Embed Size (px)
Citation preview

1
Revised to: Appl. Environ. Microbiol. 1 MS No. AEM06937-11 2
3 Characterization and Functional Analyses of 4
(R)-Specific Enoyl-CoA Hydratases in Polyhydroxyalkanoate-5 producing Ralstonia eutropha 6
7 8
Yui Kawashima1†, Wen Cheng1,2†, Jun Mifune1, Izumi Orita1, Satoshi Nakamura1, 9 and Toshiaki Fukui1* 10
11 1Department of Bioengineering, Graduate School of Bioscience and Biotechnology, Tokyo 12 Institute of Technology, 4259 Nagatsuta, Midori-ku, Yokohama 226-8501, Japan, 2Department 13 of Chemical Engineering, Tsinghua University, Beijing 100084, China. 14 15 †The first two authors contributed equally to the manuscript. 16 17 18 *Corresponding author 19 Mailing address: [email protected] 20 Department of Bioengineering, 21 Graduate School of Bioscience and Biotechnology, 22 Tokyo Institute of Technology 23 B-37 4259 Nagatsuta, Midori-ku, Yokohama 226-8501, Japan 24 Tel/Fax: +81-45-924-5766 25 26 Running title: (R)-Enoyl-CoA hydratases from Ralstonia eutropha 27 28 Key words: bioplastic, polyhydroxyalkanoate synthesis, (R)-specific enoyl-CoA hydratase, 29 β-oxidation, Ralstonia eutropha, Cupriavidus necator, metabolic engineering 30 31 32 33
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.06937-11 AEM Accepts, published online ahead of print on 11 November 2011
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 34 35
A genome survey of polyhydroxyalkanoate (PHA)-producing Ralstonia eutropha H16 36 detected the presence of 16 orthologs of (R)-specific enoyl-CoA hydratase, among which 37 three proteins shared high homologies with the enzyme specific to enoyl-CoAs of medium-38 chain-length encoded by phaJ4Pa from Pseudomonas aeruginosa. The recombinant forms of 39 the three proteins, termed PhaJ4a–cRe, actually showed enoyl-CoA hydratase activity with 40 (R)-specificity, and the catalytic efficiencies were elevated as the substrate chain length 41 increased from C4 to C8. PhaJ4aRe and PhaJ4bRe showed more than 10-fold higher catalytic 42 efficiency than PhaJ4cRe. The functions of the new PhaJ4 proteins were investigated using 43 previously engineered R. eutropha strains as host strains; these strains are capable of 44 synthesizing poly((R)-3-hydroxybutyrate-co-(R)-3-hydroxyhexanoate) [P(3HB-co-3HHx)] 45 from soybean oil. Deletion of phaJ4aRe from the chromosome resulted in significant decrease 46 of 3HHx composition in the accumulated copolyester, whereas no change was observed by 47 deletion of phaJ4bRe or phaJ4cRe, indicating that only PhaJ4aRe was one of the major enzymes 48 supplying the (R)-3HHx-CoA monomer through β-oxidation. Introduction of phaJ4aRe or 49 phaJ4bRe into the R. eutropha strains using a broad-host-range vector enhanced the 3HHx 50 composition of the copolyesters, but introduction of phaJ4cRe did not. The two genes were 51 then inserted into the pha operon on chromosome 1 of the engineered R. eutropha by 52 homologous recombination. This chromosomal modification enabled the biosynthesis of 53 P(3HB-co-3HHx) composed of a larger 3HHx fraction without a negative impact on cell 54 growth and PHA production on soybean oil, especially when phaJ4aRe or phaJ4bRe was 55 tandemly introduced with phaJAc from Aeromonas caviae. 56
57
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
INTRODUCTION 58 (R)-Specific 2-enoyl-CoA hydratase (R-ECH) is an enzyme that catalyzes the addition of 59
a water molecule across the double bond in enoyl-CoA with (R)-stereospecificity, which 60 results in the generation of the corresponding (R)-3-hydroxyacyl (3HA)-CoA. In mammals, 61 plants, and yeasts, R-ECH occurs as a domain of peroxisomal multifunctional enzyme type 2 62 and has a role in the hydration of 2-enoyl-CoA intermediates in fatty acid β-oxidation (15, 18, 63 22, 30). Monofunctional R-ECH was first identified in the Gram-negative 64 polyhydroxyalkanoate (PHA)-producing bacterium Aeromonas caviae (7, 8). The gene of R-65 ECH in A. caviae (phaJAc) is clustered with genes of PHA-granule associated protein (phaPAc) 66 and PHA synthase (phaCAc) in the order of phaP-C-J. It has been clarified that PhaJAc plays an 67 essential role in providing (R)-3HA-CoA monomers of C4–C6-chain length through β-68 oxidation for co-polymerization mediated by PhaCAc (8). R-ECHs have been further identified 69 and characterized from several PHA producers: Aeromonas hydrophila (14, 20, 26), 70 Pseudomonas aeruginosa (39, 42), P. oleovorans (6), P. putida (6, 32, 45), and 71 Rhodospirillum rubrum (31). In P. aeruginosa, four kinds of R-ECHs, PhaJ1–4Pa, were 72 characterized. PhaJ1Pa was specific to short-chain-length (scl)-2-enoyl-CoAs (C4–C6) as well 73 as PhaJAc, while PhaJ4Pa was R-ECH specific to medium-chain-length (mcl)-substrates longer 74 than C6 (42). Recent whole genome analyses detected the presence of numerous PhaJ 75 orthologs in diverse organisms regardless of the ability to synthesize PHAs. For instance, a 76 previous report demonstrated that a MaoC-like protein in a non-PHA-producing Escherichia 77 coli, of which the amino acid sequence showed a 34% identity to that of PhaJAc, was involved 78 as R-ECH in the formation of mcl-PHAs from dodecanoate in recombinant strains (27). Thus, 79 it is expected that novel R-ECHs with unique catalytic properties distinct from known 80 enzymes will be included in the homologs detected in silico. The first crystal structure of R-81 ECH has been reported for the dimeric enzyme from A. caviae with a resolution of 1.5 Å (13). 82
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
The structure of the monomer consists of a five-stranded antiparallel β-sheet and a central α-83 helix into a so-called “hot dog” fold. The active residues were confirmed to be Asp31 and 84 His36 by site-directed mutagenesis. The residues Leu65 and Val130 are situated at the bottom 85 of the substrate-binding tunnel, and define the preference of the enzyme for the substrate 86 chain length (40). 87
Bacterial polyhydroxyalkanoates (PHAs) are biodegradable thermoplastics produced 88 from renewable biomass that have drawn increased attention as possible alternatives for 89 petroleum-based polymers (21, 28, 37). Poly((R)-3-hydroxybutyrate-co-(R)-3-90 hydroxyhexanoate) [P(3HB-co-3HHx)] is one of the most practical PHAs, as it exhibits a 91 lower melting point and higher flexibility than poly((R)-3-hydroxybutyrate) [P(3HB)], the 92 most abundant PHA in nature. The above-mentioned bacterium A. caviae was originally 93 isolated as a natural producer of P(3HB-co-3HHx) from alkanoic acids of even carbon 94 numbers and olive oil, but the copolyester productivity by A. caviae was not high (~12 wt% 95 of dry cell weight on olive oil) (5, 34). Aiming for efficient microbial production of P(3HB-96 co-3HHx), several studies focused on the engineering of a well-studied P(3HB)-producing 97 bacterium Ralstonia eutropha (Cupriavidus necator). It has been demonstrated that R. 98 eutropha strain PHB–4 expressing phaC from A. caviae is capable of accumulating P(3HB-co-99 3HHx) efficiently from vegetable oils (8, 16). In these recombinant strains grown on 100 vegetable oils, (R)-3HB-CoA is mainly formed from two acetyl-CoA molecules through two-101 step reactions mediated by β-ketothiolases (PhaARe, BktBRe) and stereospecific NADPH-102 acetoacetyl-CoA reductase (PhaB1Re) (2, 24), while (R)-3HHx-CoA and partial amounts of 103 (R)-3HB-CoA are supposed to be formed from C6- and C4-intermediates in β-oxidation, 104 respectively. However, unlike A. caviae (10) and P. aeruginosa (4), the details of the linkage 105 between β-oxidation and PHA biosynthesis in R. eutropha have not been elucidated till date. 106 Recent DNA microarray analysis of R. eutropha grown on palm oil identified two loci 107
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
