C.E. P Bray C Org N total Arcilla Limo Arena Cobertura Pendiente Altitud AB TOT Riqueza pH H2O 1 1
C.E. 0.073 1 P Bray 0.103 -0.171 1 C Org -0.226 0.593 -0.428 1 N total -0.122 0.746 -0.305 0.797 1
arcilla 0.155 -0.195 0.553 -0.756 -0.475 1 limo 0.012 -0.145 -0.043 0.087 0.045 -0.445 1
arena -0.178 0.276 -0.598 0.806 0.509 -0.931 0.089 1
Cobertur 0.155 0.225 -0.455 0.300 0.114 -0.516 0.004 0.572 1
Pendient 0.044 0.397 -0.361 0.444 0.460 -0.517 0.364 0.428 0.227 1 Altitud 0.059 0.480 -0.202 0.458 0.540 -0.319 -0.127 0.405 0.166 0.036 1
AB TOT -0.380 -0.304 -0.108 0.072 -0.352 -0.190 -0.097 0.252 0.156 -0.219 -0.030 1
Riqueza 0.013 0.530 -0.357 0.451 0.557 -0.459 0.208 0.425 0.482 0.530 0.083 -0.178 1
La gráfica resultante del CCA, se dividió en dos esquemas, el que muestra la
relación transectos-ambiente y el de especies-ambiente, con el objetivo de tener
mayor claridad visual. En aquella que muestra la relación transectos-ambiente
(Fig. 13), se observa que hacia el lado izquierdo de la figura se ubican la mayoría
de los transectos (identificados como grupos A y B en la figura) realizados en
bosques y que están asociados a la variables riqueza, cobertura, AB total, C
orgánico, N, la pendiente, arena y CE. Por lo contrario del lado derecho se
encuentran la mayoría de los transectos hechos en cafetales y que están
asociados con las variables arcilla y P, lo que sugiere que el manejo de los
cafetales induce una pérdida de arena en los horizontes superficiales y un
aumento en el contenido de P por fertilización artificial.

57
-1.0 1.0Eje 1-0.8
1.0Ej
e 2
pH H2O 1
C.E.
P Bray
. C Org
. N tota
arcilla
limo
arena
Cobertur
PendientAltitud
AB TOT
Riqueza
1A
2B
3B
4S
5B
6B
7S 8S
9B
10S
11S
12B
13C
14C15B
16B
17C
18C
19C
20B
21C
22B
Figura 13. CCA de datos florísticos en bosques (diamantes) y cafetales (círculos), incluyendo
variables ambientales de suelo (pH, CE, N, P, C orgánico, arena, arcilla, limo) de condiciones
forestales (área basal total, cobertura, riqueza), pendiente y altitud. Los sitios se identifican por un
número seguido por una letra: B para bosques maduros, S para bosques secundarios y C para
cafetales.
El grupo A (figura 13) corresponde a fragmentos de BMM ubicados en sitios con
mejores condiciones de desarrollo forestal en base a las variables ambientales, sin
embargo los puntos están muy dispersos y no es clara su descripción por alguno
de los ejes.
GRUPO B
GRUPO C
GRUPO A
GRUPO B
GRUPO C
GRUPO A

58
-1.0 1.0Eje 1-0.8
1.0
Eje 2 Saur 1
Ilex 1
Oreo 1
Aster_5_
Sene 2
Telan gr
Tetra/ve
Clet 1
Visma me
Cnid mul
Calia 1
Inga 1
Inga 4
Inga 5
Pite 2
Quer ins
Quer lei
Quer sap
Quer sar
Quer xalQuer_P Liqu sty
Laur_. .
Beil mexCina eff
Ocot psy
Budd ame
Hamp 1Mico 1
Mico gla
Cedrela
Trop 1
Myrsin_1
Rapa 2
Rapa_PEuge 1
Euge 2
Euge 3
Psid guaSyzy jam
Piper 1
Piper la
Ramn 1
Pali pad
Psyc 1
Cytr 2
Zant 1
Solan_1
Cest 1
Sola sch
Turp insStyr 1
Styr 2
Styr lab Trem mic
Urer/myr
Cith 1
pH H2O 1
C.E.
P Bray
. C Org
. N tota
arcilla
limo
arena
Cobertur
PendientAltitud
AB TOT
Riqueza
Figura 14. CCA de especies leñosas (árboles y arbustos), incluyendo variables ambientales de
suelo (pH, CE, N, P, C org, arena, arcilla, limo) de condiciones forestales (Área basal total,
cobertura, riqueza), pendiente y altitud.
Así también se forman el grupo B, con fragmentos de BMM ubicados en
condiciones más pobres de nutrientes y suelos más básicos, y el grupo C, donde
se insertan los cafetales y los fragmentos de BMM más pobres en cuanto a
riqueza florística. El cafetal 13 presenta condiciones que lo definen más cercano a
las características forestales que otros sitios de BMM y justamente es un cafetal
que presentaba condiciones de abandono pues no estaba libre de maleza y los
cafetos estaban muy secos y altos.
En la gráfica que muestra la relación especies-ambiente (Fig.14), es claro que
Inga sp., Psidium guajava, Trema micrantha, Quercus xalapensis y Clethra

59
macrophylla, están relacionados (no determinados) con mayores contenidos de
arcilla y de P en el suelo que se asocian con los cafetales. Con las variables que
se asocian con los transectos de bosques (riqueza, cobertura, AB total, C
orgánico, N, la pendiente, arena y CE) se relacionan la mayoría de las especies
del género Quercus, así como Turpinia sp., Styrax sp., Eugenia sp., Liquidambar
sp., y Miconia sp., típicas de BMM.
4. DISCUSIÓN
4.1 Uso de suelo
A pesar de resultar muy poca el área ocupada por asentamientos humanos, la
situación de deterioro de la vegetación de BMM en la microcuenca es notoria por
la proporción tan alta de pastizales y zonas de cultivo (71%). La mayor parte del
cambio del uso del suelo se produjo con el auge que, desde hace treinta años,
tiene la actividad agropecuaria, principalmente la ganadería extensiva, según
comentan informantes de la región (Néstor X, Eliseo X, Ricardo Romero,
comunicación personal). Estas áreas están rodeadas en su mayoría por cafetales
y pastizales. Esto, aunado a que la imagen utilizada refleja una realidad del año
1995, revela un alto estado de fragmentación de las áreas de bosque que,
además, parecen ser de recolonización reciente pues se observó una estructura
de tamaños diamétricos muy pequeños con dominancia de especies secundarias.

60
Desde el siglo XVI se describe el paisaje de Huatusco como un pastizal rocoso,
dedicado a estancias de ganado. Esta situación de deterioro forestal está
relacionada no sólo con la tradición de uso de la tierra con objetivos de producción
y la creciente apertura de caminos desde la colonización, sino con la posterior
instauración de fábricas de fósforos de madera, que explotaban recursos
forestales de la región desde 1849 y la herencia histórica del cultivo del café desde
1858, además de contar con el apoyo del gobierno porfirista que impulsó la
Escuela Regional de Agricultura en la ciudad de Huatusco, una de las dos que
existen en el país, hacia 1880 (Tortolero 1995, Córdova-Santamaría 2003).
Casos similares reportan un cambio de uso de suelo de bosque a cultivo del 40%
(Evangelista-Oliva et al. 2010), y encuentran en los sistemas agroforestales un
recurso de restauración de los sistemas intensivos para disminuir la pérdida de
diversidad, sin obviar la imperante necesidad de proteger los remanentes
boscosos en concordancia con los sistemas productivos locales (Philpott et al.
2008).
La microcuenca Citlalapa está inmersa en un mosaico topográfico cuyas zonas
con poca pendiente son utilizadas para la ganadería y agricultura de temporal y las
laderas accesibles son utilizadas por la producción de café. En el mapa obtenido
es notorio que quedan pocos fragmentos de bosques, que no necesariamente son
consistentes con lo que se considera bosque conservado. Los bosques de la
región están rodeados en su mayoría por cafetales y pastizales. Los datos de
estructura de los bosques en este estudio, agravan aún más el estado aparente de

61
los fragmentos de bosque que pueden observarse, ya que los fragmentos se
encuentran degradados y en procesos tempranos de sucesión secundaria, como
lo indico la estructura diamétrica muy pequeña con dominancia de especies
secundarias. Los sitios mejor conservados se encuentran en pendientes más
pronunciadas, lo que muestra que el difícil acceso es uno de los factores que
permite la preservación de vegetación natural. Sin embargo, debe considerarse
que las cifras obtenidas con este mapa corresponden al paisaje de hace 16 años
(1995). Deben realizarse mapas más actuales para realizar un diagnóstico que
contribuya al ordenamiento territorial, sin embargo, este mapa es útil para generar
un antecedente del estado de la microcuenca en 1995.
4.2 Florística
El conocimiento florístico que existe de la región aún es limitado pues sólo se han
realizado levantamientos aproximados cuyos objetivos son apoyar estudios con
otras connotaciones, como el interés taxonómico o agrícola, o bien estudios
comparativos de sistemas agroforestales donde la localidad de Las Cañadas ha
sido el único punto de muestreo (Manson et al. 2008).
Los orígenes de las especies encontradas coinciden con lo reportado en la
bibliografía, pues en ella se ha mencionado que en el BMM mexicano existen tres
tipos de elementos florísticos, el templado con especies de dosel de origen
neártico, el tropical con sotobosque de origen neotropical y el endémico, cuya
importancia radica a nivel de especies (Rzedowski 1978, Luna-Vega et al. 1999,
Alcántara y Luna-Vega 2001).

