Embed Size (px)
Citation preview

Circa l’80% di tutti gli ormoni e neurotrasmettitori e neuromodulatori inducono le loro risposte combinandosi con recettori che sono accoppiati agli
effettori con proteine G

Classificazione dei GPCRs
da Gether, Endocr.Rev. 21,90,200o

Ai recettori accopiati alle proteine G si legano agonisti chimicamente molto diversi:
• Amine biogene (noradrenalina, dopamina, serotonina)
• Peptidi (sostanza P, angiotensina)
• Ormoni glicoproteici (LH, FSH, TSH)
• Fotoni (nel caso il recettore sia il pigmento visivo rodopsina)

Ciclo di funzionamento delle G-proteine
da Milligam e Kostenis, Br J. Phacol., 147, S46, 2006



La transizione tra forma inattiva e forma attiva e indotta dal recettore che agisce come promotore di scambio dei nucleotidi guanilici accendendo il segnale
Il ritorno alla forma inattiva è garantita dalle stesse proprietà intrinseche della subunità che dopo un certo tempo di interazione con l’effettore, idrolizza il GTP spegnendo il segnale


GDP
Membrana cellulare
Le proteine G sono eterotrimeri costituiti da 3 subunità di peso
molecolare decrescente
41000-45000
35000
10000

Subunità delle proteine G
Ad oggi si conoscono:
• 16 subunità
• 5 subunità
• 14 subunità
Probabilmente quindi un complesso può formare un eterotrimero con diverse subunità

Struttura subunità
• Le diverse subunità hanno una elevata omologia di sequenza che spiega le loro proprietà comuni
• Sono ad es. altamenti conservati 5 segmenti di pochi aminoacidi ( G1-5) che formano il sito di legame e di idrolisi dei nucleotidi guanilici
• Sono differenti invece le regioni che interagiscono con i recettori (carbossiterminale) ed effettori. Esse conferiscono la specificità d’azione delle subunità

Una delle principali funzioni della proteina G è quella di promuovere la formazione di secondi messaggeri cioè molecole che permettono di trasmettere il segnale dato dal primo messaggero (ligando endogeno o Farmaco)

I PRINCIPALI BERSAGLI DELLE PROTEINE G SONO:
1. L’adenilato ciclasi (AC) che catalizza la formazione di AMP-ciclico cAMP)
2. La fosfolipasi C che catalizza la produzione di inositolo-3-fosfato (IP3)
3. La guanilato ciclasi 4. I canali ionici in particolare quelli del
calcio e del potassio 5. L a Rho A/Rho chinasi che controlla
l’attività di diverse vie di segnale per la proliferazione e crescita cellulare

I secondi messaggeri derivati dall’attivazione degli enzimi bersaglio
comprendono:
• cAMP • cGMP • IP3/DAG • Ca2+
• Acido Arachidonico (AA)

I secondi messaggeri regolano molti aspetti funzionali cellulari (effettori finali) attraverso l’attivazione di proteine ad attività chinasica (PKG, PKA PKC): •Enzimi coinvolti nel metabolismo energetico •La divisione e differenziamento cellulare •Il trasporto ionico •I canali ionici •Le proteine contrattili della muscolatura liscia


Spegnimento del segnale
• In alcuni sistemi l’effettore partecipa alla fase di spegnimento del segnale accelerando l’drolisi del GTP da parte della subunità
• Un’intera famiglia di proteine denominate RGS interagiscono con la subunità legata al GTP e accelerano l’idrolisi del nucleotide

AMPLIFICAZIONE DEL SEGNALE
• Il segnale evocato dal recettore viene amplificato perché:
• Una singola molecola recettoriale può attivare più di una proteina G
• La subunità può permanere in forma attiva per un certo tempo


