Embed Size (px)
Citation preview

ISSN 0006�3509, Biophysics, 2013, Vol. 58, No. 3, pp. 366–372. © Pleiades Publishing, Inc., 2013.Original Russian Text © O.S. Ostroumova, S.S. Efimova, L.V. Schagina, 2013, published in Biofizika, 2013, Vol. 58, No. 3, pp. 474–480.
366
INTRODUCTION
Flavonoids is the name of a group of phenolic com�pounds prevalently of plant origin—derivatives ofbenzo�γ�pyrone, in the basis of which lies a skeletonconsisting of two benzene rings connected betweenthemselves by a three�carbon chain (propane skele�ton). As dependent on the degree of oxidation andhydroxylation of the propane fragment, the position ofphenyl radical and the magnitude of heterocycle, fla�vonoids are divided into several groups: flavonoidsproper, isoflavonoids, neoflavonoids and biflavonoids.The first group includes derivatives of flavane, flavone,aurone and chalcone. Flavonoids are encountered inplants, both in free state and in the form of glycosides.The interest in flavonoids is conditioned by the broadspectrum of their biological activity (P�vitamin activ�ity, hypotensive, sedative, cardioprotector, anti�inflammatory, antimicrobial action) and extraordinar�ily low toxicity, which testifies on prospects of creatingnew pharmacological preparations on the basis on fla�vonoids.
The amphihilicity of flavonoid molecules allowsthem to easily incorporate into biological membranes.Adsorption of flavonoids may be accompanied by achange in membrane properties, in particular itsdipole potential ϕd. Liberman and Topaly in 1969 sug�gested the existence of a potential jump at the mem�
Editor’s Note: I certify that this is a closest equivalent of the orig�inal publication with all its factual statements and terminology,phrasing and style; English title and Abstract provided byauthors. A.G.
brane–water solution border [1]. Hladky and Haydonin 1973 for the first time used the term “dipole poten�tial” and showed that in a bilayer lipid membrane anelectric potential jump must arise, conditioned by theorientation of membrane dipoles [2]. At present it isgenerally accepted to define the membrane dipolepotential as the potential jump conditioned by the spe�cific orientation of the dipoles of membrane lipids andnear�membrane water, as a result of which the electricpotential of the inner part of the bilayer turns out to bepositive relative to the membrane�surrounding waterphase [3] The dipole potential creates in the mem�brane an electric field on the order of 109 V/m. Itshould be noted that this magnitude substantiallyexceeds the value of intensity of a field (107 V/m) cre�ated by a potential difference of 70 mV at a bilayerthickness on the order of 5 nm [4]. Inasmuch as thevalue of dipole potential in the capacity of one of theconstituents determined the height of potential barrierfor transition of charged particles across the mem�brane, there can be no doubt about the dependency ofthe properties of membrane�incorporated ion�trans�porting systems on the dipole potential. By the presentmoment it is known that the dipole potential of modelbilayer lipid membranes influences the conductivity,gating charge, cooperativity of functioning and equi�librium number in the membrane of opened ion chan�nels formed by antimicrobial agents – gramicidin,alamethicin, syringomycin E, surfactin, HPA3,amphotericin B [5–15]. The dipole potential regulatesthe membrane permeability to organic ions [16, 17],ion transport with the aid of transporters (valinomy�
CELL BIOPHYSICS
Changes of Dipole Potential of Phospholipid Membranes Resulted from Flavonoid Adsorption
O. S. Ostroumova, S. S. Efimova, and L. V. SchaginaInstitute of Cytology, Russian Academy of Sciences, St. Petersburg, 194064 Russia
e�mail: [email protected] November 7, 2012; in final form, January 28, 2013
Abstract—The effects of flavonoids, phloridzin, quercetin, myricetin and biochanin A on the dipole poten�tial of planar lipid bilayers formed from dioleylphosphoethanolamine, dioleylphosphoserine, dioleoylphos�phocholine, and diphytanoylphosphocholine are investigated. The characteristic parameters of the Langmuiradsorption isotherm, the maximum changes in the membrane dipole potential at an infinitely large concen�tration of flavonoid and its dissociation constant, which reflects the affinity of flavonoid to the membrane lip�ids, are determined. Modifying effects of chalcones, flavonols and isoflavones are compared. The influenceof the surface charge of the lipid bilayer and the spontaneous curvature of the membrane�forming phospho�lipids on the adsorption of flavonoids on the model membranes is discussed.