62
A pesar que los relictos de vegetación resultan muy reducidos y dispersos, la flora
leñosa de la microcuenca resulta muy diversa, pues existen 158 especies en un
área de muestreo relativamente pequeña (3300 m2= 0.33 ha), lo que puede estar
explicado por la variada topografía y microclimas de la zona que cubren los
requerimientos de hábitat para tal cantidad de especies. Sin embargo, en
ecosistemas como éste la constante tala crea condiciones microclimáticas de
relativa baja disponibilidad de humedad, ocasionando condiciones más limitantes
para el crecimiento vegetal y provocando la reducción local de la diversidad por la
desaparición de taxones poco tolerantes a esos cambios.
Se encontraron mejor representadas especies de árboles medianos característicos
de una etapa sucesional secundaria, pertenecientes a las familias Asterácea,
Laurácea, Rubiácea, y Fabácea. Árboles típicos del BMM primario, como los
pertenecientes a las fagáceas, están bien representados en individuos pero sólo
hay un género (Quercus) de dicho gremio sucesional.
La proporción tan alta de especies nativas (97%) refleja la escasa invasión de
leñosas exóticas en remanentes de bosque sin manejo, y la perseverancia en el
manejo de los sistemas de cultivo cafetalero con especies de la región, aunque la
mayoría de ellas son especies pioneras. Las especies exóticas que se encontraron
son árboles en casi su totalidad y la mitad aproximadamente están en cafetales, lo
cual indica que probablemente son producto de introducción por el hombre y no un
indicio de invasión de hábitat. Como era de esperarse, en los cafetales la mayoría
de las especies son árboles pues son los únicos que les confieren el beneficio de
la sombra según el objetivo de este tipo de manejo.

63
El área basal mayor en cafetales, indica que el tipo de manejo promueve dejar
individuos con dimensiones cuya copa provea sombra, pero también refleja la
antigüedad de dicho sistema de cultivo y la posible composición del bosque
existente en tiempos remotos, pues los árboles con mayor grosor de la
microcuenca se encontraron en estos cultivos y no en las fragmentos de bosque,
lo cual sugiere que los fragmentos de bosque no son remanentes de un bosque
maduro sino que representan diferentes estados sucesionales recientes. Por otro
lado, las áreas basales son menores en los remanentes de BM y, agrupando
transectos de bosques y de cafetales, el número de individuos por transecto se
relaciona inversamente con las áreas basales. Esto indica que los remanentes de
BMM poseen una mayor densidad de individuos que en su mayoría son más
jóvenes, los cuales, a pesar de no ser removidos para abrir nuevas zonas de
cultivo, no han prosperado por la constante extracción de leña. Esta estructura en
los remanentes se puede interpretar como una etapa sucesional temprana.
El bajo número de especies compartidas se puede deber a que en los cafetales un
alto porcentaje (38%)son especies introducidas mientras que éstas son muy
pocas en el BMM (0.09%), las cuales representan apenas el 11% de la flora
muestreada total. Sin embargo se puede inferir que las compartidas-nativas, son
aquellas más tolerantes a procesos de transición y a los cambios
microambientales, ya que se encuentran bien representadas en ambos tipos de
vegetación, por lo que son éstas especies las que se recomendaría explorar en
experimentación para proyectos de restauración de la microcuenca.

64
En trabajos de restauración en bosque se recomienda utilizar una combinación de
especies de crecimiento rápido y tolerantes al sol que sirvan de nodrizas para
especies de crecimiento lento (Ramírez-Marcial et al. 2006).
El tepozán (B. americana) se encuentra recomendado por Fernando Altamirano y
José Ramírez para repoblar la vegetación local (Flores Olvera y Lindig-Cisneros
2005) se considera tolerante al sol y con una alta tasa de crecimiento y se asocia
tanto con vegetación secundaria como a bordes o laderas de bosques relictuales
(Durán-Espinosa 2006). Trema micrantha (tolerante al sol y de crecimiento rápido)
y Clethra macrophylla (medianamente tolerante y crecimiento intermedio), pueden
funcionar como protectoras para la propagación de Quercus sp (González-
Espinosa et al. 2007).
Las especies nativas de bosque mesófilo encontradas en los cafetales y no así en
los fragmentos de bosque (Quercus candicans, Quercus elliptica, Heliocarpus sp.)
es posible que pertenecieran a bosques viejos ya deforestados y que no se hayan
podido distribuir en las zonas boscosas al quedar poco representadas, por lo que
su presencia esté reducida a áreas con manejo. La especie dominante es
Palicourea padifolia, la cual es característica de este tipo de vegetación en etapa
secundaria tanto temprana como tardía. Las especies que siguen en importancia
con valores altos (Q. sapotifolia, Q. insignis, T. micrantha, Q. lancifolia, Clethra
macrophylla., T, insignis, B. mexicana y C. effusum) son especies nativas de
bosque mesófilo y se consideran componentes estructural y fisionómicamente
importantes de este tipo de bosque (Rzedowski 1996).

65
Las demás especies están por debajo de un VIR de 10%, y de éstas el 65% tienen
un VIR menor a 2%, es decir, hay una gran proporción de especies raras o con
poca frecuencia. Resultados similares para este tipo de vegetación fueron
reportados por Williams-Linera y López-Gómez (2005).
La cobertura es muy alta en general en los dos tipos de vegetación, lo que
significa que se conserva una estructura de sombra que beneficia el contenido de
humedad en el suelo.
4.3 Diversidad
La riqueza que existe en la zona es alta, sin embargo, la mayoría son especies
poco representadas pues la suma de “singletones” y “doubletones” comprende a
más de la mitad de la flora registrada. Los sitios con mayor riqueza son los
fragmentos de BMM y los de menor los cafetales, pero aun así el sitio más rico,
apenas tiene un 14% de las especies existentes, corroborando el carácter típico
del BMM de que la alta heterogeneidad y diversidad β son las que contribuyen a la
riqueza del sitio.
Sin embargo, se desconoce la diversidad de la zona que existía de manera previa
a la intervención humana, lo que limita la perspectiva histórica de las datos
obtenidos.
En las curvas de acumulación de especies no se llega a una asíntota, es decir la
diversidad sigue aumentando conforme se siguen haciendo muestreos, además
las curvas de “singletones” y “doubletones” no se sobreponen, lo que se interpreta
como un inventario incompleto. Esto se reconoce como un sub-muestreo derivado

66
del deterioro y la accesibilidad abrupta de ciertos fragmentos, no obstante los
estudios en BMM tienden a presentar este tipo de resultados en una zona tan
fragmentada (González-Espinosa et al. 2004, Williams-Linera et al. 2005, López-
Gómez y Williams-Linera 2006). Estas curvas presentan una menor pendiente
cuando se separan por tipos de vegetación, lo que indica una diversidad
diferencial entre éstos, y se observa que en los bosques la acumulación es mayor
que en los cafetales, sugiriendo una alto recambio en el primer tipo de vegetación
y un bajo número de especies compartidas entre bosques y cafetales.
En la curva de acumulación la riqueza de cafetales y bosques se sobrepone
cuando se relaciona con individuos pero no con transectos (fig. 11b), debido a la
diferencia entre las densidades entre ambas, lo que sugiere que la riqueza de los
cafetales puede estar relacionada con su área, lo que resulta del manejo que se
haya aplicado.
De acuerdo con Peters et al. (2003), la producción del cultivo en un cafetal no
tiene ventaja productiva bajo sombra de Inga sp., y bajo sombra rústica existe una
mayor producción de madera, confiriendo una mayor utilidad al sistema tradicional,
lo que permite concordancia con la conservación de la diversidad en bosques.
Estudios en BMM de Veracruz, derivados del proyecto BIOCAFE, concluyen con
respecto a la vegetación leñosa que los cafetales bajo sombra, conservan árboles
nativos de la región, es decir, son reservorios potenciales para la biodiversidad
local (Williams-Linera y López-Gómez, 2008).