In base alla sequenza aminoacidica e alla capacità di modulare l’attività di effettori differenti le subunità possono essere suddivise in tre classi:
• s stimola l’adenilato ciclasi (Gs: recettori adrenergici (β), dopaminergici (D1,D3,D5) e TSH
• i ; o inibiscono l’adenilato ciclasi (Gi: recettori adrenergici (α2), dopaminergici (D2,D4)
muscarinici (M2,M4) e somatostatina) (Go: abbondanti nel SNC) anche Gτ nel processo di fototrasduzione
• q regolazione fosfolipasi C (Gq)
• 12 e α 13 media effetti acido arachidonico

Le subunità di alcune proteine G vengono
alterate nel loro normale funzionamento dalla
tossina del colera e dalla tossina della pertosse
Entrambe sono dotate di attività ADP-Ribosil
transferasica (cioè trasferiscono un gruppo di
ADP-ribosio)

ADP-RIBOSILAZIONE


Tossina del colera • ADP-ribosila la subunità s in corrispondenza
di una arginina localizzata in G2 una delle regioni coinvolte nella idrolisi del GTP
• La conseguenza funzionale è l’inibizione della attività GTP-asica della proteina che rimane bloccata nella sua forma attiva inducendo una stimolazione persistente dell’adenilato ciclasi ed una continua produzione di cAMP

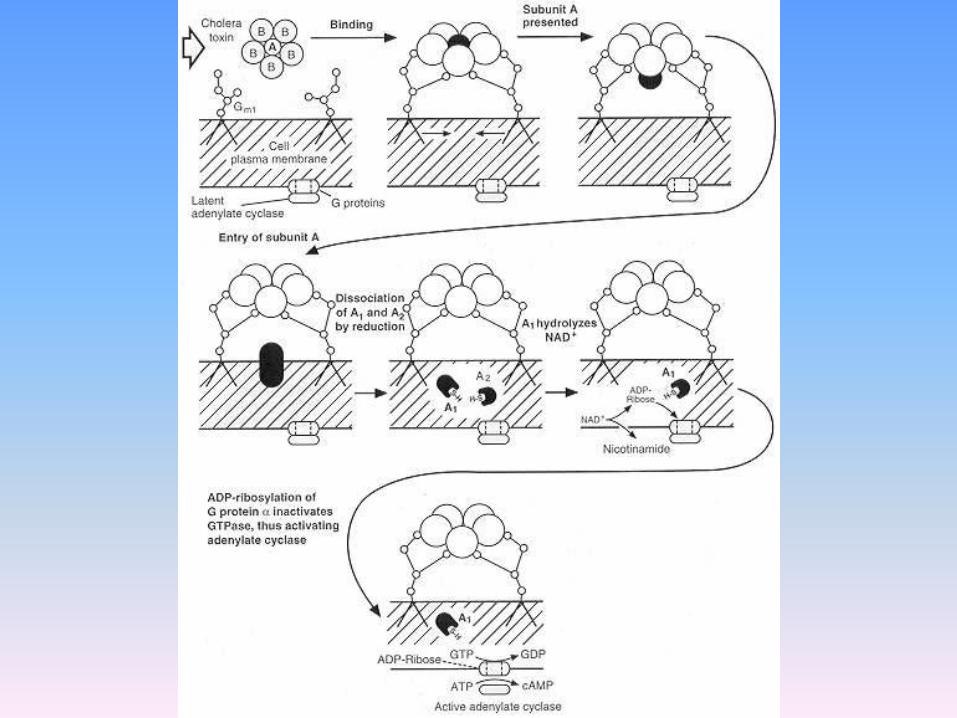

Tossina della pertosse
• ADP-ribosila le subunità i e o in corrispondenza di una Cys localizzata ad una distanza di 4 aminoacidi dal terminale carbossilico
• Tale regione è implicata nel legame con il recettore e di conseguenza la ADP-ribosilazione impedisce
l’attivazione della proteina G da parte del recettore
• La tossina della pertosse mantiene quindi la Proteina G in uno stato inattivo