Keywords: lipid bilayers, membrane dipole potential, flavonoids
DOI: 10.1134/S0006350913030123

BIOPHYSICS Vol. 58 No. 3 2013
DIPOLE POTENTIAL OF PHOSPHOLIPID MEMBRANES WITH FLAVONOIDS 367
cin, nigericin, nonactin) [18, 19] and activity ofNa+/K+�ATPase and Ca2+�ATPase [20, 21]. Peptide–lipid interactions in the membrane, i.e. sorption ofpeptides on the membrane surface and initial stage oftheir anchoring [22, 23], and also the activity of somemembrane enzymes (for example, phospholipase A2)[24] are regulated by ϕd. The dipole potential influ�ences the interaction of a series of medicinal sub�stances with cell membranes, in particular the HIVprotease inhibitor saquinavir and HIV fusion inhibitorsifuvirtide, antimicrobial agents berberine and benza�lkonium, bioactive compounds nicotinamide andpicolinamide, antibiotic bacitracin, and also neuroac�tive steroid pregnanolone [25–30]. There exist dataabout that the dipole potential of the blood�brain bar�rier determines its permeability to various substances,among them those used in brain scanning [31]. Somemedicinal substances themselves are capable of chang�ing the dipole potential of the membrane; for example,anesthetics lidocaine and articaine increase while theamyloidogenic fragment of prionic protein PrP 185�206 decreases the dipole potential of the membrane[32⎯34]. According to the latest literature data, theeffect of flavonoids on the channel�forming activity ofsome toxins and antibiotics may be connected not onlywith a change in the magnitude of the membranedipole potential but also be a result of immediateinteraction of plant polyphenols with channel�form�ing molecules [35, 36].
The essential role of membrane dipole potential inthe regulation of membrane transport and the pros�pects of using flavonoids for decreasing the magnitudeof potential jump at the bilayer–surrounding solutioninterface have determined the main aim of the presentwork: establishment of the change in the magnitude ofdipole potential of model membranes formed of vari�ous phospholipids upon adsorption of flavonoids,phloridzin, quercetin, myricetin and biochanin A.The disclosed possibilities of gradual changing thedipole potential of the membrane in a broad range,and also varying the chemical structure of the dipole�modifying agent open new prospects for investigatingthe mechanisms of membrane activity of flavonoids,conditioned both by the decrease of dipole potentialand by immediate interaction of plant polyphenolswith membrane�incorporated ion�transporting sys�tems.
EXPERIMENTAL
In the work use was made of the following reagents:KCl, HEPES, pentane, ethanol, chloroform, hexade�cane, phloridzin, biochanin A, quercetin, myricetin,and nonactin (Sigma�Aldrich, USA); 1,2�dioleyl�sn�glycero�3�phosphocholine (DOPC), 1,2�dioleyl�sn�glycero�3�phosphoserine (DOPS) and 1,2�dioleyl�sn�glycero�3�phosphoethanolamine (DOPE) (AvantiPolar Lipids, USA).
Formation of bilayer lipid membranes was con�ducted by the method of Montal and Muller [37] bymeans of bringing together condensed lipid monolay�ers on a hole in Teflon film separating the experimen�tal chamber into two compartments. The volume ofeach compartment constituted 1.5 mL, Teflon filmthickness 10 μm, hole diameter about 50 μm. Beforethe beginning of the process of membrane formationthe hole in Teflon film was treated with hexadecane.Monolayers were formed at the water–air border fromlipid solution in pentane with a concentration of1 mg/mL. For formation of monolayers use was madeof DOPC, DOPS or DOPE. Experiments were con�ducted at the same ion composition of water solutionsof electrolyte (0.1 M KCl) in both chamber compart�ments, the acidity of solutions (pH 7.4) was main�tained by a buffer mixture 5 mM HEPES�KOH.
An alcoholic solution of ionophore nonactin(7 μg/mL) was added to the water phase of both com�partments to a final concentration of 10–7–10–6 M.Electrophysiological measurements were conductedat room temperature.
Flavonoids, phloridzin, biochanin A, quercetinand myricetin (millimolar solutions in ethanol) wereintroduced into both chamber compartments to a finalconcentration in near�membrane solutions in therange from 2.5 to 100 μM.
