Embed Size (px)
Citation preview

CITOCINAS Antonio Alonso, octubre 2003
A) Introducción
Las citocinas son proteínas producidas por células de la inmunidad innata y adquirida que median muchas de las acciones de estas células.
En líneas generales las citocinas liberadas en la fase de activación de la respuesta inmune activan el crecimiento y diferenciación de los linfocitos mientras que en la fase efectora activan a las células efectoras para eliminar a los microbios u otros antígenos.
El nombre citocinas es un término genérico. Las citocinas producidas por los fagocitos mononucleares se han lla-mado clásicamente monocinas y las producidas por los linfocitos linfocinas. No obstante el desarrollo de técnicas moléculas ha permitido observar que una misma citocina puede ser sintetizada por linfocitos, monocitos y otros ti-pos de células. Por este motivo el termino citocina parece el más apropiado.
Dado que muchas citocinas son sintetizadas por leucoci-tos y actúan sobre otros leucocitos, se les llama interleuci-nas. El término es incorrecto porque algunas citocinas que son producidas por leucocitos y actúan sobre leucocitos, por razones históricas no se les llama interleucinas. A la inversa, muchas citocinas llamadas interleucinas son pro-ducidas por células no leucocitarias y actúan sobre células no leucocitarias.
A pesar de estas incongruencias el término interleucina se ha generalizado porque ha medida que se han ido carac-terizando se les ha ido dando nombres dentro de una no-menclatura standard (IL-1, IL-2, etc.).
Las distintas interleucinas han recibido en el pasado nom-bres muy diversos pero también muy útiles porque en el propio nombre iba descrito su origen o su función. Por ejemplo, la IL-2 (nombre que per se no dice nada) antes se llamaba factor de crecimiento de los linfocitos T (nom-bre que define de manera muy precisa su función más importante).
B) Propiedades generales comunes a todas las cito-cinas
1. La secreción de citocinas es un fenómeno lento, breve y autolimitado. La mayoría de las citocinas no se en-cuentran almacenadas en la célula, por lo tanto su sín-tesis requiere transcripción de su material genético y traducción del RNAm. A veces requiere además pro-cesamiento del RNA y transformaciones postraduc-ción de un péptido precursor antes de obtener la proteína activa. Esto explica la lentitud en su libe-ración. Además el RNAm es inestable lo que explica la brevedad. Las citocinas sintetizadas son in-mediatamente secretadas, aunque hay algunas excepciones.
Índice A) Introducción 1 B) Propiedades generales comunes a
todas las citocinas 1
C) Clasificación 2 D) Receptores de citocinas 3 E) Señalización vía JAKISTAT 4 F) Citocinas de la Inmunidad Innata 5
1. TNF 5 2. lnteleucina 1 8 3. Quimiocinas 9 4. lnterleucina 12 115. lnterferones tipo 1 126. lnterleucina 10 137. lnterleucina 6 148. lnterleucina 15 149. lnterleucina 18 15
G) Papel de las citocinas en la respues-ta innata y en la inflamación 15
H) Citocinas de la inmunidad adaptativa 161. Interleucina 2 172. lnterleucina 4 193. lnterleucina 5 204. lnterferón y 205. Factor β transformante del cre-
cimiento 22
6. Linfotoxina 227. lnterleucina 13 238. Otras citoc. (IL-16, IL-17, MIF) 23
I) Papel de las citocinas en la Inmuni-dad Adaptativa 23
1. Propiedades de las subpobla-ciones T CD4+ 23
2. Desarrollo de Th1 y Th2 243. Funciones efectoras de Th1 y
Th2 25
J) Citocinas de la Hematopoyesis 281. Factor de la Stem Cell 282. lnterleucina 7 293. Interleucina 3 294. GM-SCF 305. M-CSF 306. G-CSF 307. lnterleucina 9 308. lnterleucina 11 30

Citocinas Página
2
2. Las acciones de la citocinas son a menudo pleiotrópicas y redundantes. Si una citocina actúa sobre diversos tipos celulares se dice que es pleiotrópica. Esta propiedad le permite a las citocinas modu-lar varios efectos biológicos al mismo tiempo. Si varias citocinas tienen el mismo efecto biológico se dice que son redundantes.
3. Algunas citocinas influencian la síntesis y las acciones de otras citocinas. Por esto la liberación de una citocina puede disparar reacción en cadena con liberación de otras citocinas. Las citocinas libe-radas en la cascada pueden ser agonistas o antagonistas de la primera.
4. La acción de las citocinas puede realizarse localmente o a distancia. Cuando actúan localmente sobre la misma célula que la produce se dice que su efecto es autocrino, si actúan sobre células ve-cinas su efecto se llama paracrino y si pasa a la circulación y se dispersan para actuar a distancia su efecto es endocrino.
5. Las citocinas, como las hormonas polipeptídicas, inician su acción uniéndose a receptores específi-cos de la superficie de su célula diana. Esta unión se realiza con gran afinidad (Kd: 10-10-10-12 M) por lo que son necesarias cantidades muy pequeñas de citocinas para desencadenar el efecto biológi-co. La concentración de receptores de citocinas en las células diana es baja: 100-1000 receptores / célula.
6. La expresión de receptores de citocinas está regulada por señales externas a la célula. Estas seña-les determinan por tanto el nivel de respuesta de las células a las citocinas. Por ejemplo, cuando los linfocitos T reconocen a su antígeno específico expresan el receptor para IL-2. Esto focaliza la ac-ción de la IL-2 sobre los linfocitos estimulados por antígenos quedando los no estimulados insensi-bles a la citocina. De esta forma una citocina que no es antígeno específica queda condicionada por la especificidad del receptor del linfocito T por su antígeno.
7. En la mayoría de los casos la acción de una citocina sobre una célula conduce a trascripción y tra-ducción y esto conlleva la adquisición de nuevas funciones y en algunos casos a la proliferación de dicha célula. Existen tres excepciones a esta regla: las quimiocinas, el TNF y la IL-15, que actúan de forma muy rápida por encontrarse presintetizadas y almacenadas en la célula.
C) Clasificación. No existe ninguna clasificación buena de las citocinas. Lo menos malo es hablar de categorías funciona-les. Las citocinas se pueden agrupar en tres categorías:
- de la inmunidad innata - de la inmunidad adquirida - de la hematopoyesis
En general las citocinas de la inmunidad innata y la adquirida son producidas por células distintas y ac-túan sobre poblaciones celulares diferentes (tabla 1). Tabla 1 - Algunas propiedades que distinguen a las citocinas de la inmunidad innata y adquirida Inmunidad innata Inmunidad adquirida Ejemplo de citocinas TNF-α, IL-1, IL-12, IFN-γ IL-2, IL-4, IL-5, IFN-γ Producidas fundamentalmente por
Macrófagos, NK Linocitos T
Tras el estimulo de LPS, peptidoglicanos bacterianos, RNA viral, citocinas producidas por linf T (IFN-γ)
Antígenos proteicos
Función biológica principal Mediadores de la Inmunidad innata y de la inflamación (local y sistémica)
Mediadores de la inmunidad adquirida: regulación del crecimiento y diferencia-ción de los linfocitos y activación de células efectoras como macrófagos, eosinófilos y cél. cebadas.
Cantidad producida Puede ser alta, detectable en suero Generalmente baja, no detectable en suero.
Citocinas Página
3
Efectos Sistémicos y locales Habitualmente solo locales
Participan en enfermedades Sistémicas, como el shock séptico Lesión tisular local, como la inflamación granulomatosa.
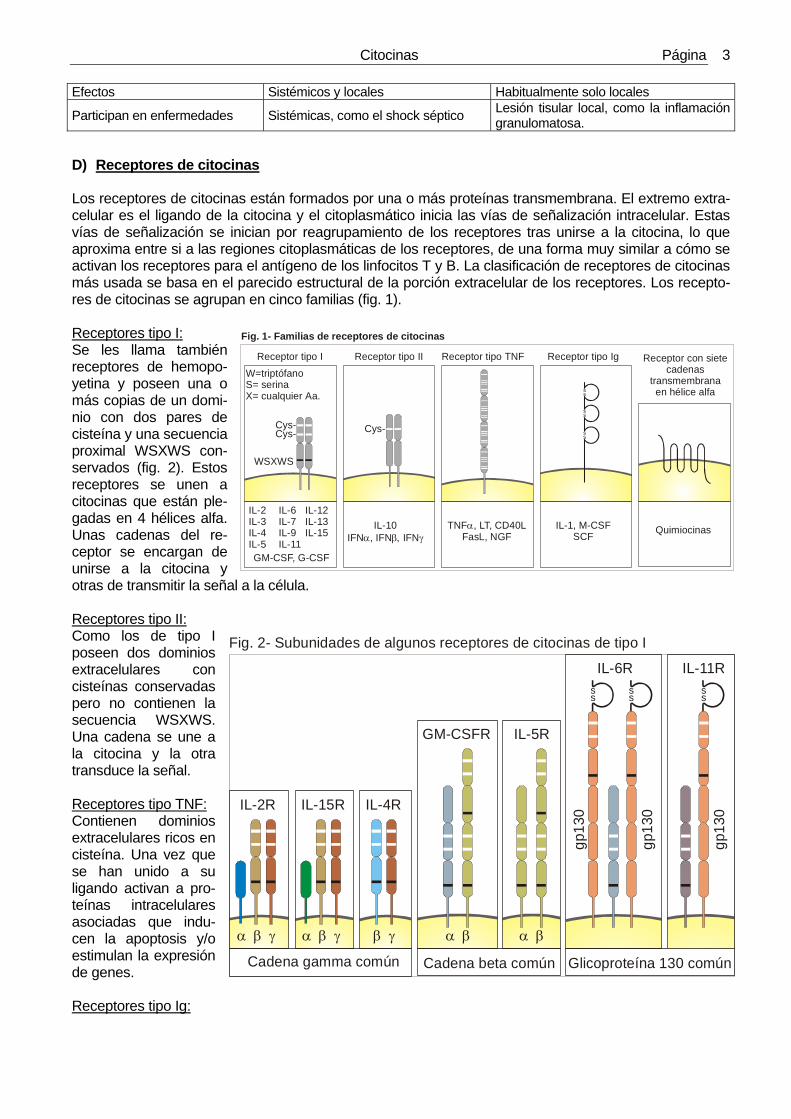
D) Receptores de citocinas Los receptores de citocinas están formados por una o más proteínas transmembrana. El extremo extra-celular es el ligando de la citocina y el citoplasmático inicia las vías de señalización intracelular. Estas vías de señalización se inician por reagrupamiento de los receptores tras unirse a la citocina, lo que aproxima entre si a las regiones citoplasmáticas de los receptores, de una forma muy similar a cómo se activan los receptores para el antígeno de los linfocitos T y B. La clasificación de receptores de citocinas más usada se basa en el parecido estructural de la porción extracelular de los receptores. Los recepto-res de citocinas se agrupan en cinco familias (fig. 1). Receptores tipo I: Se les llama también receptores de hemopo-yetina y poseen una o más copias de un domi-nio con dos pares de cisteína y una secuencia proximal WSXWS con-servados (fig. 2). Estos receptores se unen a citocinas que están ple-gadas en 4 hélices alfa. Unas cadenas del re-ceptor se encargan de unirse a la citocina y otras de transmitir la señal a la célula. Receptores tipo II: Como los de tipo I poseen dos dominios extracelulares con cisteínas conservadas pero no contienen la secuencia WSXWS. Una cadena se une a la citocina y la otra transduce la señal. Receptores tipo TNF: Contienen dominios extracelulares ricos en cisteína. Una vez que se han unido a su ligando activan a pro-teínas intracelulares asociadas que indu-cen la apoptosis y/o estimulan la expresión de genes. Receptores tipo Ig:
ss
ss
Fig. 2- Subunidades de algunos receptores de citocinas de tipo I
IL-2R
α β γ
IL-15R
α β γ
IL-4R
β γ
Cadena gamma común
GM-CSFR
α β α β
IL-5R
Cadena beta común
IL-6R
Glicoproteína 130 común
gp13
0
gp13
0
ss
IL-11R
gp13
0
Cys-Cys-
WSXWS
Receptor tipo I
Cys-
Receptor tipo II Receptor tipo TNF
ss
ss
ss
Receptor tipo Ig Receptor con siete cadenas
transmembranaen hélice alfa
IL-2IL-3IL-4IL-5
IL-6IL-7IL-9IL-11
IL-12 IL-13IL-15
IL-10IFN , IFN IFNα β, γ
GM-CSF, G-CSF
TNF , LT, CD40LFasL, NGFα IL-1, M-CSF
SCF Quimiocinas
W=triptófanoS= serinaX= cualquier Aa.
Fig. 1- Familias de receptores de citocinas

