Embed Size (px)
Citation preview

AFARI• EXPEDICIÓN AL MUNDO VETERINARIO •
AFARI• UNA PUBLICACIÓN DE LABORATORIOS CHALVER DIVISIÓN VETERINARIA • VOLUMEN 14 •
Co
lom
bia
• E
cu
ad
or •
Pa
na
má
• G
ua
te
ma
la •
Re
pú
bli
ca
Do
min
ica
na
• H
on
du
ra
s •
El
Sa
lva
do
r

Nota Del DirectorLa utilización de las diferentes ténicas para mejorar los índices reproductivos en los bovinos, se ha convertido en una estrategia para aumentar la productividad de los hatos en el mundo; actualmente se dispone de biotegnologías como la inseminación artificial a tiempo fijo, la trasferencia de embriones, el sexaje de semen, la aspiración folicular, la fertilización in vitro y la clonación. Estos procedimientos han contribuido a mejorar los parámetros reproductivos - productivos, haciendo cada vez más eficiente y competitiva la industria ganadera.
Así como es de importante analizar el entorno medio ambiental y las características genéticas de los animales buscando incrementar su producción, resulta relevante tener un conocimiento de los mecanismos fisiológicos que enmarcan el ciclo estral y los últimos avances en el entendimiento de la dinámica folicular, a fin de que el tratamiento farmacológico que se aplique para controlar el ciclo estral tenga un manejo razonable, y no se convierta en una prescripción sin sentido o simplemente en una "receta de cocina"
En está edición queremos presentar los últimos avances en el entendimiento de la dinámica folicular bovina y algunos tratamientos farmacológicos para el control del ciclo estral.
No olvide estimado lector “reproducir para producir”.
Cordial Saludo:
Leonardo Escobar Guevara M.V.Z - Esp.Gerente de MercadeoLaboratorios Chalver de Colombia S.A.
La dinámica folicular ha sido estudiada desde mediados del siglo XX, su mayor desarrollo lo ha alcanzado en las últimas dos décadas gracias a la utilización de herramientas como la ultrasonografía, este procedimiento permite estudiar el efecto de diversos tratamientos hormonales y su acción en la dinámica folicular.
La dinámica folicular comprende la ovogénesis y foliculogénesis, estos procesos en conjunto abarcan la formación y desarrollo del ovocito y del folículo respectivamente. Diversos estudios realizados coinciden en afirmar que los ovarios actúan como una sola unidad, e influyen en el desarrollo folicular por medio de vías endocrinas y sistémicas; involucrando las gonadotropinas, productos ováricos y uterinos.
3
AFARI• EXPEDICIÓN AL MUNDO VETERINARIO •
AFARIPublicación trimestral de
Laboratorios Chalver División Veterinaria
DirecciónLeonardo Escobar Guevara M.V.Z - Esp.
Gerente de Mercadeo
RedacciónLina Xiomara Ladino Medina M.V.Z - Esp.
Directora CientíficaLeonardo Escobar Guevara M.V.Z - Esp.
Gerente de Mercadeo
DiseñoChalver Publicidad
Impresión y Pre-prensaChalver S.A. Zona Franca
Para la inserción de temas o el envíode sugerencias y comentarios,
contáctenos en:Carrera 106 #15-25 Mz.6 Int.28 y 29
PBX: 7245090 / Ext: 4264Fax: 7245090 Ext. 4197
Laboratorios Chalver División VeterinariaA.A. 21424
e-mail: [email protected]
Línearmonal
ChalverReproducir para producir
DINÁMICA FOLICULAR

conectan entre sí por medio de puentes i n t e r c e l u l a r e s , formándose finalmente ovogonias. Las ovogonias se dividen por mitosis, dicho proceso es especie-específica.Una vez cesa el proceso de división mitótica, las ovogonias quedan incluidas en los cordones germinales; en este proceso las ovogonias se diferencian en ovocitos. La meiosis de la ovogonia comienza hacia los 75 a 80 días de gestación en el ganado bovino y la primera división meiótica no avanza más allá de la etapa del paquiteno de la profase-I, momento en el cual los cromosomas están descondensados y contenidos dentro de la membrana nuclear de la vesícula germinal (Adams, 2008).
5
El modelo teórico propuesto anteriormente fue confirmado con el uso de la ultrasonografía. El seguimiento ecográfico de los folículos durante el ciclo permitió establecer que la población de folículos que emergen presenta una fase de crecimiento, seguida de una fase estática y finalmente ovulan o sufren atresia. Por tanto el desarrollo folicular de la vaca abarca las etapas de reclutamiento, selección y dominancia. El crecimiento folicular en el ganado ocurre de manera ondulante y la mayoría de los ciclos estrales en el ganado constan de dos o tres ondas (Adams et al, 2008; Melquisedec, 2004).
La ovogénesis es la secuencia de fenómenos por los cuales las células germinales primordiales (ovogonias), se transforman en ovocitos maduros (óvulos). El crecimiento folicular es parte de un proceso más amplio llamado foliculogénesis, el cual comprende la formación, crecimiento y diferenciación folicular. Abarca desde el estadío de folículos primordiales hasta el folículo preovulatorio.
Los ovocitos se originan de las células germinales primordiales en el embrión, específicamente en el endodermo del saco vitelino embrionario y migran por medio de movimientos a través del mesenterio dorsal hasta la cresta gonadal del mesonefro, este proceso se desarrolla hacia el día 35 de gestación en el ganado. Dichas células se
De acuerdo con la literatura en el siglo V D.C. Hipócrates no le atribuyó ningún papel generativo al ovario, Él afirmaba que la generación de
una nueva vida era el resultado de la acción del semen del macho (eyaculación) y el de la hembra (sangre menstrual). Aristóteles, un siglo después, caracterizó el ovario como un vestigio imperfecto del testículo del macho, el cual no tenía una función aparente. Solo hasta mediados del siglo 17 la función ovárica de producir ovocitos fue reconocida. En 1827, un médico de Estonia, Karl Ernst von Baer, hizo la primera descripción del óvulo de un mamífero, a partir del estudio microscópico de las vesículas ováricas o folículos, en ovarios de una perra (Adams et al, 2008).
Los primeros estudios de la dinámica y del desarrollo folicular no se repitieron sino hasta después de 100 años. Los conocimientos sobre desarrollo folicular
eran deducidos indirectamente a través del estudio de patrones cíclicos de hormonas como las gonadotropinas y esteroides, también de forma experimental utilizando técnicas de medida y recuento de folículos en ovarios recolectados en mataderos.
Gracias a la utilización de la ultrasonografía, en el ganado vacuno se ha conseguido establecer los patrones de
crecimiento folicular en hembras en diferentes estadíos reproductivos: animales prepúberes con 2 semanas de edad,
novillas, vacas posparto, vacas gestantes, vacas anovulatorias y vacas que han sido tratadas para la sincronización del celo.(Adams et al, 2008; Marín et al, 2004).
los estudios de la dinámica folicular se hace referencia al término de ondas foliculares. Al principio se creía que el crecimiento folicular era continuo e independiente del ciclo, pero Rajakoski en 1960, basado en el estudio de ovarios recuperados en el matadero, propuso que los patrones de crecimiento y regresión folicular en cada individuo y en forma total de la población, no se desarrollaban durante todo el ciclo, sino que se relacionaba en el momento del día de éste. (Melquisedec, 2004; Alves et al, 2003).
Reseña Historica
Ovogénesis y foliculogénesis
4Saco vitelino. La gráfica ilustra la ubicación de las células germinales primordiales (naranja) (a) y los movimientos migratorios de las células germinales primordiales a lo largo del mesenterio dorsal hasta la cresta gonadal (b). (Fuente: Embryo Images On line).
Corazón
Intestino anterior
Intestino posterior
Alanfoides
Celulas germinalesprimordiales
Saco vitelino
Cloaca Mesonefro
Intestinoposterior
Cresta genital
Cresta genital

6 7
AFARIAFARI
Los ovocitos que logran alcanzar el estadío de folículo primordial quedan detenidos en la profase I. Una capa única de células epiteliales se aglomera alrededor de la inmensa mayoría de los ovocitos que sobreviven y los circunda formando los folículos primarios. Los ovocitos que no pueden ser rodeados por las células epiteliales se degeneran (Adams et al, 2008; Gigli et al, 2006).
La cantidad de folículos primordiales en bovinos de acuerdo con la literatura, varían desde 42.000 a 325.000, durante el período previo a la pubertad los folículos pasan por algunas fases de desarrollo hasta que sufren atresia. No existe relación entre el número de folículos y el tamaño de la camada así como la duración de la vida reproductiva en los animales (Gigli et al, 2006).
El crecimiento del folículo primario se basa en el aumento de tamaño del óvulo, cambios en la estructura y como se menciono anteriormente en la multiplicación de células de la granulosa.
La emergencia de ondas foliculares en el ganado se caracteriza por el crecimiento repentino (en 2 a 3 días) de folículos primordiales, algunos autores reportan de 100 a 300 según la especie (Engelhardt, 2002), hoy en día se sabe que en bovinos unos 8 a 41 folículos primordiales son reclutados y poseen un diámetro de 3 a 4 milímetros de acuerdo con los estudios ecográficos (Adams et al, 2008).
Una vez iniciado este proceso, el cual es irreversible, su crecimiento terminará en atresia o en ovulación.
AFARIAFARILa activación del crecimiento folicular comienza con la transformación de las células aplanadas del folículo primario en una capa única de células granulosas cuboidales. La proliferación de las células de la granulosa produce un incremento de 2 a 6 capas alrededor del ovocito, denominado folículo secundario, en este folículo las células tecales se diferencian en una capa externa y otra interna rodeando por fuera a las células de la granulosa. Estos folículos son conocidos como preantrales debido a la ausencia de cavidad antral (Adams et al, 2008; Gigli et al, 2006).
La proliferación de las células de la granulosa puede ser de más de 6 capas y la formación de una cavidad o antrum llena de líquido folicular constituyen el folículo terciario o antral. Estos foliculos poseen una mayor expresión de receptores para FSH en las células de la granulosa, mientras que las células de la teca expresan más receptores para LH. Existe a su vez una clasificación de estos folículos en dominantes y subordinados; la diferencia radica en la existencia de receptores para LH en los folículos dominantes a excepción de los subordinados, que solo poseen para FSH (Adams et al, 2008; Gigli et al, 2006).
Los folículos preovulatorios o De Graaf, denominados así gracias a las investigaciones realizadas por el médico Holandés, Regnier de Graaf, en 1672, donde reconoció el verdadero papel del ovario en su publicación titulada "Nuevo tratado referente a los órganos generativos de las mujeres", concluyó que estos folículos poseen la capacidad de responder al estímulo de la LH, hormona gonadotrópica encargada de desencadenar cambios morfológicos y bioquímicos que activarán la meiosis y desen-cadenaran el proceso de ovulación. El tamaño del folículo preovulatorio varía entre las especies, por ejemplo en la vaca es de 10 a 20 mm mientras que en la yegua es mayor a 35 mm (Adams et al, 2008; Gigli et al, 2006).
Células de la teca externa
Láminabasal
Célulasde lagranulosa
Células de la teca interna
Célulasque seconvierten en lacorona radiadadespués de laovulación
Amtrum llenode líquidofolicular
Cumulus oophorus

9
Dinámica folicular durante el ciclo estral
Ondas foliculares
Los estudios realizados utilizando imágenes de ecografías para monitorear las poblaciones foliculares de diferentes
tamaños, o para monitorear folículos identificados en forma individual; han demostrado categóricamente que el crecimiento folicular en el ganado ocurre de una manera ondulante, y que la mayoría de ciclos estrales en el ganado constan de dos ó tres de estas ondas; este patrón de ondas se repite además del ciclo estral, en la etapa prepúber, la preñez y en el posparto (Adams et al, 2008; Gigli, 2006 y Bo, 2002).
La emergencia de las ondas foliculares en el ganado se caracteriza por el crecimiento repentino (en 2-3 días) de 8 a 41 folículos pequeños, estos son detectados inicialmente por medio de la ultrasonografía, con un diámetro de 3 a 4 milímetros. (Referido en Adams, 1999; Adams et al, 1995).
La tasa de crecimiento es similar entre los folículos de la onda durante aproximadamente 2 días, cuando un folículo es seleccionado para continuar su crecimiento (Folículo Dominante) mientras que los demás se vuelven atrésicos y se degeneran (Folículos subordinados). Los resultados de los estudios de la dinámica folicular temprana dieron lugar a la hipótesis de que el folículo dominante inhibe el crecimiento de los subordinados en la onda existente; y la emergencia de la siguiente onda folicular. Esta hipótesis fue respaldada por una serie de estudios que implican el tratamiento sistémico con la fracción proteíca del fluido folicular y por la electrocauterización del folículo dominante (Referido en Adams, 1992).
La utilidad práctica de estos hallazgos fue inmediata y tuvieron gran repercusión, marcando una nueva era en la sincronización y la superestimulación ovárica del ganado (Bo, 1995; Adams et al, 1998).