For supplying a transmembrane potential (V) andleading the signal from the membrane use was made ofsilver chloride electrodes (Ag/AgCl) connected withchamber solutions through bridges with 1.5% agarosein solution of 2 M KCl. Measurements and digitiza�
800
600
400
200
20151050
I, pA
t, min
80
7060
504030
2015
10
5.0
2.5
Fig. 1. Change in K+�nonactin�induced equilibriumtransmembrane current after addition of quercetin intomembrane�bathing solutions in the concentration range2.5–80 μM. Membrane formed of DOPE and bathed by0.1 M KCl pH 7.4. Moments of introduction of quercetininto membrane�bathing solutions are indicated by arrows.The corresponding concentrations of flavonoid are pre�sented above the arrows. V = 50 mV.

368
BIOPHYSICS Vol. 58 No. 3 2013
OSTROUMOVA et al.
tion of transmembrane currents were conducted in thepatch clamp mode with the aid of an operationalamplifier with a discrete range of feedback resistancesfrom 107 to 1010 Ohm. The amplifier output signal wasfed through an ADC to the computer input. Observa�tion was conducted with the aid of a special programARMEX. Processing of the records of transmembranecurrents was actualized with the use of a programpackage Clampfit 9.0 (Axon Instruments, USA). Sta�tistical analysis of the obtained data was executed withthe aid of a program Origin 8.0 (OriginLab, USA).
Membrane conductance G was determined as theratio of mean current I flowing through the bilayerlipid membrane to the transmembrane potential V(50 mV). The change in membrane dipole potential(Δϕd) after introduction into membrane�bathing solu�tions of flavonoids was determined by formula:
(1)
where G0 and G – values of equilibrium K+�conduc�tance of the bilayer conditioned by nonactin beforeand after introduction of flavonoid; qe – electron
GG0
����qeΔϕd
kT������������–⎝ ⎠
⎛ ⎞ ,exp=
charge modulus; k, T – have generally acceptedmeanings [16]. For example in Fig. 1 we present thetime characteristic of the dependence of transmem�brane current induced by K+�nonactin on the concen�tration of quercetin in the range from 2.5 to 80 μM insolution bathing the DOPE membrane. The stationarytransmembrane current increases with growing fla�vonoid concentration, reaching saturation at 60 μMquercetin.
The mean magnitudes of change in membranedipole potential were determined as mean values ofΔϕd in each series of measurements conducted on 3–5bilayers (mean ±SD).
For description of flavonoid adsorption on the sur�face of lipid bilayers use was made of Langmuir iso�therm:
(2)
where Δϕd(C) – change in membrane dipole potentialat fixed concentration (C) of flavonoid in the mem�brane�bathing solution, Δϕd(∞) – maximally possible
Δϕd C( )Δϕd ∞( )C
C K+�������������������,=
125
100
75
50
25
100500 7525
–Δϕd, mV
(c)125
100
75
50
25
100500 7525
(d)
C, μM
125
100
75
50
25
100500 7525
(a)125
100
75
50
25
100500 7525
(b)–Δϕd, mV
–Δϕd, mV –Δϕd, mV
C, μM
C, μMC, μM
Fig. 2. Dependences of change in membrane dipole potential (Δϕd) on the concentration of phloridzin (a), biochanin A (b),quercetin (c) and myricetin (d) in membrane�bathing solution. Membranes formed of DOPC (dark circles), DOPE (squares) orDOPS (triangles) and bathed by 0.1 M KCl pH 7.4. V = 50 mV.
BIOPHYSICS Vol. 58 No. 3 2013
DIPOLE POTENTIAL OF PHOSPHOLIPID MEMBRANES WITH FLAVONOIDS 369
change in membrane dipole potential upon introduc�tion of modifier, K – its dissociation constant [38].
