Embed Size (px)
Citation preview

DISSERTAÇÃO DE MESTRADO
Caracterização de Proteínas do Complexo de Golgi de Tritrichomonas foetus
IVONE ROSA DE ANDRADE
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO CENTRO DE CIÊNCIAS DA SAÚDE PROGRAMA DE CIÊNCIAS MORFOLÓGICAS
RIO DE JANEIRO
2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Ivone Rosa de Andrade
Caracterização de proteínas do complexo de Golgi de Tritrichomonas foetus/ Dissertação: Mestrado em Ciências Morfológicas Universidade Federal do Rio de Janeiro, IB, CCS, Programa de Pós Graduação em Ciências Morfológicas Rio de Janeiro, 2009
xvi, 116.: il
Orientador: Dra. Marlene Benchimol
1. Tritrichomonas foetus. 2. Complexo de Golgi. 3. Ultraestrutura. 4.
Proteômica. 5. Anticorpos

CARACTERIZAÇÃO DE PROTEÍNAS DO COMPLEXO DE GOLGI DE Tritrichomonas foetus IVONE ROSA DE ANDRADE
RIO DE JANEIRO 2009
Dissertação de mestrado apresentada aoPrograma de Ciências Morfológicas �UFRJ como parte das exigências para aobtenção do grau de Mestre em BiologiaCelular Orientador: Dra. Marlene Benchimol

CARACTERIZAÇÃO DE PROTEÍNAS DO COMPLEXO DE GOLGI DE Tritrichomonas foetus IVONE ROSA DE ANDRADE
Dissertação submetida ao corpo docente do Programa de Pós-Graduação em Ciências
Morfológicas � PCM, Instituto de Ciências Biomédicas � ICB, Universidade Federal do Rio
de Janeiro � UFRJ, como parte dos requisitos necessários à obtenção do grau de Mestre.
Aprovada por:
______________________________________________________________________
Prof. Claudia Mermelstein Programa de Ciências Morfológicas � UFRJ
______________________________________________________________________
Prof. Narcisa Leal de Cunha e Silva Instituto de Biofísica � UFRJ
______________________________________________________________________
Prof. Fernando Costa e Silva Filho Instituto de Biofísica � UFRJ
______________________________________________________________________
Prof. Tecia Ulisses de Carvalho (Revisora) Instituto de Biofísica � UFRJ
______________________________________________________________________
Prof. Manoel Luis Pereira da Silva Costa (Suplente) Programa de Ciências Morfológicas � UFRJ
______________________________________________________________________
Prof. Marlene Benchimol (Orientadora) Programa de Ciências Morfológicas - UFRJ
Rio de Janeiro 2009

Essa dissertação foi realizada no Laboratório de Ultraestrutura Celular da Universidade
Santa Úrsula e na Unidade de Espectrometria de Massa e Proteômica � UEMP na
Universidade Federal do Rio de Janeiro, sob a orientação da Profa. Marlene Benchimol, com
o apoio financeiro do CNPq (Conselho Nacional de Desenvolvimento Científico e
Tecnológico), PRONEX (Programa de Núcleo de Excelência), FAPERJ (Fundação Carlos
Chagas Filho de Amparo a Pesquisa do Estado do Rio de Janeiro), CAPES (Coordenação de
Aperfeiçoamento de Pessoal de Nível Superior) e AUSU (Associação Universitária Santa
Úrsula).

v
Este trabalho é dedicado Aos meus pais, Alaide Rosa e Manoel Patrocínio, por serem os pilares da minha vida, por estarem comigo em minha caminhada, segurando minhas mãos e me apoiando. Por abrirem meus olhos e iluminarem meu caminho. Por tudo que me ensinaram, por todo carinho que possibilitou que alcançasse meus ideais.

vi
AGRADECIMENTOS
Aos familiares
Aos meus pais, por me apoiarem e me incentivarem a cada conquista. Pelo carinho e atenção que dedicaram a mim em todos os momentos.
Aos meus primos Sérgio, Eduardo, Célia, Guilherme, Alexandre e Bruna por torcerem por mim e principalmente pelas recordações que me fazem valorizar pequenos momentos.
Aos meus tios Jurimar, Gelson, Luzia, Niel (in memórian) por cuidarem de mim nos momentos difíceis e por vibrarem comigo nos momentos de alegria.
Aos meus afilhados Bruno, Lorena e Breno por serem as luzes da minha vida, por me permitirem ficar encantada a cada nova descoberta de vocês. Aos colaboradores
Ao Prof. José Morgado por me ensinar as técnicas de isolamento do complexo de Golgi, eletroforese e immunoblotting que foram essenciais para o desenvolvimento deste trabalho.
A Profª. Narcisa, pela atenção que sempre me dá nos momentos que necessito de alguma ajuda científica, por ter aceitado fazer parte da minha banca de projeto e dissertação de mestrado e principalmente pelas dicas de imunoprecipitação que aperfeiçoaram e muito os nossos resultados.
Ao Prof. Peralta, por permitir que eu acompanhasse o processamento de obtenção de anticorpos monoclonais, para que assim eu tivesse um melhor entendimento deste procedimento que foi realizado no trabalho, porém não foi desenvolvido por nós.
Ao Dr. Jorge Luiz da Chron Epigen, pela paciência em explicar as etapas do processo de obtenção dos anticorpos monoclonais e por tirar minhas dúvidas em relação ao procedimento.
A Diva da Cruz, pela atenção e dedicação que demonstrou nos dias de observação das etapas do processo de obtenção de anticorpo monoclonais.
A Ana Lúcia e Patrícia, pelas dicas e conselhos de como proceder nas técnicas proteômicas.
Aos Profs. Cláudia Mermelstein, Manoel Luis Costa e Fernando Costa e Silva por
terem aceitado o convite para fazer parte da banca julgadora da minha dissertação de mestrado.
A Profª. Tecia pelas dicas e ótima revisão da minha dissertação de mestrado.

vii
A Juliana pela paciência e pelas várias técnicas que me ensinou durante minha iniciação científica, principalmente a ultramicrotomia.
A todos que me ajudaram a romper 2 L de célula para os experimentos de isolamento
do Golgi e a fazer as inúmeras imunofluorescências durante a seleção dos anticorpos. Obrigada é pouco para vocês.
Aos Amigos
A minha amiga Marjolly por esses 7 anos de amizade, pelas conversas (às vezes monólogos nos seus momentos de raiva e que você não deixa ninguém falar, né?) pelas palavras de incentivo e muitas vezes de consolo (quando você vinha com a frase �Amiga! Você quer um consolo? O meu material não voa, o seu voou!!!! rsrsrs�) só você para me fazer rir nos momentos em que tudo estava dando errado. E principalmente muito obrigada pela atenção e dedicação que você deu a esse trabalho, estando comigo em todos os experimentos de proteômica.
Ao Antonio, por ser essa pessoa única, que me ensinou a compreender coisas essenciais para manter um bom relacionamento com as pessoas. Por ser essa pessoa bem humorada, sempre de bem com a vida que torna nossos dias no laboratório sempre mais agradáveis. Obrigada pelas discussões científicas e ajuda em experimentos. Pela energia positiva e pelo carinho que você transmite em seus abraços de todas as manhãs. Deixe que as pessoas sintam ciúmes da nossa amizade, seremos sempre �amiguinhos� e por isso você vai poder apertar quando quiser.
A Gladys, pela amizade construída no dia-a-dia que foi tomando força e hoje você se
tornou minha irmãzona. Tantos momentos que passamos juntas, muitos bons, alguns ruins, mas você sempre cuidando de mim, preocupada comigo. Mesmo distante não me esqueço de você, obrigada por tudo.
Ao Victor, meu irmãozinho adotivo, por ser esse menino moleque, às vezes ingênuo, mas isso é porque seu coração é enorme e eu admiro isso em você. Obrigada pela ajuda no laboratório, pelas conversas, pela força no momento de tristeza e pelos chopps de sexta que você agitava depois de uma semana exaustiva.
Ao Vilela (Hai!!!), por suas reações e frases inesperadas que sempre quebram o silêncio do laboratório... Mas tudo bem!!! Ás vezes um momento de descontração faz bem mesmo. Obrigada pelas ajudas nos experimentos e pelo carinho que tem por mim.
A Fernanda (ou devo dizer as Fernandas), porque às vezes você está fazendo 1001
coisas ao mesmo tempo, mas sempre tem um tempinho para nos ajudar nos experimentos, nas culturas de células, nas compras de reagentes. Obrigada por tudo, principalmente pela amizade nos últimos tempos.
A Débora (Deb), você conseguiu tirar meu posto de pequininha do laboratório... É,
realmente esse posto tem muito mais sua cara do que a minha. Mas fico muito feliz por isso, pois estou tendo a oportunidade de conhecer a cada dia mais essa pessoa meiga e sempre prestativa que você é.

viii
Ao Eli (papai), por todo carinho e cuidado que teve e continua tendo por mim, mesmo não trabalhando mais juntos. Obrigada também pelas ajudas no MET.
Ao Rodrigo, que quando estava defendendo sua dissertação de mestrado disse que era
para eu deixar de trabalhar com Giardia e passar a trabalhar com Trichomonas que era muito mais legal. Acho que isso ficou no meu subconsciente. Pois bem, não foi escolha minha, mas os caminhos me levaram a trabalhar com Trichomonas e estou muito feliz. Obrigada também pelo carinho, atenção e pela ajuda que você me deu durante as visualizações no Multifoton, que apesar de não terem sido para este trabalho, ajudaram a responder perguntas de um trabalho desenvolvido paralelamente.
Ao Fabio, por me compreender e me dar razão em alguns momentos... Hum!!!! Será
porque somos muito parecidos... Taurinos!!!! Fome sempre!!! rsrsrs Obrigada pelo companheirismo da época de faculdade e pela amizade fora dela.
A Karine, minha amiga de infância, às vezes cabeça dura demais, mas adoro esse seu
jeito. Finge que é durona nos momentos difíceis e sempre vê o que tem de melhor em cada situação. Com certeza sem você na minha vida, ela teria muito mesmo graça.
A equipe do Museu Espaço Ciência Viva, principalmente ao Pedro Persechini,
Eleonora Kurtenbach, Robson Coutinho, Dona Lurdes e Gustavo Rubini e Sônia, por me ensinarem que existe outro jeito de ensinar bem diferente das tradicionais salas de aula. Aprendi muito com vocês. A motivação de vocês em ver esse museu crescer e dar certo é contagiante. Obrigado por torcerem por mim.

ix
AGRADECIMENTOS ESPECIAIS
A Prof. Marlene Benchimol, primeiramente pela oportunidade que a senhora me deu de poder crescer profissionalmente, me aceitando para fazer parte da sua equipe. Pela confiança que depositou em mim e pelo carinho praticamente de mãe que tem demonstrado durante esses 5 anos que estamos trabalhando juntas. Por me tranqüilizar no momento difícil que passei, por todas as conversas e broncas também, que mesmo sendo poucas me fizeram refletir e contribuíram para o meu crescimento profissional e pessoal. Obrigada pela senhora ser este exemplo de seriedade e dedicação.
A Prof. Lina Zingali, por aceitar que desenvolvêssemos parte deste trabalho na UEMP, pela oportunidade de aprendizado das técnicas bioquímicas e proteômicas. Por vibrar conosco a cada resultado obtido e pelas ótimas sugestões para a otimização de nossos resultados.
Ao Rafael, meu anjo protetor que tem estado todo tempo ao meu lado me ajudando e cuidando de mim, literalmente segurando em minhas mãos nos momentos difíceis. Pessoa especial, dedicada, carinhosa e amiga, você trouxe alegria a minha vida em um momento em estava precisando. Simplesmente obrigada por você ser quem você é.

x
Sei que meu trabalho é uma gota no oceano. Mas sem ele, o oceano seria menor... (Madre Teresa de Calcutá)

xi
ÍNDICE Dedicatória v
Agradecimentos vi
Epígrafo x
Indice xi
Publicações xiii
Resumo xiv
Abstract xv
Abreviaturas xvi
1. INTRODUÇÃO 1
1.1 Tritrichomonas foetus e Tricomonoses 1
1.1.1 Tricomonose felina 1
1.1.2. Tricomonose suína 1
1.1.3 Tricomonose canina 2
1.1.4 Tricomonose bovina 2
1.1.5 Tritrichomonas foetus em humanos 4
1.1.6 Ultraestrutura Celular 5
1.1.6.1 Superfície Celular 5
1.1.6.2 Citoesqueleto 6
1.1.6.2.1 Complexo Pelta-Axóstilo 7
1.1.6.2.2 Filamentos Parabasais 8
1.1.6.2.3 Corpúsculos Basais e Filamentos Associados 9
1.1.6.2.4 Flagelos 10
1.1.6.3 Vacúolos e Lisossomos 10
1.1.6.4 Hidrogenossomos 11
1.1.6.5 Núcleo e Divisão 13
1.1.6.6 Retículo Endoplasmático 14
1.1.6.7 Complexo de Golgi 15
1.1.6.7.1 Organização Estrutural 15
1.1.6.7.2 Funções 16
1.1.6.7.3 Complexo de Golgi em tricomonadídeos 18
1.2. Justificativa 22

xii
1.2.1 Objetivo Geral 22
1.2.2 Objetivos Específicos 22
2. MATERIAL E MÉTODOS 23
2.1 Cultivo in vitro 23
2.2 Fracionamento Subcelular 23
2.3 Produção de Anticorpos Monoclonais 25
2.4 C6-NBD-Ceramida 29
2.5 Imunofluorescência 29
2.6 Microscopia Eletrônica de Transmissão 30
2.6.1 Resina Hidrofílica � Unicryl e LR-White 30
2.6.2 Resina Hidrofóbica � Epon 31
2.7 Técnicas Proteômicas 32
2.8 Dosagem de Proteínas � Método de Lowry e Peterson 33
2.9 Imunoprecipitação 34
2.10 Eletroforese em Gel de Poliacrilamida (SDS-PAGE) 34
2.11 Revelação do Gel 35
2.11.1 Coomassie G-250 35
2.11.2 Impregnação pela prata 36
2.12 Immunobloting 36
2.13 Espectrometria de Massa (MALDI-TOF e MALDI-TOF/TOF) 37
3. RESULTADOS 39
3.1 Fracionamento Subcelular 39
3.2 Caracterizações morfológicas da fração do complexo de Golgi 39
3.3 Obtenção de anticorpos monoclonais 42
3.4 Imunofluorescência 67
3.5. Imunocitoquímica 79
3.6 Eletroforese em gel de poliacrilamida 89
3.7 Immunoblotting 89
3.8 Imunoprecipitação 89
3.9 Espectrometria de Massa (MALDI-TOF/TOF) 96
4. DISCUSSÃO 100
5. CONCLUSÕES 105
6. REFERÊNCIAS 106

xiii
! Os resultados desta dissertação encontram-se no seguinte manuscrito em preparação:
DE ANDRADE ROSA, I., CARUSO, M. B., MORGADO-DÍAZ, J. A., ZINGALLI, R. B., BENCHIMOL, M. Characterization of 60 kDa and 66 kDa proteins of Golgi complex of Tritrichomonas foetus. ! Os resultados desta dissertação foram apresentados na forma de pôster no seguinte
congresso: ROSA, I. A. CARUSO, M. B., MORGADO-DÍAZ, J.A., ZINGALI, R. B., BENCHIMOL. M. Identification and characterization of a Golgi complex protein of Tritrichomonas foetus. Simpósio de Microscopia no Cerrado realizado entre os dias 24 a 26 de novembro de 2008 no Centro de Convenções da Pousada Pirineus em Pirinópolis, GO. ! Durante o desenvolvimento desta dissertação também participei dos seguintes
trabalhos: BENCHIMOL, M., DE ANDRADE ROSA, I., DA SILVA FONTES, R., BURLA DIAS, A. J. Trichomonas adhere and phagocytose sperm cells: adhesion seems to be a prominent stage during interaction. Parasitol Res. 2008.102(4):597-604. ROSA, I. DE A., EINICKER-LAMAS, M., BERNARDO, R. R., BENCHIMOL, M. Cardiolipin, a lipid found in mitochondria, hydrogenosomes and bacteria was not detected in Giardia lamblia. Exp Parasitol. 2008. 120(3):215-20.

xiv
RESUMO
Tritrichomonas foetus é um parasita do trato urogenital de gado bovino causando inúmeros problemas de saúde como infertilidade, aborto e infecções. Sabe-se atualmente que também pode infectar outros animais como: gatos, cães e porcos. Como todos os membros da Família Trichomonadidae, T. foetus apresenta diversas organelas peculiares como a costa, axóstilo, pelta, hidrogenossomos e filamentos parabasais, entre outros. Possui um aparelho Parabasal muito desenvolvido formado pelo complexo de Golgi e filamentos parabasais, indicando um papel significativo, análogo ao já descrito em outras células, embora pouco se saiba a respeito da composição de proteínas residentes desta organela em T. foetus. Portanto, procedemos à purificação do complexo de Golgi de T. foetus e a geração de anticorpos monoclonais anti-Golgi que levaram ao isolamento de proteínas de 60 e 66 kDa. Utilizamos diversos testes como: imunofluorescência, microscopia eletrônica de transmissão, imunoprecipitação, eletroforese, entre outros, para a caracterização destas proteínas. Por imunocitoquímica, verificamos que, em T. foetus, estas proteínas estão presentes no Golgi, com uma distribuição específica e em algumas vesículas. O anticorpo monoclonal 20.3 quando testado em controles como Leishmania e Trypanosoma cruzi resultou em uma marcação perinuclear, enquanto que em células da linhagem MDCK, a localização foi observada no complexo de Golgi, indicando a conservação destas proteínas. Devido à falta de banco de dados de T. foetus utilizamos banco de dados de T. vaginalis, por se tratar do organismo mais próximo filogeneticamente. Usando técnicas proteômicas foi possível identificar seis proteínas ainda não caracterizadas, repetições de anquirina e a reigião N-terminal da adaptina os quais mostraram homologia com seqüências de aminoácidos da proteína caracterizada pelo anticorpo anti-Golgi. Entretanto, outras metodologias como: a derivatização da proteína, o seqüenciamento manual e a análise por Q-TOF ainda são necessárias para a confirmação dos dados obtidos por espectrometria de massa do tipo MALDI-TOF/TOF.

xv
ABSTRACT
Tritrichomonas foetus is a parasite of the urogenital tract of cattle causing numerous health problems such as infertility, miscarriage and infection. Currently, T. foetus is recognized as a parasite that can also infect other animals such as cats, dogs and pigs. Like all members of the Family Trichomonadidae, T. foetus presents several unique organelles such as the costa, axostyle-pelta complex, hydrogenosomes and parabasal filaments, among others. The well developed parabasal apparatus formed by Golgi complex and the parabasal filament, indicates a significant role, similar to that previously described in other cells, although little is known about the resident-protein composition of this organelle. Therefore, we proceeded to the purification of the Golgi complex of T. foetus and the generation of anti-Golgi monoclonal antibody that led to the isolation of proteins of 60 and 66 kDa. We have used tests such as immunofluorescence, transmission electron microscopy, immunoprecipitation, SDS-PAGE, among others, to characterize these proteins. By immunocytochemistry, we found that these proteins are present in the Golgi of T. foetus, with a specific distribution on it and also in some vesicles. The monoclonal antibody 20.3 was also used in controls such as Leishmania and Trypanosoma cruzi and resulted in a perinuclear labeling, whereas in the MDCK cell line, the location was observed in the Golgi complex, indicating the conservation of these proteins. Due to lack of database of T. foetus, we have used the database of T. vaginalis, since it is the phylogenetically closer organism. Using proteomics techniques, it was possible to identify six uncharacterized proteins, ankyrin repeat protein and adaptin N-terminal region and which showed homology with amino acids sequences of proteins isolated by the presently studied antibody anti-Golgi. However, other methodologies such as the derivatization of the protein, the manual sequencing and analysis by Q-TOF are still needed to confirm the data obtained by mass spectrometry of type MALDI-TOF/TOF.

xvi

xvii
ABREVIAÇÕES
BSA albumina sérica bovina
cdg Camundongo
DMEM Dulbecco's Modified Eagle's Medium
DMSO Dimetilsulfóxido
DTT Ditiotreitol
FG Fração do Golgi
GTP Guanosina trifosfato
HAT Hipoxantina, Aminopterina e Timidina
HGPRT Hypoxantina Guanina Fosforibosil Transferase
HT Hipoxantina e Timidina
IAA Iodoacetamida
IP Imunoprecipitação
MALDI Matrix-assisted laser desorption/ionization
MDCK Madin-Darby Canine Kidney Cells
MS Mass spectrometry
NCBInr National Center for Biotechnology não redundante
PEG Polietilenoglicol
PNS Sobrenadante pós nuclear
Q-TOF Quadrupole time-of-flight
TTP Timidina trifosfato
TCA Ácido tricloroacético
TOF Time-of-flight mass spectrometer

1
1. INTRODUÇÃO
1.1 TRITRICHOMONAS FOETUS E TRICOMONOSES
Tritrichomonas foetus é um protozoário descrito inicialmente como um parasito do
sistema reprodutor bovino. Atualmente, os microrganismos desta espécie têm sido
encontrados também no sistema digestivo de gatos, cães e porcos.
1.1.1 Tricomonose felina
A tricomonose felina é uma doença caracterizada por afetar o intestino grosso de
gatos, principalmente de gatos jovens e filhotes (FOSTER et al., 2004; GOOKIN et al., 2001;
LEVY et al., 2003). Este tipo de tricomonose pode apresentar como sinais clínicos diarréia
crônica associada com sangue e/ou muco, flatulência e irritação anal (FOSTER et al., 2004;
STOCKDALE et al., 2006). Porém, muitos gatos infectados só apresentam o quadro de
diarréia semanas ou meses após a infecção, conseqüentemente, estes animais apresentam
problemas na eliminação do parasito, não tendo uma cura espontânea (STOCKDALE et al.,
2008).
1.1.2 Tricomonose suína
Em suínos, T. foetus foi descrito inicialmente como outra espécie de Trichomonas,
chamada de Tritrichomonas suis. Embora apresentassem hospedeiros diferentes, estudos têm
descrito T. foetus e T. suis como sendo a mesma espécie, por serem idênticos
morfologicamente. Além disso, evidências de estudos de microscopia eletrônica de
transmissão, cultura in vitro, ensaios bioquímicos e imunológicos têm demonstrado que estes
dois tricomonadídeos podem realmente se tratar da mesma espécie (MATTOS et al., 1997;
LUN et al., 2005; RIVERA et al., 2008). Este protozoário é comumente encontrado na

2
cavidade nasal, estômago, ceco, cólon e ocasionalmente no intestino delgado de porcos (LUN
et al., 2005). Porém, nestes animais T. foetus é um protozoário comensal e devido a isto não
há relatado de sinais clínicos em porcos infectados (TACHEZY et al., 2002).
1.1.3 Tricomonose canina
A descrição de tricomonadídeos em fezes de caninos com diarréia já foi relatada com
base em caracterizações morfológicas. As infecções causadas por estes organismos eram
classificadas como oportunistas ocasionadas por Pentatrichomonas hominis. No entanto, a
caracterização molecular desses tricomonadídeos só foi descrita recentemente através da
análise gênica das seqüências de rRNA, revelando assim, a identidade dos tricomonadídeos
observados nas fezes de cães com diarréia. Com base nesse estudo, os tricomonadídeos
encontrados apresentaram 100% de identidade com Tritrichomonas foetus ou P. hominis
(GOOKIN, et al., 2005).
1.1.4 Tricomonose bovina
Em bovinos, T. foetus é descrito como o agente causador da tricomonose urogenital,
uma doença sexualmente transmissível (DST) (HONIGBERG, 1978). Trata-se de um parasito
monogenético, capaz de habitar o trato reprodutivo bovino incluindo regiões como o
prepúcio, a região distal do pênis, vagina e útero (HONIGBERG, 1978). A tricomonose
urogenital bovina é transmitida através da monta ou do uso de sêmen contaminado, levando a
um grande prejuízo econômico (HONIGBERG, 1978; ALSTAD et al., 1984; BONDURANT,
1985).
O ciclo celular de T. foetus in vitro foi estimado em seis horas (RIBEIRO, 1997), onde
os trofozoítos se multiplicam por divisão binária longitudinal, através de uma mitose do tipo

3
fechada com fuso extranuclear (HEATH, 1980; CAVALIER-SMITH & CHAO, 1996;
DACKS & REDFIELDS, 1998; RIBEIRO et al., 2000).
Estudos filogenéticos realizados ao nível molecular utilizando a subunidade menor do
RNA ribossomal, foram utilizados para classificar esse organismo como sendo um dos
eucariotos mais primitivos (LEIPE et al., 1993; VISCOGLIOSI & BRUGEROLLE, 1993;
VISCOGLIOSI et al., 1999). Desta forma, foi enquadrado com a seguinte classificação
taxonômica:
Filo Parabasalia
Classe Zoomastigophorea
Ordem Trichomonadida
Família Trichomonadidae
Gênero Trichomonas
Espécie Tritrichomonas foetus
A tricomonose urogenital bovina ainda é pouco estudada devido a algumas
dificuldades encontradas, entre elas, o custo de se manter animais experimentais, ausência de
modelos de laboratório confiáveis e a ausência de linhagens celulares que possam reproduzir
características do tecido ou órgão. Células epiteliais e também não epiteliais tais como:
células epiteliais humanas (WISH) e células de rim de cachorro (MDCK) têm sido
empregadas nos estudos de interação entre tricomonas e células hospedeiras (ALDERETE &
PEARLMAN, 1984; KRIEGER et al., 1985, RASMUESSEN et al., 1986; SILVA FILHO &
DE SOUZA, 1988).
A adesão de tricomonadídeos, assim como em outros processos de interação célula-
célula, é um fenômeno muito complexo precedido pela etapa de reconhecimento celular. A
adesina TF190 (SHAIA et al., 1998) demonstrou ter capacidade de reconhecimento de
moléculas de células hospedeiras, favorecendo assim a citoaderência do parasito. Ainda pouco

4
se sabe sobre os componentes das células epiteliais envolvidos no processo de
reconhecimento pelos tricomonadídeos. Porém, alguns glicoconjugados de domínios apicais
de MCDK-I são candidatos, principalmente em interações com T. foetus (BONILHA et al.,
1995). Lectinas presentes na superfície dos trichomonadideos também parecem apresentar
uma grande importância no reconhecimento de células epiteliais (BABÁL & RUSSEL, 1999).
O mecanismo pelo qual o parasito provoca alterações às células hospedeiras ainda não
é totalmente compreendido. Entretanto, foi sugerido que citotoxinas solúveis, principalmente
cisteíno-proteases liberadas pelo parasito possam ter um papel importante no efeito citotóxico
nas células hospedeiras e na infecção (ARROYO & ALDERETE, 1995; THOMFORD et al.,
1996; PETRIN et al., 1998; MENDONZA-LOPEZ et al., 2000; QUE & REED, 2000; SAJID
& MCKERROW, 2002). Além disso, essas citotoxinas também podem agir como fatores de
virulência (ARROYO & ALDERETE, 1989; NEALE & ALDERETE, 1990; ARROYO &
ALDERETE, 1995; MALLINSON et al., 1994; MALLINSON et al., 1995; THOMFORD et
al., 1996), como fatores de adesão (CROUCH & ALDERETE, 1999; MENDONZA-LOPEZ
et al., 2000; ALVAREZ-SANCHEZ et al., 2000) e ainda como fatores que contribuem para a
patogenicidade quando liberado na superfície da mucosa hospedeira (TALBOT et al., 1991;
BASTIDA-CORCUERA et al., 2000).
Estudos recentes foram capazes de demonstrar a destruição de células epiteliais
vaginais bovinas quando em interação de T. foetus (SINGH et al., 1999). Este dano causado
às células epiteliais seria provocado pela indução de apoptose que pode ser causada por
cisteíno-protease de 30kDa (SINGH et al., 2004).
1.1.5 Tritrichomonas foetus em humanos
Recentemente, a identificação de algumas espécies de tricomonadídeos feita em
pulmões de humanos foi realizada através de estudos imunológicos e moleculares. Os

5
resultados apontaram reações positivas para T. vaginalis (DUBOUCHER et al., 2003), P.
hominis (JONGWUTIWES et al., 2000), T. tenax (MALLAT, et al., 2004), T. gallinarum
(KUTISOVA et al., 2005) e T. foetus (DUBOUCHER et al.,2006). A identificação desses
dois últimos protozoários cria dúvidas a respeito do potencial zoonótico dos tricomonadídeos,
embora a possibilidade de cepas adaptadas aos humanos não possa ser excluída. A presença
de tricomonadídeos no trato respiratório de humanos não é incomum e faz com que a real
freqüência destes microrganismos no pulmão seja questionada (DUBOUCHER et al., 2008).
A presença de tricomonadídeos como um agente coinfectante foi relata em 60% dos
pacientes com pneumonia causada por Pneumocystis jiroveci. Em casos em que a doença é
provocada por fungos essa porcentagem pode atingir 100%. Este fato pode ocorrer devido ao
ambiente propício criado pela obstrução dos alvéolos, por fungos ou por debris celulares,
gerando um local de hipóxia. Isso leva a crer que a hipóxia alveolar seja um fator que
favoreça o desenvolvimento de tricomonadídeos mais do que a imunodepressão
(DUBOUCHER et al., 2008).
1.1.6 Ultraestrutura Celular
1.1.6.1 Superfície Celular
A superfície celular compreende a membrana plasmática da célula e o glicocálice, ou
seja, a bicamada lipídica com proteínas integrais, periféricas e/ou ancoradas e os carboidratos,
associados covalentemente a proteínas ou lipídeos do lado externo da membrana, formando
glicoproteínas e glicolipídeos, respectivamente (ALBERTS et al., 2004).
A membrana plasmática de T. foetus apresenta três regiões fisiologicamente distintas:
a membrana que recobre o corpo do parasito, a membrana ondulante e os flagelos. A região
que recobre o corpo do parasito apresenta um glicocálice bastante desenvolvido, apresentando
um aspecto ondulado, quando visto por criofratura (BENCHIMOL, et al., 1992).

6
Na região flagelar é possível verificar áreas de especialização bem definidas, como a
membrana que reveste os flagelos. Esta possui uma menor densidade de partículas
intramembranosas quando observadas por criofratura, quando comparada com a membrana do
corpo da célula. Estas partículas tratam-se de proteínas integrais, as quais teriam a função de
conectar o citoesqueleto com a organização do axonema (BENCHIMOL et al., 1992). Na
membrana dos flagelos anteriores observam-se arranjos circulares de partículas
intramembranosas, formando rosetas, que poderiam ter funções sensoriais (BENCHIMOL et
al., 1992). Outra região especializada é o colar ciliar ou necklace, observada na região de
onde emergem os flagelos anteriores. Entretanto, sua função ainda é desconhecida
(BENCHIMOL et al., 1992).
A membrana ondulante, que liga o flagelo recorrente ao corpo do parasito, acompanha
o batimento deste flagelo, sugerindo que deva atuar dissipando as vibrações causadas pela
força propulsora. Concentrado nesta membrana encontra-se uma extensa trama de filamentos
estáveis do citoesqueleto deste organismo (GERMOT et al., 1996).
1.1.6.2 Citoesqueleto
Os tricomonadídeos apresentam um citoesqueleto (FIGs. 1 e 2) formado
principalmente pelo complexo pelta-axóstilo, a costa, os filamentos parabasais, os corpúsculos
basais e filamentos associados, os filamentos sigmóides, o corpo infra e supra-cinetosomal.
Entretanto, ainda não se tem descrito a funcionalidade das duas últimas estruturas citadas.

7
Figura 1. Esquema de Tritrichomonas foetus mostrando as principais estruturas: FA: flagelos anteriores; A: axóstilo; CB: corpúsculo basal; C: costa; RE retículo endoplasmático; G: Golgi; H: hidrogenossomos; L: lisossomos; N: núcleo; Nu: nucléolo; P: pelta; FP: filamento parabasal; FR: flagelo recorrente; MO: membrana ondulante; V: vacúolo; Gl: glicogênio (Retirado de BENCHIMOL, 2004).
1.1.6.2.1 Complexo Pelta-axóstilo
O complexo pelta-axóstilo é uma estrutura formada por microtúbulos estáveis
(RIBEIRO et al., 2000), embora alguns trabalhos admitam que essas estruturas se
despolimerizem (VISCOGLIOSI & BRUGEROLLE, 1994). Além disso, o complexo pelta-
axóstilo possui uma participação importante no processo de divisão celular, promovendo a
constrição do núcleo na etapa de cariocinese (RIBEIRO et al., 2000).

8
O axóstilo consiste em uma fita disposta longitudinalmente desde a porção anterior
até o final da porção posterior da célula, onde esta estrutura empurra a membrana plasmática
(BENCHIMOL et al., 2000). A pelta (FIG. 1, P), também formada por microtúbulos está
localizada na porção anterior da célula. Esta estrutura parece desempenhar o papel de
sustentação da parede do canal periflagelar, do qual os flagelos emergem.
1.1.6.2.2 Filamentos Parabasais
Os filamentos parabasais (FIGs. 1 e 2, FPs) estão presentes em todos os
tricomonadídeos e são encontrados próximos à face Cis do complexo de Golgi
(HONIGBERG & BRUGEROLLE, 1990). São estruturas formadas por polímeros que
possuem uma periodicidade com bandas claras e escuras alternadas, apresentando assim, um
aspecto de fibra estriada (VISCOGLIOSI & BRUGEROLLE, 1994). A associação destes
filamentos com o complexo de Golgi forma o aparelho parabasal (HONIGBERG &
BRUGEROLLE, 1990). Além disso, verificou-se uma conexão estrutural entre a primeira
cisterna do complexo de Golgi e esta estrutura periódica, sugerindo um papel funcional desses
filamentos na migração do Golgi, junto com os corpúsculos basais e os flagelos, durante a
mitose (BENCHIMOL et al., 2001).
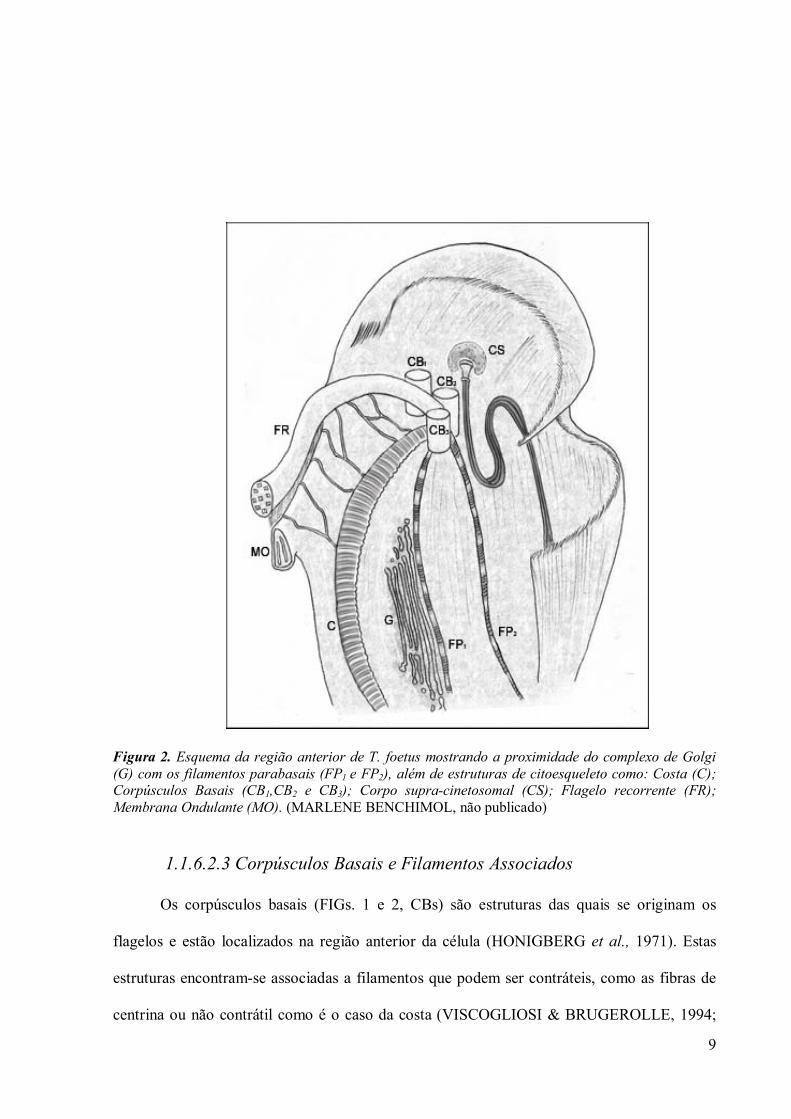
9
Figura 2. Esquema da região anterior de T. foetus mostrando a proximidade do complexo de Golgi (G) com os filamentos parabasais (FP1 e FP2), além de estruturas de citoesqueleto como: Costa (C); Corpúsculos Basais (CB1,CB2 e CB3); Corpo supra-cinetosomal (CS); Flagelo recorrente (FR); Membrana Ondulante (MO). (MARLENE BENCHIMOL, não publicado)
1.1.6.2.3 Corpúsculos Basais e Filamentos Associados
Os corpúsculos basais (FIGs. 1 e 2, CBs) são estruturas das quais se originam os
flagelos e estão localizados na região anterior da célula (HONIGBERG et al., 1971). Estas
estruturas encontram-se associadas a filamentos que podem ser contráteis, como as fibras de
centrina ou não contrátil como é o caso da costa (VISCOGLIOSI & BRUGEROLLE, 1994;

10
BRUGEROLLE et al., 2000). Muitos outros filamentos associados aos corpúsculos basais
estão dispostos em lamelas em forma de ganchos.
A costa é uma estrutura protéica estriada, que se estende da região do corpúsculo basal
do flagelo recorrente até a região posterior do parasito (BENCHIMOL et al., 1993). Está
presente somente nos tricomonadídeos que possuem membrana ondulante. Devido a isto,
acredita-se que sua função esteja relacionada com o suporte do estresse mecânico e a
sustentação da mesma (HONIGBERG et al., 1971).
1.1.6.2.4 Flagelos
Os flagelos dos tricomonadídeos apresentam variações quanto ao número dentre as
diferentes espécies desse grupo. T. foetus possui três flagelos anteriores (FIG. 1, FA) e um
recorrente (FIGs. 1 e 2, FR). Os flagelos anteriores emergem da célula pelo canal flagelar,
enquanto o flagelo recorrente surge da abertura latero-dorsal deste canal (WARTON &
HONIGBERG, 1979; BENCHIMOL, 2004).
1.1.6.3 Vacúolos e Lisossomos
A capacidade endocítica de T. foetus vem sendo investigada com o uso de diversos
marcadores como Lucifer yellow, peroxidase e proteínas conjugadas com partículas de ouro
(albumina, lactoferrina, transferrina, lectinas) (BENCHIMOL et al., 1986, 1990; AFFONSO
et al., 1994, 1997).
Duas vias endocíticas, uma mediada por receptores e outra de fase fluida, foram
caracterizadas em T. foetus através da ligação específica e da internalização da lactoferrina
(BRUGEROLLE, 1971; PETERSON & ALDERETE, 1984; AFFONSO, et al., 1997). Por
outro lado, a fagocitose em T. foetus tem sido estudada através da interação deste parasito
com outras células como hemácias (DE CARLI et al., 2004) e espermatozóides

11
(BENCHIMOL et al., 2007). Nestes trabalhos, T. foetus mostrou ser capaz de estabelecer a
adesão com estas células e posteriormente internalizá-las.
Os lisossomos (FIG. 1, L) em tricomonadídeos estão presentes como grandes
compartimentos citoplasmáticos, os quais participam da atividade endocítica, e demonstram
reação positiva em ensaios citoquímicos para fosfatase ácida (BENCHIMOL, 1999). Apesar
de não existir uma região preferencial para a ocorrência da endocitose, os lisossomos e
fagolisossomos encontram-se localizados predominantemente na região posterior dos
tricomonadídeos (AFFONSO et al., 1994, 1997; BENCHIMOL & DE SOUZA, 1995).
1.1.6.4 Hidrogenossomos
Os tricomonadídeos são organismos que não apresentam mitocôndrias, porém
apresentam organelas chamadas hidrogenossomos (FIGs.1, 3a, H), responsáveis pelo
metabolismo energético destes organismos. Esta organela foi descrita em T. foetus como um
compartimento subcelular capaz de produzir hidrogênio molecular e ATP (LINDMARK &
MÜLLER, 1973).
Os hidrogenossomos encontram-se geralmente associados a grânulos de glicogênio e a
estruturas do citoesqueleto, como o axóstilo e a costa (BENCHIMOL & DE SOUZA, 1983;
BENCHIMOL et al., 1996; BENCHIMOL et al., 2000). A associação dos hidrogenossomos
com o retículo endoplasmático também já foi reportada, o que poderia ser um indício de
fornecimento de lipídios para o crescimento dos hidrogenossomos (BENCHIMOL et al.,
1996; BENCHIMOL et al., 2000). A proximidade dos hidrogenossomos com outras estruturas
celulares poderia estar relacionada ao fornecimento de ATP, tal como ocorrem com a
associação já descrita entre mitocôndrias e microtúbulos em células de eucariotos superiores
(HEGGENESS et al., 1978).