Dos versus tres patrones de ondaLa mayoría de los ciclos reproductivos estrales de los bovinos (más de un 95%) están compuestos por dos o tres ondas foliculares (Referido en Adams, 1998). Tanto en los ciclos estrales de dos ondas como en los de tres, la aparición de la primera onda folicular se produce invariablemente en el día de la ovulación (Día 0). La aparición de la segunda onda se produce en el Día 9 ó 10 en los ciclos de dos ondas y en el Día 8 ó 9 en los ciclos de tres ondas. En los ciclos de tres ondas, aparece una tercera onda en el Día 15 ó 16. Bajo la influencia de la progesterona (por ejemplo, en el diestro) los folículos dominantes
de las ondas sucesivas experimentan atresia. El folículo dominante presente al comienzo de la luteólisis se convierte en el folículo ovulatorio, y la aparición de la siguiente onda se retrasa hasta el día de la subsiguiente ovulación. El CL comienza a degenerarse más temprano en los ciclos de dos ondas (Día 16) que en los de tres ondas (Día 19), dando lugar a un ciclo estral igualmente más corto (19 -20 días versus 22-23 días). Por lo tanto, el así llamado ciclo estral de 21 días en el ganado existe únicamente como un promedio de los ciclos de dos y de tres ondas (Figura 1). Los factores predictivos asociados a un patrón de dos ondas versus uno de tres ondas pueden aportar una idea
de los mecanismos que controlan el patrón, y tienen implicaciones importantes en
el manejo de la crianza y en el desarrollo de
protocolos efectivos de sincronización ovárica. Las tasas de preñez en el ganado con patrones de dos ondas versus las de patrones de tres ondas fueron comparadas basándose en la noción de que el folículo preovulatorio en el patrón de dos ondas crece durante un período relativamente más prolongado y puede contener un ovocito relativamente envejecido. Sin embargo, los resultados han sido contradictorios; las tazas de preñez no fueron diferentes entre los ciclos de dos ondas versus los de tres ondas en algunos estudios (Ahmad et al, 1997; Bleach et al, 2004), mientras que en otro estudio se reportó que las tasas de preñez fueron mas bajas en los ciclos de dos ondas (Townson et al, 2002). Parece que no existe una predilección específica en cuanto a la raza o la edad para determinado patrón de onda en el ganado de la especie B. taurus; un aumento de la proporción de los patrones de tres ondas ha sido asociado con un bajo nivel nutricional y estrés por calor. (Referido Adams, 1998; Bo et al 2003).
En el ganado de la especie B. Indicus, no se detectó ningún efecto estacional sobre el patrón de las ondas, pero el patrón estuvo influenciado por la paridad. (Referido en Bo et al, 2003)
4
68
10
12
14
16
0 2 4 6 8 10 12 14 16 18 0
0.5
1.510
20
30
40
FSH
(ng/
mL)
LH (n
g/m
L)
Días después de la ovulación
Diam
etro
folic
ular
(mm
)
FSH
LH
Ov OvFase Luteal
Intervalo interovulario de 2 ondas
Intervalo interovulatorio de 3 ondas
4
68
10
12
14
16
0 2 4 6 8 10 12 14 16 18 20
0.5
1.510
20
30
FSH
(ng/
mL)
LH (n
g/m
L)
Días después de la ovulación
Diam
etro
folic
ular
(mm
)
0
Ov OvFase Luteal
10 11
AFARIAFARI
Figura 1. Los folículos dominantes y los subordinados están representados en círculos vacíos (viables) o sombreados (atrésicos). Un incremento en las concentraciones de FSH circulante (línea de color azul) precede a la aparición de cada onda. Un incremento en las concentraciones de LH circulante (línea color rojo) precede a la ovulación. El incremento de LH es precedido y sucedido por un período de alta frecuencia de pulsos de LH como resultado de las bajas concentraciones de progesterona circulante (es decir, períodos de luteólisis y luteogénesis, respectivamente) (Fuente: Adams et al, 2008)

La mayoría de las novillas Nelore (65%) exhibieron un patrón de tres ondas, mientras que la mayoría de las vacas Nelore (85%) presentaron un patrón de dos ondas. Otros han informado que hasta un 27% de los ciclos estrales en las vacas B. Indicus constaban de cuatro ondas de desarrollo folicular, comparados con un 7% en las novillas B. Indicus (Referido en Bo et al, 2003). En un estudio reciente en el cual fueron incluidos los datos ultrasonográficos de 91 intervalos interovulatorios, (Jaiswal et al,2006) se compararon los patrones de dos y tres ondas de desarrollo folicular para determinar las características de repetibilidad y predictibilidad de un patrón de onda estipulado. Los ciclos de dos ondas fueron casi 3 días más cortos que los ciclos de tres ondas (19,8 ± 0,2 versus 22,5 ± 0,3; P < 0,01). La mayoría de los ciclos <21 días (88%) tuvieron un patrón de dos ondas (P < 0,05), mientras que la mayoría de los ciclos >22 días (78%) tuvieron un patrón de tres ondas (P < 0,05). La proporción de ciclos en serie en los cuales el patrón siguió siendo el mismo (es decir, repetibilidad) fue más de dos veces mayor que la de los ciclos en los cuales cambiaron los patrones (70% versus 30%; P <0,01). La repetibilidad del patrón de onda, y la proporción de los patrones de dos ondas versus los de tres ondas en el hato no se vieron afectadas por la época del año. La correlación más fuerte para el número de ondas en un intervalo interovulatorio fue la duración de la dominancia folicular de la Onda 1. La duración de la dominancia (definida como el período de la fase de crecimiento y la fase estática del folículo dominante) fue 3 días más prolongada y el inicio de la regresión fue mas tardío en los patrones de dos ondas que en los de tres ondas (P < 0,01). La dominancia de la Onda 1 se asoció con un retraso subsiguiente en el logro del diámetro máximo por el folículo dominante de la Onda 2, así como un inicio temprano de la luteólisis. Por lo tanto, los factores que influyen en el desarrollo del folículo dominante de la Onda 1 pueden ser los responsables de la regulación del patrón de las ondas. (Adams et al, 2008)
Ovarios funcionales. La fotografía ilustra la funcionalidad bilateral de los ovarios durante la selección del folículo dominante. (Fuente: www.learningradiology.com)
Los dos ovarios actúan primordialmente como una sola unidad; es decir, cada onda folicular incluye folículos de ambos ovarios, que responden al unísono. En un estudio crítico de las relaciones intraováricas, (Ginther et al, 1989) los autores concluyeron que el folículo dominante, destruye a los subordinados y a la aparición de ondas nuevas por vía sistémica (endocrina) y no por canales locales. Solamente uno de los folículos del par de ovarios es seleccionado para volverse el dominante; el lado en el cual se desarrolla el folículo dominante fue aleatorio y el folículo dominante tiene la misma probabilidad de alojarse en el mismo ovario o en el ovario contralateral al del folículo subordinado más grande.
Además el lado del CL o el folículo dominante de una onda anterior no tiene efecto sobre el lado del folículo ovulatorio.
Aunque los factores foliculares (autocrinos y paracrinos) son importantes para el crecimiento, la integridad y la destrucción de un folículo individual, no existe una documentación concluyente in vivo de que un folículo afecte directamente el estado de fortaleza/regresión de los demás foliculos de la cohorte a través de un efecto localizado. (Adams et al, 2008)
Interacción de las hormonas que controlan la dinámica de la onda, la ovulación y el desarrollo del cuerpo lúteo
Control local vs. control sistémico
12

14
Las gonadotropinas y sus receptores
La aparición de una onda folicular y la selección del folículo dominante están asociadas temporalmente a un aumento y una caída de las concentraciones circulantes de FSH (Fig. 1). (Adams et al, 1992)
La aparición de una onda folicular está precedida por un incremento repentino de las concentraciones plasmáticas de FSH, tanto en las ondas espontáneas como en las inducidas. Los productos foliculares, especialmente los del folículo dominante, son los responsables de la supresión de la liberación de FSH, y por consiguiente, de la aparición de la siguiente onda folicular (Fig. 1). Al final del período de dominancia (es decir, en la ovulación, o la fase estática media de un folículo dominante anovulatorio), las concentraciones de FSH circulante comienzan a elevarse; éstas aumentan 1,5 - 2 veces durante los 2 días siguientes y alcanzan el pico
máximo aproximadamente entre las 12 y las 24 horas anteriores a la emergencia de la onda (cuando el futuro folículo dominante tiene un diámetro de 4-5 mm). Cuando un folículo dominante es removido (es decir, ablación folicular), se inicia un incremento repentino de FSH en el término de 12 horas, dando como resultado la aparición de una nueva onda folicular dentro de las 24 horas siguientes (Bergfelt et al, 1994). La selección del folículo dominante se asocia a concentraciones sanguíneas decrecientes de FSH durante los tres primeros días de la onda. El nadir de la FSH se alcanza cuatro días después de la aparición de la onda y las concentraciones continúan siendo bajas durante los siguientes 2-3 días. Los receptores de FSH solamente están presentes en las células granulosas, mientras que los receptores de LH se encuentran localizados tanto en las células granulosas como en las tecales en la pared de los folículos antrales. El folículo dominante adquiere más receptores de LH sobre sus células granulosas que sus subordinados y por lo tanto
puede sustituir su dependencia a la gonadotropina (FSH)
por la dependencia a la
LH durante el nadir de FSH y continuar creciendo mientras se degeneran los subordinados.
Productos ováricos y uterinos
El CL es la principal fuente de progesterona; la morfología del CL y la concentración de progesterona en el plasma son buenos indicadores de la síntesis de progesterona dentro del CL (Singh et al, 2003). La angiogénesis intensa, la proliferación de las células de la granulosa y la teca de la pared del folículo que ha ovulado y su diferenciación (luteinización) durante los primeros 5-6 días después de la ovulación (metestro), originan un aumento progresivo de la concentración de progesterona en el plasma, desde 1 ng/mL a los 3 días después de la ovulación hasta aproximadamente 3 ng/mL a los 6 días. La concentración de progesterona en el plasma alcanza su pico máximo entre los días 10 y 14 pos-ovulación (> 4ng/mL), seguida por un descenso de la concentración después de 16 días debido a la luteólisis (muerte de las células hipóxicas como resultado de la hialinización de los vasos sanguíneos), inducida por la PGF2α liberada por el endometrio de la vaca no gestante. Los folículos dominantes, tanto de las ondas anovulatorias como de las ovulatorias, producen estradiol. Las células de la teca son necesarias para la conversión de la progesterona en andrógenos, mientras que la enzima aromatasa (que convierte el andrógeno en estradiol) está localizada exclusivamente en las células de la granulosa. Después de la aparición de la onda, el contenido de estradiol en el líquido folicular del folículo dominante en crecimiento aumenta por lo menos 20 veces en el día de la selección (3 días después de la aparición de la onda) y le sigue una disminución de tres veces hacia la fase estática del folículo dominante anovulatorio (6 días) antes de retornar a la línea basal en la fase regresiva temprana (11 días) (Singh et al, 2003). La concentración pico
de estradiol en el líquido folicular del folículo ovulatorio es dos veces más alta que la del pico en los folículos dominantes anovulatorios. El tratamiento con estradiol exógeno durante la fase luteínica induce la desaparición del folículo dominante existente, muy probablemente por la supresión de LH y FSH, seguida de un efecto de rebote de la FSH y una aparición sincrónica de una nueva onda folicular (Adams, 1999; Bo et al, 1995). Este tratamiento es la base de muchos protocolos actuales para la sincronización de la ovulación para IA (inseminación artificial) a tiempo fijo y la hiperestimulación. Además del estradiol, que tiene una importante acción inhibitoria sobre la FSH, los folículos en crecimiento producen otros factores tales como IGFs (factor de crecimiento de tipo insulínico), inhibinas y folistatina (Singh et al, 1999), que también regulan la liberación de la FSH y su disponibilidad. Aunque el folículo dominante desempeña un papel importante, todos los folículos de una onda emergente contribuyen a la supresión del incremento repentino de FSH que da origen a la onda (Ginther et al, 2000). Tanto la liberación de la FSH como la de LH son inducidas por pulsos de GnRH en el hipotálamo, pero debido a que la liberación de FSH está profundamente influenciada por los productos foliculares y a que su vida media en el ganado es más prolongada que la de la LH, la liberación episódica de FSH es menos aparente que la de LH. La frecuencia y la amplitud de los pulsos de LH son influenciadas por las concentraciones circulantes de progesterona y estradiol. Los altos niveles de progesterona producidos por un CL funcional durante el diestro o la gestación inhiben la frecuencia de los pulsos de LH (Fig. 1).
15