important for β-oxidation (1), but no information was reported regarding the metabolic 108 linkage. On the other hand, it has been reported that phaJ genes are useful tools for enhancing 109 3HHx composition in the copolymer synthesized by R. eutropha from vegetable oils. Mifune 110 et al. inserted phaCNSDG (a mutant gene of phaCAc (43)) and phaJAc into and deleted phaARe 111 from the pha operon on chromosome 1 of R. eutropha H16, and demonstrated that the 112 engineered strain was able to produce 79 wt% of P(3HB-co-3HHx) with a 9.9 mol% 3HHx 113 fraction from soybean oil (24). Budde et al. reported that the R. eutropha strain lacking 114 acetoacetyl-CoA reductase genes and expressing phaC2 from Rhodococcus aetherivorans and 115 phaJ1 from P. aeruginosa could accumulate 56-60 wt% of P(3HB-co-3HHx) with 24-25 116 mol% 3HHx from palm oil (3). Several phaJ genes have been also applied in attempts to 117 establish a biosynthesis pathway for PHA copolymers through β-oxidation in E. coli (6, 11, 118 20, 26, 31, 39, 40, 42). 119
The genome sequence of R. eutropha H16 (29) revealed the presence of multiple 120 homologs of genes involved in PHA metabolism, such as 15 homologs of β-ketothiolase, 3 121 homologs of NADPH-acetoacetyl-CoA reductase, and 7 homologs of PHA depolymerase, 122 suggesting complicated and robust metabolism for synthesis and degradation of PHA. Recent 123 studies revealed major contributions of three β-ketothiolases (PhaA, BktB, and H16_A0170) 124 to the dimerization of acetyl-CoA (19), and two acetoacetyl-CoA reductases (PhaB1 and 125 PhaB3) for the reduction of acetoacetyl-CoA (2) during P(3HB) biosynthesis on fructose. On 126 vegetable oil substrates, PhaB1 not only acted as a major reductase for the generation of (R)-127 3HB-CoA (2, 24), but also participated to some degree in the formation of (R)-3HHx-CoA 128 through β-oxidation (24). 129
This study focused on R-ECHs in R. eutropha, especially on the three enzymes highly 130 homologous to PhaJ4Pa which is R-ECH specific to mcl-substrates. The catalytic properties 131 were determined by using the recombinant proteins, and their roles in PHA biosynthesis 132
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
through β-oxidation were examined by gene disruption. Furthermore, the genes were applied 133 in metabolic engineering for biosynthesis of PHA copolymers with altered molar composition 134 from soybean oil. The information obtained here would be useful for the engineering of R. 135 eutropha as a platform for the efficient production of superior PHA copolymers. 136 137 138
MATREIALS AND METHODS 139 Bacterial strains and plasmids. 140
Bacterial strains and plasmids used in this study are listed in Table 1. R. eutropha strains 141 were cultivated at 30 °C in a nutrient-rich (NR) medium containing 10 g of meat extract, 10 g 142 of polypeptone and 2 g of yeast extract in 1 L of tap water. E. coli strains were grown at 37 °C 143 on a Luria-Bertani (LB) medium. Kanamycin (50 mg/L for E. coli and 300 mg/L for R. 144 eutropha strains) or ampicillin (50 mg/L for E. coli) was added to the medium when 145 necessary. 146 147 Construction of expression plasmids and strains. 148
DNA manipulations were carried out according to standard procedures, and sequences of 149 oligonucleotide primers used for PCR amplification are shown in Table 2. Recombinant form 150 of R-ECHs derived from R. eutropha with an N-terminal His6-tag was individually produced 151 by E. coli BL21(DE3) harboring a pColdII-based expression plasmid. Genomic DNA of R. 152 eutropha H16 was isolated and used as a template for PCR amplification of phaJ4aRe, 153 phaJ4bRe, and phaJ4cRe with a primer set of A1070-F/R, B0397-F/R, and B0667-F/R, 154 respectively. The PCR products were subcloned into pUC118 and the absence of unintended 155 mutations in the insert was confirmed by DNA sequencing. The fragments of phaJ4aRe, 156 phaJ4bRe, and phaJ4cRe were excised by digestion with NdeI and XhoI, and then ligated with 157
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
pColdII (cspA promoter, Ampr) (Takara Bio, Otsu, Shiga, Japan) at the corresponding sites to 158 obtain pColdII-J4a, pColdII-J4b, and pColdII-J4c, respectively. For expression of the genes in 159 R. eutropha, the NdeI-XhoI restricted fragments of phaJ4aRe, phaJ4bRe, and phaJ4cRe were 160 inserted at NdeI and SalI sites of pBPP (phaP1Re promoter) (9), resulting in the construction 161 of pBPP-J4a, pBPP-J4b, and pBPP-J4c, respectively. 162
A plasmid pK18ms∆J4a for deletion of phaJ4aRe from chromosome 1 of R. eutropha was 163 constructed as described below. First, upstream and downstream regions of phaJ4aRe were 164 individually amplified by PCR with genomic DNA of R. eutropha H16 as a template and 165 primer sets of A1070up-F/R and A1070down-F/R, respectively. Fusion PCR using the 166 amplified regions with A1070up-F/A1070down-R primers gave a fragment not including the 167 phaJ4aRe region. The resulting fragment was digested by XbaI and HindIII, and then ligated 168 with pK18mobsacB (33) at the corresponding sites to obtain pK18ms-∆J4a. pK18ms∆J4b and 169 pK18ms∆J4c for deletion of phaJ4bRe and phaJ4cRe were constructed by the same way using 170 primer sets of B0397up-F/R and B0397down-F/R, and B0667up-F/R and B0667down-F/R, 171 respectively. These vectors were applied to construct deletants of each phaJ4Re gene in R. 172 eutropha strains H16CAc (25) and NSDG∆AB (24). 173
For insertion of phaJ4aRe or phaJ4bRe between phaCNSDG and phaB1Re in R. eutropha 174 chromosome, the genes of phaJ4aRe and phaJ4bRe were individually amplified from genomic 175 DNA of R. eutropha H16 with primers A1070Ins-F/R and B0397Ins-F/R, respectively. The 176 primers A1070Ins-F and B0397Ins-F were designed to have ribosome-binding sequence for 177 efficient expression of the gene. The fragments were digested with XbaI and SpeI, and then 178 ligated with pK18msNSDG-B, previously constructed to make strain NSDG∆A (24), at the 179 corresponding sites to construct pK18msNSDG-J4aB and pK18msNSDG-J4bB. These 180 plasmids were used to make TT101 and TT102 with strain NSDG∆A as a parent strain. The 181 XbaI-SpeI fragments of phaJ4aRe and phaJ4bRe were also inserted into pK18msNSDG-JB 182
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
(24) at the corresponding sites to obtain pK18msNSDG-JJ4aB and pK18msNSDG-JJ4bB, 183 which were used to transform the strain MF03 (24) for construction of strains TT103 and 184 TT104 harboring a tandem of phaJAc-phaJ4aRe and phaJAc-phaJ4bRe, respectively, between 185 phaCNSDG and phaB1Re on the chromosome. 186
Transconjugation of the mobilizable plasmids from E. coli S17-1 (35) to R. eutropha, and 187 isolation of strains generated by pop in-pop out recombination using pK18mobsacB-based 188 suicide plasmids were performed as described previously (24, 25). 189 190 Expression of phaJ4a–cRe in E. coli and purification of the His6-tagged recombinant 191 enzymes. 192
The expression plasmid pColdII-J4a, pColdII-J4b and pColdII-J4c were used for 193 transformation of E. coli BL21(DE3). A 3 ml overnight culture of the cells was inoculated 194 into 100 ml of LB medium containing 50 mg of ampicillin per liter and cultivated in a 500 ml 195 flask on a reciprocal shaker (115 strokes per min) at 37 °C. After the OD600 reached to 0.4, the 196 culture medium was cooled down to 15 °C, and isopropyl-β-D-thiogalactopyranoside was 197 added to a final concentration of 0.5 mM. The cultivation was continued for additional 24 h at 198 15 °C. The grown cells were harvested, sonicated in buffer A (20 mM NaPB, 500 mM NaCl, 199 30 mM imidazole, pH7.4), and then subjected to centrifugation (15,000 rpm, 10 min, 4 °C). 200 The obtained soluble protein faction was filtered by 0.22 mm filter and loaded onto a HisTrap 201 FF crude 1ml column (GE Healthcare UK Ltd�, Buckinghamshire, England) equilibrated 202 with buffer A and was eluted with a linear gradient (20 ml) of 30 mM to 500 mM imidazole (1 203 ml/min). The fractions exhibiting 2-enoyl-CoA hydratase activity were combined, and then 204 dialyzed in 50 mM Tris-HCl (pH 7.5) at 0 °C for 24h. The purified enzyme was analyzed by 205 sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) with a 10-15% 206 gradient gel. 207