67
4.4 Análisis Canónico de Correspondencias
La inercia total obtenida es muy alta y el tener el 52.7% de varianza explicada en
la relación especies-ambiente es alto con respecto a lo reportado en otros trabajos
(Williams-Linera 2002, López-Gómez 2004). Esto indica que las variables
ambientales se correlacionan adecuadamente la estructura florística de la
microcuenca. Principalmente se relacionan negativamente la arcilla y con el
carbono orgánico y arena que a su vez definen cafetales y bosques
respectivamente. El manejo de cafetales (principalmente la continua eliminación
de la vegetación del sotobosque) ha provocado pérdida de porosidad y menor
retención de humedad, lo que a su vez provoca pérdida de materia orgánica,
nitrógeno total, y de iones solubles pues es común que en estos tipos de bosque
el horizonte A sea muy delgado y con el uso del cultivo se pierda totalmente por
erosión (Bautista et al. 2003). En agroecosistemas cafetaleros de Veracruz se ha
reportado mayor contenido de fósforo en el horizonte superficial de suelos que en
zonas de bosque por el uso de fertilizantes (Geissert 2008)- También se reporta la
costumbre de añadir fósforo para mejorar la calidad del sabor del producto en
Chiapas y Veracruz (Rosas 2008), lo que explicaría la asociación de este
elemento con la arcilla y cafetales.
Las especies arbóreas asociadas a las condiciones menos forestales son de
rápido crecimiento (Trema micrantha, Inga sp.) o son de condición pionera
(Rapanea sp.) o exóticas (Psidium guajava). Las especies relacionadas con
condiciones forestales son la mayoría del género Quercus sp., que son de lento
crecimiento, o características de BMM como Eugenia sp. o Styrax sp.. Entre otras

68
especies secundarias y tipicas de BMM (Psychotria sp., Miconia sp., Vismia sp.),
la especie más abundante en los transectos resultó ser Palicourea padifolia, la
cual presenta una velocidad de crecimiento intermedia, lo que sugiere que la
vegetación de la microcuenca está en su mayoría en una etapa sucesional
secundaria a secundaria-tardía.
5. CONCLUSIONES
La microcuenca del Río Citlalapa hasta 1995, presentaba un alto grado de
deterioro en su vegetación original, pues el 75% de su paisaje corresponde a
zonas intervenidas (potrero, cultivo, poblado). Por ello resulta de gran utilidad la
representación gráfica y espacial que brinda el mapa de uso de suelo, ya que sirve
como referencia para un seguimiento ecohistórico y es una herramienta para
fundamentar la inclusión del área en programas de Servidumbres Ambientales,
Fijación de Carbono, Ecoturismo, etc.
La composición florística coincide con la reportada para BMM en México y las
especies con mayor valor de importancia corresponden a etapas sucesionales
más tempranas, principalmente secundaria, de un BMM.
La diversidad de la microcuenca es alta, sin embargo la combinación de áreas
basales bajas y densidades altas refleja su condición transitoria de recuperación
luego del auge de cultivo de café y consecuente desmonte a mediados del siglo
XX. El abandono del cultivo del café y el fenómeno de emigración han permitido la
regeneración de vegetación natural por abandono de zonas que previamente
fueron explotadas.

69
Algunas condiciones ambientales se relacionan con el deterioro, las variables
edáficas resultan un espejo de la condición de manejo y la vegetación asociada
refleja la tendencia de las condiciones forestales. No solo la tala industrial y la
expansión de zonas de cultivo y ganado deterioran el sitio, sino también la
extracción de leña, o “acarreo hormiga”, desmedido disminuye la posibilidad de
progresión de la sucesión secundaria. Esto se refleja en que las zonas de
pendientes pronunciadas como barrancas, por su difícil acceso poseen una
diversidad mayor y mejores condiciones forestales, sin embargo es poca el área
que representan.
Es de extrema utilidad reconocer las especies nativas susceptibles de ser usadas
en propuestas de reforestación, así como integrar esta información con una
perspectiva espacial, es decir relacionar la diversidad de los fragmentos con
parámetros de paisaje como su ubicación con respecto a cuerpos de agua,
infraestructura, pendiente, etc., y así reconocer los parámetros espaciales que
ejercen mayor influencia en la estructura forestal.

70
A P É N D I C E

IDFamilia
Espe
cie
TOT
1A2B
3B4S
5B6B
7S8S
9B10
S11
S12
B15
B16
B20
B22
BVIR‐B
13C
14C
17C
18C
19C
21C
VIR‐C
VIR‐T
OR
FV1Acanthaceae
Acan
thaceae sp 01
10
00
00
00
10
00
00
00
02.14
00
00
00
0.00
1.56
NAr
2Acan
thaceae sp 02
70
00
00
00
00
00
00
70
02.17
00
00
00
0.00
1.85
NAr
3Odo
nton
ema callistachyum
10
00
00
00
00
10
00
00
02.42
00
00
00
0.00
1.56
NAr
4Actin
idiaceae
Saurau
ia sp
01
40
00
00
00
00
00
04
00
02.31
00
00
00
0.00
1.72
NA
5Saurau
ia leucocarpa
20
00
10
00
10
00
00
00
04.34
00
00
00
0.00
3.17
NA
6Almendrillo
20
00
00
00
00
02
00
00
02.23
00
00
00
0.00
1.64
NA
7An
acardiaceae
Man
gifera indica
10
00
00
00
00
00
00
00
00.00
00
00
10
6.32
1.63
IA
8Spon
dias sp
01
10
00
00
00
00
00
01
00
02.13
00
00
00
0.00
1.56
IA
9Aq
uifoliaceae
Ilex sp 01
80
00
00
03
00
00
30
00
26.99
00
00
00
0.00
5.14
NA
10Araliaceae
Oreop
anax echinop
s2
00
00
00
00
00
10
01
00
4.36
00
00
00
0.00
3.19
NA
11Oreop
anax xalap
ensis
10
00
00
00
00
00
00
01
02.14
00
00
00
0.00
1.56
NA
12Asteraceae
Asteraceae
sp 01
20
00
00
00
00
02
00
00
02.18
00
00
00
0.00
1.61
NAr
13Asteraceae
sp 02
10
00
00
00
01
00
00
00
02.14
00
00
00
0.00
1.57
NAr
14Asteraceae
sp 03
10
00
00
00
00
01
00
00
02.13
00
00
00
0.00
1.56
NAr
15Asteraceae
sp 04
10
00
00
00
01
00
00
00
02.13
00
00
00
0.00
1.56
NAr
16Asteraceae
sp 05
21
10
00
00
00
00
00
00
04.29
00
00
00
0.00
3.14
NAr
17Asteraceae
sp 06
10
00
01
00
00
00
00
00
02.13
00
00
00
0.00
1.56
NAr
18Eupa
toriu
m sp
01
10
00
00
00
00
01
00
00
02.14
00
00
00
0.00
1.56
NAr
19Eupa
toriu
m sp
02
20
00
00
00
02
00
00
00
02.19
00
00
00
0.00
1.61
NAr
20Eupa
toriu
m m
orifo
lium
30
00
00
00
00
03
00
00
02.25
00
00
00
0.00
1.67
NAr
21Pittocau
lon sp 01
20
00
00
00
00
00
00
20
02.19
00
00
00
0.00
1.61
NA
22Senecio sp 01
10
00
00
00
00
01
00
00
02.14
00
00
00
0.00
1.56
NA
23Simsia
sp 01
20
00
00
00
00
20
00
00
02.18
00
00
00
0.00
1.61
NAr
24Telantho
phora gran
difolia
50
01
00
02
00
10
01
00
08.65
00
00
00
0.00
6.33
NAr
25Telantho
phora sp 01
50
00
00
00
00
00
05
00
02.38
00
00
00
0.00
1.77
NAr
26Tetrachyron sp 01
41
10
20
00
00
00
00
00
06.47
00
00
00
0.00
4.74
NAr
27Ve
rbesina sp 01
10
00
00
00
00
00
00
00
00.00
00
00
01
6.11
1.56
NAr
28Bu
rseraceae
Bursera sp 01
10
00
00
00
00
00
00
10
02.16
00
00
00
0.00
1.58
IA
29Ce
lastraceae
Celastraceae
sp 01
10
00
00
00
00
00
10
00
02.16
00
00
00
0.00
1.58
NA
30Microtrop
is sp 01
10
00
00
01
00
00
00
00
02.17
00
00
00
0.00
1.59
NA
31Zino
wiewia sp
01
10
00
00
00
00
00
00
00
00.00
10
00
00
6.66
1.74
NAr
32Ch
enop
odia
Atrip
lex canescens
10
00
00
00
00
00
00
00
00.00
00
00
10
6.37
1.65
NA
33Ch
lorantaceae
Hedyosmum
mexican
um6
00
06
00
00
00
00
00
00
2.51
00
00
00
0.00
1.87
NA
34Clethraceae
Clethra macroph
ylla
344
00
210
00
30
00
03
00
011
.39
00
21
00
15.78
12.63
NA
35Clusiaceae
Vism
ia m
exican
a8
07
00
00
10
00
00
00
00
4.68
00
00
00
0.00
3.47
NA
36Co
rnaceae
Cornus florida
20
00
00
00
20
00
00
00
02.29
00
00
00
0.00
1.68
NA
37Elaeocarpaceae
Sloa
nea sp 01
10
00
00
10
00
00
00
00
02.14
00
00
00
0.00
1.57
NA
38Euph
orbiaceae
Alchornea latifolia
20
00
00
00
00
00
02
00
02.20
00
00
00
0.00
1.62
NA
39Cn
idosculus m
ultilob
us9
00
33
00
11
00
10
00
00
10.92
00
00
00
0.00
8.02
NAr
40Croton
draco
20
00
00
00
00
00
00
00
00.00
02
00
00
8.56
2.21
NA
41Fabaceae
Faba
ceae
sp 01
10
00
00
00
00
00
00
00
12.15
00
00
00
0.00
1.58
NA
42Acacia sp
01
10
10
00
00
00
00
00
00
02.17
00
00
00
0.00
1.59
NA
43Acacia cochliacantha
33
00
00
00
00
00
00
00
02.26
00
00
00
0.00
1.67
NA
44Ca
llian
dra sp 01
20
10
10
00
00
00
00
00
04.27
00
00
00
0.00
3.12
NA
45Ca
llian
dra ho
ustonian
a2
00
20
00
00
00
00
00
00
2.19
00
00
00
0.00
1.61
N1
B O S Q U E S
C A F E T A L E S
Listad
o de
especies leño
sas (con da
p ≥ 2.5 cm
) registrad
as por sitio
de mue
streo. Orig
en: N
(nativa); I (introdu
cida
). Form
a de
vida: Ar (arbu
sto); A
(árbol); VIR (Valor de Im
portan
cia
Relativ
a); B
Q (B
osqu
e); C
F (Cafetal).
71