16 17
Dinámica de los folículos pequeños
aquellos obtenidos de los folículos en crecimiento
(Día 2), en la fase estática temprana (Día 3) o en la fase tardía regresiva (Día
7). Interesantemente, los valores en pixeles en la escala de grises en las imágenes por
ultrasonido del estroma perifolicular fueron inferiores en los ovarios de los cuales fueron recogidos
los ovocitos que no produjeron embriones, comparados con aquellos que sí produjeron embriones (Vassena et al,
2003). Recientemente se hizo una comparación de la capacidad de desarrollo de los ovocitos que se desarrollaron con
concentraciones de progesterona bajas versus concentraciones altas durante la hiperestimulación con FSH (Jaiswal et al, 2006), Los
ovocitos que se desarrollaron en un medio rico en progesterona (es decir, baja frecuencia de pulsos de LH) tuvieron menor capacidad de
originar embriones. Interesantemente, los ovocitos que se desarrollaron en un bajo nivel de progesterona durante un período prolongado, (es decir,
actuando como folículos persistentes de gran tamaño) tuvieron una ovulación fracasada. Estos hallazgos les han dado ímpetu a estudios posteriores para
determinar si la exposición a la progesterona a corto plazo (es decir, alta frecuencia de pulsos de LH) durante la fase de crecimiento del desarrollo
folicular después de la hiperestimulación con FSH pueden ser utilizados para acrecentar las proporciones de embriones transferibles in vivo e in
vitro.
Capacidad de respuesta a la FSHMientras que la dinámica de desarrollo de los folículos
4 mm ha sido bien caracterizada, la dinámica de los folículos más pequeños sigue siendo un misterio. En un experimento clásico que involucraba la medición del índice mitótico de las células de la granulosa antes y después del tratamiento con colchicina (Lussier et al, 1987), se observó que los folículos pequeños crecen más lentamente (27 días para crecer de 0,13 mm a 0,67 mm) que los folículos más grandes (6,8 días para crecer de 0,68 mm
Cuando las concentraciones de progesterona en el plasma eran de aproximadamente 1,7 ng/mL, se observaron seis pulsos de LH en 8 horas, con una amplitud de 0,2 ng/mL; mientras que solamente se observaron 1,8 pulsos en 8 horas, con una amplitud promedio de 0,34 ng/mL cuando las concentraciones de progesterona eran de 5 ng/mL (Jaiswal, 2007). Por consiguiente, los folículos dominantes crecen más y continúan siendo dominantes durante un intervalo más prolongado cuando la frecuencia de los pulsos de LH se encuentra elevada (es decir, progesterona baja) (Referido en Adams, 1999). El aumento de las concentraciones de estradiol y la disminución de la progesterona después de la luteólisis aumentan aún más la frecuencia de los pulsos de LH, dando como resultado un gran incremento preovulatorio de LH.
Competencia de los ovocitos y estado folicularLos cambios morfológicos y celulares durante el desarrollo folicular y lúteo se reflejan en cambios bioquímicos a nivel tisular y plasmático, así como en las características de las imágenes por ultrasonido y la capacidad de desarrollo del ovocito (Referido en Singh et al, 2003). Los análisis asistidos por computador de las imágenes ultrasónicas del los folículos dominantes y subordinados en diferentes fases de desarrollo demostraron que la heterogeneidad de los pixeles del antro y de la pared, y la nitidez de la interfaz fluido-pared estaban correlacionadas negativamente (P < 0,01), con el estradiol y la relación estradiol:progesterona en el líquido folicular (Singh, 2003). Histológicamente, el espesor de la pared de los folículos dominantes de la Onda 1 disminuyó en forma dramática durante la fase estática tardía (60mµ) y la fase regresiva (42mµ), al compararlo con la fase de crecimiento y la fase estática temprana (120mµ). Las células de la capa granulosa del folículo dominante de la Onda 1 se desprendieron y se degeneraron durante la fase estática tardía y fue revestido por células aplanadas parecidas a los fibroblastos durante la fase regresiva. La pared de los folículos subordinados fue más delgada y tuvo un índice mitótico más bajo que la del folículo dominante 3 y 6 días después de la aparición de la onda. El folículo dominante de la Onda 1 exhibió una evidencia histológica de atresia en el momento de la aparición de la Onda 2. Con base en estos datos, la dominancia folicular alcanza su punto máximo morfológica y bioquímicamente durante la fase media de crecimiento (3 días después de la aparición de la onda) y empieza a decrecer al comienzo de la fase estática (6 días después de la aparición de la onda). Existe una correlación positiva entre la atresia folicular temprana y la competencia del ovocito (Salamone et al 1999; Vassena et al, 2003). Una proporción significativamente mayor de complejos cúmulo-ovocito extraídos de los folículos subordinados durante la fase regresiva temprana se expandió, y una proporción significativamente mayor de ovocitos presentó evidencia de maduración nuclear que aquellos extraídos durante la fase de crecimiento y la fase estática temprana. Además, los ovocitos obtenidos de los folículos subordinados al comienzo de la regresión (Día 5 de la onda folicular) fueron más aptos para desarrollarse en embriones in vitro que

1
23
4
6
8
10
0 2 4 6 8 10 12 14 16 18 0Días después de la ovulación
Diam
etro
Folic
ular
(mm
)
Onda 1
Ov OvIntervalo interovulatorio de 2 ondas
12
14
16
Onda 2{ {18 19
Figura 2. Patrón de la onda folicular ovárica detectada en folículos tan pequeños como de 1 mm de diámetro (se muestra el patrón de dos ondas). Los folículos pequeños (1-3 mm), ilustran la aparición de la onda 2,5 días antes de lo que se detectaba anteriormente (es decir, de 4-5 mm). Note que la tasa de crecimiento del folículo destinado a convertirse en el folículo dominante (línea punteada) es similar a la de los otros en la onda hasta aproximadamente 5 días después de la aparición de la onda (comenzando a 1 mm), y que el folículo destinado a ser el dominante aventaja en tamaño a aquellos destinados a ser subordinados desde su detección más temprana (1 mm) (Fuente Adams, et al. 2008).
a 3,67 mm) y que se requirieron dos ciclos estrales para que los folículos preantrales alcanzaran el estado de madurez antral. Sin embargo, estas estimaciones no arrojan luces sobre la dinámica folicular en relación a los cambios de las concentraciones circulantes de gonadotropinas y la aparición de la onda folicular.
De manera similar, el papel de la FSH en la regulación del desarrollo ondulante de los folículos 4 mm ha sido definido claramente (Adams et al, 1992), mientras que el papel de la FSH en los folículos más pequeños, incluyendo los folículos preantrales y antrales pequeños, sigue siendo incierto. Un estudio temprano demostró la unión de la FSH a las células granulosas de los folículos con una sola capa de células granulosas (Richards et al, 1998), pero se ha argumentado que éstas no pueden ser acopladas al sistema del segundo mensajero de la adenilato ciclasa durante las etapas tempranas de la foliculogénesis y pueden ser no funcionales (Wandji et al, 1992). Sin embargo, el efecto promotor del crecimiento de la FSH sobre los folículos preantrales y antrales pequeños en el ganado producido in vitro (Itoh et al, 2002) e in vivo (Fricke et al, 1997; Tanaka et al, 2001) indican que la FSH desempeña un papel en el desarrollo de la etapa temprana de los folículos preantrales y antrales pequeños.
Si los folículos pequeños son sensibles a la FSH, es lógico postular que su dinámica de desarrollo sigue un patrón ondulante en respuesta a incrementos periódicos endógenos de FSH. Hasta las épocas recientes, la referencia a una onda folicular estaba limitada a los folículos 4 mm, basándose simplemente en el límite de resolución de los equipos de ultrasonido existentes. A nivel microscópico, no hay una distinción morfológica entre los folículos antrales en las etapas media y tardía < 4 mm y aquellos 4 mm (Braw-Tal et al, 1997; Lussier et al, 1987). A nivel celular, las dos categorías de tamaño de folículos no sólo expresan receptores de FSH, sino que tienen un nivel de expresión similar en cada célula de la granulosa (Bao et al, 1998) .
La aparición periódica de ondas de los folículos 4 mm en respuesta a incrementos periódicos de las concentraciones sanguíneas de FSH (Adams et al, 1992) y la consistencia en el número de folículos 2 mm (Singh et al, 2004), o 3 mm (Burns et al 2005), reclutados en ondas sucesivas, indican que los folículos pueden ser arrastrados progresivamente a las ondas desde las primeras etapas del desarrollo.
Aparición de la onda y dominancia folicularLa disponibilidad de nuevos escáneres de ultrasonido capaces de identificar estructuras tan pequeñas como de 1 mm permitió realizar un estudio diseñado para caracterizar el patrón de desarrollo de los folículos de 1-3 mm en el ganado, y para determinar la etapa en la cual el futuro folículo dominante alcanza primero una ventaja en tamaño entre sus cohortes (Jaiswal et al, 2004).
Los resultados revelaron un cambio con el transcurso de los días (P < 0,05) en el número de folículos de 1-3 mm, con un máximo (P < 0,05) 1 o 2 días antes de la apar ic ión de la onda def in ida convencionalmente (el primer folículo dominante detectado a 4 mm), seguido 3-4 días después por un máximo (P < 0,05) número de folículos 4 mm (Fig. 2).
El futuro folículo dominante fue identificado por primera vez a un diámetro de 1 mm y emergió 6-12 horas antes que el primer folículo subordinado (P < 0,01); (Fig. 2).
Después de la detección del folículo dominante a 1 mm (0 h), su diámetro fue mayor que el del primer
y el segundo folículo subordinado a las 24 horas (P = 0,04) y a las 12 horas (P = 0,01), respectivamente, cuando el folículo dominante medía 2,4 ± 0,17 mm y 1,7 ± 0,14 mm (Fig. 2).
La tasa de crecimiento del folículo dominante fue mayor que la del primer y el segundo folículo subordinado a las 120 horas (P = 0,03) y a la 108 horas (P = 0,02) respectivamente, cuando el folículo dominante medía 9,5 ± 0,30 mm y 8,8 ± 0,49 mm.
Los autores concluyen que:
1. Los folículos de 1-3 mm maduran en una forma ondulante en asociación con incrementos repentinos en las concentraciones plasmáticas de FSH.
2. Los folículos de 1-3 mm son exquisitamente sensibles a las elevaciones transitorias de la FSH (es decir, de menos de 6 horas).
3. La selección del folículo dominante es notoria más temprano de lo que se había documentado anteriormente y se caracteriza por una progresión jerárquica durante un período que comprende la totalidad de la sobrecarga de FSH (5 días).