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
208 209 Enzyme assay. 210
Crotonyl-CoA was purchased from Sigma (St. Louis, MO), and trans-2-enoyl-CoA 211 substrates of C6 and C8 were synthesized and purified as described previously (24). The purity 212 was checked by HPLC equipped with a COSMOSIL 5C18-AR-II column (Nacalai tesque, 213 Kyoto, Japan). The CoA-thioesters were eluted by a linear gradient of acetonitrile from 1% to 214 60% (v/v) in 50 mM potassium phosphate buffer (pH 7.0) for 30 min. 215
Overall 2-enoyl-CoA hydratase activity was assayed by hydration of trans-2-enoyl-CoA 216 substrates (10, 24). A 25 μl of enzyme solution with appropriate dilution was added to 375 μl 217 mixture of 100 mM Tris-HCl (pH 7.8) containing 25 μM trans-2-enoyl-CoA in a quartz 218 cuvette, and the decrease in absorbance at 263 nm was measured at 30 °C. The ε263 of the 219 enoyl-thioester bond was 6.7 ×103 M-1 cm-1. The kinetic analyses were performed by 220 measuring the activity with various concentrations of the substrates. 221
The stereospecificity of enoyl-CoA hydratase was evaluated by monitoring hydration of 222 crotonyl-CoA coupled with (R)-specific polymerization of the resulting 3HB-CoA catalyzed 223 by crude PHA synthase (PhaCRe) from R. eutropha H16 (10). The crude PHA synthase was 224 prepared from R. europha H16 grown on fructose for 36 h at 30 °C, and was isolated as a 225 form of active enzyme associated with insoluble PHA granules. In detail, the cells in 100 ml 226 culture broth were harvested, washed once with buffer 1 (50 mM Tris-HCl, pH7.5) and then 227 subjected to sonication in 3 ml buffer 1. The sonicated cells were centrifuged with a low 228 speed (1,500 ×g, 5 min) to remove large debris and undisrupted cells, and the opaque 229 supernatant was further centrifuged with a high speed (15,000 ×g, 15 min) at 4 °C. The pellet 230 fraction of P(3HB) granules containing the crude PHA synthase was washed once with buffer 231 1 and resuspended in 3 ml buffer 1 for further use. The reaction mixture (400 μl) was 232
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
composed of 0.2 mM crotonyl-CoA, 1 mM 5,5’-dithio-bis(2-nitrobenzoic acid), and enoyl-233 CoA hydratase and the crude PHA synthase with appropriate dilutions in 50 mM pottasium 234 phosphate buffer. The reaction was monitored by increase in absorbance at 412 nm derived 235 from the release of free CoA-SH linked with the polymerization of (R)-3HB-CoA to P(3HB) 236 by PHA synthase. 237
238 PHA production by recombinant R. eutropha strains harboring phaJ4a–cRe. 239
PHA production by R. eutropha strains was carried out on a reciprocal shaker (115 240 strokes/min) at 30 °C for 72 h in 500 mL flasks with 100 mL of nitrogen-limited mineral salts 241 (MB) medium, which was composed of 0.9 g of Na2HPO412H2O, 0.15 g of KH2PO4, 0.05g 242 of NH4Cl, 0.02 g of MgSO47 H2O, and 0.1 ml of trace-element solution (17) in 100 ml of 243 distilled water. A filter-sterilized solution of fructose was added to the medium at a final 244 concentration of 0.5% (w/v) as a sole carbon source. Soybean oil was directly added to the 245 medium at 1.0% (v/v). Kanamycin was added at 300 mg/L, when necessary. After the 246 cultivation for 72 h, cells were harvested, washed once with distilled water, and then 247 lyophilized. Cellular PHA content and composition were determined by gas chromatography 248 (GC) after direct methanolysis of the dried cells in the presence of 15% sulfuric acid as 249 described previously (7). 250
251 252
RESULTS 253 Bioinformatic analysis of PhaJ-orthologous proteins in R. eutropha H16 genome 254
A BLAST homology search using the amino acid sequences of PhaJAc and PhaJ1–4Pa as 255 query sequences resulted in the detection of 16 genes encoded in R. eutropha H16 genome 256 (29) that showed homologies to the PhaJ proteins. Thirteen sequences among the 16 subjects 257
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
were composed of one MaoC-like domain (pfam01575) having lengths of 136-169 amino 258 acids such as PhaJAc, PhaJ1Pa, and PhaJ4Pa, while H16_A2250 contained two MaoC-like 259 domains in tandem. H16_A2151 contained an additional sequence of approximately 150 260 amino acids at the N-terminus, which is similar to the sequences of PhaJ2Pa and PhaJ3Pa. 261 H16_B0354 has another additional sequence of approximately 200 amino acids at the C-262 terminus. A phylogenetic tree of the sequences of the MaoC-like domains, constructed by 263 ClustalW multiple sequence alignment software, is shown in Fig. 1. In order to achieve the 264 biosynthesis of P(3HB-co-3HHx) with a larger 3HHx fraction, it is presumed that the use of 265 R-ECH with higher catalytic efficiency toward the C6 substrate than the C4 substrate would be 266 favorable for the generation of more (R)-3HHx-CoA monomers through β-oxidation cycles. 267 We therefore focused on three homologs clustered into the same branch with phaJ4Pa 268 encoding R-ECH specific to mcl-substrates, which are H16_A1070 (158 aa), H16_B0397 269 (151 aa), and H16_B0667 (151 aa) referred to as PhaJ4aRe, PhaJ4bRe and PhaJ4cRe, sharing 270 44%, 43%, and 52% identities to PhaJ4Pa, respectively. 271
The gene organizations around the three phaJ4 homologs are illustrated in Fig. 2. 272 Interestingly, all of them are clustered with acyl-CoA dehydrogenase genes. phaJ4aRe is a 273 neighbor of another phaJ homolog, H16_A1070, and clustered with two acyl-CoA 274 dehydrogenase genes in tandem (H16_A1067-1068). phaJ4bRe is located downstream of a 275 tandem of acyl-CoA dehydrogenase genes (H16_B0395-B0396). phaJ4cRe is within a large 276 gene cluster composed of multiple genes of acyl-CoA dehydrogenases (H16_B0660-B0661, 277 H16_B0664-B0665), thiolases (H16_B0662, H16_B0668), and short-chain dehydrogenases 278 (H16_B0663, H16_B0666). This gene organization suggests the probable participation of 279 phaJ4a-cRe in the metabolisms of acyl-CoA molecules. 280 281 Catalytic properties of the recombinant PhaJ4a–cRe 282
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
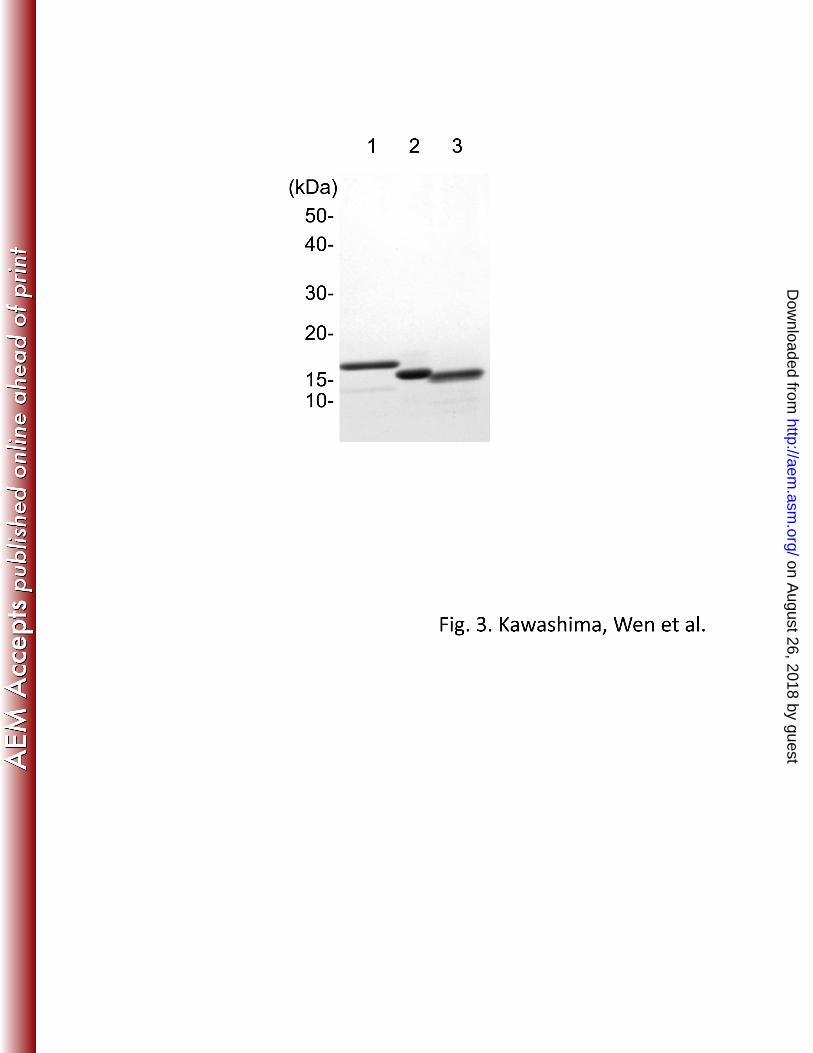
The genes of the three PhaJ4 homologs, phaJ4a–cRe, were individually overexpressed in 283 E. coli BL21(DE3), and the His6-tagged recombinant enzymes were purified to homogeneity 284 by Ni-affinity chromatography (Fig. 3). The subunit molecular weights of the recombinant 285 PhaJ4aRe, PhaJ4bRe and PhaJ4cRe were estimated to be 17.5, 17.0, and 16.5 kDa by SDS-286 PAGE, which were in agreement with those deduced from amino acid sequences (17.6, 16.7, 287 16.6 kDa), respectively. 288
The kinetic analysis of the recombinant enzymes was performed on trans-2-enoyl-CoAs 289 of chain length from C4 to C8 (Table 3). All the PhaJ4 homologs from R. eutropha followed 290 Michaelis-Menten kinetics for hydration of the substrates examined as well as PhaJAc and 291 PhaJ4Pa (10, 40, 42). The Km values decreased as the substrate chain length increased, and 292 showed kcat values one order of magnitude larger toward the C6 and C8 substrates than toward 293 the shorter C4 substrate. This information indicated that all the PhaJ4 homologs were specific 294 to mcl-substrates as expected, but they could accept the C4 substrate, unlike PhaJ4Pa. Among 295 the three PhaJ4s from R. eutropha, PhaJ4bRe showed the highest catalytic efficiency and 296 specificity toward the C6–C8 substrates (kcat/Km = 1.4–3.8 ×108 and C6–8/C4 ratios of kcat/Km = 297 67–1.8 ×102, respectively), and PhaJ4aRe had the highest affinity toward 2-hexenoyl-CoA (Km 298 = 41 μM) which was comparable with that of PhaJAc (Km = 40 μM) (40). The catalytic 299 efficiencies of PhaJ4cRe were much lower than those of the other two PhaJ4s because of the 300 smaller kcat values especially towards the short substrates (C4–C6). 301