Citocinas Página
4
Reciben este nombre por poseer dominios extracelulares tipo Ig. El modo de señalización es distinto dependiendo del receptor. Receptores con siete cadenas transmembrana en hélice α: También se les llama “serpenteantes” porque entran y salen de la célula serpenteando. El ge-noma humano codifica muchos receptores de este tipo relacionados con la recepción de estí-mulos ambientales. Los relacionados con el sistema inmune son estimulados por las quimioci-nas. La clasificación de receptores basada en sus dominios extracelulares es útil para entender la unión de sus ligandos pero no dice nada de cómo las citocinas activan a sus células diana, que depende, obvia-mente de los dominios intracelulares de los receptores. Los receptores de distintas citocinas activan vías metabólicas intracelulares diferentes (tabla 2). Tabla 2 – Vías de activación, receptores y mecanismos de señalización de las citocinas Vía Receptores que usan esta vía Mecanismo de señalización JAK/STAT Receptores tipo I y tipo II Fosforilación mediada por JAK y activación de los
factores de transcripción STAT TRAFs Familia de receptores del TNF: TNF-RII,
CD40 Unión de proteínas adaptadoras, activación de factores de transcripción.
“Dominios de muer-te”
Familia de receptores del TNF: TNF-RI, Fas
Unión de proteínas adaptadoras, activación de caspasas.
A través de recep-tor “de peaje”
IL-1R, IL-18R (homólogo al la proteína toll de la drosphila)
Unión de proteínas adaptadoras, activación de factores de transcripción.
Tirosin kinasas M-CSFR, SCFR Actividad tirosin kinasa intrínseca del receptor Proteína G Receptores de quimiocinas JAK: Janus kinase (Kinasa Jano). STAT:Signal transducer and activator of transcription (Transductor de señal y activador de trascripción) TRAFs: TNF Receptor-Associated Factors (Factores asociados al receptor de TNF) M-CSFR: Monocyte-Colony Stimulatín Factor Receptor (Receptor de Factor Estimulante de Colonias de Mono-citos) SCFR: Stem cell Factor Receptor (Receptor del Factor de la Stem Cell) E) E) Señalización de citocinas vía Kinasas Jano (JAK) y Transductores de Señales y Activado-
res de Trascripción (STAT): Uno de los mecanismos mejor entendidos por los que las citocinas trasducen señales para inducir re puestas en las células diana son aquellos en los que participan las enzimas llamadas kinasas Jano factores de trascripción llamados transductores de señales y activadores de trascripción. Las vías de señalización JAK/STAT son usadas por todos los receptores de citocinas tipos I y Il. El estudio de estas vías ha revelado la relación directa que existe entre la unión de la citocina a su receptor y la activación trascripcional de los genes implicados. El descubrimiento de las vías JAK/STAT se hizo por análisis genético y bioquímico de la vía de seña-lización de los interferones. Las regiones promotores de los genes sensibles a interferones contiene secuencias que ligan a proteínas celulares que son fosforiladas cuando la célula se trata con interfe-rón. Se observó que estas proteínas activan la trascripción de genes sensibles a la citocina y fueron llamadas transductoras de señales y activadoras de la trascripción. Se crearon líneas celulares mu-tantes insensibles a los interferones y se observó que la causa de esta insensibilidad era la ausencia de algunas de estas proteínas. Además, cuando se trasfectaron los genes de estas proteínas se restableció la sensibilidad de las líneas a los interferones. Esto estableció el papel fundamental de estas proteínas en la respuesta a estas citocinas. En otras líneas celulares mutantes se encontró que eran deficitarias en una o más tirosinkinasas. Estas kinasas poseen dos dominios de kinasa por lo que se les llamó kinas Jano, en recuerdo del dios romano con dos caras. De estos dos dominios
Citocinas Página
5
de kinasa sólo uno tiene actividad enzimática. Estudios posteriores entre los que se encuentra el análisis de ratones knockout para JAK y/o STAT han puesto de manifiesto que estas vías de señali-zación JAK/STAT son usadas por muchas citocinas y hormonas.
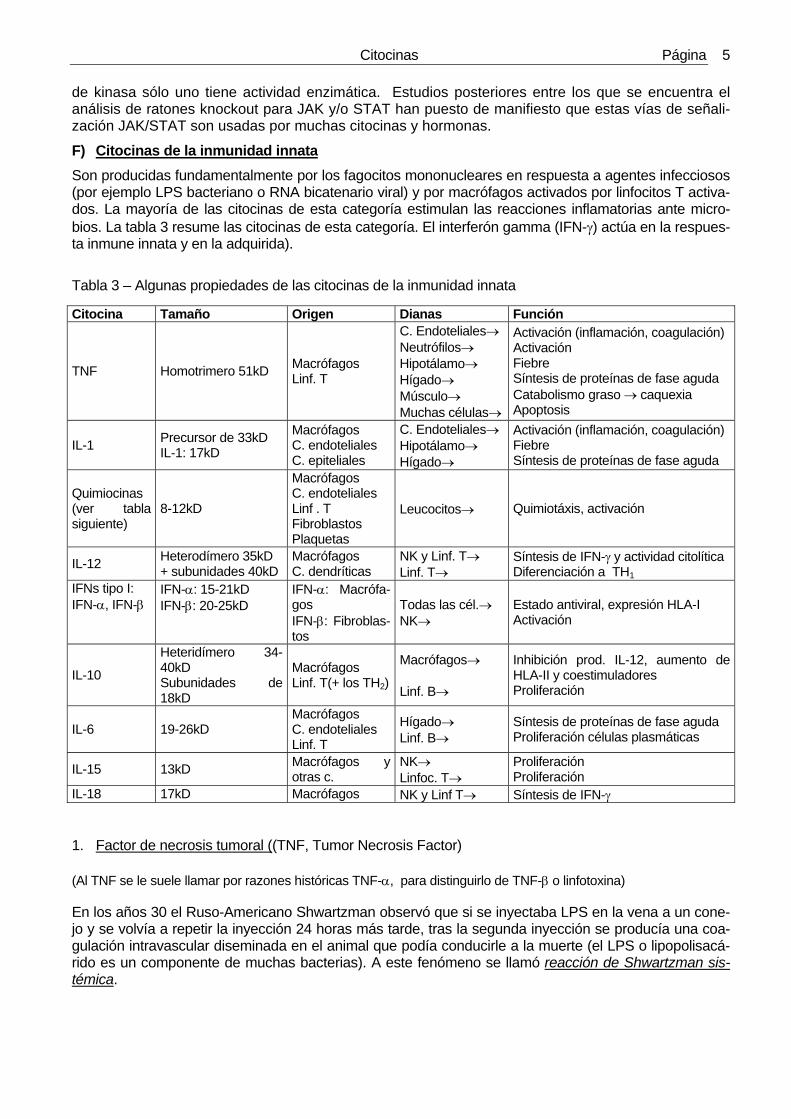
F) Citocinas de la inmunidad innata Son producidas fundamentalmente por los fagocitos mononucleares en respuesta a agentes infecciosos (por ejemplo LPS bacteriano o RNA bicatenario viral) y por macrófagos activados por linfocitos T activa-dos. La mayoría de las citocinas de esta categoría estimulan las reacciones inflamatorias ante micro-bios. La tabla 3 resume las citocinas de esta categoría. El interferón gamma (IFN-γ) actúa en la respues-ta inmune innata y en la adquirida).
Tabla 3 – Algunas propiedades de las citocinas de la inmunidad innata
Citocina Tamaño Origen Dianas Función
TNF Homotrimero 51kD Macrófagos Linf. T
C. Endoteliales→ Neutrófilos→ Hipotálamo→ Hígado→ Músculo→ Muchas células→
Activación (inflamación, coagulación) Activación Fiebre Síntesis de proteínas de fase aguda Catabolismo graso → caquexia Apoptosis
IL-1 Precursor de 33kD IL-1: 17kD
Macrófagos C. endoteliales C. epiteliales
C. Endoteliales→ Hipotálamo→ Hígado→
Activación (inflamación, coagulación) Fiebre Síntesis de proteínas de fase aguda
Quimiocinas (ver tabla siguiente)
8-12kD
Macrófagos C. endoteliales Linf . T Fibroblastos Plaquetas
Leucocitos→ Quimiotáxis, activación
IL-12 Heterodímero 35kD + subunidades 40kD
Macrófagos C. dendríticas
NK y Linf. T→ Linf. T→
Síntesis de IFN-γ y actividad citolítica Diferenciación a TH1
IFNs tipo I: IFN-α, IFN-β
IFN-α: 15-21kD IFN-β: 20-25kD
IFN-α: Macrófa-gos IFN-β: Fibroblas-tos
Todas las cél.→ NK→
Estado antiviral, expresión HLA-I Activación
IL-10
Heteridímero 34-40kD Subunidades de 18kD
Macrófagos Linf. T(+ los TH2)
Macrófagos→ Linf. B→
Inhibición prod. IL-12, aumento de HLA-II y coestimuladores Proliferación
IL-6 19-26kD Macrófagos C. endoteliales Linf. T
Hígado→ Linf. B→
Síntesis de proteínas de fase aguda Proliferación células plasmáticas
IL-15 13kD Macrófagos y otras c.
NK→ Linfoc. T→
Proliferación Proliferación
IL-18 17kD Macrófagos NK y Linf T→ Síntesis de IFN-γ 1. Factor de necrosis tumoral ((TNF, Tumor Necrosis Factor) (Al TNF se le suele llamar por razones históricas TNF-α, para distinguirlo de TNF-β o linfotoxina) En los años 30 el Ruso-Americano Shwartzman observó que si se inyectaba LPS en la vena a un cone-jo y se volvía a repetir la inyección 24 horas más tarde, tras la segunda inyección se producía una coa-gulación intravascular diseminada en el animal que podía conducirle a la muerte (el LPS o lipopolisacá-rido es un componente de muchas bacterias). A este fenómeno se llamó reacción de Shwartzman sis-témica.

Citocinas Página
6
Cuando tras la primera inyección en vena (o en piel) se realizaba otra en la piel se producía una reac-ción local que terminaba necrosando la zona de la segunda inyección. A esta se llamó reacción de Shwartzman local.
Las investigaciones de Shwartzman fueron publicadas en 1937 (Phenomenon of local tissue reactivity and its immunological and clinical significance). Murió en 1965 sin saber que había descu-bierto el TNF. Cuando se extrae sangre de un animal que ha sufrido una reacción de Shwartzman sistémica o local y el suero se inyec-ta en el interior de un tumor se produce una necrosis localiza-da del tumor en la zona de la inyección. Al “factor” del suero causante de este fenómeno se le llamó en los años 70 Factor de Necrosis Tumoral. Hoy el TNF existe puro y cuando se inyecta de forma apro-piada a animales reproduce maravillosamente bien las reac-ciones de Shwartzman sistémica y local y la necrosis de tu-mores. Además, los anticuerpos anti-TNF bloquean la apari-ción de estos fenómenos. El TNF es el mediador mas importante de muchas infeccio-nes bacterianas agudas. Es producido fundamentalmente por los macrófagos, linf. T, los linf. B, las células NK y las células cebadas. El estímulo más importante para que estas células produzcan TNF es el LPS, una endotoxina de las bacterias gram negativas. El TNF es una proteína formada por tres péptidos idénticos
(homotrímero) que adoptan la estructura de una pirámide triangular. Cada lado de la pirámide está for-mada por una subunidad de TNF. Los lugares de unión al receptor se encuentran en la base de la pirá-mide lo que permite que el TNF pueda unirse al mismo tiempo a varios receptores. Para este homotrímero existen dos tipos de receptores. Los dos son estructuras transmembrana que están presentes en casi todas las células. Cuando el TNF se acopla al receptor en el exterior de la célu-la, se transmiten señales al interior que ponen en marcha una serie de acontecimientos. Estos aconte-cimientos son distintos para los dos tipos de receptores de TNF existentes (fig. 4). El receptor I de TNF (TNF-RI o p55) posee en el citoplasma un segmento llamado dominio de muerte o death domain. Tras la activación de este receptor por el TNF se unen al dominio de muerte una proteína adaptadora llamada TRADD (TNF Receptor-Associated Death Domain), otra llamada FADD (Fas Asso-ciated Death Domain) y el precursor de la caspasa 8. De esta forma queda activada la caspasa 8 y ocu-rre una serie de eventos que conducen a la muerte programada de la célula o apoptosis. El receptor II (TNF-RII o p75) posee igualmente el dominio de muerte y se une a TRADD pero recluta a otras dos proteínas intermediarias de la señalización llamadas TRAF (TNF Receptor-Associated Factor) y RIP (Receptor Interacting Protein). TRAF activa la vía de las kinasas SAP y RIP la vía de NFκB que conducen a la generación de los factores de transcripción AP-1 y NFκB. Para el papel de AP-1 y NFκB ver la figura de acontecimientos intracelulares en la activación de los linfocitos T, en las trasparencias generales de la asignatura).
LPS(i.v.)
LPS(i.v.)
24Horas
LPS(piel o i.v.)
24Horas
LPS(piel)
Sistémica Local
Fig. 3. Las reacciones de Shwartzman

Citocinas Página
7
Funciones biológicas del TNF A) En condiciones normales las
funciones del TNF quedan res-tringidas a la zona en la que se lleva a cabo la infección. Estas circunstancias el TNF realiza sus funciones a nivel local. Es-tas funciones son:
1. Aumenta el reclutamiento de
neutrófilos y monocitos a la zona de infección y activa a estas células para erradicar a los gérmenes. Esto lo lleva a cabo:
a) aumentando la expresión
de moléculas de adhesión en el endotelio vascular próximo al foco de infec-ción, con lo que se facilita el “homing” y
b) haciendo que las células endoteliales y los macrófagos liberen quimiocinas que atraerán más
leucocitos al foco de infección (este fenómeno de atracción química se denomina quimiotáxis). 2. Induce la apoptosis de algunos tipos celulares (la significación fisiológica de este hecho es descono-
cida). 3. Induce a los fagocitos mononucleares a secretar IL-1, que funciona de forma parecida al TNF. Existen circunstancias en las cuales las funciones locales del TNF pueden causar efectos indeseables, probablemente debido a una producción local mantenida. Este es el caso de las enfermedades autoin-munes. Actualmente se está intentando minimizar estos efectos adversos neutralizando el TNF con an-ticuerpos monoclonales anti-TNF o con receptores de TNF solubles. B) En las infecciones severas se produce el TNF en tal cantidad que difunde desde los focos de infec-
ción a la sangre y causa trastornos sistémicos: 1. Aumenta la liberación de prostaglandinas por las células hipotálamicas lo que produce fiebre (por
esto al TNF se llama pirógeno endógeno). La aspirina bloquea la síntesis de prostaglandinas y dis-minuye la fiebre.
2. Aumenta la síntesis en el hígado de dos proteínas séricas: amiloide a y fibrinógeno. 3. Disminuye el apetito e inhibe la síntesis de lipoproteinlipasa, llegando a ocasionar una delgadez ex-
trema llamada caquexia. La lipoproteinlipasa libera los ácidos grasos de las lipoproteinas del suero con lo que dichos ácidos quedan disponibles para pasar a las células para su utilización.
4. Disminuye la contractilidad muscular general (→ flacidez) y cardiaca (→ disminución de la presión arterial).
5. Aumenta la expresión en las células endoteliales de “factor tisular” (un potente activador de la coa-gulación) e inhibe la expresión de trombomodudina, un inhibidor de la coagulación . Estas dos ac-ciones conducen a la aparición de trombosis vasculares. La necrosis que produce el TNF en los tu-mores es en gran medida debida a su trombosis vascular.
6. El TNF causas trastornos metabólicos severos tales como el descenso de la glucemia hasta niveles incompatibles con la vida. Esto es debido a sobreutilización por el músculo y fallo del hígado para reemplazarla.
TNF
TNF-
RI
Death domain TR
AD
D
TRA
DD
FADD Cas
pasa
8
Apoptosis
TNFTN
F-R
II
Death domain TR
ADD
TRAD
D
TRAF RIPSAP
AP-1NF Bκ
Fig. 4. Los dos tipos de receptores del TNF.