IDFamilia
Espe
cie
TOT
1A2B
3B4S
5B6B
7S8S
9B10
S11
S12
B15
B16
B20
B22
BVIR‐B
13C
14C
17C
18C
19C
21C
VIR‐C
VIR‐T
OR
FVB O S Q U E S
C A F E T A L E S
46Erythrina sp
01
10
00
00
00
00
00
01
00
02.15
00
00
00
0.00
1.57
IA
47Inga
paterno
20
01
00
00
00
00
00
00
02.14
00
00
01
7.18
3.48
IA
48Inga
sp 01
10
00
00
00
00
00
00
00
00.00
00
00
10
6.27
1.61
IA
49Inga
sp 02
10
00
00
00
00
00
00
00
00.00
00
00
01
6.98
1.84
IA
50Inga
sp 03
40
00
00
00
01
00
01
00
04.59
00
10
01
12.40
6.53
IA
51Inga
sp 04
20
00
00
00
00
00
00
00
00.00
00
00
11
12.56
3.24
IA
52Lysilom
a sp 01
10
10
00
00
00
00
00
00
02.47
00
00
00
0.00
1.79
IA
53Loncho
carpus orizab
ensis
11
00
00
00
00
00
00
00
02.14
00
00
00
0.00
1.57
IA
54Co
joba
arborea
70
00
06
00
00
00
01
00
04.93
00
00
00
0.00
3.64
NA
55Fagaceae
Quercus can
dicans
60
00
00
00
00
00
00
00
00.00
60
00
00
15.03
3.74
NA
56Quercus elliptica
10
00
00
00
00
00
00
00
00.00
10
00
00
6.27
1.61
NA
57Quercus insig
nis
190
06
00
16
01
50
00
00
018
.49
00
00
00
0.00
13.33
NA
58Quercus lancifo
lia18
00
00
00
01
10
05
00
64
15.06
00
01
00
6.61
12.67
NA
59Quercus pinna
tivenulosa
10
00
01
00
00
00
00
00
02.24
00
00
00
0.00
1.64
NA
60Quercus sa
potifolia
210
00
01
01
00
10
00
310
116
.33
00
04
00
10.00
14.32
NA
61Quercus sa
rtorii
70
00
00
10
00
00
04
00
05.17
10
01
00
14.17
7.51
NA
62Quercus xalap
ensis
30
00
00
00
00
00
00
01
02.51
00
00
02
8.46
3.99
NA
63Quercus sp
01
20
01
00
10
00
00
00
00
04.34
00
00
00
0.00
3.17
NA
64Ha
mam
elidaceae
Liqu
idam
bar styraciflu
a16
90
00
00
10
00
00
30
03
10.75
00
00
00
0.00
7.90
NA
65Hipp
ocastanaceae
Billia hipp
ocastanu
m3
00
00
00
00
00
03
00
00
2.38
00
00
00
0.00
1.75
NA
66Icacinaceae
Oecop
etalum
mexican
um1
00
00
00
00
00
00
00
00
0.00
10
00
00
6.13
1.57
NA
67Lauraceae
Lauraceae sp 01
20
00
00
00
00
00
10
10
04.45
00
00
00
0.00
3.25
NA
68Be
ilschmiedia mexican
a26
00
20
06
08
05
02
00
30
14.12
00
00
00
0.00
10.50
NA
69Cinn
amom
um effu
sum
190
05
00
90
10
10
12
00
013
.64
00
00
00
0.00
10.09
NA
70Cinn
amom
um triplinerve
30
00
00
03
00
00
00
00
02.24
00
00
00
0.00
1.66
NA
71Licaria
sp 01
10
00
00
00
00
01
00
00
02.14
00
00
00
0.00
1.56
NA
72Nectand
ra sa
licifo
lia2
02
00
00
00
00
00
00
00
2.20
00
00
00
0.00
1.62
NA
73Nectand
ra sp
01
10
00
00
00
00
00
00
10
02.14
00
00
00
0.00
1.56
NA
74Ocotea bernou
llian
a2
00
00
02
00
00
00
00
00
2.19
00
00
00
0.00
1.61
NA
75Ocotea disjun
cta
10
00
01
00
00
00
00
00
02.42
00
00
00
0.00
1.76
NA
76Ocotea psycho
trioides
100
07
00
02
00
00
01
00
06.77
00
00
00
0.00
5.01
NAr
77Persea
lon
gipes
10
00
00
00
10
00
00
00
02.14
00
00
00
0.00
1.56
IA
78Persea
american
a1
00
00
00
00
00
00
00
00
0.00
00
00
01
7.29
1.93
NA
79Loganiaceae
Budd
leja corda
ta1
00
01
00
00
00
00
00
00
2.16
00
00
00
0.00
1.58
NA
80Bu
ddleja american
a16
012
02
00
01
00
00
00
00
7.11
00
00
01
6.11
6.87
NA
81Malvaceae
Hampea integerrim
a3
01
10
00
00
10
00
00
00
8.51
00
00
00
0.00
6.32
NAr
82Melastomataceae
Micon
ia sp 01
40
00
00
20
10
00
00
10
06.46
00
00
00
0.00
4.73
NAr
83Co
nostegia sp
01
10
00
00
00
00
00
01
00
02.13
00
00
00
0.00
1.56
NAr
84Micon
ia glaberrim
a10
07
20
00
00
01
00
00
00
6.77
00
00
00
0.00
5.02
NAr
85Meliaceae
Cedrela sp 01
40
00
10
00
30
00
00
00
04.59
00
00
00
0.00
3.36
NA
86Gua
rea 01
10
00
00
00
00
00
00
00
00.00
00
00
01
6.42
1.66
NA
87Mon
imiaceae
Sipa
runa
sp 01
20
00
00
00
00
02
00
00
02.20
00
00
00
0.00
1.62
NA
88Sipa
runa
sp 02
10
00
00
00
00
10
00
00
02.13
00
00
00
0.00
1.56
NA
89Sipa
runa
sp 03
10
00
00
00
00
00
00
10
02.14
00
00
00
0.00
1.57
NA
90Moraceae
Trop
his sp 01
30
00
01
20
00
00
00
00
04.32
00
00
00
0.00
3.17
NA
91Myricaceace
Myrica sp 01
10
00
00
00
00
00
00
01
03.48
00
00
00
0.00
2.49
NAr
92Myrsin
aceae
Myrsin
aceae sp 01
20
00
00
00
00
00
01
10
04.29
00
00
00
0.00
3.14
NAr
72