The stereospecificity of the three PhaJ4s was evaluated by hydration of crotonyl-CoA 302 coupled with the (R)-specific polymerization of 3HB-CoA catalyzed by PHA synthase from 303 R. eutropha H16 (PhaCRe). As shown in Figure 4, obvious increases of absorbance were 304 observed at 412 nm when PhaCRe and DTNB were present in the reaction mixture in all the 305 cases of PhaJ4a–cRe. Apparently, the three enzymes produced 3HB-CoA with an (R)-306
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
configuration, similar to other known R-ECHs such as PhaJAc (10), PhaJ1Pa, PhaJ2Pa, PhaJ4Pa 307 (39, 42), and PhaJRr from R. rubrum (31). 308 309 Effects of disruption of phaJ4a–cRe on PHA biosynthesis by R. eutropha 310
The chromosomal phaJ4a–cRe genes were individually disrupted in R. eutropha to clarify 311 the participation of the respective gene in PHA biosynthesis through β-oxidation. We initially 312 disrupted each gene in strain H16CAc that had been constructed by replacing the original 313 phaCRe with phaCAc for P(3HB-co-3HHx) biosynthesis from vegetable oils (25). Although the 314 3HHx composition was decreased from 0.8 mol% to about half by disruption of phaJ4aRe 315 when soybean oil was used as a carbon source (data not shown), there were uncertain 316 compositional changes caused by the gene disruption because of the quite low 3HHx 317 composition in the parent strain. Therefore, the gene disruption was further conducted in 318 strain NSDG∆AB, as the previous study demonstrated that this recombinant strain 319 accumulated low amounts of P(3HB-co-3HHx) with a large 3HHx fraction (26 mol%) on 320 soybean oil due to extremely reduced provision of the (R)-3HB-CoA monomer caused by the 321 disruption of phaAB1Re (24). As shown in Table 4, the gene disruption of each phaJ4 homolog 322 in NSDG∆AB did not influence growth and P(3HB) biosynthesis on fructose. Interestingly, 323 when soybean oil was fed as a carbon source, we observed that the disruption of phaJ4aRe in 324 NSDG∆AB resulted in a significant decrease of 3HHx composition to 3.2 mol% in the 325 accumulated copolyester (Table 4), whereas PHA biosynthesis properties were not affected by 326 the disruption of phaJ4bRe or phaJ4cRe. The amount of 3HB and 3HHx units incorporated into 327 the polymer fraction are also shown in Table 4. R. eutropha NSDG∆A accumulated 3.3 g/L of 328 P(3HB-co-1.2 mol% 3HHx), corresponding to the incorporation into the polymer fraction of 329 36 mmol 3HB units and 1.2 mmol 3HHx units per liter of culture. The disruption of phaB1Re 330 in NSDG∆A resulted in the drastic decrease of 3HB units to 1.6 mmol/L-culture, 331
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
accompanied by a decrease in 3HHx units to 0.60 mmol/L-culture. The disruption of phaJ4aRe 332 in NSDG∆AB further decreased the 3HHx units to 0.048 mmol/L-culture, but did not affect 333 the incorporation of the 3HB unit. These results clearly demonstrated that phaJ4aRe plays an 334 important role in the provision of (R)-3HHx-CoA through β-oxidation in R. eutropha H16, 335 along with PhaB1Re. 336
337 PHA production by recombinant R. eutropha harboring phaJ4a–cRe on the plasmid 338
A series of recombinant plasmids for the introduction of additional copies of the 339 respective phaJ4 gene into R. eutropha was constructed based on pBPP, a broad-host-range 340 plasmid harboring a promoter region for phaP1Re. It has been reported that pBPP can act as a 341 moderately strong constitutive expression plasmid in R. eutropha (9). The resulting plasmids, 342 pBPP-J4a, pBPP-J4b, and pBPP-J4c, were individually transferred into R. eutropha strains 343 H16CAc and NSDG∆A by transconjugation, and then PHA production was examined by the 344 recombinant strains (Table 5). When fructose was used as a carbon source, all the 345 recombinant strains grew and accumulated P(3HB) homopolymer with nearly the same 346 properties as the control strains harboring the empty vector. When additional copies of 347 phaJ4aRe and phaJ4bRe were introduced, this led to an increase in 3HHx composition in 348 P(3HB-co-3HHx) synthesized from soybean oil. The 3HHx fraction was increased from 0.7 349 mol% to 1.2–1.3 mol% in H16CAc, and significantly increased from 2.7 mol% to 8.5–8.9 350 mol% in NSDG∆A by the plasmid-based expression of phaJ4aRe or phaJ4bRe. Such 351 compositional change was not seen when phaJ4cRe was introduced by the plasmid. 352
353 PHA production by recombinant R. eutropha harboring phaJ4a–cRe within chromosomal 354 pha operon 355
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
When soybean oil was used as a carbon source, the cell yields of the R. eutropha 356 strains harboring the empty vector pBPP (2.5–2.7 g dry cell mass/L) were exceedingly 357 reduced when compared with those of the parent strains (4.6–5.2 g-dry cell mass/L), probably 358 because of adverse effects of the plasmid. As the lower cell yield directly reflected to a 359 decrease in PHA productivity, our previous study overcame this problem by the engineering 360 of the pha operon on chromosome 1 of R. eutropha by homologous recombination (24, 25). 361 We therefore inserted phaJ4aRe or phaJ4bRe between phaCNSDG and phaB1Re in the pha 362 operon of NSDG∆A, and PHA production by the resulting strains was examined with soybean 363 oil as a carbon source, as shown in Table 6. The engineered strains well grew and 364 accumulated PHA at the same level as the parent strain (4.7–4.9 g-dry cell mass/L and 3.9–4.0 365 g-PHA/L), which indicated the absence of a negative impact of the introduced phaJ gene(s) 366 on growth and PHA biosynthesis. Moreover, the recombinant strains harboring an additional 367 copy of phaJ4aRe or phaJ4bRe within the pha operon were capable of synthesizing P(3HB-co-368 3HHx) with a much larger 3HHx fraction than the parent strain NASDG∆A, as the 3HHx 369 fraction was increased up to 7.3 and 8.0 mol% by introduction of phaJ4aRe and phaJ4bRe 370 (strains TT101 and TT102), respectively. However, the 3HHx compositions were lower than 371 9.1 mol% in PHA produced by the previously constructed strain MF03 harboring phaJAc. 372 Further constructed strains TT103 and TT104, having a tandem of phaJAc-phaJ4aRe and 373 phaJAc-and phaJ4bRe within the pha operon, respectively, produced P(3HB-co-3HHx) with a 374 higher 3HHx composition (10.2–10.5 mol%) than MF03. Although the increase in the 3HHx 375 composition seemed to be slight, the actual amount of the incorporated 3HHx unit was 376 significantly increased after taking into consideration the accumulation of the huge amount of 377 PHA within the cells. These results suggested that higher expression of (R)-enoyl-CoA 378 hydratase genes could enhance incorporation of the 3HHx unit into PHA synthesized through 379 β-oxidation. 380
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
381 382
DISCUSSION 383 Many previous studies have demonstrated that recombinant strains of R. eutropha 384
possessing heterologous PHA synthase with broad substrate specificity could produce PHA 385 copolyesters composed of the 3HB unit and mcl-3HA units from vegetable oils and alkanoic 386 acids (7, 8, 16, 23-25, 38, 41, 43, 44). These facts apparently indicated that R. eutropha equips 387 the native channeling pathway for supplying the (R)-3-hydroxyacyl-CoA monomer through β-388 oxidation, but the details have been unclear so far, except for the partial contribution of 389 PhaB1Re for the supply of (R)-3HHx-CoA from soybean oil (24). This study focused on R-390 ECH as another possible enzyme involved in the linkage between PHA biosynthesis and β-391 oxidation in R. eutropha. 392