12
Alguns autores consideram esta organela como uma variação das mitocôndrias que se
adaptaram a vida anaeróbica (BIAGINI et al., 1997; EMBLEY et al., 2003).
Entre as semelhanças já apresentadas pode-se destacar a participação na produção de
ATP pela degradação do piruvato (LINDMARK & MÜLLER, 1973; MÜLLER, 1993, BUI et
al., 1996), assumindo o papel energético tanto nas mitocôndrias quanto nos hidrogenossomos
e a presença da dupla membrana (BENCHIMOL et al., 1982b), aproximando estas duas
organelas em relação à mesma origem evolutiva. Além disso, a divisão dos hidrogenossomos
pode ocorrer por partição ou por segmentação, assim como nas mitocôndrias. Na partição, a
divisão se inicia com uma invaginação da membrana interna do hidrogenossomo, formando
um septo transversal que separa a matriz da organela em dois compartimentos. Já no processo
de segmentação ocorre primeiro o alongamento da organela e a formação de uma constrição
na sua porção central (BENCHIMOL et al., 1996). Recentemente, outras semelhanças foram
identificadas, tais como: a presença de translocases na membrana interna dos
hidrogenossomos de tricomonadídeos (SUTAK et al., 2004), a presença de centros de Fe-S
(Centros de ferro e enxofre, DOLEZAL et al., 2005) semelhantes aos mitocondriais e a
presença de cardiolipina (DE ANDRADE ROSA et al., 2006), fosfolipídio típico de
membranas bacterianas e da membrana mitocondrial interna.
Entretanto, os hidrogenossomos e as mitocôndrias diferem em alguns aspectos. Ainda
não foram descritas em hidrogenossomos a presença de citocromos e nem a atividade de F0-F1
ATPase (LLOYD et al., 1979). A presença de um material genético apesar de ter sido
sugerida inicialmente (CERKASOVOVÁ et al., 1976), não foi identificada por técnicas de
fluorescência (TURNER & MULLER, 1983) ou de nick-translation (CLEMENS &
JOHNSON, 2000), sugerindo que, possivelmente, tenha ocorrido uma evasão total para o
compartimento nuclear (JOHNSON et al., 1993).

13
Porém, a origem dos hidrogenossomos ainda é um assunto de grande debate, sendo
que no momento os autores apresentam duas vertentes. A primeira defende que os
hidrogenossomos teriam surgido a partir de um ancestral mitocondrial de metabolismo
aeróbico, o qual utilizava o oxigênio como aceptor final enquanto a outra sugere que essa
organela funcionava em condições anaeróbicas e, consequentemente produziam hidrogênio,
como ocorre com os hidrogenossomos (TJADEN et al., 2004). Com isso, inúmeros estudos
têm sido desenvolvidos com o intuito de procurar semelhanças entre os hidrogenossomos e as
mitocôndrias, tanto ao nível bioquímico, molecular, quanto ao nível ultraestrutural.
1.1.6.5 Núcleo e Divisão
Os tricomonadídeos quando não estão em processo de divisão apresentam um único
núcleo (FIGs. 1, 3a, N) localizado preferencialmente na região anterior da célula. O envoltório
nuclear é formado por duas membranas, as quais apresentam complexos dos poros
característicos (BENCHIMOL et al., 1982a). A matriz nuclear possui filamentos organizados
diferentes dos filamentos presentes em outros organismos, onde a região abaixo do envoltório
nuclear apresenta uma malha mais frouxa que a região central (RIBEIRO, 1997).
Estes protozoários apresentam um tipo diferente de divisão, chamada de mitose
fechada. A mitose dos tricomonadídeos é caracterizada por não apresentar fragmentação do
envoltório nuclear e pela presença de um fuso extranuclear, o qual, aparentemente, não
estabelece contato direto com os cromossomos (BRUGEROLLE et al., 1974). Segundo
Ribeiro e colaborados (2000), T. foetus possui seis fases distintas da mitose. (1) A intérfase
(G1), onde o parasito apresenta aspecto pirifome e estruturas ainda não duplicadas; (2) a pré-
mitose (S/G2) que compreende a etapa em que todas as estruturas de citoesqueleto e o
material genético são duplicados. O complexo pelta-axóstilo não desaparece durante a mitose,
pois tem um papel fundamental na mudança de forma da célula e na cariocinese; (3) na

14
prófase, ocorre um aumento de volume da célula; (4) a metáfase é caracterizada pela migração
dos corpúsculos basais para pólos opostos na célula, fazendo com que ocorra a migração dos
flagelos e dos complexos pelta-axóstilo. Devido à migração destas estruturas, o protozoário
assume uma forma triangular. A formação da placa metafásica não foi observada até o
momento, permanecendo uma incógnita sobre o que acarretaria a segregação dos
cromossomos; (5) na transição metáfase-anáfase o batimento dos flagelos auxilia no
deslocamento dos axóstilos para sentidos opostos, posicionados de forma cruzada no interior
da célula. Desta forma, seria possível promover a torção da mesma e o estrangulamento do
núcleo, acarretando a citocinese; (6) a transição anáfase-telófase: é a etapa em que ocorre o
afastamento dos núcleos e a separação das duas células-filhas.
1.1.6.6 Retículo Endoplasmático
O retículo endoplasmático de tricomonadídeos é normalmente visualizado ao redor do
núcleo formando a membrana externa do envoltório nuclear (QUEIROZ et al., 1991). Esta
organela pode ser encontrada também próxima aos hidrogenossomos (HONIGBERG, 1978;
BENCHIMOL et al., 1996, 2000), e ao axóstilo (BENCHIMOL et al., 2000). Os ribossomos
podem ser visualizados aderidos à membrana do retículo endoplasmático, formando o retículo
endoplasmático rugoso, ou livres no citoplasma (BENCHIMOL et al., 2001) (FIG. 3b).
Evidências têm demonstrado a sua participação também em processos de autofagia e
seqüestro de cálcio (BENCHIMOL et al., 1996; BENCHIMOL, 1999; DE SOUZA &
BENCHIMOL, 1988).
Durante a mitose dos tricomonadídeos, o retículo endoplasmático se alinha de modo
paralelo com os microtúbulos do fuso mitótico, sugerindo um recurso de fornecimento de
cálcio para a divisão celular do parasito (RIBEIRO et al., 2002).

15
1.1.6.7 Complexo de Golgi
1.1.6.7.1 Organização Estrutural
O complexo de Golgi é dotado de uma polaridade por possuir em cada pilha duas
faces distintas: uma face cis, também conhecida como face de entrada e uma face trans ou
face de saída. Ambas as faces apresentam-se intimamente associadas a compartimentos
especiais, compostos por uma rede interconectada de estruturas tubulares e de cisternas. Por
sua vez, as cisternas são agrupadas de acordo com a localização, morfologia e composição
química. Assim, as cisternas mais próximas do retículo endoplasmático (RE) e de
conformação convexa são denominadas cisternas cis, as posicionadas na região central do
Golgi são as cisternas médias e as mais côncavas e próximas do sítio de saída da célula, são
chamadas de cisternas trans (FIG. 3).
Além disso, existem compartimentos formados por estruturas membranosas tubulares
conectadas ou por vesículas, denominados rede Golgi cis (CGN) e rede Golgi trans (TGN). A
rede Golgi cis, também chamada de compartimento intermediário entre RE e Golgi (ERGIC)
está localizado entre o retículo endoplasmático (RE) e o sítio de entrada do Golgi. É a face de
entrada do Golgi, que recebe as proteínas recém sintetizadas do RE e as transporta para a
cisterna cis (DEAN & PELHAM, 1990; SEMENZA et al., 1990). Por outro lado, o TGN
segue as cisternas TRANS sendo o sítio de saída de sustâncias para outros compartimentos da
célula ou do meio extracelular (BERTACHINI-LOMBELLO et al., 2001).
As membranas dos diferentes compartimentos do complexo de Golgi diferem quanto a
sua composição lipídica e proteica. Entre as proteínas presentes no Golgi são encontradas
principalmente enzimas, proteínas estruturais e proteínas envolvidas na formação e
direcionamento de vesículas. Devido a esta diferença de conteúdo enzimático é possível

16
destacar algumas enzimas que são tidas como marcadoras por serem específicas de um
determinado compartimento (TEASDALE & JACKSON, 1996).
Figura 3. Esquema tridimensional do Complexo de Golgi mostrando os sub- compartimentos desta organela. A face cis corresponde a região mais próxima do retículo endoplasmático e a face trans a mais próxima da membrana plasmática (Adaptado de ALBETS et al. 2004).
1.1.6.7.2 Funções
O Complexo de Golgi é responsável por desenvolver importantes funções celulares,
tais como: (1) é um dos principais sítios de síntese de carboidratos, produzindo a maioria dos
polissacarídeos celulares. Além disso, sua posição na saída da rota do RE facilita a adição de
oligossacarídeos são adicionados como cadeias laterais em proteínas e lipídeos enviados pelo
RE; (2) é uma estação de classificação e endereçamento dos produtos sintetizados no retículo
endoplasmático, direcionando para a membrana plasmática ou para outras organelas; (3)
biogênese dos lisossomos; (4) acúmulo de cálcio (ALBERTS et al., 2004).
A adição de açúcares a proteínas e lipídeos durante o processo de glicosilação é de
extrema importância, pois é sabido que a presença de açúcares confere uma menor

17
flexibilidade e carga negativa, fundamental para a estrutura quartenária das proteínas. Além
disso, este processo é fundamental para dificultar a ação de enzimas proteolíticas (JENTOFT,
1990). A glicosilação do tipo N é iniciada no RE com a adição de uma cadeia de 14
oligossacarídeos que são transferidos em bloco único a um resíduo de asparagina. Esse
processo tem continuidade no Golgi com o objetivo de diferenciar as porções glicídicas das
diferentes glicoproteínas (ALBERTS et al., 2004). A glicosilação de oligossacarídeos O-
ligados também ocorre nesta organela, porém neste caso, os açúcares são adicionados um a
um a um radical OH lateral de um aminoácido serina ou treonina (JENTOFT, 1990).
As modificações protéicas ocorrem nos diferentes compartimentos do Golgi através do
transporte vesicular. Para isso, existem proteínas que auxiliam no direcionamento deste
transporte. O complexo de proteínas de revestimento do tipo II (COPII) gera vesículas que
realizam o movimento progressivo, ou seja, carreaiam vesículas oriundas do RE para o Golgi.
Por outro lado, o complexo de proteínas de revestimento do tipo I (COPI) permite o fluxo de
membranas através de um transporte retrógrado de vesículas que brotam do complexo de
Golgi e do ERGIC em direção ao retículo endoplasmático (HAURI & SCHWEIZER, 1992).
Normalmente, o estudo desse compartimento é realizado através da localização de
uma proteína residente, a proteína E53 (SCHWEIZER et al., 1988). Essa proteína cicla
rapidamente do RE para o ERGIC, atuando como um receptor para o transporte de
glicoproteínas realizando entre o RE e ERGIC (HAURI et al., 2000). Além disso, a fosfatase
ácida é usada como enzima marcadora deste compartimento (JENTOFT, 1990; TEASDALE
& JACKSON, 1996).
As vesículas que brotam do ERGIC se fundem com a primeira cisterna do Golgi, a
cisterna cis. Esta é caracterizada por ser o local das reações de fosforilação (ALBERTS et al.,
2004). Uma importante função desse processo está relacionada com a doação de fosfato a um
ou mais resíduos de manose das enzimas lisossomais para a formação de resíduos de manose-

18
6-fosfato. A presença de manose-6-fosfato em enzimas funciona como sinal, que ao ser
reconhecido por receptores são encaminhadas para os lisossomos. Nessa cisterna, duas
enzimas são consideradas marcadoras, as manosidases I e II (KORNFELD, 1987).
Nas cisternas da região medial a localização da manosidase III e da N-
acetilglicosamina transferase nestas cistenas permitem afirmar que a remoção de resíduos de
manose e a adição de N-acetilglicosamina são modificações restritas deste compartimento do
Golgi.
A reação de sulfatação ocorre a partir do doador de sulfato PAPS (3-fosfoadenosina-
5´-fosfosulfato) presente no lúmem das cisternas da região trans. O sulfato é adicionado aos
proteoglicanos, proteínas secretadas e a domínios extracelulares de proteínas e lipídeos da
membrana plasmática, conferindo a estes carga negativa (DICK et al.,, 2008). Assim como
nas outras cisternas, a participação da cisterna trans no processo de glicosilação foi
comprovada através da localização da galactositransferase e da sialiltransferase, enzimas que
adicionam galactose e ácido siálico, respectivamente as proteínas em processamento
(WEINSTEIN et al., 1982). Sugere-se que o lúmen da cisterna trans seja contínuo à rede trans
do Golgi, onde as proteínas são secretadas para dentro de pacotes de transporte e endereçadas
para seus destinos finais (GRIFFITHS & SIMONS,1986).
1.1.6.7.3 Complexo de Golgi em Tricomanadídeos
Sabe-se que o complexo de Golgi desempenha importantes funções celulares, como a
glicosilação do tipo N e O de proteínas e lipídios, endereçamento e biossíntese dos
lisossomos. Ainda existem poucos estudos relacionados a esta organela em tricomonadídeos
no que diz respeito à sua estrutura e função (BENCHIMOL & DE SOUZA, 1985; QUEIROZ
et al., 1991; DÍAZ & DE SOUZA, 1998; BENCHIMOL et al., 2001).

19
Nos tricomonadídeos, o complexo de Golgi é único e bastante proeminente (FIG. 4a),
ao contrário do que ocorre com outros protozoários parasitos como Toxoplasma e
Trypanosoma (HONIGBERG & BRUGEROLLE, 1990). Em tricomonas, esta organela pode
medir de 4,7-6µm de comprimento e 1-1,2µm de largura, apresentando 8 a 12 cisternas
alongadas e localiza-se na porção dorsal da célula e a direita do núcleo (FIGs. 4a - 4b).
Pequenas vesículas sem revestimento com 40 nm de diâmetro e outras com revestimento
medindo 75 nm de diâmetro foram descritas como associadas à porção lateral das cisternas do
Complexo de Golgi (BENCHIMOL et al., 2001).
Em tricomonas, os filamentos parabasais (FIGs. 2, 4a) encontram-se associados à esta
organela formando o aparelho Parabasal (HONIGBERG & BRUGEROLLE, 1990). Conexões
filamentosas foram observadas interligando a cisterna cis com os filamentos parabasais
(BENCHIMOL et al., 2001), sugerindo que este sistema possa proporcionar uma sustentação
às cisternas do Golgi (HONINGBERG & BRUGEROLLE, 1990).
Os filamentos parabasais são estruturas caracterizadas por apresentar uma
periodicidade semelhante à encontrada na costa. Entretanto, os filamentos parabasais diferem
por serem mais delgados e apresentam-se em número de dois, sendo assim denominados
filamentos parabasais 1 e 2 (FP1 e FP2) (HONINGBERG & BRUGEROLLE, 1990).
Estudos citoquímicos mostraram a presença das enzimas tiaminopirofosfatase e
fosfatase ácida nas cisternas cis e trans do Golgi de tricomonadídeos, repectivamente
(QUEIROZ et al., 1991; BENCHIMOL et al., 2001). Além disso, a técnica de Thiéry foi
capaz de demonstrar a presença de açúcares na membrana desta organela (BENCHIMOL &
DE SOUZA, 1985; BENCHIMOL et al., 2001). Posteriormente, com o uso de lectinas como
WGA (Wheat germ agglutinin) fluorescentes e conjugadas ao ouro coloidal foi possível
comprovar a presença de resíduos de N-acetil-glicosamina (BENCHIMOL et al., 2001;
BENCHIMOL & BERNARDINO, 2002). O emprego da técnica de iodeto de zinco-tetróxido

20
de ósmio (ZIO) proporcionou a visualização de fenestras nos sáculos do Golgi e
interconexões entre as cisternas do mesmo (BENCHIMOL & DE SOUZA, 1985;
BENCHIMOL et al., 2001). Por freeze-etching foi possível descrever estruturas filamentosas
conectando uma face à outra das cisternas do Golgi (BENCHIMOL et al., 1993).
Além disso, métodos bioquímicos foram capazes de revelar que o Golgi de tricomonas
desempenha um papel importante no acúmulo de cálcio, sendo o principal sítio com esta
função nestes organismos (ALMEIDA, 2000).
Ao longo do ciclo celular dos tricomonadídeos, foi observado o alongamento das
cisternas do Golgi embora o número de cisternas se mantenha constante. Antes da mitose, o
Golgi atinge um comprimento que pode variar de 4,7 a 6,0 µm e sofre uma fissão, dividindo
esta organela. A célula passa a ter duas pequenas pilhas de cisternas medindo entre 1,0 a 1,2
µm. Durante a mitose de tricomonas, o Golgi permanece íntegro e migra junto com a porção
anterior de cada axótilo, posicionando-se entre os corpúsculos basais e o núcleo. Durante esse
processo esta organela sofre um novo alongamento atingindo 3,7 µm, no final da telófase.
Além disso, proteínas, como as adesinas, foram detectadas ao longo da via secretora durante
todo o ciclo celular (BENCHIMOL et al., 2001).
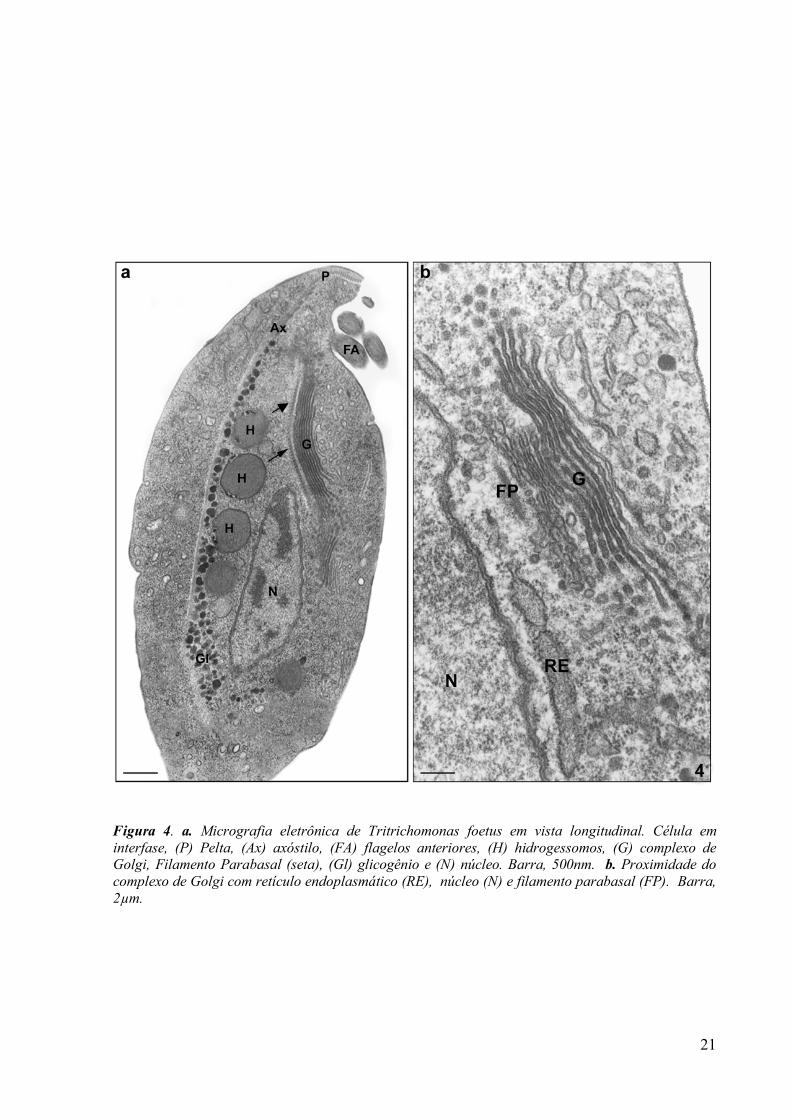
21
Figura 4. a. Micrografia eletrônica de Tritrichomonas foetus em vista longitudinal. Célula em interfase, (P) Pelta, (Ax) axóstilo, (FA) flagelos anteriores, (H) hidrogessomos, (G) complexo de Golgi, Filamento Parabasal (seta), (Gl) glicogênio e (N) núcleo. Barra, 500nm. b. Proximidade do complexo de Golgi com retículo endoplasmático (RE), núcleo (N) e filamento parabasal (FP). Barra, 2µm.

22
1.2 JUSTIFICATIVAS
O complexo de Golgi em Tritrichomonas foetus se destaca por ser bastante
desenvolvido, o que indica que deva exercer funções importantes para o parasito e na
parasitemia. Contudo, o assunto ainda é pouco estudado. Por isso, existe a necessidade de se
realizar estudos que ajudem a elucidar a funcionalidade, as proteínas aí presentes, e
comportamento desta organela e de seus componentes durante o ciclo de vida deste parasito.
1.2.1 Objetivo Geral
• Identificar pelo menos uma proteína do complexo de Golgi de T. foetus que possa servir
como proteína marcadora e assim, ser rastreada.
1.2.2 Objetivos Específicos
• Isolar o Complexo de Golgi de T. foetus
• Produzir anticorpos monoclonais contra proteínas do C. Golgi de T. foetus
• Rastrear por imunocitoquímica e imunofluorescência a sensibilidade e especificidade dos
anticorpos monoclonais gerados
• Identificar proteínas reconhecidas pelos anticorpos
• Identificar proteínas específicas de Golgi em Tricomonadídeos
• Verificar se as proteínas reconhecidas pelos anticorpos são conservadas, e, portanto, são
reconhecidas em outros organismos.

23
2. MATERIAL E MÉTODOS
2.1 Cultivo in vitro
Os protozoários da cepa K de Tritrichomonas foetus, utilizada neste trabalho, foi
isolada pelo Dr. Hélio Guida (Embrapa, Rio de Janeiro, Brasil) da cavidade prepucial de um
touro do estado do Rio de Janeiro, Brasil. A cepa JT de Trichomonas vaginalis foi isolada de
uma paciente atendida no Hospital Universitário Federal do Rio de Janeiro, Brasil.
Os trofozoítos foram cultivados axenicamente em meio de cultura TYM modificado
(Diamond, 1957), acrescido de 10% de soro fetal bovino inativado. As culturas foram
mantidas a 37ºC por 24 horas. Para o fracionamento subcelular foram cultivados 2 litros de T.
foetus, na densidade de 107 células/ml.
O meio TYM utilizado é composto por 22mg/ml de triptona, 11 mg/ml de extrato de
levedura, 5,6 mg/ml de maltose, 1mg/ml de L-cisteína, 0,2 mg/ml de ácido ascóbico, 0,9
mg/ml de fosfato de potássio monobásico e 0,9 mg/ml de fosfato de potássio dibásico. O pH é
ajustado para 6,2 com HCl 0,1M e o meio é posteriormente esterilizado por autoclação por 20
min.
As células epiteliais de rim de cachorro MDCKII foram doadas pelo Dr. José Andrés
Morgado Diaz (Departamento de Biologia Celular � INCA) e cultivadas em meio DMEM
(Sigma, USA) suplementado com 10% de soro fetal bovino. As culturas foram mantidas em
condições de 37ºC em uma atmosfera de 5% de CO2.
2.2 Fracionamento Subcelular
O isolamento do complexo de Golgi de T. foetus foi realizado de acordo com o
protocolo descrito por Díaz e De Souza (1998) (FIG. 5). Para isso foram cultivados dois litros

24
do parasito e todo o procedimento foi realizado a 4ºC. Os trofozoítos de T. foetus foram
centrifugados à 1.000x g por 10 minutos (ultracentrífuga Sorvall RCB com rotor GSA). As
células foram lavadas três vezes com tampão G (10mM tampão Tris-HCl, pH 7,4, contendo
0,25M sacarose e 2mM MgCl2) utilizando a mesma velocidade. As células foram ressuspensas
em solução hipotônica (tampão G sem sacarose) e, em seguida, foram rompidas
mecanicamente em homogenizador Potter. O monitoramento da ruptura das células foi
realizado com a observação do material ao microscópio de contraste de fase. O
homogeneizado foi centrifugado à 2.500x g por 10 minutos e o pellet contendo células não
rompidas, núcleo e hidrogenossomos foram descartados. O sobrenadante pós-nuclear (PNS)
resultante foi misturado em proporção de 1:1 a uma solução concentrada de sacarose a 2,3M
em tampão G. Essa suspensão foi colocada no fundo dos tubos da ultracentrífuga (Beckman
SW28). Soluções com concentrações decrescentes de sacarose foram adicionadas
sucessivamente ao tubo (1,2M, 1,0M e 0,8M em tampão Tris-HCl 10mM, pH 7,4). O
gradiente foi centrifugado a 95.000x g por 1 hora e 30 minutos na ultracentrífuga L8M usando
o rotor Beckman SW28 (Palo Alto, CA, USA). O pellet e as bandas formadas foram
cuidadosamente coletados com pipetas Pasteur e diluídos 10 vezes em tampão G sem
sacarose. O pellet, rico em complexos de Golgi, resultante da centrifugação a 80.000x g por
45 minutos, utilizando o rotor Beckman SW28, foi coletado para análise.
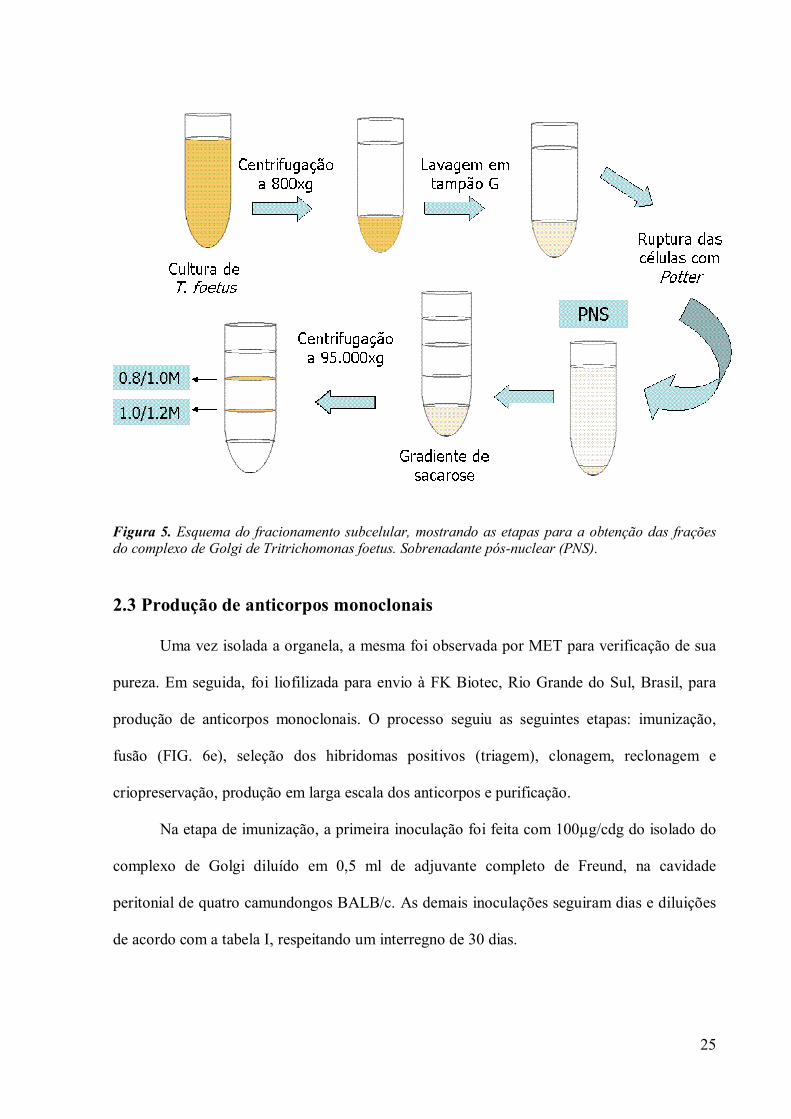
25
Figura 5. Esquema do fracionamento subcelular, mostrando as etapas para a obtenção das frações do complexo de Golgi de Tritrichomonas foetus. Sobrenadante pós-nuclear (PNS).
2.3 Produção de anticorpos monoclonais
Uma vez isolada a organela, a mesma foi observada por MET para verificação de sua
pureza. Em seguida, foi liofilizada para envio à FK Biotec, Rio Grande do Sul, Brasil, para
produção de anticorpos monoclonais. O processo seguiu as seguintes etapas: imunização,
fusão (FIG. 6e), seleção dos hibridomas positivos (triagem), clonagem, reclonagem e
criopreservação, produção em larga escala dos anticorpos e purificação.
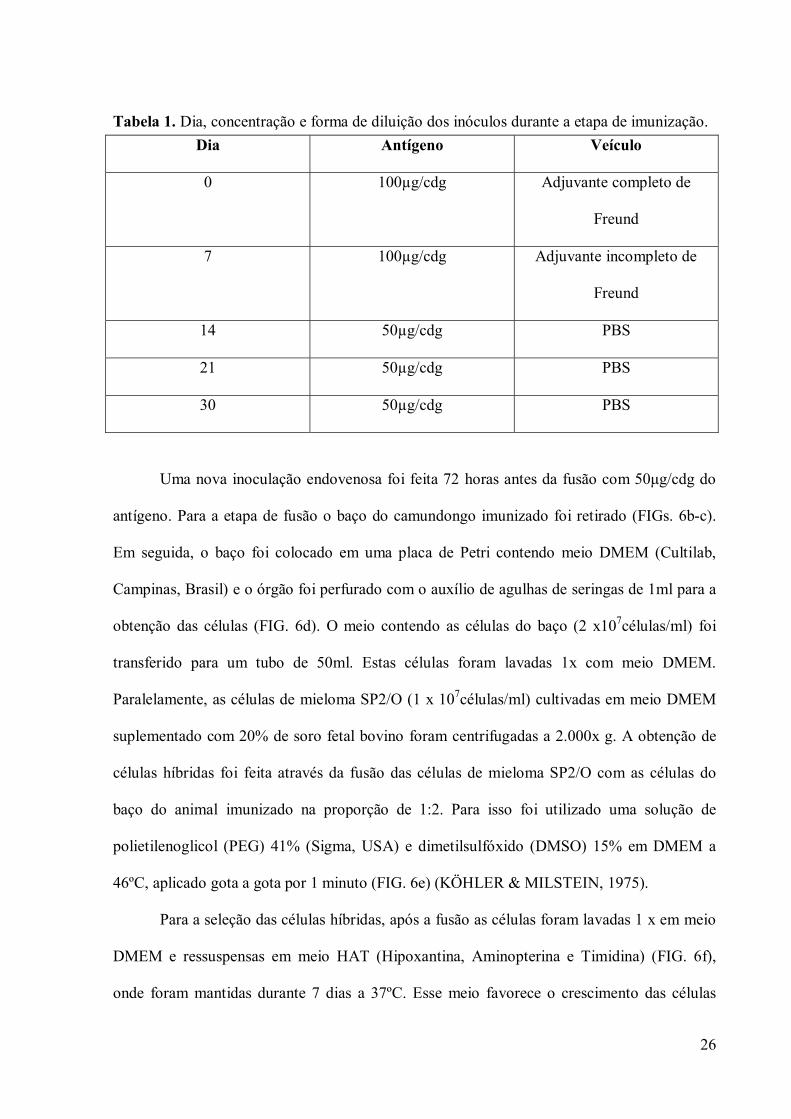
Na etapa de imunização, a primeira inoculação foi feita com 100µg/cdg do isolado do
complexo de Golgi diluído em 0,5 ml de adjuvante completo de Freund, na cavidade
peritonial de quatro camundongos BALB/c. As demais inoculações seguiram dias e diluições
de acordo com a tabela I, respeitando um interregno de 30 dias.
26
Tabela 1. Dia, concentração e forma de diluição dos inóculos durante a etapa de imunização.
Uma nova inoculação endovenosa foi feita 72 horas antes da fusão com 50µg/cdg do
antígeno. Para a etapa de fusão o baço do camundongo imunizado foi retirado (FIGs. 6b-c).
Em seguida, o baço foi colocado em uma placa de Petri contendo meio DMEM (Cultilab,
Campinas, Brasil) e o órgão foi perfurado com o auxílio de agulhas de seringas de 1ml para a
obtenção das células (FIG. 6d). O meio contendo as células do baço (2 x107células/ml) foi
transferido para um tubo de 50ml. Estas células foram lavadas 1x com meio DMEM.
Paralelamente, as células de mieloma SP2/O (1 x 107células/ml) cultivadas em meio DMEM
suplementado com 20% de soro fetal bovino foram centrifugadas a 2.000x g. A obtenção de
células híbridas foi feita através da fusão das células de mieloma SP2/O com as células do
baço do animal imunizado na proporção de 1:2. Para isso foi utilizado uma solução de
polietilenoglicol (PEG) 41% (Sigma, USA) e dimetilsulfóxido (DMSO) 15% em DMEM a
46ºC, aplicado gota a gota por 1 minuto (FIG. 6e) (KÖHLER & MILSTEIN, 1975).
Para a seleção das células híbridas, após a fusão as células foram lavadas 1 x em meio
DMEM e ressuspensas em meio HAT (Hipoxantina, Aminopterina e Timidina) (FIG. 6f),
onde foram mantidas durante 7 dias a 37ºC. Esse meio favorece o crescimento das células
Dia Antígeno Veículo
0 100µg/cdg Adjuvante completo de
Freund
7 100µg/cdg Adjuvante incompleto de
Freund
14 50µg/cdg PBS
21 50µg/cdg PBS
30 50µg/cdg PBS

27
híbridas, devido à ação da aminopterina, uma droga que age inibindo a síntese de DNA, pois
impede a produção de guanosina trifosfato (GTP) e timidina trifosfato (TTP). A produção de
TTP pode ser realizada através da adição de timidina no meio. Por outro lado, a síntese de
GTP só ocorre a partir da hipoxantina metabolizada pelos linfócitos B, através da atividade da
enzima hypoxantina guanina fosforibosil transferase (HGPRT). Apesar dos linfócitos
conseguirem sintetizar o DNA, estes morrem no período de 7 a 10 dias de cultura. Os
mielomas que não possuem esta enzima não conseguem produzir uma nova fita de DNA e não
sobrevivem. Desta forma, somente as células híbridas sobrevivem por herdarem a enzima
hypoxantina guanina fosforibosil transferase (HGPRT) dos linfócitos e a durabilidade dos
mielomas. Após o período de seleção das células híbridas, o meio HAT é substituído por meio
HT (Hipoxantina e Timidina) por mais 3 dias. As culturas seguintes foram realizadas em meio
DMEM suplementado com 20% de soro fetal bovino.
O teste de triagem foi realizado somente depois que as colônias de hibridomas
atingiram mais de 50% de crescimento em cada cavidade. Nesta etapa, os sobrenadantes das
culturas foram testados através do ensaio imunoenzimático direto (ELISA), para determinar
se as células estavam secretando imunoglobulinas de interesse. Os sobrenadantes que
apresentaram resultados positivos foram testados por imunofluorescência para verificar a
localização do reconhecimento do anticorpo e a intensidade desta fluorescência. Os
hibridomas secretores de anticorpos com especificidade para o complexo de Golgi foram
selecionados para a etapa de clonagem.
A clonagem consiste na realização de diluições seqüenciais de modo a permitir que,
matematicamente, uma única célula permanecesse em cada cavidade. Assim, os hibridomas
oriundos tratavam-se de clones de uma única célula e, conseqüentemente, os anticorpos
secretados reconheciam o mesmo epítopo, sendo, portanto monoclonais. Os melhores clones
foram selecionados após imunofluorescência e em seguida foram reclonados, repetindo as

28
diluições feitas para a obtenção dos clones. Posteriormente os mesmos foram expandidos,
congelados e armazenados. A etapa da produção de anticorpos em larga escala foi realizada in
vitro, através da coleta do sobrenadante do meio onde os hibridomas foram cultivados. Para a
etapa de purificação de anticorpos, foi necessária a utilização da técnica de cromatografia de
bioafinidade.
Figura 6. Etapas da produção de anticorpos monoclonais. a. Preparo do camundongo para a retirada do baço; b. Retirada do baço do camundongo imunizado; c. Baço do camundongo d. Extração das células do baço; e. Fusão de mielomas com células do baço; f. Plaqueamento das células híbridas.