Citocinas Página
8
Una complicación severa de la infección masiva con bacterias gram negativas es el shock séptico o shock endotóxico, una situación habitualmente mortal. Los anticuerpos monoclonales y los receptores solubles de TNF son capaces de revertir este shock séptico en animales de experimentación pero, por razones desconocidas, son ineficaces en los humanos. 2. Interleucina 1
La función principal de la IL-1 (como la del TNF) es de mediadora de la respuesta inflamatoria en las infecciones y otros estímulos inflamatorios.
La IL-1 es producida por los fagocitos mononucleares activados por productos bacterianos como el LPS u otras citocinas, como el TNF (un ejemplo de acción de citocinas en cascada). Al contrario que el TNF, la IL-1 es también producida por neutrófilos, células epiteliales como los queratinocitos y células endote-liales.
Hay dos formas de IL-1, la IL-1α y la IL-1β. La homología estructural entre ambas es menor del 30% pero se unen al mismo receptor de membrana y poseen las mismas funciones. Ambas formas se sinte-tizan como un péptido precursor de 33kD y se secretan como proteínas maduras de 17kD. El precursor de la IL-1β madura a proteína activa por la acción de una proteasa llamada enzima convertidora de IL-1β o ICE (IL-1 converting enzyme). ICE fue el primer miembro de una familia de cistin proteasas llama-das caspasas que se descubrió en mamíferos. Muchos miembros de esta familia están involucrados en muchas células en la muerte por apoptosis.
Al contrario de lo que ocurre con la mayoría de las proteínas secretadas, ni la IL-1α ni la IL-1β poseen una secuencia hidrofóbica en su péptido precursor para guiarlas al retículo endoplásmico. Se descono-ce cómo son secretadas estas proteínas. La mayor parte de la IL-1 encontrada en la circulación es IL-1β.
Se han descrito dos receptores para IL-1 y ambos son miembros de la superfamilia de Igs. El tipo I se encuentra en la mayoría de los tipos de células y es el que media la mayoría de las acciones mediadas por la IL-1. Está asociado con otra proteína llamada proteína accesoria del receptor de IL-1, que es homóloga al propio receptor de IL-1. El receptor tipo II se expresa en linf. B pero puede ser inducido en otras células. No responde a la IL-1 por lo que su función es “atrapar” para inhibir competitivamente la unión de la IL-1 al receptor tipo I. La región citoplasmática del receptor tipo I es homóloga al receptor Toll (peaje) de la drosophila, que está relacionado en la defensa contra la infección. Hasta el momento se han identificado en los mamíferos 6 moléculas homólogas a Toll y todas ellas parecen estar relacio-nadas con la defensa frente a distintos tipos de infecciones microbianas. La unión de la IL-1 al receptor tipo I conduce a la activación de los factores de transcripción NF-kB y AP-1 mediante una vía en la que parece participar un miembro de la familia TRAF (ver activación vía TNF).
Función biológica de la IL-1. Es similar a la del TNF y depende de la cantidad de citocina produci-da. • Cuando se secreta a bajas concentraciones funciona como un mediador de la inflamación loca. Ac-
túa sobre las células endoteliales para aumentar la expresión de moléculas que aumentan la ad-hesión leucocitaria tales como los ligandos para las integrinas.
• Secretada en cantidades mayores penetra en el torrente circulatorio y posee acciones endocrinas. Como el TNF, produce fiebre, induce la síntesis de proteínas de fase aguda por el hígado e inicia el desgasto metabólico que conduce a la caquexia.
El parecido en las acciones del TNF y la IL-1 parece extraño porque tanto ambas citocinas como sus receptores son muy distintos estructuralmente. La probable explicación lógica a este fenómeno del pa-recido de sus efectos biológicos es que los receptores de ambas citocinas activan proteínas homólogas y activan a los mismos factores de transcripción. No obstante, existen varias diferencias entre TNF e IL-1. Por ejemplo, la IL-1 no induce apoptosis e incluso a altas concentraciones no causa shock séptico.
Los fagocitos mononucleares producen un inhibidor natural de la IL-1. Se trata de una proteína estructu-ralmente homóloga que se une al mismo receptor que la IL-1 y por tanto compite con ella, por lo que se

Citocinas Página
9
llama antagonista del receptor de la IL-1 (IL-1ra, IL-1 receptor antagonist). Este antagonista se ha usado en ensayos clínicos para mejorar el shock séptico y la artritis reumatoide. No ha dado resultado, proba-blemente debido a la redundancia de las acciones de la IL-1 y el TNF. 3. Quimiocinas.
Las quimiocinas forman una larga familia de citocinas estructuralmente homólogas que estimulan el movimiento leucocitario y regulan su migración desde la sangre a los tejidos.
Estructura, producción y receptores de quimiocinas:
Todas las quimiocinas son polipéptidos de 8-12 kD que poseen dos puentes disulfuro. Hasta el momen-to se han identificado unas 50 quimiocinas y probablemente existan más. Existen dos familias de qui-miocinas. Las que poseen contiguas las dos cisteínas que forman el puente disulfuro se llaman quimio-cinas CC y aquellas que poseen las dos cisteínas separadas por otro aminoácido se llaman quimiocinas CXC. Estas diferencias se correlacionan con sus genes, que se organizan en dos grupos genéticos se-parados. En la inflamación, las citocinas CC actúan sobre monocitos, linfocitos y eosinófilos mientras que las CXC actúan sobre todo sobre neutrófilos. Como excepciones, existe una quimiocina con una sola cisteína y otra con dos cisteínas separadas por tres aminoácidos (CX3C).
Las quimiocinas, tanto CC como CXC, son producidas por leucocitos y por varios tipos de células tisula-res, como endoteliales, epiteliales y fibroblastos. En muchas de estas células la secreción de quimioci-nas es inducida por microbios y por citocinas inflamatorias como el TNF y la IL-1. Algunas quimiocinas CC son producidas por linf. T activados actuando de puente entre la inmunidad adaptativa y la innata. Algunas citocinas se producen de forma constitutiva, sin ningún estímulo inflamatorio, en los órganos linfoides, por lo que están relacionadas con el trafico fisiológico de linfocitos entre los órganos linfoides. Las quimiocinas de ambas familias se une en la superficie de la célula endotelial a proteoglicanos de heparán sulfato y de esta forma quedan expuestas a los leucocitos circulantes. Los leucocitos circulan-tes, al contacto con las quimiocinas, se unen al endotelio por medio de las moléculas de adhesión y aumentan su motilidad para abandonar la sangre.
Hasta el momento se han descrito 10 receptores para quimiocinas CC (CC-R1 a CC-R10) y 6 recepto-res para quimiocinas CXC (CX-R1 a CX-R6) (tabla 4).
Tabla 4 - Receptores de quimiocinas
Receptor Ligandos (algunos ejemplos) Células que lo expresan Función principal CXC-R1 IL-8 Neutrófilos Inflamación aguda CXC-R2 IL-8, Gro-α /β Neutrófilos Inflamación aguda CXC-R3 IP10, Mig (estimulado por IFN-γ Linf. T (TH1 > TH2) Migración T a sitios DTH CXC-R4 SDF-1 Linf. T, B, Monoc,, Neutróf. Algu-
nas células tisulares. Reclutamiento T. Co-receptor para HIV
CXC-R5 BLC/BCA-1 Linf T, B Migración B a folículos linf. CC-R1 MIP-1α. RANTES. MCP-2 y 3 Monoc., Basóf., Eosinóf., T Inflamación CC-R2 MCP-1, 2, 3 y 4 Monoc., Basóf., T Inflamación CC-R3 Eotaxin. MCP-2, 3 y 4.RANTES Eosinóf., Basóf., T (TH2 > TH1) Inflamación alérgica CC-R4 TARC T (TH2 > TH1) Tráfico linfocitario CC-R5 RANTES. MIP-1α. MIP-1β Monoc., T (TH1 > TH2) Inflamación. Co-receptor para
HIV CC-R6 LARC/MIP-3α T, B Tráfico linfocitario CC-R7 ELC Monoc., Cél, dendríticas, T naive Tráfico linfocitario c. Dendrít. (¿) CC-R8 I309 Monocitos, T Tráfico linfocitario

Citocinas Página
10
Solamente se dan algunas de las quimiocinas que se unen a cada receptor. Los nombres de las quimiocinas a menudo se basan en la forma en que fueron descubiertas. BLC: B lymphcyte chemokine → quimiocina de los linfocitos B ELC: Epstein-Barr Virus chemokine ligand → Ligando de la quimiocina del virus de Epstein Barr IP-10: Inducible protein 10 → proteína inducible 10. LARC: Liver and activation-regulated chemokine → Quimiocina del hígado y regulada por activación. MCP: Monocyte chemoattractan protein → proteína quimioatrayente de monocitos MIP: Macrophage inflammatory protein → Proteína inflamatoria del macrófago. RANTES: Regulated upon Activation Normal T cell Expressed and Secreted → Nomalmente expresada y secreta-
da por los linf. T y regulada tras activación. SDF-1: Stromal cell-derived factor-1 → Factor derivado de las células del estroma-1. TARC: Thymus and activation-regulated chemokine → Quimiocina del timo y regulada tras activación.
Los receptores de quimiocinas se expresan en los leucocitos alcanzando los linfocitos T las expresiones más altas. Dentro de cada familia los receptores poseen solapamiento de espe-cificidad. El patrón de expresión celular de estos receptores determina qué células responden y cuales no responden a determinadas quimiocinas. Todos los receptores de quimioci-nas tienen una estructura característica con 7 dominios transmembrana en hélice α (fig. 5).
Esta característica se puede ver también en otros receptores que están acoplados a proteínas G triméricas fijadoras de GTP. Cuando están ocupados por su ligando, estos recepto-res actúan como proteínas intercambiadoras de GTP, catali-zando el reemplazo del GDP unido por GTP. La forma aso-ciada a GTP de estas proteínas G pueden activar a varios enzimas celulares, incluidos algunos que pueden estimular el movimiento de las células. Los receptores de quimiocinas pueden dejar de expresarse rápidamente después de unirse a su ligando, lo que puede representar un mecanismo para terminar la respuesta.
Algunos receptores de quimiocinas (CC-R5 y CXC-R4) actúan como receptores para el VIH. Las citoci-nas apropiadas, liberadas por los linfocitos, compiten por los receptores de citocinas con el VIH.
Funciones biológicas de las quimiocinas:
Las quimiocinas se descubrieron gracias a sus propiedades como quimioatrayentes de los leucocitos pero hoy sabemos que poseen muchas funciones en el sistema inmune y en otros sistemas.
• Reclutan células de defensa hacia los lugares de infección. El reclutamiento de leucocitos es el re-sultado de varias acciones secuenciales de las quimiocinas sobre estas células. Las quimiocinas in-ducen rápidos cambios de forma en los leucocitos (in vitro, en segundos) resultando en la formación de lamelopodios. Estimulan la polimerización y despolimerización alternativas de los filamentos de actina que conducen al movimiento de los leucocitos y su migración hacia el gradiente químico de la citocina. Las quimiocinas también aumentan la afinidad de las integrinas leucocitarias por sus ligan-dos en el endotelio. Las distintas quimiocinas actúan sobre células diferentes, lo que condiciona el contenido del infiltrado inflamatorio. Por ejemplo, la IL-8 (una quimiocina CXC) recluta sobre todo neutrófilos mientras que la eotaxina (una quimiocina CC) recluta eosinófilos.
• Regulan el tráfico de linfocitos y otros leucocitos entre órganos linfoides periféricos. Algunos de los descubrimientos más intrigantes y sorprendentes sobre las quimiocinas han sido sus funciones en la migración normal de las células inmunes entre órganos linfoides. Se ha identificado una quimiocina que dirige la entrada de los linfocitos B en los folículos linfoides. El knockout del gen que codifica el receptor (CXC-R5) para esta quimiocina resulta en ausencia de linfocitos B en los folículos.
• Las quimiocinas están relacionadas con el desarrollo de varios órganos. Los ratones con ausencia del gen del receptor CXC-R4 tienen problemas de linfopoyesis y angiogénesis y defectos fatales en
Quimiocina
Receptor
Las regiones en hélice alfase muestran como cilindros
Fig. 5. Receptor con 7 cadenas en hélicealfa de las quimiocinas y otras citocinas.

Citocinas Página
11
el desarrollo del corazón y del cerebelo. Estas funciones de la quimiocinas no eran esperadas y abren la posibilidad para otras funciones en la morfogénesis aún no conocidas.
4. Interleucina-12
Es un mediador fundamental de la respuesta inmune innata temprana contra los microbios y es inductor de la inmunidad celulo-mediada (que es la respuesta inmune adaptativa que se pone en marcha contra estos microbios). La IL-12 fue identificada originalmente como un activador de la actividad citolítica de las células NK, pero su función más importante es la estimulación de la producción de IFN-γ por las cé-lulas NK y las T. Estructura, producción y receptores de IL-12:
La IL-12 es un heterodímero de 35kD (p35) y 40kD (p40), unidos por puentes disulfuro. La subunidad p35 tiene dos dominios globulares en hélice α. La subunidad p40 es homóloga al receptor para IL-6, conteniendo ambas un dominio tipo Ig y motivos característicos de los receptores de citocinas tipo I.
La fuente principal de IL-12 son los fagocitos mononucleares activados y las células dendríticas. La subunidad p35 parece ser sintetizada por muchas células pero solo “estas células presentadoras de antígenos profesionales” sintetizan la subunidad p40 y por lo tanto la IL-12 biológicamente activa.
Los estímulos que disparan la producción de IL-12 son: LPS, infección por bacterias intracelulares como listeria y micobacterias y la infección por virus. Además, los linfocitos T helper estimulados por su antí-geno inducen la producción de IL-12 en los macrófagos y células dendríticas fundamentalmente vía CD40L-CD40. El IFNγ producido por las células T y NK también estimula la producción de IL-12. Por lo tanto, la IL-12 es producida por las células presentadoras de antígeno profesionales cuanto éstas pre-sentan antígenos a las células T, durante las fases de inducción y efectora de las respuestas inmunes célulo-mediadas.
El receptor para IL-12 es un heterodímero compuesto de las subunidades β1 y β2. Las dos cadenas son necesarias para la unión de la IL con alta afinidad aunque sólo β2 está involucrada en la señaliza-ción. La señalización por este receptor se hace por la vía JAK/STAT. La IL-12 es la única que se conoce que active a STAT4. La expresión de la ca-dena β de señalización es au-mentada por citocinas (funda-mentalmente IFN-γ, cuya pro-ducción es estimulada por IL-12) y esto es un ejemplo de un cir-cuito positivo de amplificación de la respuesta inmune.
Funciones biológicas de la IL-12:
La IL-12 es crítica en la inicia-ción de una secuencia de res-puestas de los macrófagos, cé-lulas T y NK que resulta en la erradicación de los microbios intracelulares (fig. 6).
• La IL-12 estimula la produc-ción de IFN-γ por las células T y NK. Los macrófagos
Microbios
Macrófago infectado
Fragmentosde microbio
presentados por c. dendríticao macrófago CD4+
“inocente”
CD8+CD4+
TH1NK
El IFN- activa el killing de los macrófagos
γEl IFN- activa la
actividad citolítica de Tcit.y NK
γ
Producción de IL-12CD40
CD40L
TCR
Producción de IFN-γ
Célula infectada destruida por T citot. o NK
Fig. 6. Algunas de las funciones de la IL-12.