ý••'5f
Ovulación - Cuerpo lúteopersistente Tipo IV
Dominancia y desviaciónseguida de atresia
No desviación
22 23
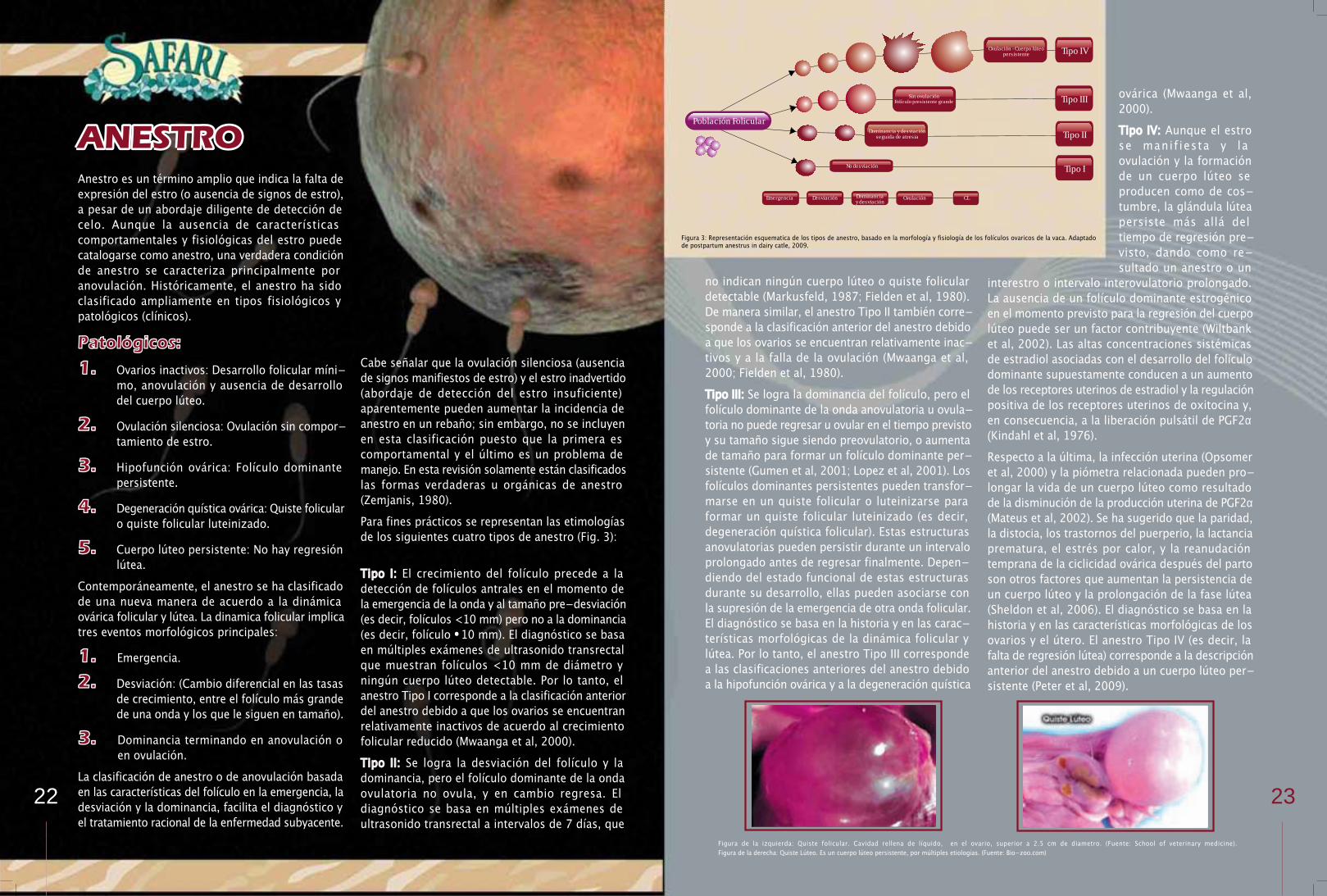
no indican ningún cuerpo lúteo o quiste folicular detectable (Markusfeld, 1987; Fielden et al, 1980). De manera similar, el anestro Tipo II también corresponde a la clasificación anterior del anestro debido a que los ovarios se encuentran relativamente inactivos y a la falla de la ovulación (Mwaanga et al, 2000; Fielden et al, 1980).
Tipo III: Se logra la dominancia del folículo, pero el folículo dominante de la onda anovulatoria u ovulatoria no puede regresar u ovular en el tiempo previsto y su tamaño sigue siendo preovulatorio, o aumenta de tamaño para formar un folículo dominante persistente (Gumen et al, 2001; Lopez et al, 2001). Los folículos dominantes persistentes pueden transformarse en un quiste folicular o luteinizarse para formar un quiste folicular luteinizado (es decir, degeneración quística folicular). Estas estructuras anovulatorias pueden persistir durante un intervalo prolongado antes de regresar finalmente. Depen-diendo del estado funcional de estas estructuras durante su desarrollo, ellas pueden asociarse con la supresión de la emergencia de otra onda folicular. El diagnóstico se basa en la historia y en las carac-terísticas morfológicas de la dinámica folicular y lútea. Por lo tanto, el anestro Tipo III corresponde a las clasificaciones anteriores del anestro debido a la hipofunción ovárica y a la degeneración quística
Cabe señalar que la ovulación silenciosa (ausencia de signos manifiestos de estro) y el estro inadvertido (abordaje de detección del estro insuficiente) aparentemente pueden aumentar la incidencia de anestro en un rebaño; sin embargo, no se incluyen en esta clasificación puesto que la primera es comportamental y el último es un problema de manejo. En esta revisión solamente están clasificados las formas verdaderas u orgánicas de anestro (Zemjanis, 1980).
Para fines prácticos se representan las etimologías de los siguientes cuatro tipos de anestro (Fig. 3):
Tipo I: El crecimiento del folículo precede a la detección de folículos antrales en el momento de la emergencia de la onda y al tamaño pre-desviación (es decir, folículos <10 mm) pero no a la dominancia (es decir, folículo • 10 mm). El diagnóstico se basa en múltiples exámenes de ultrasonido transrectal que muestran folículos <10 mm de diámetro y ningún cuerpo lúteo detectable. Por lo tanto, el anestro Tipo I corresponde a la clasificación anterior del anestro debido a que los ovarios se encuentran relativamente inactivos de acuerdo al crecimiento folicular reducido (Mwaanga et al, 2000).
Tipo II: Se logra la desviación del folículo y la dominancia, pero el folículo dominante de la onda ovulatoria no ovula, y en cambio regresa. El diagnóstico se basa en múltiples exámenes de ultrasonido transrectal a intervalos de 7 días, que
Anestro es un término amplio que indica la falta de expresión del estro (o ausencia de signos de estro), a pesar de un abordaje diligente de detección de celo. Aunque la ausencia de características comportamentales y fisiológicas del estro puede catalogarse como anestro, una verdadera condición de anestro se caracteriza principalmente por anovulación. Históricamente, el anestro ha sido clasificado ampliamente en tipos fisiológicos y patológicos (clínicos).
Patológicos:1. Ovarios inactivos: Desarrollo folicular míni
mo, anovulación y ausencia de desarrollo del cuerpo lúteo.
2. Ovulación silenciosa: Ovulación sin comportamiento de estro.
3. Hipofunción ovárica: Folículo dominante persistente.
4. Degeneración quística ovárica: Quiste folicular o quiste folicular luteinizado.
5. Cuerpo lúteo persistente: No hay regresión lútea.
Contemporáneamente, el anestro se ha clasificado de una nueva manera de acuerdo a la dinámica ovárica folicular y lútea. La dinamica folicular implica tres eventos morfológicos principales:
1. Emergencia.
2. Desviación: (Cambio diferencial en las tasas de crecimiento, entre el folículo más grande de una onda y los que le siguen en tamaño).
3. Dominancia terminando en anovulación o en ovulación.
La clasificación de anestro o de anovulación basada en las características del folículo en la emergencia, la desviación y la dominancia, facilita el diagnóstico y el tratamiento racional de la enfermedad subyacente.
ANESTROovárica (Mwaanga et al, 2000).
Tipo IV: Aunque el estro se manif iesta y la ovulación y la formación de un cuerpo lúteo se producen como de costumbre, la glándula lútea persiste más allá del tiempo de regresión previsto, dando como resultado un anestro o un
interestro o intervalo interovulatorio prolongado. La ausencia de un folículo dominante estrogénico en el momento previsto para la regresión del cuerpo lúteo puede ser un factor contribuyente (Wiltbank et al, 2002). Las altas concentraciones sistémicas de estradiol asociadas con el desarrollo del folículo dominante supuestamente conducen a un aumento de los receptores uterinos de estradiol y la regulación positiva de los receptores uterinos de oxitocina y, en consecuencia, a la liberación pulsátil de PGF2α (Kindahl et al, 1976).
Respecto a la última, la infección uterina (Opsomer et al, 2000) y la piómetra relacionada pueden prolongar la vida de un cuerpo lúteo como resultado de la disminución de la producción uterina de PGF2α (Mateus et al, 2002). Se ha sugerido que la paridad, la distocia, los trastornos del puerperio, la lactancia prematura, el estrés por calor, y la reanudación temprana de la ciclicidad ovárica después del parto son otros factores que aumentan la persistencia de un cuerpo lúteo y la prolongación de la fase lútea (Sheldon et al, 2006). El diagnóstico se basa en la historia y en las características morfológicas de los ovarios y el útero. El anestro Tipo IV (es decir, la falta de regresión lútea) corresponde a la descripción anterior del anestro debido a un cuerpo lúteo persistente (Peter et al, 2009).
Figura de la izquierda: Quiste folicular. Cavidad rellena de líquido, en el ovario, superior a 2.5 cm de diametro. (Fuente: School of veterinary medicine).Figura de la derecha: Quiste Lúteo. Es un cuerpo lúteo persistente, por múltiples etiologias. (Fuente: Bio-zoo.com)
Figura 3: Representación esquematica de los tipos de anestro, basado en la morfología y fisiología de los folículos ovaricos de la vaca. Adaptado de postpartum anestrus in dairy catle, 2009.
ANESTROTipo III
Tipo II
Tipo I
Población Folicular
Sin ovulaciónFolículo presistente grande
Emergencia Desviación Dominanciay desviación
Ovulación CL

Ovulación
Ovulación
Estro5 10 15 Estro
Reclutamiento Folicular
Coce
ntra
ción
Horm
onal
PGF2•
LH
E2
FolículoDominante
Atresia
Progesterona
FSH
24 25
Como es bien sabido cuando se manipulan las "oleadas" foliculares se mejora la fertilidad, ya que los folículos dominantes se hacen más uniformes en el tamaño y madurez en el hato. Para lograr este objetivo se usan dos métodos básicos:
1. Aplicación de una dosis ovulatoria de GnRH, la cual induce la liberación de LH, provocando la ovulación y luteinización de los folículos dominantes, y el crecimiento de una onda folicular a los 2-3 días (Aspron, 2004). Tiene como desventaja que se debe contar con folículos maduros, ya que los folículos en fase de reclutamiento no responden a la acción de la GnRH por carencia de receptores para LH.
2. Utilización del benzoato de estradiol o cipionato. Cuando hay dominancia progestacional (por acción de progestágenos endógenos o exógenos), el estradiol reduce la secreción de LH e induce atresia del folículo dominante y una nueva oleada folicular. (Bo et al, 1991; Bo, 1997) En caso contrario cuando no hay progestágenos, el estradiol induce la liberación de GnRH, causando la síntesis de LH,
En hatos con manejo intensivo se programan los celos e inseminaciones mediante manipulación hormonal del ciclo estral, buscando lograr fertilidad al primer servicio y en el caso de animales en anestro la inducción del celo fértil de manera sincronizada.
Los esquemas de sincronización de ovulaciones han sido desarrollados para resolver el problema de escasa manifestación del celo en época de estrés calórico y han dado buenos resultados en hatos con mala detección de calores.
Las hormonas que se usan para sincronizar el estro son idénticas a las hormonas reproductivas encontradas en el hipotálamo (GnRH), ovario (estradiol y progesterona) y útero (prostaglandina) o análogos de ellas. El primer método de sincronización se desarrolló en el año de 1960 con la utilización de progestágenos para bloquear la ovulación; el problema de este método fue la baja tasa de concepción debido al envejecimiento de los folículos (Bo, 2006).
Años después se desarrollaron protocolos a base de PGF2α, para lograr mejor eficiencia se combinaron los dos protocolos, sin embargo, un período muy prolongado de acción de los progestágenos deprime el porcentaje de concepción (Bo, 2006).
Con el estudio de la dinámica folicular se logro determinar que la persistencia del folículo dominante era la causa de la baja fertilidad, buscando mejorar ésto se desarrollaron programas de control folicular, a fin de controlar la fase lútea y el tiempo de ovulación. En la actualidad la mayoría de los métodos de sincronización utilizan:
- Control del crecimiento folicular.
- Prevención de la ovulación prematura y promoción de la ovulación en vacas en anestro.
- Inducción de la lisis del cuerpo lúteo en vacas que se encuentran en ciclo.
- Sincronización del celo o la ovulación al final del tratamiento o ambos.
MANEJO FARMACOLÓGICODEL CICLO ESTRAL BOVINO
provocando de este modo la ovulación y luteinización del folículo dominante. El estradiol actúa directamente sobre el folículo dominante para inducir atresia y al mismo tiempo inhibir la síntesis y liberación de FSH (Bo, 2002). También induce la regresión del cuerpo amarillo si se aplica al inicio del ciclo estral. Los estrógenos en soluciones oleosas para aplicación parenteral se absorben rápidamente, de forma general los estrógenos esterificados poseen absorción retardada después de su administración intramuscular, un éster es una cadena compuesta por átomos de carbono adheridos a la hormona esteroide matriz en la posición del carbono 17. El benzoato de estradiol es un éster sintético del estrógeno natural (estradiol). Posterior a su administración éstos ésteres son absorbidos e hidrolizados hacia el estradiol, estrógeno más activo.
En estudios realizados con el benzoato de estradiol, este suprime el desarrollo folicular cuando es administrado al momento de la inserción del implante combinado con la administración de 50 ó 100 mg, de progesterona vía imtramuscular en protocolos de superovulaciones (Bo et al, 1994). Bó y colaboradores (2007) reportan que de varias dosis evaluadas de benzoato de estradiol (1 mg, 2.5 mg y 5 mg), combinadas con progesterona, la más efectiva fue la de 2.5 mg , con un comienzo de una nueva onda folicular a los 3,9 días (con un rangode 3 a 4 días).
Principios de la sincronización de estros
Control del crecimientofolicular
Figura 4: Rol hormonal durante el ciclo estral en una hembra bovina con dos ondas foliculares.
Estrogenos Progesterona FSHR FSH LH