IDFamilia
Espe
cie
TOT
1A2B
3B4S
5B6B
7S8S
9B10
S11
S12
B15
B16
B20
B22
BVIR‐B
13C
14C
17C
18C
19C
21C
VIR‐C
VIR‐T
OR
FVB O S Q U E S
C A F E T A L E S
93Ardisia
sp 01
10
00
00
00
00
10
00
00
02.14
00
00
00
0.00
1.56
NAr
94Ardisia
sp 02
55
00
00
00
00
00
00
00
02.36
00
00
00
0.00
1.76
NAr
95Myrsin
e coria
cea
112
21
50
01
00
00
00
00
011
.10
00
00
00
0.00
8.17
NA
96Myrsin
e sp 01
40
00
20
00
00
00
02
00
04.37
00
00
00
0.00
3.21
NA
97Myrtaceae
Eugenia capu
lli11
09
00
00
00
00
02
00
00
4.75
00
00
00
0.00
3.55
NA
98Eugenia sp 01
160
20
00
00
01
00
00
00
137.23
00
00
00
0.00
5.40
NA
99Eugenia sp 02
260
00
00
00
01
00
10
00
247.61
00
00
00
0.00
5.77
NA
100
Eugenia sp 03
10
00
00
00
00
00
00
10
02.21
00
00
00
0.00
1.61
NA
101
Eugenia sp 04
10
00
00
00
00
00
00
10
02.13
00
00
00
0.00
1.56
NA
102
Psidium gua
java
43
00
00
00
00
00
00
00
02.24
00
00
10
6.42
3.32
IA
103
Syzygium
jambo
s6
00
10
00
40
00
00
01
00
6.57
00
00
00
0.00
4.83
IA
104Nyctaginaceae
Nyctagina
ceae
sp 01
10
00
00
10
00
00
00
00
02.14
00
00
00
0.00
1.56
IAr
105Pipe
raceae
Piper sp 01
20
00
01
01
00
00
00
00
04.27
00
00
00
0.00
3.12
NAr
106
Piper sp 02
10
00
00
00
00
10
00
00
02.13
00
00
00
0.00
1.56
NAr
107
Piper sp 03
10
00
00
00
00
00
00
10
02.14
00
00
00
0.00
1.57
NAr
108
Piper lap
atifo
lium
90
11
00
04
00
30
00
00
08.79
00
00
00
0.00
6.48
NAr
109
Piper sp 04
20
00
00
00
00
00
00
20
02.18
00
00
00
0.00
1.61
NAr
110Proteaceae
Macad
amia sp
01
20
00
00
00
00
00
00
00
00.00
00
02
00
6.69
1.62
IA
111Rh
amnaceae
Rham
nus sp 01
40
10
30
00
00
00
00
00
04.47
00
00
00
0.00
3.28
NA
112
Rham
nus c
apreifo
lia1
00
10
00
00
00
00
00
00
2.13
00
00
00
0.00
1.56
NA
113Ro
saceae
Prun
us brachybotrya
10
00
01
00
00
00
00
00
02.14
00
00
00
0.00
1.56
IA
114
Rubu
s sp 01
11
00
00
00
00
00
00
00
02.13
00
00
00
0.00
1.56
NAr
115Ru
biaceae
Chioccoca sp 01
10
00
01
00
00
00
00
00
02.13
00
00
00
0.00
1.56
NAr
116
Coffe
a sp 01
10
00
00
00
00
01
00
00
02.13
00
00
00
0.00
1.56
IAr
117
Depp
ea sp
01
50
00
05
00
00
00
00
00
02.34
00
00
00
0.00
1.75
NAr
118
Hamelia sp
01
10
00
01
00
00
00
00
00
02.13
00
00
00
0.00
1.56
NAr
119
Palicou
rea pa
difolia
890
213
270
05
314
80
13
120
128
.21
00
00
00
0.00
21.29
NAr
120
Psycho
tria tricho
toma
160
03
00
50
00
01
02
50
011
.25
00
00
00
0.00
8.33
NAr
121
Psycho
tria galeottiana
20
00
02
00
00
00
00
00
02.19
00
00
00
0.00
1.61
NAr
122
Rand
ia sp
01
44
00
00
00
00
00
00
00
02.29
00
00
00
0.00
1.70
NAr
123
Rond
eletia sp
01
30
00
03
00
00
00
00
00
02.28
00
00
00
0.00
1.68
NAr
124
Rond
eletia sp
02
70
00
00
70
00
00
00
00
02.48
00
00
00
0.00
1.86
NAr
125
Rond
eletia ligu
stroides
20
00
02
00
00
00
00
00
02.19
00
00
00
0.00
1.61
NAr
126
Cytrus sp
01
10
00
00
00
00
00
00
00
00.00
00
00
10
6.12
1.57
IA
127
Cytrus sp
02
20
00
00
00
00
01
00
00
02.13
00
00
10
6.14
3.14
IA
128
Zantho
xylum m
elan
ostictum
40
00
00
01
10
00
00
02
06.52
00
00
00
0.00
4.78
NA
129Sabiaceae
Meliosm
a alba
10
00
00
00
10
00
00
00
02.14
00
00
00
0.00
1.57
NA
130Sapo
taceae
Sapo
taceae
sp 01
10
00
00
00
00
00
10
00
02.18
00
00
00
0.00
1.59
NA
131Solanaceae
Solana
ceae
sp 01
40
00
00
00
20
20
00
00
04.38
00
00
00
0.00
3.22
NAr
132
Cestrum sp 01
50
30
00
00
00
02
00
00
04.43
00
00
00
0.00
3.27
NAr
133
Solanu
m sp
01
30
10
00
00
10
10
00
00
06.41
00
00
00
0.00
4.68
NAr
134
Solanu
m sc
hlechtenda
lianu
m7
21
10
20
00
10
00
00
00
10.78
00
00
00
0.00
7.90
NAr
135Staphyleaceae
Turpinia sp
01
10
00
00
00
00
00
00
00
00.00
00
00
10
6.32
1.63
NA
136
Turpinia insig
nis
120
02
00
11
00
20
42
00
015
.42
00
00
00
0.00
11.23
NA
137
Turpinia occidentalis
20
00
00
00
00
00
00
00
22.33
00
00
00
0.00
1.70
NA
138
Turpinia sp
02
10
00
00
00
00
00
00
00
12.18
00
00
00
0.00
1.59
NA
139Styracaceae
Styrax sp
01
40
00
00
10
00
10
02
00
06.47
00
00
00
0.00
4.74
NA
73