29
2.4 C6-NBD-Ceramida
C6NBD-ceramida é um análogo fluorescente da ceramida, biologicamente ativo e
permeável as membranas (ROSENWALD & PAGANO, 1993). Depois de expostos a célula
estes lipídeos são acumulados no complexo de Golgi, onde são convertidos em
glicosilcerebrosídeos e esfingomielinas fluorescentes (LIPSKY & PAGANO,1985).
Neste estudo, utilizamos a C6NBD-ceramida (Molecular Probes Co, USA) para
demonstrar a localização do complexo de Golgi de T. foetus e de células MDCK e verificar se
os anticorpos monoclonais apresentam marcação na mesma região. Para isso, incubamos os
trofozoítos com 5µM de C6NBD-ceramida por 5 minutos à 37ºC. Em seguida, as células
foram centrifugadas a 1.000x g por 5 minutos. O pellet foi fixado com formaldeído 1% em
tampão fosfato 0,1 M, pH 7,4, por 1 hora. Depois, os núcleos das células foram corados com
DAPI (5µg/ml). As amostras foram observadas ao microscópio óptico de fluorescência Zeiss
Axophot II.
2.5 Imunofluorescência
Para os testes de imunofluorescência as células foram primeiramente fixadas com
formaldeído 4% em tampão fosfato 0,1M, pH 7,2, e aderidas a lamínulas recobertas com poli-
L-lisina 0,1% (Sigma, USA). A permeabilização das células foi feita com 0,1% de Triton X-
100 em PBS/BSA a 1%, por 10 minutos. Em seguida, as células passaram por uma etapa de
bloqueio com cloreto de amônio (NH4Cl) 50mM por 15min e depois com albumina bovina
(BSA) 3% por 15 minutos. Posteriormente, as células foram incubadas por 3 horas ou
overnight com os sobrenadantes dos hibridomas 20, 93, 110 e com o anticorpo primário
purificado anti-Golgi 20.3 aqui desenvolvidos, diluído 1:10 em PBS/BSA 1%. A revelação
dos anticorpos foi feita com a incubação da amostra com o anticorpo secundário anti-
camundongo conjugado com AlexaFluor 488, diluído 1:100, por 1 hora. Os núcleos das

30
células foram corados com DAPI (5µg/ml). As amostras foram observadas ao microscópio
óptico de fluorescência Zeiss Axophot II.
Para verificar se as proteínas reconhecidas pelo anticorpo purificado eram
conservadas, foram feitas imunofluorescências com T. vaginalis, formas epimatigota do
Trypanosoma cruzi e promastigota de Leishmania amazonensis, além de células MDCKII. Os
protozoários Trypanosoma cruzi e Leishmania amazonensis foram doados fixados com 4% de
formaldeído em tampão fosfato 0,1M, pH 7,2 pela Dra. Narcisa Leal da Cunha-e-Silva
(UFRJ, Rio de Janeiro, Brasil).
O controle foi realizado incubando as células somente com o anticorpo secundário.
2.6 Microscopia Eletrônica de Transmissão
2.6.1 Resinas hidrofílicas - Unicryl e LR-White
As células foram fixadas em uma mistura de 4% de formaldeído e 0,1% de glutaraldeído
grau I em tampão cacodilato de sódio a 0,1M, pH 7,2, overnight. As amostras foram lavadas
3x em PBS, pH 7,2 e 3x em H2O destilada. Para a obtenção de um melhor contraste do
material, foi feita a coloração em bloco em uma solução de acetato de uranila a 5% em água
destilada por 2 horas, protegido da luz. Após esse período, o material foi lavado em água
destilada e centrifugado a 1.500x g por 5 minutos com o objetivo de se obter um pellet
compacto. A amostra foi transferida para uma placa de Petri com etanol 70% e cortado em
pequenos fragmentos. A desidratação do material foi feita em concentrações crescentes de
etanol 70%, 90%, 100%, super-seco, por 15 minutos em cada etapa. A infiltração foi realizada
com concentrações crescentes de resina LR-White diluída em etanol super-seco, nas
proporções de 1:2, 1:1, 2:1 e puro, depois o material foi polimerizado em resina pura à -18ºC
sob luz UV.

31
Para imunocitoquímica de materiais polimerizados em resina LR-White, foram obtidos
cortes ultrafinos de 70nm de espessura que foram coletados em grades de níquel de 300 mesh.
As etapas de bloqueio foram realizadas com cloreto de amônio 50mM, pH 8,0, por 40
minutos, PBS/BSA 1%, PBS/BSA3% e Tween 0,2%, por 10 minutos em cada solução. As
células foram incubadas com os sobrenadantes dos hibridomas 20, 93, 110 e com o anticorpo
purificado anti-Golgi 20.3 diluído 1:10 em PBS/PBS 1%, overnight. O material foi lavado
com PBS/BSA 3%, 1%, 3% e incubado com o anticorpo secundário anti-camundongo IgG
conjugado com partículas de ouro coloidal de 10nm, diluído 1:100 em PBS/BSA 1%, por 50
minutos. O controle foi feito com a amostra que teve a etapa de incubação com o anticorpo
primário ocultada. Em seguida, o material foi novamente lavado em soluções de PBS/BSA
3%, 1%, PBS puro e H2O destilada. O material foi contrastado com acetato de uranila 5% e
citrato de chumbo e, posteriormente, observado ao microscópio eletrônico de transmissão
JEOL 1210.
2.6.2 Resina Hidrofóbica � EPON
Os trofozoítos de T. foetus K foram coletados por centrifugação e fixados em
glutaraldeído a 2,5% em tampão Cacodilato de sódio 0,1M, pH 7,2, por 2 horas ou overnight à
temperatura ambiente. Depois desse período, as células foram lavadas por três vezes em PBS,
pH 7,2 e pós-fixadas em solução de tetróxido de ósmio a 1% e tampão Cacodilato de sódio
contendo ferricianeto de potássio a 0,8% acrescido de CaCl2 5mM. Posteriormente, as células
foram desidratadas utilizando concentrações crescentes de acetona 70%, 90%, 100% por 15
minutos em cada concentração. O material foi, então, infiltrado em solução de resina Epoxy
Poly Bed 812 (Epon): acetona, nas proporções de 1:2, 1:1, 2:1 e Epon puro, por 3h ou
overnight em cada mistura. Finalmente, foi feita a polimerização do material na estufa a 60ºC
por 72 horas.

32
Depois dos blocos polimerizados, foram realizados cortes ultrafinos de 70 a 100nm,
utilizando o ultramicrótomo Leica Ultracut UCT. Os cortes foram coletados em grades de
cobre de 300 mesh ou em grades de níquel de 300 mesh quando o objetivo era fazer
imunocitoquímica. As amostras coletadas em grades de cobre foram contrastadas em acetato
de uranila 5% por 30 minutos e em citrato de chumbo por 10 minutos.
Para as imunocitoquímicas realizadas em materiais incluídos em resina epon foram feitas
algumas modificações no protocolo acima descrito. Cortes ultrafinos foram obtidos com
100nm de espessura e coletados em grades de níquel de 300 mesh. Foi necessário realizar uma
etapa de etching consistindo em uma retirada parcial da resina epon e uma �desosmificação�
com H2O2 1% por 10 minutos, antes da etapa de bloqueio (revisto por SKEPPER, 2000). O
material foi lavado exaustivamente em H2O destilada, seguida da etapa de bloqueio, como já
descrita anteriormente. Os tempos de incubação com os anticorpos primário e secundário
foram de 48 h e 2 h, respectivamente.
2.7 Técnicas Proteômicas (SOLOVIEV & FINCH, 2005)
Neste trabalho, utilizamos uma estratégia para identificação das proteínas do Golgi de
T. foetus, que foi feita através da seleção da proteína usando a técnica de imunoprecipitação
(IP). Para isso, utilizou-se o anticorpo específico para uma das proteínas do Golgi e proteína-
A conjugada à resina Sepharose. Em seguida, as proteínas capturadas foram separadas por
peso molecular através de um gel de poliacrilamida 12% SDS-PAGE. As bandas
correspondentes às proteínas do complexo de Golgi imunoprecipitadas foram extraídas e
tratadas com Ditiotreitol (DTT - agente redutor de pontes dissulfeto), iodoacetamida (IAA -
responsável pela alquilação das cisteínas) e digeridas pela ação da tripsina que cliva para a
conversão das proteínas em peptídeos. Posteriormente, as amostras foram analisadas pelo
espectrômetro de massa do tipo MALDI-TOF/TOF (FIG. 7).

33
Figura. 7. Estratégia para identificação das proteínas do complexo de Golgi. As proteínas do Golgi foram imunoprecipitadas(IP) e avaliadas por SDS-PAGE. As bandas correspondentes às proteínas do Golgi foram extraídas e digeridas com tripsina para obtenção de peptídeos, que foram analisados por MALDI-TOF/TOF. As informações obtidas a partir dos espectros de massas foram comparadas com seqüências depositadas em bancos de dados do MASCOT®.
2.8 Dosagens de Proteínas - Método Lowry-Peterson (PETERSON, 1983)
Para os experimentos bioquímicos, as frações isoladas do Golgi de T. foetus foram
tratadas com tampão de lise (20mM de Tris-HCl, 2mM de EDTA, 0,1% de SDS, 0,5% de
deoxicolato, 1% de Triton X-100, 137mM NaCl, 10% de glicerol, pH 7,2) e coquetel de
inubidores de protease 1x (Sigma, USA). Porém, alguns destes componentes são considerados
interferentes para a dosagem de proteína pelo método de Lowry, devido a isso, foi necessário
fazer, primeiramente, uma precipitação das proteínas.
Em eppendorfs de 1,5ml foram aplicados o 10µl do tampão de lise (branco), 10, 20,
30, 40 e 50µl de solução de albumina 1mg/ml (padrão) e 20µl da amostra. Em seguida, foram
adicionados 50µl da solução de deoxicolato 1,5%. A mistura foi mantida por 10 minutos à
temperatura ambiente. Após esse período, foram adicionados 50µl de ácido tricloroacético
72% (TCA) que ficou reagindo por 10 minutos à temperatura ambiente. As amostras foram
centrifugadas a 4.000x g por 25 minutos. O sobrenadante foi descartado e o pellet ressuspenso
em 400µl no reagente de formação do complexo (250µl de CuSO4 0,4%, 250µl de Tartarato
de potássio 0,8%, 500 µl de Na2CO3 20%, 1ml de SDS 10%, 1ml de NaOH 0,8M e 5ml de
água destilada) por 10 minutos. Em seguida, as amostras foram incubadas com 100µl do

34
reagente de Folin (1:6) por 60 minutos. A leitura foi feita a 650nm no leitor de ELISA
TermoMax microplate reader.
2.9 Imunoprecipitação
Para a reação de imunoprecipitação, a fração total do complexo de Golgi (1mg/ml) foi
incubada com 20µl de anticorpo 20.3 (1mg/ml), overnight à 4ºC (Pré-ligação).
Simultaneamente, em outro eppendorf foi feita a reação de pré-lavagem, incubando (1mg/ml)
com 50µl de proteína A-Sepharose (Sigma, USA) durante o mesmo tempo. Após esse
período, as amostras foram centrifugadas a 1.000x g por 10 minutos à 4ºC. Em seguida, foi
feita a incubação do sobrenadante da Pré-ligação e o pellet da pré-lavagem, overnight à 4ºC,
sob leve agitação. Posteriormente as amostras foram centrifugadas a 1.000x g por 10 min. O
pellet foi ressuspenso em 90µl de tampão de amostra (300 µl de tampão de amostra Laemmli
- Bio-Rad Laboratories, USA, 15 µl de β-mercaptoetanol e coquetel de inibidores de protease
1x concentrado - Sigma, USA), e fervido por 5 minutos para dissociação do complexo
proteico. A amostra foi centrifugada a 1.000x g por 5 minutos, para separar a Sepharose, e o
sobrenadante, contendo o anticorpo e o antígeno separados, que foi aplicado no gel de
Poliacrilamida a 12% como descrito abaixo.
2.10 Eletroforese em gel de poliacrilamida (SDS-PAGE) e Eletrotransferência
A eletroforese foi realizada de acordo com o procedimento descrito por Laemmli (1970).
O homogeinizado total de T. foetus e as frações do complexo de Golgi foram diluídos em
tampão de amostra (300 µl de tampão de amostra Laemmli - Bio-Rad Laboratories, USA, 15
µl de β-mercaptoetanol e coquetel de inibidores de protease 1x concentrado - Sigma, USA)

35
para uma concentração final de 5 ou 30µg/µl, dependendo da finalidade. Para as proteínas
imunoprecipitadas, foram aplicados 30µl da preparação descrita acima por poço do gel.
As proteínas foram separadas de acordo com o peso molecular em géis de Poliacrilamida
10% ou 12%, seguindo os seguintes parâmetros: 120V e 120 minutos. O peso molecular das
proteínas das amostras foram comparadas com o padrão kaleidoscope (Bio-Rad Laboratories,
USA). Algumas das preparações com 30µg/µl foram eletrotransferidas para membrana de
nitrocelulose ou PVDF utilizando o aparelho de transferência semi-seco a 10V por 60 minutos
como descrito por Towbin e colaboradores (1979). Após a transferência, as membranas de
nitrocelulose ou de PVDF foram coradas em uma solução de 0,2% Ponceau red e 3% de
ácido tricloroacético e em seguida, guardada à 4ºC.
2.11 Revelação do Gel
O mapa protéico resultante da eletroforese pode ser visualizado através de métodos de
coloração de proteínas. As proteínas no gel corado aparecem como bandas reveladas pelo azul
de Coomassie G-250 ou através da impregnação por prata.
2.11.1 Coomassie G-250 (NEUHOFF et al., 1988)
Os géis com 30 µg/µl de proteína foram colocados em solução fixadora contendo
etanol 30%, ácido fosfórico 2% (v/v) em água por 30 minutos. Esse procedimento foi repetido
três vezes. Em seguida, os géis foram lavados 2x por 20 minutos em ácido fosfórico 2% (v/v)
em água. Essa solução foi substituída por uma solução de ácido fosfórico 2% (v/v), etanol (ou
metanol) 18%, sulfato de amônio 15% (p/v) em água durante 30 minutos. A coloração do gel
foi realizada com adição de 5 ml (1% do volume) de uma solução contendo 20g de azul de
Comassie G-250 por litro de água. Os géis foram mantidos nesta solução por 72 horas sob
agitação moderada. O excesso de corante foi removido com lavagens de água deionizada.

36
2.11.2 Impregnação pela Prata
O gel para visualização do perfil proteico de 5µg/µl de proteínas do homogeinizado
total de T. foetus e da fração do complexo de Golgi, obtida após isolamento, foi fixado em
uma solução contendo 50% de metanol, 10% de ácido acético, 10% de Fixative enhancer
(Bio-Rad Laboratories, USA) e 30% de H2O deionizada. Em seguida, o gel foi lavado duas
vezes por 10 minutos em H2O deionizada e corado pela prata com a mistura de 10 ml da
solução A (Bio-Rad Laboratories, USA) (1ml de Silver complex solution, 1ml de Reduction
moderation solution, 1ml de Image develoment reagent e 7ml de H2O deionizada) e 10ml da
solução B (5g de Development acelerator em 100ml de H2O deionizada). Quando a coloração
do gel se tornou satisfatória a reação foi interrompida com uma solução de ácido acético 5%.
2.12 Immunoblotting
A membrana de nitrocelulose com as proteínas eletrotransferidas foi lavada 2x com
tampão TBS-T (20mM de Tris-HCl, 137 mM de cloreto de sódio e 0,1% de Tween-20) para a
retirada do corante vermelho de Ponceau. Em seguida, foi bloqueada com leite desnatado
(Molico) a 5% diluído em tampão TBS-T, por 1 hora. A membrana de nitrocelulose foi
incubada com os anticorpos primários diluídos 1:1000 em TBS-T contendo 5% de leite
desnatado, overnight. Depois, foi feito um novo bloqueio e a membrana foi incubada com o
anticorpo secundário conjugado com peroxidase (Santa Cruz Biotechnology Inc, USA)
diluído 1:2000 em TBS-T contendo 5% de leite desnatado. A revelação foi feita com o kit de
quimioluminescência ECL (Amersham Biosciences, Sweden), por 3 minutos e com exposição
a um filme de Raio-X (KODAK, USA) por 60 segundos.

37
2.13 Espectrometria de Massa (MALDI-TOF e MALDI-TOF/TOF)
As bandas presentes no gel de imunoprecipitação correspondente às proteínas de
interesse foram extraídas do gel e passaram primeiramente por uma etapa de descoloração.
Nesta etapa, os fragmentos do gel foram colocados em uma solução contendo 50% de
acetonitrila e 25mM de bicarbonato de amônio por 24 hora, sob agitação. Depois, os
fragmentos do gel foram transferidos para tubos eppendorfs previamente tratados com
metanol para evitar a contaminação da amostra com polímeros. Foi adicionado 25µl da
solução A (10mM de DTT e 25mM de sulfato de amônio) por 1 hora à 56ºC, para ocorrer a
redução das pontes de disulfeto. Em seguida, a solução anterior foi removida e adicionou-se
25µl da solução B (55mM de iodoacetamida, 25mM de bicarbonato de amônio, 10mg/ml de
DTT) por 45 minutos protegido da luz, para bloquear as cisteínas livres.
Após essas etapas, os fragmentos do gel foram desidratados com 50µl de acetonitrila
100% por 10 minutos. Posteriormente, essa solução foi retirada e foi adicionado 10µl da
solução C (0,01µg/ml de tripsina diluída em 25mM de bicarbonato de amônio) por 20 horas à
37ºC. Após esse período, acrescentou-se 50µl da solução D (50% de acetonitrila e 5% de
ácido tricloroacético) por 15 minutos, no sonicador. As soluções obtidas foram reservadas em
outros tubos eppendorfs e esta etapa foi repetida mais duas vezes para que fosse evitada a
perda de qualquer material. A solução final contendo aproximadamente 110µl foi liofilizada
no SpeedVac, até que o material estivesse totalmente seco. Em seguida, as amostras foram
ressuspensas em 15 µl de água deionizada. Para concentrar a amostra e eliminar resíduos dos
solventes usados na extração de peptídeos do gel, as amostras foram passadas no Zip-Tip C18
(Millipore Corporation, USA).
As amostras foram misturadas com uma matriz sólida (alpha-cyano) nas proporções de
1:1 e somente 1µl dessas misturas foi aplicado nos spots das placas do MALDI-TOF/TOF a

38
fim de correlacionar os dados presentes nos espectros de MS/MS com uma possível
identificação protéica utilizamos o programa MASCOT® baseando as buscas no banco de
dados NCBInr (National Center for Biotechnology não redundante). Para tal, foram
estabelecidos os seguintes parâmetros: leitura dos peptídeos trípticos, permitindo uma única
falha de clivagem e sem restrições taxonômicas. A carboaminometilação em resíduos de
cisteína foi adotada como modificação fixa e a oxidação da metila como modificação
variável. A precisão das massas foi realizada dentro de 100ppm em MS/MS, estes parâmetros
foram baseados em outros trabalhos da área (DE JESUS et al., 2007; CUERVO et al., 2008).

39
3. RESULTADOS
3.1 Fracionamento Subcelular
O rompimento de T. foetus foi realizado no Potter e acompanhado por microscopia de
contraste de fase. O homogenizado foi considerado satisfatório para o fracionamento quando a
maioria das células (aproximadamente 90%) estava rompida. O sobrenadante pós nuclear
(PNS) e as frações das interfaces de 0,8-1,0M e 1,0-1,2M, correspondentes às frações
enriquecidas do complexo de Golgi foram obtidas por flotação no gradiente de sacarose.
Conseqüentemente, essas amostras foram processadas para microscopia eletrônica de
transmissão para averiguar a pureza de cada fração.
3.2 Caracterizações morfológicas da fração do complexo de Golgi
As frações referentes ao complexo de Golgi de T. foetus avaliadas por microscopia
eletrônica de transmissão apresentaram-se na sua maioria formada por membranas. Na análise
do sobrenadante pós nuclear (PNS), fração correspondente ao extrato celular antes de ser
submetido ao gradiente de sacarose, foi possível observar algumas cisternas ainda intactas
(FIG. 8). Por outro lado, nas frações obtidas das interfaces 0,8/1,0M (FIG. 9a e c) e 1,0/1,2M
(FIG. 9b e d) do gradiente de sacarose foram observadas amostras ricas em membranas, as
quais foram resseladas. Em maior aumento, foi possível observar detalhes destas membranas
e verificar que não apresentam uma forma definida (FIG. 9c e d, cabeças de seta). Nestas
frações enriquecidas no complexo de Golgi não foram observadas contaminações por
organelas como núcleo ou hidrogenossomos.
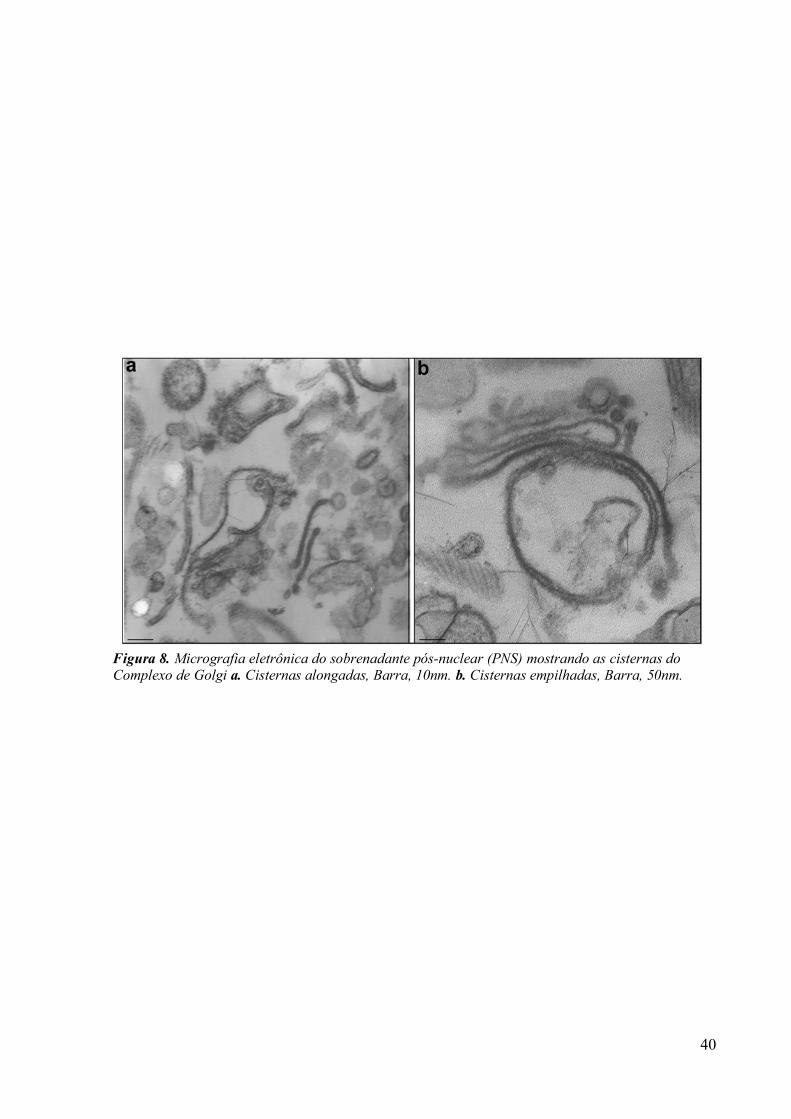
40
Figura 8. Micrografia eletrônica do sobrenadante pós-nuclear (PNS) mostrando as cisternas do Complexo de Golgi a. Cisternas alongadas, Barra, 10nm. b. Cisternas empilhadas, Barra, 50nm.
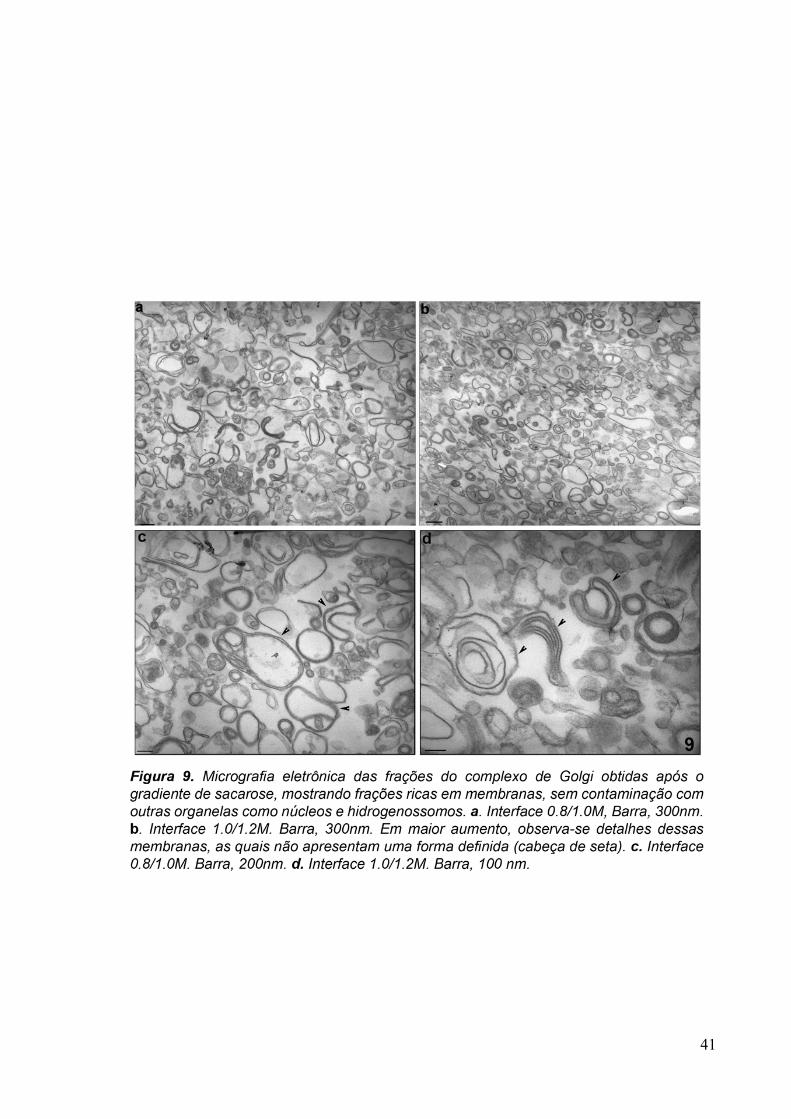
41

42
3.3 Obtenção de anticorpos monoclonais
A primeira etapa (triagem) de seleção para a obtenção dos anticorpos monoclonais foi
realizada através do teste de ELISA, pela empresa responsável pela produção destes. Nesta
etapa, o mesmo isolado do Golgi utilizado para imunizar os camundongos foi adsorvido as
microplacas. Esta análise resultou em 395 amostras positivas que foram avaliadas por
imunofluorescência para determinar a localização e a intensidade da fluorescência (Tabela 2).
Estas informações foram adicionadas a tabelas contendo o número de cada célula secretora.
Além disso, as amostras foram classificadas quanto à intensidade de fluorescência em uma
escala de um a seis (+). As amostras que não tiveram marcação foram representadas pelo
símbolo (-) e o símbolo (°) foi determinado para as marcações visualizadas em toda a célula.
Das 395 amostras positivas 94 apresentaram marcação para o complexo de Golgi com
intensidade igual ou maior que +++ que foram selecionados para a etapa de clonagem (Tabela
2, azul). Amostras (total = 5) que apresentaram marcação para o complexo de Golgi, porém
com baixa intensidade (++) também foram selecionadas devido a sua especificidade por esta
organela (Tabela 2, rosa). Amostras que apresentaram especificidade para o retículo
endoplasmático (total =12) foram escolhidas para a produção de anticorpos monoclonais
visando trabalhos futuros (Tabela 2, amarelo). O resultado desta seleção deu origem a 111
clones que passaram pelo mesmo processo de avaliação (Tabela 3). Assim, 17 sobrenadantes
que apresentaram marcação para complexo de Golgi com alta intensidade de fluorescência (>
+++) (Tabela 3, azul), 1 sobrenadante com marcação para o complexo de Golgi com baixa
intensidade fluorescência (Tabela 3, rosa) e 3 sobrenadantes com marcação para outras
estruturas celulares (membrana plasmática e retículo endoplasmático) (Tabela 3, amarelo)
foram selecionados. Como as marcações obtidas foram visualizadas em mais de uma
estrutura, foi necessário fazer uma reclonagem, com o objetivo de se obter uma única
organela marcada.

43
A etapa de reclonagem gerou 18 hibridomas reclonados, avaliados da forma
mencionada nas outras etapas (Tabela 4). Nesta etapa obtivemos 4 hibridomas com marcação
exclusivamente para o complexo de Golgi (amostras 20, 41, 78 e 93) e com a intensidade de
fluorescência alta (Tabela 4, azul). Além disso, duas outras amostras foram de nosso interesse.
A amostra (22) que apresentou marcação no retículo endoplasmático, complexo de Golgi e
vesículas (Tabela 4, rosa) e outra (69) com marcação para retículo endoplasmático e
envoltório nuclear (Tabela 4, amarelo). As células secretoras dos sobrenadantes responsáveis
por tais marcações foram selecionadas para a produção de anticorpos em larga escala. Três
hibridomas (99, 103 e 110) foram perdidos uma vez que estas células morreram ou pararam
de secretar imunoglobulinas.
Após a obtenção de 100 ml dos sobrenadantes dos hibridomas selecionados, estes
foram novamente testados para verificar se as células continuavam secretando
imunoglobulinas antes de passar para a etapa de purificação dos anticorpos (Tabela 5). Nesta
etapa selecionamos 2 sobrenadantes (20 e 69) para serem purificados. O sobrenadante 20
apresentou melhor intensidade de fluorescência com marcação para o complexo de Golgi.
Após a purificação originou o anticorpo monoclonal IgG anti-Golgi 20.3. Por outro lado, o
sobrenadante 69 deu origem aos anticorpos monoclonais IgG 69.1 e 69.2 que não foram
utilizados neste trabalho pois marcavam retículo, o que no momento não era do nosso
interesse.
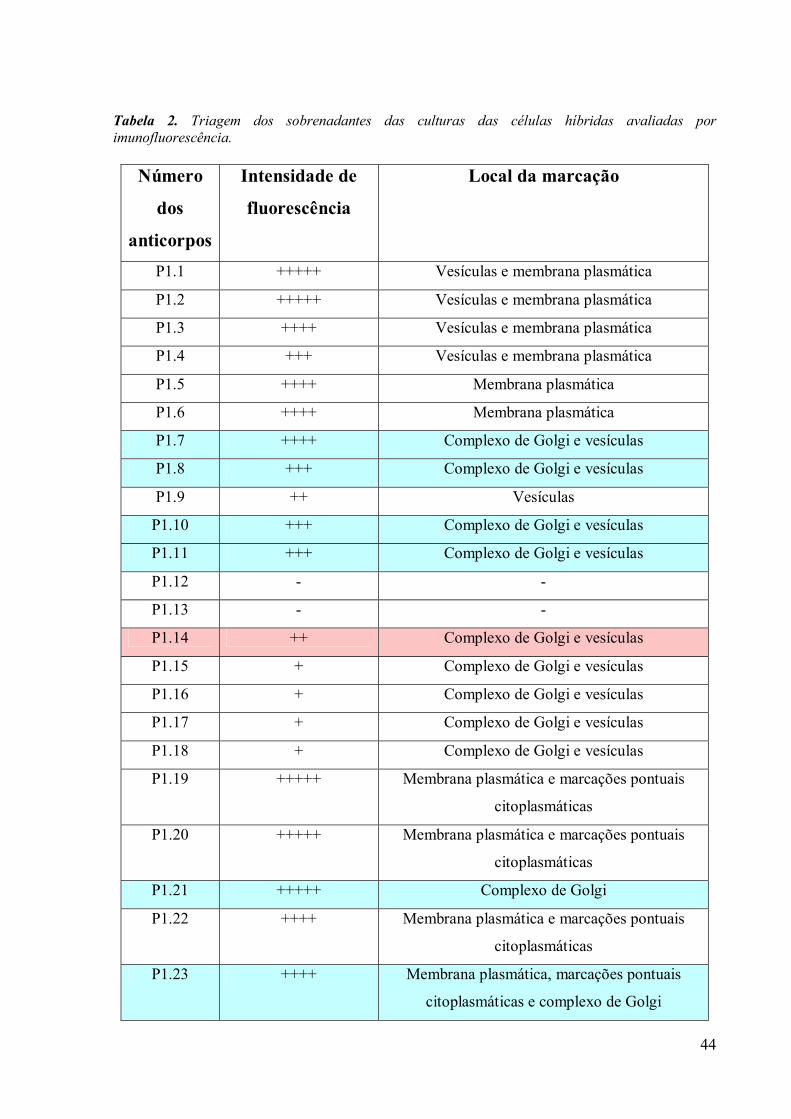
44
Tabela 2. Triagem dos sobrenadantes das culturas das células híbridas avaliadas por imunofluorescência.
Número
dos
anticorpos
Intensidade de
fluorescência
Local da marcação
P1.1 +++++ Vesículas e membrana plasmática
P1.2 +++++ Vesículas e membrana plasmática
P1.3 ++++ Vesículas e membrana plasmática
P1.4 +++ Vesículas e membrana plasmática
P1.5 ++++ Membrana plasmática
P1.6 ++++ Membrana plasmática
P1.7 ++++ Complexo de Golgi e vesículas
P1.8 +++ Complexo de Golgi e vesículas
P1.9 ++ Vesículas
P1.10 +++ Complexo de Golgi e vesículas
P1.11 +++ Complexo de Golgi e vesículas
P1.12 - -
P1.13 - -
P1.14 ++ Complexo de Golgi e vesículas
P1.15 + Complexo de Golgi e vesículas
P1.16 + Complexo de Golgi e vesículas
P1.17 + Complexo de Golgi e vesículas
P1.18 + Complexo de Golgi e vesículas
P1.19 +++++ Membrana plasmática e marcações pontuais
citoplasmáticas
P1.20 +++++ Membrana plasmática e marcações pontuais
citoplasmáticas
P1.21 +++++ Complexo de Golgi
P1.22 ++++ Membrana plasmática e marcações pontuais
citoplasmáticas
P1.23 ++++ Membrana plasmática, marcações pontuais
citoplasmáticas e complexo de Golgi
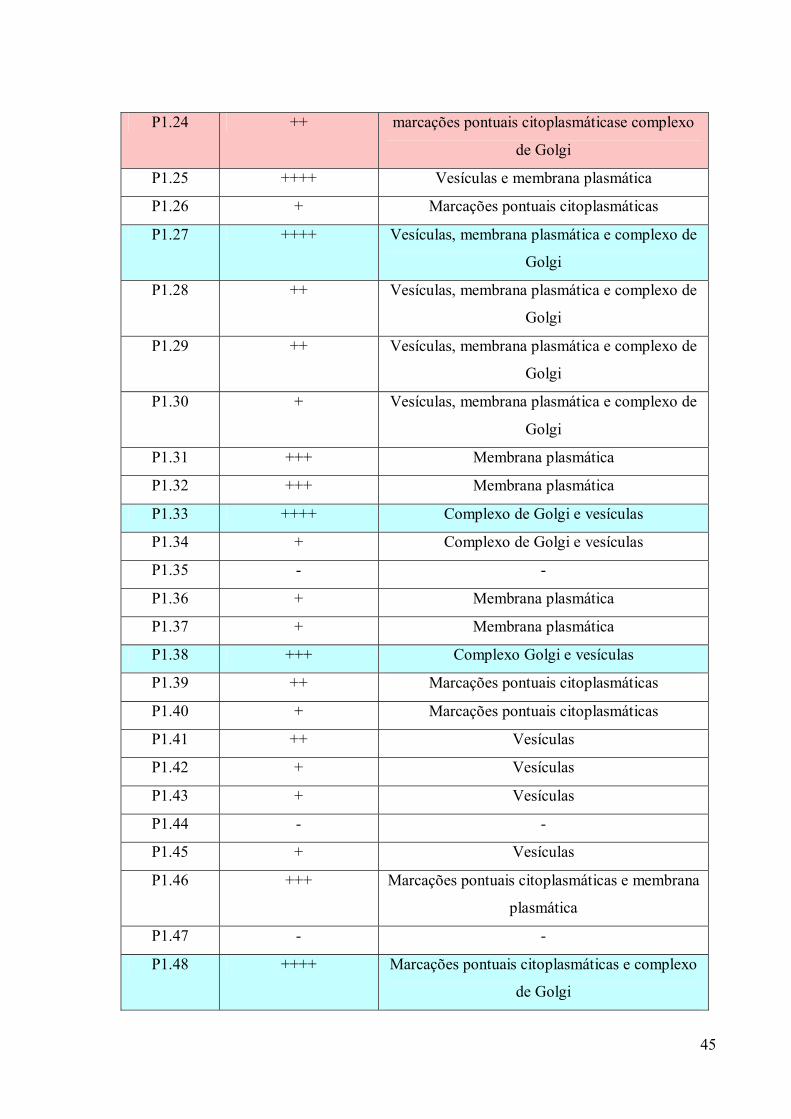
45
P1.24 ++ marcações pontuais citoplasmáticase complexo
de Golgi
P1.25 ++++ Vesículas e membrana plasmática
P1.26 + Marcações pontuais citoplasmáticas
P1.27 ++++ Vesículas, membrana plasmática e complexo de
Golgi
P1.28 ++ Vesículas, membrana plasmática e complexo de
Golgi
P1.29 ++ Vesículas, membrana plasmática e complexo de
Golgi
P1.30 + Vesículas, membrana plasmática e complexo de
Golgi
P1.31 +++ Membrana plasmática
P1.32 +++ Membrana plasmática
P1.33 ++++ Complexo de Golgi e vesículas
P1.34 + Complexo de Golgi e vesículas
P1.35 - -
P1.36 + Membrana plasmática
P1.37 + Membrana plasmática
P1.38 +++ Complexo Golgi e vesículas
P1.39 ++ Marcações pontuais citoplasmáticas
P1.40 + Marcações pontuais citoplasmáticas
P1.41 ++ Vesículas
P1.42 + Vesículas
P1.43 + Vesículas
P1.44 - -
P1.45 + Vesículas
P1.46 +++ Marcações pontuais citoplasmáticas e membrana
plasmática
P1.47 - -
P1.48 ++++ Marcações pontuais citoplasmáticas e complexo
de Golgi