Citocinas Página
12
producen IL-12 en respuesta a muchos microbios y esta IL-12 hace que las células T y NK produz-can IFN-γ que activan a los macrófagos para destruir a los microbios fagocitados. En las sepsis se-veras gram negativas se producen grandes cantidades de IL-12, resultando en la producción de IFN-γ, que sinergiza con el LPS para la producción de TNF (por los macrófagos), el mediador más importante del shock séptico. Los antagonistas de la IL-12 impiden la muerte en modelos experi-mentales de shock séptico inducido por LPS.
• La IL-12 estimula la diferenciación de las células Th a Th1, productoras de IFN-γ, que estimula a los fagocitos. La IL-12 es también un factor de crecimiento de los linf. T.
• La IL-12 aumenta las funciones citolíticas de las células T citot. y NK.
Los ratones knockout sin el componente p40 de la IL-12 muestran deficiente producción de IFN-γ y de-sarrollo de linfocitos T hacia TH1. También poseen defectos funcionales de las células T. Se han descri-to algunos pacientes con mutaciones en el receptor de la IL-12, que resultan ser muy susceptibles a infecciones por salmonella, una micobacteria atípica.
La IL-12 es un importante puente de unión entre la inmunidad innata y la adquirida: producida en las etapas tempranas de la respuesta innata frente a microbios intracelulares va a estimular a la respuesta adaptativa contra estos microbios.
La IL-12 se está usando en ensayos clínicos para potenciar reacciones célulo-mediadas en pacientes con inmunodeficiencia o cáncer, como terapia sistémica y como un componente de vacunas. 5. Interferones tipo I (IFNα/IFNβ)
Los interferones tipo I son los mediadores de la respuesta inmune innata temprana frente a las infecciones virales. La palabra “interferón” procede de su capacidad para interferir con las in-fecciones virales. Estructura producción y receptores de los IFN tipo I (desde ahora IFN-I)
Existen dos tipos de IFN-I. El IFN-I α y el IFN-I β.
IFN-α IFN-β El IFN-α es en realidad una familia de polipéptidos es-tructuralmente relacionados, cada uno de ellos codifica-do por un gen distinto. La fuente más importante de IFN-α es una subpoblación de fagocitos mononucleares. Al IFN-α también se le llama interferón leucocitario.
El IFN-β es una sol proteína, producida por muchas células, entre ellas los fibroblastos, por lo que también se le llama interferón fibroblástico.
El estímulo más potente para la síntesis de IFN-I es la infección viral. En el laboratorio se puede inducir la síntesis de IFN-I mediante RNA de doble hebra sintético que puede mimetizar una señal producida durante la replicación viral. Las células T activadas también pueden estimular en los fagocitos mononu-cleares la síntesis de IFN-I.
Aunque IFN-α e IFN-β son estructuralmente bastante diferentes, se unen a los mismos receptores de membrana e inducen respuestas biológicas similares.
El receptor del IFN-I es un heterodímero de dos péptidos estructuralmente relacionados, un péptido se une al IFN-I y el otro transduce señales por la vía JAK/STAT. Ambos péptidos son miembros de la fami-lia de receptores de citocinas tipo II.
Acciones biológicas de los IFN-I

Citocinas Página
13
Las acciones de los IFN-I protegen a las células de las infecciones virales y estimulan la inmunidad celular contra microbios intracelula-res (fig. 7).
• Los IFN-I inhiben la replicación viral. Esto lo hacen gracias a que inducen la síntesis de en-zimas tales como la 2’, 5’ adeni-lato sintetasa que impiden la transcripción del RNA o DNA viral y por tanto la replicación vi-ral. La acción del IFN-I es de ti-po paracrino, es decir actúa so-bre células vecinas no infecta-das induciendo en ellas un es-tado antiviral. También es de ti-po autocrino ya que puede ac-tuar sobre la misma célula in-fectada que lo produce para impedir en ella la replicación vi-ral.
• Los IFN-I aumentan la expre-sión de moléculas HLA de cla-se I y por tanto facilitan el reco-nocimiento de las células infectadas por virus por los linfocitos T citot. específicos para ese virus.
• Los IFN-I también estimulan la producción de células TH1. Esto es debido a que hacen que las célu-las TH0 expresen receptores para IL-12 y esta es la citocina más importante en la derivación de es-tas células hacia TH1.
• Por último, los IFN-I inhiben la proliferación de muchos tipos de células. Esto es posible que sea debido a que inducen en estas células las mismas enzimas que bloquean la replicación viral pero también pueden estar involucradas otras enzimas que alteran el metabolismo de aminoácidos tales como el triptófano. No se conoce el alcance fisiológico de este freno a la proliferación celular.
En resumen, los IFN-I actúan contra las infecciones virales. Los animales knockout para el receptor de IFN-I son muy susceptibles a las infecciones virales. El IFN-α se está evaluando en el tratamiento de algunas formas de hepatitis virales y el IFN-β se usa en el tratamiento de la esclerosis múltiple. Se des-conoce el mecanismo de acción en esta última enfermedad.
6. Interleucina 10
Hasta ahora hemos cescrito citocinas que estimulan la inmunidad innata. La IL-10, por el con-trario, es un inhibidor de los macrófagos activados y por tanto es importante en la homeostasis de las reacciones inmunes innatas. Estructura, producción y receptores de la IL-10
La IL-10 posee 4 dominios globulares en hélice α y su receptor pertenece a la familia del tipo II. La IL-10 es producida fundamentalmente por los macrófago activados y dado que inhibe a los propios macrófagos es un buen ejemplo de regulación por feedback negativo. También la producen los linfocitos T y algunas células no linfoides, como los queratinocitos. No están claros los estímulos que inducen la secreción de IL-10. Acciones biológicas de la IL-10
Célula infectadapor virus
IFN-ICélula
no infectada
Síntesis de enzimas que impiden la replicación viral
El IFN-I induce en la célula un
“estado antiviral”
Receptorpara IFN-I
Célula infectada
HLA-I
El IFN-I aumenta la expresión de
HLA-I y por tanto la presentación de Ags. Virales
T CD8+
Los linf. T cititóxicos reconocen y destruyen a las células infectadas.
Fig. 7. Funciones del IFN tipo I.

Citocinas Página
14
Las acciones biológicas de la IL-10 son el resultado de su capacidad para inhibir muchas de las accio-nes de los macrófagos activados.
• Inhibe la producción de IL-12 y TNF por los macrófagos activados. No se conocen los mecanismos bioquímicos por los que lleva a cabo estas acciones. Dado que la IL-12 es un estímulo crítico para la secreción de IFN-γ y este es inductor de las reacciones inmunes innatas y célulo-mediadas, la IL-10 inhibe estas reacciones inmunes. La IL-10, de hecho, fue descubierta como un inhibidor de la pro-ducción de IFN-γ.
• La IL-10 inhibe la expresión de coestimuladores y de moléculas HLA de clase II por los macrófagos, por lo que impide la presentación de antígenos y l activación de las células T.
• La IL-10 estimula, en cultivos, la proliferación de linf. B pero se desconoce la importancia de este hecho.
Los ratones knockout para IL-10 desarrollan enfermedad inflamatoria intestinal, probablemente por re-acción incontrolada de los macrófagos contra los gérmenes entéricos.
El virus de Epstein-Barr contiene un gen homólogo al de la IL-10 humana y la IL-10 producida por este gen posee las mismas propiedades que la humana. Esto puede hacer pensar que el virus ha adquirido el este gen durante su evolución con el fin de evitar las reacciones del huésped contra él. Si éste es el caso, no cabe duda que la adquisición de este gen le ha aportado una ventaja evolutiva.
7. IL-6
La IL-6 participa tanto en la inmunidad innata como en la adaptativa. Es producida por los fagocitos mo-nonuleares, células del endotelio vascular, fibroblastos, linfoctios T activados y otras células en respues-ta a la infección por microbios o tras la acción de otras citocinas, sobre todo IL-1 y TNF (fig. 8).
La IL-6 es un homodímero y cada subunidad tiene 4 dominios globulares en hélice α.
El receptor de la IL-6 posee dos subunidades. Una se une a la citocina y la otra transduce las señales. Las dos pertenecen a la familia de receptores de citocinas del tipo I. La subunidad transductora de seña-les es una glicoproteína de 130 kD (gp130) que activa la via de señalización JAK/STAT. También actúa como cadena trasductora en otros receptores de citocinas (p.e. la IL-11).
La IL-6 ejerce varias funciones:
• En la inmunidad innata estimula la síntesis de proteínas de fase aguda por los hepatocitos y por lo tanto contribuye a los efectos sistémicos de la inflamación (la llamada “respuesta de fase aguda”).
• En la inmunidad adaptativa estimula el crecimiento de los linfocitos que se han diferenciado a célu-las productoras de anticuerpos.
• De igual manera, la IL-6 es también factor de crecimiento para las células plasmáticas cancerosas (mielomas) y muchas células de mieloma que crecen de forma autónoma secretan IL-6 como un factor de crecimiento autocrino. Además la IL-6 puede favorecer el crecimiento de hibridomas (mie-loma + célula plasmática) productores de anticuerpos monoclonales.
8. IL-15
Es producida fundamentalmente por los fagocitos mononucleares en respuesta a infección viral, LPS y otras señales que disparan la inmunidad innata.
Estructuralmente es homóloga a la IL-2 (ver IL-2).
El receptor para la IL-15 posee tres cadenas´(α, β y γ ). La cadena α es la que se une a la IL-15 y es típica de esta citocina. Las cadenas β y γ, que son las transductoras de señales, son idénticas a las β y γ de la IL-2.
La función mejor conocida de la IL-15 es potenciar la proliferación de las células NK. Dado que la IL-15 es sintetizada tan pronto como aparece una infección viral, la acción antiviral de las células NK expandi-

Citocinas Página
15
das tiende a controlar la infección viral ya en los primeros días de infección, cuando aún no ha dado tiempo a que se desarrollen los linfocitos T citotóxicos específicos antivirales.
La IL-15 puede también actuar como un factor de crecimiento y alargador de la vida de las células T, sobre todo de las T CD8+ de memoria.
Los ratones con falta de IL-15 o de la cadena α de su receptor tienen muy disminuido el número de cé-lulas NK y algo el de células T.
9. IL-18
Es estructuralmente homóloga a la IL-1 y su receptor también es del “tipo pejaje” pero las funciones de la IL-1 y de la IL-18 son completamente diferentes.
La IL-18 es producida por los macrófagos en respuesta al LPS y otros productos microbianos.
Estimula la producción de IFN-γ por las células T y NK y sinergiza con la IL-12 en esta respuesta. Por lo tanto la IL-18 es un inductor de la inmunidad celulo-mediada, especialmente en combinación con la IL-12.
Los ratones knockout para IL-18 producen poco IFN-γ y a los que les falta la IL-18 y la IL-12 casi no pro-ducen ningún IFN-γ ni montan respuestas Th1 contra microbios intracelulares.
Como la IL-1β, la IL-18 es sintetizada como un precursor que tiene que ser escindido por la enzima convertidora de IL-1 o ICE (IL-1 Converting Enzyme). Los ratones knockout para ICE tienen una pro-ducción defectuosa de IFN-γ tras estímulo con LPS, lo que es atribuible a la falta de IL-18 funcionante.
G) Papel de las citocinas en la respuesta innata y en la inflamación Varias citocinas poseen funciones relevantes en la respuesta inmune innata frente a distintas clases de microorganismos. En las infecciones por bacterias extracelulares piógenas (formadoras de pus), los macrófagos responden a las endotoxinas bacterianas produciendo TNF, IL-1 y quimiocinas (fig. 9).
El TNF y la IL-1 actúan so-bre el endotelio vascular en el lugar de la infección indu-ciendo la expresión de mo-léculas de adhesión lo que facilita la unión estable de los monocitos y neutrófilos.
Las quimiocinas producidas por los macrófagos y por las células endoteliales estimu-lan la extravasación de los leucocitos al lugar de la infección, donde se monta la reacción inmune.
Niv
el d
e ci
toci
nas
en s
uero
Horas tras la inyección de LPS0 1 3 6
LPS
TNF
Trombosis
Quimiocinas
IL-1 IL-12
Inflamación
NK T
IFN-γ Activ. Macrófagos
Fig. 9. Citocinas más relevantes en la RI innata y la inflamación.