IDFamilia
Espe
cie
TOT
1A2B
3B4S
5B6B
7S8S
9B10
S11
S12
B15
B16
B20
B22
BVIR‐B
13C
14C
17C
18C
19C
21C
VIR‐C
VIR‐T
OR
FVB O S Q U E S
C A F E T A L E S
140
Styrax sp
02
40
00
00
00
00
20
20
00
04.41
00
00
00
0.00
3.24
NA
141
Styrax glabrescens
60
05
00
00
00
00
01
00
04.57
00
00
00
0.00
3.37
NA
142Symplocaceae
Symplocos coccinea
30
00
00
00
00
00
00
03
02.32
00
00
00
0.00
1.71
NA
143Tiliaceae
Belotia
sp 01
10
00
00
00
00
00
00
10
02.13
00
00
00
0.00
1.56
NA
144
Heliocarpus sp
01
10
00
00
00
00
00
00
00
00.00
01
00
00
6.18
1.59
NA
145
Heliocarpus app
endiculatus
30
00
00
00
00
03
00
00
03.20
00
00
00
0.00
2.32
NA
146
Tilia m
exican
a2
00
00
20
00
00
00
00
00
2.20
00
00
00
0.00
1.62
NA
147Ulm
aceae
Trem
a micrantha
240
04
00
00
02
00
00
00
05.11
010
51
20
40.55
13.32
NA
148Urticaceae
Myriocarpa sp 01
40
00
01
00
00
03
00
00
04.44
00
00
00
0.00
3.26
NA
149
Myriocarpa sp 02
10
00
00
00
00
01
00
00
02.13
00
00
00
0.00
1.56
NA
150
Urera sp
01
10
00
00
00
00
01
00
00
02.14
00
00
00
0.00
1.57
NA
151Ve
rben
aceae
Citharexylum
sp 01
10
00
01
00
00
00
00
00
02.14
00
00
00
0.00
1.56
NA
152
Citharexylum
sp 02
30
00
00
00
00
00
20
10
04.34
00
00
00
0.00
3.18
NA
153
Cornutia sp
01
20
00
00
00
00
20
00
00
02.19
00
00
00
0.00
1.61
NAr
154
Duranta repens
10
10
00
00
00
00
00
00
02.13
00
00
00
0.00
1.56
NAr
155
Lipp
ia m
yrioceph
ala
30
30
00
00
00
00
00
00
02.26
00
00
00
0.00
1.67
NA
156De
scon
ocida
Descon
ocida sp 01
180
15
20
40
00
31
00
01
014
.53
00
10
00
14.53
14.53
NAr
157
Descon
ocida sp 02
10
00
01
00
00
00
00
00
01.57
00
00
00
1.57
1.57
NAr
158
Descon
ocida sp 03
10
00
00
10
00
00
00
00
01.56
00
00
00
1.56
1.56
NAr
749
3661
6877
3445
3832
2744
2929
4344
2852
1013
910
1010
Total ind
ividuo
s
74

75
6. BIBLIOGRAFÍA
Aguilar, C., E. Martínez, y L. Arriaga. 2000. Deforestación y Fragmentación de
Ecosistemas: ¿Qué tan grave es el problema en México? Biodiversitas 30.
Alcántara, O. y I. Luna. 2001. Análisis florístico de dos áreas con bosque mesófilo
de montaña en el estado de Hidalgo, México: Eloxochitlán y Tlahuelompa.
Acta Botanica Mexicana 54:51–87.
Aldrich, M. y S. Hostettler, editors. 2000. Tropical montane cloud forest, time for
action. UNEP-World Conservation Monitoring Centre, Cambridge, UK.
Alvarez-Aquino, C., G. Williams-Linera, y A. C. Newton. 2004. Experimental native
tree seedling establishment for the restoration of a Mexican cloud forest.
Restoration Ecology 12:412-418.
Bautista-Cruz, A. y R. F. Del Castillo. 2005. Soil Changes During Secondary
Succession in a Tropical Montane Cloud Forest Area. Soil Sci. Soc. Am. J.
69:906-914.
Bautista-Cruz, A., M. C. Gutierrez-Castrena, R. F. Del Castillo-Sánchez, y J. D.
Etchevers-Barra. 2005. Cronosecuencia de un suelo y su clasificación en un
área originalmente ocupada por bosque mesófilo de montaña. TERRA
Latinoamericana 23.
Bautista, A., R. F. del Castillo, y C. Gutiérrez. 2003. Patrones de desarrollo del
suelo asociados con sucesión secundaria en un área originalmente
ocupada por bosque mesófilo de montaña. Ecosistemas Año XII.
Braak, C. J. F. t. y P. Šmilauer. 2002. CANOCO Reference Manual and CanoDraw
for Windows User's Guide: Software for Canonical Community Ordination
(version 4.5).
Bray, R. y L. Kurtz. 1945. Determination of total, organic and available forms of
phosphorus in soil. Soil Sci. Proc. 59:39-45.
Bruijnzeel, L. A. 2004. Tropical montane cloud forest: a unique hydrological case.
Pages 462-482 in M. Bonnell and S. Bruijnzeel, editors. Forests, Water and
People in the Humid Tropics. Cambridge University Press, Cambridge.

76
Bubb, P., I. May, L. Miles, y J. Sayer. 2004. Cloud Forest Agenda. Tech. Rep. 20,
UNEP-WCMC, Cambridge, UK.
Burstein, J., G. Chapela, J. Aguilar, y E. D. León. 2002. Informe sobre la Propuesta
Pago por Servicios Ambientales en México. PRISMA.
Bush, M. B., M. R. Silman, y D. H. Urrego. 2004. 48,000 Years of Climate and
Forest Change in a Biodiversity Hot Spot. Science 303:827-829.
Cartujano, S., S. Zamudio, O. Alcántara, y I. Luna. 2002. El bosque mesófilo de
montaña en el municipio de Landa de Matamoros, Querétaro, México. Bol.
Soc. Bot. México 70:13-43.
Colwell, R. K. 2000a. EstimateS: Statistical Estimation of Species Richness and
Shared Species from Samples (Software and User's Guide). Version 6.
Colwell, R. K., C. X. Mao, y J. Chang. 2004. Interpolating, extrapolating, and
comparing incidence-based species accumulation curves. Ecology 85:2717-
2727.
Córdova-Santamaría, S. 2003. El Huatusco de antaño: Retrospectiva de la
cafeticultura del siglo XIX. Universidad Autónoma Chapingo, México.
Córdova-Santamaría, S. 2005. Café y sociedad en Huatusco Veracruz: formación
de la cultura cafetalera (1870-1930). Consejo Nacional para la Cultura y las
Artes, México.
Cruz-Ruggiero, P. G., M. A. Batalha, V. R. Pivello, y S. T. Meirelles. 2002. Soil-
vegetation relationships in cerrado (Brazilian savanna) and semideciduous
forest, Southeastern Brazil. Plant Ecology 160:1-16.
Chacón, C. y R. Castro, editores. 1998. Conservación de tierras privadas en
América Central. CEDARENA, San José, Costa Rica.
Challenger, A. 1998. Utilización y conservación de los ecosistemas terrestres de
México. Pasado, presente y futuro. CONABIO/ Instituto de Biología, UNAM/
Sierra Madre, México.
Challenger, A. 2003. Conceptos generales acerca de los ecosistemas templados
de montaña en México: su estado de conservación.in O. Sánchez, E. Vega,
E. Peters, and O. Monroy-Vilchis, editors. Conservación de ecosistemas
templados de montaña en México. INE, México.

77
Chao, A. 1984. Nonparametric Estimation of the Number of Classes in a
Population. Scandinavian Journal of Statistics 11:265-270
Chao, A. y S. Lee. 1992. Estimating the Number of Classes via Sample Coverage.
Journal of the American Statistical Association 87:210-217.
Chao, A., M. C. Ma, y M. C. K. Yang. 1993. Stopping rules and estimation for
recapture debugging with unequal failure rates. Biometrika 80:193-201.
Chazdon, R. L., R. K. Colwell, J. S. Denslow, y M. R. Guariguata. 1998. Statistical
methods for estimating species richness of woody regeneration in primary
and secondary rain forests of Northeastern Costa Rica.
Davies, B. 1974. Loss-on ignition as an estimate of soil organic matter. Soil Sci.
Proc. 38.
Dirzo, R. y M. C. García. 1992. Rates of deforestation in Los Tuxtlas, a neotropical
area in southeast Mexico. Conservation Biology 6:84-90.
Doumenge, C., D. Gilmour, M. Ruiz-Perez, y B. J. 1995. Tropical montane cloud
forests:conservation status and management issues. Pages pp. 24–37 in J.
J. O. a. S. F. N. e. Hamilton L.S., editor. Tropical Montane Cloud Forests.
Springer-Verlag, New York.
Durán-Espinosa, C. 2006. Scrophulariaceae. Flora de Veracruz 139.
Evangelista-Oliva, V., J. López Blanco, J. Caballero Nieto, y M. Á. Martínez Alfaro.
2010. Patrones espaciales de cambio de cobertura y uso del suelo en el
área cafetalera de la sierra norte de Puebla. Investigaciones
Geográficas:23-38.
Fa, J. E. y L. M. Morales. 1993. Patterns of mammalian diversity in Mexico. .in T.
P. Ramamoorthy, R. Bye, A. Lot, and J. Fa, editors. Biological diversity of
Mexico: origins and distribution. Oxford University Press, New York.
FAO, editor. 2007. Situación de los Bosques del Mundo, Roma.
Finegan, B. y D. Delgado. 2000. Structural and floristic heterogeneity in a 30-year-
old costa rican rain forest restored, on pasture through natural secondary
succession. Restoration Ecology 8:380-393.
Flores-Villela, O. y P. Gerez. 1988. Conservación en México: síntesis sobre
vertebrados terrestres, vegetación y uso del suelo. INIREV-CI, México.