los efectos colaterales, como lo hace Zincrocel® (D (+) Cloprostenol 0.075mg/mL). En cualquier grupo de hembras bovinas a las que se les administra Zincrocel® (D (+) Cloprostenol 0.075mg/mL) con un intervalo de 11-14 días, responderán a la primera inyección las que se encuentran entre los días 5 y 17 del ciclo al inicio del tratamiento; mientras que en el momento de la segunda inyección la mayoría se encontrarán entre los días 8 y 11 del ciclo y responderán a la aplicación de Zincrocel® (D (+) Cloprostenol 0.075mg/mL). (Figura 5).
2627
Métodos de sincronización
Métodos con prostaglandinas (PGF2 )La posibilidad de controlar el ciclo estral bovino por aplicación de prostaglandina como agente luteolítico, abrió una nueva era en el campo del manejo reproductivo. La sincronización estral con prostaglandina F2 alfa se basa en que ésta destruye el CL con la consecuente presentación temprana del estro. En vista de que el 95% de esta prostaglandina se oxida casi de inmediato en los pulmones, y debido a sus efectos broncoconstrictores potentes, surgieron los análogos de esta hormona natural, con los que se incrementa el poder luteolítico y se disminuyen
Involución del cuerpo lúteo en vacas
Una sola inyección de PGF2α es eficaz para inducir la luteolisis a partir del día 5 a 7 del ciclo. La presentación del celo varía con el momento en que se aplica la prostaglandina o su análogo, por lo general el intervalo es corto cuando se aplica al inicio del ciclo en los días 7 a 9, o en una etapa más tardía 14 a 16, coincidiendo con la presentación de la primera y segunda oleada de crecimiento folicular. Si se aplicara la prostaglandina entre los días 10 y 12, la presentación del celo se reparte entre los tres y 10 días posteriores debido a que la primera oleada folicular se encuentra en proceso de atresia y aún está muy inmadura la segunda y no está cerca de un estado preovulatorio. Con la administración de dos dosis de prostaglandina se logra una mayor sincronía de celos con un intervalo de 11 a 14 días de diferencia o si se establece una sincronía folicular siete días antes de la inyección de prostaglandina, los celos se presentan 72 a 96 horas. (Bo et al, 2001).
Manifestación de celos y ovulación
El celo en las vacas será más evidente cuanto más estrógenos tanto exógenos como endógenos presente. Puede haber manifestación del celo sin ovulación, y una vaca con ovulación puede tener una fase lútea corta si se le aplican estrógenos únicamente. Esta demostrado que la combinación de estrógenos en una sincronización folicular utilizando progesterona mejora la tasa de concepción. Es por esto que en muchos hatos se recomienda incluir estrógenos al programa de sincronización, especialmente en aquellas vacas en anestro o anovulatorias al inicio del procedimiento. Se pueden mejorar los índices de fertilidad aplicando una dosis de GnRH ovulatoria, 48 horas después de la segunda dosis de PGF2α e inseminando a tiempo fijo. (Bo et al, 2007)
Figura 5: Sincronización de celos con PGF2α.

Rowson y Cooper informaron que más del 90% de las hembras que estaban ciclando respondían al tratamiento y presentaban celo entre 2 y 4 días después de la segunda inyección y destacaron la gran concentración de celos y rápida iniciación del mismo después de la segunda inyección con respecto a la primera. (Bo et al, 2007).
Scaramuzzi y sus colaboradores concluyeron que en la hembra bovina tratada con un análogo de PGF2α, el intervalo pos-inyección hasta el estro parecía estar relacionado con el tamaño y madurez del folículo ovárico mas grande no anéstrico presente en el momento de realizar el tratamiento o dicho de otra manera, el tiempo requerido por ese folículo para completar su desarrollo. Momont y Seguin en 1999 demostraron que la receptibilidad del CL a la PGF2α aumenta gradualmente a partir del día 6 del ciclo estral; de esta manera, para mejorar la sincronización con el tratamiento de 2 inyecciones de PGF2α se determina prolongar el intervalo entre dosis en vacas de 14 días, con lo cual se aumentaría el número de animales en fase sensible a la acción de la PGF2α. (Bo et al, 2007).
En resumen, se podría decir que la administración de una dosis luteolítica de Zincrocel® (D (+) Cloprostenol 0.075mg/mL), causará la regresión del cuerpo lúteo en la mayoría de los días del diestro,
por lo tanto inducirá el celo en mas de la mitad de los animales que están ciclando. Una inyección repetida 11-14 días mas tarde resultará en la sincronización de los estros del hato, debido a que en ese momento todas las vacas se encontrarán entre los día 6 y 16 del ciclo (Fig.5). No debe esperarse un 100% de respuesta, aún cuando todas las vacas del grupo tuvieran un CL al momento de la segunda inyección. Esto es así porque no todos los CL maduros son lisados por PGF2α, especialmente en vacas lecheras en lactancia. Por ejemplo, solo el 25% de las vacas que están en el día 6 responderán, 66% de las que están en el día 7 y menos del 100% de las que están en estadios óptimos del ciclo (día 8 en adelante) (Bo et al, 2007).
De acuerdo al protocolo inicial con 2 inyecciones de Zincrocel® (D (+) Cloprostenol 0.075mg/mL), se han propuesto modificaciones de este tratamiento:
- Alternativa 1: Se aplica Zincrocel® (D (+) Cloprostenol 0.075mg/mL) a todos los animales, inseminando los que se detecten en celo (am-pm) tras la primera inyección. Las hembras no detectadas en celo reciben una segunda inyección de Zincrocel® (D (+) Cloprostenol 0.075mg/mL) en 11 - 14 días y se someten a detección de celos e I. A. (Fig. 6)
Con este tratamiento entrarán en celo y serán inseminadas después de la primera dosis de Zincrocel® aproximadamente un 60 - 65% de las hembras que están ciclando. Este grupo está formado por las vacas que tenían un CL funcional y respondieron a la acción luteolítica del tratamiento y por aquellas que ya habían iniciado la lisis natural del cuerpo lúteo y presentaron celo espontáneamente. De modo que solo recibirán una segunda dosis las que en el día 0 tenían un CL en formación no sensible al Zincrocel® (D (+) Cloprostenol 0.075mg/mL), aquellas que estuvieran en anestro y a las que no se observaron en celo por errores en la detección.
Según resultados, en la utilización de este protocolo en novillas, se ha obtenido que el 65.6% se observan en celo durante los 7 días posteriores a la primera dosis, y un 63.6% de concepción como resultado del trabajo de IA. (Bo et al, 2007)
- Alternativa 2: Detección del celo e inseminar durante los primeros 6 días, las vacas que no entren en calor recibirán una inyección de Zincrocel® (D (+) Cloprostenol 0.075mg/mL), y posteriormente serán inseminadas a celo detectado durante los próximos 5 días. (Figura 7).
Este es un protocolo muy práctico y muy aceptado en ganaderías de carne. Se deben detectar celos durante 6 días (se pueden utilizar toros marcadores y diferenciar con algún sistema que nos permita distinguir cuales fueron las hembras que entraron en celo y fueron servidas). Si en el hato contamos con que todas
las vacas están ciclando; durante los días previos al servicio, entrarán en celo del 20% - 25% de las hembras (se encuentran en la etapa de lisis del CL, días 17 a 21 del ciclo). Durante el periodo de detección, 6 días previos al Zincrocel® (D (+) Cloprostenol 0.075mg/mL), más el día de la aplicación de la hormona las hembras que no respondieron por tener cuerpo lúteo en formación, alcanzan un estadío sensible a la hormona, de modo que al momento de la inyección todas las hembras no inseminadas y que están ciclando, tiene CL formado. (Bo et al, 2006). Con la utilización de estos protocolos y manejando una detección de celo eficaz (80%), podríamos obtener una tasa de concepción del 50% y la tasa de preñez estaría en 40%. (Bo et al, 2007)
Métodos con GnRh y prostaglandinas (PGF2 )Se puede lograr la sincronización de una oleada folicular y un celo posterior si se aplica GnRH y siete días después PGF2α. La inyección de GnRH induce la ovulación del folículo dominante y el posterior desarrollo de una nueva onda folicular 2 a 3 días después (Bo et al, 2007); posteriormente la inyección del análogo de prostaglandina 7 días después induce la luteólisis. De esta forma, dos o tres días después
de la inyección de la prostaglandina se presenta celo y la ovulación del nuevo folículo dominante. A pesar de que con este protocolo se observaban más estros sincrónicos; no se tenía la suficiente precisión al momento de la ovulación para realizar la inseminación artificial a tiempo fijo. La aplicación de una segunda dosis de GnRH, la cual causa la ovulación del folículo dominante 1.5 a 2 días después de la aplicación de la PGF2α, se está manejando para tener una mejor sincronía.(Bo et al, 2007). Hay que recalcar que se explicarán y presentarán varios tratamientos existentes. La idea no es indicar cual tratamiento es mejor que otro; creemos que todos los tratamientos presentados tienen su aplicación en distintas situaciones y ámbitos. Muchas veces por causas de manejo o costos, un tratamiento puede ser altamente beneficioso en una situación, mientras que puede no serlo en otra.
Cada profesional deberá decidir cuales son los tratamientos que más se adecúan a su situación específica. Lo que se busca desarrollar implementando un protocolo de Inseminación Artificial, es controlar el crecimiento de los folículos ováricos preovulatorios, la regresión del CL y la ovulación. El protocolo para sincronizar la ovulación se hizo conocido como "OVSYNCH" (Figura 8) (Bo et al, 2007).
28 29
Figura 6: Sincronización de celos con PGF2α. Alternativa 1.
Figura 7: Sincronización de celos con PGF2α. Alternativa 2.

La utilización de un tratamiento con Maxpren® (Acetato de Buserelina 0.0042mg/mL) u Ovulen® (Acetato de Gonadorelina), induce la ovulación del folículo dominante y el posterior desarrollo de una nueva onda folicular; la inyección de Zincrocel® (D (+) Cloprostenol 0.075mg/mL) 7 días después induce la regresión del CL; y para precisar la ovulación del folículo dominante se utiliza una segunda inyección de Maxpren® u Ovulen® 2 días después del Zincrocel® y 17-24 horas de la segunda inyección de Maxpren® u Ovulen® las vacas son inseminadas a tiempo fijo sin detectar celo (Fig. 8). Este sistema induce un estro más sincrónico que los animales tratados únicamente con PGF2α. De acuerdo con estudios realizados, el porcentaje de gestación es del 50% sin detección de estros (Bo et al, 2007). La anterior información se utilizó para desarrollar programas de manejo reproductivo para vacas en lactancia, ya que en estudios realizados se obtuvo una curiosa respuesta ovulatoria a la GnRH, observándose una respuesta diferencial entre vacas y novillas; en donde el 90% de las vacas en lactancia ovularon y solamente el 54% de las novillas lo hicieron. De allí la conclusión de que el tratamiento de GnRH funciona bien en vacas lactando pero no tan bien en novillas. (Bo et al, 2007). Las causas de esta respuesta diferencial pueden explicarse en las características foliculares diferenciales entre vacas y novillas, que puede deberse a una duración menor de las ondas foliculares y presencia de más ciclos de 3 ondas en novillas comparado con las vacas donde su recambio folicular es mas lento y presentan más ciclos de 2 ondas foliculares. (Bo et al, 2007).
La perspectiva de cómo ha ido evolucionando el conocimiento y capacidad de controlar la reproducción en el ganado bovino, a llevado a realizar modificaciones del protocolo original Ovsynch, en base a la información que sugiere que la sincronización de la ovulación depende del estadío del ciclo estral en el que se le administró la primera dosis de GnRH. (Vasconcelos, 1999). Ya que aquellos animales en los que se inició el Ovsynch durante el metaestro, el folículo dominante puede que sea muy pequeño como para responder a la primera inyección de GnRH. Es necesario para esto, recordar que los folículos dominantes adquieren la cantidad necesaria de receptores de LH para su ovulación, después de haber sido seleccionados, cuando su diámetro es superior a 8.5 mm (Ginther, 1996). Por lo tanto luego de 7 días al momento de inyectar PGF2α, el folículo probablemente ya haya regresado y una nueva onda folicular esté emergiendo y para la segunda dosis de GnRH (48 horas después), habrá
otro folículo dominante pequeño que no responderá ovulando al tratamiento. Otro de los problemas que se presentan en el Ovsynch es la ovulación de un folículo pequeño, lo que resultaría en un menor tamaño del CL y una reducción en las tasas de preñez por las bajas concentraciones de progesterona. (Bo et al, 2007).
La Pre-sincronización con una o dos dosis (con intervalo de 14 días entre una y otra) de PGF2α y comenzando el protocolo Ovsynch de 12 a 14 días después de la última aplicación de PGF2α, mejora el porcentaje de preñez (Figura 9). Este programa combinado ha sido denominado "Presynch". En una revisión de trabajos en protocolos Ovsynch, se habla que mediante el uso de un tratamiento con PGF2α para la "pre-sincronización" aumentó la tasa de preñez entre 11 y 13 puntos, es decir que paso del 40% al 51-53% (LeBlanc, 2001)
Este protocolo es muy manejado en ganado de leche, iniciando la primera aplicación de Zincrocel® (D ( + ) C l o p r o s t e n o l 0.075mg/mL) el día 37 pos-parto, el día 51 se aplica la segunda dosis de Z i n c r o c e l ® ( D ( + ) Cloprostenol 0.075mg/mL); y se debe iniciar el Ovsynch el día 63 pos-parto con la aplicación
ya sea de Maxpren® (Acetato de Buserelina 0.0042 mg/mL) u Ovulen® (Acetato de Gonadorelina 0.1mg/mL), continuando 7 días después de esta aplicación con 2 mL de Zincrocel® (D (+) Cloprostenol 0.075mg/mL); 24 horas posteriores nuevamente Maxpren® (Acetato de Buserelina
0.0042 mg/mL) u Ovulen® (Acetato de Gonadorelina 0.1mg/mL) y 17 a
24 horas después IATF. (Figura 9).
30 31Con el fin de simplificar un poco el Ovsynch se desarrolló el protocolo Cosynch (Coordinated Synchonization), en el que se reduce de 4 a 3 el número de veces que se maneja el ganado al realizar la inseminación al mismo tiempo que se aplica la segunda dosis de GnRH. (Figura 10).
Figura 8: Protocolo Ovsynch(Ovulation Synchronization)
Figura 9: Protocolo Presynch(Presincronización)
Figura 10: Protocolo Cosynch(Coordinated Synchronization)