RESULTS AND DISCUSSION
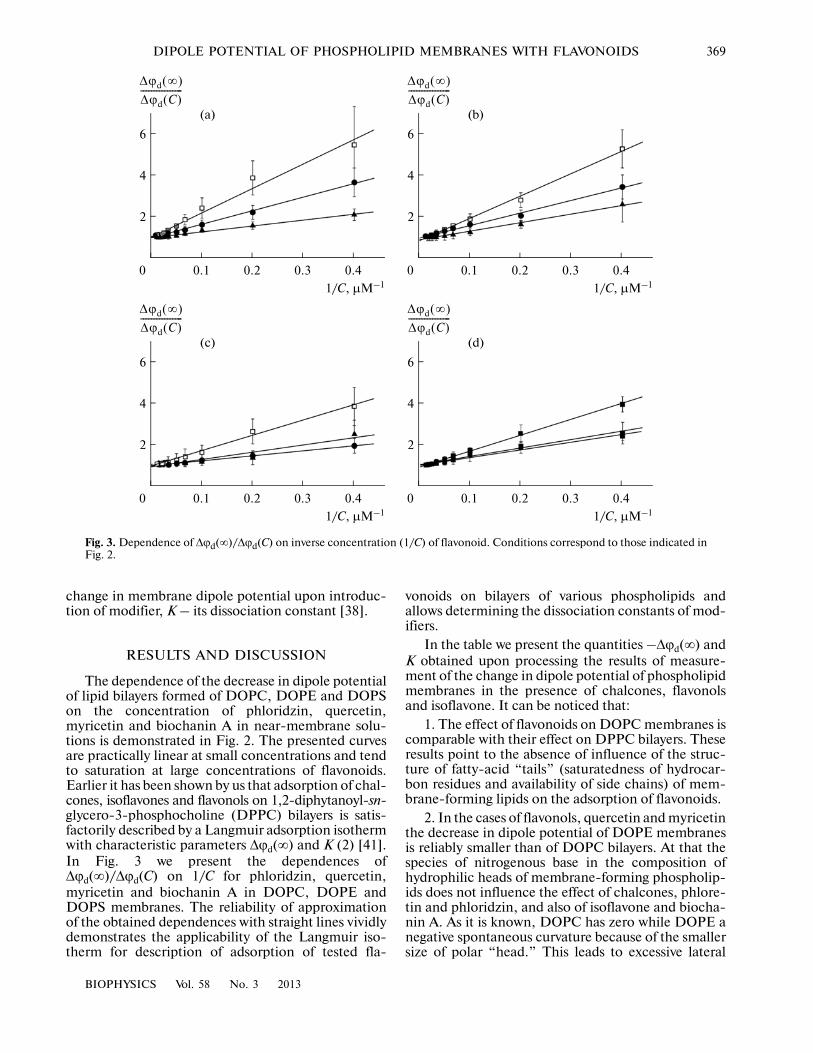
The dependence of the decrease in dipole potentialof lipid bilayers formed of DOPC, DOPE and DOPSon the concentration of phloridzin, quercetin,myricetin and biochanin A in near�membrane solu�tions is demonstrated in Fig. 2. The presented curvesare practically linear at small concentrations and tendto saturation at large concentrations of flavonoids.Earlier it has been shown by us that adsorption of chal�cones, isoflavones and flavonols on 1,2�diphytanoyl�sn�glycero�3�phosphocholine (DPPC) bilayers is satis�factorily described by a Langmuir adsorption isothermwith characteristic parameters Δϕd(∞) and K (2) [41].In Fig. 3 we present the dependences ofΔϕd(∞)/Δϕd(C) on 1/C for phloridzin, quercetin,myricetin and biochanin A in DOPC, DOPE andDOPS membranes. The reliability of approximationof the obtained dependences with straight lines vividlydemonstrates the applicability of the Langmuir iso�therm for description of adsorption of tested fla�
vonoids on bilayers of various phospholipids andallows determining the dissociation constants of mod�ifiers.
In the table we present the quantities –Δϕd(∞) andK obtained upon processing the results of measure�ment of the change in dipole potential of phospholipidmembranes in the presence of chalcones, flavonolsand isoflavone. It can be noticed that:
1. The effect of flavonoids on DOPC membranes iscomparable with their effect on DPPC bilayers. Theseresults point to the absence of influence of the struc�ture of fatty�acid “tails” (saturatedness of hydrocar�bon residues and availability of side chains) of mem�brane�forming lipids on the adsorption of flavonoids.