46
P1.49 - -
P1.50 +++ Membrana plasmática e vesículas
P1.51 - -
P1.52 - -
P1.53 - -
P1.54 + Membrana plasmática e vesículas
P1.55 ++++ Membrana plasmática e vesículas
P1.56 +++ Vesículas e Membrana plasmática
P1.57 ++++ Complexo de Golgi e vesículas
P1.58 + Complexo de Golgi e vesículas
P1.59 ++++ Complexo de Golgi, vesículas e membrana
plasmática
P1.60 - -
P1.61 - -
P1.62 ++ -
P1.63 + Marcações pontuais citoplasmáticas e membrana
plasmática
P1.64 - -
P1.65 ++++ Complexo de Golgi e membrana plasmática
P1.66 ++++ Complexo de Golgi e membrana plasmática
P1.67 +++ Marcações pontuais citoplasmáticas e complexo
de Golgi
P1.68 - -
P1.69 - -
P1.70 ++++ Complexo de Golgi, vesículas e membrana
plasmática
P1.71 ++++ Complexo de Golgi, vesículas e membrana
plasmática
P1.72 ++ Complexo de Golgi, vesículas e membrana
plasmática
P1.73 +++ Complexo de Golgi, vesículas e membrana
plasmática
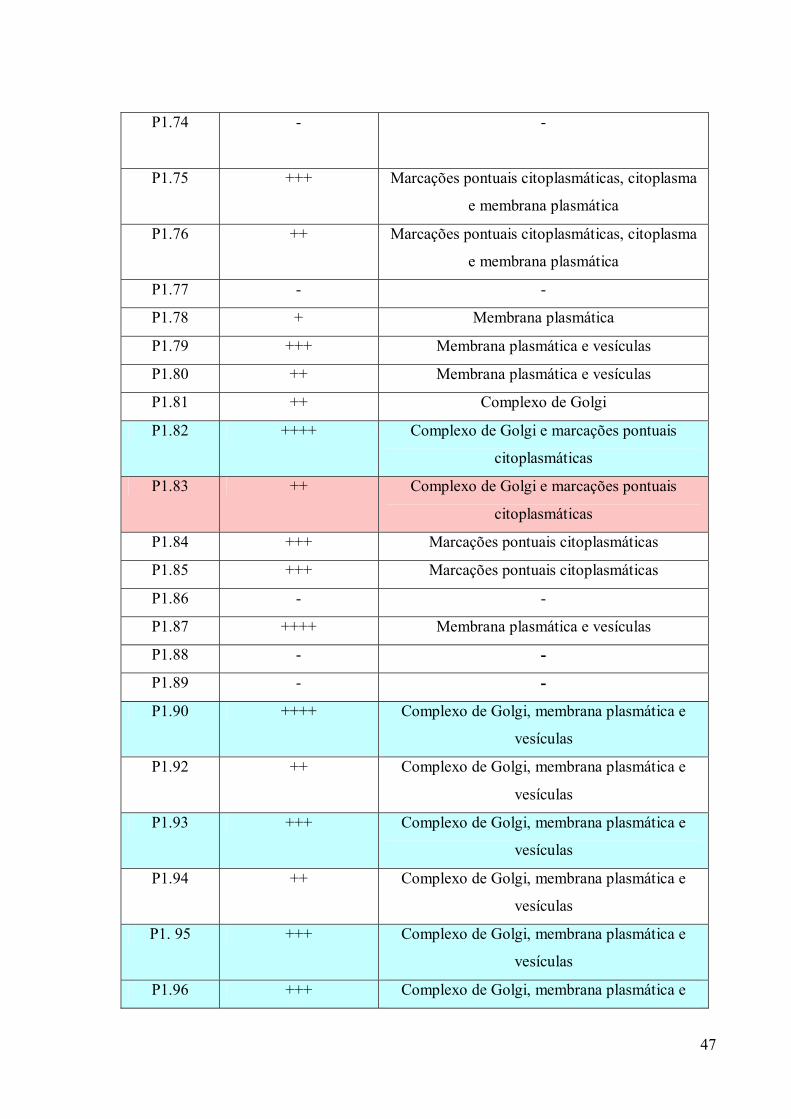
47
P1.74 - -
P1.75 +++ Marcações pontuais citoplasmáticas, citoplasma
e membrana plasmática
P1.76 ++ Marcações pontuais citoplasmáticas, citoplasma
e membrana plasmática
P1.77 - -
P1.78 + Membrana plasmática
P1.79 +++ Membrana plasmática e vesículas
P1.80 ++ Membrana plasmática e vesículas
P1.81 ++ Complexo de Golgi
P1.82 ++++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P1.83 ++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P1.84 +++ Marcações pontuais citoplasmáticas
P1.85 +++ Marcações pontuais citoplasmáticas
P1.86 - -
P1.87 ++++ Membrana plasmática e vesículas
P1.88 - -
P1.89 - -
P1.90 ++++ Complexo de Golgi, membrana plasmática e
vesículas
P1.92 ++ Complexo de Golgi, membrana plasmática e
vesículas
P1.93 +++ Complexo de Golgi, membrana plasmática e
vesículas
P1.94 ++ Complexo de Golgi, membrana plasmática e
vesículas
P1. 95 +++ Complexo de Golgi, membrana plasmática e
vesículas
P1.96 +++ Complexo de Golgi, membrana plasmática e

48
vesículas
P2.1 ++++ Membrana plasmática
P2.2 +++ Membrana plasmática e vesículas
P2.3 - -
P2.4 +++ Membrana plasmática
P2.5 ++++ º
P2.6 - -
P2.7 +++ º
P2.8 +++ Vesículas e retículo endoplasmático
P2.9 ++++ º
P2.10 ++++ º
P2.11 +++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P2.12 +++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P2.13 +++ Membrana plasmática
P2.14 ++++ º
P2.15 - -
P2.16 ++++ º
P2.17 +++++ Complexo de Golgi, membrana plasmática,
retículo endoplasmático e vesículas
P2.18 - -
P2.19 +++++ º
P2.20 +++++ º
P2.21 +++++ º
P2.22 ++ Complexo de Golgi, vesículas e retículo
endoplasmático
P2.23 + Complexo de Golgi, vesículas e retículo
endoplasmático
P2.24 +++ Complexo de Golgi, vesículas e retículo
endoplasmático
P2.25 - -
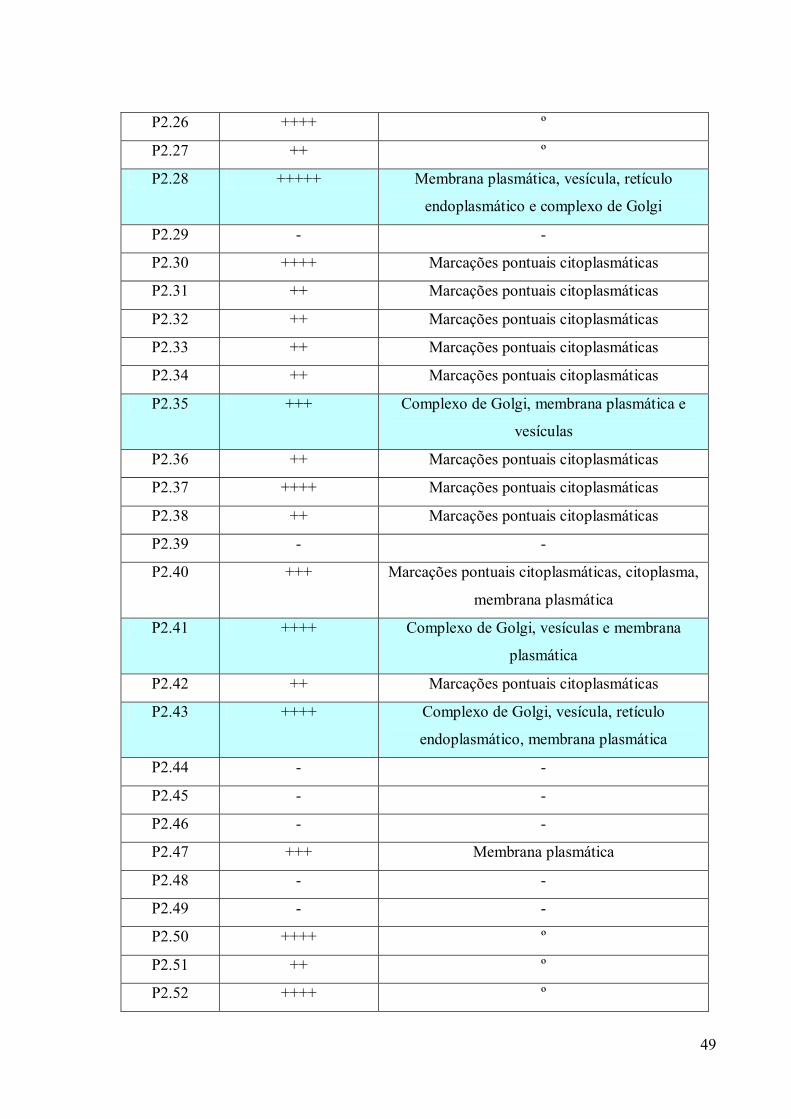
49
P2.26 ++++ º
P2.27 ++ º
P2.28 +++++ Membrana plasmática, vesícula, retículo
endoplasmático e complexo de Golgi
P2.29 - -
P2.30 ++++ Marcações pontuais citoplasmáticas
P2.31 ++ Marcações pontuais citoplasmáticas
P2.32 ++ Marcações pontuais citoplasmáticas
P2.33 ++ Marcações pontuais citoplasmáticas
P2.34 ++ Marcações pontuais citoplasmáticas
P2.35 +++ Complexo de Golgi, membrana plasmática e
vesículas
P2.36 ++ Marcações pontuais citoplasmáticas
P2.37 ++++ Marcações pontuais citoplasmáticas
P2.38 ++ Marcações pontuais citoplasmáticas
P2.39 - -
P2.40 +++ Marcações pontuais citoplasmáticas, citoplasma,
membrana plasmática
P2.41 ++++ Complexo de Golgi, vesículas e membrana
plasmática
P2.42 ++ Marcações pontuais citoplasmáticas
P2.43 ++++ Complexo de Golgi, vesícula, retículo
endoplasmático, membrana plasmática
P2.44 - -
P2.45 - -
P2.46 - -
P2.47 +++ Membrana plasmática
P2.48 - -
P2.49 - -
P2.50 ++++ º
P2.51 ++ º
P2.52 ++++ º
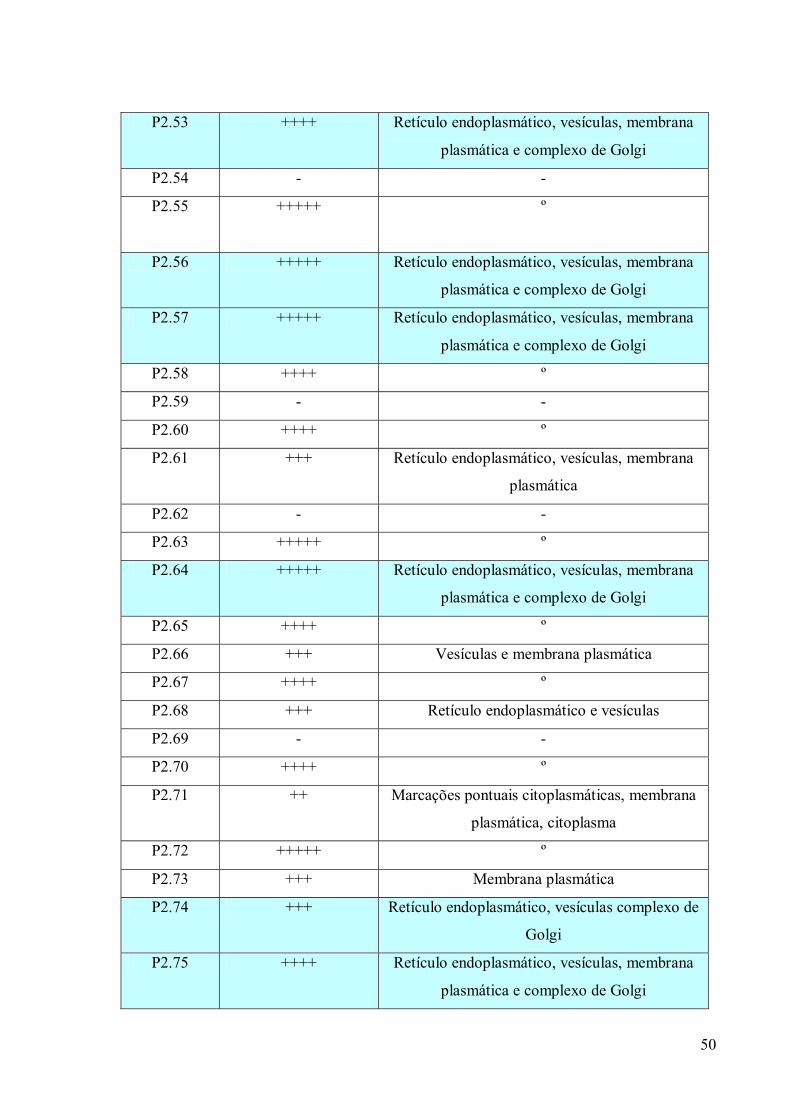
50
P2.53 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P2.54 - -
P2.55 +++++ º
P2.56 +++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P2.57 +++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P2.58 ++++ º
P2.59 - -
P2.60 ++++ º
P2.61 +++ Retículo endoplasmático, vesículas, membrana
plasmática
P2.62 - -
P2.63 +++++ º
P2.64 +++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P2.65 ++++ º
P2.66 +++ Vesículas e membrana plasmática
P2.67 ++++ º
P2.68 +++ Retículo endoplasmático e vesículas
P2.69 - -
P2.70 ++++ º
P2.71 ++ Marcações pontuais citoplasmáticas, membrana
plasmática, citoplasma
P2.72 +++++ º
P2.73 +++ Membrana plasmática
P2.74 +++ Retículo endoplasmático, vesículas complexo de
Golgi
P2.75 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
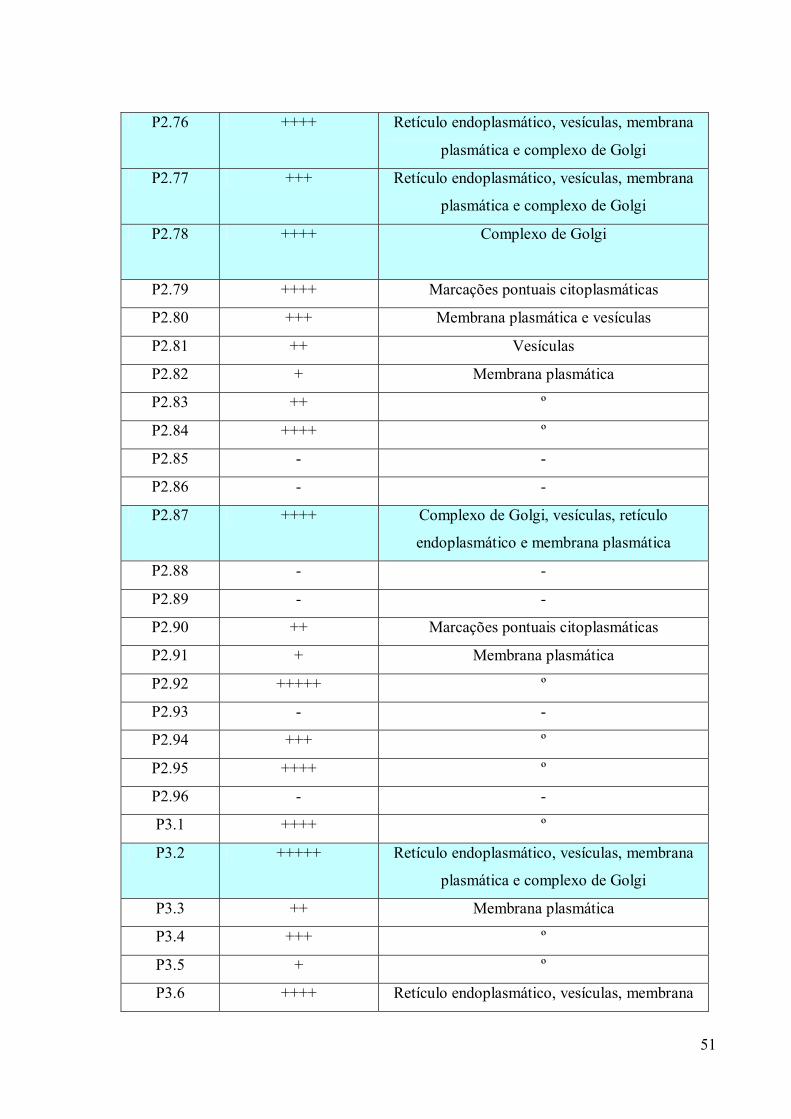
51
P2.76 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P2.77 +++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P2.78 ++++ Complexo de Golgi
P2.79 ++++ Marcações pontuais citoplasmáticas
P2.80 +++ Membrana plasmática e vesículas
P2.81 ++ Vesículas
P2.82 + Membrana plasmática
P2.83 ++ º
P2.84 ++++ º
P2.85 - -
P2.86 - -
P2.87 ++++ Complexo de Golgi, vesículas, retículo
endoplasmático e membrana plasmática
P2.88 - -
P2.89 - -
P2.90 ++ Marcações pontuais citoplasmáticas
P2.91 + Membrana plasmática
P2.92 +++++ º
P2.93 - -
P2.94 +++ º
P2.95 ++++ º
P2.96 - -
P3.1 ++++ º
P3.2 +++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.3 ++ Membrana plasmática
P3.4 +++ º
P3.5 + º
P3.6 ++++ Retículo endoplasmático, vesículas, membrana

52
plasmática
P3.7 +++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.8 - -
P3.9 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.10 - -
P3.11 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.12 +++ Marcações pontuais citoplasmáticas
P3.13 ++ Membrana plasmática
P3.14 - -
P3.15 +++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.16 +++ Marcações pontuais citoplasmáticas
P3.17 ++++ Retículo endoplasmático, vesículas e membrana
plasmática
P3.18 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.19 +++ Membrana plasmática
P3.20 +++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.21 ++ Membrana plasmática
P3.22 - -
P3.23 +++ Retículo endoplasmático, vesículas e membrana
plasmática
P3.24 ++ Vesículas
P3.25 + Marcações pontuais citoplasmáticas
P3.26 +++ Retículo endoplasmático e vesículas
P3.27 ++++ Retículo endoplasmático, vesículas, membrana
plasmática
P3.28 - -
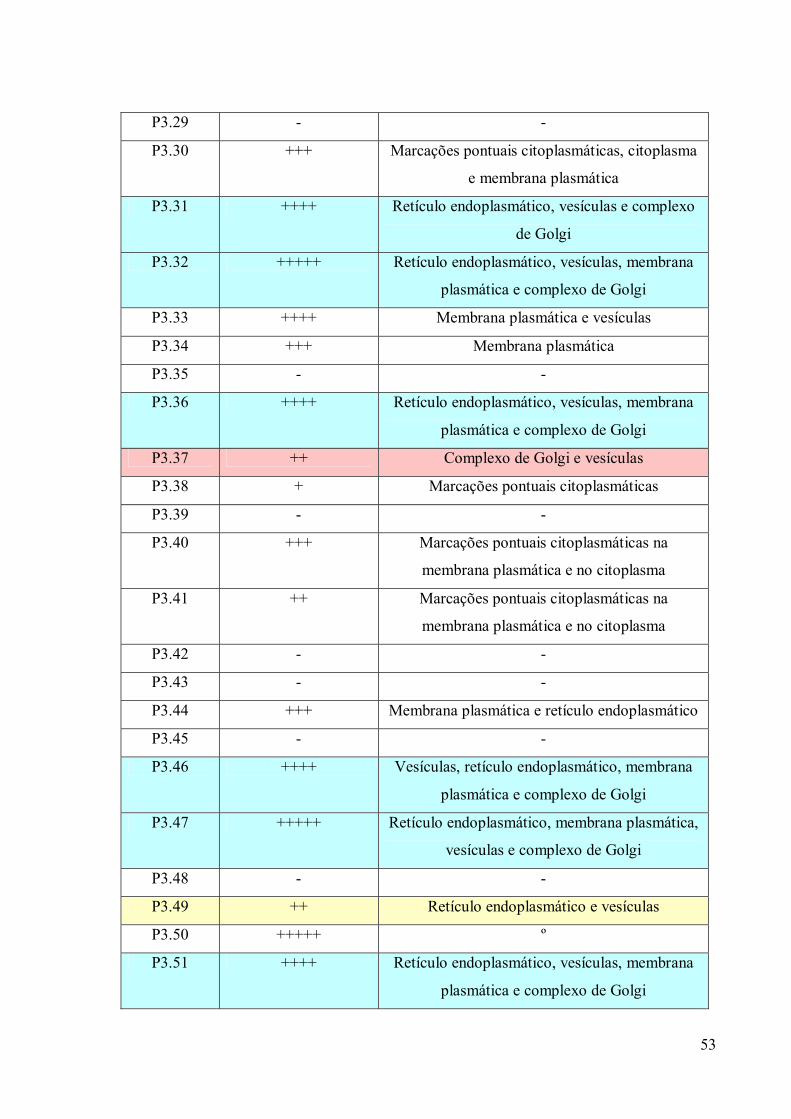
53
P3.29 - -
P3.30 +++ Marcações pontuais citoplasmáticas, citoplasma
e membrana plasmática
P3.31 ++++ Retículo endoplasmático, vesículas e complexo
de Golgi
P3.32 +++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.33 ++++ Membrana plasmática e vesículas
P3.34 +++ Membrana plasmática
P3.35 - -
P3.36 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
P3.37 ++ Complexo de Golgi e vesículas
P3.38 + Marcações pontuais citoplasmáticas
P3.39 - -
P3.40 +++ Marcações pontuais citoplasmáticas na
membrana plasmática e no citoplasma
P3.41 ++ Marcações pontuais citoplasmáticas na
membrana plasmática e no citoplasma
P3.42 - -
P3.43 - -
P3.44 +++ Membrana plasmática e retículo endoplasmático
P3.45 - -
P3.46 ++++ Vesículas, retículo endoplasmático, membrana
plasmática e complexo de Golgi
P3.47 +++++ Retículo endoplasmático, membrana plasmática,
vesículas e complexo de Golgi
P3.48 - -
P3.49 ++ Retículo endoplasmático e vesículas
P3.50 +++++ º
P3.51 ++++ Retículo endoplasmático, vesículas, membrana
plasmática e complexo de Golgi
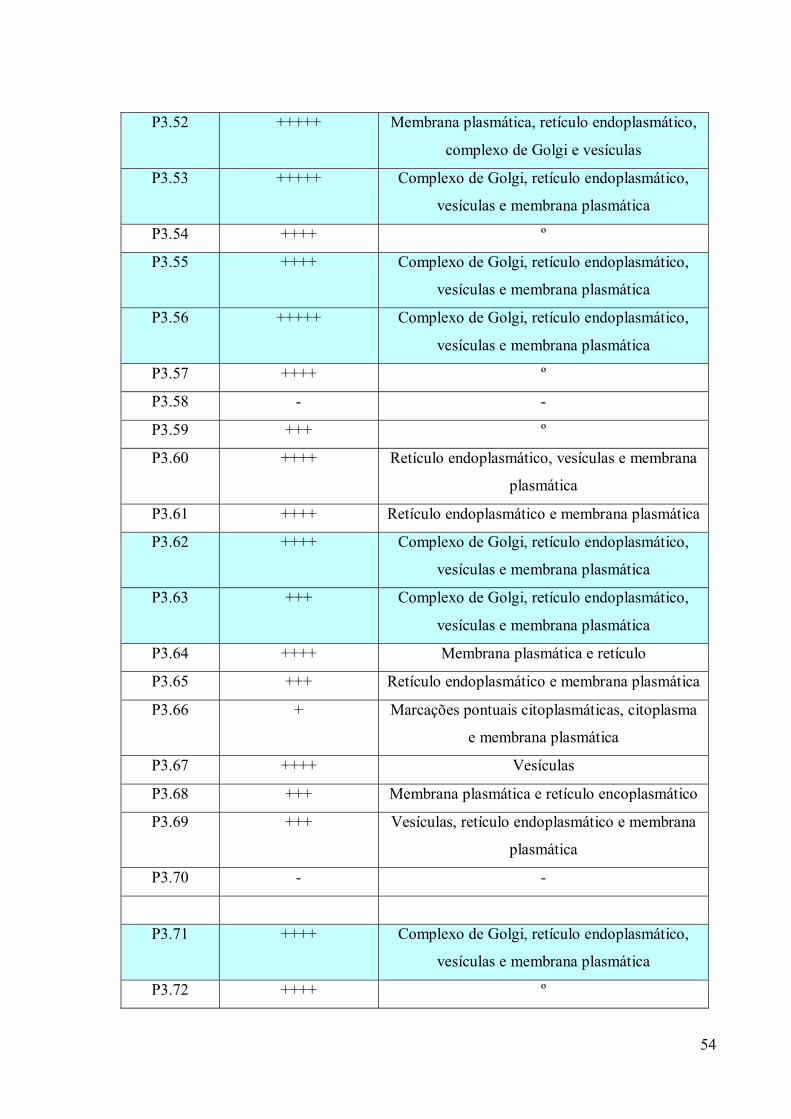
54
P3.52 +++++ Membrana plasmática, retículo endoplasmático,
complexo de Golgi e vesículas
P3.53 +++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P3.54 ++++ º
P3.55 ++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P3.56 +++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P3.57 ++++ º
P3.58 - -
P3.59 +++ º
P3.60 ++++ Retículo endoplasmático, vesículas e membrana
plasmática
P3.61 ++++ Retículo endoplasmático e membrana plasmática
P3.62 ++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P3.63 +++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P3.64 ++++ Membrana plasmática e retículo
P3.65 +++ Retículo endoplasmático e membrana plasmática
P3.66 + Marcações pontuais citoplasmáticas, citoplasma
e membrana plasmática
P3.67 ++++ Vesículas
P3.68 +++ Membrana plasmática e retículo encoplasmático
P3.69 +++ Vesículas, retículo endoplasmático e membrana
plasmática
P3.70 - -
P3.71 ++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P3.72 ++++ º
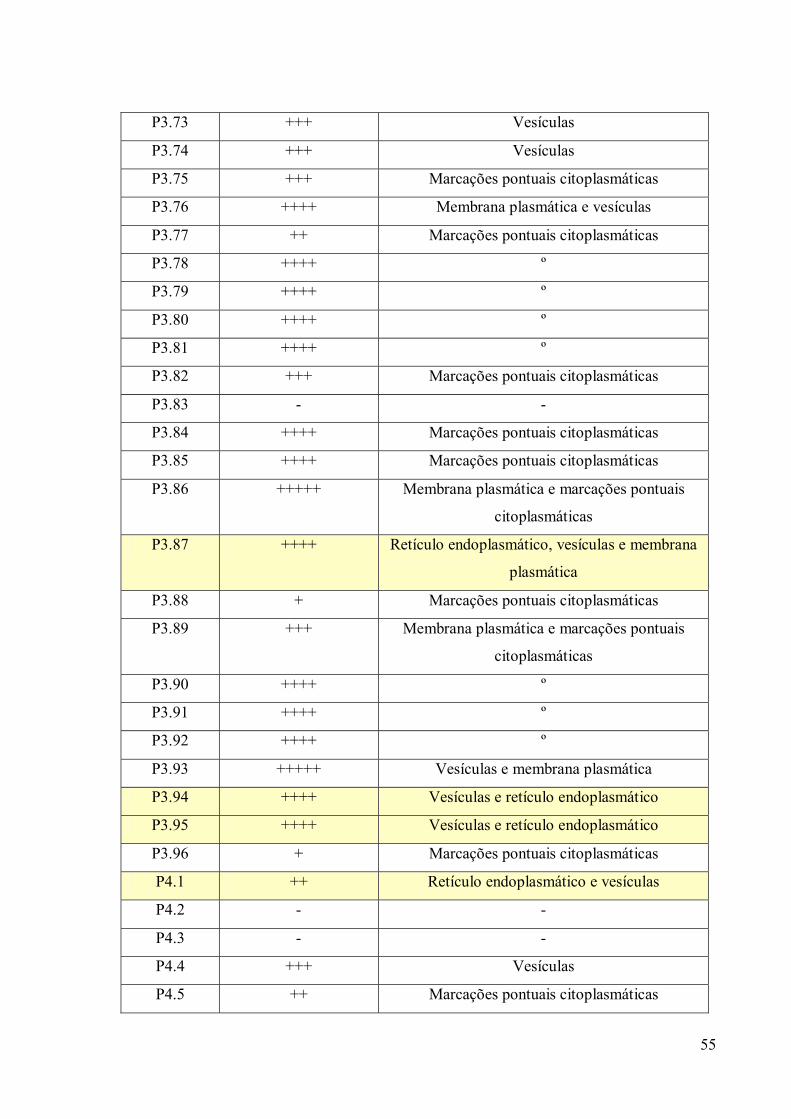
55
P3.73 +++ Vesículas
P3.74 +++ Vesículas
P3.75 +++ Marcações pontuais citoplasmáticas
P3.76 ++++ Membrana plasmática e vesículas
P3.77 ++ Marcações pontuais citoplasmáticas
P3.78 ++++ º
P3.79 ++++ º
P3.80 ++++ º
P3.81 ++++ º
P3.82 +++ Marcações pontuais citoplasmáticas
P3.83 - -
P3.84 ++++ Marcações pontuais citoplasmáticas
P3.85 ++++ Marcações pontuais citoplasmáticas
P3.86 +++++ Membrana plasmática e marcações pontuais
citoplasmáticas
P3.87 ++++ Retículo endoplasmático, vesículas e membrana
plasmática
P3.88 + Marcações pontuais citoplasmáticas
P3.89 +++ Membrana plasmática e marcações pontuais
citoplasmáticas
P3.90 ++++ º
P3.91 ++++ º
P3.92 ++++ º
P3.93 +++++ Vesículas e membrana plasmática
P3.94 ++++ Vesículas e retículo endoplasmático
P3.95 ++++ Vesículas e retículo endoplasmático
P3.96 + Marcações pontuais citoplasmáticas
P4.1 ++ Retículo endoplasmático e vesículas
P4.2 - -
P4.3 - -
P4.4 +++ Vesículas
P4.5 ++ Marcações pontuais citoplasmáticas

56
P4.6 +++++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.7 +++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.8 ++++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.9 ++++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.10 ++++ Marcações pontuais citoplasmáticas
P4.11 ++++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.12 +++ Retículo endoplasmático e marcações pontuais
citoplasmáticas
P4.13 +++ Marcações pontuais citoplasmáticas
P4.14 ++++ Marcações pontuais citoplasmáticas
P4.15 ++++ Marcações pontuais citoplasmáticas
P4.16 +++ Retículo endoplasmático e marcações pontuais
citoplasmáticas
P4.17 +++ Marcações pontuais citoplasmáticas
P4.18 ++ Marcações pontuais citoplasmáticas
P4.19 - -
P4.20 - -
P4.21 - -
P4.22 - -
P4.23 - -
P4.24 ++++ Marcações pontuais citoplasmáticas
P4.25 ++++ Retículo endoplasmático e marcações pontuais
citoplasmáticas
P4.26 ++++ Membrana plasmática, complexo de Golgi e
marcações pontuais citoplasmáticas
P4.27 +++ Marcações pontuais citoplasmáticas e membrana
plasmática
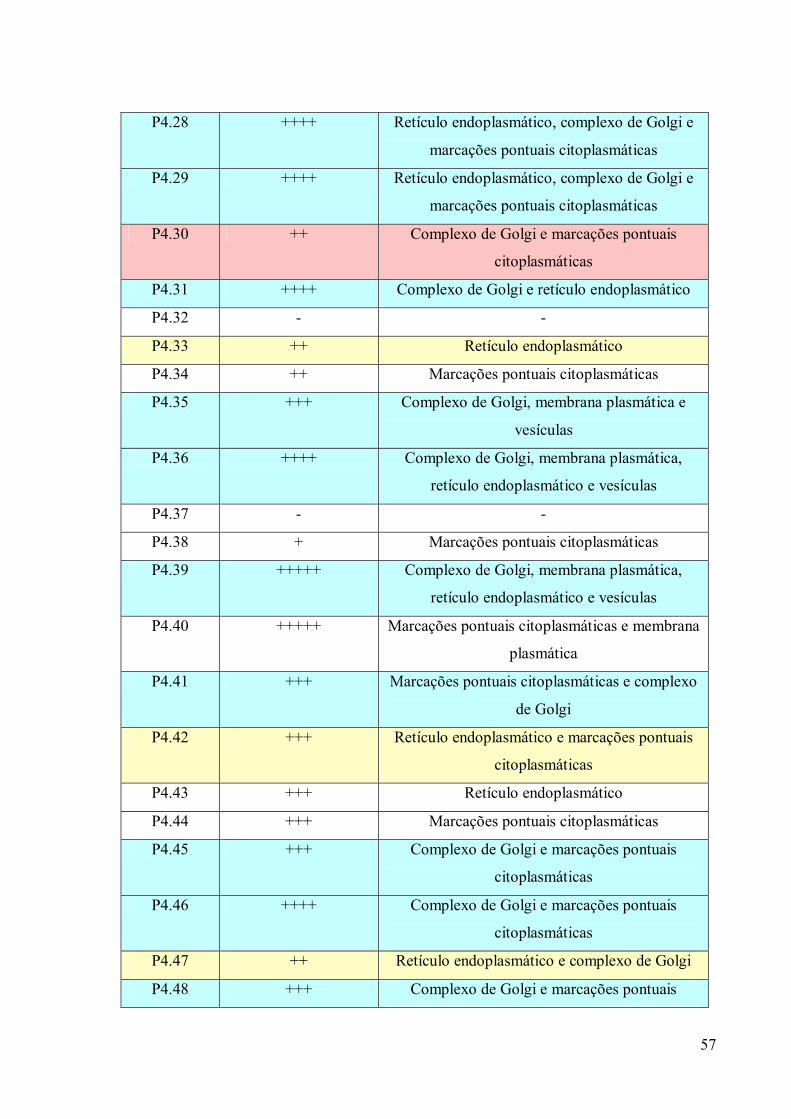
57
P4.28 ++++ Retículo endoplasmático, complexo de Golgi e
marcações pontuais citoplasmáticas
P4.29 ++++ Retículo endoplasmático, complexo de Golgi e
marcações pontuais citoplasmáticas
P4.30 ++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.31 ++++ Complexo de Golgi e retículo endoplasmático
P4.32 - -
P4.33 ++ Retículo endoplasmático
P4.34 ++ Marcações pontuais citoplasmáticas
P4.35 +++ Complexo de Golgi, membrana plasmática e
vesículas
P4.36 ++++ Complexo de Golgi, membrana plasmática,
retículo endoplasmático e vesículas
P4.37 - -
P4.38 + Marcações pontuais citoplasmáticas
P4.39 +++++ Complexo de Golgi, membrana plasmática,
retículo endoplasmático e vesículas
P4.40 +++++ Marcações pontuais citoplasmáticas e membrana
plasmática
P4.41 +++ Marcações pontuais citoplasmáticas e complexo
de Golgi
P4.42 +++ Retículo endoplasmático e marcações pontuais
citoplasmáticas
P4.43 +++ Retículo endoplasmático
P4.44 +++ Marcações pontuais citoplasmáticas
P4.45 +++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.46 ++++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.47 ++ Retículo endoplasmático e complexo de Golgi
P4.48 +++ Complexo de Golgi e marcações pontuais
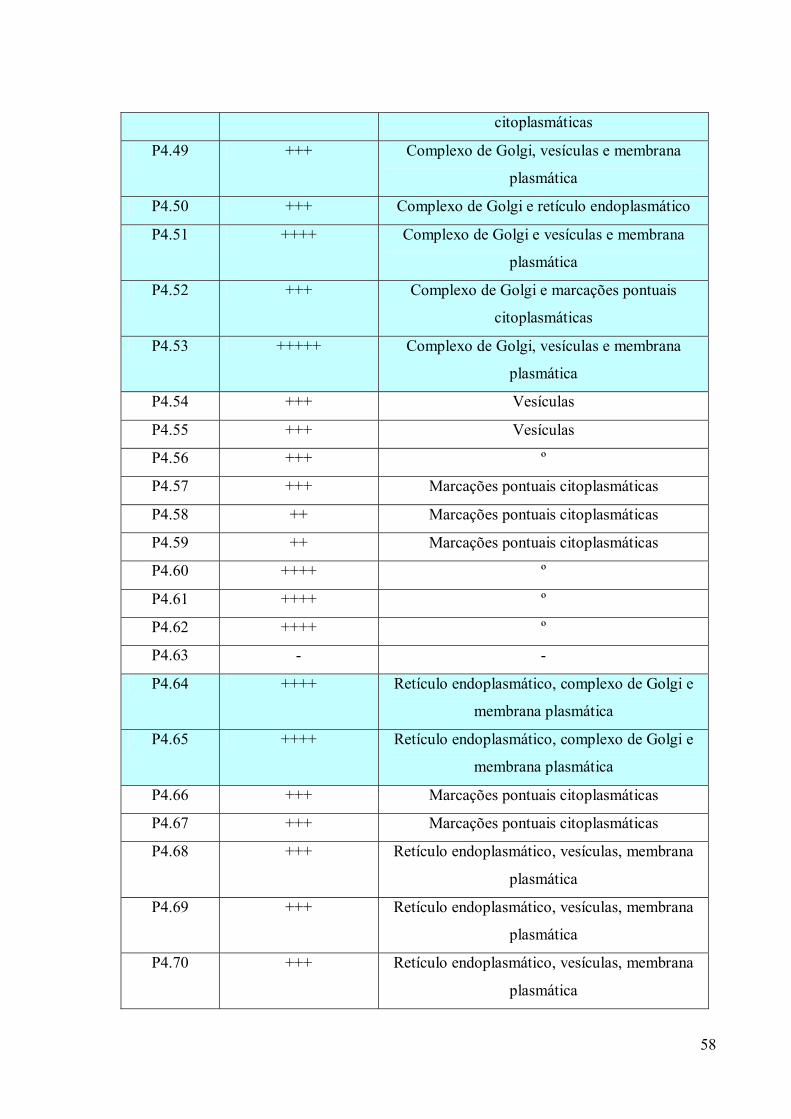
58
citoplasmáticas
P4.49 +++ Complexo de Golgi, vesículas e membrana
plasmática
P4.50 +++ Complexo de Golgi e retículo endoplasmático
P4.51 ++++ Complexo de Golgi e vesículas e membrana
plasmática
P4.52 +++ Complexo de Golgi e marcações pontuais
citoplasmáticas
P4.53 +++++ Complexo de Golgi, vesículas e membrana
plasmática
P4.54 +++ Vesículas
P4.55 +++ Vesículas
P4.56 +++ º
P4.57 +++ Marcações pontuais citoplasmáticas
P4.58 ++ Marcações pontuais citoplasmáticas
P4.59 ++ Marcações pontuais citoplasmáticas
P4.60 ++++ º
P4.61 ++++ º
P4.62 ++++ º
P4.63 - -
P4.64 ++++ Retículo endoplasmático, complexo de Golgi e
membrana plasmática
P4.65 ++++ Retículo endoplasmático, complexo de Golgi e
membrana plasmática
P4.66 +++ Marcações pontuais citoplasmáticas
P4.67 +++ Marcações pontuais citoplasmáticas
P4.68 +++ Retículo endoplasmático, vesículas, membrana
plasmática
P4.69 +++ Retículo endoplasmático, vesículas, membrana
plasmática
P4.70 +++ Retículo endoplasmático, vesículas, membrana
plasmática
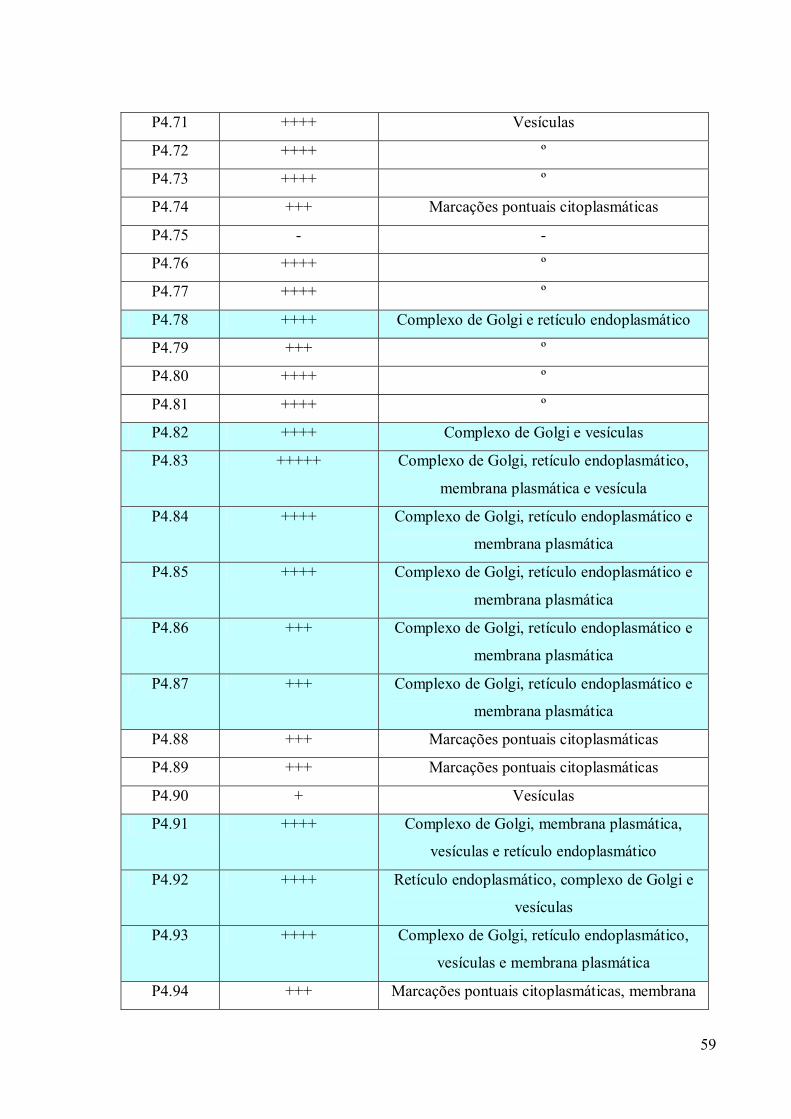
59
P4.71 ++++ Vesículas
P4.72 ++++ º
P4.73 ++++ º
P4.74 +++ Marcações pontuais citoplasmáticas
P4.75 - -
P4.76 ++++ º
P4.77 ++++ º
P4.78 ++++ Complexo de Golgi e retículo endoplasmático
P4.79 +++ º
P4.80 ++++ º
P4.81 ++++ º
P4.82 ++++ Complexo de Golgi e vesículas
P4.83 +++++ Complexo de Golgi, retículo endoplasmático,
membrana plasmática e vesícula
P4.84 ++++ Complexo de Golgi, retículo endoplasmático e
membrana plasmática
P4.85 ++++ Complexo de Golgi, retículo endoplasmático e
membrana plasmática
P4.86 +++ Complexo de Golgi, retículo endoplasmático e
membrana plasmática
P4.87 +++ Complexo de Golgi, retículo endoplasmático e
membrana plasmática
P4.88 +++ Marcações pontuais citoplasmáticas
P4.89 +++ Marcações pontuais citoplasmáticas
P4.90 + Vesículas
P4.91 ++++ Complexo de Golgi, membrana plasmática,
vesículas e retículo endoplasmático
P4.92 ++++ Retículo endoplasmático, complexo de Golgi e
vesículas
P4.93 ++++ Complexo de Golgi, retículo endoplasmático,
vesículas e membrana plasmática
P4.94 +++ Marcações pontuais citoplasmáticas, membrana
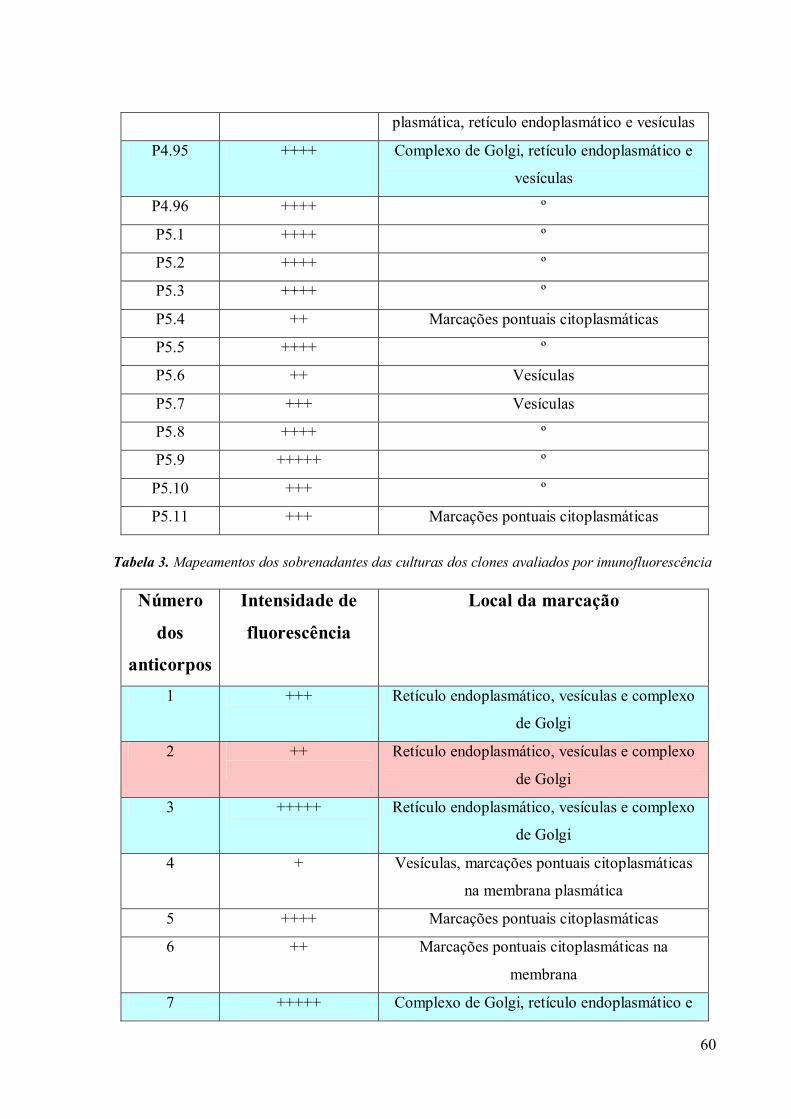
60
plasmática, retículo endoplasmático e vesículas
P4.95 ++++ Complexo de Golgi, retículo endoplasmático e
vesículas
P4.96 ++++ º
P5.1 ++++ º
P5.2 ++++ º
P5.3 ++++ º
P5.4 ++ Marcações pontuais citoplasmáticas
P5.5 ++++ º
P5.6 ++ Vesículas
P5.7 +++ Vesículas
P5.8 ++++ º
P5.9 +++++ º
P5.10 +++ º
P5.11 +++ Marcações pontuais citoplasmáticas
Tabela 3. Mapeamentos dos sobrenadantes das culturas dos clones avaliados por imunofluorescência
Número
dos
anticorpos
Intensidade de
fluorescência
Local da marcação
1 +++
Retículo endoplasmático, vesículas e complexo
de Golgi 2 ++
Retículo endoplasmático, vesículas e complexo
de Golgi 3 +++++
Retículo endoplasmático, vesículas e complexo
de Golgi 4 + Vesículas, marcações pontuais citoplasmáticas
na membrana plasmática 5 ++++ Marcações pontuais citoplasmáticas 6 ++ Marcações pontuais citoplasmáticas na
membrana 7 +++++ Complexo de Golgi, retículo endoplasmático e