A homology search of the R. eutropha H16 genome identified 16 genes homologous to 393 known PhaJ proteins from A. caviae and P. aeruginosa. Among them, H16_A1070 (PhaJ4aRe), 394 H16_B0397 (PhaJ4bRe), and H16_B0667 (PhaJ4cRe) were investigated in this study because 395 the three proteins share high homology with PhaJ4Pa which is an mcl-R-ECH. An enzyme 396 assay using the recombinant proteins demonstrated that all the three enzymes actually showed 397 R-ECH activity towards 2-enoyl-CoAs of C4–C8. They could accept crotonyl-CoA (C4) as a 398 substrate unlike PhaJ4Pa but the activity levels toward crotonyl-CoA were much lower than 399 those towards the longer substrates, indicating that all three PhaJ4s from R. eutropha are R-400 ECH specific to mcl-substrates. Particularly, PhaJ4aRe and PhaJ4bRe had high catalytic 401 efficiency and high specificity to the C6–C8 substrates. 402
Gene disruption for the newly identified PhaJ4s from R. eutropha clarified that the three 403 R-ECHs have no critical role in growth and PHA biosynthesis on fructose, while PhaJ4aRe 404 participated in the provision of (R)-3HHx-CoA for PHA biosynthesis from soybean oil along 405
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
with PhaB1Re (Table 4). The contribution of PhaB1Re and PhaJ4aRe to the monomer supply for 406 P(3HB-co-3HHx) biosynthesis through β-oxidation was roughly quantified in R. eutropha 407 NSDG∆A, as shown in Figure 5. As described previously, PhaB1Re played a major role in the 408 formation of (R)-3HB-CoA on soybean oil (24). It was estimated that the 3HB unit produced 409 by PhaB1Re from acetoacetyl-CoA occupied 95.5% of the total 3HB unit within the polymer, 410 while PhaJ4a–cRe minimally contributed to the 3HB unit formation despite the activity to 411 crotonyl-CoA. This was probably because of the dominant formation of (R)-3HB-CoA by 412 PhaB1Re having much higher affinity toward acetoacetyl-CoA (Km = approx. 5 μM) (36) than 413 PhaJ4a-cRe toward crotonyl-CoA (Km = 97–311 μM). With respect to the generation of (R)-414 3HHx-CoA, PhaB1Re produced 50% of the total C6-unit from 3-oxohexanoyl-CoA during β-415 oxidation. The ratio of C6/C4 units provided by PhaB1Re was 1.7%, which was consistent with 416 the observation that NADPH-acetoacetyl-CoA reductase activity toward 3-oxohexanoyl-CoA 417 was 3.6% of that toward acetoacetyl-CoA in a crude extract of R. eutropha (12). We further 418 revealed that, among the three R-ECHs, only PhaJ4aRe mediated the formation of (R)-3HHx-419 CoA from 2-hexenoyl-CoA during β-oxidation, and the contribution was estimated to be 45% 420 of the total 3HHx unit incorporated into PHA. Although PhaJ4bRe exhibited higher catalytic 421 efficiency than PhaJ4aRe toward 2-hexenoyl-CoA, an apparent role for PhaJ4bRe in the 422 formation of the C6-unit was not observed, possibly because of low expression of the gene in 423 the cells grown on soybean oil. The results also indicated that small portions of C4 and C6 424 monomers (4.5–5%) were generated by unknown enzymes other than PhaB1Re or PhaJ4aRe, 425 which could likely be other PhaB-FabG-related short-chain dehydrogenases or other R-ECHs. 426
Previous reports have demonstrated that expression of phaJ in PHA-producing 427 bacteria enriched 3HA fractions longer than C6 in copolyesters synthesized from vegetable 428 oils or alkanoic acids (3, 24). In this study, expression of plasmid-borne phaJ4aRe and 429 phaJ4bRe in R. eutropha strains H16CAc and NSDG∆A tended to increase the 3HHx 430
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
composition (Table 5). Additional copies of phaJ4cRe did not affect the copolymer 431 biosynthesis, most likely due to the much lower catalytic efficiency of PhaJ4cRe than that of 432 PhaJ4aRe and PhaJ4bRe. Further engineering was performed by homologous recombination in 433 order to avoid poor growth caused by plasmid-based transformation. The strains TT101 and 434 TT102, having an additional copy of phaJ4aRe or phaJbRe downstream of phaCNSDG on 435 chromosome 1 of R. eutropha NSDG∆A, accumulated P(3HB-co-3HHx) composed of larger 436 3HHx fractions (7.2–8.0 mol%) than the parent strain NSDG∆A (3.0 mol%) with comparably 437 high cell yield and PHA content on soybean oil (Table 6). Unexpectedly, the 3HHx fractions 438 in the copolyester synthesized by TIT101 and TIT102 were smaller than that by MF03 439 harboring phaJAc despite the high activity and specificity of PhaJ4aRe and PhaJ4bRe to longer 440 substrates (Table 3). However, increased expression of phaJ genes was still effective in 441 channeling more of the C6-unit from β-oxidation to PHA, as the strains TT013 and TT014 442 harboring a tandem of phaJAc with phaJ4aRe or phaJ4bRe could accumulate P(3HB-co-3HHx) 443 with a larger 3HHx fraction from soybean oil. These results demonstrated that the new phaJ4s 444 from R. eutropha, as well as phaJAc, are useful genes for the compositional regulation of 445 P(3HB-co-3HHx) produced through β-oxidation. They are also expected to be applied to 446 further engineering of PHA biosynthesis and β-oxidation pathways in various PHA-producing 447 bacteria to achieve wider compositional regulation in the biosynthesis of PHA copolymers. 448
449 450
Acknowledgements 451 This study was supported by KAKENHI (Grant-in-Aid for Scientific Research) on 452
Priority Areas “Applied Genomics” from the Ministry of Education, Culture, Sports, Science 453 and Technology (MEXT) of Japan. 454
455
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
REFERENCES 456 457
1. Brigham, C. J., C. F. Budde, J. W. Holder, Q. Zeng, A. E. Mahan, C. Rha, and A. 458 J. Sinskey. 2010. Elucidation of β-oxidation pathways in Ralstonia eutropha H16 by 459 examination of global gene expression. J. Bacteriol. 192:5454-5464. 460
2. Budde, C. F., A. E. Mahan, J. Lu, C. Rha, and A. J. Sinskey. 2010. Roles of 461 multiple acetoacetyl coenzyme A reductases in polyhydroxybutyrate biosynthesis in 462 Ralstonia eutropha H16. J. Bacteriol. 192:5319-5328. 463
3. Budde, C. F., S. L. Riedel, L. B. Willis, C. Rha, and A. J. Sinskey. 2011. Production 464 of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from plant oil by engineered 465 Ralstonia eutropha strains. Appl. Environ. Microbiol. 77:2847-2854. 466
4. Davis, R., A. Chandrashekar, and T. R. Shamala. 2008. Role of (R)-specific enoyl 467 coenzyme A hydratases of Pseudomonas sp in the production of 468 polyhydroxyalkanoates. Antonie Van Leeuwenhoek 93:285-296. 469
5. Doi, Y., S. Kitamura, and H. Abe. 1995. Microbial synthesis and characterization of 470 poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules 28:4822-4828. 471
6. Fiedler, S., A. Steinbüchel, and B. H. Rehm. 2002. The role of the fatty acid β-472 oxidation multienzyme complex from Pseudomonas oleovorans in 473 polyhydroxyalkanoate biosynthesis: molecular characterization of the fadBA operon 474 from P. oleovorans and of the enoyl-CoA hydratase genes phaJ from P. oleovorans 475 and Pseudomonas putida. Arch. Microbiol. 178:149-160. 476
7. Fukui, T., and Y. Doi. 1997. Cloning and analysis of the poly(3-hydroxybutyrate-co-477 3-hydroxyhexanoate) biosynthesis genes of Aeromonas caviae. J. Bacteriol. 179:4821-478 4830. 479
8. Fukui, T., and Y. Doi. 1998. Efficient production of polyhydroxyalkanoates from 480 plant oils by Alcaligenes eutrophus and its recombinant strain. Appl. Microbiol. 481 Biotechnol. 49:333-336. 482
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
9. Fukui, T., K. Ohsawa, J. Mifune, I. Orita, and S. Nakamura. 2011. Evaluation of 483 promoters for gene expression in polyhydroxyalkanoate-producing Cupriavidus 484 necator H16. Appl. Microbiol. Biotechnol. 89:1527-1536. 485