2. In the cases of flavonols, quercetin and myricetinthe decrease in dipole potential of DOPE membranesis reliably smaller than of DOPC bilayers. At that thespecies of nitrogenous base in the composition ofhydrophilic heads of membrane�forming phospholip�ids does not influence the effect of chalcones, phlore�tin and phloridzin, and also of isoflavone and biocha�nin A. As it is known, DOPC has zero while DOPE anegative spontaneous curvature because of the smallersize of polar “head.” This leads to excessive lateral
6
4
2
0.40.20 0.30.1
1/C, μM–1
Δϕd ∞( )
Δϕd C( )����������������
(c)
6
4
2
0.40.20 0.30.1
1/C, μM–1
Δϕd ∞( )
Δϕd C( )����������������
(d)
6
4
2
0.40.20 0.30.1
1/C, μM–1
Δϕd ∞( )
Δϕd C( )����������������
(a)
6
4
2
0.40.20 0.30.1
1/C, μM–1
Δϕd ∞( )
Δϕd C( )����������������
(b)
Fig. 3. Dependence of Δϕd(∞)/Δϕd(C) on inverse concentration (1/C) of flavonoid. Conditions correspond to those indicated inFig. 2.

370
BIOPHYSICS Vol. 58 No. 3 2013
OSTROUMOVA et al.
pressure in the region of hydrocarbon tails [42]. It maybe thought that the plane of adsorption of flavonoidsresides near the region of pressure jump, while otherflavonoids as dependent on hydrophilicity of theirmolecules have a greater or smaller degree of submer�sion into the bilayer.
3. The effects of flavonoids on the dipole potentialof negatively charged DOPS membranes are substan�tially less than for uncharged DOPC bilayers. An anal�ogous result was obtained upon studying the adsorp�tion of phloretin on neutral and negatively chargedmonolayers of dimyristoyl phosphatidyl choline anddimyristoyl phosphatidyl glycerol respectively [43]. Inall probability, electrostatic repulsion of negativelycharged serine residues leads to a decrease in the sur�face density of dipoles of membrane�forming lipids.The membrane dipole potential is tied with the surfacedensity by an equation of plane capacitor:
(3)ϕdμ⊥n
ε0ε�������,=
where μ⊥– mean projection of the dipole moment oflipid molecules (with account being taken of associ�ated water) onto the normal to the membrane plane;n – surface density of dipoles, ε – dielectric permittiv�ity of the membrane [44–46]. It should be expectedthat a decrease in the surface density of dipoles in con�sequence of electrostatic repulsion of lipid “heads”would lead to a decrease in the value of dipole poten�tial. According to [38] the Δϕd evoked by adsorption ofphloretin or its analogs may depend on the initialdipole potential of the membrane.
Nonetheless the negative surface charge of themembrane does not tell on the adsorption of phlo�ridzin. Similar results were obtained by Sokolov et al.[47] upon measurement by the method of intramem�brane field compensation of the change in potentialjump upon adsorption of phloridzin on negativelycharged membranes of azolectin and neutral bilayersof DOPC. Inasmuch as the water solubility of glyco�sides is significantly higher than of correspondingaglycons, it may be expected that among all studiedflavonoids the degree of bilayer submersion of phlo�
Characteristic parameters of Langmuir adsorption isotherm for flavonoids in phospholipid bilayers
Flavonoid Chemical structureDipole
moment#, DParameter
Membrane composition
DPPC DOPC DOPE DOPS
Phloretin
5.60 [16]
–Δϕd(∞), mV 147 ± 7* 140 ± 8* 128 ± 8* 90 ± 6*
K, μM 2.0 ± 0.5* 0.7 ± 0.2* 2.2 ± 0.4* 2.7 ± 0.8*
Phloridzin
–Δϕd(∞), mV 92 ± 4* 91 ± 10 88 ± 8 99 ± 12
K, μM 5.1 ± 0.2* 6.2 ± 1.1 13.0 ± 0.9 3.0 ± 0.6
Biochanin A 7.83 [39]
–Δϕd(∞), mV 92 ± 11* 112 ± 8 84 ± 9 54 ± 7
K, μM 2.1 ± 0.3* 6.4 ± 0.9 9.9 ± 1.1 3.2 ± 0.5
Quercetin 6.56 [40]
–Δϕd(∞), mV 104 ± 7* 100 ± 9 60 ± 11 54 ± 9
K, μM 3.3 ± 0.5* 2.5 ± 0.6 7.5 ± 1.7 2.3 ± 1.1
Myricetin 6.35 [40]
–Δϕd(∞), mV 111 ± 11* 89 ± 9 39 ± 4 36 ± 6
K, μM 3.3 ± 0.2* 4.8 ± 0.9 8.1 ± 0.7 3.7 ± 0.8
Note: # – dipole moment of flavonoid in water phase; * – results taken from [41].