61
vesículas 8 +++ Marcações pontuais citoplasmáticas 9 + Marcações pontuais citoplasmáticas
10 +++ Marcações pontuais citoplasmáticas 11 +++
Marcações pontuais citoplasmáticas na
membrana plasmática 12 +++ Marcações pontuais citoplasmáticas e flagelos 13 ++++ Flagelos, marcações pontuais citoplasmáticas na
membrana plasmática, vesículas, retículo
endoplasmático 14 ++++
Marcações pontuais citoplasmáticas na
membrana plasmática 15 +++++
Membrana plasmática, retículo endoplasmático e
muitas vesículas
16 +++
Membrana plasmática e vesículas
17 +++
Retículo endoplasmático, vesículas e complexos
de Golgi 18 +++
Retículo endoplasmático, alguns complexos de
Golgi e vesículas 19 +++ Membrana plasmática 20 +++++ Membrana plasmática, retículo endoplasmático e
complexo de Golgi 21 +++++ Membrana plasmática e vesículas 22 ++++ Membrana plasmática, vesículas e complexo de
Golgi 23 ++++ Membrana plasmática e vesículas 24 ++++ Marcações pontuais citoplasmáticas na
membrana plasmática 25 +++++ Marcações pontuais citoplasmáticas na
membrana plasmática e vesículas 26 +++ Vesículas
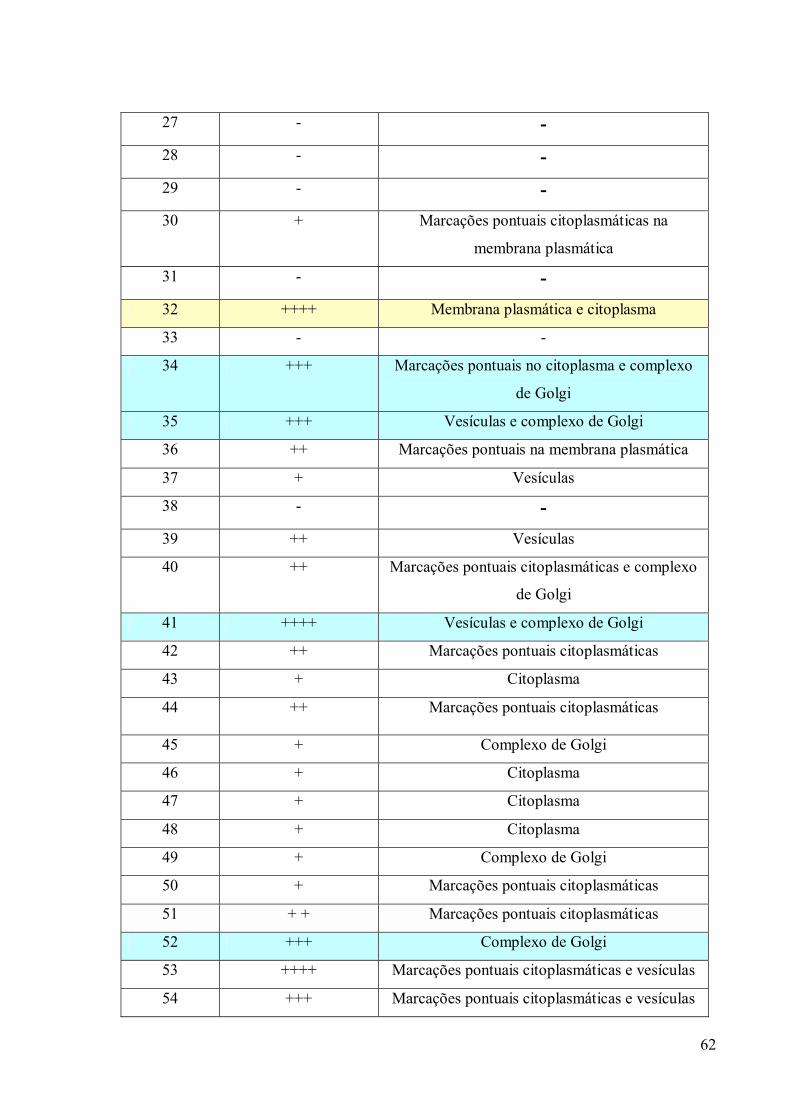
62
27 - - 28 - - 29 - - 30 + Marcações pontuais citoplasmáticas na
membrana plasmática 31 - - 32 ++++ Membrana plasmática e citoplasma 33 - - 34 +++ Marcações pontuais no citoplasma e complexo
de Golgi 35 +++ Vesículas e complexo de Golgi 36 ++ Marcações pontuais na membrana plasmática 37 + Vesículas 38 - - 39 ++ Vesículas 40 ++ Marcações pontuais citoplasmáticas e complexo
de Golgi 41 ++++ Vesículas e complexo de Golgi 42 ++ Marcações pontuais citoplasmáticas 43 + Citoplasma 44 ++
Marcações pontuais citoplasmáticas
45 + Complexo de Golgi 46 + Citoplasma 47 + Citoplasma 48 + Citoplasma 49 + Complexo de Golgi 50 + Marcações pontuais citoplasmáticas 51 + + Marcações pontuais citoplasmáticas 52 +++ Complexo de Golgi 53 ++++ Marcações pontuais citoplasmáticas e vesículas 54 +++ Marcações pontuais citoplasmáticas e vesículas
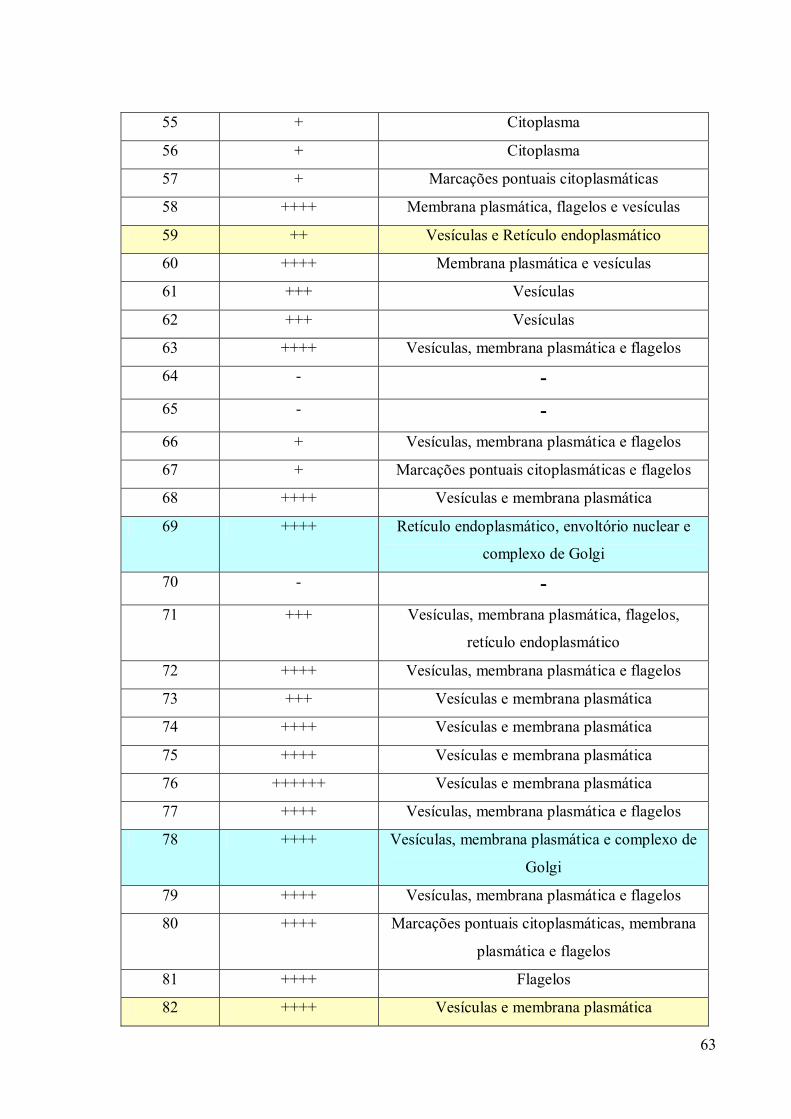
63
55 + Citoplasma 56 + Citoplasma 57 + Marcações pontuais citoplasmáticas 58 ++++ Membrana plasmática, flagelos e vesículas 59 ++ Vesículas e Retículo endoplasmático 60 ++++ Membrana plasmática e vesículas 61 +++ Vesículas 62 +++ Vesículas 63 ++++ Vesículas, membrana plasmática e flagelos 64 - - 65 - - 66 + Vesículas, membrana plasmática e flagelos 67 + Marcações pontuais citoplasmáticas e flagelos 68 ++++ Vesículas e membrana plasmática 69 ++++ Retículo endoplasmático, envoltório nuclear e
complexo de Golgi 70 - - 71 +++ Vesículas, membrana plasmática, flagelos,
retículo endoplasmático 72 ++++ Vesículas, membrana plasmática e flagelos 73 +++ Vesículas e membrana plasmática 74 ++++ Vesículas e membrana plasmática 75 ++++ Vesículas e membrana plasmática 76 ++++++ Vesículas e membrana plasmática 77 ++++ Vesículas, membrana plasmática e flagelos 78 ++++ Vesículas, membrana plasmática e complexo de
Golgi 79 ++++ Vesículas, membrana plasmática e flagelos 80 ++++ Marcações pontuais citoplasmáticas, membrana
plasmática e flagelos 81 ++++ Flagelos
82 ++++ Vesículas e membrana plasmática
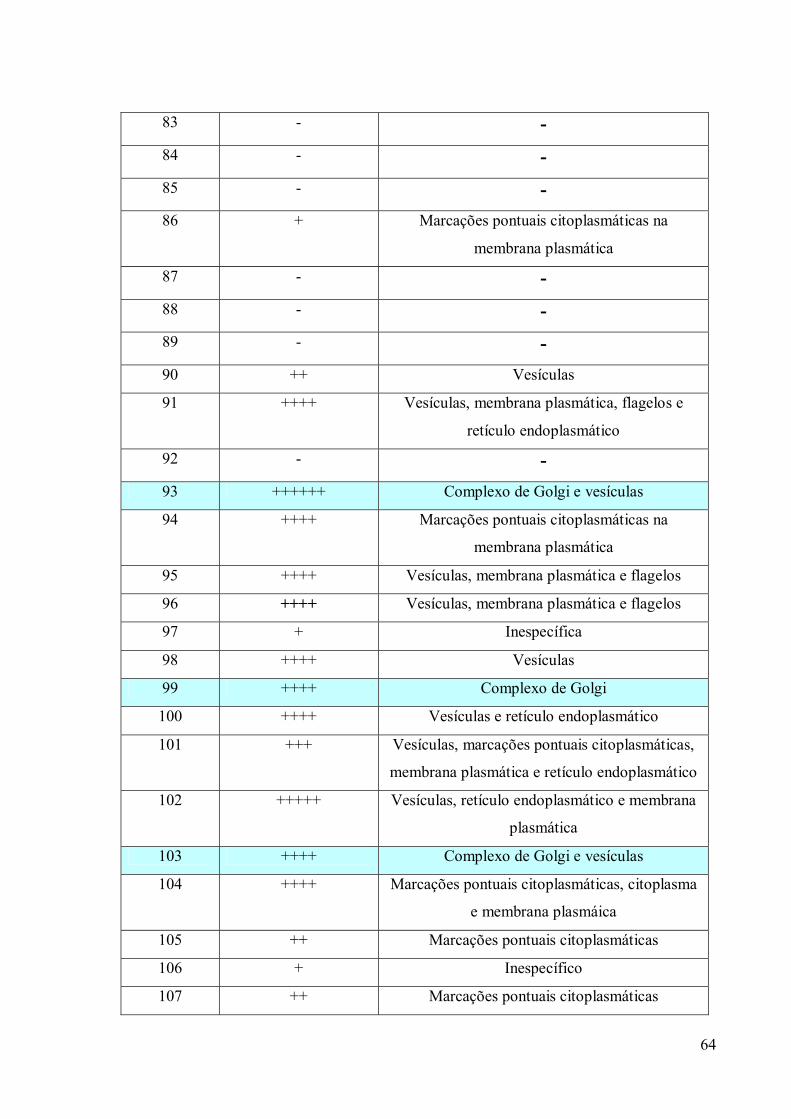
64
83 - - 84 - - 85 - - 86 + Marcações pontuais citoplasmáticas na
membrana plasmática 87 - - 88 - - 89 - - 90 ++ Vesículas 91 ++++ Vesículas, membrana plasmática, flagelos e
retículo endoplasmático 92 - - 93 ++++++ Complexo de Golgi e vesículas
94 ++++ Marcações pontuais citoplasmáticas na
membrana plasmática 95 ++++ Vesículas, membrana plasmática e flagelos 96 ++++ Vesículas, membrana plasmática e flagelos 97 + Inespecífica
98 ++++ Vesículas
99 ++++ Complexo de Golgi
100 ++++ Vesículas e retículo endoplasmático
101 +++ Vesículas, marcações pontuais citoplasmáticas,
membrana plasmática e retículo endoplasmático
102 +++++ Vesículas, retículo endoplasmático e membrana
plasmática
103 ++++ Complexo de Golgi e vesículas
104 ++++ Marcações pontuais citoplasmáticas, citoplasma
e membrana plasmáica
105 ++ Marcações pontuais citoplasmáticas
106 + Inespecífico
107 ++ Marcações pontuais citoplasmáticas
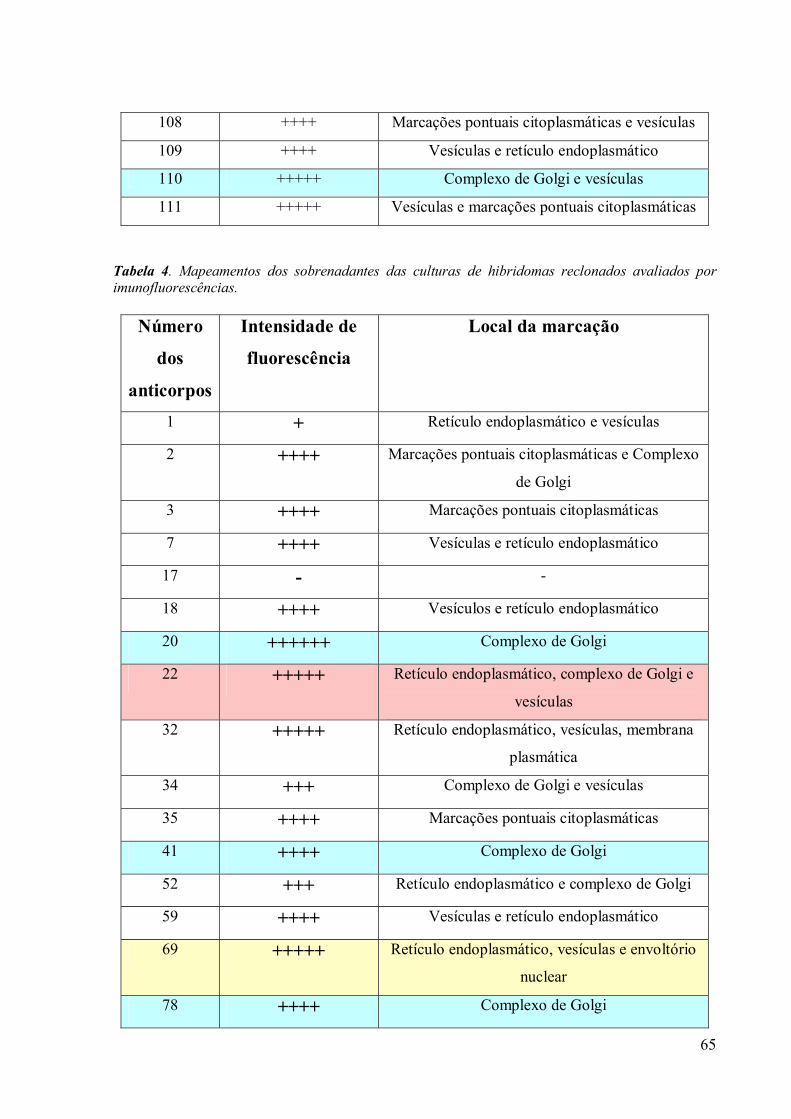
65
108 ++++ Marcações pontuais citoplasmáticas e vesículas
109 ++++ Vesículas e retículo endoplasmático
110 +++++ Complexo de Golgi e vesículas
111 +++++ Vesículas e marcações pontuais citoplasmáticas
Tabela 4. Mapeamentos dos sobrenadantes das culturas de hibridomas reclonados avaliados por imunofluorescências.
Número
dos
anticorpos
Intensidade de
fluorescência
Local da marcação
1 + Retículo endoplasmático e vesículas
2 ++++ Marcações pontuais citoplasmáticas e Complexo
de Golgi
3 ++++ Marcações pontuais citoplasmáticas
7 ++++ Vesículas e retículo endoplasmático
17 - -
18 ++++ Vesículos e retículo endoplasmático
20 ++++++ Complexo de Golgi
22 +++++ Retículo endoplasmático, complexo de Golgi e
vesículas
32 +++++ Retículo endoplasmático, vesículas, membrana
plasmática
34 +++ Complexo de Golgi e vesículas
35 ++++ Marcações pontuais citoplasmáticas
41 ++++ Complexo de Golgi
52 +++ Retículo endoplasmático e complexo de Golgi
59 ++++ Vesículas e retículo endoplasmático
69 +++++ Retículo endoplasmático, vesículas e envoltório
nuclear
78 ++++ Complexo de Golgi
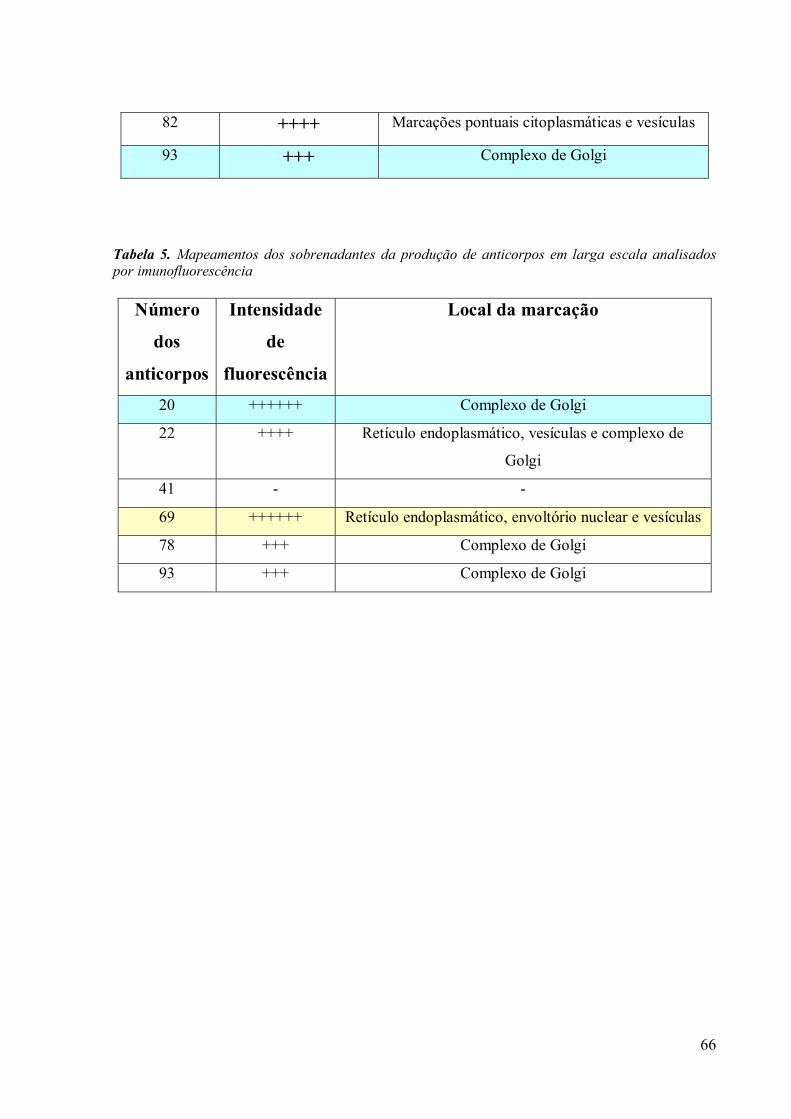
66
82 ++++ Marcações pontuais citoplasmáticas e vesículas
93 +++ Complexo de Golgi
Tabela 5. Mapeamentos dos sobrenadantes da produção de anticorpos em larga escala analisados por imunofluorescência
Número
dos
anticorpos
Intensidade
de
fluorescência
Local da marcação
20 ++++++ Complexo de Golgi
22 ++++ Retículo endoplasmático, vesículas e complexo de
Golgi
41 - -
69 ++++++ Retículo endoplasmático, envoltório nuclear e vesículas
78 +++ Complexo de Golgi
93 +++ Complexo de Golgi

67
3.4 Imunofluorescência
Durante as etapas de seleção dos hibridomas foram feitas análises de
imunofluorescência para determinar a localização e intensidade de fluorescência. Na etapa de
clonagem o anticorpo 110 (Tabela 3) mostrou uma ótima marcação para complexo de Golgi.
(FIG. 10). Entretanto este clone perdeu a capacidade de secretar imunoglobulinas.
Na etapa de reclonagem, o anticorpo 93 (Tabela 4) marcou o complexo de Golgi de T.
foetus (FIG. 11), assim como o 20 (Tabela 4), entretanto este último demonstrou marcação
mais intensa (FIG. 12). A marcação e a intensidade de fluorescência deste anticorpo se
mantiveram na etapa de produção de anticorpo em larga escala. Desta forma, o anticorpo 20
foi purificado, dando origem ao anticorpo 20.3 que em trofozoítos de T. foetus observamos
uma marcação restrita ao complexo de Golgi (FIG. 13). Para confirmar que a localização a
qual nos referimos tratava-se realmente do complexo de Golgi, tratamos trofozoítos de T.
foetus com C6-NBD-Ceramida, um marcador específico para esta organela (FIG. 14). A
marcação obtida com este marcador foi semelhante ao descrito nas imunofluorescências com
os anticorpos anti-Golgi aqui produzidos.
Apesar dos anticorpos terem sido produzidos contra T. foetus, nos despertou a
curiosidade de saber se a proteína reconhecida pelo anticorpo anti-Golgi 20.3 poderia ser
conservada e assim reagir com proteínas de outros organismos. Desta forma, realizamos testes
de imunofluorescência em protozoários como T. vaginalis, também membro da Família
Trichomonadidae e nas formas promatigota Leishmania amazonensis e epimastigota de T.
cruzi. Células de mamíferos, representadas aqui pela linhagem de células MDCK também
foram usadas nestes testes.
Em Trichomonas vaginalis (FIG. 15), a marcação obtida no complexo de Golgi foi
semelhante à descrita em T. foetus. Por outro lado, em promastigotas de Leishmania
amazonensis, foi possível verificar que muitos destes parasitos apresentaram marcações