10. Fukui, T., N. Shiomi, and Y. Doi. 1998. Expression and characterization of (R)-486 specific enoyl coenzyme A hydratase involved in polyhydroxyalkanoate biosynthesis 487 by Aeromonas caviae. J. Bacteriol. 180:667-673. 488
11. Fukui, T., S. Yokomizo, G. Kobayashi, and Y. Doi. 1999. Co-expression of 489 polyhydroxyalkanoate synthase and (R)-enoyl-CoA hydratase genes of Aeromonas 490 caviae establishes copolyester biosynthesis pathway in Escherichia coli. FEMS 491 Microbiol. Lett. 170:69-75. 492
12. Haywood, G. W., A. J. Anderson, L. Chu, and E. A. Dawes. 1988. The role of 493 NADH-linked and NADPH-linked acetoacetyl-CoA reductases in the poly-3-494 hydroxybutyrate synthesizing organism Alcaligenes eutrophus. FEMS Microbiol. Lett. 495 52:259-264. 496
13. Hisano, T., T. Tsuge, T. Fukui, T. Iwata, K. Miki, and Y. Doi. 2003. Crystal 497 structure of the (R)-specific enoyl-CoA hydratase from Aeromonas caviae involved in 498 polyhydroxyalkanoate biosynthesis. J. Biol. Chem. 278:617-624. 499
14. Hu, F., Y. Cao, F. Xiao, J. Zhang, and H. Li. 2007. Site-directed mutagenesis of 500 Aeromonas hydrophila enoyl coenzyme A hydratase enhancing 3-hydroxyhexanoate 501 fractions of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Curr. Microbiol. 55:20-502 24. 503
15. Huyghe, S., G. P. Mannaerts, M. Baes, and P. P. Van Veldhoven. 2006. 504 Peroxisomal multifunctional protein-2: the enzyme, the patients and the knockout 505 mouse model. Biochim. Biophys. Acta. 1761:973-994. 506
16. Kahar, P., T. Tsuge, K. Taguchi, and Y. Doi. 2004. High yield production of 507 polyhydroxyalkanoates from soybean oil by Ralstonia eutropha and its recombinant 508 strain. Polym. Degrad. Stab. 83:79-86. 509
17. Kato, M., H. J. Bao, C. K. Kang, T. Fukui, and Y. Doi. 1996. Production of a novel 510 copolyester of 3-hydroxybutyric acid and medium chain length 3-hydroxyalkanaic 511
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
acids by Pseudomonas sp. 61-3 from sugars. Appl. Microbiol. Biotechnol. 45:363-512 370. 513
18. Kunau, W. H., and A. Hartig. 1992. Peroxisome biogenesis in Saccharomyces 514 cerevisiae. Antonie Van Leeuwenhoek 62:63-78. 515
19. Lindenkamp, N., K. Peplinski, E. Volodina, A. Ehrenreich, and A. Steinbüchel. 516 2010. Impact of multiple β-ketothiolase deletion mutations in Ralstonia eutropha H16 517 on the composition of 3-mercaptopropionic acid-containing copolymers. Appl. 518 Environ. Microbiol. 76:5373-5382. 519
20. Lu, X. Y., Q. Wu, W. J. Zhang, G. Zhang, and G. Q. Chen. 2004. Molecular 520 cloning of polyhydroxyalkanoate synthesis operon from Aeromonas hydrophila and its 521 expression in Escherichia coli. Biotechnol Prog 20:1332-1336. 522
21. Madison, L. L., and G. W. Huisman. 1999. Metabolic engineering of poly(3-523 hydroxyalkanoates): from DNA to plastic. Microbiol. Mol. Biol. Rev. 63:21-53. 524
22. Mannaerts, G. P., P. P. Van Veldhoven, and M. Casteels. 2000. Peroxisomal lipid 525 degradation via β- and α-oxidation in mammals. Cell Biochem. Biophys. 32 526 Spring:73-87. 527
23. Matsusaki, H., H. Abe, K. Taguchi, T. Fukui, and Y. Doi. 2000. Biosynthesis of 528 poly(3-hydroxybutyrate-co-3-hydroxyalkanoates) by recombinant bacteria expressing 529 the PHA synthase gene phaC1 from Pseudomonas sp. 61-3. Appl. Microbiol. 530 Biotechnol. 53:401-409. 531
24. Mifune, J., S. Nakamura, and T. Fukui. 2010. Engineering of pha operon on 532 Cupriavidus necator chromosome for efficient biosynthesis of poly(3-533 hydroxybutyrate-co-3-hydroxyhexanoate) from vegetable oil. Polym. Degrad. Stab. 534 95:1305-1312. 535
25. Mifune, J., S. Nakamura, and T. Fukui. 2008. Targeted engineering of Cupriavidus 536 necator chromosome for biosynthesis of poly(3-hydroxybutyrate-co-3-537 hydroxyhexanoate) from vegetable oil. Can. J. Chem. 86:621-627. 538
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
26. Park, S. J., W. S. Ahn, P. R. Green, and S. Y. Lee. 2001. Production of poly(3-539 hydroxybutyrate-co-3-hydroxyhexanoate) by metabolically engineered Escherichia 540 coli strains. Biomacromolecules 2:248-254. 541
27. Park, S. J., and S. Y. Lee. 2003. Identification and characterization of a new enoyl 542 coenzyme A hydratase involved in biosynthesis of medium-chain-length 543 polyhydroxyalkanoates in recombinant Escherichia coli. J. Bacteriol. 185:5391-5397. 544
28. Philip, S., T. Keshavarz, and I. Roy. 2007. Polyhydroxyalkanoates: biodegradable 545 polymers with a range of applications. J. Chem. Technol. Biotechnol. 82:233-247. 546
29. Pohlmann, A., W. F. Fricke, F. Reinecke, B. Kusian, H. Liesegang, R. Cramm, T. 547 Eitinger, C. Ewering, M. Pötter, E. Schwartz, A. Strittmatter, I. Voß, G. 548 Gottschalk, A. Steinbüchel, B. Friedrich, and B. Bowien. 2006. Genome sequence 549 of the bioplastic-producing "Knallgas" bacterium Ralstonia eutropha H16. Nat. 550 Biotechnol. 24:1257-1262. 551
30. Poirier, Y., V. D. Antonenkov, T. Glumoff, and J. K. Hiltunen. 2006. Peroxisomal 552 β-oxidation--a metabolic pathway with multiple functions. Biochim. Biophys. Acta 553 1763:1413-1426. 554
31. Reiser, S. E., T. A. Mitsky, and K. J. Gruys. 2000. Characterization and cloning of 555 an (R)-specific trans-2,3-enoylacyl-CoA hydratase from Rhodospirillum rubrum and 556 use of this enzyme for PHA production in Escherichia coli. Appl. Microbiol. 557 Biotechnol. 53:209-218. 558
32. Sato, S., H. Kanazawa, and T. Tsuge. 2011. Expression and characterization of (R)-559 specific enoyl coenzyme A hydratases making a channeling route to 560 polyhydroxyalkanoate biosynthesis in Pseudomonas putida. Appl. Microbiol. 561 Biotechnol. 90:951-959. 562
33. Schäfer, A., A. Tauch, W. Jäger, J. Kalinowski, G. Thierbach, and A. Pühler. 1994. 563 Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli 564 plasmids pK18 and pK19: selection of defined deletions in the chromosome of 565 Corynebacterium glutamicum. Gene 145:69-73. 566
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

23
34. Shimamura, E., K. Kasuya, G. Kobayashi, T. Shiotani, Y. Shima, and Y. Doi. 567 1994. Physical properties and biodegradability of microbial poly(3-hydroxybutyrate-568 co-3-hydroxyhexanoate). Macromolecules 27:878-880. 569
35. Simon, R., U. Priefer, and A. Pühler. 1983. A broad host range mobilization system 570 for in vivo genetic engineering. Transposon mutagenesis in gram negative bacteria. 571 Bio/Technology 1:784-791. 572
36. Steinbüchel, A., and H. G. Schlegel. 1991. Physiology and molecular genetics of 573 poly(β-hydroxyalkanoic acid) synthesis in Alcaligenes eutrophus. Mol. Microbiol. 574 5:535-542. 575
37. Sudesh, K., H. Abe, and Y. Doi. 2000. Synthesis, structure and properties of 576 polyhydroxyalkanoates: biological polyesters. Prog. Polym. Sci. 25:1503-1555. 577
38. Timm, A., and A. Steinbüchel. 1992. Cloning and molecular analysis of the poly(3-578 hydroxyalkanoic acid) gene locus of Pseudomonas aeruginosa PAO1. Eur. J. 579 Biochem. 209:15-30. 580
39. Tsuge, T., T. Fukui, H. Matsusaki, S. Taguchi, G. Kobayashi, A. Ishizaki, and Y. 581 Doi. 2000. Molecular cloning of two (R)-specific enoyl-CoA hydratase genes from 582 Pseudomonas aeruginosa and their use for polyhydroxyalkanoate synthesis. FEMS 583 Microbiol. Lett. 184:193-198. 584
40. Tsuge, T., T. Hisano, S. Taguchi, and Y. Doi. 2003. Alteration of chain length 585 substrate specificity of Aeromonas caviae R-enantiomer-specific enoyl-coenzyme A 586 hydratase through site-directed mutagenesis. Appl. Environ. Microbiol. 69:4830-4836. 587
41. Tsuge, T., Y. Saito, Y. Kikkawa, T. Hiraishi, and Y. Doi. 2004. Biosynthesis and 588 compositional regulation of poly[(3-hydroxybutyrate)-co-(3-hydroxyhexanoate)] in 589 recombinant Ralstonia eutropha expressing mutated polyhydroxyalkanoate synthase 590 genes. Macromol. Biosci. 4:238-242. 591