Citocinas Página
16
Los macrófagos responden a muchos microbios, incluidos los intracelulares y los que producen LPS, produciendo IL-12, que induce la producción local de IFN-γ por las células T y las NK. El IFN-γ activa entonces a los macrófagos para que destruyan a los microbios fagocitados. La IL-12 estimula la res-puesta inmune adaptativa y dirige la diferenciación T hacia TH1 que son las mediadoras de la inmuni-dad celular y las más efectivas para la destrucción de bacterias intracelulares. Estas acciones de le IL-12m son complementadas por la IL-18.
El reclutamiento y activación de leucocitos mediados por citocinas son los responsables del daño que se origina en los tejidos normales en reacciones de la inmunidad innata contra la infección. Estas citocinas derivadas de los macrófagos, especialmente TNF, IL-1 e IL-12 son los responsables de las manifesta-ciones sistémicas de la infección.
En las infecciones virales los IFN-I, secretados por células infectadas y por macrófagos, inhiben la repli-cación viral. La IL-15 induce la expansión de las células NK y la IL-12 estimula su actividad citolítica. Con la lisis de las células infectadas, se eliminan los reservorios del virus.
De los patrones de citocinas que se producen en respuesta a diferentes microbios depende el tipo de reacción que se pondrá en marcha contra ellos. Por ejemplo, la respuesta temprana contra bacterias piógenas la llevan a cabo fundamentalmente los neutrófilos, la respuesta contra bacterias intracelulares es dominada por los macrófagos y las reacciones antivirales las llevan a cabo las células NK y otras células. No obstante pueden existir solapamientos y todos estos tipos de reacciones, con más o menos intensidad, pueden ponerse de manifiesto en infecciones muy distintas.
H) Citocinas de la inmunidad adaptativa o adquirida Estas citocinas participan en la proliferación y diferenciación de los linfocitos (una vez que han reconoci-do al antígeno) en la fase de activación así como en la activación de células efectoras especializadas durante la fase efectora. Fase de la RI adquirida → Reconocimiento Activación Efectora
Acción de las citocinas → - Proliferación y diferencia-ción de linfocitos
Activación de células efectoras especializadas
Una de las principales consecuencias del reconocimiento del antígeno por los linfocitos es la producción de citocinas por parte de éstos. Tras el reconocimiento antigénico, los linfocitos T pueden derivar a Th1 o Th2 y estas dos subpoblaciones T producen patrones de citocinas diferentes que llevan a cabo fun-ciones distintas.
La tabla 5 resume las citocinas que participan en la RI adquirida o adaptativa.
Tabla 5 – Algunas propiedades de las citocinas de la inmunidad adquirida
Citocina Tamaño Origen Dianas Acciones T Proliferación, aumento sínt. Citocinas, potencia apoptosis por Fas NK Proliferación, activación IL-2 14-17* T B Proliferación, síntesis de Ac. T Proliferación, diferenciación a TH2 B Cambio de isotipo a IgE IL-4 18 TH2
Cebadas Cebadas Proliferación B Proliferación, producción de IgA IL-5 45-50** TH2 Eosinófilos Aumento de la producción, Activación Macrófagos Incremento actividad microbicida Endotel. Activación IFN-γ 50+
TH1 CD8 NK Otras Amento expresión HLA (I y II) Aumento procesam. y pres. Ags.
T Inhibición proliferación y funciones efectoras B Inhibición proliferación, producción de IgA TGF-β 25++
T Macróf Otras Macrófagos Inhibición

Citocinas Página
17
Neutrófilos Reclutamiento y activación LT 21-24# T Org. Linf. Organogénesis
IL-13 15 TH2 Macrófagos Inhibición de la activación * Todos los pesos se dan en kD ** Es un homodímero. + 50 kD ya glicosilada. Es un homodímero de 21 a 24 kD cada monómero. ++ Homodímero de 12.5 kD cada monómero. # Es un homotrímero o un homodímero asociado a LT β2 -----------------------------------------------------------------------------------------------------------------------------------------------------------LGF-β: Factor de crecimiento transformante β LT: Linfotoxina
1) IL-2 Es el factor responsable de la expansión clonal de los linfocitos T después de haber reconocido a su antígeno. Por este motivo la IL-2 se llamó inicialmente factor de crecimiento de los linfocitos T. La IL-2 actúa fundamentalmente sobre la misma célula que la produce por lo que su función es autocrina.
Producción, estructura y receptores
Es producida por los linfocitos T CD4+ y en menor medida por los T CD8+. Tras la activación de los linfocitos por el antígeno específico, con la colaboración de los coestimuladores, se inicia la transcripción del gen de la IL-2, la traducción y la secreción de la Interleucina. Estos fenómenos son transitorios ocu-rriendo la máxima secreción a las 8-12 horas después de la activación.
La IL-2 es una glicoproteína globular de 14-17 kD plegada en 4 dominios en hélice α. Es el prototipo de la citocinas con 4 hélices α que interactúan con los receptores de citocinas del tipo I.
El receptor de la IL-2 (IL-2R) está formado por tres proteínas asociadas no covalentemente llamadas α, β, y γ. Las cadenas β y γ son miembros de la familia de receptores de citocinas del tipo I. Las cadenas α y β son las que se unen la IL-2 y las β y γ trasducen las señales al interior de la célula (fig. 10).
La cadena α es un polipéptido de 55kD (también llamado p55) que aparece tras la activación de los linfocitos T y que se llamó originalmente Tac (T activation). Esta cadena α posee poca afinidad por la IL-2 (Kd=1x10-8M). La cadena β se expresa a bajo nivel en las células T en reposo y en las NK y está aso-ciada con otro polipéptido, la cadena γ. Las cadenas β y γ forman un receptor de afinidad intermedia por la IL-2 (Kd=1x10-9M). La IL-2 induce el crecimiento (división) de las células que expresan este receptor βγ de afinidad intermedia. A la cadena γ también se la llama γc (γ common, γ común) porque está presente en otros receptores de interleucinas, como la IL-4, IL-7 e IL-15. El receptor αβγ une a la IL-2 con mucha más afinidad (Kd=1x10-11M) y por tanto los requerimientos de IL-2 para estimular el crecimiento son me-nores.
El hecho de que tras la estimulación antigénica se produzca IL-2 y además se exprese el receptor para IL-2 garantiza que solamente la células que han respondido al antígeno van a proliferar y montar una respuesta inmune.
La unión de la IL-2 a su receptor activa en el lif. T varias vias de transducción de señales. Entre estas están la JAK/STAT, que es exclusiva para citocinas, y las vias de la kinasa de fosfatidilinositol y la de Ras, que son activadas por muchos tipos de receptores. La estimulación mantenida (crónica) de los linfocitos T hace que estos se desprendan de la cadena α, de hecho la presencia d niveles altos de ca-dena α en el suero es sinónimo de estimulación antigénica intensa, como ocurre, por ejemplo, en el rechazo agudo de órganos trasplantados.
Acciones biológicas.
Citocinas Página
18
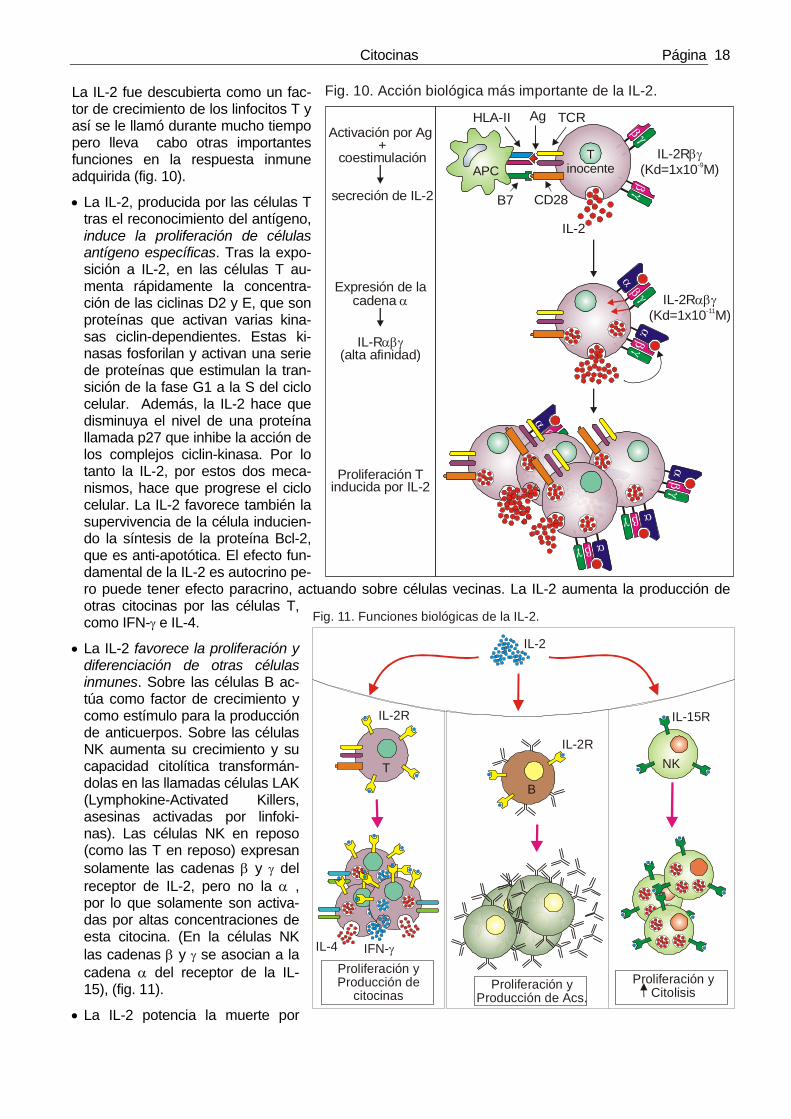
La IL-2 fue descubierta como un fac-tor de crecimiento de los linfocitos T y así se le llamó durante mucho tiempo pero lleva cabo otras importantes funciones en la respuesta inmune adquirida (fig. 10).
• La IL-2, producida por las células T tras el reconocimiento del antígeno, induce la proliferación de células antígeno específicas. Tras la expo-sición a IL-2, en las células T au-menta rápidamente la concentra-ción de las ciclinas D2 y E, que son proteínas que activan varias kina-sas ciclin-dependientes. Estas ki-nasas fosforilan y activan una serie de proteínas que estimulan la tran-sición de la fase G1 a la S del ciclo celular. Además, la IL-2 hace que disminuya el nivel de una proteína llamada p27 que inhibe la acción de los complejos ciclin-kinasa. Por lo tanto la IL-2, por estos dos meca-nismos, hace que progrese el ciclo celular. La IL-2 favorece también la supervivencia de la célula inducien-do la síntesis de la proteína Bcl-2, que es anti-apotótica. El efecto fun-damental de la IL-2 es autocrino pe-ro puede tener efecto paracrino, actuando sobre células vecinas. La IL-2 aumenta la producción de otras citocinas por las células T, como IFN-γ e IL-4.
• La IL-2 favorece la proliferación y diferenciación de otras células inmunes. Sobre las células B ac-túa como factor de crecimiento y como estímulo para la producción de anticuerpos. Sobre las células NK aumenta su crecimiento y su capacidad citolítica transformán-dolas en las llamadas células LAK (Lymphokine-Activated Killers, asesinas activadas por linfoki-nas). Las células NK en reposo (como las T en reposo) expresan solamente las cadenas β y γ del receptor de IL-2, pero no la α , por lo que solamente son activa-das por altas concentraciones de esta citocina. (En la células NK las cadenas β y γ se asocian a la cadena α del receptor de la IL-15), (fig. 11).
• La IL-2 potencia la muerte por
Activación por Ag +
coestimulación
secreción de IL-2
APC
HLA-II TCR
B7 CD28
Tinocente
Ag
IL-2R(Kd=1x10 M)
βγ-9
IL-2IL-2
Expresión de lacadena
IL-R
(alta afinidad)
α
αβγ
Proliferación Tinducida por IL-2
IL-2R(Kd=1x10 M)
αβγ-11
Fig. 10. Acción biológica más importante de la IL-2.
BB
Proliferación yProducción de Acs.
NK
IL-15RIL-2R
T
Proliferación yProducción de
citocinasProliferación y
Citolisis
IL-4 IFN-γ
B
IL-2R
IL-2
Fig. 11. Funciones biológicas de la IL-2.

Citocinas Página
19
apoptosis de linfocitos T activados por antígeno. Cuando la activación de los linfocitos por su antígeno se produce en presencia de IL-2 los linfocitos se hacen muy sensibles a la muerte por apoptosis por la vía de Fas. Es llamativo que la misma citocina pueda por un lado estimular la proliferación y supervi-vencia por un lado y por otro la apoptosis. Parece que si la respuesta inmune es persistente y las célu-las T son expuestas a cantidades crecientes de IL-2 las acciones proapoptoticas se hacen dominan-tes. Los ratones knockout para IL-2, IL-2Rα o IL-2Rβ desarrollan autoinmunidad. Esto parece indicar la necesidad de la acción de la IL-2 para eliminar las células que son estimuladas repetidamente, por ejemplo, por antígenos propios. Por el contrario los ratones knockout a los que le falta la cadena γ y los humanos con mutación en esta cadena presentan defectos de maduración linfocitaria. Esto parece ser debido a la incapacidad de los linfocitos T para responder a la IL-7, que es linfopoyética, dado que el receptor de esta Interleucina también usa la cadena γ.
2) IL-4 Es el mayor estimulo para la producción de IgE y para la diferenciación de CD4+ inocentes (Th0) hacia Th2.
Estructura, producción y receptores de la IL-4.
La Il-4 es un miembro de la familia de citocinas con 4 hélices α. La producen los linfocitos Th2, las célu-las cebadas activada y los basófilos.
El receptor de la IL-4 posee una cadena α (que pertenece a la familia de receptores tipo I y que es la que se une a la IL-4) y una cadena γ idéntica a la del IL-2R. Este receptor señaliza a través de la vía JAK/STAT y a través de una vía en la que participa el llamado sustrato de respuesta de insulina, concre-tamente el IRS-2 (Insulin Response Substrate-2). La IL-4 es la única citocina que activa a la proteína STAT6, que es la responsable de muchas de las acciones de la IL-4, tales como diferenciación a Th2 y el cambio de clase a IgE. La activación de la vía IRS-2 es la responsable de la proliferación celular que induce la IL-4. En las células no linfoides la cadena α no se une a la cadena γ mencionada del receptor de la IL-2 sino con otra cadena que también forma parte del receptor de la IL-13.
Acciones biológicas de la IL-4.
• La IL-4 es el estimulo más importante para que los linfo-citos B hagan el cambio de isotipo hacia IgE. Los ratones knockout que no producen IL-4 poseen menos del 10% de la cifra normal de IgE. Los Ac. IgE participan en la defensa contra helmintos y artrópodos mediada por eosinófilos. Pro-ducir la IL-4 necesaria para que se produzca este cambio de clase de Ig. es la función más importante de las células Th2 en la defensa del hués-ped. La IgE es la mediadora más importante de las reac-ciones alérgicas. En el ratón la IL-4 inhibe el cambio de clase hacia IgG2a e IgG3, jus-to lo contrario de lo que hace el IFN-γ. Este es un ejemplo de acción antagónica entre ci-tocinas (fig. 12).
BB
Cambio de isotipoa IgE / IgG4
B
IL-4
IgMm
IgEIgG4
Macrófago
Inhibición dela activación
de macrófagos
TCD4+Th0
IL-4R IL-4R
TCD4+Th2
Derivación y expansión a Th2
Fig 12. Funciones biológicas de la IL-4