O OH
HO OHHO
HO OH
O
OH
OO
OHOH
OH
HO
OHO
OH OOCH3
O
OH
HO
OH O
OH
OH
O
OH
HO
OH O
OH
OH
OH

BIOPHYSICS Vol. 58 No. 3 2013
DIPOLE POTENTIAL OF PHOSPHOLIPID MEMBRANES WITH FLAVONOIDS 371
ridzin is minimal [48]. The results of fluorescent mea�surements of the values of dipole potential of liposo�mal membranes formed with participation of neutraland negatively charged phospholipids testifies to thatthe negative charge of serine resides at a sufficientremoval from the membrane�surrounding water phase[46, 49]. In all probability, it does on influence theinteraction of glycoside with the membrane surface.
4. The dissociation constant of all indicated fla�vonoids with the exception of phloretin is substantiallygreater in the case of DOPE membranes as comparedwith bilayers formed of other phospholipids. Inas�much as 1/K characterizes the affinity of modifier forlipid phase, the obtained results testify to the smallestmagnitudes of the coefficients of partition of fla�vonoids between the DOPE membrane and the bath�ing solution. At that the closeness of the values ofphloretin dissociation constants for DPPC, DOPC,DOPE and DOPS membranes allows asserting thatthe coefficient of partition of phloretin practicallydoes not depend on the phospholipid composition ofthe membrane. According to [50], the coefficient ofpartition of phloretin and its analogs between themembrane and water solution is under significantinfluence of dipole–dipole interactions at the phaseboundary. It may be supposed that the partition coeffi�cient depends not only on the type of membrane�forming lipid but also on the dipole moment, plane ofadsorption, and also the possible orientation ofbilayer�sorbing molecules of the dipole modifier. Thedipole moments of the tested flavonoids are close (seetable). However the unique cap of phloretin to assumea “hairpin”�like conformation [48] may facilitate itsincorporation into DOPE membranes. In all probabil�ity, submersion of more rigid rod�like molecules ofother aglycons into the zone of excessive lateral pres�sure emerging in DOPE bilayers is hindered. Thelarger size of glycoside as compared with the corre�sponding aglycon may also prevent phloridzin interac�tion with the membrane.
ACKNOWLEDGMENTS
The authors express their gratitude to V.V. Malevfor fruitful discussion of results.
The work was supported by the Russian Founda�tion for Basic Research (12�04�31332), presidentialprogram (MK�1813.2012.4), RAS Presidium program“Molecular and Cell Biology” and State contract 8119(MSE RF, FTP “SSPCIR”).
REFERENCES
1. E. A. Liberman and V. P. Topaly, Biofizika 14, 452(1969).
2. S. B. Hladky and D. A. Haydon, Biochim. Biophys.Acta 318, 464 (1973).
3. H. Brockmann, Chem. Phys. Lipids 73, 57 (1994).
4. D. T. Warshaviak, M. J. Muellner, and M. Chachisvilis,Biochim. Biophys. Acta 1808, 2608 (2011).
5. R. Latorre and J. J. Donovan, Acta Physiol. Scand.Suppl. 481, 37 (1980).
6. D. D. Busath, C. D. Thulin, R. W. Hendershot, et al.,Biophys. J. 75 (6), 2830 (1998).
7. T. C. Hwang, R. E. Koeppe, and O. S. Andersen, Bio�chemistry 42 (46), 13646 (2003).
8. R. L. Duffin, M. P. Garrett, K. B. Flake, et al., Lang�muir 19, 1439 (2003).
9. T. Luchian and L. Mereuta, Langmuir 22, 8452 (2006).10. O. S. Ostroumova, L. V. Schagina, and V. V. Malev,
Membr. Cell Biol. (Moscow) 2 (3), 259 (2008).11. L. Mereuta, T. Luchian, Y. Park, et al., Biochem. Bio�
phys. Res. Commun. 373, 467 (2008).12. O. S. Ostroumova and L. V. Schagina, Membr. Cell
Biol. (Moscow) 3 (3), 281 (2009).13. O. S. Ostroumova, V. V. Malev, M. G. Ilin, et al., Lang�
muir 26 (19), 15092 (2010).14. A. Apetrei, L. Mereuta, and T. Luchian, Biochim. Bio�
phys. Acta 1790, 809 (2009).15. O. S. Ostroumova, S. S. Efimova, and L. V. Schagina,
PLoS One 7 (1), e30261 (2012).16. O. S. Andersen, A. Finkelstein, I. Katz, et al., J. Gen.