42. Tsuge, T., K. Taguchi, T. Seiichi, and Y. Doi. 2003. Molecular characterization and 592 properties of (R)-specific enoyl-CoA hydratases from Pseudomonas aeruginosa: 593 metabolic tools for synthesis of polyhydroxyalkanoates via fatty acid β-oxidation. Int. 594 J. Biol. Macromol. 31:195-205. 595
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

24
43. Tsuge, T., S. Watanabe, D. Shimada, H. Abe, Y. Doi, and S. Taguchi. 2007. 596 Combination of N149S and D171G mutations in Aeromonas caviae 597 polyhydroxyalkanoate synthase and impact on polyhydroxyalkanoate biosynthesis. 598 FEMS Microbiol. Lett. 277:217-222. 599
44. Tsuge, T., T. Yamamoto, K. Yano, H. Abe, Y. Doi, and S. Taguchi. 2009. Evaluating 600 the ability of polyhydroxyalkanoate synthase mutants to produce P(3HB-co-3HA) 601 from soybean oil. Macromol. Biosci. 9:71-78. 602
45. Vo, M. T., K. W. Lee, Y. M. Jung, and Y. H. Lee. 2008. Comparative effect of 603 overexpressed phaJ and fabG genes supplementing (R)-3-hydroxyalkanoate monomer 604 units on biosynthesis of mcl-polyhydroxyalkanoate in Pseudomonas putida 605 KCTC1639. J. Biosci. Bioeng. 106:95-98. 606
607 608 609
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

25
LEGENDS TO FIGURES 610 Fig. 1. A phylogenetic tree of MaoC-like domains in PhaJ homologs encoded in R. eutropha 611 H16 genome and the domains in the known PhaJ proteins from A. caviae and P. aeruginosa. 612 The proteins derived from R. eutropha were represented as the locus tags assigned in the 613 genome (29). The “_N” and “_C” at the end of the locus tags indicate N-terminal and C-614 terminal domains of the proteins, respectively. Ac-PhaJ; PhaJ from A. caviae FA440 615 (BAA21816), and Pa-PhaJ1, Pa-PhaJ2, Pa-PhaJ3, and Pa-PhaJ4; PhaJ1 (AAG06690), PhaJ2 616 (AAG04407), PhaJ3 (AAG08174), and PhaJ4 (AAG07402) from P. aeruginosa PAO1. 617 618 Fig. 2. Organization of phaJ4a (upper), phaJ4b (middle), and phaJ4c (bottom) loci in R. 619 eutropha H16 genome. The phaJ4 genes are indicated by dark gray, and the genes with 620 predicted functions in β-oxidation are indicated by light gray. The gene are represented as the 621 locus tags. 622 623 Fig. 3. SDS-PAGE analysis of the recombinant PhaJ4 ptoteins purified from E. coli 624 BL21(DE3) harboring the respective expression vector. Lanes 1, PhaJ4aRe; 2, PhaJ4bRe; 3, 625 PhaJ4cRe. 626 627 Fig. 4. Evaluation of the stereospecificity of PhaJ4aRe, PhaJ4bRe, and PhaJ4cRe. Hydration of 628 crotonyl-CoA by the recombinant forms of PhaJ4 proteins coupled with (R)-specific 629 polymerization catalyzed by the crude PHA synthase from R. eutropha was 630 spectrophotometrically monitored. Open circle, addition of PhaJ4aRe; open triangle, addition 631 of PhaJ4bRe; open square, addition of PhaJ4cRe; closed circle, no addition of enoyl-CoA 632 hydratase. 633 634
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

26
Fig. 5. Proposed pathway of P(3HB-co-3HHx) biosynthesis from soybean oil by the 635 recombinant strain of R. eutropha NSDG∆A, along with estimated contribution of monomer-636 supplying enzymes to biosynthesis of the copolyester through β-oxidation. 637 638 639 640 641
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 1. Bacterial strains and plasmids used in this study
Strains or plasmids Relevant markers Reference or source
Escherichia coli
DH5α deoR, endA1, gyrA96, hsdR17 (rK- mK
+), recA1, relA1, supE44, thi-l, ∆(lacZYA-argFV169), φ80lacZ∆M15, F-, λ- Clontech
S17-1 thi, pro, hsdR, recA, chromosomal RP4; Tra+; Tmpr Str/Spcr (35) BL21(DE3) E. coli B, F-, dcm, ompT, hsdS(rB- mB-), gal, λ(DE3) Stratagene
Ralstonia eutropha
H16 Wild strain DSM428 H16CAc H16 derivative; ΔphaCRe::phaCAc (25) NSDGΔA H16 derivative; ΔphaCRe::phaCNSDG, ΔphaARe (24) NSDGΔAB H16 derivative; ΔphaCRe::phaCNSDG, ΔphaAB1Re (24) H16CAcΔJ4a H16CAc derivative; ΔphaJ4aRe This study
H16CAcΔJ4b H16CAc derivative; ΔphaJ4aRe This study
H16CAcΔJ4c H16CAc derivative; ΔphaJ4cRe This study NSDGΔABJ4a NSDGΔAB derivative; ΔphaJ4aRe This study NSDGΔABJ4b NSDGΔAB derivative; ΔphaJ4bRe This study NSDGΔABJ4c NSDGΔAB derivative; ΔphaJ4cRe This study MF03 ∆phaCRe::phaCNSDG-phaJAc, ΔphaARe (24) TT101 ∆phaCRe::phaCNSDG-phaJ4aRe, ΔphaARe This study TT102 ∆phaCRe::phaCNSDG-phaJ4bRe, ΔphaARe This study TT103 ∆phaCRe::phaCNSDG-phaJAc-phaJ4aRe, ΔphaARe This study TT104 ∆phaCRe::phaCNSDG-phaJAc-phaJ4bRe, ΔphaARe This study
Plasmids
pColdII ColE1 ori, Ampr, lacI, cspA promoter Takara Bio pColdII-J4a pColdII carrying phaJ4aRe This study pColdII-J4b pColdII carrying phaJ4bRe This study pColdII-J4c pColdII carrying phaJ4cRe This study pBPP pBBR ori, mob, Kanr, phaP1Re promoter (9) pBPP-J4a pBPP carrying phaJ4aRe This study pBPP-J4b pBPP carrying phaJ4bRe This study pBPP-J4c pBPP carrying phaJ4cRe This study pK18mobsacB pMB1 ori, mob, Kanr, sacB (33) pK18ms∆J4a pK18mobsacB carrying phaJ4aRe dela This study pK18ms∆J4a pK18mobsacB carrying phaJ4bRe del This study pK18ms∆J4a pK18mobsacB carrying phaJ4cRe del This study pK18msNSDG-J4aB pK18mobsacB carrying phaCNSDG-phaJ4aRe-phaB1Re This study pK18msNSDG-J4bB pK18mobsacB carrying phaCNSDG-phaJ4bRe-phaB1Re This study pK18msNSDG-JJ4aB pK18mobsacB carrying phaCNSDG-phaJAc-phaJ4aRe-phaB1Re This study pK18msNSDG-JJ4bB pK18mobsacB carrying phaCNSDG-phaJAc-phaJ4bRe-phaB1Re This study
aThe postfix del indicates inserts used for targeted gene deletion.
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 2. Sequences of primers used in this study
Primers Sequences A1070-F TCCATATGCGTACCATCGCATCGCTG (underline: NdeI) A1070-R CGCTCGAGTCACCCGTAGCGGCGCGTGA (underline: XhoI) B0397-F CACATATGAAGACCTACGAGAACATC (underline: NdeI) B0397-R CTCGAGTCAGGGAAAGCGCCGCAGGA (underline: XhoI) B0667-F CACATATGCCAAGAATCTTCCGTTCT (underline: NdeI) B0667-R ATCTCGAGTCAGAAGTAATAGCGGCTG (underline: XhoI) A1070up-F CAGGCGTCTAGACACCGCATGGTGGAGATGTT (underline: XbaI) A1070up-R ACTGCGTATCCGTCTTGACGATAAAGTGTTCT A1070down-F TTATCGTCAAGACGGATACGCAGTTTGACGCGGTGC A1070down-R ACCTCCAAGCTTGTGACGGGCTTCTTGCTGGAT (underline: HindIII) B0397up-Fw CGTGCGGAATTCTGTCGACAAGGAATACGACC (underline: EcoRI) B0397up-Rv GGGCGGCCTGCCCGACGGCGCGCTTGAAGTAATGG B0397down-F AGCGCGCCGTCGGGCAGGCCGCCCGTCTTTTGCCAT B0397down-R GCTGCTAAGCTTCATTGAGCTTGCCCGGATTG (underline: HindIII) B0667up-F CACCATGAATTCCGGCACCAACGAAATCCAGC (underline: EcoRI) B0667up-R CTGGATTCCTGTCCCGTCATGTACGAGGCATAGTCG B0667down-F GTACATGACGGGACAGGAATCCAGAATGAAAG B0667down-R TTATCAAAGCTTTCATGATGTCTCCTCCGGTT (underline: HindIII) A1070Ins-F TTGACACTAGTTCTAGAAGGAGGAGTATATACATATGCGTACCATCGCATCGC
(underline: SpeI-XbaI) A1070Ins-R GCCAAGCGGCCGCTTCGAAACTAGTTGCAGCTCGACTCACCCGTAG
(underline: NotI-Csp45I-SpeI) B0397Ins-F GACCCACTAGTTCTAGAAGGAGGAATATACATATGAAGACCTACGAGAACAT
(underline: SpeI-XbaI) B0397Ins-R CAAGCGCGGCCGCTTCGAAACTAGTTCAGGGAAAGCGCCGCAGGAT
(underline: NotI-Csp45I-SpeI)
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 3. Kinetic parameters of recombinant PhaJ4a–cRe for hydration towards trans-2-enoyl-CoAs of C4–C8 and comparison with those of PhaJ4Pa and PhaJAc
Enzyme (Ref) Length of substrate
Km (µM)
kcat (s-1)
kcat/Km
(s-1 M-1)
kcat/Km
ratio (Cx/C4)
Ref
C4 97 ±5 3.8 ±0.09 ×102 4.0 ×106 1 C6 41 ±7 3.3 ±0.24 ×103 8.0 ×107 20
PhaJ4aRe
C8 24 ±5 3.1 ±0.32 ×103 1.3 ×108 33
C4 311 ±15 6.4 ±0.45 ×102 2.1 ×106 1 C6 77 ±14 1.1 ±1.38 ×104 1.4 ×108 67
PhaJ4bRe
C8 23 ±4 8.7 ±0.44 ×103 3.8 ×108 1.8 ×102
C4 178 ±46 5.8 ±0.89 ×101 3.0 ×105 1 C6 110 ±16 4.8 ±0.44 ×102 4.3 ×106 14
PhaJ4cRe
C8 38 ±12 1.3 ±0.23 ×103 3.3 ×107 1.1 ×102 PhaJ4Pa C4 - - - C6 198 ±12 1.1 ±0.05 ×103 5.7 ×106 (41) C8 118 ±11 9.5 ±0.50 ×102 8.0 ×106 PhaJAc C4 24 ±2 1.9 ±0.09 ×103 8.0 ×107 1 C6 40 ±5 2.9 ±0.2 ×102 7.4 ×106 9.3 ×10-2 (39) C8 42 ±5 0.58 ±0.04 1.4 ×104 1.8 ×10-4
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 4. Effects of deletion of phaJ4a–cRe on PHA production by R. eutropha strains from fructose and soybean oil
Carbon source / Strain Dry cell
mass [g/L]
PHA cont.