Citocinas Página
20
• La IL-4 estimula la derivación de células TCD4+ inocentes hacia Th2 y sobre estas Th2 actúa como factor de crecimiento. Los ratones knockout a los que les falta IL-4 o STAT6 son incapaces de generar y mantener cifras aceptables de células Th2 incluso en presencia de estímulos que actúan como in-ductores potentes de estas células, como son las infecciones por helmintos.
• La IL-4 antagoniza el efecto estimulante del IFN-γ sobre los macrófagos por lo que inhibe las reaccio-nes celulo-mediadas. Este puede ser uno de los mecanismos por los que las células actúan como in-hibidoras de la inflamación.
3) IL-5 La IL-5 es un activador de los eosinófilos y actúa como puente de unión entre la activación de las célu-las T y la inflamación esosinofílica.
Estructura, producción y receptores de la IL-5.
La IL-5 es un homodímero. Cada subunidad contiene un dominio en hélice α.
La IL-5 es producida por las células Th2 y por los mastocitos activados.
El receptor de la IL-5 es del tipo I de la familia de receptores de citocinas. Está asociado con una cade-na que actúa como transductora de señales y que está presente también el receptor de la IL-3 y en el de GM-CSF (Granulocyte Macrophage-Colony Stimulating Factor, factor estimulante de colonias granu-locito-macrofágicas). Por este motivo a esta subunidad se le llama a veces cadena β común. La IL-5 envía sus señales a través de la vía JAK/STAT.
Acciones biológicas de la IL-5
• La IL-5 estimula la diferenciación y el crecimiento de los eosinófilos y activa a los eosinófilos maduros. Estos eosinófilos activados pueden matar helmintos. Los eosinófilos expresan receptores para el fragmento Fc de la IgE y pueden por tanto unirse a la IgE una vez esta ha reaccionado con los helmin-tos. Por lo tanto las dos citocinas más importantes de las células Th2 funcionan de horma coordinada: la IL-4 estimula producción de IgE, que opsoniza a los helmintos para que sean reconocidos por los eosinófilos y la IL-5 activa a los eosinófilos para que destruyan a los parásitos. Los ratones knockout sin IL-5 poseen respuestas defectuosas de los eosinófilos y son susceptibles a ciertas infecciones por helmintos.
• La IL-5 estimula la proliferación de linfocitos B y la producción de anticuerpos IgA. De hecho, la IL-5 se identificó inicialmente en ratones como una citocina que estimulaba el crecimiento de los linfocitos B. No obstante esta función deben llevarla a cabo también otras citocinas porque los ratones knockout para IL-5 poseen cifras normales de estas células.
4) IFN-γ El IFN-γ es la principal citocina activadora de los macrófagos y tiene un papel fundamental tanto en la inmunidad innata como en la inmunidad específica mediada por células. Es la citocina estrella de las células Th1. El IFN-γ también se llama IFN tipo II o IFN inmune. Aunque tiene alguna actividad antiviral actúa fundamentalmente como citocina efectora de las respuestas inmunes.
Estructura, producción y receptores del IFN-γ
El IFN-γ es una proteína homodimérica producida por las células NK, CD4+Th1 y CD8+.
Las células NK secretan IFN-γ tras el reconocimiento en los microbios de componentes aún desconoci-dos o en respuesta a la IL-12. En este aspecto el IFN-γ funciona como un mediador de la inmunidad innata.
En la inmunidad adaptativa los linfocitos T producen IFN-γ tras el reconocimiento del Ag. Esta produc-ción es incrementada por la IL-12. La secuencia de reacciones en las que participan la IL-12 y el IFN-γ es vital en las reacciones contra microbios intracelulares.
El receptor para IFN-γ está formado por dos polipéptidos estructuralmente homólogos que pertenecen al tipo II de la familia de receptores de citocinas, uno de ellos se une a la citocina y otro transduce las se-ñales. Tras la unión de la citocina se activa STAT1 que estimula la transcripción de genes de respuesta

Citocinas Página
21
del IFN-γ, incluidos genes que codifican a moléculas del MHC, enzimas que sintetizan sustancias micro-bicidas como el óxido nítrico y citocinas como la subunidad p40 de la IL-12. Tras la reacción del IFN-γ con su receptor se activan varios genes de respuesta de IFN-γ, bien por la acción de STAT solamente o por STAT en combinación con otros factores de transcripción que también son inducidos por STAT1, tales como el factor 1 de respuesta de IFN y el transactivador de clase II. Por lo tanto STAT1 es un re-gulador “maestro” de las acciones del IFN-γ. Los ratones knockout para STAT1 son totalmente insensi-bles a la acción de IFN-γ.
Acciones biológicas del IFN-γ
• El IFN-γ es la citocina que usan las células T y las NK para activar a los macrófagos para que elimi-nen a los microbios que han fa-gocitado (fig. 13).
El IFN-γ estimula la síntesis de in-termediarios de oxígeno reactivos y de oxido nítrico con lo cual au-menta la capacidad microbicida de los macrófagos. Esto lo lleva a cabo el IFN-γ aumentando la transcripción y/o el ensamblaje de las encimas necesarias para lle-var a cab dichos cambios meta-bólicos. Estas enzimas son la oxidasa de los fagocitos y la oxido nítrico cintazas inducible. Las mo-léculas activas se producen en los lisosomas y la destrucción de los microbios se produce en los fagolisosomas.
• El IFN-γ aumenta la expresión de moléculas del MHC de clase I y clase II y de coestimuladores en las CPA. Además, aumenta la expre-sión de muchas proteínas relacionadas con el procesamiento de antígenos como TAP, LMP-2, LMP-7 y HLA-DM por lo que mejora esta presentación y por tanto el reconocimiento de antígenos por las cé-lulas T.
El IFN-γ activa a las células endoteliales de los vasos sanguíneos, y potencia muchas de las acciones del TNF sobre estas células, con lo que potencia la adhesión y la extravasación de los linfocitos T hacia los lugares de infección.
• El IFN-γ favorece la diferenciación de las células T CD4+ “inocentes”(Th0) hacia Th1 e inhibe su dife-renciación a Th2. El efecto sobra la diferenciación hacia Th1 lo ejerce el IFN-γ de una forma indirecta: el IFN-γ induce la secreción de IL-12 por los fagocitos mononucleares y es la IL-12 la que favorece la diferenciación de Th0 hacia Th1. En los ratones, el IFN-γ aumenta la expresión de la cadena de seña-lización del receptor de la IL-12.
• El IFN-γ actúa sobre los linfocitos B para potenciar el cambio de clase hacia algunos isotipos de Ig. e inhibir el cambio hacia otros isotipos. Estas acciones no han sido exploradas en profundidad en humanos pero en los ratones el IFN-γ induce el cambio hacia IgG2a y lo inhibe hacia IgG1 e IgE, que son isotipos dependientes de la IL-4. Las Igs inducidas por el IFN-γ se unen a los receptores Fc de los fagocitos y fijan complemento y estos dos hechos facilitan la fagocitosis de los microbios opsonizados.
• El IFN-γ activa a los neutrófilos y activa la capacidad citolítica de las células NK.
En resumen, la acción del IFN-γ es potenciar las reacciones inflamatorias ricas en macrófagos e inhibir las reacciones inflamatorias dependientes de IgE ricas en eosinófilos. Los ratones knockout para IFN-γ
Macrófagoinfectado
Activación yCapacidad
microbicida
B
IgMm
Cambio declase a Acs.opsonizantes
CD4+Th0
TCR
CD4+Th1
Derivación de Th0
hacia Th1
Expres. HLA
Present. Ags.
CPA
HLA-II HLA-I
IFN-γFig. 13. Aciones biológicas del IFN-γ

Citocinas Página
22
o su receptor son susceptibles a infecciones por microbios intracelulares tales como micobacterias, de-bido a defectos de activación de los macrófagos.
5) 5) Factor β Transformante del Crecimiento (TGF- β, Transforíning Growth Factor β)
El efecto más importante de esta citocina es inhibir la proliferación y activación de los linfocitos y otro leucocitos.
El TGF-β se descubrió como un producto que prolongaba la supervivencia de células en agar. En reali-dad se trata de una familia de moléculas íntimamente relacionadas (TGF-β1, TGF-β2 y TGF-β3) codifi-cadas por genes diferentes. Las células del sistema inmune sintetizan sobre todo TGF-β1.
El TGF-β1 es una proteína homotrimérica que se sintetiza como un precursor y se activa por escisión proteolítica. Es secretado por los linfocitos T activados por antigeno, por fagocitos mononucleares acti-va dos por LPS y por otras muchas células. Las células T que producen TGF-β algunas veces produ-cen también IL-4 e IL-10, las dos con propiedades inmunosupresoras como el TGF-β. Esta subpobla-ción de células reguladores se ha llamado Th3, pero aún se sabe muy poco de ella. Existen dos recep-tores polipeptídicos de alta afinidad para TGF-β, llamados tipo I y tipo II) que señalizan mediante un do-minio de serin-treonin kinasa. Acciones biológicas.
• El TGF-β inhibe la proliferación y diferenciación de los linf. T y la activación de los macrófagos. TGF-β también actúa sobre leucocitos polimorfonucteares y células endoteliales para contrarrestar lo efec-tos de las citocinas proinflamatorias. Mediante estas acciones el TGF-β inhibe las respuestas inflama-torias. Los ratones que han sido knocked out para el gen de TGF-β desarrollan reacciones inflama-trias incontroladas.
• El TGF-β estimula en los ratones el cambio de isotipo hacia IgA. IgA es el anticuerpo de las secre-ciones y las mucosas.
6) Linfotoxina (LT)o TNF-β Es una citocina producida, en diferentes formas, por los linfocitos y otras células. Posee un 30% de homología con el TNF de los macrófagos y comparte muchas de sus funciones a lo que debe su se-gundo nombre, TNF-β. La forma secretada de la linfotoxina, a veces llamada LT-α, es un homotrí-mero similar al TNF y se une a los receptores del TNF. La LT también se expresa como una proteí-na de membrana y en esta forma una cadena de la forma secretada se asocia con dos subunidades de una proteína de membrana llamada LTβ. Esta forma de membrana se une a un receptor distinto pero que también pertenece a la familia de receptores del TNF. La LT activa a las células endoteliales y a los neutrófilos actuando como mediador de la respuesta inflamatoria aguda y es por tato un nexo de unión entre la activación de las células T y la inflamación. Estas acciones biológicas de la LT son las mismas que las del TNF, dado que se unen al mismo re-ceptor. Sin embargo, dado que la cantidad de LT sintetizada por los linfocitos T activados por antí-geno es mucho menor que la cantidad de TNF sintetizado por los fagocitos mononucleares estimula-dos por LPS, la LT no es detestable en la circulación. Por lo tanto, la LT es una citocina de acción local y no un mediador del reacciones sistémicas. Estudios realizados en ratones transgénicos y knockout han implicado a la LT en el desarrollo de los órganos linfoides. Los ratones knockout a los que les falta la LTβ de membrana o su receptor forman sus nódulos linfoides, las placas de Peyer y la pulpa blanca esplénica de forma defectuosa, por ejemplo las áreas B no se forman normalmente y no existen centros germinales en el bazo. La LT

Citocinas Página
23
puede esta involucrada en estos trastornos por inducir la producción de quimiocinas que guiarían a estas células a áreas determinadas de los órganos linfoides.
7) IL-13 Es una citocina estructuralmente similar a la IL-4 que es producida por las células Th2 y algunas cé-lula epiteliales. El receptor para 1L-13 se encuentra sobre todo en células no linfoides, como los macrófagos y se puede activar tanto por la IL-13 como por la IL-4. Por otro lado, aunque la IL-13 puede mimetizar efecto de la IL-4 en células no linfoides como los macrófagos, su efecto sobre linfo-citos T y B es mucho menor que el de la IL-4. De cualquier manera el grado de redundancia entre IL-13 e IL-4 no está bien establecido. El papel fundamental de la IL-13 sobre los macrófagos es inhibir su activación y antagonizar al IFN-γ Los ratones knockout para IL-13 muestran descenso de la producción de IL-13 y de reacciones alér-gicas.
8) Otras citocínas.
Se han descrito otras 3 citocinas pero su función biológica no está aún bien establecida. La IL-16 es producida por los linfocitos T y actúa como atrayente para eosinófilos. Sin embargo, ha resultado ser un fragmento de una proteína intracelular por lo que no se sabe si su producción es fisiológica o es el resultado de la fragmentación de otra molécula. La IL-17 es producida por los linfocito T y mimetiza muchas de las acciones proinflamatorias del TNF la LT. Su función in vivo es desconocida. El Factor de Inhibición de la Miqración (MIF, migration inhibition factor) es igualmente producido por los linfocitos T- MIF inmoviliza a los fagocitos mononucleares, con lo que quedarían retenidos en los lugares de inflamación. Curiosamente esta fue la primera actividad de citocina de todas las descri-tas, pero solo se puede hablar de 'actividad de citocina" y no de "citocina' porque todavía no se ha podido caracterizar a la molécula o moléculas que poseen esta actividad.
I) Papel de las citocinas T en la inmunidad adaptativa: desarrollo y funciones de las subpobla-ciones Th1 y Th2.
Las células T CD4+ inocentes (Th0), en respuesta a la estimulación por antígenos proteicos, puede diferenciarse a subpoblaciones efectoras que producen patrones diferentes de citocinas y que, por lo tanto, llevan a cabo funciones efectoras distintas. Las dos subpoblacíones mejor definidas de linfocitos T CD4 son Th1 y Th2. El IFN-γ es la citocina que define a la población Th1 y la IL-4 y la IL-5 son las que definen a la Th2 (tabla 6) Estas subpoblacíones de linfocitos T CD4+, que podían ser distinguidas por su producción de citoci-nas fueron descubiertas en la década de los 80 al estudiar grandes poblaciones de clonos de linfoci-tos T CD4 derivados de linfocitos T CD4+ de ratón estimulados por antígenos. Hoy es evidente que cada célula T puede segregar una mezcla de citocinas y que pueden existir muchas más subpobla-cíones de las descritas. Sin embargo las reacciones inmunes crónicas están dominadas o por Th1 o por Th2. La proporción relativa de estas dos subpoblacíones en una reacción inmune determina en gran medida las funciones protectoras y las consecuencias patológicas de tal reacción. 1. Propiedades de las subpoblaciones T CD4+