Physiol. 67, 749 (1976).17. E. Melnik, R. Latorre, J. E. Hall, et al., J. Gen. Physiol.
69 (2), 243 (1977).18. S. B. Hladky, Biochim. Biophys. Acta 352 (1), 71
(1974).19. S. Bala, M. H. Kombrabail, and B. S. Prabhananda,
Biochim. Biophys. Acta 1510, 258 (2001).20. D. Y. Malkov and V. S. Sokolov, Biochim. Biophys.
Acta 1278 (2), 197 (1996).21. J. Karlovska, D. Uhrikova, N. Kucerka, et al., Biophys.
Chem. 119 (1), 69 (2006).22. Y. Yano and K. Matsuzaki, Biochemistry 41, 12407
(2002).23. V. Buzon and J. Cladera, Biochemistry 45, 15768
(2006).24. B. Maggio, J. Lipid Res. 40 (5), 930 (1999).25. T. Asawakarn, J. Cladera, and P. O’Shea, J. Biol. Chem.
276, 38457 (2001).26. I. I. Severina, M. S. Muntyan, K. Lewis, et al., IUBMB
Life 52 (6), 321 (2001).27. J. Cladera, P. O’Shea, J. Hadgraft, et al., J. Pharm. Sci.
92 (5), 1018 (2003).28. M. Alakoskela, T. Soderlund, J. M. Holopainen, et al.,
Mol. Pharmacol. 66, 161 (2004).29. A. Borba, F. Lairion, A. Disalvo, et al., Biochim. Bio�
phys. Acta 1788 (12), 2553 (2009).30. P. M. Matos, T. Freitas, M. A. Castanho, et al., Bio�
chem. Biophys. Res. Commun. 403 (3–4), 270 (2010).31. J. Cattelotte, N. Tournier, N. Rizzo�Padain, et al.,
J. Neurochem. 108 (3), 767 (2009).32. C. J. Hogberg and A. P. Lyubartsev, Biophys. J. 94 (2),
525 (2008).33. E. H. Mojumdar and A. P. Lyubartsev, Biophys. Chem.
153 (1), 27 (2010).

372
BIOPHYSICS Vol. 58 No. 3 2013
OSTROUMOVA et al.
34. S. Sonkina, I. I. Tukhfatullina, N. Benseny�Cases,et al., J. Pept. Sci. 16 (7), 342 (2010).
35. O. S. Ostroumova, S. S. Efimova, and L. V. Schagina,Biochim. Biophys. Acta Biomembr. 1808 (8), 2051(2011).
36. O. S. Ostroumova, S. S. Efimova, E. G. Chulkov, et al.,PLoS One. 7 (9), e45135 (2012).
37. M. Montal and P. Muller, Proc. Nat. Acad. Sci. USA 69(12), 3561 (1972).
38. R. Cseh, M. Hetzer, K. Wolf, et al., Eur. Biophys. J. 29(3), 172 (2000).
39. E. A. Armelin, P. M. Donate, and S. E. Galembeck,Tetrahedron. 56, 5105 (2000).
40. G. C. Justino and A. J. S. C. Vieira, J. Mol. Model. 16,863 (2010).
41. S. S. Efimova and O. S. Ostroumova, Langmuir 28(26), 9908 (2012).
42. S. M. Bezrukov, Current Opinion in Colloid InterfaceSci. 5, 237 (2000).
43. F. Lairion and E. A. Disalvo, Langmuir 20, 9151(2004).
44. R. F. Flewelling and W. L. Hubbell, Biophys. J. 49, 541(1986).
45. S. A. Simon, T. J. McIntosh, A. D. Magid, et al., Bio�phys. J. 61 (3), 786 (1992).
46. T. Starke�Peterkovic and R. J. Clarke, Eur. Biophys. J.39, 103 (2009).
47. V. S. Sokolov, V. V. Cherny, and V. S. Markin, Biofizika29 (3), 424 (1984).
48. Y. S. Tarahovsky, E. N. Muzafarov, and Y. A. Kim, Mol.Cell Biochem. 314, 65 (2008).
49. E. Gross, R. S. Bedlack, and L. M. Loew, Biophys. J.67, 208 (1994).
50. R. Awiszus and G. Stark, Eur. Biophys. J. 15, 321(1988).





