Se inicia el protocolo con la aplicación de 2.5 mL de Maxpren® (Acetato de Buserelina 0.0042mg/mL) o 1 mL de Ovulen® (Acetato de Gonadorelina 0.1mg/mL), lo que causa la ovulación del folículo dominante y el posterior desarrollo de una nueva onda folicular; 2.0 mL de Zincrocel® (D (+) Cloprostenol 0.075mg/mL) deben ser aplicados 7 días después, desencadenado la regresión del CL; y 2 días después de esta aplicación, se utiliza una segunda inyección de Maxpren® u Ovulen® a la misma dosis realizando la IATF el mismo día. (Figura 10).
Este protocolo es más utilizado en ganado de carne, aunque se puede manejar en ganado lechero, con o sin presincronización. Los resultados son similares al Ovsynch, siendo posible obtener 40 - 45% de gestación. Hay que tener en cuenta que en protocolos como el Ovsynch y Cosynch la fertilidad es mejor cuando se insemina a celo detectado (am-pm).(Asprón, 2004).
El método Selectsynch (Selective Synchronization) se desarrolló con base en los resultados obtenidos al inseminar a calor detectado las vacas sometidas a los tratamientos anteriores. El protocolo se maneja aplicando 2.5 mL de Maxpren® (Acetato de Buserelina 0.0042mg/mL) o 1 mL de Ovulen® (Acetato de Gonadorelina 0.1mg/mL) (causando la ovulación del folículo dominante y el posterior desarrollo de una nueva onda folicular); 2.0 mL de Zincrocel® (D (+) Cloprostenol 0.075mg/mL) (aplicados 7 días después), inicia la regresión del CL; y se insemina a celo detectado, haciendo la observación de celos desde las 24 - 48 horas anteriores a la aplicación del Zincrocel® y continuándose por 5 - 7 días (Figura 11). Si se presenta celo antes de la aplicación de Zincrocel® se debe suspender el tratamiento e inseminar (a.m. - p.m.).
Como algunas hembras no manifiestan el celo después del tratamiento la alternativa para estos casos es la de detectar celos hasta 48 - 60 horas después del Zincrocel®, inseminar las que manifiesten celo y las que no lo hicieron se inseminan a las 60 - 72 horas aplicándoles 2.5 mL de Maxpren® al momento de la I.A., convirtiendo el tratamiento en un Cosynch con unas horas más entre la prostaglandina y el servicio.
El Selectsynch es más sencillo y barato que el Ovsynch y Cosynch, pues solo 30-40% de las vacas reciben la segunda dosis de GnRH, pero requiere detección de celos. La mayoría de las vacas muestran calor 2 - 4 días después de la PGF2α, con una presentación de celos del 70-75% y fertilidad de 60-70% (mayor que en otros métodos por el hecho de inseminar en un mejor momento) obteniendo así un porcentaje de preñez del 40-45% (Asprón, 2004)
33
Figura 11: Protocolo Selectsynch(Selective Synchronization)

En el año 2001, Fernándes y Barros desarrollaron un nuevo esquema llamado Heatsynch (Sincronización de calores), que es similar al Ovsynch pero cambiando la segunda aplicación de GnRH por benzoato de estradiol (1 mg). (Figura 12).
El esquema es: 2.5mL de Maxpren® (Acetato de Buserelina 0.0042mg/mL) o 1mL de Ovulen® (Acetato de Gonadorelina), induciendo la ovulación del folículo dominante y desarrollo de una nueva onda folicular; la inyección de Zincrocel® (D (+) Cloprostenol 0.075mg/mL) 7 días después que causa la lisis CL; y para precisar la ovulación del folículo dominante se utiliza 1mL de Tradiovet® (Benzoato de estradiol 1mg/mL) 24 horas después del Zincrocel®, 30-34 horas después de la aplicación de Tradiovet® las vacas serían inseminadas sin necesidad de detección de celo. (Figura 12).
En el método Ovsynch la segunda GnRH estimula el pico de LH que provoca la ovulación; en condiciones fisiológicas la secreción de GnRH que va a causar el pico ovulatorio de LH se produce por retroalimentación del estradiol producido en el folículo dominante. La razón para utilizar el estradiol es que es mucho más barato que la GnRH, pero su efecto es más lento. "La GnRH provoca el pico de LH una hora después de su aplicación, mientras que el estradiol tarda 16 horas en ejercer el mismo efecto" (Bo et al, 2007). Por lo cual el Tradiovet® (Benzoato de estradiol 1mg/mL) se aplica solo 24 horas después del Zincrocel® (D (+) Cloprostenol 0.075mg/mL) (día 8) y la inseminación se hace hasta 30 - 34 horas después de inyectarlo. Un estudio en 53 vacas Nelore lactantes (60 a 90 días pos-parto) que estaban ciclando, resulto en una tasa de preñez del 43,3%. (Bo et al, 2007).
Métodos con progestágeno, estradiol, prostaglandina y estradiol (P.E.P.E)El empleo de progestágenos (progesterona o sus análogos) se fundamenta en la acción inhibitoria de la progesterona para la manifestación del celo. Al administrarlos en dispositivos vaginales (durante 7
días), implantes subcutáneos (9 días) o por vía oral (14 días) actúan como un CL artificial y mientras ejercen su acción la vaca no manifiesta estro y al retirarlos permiten la presentación del celo.
En la actualidad se utilizan los dispositvos intravaginales de progesterona. Existen diferentes dispositivos con progesterona: PRID (Sanofi), CIDR (Pfizer), DIB (Syntex), Cue-Mate (Bioniche) y los TRIU-B (Biogénesis), cada uno varia en la forma y concentración de progesterona, por ejemplo el CIDR contiene 1.38 g de progesterona y el DIB 1 g de progesterona (Ver figura 13).
Con la creación de estos dispositivos se han desarrollado una gran variedad de protocolos; al principio se utilizaban durante un periodo de 14 a 21 días logrando una buena sincronía de celos pero una baja fertilidad, por la formación de folículos persistentes.
Con el fin de inducir regresión luteal se combino el protocolo con benzoato de estradiol al momento de la inserción del dispositivo, utilizándose en ese momento una capsula de 10 mg. Posteriormente con el avance de la ciencia se combino la aplicación de PGF2α, pero los resultados obtenidos fueron variables ya que cuando se utilizaban en una fase luteal tardía (después del día 14) se reportaba una baja fertilidad. (Macmillan et al, 1998; Roche, 1974).En la actualidad se han desarrollado protocolos donde se sincronizan los celos utilizando los dispositivos intravaginales con progesterona combinados con: estradiol y prostaglandina. Este protocolo busca sincronizar el desarrollo folicular de forma que los animales tengan un folículo en crecimiento y con capacidad de ovular al momento de retirar el dispositivo. La aplicación de benzoato de estradiol más progestágeno en estudios realizados resulta en un comienzo sincrónico de una nueva onda folicular (en promedio 4.3 días) independiente del momento del ciclo estral en el cual se administre (Bó et al, 2007).
En estudios realizados en Nueva Zelanda, se recomienda inducir el pico preovulatorio de LH con la administración de estradiol, esto produce un feed-back positivo sobre la GnRH, y la dosis utilizada fue de 0.75 mg ó 1 mg de EB a las 24 horas de removido el CIDR. Los resultados obtenidos demostraron que el pico de LH se da en promedio a las 16,1 horas después de la aplicación del benzoato y la ovulación 40 horas después (64 horas promedio después de removido el dispositivo). (Fig. 14). Considerando que los espermatozoides tardan entre 8 a 12 horas en llegar al sitio de la fertilización se recomienda entonces realizar la inseminación entre las 52 y 56 horas de haber retirado el dispositivo (Bo et al, 2007).
En algunos protocolos se ha recomendado la utilización de cipionato de estradiol en vez de benzoato de estradiol para inducir la ovulación y reducir el número de veces que se maneja a los animales (administrándolo el día 7). Desde el punto de vista farmacológico la diferencia del cipionato con el benzoato de estradiol radica en que el primero tiene una vida media mayor y por lo tanto no se sincroniza adecuadamente la emergencia de una nueva onda folicular (Bo et al, 2007), es por esto que se prefiere utilizar estrógenos con una vida media corta como la del Tradiovet® (Benzoato de estradiol 1mg/mL).
A continuación se describen diferentes protocolos recomendados en vacas y novillas a celo detectado o a tiempo fijo:
34 35
Figura 14: Protocolo para Vacas - inseminación a tiempo fijo
Figura 13: Dispositivos intravaginales bovinos de progesterona.
Figura 12: Protocolo Heatsynch(Sincronización de calores)

Los porcentajes de preñez que se manejan en estos protocolos, varían entre un 60 - 70% (Bo et al, 2007). Este es el protocolo más utilizado en vacas que consiste en administrar 2 mg de Tradiovet® (Benzoato de estradiol 1mg/mL) al momento de la inserción del dispositivo en el Día 0, para de esta manera sincronizar el desarrollo folicular; y remover el dispositivo el día 7 junto con la administración de Zincrocel® (D (+) Cloprostenol 0.075mg/mL), para inducir la luteólisis; 24 horas después se administra 1mg de Tradiovet® (Benzoato de estradiol 1mg/mL) para sincronizar la ovulación y la IATF se realiza entre las 52 a 56 horas post-remoción del dispositivo, o 30 - 34 horas después de la última aplicación de Tradiovet® (Figura 14) (Bo et al, 1999). También se a optado por remover el dispositivo en el día 8 en lugar del día 7 y se han encontrado que resultan en porcentajes de preñez similares. (Bo et al, 2001). Esta posibilidad tiene una gran aplicación práctica, ya que se pude tratar un grupo grande de animales un mismo día y después variar el retiro del dispositivo para IATF en dos días distintos. Trabajos realizados por Macmillan proponen la utilización de tratamientos de 8 días en vacas lecheras en lactancia, basándose principalmente en la teoría de que al alargar el tratamiento por un día se permitirá un mayor crecimiento del folículo dominante. (Macmillan et al, 1999).
El mismo protocolo pero con la utilización de media dosis de BE también ha sido utilizado en novillas; pero con base en un experimento realizado por Bo y sus colaboradores (2007), en novillas cruza cebú para determinar el efecto de la utilización de media dosis de Benzoato de estradiol en el momento de colocar el dispositivo; consideraron que no se justifica recomendar la dosis menor a 2 mg; pues aunque no se encontraron diferencias significativas en los porcentajes de preñez al aplicar 1 ó 2 mg (54,84% vrs. 65.08% respectivamente), concluyen que el bajo costo del BE, utilizando 2 mg, puede aumentar de alguna forma estos resultados. (Figura 15). (Bo et al, 2007).
Otra alternativa es la utilización de GnRH después de removido el dispositivo. Este protocolo a mostrado en trabajos de campo una efectividad hasta de 76% (Anderson, 1999); por lo tanto, el tratamiento con 2 mg de Tradiovet® (Benzoato de estradiol 1mg/mL) el día de la inserción del dispositivo, 7 días después 2 mL de Zincrocel® (D (+) Cloprostenol 0.075mg/mL), y la aplicación de 2.5mL de Maxpren® (Acetato de Buserelina 0.0042mg/mL) en el momento de I.A. a celo detectado;es otra alternativa para sincronizar la ovulación (Figura 16). La opción de usar GnRH en lugar de Benzoato de estradiol, sería interesante, si queremos ahorrarnos un encierre de las vacas.
37
Realizando y experimentando varios cambios en protocolos con dispositivo intravaginal, se está manejado la utilización conjunta con GnRH; en donde se aplicarían junto con del dispositivo el día 0; 2.5mL de Maxpren® (Acetato de Buserelina 0.0042mg/mL) o 1mL de Ovulen® (Acetato de Gonadorelina); 7 días después se retira el dispositivo y se aplican 2 mL de de Zincrocel® (D (+) Cloprostenol 0.075mg/mL); inseminando a celo detectado, del día 8 al 10, aplicando Maxpren® u Ovulen® en el momento de la inseminación (Figura 17). Los tratamientos que utilizan Benzoato de Estradiol o GnRH en combinación con el dispositivo intravaginal liberador de P4, resultan en similares tasas de preñez. (Balla et al, 2006)
Figura 17: Protocolo en Novillas a celo detectado
Figura 16: Protocolo en Vacas combinando BE y GnRH e inseminando a celo detectado.
36
Figura 15: Protocolo para Novillas I.A.T.F.