Citocinas Página
24
La mejor característica diferencial entre Thl y Th2 es las cito-cinas que producen, dado que hasta el momento no existe ningún marcador fenotípico distintivo que permita su identifi-cación. Estas subpoblaciones muestran también diferencias en la expresión de receptores de citocinas pero esto puede variar con su estado de activación y puede no ser estable. Las citocinas producidas por estas células no solo determinan sus funciones efectoras sino que también participan en el desarrollo y la expansión de las respectivas subpoblaciones. Por ejemplo, el IFN-γ secretado por las células Th1 estimula la diferenciación de células Th0 hacia Th1 e inhibe la diferen-ciación de Th0 hacia Th2. La IL-4 producida por Th2 aumen-ta la diferenciación hacia Th2 y la IL-10 producidas por la cé-lulas Th2 y otras inhibe la activación de células Th1. Por lo tanto, cada subpoblación se estimula a si misma e inhibe a la otra subpoblación. Por esta razón, una vez que una reacción inmune se desarrolla en una dirección, las propias citocinas hacen que se polarice más en esa dirección. Esto se observa mejor en infecciones crónicas, cuando la exposición al antí-geno es persistente. Estas subpoblacione de células T mues-tra patrones de migración distintos hacia los lugares de infec-ción debido a su distint capacidad para unirse a las selectinas endoteliales y su diferente respuesta a las quimiocinas. Ver el cuadro adjunto. 2. Desarrollo de las subpoblaciones Th1 y Th2 Las subpoblaciones Th1 y Th2 de desarrollan de un precursor único, la célula T CD4+ "inocente" y la diferenciación en una u otra dirección está determinada por estímulos generados en la fase tempra-na de la respuesta inmune. Los estímulos más importantes para la diferenciación son las citocinas. La IL-12 es el inductor más importante de Th1 y la IL-4 el de Th2. estas mismas citocinas actúan como factores de crecimiento para expandir sus poblaciones respectivas (fig. 14). La vía de diferenciación de Th1 es la respuesta a los microbios que infectan o activan a los macrófa-go y a los microbios que activan a las células NK. La diferenciación hacia Th1 es estimulada por agente que infectan a los macrófagos como muchas bacterias intracelulares (listeria y micobactedas) y alguno parásitos (leishmania). También es activada por los virus. Una característica común de todas estas infecciones es la producción de IL-12 durante la respuesta inmune innata. Algunos mi-crobios activan directamente a los macrófagos a producir IL-12 y otros pueden activarlos de forma indirecta. Por ejemplo, los virus y algunas bacterias activan a las células NK para producir IFN-γ, que induce en los macrófago la producción de IL-12. La IL-12 se une a sus receptores en las células CD4+, activa la vía STAT4, induce su diferenciación hacia Th1. El IFN-γ promueve el desarrollo hacia Th1 estimulando la producción de IL-12 por los macrófagos y la expresión de receptores para IL-12 en los linfocitos T.
Tabla 6 – Propiedades de los lin-focitos Th1 y Th2 Th1 Th2 Citocinas IFN-γ, IL-2, TNF IL-4, IL-5, IL-13 IL-10 IL-3, GM-CSF
+++ - ? ++
- +++ ++ ++
Receptores IFN-γ, cadena β IL-12, cadena β2
- ++
++ -
Quimiocinas CCR3 CCR4 CSCR3
? ? ++
++ ++ ?
Ligandos para selectinas E y P ++ ?
Anticuerpos es-timulados (ratón) IgG2a IgE, IgG1
(*) Activación macrófagos +++ -
(*) IgE e IgG4 en humanos
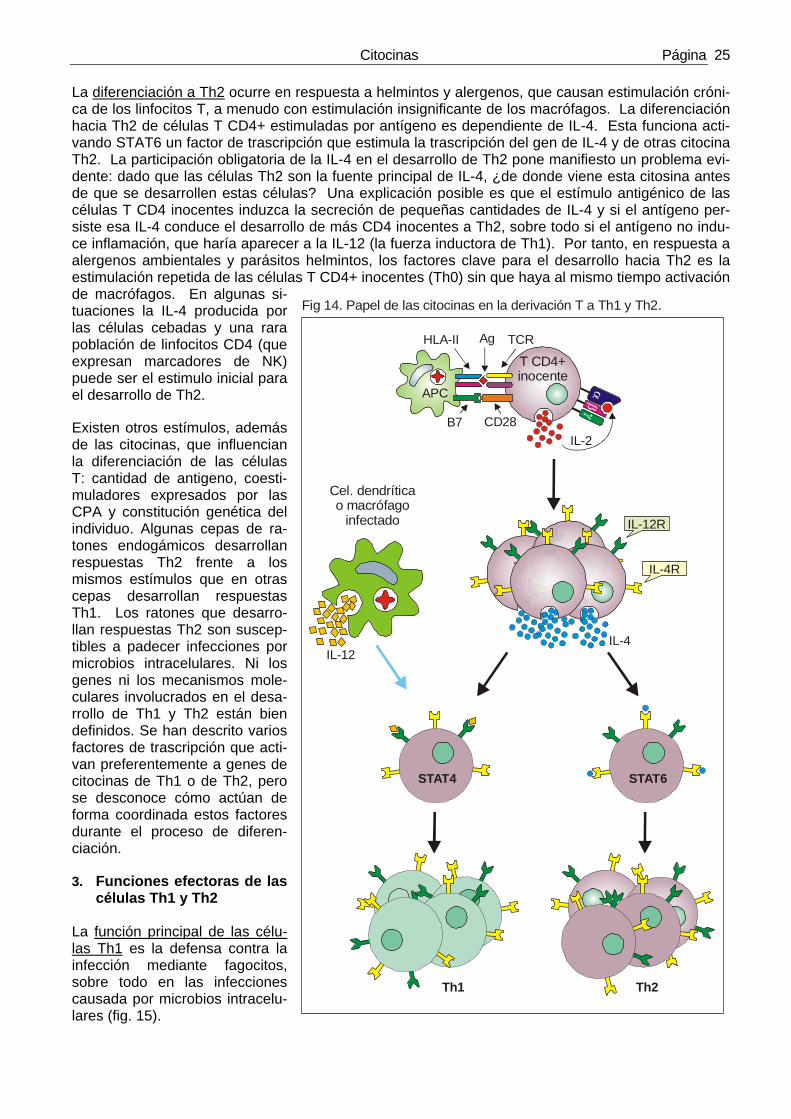
Citocinas Página
25
La diferenciación a Th2 ocurre en respuesta a helmintos y alergenos, que causan estimulación cróni-ca de los linfocitos T, a menudo con estimulación insignificante de los macrófagos. La diferenciación hacia Th2 de células T CD4+ estimuladas por antígeno es dependiente de IL-4. Esta funciona acti-vando STAT6 un factor de trascripción que estimula la trascripción del gen de IL-4 y de otras citocina Th2. La participación obligatoria de la IL-4 en el desarrollo de Th2 pone manifiesto un problema evi-dente: dado que las células Th2 son la fuente principal de IL-4, ¿de donde viene esta citosina antes de que se desarrollen estas células? Una explicación posible es que el estímulo antigénico de las células T CD4 inocentes induzca la secreción de pequeñas cantidades de IL-4 y si el antígeno per-siste esa IL-4 conduce el desarrollo de más CD4 inocentes a Th2, sobre todo si el antígeno no indu-ce inflamación, que haría aparecer a la IL-12 (la fuerza inductora de Th1). Por tanto, en respuesta a alergenos ambientales y parásitos helmintos, los factores clave para el desarrollo hacia Th2 es la estimulación repetida de las células T CD4+ inocentes (Th0) sin que haya al mismo tiempo activación de macrófagos. En algunas si-tuaciones la IL-4 producida por las células cebadas y una rara población de linfocitos CD4 (que expresan marcadores de NK) puede ser el estimulo inicial para el desarrollo de Th2. Existen otros estímulos, además de las citocinas, que influencian la diferenciación de las células T: cantidad de antigeno, coesti-muladores expresados por las CPA y constitución genética del individuo. Algunas cepas de ra-tones endogámicos desarrollan respuestas Th2 frente a los mismos estímulos que en otras cepas desarrollan respuestas Th1. Los ratones que desarro-llan respuestas Th2 son suscep-tibles a padecer infecciones por microbios intracelulares. Ni los genes ni los mecanismos mole-culares involucrados en el desa-rrollo de Th1 y Th2 están bien definidos. Se han descrito varios factores de trascripción que acti-van preferentemente a genes de citocinas de Th1 o de Th2, pero se desconoce cómo actúan de forma coordinada estos factores durante el proceso de diferen-ciación. 3. Funciones efectoras de las
células Th1 y Th2 La función principal de las célu-las Th1 es la defensa contra la infección mediante fagocitos, sobre todo en las infecciones causada por microbios intracelu-lares (fig. 15).
IL-2
APC
HLA-II TCR
B7 CD28
T CD4+inocente
Ag
IL-4
IL-12R
IL-4R
Cel. dendríticao macrófago
infectado
IL-12
STAT6
Th2
STAT4
Th1
Fig 14. Papel de las citocinas en la derivación T a Th1 y Th2.
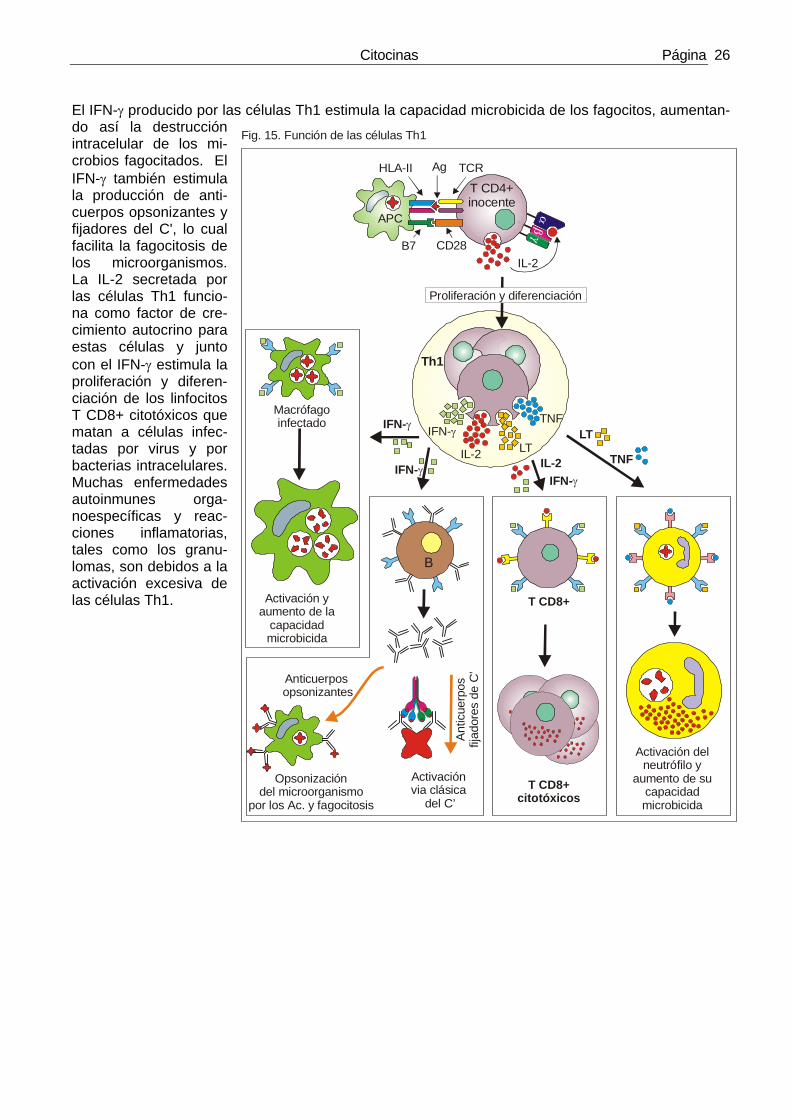
Citocinas Página
26
El IFN-γ producido por las células Th1 estimula la capacidad microbicida de los fagocitos, aumentan-do así la destrucción intracelular de los mi-crobios fagocitados. El IFN-γ también estimula la producción de anti-cuerpos opsonizantes y fijadores del C', lo cual facilita la fagocitosis de los microorganismos. La IL-2 secretada por las células Th1 funcio-na como factor de cre-cimiento autocrino para estas células y junto con el IFN-γ estimula la proliferación y diferen-ciación de los linfocitos T CD8+ citotóxicos que matan a células infec-tadas por virus y por bacterias intracelulares. Muchas enfermedades autoinmunes orga-noespecíficas y reac-ciones inflamatorias, tales como los granu-lomas, son debidos a la activación excesiva de las células Th1.
IL-2
APC
HLA-II TCR
B7 CD28
T CD4+inocente
Ag
IL-2
Th1
TNF
LTIFN-γ
Proliferación y diferenciación
IFN-γMacrófagoinfectado
Activación y aumento de la
capacidadmicrobicida
B
Ant
icue
rpos
fijad
ores
de
C’
Anticuerpos opsonizantes
Opsonizacióndel microorganismo
por los Ac. y fagocitosis
Activación via clásica
del C’
IFN-γ
T CD8+
T CD8+citotóxicos
IFN-γ
LT
TNFIL-2
Activación delneutrófilo y
aumento de sucapacidadmicrobicida
Fig. 15. Función de las células Th1
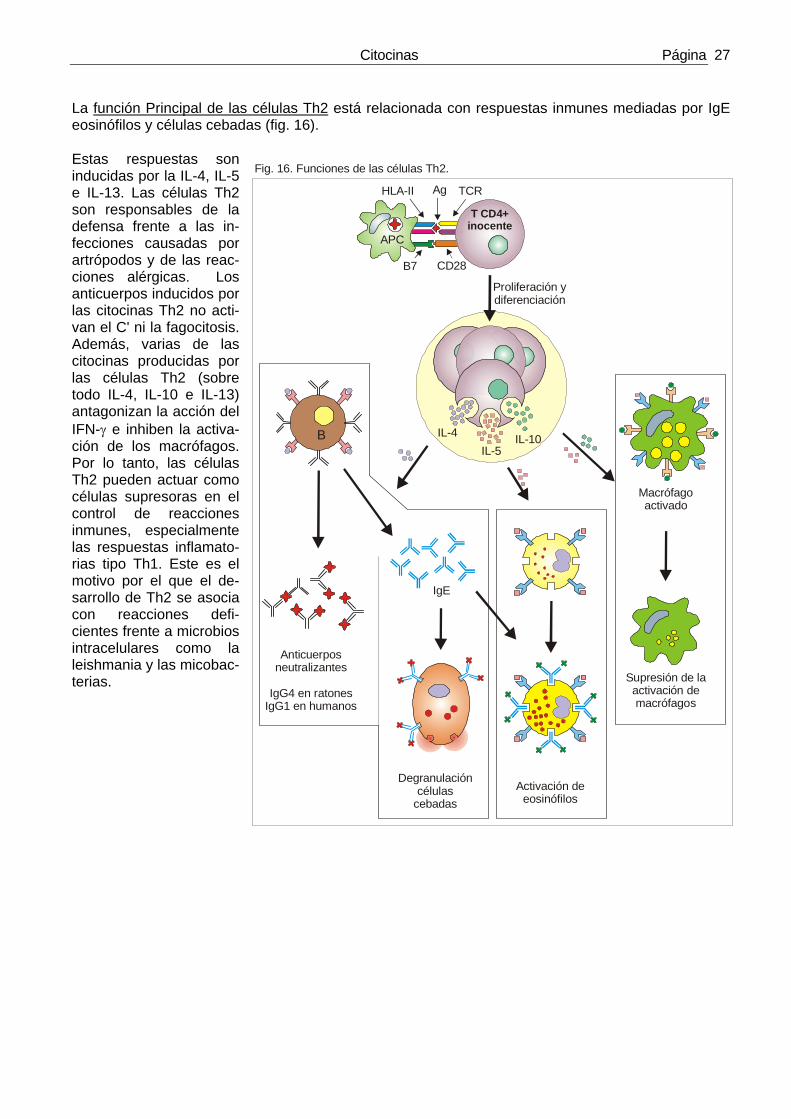
Citocinas Página
27
La función Principal de las células Th2 está relacionada con respuestas inmunes mediadas por IgE eosinófilos y células cebadas (fig. 16). Estas respuestas son inducidas por la IL-4, IL-5 e IL-13. Las células Th2 son responsables de la defensa frente a las in-fecciones causadas por artrópodos y de las reac-ciones alérgicas. Los anticuerpos inducidos por las citocinas Th2 no acti-van el C' ni la fagocitosis. Además, varias de las citocinas producidas por las células Th2 (sobre todo IL-4, IL-10 e IL-13) antagonizan la acción del IFN-γ e inhiben la activa-ción de los macrófagos. Por lo tanto, las células Th2 pueden actuar como células supresoras en el control de reacciones inmunes, especialmente las respuestas inflamato-rias tipo Th1. Este es el motivo por el que el de-sarrollo de Th2 se asocia con reacciones defi-cientes frente a microbios intracelulares como la leishmania y las micobac-terias.
APC
HLA-II TCR
B7 CD28
T CD4+inocente
Ag
Proliferación ydiferenciación
IL-4IL-5
IL-10B
Anticuerposneutralizantes
IgG4 en ratonesIgG1 en humanos
IgE
Degranulacióncélulas
cebadasActivación de
eosinófilos
Supresión de laactivación demacrófagos
Macrófagoactivado
Fig. 16. Funciones de las células Th2.
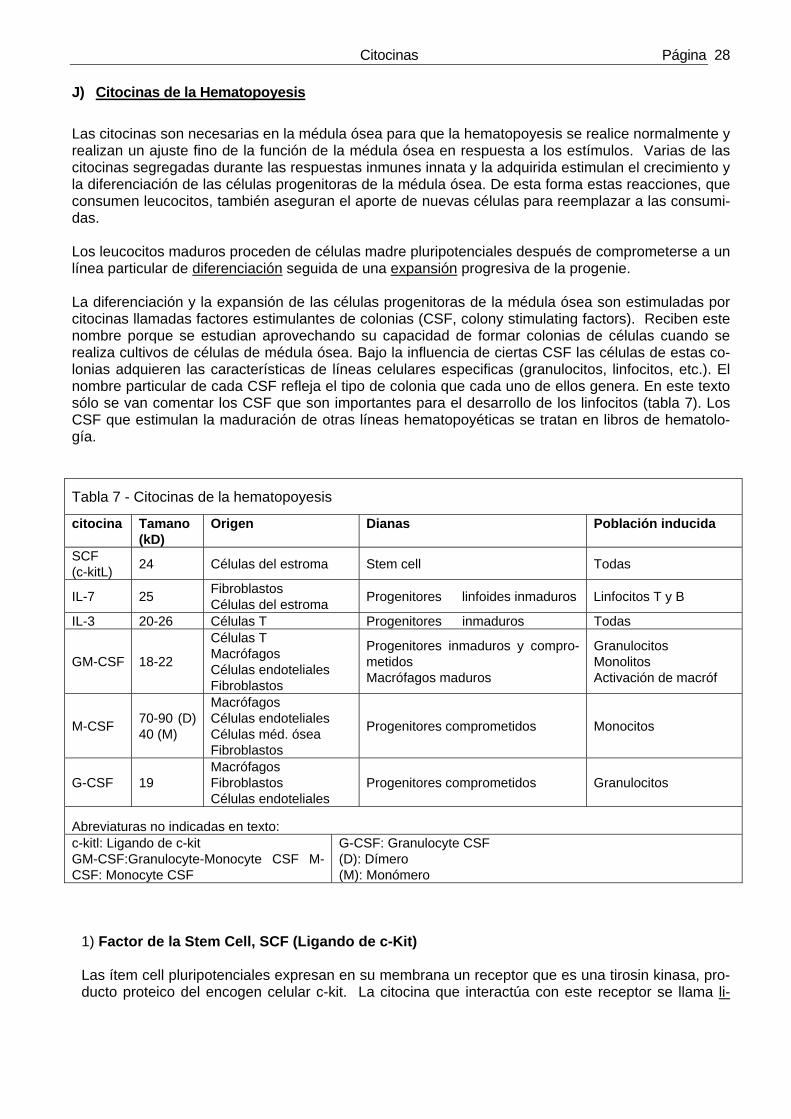
Citocinas Página
28
J) Citocinas de la Hematopoyesis Las citocinas son necesarias en la médula ósea para que la hematopoyesis se realice normalmente y realizan un ajuste fino de la función de la médula ósea en respuesta a los estímulos. Varias de las citocinas segregadas durante las respuestas inmunes innata y la adquirida estimulan el crecimiento y la diferenciación de las células progenitoras de la médula ósea. De esta forma estas reacciones, que consumen leucocitos, también aseguran el aporte de nuevas células para reemplazar a las consumi-das. Los leucocitos maduros proceden de células madre pluripotenciales después de comprometerse a un línea particular de diferenciación seguida de una expansión progresiva de la progenie. La diferenciación y la expansión de las células progenitoras de la médula ósea son estimuladas por citocinas llamadas factores estimulantes de colonias (CSF, colony stimulating factors). Reciben este nombre porque se estudian aprovechando su capacidad de formar colonias de células cuando se realiza cultivos de células de médula ósea. Bajo la influencia de ciertas CSF las células de estas co-lonias adquieren las características de líneas celulares especificas (granulocitos, linfocitos, etc.). El nombre particular de cada CSF refleja el tipo de colonia que cada uno de ellos genera. En este texto sólo se van comentar los CSF que son importantes para el desarrollo de los linfocitos (tabla 7). Los CSF que estimulan la maduración de otras líneas hematopoyéticas se tratan en libros de hematolo-gía.
Tabla 7 - Citocinas de la hematopoyesis
citocina Tamano (kD)
Origen Dianas Población inducida
SCF (c-kitL) 24 Células del estroma Stem cell Todas
IL-7 25 Fibroblastos Células del estroma Progenitores linfoides inmaduros Linfocitos T y B
IL-3 20-26 Células T Progenitores inmaduros Todas
GM-CSF 18-22
Células T Macrófagos Células endoteliales Fibroblastos
Progenitores inmaduros y compro-metidos Macrófagos maduros
Granulocitos Monolitos Activación de macróf
M-CSF 70-90 (D) 40 (M)
Macrófagos Células endoteliales Células méd. ósea Fibroblastos
Progenitores comprometidos Monocitos
G-CSF 19 Macrófagos Fibroblastos Células endoteliales
Progenitores comprometidos Granulocitos
Abreviaturas no indicadas en texto: c-kitl: Ligando de c-kit GM-CSF:Granulocyte-Monocyte CSF M-CSF: Monocyte CSF
G-CSF: Granulocyte CSF (D): Dímero (M): Monómero
1) Factor de la Stem Cell, SCF (Ligando de c-Kit)
Las ítem cell pluripotenciales expresan en su membrana un receptor que es una tirosin kinasa, pro-ducto proteico del encogen celular c-kit. La citocina que interactúa con este receptor se llama li-