[wt%] PHA [g/L]
3HHx comp.
[mol%]
3HB unit in PHAa [mmol/
L-culture]
3HHx unit in PHAa [mmol/
L-culture] Fructose NSDG∆Ab 1.8 ±0.08 47 ±3.0 0.85 0 8.1 - NSDGΔAB 1.5 ±0.04 45 ±1.6 0.68 0 6.5 - NSDGΔABJ4a 1.4 ±0.10 42 ±3.0 0.59 0 5.8 - NSDGΔABJ4b 1.5 ±0.05 44 ±0.5 0.66 0 6.1 - NSDGΔABJ4c 1.5 ±0.05 47 ±1.8 0.71 0 6.9 -
Soybean oil
NSDG∆A 4.7 ±0.13 82 ±0.5 3.9 3.0 ±0.03 36 1.1 NSDGΔABb 0.94 ±0.14 26 ±0.6 0.20 26 ±0.2 1.6 0.60 NSDGΔABJ4a 0.73 ±0.06 22 ±0.7 0.16 3.2 ±0.2 1.5 0.048 NSDGΔABJ4b 0.92 ±0.05 30 ±1.6 0.27 25 ±1.2 1.8 0.61 NSDGΔABJ4c 0.88 ±0.04 30 ±1.6 0.26 25 ±0.9 1.8 0.59
Cells were cultivated at 30°C for 72 hours in an MB medium containing 0.5% (w/v) fructose or 1% (v/v) soybean oil. aAmount of each the monomer unit incorporated into PHA. bData from Ref. (24)
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from
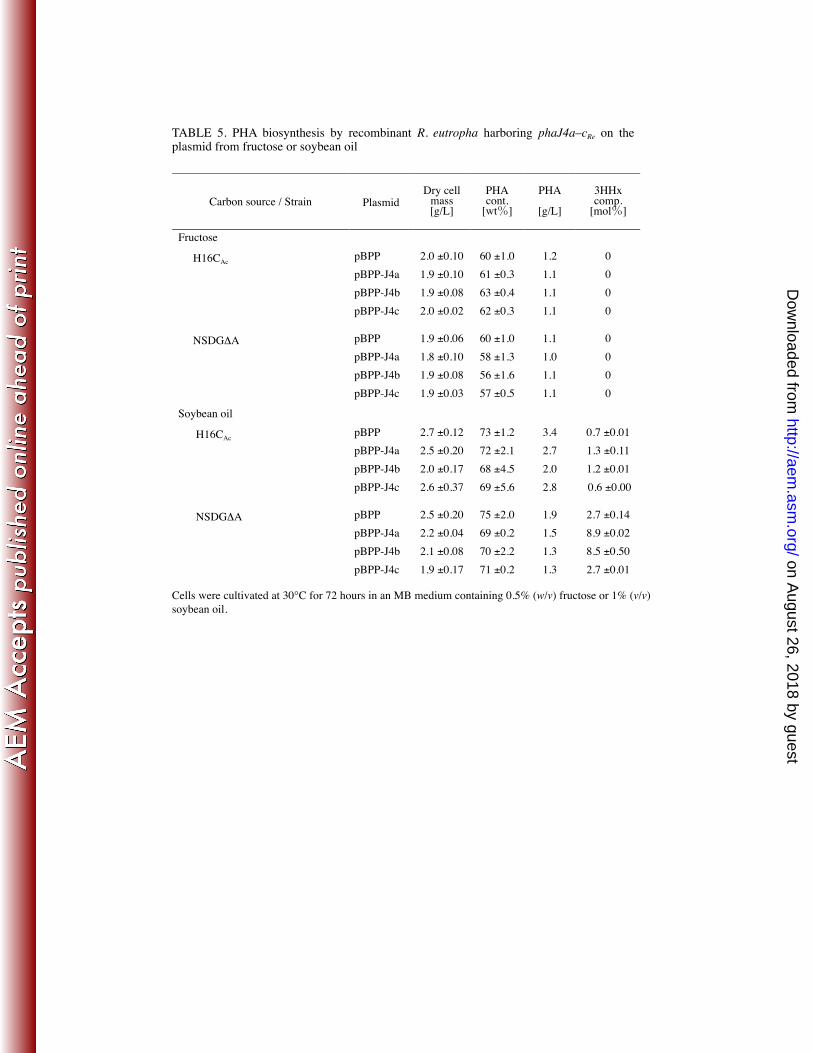
TABLE 5. PHA biosynthesis by recombinant R. eutropha harboring phaJ4a–cRe on the plasmid from fructose or soybean oil
Cells were cultivated at 30°C for 72 hours in an MB medium containing 0.5% (w/v) fructose or 1% (v/v) soybean oil.
Carbon source / Strain Plasmid Dry cell
mass [g/L]
PHA cont.
[wt%]
PHA
[g/L]
3HHx comp.
[mol%]
Fructose pBPP 2.0 ±0.10 60 ±1.0 1.2 0 pBPP-J4a 1.9 ±0.10 61 ±0.3 1.1 0 pBPP-J4b 1.9 ±0.08 63 ±0.4 1.1 0
H16CAc
pBPP-J4c 2.0 ±0.02 62 ±0.3 1.1 0
pBPP 1.9 ±0.06 60 ±1.0 1.1 0 pBPP-J4a 1.8 ±0.10 58 ±1.3 1.0 0 pBPP-J4b 1.9 ±0.08 56 ±1.6 1.1 0
NSDGΔA
pBPP-J4c 1.9 ±0.03 57 ±0.5 1.1 0
Soybean oil
pBPP 2.7 ±0.12 73 ±1.2 3.4 0.7 ±0.01 pBPP-J4a 2.5 ±0.20 72 ±2.1 2.7 1.3 ±0.11 pBPP-J4b 2.0 ±0.17 68 ±4.5 2.0 1.2 ±0.01
H16CAc
pBPP-J4c 2.6 ±0.37 69 ±5.6 2.8 0.6 ±0.00
pBPP 2.5 ±0.20 75 ±2.0 1.9 2.7 ±0.14 pBPP-J4a 2.2 ±0.04 69 ±0.2 1.5 8.9 ±0.02 pBPP-J4b 2.1 ±0.08 70 ±2.2 1.3 8.5 ±0.50
NSDGΔA
pBPP-J4c 1.9 ±0.17 71 ±0.2 1.3 2.7 ±0.01
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 6. PHA biosynthesis by recombinant R. eutropha harboring phaJ(s) within chromosomal pha operon from soybean oil
Strain pha operon organization Dry cell mass [g/L]
PHA cont.
[wt%]
PHA
[g/L]
3HHx comp.
[mol%]
NSDGΔA phaCNSDG -BRe 4.7 ±0.13 82 ±0.54 3.9 3.0 ±0.03 MF03 phaCNSDG -JAc-BRe 4.8 ±0.14 82 ±1.9 3.9 9.1 ±0.06 TT011 phaCNSDG -J4aRe-BRe 4.7 ±0.10 81 ±0.72 3.9 7.2 ±0.06 TT012 phaCNSDG -J4bRe-BRe 4.9 ±0.18 82 ±1.9 4.0 8.0 ±0.08 TT013 phaCNSDG-JAc-J4aRe-BRe 4.7 ±0.08 82 ±2.4 3.9 10.5 ±0.15 TT014 phaCNSDG-JAc-J4bRe-BRe 4.7 ±0.09 81 ±0.77 3.8 10.1 ±0.20
Cells were cultivated at 30°C for 72 hours in an MB medium containing 1% (v/v) soybean oil.
on August 26, 2018 by guest
http://aem.asm
.org/D
ownloaded from