39
Composición:Cada mL de Tradiovet® solución inyectable contiene:
Benzoato de estradiol 1 mg
Excipientes c.s.p. 1 mL
Descripción:Tradiovet® es un éster de estradiol, equivalente natural de la hormona sexual femenina estradiol (estrógenos). Los estrógenos son hormonas esteroideas secretados por las células de la granulosa del ovario estimuladas por la FSH, que favorece la conversión de andrógenos en estrógenos por medio de la expresión de la enzima P-450 aromatasa. La síntesis y secreción de estrógenos está regulada durante el ciclo estral por el efecto de las gonadotropinas.
Los estrógenos son hormonas sexuales encargadas de estimular y mantener el normal funcionamiento del tracto reproductivo de las hembras; estimulan el crecimiento, controlan la ovulación, preparan el aparato reproductor para la fecundación e implantación, y poseen efectos metabólicos sobre los minerales, los glucósidos, las proteínas y los lípidos.
Mecanismo de acción:Los estrógenos actúan por medio de la regulación de la expresión génica. Por tanto esta hormona atraviesa la membrana citoplasmática y se difunden en el citoplasma hasta unirse al receptor nuclear. Existen dos isoformas del receptor de estrógenos: el receptor de estrógenos β (ER β) y el receptor de estrógenos α (ER α), los cuales se ubican en diferentes cromosomas. Los estrógenos aumentan la síntesis celular de cromatina de ARN y de diversas proteínas
en los tejidos sensibles. El receptor de estrógenos β (ER β) se expresa en tejidos como el sistema nervioso central, sistema cardiovascular, sistema inmunitario, tracto urogenital, tracto gastrointestinal, riñones, pulmones y glándula mamaria, aunque en menor proporción ya que el receptor estrógenos α (ER α) predomina en este tejido y en el útero.
En el ciclo estral los estrógenos están involucrados activamente en el proceso reproductivo, ya que son necesarios para la liberación del pico preovulatorio de LH, este incremento de gonadotropina, causa ruptura del folículo y la liberación del ovocito. Su uso prolongado inhibe los centros hipotalámicos e hipofisiarios inhibiendo la producción de GnRH y FSH.
El estradiol es la fracción estrogénica activa fisiológicamente más importante en los rumiantes. Es producido por el folículo ovárico en la hembra, siguiendo un patrón cíclico y provocando los cambios de conducta observados durante el estro. En el macho es producido en pequeñas cantidades por las glándulas adrenales.
Los estrógenos en general parecen actuar a través de las diversas hormonas que controlan el metabolismo de la energía. Después del tratamiento de rumiantes con estrógenos se elevan las concentraciones plasmáticas de insulina y de hormona del crecimiento. Esto resulta en aumento de la síntesis de proteína muscular, lo que se evidencia por un incremento de la captación de aminoácidos. Por tanto, los efectos de los estrógenos no serían directos sobre la célula muscular, sino mediados a través principalmente de la hormona del crecimiento, resultando en un balance nitrogenado positivo, aumento de la deposición de proteínas y una mayor retención de calcio y fósforo.

40 41
Indicaciones:Tradiovet® está indicado para la inducción y sincronización del ciclo estral, participando de forma activa en el proceso reproductivo. Los estrógenos son necesarios para la liberación del pico preovulatorio de LH, con lo cual se causa la ruptura del folículo y la liberación del ovocito. Tradiovet® se administra para inducir el estro, la liberación de LH y la ovulación en vacas. En estudios de campo, se administra Tradiovet® al momento de insertar el Dispositivo intravaginal Bovino para sincronizar la emergencia de una onda folicular, y 24 horas después de retirar el dispositivo para sincronizar la ovulación (Martínez, et al 2007).
Tradiovet® está indicado en anestro o ausencia de celos por deficiencias de estrógenos, para aumentar los signos del celo, estro o calor en las hembras que presenten calores silentes.
Tradiovet® está indicado para la sensibilización del músculo uterino a la oxitocina para facilitar la acción de ésta en el tratamiento de retención placentaria, expulsión de fetos momificados y material purulento en caso de piómetra abierto, en estos casos los estrógenos tienen capacidad de incrementar el flujo de sangre, la excitabilidad del miometrio, la permeabilidad para los leucocitos y la secreción de inmunoglobulinas. Este tratamiento puede asociarse al uso de antibióticos y oxitócicos. En el posparto estimula la involución uterina.
Tradiovet® en perras y gatas, esta indicado para evitar la gestación como anticonceptivos tempranos,
por tener la capacidad de interrumpir el transporte de los óvulos o cigotos a través del oviducto impidiendo de esta forma la implantación y el inició de la preñez.
Tradiovet® se utiliza en la deficiencia de estrógenos, vaginitis atrófica, hipogonadismo femenino, insuficiencia ovárica primaria, hemorragia uterina inducida por desequilibrio hormonal.
Dosificación:Vacas:
Para la sincronización de la ovulación en los programas de inseminación artificial a tiempo fijo, aplicar 2 mg (2 mL) al momento de colocar el dispositivo de liberación controlada de progesterona, y 1 mg (1mL) a las 24 horas de retirarlo.
En anestro 3 - 5 mg (3 - 5 mL), piómetra 5 - 10 mg (5 - 10 mL), retención placentaria 5 - 10 mg (5 - 10 mL), fetos momificados 5 - 10 mg (5 - 10 mL) y cuerpo lúteo persistente 2.5 - 5 mg (2.5 - 5 mL).
Yeguas:
En la inducción del estro en yeguas con calor silente 1 mg (1 mL). Incremento de los mecanismos de defensa uterinos: 1 - 2 mg, (1 - 2 mL) diariamente durante 3 a 5 días seguidos.
Cerdas y ovejas:
5 - 10 mg ó 5 - 10 mL.
Perras:
Para evitar la gestación por montas indeseadas, 0.02 mg por Kg (0.5 mL x 25 Kg) dentro de las 72 horas siguientes a la monta y repetir la misma dosis a las 48 horas.
Gatas:
Para evitar la gestación por montas indeseadas, 0.125 - 0.25 mg (0.125 - 0.25 mL dosis total) dentro de las 40 horas siguientes a la monta y repetir la misma dosis a las 48 horas.
Vía de administración:Aplicación intramuscular.
Contraindicaciones:No aplicar en hembras gestantes ni en machos, no aplicar en
animales sexualmente inmaduros , n i en
animales con signos clínicos de anemia.
Precauciones:Las que el Médico Veterinario considere de acuerdo al estado del tracto reproductivo de la hembra, no exceder la dosis prescrita por el Médico Veterinario.
La respuesta de los tratamientos hormonales es individualizada por lo tanto queda a criterio del Médico Veterinario establecer la dosis requerida.
Tiempo de Retiro:Los animales tratados no deben sacrificarse para el consumo humano hasta 5 días después de finalizado el tratamiento. La leche producida durante el tratamiento y 72 horas después de finalizado el mismo no debe darse al consumo humano.
Presentacion comercial:Frasco vial de vidrio con agrafe Flip Off por 20 y100 mL.