68
pontuais no citoplasma na região posterior da célula e no envoltório nuclear (FIG.16). Quando
testado em Trypanosoma cruzi (FIG.17), encontramos uma marcação de forma difusa em
apenas algumas células, restrita à região posterior (FIG.17b e f) e ao redor do núcleo
(FIG.17d), sugerindo uma marcação de retículo endoplasmático.
As células MDCKII foram eleitas como representantes de células de mamíferos. Em
células em interfase, observamos uma intensa marcação na região perinuclear e em algumas
vesículas no citoplasma (FIG. 18a). Por outro lado, células em mitose apresentaram uma
marcação disposta em pontos em toda célula, o que é natural em células em mitose (FIG.
18b). Ambas as marcações encontradas são características de complexo de Golgi de
mamíferos, o que nos leva a crer que o anticorpo anti-Golgi 20.3 possa estar reconhecendo
também uma proteína desta organela. As células MDCK também foram tratadas com C6-
NBD-Ceramida para verificar se a marcação obtida com o anticorpo anti-Golgi 20.3 era
coerente com a localização do complexo de Golgi desta célula (FIG. 19).
Os controles realizados somente com o anticorpo secundário anti-camudongo não
apresentaram marcações para nenhum dos tipos celulares utilizados nas imunofluorescências.
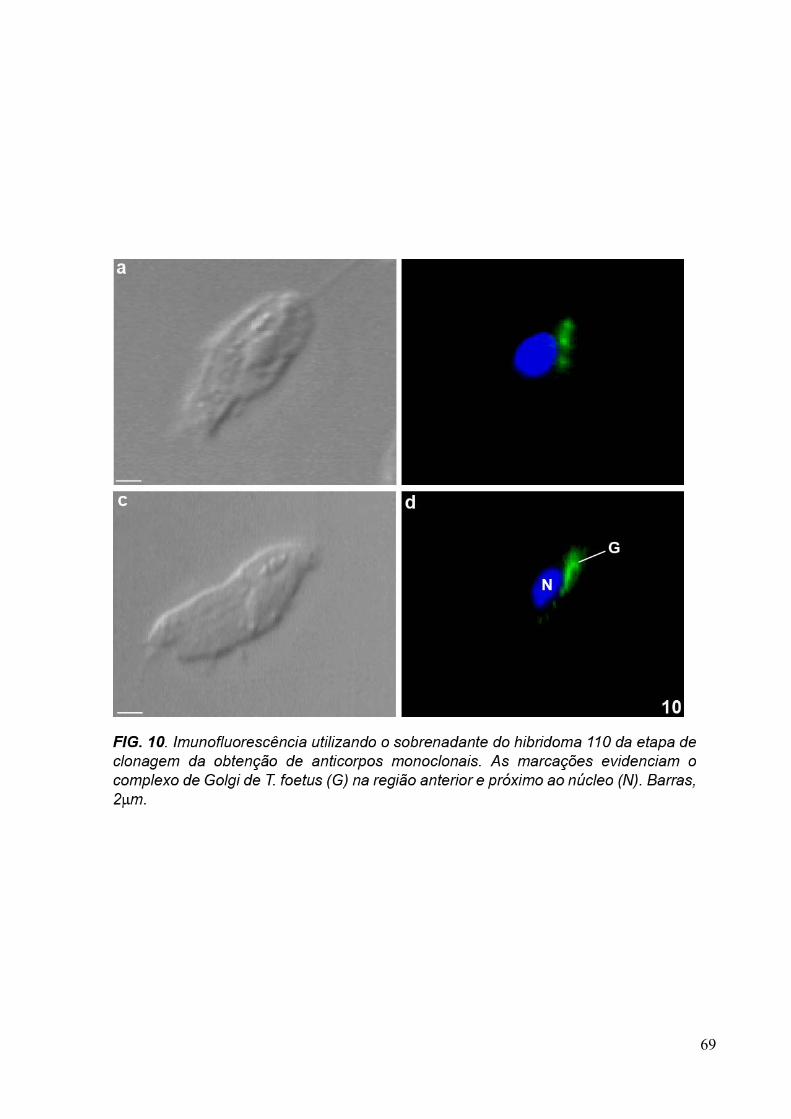
69
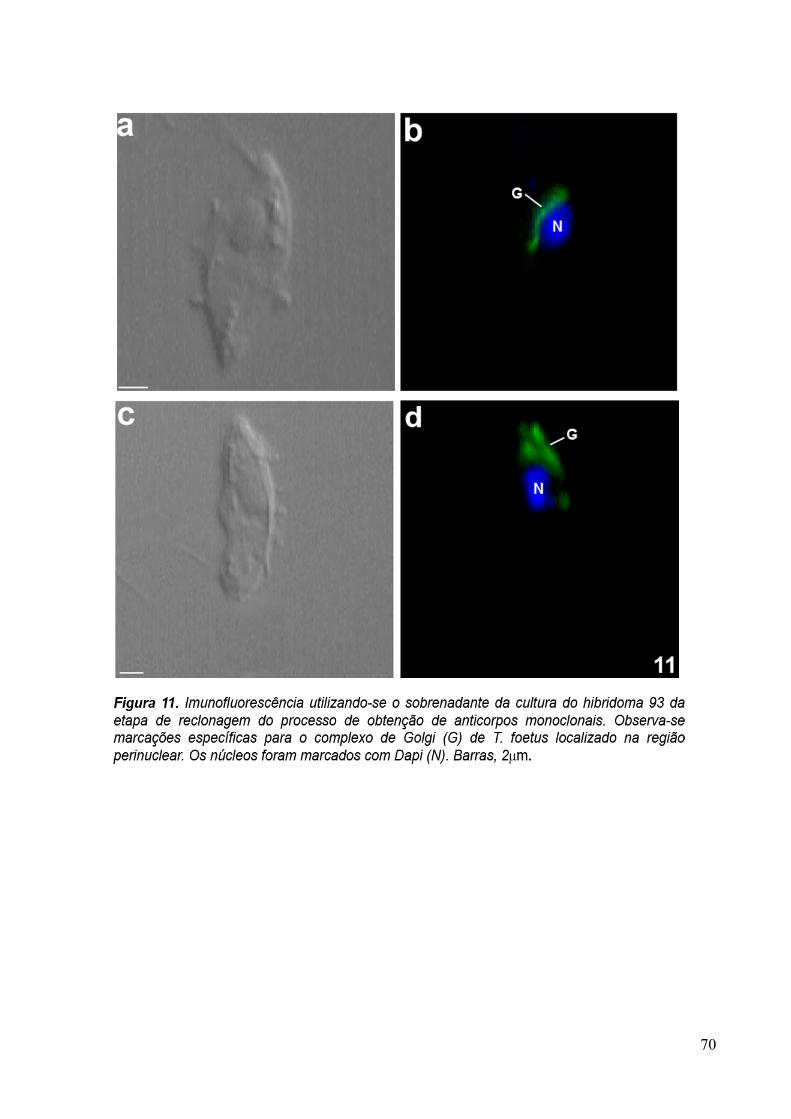
70
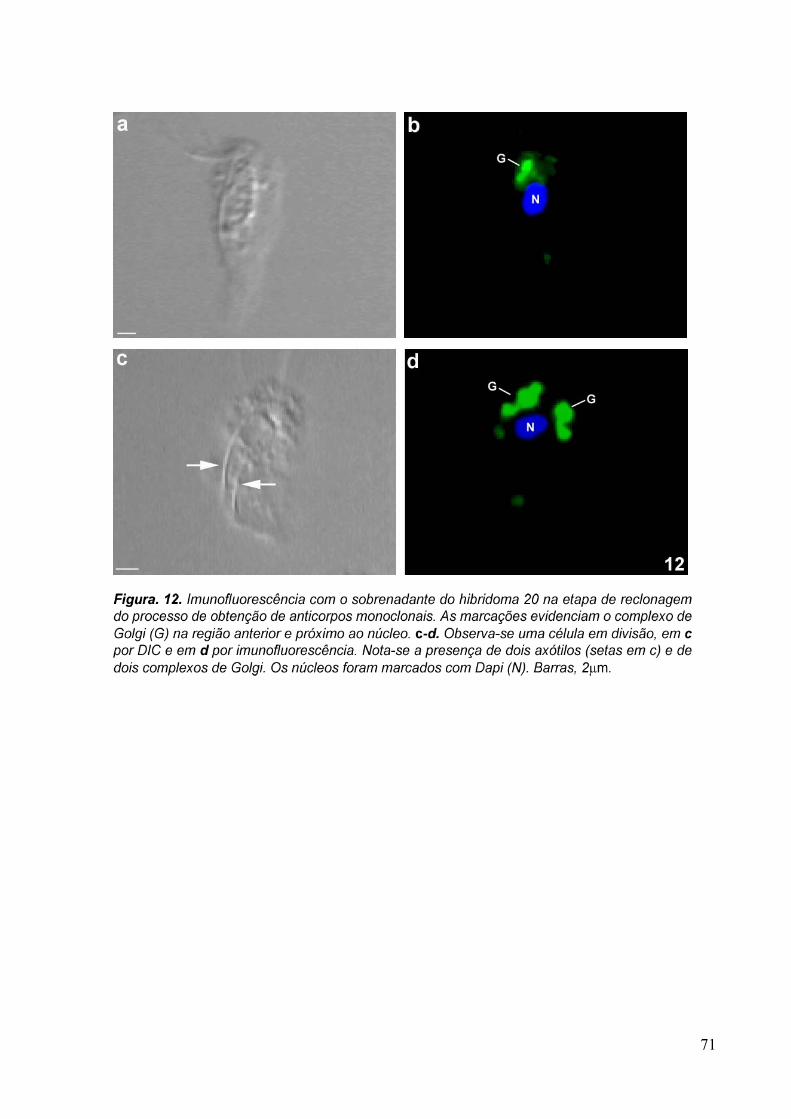
71
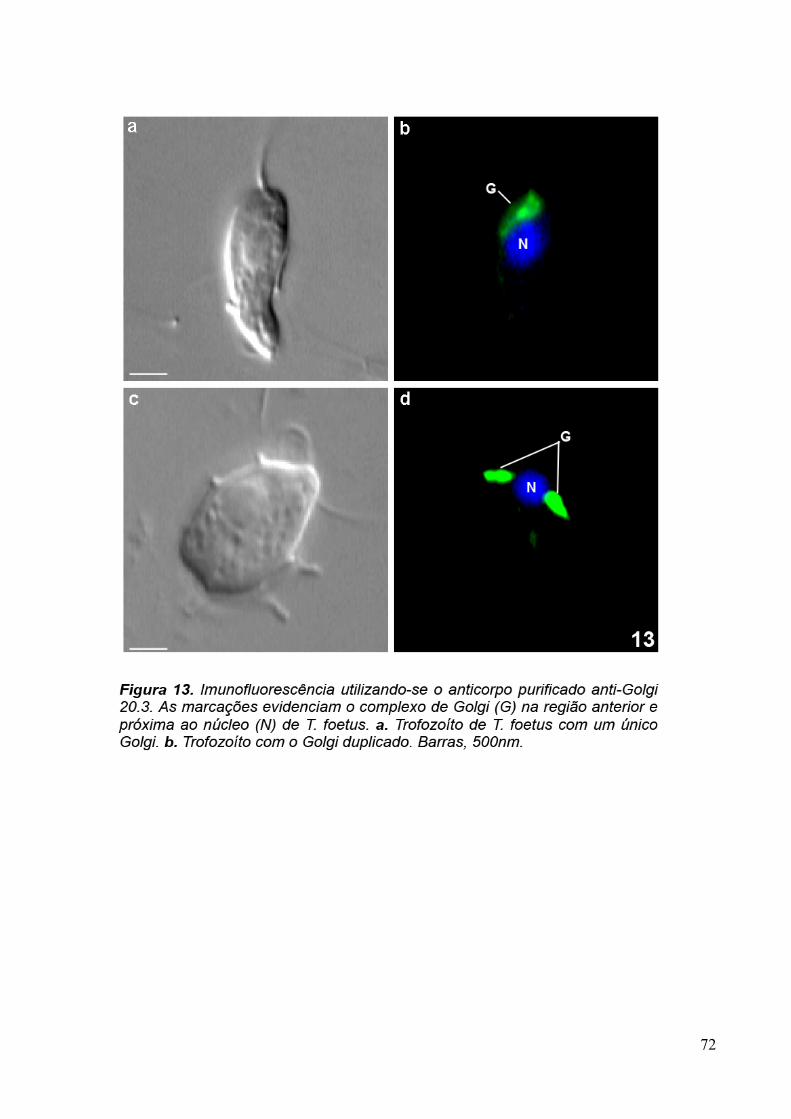
72
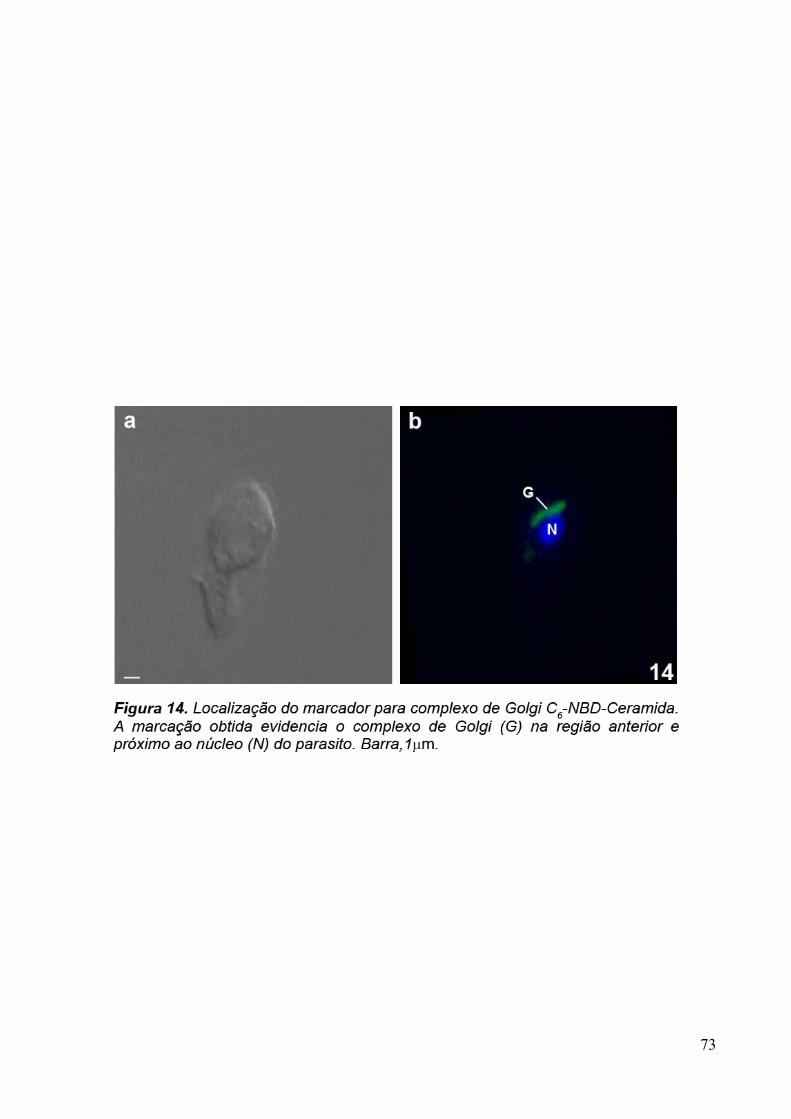
73
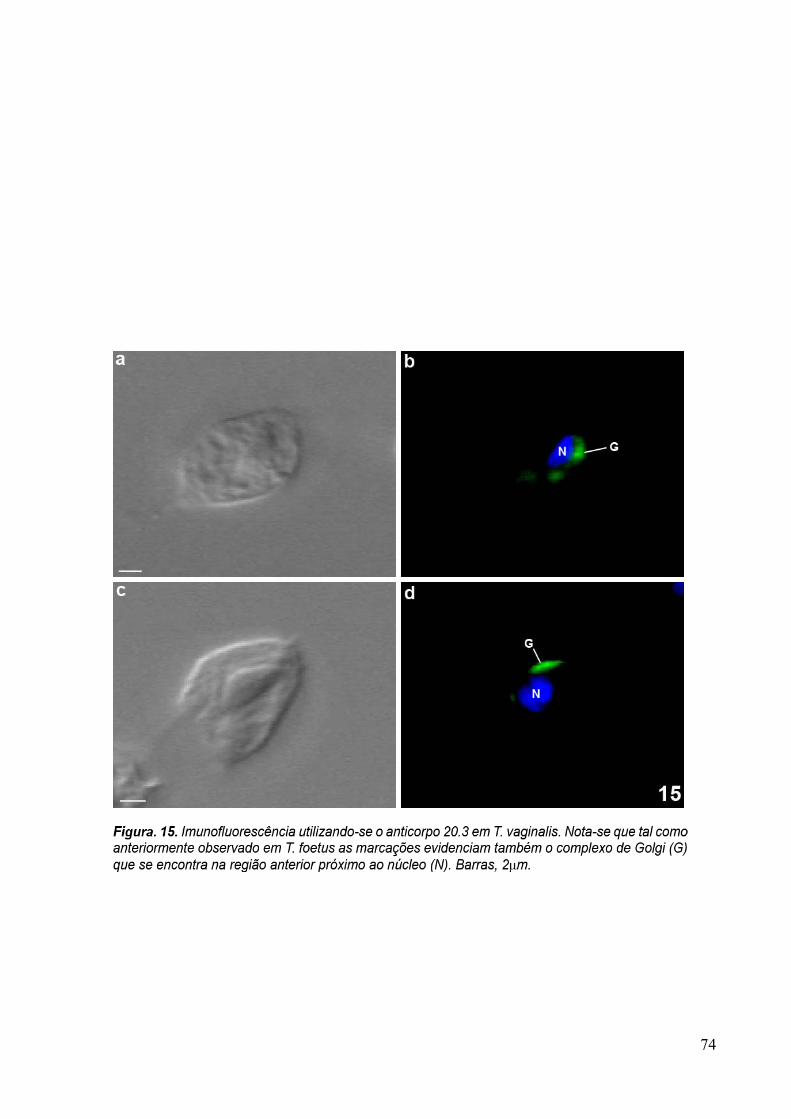
74
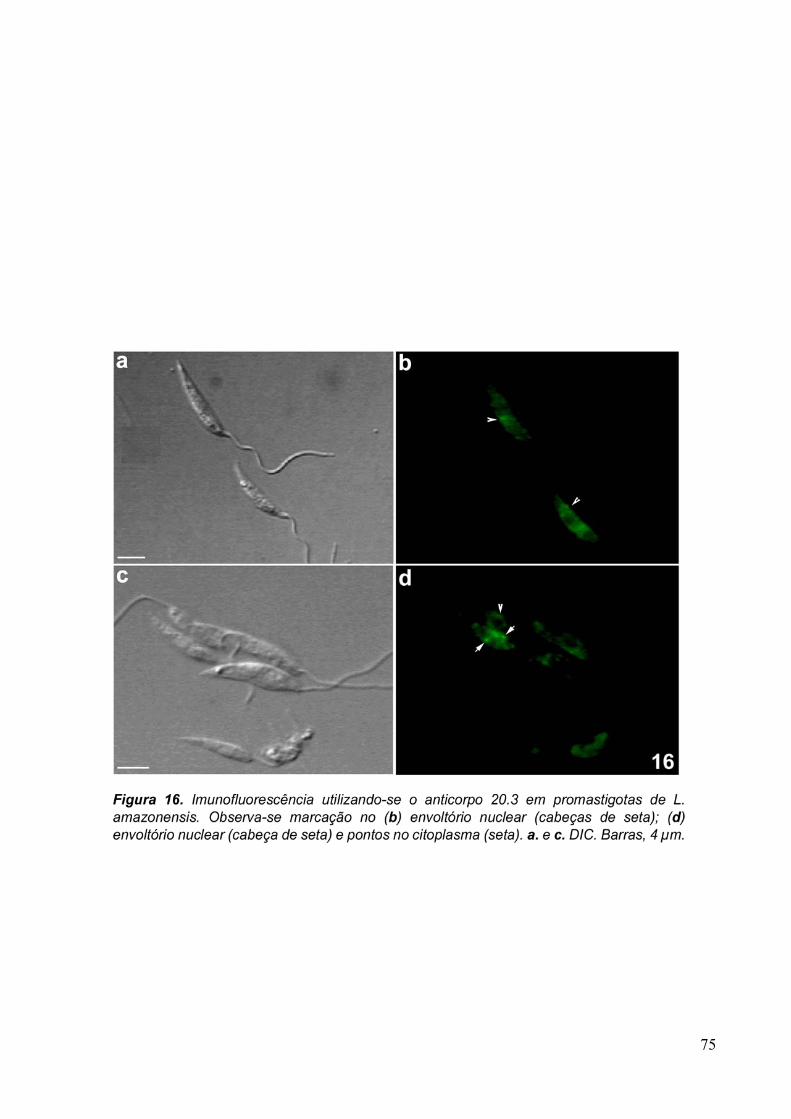
75
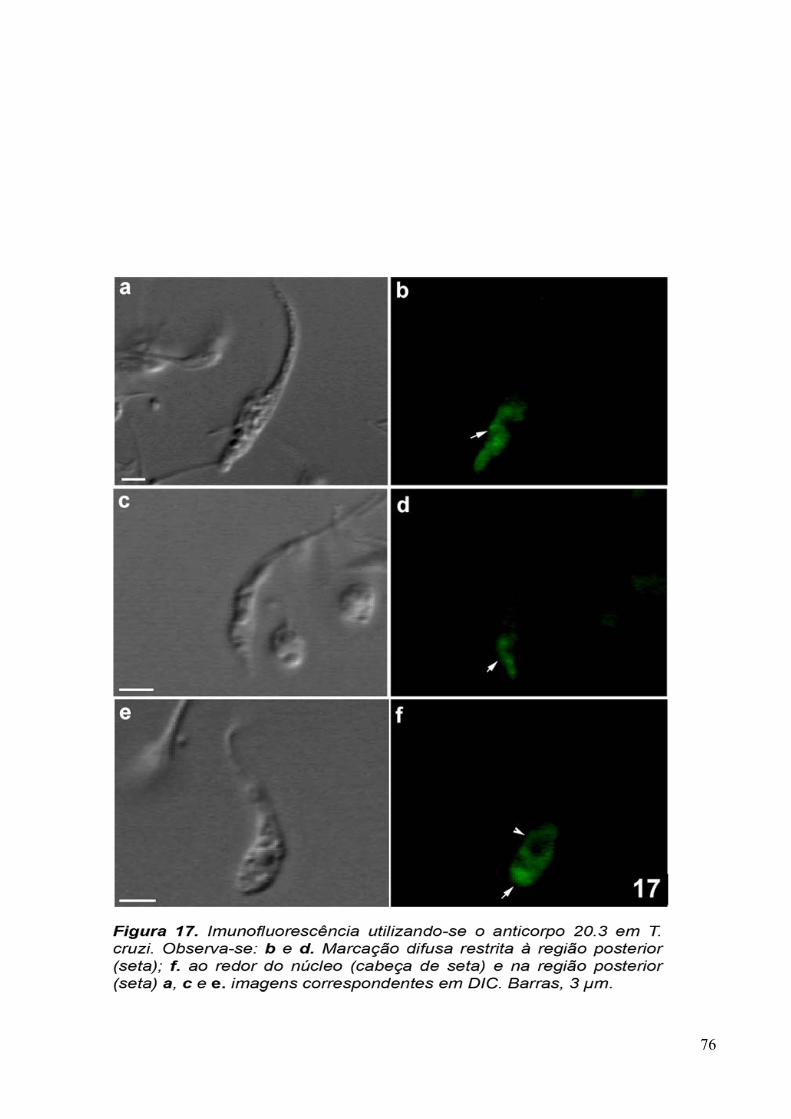
76
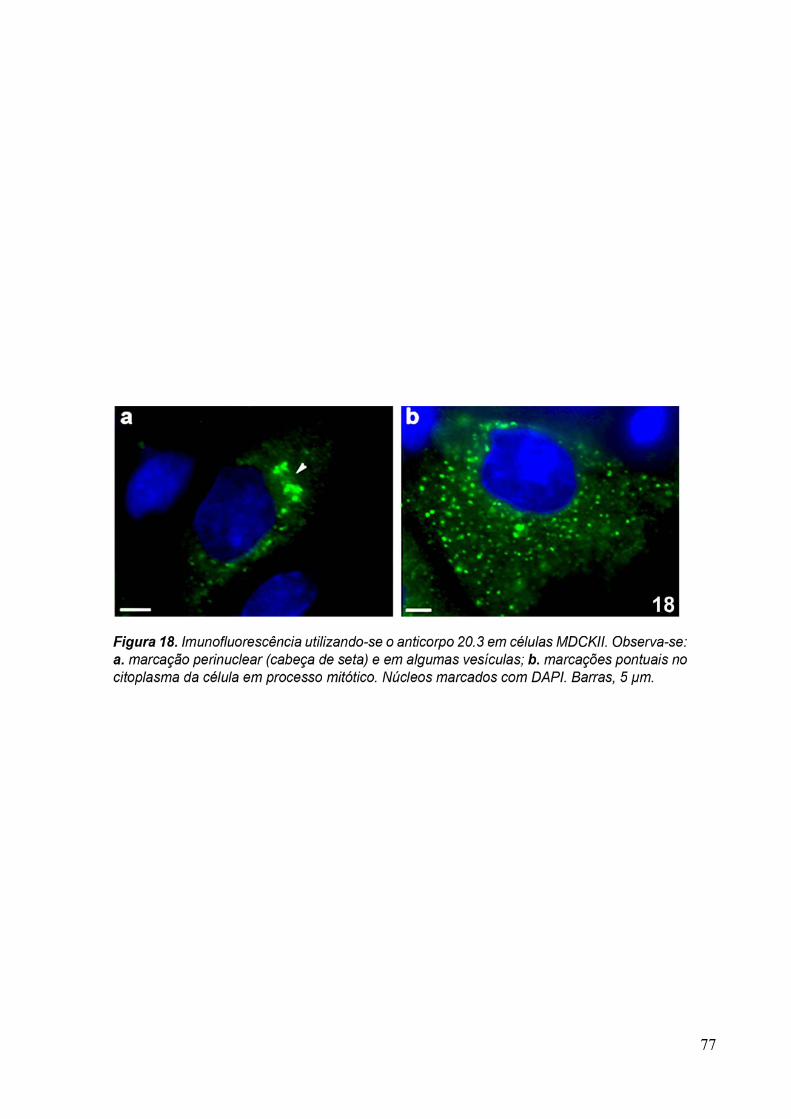
77
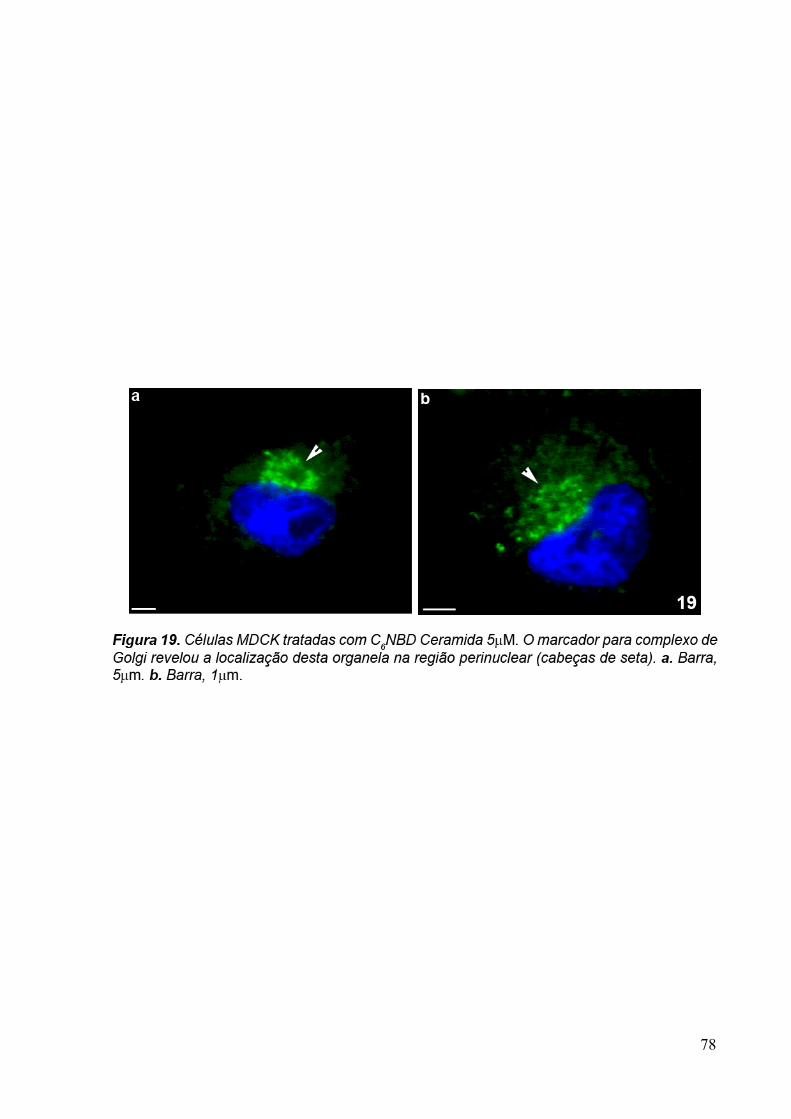
78

79
3.5. Imunocitoquímica
Durante a seleção dos anticorpos, foram realizadas imunocitoquímicas (em
microscopia eletrônica de transmissão) utilizando-se alguns anticorpos que apresentaram boa
marcação por imunofluorescência. Na FIG. 20, observamos uma nítida marcação bem
distribuída entre as cisternas do complexo de Golgi de T. foetus utilizando o anticorpo 93
(Tabela 3).
O anticorpo 110 (Tabela 4) apresentou uma marcação intensa na membrana plasmática
e nas cisternas das regiões medial e Trans do complexo de Golgi (FIG. 21). A rede Trans do
Golgi, vesículas e perfis de membrana distribuídos na célula também foram identificados por
este anticorpo. Este tipo de marcação nos leva a crer que a proteína tenha sido marcada ao
longo de seu tráfego pelo Golgi até o seu destino final, na membrana plasmática (FIG. 21). O
fato das primeiras cisternas não apresentarem marcação, pode ser devido a modificações pós-
translacionais sofridas pelas proteínas nas cisternas da região medial. Estas modificações
seriam essenciais para o reconhecimento pelo anticorpo.
Com o anticorpo 20, a marcação em T. foetus se apresentou de forma intensa no
complexo de Golgi, principalmente nas cisternas da região medial, onde se concentrou a
maioria das partículas de ouro (FIGs. 22 e 23). Uma leve marcação nas redes Cis, Trans e em
algumas vesículas também pode ser observada (FIGs. 22 e 23). Na figura 24, observam-se
ainda três Golgi marcados de forma homogênea, sem uma localização preferencial das
partículas de ouro.
Com o anticorpo 20.3 em trofozoítos de T. foetus, a marcação obtida manteve-se
restrita ao complexo de Golgi. (FIGs. 25 e 26).
A imunocitoquímica realizada com as frações do Golgi, utilizando-se os anticorpos
20.3 apresentou a marcação na membrana desta organela. (FIG. 27).

80
As imunocitoquímicas feitas como controle, sem a presença do anticorpo primário não
tiveram marcação.
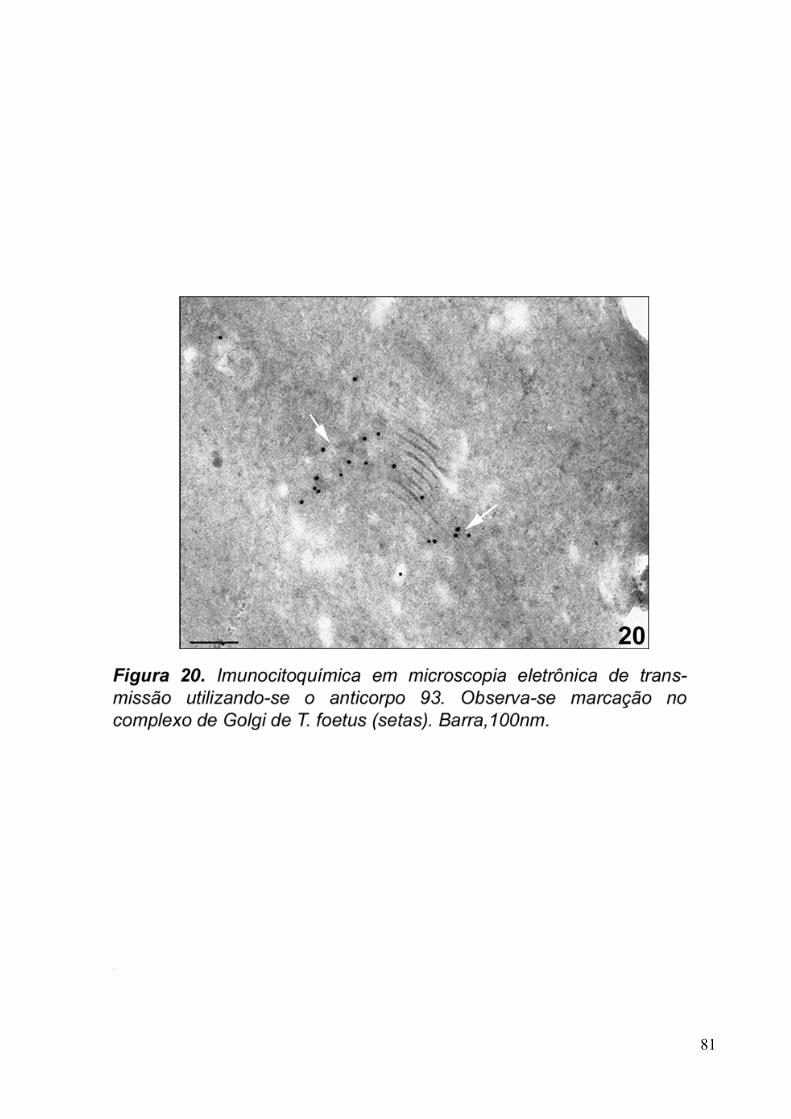
81

82
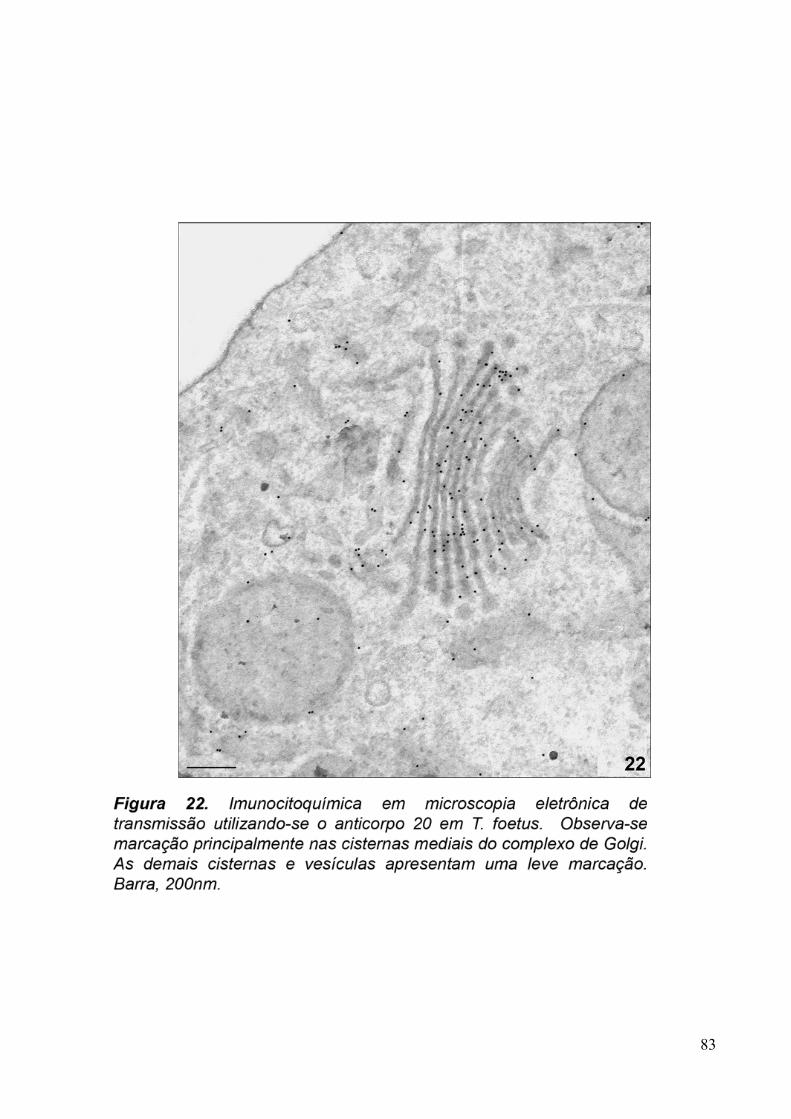
83
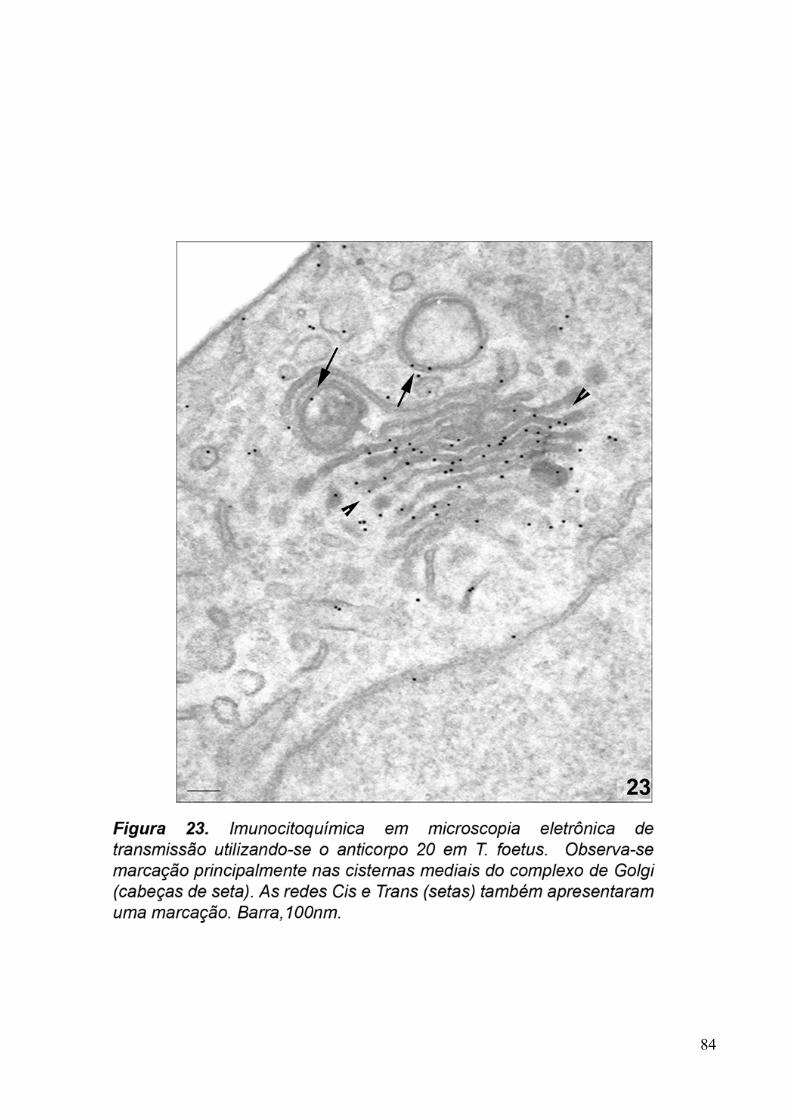
84
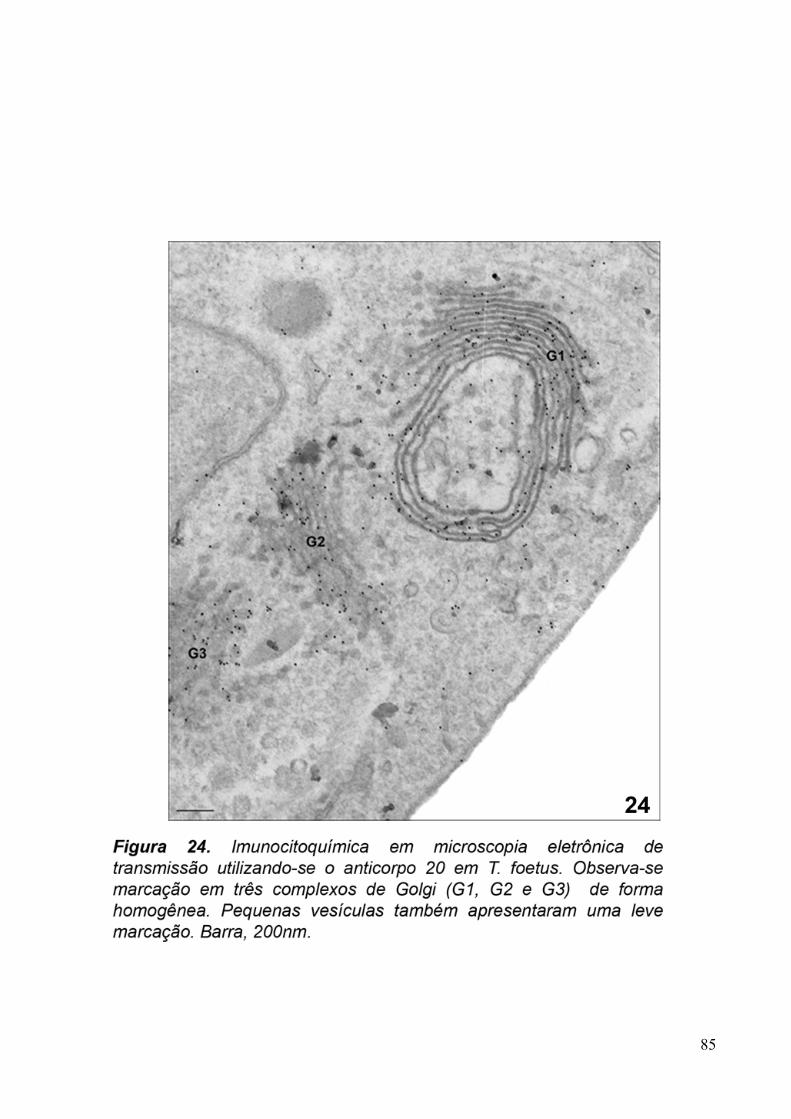
85
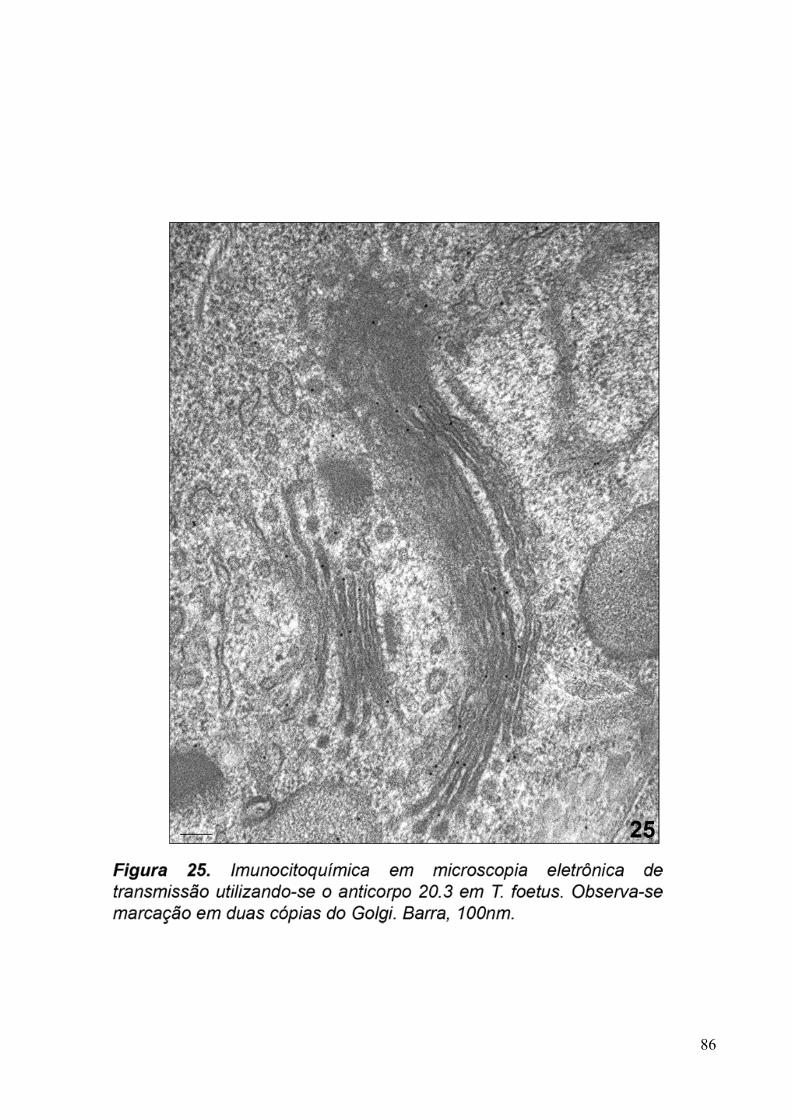
86
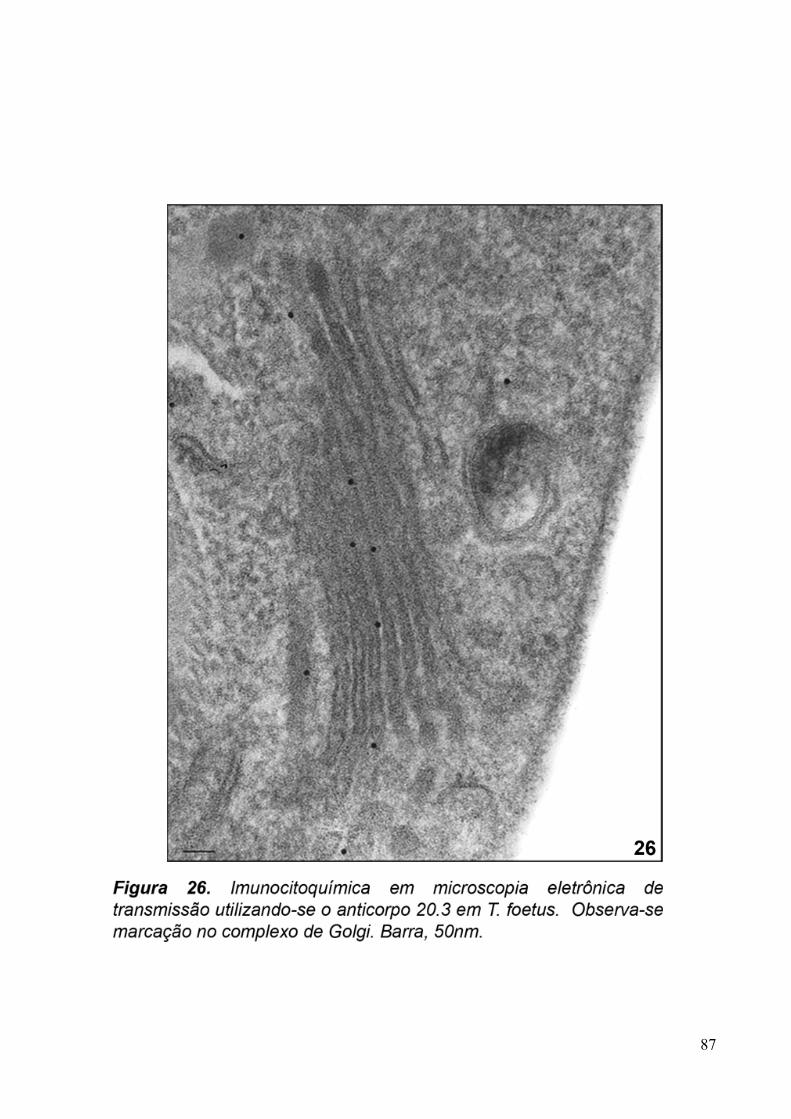
87
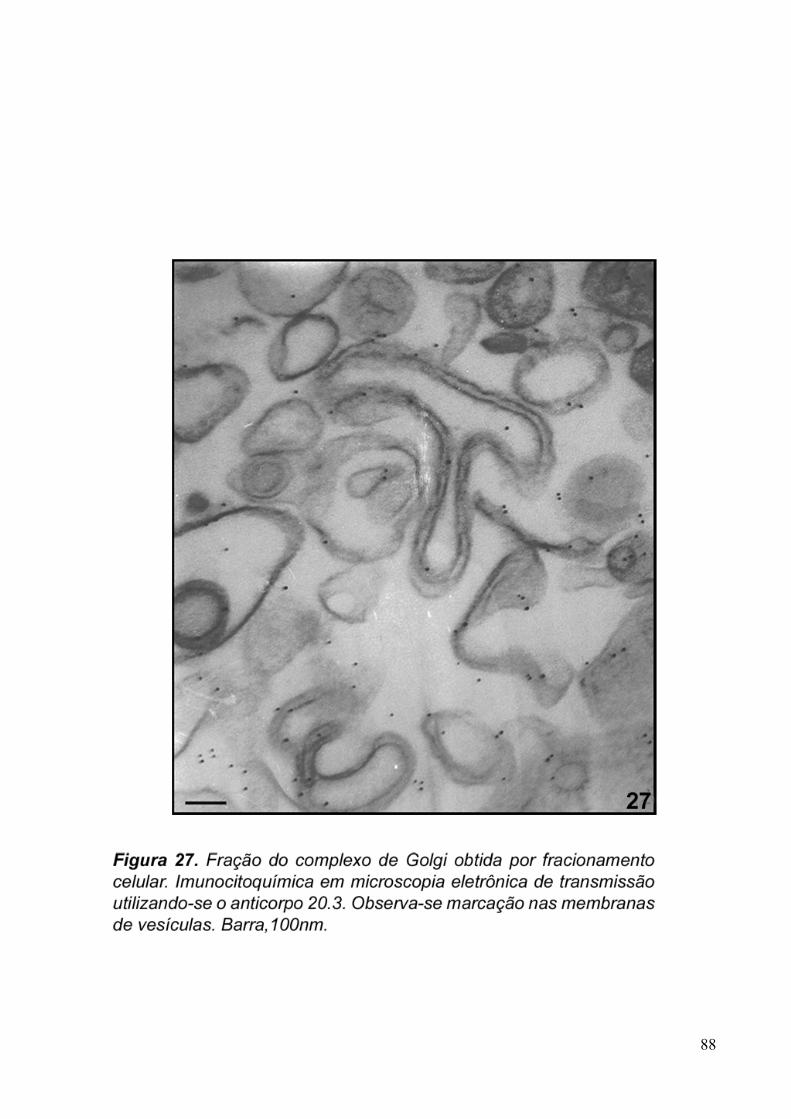
88