Citocinas Página
29
gando de c-kit o factor de la stem cell (SCF) porque actúa en células stem cell inmaduras. A c-kit también se le llama receptor de la stem cell.
El SCF es sintetizado por las células del estroma de la médula ósea como una proteína que puede ser transmembrana o secretada, ambas producidas por splicing alternativo del RNA pero codifica-das por mismo gen. Se cree que el SCF es necesario para hacer a las células madre de la médula ósea sensibles a otros CSF pero que por si mismo no induce la formación de colonias (que es lo que precisamente indica su nombre). Puede también jugar algún papel en el mantenimiento de la viabilidad y la capacidad proliferativa de las células T en el timo y de las células cebadas en los te-jidos mucosos. La forma soluble del CSF está ausente en una cepa mutante de ratón llamada “s-teel". Los ratones steel tienen defectos de producción de células cebadas pero no de otras líneas lo que sugiere que la forma transmembrana es más importante que la soluble para el mantenimiento de varias líneas hematopoyéticas. La eliminación de ambas formas del CSF por knockout genético resulta letal.
2) lnterleucina-7 Es un citocina en hélice α secretada por las células del estroma de la médula ósea que estimula la supervivencia y la expansión de los precursores inmaduros comprometidos hacía las líneas de linfo-citos T y B. El receptor de la IL-7 consiste en una cadena α (que se une a la IL-7) unida a una cade-na γ señalizadora. Esta cadena γ es la misma que forma el receptor de la IL-2 y es en este receptor donde se identificó inicialmente esta cadena. Los ratones knockout para la IL-7, la cadena α de su receptor o la kinasa JACK3 que lleva asociada, son linfopénicos, con descenso del número de linfocitos T y B. Es probable que la inmunodeficiencia que se observa en ratones knockout para la cadena γ o para la kinasa asociada JAK3 (así como la inmunodeficiencia que se observa en humanos con mutaciones de estos genes) pueda ser el resul-tado de defectos de señalización vía IL-7. In vitro, la IL-7 es también un factor de crecimiento para las células T maduras, mimetizando la ac-ción de la IL-2. 3) lnterfeucina-3 También se le conoce como factor estimulante de colonias multilínea (multi-CSF) y es un producto de las células T CD4+ que actúa sobre los progenitores de médula inmaduros favoreciendo la ex-pansión de estas células, que se diferencian hacia todos los tipos de células maduras conocidos. La IL-3 es u miembro de la familia de receptores de citocinas con cuatro cadenas en hélice α. En los humanos receptor de la IL-3 está formado por un componente que se une a la IL-3 que es un miem-bro de la familia de receptores de tipo I y una subunidad transductora de señales que también está presente en receptor de la IL-5 y en el de GM-CSF. En el ratón la subunidad transductora de señales es típica de la IL-3. En ambas especies la transducción de la señal se hace por la vía JAK/STAT. La mayoría de los análisis funcionales sobre IL-3 se han llevado a cabo en ratones. Además de sus efectos sobre múltiples líneas celulares, la IL-3 induce el crecimiento y la diferenciación de progenito-res medulares hacia células cebadas, una acción reforzada por la IL-4. Sin embargo, el knockout del receptor de IL-3 o de su subunidad transductora de señales, en ratones, no causa alteraciones nota-bles de la hematopoyesis. La IL-3 humana se ha identificado por clonación del DNA complementario de una molécula homóloga a la IL-3 de ratón, pero ha sido difícil establecer la función de esta citoci-na en la hematopoyesis humana. De hecho, muchas de las acciones atribuidas a la IL-3 en el ratón parecen ser llevadas a cabo en el humano por GM-CSF. 4) GM-CSF (Granulocyte-Macrophage Colony Stimulating Factor) 5) M-CSF (Monocyte Colony Stimulating Factor)

Citocinas Página
30
6) G-CSF (Granulocyte-Colony Stimulating Factor) Son citocinas producidas por los linfocitos T activados, los macrófagos, las células endoteliales y las células del estroma de la médula ósea que actúan sobre los progenitores de la médula ósea para au-mentar la producción de leucocitos. El G-CSF es producido en los focos de infección y actúa como una hormona para movilizar los neu-trófilos de la médula ósea para reemplazar a los consumidos en las reacciones inflamatorias. GM-CSF es también un activador de macrófagos y activa la diferenciación de células de Langerhans a células dendríticas. Cada vez se usa más el GM-CSF recombinante en la clínica para facilitar la recuperación de la médula ósea después del tratamiento del cáncer y en los trasplantados de médu-la ósea. Este quizá sea el mejor ejemplo del uso clínico de las citocinas. 7) lnterleucina-9 Es una citocina que facilita el crecimiento de algunas líneas celulares T y de los progenitores de célu-las cebadas en la médula ósea. La IL-9 usa la cadena γ como señalizadora y la vía JAK/STAT. Su función fisiológica es desconocida. 8) lnterleucina-11 Es producida por las células del estroma de la médula ósea. Usa la misma subunidad de señaliza-ción que la IL-6, la gp130 y señaliza por la vía JAK/STAT. Estimula la megacariocitopoyesis y se usa en clínica para tratar a pacientes con cifras insuficientes de plaquetas como resultado del tratamiento antineoplásico.





