42 43
Conclusiones:1. Los folículos del ovario del bovino se desarrollan en ondas. Una onda de crecimiento folicular, está constituida por un gran número de folículos que se desarrollan al mismo tiempo, seguido de la selección y crecimiento del folículo dominante y la regresión o atresia de los folículos subordinados. En ausencia de luteólisis, el folículo dominante dejará de crecer e iniciará la regresión, dando lugar al crecimiento de una nueva onda folicular. Se ha demostrado que existen 2 o 3 ondas del desarrollo folicular durante el ciclo estral.
2. El desarrollo folicular está controlado por la secreción de hormonas provenientes de la hipófisis, el CL y de los folículos. Se ha observado que cada onda folicular está siempre precedida por un pico de FSH. Se cree que este pico es fundamental para que los folículos antrales ingresen al pool de folículos grandes (> 4 mm) que observamos en la onda folicular. Luego del comienzo de la onda el folículo dominante comienza a producir grandes cantidades de estradiol e inhibina que actúan inhibiendo la liberación de FSH. Con poca FSH circulante los folículos subordinados comienzan a regresar. El folículo dominante puede seguir creciendo debido a la adquisición de receptores de LH en las células de la granulosa (además de los que se encuentran en las de la teca). Durante el diestro, los altos niveles de progesterona afectarán adversamente la frecuencia de los pulsos de LH e inducirán la regresión del folículo dominante.
3. La propiedad luteolítica de la prostaglandina la hace y la hará por mucho tiempo imprescindible en los tratamientos para sincronizar los ciclos estrales en bovinos. Sin embargo la necesidad de perfeccionar los tratamientos de sincronización con PGF deja claro que para lograr una precisa sincronización sería necesario trabajar no sólo sobre la función del cuerpo lúteo, sino también sobre la dinámica folicular.
4. Los avances logrados en cuanto a la compresión de los mecanismos neuroendocrinos de regulación del ciclo estral, han permitido profundizar el conocimiento de la dinámica folicular y luteal, y la disponibilidad de hormonas sintéticas naturales o análogos a nivel comercial, han logrado realizar tratamientos de sincronización de celos que permiten la IATF, siendo esta una interesante alternativa cuando la detección de celos es un problema serio de manejo.
5. Para obtener una máxima fertilidad en los esquemas de IATF es necesario controlar el CL, el desarrollo folicular y la ovulación. Para controlar la fase luteal se puede inducir la regresión del CL con PGF o estrógenos, o prolongarla mediante dispositivos con liberación lenta de progestágenos. Para controlar el desarrollo folicular se puede utilizar estrógenos más progestágenos que suprimen. Mediante un feed-back negativo sobre las gonadotrofinas, el desarrollo folicular y resulta en el comienzo de una nueva onda folicular.
Referencias Bibliograficas:
" ASPRÓN, M.A. Curso de actualización-Manejo reproductivo del Ganado Bovino. IVIS. 2004. En Internet: www.ivis.org.
" ADAMS GP, MATTERI RL, KASTELIC JP, KO JC, GINTHER OJ. Association between surges of FSH and the emergence of follicular waves in heifers. J Reprod Fertil 1992;94:177-88.
" ADAMS GP, PIERSON RA. Bovine model for study of ovarian follicular dynamics in humans. Theriogenology 1995;43: 113-20.
" ADAMS GP. Comparative patterns of follicle development and selection in ruminants. J Reprod Fertil Suppl 1999;54:17-32.
" ADAMS GP. Control of ovarian follicular wave dynamics in mature and prepubertal cattle for synchronization and superstimulation. In: roceedings of the XX congress of the world association for buiatrics, vol. 2; 1998. p. 595-605.
" ADAMS, G.P. JAISWAL, R. SINHG, J. MALHI, P. Progress in understanding ovarian follicular dynamics in cattle. Theriogenology. Vol. 69. p: 72-80. 2008,
" AHMAD N, TOWNSEND EC, DAILEY RA, INSKEEP EK. Relationship of hormonal patterns and fertility to occurrence of two or three waves of ovarian follicles, before and after breeding, in beef cows and heifers. Anim Reprod Sci 1997;49:13-28.
" ALLEN, WR. Luteal deficiency and embryo mortality in the mare. Reproduction Domestic Animal. 2001. Vol. 36. p: 121-131.
" ALVES. Pulsatile secretion of gonadotrophins, ovarian steroids and ovarian oxytocin during the luteal phase of oestrus cycle in the cow. J.Reprod Fert,. 479 - 491.
" ANDERSON J. Resultados de trabajos util izando CIDR. Segundo Workshop de Repoducción Bovina Boehringer Ingelheim S.A., Tandil, 1999; 1-17
" BALLA E. Control of the bovine estrous. 2006.
" BAO B, GARVERICK HA. Expression of steroidogenic enzyme and gonadotropin receptor genes in bovine follicles during ovarian follicular waves: a review. J Anim Sci 1998;76:1903-21.
" BARRAGRY, T. Veterinary Drug Therapy. Lea & Febiger. 1994. p 696-700.
" BARUSELLI, P,S,;REIS, E.L.; MARQUES M.O. Técnicas de manejo para optimizar a eficiencia reprodutiva em femeas bos indicus. Grupo de estudio de nutricao de rumiantes - Departamento de melhoramiento e nutriciao animal FCA -FMVZ - Unesp Botucatu, Sao Paulo, 2004.
" BERETTA, C. CAVALLI M. A sheer pharmacologic approach to compare the contractile effects of PGF2a , DL-cloprostenol y D-cloprostenol on isolates uterine, trácheal, ileal and arterial smooth muscle preparations. Theriogenology. 2004, 62: 837-846.
" BERGFELT DR, LIGHTFOOT KC, ADAMS GP. Ovarian synchronization following ultrasound-guided transvaginal follicle ablation in heifers. Theriogenology 1994;42:895-907.
" BLEACH ECL, GLENCROSS RG, KNIGHT PG. Association between ovarian follicle development and pregnancy rates in dairy cows undergoing spontaneous oestrous cycles. Reproduction 2004;127:621-9.
" BO G. ALONSO A, CACCIA M, CARCEDO J, CUTAIA L., MORENO D. Manual de Sincronización de celos e inseminación artificial. Especilización en Reproducción Bovina. Instituto de Reproducción Animal Córdoba (IRAC). 2007.
" BO G. ALONSO A, CACCIA M, CARCEDO J, CUTAIA L., MORENO D. Manual de Tansferencia de Embriones y nuevas Tecnologías. Especilización en Reproducción Bovina. Instituto de Reproducción Animal Córdoba (IRAC). 2007.
" BO G. Dinámica Folicular y tratamientos hormonales para sincronizar la ovulación en el ganado bovino. Memorias. XI Congreso Venezolano de Producción e industria animal. ULA. Trujillo. 2002. p:1-17.
" BO G., ILLUMINANTE H., , MEGLIA G., SCHMIDT E., BARTOLOMÉ J., 1999.
" BÓ G.. ALONSO A, CACCIA M, CARCEDO J, CUTAIA L., MORENO D. Manual de Fisiología de la reproducción de la vaca. Especilización en Reproducción Bovina. Instituto de Reproducción Animal Córdoba (IRAC). 2007.
" BO GA, BARUSELLI PS, MARTINEZ MF. Pattern and manipulation of follicular development in Bos indicus cattle. Anim Reprod Sci 2003;78:307-26.
" BOTANA, L.M. LANDONI, F. MARTÍN, T. Farmacología y terapéutica veterinaria. Ed. Mc. Graw Hill. 2002. p:411-434
" BRAW-TAL R, YOSSEFI S. Studies in vivo and in vitro on the initiation of follicle growth in the bovine ovary. J Reprod Fertil 1997;109:165-71.
" BURGOS, R. HANCKE, J. Estrógenos, andrógenos y progetágenos. Capítulo 31 y DE LA SOTA, R.L, SOTTO A.T. GOBELLO M.C. Capítulo 32. En: BOTANA, L.M. LANDONI, F. MARTÍN-JIMÉNEZ, T. Farmacología y Terapéutica Veterinaria. Editorial Mc. Graw Hill. 2002. p. 411-433.
" BURNS DS, JIMENEZ-KRASSEL F, IRELAND JL, KNIGHT PG, IRELAND JJ. Numbers of antral follicles during follicular waves in cattle: evidence for high variation among animals, very high repeatability in individuals, and an inverse association with serum follicle-stimulating hormone concentrations. Biol Reprod 2005;73:54-62.
" DAELS, P. Progesterone therapy and pregnancy loss. 8th. AAEP. Annual Resort Symposium. Rome, Italy - January 19-21, 2006.
" DRIANCOURT, M.A. Regulation of ovarian folicular dynamics in farm animals. Implications for manipulation of reproduction. Therogenology . 1211 - 1239, 2001
" ENGELHARDT BH. Control of follicular growth and developmente Reprod Fert. 2002.
" ERICKSON BH. Development and radio-response of the prenatal bovine ovary. J Reprod Fertil 1966;10:97-105.
" ERICKSON BH., Ovarian follicular and corpus luteum changes, progesterone concentrations, estrus and ovulation following estradiol benzoate/progesterone based treatment protocol in cross-bred cows. Animal Reproduction Science. 2007.
" ESPINOZA, J.L. ORTEGA, R., PALACIOS, A., VLAENCIA, J. ARÉCHIGA, C. Crecimiento folicular ovárico en animales domésticos: Una revisión. Iterciencia. 2007. Vol. 32. No. 2. p 93-99.
" FIELDEN ED, HARRIS RE, MACMILLAN KL, SHRESTHA SL. Some aspects of reproductive performance in selected town-supply dairy herds. NZ Vet Journal 1980;131/132:141-2.
" FRICKE PM, AL-HASSAN MJ, ROBERTS AJ, REYNOLDS LP, REDMER DA, FORD JJ. Effect of gonadotropin treatment on size, number, and cell proliferation of antral follicles in cows. Domest Anim Endocrinol 1997;14:171-80.
" GIGLI, I. RUSSO, A. AGÜERO, A. Consideraciones sobre la dinámica ovárica en equino, bovino y camélidos sudamericanos. In. Vet. 2006. Vol. 6. 183-204.
" GINTHER OJ, BERGFELT DR, KULICK LJ, KOT K. Selection of the dominant follicle in cattle: role of two-way functional coupling between follicle-stimulating hormone and the follicles. Biol Reprod 2000;62:920-7.
" GINTHER OJ, KASTELIC JP, KNOPF L. Intraovarian relationships among dominant and subordinate follicles and the corpus luteum in heifers. Theriogenology 1989;32:787-95.
" GU¨MEN A, WILTBANK MC. An alteration in the hypothalamic action of estradiol due to lack of progesterone exposure can cause follicular cysts in cattle. Biol Reprod 2002;66:1689-95.
" ITOH T, KACCHI M, ABE H, SENDAI Y, HOSHI H. Growth, antrum formation, and estradiol production of bovine preanatral follicles cultured in a serum-free medium. Biol Reprod 2002;67: 1099-105.
" JAISWAL R. Regulation of follicular wave patterns in cattle. PhD Thesis. University of Saskatchewan; 2007. p. 169. [30] Salamone DF, Adams GP, Mapletoft RJ. Changes in the cumulus- oocyte complex of subordinate follicles relative to follicular wave status in cattle. Theriogenology 1999;52:549-61.
" JAISWAL RS, SINGH J, ADAMS GP. Developmental pattern of small antral follicles in the bovine ovary. Biol Reprod 2004;71:1244- 51.
" JAISWAL RS, SINGH J, MARSHALL L, ADAMS GP. Repeatability of 2- and 3-wave patterns during the bovine estrous cycle. Theriogenology; submitted for publication.
" JAISWAL RS, SINGH J, NAGRA HS, GRAFTON T, RATTO MH, MALHI PS, et al. Oocyte competence under different progestational environments. Biol Reprod 2006;109:166.
" KINDAHL H, EDQVIST LE, GRANSTROM E, BANE A. The release of prostaglandin F2a as reflected by 15-keto-13, 14-dihydroprostaglandin F2a in the peripheral circulation during normal luteolysis in heifers. Prostaglandins 1976;11:871-6.
" LEBLANC S. The Ovsynch Breeding Program for Dairy Cows - A Review and Economic Perspective. The Bovine Practitoner 2001: 35:13-21
" LOPEZ-GATIUS F, SANTOLARIA P, YANIZ J, RUTLLANT J, LOPEZ-BEJAR M. Persistent ovarian follicles in dairy cows: a therapeutic approach. Theriogenology 2001;56:649-59.
" LUSSIER JG, MATTON P, DUFOUR JJ. Growth rates of follicles in the ovary of the cow. J Reprod Fertil 1987;81:301-7.
" MACMILLAN KL., COLSON DD., EAGLES VM. Modifications to improve whole herd synchrony programs in seasonal dairy herds. Proc Autralian Assoc of Cattle Vet; 1999; 121-129.
" MARKUSFELD O. Inactive ovaries in high-yielding dairy cows before service: etiology and effect on conception. Vet Rec 1987;121:149-53.
" MARTIN TL., FOGWELL RL., IRELAND JJ., Concentrations of inhibins and steroids in follicular fluid diruing development of dominant follicles in heifers. Biol Reprod 1991; 44:693-700.
" MATEUS L, DA COSTA LL, BERNARDO F, SILVA JR. Influence of puerperal uterine infection on uterine involution and postpartum ovarian activity in dairy cows. Reprod Domest Anim 2002;37:31-5.
" MELQUISEDEC A., Oogenesis in cattle. 2004.
" MWAANGA ES, JANOWSKI T. Anoestrus in dairy cows: causes, prevalence and clinical forms. Reprod Domest Anim 2000;35: 193-200.
" OPSOMER G, GROHN YT, HERTL J, CORYN M, DELUYKER H, DE KRUIF A. Risk factors for post partum ovarian dysfunction in high producing dairy cows in Belgium: a field study. Theriogenology 2000;53:841-57.
" PETER AT, VOS PLAM, AMBROSE DJ. Postpartum anestrus in dairy catte. Theriogenology 2009;71:1333-42.
" PLUMB, D. Veterinary drug handbook. Fourth Edition. Iowa State Press. 2002. p. 203-204.
" RAJAKOSKI E. The ovarian follicular system in sexually mature heifers with special reference to seasonal, cyclical, and left-right variations. Acta Endocrinol 1960;34:7-68.
" RICHARDS JS, MIDGLEY AR. Protein hormone action: a key to understanding ovarian follicular and luteal cell development. Endocrinology 1976;98:929-34.
" SHELDON IM, WATHES DC, DOBSON H. The management of bovine reproduction in elite herds. Vet J 2006;171:70-8.
" SINGH J, BROGLIATTI GM, CHRISTENSEN CR, ADAMS GP. Active immunization against follistatin and its effect on FSH, follicle development and superovulation in heifers. Theriogenology 1999;52:49-66.
" SINGH J, DOMINGUEZ M, JAISWAL R, ADAMS GP. A simple ultrasound test to predict superovulatory response in cattle. Theriogenology 2004;62:227-43.
" TANAKA Y, NAKADA K, MORIYOSHI M, SAWAMUKAI Y. Appearance and number of follicles and change in serum FSH in female bovine fetuses. Reproduction 2001;121:777-82.
" TOWNSON DH, TSANG PCW, BUTLER WR, FRAJBLAT M, GRIEL JR LC, JOHNSON CJ, et al. Relationship of fertility to ovarian follicular waves before breeding in dairy cows. J Anim Sci 2002;80: 1053-8.
" VASCONCELOS JLM., SILCOX RW., ROSA GJM., PURSLEY JR., WILTBANK MC., Synchronization rate, size of the ovulatory follicle, and pregnancy rate after
" VASSENA R, MAPLETOFT RJ, ALLODI S, SINGH J, ADAMS GP. Morphology and developmental competence of bovine oocytes relative to follicular status. Theriogenology 2003;60: 923-32.
" WANDJI SA, FORTIER MA, SIRARD MA. Differential response to gonadotrophins and prostaglandin E2 in ovarian tissue during prenatal and postnatal development in cattle. Biol Reprod 1992;46:1034-41.
" WILTBANK M, GUMEN A, SARTORI R. Physiological classification of anovulatory conditions in cattle. Theriogenology 2002;57: 21-52.
" ZEMJANIS R. Anestrus in cattle. In: Morrow DA, editor. Current therapy in theriogenology. WB Saunders: Philadelphia; 1980. p. 193-9.





