89
3.6 Eletroforese em gel de poliacrilamida
O perfil protéico da fração do complexo de Golgi de T. foetus (FIG. 28), observou-se
24 andas de polipeptídeos com peso molecular entre 14 e 97 kDa. Além disso, foi possível
verificar 5 bandas mais evidentes, as quais apresentam pesos moleculares de 16, 28, 38 74 e
79 kDa (#).
3.7 Immunoblotting
As proteínas do homogeneizado total foram corridas em gel, transferidas para
membranas de nitrocelulose e incubadas com o anticorpo 20.3. O anticorpo 20.3 reconheceu
duas bandas de peso molecular de 60 e 66kDa (FIG. 29).
3.8 Imunoprecipitação
O complexo proteína-anticorpo-proteína A Sepharose foi preparado misturando-se ao
tampão de amostra e aquecido a 95ºC por cinco minutos para desfazer este complexo. Depois,
as amostras foram centrifugadas por 13.000x g, por 2 minutos, para separar as proteínas da
resina, para que pudessem ser aplicadas no gel de poliacrilamida a 12%.
O gel corado com Comassie blue G-250 (FIG. 30) revelou as proteínas resultantes do
processo de imunoprecipitação (IP). As IPs realizadas com o anticorpo 20.3 nas frações PNS,
FG1 (interface 0.8/1.0M) e FG2 (interface 1.0/1.2M), revelaram 5 bandas em cada amostra
(FIG. 30). As proteínas de 60 e 66 kDa, reconhecidas no immunoblotting do homogeneizado
de T. foetus, também foram detectadas nas IPs. Além destas, foram identificadas outras três
proteínas de pesos moleculares de 25, 50 e 73 kDa. No entanto, quando as amostras das IPs
foram submetidas ao immnoblotting, somente quatro bandas puderam ser localizadas nas

90
amostras PNS e FG2 (25, 50, 60 e 66 kDa) e três na fração FG1 (onde a banda de 66 kDa não
foi revelada) (FIG. 31). Em nenhuma das frações, a banda de 73 kDa foi identificada.
O anticorpo 20.3 foi aplicado no gel de poliacrilamida sem passar pela etapa de
imunoprecipitação, e em seguida, as proteínas foram coradas pela prata, para que servissem
de padrão de peso molecular de IgG (FIG. 32). Assim, as bandas de aproximadamente 25 e 50
kDa presentes em todas as amostras de IP foram classificadas como sendo referentes à cadeia
leve e pesada do anticorpo. Outra banda visível no perfil do anticorpo (FIG. 32) também
apareceu na reação de imunoprecipitação do anticorpo anti-Golgi 20.3 (FIG. 30), que mostrou
uma proteína de peso molecular de 73 kDa.
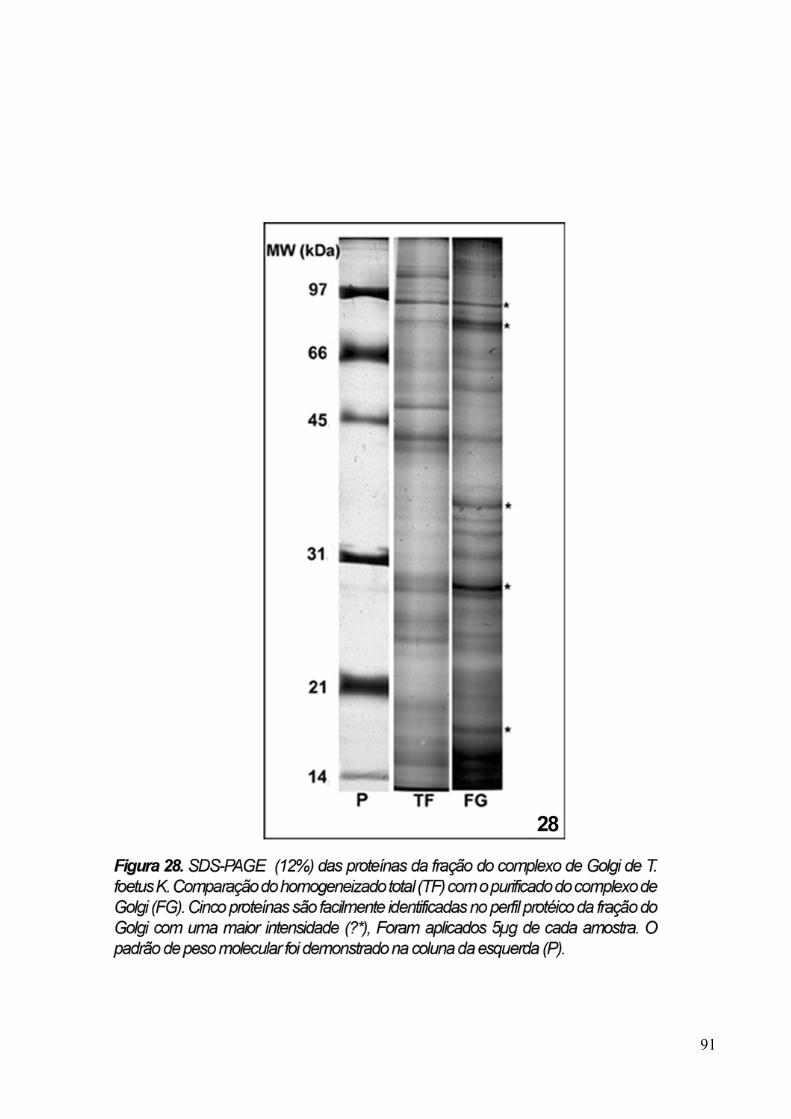
91
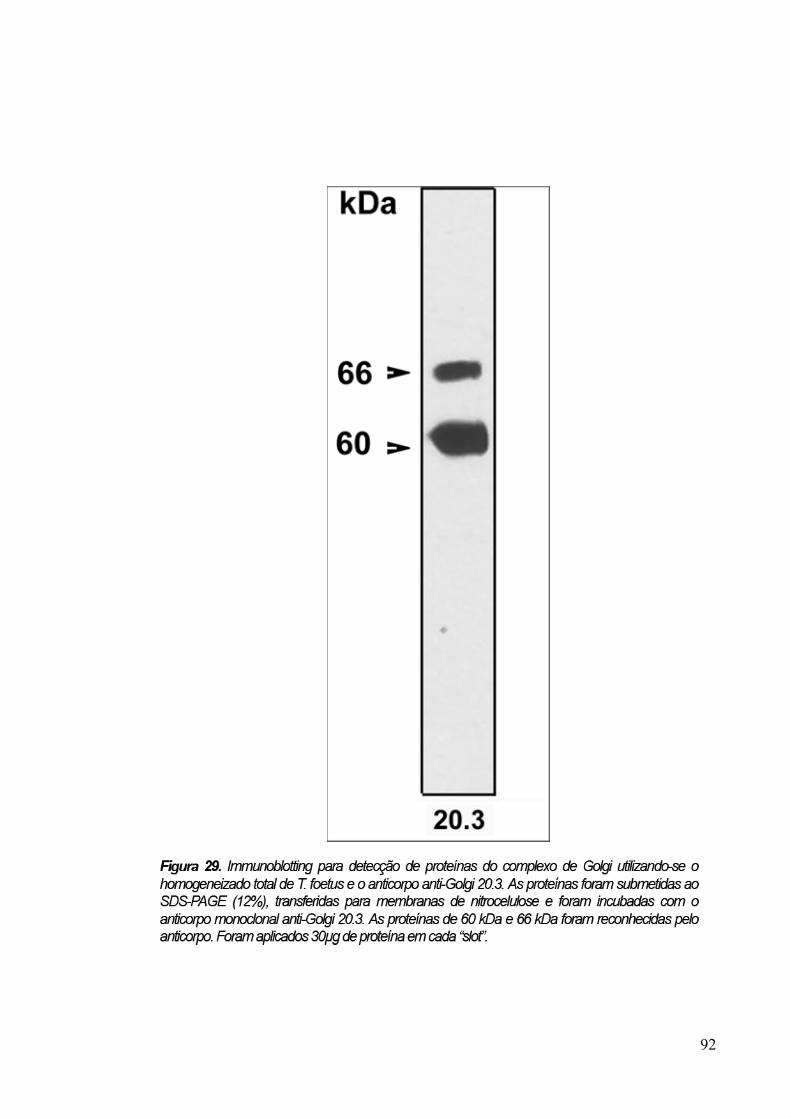
92
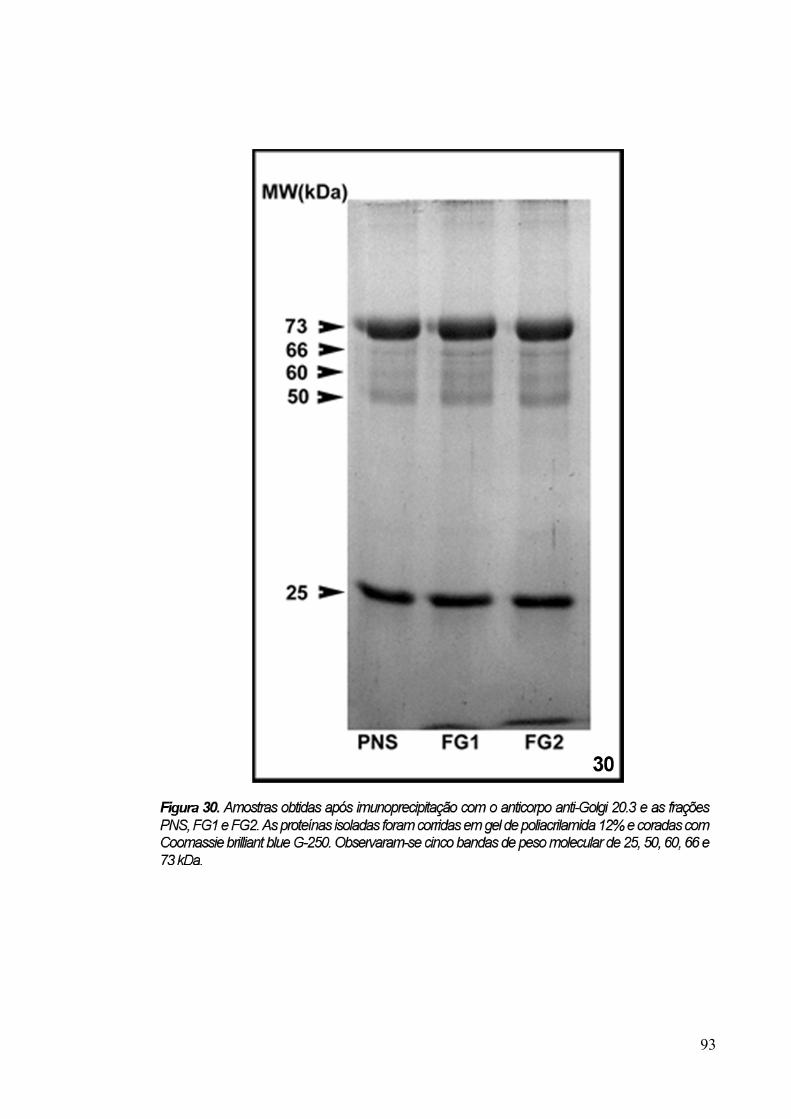
93

94
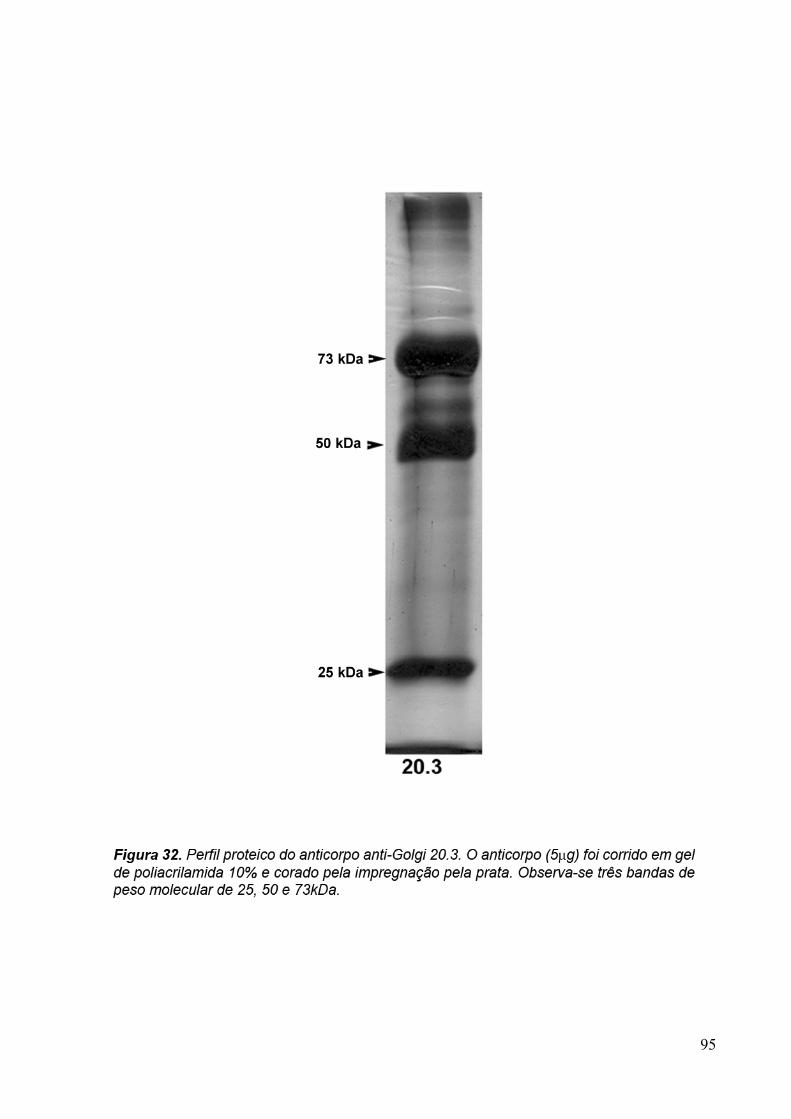
95

96
3.9 Espectrometria de Massa (MALDI-TOF/TOF)
As proteínas imunoprecipitadas pelo anticorpo anti-Golgi 20.3 foram corridas em gel
de poliacrilamida 12% e coradas com Commasie Blue G-250. As bandas obtidas foram
cortadas e os peptídeos foram extraídos do gel de acordo como descrito no material e
métodos.
As bandas referentes ao peso molecular 25 e 73 kDa, foram identificadas por
espectrometria de massa como sendo as cadeias leve (Tabela 6) e pesada (Tabela 7) da
imunoglobulina.
As massas dos peptídeos da proteína de 66 kDa foram analisadas na forma de espectro
(FIG. 33). Algumas dessas massas foram capazes de gerar seqüências de aminoácidos (Tabela
5), as quais foram analisadas no MASCOT® usando o banco de dados do NCBInt. A
comparação dessas sequências de aminoácidos com os dados do NCBInt, mostrou
similaridade com oito proteínas de T. vaginalis (Tabela 8). Além disso, estas seqüências de
aminoácidos também apresentaram similaridade com de proteínas de outros protozoários
como Leishmania, T. cruzi, T. brucei e Entamoeba dispar (Tabela 9).
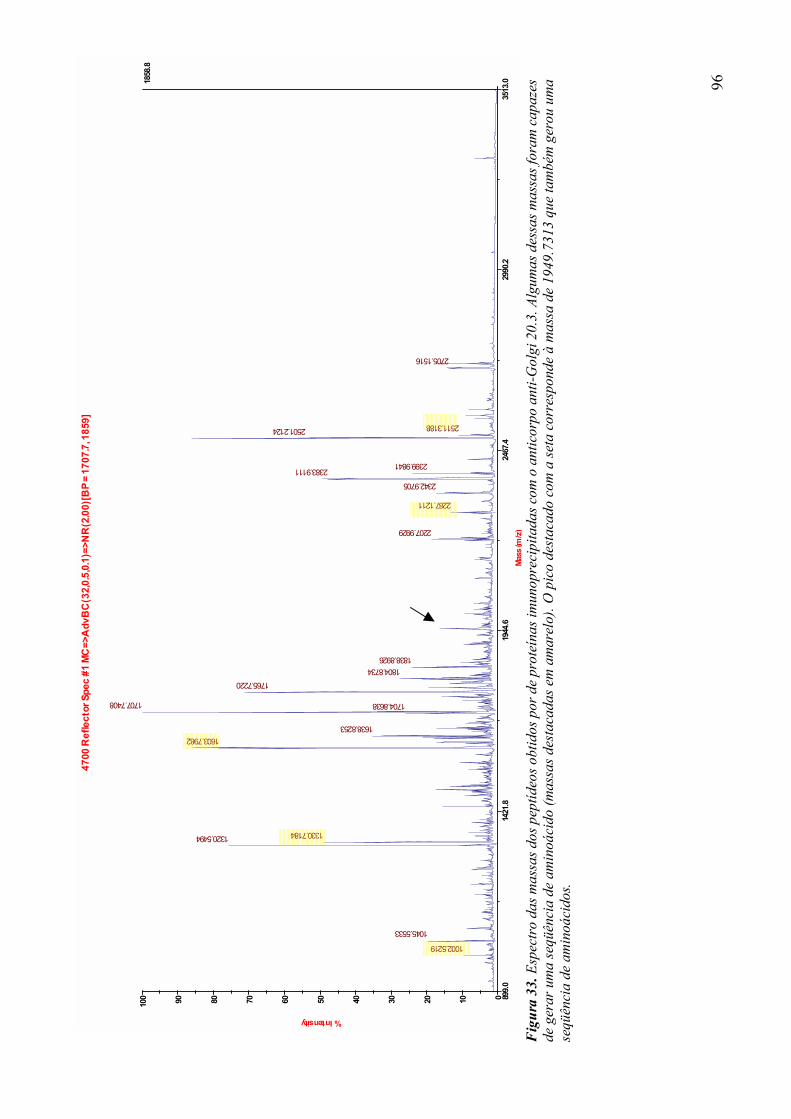
96
Fig
ura
33. E
spec
tro
das m
assa
s dos
pep
tídeo
s obt
idos
por
de
prot
eína
s im
unop
reci
pita
dasc
om o
ant
icor
po a
nti-G
olgi
20.
3. A
lgum
as d
essa
s mas
sas f
oram
cap
azes
de
ger
ar u
ma
seqü
ênci
a de
am
inoá
cido
(mas
sas d
esta
cada
s em
am
arel
o). O
pic
o de
stac
ado
com
a se
ta c
orre
spon
de à
mas
sa d
e 19
49.7
313
que
tam
bém
ger
ou u
ma
seqü
ênci
a de
am
inoá
cido
s.
899.
014
21.8
1944
.624
67.4
2990
.235
13.0
Mas
s (m
/z)
1858
.8
0102030405060708090100
% Intensity47
00 R
efle
ctor
Spe
c #1
MC
=>A
dvB
C(3
2,0.
5,0.
1)=>
NR
(2.0
0)[B
P =
1707
.7, 1
859]
1707.7408
1603.7982
1320.5494
1765.7220
2501.2124
1330.7184
2383.9111
1638.8253
1804.8734
1704.8638
1838.8926
2399.9841
1045.5533
2207.9929
2342.9705
2705.1516
2287.1211
2511.3188
1002.5219
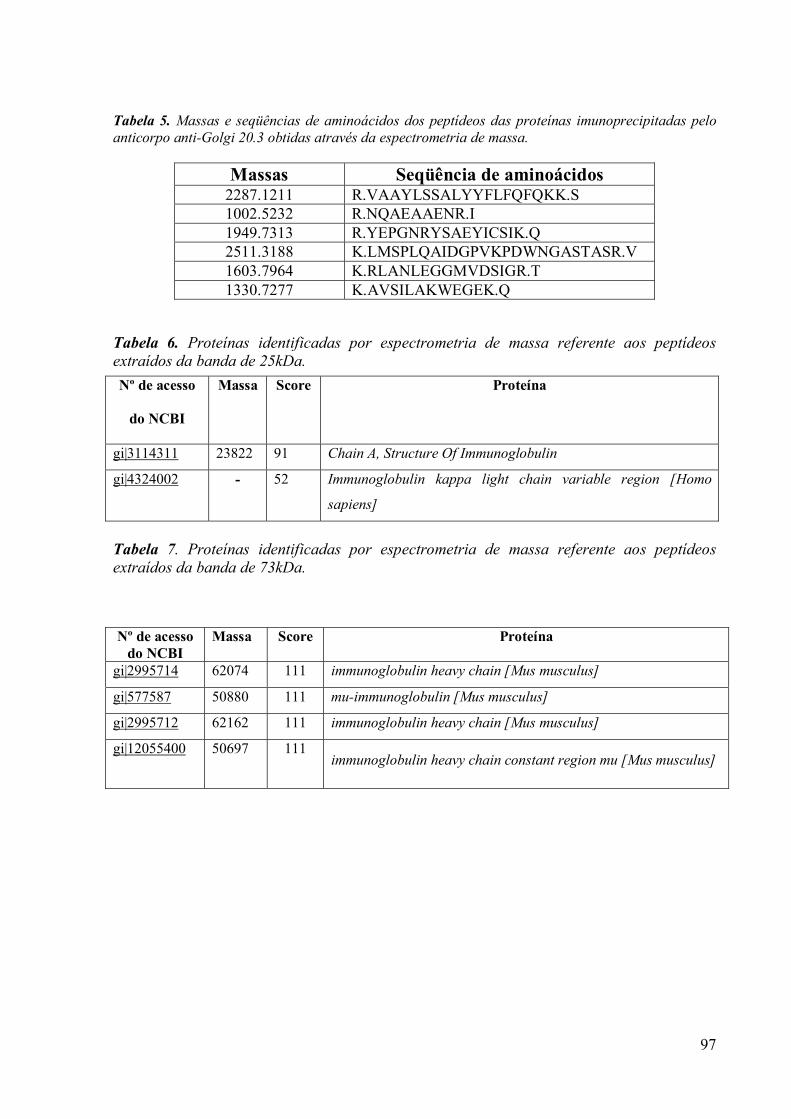
97
Tabela 5. Massas e seqüências de aminoácidos dos peptídeos das proteínas imunoprecipitadas pelo anticorpo anti-Golgi 20.3 obtidas através da espectrometria de massa.
Massas Seqüência de aminoácidos 2287.1211 R.VAAYLSSALYYFLFQFQKK.S 1002.5232 R.NQAEAAENR.I 1949.7313 R.YEPGNRYSAEYICSIK.Q 2511.3188 K.LMSPLQAIDGPVKPDWNGASTASR.V 1603.7964 K.RLANLEGGMVDSIGR.T 1330.7277 K.AVSILAKWEGEK.Q
Tabela 6. Proteínas identificadas por espectrometria de massa referente aos peptídeos extraídos da banda de 25kDa.
Tabela 7. Proteínas identificadas por espectrometria de massa referente aos peptídeos extraídos da banda de 73kDa.
Nº de acesso
do NCBI
Massa Score Proteína
gi|3114311 23822 91 Chain A, Structure Of Immunoglobulin
gi|4324002 - 52 Immunoglobulin kappa light chain variable region [Homo
sapiens]
Nº de acesso do NCBI
Massa Score Proteína
gi|2995714 62074 111 immunoglobulin heavy chain [Mus musculus]
gi|577587 50880 111 mu-immunoglobulin [Mus musculus]
gi|2995712 62162 111 immunoglobulin heavy chain [Mus musculus]
gi|12055400 50697 111 immunoglobulin heavy chain constant region mu [Mus musculus]
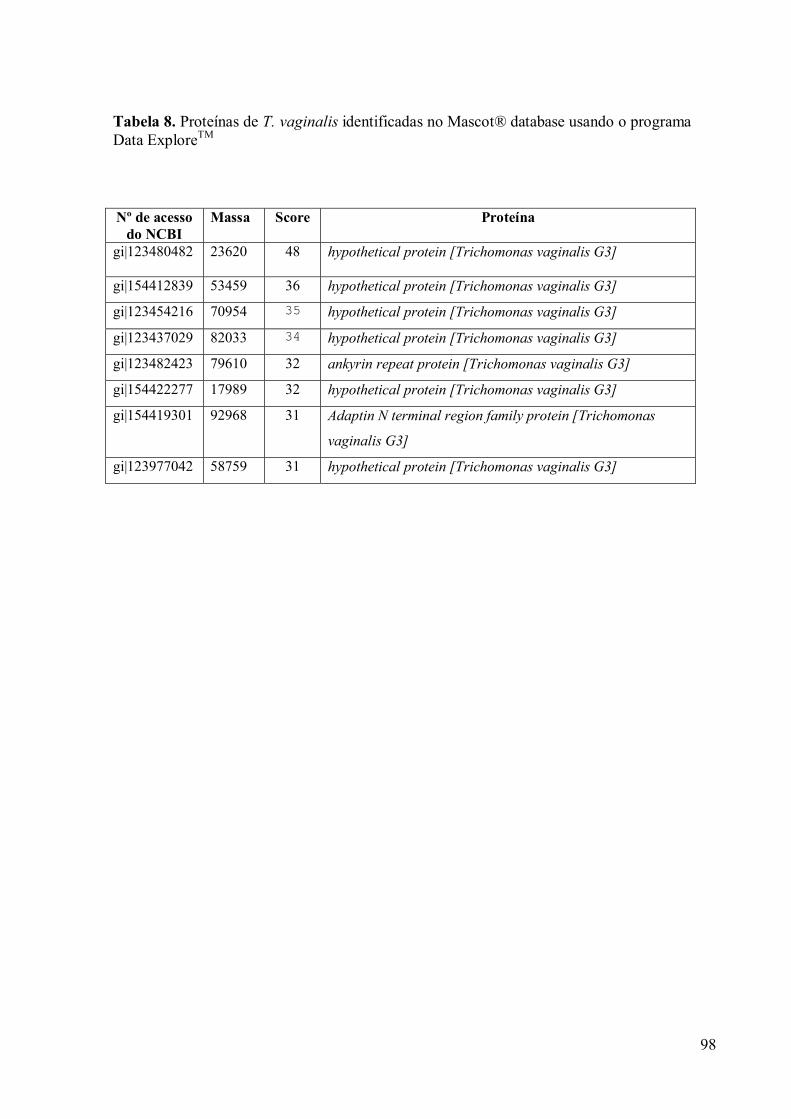
98
Tabela 8. Proteínas de T. vaginalis identificadas no Mascot® database usando o programa Data ExploreTM
Nº de acesso do NCBI
Massa Score Proteína
gi|123480482
23620 48 hypothetical protein [Trichomonas vaginalis G3]
gi|154412839 53459 36 hypothetical protein [Trichomonas vaginalis G3]
gi|123454216 70954 35 hypothetical protein [Trichomonas vaginalis G3]
gi|123437029 82033 34 hypothetical protein [Trichomonas vaginalis G3]
gi|123482423 79610 32 ankyrin repeat protein [Trichomonas vaginalis G3]
gi|154422277 17989 32 hypothetical protein [Trichomonas vaginalis G3]
gi|154419301 92968 31 Adaptin N terminal region family protein [Trichomonas
vaginalis G3]
gi|123977042 58759 31 hypothetical protein [Trichomonas vaginalis G3]
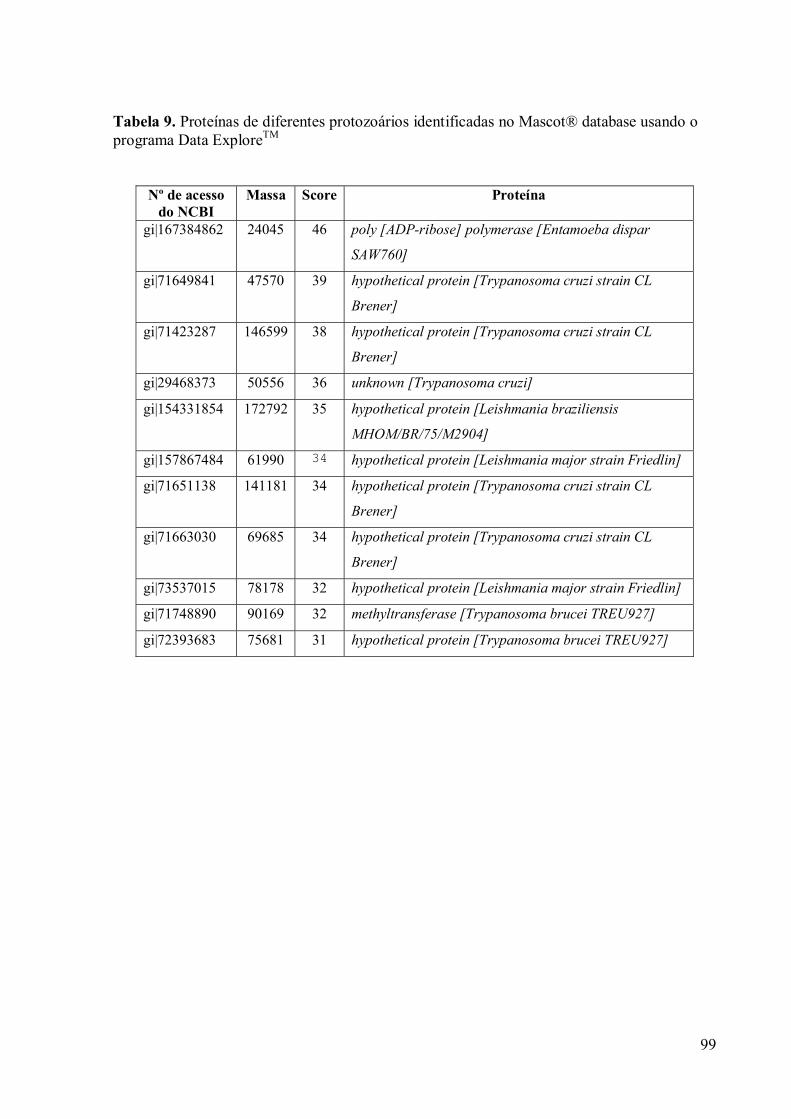
99
Tabela 9. Proteínas de diferentes protozoários identificadas no Mascot® database usando o programa Data ExploreTM
Nº de acesso do NCBI
Massa Score Proteína
gi|167384862 24045 46 poly [ADP-ribose] polymerase [Entamoeba dispar
SAW760]
gi|71649841 47570 39 hypothetical protein [Trypanosoma cruzi strain CL
Brener]
gi|71423287 146599 38 hypothetical protein [Trypanosoma cruzi strain CL
Brener]
gi|29468373 50556 36 unknown [Trypanosoma cruzi]
gi|154331854 172792 35 hypothetical protein [Leishmania braziliensis
MHOM/BR/75/M2904]
gi|157867484 61990 34 hypothetical protein [Leishmania major strain Friedlin]
gi|71651138 141181 34 hypothetical protein [Trypanosoma cruzi strain CL
Brener]
gi|71663030 69685 34 hypothetical protein [Trypanosoma cruzi strain CL
Brener]
gi|73537015 78178 32 hypothetical protein [Leishmania major strain Friedlin]
gi|71748890 90169 32 methyltransferase [Trypanosoma brucei TREU927]
gi|72393683 75681 31 hypothetical protein [Trypanosoma brucei TREU927]

100
4. DISCUSSÃO
O complexo de Golgi bastante desenvolvido em tricomonadídeos é um provável
indicador de funções importantes análogas às descritas em outras células (Müller, 1990).
Porém, ainda são poucos os estudos de isolamento e/ou análises bioquímicas voltados para
esta organela. Em 1996, Díaz e colaboradores descreveram uma técnica de isolamento do
complexo de Golgi de T. foetus. Os autores apontaram o fracionamento celular como sendo
uma técnica essencial para a obtenção de dados bioquímicos e para um melhor entendimento
da organização e função das estruturas intracelulares. Foi a partir desta técnica que também se
obtiveram os primeiros resultados da caracterização bioquímica do perfil protéico da fração
do Golgi (DÍAZ & DE SOUZA, 1998). Posteriormente, Almeida e colaboradores (2003)
sugeriram a presença de uma Ca++ ATPase no Golgi de T. foetus, após estudos utilizando o
fracionamento celular e inibidores para esta enzima. Estes resultados serviram para confirmar
a presença desta proteína no complexo de Golgi, já demonstrada anteriormente através de
estudos citoquímicos (BENCHIMOL et al., 2001). Isso corroborou para demonstrar o
envolvimento desta organela na regulação de Ca++ na célula junto com os hidrogenossomos e
o retículo endoplasmático (BENCHIMOL et al., 2001).
Em estudos do Golgi em células de mamífero, o uso da técnica de fracionamento
celular utilizando gradiente de sacarose para obtenção de uma fração purificada, demonstrou a
capacidade de isolar a maioria das proteínas identificadas do complexo de Golgi
(WEINSTEIN et al., 1982; MOREMAN et al., 1991; DUDEN et al., 1991). Entre estas
proteínas destacam-se enzimas envolvidas no processamento de oligossacarídeos, proteínas de
transporte vesicular e proteínas integrais e associadas à membrana, como: a
galactosiltransferase (GERBER et al., 1979), sialiltransferases (WEINSTEIN et al., 1982), α-
manosidade II (MOREMAN et al., 1991) e a βCOP, uma proteína periférica da membrana do
Golgi (DUDEN et al., 1991).