Esistono 10 isoforme di AC: •9 di membrana •1 solubile
Sono divisibili i 5 famiglie : Forme sensibili alla Ca2+ -Calmodulina: AC1, AC3, AC8 Le stimolate dai recettori β :AC2, AC4, AC7 Quella caratterizzate da inibizione da Ca 2+ : A5, A6 Quella insensibile al diterpene Forskolina: AC9 La più simile alle forme batteriche: sAC

Struttura dell’adenilato ciclasi (AC)
da Cooper, Biochem. J. , 375:517-529, 2003

Molti effetti trasduzionali del cAMP sono mediati dalla Proteinchinasi A (PKA)
PKA in forma inattiva è formata da due subunità: una catalitica e una regolatrice. Il legame con cAMP libera l’unità catalitica che esplica così la sua attività chinasica su numerosi specifici substrati proteici a livello dei residui serina/treonina.

cAMP ha anche altri bersagli cellulari in maniera non mediata da PKA, in particolare alcuni canali ionici coinvolti in processi sensoriali di fototrasduzione e di recezione olfattiva

L’AMPc prodotto dell’AC viene idrolizzato a AMP dalle Fosfodiesterasi (PDE) Se ne conoscono 11 isoforme ( alcuni farmaci specifici come le metilxantine utilizzate nella profilassi e trattamento dell’asma bronchiale sono inibitori di molte PDE)

Le fosofodiesterasi
da , Bender e Beavo, Pharmacol Rev., 58, 488, 2006

cGMP come secondo messaggero
La ciclizzazione del cGMP è ad opera di una Guanidilato Ciclasi GC a partire dal GTP. Ci sono due forme di GC: le solubili attivate direttamente dal NO e quelle di membrana (classe di circa 7 isoenzimi). cGMP attiva la Proteinchinasi G (PKG) Molti degli effetti sono ancora poco conosciuti

Complesso • Nell’eterotrimero il complesso inibisce la
dissociazione del GDP garantendo che, in assenza di stimolazione recettoriale, l’ subunità mantenga la conformazione inattiva
• Il complesso è necessario per l’interazione della proteina G con il recettore. Il recettore attivato è in grado di legare con alta affinità solamente l’eterotrimero e non l’ -GDP
• Il complesso liberato dall’attivazione recettoriale agisce anche come molecola segnale capace di interagire direttamente con effettori specifici

Effettori delle proteine G
Comprendono:
• effettori enzimatici (portano alla formazione nel citoplasma di sostanze biologicamente attive)
• effettori canali (portano a modificazioni della concentrazione ionica intracellulare)

Effettori delle subunità
• Attivano i canali ionici per il K
• Inibiscono specifici canali del Ca
• Stimolano alcune forme di fosfolipasi C
• Attivano l’enzima fosfatidilinositolo 3-chinasi
• Innescano la cascata delle chinasi MAP e ERK


I recettori legati alle proteine G possono direttamente controllare i canali ionici oltre che con il complesso βϒ anche attraverso l’interazione diretta con la subunità α attiva della proteina G senza l’intervento dei secondi messaggeri

CIÒ CHE ACCOMUNA GLI EFFETTORI ENZIMATICI È
La produzione di metaboliti che prendono il nome di secondi messaggeri in quanto fungono da anello di congiunzione tra l’attivazione recettoriale e la risposta della cellula

I secondi messaggeri sono definiti da due caratteristiche:
• Generano una o più risposte legandosi all’interno della cellula a substrati specifici dei quali ne modulano l’attività
• Hanno una attività transitoria e dipendente dallo stimolo primario che l’ha generata, in quanto ci sono sistemi deputati alla loro degradazione o rimozione

Gli effettori delle proteine G quindi possono essere, con o senza secondo messaggero:
• Enzimi
• Canali ionici