78
Flores-Villela, O. y P. Gerez. 1994. Biodiversidad y conservación en México:
Vertebrados, vegetación y uso del suelo. CONABIO-UNAM, México.
Flores Olvera, M. H. y R. Lindig-Cisneros. 2005. La Lista de nombres vulgares y
botánicos de árboles y arbustos propicios para repoblar los bosques de la
República de Fernando Altamirano y José Ramírez a más de 110 años de
su publicación. Revista Mexicana de Biodiversidad 76:11-35.
Foster, P. 2001. The potential negative impacts of global climate change on
tropical montane cloud forests. Earth-Science Reviews 55:73-106.
Garcia-Franco, J., G. CastilloCampos, K. Mehltreter, M. L. Martínez, y G. Vázquez.
2008. Composición florística de un bosque mesófilo del centro de Veracruz,
México. Boletín de la Sociedad Botánica de México [en línea].
Gee, G. W. y J. W. Bauder. 1986. Particle size analysis. Pages 383–409 in A.
Klute, editor. Methods of Soil Analysis, Part 1. Physical and Mineralogical
Methods. Agronomy 9, Madison, Wisconsin, USA.
Geissert, D. y A. Ibañez. 2008. Calidad y ambiente físico-químico de los suelos.
Pages 213-221 in R. H. Manson, V. Hernández-Ortiz, S. Gallina, and K.
Mehltreter, editors. Agroecosistemas cafetaleros de Veracruz:
biodiversidad, manejo y conservación. Instituto de Ecología A.C. (INECOL),
Instituto Nacional de Ecología (INE-SEMARNAT), Xalapa, Ver.
Gentry, A. H. 1982. Patterns of Neotropical plant species diversity. Evol. Biol. 15:1-
84.
Gentry, A. H. 1988. Changes in plant community diversity and floristic composition
on environmental and geographical gradients. Annals of the Missouri
Botanical Garden (USA) 75:1-34.
González-Espinosa, M., N. Ramírez-Marcial, A. Camacho-Cruz, S. C. Holz, J. M.
Rey-Benayas, y M. R. Parra-Vázquez. 2007. Restauración de bosques en
territorios indígenas de Chiapas: modelos ecológicos y estrategias de
acción. Boletín de la Sociedad Botánica de México:11-23.
González-Espinosa, M., J. M. Rey-Benayas, N. Ramírez-Marcial, M. A. Huston, y
D. Golicher. 2004. Tree diversity in the northern Neotropics: regional
patterns in highly diverse Chiapas, Mexico. Ecography 27:741-756.

79
Guerrero, A. I. S. y M. Equihua. 2005. Experimental tree assemblages on the
ecological rehabilitation of a cloud forest in Veracruz, Mexico. Forest
Ecology and Management 218:329-341.
Hajabbasi, M. A., A. Jalalian, y H. R. Karimzadeh. 1997. Deforestation effects on
soil physical and chemical properties, Lordegan, Iran. Plant and Soil
190:301-308.
Hamilton, L. S. 1995. Mountain Cloud Forest Conservation and Research: A
Synopsis. Mountain Research and Development 15:259-266.
Hartshorn, G. S. 2002. Biogeografía de los bosques neotropicales. Pages 59-81 in
M. R. G. Urbano and G. H. Kattan, editors. Ecología y conservación de
bosques neotropicales. Libro Universitario Regional (EULAC-GTZ),
Cartago.
Harvey, C. A., O. Komar, R. Chazdon, B. G. Ferguson, B. Finegan, D. M. Griffith,
M. MartÍNez-Ramos, H. Morales, R. Nigh, L. Soto-Pinto, M. Van Breugel,
and M. Wishnie. 2008. Integrating Agricultural Landscapes with Biodiversity
Conservation in the Mesoamerican Hotspot. Conservation Biology 22:8-15.
Lambin, E. F., B. L. Turner, H. J. Geist, S. B. Agbola, A. Angelsen, J. W. Bruce, O.
T. Coomes, R. Dirzo, G. Fischer, C. Folke, P. S. George, K. Homewood, J.
Imbernon, R. Leemans, X. B. Li, E. F. Moran, M. Mortimore, P. S.
Ramakrishnan, J. F. Richards, H. Skanes, W. Steffen, G. D. Stone, U.
Svedin, T. A. Veldkamp, C. Vogel, y J. C. Xu. 2001. The causes of land-use
and land-cover change: moving beyond the myths. Global Environmental
Change-Human and Policy Dimensions 11:261-269.
López-Gómez, A. M. 2004. Los cafetales de sombra como reservorio de la
biodiversidad de plantas leñosas del bosque mesófilo de montaña del
centro de Veracruz. Instituto de Ecologia, A.C., Xalapa, Ver.
López-Gómez, A. M. y G. Williams-Linera. 2006. Evaluación de métodos no
paramétricos para la estimación de riqueza de especies de plantas leñosas
en cafetales. Boletín de la Sociedad Botánica de México:7-15.
Luna-Vega, I. L., A. Othon Alcantara, D. E. Organista, y J. J. Morrone. 1999.
Historical Relationships of the Mexican Cloud Forests: A Preliminary

80
Vicariance Model Applying Parsimony Analysis of Endemicity to Vascular
Plant Taxa. Journal of Biogeography 26:1299-1305.
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton
University Press, Princeton, N.J.
Manson, R. H. 2004. Los servicios hidrológicos y la conservación de los bosques
de México. Madera y Bosques 10(1):3-20.
Manson, R. H., V. Hernández-Ortiz, S. Gallina, y K. Mehltreter, editors. 2008.
Agroecosistemas cafetaleros de Veracruz: Biodiversidad, Manejo y
Conservación. Instituto de Ecología, A.C. (INECOL) e Instituto Nacional de
Ecología (INE), Ciudad de México, México.
Martínez, M. L., O. Pérez-Maqueo, G. Vázquez, G. Castillo-Campos, J. García-
Franco, K. Mehltreter, M. Equihua, y R. Landgrave. 2009. Effects of land
use change on biodiversity and ecosystem services in tropical montane
cloud forests of Mexico. Forest Ecology and Management 258:1856-1863.
Montaña, C. 1990. A floristic-structural gradient related to land forms in the
southern Chihuahuan Desert. Journal of Vegetation Science 1:669-674.
Montaña, C. y P. Greig-Smith. 1990. Correspondence Analysis of species by
environmental variable matrices. Journal of Vegetation Science 1:453-460.
Mostacedo, B. y T. S. Fredericksen. 2000. Manual de métodos básicos de
muestreo y análisis en ecología vegetal. BOLFOR, Santa Cruz, Bolivia.
Mueller-Dombois, D. y H. Ellenberg. 1974. Aims and methods of vegetation
ecology. Wiley, New York,.
Muñiz-Castro, M. A., G. Williams-Linera, y J. M. R. Benayas. 2006. Distance effect
from cloud forest fragments on plant community structure in abandoned
pastures in Veracruz, Mexico. Journal of Tropical Ecology 22:431-440.
Muñoz-Villers, L. E. y J. López-Blanco. 2008. Land use/cover changes using
Landsat TM/ETM images in a tropical and biodiverse mountainous area of
central-eastern Mexico. International Journal of Remote Sensing 29:71 - 93.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca, y J. Kent.
2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858.