101
Em T. cruzi, protozoário parasito que apresenta um complexo de Golgi pouco
desenvolvido, o gradiente de sacarose se mostrou eficiente para o isolamento desta organela.
Isto tornou possível a identificação de uma proteína de 58 kDa, a qual os autores sugerem que
a sua função in vivo esteja relacionada ao ancoramento do complexo de Golgi a microtúbulos
do citoesqueleto (MORGADO-DÍAZ et al., 2001), como já descrito em células de mamífero
(BLOOM & BRASHEAR, 1989).
O uso de anticorpos monoclonais tem sido empregado como uma ferramenta
importante para a caracterização de diversas estruturas celulares em protozoários. Em T.foetus
anticorpos monoclonais foram usados para purificar e caracterizar proteínas de superfície
envolvidas na adesão do parasito a célula hospedeira (HODGSON et al.,1990; SHAIA et al.,
1998). Uma nova proteína de atractóforo (estrutura semelhante ao centrossomo), a P477,
também foi descrita após a seleção de anticorpos monoclonais produzidos a partir de um
isolado do citoesqueleto de T. vaginalis (BRIECHEUX et al., 2007). Com o anticorpo
específico os autores conseguiram caracterizar a estrutura da proteína e estabelecer sua
funcionalidade.
Para estudos relacionados com o complexo de Golgi de tricomonadídeos, até o
momento só foram utilizados anticorpos produzidos contra outros organismos ou proteínas
não específicas do Golgi, mas que apresentassem reação cruzada com esta organela. Desta
forma, foi possível determinar através de imunocitoquímica utilizando um anticorpo
policlonal para DNaK de Eschirichia coli, a localização de Hsp70 e Hsp60 no complexo de
Golgi, retículo endoplasmático e nos hidrogenossomos (BOZNER, 1997). Além disso, o
comportamento do Golgi durante a divisão celular de T. vaginalis foi acompanhado por
imunofluorescência utilizando os anticorpos anti-adesina AP51 e AP65 (BENCHIMOL et al.,
2001). Assim, anticorpos produzidos contra o complexo de Golgi de tricomonadídeos são
necessários para se obter informações mais específicas sobre esta organela, como sua

102
composição protéica e funcionalidade. Em 1999, Lingnau e colaboradores realizaram a
técnica de fracionamento celular para obter vesículas endossomais e lisossomais de T. brucei.
Esta fração foi utilizada para produzir anticorpos monoclonais com a finalidade de
caracterizar uma proteína de 85kDa, chamada proteína 1 do Golgi/Lisossomo. Por estudos
imunocitoquímicos foi verificada a localização desta proteína no complexo de Golgi e
lisossoma de T. brucei. Além disso, o anticorpo produzido apresentou reatividade com
Leishmania amazonensis e Toxoplasma gondii, demonstrando que esta proteína possa ter a
função conservada. Neste trabalho seguimos metodologias semelhantes às descritas por
Lingnau e colaboradores (1999), com o objetivo de caracterizar uma proteína do complexo de
Golgi de T. foetus. Assim, obtivemos anticorpos produzidos a partir do isolado do Golgi, que
por sua vez teve sua eficiência comprovada nos ensaios de imunofluorescência e
imunocitoquímica, onde observamos marcações restritas ao complexo de Golgi de
tricomonas, semelhante à marcação observada por Benchimol e colaboradores (2001). Além
disso, o anticorpo anti-Golgi 20.3 reconheceu proteínas de T. vaginalis, L. amazonensis,
T.cruzi e MDCK. Entretanto, a marcação encontrada nos tripanosomatídeos foi bastante
distinta, localizada na região posterior da célula e ao redor do núcleo. Comparado nossos
resultados com aqueles obtidos em estudos utilizando marcadores de retículo endoplasmático
em Leishmania major (KNUEPFER et al.,2001), sugerimos que o anticorpo produzido esteja
reconhecendo uma proteína desta organela. A marcação no retículo endoplasmático destes
protozoários pode ser devido à íntima relação desta organela com o complexo de Golgi. Esta
mistura entre os componentes do RE-Golgi inclusive com o envelope nuclear também foi
observada em Giardia lamblia (SOLTYS et al., 1996), leveduras (VORISEK, 1998) e
Toxoplasma gondii (LINGNAU et al., 1999).
A afinidade dos anticorpos por proteínas do Golgi foi satisfatória para os estudos
bioquímicos, uma vez que estas moléculas foram capazes de reconhecer proteínas na fração

103
purificada desta organela. O fato de o anticorpo mostrar reação em duas bandas no gel de
eletroforese (60 e 66kDa), pode indicar duas proteínas distintas ou uma mesma proteína com
diferentes modificações co- e pós-translacionais, as quais conferem diferentes pesos
moleculares. Estas modificações podem acarretar mudanças na localização, nas interações
moleculares ou ainda nas atividades das proteínas (LARSEN & ROEPSTORFF, 2000). A
presença de cinco bandas nos géis de imunoprecipitação tornou necessária a distinção das
bandas referentes as proteínas do complexo de Golgi e das bandas do anticorpo. Por isso,
realizamos o SDS-PAGE do anticorpo 20.3, onde foi possível verificar que além das bandas
de 25 e 50kDa, a banda de 73kDa também estava presente nesta amostra. Esses dados foram
então confirmados por espectrometria de massa, que identificou as bandas de 25 e 50kDa
como sendo as cadeias leve e pesada da imunoglobulina de camundongo, respectivamente.
Entretanto, a análise da banda de 73kDa também foi identidicada como sendo a cadeia pesada
da imunoglobulina de camundongo. Assim, acreditamos que esta banda possa ser de
imunoglobulinas que não foram totalmente dissociadas ou que estejam conjugadas a alguma
proteína usada na solução onde os anticorpos foram ressuspensos após a etapa de purificação
no processo de obtenção de anticorpos monoclonais.
Para a identificação da proteína reconhecida pelo anticorpo foi necessário combinar a
técnica de fracionamento celular com a espectrometria de massa, obtendo assim, uma
poderosa abordagem para este fim. Normalmente, a proteômica de organelas faz uso de géis
uni- e bidimensionais acompanhados da identificação da proteína por espectrometria de massa
(WU et al., 2004). Entretanto, neste trabalho utilizamos o anticorpo produzido como
ferramenta para isolar a proteína do Golgi por meio da imunoprecipitação, por isso, não foi
necessária a realização do gel bidimensional. Por se tratar de um trabalho novo em T. foetus,
encontramos dificuldades para estabelecer protocolos de purificação e identificação através de
técnicas proteômicas. Uma das maiores dificuldades durante a identificação das proteínas

104
reconhecidas pelo anticorpo do Golgi foi o fato de não existir um banco genômico de T. foetus
disponível. Assim, nossas análises tiveram que ser realizadas com base no genoma de T.
vaginalis, por se tratar do organismo mais próximo filogeneticamente, e que apresenta o
genoma já seqüenciado. Além disso, verificamos neste trabalho que o anticorpo produzido
reconhece também o complexo de Golgi deste protozoário. Desta forma, utilizando os
parâmetros descritos em estudos proteômicos de T. vaginalis (DE JESUS et al., 2007;
CUERVO et al., 2008), conseguimos identificar oito proteínas de T. vaginalis que
apresentaram similaridade com as seqüência de aminoácidos gerados durante a análise por
espectrometria de massa. As proteínas hipotéticas, as quais apresentam funções
desconhecidas, representam grandes candidatas para a exploração de novas funções ou até
mesmo para integrar informações a esta organela. Além disso, a identificação por
espectrometria de massa apontou similaridade com a região N-terminal da adaptina e com
repetições de anquirina. Estas regiões são tidas como domínios conservados de proteínas
presente em todos os organismos, incluindo procariontes, alguns vírus e principalmente em
eucariotos (MOSAVI et al., 2004). As repetições de anquirina agem como suporte para a
interações moelulares, que são extremamente importantes, pois medeiam a interação proteína-
proteína, além de interagir com muitas ouras proteínas como: canais iônicos, moléculas de
adesão celular, tubulina, clatrina e outras (BENNET & CHEN, 2001). Proteínas com
repetições de anquirina localizadas no retículo endoplasmático, sugerem que sua função esteja
relacionada com mudanças morfológicas desta organela ou esteja envolvida na síntese e
transporte de proteínas para o complexo de Golgi (KURIYAMA et al., 1999). Entretanto,
outras abordagens são necessárias para que possamos confirmar estes dados. A derivatização
da proteína, o seqüenciamento manual e a utilização do Q-TOF são metodologias que serão
adotadas para este fim.

105
5. CONCLUSÕES
No presente estudo obtivemos um anticorpo monoclonal que reconhece o complexo de
Golgi de T. foetus, que pode ser usado em trabalhos futuros como uma ferramenta que ajude a
elucidar questões sobre esta organela. Além disso, estabelecemos protocolos de métodos
bioquímicos e proteômicos que podem ser usados não somente no estudo do complexo de
Golgi, mas também de outras estruturas celulares de T. foetus.

106
6. REFERÊNCIAS AFFONSO, A. L., BENCHIMOL, M., RIBEIRO, K. C., LINS, U., DE SOUZA, W. Further
studies on the endocytic activity of Tritrichomonas foetus. Parasitol Res. v. 80, p. 403-413. 1994.
AFFONSO, A. L., DE ALMEIDA, J. C., BENCHIMOL, M. Partial characterization of
cytoplasmic compartments involved in the endocytic process of Tritrichomonas foetus. Eur J Cell Biol. v. 72, p. 247-256. 1997.
ALBERTS, B. JOHNSON, A., LEWIS, J., RAFF, M., ROBERTS, K., WALTER, P. Biologia
Molecular da Célula. Atmed 4ªed. Porto Alegre. 2004. p. 731. ALDERETE, J. F., PEARLMAN, E. Pathogenic Trichomonas vaginalis cytotoxicity to cell
culture monolayers. Br J Vener Dis. v. 60, p. 99-105. 1984. ALMEIDA, J. C. Aspectos bioquímicos e ultraestrututrais das organelas envolvidas nos
sistemas de captação de cálcio de Tritrichomonas foetus e Neocallimastix frontalis: o papel do hidrogenossomo e do complexo de Golgi. Rio de Janeiro: Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro. p. 52, 2000.
ALMEIDA, J. C., BENCHIMOL, M., DE SOUZA, W., OKOROKOV, L. A. Ca(2+)
sequestering in the early-branching amitochondriate protozoan Tritrichomonas foetus: an important role of the Golgi complex and its Ca(2+)-ATPase. Biochim Biophys Acta. v. 1615, p. 60-68. 2003.
ALSTAD. A. D., KROGH, D., FISCHER, K., GUSTAFSON, S., CASSEL, G., REICHERT,
BELDEN, W. J., BASZLER, T. Trichomoniasis in a beef herd. Vet Med. v.79, p.708-709. 1984.
ALVAREZ-SÁNCHEZ ME, AVILA-GONZÁLEZ L, BECERRIL-GARCÍA C, FATTEL-
FACENDA LV, ORTEGA-LÓPEZ J, ARROYO R. A novel cysteine proteinase (CP65) of Trichomonas vaginalis involved in cytotoxicity. Microb Pathog. v. 28, p. 193-202. 2000.
ARROYO, R., ALDERETE, J. F. Trichomonas vaginalis surface proteinase activity is
necessary for parasite adherence to epithelial cells. Infect Immun. v. 57, p. 2991-2997. 1989.
ARROYO, R., ALDERETE, J. F. Two Trichomonas vaginalis surface proteinases bind to
host epithelial cells and are related to levels of cytoadherence and cytotoxicity. Arch Med Res. v. 26, p. 279-285. 1995.
BABÁL, P., RUSSELL, L. C. Sialic acid-specific lectin-mediated adhesion of Tritrichomonas
foetus and Tritrichomonas mobilensis. J Parasitol. v. 85, p. 33-40. 1999. BASTIDA-CORCUERA F, BUTLER JE, HEYERMANN H, THOMFORD JW, CORBEIL
LB. Tritrichomonas foetus extracellular cysteine proteinase cleavage of bovine IgG2 allotypes. J Parasitol. v. 86, p. 328-332. 2000.

107
BENCHIMOL, M., ELIAS, C. A., DE SOUZA, W. Tritrichomonas foetus: ultrastructural
localization of basic proteins and carbohydrates.Exp Parasitol. v. 54, p.135-144. 1982a.
BENCHIMOL, M., ELIAS, C. A., DE SOUZA, W. Tritrichomonas foetus: ultrastructural
localization of calcium in the plasma membrane and in the hydrogenosome. Exp Parasitol. v. 54, p. 277-284. 1982b.
BENCHIMOL, M., DE SOUZA, W. Fine structure and cytochemistry of the hydrogenosome
of Tritrichomonas foetus. J Protozool. v. 30, p. 422-425. 1983. BENCHIMOL, M., DE SOUZA, W. Tritrichomonas foetus: cytochemical visualization of the
endoplasmic reticulum-Golgi complex and lipids. Exp Parasitol. v. 59, p. 51-58. 1985. BENCHIMOL, M., DA CUNHA E SILVA, N. L., ELIAS, C. A., DE SOUZA, W.
Tritrichomonas foetus: ultrastructure and cytochemistry of endocytosis. Exp Parasitol. v. 62, p. 405-415. 1986.
BENCHIMOL, M., KACHAR, B., DE SOUZA, W. Surface domains in the pathogenis
protozoan Tritrichomonas foetus. J. Protozool. v. 34, p.480-484, 1992. BENCHIMOL, M., KACHAR, B., DE SOUZA, W. The structural organization of the
pathogenic protozoan Tritrichomonas foetus as seen in replicas of quick frozen, freeze-fractured and deep etched cells. Biol Cell. v. 77, p.289-295. 1993.
BENCHIMOL, M., DE SOUZA W. Carbohydrate involvement in the association of a
prokaryotic cell with Trichomonas vaginalis and Tritrichomonas foetus. Parasitol Res. v. 81, p. 459-464. 1995.
BENCHIMOL, M., ALMEIDA, J. C., DE SOUZA, W. Further studies on the organization of
the hydrogenosome in Tritrichomonas foetus. Tissue Cell. v. 28, p. 287-299. 1996. BENCHIMOL, M., JOHNSON, P. J., DE SOUZA, W. Morphogenesis of the
hydrogenosome: an ultrastructural study. Biol Cell. v. 87, p. 197-205. 1996. BENCHIMOL M. Hydrogenosome autophagy: an ultrastructural and cytochemical study.
Biol Cell. v. 91, p. 165-174. 1999. BENCHIMOL, M., DINIZ, J. A., RIBEIRO, K. The fine structure of the axostyle and its
associations with organelles in Trichomonads. Tissue Cell. v. 32, p. 178-187. 2000. BENCHIMOL, M., RIBEIRO, K. C., MARIANTE, R. M., ALDERETE, J. F. Structure and
division of the Golgi complex in Trichomonas vaginalis and Tritrichomonas foetus. Eur J Cell Biol. v. 80, p. 593-607. 2001.
BENCHIMOL, M., BERNARDINO, M. V. Ultrastructural localization of glycoconjugates in
Tritrichomonas foetus. Parasitol Res. v. 88, p. 134-143. 2002.

108
BENCHIMOL, M. Trichomonads under microscopy. Microsc Microanal. v.10, p. 528-550. 2004.
BENCHIMOL, M., DE ANDRADE ROSA, I., DA SILVA FONTES, R., BURLA DIAS, A.
J. Trichomonas adhere and phagocytose sperm cells: adhesion seems to be a prominent stage during interaction. Parasitol Res. v. 102, p. 597-604. 2008.
BENNETT, V., CHEN, L. Ankyrins and cellular targeting of diverse membrane proteins to
physiological sites. Curr Opin Cell Biol. v. 13, p: 61-67. 2001. BERTACHINI-LOMBELLO, C., LOURENÇO, L. B., CARVALHO, H. F. Complexo de
Golgi. In: Hernandes F. Carvalho; Shirlei M. Recco-Pimentel. (Org.). A célula 2001. 1 ed.: Manole, 2001, v.12, p. 138-148.
BIAGINI, G. A., FINLAY, B. J., LLOYD, D. Evolution of the hydrogenosome. FEMS
Microbiol Lett. v. 155, p. 133-140. 1997. BLOOM, G. S., BRASHEAR, T. A. A novel 58-kDa protein associates with the Golgi
apparatus and microtubules. J Biol Chem. v. 264, p. 16083-16092. 1989. BONDURANT, R. H. Diagnosis, treatment and control of bovine trichomoniaisis. Comp
Cont Ed Pratc Vet Res. v.7, p.179-188. 1985. BONILHA, V. L., CIAVAGLIA, M. D. O. C., DE SOUZA, W., SILVA FILHO, F. The
involvement of terminal carbohydrates of the mammalian cell surface in the cytoadhesion of trichomonads. Parasitol Res. v. 81, p. 121-126. 1995.
BOZNER P. Immunological detection and subcellular localization of Hsp70 and Hsp60
homologs in Trichomonas vaginalis. J Parasitol. v. 83, p. 224-229. 1997. BRICHEUX, G., COFFE, G., BRUGEROLLE, G.. Identification of a new protein in the
centrosome-like "atractophore" of Trichomonas vaginalis. Mol Biochem Parasitol. v. 153, p. 133-140. 2007.
BRUGEROLLE, G. Demonstration of the endocytosis process and lysosome structures in
Trichomonas vaginalis. C R Acad Sci Hebd Seances Acad Sci D. v. 272, p. 2558-2560. 1971.
BRUGEROLLE, G., BRICHEUX, G., COFFE, G. Centrin protein and genes in Trichomonas
vaginalis and close relatives. J Eukaryot Microbiol. v. 47, p. 129-138. 2000. BUI, E. T., BRADLEY, P. J., JOHNSON, P. J. A common evolutionary origin for
mitochondria and hydrogenosomes. Proc Natl Acad Sci USA. v. 93, p. 9651-9656. 1996.
CAVALIER-SMITH, T., CHAO, E. E. Molecular phylogeny of the free-living archezoan
Trepomonas agilis and the nature of the first eukaryote. J Mol Evol. v. 43, p. 551-562. 1996.

109
CERKASOVOVÁ, A., CERKASOV, J., KULDA, J., REISCHIG, J. Circular DNA and cardiolipin in hydrogenosomes, microbody-like organelles of trichomonads. Folia Parasitol (Praha). v. 23, p. 33-37. 1976.
CLEMENS, D. L., JOHNSON, P. J. Failure to detect DNA in hydrogenosomes of
Trichomonas vaginalis by nick translation and immunomicroscopy. Mol Biochem Parasitol. v. 106, p. 307-313. 2000.
CROUCH, M. L., ALDERETE, J. F. Trichomonas vaginalis interactions with fibronectin and
laminin. Microbiology. v. 145, p. 2835-2843. 1999. CUERVO, P., CUPOLILLO, E., BRITTO, C., GONZÁLEZ, L. J., E SILVA-FILHO, F. C.,
LOPES, L. C., DOMONT, G. B., DE JESUS, J. B. Differential soluble protein expression between Trichomonas vaginalis isolates exhibiting low and high virulence phenotypes. J Proteomics. v. 30, p. 109-122. 2008.
DACKS, J. B., REDFIELD, R. J. Phylogenetic placement of Trichonympha. J Eukaryot
Microbiol. v. 45, p. 445-447. 1998. DEAN, N., PELHAM, H. R. Recycling of proteins from the Golgi compartment to the ER in
yeast. J Cell Biol. v. 111, p: 369-377. 1990. DE ANDRADE ROSA, I., EINICKER-LAMAS, M., RONEY BERNARDO, R.,
PREVIATTO, L. M., MOHANA-BORGES, R., MORGADO-DÍAZ, J.A., BENCHIMOL, M. Cardiolipin in hydrogenosomes: evidence of symbiotic origin. Eukaryot Cell. v. 5, p: 784-787. 2006.
DE CARLI, G. A., TASCA, T., PIRES BORGES, F. Tritrichomonas foetus: a scanning
electron microscopy study of erythrocyte adhesion associated with hemolytic activity. Vet Res. v. 35, p. 123-130. 2004.
DE JESUS, J. B., CUERVO, P., JUNQUEIRA, M., BRITTO, C., SILVA-FILHO, F. C.,
SOARES, M. J., CUPOLILLO, E., FERNANDES, O., DOMONT, G. B. A further proteomic study on the effect of iron in the human pathogen Trichomonas vaginalis. Proteomics. v. 7, p. 1961-1972. 2007.
DE SOUZA, W., BENCHIMOL, M. Electron spectroscopic imaging of calcium in the
hydrogenosomes of Tritrichomonas foetus. J Submicrosc Cytol Pathol. v. 20, p. 619-621. 1988.
DIAMOND, L. S. The estabilishment of various trichomonads of animals and man in axenic
cultures. J Parasitol. v. 43, p. 488-490. 1957. DÍAZ, J. A., MONTEIRO-LEAL, L. H., DE SOUZA W. Tritrichomonas foetus: isolation and
characterization of the Golgi complex. Exp Parasitol. v. 83, p. 174-183. 1996. DÍAZ, J. A., DE SOUZA, W. Biochemical characterization of the Golgi-complex protein of
Tritrichomonas foetus Parasitol Res. v. 8, p. 760-762. 1998.

110
DICK, G., GRØNDAHL, F., PRYDZ, K. Overexpression of the 3'-phosphoadenosine 5'-phosphosulfate (PAPS) transporter 1 increases sulfation of chondroitin sulfate in the apical pathway of MDCK II cells. Glycobiology. v. 18, p. 53-65. 2008.
DOLEZAL, P., SMÍD, O., RADA, P., ZUBÁCOVÁ, Z., BURSAĆ, D., SUTÁK, R.,
NEBESÁROVÁ, J., LITHGOW, T., TACHEZY, J. Giardia mitosomes and trichomonad hydrogenosomes share a common mode of protein targeting. Proc Natl Acad Sci USA. v. 102, p. 10924-10929. 2005.
DUBOUCHER, C., NOËL, C., DURAND-JOLY, I., GERBOD, D., DELGADO-
VISCOGLIOSI, P., JOUVESHOMME, S., LECLERC, C., CARTOLANO, G. L., DEI-CAS, E., CAPRON, M., VISCOGLIOSI, E. Pulmonary coinfection by Trichomonas vaginalis and Pneumocystis sp. as a novel manifestation of AIDS. Hum Pathol. v. 34, p. 508-511. 2003.
DUBOUCHER, C., CABY, S., DUFERNEZ, F., CHABÉ, M., GANTOIS, N., DELGADO-
VISCOGLIOSI, P., BILLY, C., BARRÉ, E., TORABI, E., CAPRON, M., PIERCE, R. J., DEI-CAS, E., VISCOGLIOSI, E. Molecular identification of Tritrichomonas foetus-like organisms as coinfecting agents of human Pneumocystis pneumonia. J Clin Microbiol. v. 44, p. 1165-1168. 2006.
DUBOUCHER, C., PIERCE, R. J., CAPRON, M., DEI-CAS, E., VISCOGLIOSI, E. Recent
advances in pulmonary trichomonosis. Trends Parasitol. v. 24, p 201-202. 2008. DUDEN, R., GRIFFITHS, G., FRANK, R., ARGOS, P., KREIS, T. E. Beta-COP, a 110 kd
protein associated with non-clathrin-coated vesicles and the Golgi complex, shows homology to beta-adaptin. Trends Cell Biol. v. 14, p. 14-19. 1991.
EMBLEY, T. M., VAN DER GIEZEN, M., HORNER, D. S., DYAL, P. L., BELL, S.,
FOSTER, P. G. Hydrogenosomes, mitochondria and early eukaryotic evolution. IUBMB Life. v. 55, p. 387-395. 2003.
FOSTER, D. M., GOOKIN, J. L., POORE, M. F., STEBBINS, M. E., LEVY, M. G. Outcome
of cats with diarrhea and Tritrichomonas foetus infection. J Am Vet Med Assoc. v. 225, p. 888-892. 2004.
GERBER, A. C. H., KOZDROMSKI, I., WYSS, S. R., BERGER, E. G. The change
heterogeneity of soluble human galactosyltranferases isolated from milk, amniotic fluid and malignant ascites. v. 93, p. 453-460.1979.
GERMOT, A., PHILIPPE, H., AND LE GUYADER, H. Presence of a mitochondrial-type 70-
kDa heat shock protein in Trichomonas vaginalis suggests a very early mitochondrial endosymbiosis in eukaryotes. Proc Natl Acad Sci USA. v. 93, p. 14614-14617. 1996.
GOOKIN, J. L., LEVY, M. G., LAW, J. M., PAPICH, M. G., POORE, M. F.,
BREITSCHWERDT, E.B., Experimental infection of cats with Tritrichomonas foetus. Am J Vet. v. 62, p. 1690-1697. 2001.

111
GOOKIN, J. L., BIRKENHEUER, A. J., ST JOHN, V., SPECTOR, M., LEVY, M. G. Molecular characterization of trichomonads from feces of dogs with diarrhea. J Parasitol. v. 91, p. 939-943. 2005.
GRIFFITHS, G., SIMONS, K. The trans Golgi network: sorting at the exit site of the Golgi
complex. Science. v. 234, p: 438-943. 1986. HAURI, H. P., SCHWEIZER, A. The endoplasmic reticulum-Golgi intermediate
compartment. Curr Opin Cell Biol. v. 4, p. 600-608. 1992. HAURI, H., APPENZELLER, C., KUHN, F., NUFER, O. Lectins and traffic in the secretory
pathway. FEBS Lett. v. 476, p. 32-37. 2000. HEATH, B. Variant mitosis in lower eukaryotes: indicators of the evolution of mitosis? Int
Ver Cytol. v. 64, p. 6-81. 1980. HODGSON, J. L., JONES, D. W., WIDDERS, P. R., CORBEIL, L. B. Characterization of
Tritrichomonas foetus antigens by use of monoclonal antibodies. Infect Immun. v. 58, p. 3078-3083. 1990.
HEGGENESS, M. H., SIMON, M., SINGER, S. J. Association of mitochondria with
microtubules in cultured cells. Proc Natl Acad Sci USA. v. 75, p. 3863-3866. 1978. HONIGBERG, B. M., MATTERN, C. F., DANIEL, W. A. Fine structure of the mastigont
system in Tritrichomonas foetus (Riedmüller). J Protozool. v. 18, p. 183-198. 1971. HONIGBERG, B. M. Trichomonad of veterinary importance In: Parasitic Protozoa (ed.
Kreier, J. P). Academic Press, New York, San Francisco, London. v.II, p. 163-273. 1978.
HONIBERG, M. B., BRUGEROLLE, G. Structure in trichomonads parasitic in humans,
Honiberg, M. B. (ed), Spreinger-Verlang, pp. 5-35. 1990. JENTOFT, N. Why are proteins O-glycosylated? Trends Biochem Sci. v. 15, p. 291-294.
1990. JONGWUTIWES. S., SILACHAMROON, U., PUTAPORNTIP, C. Pentatrichomonas
hominis in empyema thoracis. Trans R Soc Trop Med Hyg. v. 94, p. 185-186. 2000. JOHNSON, P. J., LAHTI, C. J., BRADLEY, P. J.. Biogenesis of the hydrogenosome in the
anaerobic protist Trichomonas vaginalis. J Parasitol. v. 79, p. 664-670. 1993. KNUEPFER, E., STIERHOF, Y. D., MCKEAN, P. G., SMITH, D. F. Characterization of a
differentially expressed protein that shows an unusual localization to intracellular membranes in Leishmania major. Biochem J. v. 356, p. 335-344. 2001.
KÖHLER, G., MILSTEIN, C. Continuous cultures of fused cells secreting antibody of
predefined specificity. Nature. v. 256, p. 495-497. 1975. KORNFELD S. Trafficking of lysosomal enzymes. FASEB J. v. 1, p. 462-468. 1987.

112
KRIEGER, J. N., RAVDIN, J. I., REIN, M. F. Contact-dependent cytopathogenic mechanisms of Trichomonas vaginalis. Infect Immun. v. 50, p. 778-786. 1985.
KURIYAMA, H., TAKANO, H., SUZUKI, L., UCHIDA, H., KAWANO, S., KUROIWA,
H., KUROIWA, T. Characterization of Chlamydomonas reinhardtii zygote-specific cDNAs that encode novel proteins containing ankyrin repeats and WW domains. Plant Physiol. v. 119, p. 873-884. 1999.
KUTISOVA, K., KULDA, J., CEPICKA, I., FLEGR, J., KOUDELA, B., TERAS, J.,
TACHEZY, J. Tetratrichomonads from the oral cavity and respiratory tract of humans. Parasitology. v. 131, p. 309-319. 2005.
LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature. v. 227, p. 680-685. 1970. LARSEN, A. K., ESCARGUEIL, A. E., SKLADANOWSKI, A. Resistance mechanisms
associated with altered intracellular distribution of anticancer agents. Pharmacol Ther. v. 85, p. 217-29. 2000.
LEIPE, D. D., GUNDERSON, H. J., NERAD, A. T., SOGIN, L. M., Small subunit ribosomal
RNA+ of Hexamita inflata and the quest for the first eukaryotic tree. Mol Biochem Parasitol. v. 59, p. 41-48. 1993.
LEVY, M.G., GOOKIN, J. L., POORE, M., BIRKENHEUER, A. J. DYKSTRA, M. J.,
LITAKER, R. W., Tritrichomonas foetus and not Pentatrichomonas hominis is the etiologic agent of feline trichomonal diarrhea. J Parasitol. v. 89, p 99-104. 2003.
LINGNAU, A., ZUFFEREY, R., LINGNAU, M., RUSSELL, D. G.. Characterization of
tGLP-1, a Golgi and lysosome-associated, transmembrane glycoprotein of African trypanosomes. J Cell Sci. v. 112, p. 3061-3070. 1999.
LINDMARK, D. G., MÜLLER, M. Hydrogenosome, a cytoplasmic organelle of the
anaerobic flagellate Tritrichomonas foetus, and its role in pyruvate metabolism. J Biol Chem. v. 248, p. 7724-7728. 1973.
LIPSKY, N. G., PAGANO, R. E. A vital stain for the Golgi apparatus. Science. v.228, p. 745-
747. 1985. LLOYD, D., LINDMARK, D. G., MÜLLER, M. Adenosine triphosphatase activity of
Tritrichomonas foetus. J Gen Microbiol. v. 115, p. 301-307. 1979. LUN, Z. R., CHEN, X. G., ZHU, X. Q., LI, X. R., XIE, M. Q. Are Tritrichomonas foetus and
Tritrichomonas suis synonyms? Trends Parasitol. v. 21, p. 122-125. 2005. MALLAT, H., PODGLAJEN, I., LAVARDE, V., MAINARDI, J. L., FRAPPIER, J.,
CORNET, M. Molecular characterization of Trichomonas tenax causing pulmonary infection. J Clin Microbiol. v. 42, p. 3886-3887. 2004.

113
MALLINSON, R. H., GOLDSMITH, D. J., HIGGINS, R. M., VENNING, M. C., ACKRILL, P. Acute swollen legs due to rhabdomyolysis: initial management as deep vein thrombosis may lead to acute renal failure. BMJ. v. 309, p. 1361-1362. 1994.
MALLINSON, D. J., LIVINGSTONE, J., APPLETON, K. M., LEES, S. J., COOMBS, G. H.,
NORTH, M. J. Multiple cysteine proteinases of the pathogenic protozoon Tritrichomonas foetus: identification of seven diverse and differentially expressed genes. Microbiology. v. 141, p. 3077-3085. 1995.
MATTOS, A., SOLÉ-CAVA, A. M., DECARLI. G., BENCHIMOL, M. Fine structure and
isozymic characterization of trichomonadid protozoa. Parasitol Res. v. 83, p. 290-295. 1997.
MENDOZA-LÓPEZ, M. R., BECERRIL-GARCIA, C., FATTEL-FACENDA, L. V.,
AVILA-GONZALEZ, L., RUÍZ-TACHIQUÍN, M. E., ORTEGA-LOPEZ, J., ARROYO, R. CP30, a cysteine proteinase involved in Trichomonas vaginalis cytoadherence. Infect Immun. v. 68, p. 4907-4912. 2000.
MOREMAN, K. W., TOUSTER, O., ROBBINS, P. W. Novel purification of the catalitic
domain of the Golgi (α-mannosidase II. Characterization and comparison with the intact enzyme. J Biol Chem. v. 266, p. 16876-16885. 1991.
MORGADO-DÍAZ, J. A., NAKAMURA, C. V., AGRELLOS, O. A., DIAS, W. B.,
PREVIATO. J. O., MENDONÇA-PREVIATO, L., DE SOUZA, W. Isolation and characterization of the Golgi complex of the protozoan Trypanosoma cruzi. Parasitology. v. 123, p.33-43. 2001.
MOSAVI, L. K., CAMMETT, T. J., DESROSIERS, D. C., PENG, Z. Y. The ankyrin repeat
as molecular architecture for protein recognition. Protein Sci. v. 13, p. 1435-1448. 2004.
MÜLLER, M. Biochemistry of Trichomonas vaginalis. In �Trichomonads Parasitic in
Humans� (B. M. Honigberg, Ed.). Springer-Veriag, New York. pp. 54-83.1990. MÜLLER, M. The hydrogenosome. J Gen Microbiol. v. 139, p. 2879-2889. 1993. NEALE, K. A., ALDERETE, J. F. Analysis of the proteinases of representative Trichomonas
vaginalis isolates. Infect Immun. v. 58, p. 157-162. 1990. NEUHOFF, V., AROLD, N., TAUBE, D., EHRHARDT, W. Improved staining of proteins in
polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis. v. 9, p. 255-262. 1988.
PETERSON, K. M., ALDERETE, J. F. Iron uptake and increased intracellular enzyme
activity follow host lactoferrin binding by Trichomonas vaginalis receptors. J Exp Med. v. 160, p. 398-410. 1984.
PETERSON, G. L. Determination of total protein. Methods Enzymol. v. 91, p. 95-121. 1983.

114
PETRIN, D., DELGATY, K., BHATT, R., GARBER, G. Clinical and microbiological aspects
of Trichomonas vaginalis. Clin Microbiol Rev. v. 11, p. 300-317. 1998. QUE, X., REED, S. L. Cysteine proteinases and the pathogenesis of amebiasis. Clin
Microbiol Rev. v. 13, p.196-206. 2000. QUEIROZ, R. C. B., SANTOS, L. M. S., BENCHIMOL, M., DE SOUZA, W. Cytochemical
localization of enzyme markers in Tritrichomonas foetus. Parasitol Res. 77: 561-566. 1991.
RASMUSSEN, S. E., NIELSEN, M. H., LIND, I., RHODES, J. M. Morphological studies of
the cytotoxicity of Trichomonas vaginalis to normal human vaginal epithelial cells in vitro. Genitourin Med. v. 62, p. 240- 246. 1986.
RIBEIRO, K. C. Estudo do processo mitótico em Tritrichomonas foetus. Rio de Janeiro:
CBB, Universidade Estadual do Norte Fluminense. 178p. 1997. RIBEIRO, K. C., MONTEIRO-LEAL, L. H., BENCHIMOL, M. Contributions of the
axostyle and flagella to closed mitosis in the protists Tritrichomonas foetus and Trichomonas vaginalis. J Eukaryot Microbiol. v. 47, p. 481-492. 2000.
RIBEIRO, K. C., PEREIRA-NEVES, A., BENCHIMOL, M. The mitotic spindle and
associated membranes in the closed mitosis of trichomonads. Biol Cell. v. 94, p. 157-172. 2002.
RIVERA, W. L., LUPISAN, A. J., BAKING, J. M. Ultrastructural study of a
tetratrichomonad isolated from pig fecal samples. Parasitol Res. v. 103, p. 1311-1316. 2008.
ROSENWALD, A. G., PAGANO, R. E. Intracellular transport of ceramide and its
metabolites at the Golgi complex: insights from short-chain analogs. J Biol Chem. v. 268, p. 4577-4579. 1993.
SAJID, M., MCKERROW, J. H. Cysteine proteases of parasitic organisms. Mol Biochem
Parasitol. v. 120, p. 1-21. 2002. SEMENZA JC, HARDWICK KG, DEAN N, PELHAM HR. ERD2, a yeast gene required for
the receptor-mediated retrieval of luminal ER proteins from the secretory pathway. Cell. v. 61. p. 1349-1357. 1990.
SCHWEIZER, A., FRANSEN, J. A., BÄCHI, T., GINSEL, L., HAURI, H. P. Identification,
by a monoclonal antibody, of a 53-kD protein associated with a tubulo-vesicular compartment at the cis-side of the Golgi apparatus. J Cell Biol. v. 107, p.1643-1653. 1988.
SILVA FILHO, F. C., DE SOUZA, W. The interaction of Trichomonas vaginalis and
Tritrichomonas foetus with epithelial cells in vitro. Cell Struct Funct. v. 13, p.301-310.1988.

115
SILVA FILHO, F., BREIER-SARAIVA, E. M., TOSTA, M. X., DE SOUZA W. Trichomonas vaginalis and Tritrichomonas foetus secrete neuraminidase into the culture medium. Mol Biochem Parasitol. v. 35, p. 73-78. 1989.
SINGH, B. N., LUCAS, J. J., BEACH, D. H., SHIN, S. T., GILBERT, R. O. Adhesion of
Tritrichomonas foetus to bovine vaginal epithelial cells. Infect Immun. v. 67, p. 3847-3854. 1999.
SINGH, B. N., LUCAS, J. J., HAYES, G. R., KUMAR. I., BEACH, D. H., FRAJBLAT, M.,
GILBERT, R. O., SOMMER, U., COSTELLO, C. E. Tritrichomonas foetus induces apoptotic cell death in bovine vaginal epithelial cells. Infect Immun. v. 72, p. 4151-4158. 2004.
SHAIA, C. I., VOYICH, J., GILLIS, S. J., SINGH, B. N., BURGESS, D. E. Purification and
expression of the Tf190 adhesin in Tritrichomonas foetus. Infect Immun. v. 66, p. 1100-1005. 1998.
SKEPPER, J. N. Immunocytochemical strategies for electron microscopy: choice or
compromise. J Microsc. v. 199, p. 1-36. 2000. SOLOVIEV, M., FINCH, P. Peptidomics, current status. J Chromatogr B Analyt Technol
Biomed Life Sci. v. 815, p. 11-24. 2005. SOLTYS, B. J., FALAH, M., GUPTA, R. S. Identification of endoplasmic reticulum in the
primitive eukaryote Giardia lamblia using cryoelectron microscopy and antibody to Bip. J Cell Sci. v. 109, p. 1909-1917. 1996.
STOCKDALE, H. D., SPENCER, J. A., DYKSTRA, C. C. WEST, G. S., HANKES, T.,
MCMILLAN, K. L., WHITLEY, M., BLAGBURN, B. L., Feline Trichomoniais: an emerging disease. Compend. Contin. Educ Pract Vet. v. 28, p. 463-471. 2006.
STOCKDALE H. D., GIVENS, M. D., DYKSTRA. C. C., BLAGBURN, B. L.
Tritrichomonas foetus infections in surveyed pet cats. Vet Parasitol. 2008, doi:10.1016/j.vetpar.2008.10.091.
SUTAK, R., DOLEZAL, P., FIUMERA, H. L., HRDY, I., DANCIS, A., DELGADILLO-
CORREA, M., JOHNSON, P. J., MÜLLER, M., TACHEZY, J. Mitochondrial-type assembly of FeS centers in the hydrogenosomes of the amitochondriate eukaryote Trichomonas vaginalis. Proc Natl Acad Sci USA. v. 101, p. 10368-10373. 2004.
TACHEZY, J., TACHEZY, R., HAMPL, V., SEDINOVÁ, M., VANACOVÁ, S., VRLÍK,
M., VAN RANST, M., FLEGR, J., KULDAA, J. Cattle pathogen Tritrichomonas foetus (Riedmüller, 1928) and pig commensal Tritrichomonas suis (Gruby & Delafond, 1843) belong to the same species. J Eukaryot Microbiol. v. 49, p 154-163. 2002.
TALBOT, J. A., NIELSEN, K., CORBEIL, L. B. Cleavage of proteins of reproductive
secretions by extracellular proteinases of Tritrichomonas foetus. Can J Microbiol. v. 37, p. 384-390.1991.

116
TEASDALE, R. D., JACKSON, M. R. Signal-mediated sorting of membrane proteins between the endoplasmic reticulum and the golgi apparatus. Annu Rev Cell Dev Biol. v. 12, p. 27-54.1996.
THOMFORD, J. W., TALBOT, J. A., IKEDA, J. S., CORBEIL, L. B. Characterization of
extracellular proteinases of Tritrichomonas foetus. J Parasitol. v. 82, p.112-117. 1996. TJADEN, J., HAFERKAMP, I., BOXMA, B., TIELENS, A. G., HUYNEN, M.,
HACKSTEIN, J. H. A divergent ADP/ATP carrier in the hydrogenosomes of Trichomonas gallinae argues for an independent origin of these organelles. Mol Microbiol. v. 51, p. 1439-1446. 2004.
TOWBIN, H., STAEHELIN, T., GORDON, J. Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A. v. 76, p.4350-4354. 1979.
TURNER, G., MÜLLER, M. Failure to detect extranuclear DNA in Trichomonas vaginalis
and Tritrichomonas foetus. J Parasitol. v. 69, p. 234-236. 1983. VISCOGLIOSI, E., BRUGEROLLE, G. Cytoskeleton in Trichomonads. II. Immunological
and biochemical characterization of preaxostylar fibers and undulating membranes in genus Tritrichomonas. Eur J Protistol. v. 29, p. 160-170. 1993.
VISCOGLIOSI, E., BRUGEROLLE, G. Striated fibers in trichomonads: costa proteins
represent a new class of proteins forming striated roots. Cell Motil Cytoskeleton. v. 29, p. 82-93. 1994.
VISCOGLIOSI, E., EDGCOMB, V. P., GERBOD, D., NOËL, C., DELGADO-
VISCOGLIOSI, P. Molecular evolution inferred from small subunit rRNA sequences: what does it tell us about phylogenetic relationships and taxonomy of the parabasalids? Parasite. v. 6, p. 279-291. 1999.
VORÍSEK J. Cytochemical images of secretion in Saccharomyces cerevisiae and animal cells
are different. Acta Histochem. v. 100, p: 419-438. 1998. WARTOŃ, A., HONIGBERG, B. M. Structure of trichomonads as revealed by scanning
electron microscopy. J Protozool. v. 26, p. 56-62. 1979. WEINSTEIN, J., SOUZA-E-SILVA, U. PAULSON, J. C. Purification of a Gal β1→
4GlcNAc (2(6 sialyltransferase and a Gal β1 → 3(4) GlcNAc (α2→3 sialyltranferase to homogeneity from rabbit liver. J Biol Chem. v. 257, p. 13835-13844. 1982.
WU, C. C., MACCOSS, M. J., MARDONES, G., FINNIGAN, C., MOGELSVANG, S.,
YATES, JR. 3RD., HOWELL, K. E. Organellar proteomics reveals Golgi arginine dimethylation. Mol Biol Cell. v. 15, p: 2907-2919. 2004.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