Effettori enzimatici delle Proteine G
Comprendono:
• L’adenilato ciclasi
• La guanilato ciclasi
• La fosfolipasi C
• La fosfolipasi A
• La fosfolipasi D

Tra gli effettori delle proteine G con funzione di canale ionico i più studiati
sono:
• I canali del calcio
• I canali del potassio tra cui quello presente nelle cellule pace-maker cardiache



I secondi messaggeri FOSFOINOSITIDI (PI)
Il Fosfatidil inositolo- 4,5- bifosfato (PIP2) è substrato dell’enzima di membrana Fosfolipasi C (PLC)
La reazione origina:
1. Diaclicilglicerolo (DAG)
2. Inositolo 1,4,5-trifosfato (IP3)

IP3 è un secondo messaggero solubile che ha come recettore specifico un canale del Ca attivato da ligando e localizzato nella membrana del REL. Controlla il rilascio di Ca2+ dai siti di accumulo intracellulare

Il ciclo dei polifosfoinositidi e le fosfolipasi C
da Balla, J. Endocrinol. 188, 135, 2006

DAG ha come azione principale l’attivazionedella Protein-Chinasi C (PKC) PKC a sua volta fosforila varie proteine intracellulari a livello serina/treonina analogamente a PKA Contribuisce in particolare alla regolazione di: •processi di secrezione ghiandolare •attivazione delle piastrine e dei neutrofili •regolazione dell’espressione genica •crescita cellulare •metabolismo

L’Acido Arachidonico (AA)
AA è un acido grasso che in forma esterificata è localizzato nei fosfolipidi di membrana
È precursore della formazione di acidi grassi che in modo autocrino o paracrino modulano un ampia varietà di tipi cellulari.
È un mediatore di risposte cellulari in risposta a:
• stimoli fisiologici a(istamina, bradichinina, angiotensina II),
• fisici (es. ischemia) e
•farmacologici (es, sostanze ionofore del Calcio)

Molte sostanze generate dal metabolismo dell’AA tra cui Eicosanoidi endogeni come prostaglandine, trombossani, esercitano la loro azione attivando specifici GPCRs

Acidi grassi essenziali
Acidi grassi esterificati (fosfolipidi)
Fosfolipasi A2
Acido arachidonico
Ciclossigenasi Lipossigenasi Fans
Endoperossidi ciclici
Eicosanoidi
Stimolo patologico
Lipocortina
Prostaglandine
-Dolore
-Febbre
-Infiammazione
-Acido gastrico
-Mucosa gastrica
-Doglie
-Reni: escrezione di
Na+ e H2O
Prostaciclina
-Vasodilatazione
-Inibizione della aggregazione piastrinica dei trombociti
Trombossani
-Vasocostrizione
-Aumento della aggregazione dei trombociti
Leucotrieni
-Reazioni allergiche (per es. asma bronchiale), leucotassi


DESENSITIZZAZIONE E TOLLERANZA NEI RECETTORI ACCOPPIATI A PROTEINE G
•La desensitizzazione non è intrinseca •Segue un trattamento cronico con famaci agonisti •I meccanismi alla base sono: 1. processi di fosforilazione su residui di
serina e treonina 2. internalizzazione del complesso legato

Tempo
Ris
po
sta
AM
Pc
Agonista
IL FENOMENO DI DESENSITIZZAZIONE SI OSSERVA ad.es esponendo una linea cellulare che esprime recettori beta adrenergici ad un agonista beta-adrenergico
In queste condizioni se si misura l’incremento di AMPc nel tempo si osserva quanto segue:


La desensitizzazione del recettore adrenergico si attua mediante tutti i tre
meccanismi generali di desensitizzazione:
• Perdita di affinità per l’agonista
• Riduzione della capacità di attivare la proteina G
• Riduzione del numero(downregulation)
• I tre eventi hanno caratteristiche cinetiche diverse ma sono tutti dipendenti da fosforilazione del recettore