81
Nadkarni, N. M., D. Schaefer, T. J. Matelson, y R. Solano. 2002. Comparison of
arboreal and terrestrial soil characteristics in a lower montane forest,
Monteverde, Costa Rica. Pedobiologia 46:24-33.
Palik, B. J., P. C. Goebel, L. K. Kirkman, y L. West. 2000. Using landscape
hierarchies to guide restoration of disturbed ecosystems. Ecological
Applications 10:189-202.
Palmer, M. A., R. F. Ambrose, y N. L. Poff. 1997. Ecological theory and community
restoration ecology. Restoration Ecology 5:291-300.
Palmer, M. A., E. S. Bernhardt, J. D. Allan, P. S. Lake, G. Alexander, S. Brooks, J.
Carr, S. Clayton, C. N. Dahm, J. F. Shah, D. L. Galat, S. G. Loss, P.
Goodwin, D. D. Hart, B. Hassett, R. Jenkinson, G. M. Kondolf, R. Lave, J. L.
Meyer, T. K. O'Donnell, L. Pagano, y E. Sudduth. 2005. Standards for
ecologically successful river restoration. Journal of Applied Ecology 42:208-
217.
Pedraza, R. A. y G. Williams-Linera. 2003. Evaluation of native tree species for the
rehabilitation of deforested areas in a Mexican cloud forest. New Forests
26:83-99.
Peeters, L. Y. K., L. Soto-Pinto, H. Perales, G. Montoya, y M. Ishiki. 2003. Coffee
production, timber, and firewood in traditional and Inga-shaded plantations
in Southern Mexico. Agriculture, Ecosystems & Environment 95:481-493.
Philpott, S. M., W. J. Arendt, I. Armbrecht, P. Bichier, T. V. Diestch, C. Gordon, R.
Greenberg, I. Perfecto, R. Reynoso-Santos, L. Soto-Pinto, C. Tejeda-Cruz,
G. Williams-Linera, J. Valenzuela, y J. M. Zolotoff. 2008. Biodiversity loss in
Latin American coffee landscapes: review of the evidence on ants, birds,
and trees. Conserv Biol 22:1093-1105.
Pimentel, D. y N. Kounang. 1998. Ecology of soil erosion in ecosystems.
Ecosystems 1:416-426.
Pineda, E., C. Moreno, F. Escobar, y G. Halffter. 2005. Frog, Bat, and Dung Beetle
Diversity in the Cloud Forest and Coffee Agroecosystems of Veracruz,
Mexico. Conservation Biology 19:400-410.

82
Ponce-Vargas, A., L.-V. I., A.-A. O., y R.-J. C.A. 2006. Florística del bosque mesófi
lo de montaña de Monte Grande, Lolotla, Hidalgo, México. Revista
Mexicana de Biodiversidad 77:177-190.
Ramírez-Marcial, N. 2001. Diversidad de árboles y arbustos del bosque mesófilo
en el norte de Chiapas y su relación con México y Centroamérica. Boletín
de la Sociedad Botánica de México 69:63-76.
Ramírez-Marcial, N., A. Camacho-Cruz, y M. González-Espinosa. 2005. Potencial
florístico para la restauración de bosques en Los Altos y Montañas del
Norte de Chiapas. Pages Pp. 325-369 in M. González-Espinosa, N.
Ramírez-Marcial, and L. Ruiz-Montoya, editors. Diversidad biológica en
Chiapas. Plaza y Valdéz/ECOSUR/COCyTECH, México, D.F.
Ramírez-Marcial, N., A. Camacho-Cruz, M. Gonzalez-Espinosa, y R. Lopez-
Barrera. 2006. Establishment, survival and growth of tree seedlings under
successional montane oak forests in Chiapas, Mexico. Ecology and
Conservation of Neotropical Montane Oak Forests 185:177-189.
Ramírez-Marcial, N., M. Gonzalez-Espinosa, y G. Williams-Linera. 2001.
Anthropogenic disturbance and tree diversity in Montane Rain Forests in
Chiapas, Mexico. Forest Ecology and Management 154:311-326.
Romero-Alvarado, Y., L. Soto-Pinto, L. García-Barrios, y J. Barrera-Gaytán. 2002.
Coffee yields and soil nutrients under the shades of <i>Inga</i> sp.
vs. multiple species in Chiapas, Mexico. Agroforestry Systems 54:215-224.
Romero, R. 2006. Comunicación verbal. Huatusco, Veracruz.
Rosas, J., E. Escamilla, y O. Ruiz. 2008. Relación de los nutrimentos del suelo con
las características físicas y sensoriales del café orgánico. TERRA
Latinoamericana vol. 26
Rzedowski, J. 1978. Vegetación de México. 2006 edition. Limusa, México.
Rzedowski, J. 1996. Análisis preliminar de la flora vascular de los bosques
mesófilos de montaña de México. Acta Botanica Mexicana 35:24-44.
Soto-Pinto, L., Y. Romero-Alvarado, J. Caballero-Nieto, y G. Segura-Warnholtz.
2001. Woody plant diversity and structure of shade-grown-coffee plantations
in Northern Chiapas, Mexico. Rev. biol. trop 49:977-987.

83
Temperton, V. M. 2004. Assembly rules and restoration ecology : bridging the gap
between theory and practice. Island, Washington, D.C. ; London.
Toledo-Aceves, T., editor. 2010. El bosque mesófilo de montaña en México:
Amenazas y oportunidades para su conservación y manejo sostenible.
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. ,
México, DF.
Toledo-Aceves, T., J. A. Meave, M. González-Espinosa, y N. Ramírez-Marcial.
2011. Tropical montane cloud forests: Current threats and opportunities for
their conservation and sustainable management in Mexico. Journal of
Environmental Management 92:974-981.
Toledo, V. M. y M. Ordóñez. 1993. The biodiversity scenario of Mexico: a review of
terrestrial habitats.in T. P. Ramamoorthy, R. Bye, A. Lot, and J. Fa, editors.
Biological diversity of Mexico: Origins and distributions. Oxford University
Press, Nueva York.
Tortolero, A. 1995. De la coa a la máquina de vapor: Actividad agrícola e
innovación tecnológica en las haciendas mexicanas (1880-1917). Siglo XXI,
México.
UNEP-WCMC. 2004. Potential cloud forest distribution. En http://www.unep-
wcmc.org/forest/cloudforest/maps.cfm (acceso en 2009).
Valverde, P. L., J. A. Zavala-Hurtado, C. Montaña, y E. Ezcurra. 1996. Numerical
analyses of vegetation based on environmental relationships in the southern
Chihuahuan Desert. The Southwestern Naturalist 41:424-433.
Williams-Linera, G. 2002. Tree species richness complementarity, disturbance and
fragmentation in a Mexican tropical montane cloud forest. Biodiversity and
Conservation 11:1825-1843.
Williams-Linera, G. 2007. El bosque de niebla del centro de Veracruz: ecología,
historia y destino en tiempos de fragmentación y cambio climático.
CONABIO-Instituto de Ecologia, AC, Xalapa, Veracruz, México.
Williams-Linera, G. y A. López-Gómez. 2008. Estructura y diversidad de la
vegetación leñosa. Pages 55-68 in R. H. Manson, V. Hernández-Ortiz, S.

84
Gallina, and K. Mehltreter, editors. Agroecosistemas Cafetaleros de
Veracruz: Biodiversidad, Manejo y Conservación, Xalapa, Ver.
Williams-Linera, G., A. M. López-Gómez, y M. A. Muñiz-Castro. 2005.
Complementariedad y patrones de anidamiento de especies de árboles en
el paisaje de bosque de niebla del centro de Veracruz (México). Page 153
−164 in G. Halffter, J. Soberón, P. Koleff, and A. Melic, editors. Sobre
diversidad biológica: El significado de las deiversidades alfa, beta y gamma.
Monografías Tercer Milenio, Zaragoza, España.
Young, T. P., D. A. Petersen, y J. J. Clary. 2005. The ecology of restoration:
historical links, emerging issues and unexplored realms. Ecology Letters
8:662-673.
Yuen, S. H. y A. G. Pollard. 1953. Determination of nitrogen in soil and plant
materials: Use of boric acid in the micro-kjeldahl method. Journal of the
Science of Food and Agriculture 4:490-496.
Zar, J. 1996. Biostatistical Analysis. Third edition. Prentice Hall, Upper Saddle
River, NJ.








![Dizajn Relacione Baze - rti.etf.bg.ac.rs6].pdf · Dizajn Relacione Baze BAZE PODATAKA. 6.2 Dizajn Relacione Baze Karakteristike Dobrog Dizajna Relacione Baze Atomski Domeni i Prva](https://img.pdfslide.tips/doc/110x75/5e0fa9b06df8a816bd173714/dizajn-relacione-baze-rtietfbgacrs-6pdf-dizajn-relacione-baze-baze-podataka.jpg)




















